Embed Size (px)
DESCRIPTION
que son los lipidos y sus sintesis
Citation preview

Bolilla 5
Metabolismo de los lípidos
- Digestión y absorción de lípidos.
- Transporte de lípidos.
- Degradación de Ácidos Grasos.
- Cuerpos cetónicos.
- Biosíntesis de lípidos
Metabolismo del colesterol.
Regulación molecular.

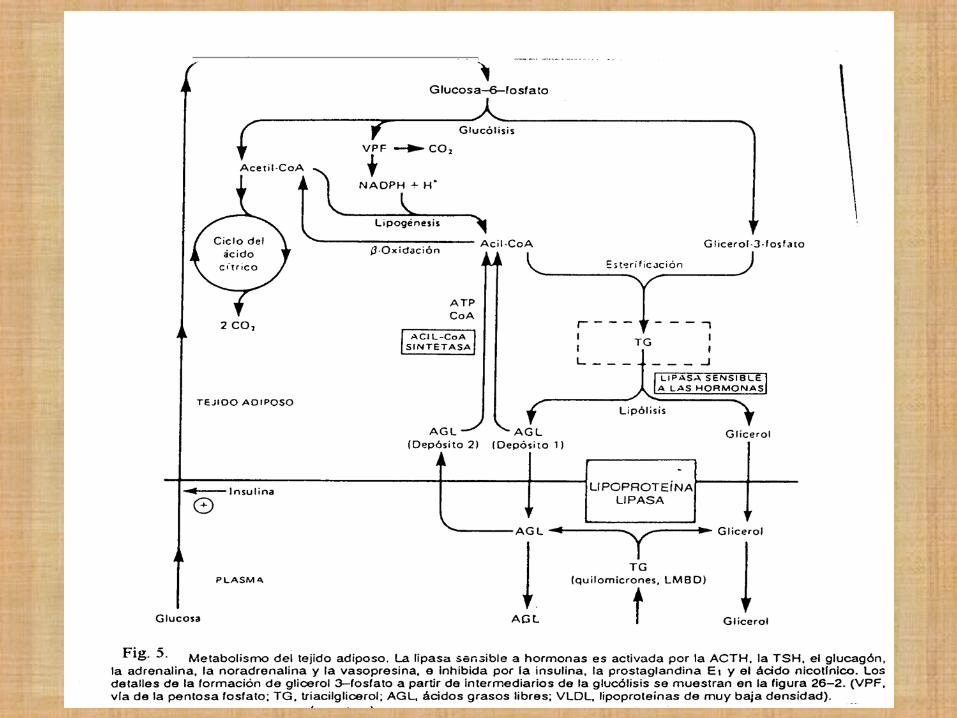
LAS HORMONAS ACTIVAN LA MOVILIZACIÓNN
DE TRIGLICERIDOS ALMACENADOS
- Cuando existe necesidad de energía
metabólica se movilizan las reservas de TG
almacenadas en el tejido adiposo.
- Así se transportan hacia los tejidos: músculo
esquelético, corazón, corteza renal.
- En estos órganos los ácidos grasos de los TG
se oxidan para producir Energía.
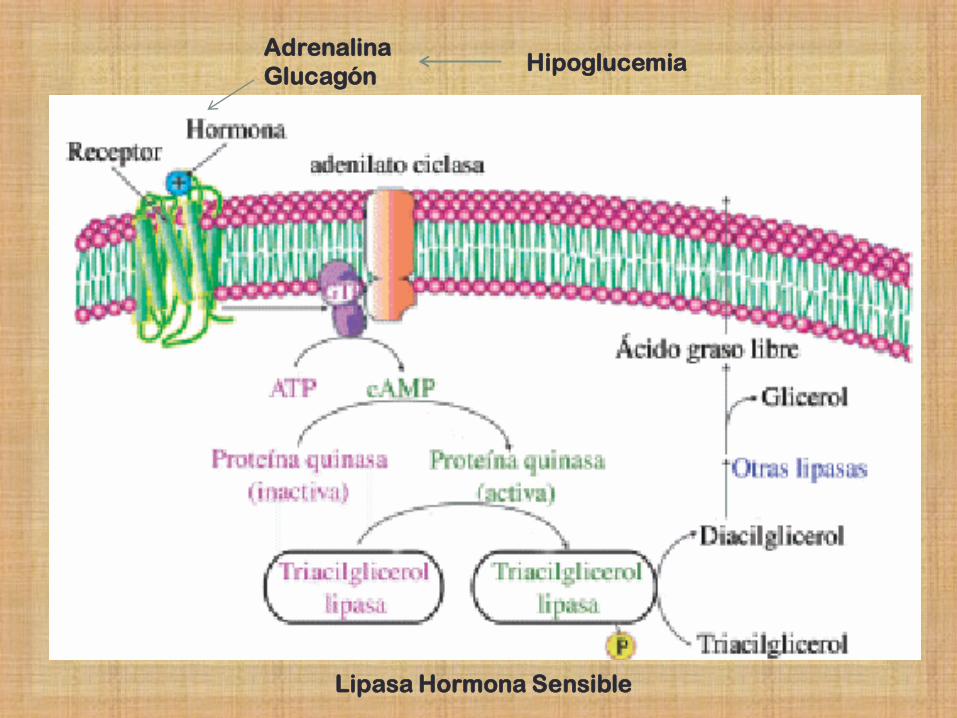
Adrenalina
Glucagón Hipoglucemia
Lipasa Hormona Sensible
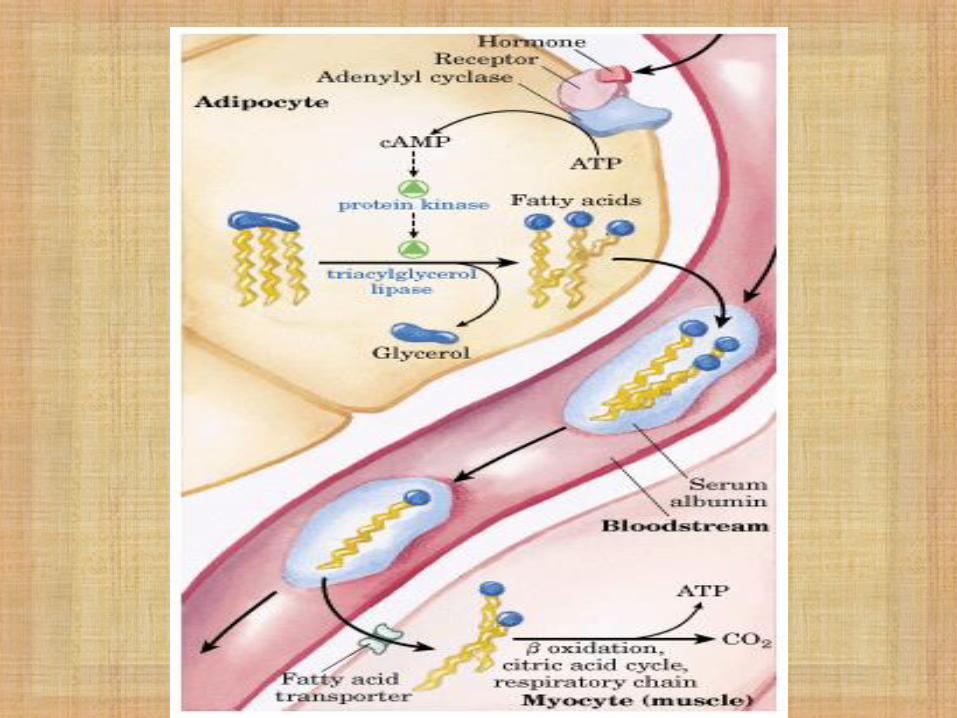

Degradación de Ácidos Grasos
- Los AG cubren hasta el 40% de las
necesidades totales de combustible en una
dieta normal.
