Embed Size (px)
Citation preview

Cancer Biology Course
Tumor suppressor and Apoptosis:
Master Guardian and Executioner
本簡報內容部份取材自Garland Science, Taylor &
Francis Group 出版社所出版之The Biology of
Cancer ,僅供本課程教學使用,除上課目的外,
請勿以任何形式使用全部或部分內容
徐欣伶 (Hsin-Ling Hsu)
國家衛生研究院
03/31/2016

Cancer Pathogenesis Bozcuk et al., Breast Cancer Res Treat. 2001. 68(3):239-48.
• Hyper-activation of oncogenes
• Dysfunction of suppressor genes
• Loss of cell differentiation
• Augmentation of proliferate activity
• Alteration of hormone receptor status
• Increment of metastatic potential

Cell cycle progression
Differentiation
Apoptosis
Senescence
Genomic instability
Tumorigenesis
Oncogene
(proliferation) Tumor suppressor
(prevention)

Oncogene
(proliferation)
Cell growth
Differentiation
Apoptosis
Senescence
Genomic instability
Tumorigenesis
Tumor suppressor
(prevention)

p53 was identified in 1979 by Lionel Crawford, David P. Lane, Arnold
Levine, and Lloyd Old, It had been hypothesized to exist before as the target
of the SV40 virus, a strain that induced development of tumors.
The TP53 gene from the mouse was first cloned by Peter Chumakov of the
Russian Academy of Sciences in 1982, and independently in 1983 by Moshe
Oren (Weizmann Institute).
The human TP53 gene was cloned in 1984.
p53 was initially presumed to be an oncogene due to the use of mutated
cDNA following purification of tumor cell mRNA. Its character as a tumor
suppressor gene was finally revealed in 1989 by Bert Vogelstein working at
Johns Hopkins School of Medicine.
p53 is “The Guardian Angel of the Genome”-is a gene that, when a cell's
DNA is damaged, acts as an "emergency brake" to halt the resulting cycle of
cell division that can lead to tumor growth and cancer.
Discovery of p53
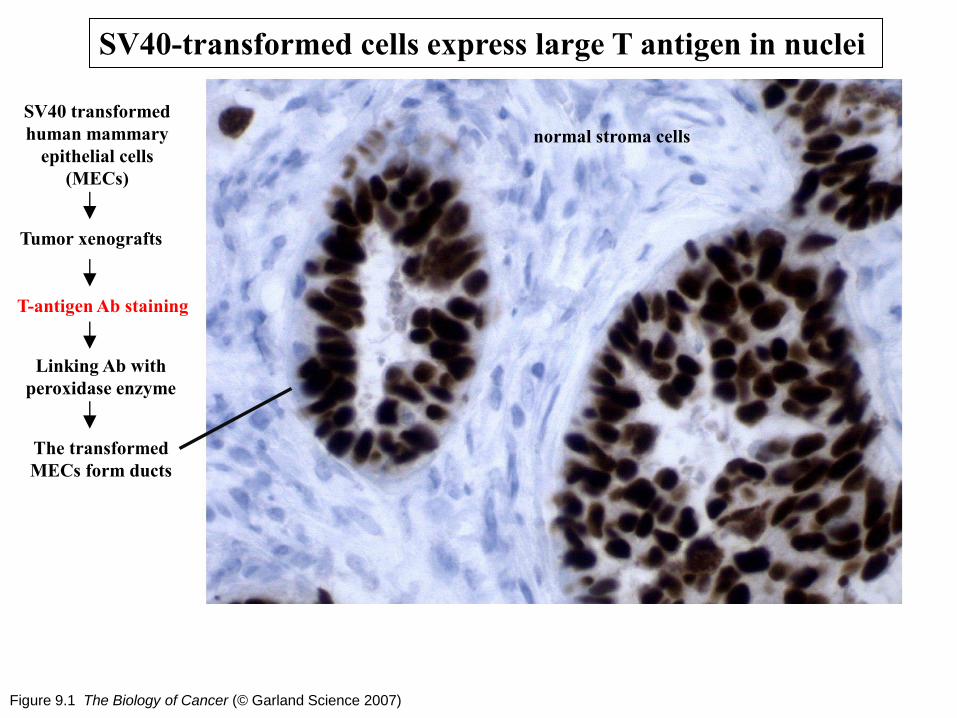
Figure 9.1 The Biology of Cancer (© Garland Science 2007)
SV40-transformed cells express large T antigen in nuclei
SV40 transformed
human mammary
epithelial cells
(MECs)
T-antigen Ab staining
Linking Ab with
peroxidase enzyme
Tumor xenografts
The transformed
MECs form ducts
normal stroma cells
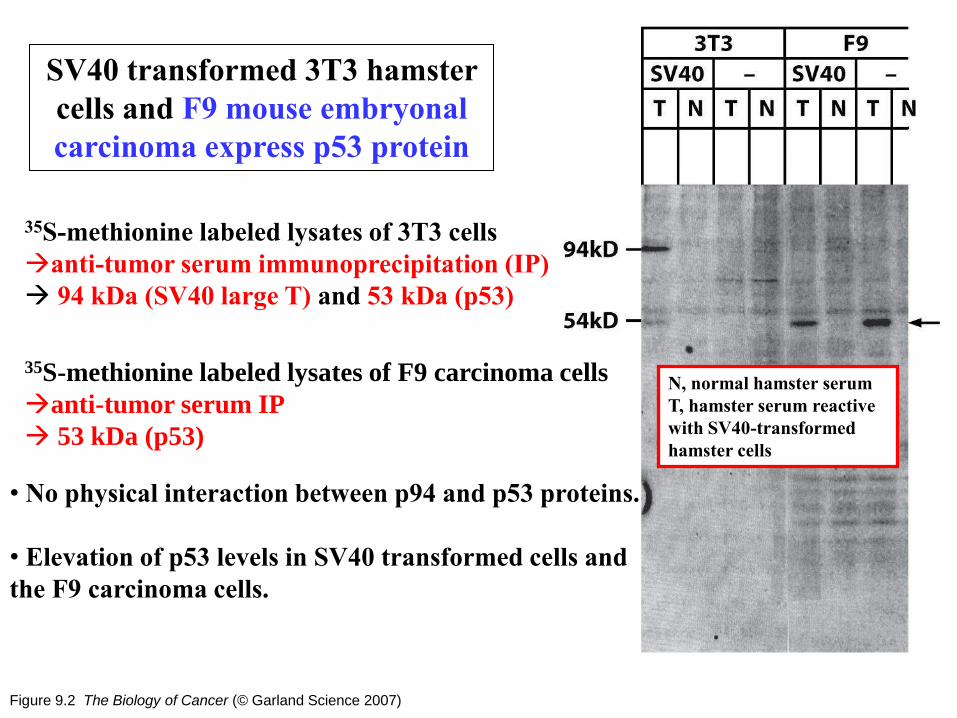
Figure 9.2 The Biology of Cancer (© Garland Science 2007)
SV40 transformed 3T3 hamster
cells and F9 mouse embryonal
carcinoma express p53 protein
35S-methionine labeled lysates of 3T3 cells
anti-tumor serum immunoprecipitation (IP)
94 kDa (SV40 large T) and 53 kDa (p53)
35S-methionine labeled lysates of F9 carcinoma cells
anti-tumor serum IP
53 kDa (p53)
• No physical interaction between p94 and p53 proteins.
• Elevation of p53 levels in SV40 transformed cells and
the F9 carcinoma cells.
N, normal hamster serum
T, hamster serum reactive
with SV40-transformed
hamster cells

Table 9.1 The Biology of Cancer (© Garland Science 2007)

Figure 9.12 The Biology of Cancer (© Garland Science 2007)
Specialized domains of p53
and the consensus p53-binding DNA sequence
p53-IP assays identify 452 sites in the human genome which binds p53
p53 phosphorylation blocks MDM2
binding and saves p53 from
ubiquitylation and degradation
Immunoprecipitation
of p53-DNA complex

Figure 9.35 The Biology of Cancer (© Garland Science 2007)
Multiple types of post-translational modifications of p53

