Embed Size (px)
Citation preview

December 2001 Vol 51 No 12 BioScience 1005
Features
Species diversity is being lost in habitats that areincreasingly diminished by development fragmenta-
tion and urban runoff the sensitive species drop out and afew aggressive ones persist at the expense of others Alarmedby declining biodiversity many conservationists and re-searchers are asking what happens to ecosystem functioningif we lose species how diverse communities can be restoredwhich (if any) particular species are critical for performingecosystem services and which functions are most critical toecosystem sustainability In southern California 90 of thecoastal wetland area has been destroyed and remaining wet-lands continue to be damaged even the regionrsquos protected re-serves are threatened by highway and utility-expansion pro-jects The fate of biodiversity in these diminished wetlandsserves to warn other regions of the need for continual as-sessment of the status and function of both common and rarespecies as well as the need for experimental tests of theirimportancemdashbefore they are lost
This article synthesizes data for tidal marshes of the Cali-fornian biogeographic region which stretches from PointConception near Santa Barbara south to Bahiacutea San Quintiacutenin Baja California We focus on the broad marsh plain whichis dominated by eight species of halophytes (salt-tolerantplants Figure 1) From regional censuses we document therecent loss of short-lived species from several wetlands Fromeyewitness accounts of tidal-exclusion events at Estero dePunta Banda and Tijuana Estuary we link species loss to theinterruption of tidal influence And from experimental plant-ings of marsh halophytes in a bare restoration site we docu-ment the difficulty of restoring plant diversity demonstrate
Joy B Zedler Aldo Leopold Chair of Restoration Ecology Botany De-
partment and Arboretum 430 Lincoln Drive University of Wisconsinndash
Madison Madison WI 53706 is a wetland ecologist who has studied
southern California coastal wetlands for about 30 years John C Call-
away assistant professor in the Department of Environmental Science
University of San Francisco San Francisco CA 94117 conducts field
research at Tijuana Estuary and San Francisco Bay he is a wetland
ecologist whose research focuses on sediment and vegetation dynamics
in restored wetlands Gary Sullivan is a wetland ecologist with expe-
rience in freshwater lakes and streams estuaries and salt marshes
his current focus is on restoring large wetlands along the Illinois
River for a nonprofit organization The Wetlands Initiative Chicago IL
60604-3703 copy 2001 American Institute of Biological Sciences
Declining Biodiversity Why Species Matter andHow Their Functions MightBe Restored in CalifornianTidal MarshesJOY B ZEDLER JOHN C CALLAWAY AND GARY SULLIVAN
BIODIVERSITY WAS DECLINING BEFORE
OUR EYES BUT IT TOOK REGIONAL
CENSUSES TO RECOGNIZE THE PROBLEM
LONG-TERM MONITORING TO IDENTIFY
THE CAUSES AND EXPERIMENTAL
PLANTINGS TO SHOW WHY THE LOSS OF
SPECIES MATTERS AND WHICH RESTORA-
TION STRATEGIES MIGHT REESTABLISH
SPECIES
Editorrsquos note This article was derived from the plenary address DrZedler made at the March 2001 AIBS annual meeting
1006 BioScience December 2001 Vol 51 No 12
Features
what each species potentially contributes to ecosystem func-tioning and develop recommendations for restoring biodi-versity Our experimental tests of the relationship of speciesrichness to ecosystem functioning at Tijuana Estuary con-tribute to the debate about whether ecosystem functioning isinfluenced more by the presence of many species or by thepresence of one or two high-performance species We eval-uate the contribution of multiple and individual species andshow that individual species can match the performance ofsix-species assemblages (sextets) but no single species excelsin all functions Our experimentation with increasing levelsof biodiversity in an actual restoration site (a bare marshplain) provided results of immediate use to biodiversity en-hancement projects while simultaneously vegetating the site
Does diversity matter in restoration Questions about species number species identity and specificfunctions are especially relevant to the restoration of ecosys-tems When confronted with a bare restoration site one be-gins by asking if planting is necessary and if so of which andhow many species The answers may well depend on therestoration targetmdashwhat level of diversity and function is de-sired and when The goal of many restoration projects is tomatch the structural and functional attributes of regional ref-erence ecosystems as quickly as possible Thus any plantingpractice that speeds the development of species-rich vegeta-tion is of interest as are vegetation attributes that support moreanimal species and confer greater resilience to the system
The role of species richness has been explored in several di-versity experiments by planting randomly drawn assem-blages of herbaceous plants and evaluating a range of func-tional attributes most notably biomass accumulation Ingeneral increasing numbers of plant species lead to increasesin productivity (Naeem et al 1994 1995 1996) nutrient re-tention (Ewel et al 1991) resiliency (Tilman and Downing1994) reliability (Johnson et al 1996 Naeem and Li 1997)and decreases in invasibility of other species (Tilman 1997Symstad 2000) In a series of diversity experiments con-ducted in eight European countries Hector and colleagues(1999) reported that a 50 decline in species richness reducedaboveground biomass by 10ndash20 A recent review of thediversity literature confirmed that many functions increase inconcert with a rise in the number of plant species althoughthere was little support for a continually increasing linear re-lationship between species diversity and function (Schwartzet al 2000) In addition diversity effects reach across trophiclevels mycorrhizal diversity affects plant diversity (van der Hei-jden et al 1998) and insect diversity is related to plant diversity(Knops et al 1999) although weakly in one case (Siemann etal 1998)
Although a variety of functions respond positively togreater diversity it remains unclear whether the response isdue to the number of species or the chance inclusion of anoutstanding stress-tolerator colonizer competitor or otherldquosuperplantrdquoTo restore habitat most effectively managers needto know how many and which species to plant so that the veg-
etation will perform multiple functions such as persistencecanopy development and retention of carbon and nutri-ents If the target community has only one species for exampleSpartina alterniflora monotypes along the Atlantic Coastthen restoration is greatly simplified If the reference systemincludes many species a subset of plantings might suffice foraccelerating the development of vegetation To simplifychoices some researchers classify species into functionalgroups based on life span (annual versus perennial) growthform (succulent or nonsucculent upright or trailing grass orforb) or presence of nitrogen-fixing symbionts Experimentswith species drawn randomly from functional groups have un-covered effects on productivity nutrient availability andother functions (Ewel et al 1991 Chapin et al 1992 Silver etal 1996 Hooper and Vitousek 1997 Huston et al 2000)Thus introducing one or more representatives of a func-tional group to a restoration site might suffice for providingcritical ecosystem functionsmdashfor example nitrogen-fixersto accelerate nitrogen accumulation (Crocker and Major1955Vitousek and Reiners 1975) or highly productive speciesto increase overall grassland productivity (Symstad et al1998) Restoration efforts could then be streamlined by lim-iting initial plantings to a few key members of the targetcommunity In southern California salt marshes howeverfunctional groups have proven elusive (Sullivan and Zedler1999)
Californian salt marshesThe Californian salt marsh (Figure 1) is a useful model sys-tem for testing the effect of diversity on ecosystem functionfor several reasons
It includes a small number of species (eight dominatethe marsh plain)
Figure 1 Salt marsh vegetation at Bahiacutea San Quintiacuten BajaCalifornia Norte Mexico
It has naturally occurring assemblages that range fromone to many species
It includes various growth forms (Table 1)
The plants are readily grown from seed and all becomereproductive in their first or second year
As recommended by Vitousek and Hooper (1993 p 14) weldquomanipulate diversity as an experimental treatment and em-phasize the range of diversity from one to ten speciesrdquo Con-cern about experimentation with ldquoimmature vegetationrdquo(Huston et al 2000) is not an issue for restoration sites be-cause the establishment of vegetation on newly graded sub-strate always involves immature plants and any approachesthat can accelerate the maturation process are immediatelyuseful to ecological restoration In other words we seek waysto increase the rate of biomass accumulation and the devel-opment of complex canopies in sites with coarse soils and lim-ited nutrients especially nitrogen (N) We ask whether plant-ing is necessary and if so how many and which species needto be planted to accelerate the development of complexcanopies and the accumulation of soil N
The Californian tidal marsh (Figure 2) has a fringe ofSpartina foliosa a broad marsh plain and a high marsh tran-sition to upland Tides inundate S foliosa twice daily and
the marsh plain at least once daily with water depth varyingby season Tidal amplitudes are narrowest and marsh plaininundation least frequent in March and April about the timevegetation begins to grow exponentially (Winfield 1980)The marsh plain is riddled with tidal creeks and spans a ~30-cm elevation range at our Bahiacutea San Quentiacuten reference sys-tem (Zedler et al 1999) and a ~20-cm range at Tijuana Es-tuary (approximately 60ndash80 cm NGVD [National GeodeticVertical Datum] Figures 2 3 Zedler et al 1992) Marshes inMexicorsquos Bahiacutea San Quintiacuten have large tidal channels with un-interrupted seawater flows (Figure 3)
The regular influence of tides is punctuated by river flood-ing and sea storms over the long term Annual rainfall aver-ages only 25 cm in San Diego and river flows are corre-spondingly low however high interannual variation leads tocatastrophic flooding Wetlands with large watersheds orwith disturbed lands (agricultural fields urbanization) up-stream are subject to major inflows of sediments for exam-ple Mugu Lagoon lost 40 of its low-tide volume during twofloodingndashsedimentation events (Onuf 1987) Such extremesedimentation episodes are attributable to disturbance ofvegetation and soils upstream of the estuaries Tijuana Estu-ary has a large watershed but the Tijuana River did not floodfrom 1944 to 1977 Since then however flows exceeded 70 cu-
bic meters per second during the win-ters of 1978 1980 1983 1986 19881993 1995 and 1997 In addition amajor sea storm overwashed thedunes during the 1983 El Nintildeo eventfilling tidal channels with sand and re-ducing tidal inflows (Zedler et al1992) The estuary mouth subse-quently closed in April 1984
Marshes north of the USndashMexicoborder are still numerous (Figure 4)but their loss of area and habitat qual-ity has caused many wetland-depen-dent species to become threatenedwith extinctionmdashfor example an en-dangered hemiparasitic annual plantknown as salt marsh birdrsquos-beak(Cordylanthus maritimus ssp mari-tumus) the wandering skipper(Panoquina errans) five species oftiger beetle (Cicindella spp) the light-footed clapper rail (Rallus longirostrislevipes) and the Beldingrsquos Savannahsparrow (Passerculus sandwichensisbeldingi) These rare species and sev-eral more abundant wading birds andwaterfowl species are highly valuedby the public But these charismaticspecies are not the only victims ofhabitat loss and degradation Themarsh plain halophytes although notvery showy (Table 1) are still at risk
December 2001 Vol 51 No 12 BioScience 1007
Features
Table 1 Life history and growth forms of eight halophytes common to the marshplain of Californian coastal wetlands Illustrations (by Donovan McIntire) showrelative proportions of biomass above and below ground as well as rooting depthsbased on greenhouse experiments Regional distributions are for 23 wetlands withinthe region (Table 2) fewer occurrences indicate sensitivity to tidal closure Recentdeclines (Table 2) were recorded from selected wetlands (mostly San Diego County)Recruitment data (H=high M=medium L=low) are from Table 3 Species codes areSv Salicornia virginica Fs Frankenia salina Jc Jaumea carnosa Lc Limoniumcalifornicum Se Suaeda esteroa Tc Triglochin concinna Sb Salicornia bigeloviiBm Batis maritima