- En períodos de ayuno o durante la
hibernación en ciertos animales, los AG son
la única fuente de energía.

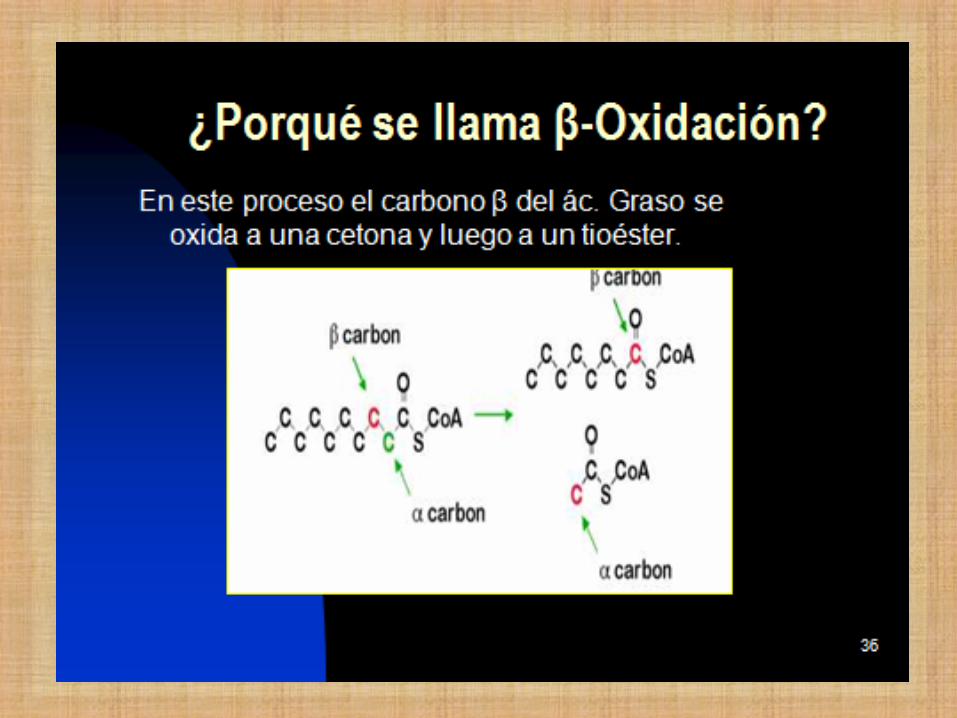
DEGRADACIÓN DE LÍPIDOS β-Oxidación de Ácidos Grasos
Ocurre en tejidos como: Hígado, músculo
esquelético, corazón, riñón, tejido adiposo,
etc.
Comprende la oxidación del carbono β del
ácido graso.
Se lleva a cabo en las MITOCONDRIAS.
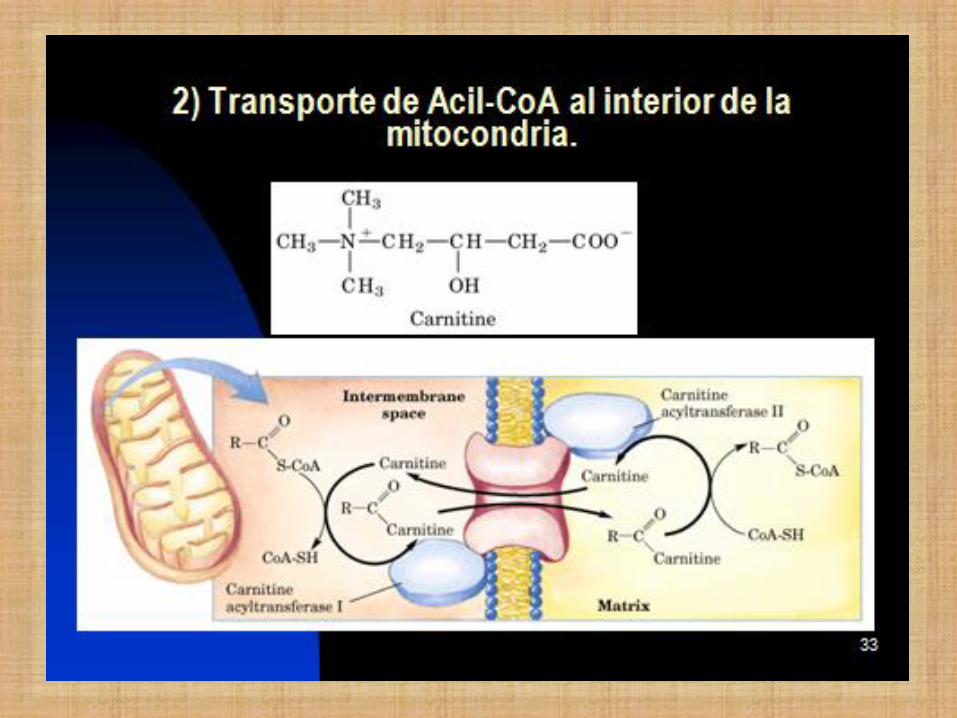
Antes debe ocurrir:
1. Activación del ácido graso (requiere energía
en forma de ATP)
2. Transporte al interior de la mitocondria
7
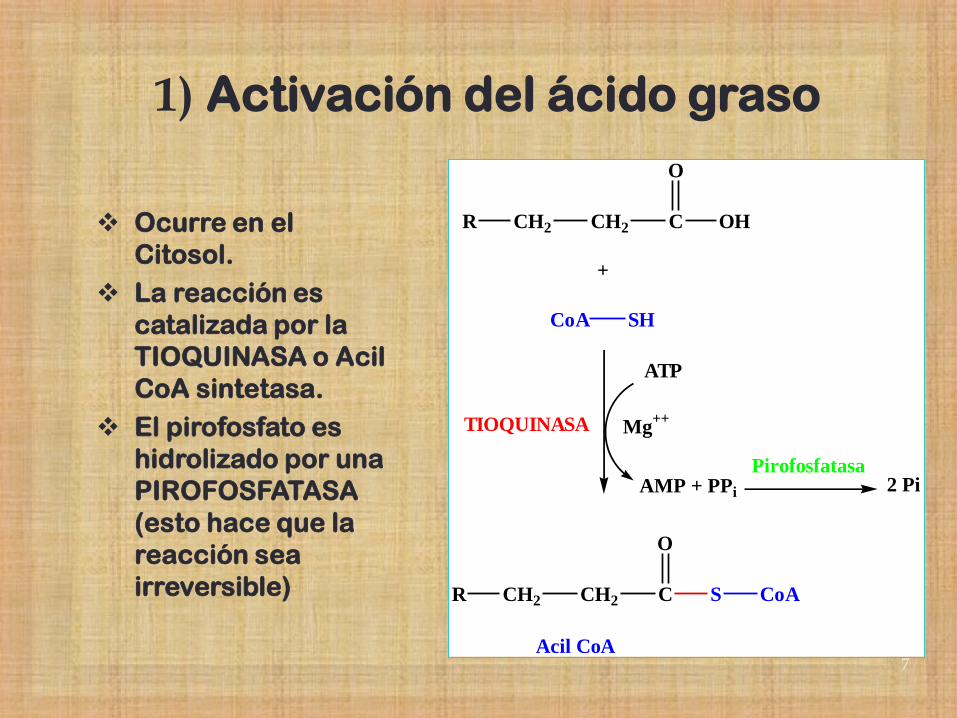
1) Activación del ácido graso
Ocurre en el
Citosol.
