Embed Size (px)
Citation preview

ヒト単球性白血病細胞株(THP-1)における
樹状細胞様の感作性物質応答と
その作用機構に関する研究
雨滴50号
宮澤 正明

ヒト単球性白血病細胞株(THP-1)における
樹状細胞様の感作性物質応答と
その作用機構に関する研究
宮澤 正明

略語
緒言
第1編
第1節
L
2.
3.
4.
5.
第2節
1.
2
3
第3節
第4節
目次
1
2
THP-1細胞における細胞表面抗原及びサイ
トカインの感作性物質応答 ・・・・・… 6
実験の部 ・・・・・・・・・… ●●
細胞と培養液 ・・・・・・・・・…
被験物質とTHP-1細胞に対する処埋方法
細胞表面抗原の解析 ・・・・・・…
サイトカイン産生量の解析 ・・・…
統計学的解析 ・・・・・・・・・…
結果・・・・・・・・・・・・・・・…
D:NcB、:Niso4、 sLsによる細胞表面抗原
の発現解析 ・・・・・・・・・・・…
DNCB、 Niso4、 SLSによるサイトカイン
産生量変化 ・・・・・・・・・・…
細胞表面抗原の発現とサイトカイン産生
量の感作性物質応答性 ・・・・・・…
考察・・・・・・・・・・・・・・・…
小括・・・・・・・・・・・・・・・…
1
10
14
1(∠(∠

第2編
第1章
第1節
1.
2.
3.
4.
5.
第2節
1.
2
3
4ζ」
6
第3節
第4節
感作性物質によるTHP・・1細胞活性化の作
用機構の解明 ・・・・・・・・・…
THP-1細胞活性化に及ぼすTNF一αと細胞
外Arpの役割 ・・・・・・・・・…
実験の部 ・・・・・・・・・・・…
細胞と培養液 ・・・・・・・・・…
被験物質とTHP-1細胞に対する処理方法
細胞表面抗原の解析・・・・・・…
サイトカイン産生量の解析 ・・・…
統計学的解析 ・・・・・・・・・…
結果・・・・・・・・・・・・・…
D:NcB、 Niso4及びsLsによるサイトカ
イン産生と細胞表面抗原発現の経時変化
CD86、 CD54及びCD40の発現増強に及
ぼすrhTNF一αの効果 ・・・・・…
CD86、 CD54及びCD40の発現増強に及
ぼすrhIL-8の効果 ・・・・・・…
TNF一αの中和抗体による影響 ・…
細胞外ATPがCD86及びCD54の発現量
へ及ぼす影響 ・・・… ◎・…
suraminがD:NCB、 NiSO4及びEUによる
CD86、 CD54の発現へ及ぼす影響 ・・
考察 ・・・・・・・・・・・・…
小泊 ・・・・・・・・・・・・…
27
33
38
024446

第2章
第1節
L 2.
3.
4.
5.
第2節
L
2
3
4
第3節
第4節
THP-1細胞活性化に及ぼすシグナル伝達
経路の役割 ・・・・・・・・・・・…
実験の部 ・・・・・・・・・・・…
細胞と培養液 ・・・・・・・・・…
p38 MAPKとERKのWestem BIot:解析
細胞表面抗原の解析・・・・・・…
サイトカイン産生量の解析 ・・・…
統計学的解析 ・・・・・・・・・…
結果・・・・・・・・・・・・・…
DNCB及びNiSO4によるP38 MAPK、
ERKのリン酸化とp38 MAPK及びERK
に対する阻害剤の効果 ・・・・・…
DNCB及びNisO4によるTNF一αの産生
に対するP38 MAPK、 ERKの影響 ・・
p38 MAPKがDNcB及びNiso4による
CD86、 CD54及びCD40の発現に及ぼす
影響 ・・・・・・・… σ・…
ERKがDNcB及びNiso4によるcD86、
CD54及びCD40の発現に及ぼす影響・・
考察 ・・・・・・・・・・・・・…
小括 ・・・・・・・・・・・・・…
58
61
63
第3編 感作性物質によるIL-8産生の特異性と
発現機構の解明 ・・・・・・・・… 72

第1節
L 2.
3.
4.
5.
第2節
L
2
3
第3節
第4節
実験の部 ・・・・・・・・・・・…
細胞と培養液 ・・・・・・・・・…
被験物質とTHP-1細胞に対する処理方法
細胞表面抗原の解析・・・・・・…
サイトカイン産生量の解析 ・・・…
統計学的解析 ・・・・・・・・・…
結果・・・・・・・・・… ④…
感作性物質及び非感作性物質に対する
IL-8の応答性 ・・・・・・・・・…
感作性物質による1レ8の産生における
細胞内シグナル伝達系の影響 ・…
感作性物質によるIL-8の産生における
TNF一αの影響 ・e・・・・・…
考察 ・・・・・・・・・・・・・…
如露 ・・・・・・・・・・・・…
76
78
∩60000結論 86
謝辞 94
引用文献 96

CD34一一DC
DMSODNCB
EU
ERKFA
HQh-CLAT
HCIL一一1P
IL-6
1L”8
JNK
LPS
MAPKMo-DC
MPMFI
MHC class II
NiSO4
PAA
PGPI
RFI
rhlし8
rhTN F-ct
SLS
TNF-ct
略語
CD34+ progenitor cell derived dendritic cell
dimethyl sulfoxide
2.4-dinitrochlorobenzene ’
eugeno玉
extracellular signal-regulated kinase
foftnaldehyde
hydroquinone
human Cell Line Activation Test
hydroxycitronellal
interleukin-m l B
interleukin-6
interleukin-8
c-Jun N-terrninal kinase
lipopolysaccharide
mitogen-activated protein kinase
inoRocyte-derived dendritic cell
methyl paraben
mean fiueresceRce intensity
class II maj or histocompatibility complex
nickel sulfate
phenylacetaldehyde
propyl gallate
propidium iodide
relative fluorescence intensity
recombinant human interleukin-8
recombinant human tumor necrosis factor-ct
sodium laury1 sulfate
tumor Recrosis factor-oc
1

緒言
堕罪品を開発する上で、皮膚に対する安全性を確認することは
極めて重要で、中でも皮膚感作性の評価を行うことは、安全な製
品を開発する上で欠かせない項目の1つである。皮膚感作性とは
遅延性過敏反応の一つで、化学物質の接触により過剰な免疫反応
により皮膚にかぶれが起こる現象であり、外来の化学物質や物理
的因子などのダメージによる皮膚刺激性のかぶれとは、その発症
機構が大きく異なる。従来、化学物質の皮膚感作性評価は、モル
モットやマウスなどの動物を用いた試験が行われてきた12)。一
方、動物愛護の観点から動物実験に対する批判の高まりを受け、
EUにおいては化粧品指令第7次改正3)により、2013年3月まで
に化粧品開発のための動物実験の禁止が施行されるに至ってい
る。このような社会的背景から、動物実験が不可欠であった化学
物質の全ての安全性評価において、動物実験代替法の必要性が明
確となっている。これまでにOECDでは、経皮吸収性試験4)や
局所刺激性を評価する光毒性試験5)及び皮膚腐食性試験6・7)が、
安全性評価における動物実験代替法としてガイドライン化され
ている。また、皮膚感作性に関しても、動物を用いない試験法に
対する関心が高まり、試験法開発に向けた研究が世界的に行われ
ているが、ガイドライン化された試験法はない。この埋由として、
皮膚刺激性などの局所刺激性に比べ、皮膚感作性は免疫系が関与
する全身反応であり、加痂mで全ての反応を具現化することが極
めて難しいことが挙げられる。よって、動物を用いない皮膚感作
性試験を開発するためには、皮膚感作性の毒性発現機構を十分に
2

埋解し、in・vitroで具現化できる鍵となる反応に着目することが重
要である。
感作性物質が皮膚に接触すると、皮膚中のタンパク質やペプチ
ドなどと結合することにより完全抗原となり、表皮中の樹状細胞
(ランゲルハンス細胞)によって認識される。その後、ランゲル
ハンス細胞は活性化し、皮膚から所属リンパ節へ移行し、T細胞
に対して抗原情報を提示する。そして、抗原特異的なT細胞が増
殖すると、感作が成立する8・9)。ここまでの感作誘導反応により、
感作性物質の情報は体に記憶される。さらに、再び、皮膚に同じ
物質が触れることによって、T細胞が活性化し、紅斑や浮腫とい
った炎症反応が皮膚に起こる。これら一連の反応は極めて複雑で
あるが、感作が成立するまでに鍵となるいくつかの反応(経皮吸
収性、タンパク結合性、ランゲルハンス細胞の活性化、抗原特異
的なT細胞増殖)を個別にin vitroで再現する試みがなされてい
るIo)。この中でもランゲルハンス細胞の活性化では、 MHC class
IIやCD86、 CD54のような共刺激性分子の発現増強ll・12)やIL-1β
のようなサイトカインの産生B・14)が認められており、これらの
変化に着目した研究が積極的に進められている。一・方で、ランゲ
ルハンス細胞は表皮細胞中に1~3%しか存在しないため、表皮細
胞からの分離が難しく、また分離後に性質が変化することも知ら
れており、これらがランゲルハンス細胞を用いた研究の課題とな
っている。そこで、近年、ランゲルハンス細胞の代替として、ヒ
ト末梢血CD14陽性細胞15)あるいはCD34陽性造血前駆細胞16>
から種々のサイトカイン処理により誘導した樹状細胞を使用し
た検討がなされている。これらの細胞は表現型及び機能上の特性
3

がランゲルハンス細胞と類似しているだけでなく、∫η痂m実験に
おいて、皮膚感作性物質によりランゲルハンス細胞と同様に、抗
原提示に関わる種々の細胞表面抗原の発現やサイトカインの産
生が増強することから、生体での反応をin vitroで再現できるモ
デルとして、皮膚感作性の発症機構の研究に用いられている
17“’22)。一方で、このような樹状細胞の活性化を指標にすることで、
in vitro皮膚感作性試験法へ応用できる可能性も高まってきてい
る23>。しかしながら、安全なヒト血液の供給や用いる細胞の個
人差の問題19’24>などがあるため、これら細胞を用いた試験法開
発には実用面での課題がある。
著者らはそうした問題を克服するために、入手が容易で、性状
が安定な、ヒト細胞株であるTHP-1細胞(human monocytic
leukemia ceU line)25)に着目した。これまでに、 THP・一1細胞に対
していくつかの皮膚感作性物質を曝露すると、抗原提示の際に樹
状細胞で認められる細胞表面抗原(MHC class II、 CD86、 CD54)
の発現上昇やIL-1βの産生増強が報告されている26・27)。さらに、
THP4細胞を用いたin vitro皮膚感作性試験法(human Cell Line
Activation Test;h-CLAr)の開発を進め28・29)、抗原提示の際に共刺
激分子として機能する細胞表面抗原(CD86とCD54)の発現変化
を指標とすることで、一般化学物質64物質の感作性の有無を良
好に評価できることを確認した30)。一方で、試験法に用いる
THP-1細胞の感作性物質応答やその作用機構は十分に解明され
ておらず、試験法の実用化に向けての課題となっている。
そこで、本研究では、加痂m皮膚感作性試験法に用いる細胞と
してTHP-1細胞の有用性を高めるために、THP-1細胞の感作盛物
4

質応答とその作用機構について解析した。本研究論文の第1編で
は、感作成立において樹状細胞で認められる種々の細胞表面抗原
やサイトカインの発現変化を指標に、THP-1細胞の感作性物質及
び非感作性物質に対する応答を調べ、樹状細胞との同等性を検討
した。また、第2編では、感作性物質による種々の細胞表面抗原
やサイトカインの発現機構に関して、TNF一αのオートクライン作
用、細胞外Arpの効果及び、細胞内シグナル伝達系の役割につい
て検討した。さらに、第3編では、THP4細胞を用いて、樹状細
胞において十分に解明されていない1レ8産生の感作性物質特異
性や発現機構を検討した。
5

第1編 THP-1細胞における細胞表面抗原及びサ
イトカインの感作性物質応答3D
加痂邦。実験において、ヒト由来の樹状細胞が感作性物質により
活性化し、抗原提示の際に重要な種々の細胞表面抗原やサイトカ
インの発現が充進ずることが報告されている17”22)。最近では、
これら活性化マーカーを指標とした、加痂m皮膚感作性試験の開
発研究がなされている23)。しかしながら、血液の安定供給や均
一な細胞集団の確保、さらに細胞ドナー間のばらつき等の点19’24)
から、血液由来の樹状細胞を研究に汎用できる細胞として用いる
には課題があるのが現状である。
そこで、本研究では、これら課題を克服することを目的にヒト
単球由来の細胞株であるTHP-1細胞(human m.onocytic leukemia
cell line)を用い、 THP-1細胞が感作性物質処理によって樹状細胞
様の応答を示すかを検討した。感作性物質による樹状細胞の活性
化指標としては、細胞表面抗原のCD86、 CD54、 CD40、 CD83、
CD58、 CD l a及び、サイトカインのTNF一α、 IL-6、 IL一一8を用いて、
検討した。
6

第1節 実験の部
1.細胞と培養液
THP-1細胞はAmerican Type Culture Collection (Manassas, VA,
USA)より購入した。THP-1細胞用の培養液として、25mM HEPES
バッファーとLグルタミンを含むRPMI1640 (invitrogen Corp.,
Carlsbad, CA USA)に、10%牛胎児血清(JRH Biosciences, Lenexa,
KS, USA)と、 O.05mM 2一メルカプトエタノール及び1%ペニシリ
ンーストレプトマイシン(hnvitrogen Corp., Carlsbad, CA USA)を
それぞれ添加した培養液を用いた。
2.被験物質とTHP-1細胞に対する処理方法
被験物質は、感作性物質として2,4-dinitrochlorobenzene
(DNCB),nickel sulfate hexahydrate (NiSO4),hydroquinone (HQ),
propyl gallate (PG), formaldehyde (FA), eugenol (EU),
phenylacetaldehyde (PAA)、 hydroxycitronellal (HC)を用いた。
また、非感作性物質としては、sodium lauryl sulfate(SLS)、methyl
paraben (MP)、1actic acid(LA)を用いた。全ての被験物質と、
陽性対照として用いたサルモネラアボルタス由来の1ipopolysaccharide(LP S)は、 Sigma-Aldrich(St. Louis, MO, USA)
より購入した。NisO4、 HQ、 FA、 sLs、 LA、 LPsは予め生理食塩
水に溶解させ、また、DNCB、 PG、 PAA、 EU、 HCはdimethylsulfbxide
(DMSO;Simga-Aldrich)に溶解させた後に、培養液に添加した。
DMSOは培養液中の濃度が最高0.2%になるように添加した。
THP-1細胞は24ウェルプレートに1x106cells/mL/ウェルにてi播
7

種し、被験物質を添加した後、37℃、5%CO2条件下で24時間培
養した。被験物質の処理濃度は予め細胞毒性試験を行い、24時間
後に70~95%の細胞生存率になる3濃度を設定した。各被験物質
に対して、実験は少なくとも3回実施した。
3.細胞表面抗原の解析
細胞に化学物質を24時間処理した後、細胞を回収し、0.1%
bovine serum albumine添加PBSで洗浄した。さらに、 Fcレセプタ
ーのブロックキングとして0.01%Globlins Cohn Fraction II, III
(Sigam-Aldrich)添加PBSを10分、4℃条件下で処理した後、抗
体染色を行った。細胞表面抗原の解析に用いたFITC標識モノク
ロナール抗体を以下に示す;抗ヒトCD54(clone;65B5)(DAKO;
Glostrup, De㎜ark)、抗ヒトCD86(clone;Fun-1)、抗ヒトCD40
(clone;5C3)、抗ヒトCD83(clone;HB I 5e)、抗ヒトCD58(clone;
1C3)、抗ヒトCD l a(clone;HI149) (以上、 BD PharMingen;San
Diego, CA)。また、 Isotype controlとして、マウスIgG1をDAKO
より、マウスIgG 1,K及びマウスIgG2,KをBD PharMingenよりそれ
ぞれ購入した。抗体染色は遮光、4℃条件で、30分間行った。
細胞表面抗原の発現量測定にはFAcscaliburとcellQuest
software(Beckton Dickinson, San Jose, CA, USA)を用いた。死細
胞を染め分けるため、細胞浮遊液にpropidium iodide (PI;
Sigma-Aldrich)を最終濃度0.625μg/mしで添加し、生細胞が10,000
個になるまで測定した。細胞表面抗原の発現量の評価方法として、
平均蛍光強度(Mean fiuorescence intensity (MFI))を用いた。
8

4.サイトカイン産生量の解析
細胞に化学物質を24時間曝露した後、培養上清を回収し、
ELISA kit (R&D Systems, Mi㎜.eapolis, MN)を用いて、 TNF-ct、
IL-6、 IL-8の産生量を測定した。
5.統計学的解析
解析したマーカーの増強変化を評価するためにMFIを用いて、
溶媒処理群と化学物質処理群との間の統計学的な有意差を
Student’s・t-testにより解析した。
9

第2節 結果
1.DNcB、 Niso4、 sLsによる細胞表面抗原の発現変化
本研究では、THP-1細胞に対し、細胞生存率が70~95%を示す
3濃度でDNcB、 Niso4、 sLsを24時間処理した後、細胞表面抗
原の発現変化を調べた。Fig。1には、無処理及び、溶媒のDMso、
DNCB (2.5 ug/mL), NiSO4 (85 ng/mL), SLS (45 pg/mL),
それぞれを処理した際の細胞表面抗原の発現変化を示した。
DMSO処理群は無処理群とほぼ同じ結果だったことから、細胞表
面抗原に及ぼすDMSOの影響はほとんどないと考えられた。
DNCB (2.5μg/mL)を処理した細胞ではCD58を除き、全ての
細胞表面抗原で発現が増強した。NiSO4(85μg/mL)でもCD86、
CD54、 CD40、 CD83の発現が増強した。一方で、 SLS(45μg/mL)
処理群では、無処理群と同様に、いずれの細胞表面抗原の変化も
起こらなかった。
Fig.2にはDNcB、:Niso4、 sLsを24時間処理した後の、cD86、
CD54、 CD40、 CD83、 CD58、 CD laのMFIの値を示した。 Table 1
には統計学的解析の結果をまとめた。その結果、DNCB及びNiso4
ではDMSO及び無処理群それぞれに対してCD86、CD54、CD40、
CD83の発現を有意に増強した。加えて、 D:NCBではCDlaの発
現が増強した。一方、cD58はD:NcB及びNiso4において有意な
変動は認められなかった。また、SLSではいずれのマーカーも変
化を示さなかった。陽性対照として用いたLPSでは、 CDlaを除
く全てのマーカーの発現が強く誘導された。
10

DNCB
O翼
CD86
湧禽琴9
茎)懸SO
韮)NCB
ca
rie tti1 1ti2量
ld3
⊃窯
CD54
NisO4馨
O享
lr Zir・
亙
ldt ldi2 rd3
CD4e
瀞丁
⇔ ゆ彰
静「
ま匿障
Non-treated
NiSO4
SLS
et5e ttii 一,iij?,
墜
rd3
蟄環蓼り
にUO4 撞5
鍵
1ti1 1e’2 rd3
1dt 葉02 狛3 10司
F至ト
Nen-trea{ed
SLS
cat60 rdi sda rd3
靴筆唱
ゆld9 隻dO atii id2 ad3 罫LI-H
魯一霊
「礎
門稲
拍2 vi3 g
り4 too 竃ol ’曳02 兎03 FLIH
導
DNCB 臆憲蓼9
零”
CD83
魯
160 ui1 1ti2豊
rd3
ゆヌ
CDIa
NisO4 慧 v
SLS
群ゴ
O Φコ
國4
1ti1 重02 糠3
⇔耳
O響一
」4
徽11」4
婁
CD58
鰺§δ
卯
uie 1ti1 1ti2 1tiS 蓄ア
O噛“
O弩
1ri1 1ti2 葉03
垂ミ腎
『唱
@冨 当
@ 「
@1 旨
@ 紆
S
き
曝oo loI 1。2 1⑪3 lo4
毒薦
罫
、
‘ 一
曹
loo lo望 lo2 103 10 咤
FLIお
ゆ ゆ
rde lo1 si2 、■⑪3 1ti9雪。。 ドいお
F韮UOreSCenCe i厩e獄Sity
1ti1 1ti2 ui3 FLI-H
4
Fig. 1 Phenotypic alterations iRduced by DMSO (vehicle) ,
DNCB (2.5 ptg/mL),NiSO4 (85 pg/mL),aAd SLS (45 pg/mL).
The data showfl represent middle concentrations used in the study.
These concentrations were selected as representative data that
provide the maximum effect. Histograms for each surface marker
evaluated in THP-1 cells: lsotype control (dotted line) ,
vehicle-control (either DMSO or vintreated; shaded peak) , and
chemical-treated (dark solid line) . Since untreated control and
DMSO-treated control gave the same results, untreated and
DMSO-treated results were interchaRged. These are representative
data frem three independent experiments. MFI : meanfiuorescence inteRsity.
11

一駈芝
000000000008530
432- 432茎 茎
432{蓋
000000853005050
聾
AU7」濫」2
茎
「器翔㌔
b噛轟ltttt
一笹一 @v韮ab羅i重y
7
臨.
75㈱噛う9樹1..,,mu,.. .ma11
1eg
80
6e
4e
20
e
<鍵茎凶ξ(ま)
CDla『…
箋8R壽m 壌es ge羨老顧 サ リ
1 M DNcB N藍SO・
灘盤
O肖
簸
マ鴫
鴇貯
嫡.
m執
SPLSLS量曇奨霞嫡eSS調マ富㍉・9/mL)
g・aDNCB NiSO, SLS LPS
Fig. 2 Alterations of surface phenotypes by DNCB, NiSO4,
SLS and LP S at the indicated concentrations. The results were
expressed as MFI. Mean±S.D. was given fer at least three
independent experiments. Statistical significance is reported in
Table 1.
12