Anti-cancer function of p53
• The normal p53 works through several mechanisms:
--- Activates DNA repair proteins in the response to DNA damage
--- Induces growth arrest by holding the cell cycle at the G1/S checkpoint
--- Allow DNA repair lesions to be fixed before returning to the cell cycle
--- Initiates apoptosis, if the DNA damage is irreparable
--- Mitotic aberrations and chromosomal missegregation
• The mutation of p53 can lose its suppressive powers or have the devastating
effect of actually promoting abnormal cell growth
• Discovering a compound that could restore function to a mutant p53 may
benefit to the development of anti-cancer drugs
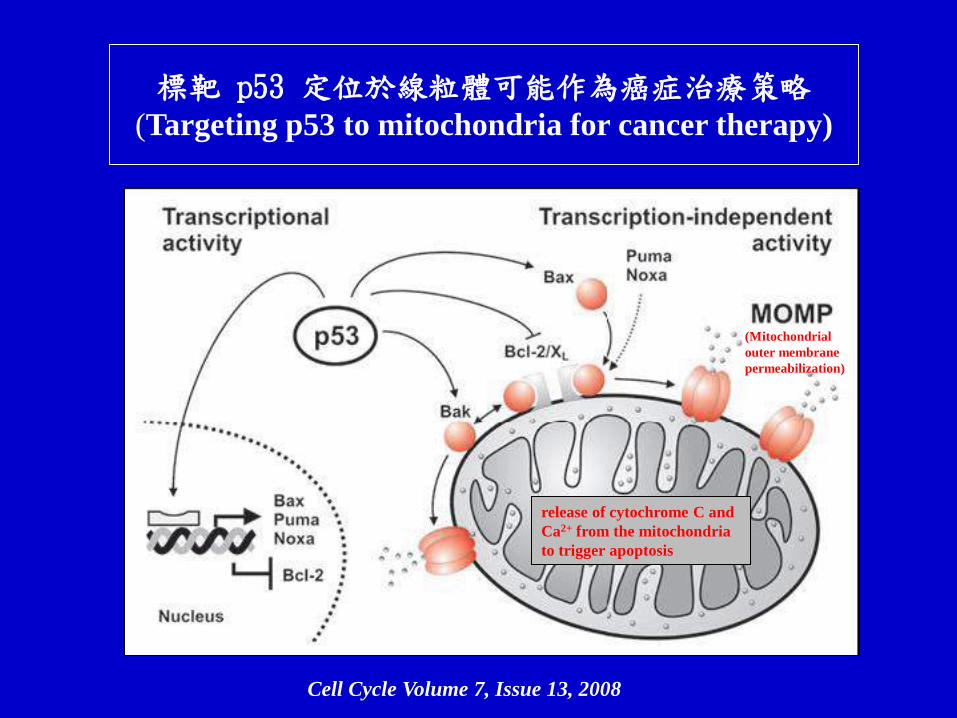
標靶 p53 定位於線粒體可能作為癌症治療策略 (Targeting p53 to mitochondria for cancer therapy)
Cell Cycle Volume 7, Issue 13, 2008
release of cytochrome C and
Ca2+ from the mitochondria
to trigger apoptosis
(Mitochondrial
outer membrane
permeabilization)
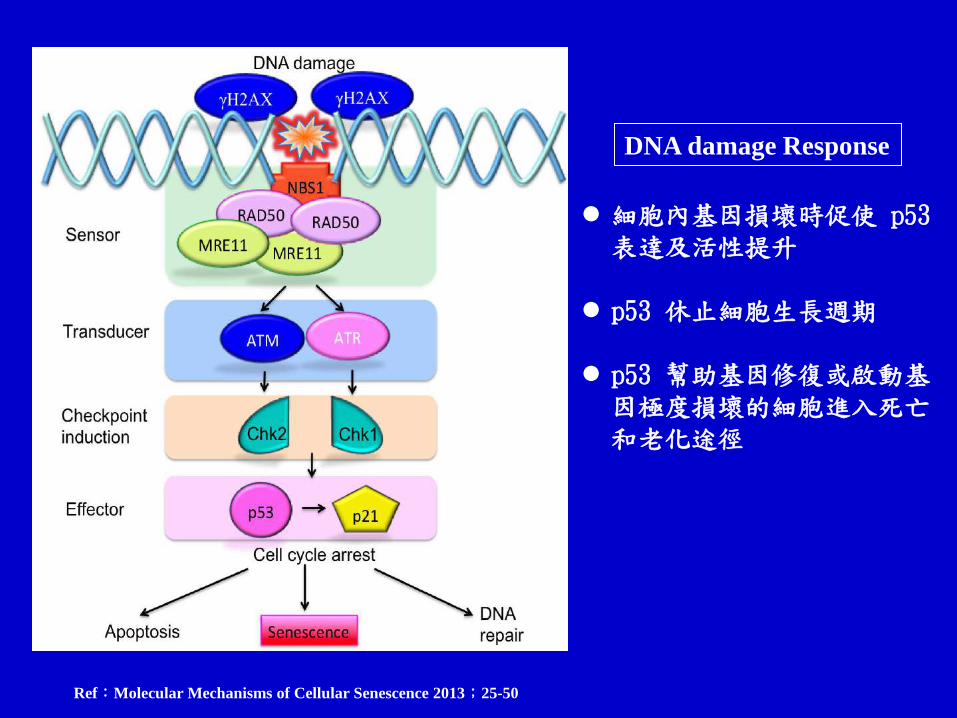
Ref:Molecular Mechanisms of Cellular Senescence 2013;25-50
細胞內基因損壞時促使 p53 表達及活性提升
p53 休止細胞生長週期 p53 幫助基因修復或啟動基
因極度損壞的細胞進入死亡和老化途徑
DNA damage Response

Figure 9.8 The Biology of Cancer (© Garland Science 2007)
p53-activating signals and p53 downstream effects
Under the cell-physiologic stress
• p53 accumulation
• p53 post-translational modification
• p53 oligomerization
• induces a number of cellular responses
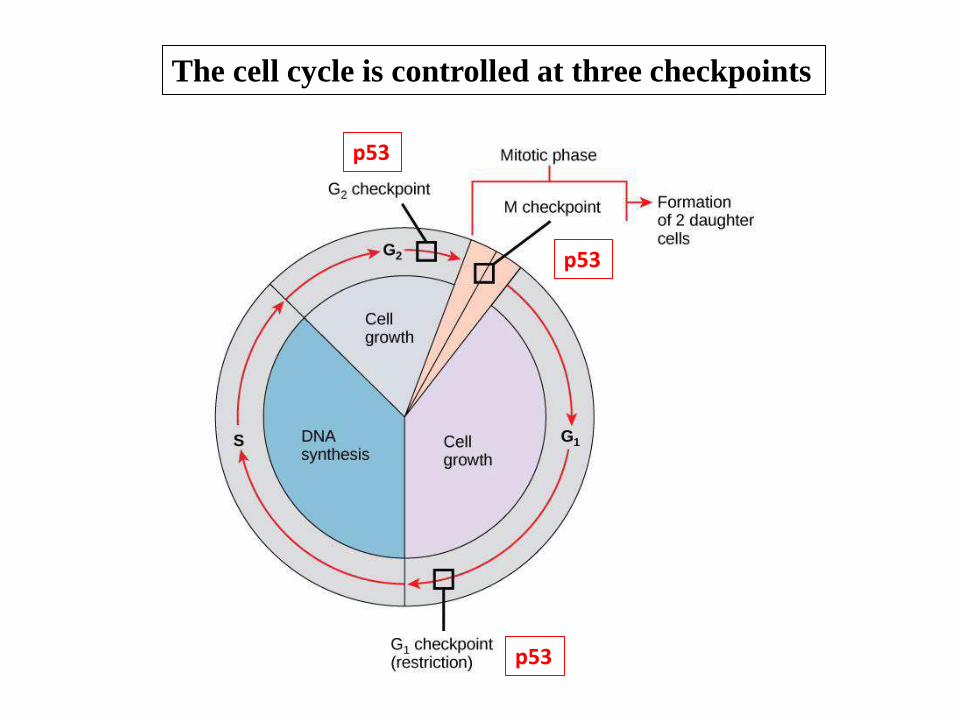
The cell cycle is controlled at three checkpoints
p53
p53
p53
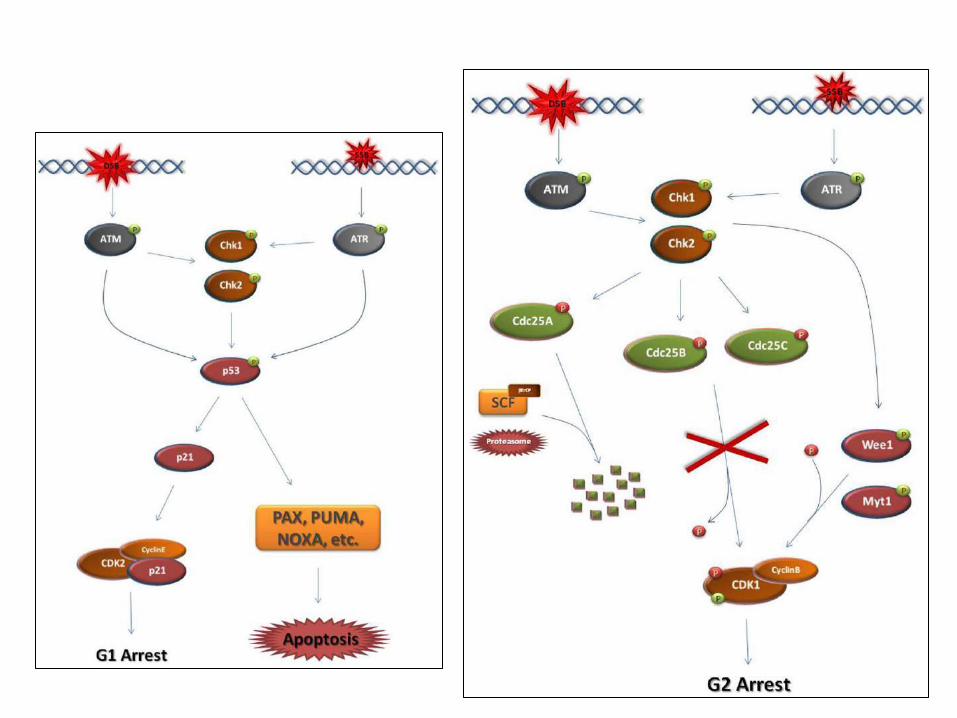
The major steps of the G1 & G2 DNA damage checkpoint
Front Genet. 2011; 4: 117.