La reacción es
catalizada por la
TIOQUINASA o Acil
CoA sintetasa.
El pirofosfato es
hidrolizado por una
PIROFOSFATASA
(esto hace que la
reacción sea
irreversible)
R CH2 CH2 C
O
OH
+
CoA SH
ATP
AMP + PPi
Mg++TIOQUINASA
R CH2 CH2 C
O
S CoA
Acil CoA
2 PiPirofosfatasa

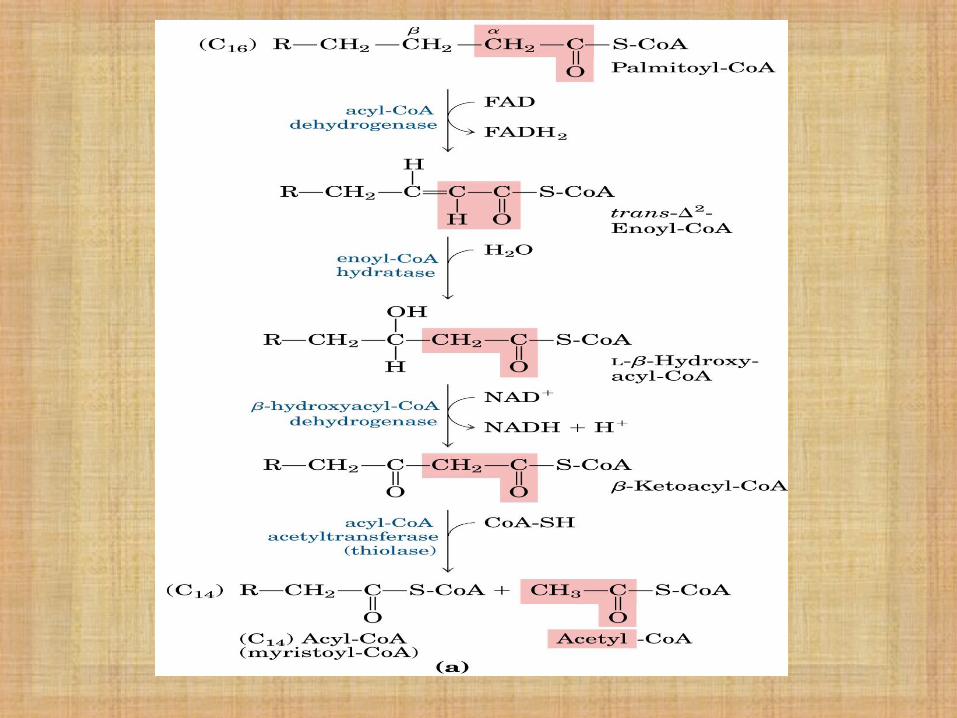
β- Oxidación de Ac. Grasos
Los AG de cadena larga son procesados por las mismas 4 etapas cíclicas.
Por ciclo, se eliminan por oxidación sucesiva, 2 carbonos a partir del extremo carboxílico.
Se produce una molécula de Acetil-CoA en cada ciclo.
El acetil-CoA producido entra en el ciclo de Krebs para producir energía, oxidándose a CO2 y H2O.