Table 1 Student’s t-test was used to establish a statistical significance
between MFI values for vehicle contrel cells and MFI values for chemical
treated cells.
㌔\@\\ \、 \
DNCB NisO4 HQ PG FA PAA EU HC SL,S MP LACD86 醸蹴購懸蕪灘灘 ‡ 織撒熱灘麟購懸鍵灘i灘
鍛.S. 霊LS。 n.S.
CD54 撫継纐鑓 ‡ 縫購 ‡ 蟻灘総磯灘鱗 n。S. 難.S。 n.S.
CI)40 慧織馨灘醸灘灘灘繋灘欝1 難灘鰹鱗糠難臨& ‡ n.s.
CD83 麟織欝講襲羅難繕灘灘灘繋灘灘蕪灘講 獄.S。 麟懸ln.& n.s。 薮.S.
TNF一α 薫響灘譲轟熱難懸灘灘 ‡ 蟻蘂欝轍灘灘 n.S。 灘騰臨& ILS. 聡.S.
IL-8 蟻慧繁鍵懸欝繊灘灘灘懸灘繕 ‡懸灘§ n.S. 灘鎌臨& 鎗.S. n.S.
Results were obtained from representative concentrations that induced a
maximum effect. (**;p〈O.Ol,*;p〈O.05,*;p〈O.1 compared with
control, n.s.; not significant) .
13

2.DNCB、 NiSO4、 SLSによるサイトカイン産生量変化
THP-1細胞に対し細胞生存率が70~95%を示す3濃度でDNCB、
Niso4、 SLSを24時間処理した後、培養上清を回収し、 TNF一α、
IL-6、 IL-8の産生量を測定した。 DNcB及びNiso4では濃度依存
的に、有意にTNF一αとIL-8の産生量が増加した。 IL一・6に関して
は、DNCBやSLSでは変化が認められないが、170μg/mL NiSO4で
は有意な産生が認められた。検討した3つのサイトカインの中で、
IL-8の産生量が最も多く、NiSO4では最大57,000 pg/mL以上に達
した。一方、SLSではいずれのサイトカインの産生も認められな
かった。陽性対照のLPSでは全てのサイトカインで多量の産生が
認められた。
14

(臼還bの畠言。竃ぞ。益
0000005050
0505 0752
2蓋董長 -
AUnUOAUO
nUO(UnU
AUO凸U凸U
4今」2玉
TNF-0し
2/”’i’li”1”i”’l1
2766
軸2..一..ぎ”’ 潤f
IL-6 .!序!・
tttt
蛛r一孚2…2玉
一尽興犠i
Fig. 3
1. L.mar8.....
こ_凱旋。
翻8km 聯霞 :臨9 翼芝【 tt - N
lnDNCB NiSO4 SLS LPS 鷲 (鮭9/mL)
Changes of cytokine production by DNCB,
瞳
NiSO4, SLS and LP S atthe indicated concentrations.
The culture supernatant was measured for TNF-ct, IL-6
and IL-8 secretion by ELISA. Mean±S.D. was given
for at least three independent experiments. Statistical
significance is reported in Table 1.
15

3.細胞表面抗原の発現とサイトカイン産生量の感作性物質応答
性
THP-1細胞は種々の細胞表面抗原の発現やサイトカイン産生
量において、感作性物質のDNCBと:Niso4に特異的な応答を示し
た。そこで、このような細胞の活性化がより多くの感作植物質に
対しても特異的かどうかを検証するために、CD86、 CD54、 CD40、
CD83に関して、6つの感作性物質と2つの非感作性物質の影響
を調べた。各被験物質の処理濃度は予め細胞毒性試験を行い、24
時間後に70~95%の細胞生存率になる3濃度を設定した。その結
果、Fig 4a-d及びTable 1に示すように、 PAAとHCは検討した
全ての細胞表面抗原において、有意に発現増強した。HQ及びFA
では、CD86、 CD40、 CD83において、濃度依存的に有意な発現
増強が認められた。また、PGとEUにおいても、4つの細胞表面
抗原のうち少なくとも3つで有意な発現増強が認められた。一方
で、非感作性物質のMP及びLAではいずれの細胞表面抗原にお
いても発現の変化は認められなかった。CD54に関しては、全て
の感作性物質で、濃度依存的に発現が増強した。CD40でも全て
の感作性物質で有意な発現増強が認められたが、必ずしも濃度依
存的ではなかった。感作性物質によるCD83の発現増強は、他の
細胞表面抗原に比べて小さいが、EUを除く全ての感作性物質で
有意に増強した。
さらに、被験物質を処理した際の培養上清を回収し、TNF・一Ctと
IL-8の産生量を調べた。 Fig.5a, b及びTable 1で示すように、 FA
とHCでは濃度依存的に、かつ、有意なTNF一・Ctの産生が認められ、
16

最高濃度での産生量は他の感作性物質に比べても多かった。PAA
もまたTNF一αの産生を強く誘導したが、濃度非依存的であった。
HQではTNF一αの産生量は僅かであるが有意な増強変化が認めら
れた。PG、 EU及び非感作性物質(MPとしA)ではTNFα産生量
は少なかった。また、HQ、 PAA、 HCは濃度依存的にIL,.8の有意
な産生を誘導した。PGは最高濃度でのみIL-8の有意な産生が認
められた。EUとFAでは共に、 IL-8を有意に産生しなかったが、
FAで濃度依存的な増強変化が認められた。一方で、 MPとしAは
どちらも1レ8産生量を有意に増強しなかった。
17

肖鼠】≧
(a) CD86 +v嚢ab垂1髭y
nり尽げ
AU登」
〆04弓」茎0
6431 54321
匪
=■,■.=,冒}・葦辱,;,,…,…曾乳,
『\\\、\ \←~
鷹
鋸 塚
(b)CD54
1\くぐ…_、\……\『
“...,”.“,..・一一一…一一・…K”’”’一“”” w’“’一”“’“”““”““’“’“W
(d)CD83
“’
煤c
鶴.
尾自【う藻§鱒繍遷回帰響爺欝聴器§霞§ ま
蓬n HQ PG FA PAA EU HC MP
Fig.4 The effect of six a11ergens and two n.on-allergens on
expression of (a) CD86, (b) CD54, (c) CD40 and
(d) CD83. Cells were treated fbr 24h at the indicated
concentration。 Mean士S.D. was given fbr at least three
independen.t experiments. Statistical significance is reported
in Table 1.
001
雨80
60
40
20
0塚
00霊
80
60
40
≦帥霞ξ(ま)
O nU
2 0 nU
1
80
60
40
20
…㌧
0
…、
00蓋
80
60
40
20
0
~
“ 影 霧
ラ
義
@ §㌔
§点
OO噂一
18

(舜章⇔幻亀言。碧ぞ。鼠
(a) TNF-or
ec 1一
20
o
+ viability
一}}ーミヒ.一.1噛……ミ.ヒ
罷\\
刷
%\… @」
(b) IL-8200{}
茎500
1eoo
seo
綜0
即♂\瀬\朋謝\割 .7
@ 一
:鱒謡遷 299審雛R$8縮§PG FA PAA EU HC MP
100
so
6e
2e
o
00韮
㌧ 08
60
鱒
02
0
繭OOON
OOべ軸
OO三 D加㎎(AL
≦呂罠々(ま)
Fig. 5 The effect of six allergens and two nen-allergens on
production of (a) TNFct and (b) IL-8. Cells were treated
fbr 24h at the indicated concentration. Mean土S.D。 was given
foi” at least three independent experiments. Statistical
significance is reported in Table 1.
19

第3節 考察
本章では第一に、THP-1細胞において、樹状細胞の活性化に伴
い発現増強することが知られている種々の細胞表面抗原の発現
やサイトカイン産生量の応答性を、感作性物質のD:NCB、 Niso4
と非感作性物質のSLSに検討した。 DNCB、 NisO4は共に代表的
な感作性物質であり、∫η痂m実験において、それぞれをヒト血液
由来の樹状細胞(Mo-DC;monocyte derived dendritic cell)あるい
はCD34陽性の樹状細胞(CD34-DC;CD34+ progenitor cell derived
dendritic ce11)に曝露すると、様々な細胞表面抗原の発現やサイト
カイン産生量を増強させることが報告されている17“22)。本研究
により、THP-1細胞においても、 CD86、 CD54、 CD40、 CD83、
TNF一α、 IL一・8がDNcB及び:Niso4に対して有意な発現増強を誘導
することが明らかとなった。一方で、SLSではいずれのマーカー
の変化も認められなかった。そこで、より多くの感作性物質と非
感作性物質が細胞へ及ぼす影響を調べることにより、これら活性
化マーカーの感作性物質特異性を検討した。その結果、THP4細
胞は様々な感作性物質に対し、CD86、 CD54、 CD40、 CD83の発
現を増強し、TNF一α、 IL-8を産生した。一方で、非感作性物質に
対しては有意な応答を示さなかった。さらに、これらの変化は、
樹状細胞(Mo-DC及びCD34一・DC)で認められる応答と類似する
ことが確認できた。
CD40は腫瘍壊死因子受容体ファミリー(TNF receptor family)
に属する膜タンパクで、B細胞や抗原提示細胞上に発現している
32・33)。そして、活性化T細胞上のCD40しによって架橋されると、
20

B細胞や抗原提示細胞の活性化を誘導するシグナルが伝達され、
CD86やCD54の発現増強を促進させる働きを有することが報告
されている34)。このように、CD40は樹状細胞によるT細胞活性
化に強く関与する分子である。Mo-DCにおいては、 DNCB及び
Niso4を含む、種々の感作性物質によるCD40の発現増強が報告
されている19’22)。Mo-DCと同様に、 THP4細胞においては、検
討した全ての感作性物質でCD40は有意に発現増強した。一方で、
非感作性物質では増強しなかった。Niso4とHCにおいては、細
胞生存率が79~94%の範囲で、濃度依存的にCD40の発現を増強
し、特に、最高濃度では無処理細胞に対してMFIの値は3倍以上
に達した。HCは、動物を用いた皮膚感作性試験で弱感作性であ
ると判定される物質35)であるが、THP-1細胞においては、検討
した全てのマーカーで強く発現を誘導した。このことから、HC
によるTHP-1細胞活性化は、他の感作性物質とは異なったシグナ
ル伝達系を介して誘導されることを示唆するのかもしれない。以
上より、THP4細胞は感作性物質によりCD40の発現を増強する
可能性が高いことが示唆された。
CD83は樹状細胞系に発現する分子である。皮膚感作の誘導に
おけるCD83の機能はまだ明らかになっていないが、 CD83の発
現挙動解析により、抗原提示や細胞間相互作用に重要な役割を果
たすことが示唆されている36)。THP-1細胞においては、 EUを除
く、検討した全ての感作性物質でCD83が有意に発現増強した。
DNCBでは25μg/mしで、CD83の有意な発現増強が認められた。
De・Smedtらは、本研究と同様の処理濃度でDNCB、EUをCD34-DC
に24時間処理しても、CD83の発現は増強しないことを報告して
21

いる37)。以上より、THP-1細胞におけるCD83は、 CD34.DCに
おけるCD83に比べて、感作性物質に対する反応性が高い可能性
が示唆された。
TNF一αはランゲルハンス細胞の活性化及び遊走に重要な役割
を果たすことが知られている38・39)。本研究では、PGとEUを除
く感作性物質でTNF一・orの有意な産生増強が認められた。 Mo-DC
において、DNCBを含む化学物質14物質がTNFαの産生量に及
ぼす影響を調べた検討では、Nic12とMnC12のみが有意な産生を
誘導することが報告されている40)。よって、感作性物質に対す
るTHP-1細胞のTNF一α応答性は、 Mo-DCよりも反応性が高く、
THP4細胞に特徴的な性質の可能性が示唆された。
ケモカインのIL-8は種々の炎症反応に関与することが知られ
ており41)、皮膚感作性における役割についてはまだ十分に解明
されていない。一方、本研究では、THP-1細胞からのIL-8産生は、
検討したほぼ全ての感作性物質で認められた。近年、Mo-DCに
おいてもDNcB及びNic12によってIL-8の産生量が増強すること
が報告されている18)。また、CD34-DCでも、 EUでは変化しな
いものの、DNCB及びNiSO4によってIL-8 mRNAレベルが有意
に増強することが報告されている42)。本研究で検討したTNF一α
及びIL-8は感作性物質処理によって細胞から産生することが示
されたが、動物試験による感作性強度の結果とは関連がないよう
に見られた。産生量と感作性物質の感作性強度の関係、また、産
生したサイトカインが細胞活性化に及ぼす影響の解明について
は、さらに検討が必要である。
CDlaはランゲルハンス細胞に発現する分子で、 MHC class II
22

と同様に、T細胞に結合し、脂質抗原を提示することが知られて
いる。また、ランゲルハンス細胞の活性化に伴い、発現が減少
することが知られている43)。THP-1細胞においては、 DNCB(2.5
μg/mL)によりCDlaの発現が増強するものの、 NiSO4や陽性対
照のLPSでさえ、 CDlaの発現増強は起きなかった。 DNCBまた
はNiSO4を処理したMo-DCにおいても、CDlaの発現に有意な変
化は認められていない17)。THP-1細胞において感作性物質が
CDlaの発現に及ぼす影響に関しては、さらに検討が必要である。
IL-6は適切なT細胞活性化に必要であることが知られている
44)。本研究では、THP-1細胞において、 Niso4(170μg/mL)に
よりIL-6の有意な産生が認められたが、 DNCBではいずれの処埋
濃度でも産生は認められなかった。Mo-DCでも、 DNCBでは変
化しないものの、NiC12によってIL-6の産生が有意に増強するこ
とが報告されている17)。また、CD34-DCにおいては、 IL-6 mR:NA
レベルではNiso4でのみ増強が認められるが42)、産生量では
DNcB及びNiso4で増強が認められている37)。このようにIL・・6
の感作性物質応答性は、THP-1細胞、 Mo-DC、 CD34-DCで類似
するが、他のマーカーに比べてその反応性は弱いことが示唆され
た。
CD58はランゲルハンス細胞の活性化に伴い、発現増強するこ
とで、強力な抗原提示やT細胞活性化に寄与することが知られて
いる45)。本研究では、THP-1細胞において、 DNcB及びNiso4
による有意な発現増強は認められなかった。これは、他の細胞表
面抗原に比べて、CD58のベースの発現が高いことに起因する可
能性が考えられた。
23

本研究では、DNcBと比べて、 Niso4によるcD54やcD40の
発現増強、並びに、TNF一・CtやIL・一8の産生増強は顕著に誘導された。
同程度の細胞毒i生を誘導するDNCB (5μg/m.L)とNiSO4(85
μg/mL)とを比較すると、検討した活性化マーカーの発現強度は
およそ2倍の差があった。Mizuashiらは、 Mo-DCを用いた検討
から、DNCBとNiC12は、細胞内レドックスのインバランス誘導
機i構が異なることを報告している46)。すなわち、DNCBは還元
型グルタチオン(GSH)に直接結合することで、細胞内のGSH/
酸化型グルタチオン(GSSG)の比を減少させるのに対し、 NiC12
では、活性酸素種を生じることによりGSH/GSSG比を減少させ
ることが示唆されている。このことから、DNCB及びNisO4によ
るTHP-1細胞の応答1生の違いは、細胞内の異なるシグナル伝達系
の存在を示唆しているのかもしれない。
THP4細胞は様々な感作性物質に対し、機能的に異なる細胞表
面抗原やサイトカインの発現を増強させることが明らかとなっ
た。また、感作性物質による細胞活性化は僅かな細胞毒性(細胞
生存率70~95%)と関連しているように見られた。一方で、細胞
毒性を誘導する処理濃度で非感作性物質を適用しても、THP4細
胞の活性化は誘導されなかった。この結果は、Huletteらの結果と
一致している47)。Huletteらは、 Mo-DCにおいて、10~15%の細
胞毒性が誘導される濃度で感作性物質を処理した際にCD86の発
現増強が認められると報告している。このことから、THP-1細胞
と樹状細胞、いずれにおいても、感作性物質による細胞活性化と
細胞毒性との間には、密接な関係があることが示唆された。
Manomeらは、 D:NCBやNiC12を含む、いくつかの感作性物質に
24

よるMo-DCの細胞死に関して報告している40)。それによると、
NiC12は1mM以上の濃度で、アポトーシスを起こす細胞が有意に
多く、DNCBでは検討した全濃度でネクローシスを起こす細胞が
有意に多いことを報告している。すなわち、細胞死の状態は化学
物質によって大きく異なることが示唆されている。THP-1細胞に
おける、細胞活性化と細胞毒性との関係については、さらに検討
が必要である。
本研究では、THP4細胞の活性化がT細胞への抗原提示に及ぼ
す影響に関しては明らかになっていないし、今後、解決すべき課
題である。抗原提示を担う樹状細胞においては、感作性物質によ
ってCD86やCD83の発現が増強することはよく知られている。
本研究により、THP-1細胞においても、樹状細胞の活性化に関わ
る種々のマーカーの発現が増強することが明らかとなった。これ
ら共通の現象からすると、THP-1細胞の活性化がT細胞への抗原
提示に対して、何らかの役割を担う可能性はあると考えられる。
25

第4節小括
THP一・1細胞が皮膚感作性物質により、樹状細胞の活性化を反映
する様々な応答を示し、それらを指標にすることで、感作性物質
と非感作性物質を判別できることが示された。また、THP-1細胞
は入手が容易で、均一な細胞集団としての特徴から研究に汎用で
きることに加え、樹状細胞の代替として化学物質の感作性の有無
を評価可能な細胞モデルであることが示唆された。
26

第2編感作読物質によるTHP-1細胞活性化の作用機構の解明48・49)
第1章 THP-1細胞活性化に及ぼすTNF一αと細胞外ArP
の役割48)
皮膚に感作性物質が接触すると、表皮中の樹状細胞(ランゲル
ハンス細胞)は活性化し、所属リンパ節へ遊走する。さらに、抗
原情報をT細胞へ提示し、抗原特異的なT細胞の増殖を誘導する。
樹状細胞はこの活性化の過程で、種々の細胞表面抗原の発現や炎
症性サイトカインの産生を増強することが知られている。最近で
は、ヒト末梢血CD 14陽性細胞由来の樹状細胞(Mo-DC)やCD34
陽性造血前駆細胞(CD34-DC)が、感作性物質処理により種々の
細胞表面抗原やサイトカインの産生を増強することがin・vitro実
験で明らかとなっている。すなわち、このような樹状細胞の特徴
を利用することで、加腕mでも化学物質の感作性を評価できる可
能性が示唆された。一方で、これらin・vitroで誘導する樹状細胞
は、細胞の安定な供給、均一一な細胞集団の確保、ドナー間のばら
つき等の課題があり、実用的な試験法の開発はまだ難しいのが現
状である。
そこで、こうした課題を克服するために、ヒト由来の細胞株で
あるTHP4細胞、 U937細胞(human histiocytic lymphoma cell line)
50・51)、MUTZ-3細胞(human acute myelomonocytic leukemia cell line)
52)において、樹状細胞の代替としての有用性が検討されている。
第1編で示したように、THP4細胞では、感作性物質により樹状
27

細胞の活性化の際に認められる様々な細胞表面抗原やサイトカ
インの発現が充進ずることが明らかとなった。さらに、感作性物
質により誘導されるCD86及びCD54の発現変化を指標とする、
in・vitro皮膚感作性試験法(human・Cell・Line・Activation・Test;
h-CLAT)の開発が進んでおり28・29)、これまでに64化合物を評価
した結果、動物実験とも良好の相関性(一致率86%)を示し、有
用な試験法である可能性が示唆されている30)。
しかしながら、感作性物質によるTHP-1細胞活性化の作用機構
に関してはよく分かっていない。そこで、炎症や免疫反応の制御
において重要な役割を果たすT:NF一・oc、 IL-8及びArPがTHP-1細
胞活性化の作用機構に及ぼす影響に着目した。
TNF一・Ctは樹状細胞の遊走に関与する38)だけでなく、∫η痂m実
験においてマウスの皮膚から高純度に精製したランゲルハンス
細胞のCD40やCD80の発現を主動させる働きがある53)ことが
報告されている。このことから、TNF一αが細胞表面抗原の発現変
化に影響する可能性が示唆される。
IL・一8に関しては、種々の炎症反応に関与することはよく知られ
ているが41)、皮膚感作性における役割は明らかになっていない。
一方で、Mo-DCやTHP-1細胞、 U937細胞において、感作性物質
で優位なIL-8の産生が報告されている18’31’51)。
また、ほとんどの細胞の細胞質内に5~10mM貯蔵されるATP
に関しては、細胞膜の損傷等によって細胞外へ放出され54)、細
胞膜上のP2 purinergicレセプターを介して細胞外シグナル分子と
して作用することが報告されている55)。P2レセプターのサブフ
ァミリーには、細胞膜チャンネルとして同定されたP2Xレセプタ
28