Spatial and temporal patterns of halophyte di-versity on the marsh plain At Bahiacutea San Quintiacuten themarsh plain supports nine species that are both common andabundant (Zedler et al 1999) The species include the eightspecies that dominated the marsh plain of Tijuana Estuary
before 1984 (Zedler et al 1992) The latter 1000-hasite is San Diego Countyrsquos largest tidal wetland anda National Estuarine Research Reserve
Southern California marshes differ in their bio-diversity Historical records of eight halophyte dis-tributions in 23 coastal salt marshes (Table 2) showthat only three species are very widespread (Sv FsJc species codes are in Table 1) Of these Salicorniavirginica has broad ecological tolerance (as indi-cated by its elevation range Figure 2) and is domi-nant in most of the regionrsquos wetlands (Zedler 1982Barbour and Major 1988) including nontidal sys-tems (Zedler et al 1980) Lc is more restricted thanSv Fs and Jc while the remaining four species areregionally restricted in their distribution especiallythe two short-lived species (Sb and Se) The eightspeciesrsquo regional occurrences indicate that speciesrichness is greatest in wetlands with long histories ofgood tidal flow and lowest in wetlands that havebeen closed to tidal flow for one or more lengthy pe-riods For example Los Pentildeasquitos Lagoon wasclosed for 5 years in the 1950s (Carpelan 1969) itlacks three of the eight marsh plain species San ElijoLagoon (Figure 5) is usually closed to tidal action andit lacks five of the marsh plain halophytes In each ofthese wetlands tidal flows are constrained by botha railroad bridge and at the tidal inlet a coastalhighway
The spatial pattern of fewer species in wetlandswith impaired tidal flow suggests that mouth closure has re-duced biodiversity Two lines of evidence support this cause-and-effect relationship First a 1998 resurvey of San DiegoCountyrsquos tidal marshes documented the loss of species atsites with impaired tidal flows Species Se was lost at four sites
and nearly lost at two additionalsites Lc was lost at two sites and Sbwas lost at one site and nearly lostat Tijuana Estuary (Table 2) Sec-ond the species declines were wit-nessed during two 1984 tidal clo-sure episodes one at Estero dePunta Banda in Baja Californiafollowing the permanent diking ofa large marsh area (Ibarra-Obandoand Poumian-Tapia 1991) and oneat Tijuana Estuary during a tem-porary (8-month) closure (Zedleret al 1992) At Estero de PuntaBanda monthly sampling docu-mented that Sb and Se persistedonly 13 months after a large por-tion of the salt marsh was diked AtTijuana Estuary the mouth closedin April 1984 after the 1983 El Nintildeostorm filled the tidal channels Af-ter closure there was no rainfall
1008 BioScience December 2001 Vol 51 No 12
Features
Table 2 Occurrence of eight marsh-plain halophytes in southern California coastalwetlands Species are ranked according to the number of sites in which they occurwetlands are in order of the number of species they support Data in boxes representchanges in the status of a species at a site determined by G Sullivanrsquos 1998 census ( = now rare 0 = now absent)
Figure 2 Topographic profile of Volcano Marsh at Bahiacutea San Quintiacuten
Transects extend from the upland (left) to the bay (right) crossing tidal
creeks and channels as well as hummocks formed by ancient lava
deposits Elevations are relative to the lowest occurrence of Spartina
foliosa (Zedler et al 1999) Occurrences of three species are given + is a
high-marsh indicator Salicornia subterminalis X is S virginica and
the third row of symbols under transect b is cordgrass S foliosa
in an unusually dry growing season Channel salinities roseto about 60 parts per thousand (ppt) and soil salinities roseto an average of 104 ppt (Zedler et al 1992) Species Sb andSe were virtually extirpated and did not recover from the 8-month nontidal catastrophe (Table 2)
The pattern of Sb and Se loss with reduced tidal influenceis consistent across the region in both space and time and atboth Estero de Punta Banda and Tijuana Estuary The patternis also obvious from monitoring small-scale (025 m2) quadrats(Figure 6) Prior to tidal closure the marsh plain of TijuanaEstuary supported a mean of 45 species during closure themean dropped below 20 Little recovery was evident in sub-sequent years despite the return of tidal influence Our May1999 data from Bahiacutea San Quintiacuten had a mean of 47 speciesper 025-m2 quadrat (n = 224) We conclude that biodiver-sity has declined at Punta Banda and Tijuana estuaries as aresult of interrupted tidal influence
Testing speciesrsquo roles in ecosystem function Given the 90 loss of wetland habitat in southern Californiarestoration is a high priority Early attempts sought only toprovide vegetation cover It was common to plant just one ortwo easily grown species (S foliosa in low elevations Sv on themarsh plain Zedler 2001) We hypothesize that ecosystemfunctioning might be impaired if too few species are re-stored No one knows how many are needed for the systemto develop at maximum speed or to achieve desired levels ofecosystem functions such as the development and mainte-nance of natural community composition the developmentof appropriate canopy structure and the accumulation of or-ganic matter and nitrogen These span the range of indica-tors that Noss (1990) recommends for monitoring biodi-versity The assessment of just one function such as netprimary productivity above ground (or its surrogate struc-tural attribute biomass at the end of the growing season) isnot likely to serve ecosystem managers or settle internationaldebates about the importance of diversity to ecosystem func-tioning Hence we tested the effects of species-rich plantingson the development of multiple functions that are critical inrestoration sites
In 1997 we established a large field experiment at TijuanaEstuary to test the effects of species richness on three specificresponse variables
1 Recruitment Which species need to be planted andwhich readily establish and spread
2 Canopy complexity Which grow taller and producemore layers as might be needed by insects and birds
3 Belowground biomass and N accumulation Whichplots store the most N in roots and rhizomes
Our experimental site was a graded marsh plain thathad room for 87 2 x 2-m plots distributed among fiveblocks along an excavated tidal channel (Figure 7) Our ex-perimental conditions included single-species plots of eachof the eight species of the nearby reference marsh plusrandomly chosen six- and three-species assemblages whichroughly correspond to small-scale richness levels beforeand after tidal closure at Tijuana Estuary (Figure 6) Sev-eral of our experimental assemblages occurred in TijuanaEstuary in 1974 and Bahiacutea San Quintiacuten in 1999 but ran-dom draws missed the most common trio (Bm Sv Sb)and sextet (Bm Sv Sb Fs Jc Se) from Tijuana Estuaryrsquos025-m2 plots Hence we added one plot with Bm Sv Sband one with Bm Sv Sb Fs Jc Se The 87 field experimentalplots were planted in April 1997 and replanted and weededas needed that summer to maintain planting treatments andcompensate for early mortality The assemblages planted re-sulted in similar numbers of plants introduced within theexperimental site Thus differential starting densities werenot of concern in comparing recruitment Assemblagecomposition was maintained during 1998 by removing
December 2001 Vol 51 No 12 BioScience 1009
Features
Figure 3 A remotely sensed image of Volcano Marshshowing tidal channels Mosaic compiled by BruceNyden Pacific Estuarine Research Lab San Diego StateUniversity
seedlings of other species thereafter recruits were allowedto persist
Seedlings were censused by Roberto Lindig-Cisneros(2001) canopy architecture was documented by Keer andZedler (forthcoming) and biomass and N accumulationwere assessed by J Callaway Because we were most con-cerned about the increasing ability of the site to retain Nwe assessed biomass and N accumulation in midwinter af-ter most plants had slowed growth Furthermore becausewe wanted to minimize destructive sampling we removedsmall root cores in year 2 and root and shoot samples at theend of year 3 Both biomass components were analyzed forabove- and belowground N content
We complemented the field study with an experiment inthe greenhouse where we were able to compare individual as-semblages We replicated all 15 randomly drawn trios and sex-
tets 8-fold all solos 16-fold and the common trioand sextet 16-fold for a total of 400 microcosmsPlanting treatments were maintained in the potsthrough vegetative growth and seedling recruit-ment but assemblage composition did not changeAll but 80 pots were grown for 2 years and as-sessed for canopy attributes and harvested for bio-mass and N content above and below ground
The importance of diversity to restorationResults from the field and greenhouse experimentssupport the general hypothesis that diversity mat-ters in restoration sites Our plantings with sixspecies enhanced recruitment made canopiesmore complex increased biomass and increasedN accumulation
Recruitment Only three species (Sv Sb andSe) recruited readily (Table 3) Of these only one(Sv) is widely distributed (Table 2) and available todisperse into restoration sites It germinates in sea-water and produces abundant seedlings in bothnatural and restored wetlands including our un-planted plots it does not need to be planted torestoration sites if a seed source exists nearby Theother two abundant recruiters Sb and Se are themarsh plainrsquos only short-lived species Personalobservation revealed that Se recruitment potentialwas much lower than that of Sb reflecting its lowerdensity in regional wetlands and differences inseedling mortality within this experiment Thefact that both recruited readily in the bare restora-tion site and poorly in the salt marsh where it oc-curred historically is evidence that the Tijuana Es-tuary marsh plain no longer provides suitablehabitat for these species The remaining five speciesdid not readily colonize either our planted or un-planted plots Their inability to recruit may be at-
1010 BioScience December 2001 Vol 51 No 12
Features
Table 3 Seedling recruitment of eight marsh-plainspecies into experimental plots Data are for the 1998growing season Note that the number of plantsintroduced to the site as seedlings in April 1997(column 2) was similar but only three speciesrecruited extensively Species codes are in Table 1
Code Number planted Number of recruits
Sv 840 17703Sb 735 15978Se 795 1668Tc 810 73Jc 840 33Bm 720 26Lc 795 24Fs 765 2
Figure 4 Coastal wetlands of southern California 1Tijuana Estuary 2Sweetwater Marsh 3 Mission Bay Reserve 4 Los Pentildeasquitos Lagoon 5San Dieguito Lagoon 6 San Elijo Lagoon 7 Batiquitos Lagoon 8 AguaHedionda Lagoon 9 Buena Vista Lagoon (brackish wetland) 10 SanLuis Rey River Mouth 11 Santa Margarita Estuary 12 Las FloresMarsh 13 San Mateo Marsh 14 Upper Newport Bay 15 Bolsa ChicaWetland 16 Anaheim Bay 17 Los Cerritos Marsh (mostly filled anddeveloped) 18 Ballona Wetland 19 Malibu Lagoon 20 Mugu Lagoon21 McGrath Lake 22 Santa Clara River Estuary 23 Ventura River(brackish) 24 Carpinteria Marsh 25 Goleta Slough 26 DeverauxLagoon
tributable to small seed sup-ply poor germination or un-suitable conditions for es-tablishment
Canopy architectureKeer and Zedler (forthcom-ing) found that all plotsrapidly achieved high coverbut that canopy height andlayering were slow to de-velop Cover is a poor mea-sure of canopy complexity itdoes not differentiate speciesor species-richness treat-ments Maximum heightswere greatest for trios andsextets reaching about 60cm by year 3 Sv grew thetallest in the first 18 months of study followed by Sb and SeLayering (the number of times a vertical rod encountersplant parts as it is lowered through the canopy) was greaterfor trios and sextets both of which achieved three layerswithin 18 months The effects of individual species werelikely to be seen early in the development of the plots Fs andSe had six and five layers respectively with Jc at four layersLayering is hypothesized to enhance habitat for both arthro-pods and birds These canopies are far from fully developedreference marshes with monotypic Sv averaged 45 cm tall withseven layers and areas with multiple species were 55 cm talland had 13 layers (Keer and Zedler forthcoming)