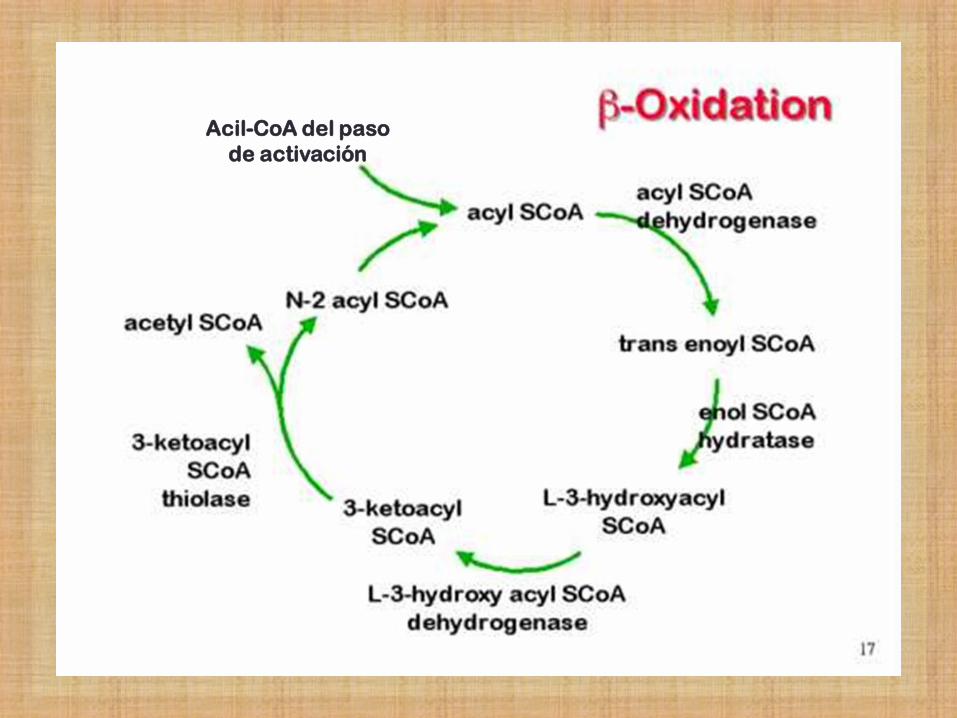
Acil-CoA del paso
de activación
12
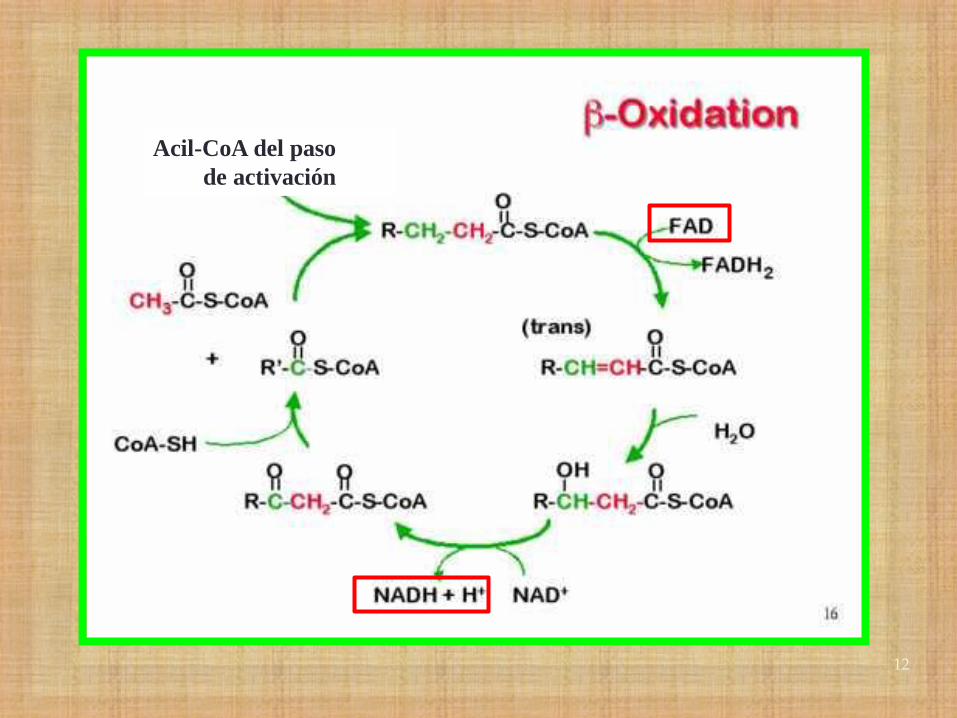
Acil-CoA del paso
de activación
14
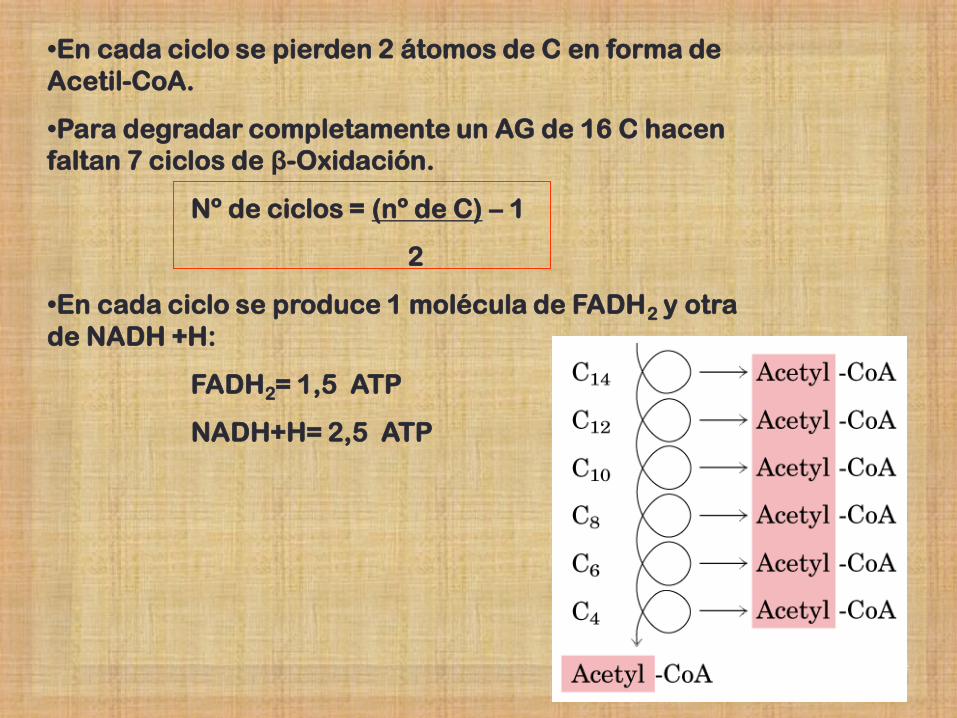
•En cada ciclo se pierden 2 átomos de C en forma de
Acetil-CoA.