細胞試驗:
p53 轉換癌症細胞趨向正常細胞之特性
高表現 p53 降低細胞內過氧化物的生成
提升細胞內過氧化物促使癌細胞的爬行能力和侵襲能力
調節 p53 活性能抑制癌細胞轉移和侵犯能力
Oncotarget 2014; 5(18):8452-65.
動物試驗與臨床證據:
動物試驗發現,p53 能抑制癌細胞的轉移並降低癌細胞侵襲血管的能力
從癌症患者檢體中發現 ROS 的生成增強癌細胞的侵襲能力
了解 p53 在抑制癌症形成的新角色,將利於針對 p53 有突變的癌症患者
選擇一個最有效的治療策略。
p53 在線粒體(製造能量)扮演重要功能
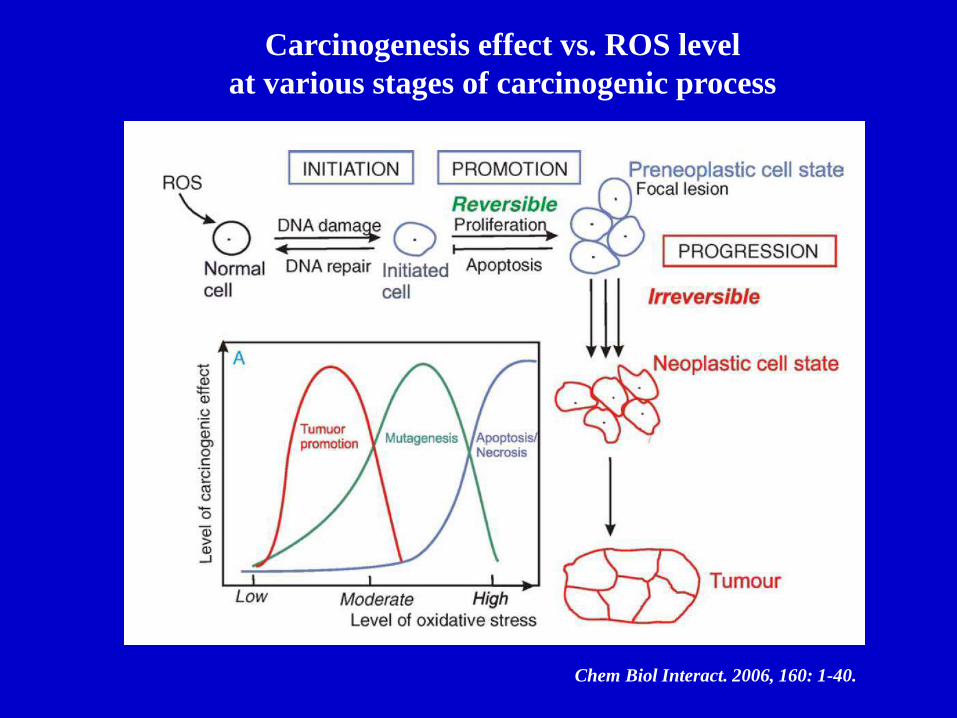
Chem Biol Interact. 2006, 160: 1-40.
Carcinogenesis effect vs. ROS level
at various stages of carcinogenic process

Ageing Research Reviews, 12: 376 – 390, 2013 Tumor-associated
Macrophage (M2)
Tumor microenvironment (腫瘤微環境) influences
angiogenesis (血管生成) and tumor progression (腫瘤增生)

CCL2/MCP-1 and IL-6 affect tumor progression (腫瘤進展)
and microenvironment (腫瘤微環境)
J Biol Chem.
2009, 284(49):34342-54

Table 9.2 The Biology of Cancer (© Garland Science 2007)
+
-
+
+
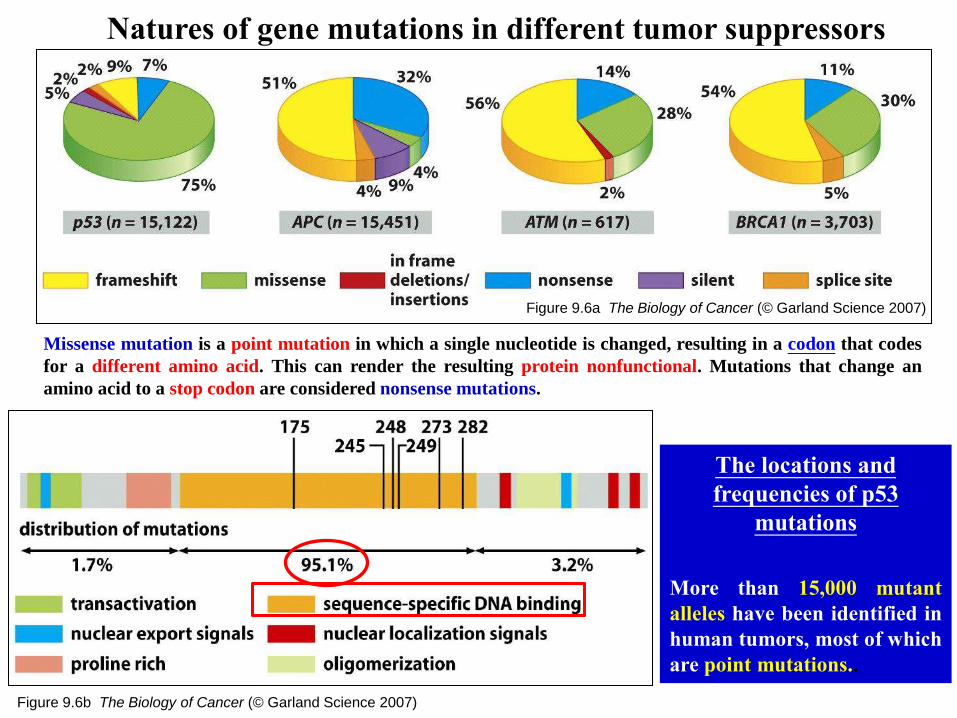
Figure 9.6a The Biology of Cancer (© Garland Science 2007)
Natures of gene mutations in different tumor suppressors
Figure 9.6b The Biology of Cancer (© Garland Science 2007)
The locations and
frequencies of p53
mutations
More than 15,000 mutant
alleles have been identified in
human tumors, most of which
are point mutations..
Missense mutation is a point mutation in which a single nucleotide is changed, resulting in a codon that codes
for a different amino acid. This can render the resulting protein nonfunctional. Mutations that change an
amino acid to a stop codon are considered nonsense mutations.