Biomass and N accumulation In the field plots above-ground biomass averaged under 300 g per m2 for solos andover 400 g per m2 for sextets Belowground biomass also in-creased with species richness and over timemdashmeans doubledbetween years 2 and 3 (to 500g per m2 for sextets) The rapidincrease in belowground biomass is highly desirable for barerestoration sites To determine whether N crops increased wemeasured biomass and tissue N concentrations The result-ing estimates of accumulated N showed a higher crop for sex-tets than for unplanted and solo plots The rapid accumula-tion of N is beneficial for restoration sites that have coarse soiland where N is the limiting factor (a common problem in cre-ated and restored wetlands Langis et al 1991 Boyer andZedler 1996 1998 Zedler and Callaway 1999 2000 Boyer etal 2000) Our greenhouse experiment revealed similar pat-terns in a more benign environment (that is one with warmertemperatures) Total biomass was higher for sextets becausebelowground biomass increased with species richness As inthe field N crops tracked biomass with greater total N ac-cumulation and greater belowground accumulation at year2A species composition effectmdashnamely the chance inclusionof high biomass producers in the randomly drawn assem-blagemdashis indicated because Jc Fs and Lc solos all accumu-lated more N than the mean for sextets and Sv and Bm
solos both accumulated as much as the triosAlthough Tc Seand Sb solos all had below-average N crops Sb and Tc accu-mulated the most N per unit biomass
These findings were used to design the planting of the 8-ha ldquoFriendship Marshrdquo a $3 million project that aims to sup-port rare species of plants and animals (Figure 8) Located inthe south arm of Tijuana Estuary this site was opened to tidalflushing in February 2000 after floodborne sediments (1ndash2 m)were excavated and the historical marsh plain uncoveredBecause Sv dominates the nearby wetlands and readily es-tablishes on its own it was not introduced as seeds or plantsInstead we planted five species for which we had abundantseedlings (Fs Jc Bm Lc and Se) to help provide the functionsmentioned abovemdashnamely recruitment complex canopiesand biomass and N accumulation The protocol of not plant-ing Sv (to slow time to dominance) and adding multispeciesclusters (to accelerate the development of species-rich
December 2001 Vol 51 No 12 BioScience 1011
Features
Figure 5 San Elijo Lagoon San Diego County California
Figure 6 Historical patterns of species richness atTijuana Estuary data (means and standard errors) arefrom 025-m2 circular plots
canopies) is new to the region Previous marsh plain restora-tion efforts have focused on planting Sv
Although the field and greenhouse experiments taught usthat species-rich plots promote recruitment develop morecomplex canopies and accumulate more biomass and Nthe performance of sextets could be attributable to the com-bined performance of the six species or to the chance inclu-sion of a single high-performance species (a species compo-sition effect) Data from the replicated assemblages in thegreenhouse allowed us to evaluate these alternatives In the mi-crocosms species composition was stable over time (whereasrecruitment increased richness of solo plots in the field) Wecompared the eight species and found that Jc produced themost shoot biomass Fs the most root biomass and Sv themost total biomass while Tc had the highest rootshoot bio-mass But their rankings changed when ash-free biomass
was considered because Sv had the most ash per plant andSb the most ash per unit biomass For ash-free biomass Jc washighest for shoots Fs for roots and Jc for total with Tc stillhaving the highest rootshoot ratio The N concentration ofshoots was highest for Sb for roots it was highest for Tc Jchad the highest crop of N in its shoots and Fs the highest rootN crop while Jc had the highest total N crop Height was great-est for Se and layering was greatest for Fs Interestingly theregionrsquos ldquosuperplantrdquo Sv achieved its first place overall rank-ing not by excelling in multiple attributes but by scoring sec-ond or third in most attributes It scored highest only in to-tal biomass but it achieved high biomass by accumulating alarge volume of salts in its tissues in addition to accumulat-ing considerable carbon
This meta-analysis supports both alternatives Speciescomposition effects (Symstad et al 1998) were indicated forevery attribute considered (there is at least one species thatcan match the performance of the average sextet) but it is notthe same species for all response variables (that is all speciesare needed to achieve high levels of all functions) Further-more each species excelled in at least one attribute Thus weconclude that diversity matters a great deal and the onlyway to ensure that multiple functions are provided is to plantand sustain all species Retaining diversity results in less riskthat functions will be eliminated The planting of species-richassemblages in restoration sites would help ensure that morefunctions are restored
There is support for the suggestion that attributes measuredin the greenhouse translate into performance in the field Thespecies that performed best in the greenhouse are most wide-spread geographically Overall the eight species ranked in vir-tually the same order as in their distributional ranking (thatis the number of southern California wetlands in which theyoccur Table 1) The top three performers in the greenhousewere Sv Jc and Fs and they all occur in 22 or 23 wetlandsLc ranked fourth in greenhouse performance and fourth inregional distribution occurring in 13 wetlands in the historicalrecord and in 11 wetlands in 1998 The others ranked fifththrough eighth in performance they are the most restrictedin their distributions now occurring in only six to eight sitesJc Fs and Sv are all highly productive long-lived perennialsthat reproduce vegetatively and form monotypic patches(Zedler et al 1980 Ferren 1985) It is not surprising thatthey persist in wetlands that experience closure to tidal in-fluence
What functions were lost when short-lived species dropped out of Tijuana Estuary Sb and Se are short-lived halophytes that do not reproducevegetatively Their roles in the ecosystem are arguably uniquefrom these perspectives alone Their annual recruitment ofseedlings following winter rainfall adds seasonality to thecanopy and soil perhaps changing food availability bothabove and below ground Regrettably these functions have notbeen quantified
1012 BioScience December 2001 Vol 51 No 12
Features
Figure 7 Adaptive restoration exemplified by theexperimental planting of 87 2-by-2-meter plots with 0 13 and 6 species in a newly excavated marsh plain atTijuana Estuary Photograph taken in June 1997 2months after planting greenhouse-grown seedlings (90per plot)
As our field data docu-ment Sv Sb and Se are allgood recruiters capable offilling canopy openings withseedlings and rapidly reveg-etating bare sites Howeverthe 1998 recruitment data(Lindig-Cisneros 2001) showthat Sb and Se producedseedlings earlier than Sv Inearly April 1998 there were2199 Sb and 220 Se seedlingsbut only 89 Sv seedlings Inour experimental design wepulled seedlings of invadingspecies to retain assemblagecomposition in 1997ndash1998so there was ample oppor-tunity for Sv seedlings to re-cruit later In early May 1998there were 9261 Sv seedlingsbut only 842 Sb and 50 Se re-cruitment then declined forall species over the late Mayand July sampling dates despite the fact that seedlings werebeing pulled and space remained available for establishmentWe infer from these results that Sb and Se have important rolesin filling gaps that is space opened late in the year is filled bySb and Se the following winter and spring
Both Sb and Se are capable of rapid height growth asdemonstrated in the greenhouse Se might be the first speciesto provide perch sites for the endangered Beldingrsquos Savannahsparrow which defends its nesting territories by singing whileperching on the tallest plants (Powell 1993) The verticalstructure afforded by Se differs from that of Sv in having amore horizontal branching angle and more lay-ers attributes that may increase its attraction ofsmall birds It is not uncommon to see sparrowsperching on tall branches of Se in the fieldwhether the plants are alive or dead We concludethat the loss of Sb and Se could impair the com-munityrsquos ability to recover from disturbanceand to support an endangered bird These func-tions are not entirely eliminated by the demiseof Sb and Se but the ecosystemrsquos functional po-tential might not be realized in their absence
Is the loss of species reversibleTijuana Estuaryrsquos current salt marsh communityis certainly different from that prior to 1984when an open canopy supported thousands ofSb seedlings per square meter and Se was com-mon throughout the tidal marsh Today thesespecies persist in a tiny fraction of the area theyoccupied historicallyAs short-lived species bothSb (annual) and Se (lives only a few years)
depend on recruitment for their persistence while all of theother marsh plain species reproduce vegetatively Seedlingswere young when Tijuana Estuary became nontidal in April1984 and one likely cause of their mortality was the dryingof surface soil and rise in salinity to an average of greater than100 ppt (Figure 9) Moderate hypersalinity (40ndash45 ppt) is typ-ical for marsh plain soils but extreme hypersalinity is morecharacteristic of higher elevations with less frequent tidal in-undation (Noe and Zedler 2001) Sb and Se can germinate inseawater but higher salinities reduce germination rates (Zedlerand Beare 1986) Had their seed banks persisted through the
December 2001 Vol 51 No 12 BioScience 1013
Features
Figure 8 An 8-ha marsh plain excavation at Tijuana Estuary Because of the large size of thissite plantings emphasized species that would not likely recruit on their own
Figure 9 Soil salinity at Tijuana Estuary (data from the Pacific EstuarineResearch Lab) Means before A on the x-axis include Spartina marsh andthe marsh plain means after A are marsh plain only Means before B arefor interstitial soil water after B are for soil pastes (adding deionized waterunderestimates interstitial salinity)
period of tidal closure Sb and Se should have germinated in1985 they did not
A recent reintroduction experiment by Vivian-Smith andZedler (2001) indicated that neither species retains a marshplain seed bank at Tijuana Estuary Vivian-Smith added 2000seeds of each species in 1998 Sb produced 272 seedlings andSe produced 2 with seedlings establishing only where seedswere added Also the current Sv-dominated canopy reducedrecruitment with Sb seedling density twice as high where thecanopy was removed (Table 4) Hence we learned that a lackof seeds and shading by Sv restricted recovery of the Sb pop-ulation And because H N Morzaria-Luna (University ofWisconsinndashMadison unpublished data) could not locate ei-ther species during her search in 2001 we suspect that changesin both the physical and biological conditions of this marshfurther inhibit recovery We explore each of these in turn
Irreversible physical changes According to recent in-formation both elevation and soil salinities have changed inthe 16 years since tidal closure A study of marsh accretion rates(Weis et al forthcoming) using cesium-137 (137Cs a globallydispersed radioisotope) suggests that since 1984 (the yearwhen Sb and Se declined) the marsh plain elevation has risen13 cm based on two soil cores At three additional locationsbenchmarks placed on the marsh plain in the 1970s by the USArmy Corps of Engineers were relocated there we have mea-sured an average of 6 cm accretion since 1984 (Morzaria-Lunaunpublished data) Further sedimentation data from TijuanaEstuary corroborate this rapid rise in elevation Cahoon andcolleagues (1996) measured 20ndash85 (mean 59) cm accretionin the S foliosa marsh between 1992 and 1993 and in anearby tidal pond (lower elevation) Ward (2000) found4ndash127 (mean 88) cm of sediment accretion from 1997 to1998 The marsh plain elevation has risen because of sedi-mentation and the recent record of flooding provides anexplanation
Tijuana Estuary has a large watershed with highly erodablesoils and major slope failures that are visible both north andsouth of the USndashMexico border The 1993 floodwaters cut anentirely new river channel just inland of the estuary Thehuge volume of sediment that was mobilized could havecontributed to a rise in elevation on the marsh plain In-creased soil salinities (now averaging 60 ppt Figure 9) cor-
roborate a shift toward a higher and hence drier marshplain For intertidal species that are sensitive to 5ndash10-cm dif-ferences in elevation the accretion rates and associated salin-ities documented at Tijuana Estuary would seem to be fatalPersistent hypersaline conditions would reduce Sb and Se es-tablishment drier conditions would favor Sv and densercanopies of Sv would reduce the persistence of seedlings