一56)と、Gタンパクに結合するP2Yレセプター57)の2つがあ
る。近年、樹状細胞においては、細胞膜上にP2レセプターを発
現する58)ことに加え、細胞外ATPによるP2レセプターを介し
たCD86、 CD83、 CD54の発現増強が報告されている59)。また、
THP-1細胞でも定常的にP2レセプターmRNAが発現しているこ
とや、ATPが細胞外へ放出されていることが報告されている60)。
そこで、本研究では、感作性物質による細胞表面抗原の充進に
及ぼす、TNF一α、 IL-8及び細胞外Arpの影響を検討した。
29

第1節 実験の部
1細胞と培養液
細胞、培養液ともに第1編、第1節と同じものを用いた。
2.被験物質とTHP-1細胞に対する処理方法
被験物質は、感作性物質として2,4-dinitrochlorobenzene
(DNCB).nickel sulfate hexahydrate (NiSO4),formaldehyde (FA),
propyl galla重e(PG)、eugenol(EU)、非感作性物質として、 sodium
lauryl sulfate (SLS)を用いた。全ての化学物質はSigma-Aldrich
(St. Louis, MO, USA)より購入した。 NiSO4、 FA、 SLSは予め生
理食塩水に溶解させ、また、D:NCB、 PG、 EUはdimethylsulfoxide
(DMSO;Simga-Aldrich)1こ溶解させた後に、培養液に添加した。
DMSOは培養液中の濃度が最高0.2%になるように添加した。
THP・一1細胞は24ウェルプレートに1x106cells/mL/ウェルにて播
種し、被験物質を添加した後、37℃、5%CO2条件下で24時間培
養した。各被験物質の処理濃度は、24時間培養後に70~95%の細
胞生存率になる濃度を設定した。DNCBは2.5μg/mしと5μg/mL、
NiSO4は85μg/mしと170μg/mL、 FAは15μg/mしと16.5μg/mL、
PGは29μg/mしと45μg/mL、 EUは130μg/mしと150μg/mL、 SLS
は54μg/mLを、それぞれ設定した。各被験物質に関して、実験
は少なくとも3回実施した。
また、細胞外のTNF一α、 IL-8あるいはATPがTHP-1細胞に及
ぼす影響を調べるために、24ウェルプレートに1x106cells/mL/ウ
ェルにてTHP一・1細胞を播種し、9.8~2500 pg/mしのリコンビナント
30

human TNF-ct (rhTNF-or; R&D Systems, Abingdon, UK), 80-50000
pg/mしのリコンビナントhuman lL-8 (rhlL-8;R&D Systems)、あ
るいは、30 ptMと250μMのATP (Molecular Probes, Eugene, OR,
USA)をそれぞれ添加した後、37℃、5%CO2条件下で24時間培
養した。
さらに、TNF一α中和抗体と化学物質の共処理による影響を調べ
るために、0.625 pg/mL抗TNF・一ct抗体(R&D Systems)を1時間、
細胞に適用した後に化学物質を24時間培養した。また、CD86及
びCD54の発現増強にP2レセプターが関与するかを調べるため
に、P2レセプターの競合的阻害剤であるsuramin(Sigma-Aldrich)
を1時間、細胞に適用した後に化学物質を添加し、24時間培養し
た。
3.細胞表面抗原の解析
化学物質を所定の時間、処理した後、細胞を回収し、O.1%bovine
serum albumine添加PBSで洗浄した。さらに、 Fcレセプターのブ
ロックキングとして0.Ol%Globlins Cohn Fraction ll, III
(Sigam-Aldrich)添加PBSを10分、4℃条件下で処理した後、抗
体染色を行った。細胞表面抗原の解析に用いたFITC標識モノク
ロナール抗体を以下に示す;抗ヒトCD54 (clone;6.5B5)
(DAKO)、抗ヒトCD86 (clone;Fun-1)、抗ヒトCD40 (clone;
5C3) (以上、 BD PharMingen)。また、 Isotype controlとして、
マウスIgG1をDAKOより、マウスIgG 1,KをBD PharMingenより
それぞれ購入した。抗体染色は遮光、4℃条件で、30分間行った。
細胞表面抗原の発現量測定にはFAcscaliburとcellQuest
31

software (Beck重on Dickinson)を用いた。死細胞を染め分けるた
め、細胞浮遊液にpropidium iodide (PI;Sigma-Aldrich) を最終
濃度0.625μg/mしで添加し、生細胞が10,000個になるまで測定し
た。細胞表面抗原の発現量の評価方法として、次式で表される相
対蛍光強度(Relative Fluorescence Intensity;RFI)を用いた。
RJFI (e/,) ==
MFI of chemical-treated cells 一 MFI of chemical-treated isotype control cells× 100
MFI of vehicle control cells 一 MFI of vehicle isotype control cells
MFI;平均蛍光強度
4.サイトカイン産生量の解析
細胞に化学物質を24時間処理した後、培養i上清を回収し、
ELISA kit(R&D Systems)を用いて、 TNF一αとIL-8の産生量を
側定した。
5.統計学的解析
解析したマーカーの発現変化を評価するために、統計学的な有
意差をStudent’s t-testにより解析した。
32

第2節 結果
1.DNCB、 NiSO4及びSLSによるサイトカイン産生と細胞表面抗
原発現の経時変化
本研究では、THP-1細胞に対し、DNCB(25μg/mしと5レしg/mL)、
NiSO4(85μg/mしと170μg/mL)、 SLS(54μg/mL)をそれぞれ適
用し、TNF一α及びIL-8の産生量、また、 CD86、 CD54及びCD40
の発現量の時間変化を、処理開始から1、3、6、9、12、24時間
後に調べた(Table 2)。無処理細胞の細胞生存率は24時間後でも
98%で変化しなかったが、高濃度のDNcB(5 pg/mL)及びNiso4
(170 pg/mL)における細胞生存率の減少は12~24時間の間に顕
著に起こり、24時間後には共に82%程度であった。DNCB(25
μg/mL)、 NiSO4(85μg/mL)及びSLS(54μg/mL)における細胞
生存率もまた、12~24時間の間に顕著に減少し、24時間後には、
いずれの場合も91~93%を示した。
DNCB、 NiSO4及びSLSによるTNF一αとIL-8の産生量の時間依
存性をFig.6に示した。高濃度のDNCB(5μg/mL)及びNiSO4
(170μg/mL)では、より多量のTNF一αの産生が認められ、6~12
時間で最大に達した。一方で、SLS、無処理群では顕著な変化は
認められなかった。また、Niso4によるIL・・8の産生は、検討した
2濃度共に、同じような時間依存的な増加が認められた。すなわ
ち、処理開始3時間では急激に産生量が増加し、6時間後以降は
緩やかに増加した。DNCBによるIL-8の産生量では、2濃度共に、
処理開始6時間までは急激に産生量が増え、それ以降はプラトウ
33

な挙動を示した。少なくとも処理開始3時間後には、溶媒処理群
に対して、DNcB及びNiso4によるTNF一一ctとIL-8の産生量は有
意に増加した(Table 3)。非感作性物質のSLSではいずれの処理
時間においても、TNF一αとIL・一8において有意な変化は認められな
かった。
さらに、DNCB、 NiSO4及びSLSによるCD86、 CD54、 CD40
の発現量の時間依存性をFig.6に示した。 DNcB及びNiso4によ
るCD86、 CD54、 CD40の発現量の増加は、 TNF・一ctとIL一・8の産生
に比べて、より遅れて誘導された。各細胞表面抗原の発現増加は、
処理開始3時問から6時間後に始まり、測定終了の24時間後ま
で継続した。24時間処病後には、DNcB及び:Niso4により、cD86、
CD54、 CD40全ての発現量において、統計学的に有意な増強が認
められた(Table 2)。 NiSO4では3時間後以降、 CD54の発現が有
意に増加し、また、9時間後以降、CD40の発現が有意に増加し
た。一方で、SLSではいずれのマーカーにおいても発現の増加は
認められなかった。
34

Table 2 Time dependent eiifects on cell viability indnced by DNCB,
NiSO4, and SLS.
Chemicals Testedconcentration
(ptg/mL)
Mean perceAt viability ± SD (e/o)
lh 3h 6h 9h 12 h 24 h
non-treated 98±1 98±1 98±1 98±1 98±1 98±1
DNCB5
コ
2
5
98±1
98i1
97 t1
96±1
96 ic 1
96±1
96±1
96土1
95i1
95±1
91±1
83士1
NiSO485
i70
98土1
98土1
97±1
96土1
97±1
96土1
96i1
95土2
96±2
94土2
91土2
82±3
SLS 54 98±1 96土1 97±1 96士1 96±1 93±3
Cells wer¢ treated with chemicals for 1, 3, 6, 9, 12, and 24 h. After-
treastnent, cell viability was measured by flow cytometry using PI
stak血g. Mean percent viability (土S.D.)was given fbr at least three
independent experiments and is sbown for each test concentration for each
chemical.
35
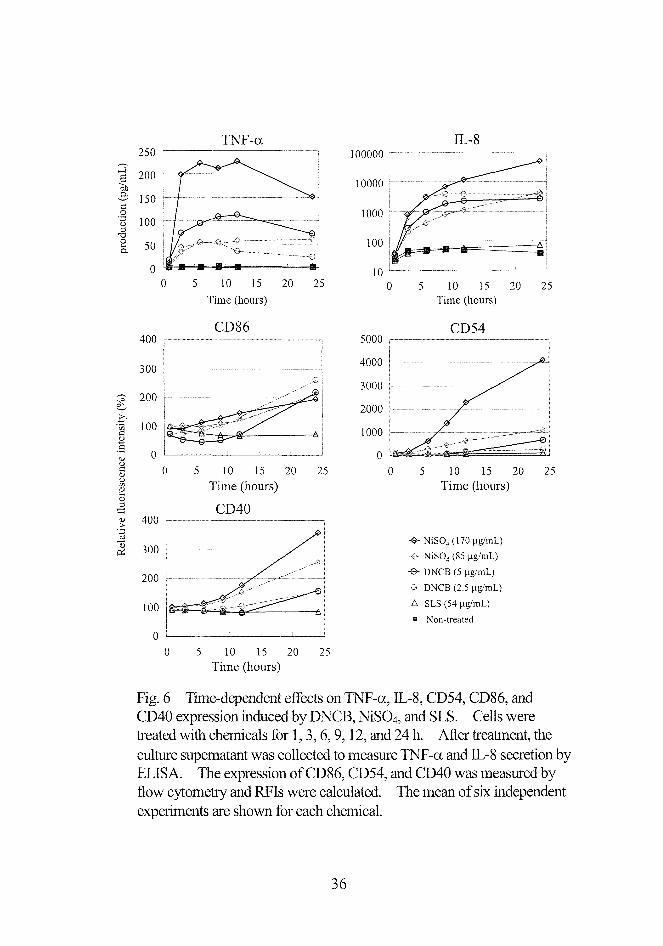
250TNF-ct
100000 ,
IL一一8
(貞量。D似)8刷ぢぞoa
(ま)意のお薫8碁。の戴◎ξω≧罵倒。
20e i-
1501由..
韮ooi
lOOOO i-
1000卜、
5Ψグ画くご1二。
400
fi)螺gi
董00
IO
鵡淵欝}一…V難献♪憂
。 5 10 15 2e 25 Time (hours)
CD86
300
200
100
o
繕
物艶肺病/a“一.e Ttr’“”
5 10 15 20 25 Time (hours)
CD40
5000
o 5 10 15 Time (hours)
CD54
20 25
4000
3000
2000
100e
o
o..
C簿冷 磨” �D一.
400
一一一ョ、
P
o 5 le 15 2e 25 Time (hours)
300
2eo
10e
o
,ノ;
ノル ア=二1鷲:講,,,,一’‘ ’N
5 10 15 20 25 Time (hours)
合NisO4(170鮮g/mL)
’“・’ NjSO, (85 pg/mL)
e 1)NCB (5 #g/mL)
ぞ桑一』1)NCB(25露9/mL)
Ax一 SLS (54 pg/mL)
’“ 垂≠高普@Non-treated
o
Fig.6 Time-dependent effects en TNF-or, IIL-8, CD54, CD86, and
CD40 expression induced by DNCB, NiSO4, and SLS. Cells were
treated with chemicals for 1,3,6,9,12, and 24 h. After treatment, the
cultUre supernatan:t was collected to measure TNF一αand正、一8 secretion by
ELISA. ”lhe expression ofCD86, CD54, and CD40 was measured by
且ow cytometry and RFIs were calculated. The mean ofsix independe凱
expetments are shown for each chemical.
36

Table 3 Statistical analysis of svllrface cell marker
up-regulatioR induced by DNCB, NiSO4, and SLS.
ChemicalsTested
むむおむ む アおユまむけ
(9/mL) 1h
P value (TNF-ct, n= 6)
3h 6h 9h 12 h 24 h
DNCB 52だ」
<0.05 く0.05 く0.Ol
<0,05 く0.Ol <0.Ol
<0.Ol く0.Ol くO.01
く0.Ol <0.01 くO.Ol
NSO4 85
170
ans
a閃
く0.0茎 く0.Ol <0.Oi <O.O蓋 くO.Ol
〈0.Ol <0.01 くO.Ol く0.◎1 <0.Ol
SLS 54 ans a闘
ans ans a無S ans
Chem紀a㎏
Tcsted
むけむむほ ねむむ
( ./inL) 韮h
1)vakエe(IL-8,貸謹6)
3h 6h 9h 玉2h 24 h
韮)NCB 52押)
お ユ
ns
獅
く0.05 〈O.Ol く0.Ol 〈0.0玉 くO.e5
く0.05 <0.05 く0.05 く0.01 〈O.O韮
NSO4 85
170
a薦
a聡
<0.Ol く0.05 く0.05 <0.05 <0。01
〈0.Ol <0.Ol く0.05 <0.Ol <0.Ol
SLS 54 ans ns ans a聡 ns
ntns
Che1揺ca㎏
Tested
むまユ をまユしじきなむお
( 9〆n正) l h
P value (CD86, n=3)
3h 6h 9h 12 h 24 h
DNCB 5(∠5
a a
ns
va a
ns
r
む む
麗郎
ぱ お
ns a a
聡s 〈o.el
〈O.Ol
N604 85
170
a鵬
a聡
a聡
a聡
ams
a聡
ans
a稲
く0.05 く0.Ol
ほns 〈0,01
SLS 54 a聡
a薦
a聡
a鵬
ans ams
ChemicalsTcstcd
むけむむけま ユむけ
(し・/mL) 韮h
P value (CD54, n=3)
3h 6h 9h 12 h 24 h
DNCB 52くノ
alas
allS
ns
ds
a a
遣捻
1 1
ヨ お
ns
T
〈O.Ol 〈O.Ol
nsa 〈o.el
NtsO4 85
170
ans
a照
<0.05 く0.05 <O.Ol 〈0.05 〈0,01
〈0,01 <0.Ol <0.01 く0.Ol <0.Ol
SLS 54 a11S ans ans ans llS ans
Chem沁a沁
Tcsted
むむめむじほセマおをユむ
(μ’/mL) Ih
P value (CD4e, n=:3)
3h 6h 9h 12 h 24 h
DNCB 52《」
な ね
絡㎎
き お
ns
曹
a a
ns
獅 aa
ns a 臼
ns 〈O.Ol
〈O.Ol
Ntse4 85
170
逸
1
a11S
ans
貧11S
acllS
ans
く0.01 <0.05 く0.Ol
<0.05 くO.Ol <0.Ol
S,LS 54 a11S ans ns a稲
a11S ans
P values were calculated with a Student’s t-test in order to
evaluate a statistical significafice between the relevant
vehicle cor血:℃1 cells and che㎡ca1-treated cells. Pvalues
were calculated for each tme. a not significant.
37

2.CD86、 CD54及びCD40の発現増強に及ぼすrhTNFαの効果
TNF一αがTHP-1細胞の活性化に関与するかを調べるために、 THP4
細胞に対して5濃度のrhTNF一αを24時間適用し、 CD86、 CD54、 CD40
の発現増強に及ぼすrhTNF-orの影響を調べた(Fig.7)。無処理細胞に
比べて、rhTNFαによる細胞生存率への影響はなかった。さらに、 CD54
に関しては9.8P9/血L rhTNF一αから、 CD40に関しては39.l p9/血L
rhTNF一αから、発現の有意な増強が認められた。 CD54での発現増強の
方が、CD40に比べて、 rhTNFαの影響は強力であった。 rhTNF一αによ
る濃度依存的な影響はCD54、 CD40で認められる一方で、 CD86では
認められなかった。
38

Gov’
CD86臣
CD54
CD40
(ま)属臣
(承)囲象
150
1.00
se i-
o1
3000 rwwwwnvwwwwwwww 120ee 1’
ieoo 1一
。
**
*
**
**
500
400
300
200
10e
o
一一g一一一 Cell viability
O g$ 多\βρヂ
<凶自。σ厳蔓(ま)
㎜8060鵜200rhTNF-ct concentration (pg/mL)
Fig.7 Effect ofrhrllNF-ct on CD86, CD54, and CD40
expresslon.
Cells were treated with rhnv-oc aloRe with five
concentrations for 24 h. rlThe 1ine graph represents cell
viability. Mean ±S.D. of CD86, CD54, and CD40
expression, as indicated by percent RJFI iRcrease, for tiitree
independent expeiiments is shown for each concentration
ofrlirlNFoc tested.
39

3.CD86、 CD54及びCD40の発現増強に及ぼすrh lL-8の効果
続いて、IL-8がTHP-1細胞の活性化に及ぼす影響を調べるために、
THP-1細胞に対して5濃度のrhlL-8を適用し、24時間培養し、 CD86、
CD54の発現増強に及ぼすrhlL-8の影響を調べた(Fig. 8、 Table 4)。無
処理細胞に比べて、rhlL-8による細胞生存率への影響はなかった。さ
らに、rhlL-8はCD86の発現量及びTNF一αの産生に対し、全く影響を
及ぼさなかった。同様の結果は、CD54においても確認された(図は
示さず)。
40

(承ご罠
150
10e
50
o
CD86 一一一g一一一 Cell viability
〈〉%“
タずずボ
lOO
80
60
40
20
0
<冨σ葭ぞ(ま)
(pg/mL)
rhlL-8 concentration
Fig.8 Effect ofrhl]L-80n CD86 expression.
Cells were trea紀ed with rh正、一8 at five
concentrations for 24 h. The line graph
represents cell viability. Mean ±S.D. of three
independe耐 exper㎞ents is shown :f()r each
concentration ofrllL-8.
Table 4 Effect of rhlL-80n TNF-oc production.
rh正、一8 conc.
@ (P9/血L)
TNF-oしproduction
@ (P9/mL)
0 0.8
80 0.7
400 0.9
2000 LO10000 LO50000 0.9
Cells were treated with rh正,一8 with five
conce加・ations fbr 24 h. After treatment the ラcultUre supernatant was measured f()r TNFα
secretion by ELISA.
41

4.TNF一αの中和抗体による影響
rhTNF一αによりCD54及びCD40の発現が増強したことから、感作
性物質によりTHP-1細胞から産生されるTNF一αが、 CD54及びCD40
の発現に影響する可能性が示唆された。そこで、本研究では、TNFα
の中和抗体とD:NCBあるいはNisO4の共処埋による影響を調べた。
TNFαの中和効果を確認するために、 ELISAによってTNF一α産生量を
定量した。その結果、Fig.4に示したように、 TNF一α抗体によって、
DNcB及びNiso4により産生したTNF一αは完全ではないものの、強力
に抑制された。5μg/mしのDNCB (P<0.01)及び、85μg/血L(P<O.01)
と170 pgtML (P<o.05)のNiso4においてそれぞれ、有意な抑制効果
が認められた。さらに、TNF一αの抑制によって、 D:NCBによるCD54
の発現が有意に抑制された。Niso4によるCD54の発現増強も、有意
に抑制された。CD54の発現抑制に加えて、 CD40の発現においても、
5μg/hiLのD:NcB及び:Niso4の2濃度で、有意な発現抑制が認められ
た。
TNF一αによる細胞活性化の効果をさらに検証するために、代表的な
感作性物質であるFA、 PG及びEUによるCD54の発現量への影響を
検討した(Fig.10)。その結果、 FA及びPGに関しては、 TNF-ocの顕著
な産生が認められ、16.5μg/mしのFA及び、29μg/㎡しと45μg/rnLの
PGでは、溶媒処理群と比べて、統計学的にも有意な増加であった。
一方で、EUではTNF-ocの産生は認められなかった。 CD54においては、
FA、 PG及びEUでいずれも有意な発現増強が認められた。さらに、
TNF一αの阻害によって、 FA及びPGによるTNF一αの産生とCD54の発
現増強は強く抑制された。EUにおいては、 TNFαの産生が無処理細胞
42

と変わらず、ほとんど認められず、また、CD54の発現増加が抗TNFα
抗体の影響を受けることもなかった。
43

0 0 0 0
2 8 4
玉(日≦鹿言○哨ぢぞ。a
seo
{.Fo 600
ご400臣 200
0
TNF一一ct 一一”垂香f Cell viabiHty
g 2.5 5
き 乞 DNCB彦
CD54
2。5 5
DNCB
ユ
75垂 窪
50ξ
25ま 0 )
竺(越9/mL) NisO4
soeo
4000
30eo
2000
10eo
o85 170 (緋g/mL)
NisO4
CD40450 「至誠
§3・・レ 轡1*蔑1繍麹幽幽胤,
醗翻a!1振一TNF一αnhへb(一)
隔anti-TNF一αmAb(+)
25 5 85 170 (魅g/mL)
しノ DNCB NisO4
Fig.9 Neutralization effect ofTNF-ct bioactivity in TH]P-1 cells
induced with DNCB and NiSO4. Cells were pre-treated with and
without anti一”IINF-or antibody for 1 h and then treated with DNCB and
NiSO4 for 24 h. After cultui“e, cells were analyzed for TNF-ct by
ELISA and for CD54 and CD40 expression by fiow cytometry.
Mean ±S.D. ofTNF-ct production and CD54 and CD40 expression
for three indqpendent experiments is shown for each chernical.
Statistical signficance was calculated with a Student’s 1-test
(**P〈O.Ol; *P〈O.05) .
44