Figure 9.4 The Biology of Cancer (© Garland Science 2007)
Frequency of mutant p53 alleles in human tumor cell genomes
17,689 somatic mutations
and 225 germ-line mutations
IARC release data, 2002

Figure 9.17 The Biology of Cancer (© Garland Science 2007)
Accumulation of p53 in p53-mutant epithelial cells
of ovarian carcinoma
(ovarian surface epithelial)
(abnormality of development cells, early neoplastic process)
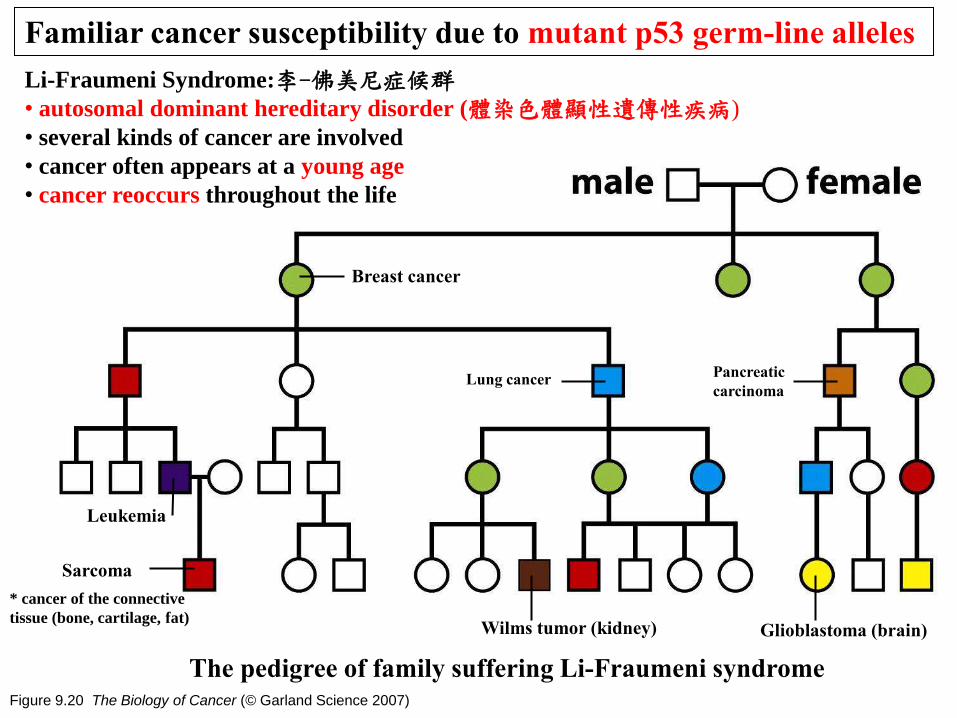
Figure 9.20 The Biology of Cancer (© Garland Science 2007)
Familiar cancer susceptibility due to mutant p53 germ-line alleles
The pedigree of family suffering Li-Fraumeni syndrome
Breast cancer
Sarcoma
Lung cancer
Leukemia
Pancreatic
carcinoma
Wilms tumor (kidney) Glioblastoma (brain)
Li-Fraumeni Syndrome:李-佛美尼症候群 • autosomal dominant hereditary disorder (體染色體顯性遺傳性疾病)
• several kinds of cancer are involved
• cancer often appears at a young age
• cancer reoccurs throughout the life
* cancer of the connective
tissue (bone, cartilage, fat)
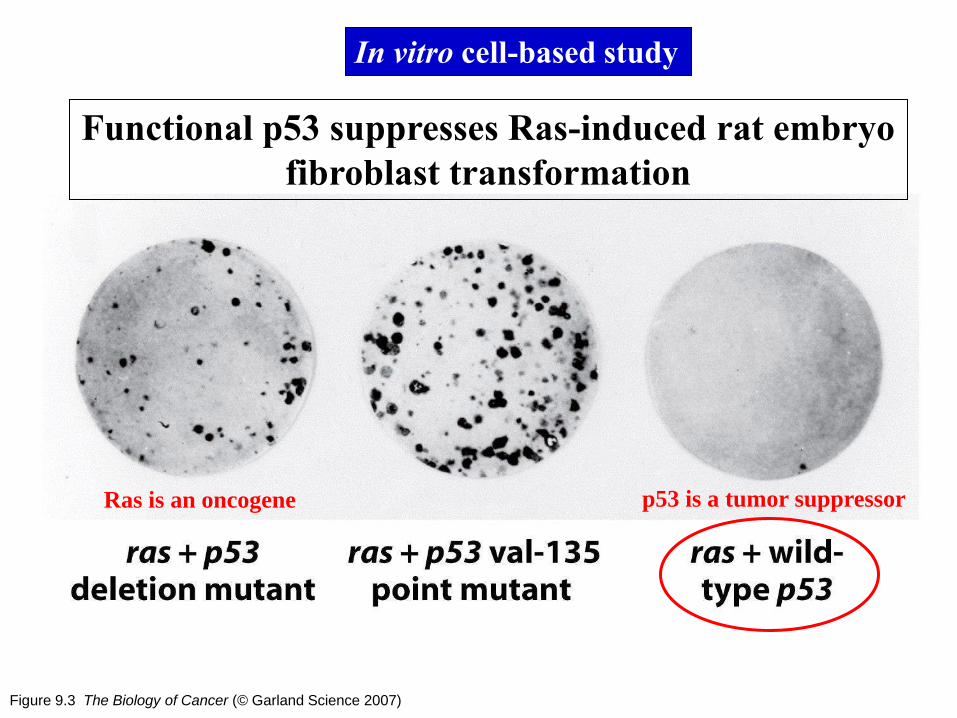
Figure 9.3 The Biology of Cancer (© Garland Science 2007)
Functional p53 suppresses Ras-induced rat embryo
fibroblast transformation
In vitro cell-based study
Ras is an oncogene p53 is a tumor suppressor
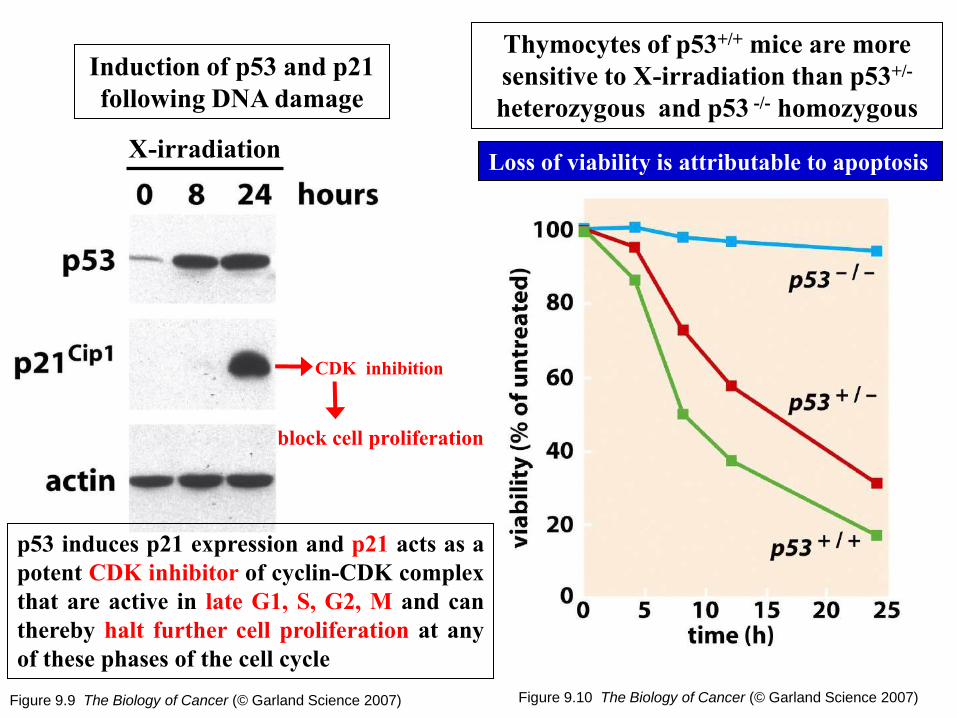
Figure 9.9 The Biology of Cancer (© Garland Science 2007)
Induction of p53 and p21
following DNA damage
Figure 9.10 The Biology of Cancer (© Garland Science 2007)
Thymocytes of p53+/+ mice are more
sensitive to X-irradiation than p53+/-
heterozygous and p53 -/- homozygous
X-irradiation
p53 induces p21 expression and p21 acts as a
potent CDK inhibitor of cyclin-CDK complex
that are active in late G1, S, G2, M and can
thereby halt further cell proliferation at any
of these phases of the cell cycle
Loss of viability is attributable to apoptosis
CDK inhibition
block cell proliferation
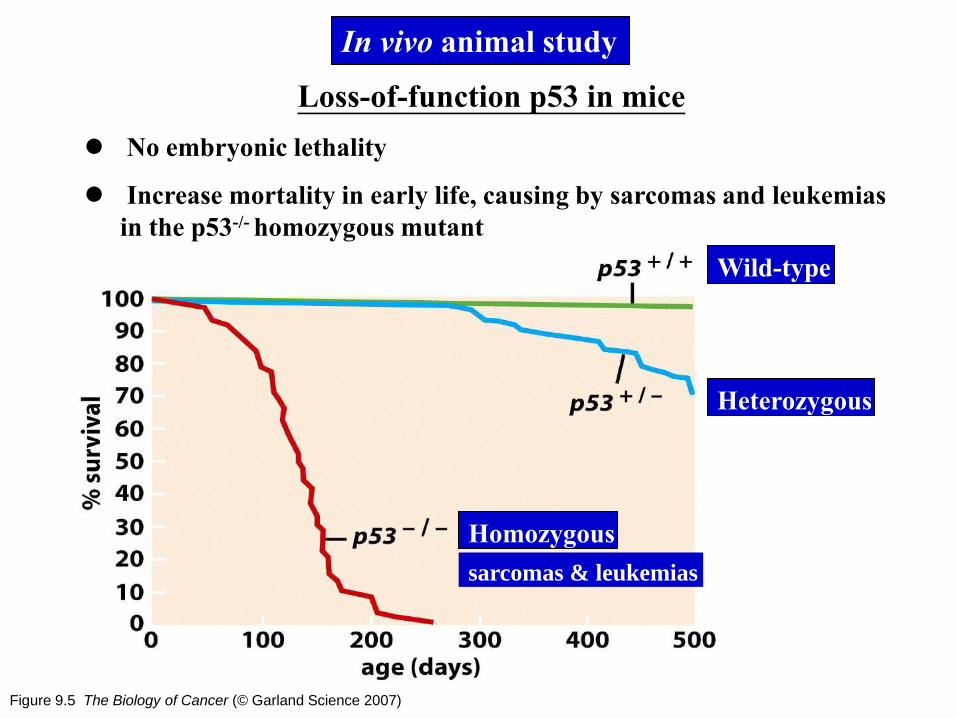
Figure 9.5 The Biology of Cancer (© Garland Science 2007)
Loss-of-function p53 in mice
No embryonic lethality
Increase mortality in early life, causing by sarcomas and leukemias
in the p53-/- homozygous mutant
In vivo animal study
Wild-type
Heterozygous
Homozygous
sarcomas & leukemias