If rising elevation and increasing salinity prevent reestab-lishment of Sb and Se then reversing these changes would re-quire a massive sediment removal effortmdashthe marsh plainwould have to be bulldozed to lower the elevation by 6ndash13 cmor more Such a disruption to the vegetation would not be al-lowed by resource managers because the plant canopy isused by light-footed clapper rails for cover and Beldingrsquos Sa-vannah sparrows for nesting
There is an alternative Although these recent physicalchanges are not reversible it is possible to excavate sedi-ments and restore areas where sediment loads have shiftedtopography well out of the tidal range Exactly this procedureis planned for up to 200 ha of former tidal wetlands in thesouthern arm of the estuary (Entrix et al 1991) The re-moval of sediments in the south arm can conceivably com-pensate for some of the accretion that is occurring in the es-tuary and in the region Attempts are also being made tocontrol the flow of sediments upstream
Irreversible biological changes When the full com-plement of species is present the community presumably hasmaximum biocomplexitymdashthat is a variety of interspecificinteractions and feedback mechanisms Its biodiversity wouldlead to biocomplexity It is possible that the marshrsquos bio-complexity helped sustain its biodiversity If some interactionsfacilitated rare species and others kept dominants from out-competing subordinates then more species would persist Inthat event losing some species would make the communityless able to support all the naturally occurring species bothplant and animalWhere dense canopy cover reduces seedlingrecruitment any species that reduces overall biomass above-ground could reduce shading and enhance recruitment andspecies richness The greenhouse experiments with repli-cated assemblages suggest such a species effect
We explored effects of selected species by comparing mi-crocosms with and without such a species We found threespecies (Sv Fs and Jc) that tended to increase whole-potbiomass (a typical effect of productive species) and one (Tc)that tended to decrease whole-pot biomass (an unusual effect)Assemblages with Tc had lower shoot biomass than assem-blages without it Tc produces little biomass when grownalone it is short with an unimpressive canopy Althoughlow in biomass Tc accumulates a disproportionate concen-tration of N twice that for Sv In the microcosms total ash-free biomass values for Tc and Sv were significantly negativelycorrelated (r = ndash043 p lt 01 n = 72 sextets) Despite itssmall size Tc seems capable of reducing the biomass of SvField data from Bahiacutea San Quintiacuten also suggest a negative re-lationship between Tc and Sv cover (r = ndash0133 p = 05 n =224) We conclude that Tc has some mechanism for reduc-
1014 BioScience December 2001 Vol 51 No 12
Features
Table 4 Experimental attempt to reestablish Sb and Seat Tijuana Estuary in 1998 (unpublished datacollected by Gabrielle Vivian-Smith Pacific EstuarineResearch Lab) Species codes are Sb Salicorniabigelovii Se Suaeda esteroa The ldquoseeds addedrdquotreatment had 2000 seeds added per speciesldquoRecruitmentrdquo is the total number of seedlings found
Treatment Recruitment
No seeds added 0 Sb 0 SeSeeds added canopy left intact 92 Sb 0 SeSeeds added canopy cleared 180 Sb 2 Se
ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features

1006 BioScience December 2001 Vol 51 No 12
Features
what each species potentially contributes to ecosystem func-tioning and develop recommendations for restoring biodi-versity Our experimental tests of the relationship of speciesrichness to ecosystem functioning at Tijuana Estuary con-tribute to the debate about whether ecosystem functioning isinfluenced more by the presence of many species or by thepresence of one or two high-performance species We eval-uate the contribution of multiple and individual species andshow that individual species can match the performance ofsix-species assemblages (sextets) but no single species excelsin all functions Our experimentation with increasing levelsof biodiversity in an actual restoration site (a bare marshplain) provided results of immediate use to biodiversity en-hancement projects while simultaneously vegetating the site
Does diversity matter in restoration Questions about species number species identity and specificfunctions are especially relevant to the restoration of ecosys-tems When confronted with a bare restoration site one be-gins by asking if planting is necessary and if so of which andhow many species The answers may well depend on therestoration targetmdashwhat level of diversity and function is de-sired and when The goal of many restoration projects is tomatch the structural and functional attributes of regional ref-erence ecosystems as quickly as possible Thus any plantingpractice that speeds the development of species-rich vegeta-tion is of interest as are vegetation attributes that support moreanimal species and confer greater resilience to the system
The role of species richness has been explored in several di-versity experiments by planting randomly drawn assem-blages of herbaceous plants and evaluating a range of func-tional attributes most notably biomass accumulation Ingeneral increasing numbers of plant species lead to increasesin productivity (Naeem et al 1994 1995 1996) nutrient re-tention (Ewel et al 1991) resiliency (Tilman and Downing1994) reliability (Johnson et al 1996 Naeem and Li 1997)and decreases in invasibility of other species (Tilman 1997Symstad 2000) In a series of diversity experiments con-ducted in eight European countries Hector and colleagues(1999) reported that a 50 decline in species richness reducedaboveground biomass by 10ndash20 A recent review of thediversity literature confirmed that many functions increase inconcert with a rise in the number of plant species althoughthere was little support for a continually increasing linear re-lationship between species diversity and function (Schwartzet al 2000) In addition diversity effects reach across trophiclevels mycorrhizal diversity affects plant diversity (van der Hei-jden et al 1998) and insect diversity is related to plant diversity(Knops et al 1999) although weakly in one case (Siemann etal 1998)
Although a variety of functions respond positively togreater diversity it remains unclear whether the response isdue to the number of species or the chance inclusion of anoutstanding stress-tolerator colonizer competitor or otherldquosuperplantrdquoTo restore habitat most effectively managers needto know how many and which species to plant so that the veg-
etation will perform multiple functions such as persistencecanopy development and retention of carbon and nutri-ents If the target community has only one species for exampleSpartina alterniflora monotypes along the Atlantic Coastthen restoration is greatly simplified If the reference systemincludes many species a subset of plantings might suffice foraccelerating the development of vegetation To simplifychoices some researchers classify species into functionalgroups based on life span (annual versus perennial) growthform (succulent or nonsucculent upright or trailing grass orforb) or presence of nitrogen-fixing symbionts Experimentswith species drawn randomly from functional groups have un-covered effects on productivity nutrient availability andother functions (Ewel et al 1991 Chapin et al 1992 Silver etal 1996 Hooper and Vitousek 1997 Huston et al 2000)Thus introducing one or more representatives of a func-tional group to a restoration site might suffice for providingcritical ecosystem functionsmdashfor example nitrogen-fixersto accelerate nitrogen accumulation (Crocker and Major1955Vitousek and Reiners 1975) or highly productive speciesto increase overall grassland productivity (Symstad et al1998) Restoration efforts could then be streamlined by lim-iting initial plantings to a few key members of the targetcommunity In southern California salt marshes howeverfunctional groups have proven elusive (Sullivan and Zedler1999)
Californian salt marshesThe Californian salt marsh (Figure 1) is a useful model sys-tem for testing the effect of diversity on ecosystem functionfor several reasons
It includes a small number of species (eight dominatethe marsh plain)
Figure 1 Salt marsh vegetation at Bahiacutea San Quintiacuten BajaCalifornia Norte Mexico
It has naturally occurring assemblages that range fromone to many species
It includes various growth forms (Table 1)
The plants are readily grown from seed and all becomereproductive in their first or second year
As recommended by Vitousek and Hooper (1993 p 14) weldquomanipulate diversity as an experimental treatment and em-phasize the range of diversity from one to ten speciesrdquo Con-cern about experimentation with ldquoimmature vegetationrdquo(Huston et al 2000) is not an issue for restoration sites be-cause the establishment of vegetation on newly graded sub-strate always involves immature plants and any approachesthat can accelerate the maturation process are immediatelyuseful to ecological restoration In other words we seek waysto increase the rate of biomass accumulation and the devel-opment of complex canopies in sites with coarse soils and lim-ited nutrients especially nitrogen (N) We ask whether plant-ing is necessary and if so how many and which species needto be planted to accelerate the development of complexcanopies and the accumulation of soil N
The Californian tidal marsh (Figure 2) has a fringe ofSpartina foliosa a broad marsh plain and a high marsh tran-sition to upland Tides inundate S foliosa twice daily and
the marsh plain at least once daily with water depth varyingby season Tidal amplitudes are narrowest and marsh plaininundation least frequent in March and April about the timevegetation begins to grow exponentially (Winfield 1980)The marsh plain is riddled with tidal creeks and spans a ~30-cm elevation range at our Bahiacutea San Quentiacuten reference sys-tem (Zedler et al 1999) and a ~20-cm range at Tijuana Es-tuary (approximately 60ndash80 cm NGVD [National GeodeticVertical Datum] Figures 2 3 Zedler et al 1992) Marshes inMexicorsquos Bahiacutea San Quintiacuten have large tidal channels with un-interrupted seawater flows (Figure 3)
The regular influence of tides is punctuated by river flood-ing and sea storms over the long term Annual rainfall aver-ages only 25 cm in San Diego and river flows are corre-spondingly low however high interannual variation leads tocatastrophic flooding Wetlands with large watersheds orwith disturbed lands (agricultural fields urbanization) up-stream are subject to major inflows of sediments for exam-ple Mugu Lagoon lost 40 of its low-tide volume during twofloodingndashsedimentation events (Onuf 1987) Such extremesedimentation episodes are attributable to disturbance ofvegetation and soils upstream of the estuaries Tijuana Estu-ary has a large watershed but the Tijuana River did not floodfrom 1944 to 1977 Since then however flows exceeded 70 cu-
bic meters per second during the win-ters of 1978 1980 1983 1986 19881993 1995 and 1997 In addition amajor sea storm overwashed thedunes during the 1983 El Nintildeo eventfilling tidal channels with sand and re-ducing tidal inflows (Zedler et al1992) The estuary mouth subse-quently closed in April 1984
Marshes north of the USndashMexicoborder are still numerous (Figure 4)but their loss of area and habitat qual-ity has caused many wetland-depen-dent species to become threatenedwith extinctionmdashfor example an en-dangered hemiparasitic annual plantknown as salt marsh birdrsquos-beak(Cordylanthus maritimus ssp mari-tumus) the wandering skipper(Panoquina errans) five species oftiger beetle (Cicindella spp) the light-footed clapper rail (Rallus longirostrislevipes) and the Beldingrsquos Savannahsparrow (Passerculus sandwichensisbeldingi) These rare species and sev-eral more abundant wading birds andwaterfowl species are highly valuedby the public But these charismaticspecies are not the only victims ofhabitat loss and degradation Themarsh plain halophytes although notvery showy (Table 1) are still at risk
December 2001 Vol 51 No 12 BioScience 1007
Features
Table 1 Life history and growth forms of eight halophytes common to the marshplain of Californian coastal wetlands Illustrations (by Donovan McIntire) showrelative proportions of biomass above and below ground as well as rooting depthsbased on greenhouse experiments Regional distributions are for 23 wetlands withinthe region (Table 2) fewer occurrences indicate sensitivity to tidal closure Recentdeclines (Table 2) were recorded from selected wetlands (mostly San Diego County)Recruitment data (H=high M=medium L=low) are from Table 3 Species codes areSv Salicornia virginica Fs Frankenia salina Jc Jaumea carnosa Lc Limoniumcalifornicum Se Suaeda esteroa Tc Triglochin concinna Sb Salicornia bigeloviiBm Batis maritima