(口§bの篇言。著ぞ。占
0000000
〆054、丙」(∠-
TNF-or87土28
lOOOG.o 800
Y. 600
臣400
2eo
o
:“1 th
CD54 +Ce韮l v藍ab霊Ilty
GPUE
lOO
000(U
8〆04。(∠0
≦9羨《(雰)
15 16.5
一ノ(#9/mL) FA
醗翻anti-TNF一(x mAb(一)
国anti-TNF-Ct mAb(+)
Fig.10 Neutralization effect ofTNF-ct bioactivity in THP-1
cells induced by EU, Pq Emd FA. Cells were pre-treated
with and without anti一一TNF一一ct antibody for 1 h and then treated
with EU, Pq and FA for 24 h. Mean±S.D. of”1]wtF-ct
production and CD54 expression for im’ee independent
experknents is shown for each chemical. Statisticai
significance was calcula愈ed with a S加dent’s減es£(**1)<0.01;
*P〈O.05) .
45

5.細胞外ArPがcD86及びcD54の発現量へ及ぼす影響
細胞外ArpがCD86及びCD54の発現量へ及ぼす影響を調べるため
に、THIP-1細胞に対して30μMあるいは250μMのArPを添加し、24
時間培養した。Arpはいずれの処理濃度においても、無処理細胞と比
べて、細胞生存率には影響しなかった。Fig.11には、 Arpを処理した
細胞のCD86及びCD54の発現:量のヒストグラムを示した。その結果、
外的なArPの適用により、 CD86及びCD54の発現が濃度依存的に増
強することが明らかとなった。
46

CD86
自マ} 櫛朝脚 樽昏F
富 言
嫡釜聾骨儲
⇔讐
爲
O
ゆW脚 櫛朝脚 価啓暦
尽 宕
埠蚤魯登
心⊃
。⇔偲
穏
1tie
一μ
ノ・~1
鎚
β~
ni 1 樋2FLt一}一lei2ht
樋3 擁IS
CD54
樋。
、蔭
5巳♂
齢、
ζ轟
ず」肇
ui 1 zi2 l ti3Fluorescence intensity
1詔
Fig.11 Effect of extracellular ATP on CD86 and CD54
expression. Cells were treated with ATP for 24 h. CD86
and CD54 expression was analyzed wnh fiow cytometry.
The histograms are for isotype control (dotted line) ,
nen-treated (shaded peak),30 pM ofATP-treated (thick
solid line),or 250μM ofArP-treated (d誼solid血}e).
[IJhese are representative data ofthree independent
expelrmeRts.
47

6.sura h3がDNCB、 NiSO4及びEUによるCD86、 CD54の発現へ及ぼ
す影響
細胞外ArpによりCD86及びCD54の発現が増強したことから、感
作性物質によって、THP-1細胞から放出されるATPが、 CD86及び
CD54の発現に影響する可能性が示唆された。そこで、本研究では、
P2レセプターアンタゴニストであるsurarr血と感作性物質の共処理に
よる影響を調べた。細胞に対して100 pMあるいは200μMのsuramiR
を1時間処理後、DNCB、 Niso4及びEUを添加して、24時間培養し
た。その結果、suraminの存在下では、 NiSO4によるCD86及びCD54
の発現が有意に抑制された。また、DNCBでは、統計学的な有意差は
得られないものの、suramin処理により、 CD86の発現増強の抑制が認
められた。EUにおいても、 suraminによって、 CD86の発現増強が抑
制される傾向が認められたが、CD54の発現増強には影響を与えなか
った。
48

CD86
(ま)H臣
3so r’””
3eo 1一!
25e
200
150
100
50
04500
+Ce難v萱ab1撫y
3.
塑内 gsc・
一一P IOO
-ii gs
xig
≦帥竃身(ま)
CD54
3000
1500
o
魯
D Suramin (一)
翻100 yM Suramin(+)
團200 geM Suramin(+)
2。5 5 85 170
しノ しノ DNCB NisO4
._[蚕_,..亡贈
130 150
EU
(pg/mL)
Fig.12 Effect of smm on CD86 and CD54 expression
indnced by DNCB, NiSO4, and EU. Cells were pre-cultui”ed
with or without P2 receptor antagonist suramin 1 h before
treatment with DNCB, NiSO4, and EU. Mean±S.D. efthree
independent experknents is shown for each cheinical. Statistical
signficance was calculated with a Student’s t-test (**P〈O.Ol;
*P〈O.05) .
49

第3節考察
生体での皮膚感作性発現の初期において、表皮の樹状細胞(ランゲ
ルハンス細胞)の活性化は重要な役割を果たしている。これまでに
Mo-DC、 CD34-DCに対して、感作性物質を処理すると、 in v∫加にお
いてもランゲルハンス細胞と同様な活性化が起こることが報告されて
いる17“22)。前章では、ヒト由来細胞株であるTHI)一・1細胞が、様々な感
作性物質によって種々の細胞表面抗原の発現やサイトカインの産生量
を増加させ、樹状細胞に類似の応答性を示すことを明らかにした。そ
こで本章では、感作性物質によるTHP-1細胞の活性化の作用機序を解
明するために、TNF一α、 IL-8及び細胞外Arpの関与について調べた。
その結果、キネティクス:解析により、DNCB及びNiso4により産生
されるTNFαやIL-8が、処理開始3時間で有意に増強し、それに続い
てCD54、 CD86、 CD40の発現が増強することを明らかにした。そこ
で、TNFαが細胞活性化に与える影響を調べた結果、 rhTNFαは濃度
依存的にCD54及びCD40の発現を増強した。さらに、抗TNF一一〇c抗体
より細胞外の”INF一一〇cを中和すると、感作性物質によるCD54及びCD40
の発現増強が顕著に抑制された。このことから、感作性物質により産
生するTNFαがオートクラインに細胞に作用し、 CD54及びCD40の
発現増強に寄与していることが示唆された。
樹状細胞においては、感作性物質によるCD54及びCD40の発現増
強に、TNF一一〇cがオートクラインに作用することはこれまで知られてい
なかった。しかしながら、最近、CD34-DCにおいて、 DNCBによる
CCR7の発現増強にTNFαのオートクライン作用が関与していること
が報告されている6i)。よって、 TNF.αを介した細胞表面抗原の発現変
50

化はTHP-1細胞だけでなく、樹状細胞でも起こり得る作用機構である
ことが示唆された。
マウスを用いた検討より、TNRαが樹状細胞の遊走や活性化に重要
なことはよく知られている62)。また、ヒト皮膚から分離したランゲル
ハンス細胞を用いて、rhTNF一αの影響を調べた報告では、 CD40及び
CD54の発現増強が認められており、 TNF一αがランゲルハンス細胞の
活性化に関与することが報告されている63)。本研究でも、THP-1細胞
ではthTNF一αによりCD40及びCD54の発現増強が認められたことか
ら、THP-1細胞のTNF-orに対する応答性はランゲルハンス細胞と類似
していた。一般に、生体において、ランゲルハンス細胞は、炎症刺激
によりTNFαを産生するが、一方で、ケラチノサイトも多量のTNFα
やGM-CSFを産生することが報告されている38)。よって、皮膚から
のしCの分離精製が不十分だと、コンタミしたケラチノサイト由来
のサイトカインの影響を無視できなかった。しかしながら、近年、マ
ウスの表皮から高純度にランゲルハンス細胞を精製し、ケラチノサイ
トの影響を排除できる条件下では、rhTNF一αによりCD40の発現が顕
著に増強するが、CD86の発現はrhTNFocの処理濃度、時間に関係な
く、変化しないということがSalgadoらによって報告されている53)。
これは、THP-1細胞において、 TNF-CtがCD86発現に関与しないとい
う結果とよく似ていた。TNF一αに対するCD86の不応答性がTHI)・一1細
胞だけでなく、ランゲルハンス細胞でも起こることが示唆された。
本研究では、DNcB及び:Niso4によるcD54の顕著な発現増強には、
TNF一αが強く関与することが示された。一方面、 EUでは、 TNF一αの産
生が無処理群と同様に増加しないにもかかわらず、CD54の発現増強
が認められた。また、抗TNFα抗体によってCD54の発現に変化が影
51

響を受けることもなかった。これは、EUに特異的な反応の可能性も
考えられるが、CD54の有意な発現増強にTNFαが関与しない経路が
存在することを示唆しているのかもしれない。近年、BoiSleveらは、
cD34-Dcにおいて、Niso4によるccR7の発現増強がTNF-ctを介さず、
p38 mitogen-ac轍ed prote血㎞ase(p38 MAPK)や。・一jun N-te㎜mal
kinase (JNK)のリン酸化を活性化して、直接的に誘導されると報告
している6D。このことから、 THP-1細胞においても、 CD54の発現増
強を直接的に誘導するシグナル伝達経路が存在する可能性があるかも
しれない。
さらに、TNF一α同様に短時間の感作性物質処理で多量に産生する
IL-8においても、細胞活性化への影響を調べた。 DNCBやNisO4によ
り産生するIL-8よりもはるかに高濃度のrhlL-8を適用しても、細胞表
面抗原の発現やTNFαの産生量に明らかな変化は認められなかった。
このことから、IL-8は、感作性物質による細胞活性化に伴う結果とし
て産生されるものの、オートクラインに作用して細胞の活性化へ寄与
する可能性は低いと考えられた。
また、細胞外ArPによりTHP4細胞のcD86及びcD54の発現が増
強することが分かった。また、P2レセプターアンタゴニストである
suraminを用いることで、感作純物質によるCD86とCD54の発現増強
が部分的に抑制されたことから、細胞外Arpを介した細胞表面抗原の
発現増強の可能性が示唆された。ヒト樹状細胞では、P2X及びP2Yレ
セプターが発現すること58)、さらに、細胞外ArpによってCD 86、
CD80、 CD54及びCD83といった細胞表面抗原の発現が増強すること
59)が報告されている。また、近年、THP-1細胞においては、 P2レセプ
ターの発現や細胞内Arpの僅かな放出が認められている60)。 Arpは
52

ほとんどの細胞で細胞質内に5~10mM存在し、細胞膜の傷害により放
出されることが示されている54)。以上より、細胞外Arpが感作性物
質による細胞活性化に重要な役割を担っている可能性が示唆された。
前章において、感作性物質による細胞の活性化には若干、細胞毒性
が誘導される処理濃度が必要である可能性が考えられた。本章のキネ
テイクス解析においても、DNCB及びNiSO4によるCD86、 CD54、 CD40
の有意な発現増強は、僅かに細胞生存率が低下する24時間処理後に誘
導された。しかしながら、DNcB及びNiso4によるTNNF-ctとIL-8の
有意な産生は、細胞生存率の低下がない3時間から12時間の処理の間
でも認められた。このことから、少なくともTNF一αとIL-8の産生は、
細胞生存率の低下に依存せず、細胞毒性とは独立した細胞応答である
可能性が示唆された。サイトカイン産生や細胞表面抗原の発現増強を
伴う細胞の活性化と、細胞毒性との関係のさらなる理解に向け、今後
の研究が期待される。
53

第4:節 小活
本章では、感作性物質によるTHP-1細胞の活性化の作用機序を:解明
するために、TNFα、 IL-8及び細胞外Arpの関与について調べた。そ
の結果、感作性物質により早期に産生するTNFαの産生がオートクラ
インに細胞に作用し、CD54及びCD40の発現増強に寄与することが
示唆された。一方で、感作性物質によるCD86の発現増強には、少な
くともTNFα、 IL-8は関与せず、細胞外Arpが部分的に関与する可能
性が示唆された。以上より、感作性物質によるTHP-1細胞の活性化は
TNF一αのオートクライン作用及び細胞外Arpによって部分的に制御さ
れ、樹状細胞における細胞活性化と同様の作用機構を介して誘導され
ることが示唆された。
54

第2章割49)
THP-1細胞活性化に及ぼすシグナル伝達経路の役
近年、感作性物質による樹状細胞の活性化の作用機構に関して、細
胞内シグナル伝達経路、特に㎡沁gen-activa綴prote血㎞おe(MAPK)
の研究がいくつか報告されている。MAPKカスケードは細胞の増殖や
分化を制御するシグナルの伝達機構としてよく知られており、MAPK
ファミリーには、p38 MAPK、 exなace11u1飢si帥a1-re即1a面㎞ase(ERK)、
c-J㎜N£e㎜inal㎞}ase(JNK)が存在する64・65)。 A㎡g㎞らば樹状細
胞に対しDNcB及びNiso4を処理するとp38 MAPKのリン酸化が誘
導され、また、p38 MAPKの活性阻害によって、 Niso4処理における
CD86発現増強が抑制されることを報告している66)。さらに、 Bois16ve
らは、樹状細胞において、DNCBによるTNF一αの産生にp38 MAPKの
活性化が関与することを報告している61)。しかしながら、DNCBや
NisO4を処理した際のTNF一αの産生や細胞表面抗原の発現増強におけ
るシグナル伝達系の役割に関しては十分に明らかになってない。また、
樹状細胞の活性化におけるp38 MAPKに比べ、 ERKの役割に関しても
ほとんど明らかになっていない。それゆえ、本研究ではTHP4細胞に
おいて、DNCBとNiso4によるTNFαの産生や種々の細胞表面抗原の
発現増強に関して、p38 MAPKとERKの役割について検討した。
55

第1節 実験の部
1 細胞と培養液
細胞、培養液ともに第1編第1節と同じものを用いた。
2、p38 MAPKとERKのWestern Blot解析
24ウェルプレートにlx106cells/m L/ウェルにてTHP-1細胞を播種し、
5 yg/丈nL D:NCBあるいは170μg/血L NiSO4(共にSigma-Aldkich)を添加
した後、37℃、5%CO2条件下で1時間培養した。また、 p38 MAPK及
びERKのリン酸化を阻害する実験では、 DNCB及びNiso4を添加す
る前に、p38 MAPK阻害剤としてSB203580(Calbiochem, La Jolla, USA)、
ERK阻害剤としてPD98059(Calbiochem)を1時間、前培養した。 DNCB
及びNiso4は、それぞれDMso、生理食塩水に溶解させた後に、培養
液に添加した。DMSOは培養液中の濃度が最高0.2%になるように添
加した。
その後、細胞ライセートを調製するために、細胞を回収、洗浄した
後、SLSサンプルバッファー(0.5M Ths-HCI、 pH6.8、10%Glycerol、
2%SLS、5%2一メルカプトエタノール、 O.0025%プロモフェノールブル
ー)を加え、100℃、5分間加熱することにより可溶化した。さらに、
可溶化液を遠心分離して上清を回収し、細胞ライセートとした。細胞
ライセートは10%sDs一ポリアクリルアミドゲル(TEFco, Tokyo,
JAPAN)により泳動後、 polyvinylidene difiuoride(PVDF;Millipore,
Bedford, USA)膜に転写した。 PVDF膜は、 phospho-p38 MAPK(Cell
Signaling Technology, Danver, USA)及びphospho-ERK(Cell Signaling)
に対する一次抗体(1000倍希釈)と一晩、4℃でインキュベートした。
56

一次抗体反応の膜は十分に洗浄し、phospho-p38 MAPK(3000倍希釈)
及びphospho-ERK(lOOO倍希釈)に対するHRP標識ヤギ抗ウサギIgG
抗体と共に、室温で1時間インキュベートした。二次抗体反応の膜は
ECL solut:ion (Amercha血, Zurich, SwitZerland)を用いて、化学発光検
出を行った。
3.細胞表面抗原の解析
24ウェルプレートに1x106cells/m L/ウェルにてTHP-1細胞を播種し、
5 P9/mlL DNCBあるいは170μ9/mL NiSO4を添加した後、37℃、5%CO2
条件下で24時間、培養した。また、p38 MAPKあるいはERKがCD86、
CD54及びCD40の発現に及ぼす影響を調べるために、 SB203580ある
いはPD98059を1時間前処理し、 DNCB及び:NiSO4を24時間、培養
した。細胞表面抗原の発現解析方法は、第2編、第1章、第1節に示
したとおりである。細胞表面抗原の発現量の評価方法として、MFIを
用いた。
4.サイトカイン産生量の解析
第2編、第1章、第1節に示したとおりである。
5.統計学的解析
第2編、第1章、第1節に示したとおりである。
57

第2節結果
1.DNcB及びNiso4によるp38 MAPK、 ERKのリン酸化とp38 MAPK
及びERKに対する阻害剤の効果
THP-1細胞に対して、 DNcB及び:Niso4を1時間処理することによ
るp38 MAPK、 ERKのリン酸化をウェスタンプロットにより:解析した。
その結果、Fig.13及びFig.14に示すように、 DNcB及びNiso4は、
DMSO及び無処理群に比べてp38 MAPKとERKのリン酸化を強く活
性化した。このことから、THP-1細胞において、 p38 MAPKとERKの
シグナル伝達系は存在し、感作性物質により活性化することが示され
た。さらに、P38 MAPK及びERKのリン酸化に関して、 P38 MAPKの
阻害剤であるSB203580と、 ERKの上流因子であるMEKの特異的阻
害剤であるPD98059の効果を検討した。その結果、阻害剤を前処理す
ることにより、阻害剤濃度依存的に、p38 MAPK及びERKのリン酸化
が抑制された。
58

a)
total-p38 es・ ・・一・・…,・…i一・….・・・…一一,・・
ボ総評“
ぎ轟評評
1 1 0, α
.寧
穿譜
届 L o・ α
(℃Φ細邸O靹細一qO9\()のΣ(瞬触O承)
智の器輯嶺$。〉慧笛葭
切
Fig.13 DNCB-and NiSO4-induced phosphorylation ofp38
MAPK and dose dependent effcct ofp38 MAPK inhibitor.
Cells were pre-incubated with or wnheut SB203580 or
PD98059 at the concentrations indicated for 1 h and followed
with DNCB at s Ftg/ml or NiSO4 at 170 pg/mi for l h. (a)
After treatment, the phospho-p38 MAPK and total-p38 were
analyzed by Western Blot analysis. Sirnilar results were
obtained in three different expeiimeRts. (b) The relative
band imensities ofthe phosphorylation of p38 MAPK in
DNCB一 or NiSO4-treated cells relatively to DMSO-treated or
non-treated cells were detemimed by densitomeuic analysis.
59

phospho-ERK 1/2 wwww・wwwwwwew.・…一一・…,...・一ti,・一・・・…v・.,,
1.’,.v!・””tl麟騨騨縮.
ぐ ド じ
識盤 、
の
群“鮮
ぱ
1.61
1 0. α
ぜ
-. L α α
(℃O輯鱒O』↑一d【O葭\○のΣ(照触Oま)
盈。。器朝三℃竃の。ξ滋。飯
切
回 ぶ獣
Fig.14 DNCB-and NiSO4-induced phosphorylation of
ERKL and dose dependent effect of ERKL inhibitor. Cells
were pre-incubated with or without PD98059 at the
concentrations indicatod fer 1 h etad followed with DNCB at
5μg/ml or NiSO4 at 170 ltg/血1 fbr l h. (a) After
treatment, the phospho-ERK I/2 and total-ERK I/2 were
analyzed by Westein Blot analysis. Similar results were
obtained in imree different expertments. (b) The relative
band h並ensities ofthe phosphorylatioll of ERK.一つNCB-or
NiSO4-treated cells relatively to DMSO-treated or
non-treated cells were detei抽ed by densitometric analysis.
60

2.・DNcB及び:Niso4によるTNF-ctの産生に対するp38 MAPK、 ERKの
影響
THP-1細胞において、 DNcB及び:Niso4の1時間処理により、 p38
MAPK及びERKの活性化が認められたことから、続いて、 P38 MAPK、
ERKがDNcB及びNiso4により早期に産生するTNFαに及ぼす影響
を調べた(Fig 15)。その結果、 DNcB(P<o.ol)及びNiso4(P<o.01)
では溶媒処理群に対し、TNF一αの有意な産生が認められた。さらに、
DNCBで認められるTNF一αの産生は、 p38 MAPKの阻害剤である
SB203580の濃度依存的に抑制され、0.3μM及び3μM SB203580では、
60%以上の抑制効果が認められた。一方面、:Niso4によるTNFαの産
生ではSB203580の影響はなかった。また、 DNCBによるTNF-ccの産
生に対し、ERKカスケードの阻害剤であるPD98059の影響はなかっ
た。しかしながら、PD98059の濃度依存的に、 Niso4によるTNFαの
産生が抑制され、また、その抑制効果は3 pM PD98059で57%、30μM
PD98059で82%に達した。このことから、 DNcB及びNiso4により
早期に産生するTNF一αはそれぞれ、 p38 MAPK及びERKによって強
く制御されていることが示唆された。
61