Figure 9.7a The Biology of Cancer (© Garland Science 2007)
Tetramerization domain of p53
Figure 9.7b The Biology of Cancer (© Garland Science 2007)
Mechanism of p53 dominant-negative mutations
15/16 are mutants
1/16 is wild-type
Negative dominant effect
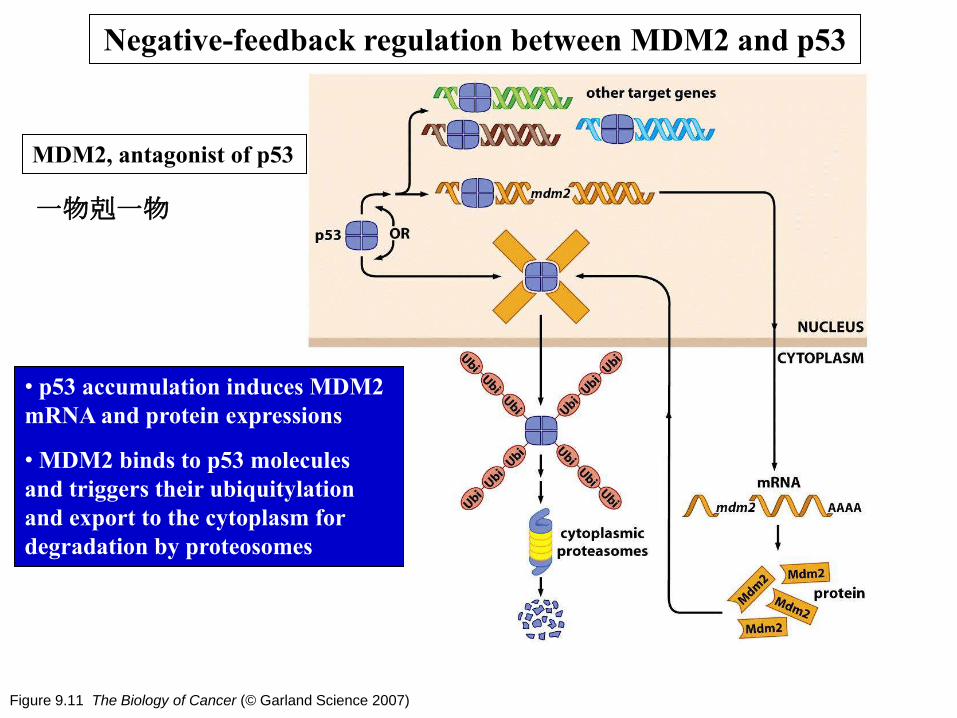
Figure 9.11 The Biology of Cancer (© Garland Science 2007)
• p53 accumulation induces MDM2
mRNA and protein expressions
• MDM2 binds to p53 molecules
and triggers their ubiquitylation
and export to the cytoplasm for
degradation by proteosomes
Negative-feedback regulation between MDM2 and p53
MDM2, antagonist of p53
一物剋一物

Figure 9.14 and 9.15 The Biology of Cancer (© Garland Science 2007)
Control of apoptosis by ARF
The gene encoding p14/p19 protein uses
an alternative transcription promoter
Protect p53
from degradation
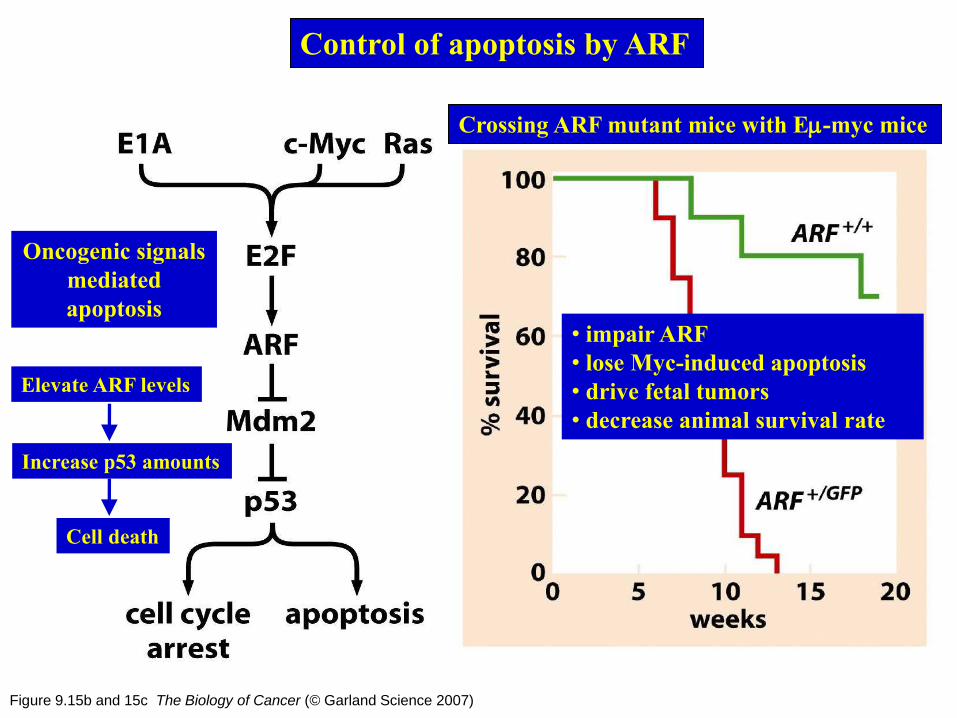
Figure 9.15b and 15c The Biology of Cancer (© Garland Science 2007)
Control of apoptosis by ARF
Oncogenic signals
mediated
apoptosis
Elevate ARF levels
Increase p53 amounts
Crossing ARF mutant mice with Em-myc mice
• impair ARF
• lose Myc-induced apoptosis
• drive fetal tumors
• decrease animal survival rate
Cell death