Spatial and temporal patterns of halophyte di-versity on the marsh plain At Bahiacutea San Quintiacuten themarsh plain supports nine species that are both common andabundant (Zedler et al 1999) The species include the eightspecies that dominated the marsh plain of Tijuana Estuary
before 1984 (Zedler et al 1992) The latter 1000-hasite is San Diego Countyrsquos largest tidal wetland anda National Estuarine Research Reserve
Southern California marshes differ in their bio-diversity Historical records of eight halophyte dis-tributions in 23 coastal salt marshes (Table 2) showthat only three species are very widespread (Sv FsJc species codes are in Table 1) Of these Salicorniavirginica has broad ecological tolerance (as indi-cated by its elevation range Figure 2) and is domi-nant in most of the regionrsquos wetlands (Zedler 1982Barbour and Major 1988) including nontidal sys-tems (Zedler et al 1980) Lc is more restricted thanSv Fs and Jc while the remaining four species areregionally restricted in their distribution especiallythe two short-lived species (Sb and Se) The eightspeciesrsquo regional occurrences indicate that speciesrichness is greatest in wetlands with long histories ofgood tidal flow and lowest in wetlands that havebeen closed to tidal flow for one or more lengthy pe-riods For example Los Pentildeasquitos Lagoon wasclosed for 5 years in the 1950s (Carpelan 1969) itlacks three of the eight marsh plain species San ElijoLagoon (Figure 5) is usually closed to tidal action andit lacks five of the marsh plain halophytes In each ofthese wetlands tidal flows are constrained by botha railroad bridge and at the tidal inlet a coastalhighway
The spatial pattern of fewer species in wetlandswith impaired tidal flow suggests that mouth closure has re-duced biodiversity Two lines of evidence support this cause-and-effect relationship First a 1998 resurvey of San DiegoCountyrsquos tidal marshes documented the loss of species atsites with impaired tidal flows Species Se was lost at four sites
and nearly lost at two additionalsites Lc was lost at two sites and Sbwas lost at one site and nearly lostat Tijuana Estuary (Table 2) Sec-ond the species declines were wit-nessed during two 1984 tidal clo-sure episodes one at Estero dePunta Banda in Baja Californiafollowing the permanent diking ofa large marsh area (Ibarra-Obandoand Poumian-Tapia 1991) and oneat Tijuana Estuary during a tem-porary (8-month) closure (Zedleret al 1992) At Estero de PuntaBanda monthly sampling docu-mented that Sb and Se persistedonly 13 months after a large por-tion of the salt marsh was diked AtTijuana Estuary the mouth closedin April 1984 after the 1983 El Nintildeostorm filled the tidal channels Af-ter closure there was no rainfall
1008 BioScience December 2001 Vol 51 No 12
Features
Table 2 Occurrence of eight marsh-plain halophytes in southern California coastalwetlands Species are ranked according to the number of sites in which they occurwetlands are in order of the number of species they support Data in boxes representchanges in the status of a species at a site determined by G Sullivanrsquos 1998 census ( = now rare 0 = now absent)
Figure 2 Topographic profile of Volcano Marsh at Bahiacutea San Quintiacuten
Transects extend from the upland (left) to the bay (right) crossing tidal
creeks and channels as well as hummocks formed by ancient lava
deposits Elevations are relative to the lowest occurrence of Spartina
foliosa (Zedler et al 1999) Occurrences of three species are given + is a
high-marsh indicator Salicornia subterminalis X is S virginica and
the third row of symbols under transect b is cordgrass S foliosa
in an unusually dry growing season Channel salinities roseto about 60 parts per thousand (ppt) and soil salinities roseto an average of 104 ppt (Zedler et al 1992) Species Sb andSe were virtually extirpated and did not recover from the 8-month nontidal catastrophe (Table 2)
The pattern of Sb and Se loss with reduced tidal influenceis consistent across the region in both space and time and atboth Estero de Punta Banda and Tijuana Estuary The patternis also obvious from monitoring small-scale (025 m2) quadrats(Figure 6) Prior to tidal closure the marsh plain of TijuanaEstuary supported a mean of 45 species during closure themean dropped below 20 Little recovery was evident in sub-sequent years despite the return of tidal influence Our May1999 data from Bahiacutea San Quintiacuten had a mean of 47 speciesper 025-m2 quadrat (n = 224) We conclude that biodiver-sity has declined at Punta Banda and Tijuana estuaries as aresult of interrupted tidal influence
Testing speciesrsquo roles in ecosystem function Given the 90 loss of wetland habitat in southern Californiarestoration is a high priority Early attempts sought only toprovide vegetation cover It was common to plant just one ortwo easily grown species (S foliosa in low elevations Sv on themarsh plain Zedler 2001) We hypothesize that ecosystemfunctioning might be impaired if too few species are re-stored No one knows how many are needed for the systemto develop at maximum speed or to achieve desired levels ofecosystem functions such as the development and mainte-nance of natural community composition the developmentof appropriate canopy structure and the accumulation of or-ganic matter and nitrogen These span the range of indica-tors that Noss (1990) recommends for monitoring biodi-versity The assessment of just one function such as netprimary productivity above ground (or its surrogate struc-tural attribute biomass at the end of the growing season) isnot likely to serve ecosystem managers or settle internationaldebates about the importance of diversity to ecosystem func-tioning Hence we tested the effects of species-rich plantingson the development of multiple functions that are critical inrestoration sites
In 1997 we established a large field experiment at TijuanaEstuary to test the effects of species richness on three specificresponse variables
1 Recruitment Which species need to be planted andwhich readily establish and spread
2 Canopy complexity Which grow taller and producemore layers as might be needed by insects and birds
3 Belowground biomass and N accumulation Whichplots store the most N in roots and rhizomes
Our experimental site was a graded marsh plain thathad room for 87 2 x 2-m plots distributed among fiveblocks along an excavated tidal channel (Figure 7) Our ex-perimental conditions included single-species plots of eachof the eight species of the nearby reference marsh plusrandomly chosen six- and three-species assemblages whichroughly correspond to small-scale richness levels beforeand after tidal closure at Tijuana Estuary (Figure 6) Sev-eral of our experimental assemblages occurred in TijuanaEstuary in 1974 and Bahiacutea San Quintiacuten in 1999 but ran-dom draws missed the most common trio (Bm Sv Sb)and sextet (Bm Sv Sb Fs Jc Se) from Tijuana Estuaryrsquos025-m2 plots Hence we added one plot with Bm Sv Sband one with Bm Sv Sb Fs Jc Se The 87 field experimentalplots were planted in April 1997 and replanted and weededas needed that summer to maintain planting treatments andcompensate for early mortality The assemblages planted re-sulted in similar numbers of plants introduced within theexperimental site Thus differential starting densities werenot of concern in comparing recruitment Assemblagecomposition was maintained during 1998 by removing
December 2001 Vol 51 No 12 BioScience 1009
Features
Figure 3 A remotely sensed image of Volcano Marshshowing tidal channels Mosaic compiled by BruceNyden Pacific Estuarine Research Lab San Diego StateUniversity
seedlings of other species thereafter recruits were allowedto persist
Seedlings were censused by Roberto Lindig-Cisneros(2001) canopy architecture was documented by Keer andZedler (forthcoming) and biomass and N accumulationwere assessed by J Callaway Because we were most con-cerned about the increasing ability of the site to retain Nwe assessed biomass and N accumulation in midwinter af-ter most plants had slowed growth Furthermore becausewe wanted to minimize destructive sampling we removedsmall root cores in year 2 and root and shoot samples at theend of year 3 Both biomass components were analyzed forabove- and belowground N content
We complemented the field study with an experiment inthe greenhouse where we were able to compare individual as-semblages We replicated all 15 randomly drawn trios and sex-
tets 8-fold all solos 16-fold and the common trioand sextet 16-fold for a total of 400 microcosmsPlanting treatments were maintained in the potsthrough vegetative growth and seedling recruit-ment but assemblage composition did not changeAll but 80 pots were grown for 2 years and as-sessed for canopy attributes and harvested for bio-mass and N content above and below ground
The importance of diversity to restorationResults from the field and greenhouse experimentssupport the general hypothesis that diversity mat-ters in restoration sites Our plantings with sixspecies enhanced recruitment made canopiesmore complex increased biomass and increasedN accumulation
Recruitment Only three species (Sv Sb andSe) recruited readily (Table 3) Of these only one(Sv) is widely distributed (Table 2) and available todisperse into restoration sites It germinates in sea-water and produces abundant seedlings in bothnatural and restored wetlands including our un-planted plots it does not need to be planted torestoration sites if a seed source exists nearby Theother two abundant recruiters Sb and Se are themarsh plainrsquos only short-lived species Personalobservation revealed that Se recruitment potentialwas much lower than that of Sb reflecting its lowerdensity in regional wetlands and differences inseedling mortality within this experiment Thefact that both recruited readily in the bare restora-tion site and poorly in the salt marsh where it oc-curred historically is evidence that the Tijuana Es-tuary marsh plain no longer provides suitablehabitat for these species The remaining five speciesdid not readily colonize either our planted or un-planted plots Their inability to recruit may be at-
1010 BioScience December 2001 Vol 51 No 12
Features
Table 3 Seedling recruitment of eight marsh-plainspecies into experimental plots Data are for the 1998growing season Note that the number of plantsintroduced to the site as seedlings in April 1997(column 2) was similar but only three speciesrecruited extensively Species codes are in Table 1
Code Number planted Number of recruits
Sv 840 17703Sb 735 15978Se 795 1668Tc 810 73Jc 840 33Bm 720 26Lc 795 24Fs 765 2
Figure 4 Coastal wetlands of southern California 1Tijuana Estuary 2Sweetwater Marsh 3 Mission Bay Reserve 4 Los Pentildeasquitos Lagoon 5San Dieguito Lagoon 6 San Elijo Lagoon 7 Batiquitos Lagoon 8 AguaHedionda Lagoon 9 Buena Vista Lagoon (brackish wetland) 10 SanLuis Rey River Mouth 11 Santa Margarita Estuary 12 Las FloresMarsh 13 San Mateo Marsh 14 Upper Newport Bay 15 Bolsa ChicaWetland 16 Anaheim Bay 17 Los Cerritos Marsh (mostly filled anddeveloped) 18 Ballona Wetland 19 Malibu Lagoon 20 Mugu Lagoon21 McGrath Lake 22 Santa Clara River Estuary 23 Ventura River(brackish) 24 Carpinteria Marsh 25 Goleta Slough 26 DeverauxLagoon
tributable to small seed sup-ply poor germination or un-suitable conditions for es-tablishment
Canopy architectureKeer and Zedler (forthcom-ing) found that all plotsrapidly achieved high coverbut that canopy height andlayering were slow to de-velop Cover is a poor mea-sure of canopy complexity itdoes not differentiate speciesor species-richness treat-ments Maximum heightswere greatest for trios andsextets reaching about 60cm by year 3 Sv grew thetallest in the first 18 months of study followed by Sb and SeLayering (the number of times a vertical rod encountersplant parts as it is lowered through the canopy) was greaterfor trios and sextets both of which achieved three layerswithin 18 months The effects of individual species werelikely to be seen early in the development of the plots Fs andSe had six and five layers respectively with Jc at four layersLayering is hypothesized to enhance habitat for both arthro-pods and birds These canopies are far from fully developedreference marshes with monotypic Sv averaged 45 cm tall withseven layers and areas with multiple species were 55 cm talland had 13 layers (Keer and Zedler forthcoming)