奴
獣
綻鷺、:こ鍍㌧
蟻
鑓溶
断/賊(
**
****
a)
2ee
AU O O
ご」 0 《ゾ
ー
重■■
(貞謹b。瓢言ε。ぞ。ζ
8一
。
ノジぐ 禽麟i遜続。・〉餅 ウ描灘㌧謙繋1ら藤鑓ttt rt・ ㌧… 詩田
b) 200貧
章150彗.: 100
筥麗5・繕 .
評 評
**
** *
wh
*
50
o
ぷ
山 高州川州へ
試評“らシ鞭“ ボ鮮“o 試鍵
討
Fig.15 implication of p38 MAPK and ERK on TNF一一〇c release
induced by DNCB Emd NiSO4. Cells were pre-cultured wnh or
wnhout (a) SB2035800r (b) PD98059 at the concentrations
indicated for 1 h and followed with DNCB at 5 pg/ml or NiSO4 at
170pgtml for 24 h. After treatment, the supematants were
recovered and exainined for TNF-ct release by ELISA. The
results were shown as the Mean±S.D. (n=3). Statistical
significance for the inhibition effect was calculated by Student’s
t一一test (**p〈o.ol;*p〈o.os).
62

3.p38 MAPKがDNcB及びNiso4によるcD86、 cD54及びcD40の
発現に及ぼす影響
続いて、SB203580がDNCB及びNiSO4によるCD86、 CD54及び
cD40の発現増強に及ぼす影響を調べた(Fig.16)。その結果、 DNcB
及びNiso4いずれにおいても、溶媒対照群に対して、 cD86、 cD54及
びCD40の発現の有意な増強が認められた。また、 SB203580を適用し
たことによる細胞生存率の顕著な変動は、いずれの処埋濃度において
も認められなかった。さらに、DNcB及びNiso4によるcD86の発現
は共に、30 pM SB203580により、40%以上の抑制が認められた。一
方、CD54及びCD40においては、 SB203580の濃度依存的に、ベース
の発現が増強する傾向が認められたため、この影響を無視できる濃度
範囲(0.03μM~3μM)で検討を行った。その結果、D:NCBによるCD54
及びCD40の発現増強は、 SB203580の濃度依存的に抑制されたが、
Niso4では抑制効果は認められなかった。
63

it ’g,o
cellviability tijrr 2g
諺21
㌦
@㌧@ \
@く \
A \
@ ㍉@ \A ミ
A い_v≒\博 、 曳 藍、 藍 ㌦ ㌦ \
@ 忌磨@ \ \
@樗_ ‘ \
_ ㌧
@\@\ \
w ㌧
@ \
ユ 、
A 、、 \
堰@糠
@ \ ピrj マ険㍉ \♂
\
@ 、メ@\@ビ 、 喧
ヨ ㌧
@\m ㍉
@、 〉@㌧ ㌧ く
@ぐ ノ \@㌧ 、 ‘ \
d ㌧ へ
@く ビ
@ ㌧_ \@卜 ㌧
@、 、ロ曳腰く、、
A \_ ㌧ 〉
@“ 〉
\
@ \@、@\@\ 〉ュ 究 尺
@ 、 \A 、
@ \
、 <
@\@\ xメ@ ㌧
@ 悔 曳@♂モ@ピ \
�@ \ <
@ 、 @ 美
_ v \へ\
堰@ 斗
j く ♂
、 ぐ
@\ \く
@く_瓦 \
@ \磨@\ 、 樗
@、@、く 、
@、_ ㍍
@ 邑@ぐ\\玄 外㌦\・
、 、
@‘ う \
@、 、
@\@ !@、_ \
_ 、 \
C 、
d、 、 Ψ
_} {
w \
? 》
@ 斗t、 イ \⊂
ュ ㌔
?
曳 ㌧
@、d \
@\@、 c
@ぜ@ ’
@、
CD86
50
O AU O O
4 3 2 韮
{』Σゆ◎OQ⇔
0
ぷぷ雇滋
ノ も
懸驚
*
騨も潤一こ1~
麟愚
讐蝶:
越謡雛勢掌潔昏
磐い談厩響癬轡ヴ
ぷ
[一**
㌧$雰こ〉ンヅビう
\む&ノ勢誠3\
収獄壕噸
鷲繍、
療
、顧い鼠へ厨ミ暮、V娘
Nピ〆い㌧ 磨ミ/
購翻蕊ひ
・濾。
曝藤:∴
脇姶\
漁∴
叡綜㌧ぐ一ケぐ
鳶こ.~
ン勲も
CD54
、ψ飯
騨メ岬認
伽
0 0 0
Qノ 〆0 3
H』Σ寸いQQ 0
40 0 0 ◎
3 2 ・蓋
類Σ○寸OQ
0
****
ふ。類∬捨eへ細へ旧
鴨訳難いバ吻\・“
i吻
1醐
1繭
黙脳憲魑
警懇隆簿蒸、覧
霧∴窯翁熱燃
謡賠齢獄潟
CD40
**
セ
r”ltxll.
獄然:㌧〕
1’i’・
添’1’堰fi’t”’
ぴぎきぽ
遥、
総糧
“\
》ハ
劉∫甕
:髄
:憲
1慈i憶
鰯\ぐ
雪\・播〉
誉∴
繕劉
粥㌧/ _《
(’ij...
’i’)’i
硬らρず()sも⑩ψ
粛囲、
試
。避x$o・紳ψ ち
譜瀞Fig.16 implication ofp38 MAPK oR CD86, CD54, and CD40
expression induced by DNCB and NiSO4. Cells were treated with or
without SB203580 at the concenmations indicated for 1 h, followed with
DNCB or NiSO4. in cell viability and CD86, the results ofO.03 FtM
SB203580 were omitted since they were sirnilar wnh those of O.3 ptM.
The results were shown as the Mean ±S.D. from at least three
independent expeiiments. Statistical sigriificance forthe initibition effect
was calculated by Studenfs t一一test (**P〈O.Ol;*P〈O.05).
64

4.ERKがDNcB及び:Niso4によるcD86、 cD54及びcD40の発現に
及ぼす影響
さらに、PD98059がDNcB及びNiso4によるcD86、 cD54及びcD40
の発現増強に及ぼす影響を調べた(Fig.17)。 PD98059を適用したこと
による細胞生存率の顕著な変動は、いずれの処埋濃度においても認め
られなかった。さらに、DNcB及びNiso4によるcD86、 cD54及び
CD40の発現増強はいずれも、 PD98059の濃度依存的に抑制された。
DNCBによるCD86、 CD54及びCD40の発現増強に対する30μM
PD98059の抑制効果はそれぞれ、40%(P<0.01)、40%(P<0.01)及び
25%(P<o.01)であった。また、Niso4によるcD86、 cD54及びcD40
の発現増強に対する30μMPD98059の抑制効果はいずれも36%
(P<0.01)以上であった。
65

Cell viability
0 0 0 0 0 0
0 8 6 4・ 2
ま(ま)眺起【唱ρ邸刷〉=のQ
ウ 犠
@臥・驕@ り@㌧ \q 、
@、@〈@ ㍉ \
@ \ 、「 『
@ く@Ψ 〈
@ 、@、
\ 、
@く Nw \
@㌧驕@~ \
_ ÷ ㌦
@ ’@ ’@}A 、、’
@㌧刀@ 、\
秩@ へ
@、ュ ㌧
@ へ
@\p へ
@ 、@ \@く磨@ 、
@♂A ㌧ リ
@ 《 \
仲n
@\ 瓦
…
軋
�@ 、
_ ㍉ \ \
?や, 0 9、 、 》、 、
dへ㌦\
…
㌧ 、耳㍉曳 ㌻、 、 鳥
、\ ㌧\、、
_\㌦㌧\
CD86
40
0 0 0
つ」 2 1
嗣』≧ゆ◎OQQ 0
**
** **
鍵幾
1
μ.
覧J蟻陰
蝋燃
潜諺
撫麟
藻凝響
塾藁ゼ
へづミモふ
誌麟蘇嘆脳・
**
ぎき誉&N
\《鉱ρごノ畿、心 ゆ与 ♪重\逡鄭 、\、A、こN v\
、♪へ / 、/~、 \\
ル党ぴ“襟き討議
ε夢ヤΣ嘆ノ暴ソ款 }ぴ
》
与ミ暴灘譲
N・く、でミ、碧謡〆誉鐘濾惣
\へ > V\ ㌧
『翻旨ざい源誉
♪ ㌔
妻“寒ぐ 獣竣Ψ
多爵携㍉竪㌧
鑓騨
「
㌧ \
CD54
伽 0 0 0
Q/ 6 3
鼠Σ尋いOQ
0
40@30 20 憩
起】≧○ぐOQ
04DC
砥掌“買いΨ駐5ψ艶メ
5“
翻認絆
マ
臼評
、ボ
0 9 評ポ
ぜボげ
9
ぐ \ ノ㌧
・寸さい’\\/》へ
\ \\ 〉 \
ピ㌧騎
)ミ》\\ ノへ \ / \
へ》㌧
湿嫡期
**
2
謡ぐ繰認
/嚇ぐ八\\\
\/ハ
”\、ミ腰 \
ぺ
N〈鄭N
}、\ へ
@ソノ浸
\、路凶 植
\邸3
2軽ハ
、馬 や
☆で慈導\
璽“
**
-/\〔蓉じ ㌔ノ・β\
〉 次こしノ\ \ 斗
r、》》 藍\蒜暴》瓢ごへ
ごV \\ \ \ご“眠\%へ鳶、∴
詮7,冬・惚・惑ゾψ㌦、 ㌔“
八
/\ /\r
****
λ\〉\\ /マ、、y・
∵も、“ 〉\‘
?\\\ ・ \
Fig.17 impiication ofERK on CD86, CD54 and CD40 expression
indviced by DNCB and NiSO4. Cells were treated with or witheut
PD98059 at the concentratiofls indicated for 1 h, followed with DNCB
or NiSO4. The results were shown as the Mean ±S.D.丘om at least
thiAee indePendent expel士nents. Statistical signi且cance:for the
ir血ibition effect was calculated by StUden:t’sかtest (**P<0.01;
*P〈O.05) .
66

第3節考察
本章では、感作性物質による細胞活性化の作用機構の解明を目的に、
細胞内シグナル伝達系であるMAPKに着目した。 MAPKファミリー
には、p38 MAPK、 ex甑ce11曲si帥al-re毅lat(対㎞ase(ERK)、 c-J㎜
N・一teirninal㎞ase (JNK)が存在する。一般的に、 ERKは細胞増殖や
分化、p38 MAPKや∫NKはストレス応答やアポトーシスに関与するこ
とが知られているが、免疫反応においても多くの重要な役割を果たす
ことが明らかとなっている67)。本研究ではp38 MAPK及びERKの働
きについて検討した。
その結果、5μg/mL DNcB及び170μg/ML Niso4はいずれも、
p38MAPK、 ERKのリン酸化を活性化した。さらに、これらのリン酸
化がp38 MAPKの阻害剤SB203580及びERKの阻害剤PD98059によ
って、効果的に抑制された。これまでの研究で、THP-1細胞において
は、DNCB(05μ9/m L、25μ9/血L及び5μ9/血L)とNiSO4(17μ9/血L、
85μg/血L及び170 ptgtML)によるp38 MAPKとERKのリン酸化が濃
度依存的に誘導されるのに対し、S:LS(45μg/mL、54μg/血L>では変
化しないことが示されている(図は示さず)68)。本章では、さらに、
DNCBによるTNF一αの産生がSB203580により選択的に抑制され、同
様にして、Niso4によるTNF一αの産生がPD98059により選択的に抑制
されることを明らかにした。いずれの場合も、TNF一αの産生の抑制効
果は最大で60%に達し、SB203580あるいはPD98059がTNFαの産生
に対して優れた抑制剤であることが示された。
近年、CD34-DCやMo-DCを用いて同様の検討がなされている。
DNCBを処理したCD34-DCにおいては、 P38 MAPKが活性化し、20μM
67

SB203580によりTNFαの産生が抑制した61)。また、 NiSO4を適用し
たMo-DCでは、 TNF一・ocの産生が10μM PD98059により50%、抑制さ
れることが報告されている18)。本研究及びこれら知見から、感作性物
質によるTNF一αの産生にP38 MAPK及びERKは強く関与するものの、
それらの役割は感作性物質によって大きく異なることが示唆された。
さらに、THP4細胞において、 D:NcB及びNiso4によるcD86の発
現増強にp38 MAPKが関与することが明らかとなった。 Mo-DC i 8)や
CD34-DC 61)でも、全く同じことが報告されていることから、 THP-1
細胞がMo-DCやCD34-DCと同様の作用機構でCD86を発現増強する
可能性が示唆された。本研究ではさらに、ERKの活性化阻害により、
DNCB及びNiSO4によるCD86の発現増強が抑制された。この結果と
同様に、:Niso4を適用したcD34・・Dcにおいて、 cD86の発現増強が
PD98059により部分的に抑制されることがBoisleveらによって報告さ
れている61)。このことから、ERKはp38 MAPKと同様に、 DNCB及
びNisO4によるCD86の発現に寄与する可能性が示唆された。一方で、
本研究及びBoisleveらの結果とは矛盾する結果が、 Aibaらによって報
告されている。Aibaらは、二人のドナーからMo.DCをそれぞれ誘導
し、NiC12及びDNCBによる細胞活性化の作用機構を調べた結果、 ERK
が細胞活性化に対し抑制的に働く可能性を示している18)。ERKの働
きの違いに対し明確な理由を説明することは難しいが、De・Smedtらは
感作性物質によるMo-DCの細胞表面抗原の発現やサイトカインの産
生量は、ドナー間によって全く異なることを報告している2め。そのた
め、Mo-DCにおけるERKの役割については、より多くのドナーの
Mo-DCを用いた検証が必要かもしれない。
DNcB、 Niso4によるcD54及びcD40の発現増強におけるMAPK
68

の役割はこれまでにMo-DCを用いて検討されている。しかしながら、
p38 MAPK阻害剤はいずれの細胞表面抗原の発現増強を抑制しない18)。
また、ERK阻害剤もNisO4によるCD40の発現増強に、ほとんど影響
しないことが報告されている66>。これら検討では、DNCB、 Niso4に
よるCD54及びCD40の発現が顕著に増強しておらず、さらにドナー
を増やして検討する必要があるように考えられる。THP-1細胞におい
ては、SB203580存在下でDNCBによるCD54及びCD40の発現増強
が抑制された。一方、PD98059では、 DNcB、 Niso4によるcD86、
CD54及びCD40の発現増強が部分的に抑制された。本研究結果は、
Mo-DC及びCD34-DCを含め、 CD54とCD40の発現増強におけるERK
の役割を証明した初めての報告である。
MAPKの特異的な阻害剤を用いることにより、細胞活性化は阻害さ
れるものの、DNCB及びNiso4によるTNF-Ctの産生や細胞表面抗原の
発現増強は完全に抑制されていない。すなわち、p38MAPKやERK以
外の他のシグナル伝達系が感作性物質による細胞活性化に関与してい
る可能性も考えられる。Boislさveらは、 CD34-DCにおいて、もう一つ
のMAPKカスケードであるJNKの阻害剤が、 DNcB及びNiso4によ
るCD86の発現増強を部分的に抑制することを報告している。よって、
感作性物質によるTHP-1細胞の活性化には、 JNKもまた関与している
可能性が示唆される。
69

第4節小活
本章では、感作性物質によるTHP-1細胞の活性化の作用機i構を解明
するために、細胞内シグナル伝達系の役割について調べた(Fig.18)。
その結果、DNcBとNiso4は共にp38 MAPK、 ERKをリン酸化するも
のの、TNFα及び細胞表面抗原の発現に及ぼすp38 MAPK及びERK
の役割は化学物質によって大きく異なることが分かった。D:NCBは
p38 MAPKに対して優位に作用し、 TNF一αの産生や細胞表面抗原の発
現を最大限に誘導する上で、P38 MAPKが重要な役割を担っているこ
とが明らかとなった。また、Niso4によるTNF一αの産生や細胞表面抗
原の発現では、P38 MAPKよりもERKの方が優位に作用する可能性が
示唆された。DNCBとNiso4は共にTNF一αの産生を誘導するが、その
産生機構に関しては、DNCBではP38 MAPKを介し、また、 NiSO4で
はERKを介して誘導された。さらに、前章の結果より、産生された
TNFαはオートクラインに細胞に作用して、 CD54及びCD40の発現増
強に寄与した。また、:Niso4の場合、 cD86の発現の作用機i構1はp38
MAPK及びERKを介するが、 CD54及びCD40の発現はERKのみを
介した作用機構であった。このように、感作性物質によるTNF一αの産
生と細胞表面抗原の発現増強はそれぞれ、異なる細胞内シグナル伝達
系を介して起こり、また、化学物質によって活性化の経路が異なるこ
とが示唆された。また、感作性物質による樹状細胞活性化の作用機構
との同等性が高いことから、THP4細胞が樹状細胞の感作性物質応答
を再現する有用な細胞モデルであることが示唆された。
70

ERK p38
2く! x
TNF-ct
CD86@CD40/
NiSO4
Fig.18 THP-1 cell activation by DNCB and NiSO4 is regulated
through MAPK signaling pathways and autoc血e ef驚ct ofTNFα.
71

第3編 感作性物質によるIL・・8産生の特異性と発現
機構の解明69)
第1編において、THP-1細胞が、 EUを除く感作性物質7物質でIL-8
を顕著に産生することを示した。IL-8は、炎症部位への好中球やTリ
ンパ球の遊走に関与するケモカインとして知られる41)。また、ツタウ
ルシに感作されているヒトの皮膚に、ツタウルシ抽出液を適用すると、
皮膚中のIL-8の産生増強が認められている70)。また、樹状細胞を用
いたiH・vitro実験では、いくつかの代表的な感作性物質により、 IL8の
産生が報告されている71)。しかしながら、樹状細胞が様々な感作性物
質に対し、特異的に応答してIL-8を産生するかは十分に検討されてい
ない。また、その産生機構についても不明である。
そこで、本研究では、樹状細胞様の作用機購を介して感作性物質に
応答することが明らかとなったTHP4細胞を用いて、感作性強度が異
なる種々の感作性物質、及び非感作性物質からなる、23物質に関して、
IL-8の産生量を測定した。それにより、樹状細胞から産生されるIL 8
の感作性物質特異性を調べた。さらに、代表的な感作性物質である
DNCB及びNiso4によるIL8産生における、細胞内シグナル伝達系及
び、TNF-ocの影響を調べることで、 IL-8の産生機構について検討した。
72

第1節 実験の部
L 細胞と培養液
細胞、培養液ともに第1編第1節と同じものを用いた。
2.被験物質とその調製方法
感作性物質としては、以下の16物質を用いた。1-benzoylacetone
(1-BA),5-chloro-2-methy1-4-iso仕hiazolin-3-one/2-methyl-4-isothiazolin-3-
one (MCIAV{1),DNCB, glutaraldehyde (GA),phthalic anhydnde (PA),
benzoyl peroxide (BA),ethylenediamine (ED),2-mercaptobenzothiasole
(2-IVII3T),NiSO4, diethyl maleate (DEM),methyl-2-nonynoate (MN),
hexylcinnamaldehyde (HCA),citral (CI),eugenol (EU),aniline (AN)
a簸dgera㎡ol(GE)。非感作性物質としては、以下の7物質を用いた。
1一一bromobutEme (1-BB),diethyl phthalate (DEP),benzalkonium chloride
(BKC) , vanillin (VA) ,2-hydroxypropyl methacrylate (2-HPM) ,
acetoanisole(AA)and 6-metlryicoumartn(6-MC)。これらの物質は、
これまでにマウスを用いた皮膚感作性試験での評価が行われており、
感作性の有無及びその強度が調べられている(TabIe・5)72・73・74)。また、
2-HPMを除く、全ての化学物質はSigma-Aldnchより購入した。2-HPM
はWako・Pure・Chemicals(Osaka, Japan)より購入した。水溶性の化学物
質(MCI/MI, GA, ED、:NiSO4、 BKC)は生理食塩水に溶解させ、水不
溶性の化学物質(1-BA, DNCB, PA, BA,2-MBT;DEM, MN, HCA, CI, EU,
AN, GE,1一・BB, DEP, VA,2-HPM, AA、6-MC)はDMSOに溶解させた後、
培養液に添加した。
73

Table 5 [lhe list of chemicals tested and the value of CV75.
Cheinicals AbbreviatioRLLNAPotency
categerv*
CV75(ptg/ML) *“
1-Benzoylacetone
5-Chloro一一2一一methy14-
isothiazolin-3-one Solution
2,4-Dinitrochlorobenzene
Gl厩alaldehyde
Benzoyl peroxide
Phthalic anhydride
2-Mercaptobenzothiasole
Ethylene dian:血e
Methyl-2-nonynoate
Nickel suifate
Die血yl malea重e
Hexylcinndinaidehyde
Cimal
Eugenol
Ge1緬01
Aniline
1-BA
MCIAN41
DNCB GA BPPA
2-PvBT
ED
MNNiSO4
DEMHCA
CI
EU GEsu
Nlergems (16)
Extreme
Extreme
Extreme
Strong
Strong
StroAg
Moderate
Moderate
Moderate
Modera重e
Moderate
Weak
Weak
Weak
Weak
Weak
82
140
5 11
42
ND***
150
273
23e
150
i20
37
20
150
167
930
1-Broiinobutane
Diethyl phthalate
Benzalkonimn chlonde
varrilko
2-hydroxypropyl methacrylate
Acetoanisole
6-rr}ethvlcomnark}
1一一BB
DEP
BKCVA
2-HPM
触6-MC
NoR-
allergens
(7)
Non-
sensltrzer
500
600
3 650
ND***
320
277
* The potency of skin sensitization was estimated using the animal test
(Gerberick et al.,1992,2005;KiMber el al., i 998). All chemicals were
ranked from extreme to noR-sensitizer.
““ CV750f each chemical was estimated by fiow cytometry.
*** NDs not decided
74