Figure 9.13 The Biology of Cancer (© Garland Science 2007)
Control of p53 levels by
various kinases
Chk2
ATM
ATR
Phosphorylation
of p53 blocks
MDM2 binding
AKT/PKB
Phosphorylation
of MDM2
enhances
binding to p53
p53 ubiquitylation and
proteosome-mediated
destruction
Positively regulates p53
Negatively regulates p53
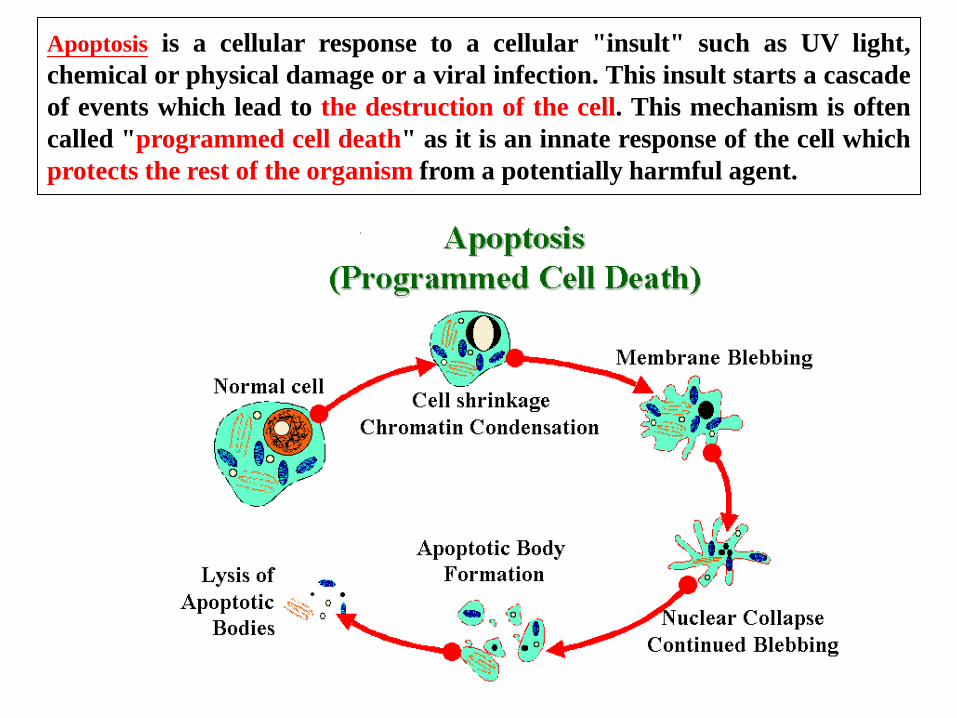
Apoptosis is a cellular response to a cellular "insult" such as UV light,
chemical or physical damage or a viral infection. This insult starts a cascade
of events which lead to the destruction of the cell. This mechanism is often
called "programmed cell death" as it is an innate response of the cell which
protects the rest of the organism from a potentially harmful agent.
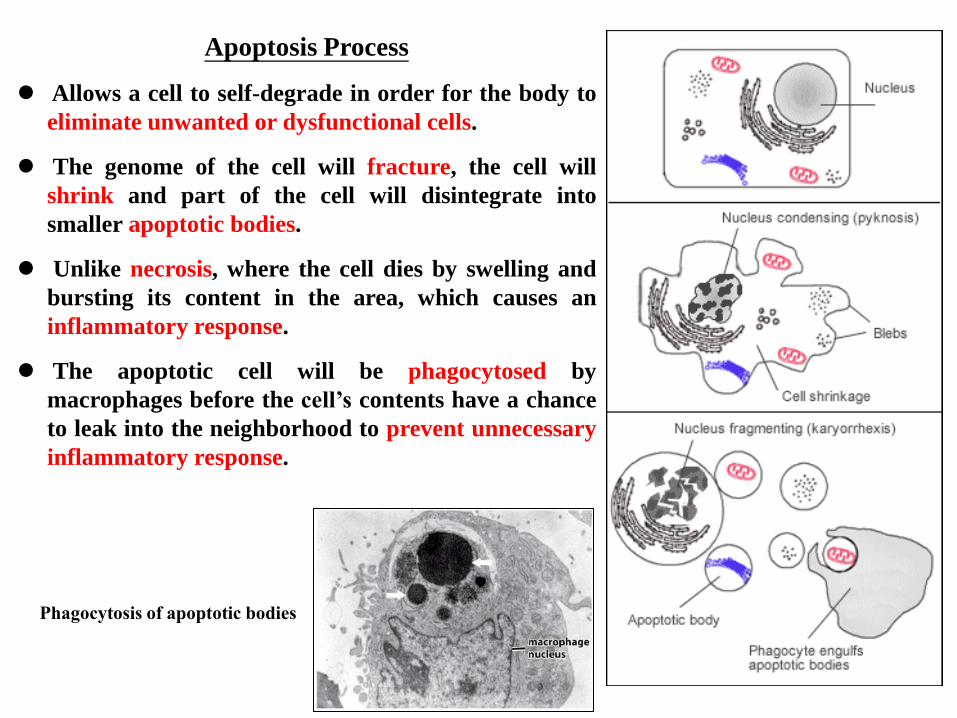
Apoptosis Process
Allows a cell to self-degrade in order for the body to
eliminate unwanted or dysfunctional cells.
The genome of the cell will fracture, the cell will
shrink and part of the cell will disintegrate into
smaller apoptotic bodies.
Unlike necrosis, where the cell dies by swelling and
bursting its content in the area, which causes an
inflammatory response.
The apoptotic cell will be phagocytosed by
macrophages before the cell’s contents have a chance
to leak into the neighborhood to prevent unnecessary
inflammatory response.
Phagocytosis of apoptotic bodies
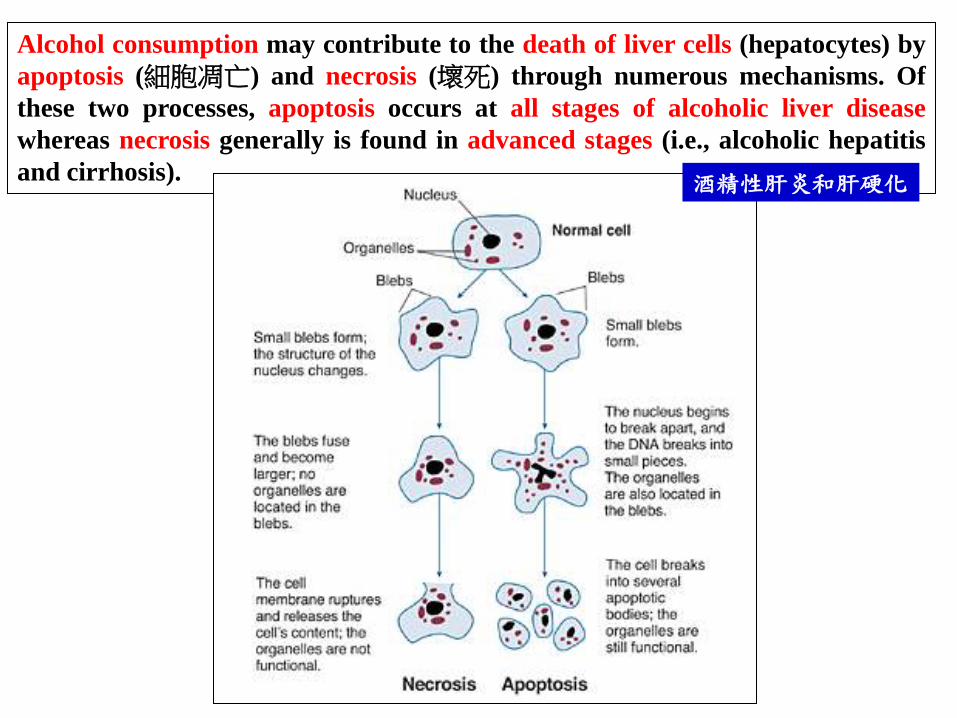
Alcohol consumption may contribute to the death of liver cells (hepatocytes) by
apoptosis (細胞凋亡) and necrosis (壞死) through numerous mechanisms. Of
these two processes, apoptosis occurs at all stages of alcoholic liver disease
whereas necrosis generally is found in advanced stages (i.e., alcoholic hepatitis
and cirrhosis). 酒精性肝炎和肝硬化

Table 9.3 The Biology of Cancer (© Garland Science 2007)