Biomass and N accumulation In the field plots above-ground biomass averaged under 300 g per m2 for solos andover 400 g per m2 for sextets Belowground biomass also in-creased with species richness and over timemdashmeans doubledbetween years 2 and 3 (to 500g per m2 for sextets) The rapidincrease in belowground biomass is highly desirable for barerestoration sites To determine whether N crops increased wemeasured biomass and tissue N concentrations The result-ing estimates of accumulated N showed a higher crop for sex-tets than for unplanted and solo plots The rapid accumula-tion of N is beneficial for restoration sites that have coarse soiland where N is the limiting factor (a common problem in cre-ated and restored wetlands Langis et al 1991 Boyer andZedler 1996 1998 Zedler and Callaway 1999 2000 Boyer etal 2000) Our greenhouse experiment revealed similar pat-terns in a more benign environment (that is one with warmertemperatures) Total biomass was higher for sextets becausebelowground biomass increased with species richness As inthe field N crops tracked biomass with greater total N ac-cumulation and greater belowground accumulation at year2A species composition effectmdashnamely the chance inclusionof high biomass producers in the randomly drawn assem-blagemdashis indicated because Jc Fs and Lc solos all accumu-lated more N than the mean for sextets and Sv and Bm
solos both accumulated as much as the triosAlthough Tc Seand Sb solos all had below-average N crops Sb and Tc accu-mulated the most N per unit biomass
These findings were used to design the planting of the 8-ha ldquoFriendship Marshrdquo a $3 million project that aims to sup-port rare species of plants and animals (Figure 8) Located inthe south arm of Tijuana Estuary this site was opened to tidalflushing in February 2000 after floodborne sediments (1ndash2 m)were excavated and the historical marsh plain uncoveredBecause Sv dominates the nearby wetlands and readily es-tablishes on its own it was not introduced as seeds or plantsInstead we planted five species for which we had abundantseedlings (Fs Jc Bm Lc and Se) to help provide the functionsmentioned abovemdashnamely recruitment complex canopiesand biomass and N accumulation The protocol of not plant-ing Sv (to slow time to dominance) and adding multispeciesclusters (to accelerate the development of species-rich
December 2001 Vol 51 No 12 BioScience 1011
Features
Figure 5 San Elijo Lagoon San Diego County California
Figure 6 Historical patterns of species richness atTijuana Estuary data (means and standard errors) arefrom 025-m2 circular plots
canopies) is new to the region Previous marsh plain restora-tion efforts have focused on planting Sv
Although the field and greenhouse experiments taught usthat species-rich plots promote recruitment develop morecomplex canopies and accumulate more biomass and Nthe performance of sextets could be attributable to the com-bined performance of the six species or to the chance inclu-sion of a single high-performance species (a species compo-sition effect) Data from the replicated assemblages in thegreenhouse allowed us to evaluate these alternatives In the mi-crocosms species composition was stable over time (whereasrecruitment increased richness of solo plots in the field) Wecompared the eight species and found that Jc produced themost shoot biomass Fs the most root biomass and Sv themost total biomass while Tc had the highest rootshoot bio-mass But their rankings changed when ash-free biomass
was considered because Sv had the most ash per plant andSb the most ash per unit biomass For ash-free biomass Jc washighest for shoots Fs for roots and Jc for total with Tc stillhaving the highest rootshoot ratio The N concentration ofshoots was highest for Sb for roots it was highest for Tc Jchad the highest crop of N in its shoots and Fs the highest rootN crop while Jc had the highest total N crop Height was great-est for Se and layering was greatest for Fs Interestingly theregionrsquos ldquosuperplantrdquo Sv achieved its first place overall rank-ing not by excelling in multiple attributes but by scoring sec-ond or third in most attributes It scored highest only in to-tal biomass but it achieved high biomass by accumulating alarge volume of salts in its tissues in addition to accumulat-ing considerable carbon
This meta-analysis supports both alternatives Speciescomposition effects (Symstad et al 1998) were indicated forevery attribute considered (there is at least one species thatcan match the performance of the average sextet) but it is notthe same species for all response variables (that is all speciesare needed to achieve high levels of all functions) Further-more each species excelled in at least one attribute Thus weconclude that diversity matters a great deal and the onlyway to ensure that multiple functions are provided is to plantand sustain all species Retaining diversity results in less riskthat functions will be eliminated The planting of species-richassemblages in restoration sites would help ensure that morefunctions are restored
There is support for the suggestion that attributes measuredin the greenhouse translate into performance in the field Thespecies that performed best in the greenhouse are most wide-spread geographically Overall the eight species ranked in vir-tually the same order as in their distributional ranking (thatis the number of southern California wetlands in which theyoccur Table 1) The top three performers in the greenhousewere Sv Jc and Fs and they all occur in 22 or 23 wetlandsLc ranked fourth in greenhouse performance and fourth inregional distribution occurring in 13 wetlands in the historicalrecord and in 11 wetlands in 1998 The others ranked fifththrough eighth in performance they are the most restrictedin their distributions now occurring in only six to eight sitesJc Fs and Sv are all highly productive long-lived perennialsthat reproduce vegetatively and form monotypic patches(Zedler et al 1980 Ferren 1985) It is not surprising thatthey persist in wetlands that experience closure to tidal in-fluence
What functions were lost when short-lived species dropped out of Tijuana Estuary Sb and Se are short-lived halophytes that do not reproducevegetatively Their roles in the ecosystem are arguably uniquefrom these perspectives alone Their annual recruitment ofseedlings following winter rainfall adds seasonality to thecanopy and soil perhaps changing food availability bothabove and below ground Regrettably these functions have notbeen quantified
1012 BioScience December 2001 Vol 51 No 12
Features
Figure 7 Adaptive restoration exemplified by theexperimental planting of 87 2-by-2-meter plots with 0 13 and 6 species in a newly excavated marsh plain atTijuana Estuary Photograph taken in June 1997 2months after planting greenhouse-grown seedlings (90per plot)
As our field data docu-ment Sv Sb and Se are allgood recruiters capable offilling canopy openings withseedlings and rapidly reveg-etating bare sites Howeverthe 1998 recruitment data(Lindig-Cisneros 2001) showthat Sb and Se producedseedlings earlier than Sv Inearly April 1998 there were2199 Sb and 220 Se seedlingsbut only 89 Sv seedlings Inour experimental design wepulled seedlings of invadingspecies to retain assemblagecomposition in 1997ndash1998so there was ample oppor-tunity for Sv seedlings to re-cruit later In early May 1998there were 9261 Sv seedlingsbut only 842 Sb and 50 Se re-cruitment then declined forall species over the late Mayand July sampling dates despite the fact that seedlings werebeing pulled and space remained available for establishmentWe infer from these results that Sb and Se have important rolesin filling gaps that is space opened late in the year is filled bySb and Se the following winter and spring
Both Sb and Se are capable of rapid height growth asdemonstrated in the greenhouse Se might be the first speciesto provide perch sites for the endangered Beldingrsquos Savannahsparrow which defends its nesting territories by singing whileperching on the tallest plants (Powell 1993) The verticalstructure afforded by Se differs from that of Sv in having amore horizontal branching angle and more lay-ers attributes that may increase its attraction ofsmall birds It is not uncommon to see sparrowsperching on tall branches of Se in the fieldwhether the plants are alive or dead We concludethat the loss of Sb and Se could impair the com-munityrsquos ability to recover from disturbanceand to support an endangered bird These func-tions are not entirely eliminated by the demiseof Sb and Se but the ecosystemrsquos functional po-tential might not be realized in their absence
Is the loss of species reversibleTijuana Estuaryrsquos current salt marsh communityis certainly different from that prior to 1984when an open canopy supported thousands ofSb seedlings per square meter and Se was com-mon throughout the tidal marsh Today thesespecies persist in a tiny fraction of the area theyoccupied historicallyAs short-lived species bothSb (annual) and Se (lives only a few years)
depend on recruitment for their persistence while all of theother marsh plain species reproduce vegetatively Seedlingswere young when Tijuana Estuary became nontidal in April1984 and one likely cause of their mortality was the dryingof surface soil and rise in salinity to an average of greater than100 ppt (Figure 9) Moderate hypersalinity (40ndash45 ppt) is typ-ical for marsh plain soils but extreme hypersalinity is morecharacteristic of higher elevations with less frequent tidal in-undation (Noe and Zedler 2001) Sb and Se can germinate inseawater but higher salinities reduce germination rates (Zedlerand Beare 1986) Had their seed banks persisted through the
December 2001 Vol 51 No 12 BioScience 1013
Features
Figure 8 An 8-ha marsh plain excavation at Tijuana Estuary Because of the large size of thissite plantings emphasized species that would not likely recruit on their own
Figure 9 Soil salinity at Tijuana Estuary (data from the Pacific EstuarineResearch Lab) Means before A on the x-axis include Spartina marsh andthe marsh plain means after A are marsh plain only Means before B arefor interstitial soil water after B are for soil pastes (adding deionized waterunderestimates interstitial salinity)
period of tidal closure Sb and Se should have germinated in1985 they did not
A recent reintroduction experiment by Vivian-Smith andZedler (2001) indicated that neither species retains a marshplain seed bank at Tijuana Estuary Vivian-Smith added 2000seeds of each species in 1998 Sb produced 272 seedlings andSe produced 2 with seedlings establishing only where seedswere added Also the current Sv-dominated canopy reducedrecruitment with Sb seedling density twice as high where thecanopy was removed (Table 4) Hence we learned that a lackof seeds and shading by Sv restricted recovery of the Sb pop-ulation And because H N Morzaria-Luna (University ofWisconsinndashMadison unpublished data) could not locate ei-ther species during her search in 2001 we suspect that changesin both the physical and biological conditions of this marshfurther inhibit recovery We explore each of these in turn
Irreversible physical changes According to recent in-formation both elevation and soil salinities have changed inthe 16 years since tidal closure A study of marsh accretion rates(Weis et al forthcoming) using cesium-137 (137Cs a globallydispersed radioisotope) suggests that since 1984 (the yearwhen Sb and Se declined) the marsh plain elevation has risen13 cm based on two soil cores At three additional locationsbenchmarks placed on the marsh plain in the 1970s by the USArmy Corps of Engineers were relocated there we have mea-sured an average of 6 cm accretion since 1984 (Morzaria-Lunaunpublished data) Further sedimentation data from TijuanaEstuary corroborate this rapid rise in elevation Cahoon andcolleagues (1996) measured 20ndash85 (mean 59) cm accretionin the S foliosa marsh between 1992 and 1993 and in anearby tidal pond (lower elevation) Ward (2000) found4ndash127 (mean 88) cm of sediment accretion from 1997 to1998 The marsh plain elevation has risen because of sedi-mentation and the recent record of flooding provides anexplanation
Tijuana Estuary has a large watershed with highly erodablesoils and major slope failures that are visible both north andsouth of the USndashMexico border The 1993 floodwaters cut anentirely new river channel just inland of the estuary Thehuge volume of sediment that was mobilized could havecontributed to a rise in elevation on the marsh plain In-creased soil salinities (now averaging 60 ppt Figure 9) cor-
roborate a shift toward a higher and hence drier marshplain For intertidal species that are sensitive to 5ndash10-cm dif-ferences in elevation the accretion rates and associated salin-ities documented at Tijuana Estuary would seem to be fatalPersistent hypersaline conditions would reduce Sb and Se es-tablishment drier conditions would favor Sv and densercanopies of Sv would reduce the persistence of seedlings