•Para degradar completamente un AG de 16 C hacen
faltan 7 ciclos de β-Oxidación.
Nº de ciclos = (nº de C) – 1
2
•En cada ciclo se produce 1 molécula de FADH2 y otra
de NADH +H:
FADH2= 1,5 ATP
NADH+H= 2,5 ATP

BALANCE ENERGÉTICO
PRODUCCIÓN DE ATP EN LA BETA- OXIDACIÓN
7 MOLÉCULAS DE FADH2 2 ATP X 7 = 14 ATP
7 MOLÉCULAS DE NADH 3 ATP X 7 = 21 ATP
SUBTOTAL 35 ATP
8 MOLECULAS ACETIL CoA 12ATP X 8 = 96 ATP
SUBTOTAL = 131 ATP
OXIDACIÓN DE PALMITATO A PALMITOIL CoA = - 2 ATP
TOTAL 129 ATP
16
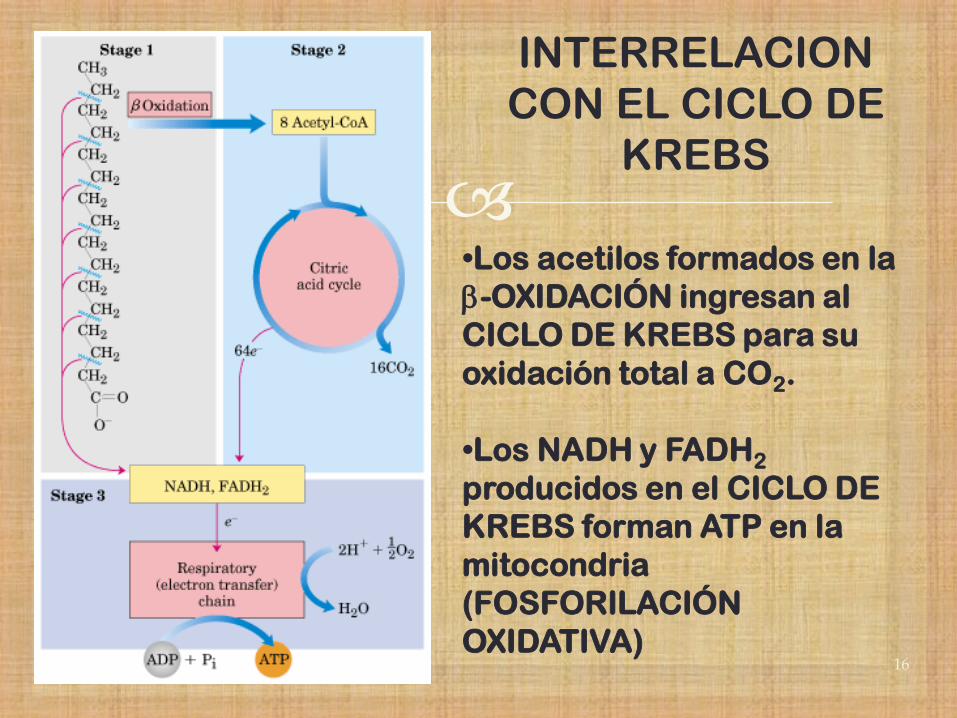
INTERRELACION
CON EL CICLO DE
KREBS
•Los acetilos formados en la
b-OXIDACIÓN ingresan al
CICLO DE KREBS para su
oxidación total a CO2.
•Los NADH y FADH2
producidos en el CICLO DE
KREBS forman ATP en la
mitocondria
(FOSFORILACIÓN
OXIDATIVA)
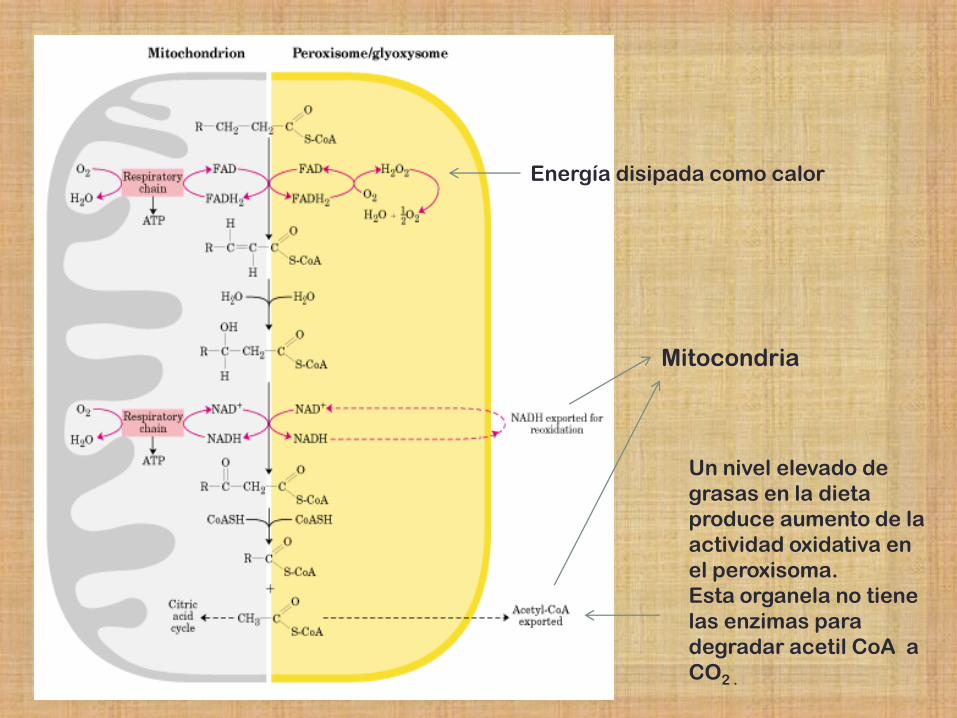
Energía disipada como calor
Un nivel elevado de
grasas en la dieta
produce aumento de la
actividad oxidativa en
el peroxisoma.