Morphological changes of apoptotic cells
Apoptotic bodies
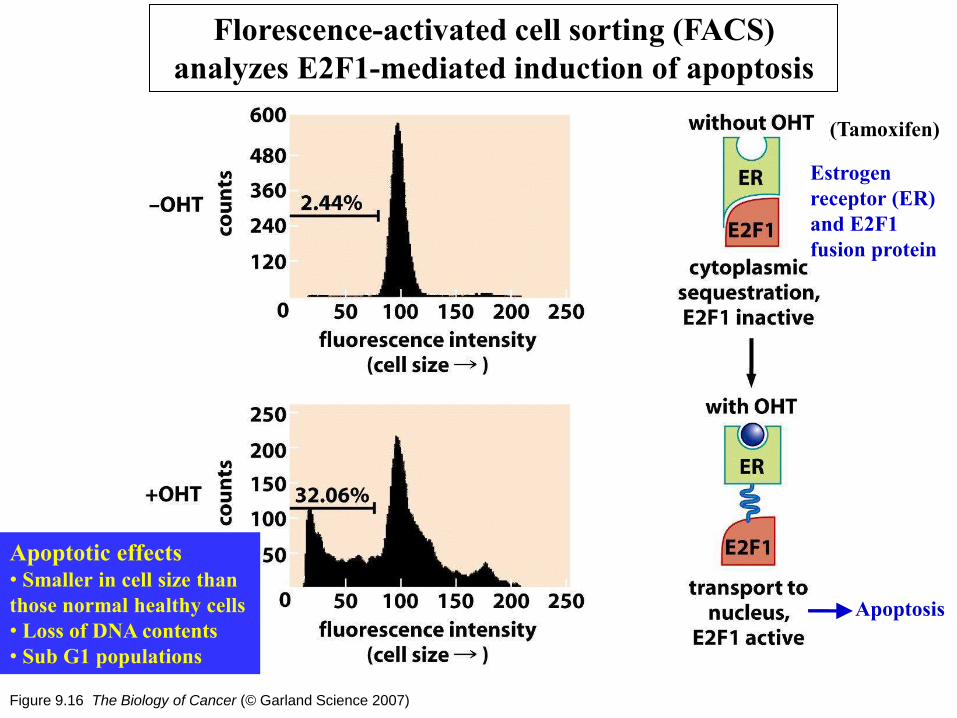
Figure 9.16 The Biology of Cancer (© Garland Science 2007)
Florescence-activated cell sorting (FACS)
analyzes E2F1-mediated induction of apoptosis
((Tamoxifen)
Measure the size
of individual cell
Apoptosis
Apoptotic effects • Smaller in cell size than
those normal healthy cells
• Loss of DNA contents
• Sub G1 populations
Estrogen
receptor (ER)
and E2F1
fusion protein

Figure 9.18a-f The Biology of Cancer (© Garland Science 2007)
Diverse manifestations of the apoptotic program Scanning electron microscopy
Chromatin condensation and nuclear collapse
DNA laddering
Golgi (green) body fragmentation
and nuclear PARP cleavage (orange)
Phospho-histone 2B (serine 14)
stains in apoptotic nuclei
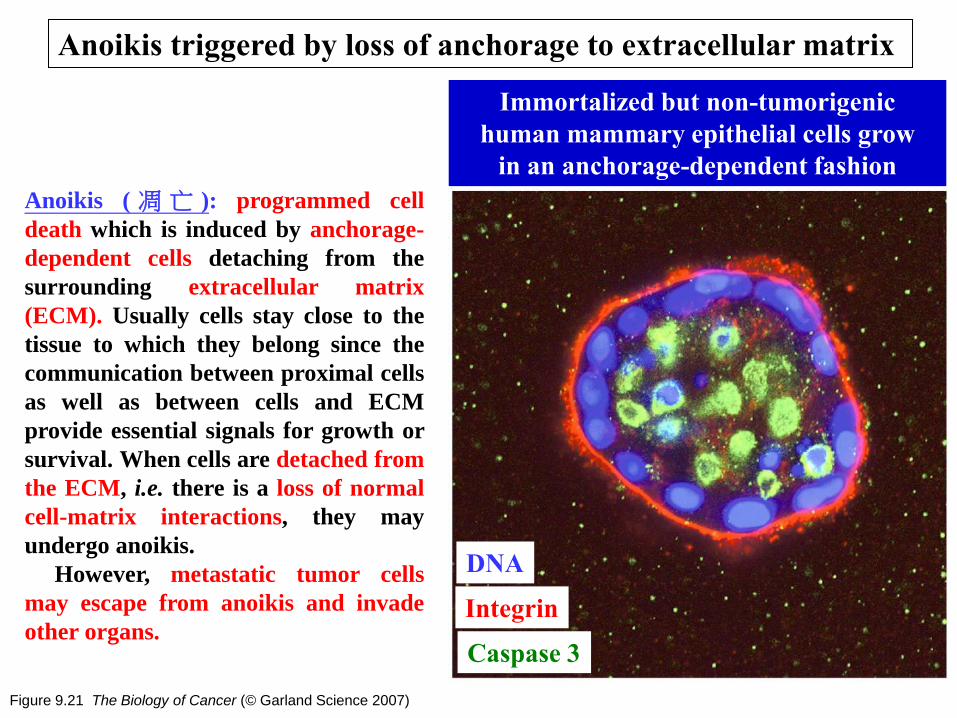
Figure 9.21 The Biology of Cancer (© Garland Science 2007)
Anoikis triggered by loss of anchorage to extracellular matrix
Immortalized but non-tumorigenic
human mammary epithelial cells grow
in an anchorage-dependent fashion
Integrin
DNA
Caspase 3
Anoikis ( 凋 亡 ): programmed cell
death which is induced by anchorage-
dependent cells detaching from the
surrounding extracellular matrix
(ECM). Usually cells stay close to the
tissue to which they belong since the
communication between proximal cells
as well as between cells and ECM
provide essential signals for growth or
survival. When cells are detached from
the ECM, i.e. there is a loss of normal
cell-matrix interactions, they may
undergo anoikis.
However, metastatic tumor cells
may escape from anoikis and invade
other organs.