If rising elevation and increasing salinity prevent reestab-lishment of Sb and Se then reversing these changes would re-quire a massive sediment removal effortmdashthe marsh plainwould have to be bulldozed to lower the elevation by 6ndash13 cmor more Such a disruption to the vegetation would not be al-lowed by resource managers because the plant canopy isused by light-footed clapper rails for cover and Beldingrsquos Sa-vannah sparrows for nesting
There is an alternative Although these recent physicalchanges are not reversible it is possible to excavate sedi-ments and restore areas where sediment loads have shiftedtopography well out of the tidal range Exactly this procedureis planned for up to 200 ha of former tidal wetlands in thesouthern arm of the estuary (Entrix et al 1991) The re-moval of sediments in the south arm can conceivably com-pensate for some of the accretion that is occurring in the es-tuary and in the region Attempts are also being made tocontrol the flow of sediments upstream
Irreversible biological changes When the full com-plement of species is present the community presumably hasmaximum biocomplexitymdashthat is a variety of interspecificinteractions and feedback mechanisms Its biodiversity wouldlead to biocomplexity It is possible that the marshrsquos bio-complexity helped sustain its biodiversity If some interactionsfacilitated rare species and others kept dominants from out-competing subordinates then more species would persist Inthat event losing some species would make the communityless able to support all the naturally occurring species bothplant and animalWhere dense canopy cover reduces seedlingrecruitment any species that reduces overall biomass above-ground could reduce shading and enhance recruitment andspecies richness The greenhouse experiments with repli-cated assemblages suggest such a species effect
We explored effects of selected species by comparing mi-crocosms with and without such a species We found threespecies (Sv Fs and Jc) that tended to increase whole-potbiomass (a typical effect of productive species) and one (Tc)that tended to decrease whole-pot biomass (an unusual effect)Assemblages with Tc had lower shoot biomass than assem-blages without it Tc produces little biomass when grownalone it is short with an unimpressive canopy Althoughlow in biomass Tc accumulates a disproportionate concen-tration of N twice that for Sv In the microcosms total ash-free biomass values for Tc and Sv were significantly negativelycorrelated (r = ndash043 p lt 01 n = 72 sextets) Despite itssmall size Tc seems capable of reducing the biomass of SvField data from Bahiacutea San Quintiacuten also suggest a negative re-lationship between Tc and Sv cover (r = ndash0133 p = 05 n =224) We conclude that Tc has some mechanism for reduc-
1014 BioScience December 2001 Vol 51 No 12
Features
Table 4 Experimental attempt to reestablish Sb and Seat Tijuana Estuary in 1998 (unpublished datacollected by Gabrielle Vivian-Smith Pacific EstuarineResearch Lab) Species codes are Sb Salicorniabigelovii Se Suaeda esteroa The ldquoseeds addedrdquotreatment had 2000 seeds added per speciesldquoRecruitmentrdquo is the total number of seedlings found
Treatment Recruitment
No seeds added 0 Sb 0 SeSeeds added canopy left intact 92 Sb 0 SeSeeds added canopy cleared 180 Sb 2 Se
ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features

It has naturally occurring assemblages that range fromone to many species
It includes various growth forms (Table 1)
The plants are readily grown from seed and all becomereproductive in their first or second year
As recommended by Vitousek and Hooper (1993 p 14) weldquomanipulate diversity as an experimental treatment and em-phasize the range of diversity from one to ten speciesrdquo Con-cern about experimentation with ldquoimmature vegetationrdquo(Huston et al 2000) is not an issue for restoration sites be-cause the establishment of vegetation on newly graded sub-strate always involves immature plants and any approachesthat can accelerate the maturation process are immediatelyuseful to ecological restoration In other words we seek waysto increase the rate of biomass accumulation and the devel-opment of complex canopies in sites with coarse soils and lim-ited nutrients especially nitrogen (N) We ask whether plant-ing is necessary and if so how many and which species needto be planted to accelerate the development of complexcanopies and the accumulation of soil N
The Californian tidal marsh (Figure 2) has a fringe ofSpartina foliosa a broad marsh plain and a high marsh tran-sition to upland Tides inundate S foliosa twice daily and
the marsh plain at least once daily with water depth varyingby season Tidal amplitudes are narrowest and marsh plaininundation least frequent in March and April about the timevegetation begins to grow exponentially (Winfield 1980)The marsh plain is riddled with tidal creeks and spans a ~30-cm elevation range at our Bahiacutea San Quentiacuten reference sys-tem (Zedler et al 1999) and a ~20-cm range at Tijuana Es-tuary (approximately 60ndash80 cm NGVD [National GeodeticVertical Datum] Figures 2 3 Zedler et al 1992) Marshes inMexicorsquos Bahiacutea San Quintiacuten have large tidal channels with un-interrupted seawater flows (Figure 3)
The regular influence of tides is punctuated by river flood-ing and sea storms over the long term Annual rainfall aver-ages only 25 cm in San Diego and river flows are corre-spondingly low however high interannual variation leads tocatastrophic flooding Wetlands with large watersheds orwith disturbed lands (agricultural fields urbanization) up-stream are subject to major inflows of sediments for exam-ple Mugu Lagoon lost 40 of its low-tide volume during twofloodingndashsedimentation events (Onuf 1987) Such extremesedimentation episodes are attributable to disturbance ofvegetation and soils upstream of the estuaries Tijuana Estu-ary has a large watershed but the Tijuana River did not floodfrom 1944 to 1977 Since then however flows exceeded 70 cu-
bic meters per second during the win-ters of 1978 1980 1983 1986 19881993 1995 and 1997 In addition amajor sea storm overwashed thedunes during the 1983 El Nintildeo eventfilling tidal channels with sand and re-ducing tidal inflows (Zedler et al1992) The estuary mouth subse-quently closed in April 1984
Marshes north of the USndashMexicoborder are still numerous (Figure 4)but their loss of area and habitat qual-ity has caused many wetland-depen-dent species to become threatenedwith extinctionmdashfor example an en-dangered hemiparasitic annual plantknown as salt marsh birdrsquos-beak(Cordylanthus maritimus ssp mari-tumus) the wandering skipper(Panoquina errans) five species oftiger beetle (Cicindella spp) the light-footed clapper rail (Rallus longirostrislevipes) and the Beldingrsquos Savannahsparrow (Passerculus sandwichensisbeldingi) These rare species and sev-eral more abundant wading birds andwaterfowl species are highly valuedby the public But these charismaticspecies are not the only victims ofhabitat loss and degradation Themarsh plain halophytes although notvery showy (Table 1) are still at risk
December 2001 Vol 51 No 12 BioScience 1007
Features
Table 1 Life history and growth forms of eight halophytes common to the marshplain of Californian coastal wetlands Illustrations (by Donovan McIntire) showrelative proportions of biomass above and below ground as well as rooting depthsbased on greenhouse experiments Regional distributions are for 23 wetlands withinthe region (Table 2) fewer occurrences indicate sensitivity to tidal closure Recentdeclines (Table 2) were recorded from selected wetlands (mostly San Diego County)Recruitment data (H=high M=medium L=low) are from Table 3 Species codes areSv Salicornia virginica Fs Frankenia salina Jc Jaumea carnosa Lc Limoniumcalifornicum Se Suaeda esteroa Tc Triglochin concinna Sb Salicornia bigeloviiBm Batis maritima
Spatial and temporal patterns of halophyte di-versity on the marsh plain At Bahiacutea San Quintiacuten themarsh plain supports nine species that are both common andabundant (Zedler et al 1999) The species include the eightspecies that dominated the marsh plain of Tijuana Estuary
before 1984 (Zedler et al 1992) The latter 1000-hasite is San Diego Countyrsquos largest tidal wetland anda National Estuarine Research Reserve
Southern California marshes differ in their bio-diversity Historical records of eight halophyte dis-tributions in 23 coastal salt marshes (Table 2) showthat only three species are very widespread (Sv FsJc species codes are in Table 1) Of these Salicorniavirginica has broad ecological tolerance (as indi-cated by its elevation range Figure 2) and is domi-nant in most of the regionrsquos wetlands (Zedler 1982Barbour and Major 1988) including nontidal sys-tems (Zedler et al 1980) Lc is more restricted thanSv Fs and Jc while the remaining four species areregionally restricted in their distribution especiallythe two short-lived species (Sb and Se) The eightspeciesrsquo regional occurrences indicate that speciesrichness is greatest in wetlands with long histories ofgood tidal flow and lowest in wetlands that havebeen closed to tidal flow for one or more lengthy pe-riods For example Los Pentildeasquitos Lagoon wasclosed for 5 years in the 1950s (Carpelan 1969) itlacks three of the eight marsh plain species San ElijoLagoon (Figure 5) is usually closed to tidal action andit lacks five of the marsh plain halophytes In each ofthese wetlands tidal flows are constrained by botha railroad bridge and at the tidal inlet a coastalhighway
The spatial pattern of fewer species in wetlandswith impaired tidal flow suggests that mouth closure has re-duced biodiversity Two lines of evidence support this cause-and-effect relationship First a 1998 resurvey of San DiegoCountyrsquos tidal marshes documented the loss of species atsites with impaired tidal flows Species Se was lost at four sites
and nearly lost at two additionalsites Lc was lost at two sites and Sbwas lost at one site and nearly lostat Tijuana Estuary (Table 2) Sec-ond the species declines were wit-nessed during two 1984 tidal clo-sure episodes one at Estero dePunta Banda in Baja Californiafollowing the permanent diking ofa large marsh area (Ibarra-Obandoand Poumian-Tapia 1991) and oneat Tijuana Estuary during a tem-porary (8-month) closure (Zedleret al 1992) At Estero de PuntaBanda monthly sampling docu-mented that Sb and Se persistedonly 13 months after a large por-tion of the salt marsh was diked AtTijuana Estuary the mouth closedin April 1984 after the 1983 El Nintildeostorm filled the tidal channels Af-ter closure there was no rainfall
1008 BioScience December 2001 Vol 51 No 12
Features
Table 2 Occurrence of eight marsh-plain halophytes in southern California coastalwetlands Species are ranked according to the number of sites in which they occurwetlands are in order of the number of species they support Data in boxes representchanges in the status of a species at a site determined by G Sullivanrsquos 1998 census ( = now rare 0 = now absent)
Figure 2 Topographic profile of Volcano Marsh at Bahiacutea San Quintiacuten
Transects extend from the upland (left) to the bay (right) crossing tidal
creeks and channels as well as hummocks formed by ancient lava
deposits Elevations are relative to the lowest occurrence of Spartina
foliosa (Zedler et al 1999) Occurrences of three species are given + is a
high-marsh indicator Salicornia subterminalis X is S virginica and
the third row of symbols under transect b is cordgrass S foliosa
in an unusually dry growing season Channel salinities roseto about 60 parts per thousand (ppt) and soil salinities roseto an average of 104 ppt (Zedler et al 1992) Species Sb andSe were virtually extirpated and did not recover from the 8-month nontidal catastrophe (Table 2)
The pattern of Sb and Se loss with reduced tidal influenceis consistent across the region in both space and time and atboth Estero de Punta Banda and Tijuana Estuary The patternis also obvious from monitoring small-scale (025 m2) quadrats(Figure 6) Prior to tidal closure the marsh plain of TijuanaEstuary supported a mean of 45 species during closure themean dropped below 20 Little recovery was evident in sub-sequent years despite the return of tidal influence Our May1999 data from Bahiacutea San Quintiacuten had a mean of 47 speciesper 025-m2 quadrat (n = 224) We conclude that biodiver-sity has declined at Punta Banda and Tijuana estuaries as aresult of interrupted tidal influence