3.被験物質の処理濃度の設定
各被験物質に対して細胞毒性試験を行い、24時間後に細胞生存率が
75%になると推定される濃度(CV75;cell viability 75%)を算出した
(Table 5)。そして、 CV75を基準に公比12で、5濃度(1.2・x,1x,1/1.2
x,1/1.22 x,1/123xCV75)を設定した。この濃度設定により、中程度の
細胞毒1生からほとんど無毒性の範囲となる、およそ50~95%の細胞生
存率の範囲でIL-8の産生量を測定した。被験物質の最高濃度は、溶媒
がDMSOならば1mg/mL、生理食塩水ならば5mg/m Lを越えない濃
度とした。さらに、MAPK及びTNFαの阻害実験では、5 P9/ML DNCB
及び5μ9/rr}L Nis 04を用いた。
4.THP-1細胞に対する化学物質の処理方法
THP-1細胞は24ウェルプレートにlx106cells/mL/ウェルにて播種し、
5μg/血LDNcB及び5μg/tnL Niso4を添加した後、37℃、5%co2条件
下で24時間培養した。p38 MAPKあるいはERKがIL-8の産生量に及
ぼす影響は第2編、第2章、第1節の3と同様の方法で調べた。また、
細胞外のTNF一αがIL-8の産生量に及ぼす影響は第2編、第1章、第1
節の2と同様の方法で調べた。
4.サイトカイン産生量の解析
第2編、第1章、第1節に示したとおりである。
5.統計学的解析
第2編、第1章、第1節に示したとおりである。
75

第2節 結果
L 感作性物質及び非感作性物質に対するIL-8の応答性
異なる感作性強度を有した感作性物質、及び非感作性物質、合わせ
て23物質をそれぞれ、THP-1細胞に処理しIL・一8の産生量を調べた(Fig.
19)。無処理あるいはDMSO処理により産生するIL-8は、それぞれ47
±・21.3pg/mL、40.8±22.O pg/血しであった。感作性物質16物質中、13
物質では濃度依存的、かつ有意なIL-8の産生増強が認められた。さら
に、このうち10物質では、無処理あるいはDMSOに対して、10倍以
上のIL8産生増強が認められた。特に、感作性強度がextr㎝e及び
sSt・ongの感作性物質では、低い処理濃度でも有意な産生が認められる
傾向があった。一方で、細胞生存率は被験物質の濃度依存的に低下し
た。IL-8の有意な産生増強は、細胞生存率が60~90%となる範囲で顕
著に認められた。感作性物質のうち3物質(PA、 ED及びGE)では、
IL-8の産生量の有意な変化が認められなかった。 EDとGEでは細胞生
存率の低下が起こるものの、PAでは細胞毒性が誘導されなかった。
非感作性物質においては、BKC、 VN、2-HPM、 AA及び6-MCで、
IL-8の有意な産生は認められなかった。これらの非感作性物質では、
多くの感作性物質と同様に、細胞生存率の低下が認められた。一方で、
1-BB及びDEPによりIL-8の産生量は顕著に増強した。特に、1-BB
では、多くの感作性物質と同様に、DMSOに対して10倍以上の産生
量が認められた。
76

a.
b.
へ 晒、N
・丁翻灘一,N
}磯。。.肖
丁照麟い.一
工灘耐.一
エMyCM
ワ怖*3
り
・醜
丁羅麟
…………--凱幽
0 0 0
0 0
0 《」
(口巳画O)
蓉咽ぢ毛ρ乙。。山圃
oO
ミNoQ
。◎
nトい
ト噂幽い
ABレ
ド…酢幽灘
㎜ 鋤 、
@
@(減套・。儀)
琴鷺ぞ。乙。。ぬ圃
み き 3讐。。35㌦。
へ露幽
趨・臨麟
i, “”‘sx
面面鵬磯醜....
欝篇鐸課穿訟駅篶篶論欝9 DNCB GA 47.,99,02fy]フ5砿
凵H・・**脚寧.一...__….
PA BP ’f”
讐肝
幽登M
幽叢込
幽卜。。
戯・。黛
繋側ED
醜Oひ一
曲。。鴇
ヒヨユヰ
・K@5・1喫1。。
80 §
60葛 秀 40窒
2G
O
c.
謝叫㈱醐脚糊0寸貰O
磐
OO糊
。腎
B◎
ひゆ淘損い
MED聖霊麟oolo
(減養田3
篇O鴇Q灘℃O』鳥oOl績一
、臨繭
癩
.薩
壷轟露露O酬瓢
Oま N
いトトA
O寸O
ooエ頃
Oσσ一
〇コ
リN蝋
瞥O[
卜。◎
UE
臨 、繭灘
、繍醗
騰・工麟幽、
麟翻
O.
oN
O.
nN
卜.
Qひ.O,寸.
。噂
O.
m笛
oo、
n翰
卜.
シ61
繍躍寸.漏
一
OoD一
〇2
い製
燈白
卜。◎
エCACH イOBN
・繭翻
鹸
薩
O卜N
O【うN
Nひ蝋0
2
斡葭
襟 Ii
(鉢9〆mL》
∴鮭
ON卜
OOO
OOい
ト守
卜寸翰
?BD
翻
OOゆ
OOい
ト守
卜燈の
ひoo創
BB㌔
壌ξ_iiO
-ii lo,
..一、藍麺麟。
躇銭§
GE
匪00
“X “N “tw “& “
.、醜盤幽
い、騨贈woo㌧◎
日NN出6
BKC
壷_ 四脚盤..醜継寸σ◎の
ON的
卜ON
N創d
賂◎○糊
触
OO9
翰笛◎o
寸ひゆ
ひトい
ペウσ噂
MPH芸
ゆσ腎ひ
O◎σ卜
O$
囚寸頃
旧い博
脳
08
06
04
02
0
、』弓
懸轟々熱d餓
卜卜N
嵩N
Nひ桝0
2 CM6.
(零)あ㍊[蝋O邸喘〉
Fig. 19 The alteration of IL-8 production in THP-1 cells exposed to 16
contact allergens (a, b) aiid 7 non-allergens (c). Each data poir}t
represents the mean ± S.D. for tlMree independent experiments. Bars
indicate extent of IL-8 production and circles provide cell viability
StatiStiCal Significance WaS CalCUIated by StUdentラS t-teSt (*」)く0.05,
**p〈e.ol) .
(醜)・ハ農巖』邸哨〉
77

2.感作性物質によるIL-8の産生における細胞内シグナル伝達系の影
響
DNcB及び:Niso4によるIL-8の産生における細胞内シグナル伝達系
の影響を調べるために、ERK及びp38 MAPKに対する阻害剤の影響を
調べた(Fig 20)。5 pgtML D:NcB及び170 pg/血L Niso4を24時間処理
すると細胞生存率はそれぞれ84%、74%となったが、ERK阻害剤であ
るPD98059、 p38 MAPK阻害剤であるSB203580を共存させても、細
胞生存率への影響はなかった。さらに、DNcB及び:Niso4によるIL-8
の産生量は、PD98059濃度依存的に、部分的に抑制された。一方、
SB203580を処理すると、DNCBによるIL-8産生量の抑制が認められ、
o.3 pM sB203580では有意に抑制された。しかしながら、 Niso4による
rし8の産生量は抑制されなかった。
78
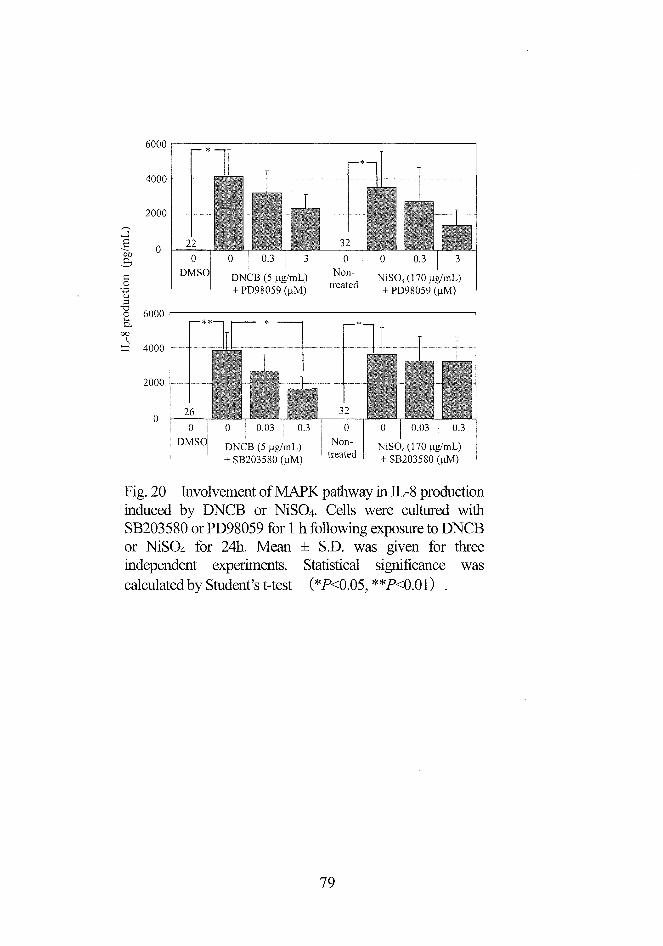
(日§\b◎瓢)8唇ぞ。a。。山圃
6000
4000
2eoo
o22
6000
1*
oD)gfso
*
32
O 1 O.3 1 3
DNCB (5 pg/mL)
+ PD98059 (#M)
oNon-
treated
O l O.3 1 3
NiSO,(170 pgtmL)
+ PD98059 (#M)
4000
2eoo
o
ユ「]一
26
o
DMSOo O.03
tttttt-tttt
撃狽獅狽窒狽狽狽狽狽狽
]
32
O.3
DNCB (5 pg/mL)
率SB203580(ptM)
o
Non-
trea{ed
o o.e3 03
NiSO, (170 pg/mL)
+ SB203580 (ptM)
Fig.20 ㎞volv㎝ent of MAPK pathway in皿.一8 productio慧
induced by DNCB or NiSO4. Celis were cultui’ed with
SB203580 or PD98059 for 1 h following exposure to DNCB
or NiSO4 for 24h. Mean ± S.D. was given for three
independent experiments. Statistical significance wascalcula:ted by Student’s t-test (*P・く0.05,**1)<0.01) .
79

3。感作性物質によるIL-8の産生におけるTNF一αの影響
TNF・一〇cがrL-8の産生に影響するかを調べるために、 THP一・1細胞に対
して5濃度のrhTNF一αを24時間適用し、 IL-8の産生量増加に及ぼす
rhTNFocの影響を調べた(Fig.22)。その結果、 fhTNF一αは濃度依存的
にIL-8の産生量を増加した。
続いて、5 ptg/血L DNcB及び170 ptg/mL Niso4により産生するTNF一一〇c
がIL-8の産生に影響するかを調べた。その結果、抗TNFα抗体により、
DNCB及び:Niso4により産生するTNFαを申和することによって、
1し8の産生量は抑制された。
80

*
200
0 0 0
く」 0 《4
1 1
(日容bo自)
8刷ぢぞ。a。。山圃
OOいN
いNO
6ら.ゆ鴇
門ひ6へ
◎○.ひ
0
0
D加温羽州齢⑳諭瞭榔C
Fig. 21 Tlie effect of it-8 on the production of
rhTNF-ct is presented. [flie culture supematants were
collected afld extent of IL-8 prodnction was detetmed.
Mean ± S.D. was given for three independent
expe血nents. Statistical sig掘ficance was calculated by
Student’s t-test (“P〈O.05) .
(譲餐b。鳥)8唇ぞ。a。。山}
6000
4000
2000
o
E.....r..=..
33
60000
DMSO DNCB (5 ptg/mL)
40000
20000
o
ご「.
35
Non-treate N至SO4(170μg/m】し)
mo aRti-TNF-or mAb (一)
va anti-TNF-or mAb(+)
Fig.22 Neutralization effect ofTNF-ct activity on llL-8
production indnced by DNCB and NiSO4. Cells were pre-treated
with anti-TNF-ct amibody for 1 h followed by exposui‘e to DNCB
or NiSO4 for 24 h. Mean土S.D. was given f()r thi●ee independent
expeiiments for each graph. Statistical sigrtificance was
calculated by StUdent’s t-test (*Pく0.05,**」P<0.01) .
81

第3節考察
これまでに樹状細胞だけでなく、THP-1細胞においても、代表的な
感作性物質によって、IL-8の産生増強が起こることが示されている
18・71)。しかしながら、樹状細胞が様々な感作性物質に対して、特異的
にIL-8を産生するかは十分に検討されていない。また、その産生機船
についても不明である。
そこで、本章でば樹状細胞様の作用機構を介して感作性物質に応
答することが明らかとなったTHP4細胞を用いて、感作性強度が異な
る種々の感作性物質16物質、及び非感作性物質7物質、合わせて23
物質に関して、rL-8の産生量を測定した。その結果、 PA、 ED及びGE
を除く感作性物質(13物質)で1し8の有意な産量の増加が認められ
た。また、このうち10物質では、溶媒対照に比べて10倍以上の産生
増加が認められた。このことから、THP-1細胞におけるIL8の応答性
は感作性物質に特異的な応答である可能性が示唆された。Tbebakらは、
DNCB、パラフェニレンジアミン、 Niso4を含む金属塩等の代表的な
感作性物質によって、樹状細胞からIL-8が産生誘導することを報告し
ている71)。また、THP-1細胞と同様にヒト細胞株で樹状細胞と類似の
感作性物質応答を有することが知られるU937細胞でも、感作性物質
13物質、非感作性物質5物質を適用すると、細胞生存率が85~90%の
範囲で、感作性物質に優位にrL-8 mR:NAの発現レベルの増強が認めら
れている75)。以上より、THP,.1細胞で認められる、 IL 8の感作性物質
に特異的な応答は、樹状細胞における応答を反映している可能性が示
唆された。
本研究で検討した感作性物質のうち、3つの感作性物質ではrし8の
82

産生が認められなかった。PAに関しては、最高処理濃度においても細
胞生存率の低下が認められなった。無処理での細胞生存率が95土2%
(n・・18)なのに対し、PAの最高処理濃度では95士3% (n・・3)であっ
た。PAの生理食塩水あるいはDMSOに対する溶解性は、本研究で検
討した他の化学物質に比べて低かった。そのため、PAの溶解性が低く、
細胞に十分に曝露されていないために、IIL-8の産生が誘導されなかっ
た可能性が示唆された。GEが属するモノテルペン類は、空気に曝さ
れると自然酸化し、感作性を有した酸化物が生成する場合があること
が知られている。GEに関しても、自然酸化による生成物に感作性が
あることが報告されている76)。よって、本実験条件下では、GEの酸
化物が十分に生成されないために、1レ8の産生が誘導されない可能性
が考えられた。EDもまた、モノアミンオキシダーゼによって、感作
性を有したジアルデヒドのglyoxa1に代謝されることが報告されてい
る77)。よって、THP-1細胞を用いた本実験系では、 EDの代謝が適切
に誘導されていないために、IL8の産生が認められない可能性が考え
られた。しかしながら、いずれの場合も、仮説を検証のための更なる
検討が必要であろう。
一方、非感作性物質のうち5物質では、IL-8の産生は認められなか
った。しかしながら、LBB及びDEPの2物質では、 IL-8の産生増強
が認められた。どちらの物質も、マウスを用いた皮膚感作性試験では
感作性はない物質として分類されている73)。しかしながら、著者らの
検討では、モルモットを用いた皮膚感作性試験法で1一・BBが陽性とな
る結果を得ている(データは示さず)。加えて、ステインやリジンを含
む合成ペプチドを用いて、化学物質とタンパクの結合性を評価する、
ペプチド反応性試験において、1BBは感作性陽性と判定されている
83

78)。また、DEPにおいては、強度は弱いものの、感作性陽性であるこ
とがヒトデータから結論付けられている79)。これら知見から考えて、
1・一BB及びDEPはいずれも感作性を有するために、 THP-1細胞におい
てIL-8の産生を増強させる可能性が考えられる。よって、1-BB及び
DEPで認められたIL-8の産生は偽陽性ではなく、化学物質本来の感作
性を示しているのかもしれない。
感作性の発症において、細胞表面抗原の発現やサイトカインの産生
を伴う樹状細胞の活性化は重要である。本章では、この過程における
一つの反応として、IL-8の産生が増強することが示唆された。続いて、
代表的な感作性物質であるDNcB及びNiso4によるIL-8産生におけ
る、細胞内シグナル伝達系及び、TNF一αの影響を調べることで、 IL-8
の産生機i構について検討した。その結果、ERK及びp38MAPKに対す
る阻害剤を用いた検討より、DNCBによるIL-8産生にはERK及びp38
MAPKが部分的に関与した。また、 Niso4によるIL-8産生にはp38
MAPKは寄与せず、 ERKのみが強く関与した。 Mo-DCでは、 DNCB
によるIL8の産生増強にp38 MAPKが寄与することが報告されている
18)。一方で、ERKがNiC12によるIL-8 mR:NAの発現に関与しているこ
とが明らかとなっている18)。このことは、THP-1細胞及び樹状細胞の
いずれにおいても、感作性物質によるIL-8の産生増強がp38 MAPK及
びERKを介して起こることを示唆している。さらに、DNCB及び:NisO4
により産生するTNFαが、 IL-8の産生に部分的に関与していることが
明らかとなった。すなわち、感作性物質によるrL-8の産生は、 CD54
及びCD40の発現増強と同様に、細胞内シグナル伝達系やTNF一一〇cのオ
ートクライン作用によって制御されていることが示唆された。
84

第4:節 小活
本章では、、樹状細胞様の作用機構を介して感作性物質に応答する
THP-1細胞を用いて、 IL-8産生の感作性物質特異性とその作用機構に
ついて検討した。その結果、IL-8の産生が感作性物質で優位に認めら
れることが明らかとなった。また、感作性物質によるIL-8の産生が、
細胞表面抗原の発現機i構と同様に、細胞内ではp38 MAPKとERKを
介し、細胞外ではllNFocを介して制御されることが明らかとなった。
THP-1による本結果は、樹状細胞におけるIL-8産生の作用機構の解明
に役立つものと期待される。さらに、樹状細胞の活性化及びその作用
機構を理解することによって、皮膚感作性の発症機構の解明に繋げて
いきたい。
85

結論
香粧品を開発する上で、皮膚に対する安全性の確認をすることは極
めて重要で、中でも皮膚感作性の評価を行うことは、安全な製品を開
発する上で欠かせない項目の1つである。従来、化学物質の皮膚感作
性評価は、モルモットなどの動物を用いた試験が行われてきた。しか
しながら、近年、動物愛護の観点から動物実験に対する批判が高まり、
EUにおいては化粧品指令第7次改正の施行により、2013年3月まで
に化粧品開発のための動物を用いた感作性試験が禁止される。こうし
た状況から、動物を用いない皮膚感作性試験に関する研究が世界的に
行われている。生体での感作反応は極めて複雑なため、その全てをin
vitroで再現することは困難であるが、感作性物質を皮膚に曝露した際
に生じる樹状細胞(ランゲルハンス細胞)の変化に着目した検討がい
くつか報告されている。その1つとして、ランゲルハンス細胞の代わ
りにヒト血液から調製された樹状細胞を使用した検討がなされている
が、安全なヒト血液の供給や用いる細胞の個人差の問題などから、未
だ実用に足る試験法は開発されていない。
著者らはそうした問題を克服することを目的として、血液由来細胞
の代わりにヒト細胞株であるTHP-1細胞を用いたin・vitro皮膚感作性
試験法(h-CLAr)の開発を進めている。 h-CLArでは、抗原提示の際
に補助刺激分子として機能する細胞表面抗原(CD86とCD54)の発現
変化を指標とすることで、一般化学物質64物質の感作性の有無を良好
に評価できることを確認した。一方で、試験法に用いるTHIP一・1細胞の
感作性物質応答やその作用機構は十分に解明されておらず、樹状細胞
と生物学的に同一の機序かを証明することが、試験法の実用化に向け
86

ての課題となっている。
そこで、本研究では、in・vitTO皮膚感作性試験法に用いる細胞として
THP-1細胞の有用性を高めるために、 THP-1細胞の感作性物質応答と
その作用機構について解析した。
1.THP-1細胞における細胞表面抗原及びサイトカインの感作性物質
応答
本研究では、THP-1細胞が感作性物質によって樹状細胞様の応答を
示すかを検討した。実験では、樹状細胞の活性化を特徴付ける、細胞
表面抗原(CD86/CD54/CD40/CD83/CD58/CD l a)とサイトカイン
(TNF一(∀IL-8/IL-6)の発現挙動を検討した。
被験物質は、代表的な感作性物質であるDNCB、 NisO4、と非感作
性物質であるSLSを用い、それぞれ細胞生存率が70~95%となる3濃
度で細胞に24時間処理した。その結果、DNCB及びNiso4では溶媒
のDMSO及び無処理群それぞれに対してCD86/CD54/CD40/CD83の発
現が有意に増強し、TNF一α/IL-8の産生も濃度依存的に増強した。加え
て、DNCBではCD l aの発現、 NisO4ではH!6の産生が増強した。一
方、SLSではいずれのマーカーも変化を示さなかった。陽性対照とし
て用いたLPSでは、 CDlaを除く全てのマーカーの発現が強く誘導さ
れた。
続いて、DNCBとNiso4に対して顕著な応答を示した
CD 86/CD54/CD40/CD83とTNF一ωIL-8に関して、感作性物質6物質
(hydroquinonc (HQ).propyl gallate (PG),foimaldehyde (FA).eugenol
(EU)、 phenylacetaldehyde (PAA)、 hydroxycitronellal(HC))と非感
作性物質2物質(methy1 paraben (MP)、1actic acid (LA))の影響を
87