Esta organela no tiene
las enzimas para
degradar acetil CoA a
CO2 .
Mitocondria
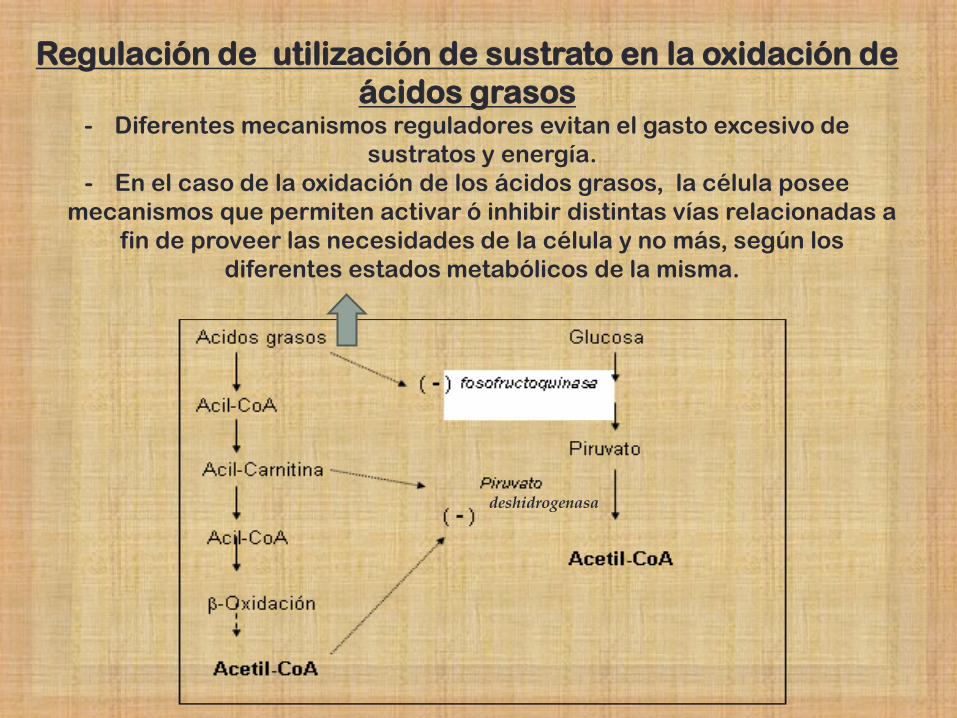
Regulación de utilización de sustrato en la oxidación de
ácidos grasos - Diferentes mecanismos reguladores evitan el gasto excesivo de
sustratos y energía.
- En el caso de la oxidación de los ácidos grasos, la célula posee
mecanismos que permiten activar ó inhibir distintas vías relacionadas a
fin de proveer las necesidades de la célula y no más, según los
diferentes estados metabólicos de la misma.
deshidrogenasa
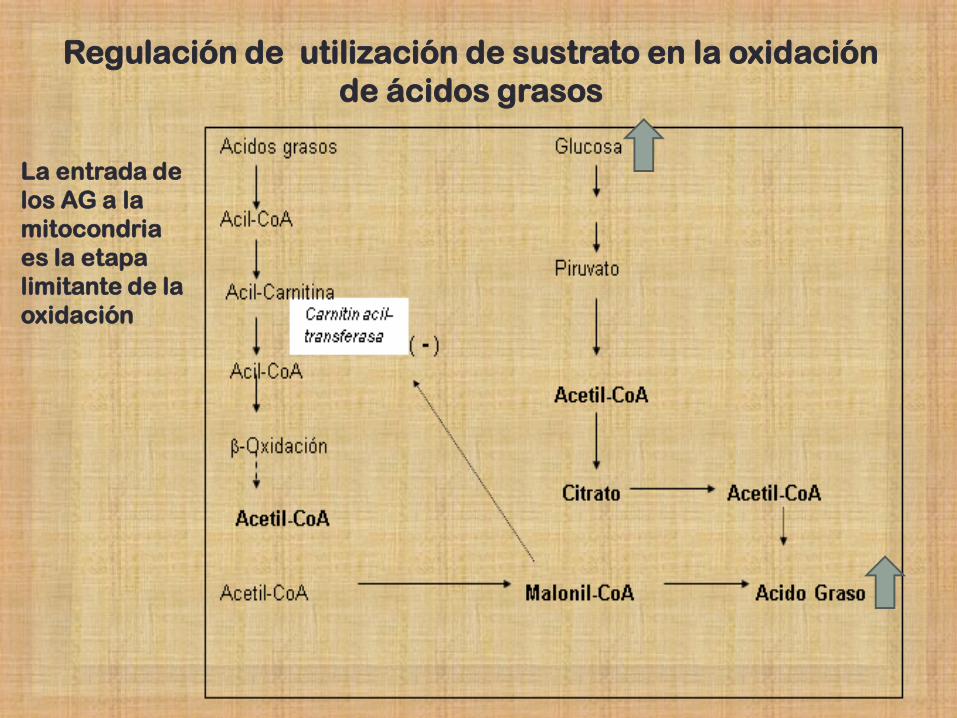
La entrada de
los AG a la
mitocondria
es la etapa
limitante de la
oxidación
Regulación de utilización de sustrato en la oxidación
de ácidos grasos

Durante la combustión completa de
un AG se produce gran cantidad de
agua, que deriva de las reacciones
de fosforilación asociadas.
Así, los camellos obtienen agua por
degradación de sus reservas
grasas.
El oso pardo hiberna por
períodos de hasta 7 meses.
Mantiene su temperatura
corporal cercana a la
normal.
Consume 6000Kcal/día
durante la hibernación.

22
CUERPOS CETÓNICOS
Son compuestos derivados de acetil CoA que al no entrar al
ciclo de Krebs son convertidos en cuerpos cetónicos.
El hígado es el principal productor ya que posee todas las
enzimas necesarias. Es incapaz de usarlos como combustible.
Los órganos que los usan son: cerebro, músculo esquelético,
corazón y otros.