Aberrant acinar formation induced by overexpressing MCT-1
in MCF-10A cells
E F
G H
A B
C D
control MCT-1
DNA EGFR DNA EGFR
Acinar formation at day 10
Round
Organized nuclei
Robust cell - cell adhesion
- Grape like
Disorganized nuclei
Poor cell - cell adhesion In vitro mammary tumorigenesis assay
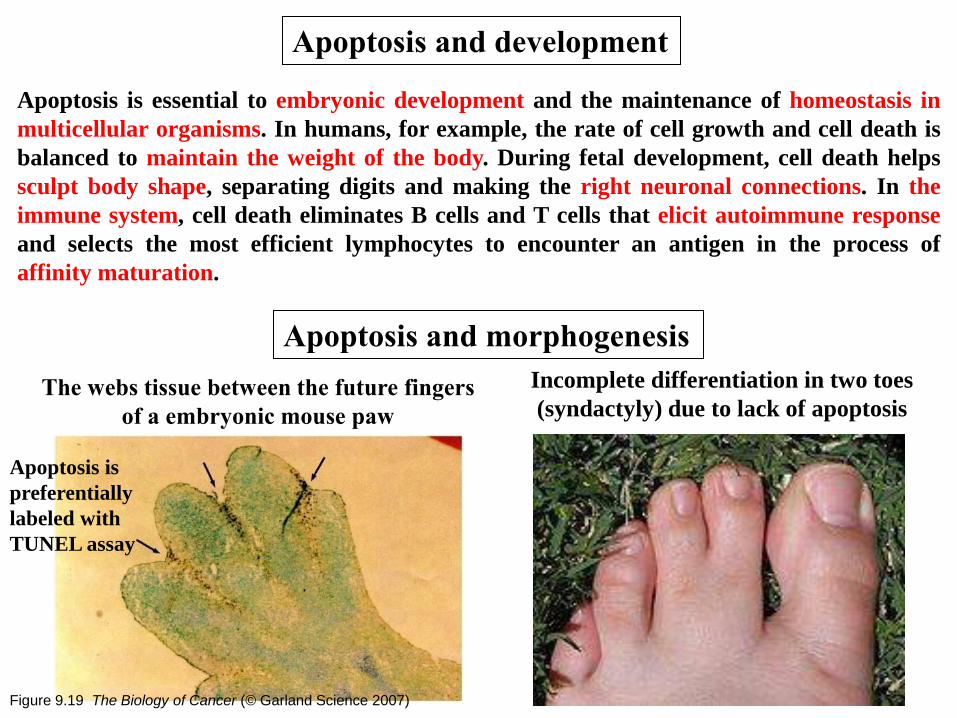
Apoptosis is essential to embryonic development and the maintenance of homeostasis in
multicellular organisms. In humans, for example, the rate of cell growth and cell death is
balanced to maintain the weight of the body. During fetal development, cell death helps
sculpt body shape, separating digits and making the right neuronal connections. In the
immune system, cell death eliminates B cells and T cells that elicit autoimmune response
and selects the most efficient lymphocytes to encounter an antigen in the process of
affinity maturation.
Apoptosis and development
Apoptosis and morphogenesis
The webs tissue between the future fingers
of a embryonic mouse paw
Incomplete differentiation in two toes
(syndactyly) due to lack of apoptosis
Figure 9.19 The Biology of Cancer (© Garland Science 2007)
Apoptosis is
preferentially
labeled with
TUNEL assay
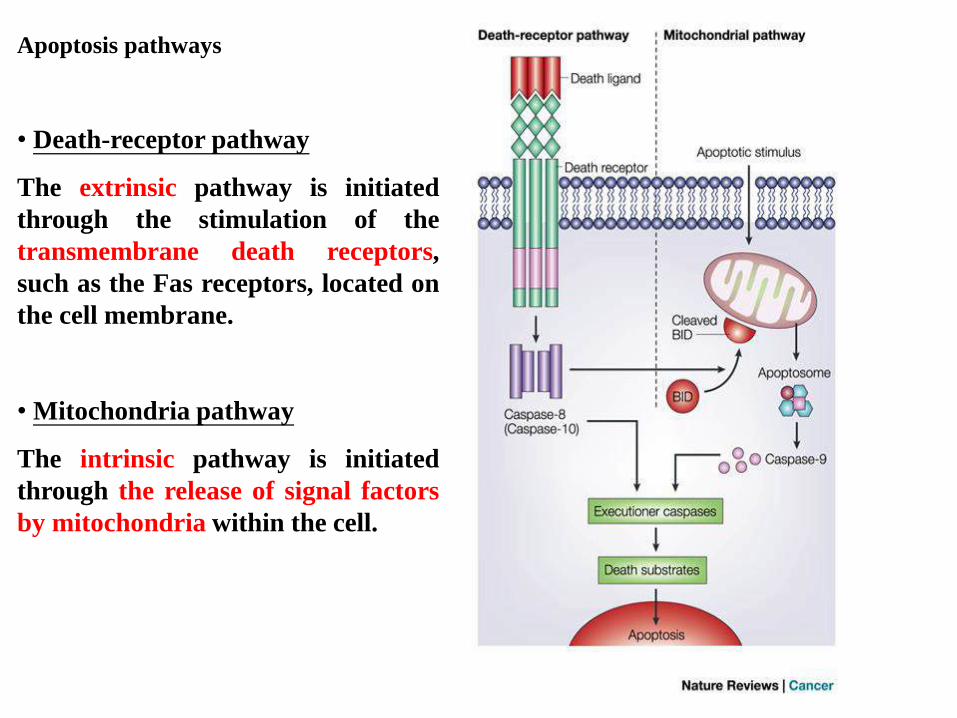
Apoptosis pathways
• Death-receptor pathway
The extrinsic pathway is initiated
through the stimulation of the
transmembrane death receptors,
such as the Fas receptors, located on
the cell membrane.
• Mitochondria pathway
The intrinsic pathway is initiated
through the release of signal factors
by mitochondria within the cell.

Figure 9.31c The Biology of Cancer (© Garland Science 2007)
FasL (Fas receptor ligand) is used by cytotoxic T lymphocyte to kill cancer cells
Cytotoxic T lymphocyte
Apoptotic cancer cells
(as evidenced by numerous
blebs on its surface)
Colorized
scanning
electron
micrograph
Death-receptor pathway

Figure 9.23a-b The Biology of Cancer (© Garland Science 2007)
Mitochondria in human liver cell
Function of mitochondria
• Oxidative phosphorylation
• Biosynthesis of metabolites
• Power producers
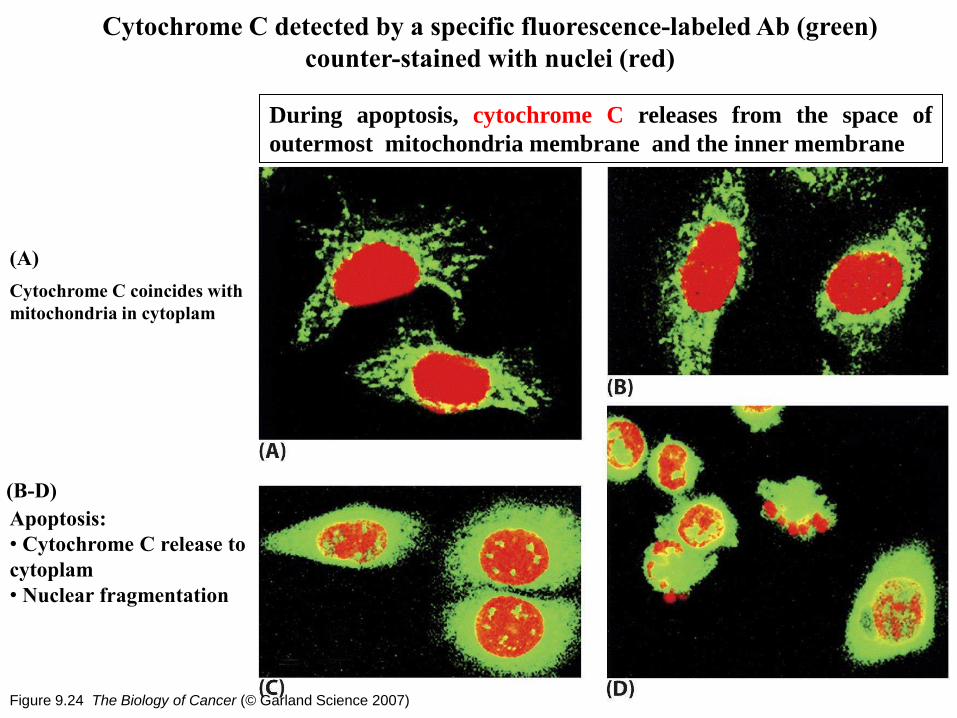
Figure 9.24 The Biology of Cancer (© Garland Science 2007)
Cytochrome C detected by a specific fluorescence-labeled Ab (green)
counter-stained with nuclei (red)
Cytochrome C coincides with
mitochondria in cytoplam
(A)
(B-D)
Apoptosis:
• Cytochrome C release to
cytoplam
• Nuclear fragmentation
During apoptosis, cytochrome C releases from the space of
outermost mitochondria membrane and the inner membrane

Important role of mitochondria in apoptosis
• Directly activated by apoptotic signals such as cell stress, free radical damage
or growth factor deprivation
• Mitochondria contain many pro-apoptotic proteins such as Apoptosis
Inducing Factor (AIF), Smac/DIABLO and cytochrome C
• Release these factors from the mitochondria following the formation of the
Permeability Transition pore, or PT pore
• Pores are formed through the action of the pro-apoptotic members of the bcl-
2 family of proteins

Apoptosome assembly
• Bcl-2 family proteins act as sensors of cellular damage or stress in the cytoplasm
• Relocate to the surface of the mitochondria
• Interact between pro- and anti-apoptotic proteins
• Disrupt the normal function of the anti-apoptotic bcl-2 proteins
• Form the pores in the mitochondria
• Release cytochrome C and other pro-apoptotic molecules from the
intermembrane space
• Release Cytochrome C into the cytosol from mitochondria
• Interact with Apaf-1 protein and then recruits pro-caspase 9 to form a multi-
protein complex with cytochrome C and Apaf-1 called the apoptosome
• Formation of the apoptosome leads to activation of caspase 9 and the induction
of apoptosis.
• Activate the caspase cascade


Figure 9.28 The Biology of Cancer (© Garland Science 2007)
The wheel of death (apoptosome)
• Cytochrome C release from mitochondria
into cytoplasm and associates with Apaf-1
• Attracts procaspase 9 to the hub of the wheel
• Cleaves and activates other caspase molecules
• Triggers the apoptotic cascade

Figure 9.29 The Biology of Cancer (© Garland Science 2007)
The apoptotic caspase cascade
Apoptosome





