調べ、THP-1細胞が感作性物質に特異的な応答を示すかについて検討
した。その結果、EUを除く、評価した全ての感作性物質で、6つのマ
ーカーのうち少なくとも5っで有意な増強が認められた。EUでは
CD 86、 CD54、 CD40の発現が有意に増強した。一方で、非感作性物
質では顕著な変化は認められなかった。また、感作性物質による活性
化マーカーの発現増強は細胞毒性と関係があるのに対し、同様に細胞
毒性が認められる非感作性物質では発現変動が認められなかった。さ
らに、樹状細胞でも、10~15%の細胞毒性が認められる濃度で感作性
物質を処理した際にCD86の発現増強が報告されている。このことか
ら、感作性物質による細胞の活性化は若干の細胞毒性を起こす濃度で
誘導される傾向があり、活性化マーカーの発現と細胞毒性との間には
感作性物質に特異的な関係があることが示唆された。
以上より、THP・一1細胞が皮膚感作性物質に対して樹状細胞の活性化
を反映する様々な応答を示し、それらを指標にすることで、感作性物
質と非感作性物質を判別できることが示された。それゆえ、THP-1細
胞は入手が容易で、均一な細胞集団としての特徴から研究に汎用でき
ることに加え、樹状細胞の代替として化学物質の感作性の有無を評価
可能な細胞モデルであることが示唆された。
2.感作性物質によるTHIP一・1細胞活性化の作用機構の解明
2-1.THP-1細胞活性化に及ぼすTNF一αと細胞外Arpの役割について
これまでの研究から、感作性物質を処理したTHP-1細胞において、
樹状細胞の活性化の際に認められる、様々な細胞表面抗原やサイトカ
インの発現が充進ずることが示された。一方面、THP-1細胞活性化の
作用機構に関してはよく分かっていない。そこで、炎症や免疫反応の
88

制御において重要な役割を果たすTNFα及びArpがTHP-1細胞活性
化の作用機構に及ぼす影響に着目し、感作性物質による細胞表面抗原
の発現追尋に及ぼす、TNF一αと細胞外Arpの影響を検討した。
まず初めにDNcB、 Niso4、 sLsをTHI’・一1細胞に処理し、経時的に
細胞生存率、TNF-ct、 IL-8、 CD86、 CD54、 CD40の発現挙動を解析し
た。無処理細胞の生存率は、処理後24時間はほぼ変化せず、98%であ
った。一方、DNCB及び:Niso4を高濃度処理した細胞の生存率の減少
は12~24時間の間に顕著に起こり、24時間後には共に82%程度であ
った。また、DNcBとNiso4では処理開始3時間後にTNF-ocとIL-8
の有意な産生増強が認められ、その後遅れてCD54、 CD86、 CD40の
発現増強が徐々に誘導された。一方で、SLSではいずれの時間におい
ても活性化マーカーの発現に顕著な変化は認められなかった。DNCB
やNiso4処理によるTNFαやrL-8の顕著な産生は、細胞生存率が低下
する24時間後だけでなく、細胞毒性がほとんど認められない処理時間
(3時間後~)でも認められた。これにより、感作性物質を24時間処
理した後のサイトカイン産生と細胞毒性は、それぞれ独立した細胞応
答として誘導されている可能性が示唆された。
続いて、TNF一一(x及びIL-8がTHI)一1細胞活性化に関与するかを調べる
ために、CD86、 CD54、 CD40に対するリコンビナントh㎜鋤TNFα
(d∬NFα)及びリコンビナントhuman M-8(rhlL-8)の影響を調べた。
その結果、CD86では変化は認められないが、 CD54、 CD40ではrhTNFα
の濃度依存的に発現が有意に充溢した。一方、rlalL-8はいずれの活性
化マーカーにも影響しなかった。そこで、感作性物質によりTHP-1細
胞から早期に産生されるTNF一αの影響を調べるために、 DNCB及び
Niso4処理1時間前に抗TNF一α抗体を添加した。その結果、抗TNFα
89

抗体の処理によりDNCB、:NisO4共にTNF一αの産生量は有意に抑制さ
れた。また、CD54及びCD40の発現充進が有意に抑制された。この
ことから、感作性物質によるCD54及びCD40の発現増強には、細胞
から早期に産生されるTNFαがオートクラインに強く寄与している
ことが示唆された。
さらに、cD86とcD54に対する細胞外ArPの影響を調べるために、
THP-1細胞にArPを24時間処理すると、 ArP濃度依存的にcD86、
CD54の発現が増強した。そこで、感作性物質によるCD86及びCD54
の発現に及ぼす細胞外Arpの影響を調べるために、 P2レセプターア
ンタゴニストであるsuraminを1時間処理した後に、 DNCB、:NiSO4、
EUを適用した。その結果、 NiSO4によるCD86及びCD54の発現増強
が有意に抑制された。DNCB、 EUによるCD86の発現増強において
もsurar血による抑制傾向が認められた。このことから、細胞外Arp
が感作性物質による細胞活性化に関与する可能性が示唆された。
以上より、感作性物質によるTHP・一1細胞の活性化は、 TNF一αのオー
トクライン作用及び、細胞外ATPによって部分的に制御され、樹状細
胞における細胞活性化と同様の作用機構を介して誘導されることが示
唆された。
2-2.THP-1細胞活性化に及ぼすシグナル伝達系の役割
近年、感作性物質による樹状細胞の活性化に関して、細胞内シグナ
ル伝達機構の研究がいくつか報告されている。しかしながら、DNCB
やNiso4を処理した際のサイトカイン産生やcD54及びcD40の増強
におけるシグナル伝達系の役割に関しては十分に明らかになってない。
また、樹状細胞の活性化に関して、P38 MAPKに比べ、 ERKの役割は
90

ほとんど明らかになっていない。それゆえ、本研究ではTHP-1細胞に
おいて、DNCBとNiso4によるTNF一α産生や種々の細胞表面抗原の発
現充進に関して、p38 MAPKとERKの役割について検討した。
その結果、溶媒処理群に比べて、DNcB及びNiso4によるp38 MAPK、
ERKのリン酸化が強く誘導された。さらにDNCB及びNiso4を処理
する1時間前に、p38 MAPK及びERK pathwayのリン酸化阻害剤を添
加することで、リン酸化活性の増強が抑制された。そこで、TNF一・ocへ
の影響を調べると、DNCBにより産生するTNFαがP38 MAPK阻害剤
(SB203580)でのみ有意に抑制され、一方でNiSO4により産生する
TNFαはERK阻害剤(PD98059)でのみ有意に抑制された。すなわち、
DNcB及び:Niso4の処理により産生するTNFαは、それぞれ全く異な
るシグナル伝達経路を介して誘導されることが示唆された。さらに、
表面抗原の発現増強に及ぼすp38 MAPKとERKの影響をそれぞれの
阻害剤を用いて検討した。その結果、DNcB及びNiso4によるcD86
の発現増強は共に、SB203580及びPD98059で有意に抑制されること
から、樹状細胞と同様にp38 MAPKとERKを介した作用機構により
誘導されることが示唆された。さらに、SB203580はD:NCBによるCD54
及びCD40の発現増強を一部抑制するが、 NiSO4によるCD54及び
CD40の発現には影響しなかった。また、 PD98059はDNCB及びNiSO4
によるCD54とCD40の発現増強を一部抑制した。以上より、 DNCB
とNiso4は共にp38 MAPK、 ERKをリン酸化するが、 TNFα及び細胞
表面抗原の発現に及ぼすp38 MAPK及びERKの役割は大きく異なる
ことが分かった。またこれは、感作性物質による樹状細胞活性化の作
用機構とも同等性が高いことから、THP-1細胞が樹状細胞の感作性物
質応答を再現する有用な細胞モデルであることが示唆された。
91

3.感作性物質によるIL-8産生の特異性と発現機構の解明
第1章において、THP-1細胞が感作性物質によりIL8を顕著に産生
することを示した。また、樹状細胞では加痂π》実験において感作性物
質によりIL 8の産生が報告されている。 IL-8は、炎症部位への好中球
やTリンパ球の遊走に関与するケモカインとして知られる。しかしな
がら、樹状細胞から産生されるIL-8の感作性物質特異性や発現機構に
ついては十分に解明されていない。そこで本研究では、THP-1細胞を
樹状細胞様の細胞モデルとして用い、23物質についてIL-8の産生量
を評価した。その結果、感作性物質に関しては、3物質を除く13物質
でIL-8の有意な産生が認められた。非感作性物質に関しては、2物質
で有意な産生が認められるものの、5物質でIL8の顕著な産生は示さ
なかった。このことから、IL-8は感作性物質で優位に産生されること
が示唆された。
続いて、IL-8産生の作用機構を調べた結果、 P38 MAPK及びERKに
対する阻害剤を用いた検討より、DNCBによるIL8の産生にはP38
MAPK及びERKが部分的に関与するのに対し、 NiSO4によるIL・一8の
産生にはp38 MAPKは寄与せず、 ERKのみが強く関与することが示唆
された。また、DNcBとNiso4共に、産生したTNF一αがIL-8産生に
部分的に関与することが示唆された。以上のように、感作性物質によ
るIL-8産生は、細胞表面抗原の発現機i構と同様に、細胞内ではp38
MAPKとERKを介し、細胞外ではTNF一αを介して制御されることが
示唆された。THP-1による本結果は、樹状細胞におけるIL-8産生の作
用機構の解明に役立つものと期待される。
本研究では、THP-1細胞の感作性物質応答とその作用機構について検
92

討した結果、TH])・・1細胞が樹状細胞で認められる種々の細胞表面抗原
やサイトカインに関して感作性物質特異的な応答を示し、樹状細胞の
代替として有用なことが示唆された。また、種々の細胞表面抗原やサ
イトカインの発現が、樹状細胞と同様の作用機構を介して起こること
が示唆された。さらに、THP-1細胞を用いた検討から、感作性物質に
より樹状細胞から産生されるH!8の産生機構解明に繋がる知見を得
た。以上のことから、THP-1細胞を用いることで、化学物質の皮膚感
作性の有無を良好に判別できるだけでなく、樹状細胞活性化の作用機
構を解明できる可能性が示唆された。本研究が感作性物質による樹状
細胞の活性化を具現化する加嘘π)皮膚感作性試験法(h-CLAT)の開
発・確立に向けて、試験法の生物学的機序についての根拠を示す一助
となることを期待したい。
皮膚感作の成立には、樹状細胞の活性化だけでなく、化学物質の経
皮吸収性やタンパク結合性、及び抗原特異的なT細胞の増殖など、い
くつかの鍵となる反応が関与する。すなわち、皮膚感作性を評価する
のに樹状細胞の活性化を再現したin vitro試験法は有用な方法である
が、それだけでは完全ではないかもしれない。今後さらに、いくつか
の反応を具現化して、h-CLATと他の試験法を組み合わせた評価が、
ヒトにおける皮膚感作性の予測の向上に繋がるものと考えられる。
93

謝辞
本研究を遂行し、まとめるにあたって、実に多くの方にお世話にな
りました。この場を借りて、感謝の意を述べさせて頂きたいと思いま
す。
本論文の審査に際して貴重な御指導を賜り、御審査頂きました城西
大学薬浴品動態制御学講座教授 杉林 堅次先生、日比野 康英先生、
小林 順先生に衷心より深甚なる謝意を表します。
本研究の遂行にあたり終始有益な御指導、御助言を賜りました花王
㈱安全性評価研究所坂[コ斉副主席研究員に心から感謝を申し上
げます。投稿論文の執筆や数多くの学会発表、日頃の研究内容に渡っ
て、手厚いご指導を頂きました。博士(薬学)の学位論文を執筆する
ことができたのも、坂口さんのご指導ご助言のお陰に他なりません。
また、本研究の遂行にあたり、多大なご協力、ご支援を頂いた同研究
所 額田 祐子研究員に心から感謝の意を表します。本論文が完成し
たのも額田さんが一緒に研究を進め、多くの成果を見出して下さった
お陰です。深く感謝申し上げます。
また、Top Tox Consulting, Javier Avalos, Ph.D.には、本研究の主論文全
てに関して適切な御助言を賜りましたこと、厚くお礼申し上げます。
花ヨ㈱ 品質保証本部 品質保証センター 安全管理室 鈴木 尋
之室長、安全性評価研究所 西山 直宏室長には、日頃より教育およ
び研究における多くのご助言やご支援を頂きました。深く感謝いたし
ます。そして、同研究所 伊藤 勇一主任研究員、吉田 雪子研究員
には、研究の進め方や悩みについて親身になって相談に乗って頂き、
様々な面で見守って下さいました。深くお礼申し上げます。また、同
94

研究所 小坂 七重研究員には、有益なご意見ならびに実験のご協力
を頂きましたこと、感謝しております。実際の実験で非常に丁寧かつ
正確に補助をして下さった須藤 里実さん、夏梅 和子さんにも大変
感謝致します。さらに、安全性評価研究所の皆様に深く感謝致します。
最後に、論文をまとめるにあたり、これまで私を温かく応援してく
れた両親と祖母、そして、明るく励まし続け、時間を与えてくれた妻
智恵子と娘 佑奈に心から謝意を記します。
以上の皆様の助言・支援・協力・励ましに対し、深く感謝申し上げ
ます。
95

引用文献
1) Magnusson, B., Kligrnan, A.M. (1969): Tlie idemification of contact
allergens by animal assay. The guinea pig maximizatioA test. J. invest.
Dermatol.. 52. 268-276. ) 一.r一一d)
2) Kimber, 1., Mitchell, J.A., Grifiin, A.C. (1986): Development of a murine
local lymph node assay fbr the deter曲a怠on of sensitセing Poten廿al.
Food Chem. Toxicol.. 24. 585-586. ) nv 1)
3) Directive 2003/15/EC of tihe european parliamept and of the council,
Februairy 27, (2003): Official J. European Union, L66, POO26-0035 B.
4) OECD, (2004): Test guideline, 428.
5) OECD, (2004): Test guideline, 432.
6) OECD, (2004): Test guideline, 430.
7) OECD, (2004): Test guideline, 431.
8) Hart, D.N.J., (1997): Dendritic cells: urrique leukocyte population which
control血e pr㎞ary辻nmune response. Blood,90,3245-3287.
9) Banchereau, J., SteiumEm, R.M. (1998): Dendrkic cells aRd the control of
immunity. Nature, 392, 245-252.
10) Jowsery, 1.R., Basketter, D A, Westmoreland, C., KiMber, 1. (2006): A
fUtUre apProach to m.easu血g relative skin sensi廿sing Potency:aproposal.
J. Appl. Toxicol., 26, 341-350.
i1) Ozawa, H., Nakagawa, S., Tagami, H., Aiba, S. (1996): lnterleulcin-1 beta
and granulocyte macrophage colony-stimulating factor mediate
Langerhans cell maturation differently. J. invest. Dermatol., 106, 44 i-445.
12) Venier, A.C., Schmitt, D., Staquet, M.J. (1999): Fragrance and contact
96

allergens in vitro modulate the HLA-DR and E-cadherin expression on
human epidermal Langerhans cells. im. Arch. Allei’gy. lmmunol., 120,
5662.
13) Enk, A.H., Katz, S.1. (1992): Early molecular events in the induction phase
ofcontact sensitivity Proc. Nat. Acad. Sci. U.S.A., 89, 1398-1402.
14)C㎜berbatch, M, Deatrnan, R.J.,㎞ber,1.(1996):Co翻加亘ve and
inducible expression of interleuldii-6 by Langerhans cells and 1ymph node
dendritic cells. lmmunology, 87, 513-518.
15) Sallusto, F., Laiiizavecchio, A. (1994): Efficient presentation of soluble
antigeR by culuted human dendritic cells is maintained by
granulocyte/macrophage colony st㎞uIa血g factor plus imerleulcin-4 and
down regulated by mmor necrosis factor ct. J. Exp. Med., 179, 1109-1118.
16) Caux, C., Dezuttt¢r-Dambuyant, C., Schmitt, D., Banchereau, J. 1992):
GM-CSF and [[NF一 ct cooperate in the generation of dendritic Langerhans
cells. Nature. 360. 258-261. )vvvp
17) Aiba, S., Terurtuma, A., Manome, H., Tagami, H. (1997): Dendritic cells
differendy respoRded to haptens and irrttants by their production of
cytoknes and expression of co-stmulatory molecules. Eur. J. IMmunol.,
27, 3031-3038.
18) Aiba, S., Manome, H., Nakagawa, S., Mollah, Z.U., Mizutashi, M., Ohtani,
T., Yoshno, Y., Tagam, H. (2003): p38 mitogen-activated protein kinase
aiid extracellular signal-regulated kiRases play distinct roles in the
activation of defidritic cellsby twe representative haptens, NiC12 and
DNCB. J. invest. Dermatol., 120, 39e-398.
19) Coutant, KD., de Fraissinette, A.B., Cordier, A., Ulrich, P. (1999):
97

Modulation of the activity of hurnan monocyte-derived dendritic cells by
chemical haptens, a metai allergen, and a staphyloceccal superantigen.
Toxicol. Sci., 52, 189一・198.
20) De Smedt, A.C.A., Van Den Heuvel, R. L., Bememan, Z.N., Schoeters,
GE.R. (2001): Modulatien of phenotype, cytokne production and
stimulatory function of CD34+ derived DC by NiC12 and SDS. Toxicol.
in Vitro. 15. 319一一325. , ラ
21) De Smedt, A.C., Van Den Heuvel, R.L., VaA Tendeloo, V.E, Bememan,
ZN., S choeters, GE., Weber, E., Tuschl, H. (2002): Phenotypic
alterations and IL-1 beta prodnctioR in CD34+ progenitor一 and
monocyte-derived den(hitic cells aftei” exposure to allergens: a
comparative analysis. Ai”ch. Deirnatol. Res., 294,109-116.
22)Staquet, MJ., Sportouch, M., Jacquet, C., Sch【磁, D., Guesnet,工,
Peguet-Navarro, J. (2004): Modor-ate skiR sensitizers can induce
phenotypic changes on in vitro generated dendritic celis. Toxicol. IA Vitre,
18. 493-500. ’
23) Ryan, C A, Hulette, B.C., Gerberick, GE (2001): Approaches for the
developmeltt of cell-based ln vitro methods for contact sensitization.
Toxicol. ln vatro. 15. 43一一45. ) sv)
24) Tuschl, H., Kevac, R., Weber, E. (2000): The expression of sinface
markers on dendritic cells as indicators for the sensitizing potential of
chemicals. Toxicol. in Vnto. 14. 541-549. ) A. t)
25) Tsuchiya, S., Yamabe, M., Yamaguchi, Y., Kobayashi, Y., Kenno, T., Tada,
K. (1980): Establisiment and characteiization of a human acute
menocytic leukemia cell 1ine (TH]?一1). int. J. Cancer, 26, 171-176.
98