Solo se usan como fuente de energía en situaciones metabólicas
especiales. Ej.: Diabetes, ayuno prolongado, dieta grasa.
El aumento de estos provoca Acidosis Metabólica.
La cetogénesis se produce en las mitocondrias hepáticas.

23
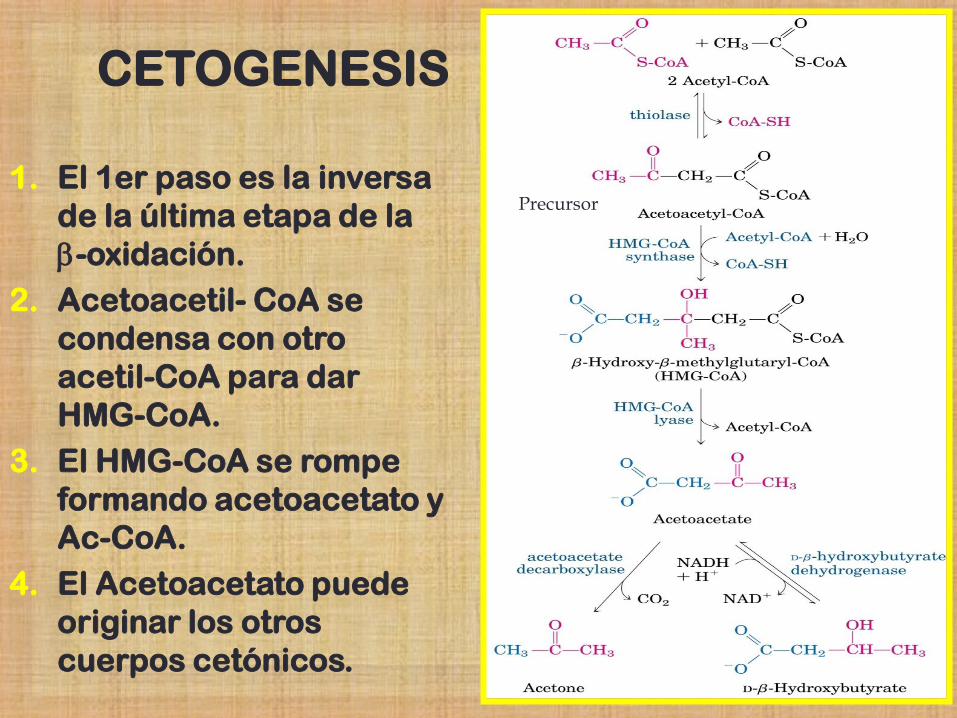
Formación de Cuerpos Cetónicos
Cetogénesis
Después de la degradación de los AG, Acetil-CoA es oxidado en el Ciclo de Krebs.
Para esto es necesaria la presencia de oxalacetato (1er intermediario del ciclo de Krebs).
Si la cantidad de este es insuficiente, las unidades de acetil-CoA son utilizadas mediante una vía alternativa en la que se producen los Cuerpos Cetónicos
Estos compuestos se forman principalmente en el hígado, a partir de acetil-CoA mediante una serie de etapas.
H3C C
O
CH3
acetona
H3C C
O
CH2 C
O
O-
acetoacetato
H3C CH
OH
CH2 C
O
O-
3-OH-butirato
24
CETOGENESIS
1. El 1er paso es la inversa
de la última etapa de la
b-oxidación.
2. Acetoacetil- CoA se
condensa con otro
acetil-CoA para dar
HMG-CoA.
3. El HMG-CoA se rompe
formando acetoacetato y
Ac-CoA.
4. El Acetoacetato puede
originar los otros
cuerpos cetónicos.
Precursor
25
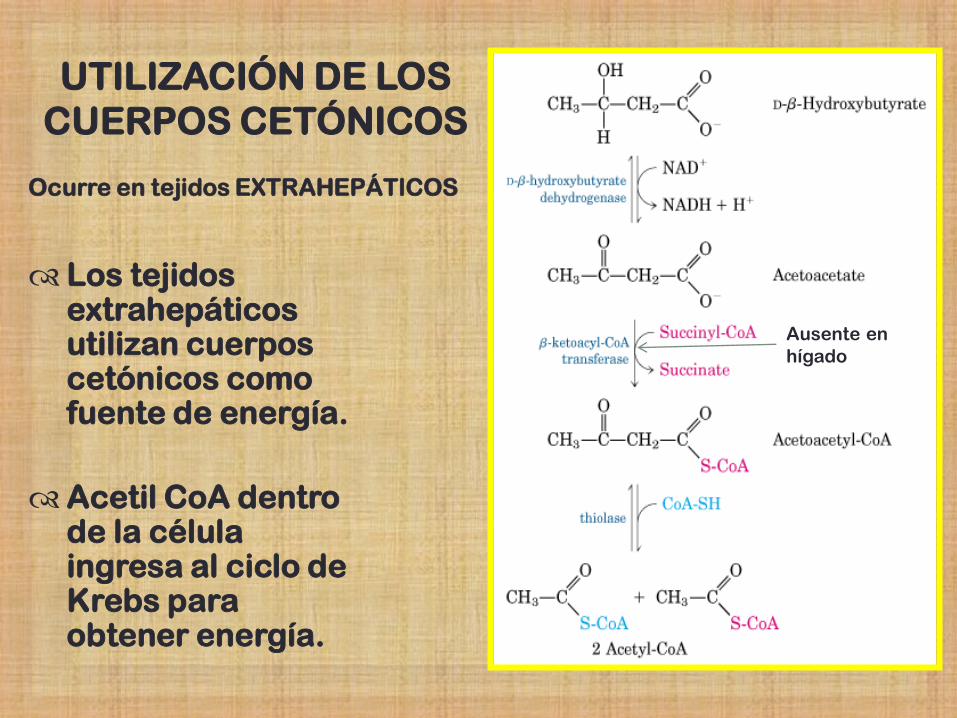
UTILIZACIÓN DE LOS
CUERPOS CETÓNICOS
Los tejidos extrahepáticos utilizan cuerpos cetónicos como fuente de energía.
Acetil CoA dentro de la célula ingresa al ciclo de Krebs para obtener energía.
Ocurre en tejidos EXTRAHEPÁTICOS
Ausente en
hígado
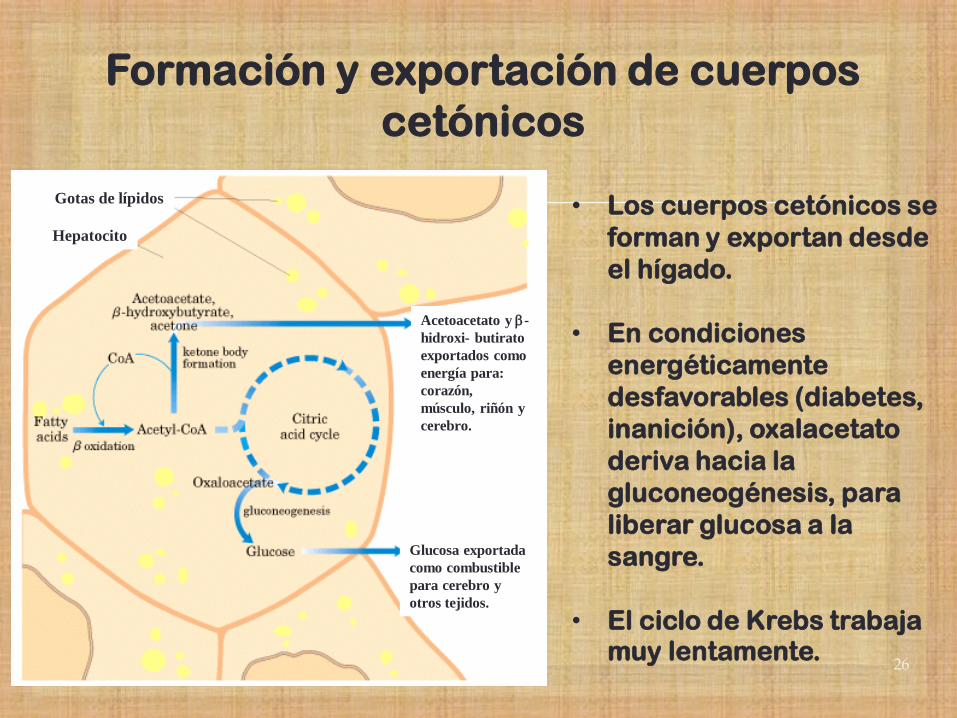
26
Formación y exportación de cuerpos
cetónicos
• Los cuerpos cetónicos se
forman y exportan desde
el hígado.
• En condiciones
energéticamente
desfavorables (diabetes,
inanición), oxalacetato
deriva hacia la
gluconeogénesis, para
liberar glucosa a la
sangre.
• El ciclo de Krebs trabaja muy lentamente.
Gotas de lípidos
Hepatocito
Acetoacetato y b-
hidroxi- butirato
exportados como
energía para:
corazón,
músculo, riñón y
cerebro.
Glucosa exportada
como combustible
para cerebro y
otros tejidos.

En las personas sanas la producción de acetona es muy baja.
Aumenta significativamente en las personas diabéticas no
tratadas.
Al ser un compuesto volátil la acetona se elimina a través de la
respiración y por orina confiriendo un olor característico que
permite diagnosticar esta patología.
Cetólisis: el hígado no puede degradar los cuerpos cetónicos ya
que no posee la enzima TIOQUINASA O CETOACIL -COA TRANSFERASA para activarlos.
Cuando se produce un desequilibrio entre la CETOGÉNESIS y la
CETÓLISIS ocurre lo que se denomina CETONEMIA, que en
condiciones normales es baja.
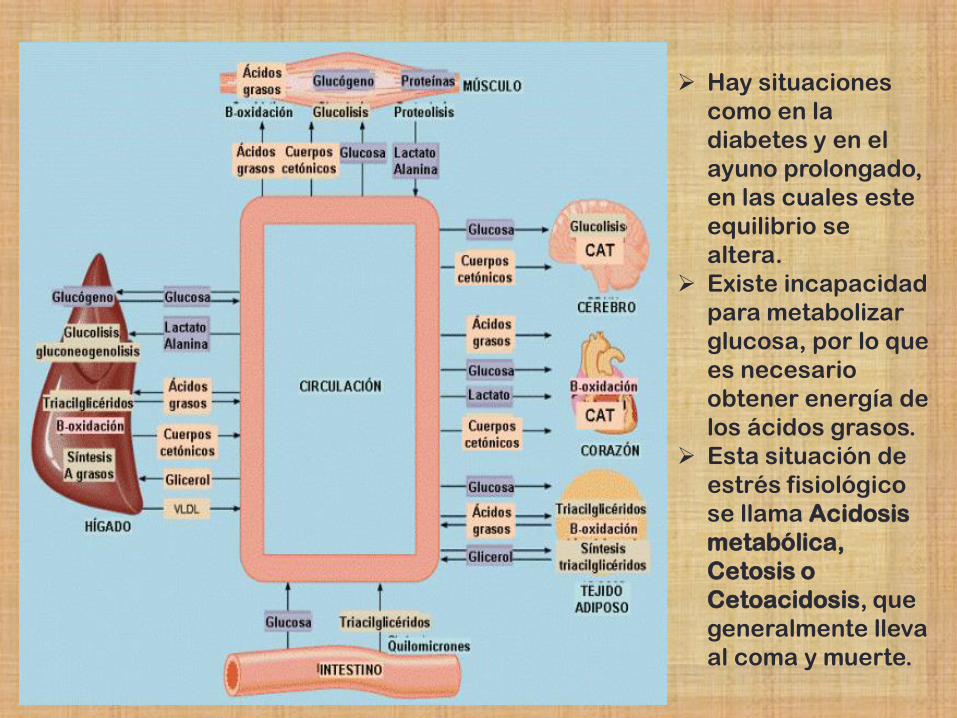
Hay situaciones
como en la
diabetes y en el
ayuno prolongado,
en las cuales este
equilibrio se
altera.
Existe incapacidad
para metabolizar
glucosa, por lo que
es necesario
obtener energía de
los ácidos grasos.
Esta situación de
estrés fisiológico
se llama Acidosis
metabólica,
Cetosis o
Cetoacidosis, que
generalmente lleva
al coma y muerte.





