26) Ashikaga, T., Hoya, M., ltagaki, H., Katurnura, Y., Aiba, S. (2002):
Evaluation of CD86 expressien and MHC class ll molecule
internalization in THP-1 huinan monocyte cells as predictive endpoints
for contact sensitizers. Toxicolo. in Vitro. 16. 711一一716. ) LVe
27) Yoshida, Y., S akaguchi, H., lto, Y., Okuda, M., Suzulci, H. (2003):
Evalua廿on o紬e s㎞se粥itセation pote翻of chemicals us血g expression
ofco-stimulatory molecules, CD54 and CD86, on the naYve THP-1 cell
l辻le. Tbxicol。 in Vitro 17 221-228. ) A. 1)
28) Ashikaga, T., Yoshida, Y., Hirota, M., Yoneyama, K., ltagaki, H.,
Sakaguchi, H., Miyazaweg M., lto, Y., Si」rzulci, H., Toyod& H. (2006):
Development of an in vitro skm sensitization test using haman cell lines:
The human CelI L血e Activation Test(h-CLAr)1. Qpttmization of the
h一一CLAT protocol. Toxicol. in Vitro, 20, 767一・773.
29) Sakaguchi, H., Ashikaga, T., Miyazawa, M., Yoshida, Y., lto, Y.,
Yoneyama, K, Hirota M., ltagaki, H., Toyoda, H., Suzulci, H. (2006):
Development of aA in vitro skn sensitization test using human cell lines;
h㎜an CeIl L血e Activation Test (h一一CLAr)H. An虚一1aboratory study of
血eh-CLAr. Tbxico1. in Vltro 20767-773. ) “v)
30) Kosaka, N., Ashikaga, T., S akaguchi, H., Sono, S., ltagaki, H., Nishiyama,
N. (2007): Evaluation of the in vitro skn sensitizatioA test; human cell 1ine
activation test (h-CLAT) using the modified prediction model. A bstracts
・f♂~%献加解∬・副彪…atives &Animal Use in theLofe Scien㈱.
31) Miyazawa, M., lto, Y., Yoshida, Y, Sakaguchi, H., Suzulci, H. (2007):
Phen卿ic alterations and cytokne pro(沁。廿on血THP-1 cells㎞
response to allergens. Toxicol. in Vist’o, 21, 428-437.
32) Van Kooten, C., Banchereau, J. (1997): Functions of CD40 on B cells,
99

dendntic cells and other cells. Curr. Qpin. lmmunol., 9, 330-337.
33) Pinchuk, L.M., Klaus, S.J., Magaletti, D.M., Pinchuk, GV., Norsen, J.P.,
Clat4k, E.A. (1996): Functional CD40 ligand expressed by humaA blood
dendritic cells is up-regulated by CD40 ligation. J. immunol., 157,
43634370.
34) Peguet-Navano, J., Dalbiez-Gauthier, C., Rattis, F.M., Van Koote, C.,
Banchereau, J., Schmitt, D., (1995) : Functional expression of CD40
antigen on hirman epidermal Langethatts cells. J. lmmunol., 155,
42414247.
35) Basketter, D.A., Scholes, E.W., Kimber, 1. (1994): [lhe performance of the
local lymph node assay with chemicals idemified as contact allergens in
the human maximization test. Food Chem. Toxicol., 32, 543-547.
36) Zhou, L.J., Tedder, T.F. (1995): Human blood dendritic cells selectively
express CD83, a member of the immunoglobulin superfamily. J.
IMmimol., 154, 3821-3835.
37) De Smedt, A.C., Van Den Heuvel, RL., Van Tendeloo, V.F., BememaR,
Z.N., Schoeters, GE. (2005): Capacity of CD34+ progeniterdetwed
dendritic cells to distinguish between sensitizers and initants. Toxicol.
Lett.. 156. 377-389. ) 一.vv)
38) Wang, B., Amerio, P., S auder, D.N. (1999): Role of cytokines in epideimal
Langerhans cell migration. 」. Leukoc. Biol., 66, 33-39.
39) Salamero, J., Bausinger, H., Mommaas, A.M., Lipsker, D., Proamer, E,
Cazenave, J.P., Goud, B., de la Salle, H., HaAau, D. (2001): CD I a
molecules maffic through the early recycling endosomal pathway in
human Langerhans cells. J. IAvest. Deimatol., 116, 401-408.
40) Manome, H., Aiba, S. (1999): Simple chemicals can indvice maturation
100

and apoptosis ofdendritic cells. iminunology 98, 481490.
41) Sebastiani, S., Albanesi, C., De, P.O., Puddn, P., Cavani, A. and
Girolomoni, G (2002): The role of chemokines in allergic.contact
dermatitis. Arch. Dermatol. Res.. 293. 552-559. ラ ,
42)Ve血eyen, GR., Schoeters, E., N両it㎝, J.M, Van Den Heuvel, R.L.,
Nelissea 1.. Witters. H.. Van Den Heuvel. R.L.. Nelissen. 1.. Witters. H ) L-p TT-UVV-U) A一一) T 1.一k]L .Lt V#一 A.一V-TVJLp 一一一A“一) 一IVJLJLVVV一“L) .L一) 1 T iPVVkV) 一)
Van Tendeloo, V.F.1., Berneman, Z.N., Schoeters, GE.R. (2eO5): Cytokine
traAscript profiling in CD34+一progenitor derived dendritic cells exposed to
contact aliergens and imtmts. Toxicol. Lett., 155, 187-194.
43)S舳ero, J., Baus血ger, H., Mommaas, A.M, Lipskeゆ., Proameiz, F.,
Cazenave, J.P., Goud, B., de la Saile, H., Hanau, D. (2001): CD I a
molecules mafiic through the early recyclmg endesomal pathway in
human Langethatis cells. J. invest. Deirnatol., 116, 401-408.
44)Hope, J.C., Campbell, F., Hop㎞s, S J.(2000):Deficiency of皿.一20r IL-6
reduces 1ymphocyte proliferation, but only llL-6 deficiency decreases the
contact hypersensitivity response. Eur. J. hmunol., 30, 197-203.
45) Temissen, M.B., Rongen, H.A., Bos, J.D. (1994): Function of adhesion
molecules 1ytnphocyte imction-associated antigen-3 Emd intercellular
adhesion molecule-1 on human epideimal Langethans cells in
antigen-specific T cell activation. J. IM[nunol., 152, 3400-3409.
46) Mizuashi, M., Ohtani, T., Nakagawa, S., Aiba, S. (2005): Redox
iMbalance induced by contact sensitizers niggers the matt}iation of
dendritic cells. J. invest. Dermatol., 124, 579-586.
47) Hulette, B.C., Ryan, C.A., Gildea, L.A., Gerberick, F. (2005): Relationship
of CD 86 surface marker expression and cytotoxicity on dendritic cells
101

exposed to chemical allergen. Toxicol. Appl. Pharmacol., 209, 159-166.
48)Miyzawa, M., Ito, Y, Kosaka, N, NUkada, Y, SakagUchi, H., SuzUICi,旺,
Nishiyama, N. (in press): Role of TN}F-ct and extracellular ATP in THP-1
cell activation following allergen exposure.
49) Miyzawa, M., lto, Y., Kosaka, N., Nukada, Y., Sakaguchi, H., Suzuki, H.,
Nishiyama, N. (in press): Role of MAPK signaling pathway in the
activation of dendntic type cell liAe, THP-1, induced by DNCB and
NiSO4.
5e) Ade, N., Mdirtinozzi-Teissier, S., Pallardy, M., Reusset, F. (2006):
Activation of U937 cells by contact sensitizers: CD 86 expression is
independent ofApoptosis. J. lmmunotoxicol., 3, 189-197.
51) Python, E, Goebel, C., Aeby, P. (2007): Assessment ofthe U937 cell line
for the detection of contact allergens. Toxicol. Appl. Pharmacol., 220,
113-24.
52) Azam, P., Peiffer, J.L., Chamousset, D., Tissier, M.H., Bonnet, P.A., Vwn,
L., Fabre, 1., dwlin, J.C. (2006): The cytokne-dependent MUTZ-3 cell
line as an in vitro model for the screening of contact sensitizers. Toxicol.
Appl. Pharmacol., 212, 14-23.
53) Salgado, C.G, Nakamura, K., Sugaya, M., Ta(la, Y., Asahina, A., Fukuda,
S., Koyama, Y., hie, S., Tamaki, K (1999): Differemial effects of
cytol〈ines and imniunosuppressive drugs on CD40, B7-1, and B 7-2
expression on purified epidei mal Lange血ans cells1. J. Invest. Dermatol.,
113, 1021一一1027.
54) Di Vrrgilio, F., Ferrari, D., Chiozzi, P., Falzoni, S., Sanz, J.M., dal Susino,
M,Mu㎞i, C, Hanau, S., Badcordi, OR.(1996):Pu血oc(聯or f㎞ction
102

in the immune system. Drug. Dev. Res., 39, 319-329.
55) Di Vrrgilio, R, Chiozzi, P., Feirmi, D. (2001): Nucleotide receptors: Em
emergng famly of regulatory molecules in blood cells. Blood, 97,
587-600.
56) Abbmechio, M.P., BumstocK G (1994): Puuinocqptors: are there families of P2X
and MY purinoceptors? Pharmaco1. Ther, 64, op5475.
57) Fredholm, B.B., Abbracchio, M.P., BumstocK G (1994): Nomenclature
and classification ofpu曲oceptors. Phamlaco1. Rev.,46,143-56.
58) Ferrari, D., la S ala, A., Chiozzi, P., Morelli, A., Faizoni, S., Girolomoni, G,
Idzko, M., Dichmarm, S., Norgauer, J., Di Vlt’gilio, F. (2000): The P2
purinergic receptors of human dendritic cells: idemification and coupling
to cytokne release. FASEB, J., 14, 2466-2476.
59)La Sala, A., Ferrah, D., Co血ti, S., Cavalti, A., Di Virgilio, E, Girolomoni,
G (2001): Extracellular ATP iAduces a distorted maturation of dendntic
cells and inhibits their capacity to initiate Th l responses. J. lmmunol., 166,
1611-1617.
60) into, T., Fujiteg M., Okusawa, T., Hasebe, A., Morita, M., Shibata, K.
(2002): Synergic effects of mycoplasmaHipopeptides and extracellular
ATP on activation of macrephages. kifect. lmmun., 70, 3586-3591.
61) Boisleve, F., KerdirierR6mer, S., Rougier-lanats N., Pallatdy, M. (2004): Nickel
and DNCB induce CCR7 expression on human den(痂。 cells血ou帥
different signallmg pathways: role of ”1 NF-or enid IVkeV’K. J. invest.
Dermatol.. 123. 494-502. , ラ
62) Cumberbatch, M., KiMber, 1. (1995): Tumor necrosis factor-ct is required
for accumulation of dendritic cells in draming lymph nodes and for
103

optimal contact sensitization. lmmunol., 84, 31一一35.
63)Be曲ier一~Tergries,0., Bermond, F., FIacher, V:, Massac職C., Sc㎞i廿, D.,
P6gue毛一Navarro, J.(2005):TNF一αer血ances phenotyl)ic and fUnctional
ma加ration of human QPidermal Langerhans cells and induces皿!12P40
and IP-10/CXCL-10 prodnction. FEBS, Lett., 579, 3660-3668.
64) Flavell, R.A. (1999): [IJhe molecular basis of T cell differendatioR.
Inrmiunol. Res., 12, 159-168.
65) Nakahara, T., Moroi, Y, Uchi, H. and Fume, M. (2006): Differemial role
of MAPK signaling in human dendritic cell matwation and Th l/Th2
engagement. J. Dermatol. Sci., 42, 1-11.
66) Airighi, J.E, Rebsamelt, M., Rousset, F., Kindler, V. and Hauser, C.
(2001): A crkical role fer p38 mitogen-activated protein kinase in the
activation of human blood-derived dendritic cells induced by
lipopolysaccharide, TNF一一ct, and contact sensitizers. J. tmunol., 166,
3837-3845.
67) Dong, C., Davis, R.J., Flavell, R.A. (2002): MAP knases in the immune response.
舳u.Rev.㎞澱ユol.2055-72. ) “w)
68) lto, Y., Miyazawa, M., Yoshida, Y., Kosaka N., Sakaguchi, H., Suzulci, H.
(2005): Role of mitogen-activated protein knase in CD 86/CD54
expression induced by centact sensitizers on THP-1 cells. Proceedings of
the l gth annual mee血g of Japanese Society of Altemative to An㎞al
Expeiments.
69) Nukada, Y., Miyazawa, M., Kosaka, N., lto, Y., Sakaguchi, H., Nishiyama,
N。(sUbmitted):Production of皿!8 in THP-1 cells following t㎜or
necrosis factor一一ct production or contact allergen stmulation via
mitogen-activated protein knase pathway.
104

70) Griffiths, C.E., Barker, J.N., Kiml〈el, S. and Nickoloff, B.J. (1991):
Modulation of leucocyte adhesion molecules, a T-cell chemotaxin (IL-8)
and a regulato1y cytokne(TNFalpha)in ailergic contact dermatitis(血Is
dermatitis). Br. J. Dermatol., 124, 519-526.
71) Toebak, M.J., Pohlmann, P.R, Sampat-Sardj oepersad, S.C., von Blomberg,
B.M.E, Bruynzeel, D.P., Scheper, R.J., Rustemeyer, T. aRd Gibbs, S.
(20e6): CXCL8 Secretion by dendritic cells predicts contact allergens
丘om㎞itants. Toxico1. in Vttro 20117-124. 」 i-d v)
72) Gerberick, GF., House, RV., Fletcher, E.R. and Ryan, C.A. (1992):
Exarnination ofthe locaHymph node assay for use in contact sensitization
risk assessment. Fundam. Appl. Toxicol., 19, 438-445.
73) Gerberick, GF., Ryan, C.A., Kern, P. S., Schlatter, H., Deaman, R.J.,
KiAber, 1., Patlewicz, GY. and Basketter, D.A. (2005): Compilation of
Historical local lymph node data for evaluation of skn sensitization
alternative methods. Dermatitis., 16, 152-202.
74)Ki血ber,1., Mlton, J., Dea㎜an, R.J., Gerberick, GF, Ryan, C.A.,
Basketter, D.A., Lea, L., House, R.V., Ladics, GS., Loveless, S.E. and
Hastings, K.L. (1998): Assessment of the skn sensitization potemial of
topical medicaments using the local lymph node assay: an interlaboratory
evaluation. J. Toxicol. Environ. Health., S 3, 563-579.
75) Python, F., Goebei, C. and Aeby, P. (2007): Assessment of the U937 cell
line for the detection of contact allergens. Toxicol. Appl. Pharmacol., 220,
113-124.
76) Hagvall, L., Backtorp, C., Svenssen, S., Nyman, G, Bolje, A. and
Karlberg, A.T. (2007): Fregrance compound Geraniol forms contact
105

allergens air exposure. ldentification and quantification of oxidation
prodncts and effect on skm sensitzation. Chem. Res. Toxicoi., 20,
807-814.
77) Foussereau, J., Samsoen, M. and Hecht, M.T. (1983): Occupabonal
dermatitis to Ampholyt G in hospital personnel. Contact Deirmatitis., 9,
233-234.
78) Gerberick, GF., Vassal}o, J.D., Foertsch, LM., Price, B.B., Chaney, J.G
and Lepoittevin. J.P. (2007): Quantification of chemical peptide reactivity
for screening contact allergens; a classification tree medel approach.
Toxicol. Sci., 97, 417-427.
79) NIH Publication. (1999): The mmine local lymph node assay: A test
method for assessing the allergic contact deirnatitis potential of
chemicals/compounds. No. 99-4494.
106

d“タ
‘
‘ン
3’ゾ
、
、
」 】 -{ 一 ・一
、、
、ぐ
吟 、
ロ
, 、
べ
与つ ㌦
、「Jt:
へ
㌧ぜ
L
「
レ2
♪直、
、
、
「
♪’一
ぐ’ 卜
も
・ぐ》} 、
’ギー
“剣
し、
“響
, ,、 ’
b
■’
,
.’ ’ギ
㌧「
’,
P }
14
→
,ノ
」
,卵膚
㌦
長,’
一t↓
昌
、‘努
、で、
幽
り‘
s”‘
、し 一
「 ㌻
,瓢
1 1 」「一
’ち」 脅
戦
’「
」
閏
鴫 べ鴫
㌔ ’ ぴ
r 、
到
㌔’
κ
’
ケ
p’
F
辱詔
し
P ↓
、
♂
’「爵
7メ
’ げ
{ ρ
δ“
〔
}甲
し炉
ノや弛
、’
幽甲
角 」
f
’
旨 ’)
一J
i ’i tい
・,
?ヂ: ・卜 嘆
咤
“)ets J
し
1一
lt ’一 “tb.1
’げら
{
ミ亀
一l
t
l i-L
(
“一 “
i5f・ .’ ’;
.
’
筆「
~
t
s)r4
’
即
’
且〆 ’
1
一 li
1
1沈や一
しノ ㌦L
、 r
一軌● 伊
t一
“
ハヤ
ー
,
1.‘
1
t
1
1
‘ヤ 一
煽 n j/
i
T ;S
’物~
s tt
一
毫} L
1
ご
鷺5、
, 》
’
一
や㍉ 一夢
ρ 」 、
でr
馬㌔」
しあ、、ピーしへ・「
』『♂ 戸 」ドノ
へ物、メ ’
》
「
v tt rt”
el T r te ’ tl 戸’ ; “t
’㌃㌦
,
ko♪
二 旨西
1
げ 、㌧
だ
、、
〒
r4
ダ’
!
7、
,痴、凋
8
5
「
砂、
噌し叩
必
戸1
ウ ’
、 些 、
」.ρ召、’
て
{郭{と弊与
マ艦
@
@囲、↑コ
、、
把N’心避雷ら ノ ヤ 戸
∫. 1ハ
1㌘壕,㌦
t e
㌦♂弩ttf t D1”ttl 1’ Ls
海霧r・ ヒ戸r二〕9,Hl
,
lr.
i
Ltl’
t t L 叶L lf.” tL じ s;.
い㌻
窪p
辱戸学ρ
P、
」戸ト
4’
’
↓雪
敏パ
鴻㌧
、
ノ
もほヨ
でハ
ト
も
p 聖
9
5τ 星 r㌻戸
「 与
イ」
着、
@‘嵐∴
め’2 0
、暖磯塵・
’ 7吃’
を-ノ
sltr
Jイ
T
’帰
r
’n’
( v ’
it
ot
ノ 、] H
lsX,. r
、1駄既・・
ヤい㌔1
r 予 へ
、
エ
ぞ 、気
」
ユ
、宝、
噛
へ
’ SN}ド
5 r
リ ノ、
71㌧(
tvl 一’ L ’
r
ずlr
⊂
♪F紬
イ辱
ttt ll n.!”:s::ftli’
メ
」
’,
’ 渇
「 , ヴ
V、 ■
刃”
ρ㌧ ド!セ
.》奪㍉宝て
@
d『4婚弓’
ρ
) s 勺 ゴa「 t’2償
且話掌L‘!窄 ’イ ‘ ’、噂
’薄いi
’篁
ハ
~ 山㌧「、了覧 ㌧
, 語
二戸
、・≠ゾ、ー四五♪。い.
@
`轟畢虞
豹 論「
@
@芦 嘆三許
む娠な妻・妥,・.
㌧‘
毎
ρシぐ
へ漏アノ
’
、
鼻 ㍗
一 ぜ
’
ユ ’
㌧
r
サ’
A
〃傷・
ρ
’
、
@」競的
乳.一 罫
撃
∂、 ,
、メー【,
rデ
威 聾・いぐ、
寧 !
イし
㍗ f
〃 、 p
μ
’ ,’
「
5
グ“「炉徽
鵠曳ソ’ 冨守 ド
μ{聴、 ’
ヨ げ{ 、
7 己、
ρ
㌔ヤ争
ド}g
’ぎ
’
どtt用
,2跨~吉員「
「 再
阜
、 ≒
「㌧ト
藩’ ,
匂ぐ
、」
,
、
イど
蹄,“s”F w-e
ノノ
’初、ゲい
み五;
~。ノ譜ム
,・㌻誤4
’ + 、 「
評
辱~
く㌦,㌔
㌔
、 ρノ
詑.
【 5
隔
産‘
唱
電㌦・ネ
蒐
r
転 trLs
へ、
tTt
「
風、’
藻 (5
7{㊥
ヤ
♂
7 ’
、, ㌧ 「、
ト レ ろ
卸’ ♂
「’ 」7 ’㍗
ぜノ㌔・ え収
R9厘 もrf
, の竪
く ぜ7’}
り雪” 5欝ヤマ}郭
ダ ヘ
ワ
の曳Ψ-. ん「にど
t4
;
噂』
1一 一t
隔 L
tl
fx tt l, u
Lh XJヤ レ
r vら ’t レ、L
ぐfN㌧、唯 lt
弓z
e
,
ノ、 ”d
s
t
」 み
s t 4 “
へρ_
「 、㌧
ず
ノ
覧
、
尋職
し, ヤ
’ 一’」
ρレ
一一
L一十 L
x(i一 X
♂ぞ”押
5 参ド ーー
、、㌧ ㍗^♂♂㌧ー
デ .♂ぎ・闘い
ヌ
’^,
㌔、
{
し ♂「
「吊」 、
覗 (
ρ
’
sNt
ザ虫’
t) “
1雷‘
~
t二一 へ
、う 冒
甘
τ‘ノ ㌧鼻,
’〆」
甲勘♂
’
,
%
eム
’ r
匹
, 航
マ
㍉,旨e㌧箆
3貞
HL
rl
一」 x 一へf7 Tミ
㌧
」
デ
v吟Lls
一
t 一 :r :t
r 一
r
」’
擁
へ
ず
覧 障’
Sl
f
E
三つ’・hジこ
’、ぐ ’ 一
L. vH
N
十し
しh咲
、
唱
’T 一t) s.j
J; Y へ
,
NAtt
’
r
」
r
愉ぐ一r
一宰て、 、
匹 ’
ド
」、
1
」》
■
〆
’
ノ’ 隔
”’ ,
’ 贈
、 「
r‘
「
,
.J
l一 」lt掃
(ノ
♂〉
”
ゾ、 ,
ン 野,
T’
L㌔
へ r
㌧「キ
,」r
、
,
、
ピ
嚇 ))
㍉’
「φ
レt
J
〆
r
一、
’
’
」
㍉}
4 ~
’、、
’
一
ttびソ “♂
臨 よ’
しtt
i
!1 l
i?
F
F
,
しし. v
Ψ
ほマル
戸
,㍗「ナ
博 マ
ゴ
占一 覇γ、U
tt
’
N )L .” it““
’tノ㌔
・朝㌔、,t
」
七樽
陰o
nへ
温ゲ
’蟷」【の r
「}・へ∵
t vプb州
楠詩
’
レ
i
’
甲
噛
h
1
ビ
s
へ’1“s
tl 卜 N・1’・ぐガ
’
7
r”〆㌻勲,P㌧Cパ電
財ボ、sl
少君 ぎl n ,i’t
「
‘ ’81
ら
㌃ゴ.
,虞へ
監』’r
Ψタ,
、
’
ご
「
ムぽ
’
tし
く女
hEt
二製畠
コr縞
や♂匂
、、
話
7㌦し・、㌧
でへ.♂.
い ド♂
ユ
ワ ㌔♂▼
》轟,」 、
.詩、
ζ㌃舌
【
ヤ ↓
門’ 、
ノ
”
s
t
済
’ ノ
ー
一身
エ
φρ
?
写b
’
うへ
、
、
’
=
一4, ノ・
し
湿しム曇⊥㌔
琴奪’釜
“f
、誤
ノ
,吾3 ~
@
E墨ゴ詣勲・
,
、昌1塩ゲる」
ノ曳み 一L- 、
〆! 戸梅ゴー・
ゆ ゐヒ ド
梶ン飽一り
’
x
-L
’r一 ”
L
tv j
む二.L s.x
ノし. tss
’唱
~㌧、, 、寸 一、 巾





