Testing speciesrsquo roles in ecosystem function Given the 90 loss of wetland habitat in southern Californiarestoration is a high priority Early attempts sought only toprovide vegetation cover It was common to plant just one ortwo easily grown species (S foliosa in low elevations Sv on themarsh plain Zedler 2001) We hypothesize that ecosystemfunctioning might be impaired if too few species are re-stored No one knows how many are needed for the systemto develop at maximum speed or to achieve desired levels ofecosystem functions such as the development and mainte-nance of natural community composition the developmentof appropriate canopy structure and the accumulation of or-ganic matter and nitrogen These span the range of indica-tors that Noss (1990) recommends for monitoring biodi-versity The assessment of just one function such as netprimary productivity above ground (or its surrogate struc-tural attribute biomass at the end of the growing season) isnot likely to serve ecosystem managers or settle internationaldebates about the importance of diversity to ecosystem func-tioning Hence we tested the effects of species-rich plantingson the development of multiple functions that are critical inrestoration sites
In 1997 we established a large field experiment at TijuanaEstuary to test the effects of species richness on three specificresponse variables
1 Recruitment Which species need to be planted andwhich readily establish and spread
2 Canopy complexity Which grow taller and producemore layers as might be needed by insects and birds
3 Belowground biomass and N accumulation Whichplots store the most N in roots and rhizomes
Our experimental site was a graded marsh plain thathad room for 87 2 x 2-m plots distributed among fiveblocks along an excavated tidal channel (Figure 7) Our ex-perimental conditions included single-species plots of eachof the eight species of the nearby reference marsh plusrandomly chosen six- and three-species assemblages whichroughly correspond to small-scale richness levels beforeand after tidal closure at Tijuana Estuary (Figure 6) Sev-eral of our experimental assemblages occurred in TijuanaEstuary in 1974 and Bahiacutea San Quintiacuten in 1999 but ran-dom draws missed the most common trio (Bm Sv Sb)and sextet (Bm Sv Sb Fs Jc Se) from Tijuana Estuaryrsquos025-m2 plots Hence we added one plot with Bm Sv Sband one with Bm Sv Sb Fs Jc Se The 87 field experimentalplots were planted in April 1997 and replanted and weededas needed that summer to maintain planting treatments andcompensate for early mortality The assemblages planted re-sulted in similar numbers of plants introduced within theexperimental site Thus differential starting densities werenot of concern in comparing recruitment Assemblagecomposition was maintained during 1998 by removing
December 2001 Vol 51 No 12 BioScience 1009
Features
Figure 3 A remotely sensed image of Volcano Marshshowing tidal channels Mosaic compiled by BruceNyden Pacific Estuarine Research Lab San Diego StateUniversity
seedlings of other species thereafter recruits were allowedto persist
Seedlings were censused by Roberto Lindig-Cisneros(2001) canopy architecture was documented by Keer andZedler (forthcoming) and biomass and N accumulationwere assessed by J Callaway Because we were most con-cerned about the increasing ability of the site to retain Nwe assessed biomass and N accumulation in midwinter af-ter most plants had slowed growth Furthermore becausewe wanted to minimize destructive sampling we removedsmall root cores in year 2 and root and shoot samples at theend of year 3 Both biomass components were analyzed forabove- and belowground N content
We complemented the field study with an experiment inthe greenhouse where we were able to compare individual as-semblages We replicated all 15 randomly drawn trios and sex-
tets 8-fold all solos 16-fold and the common trioand sextet 16-fold for a total of 400 microcosmsPlanting treatments were maintained in the potsthrough vegetative growth and seedling recruit-ment but assemblage composition did not changeAll but 80 pots were grown for 2 years and as-sessed for canopy attributes and harvested for bio-mass and N content above and below ground
The importance of diversity to restorationResults from the field and greenhouse experimentssupport the general hypothesis that diversity mat-ters in restoration sites Our plantings with sixspecies enhanced recruitment made canopiesmore complex increased biomass and increasedN accumulation
Recruitment Only three species (Sv Sb andSe) recruited readily (Table 3) Of these only one(Sv) is widely distributed (Table 2) and available todisperse into restoration sites It germinates in sea-water and produces abundant seedlings in bothnatural and restored wetlands including our un-planted plots it does not need to be planted torestoration sites if a seed source exists nearby Theother two abundant recruiters Sb and Se are themarsh plainrsquos only short-lived species Personalobservation revealed that Se recruitment potentialwas much lower than that of Sb reflecting its lowerdensity in regional wetlands and differences inseedling mortality within this experiment Thefact that both recruited readily in the bare restora-tion site and poorly in the salt marsh where it oc-curred historically is evidence that the Tijuana Es-tuary marsh plain no longer provides suitablehabitat for these species The remaining five speciesdid not readily colonize either our planted or un-planted plots Their inability to recruit may be at-
1010 BioScience December 2001 Vol 51 No 12
Features
Table 3 Seedling recruitment of eight marsh-plainspecies into experimental plots Data are for the 1998growing season Note that the number of plantsintroduced to the site as seedlings in April 1997(column 2) was similar but only three speciesrecruited extensively Species codes are in Table 1
Code Number planted Number of recruits
Sv 840 17703Sb 735 15978Se 795 1668Tc 810 73Jc 840 33Bm 720 26Lc 795 24Fs 765 2
Figure 4 Coastal wetlands of southern California 1Tijuana Estuary 2Sweetwater Marsh 3 Mission Bay Reserve 4 Los Pentildeasquitos Lagoon 5San Dieguito Lagoon 6 San Elijo Lagoon 7 Batiquitos Lagoon 8 AguaHedionda Lagoon 9 Buena Vista Lagoon (brackish wetland) 10 SanLuis Rey River Mouth 11 Santa Margarita Estuary 12 Las FloresMarsh 13 San Mateo Marsh 14 Upper Newport Bay 15 Bolsa ChicaWetland 16 Anaheim Bay 17 Los Cerritos Marsh (mostly filled anddeveloped) 18 Ballona Wetland 19 Malibu Lagoon 20 Mugu Lagoon21 McGrath Lake 22 Santa Clara River Estuary 23 Ventura River(brackish) 24 Carpinteria Marsh 25 Goleta Slough 26 DeverauxLagoon
tributable to small seed sup-ply poor germination or un-suitable conditions for es-tablishment
Canopy architectureKeer and Zedler (forthcom-ing) found that all plotsrapidly achieved high coverbut that canopy height andlayering were slow to de-velop Cover is a poor mea-sure of canopy complexity itdoes not differentiate speciesor species-richness treat-ments Maximum heightswere greatest for trios andsextets reaching about 60cm by year 3 Sv grew thetallest in the first 18 months of study followed by Sb and SeLayering (the number of times a vertical rod encountersplant parts as it is lowered through the canopy) was greaterfor trios and sextets both of which achieved three layerswithin 18 months The effects of individual species werelikely to be seen early in the development of the plots Fs andSe had six and five layers respectively with Jc at four layersLayering is hypothesized to enhance habitat for both arthro-pods and birds These canopies are far from fully developedreference marshes with monotypic Sv averaged 45 cm tall withseven layers and areas with multiple species were 55 cm talland had 13 layers (Keer and Zedler forthcoming)
Biomass and N accumulation In the field plots above-ground biomass averaged under 300 g per m2 for solos andover 400 g per m2 for sextets Belowground biomass also in-creased with species richness and over timemdashmeans doubledbetween years 2 and 3 (to 500g per m2 for sextets) The rapidincrease in belowground biomass is highly desirable for barerestoration sites To determine whether N crops increased wemeasured biomass and tissue N concentrations The result-ing estimates of accumulated N showed a higher crop for sex-tets than for unplanted and solo plots The rapid accumula-tion of N is beneficial for restoration sites that have coarse soiland where N is the limiting factor (a common problem in cre-ated and restored wetlands Langis et al 1991 Boyer andZedler 1996 1998 Zedler and Callaway 1999 2000 Boyer etal 2000) Our greenhouse experiment revealed similar pat-terns in a more benign environment (that is one with warmertemperatures) Total biomass was higher for sextets becausebelowground biomass increased with species richness As inthe field N crops tracked biomass with greater total N ac-cumulation and greater belowground accumulation at year2A species composition effectmdashnamely the chance inclusionof high biomass producers in the randomly drawn assem-blagemdashis indicated because Jc Fs and Lc solos all accumu-lated more N than the mean for sextets and Sv and Bm
solos both accumulated as much as the triosAlthough Tc Seand Sb solos all had below-average N crops Sb and Tc accu-mulated the most N per unit biomass
These findings were used to design the planting of the 8-ha ldquoFriendship Marshrdquo a $3 million project that aims to sup-port rare species of plants and animals (Figure 8) Located inthe south arm of Tijuana Estuary this site was opened to tidalflushing in February 2000 after floodborne sediments (1ndash2 m)were excavated and the historical marsh plain uncoveredBecause Sv dominates the nearby wetlands and readily es-tablishes on its own it was not introduced as seeds or plantsInstead we planted five species for which we had abundantseedlings (Fs Jc Bm Lc and Se) to help provide the functionsmentioned abovemdashnamely recruitment complex canopiesand biomass and N accumulation The protocol of not plant-ing Sv (to slow time to dominance) and adding multispeciesclusters (to accelerate the development of species-rich
December 2001 Vol 51 No 12 BioScience 1011
Features
Figure 5 San Elijo Lagoon San Diego County California
Figure 6 Historical patterns of species richness atTijuana Estuary data (means and standard errors) arefrom 025-m2 circular plots
canopies) is new to the region Previous marsh plain restora-tion efforts have focused on planting Sv
Although the field and greenhouse experiments taught usthat species-rich plots promote recruitment develop morecomplex canopies and accumulate more biomass and Nthe performance of sextets could be attributable to the com-bined performance of the six species or to the chance inclu-sion of a single high-performance species (a species compo-sition effect) Data from the replicated assemblages in thegreenhouse allowed us to evaluate these alternatives In the mi-crocosms species composition was stable over time (whereasrecruitment increased richness of solo plots in the field) Wecompared the eight species and found that Jc produced themost shoot biomass Fs the most root biomass and Sv themost total biomass while Tc had the highest rootshoot bio-mass But their rankings changed when ash-free biomass
was considered because Sv had the most ash per plant andSb the most ash per unit biomass For ash-free biomass Jc washighest for shoots Fs for roots and Jc for total with Tc stillhaving the highest rootshoot ratio The N concentration ofshoots was highest for Sb for roots it was highest for Tc Jchad the highest crop of N in its shoots and Fs the highest rootN crop while Jc had the highest total N crop Height was great-est for Se and layering was greatest for Fs Interestingly theregionrsquos ldquosuperplantrdquo Sv achieved its first place overall rank-ing not by excelling in multiple attributes but by scoring sec-ond or third in most attributes It scored highest only in to-tal biomass but it achieved high biomass by accumulating alarge volume of salts in its tissues in addition to accumulat-ing considerable carbon
This meta-analysis supports both alternatives Speciescomposition effects (Symstad et al 1998) were indicated forevery attribute considered (there is at least one species thatcan match the performance of the average sextet) but it is notthe same species for all response variables (that is all speciesare needed to achieve high levels of all functions) Further-more each species excelled in at least one attribute Thus weconclude that diversity matters a great deal and the onlyway to ensure that multiple functions are provided is to plantand sustain all species Retaining diversity results in less riskthat functions will be eliminated The planting of species-richassemblages in restoration sites would help ensure that morefunctions are restored
There is support for the suggestion that attributes measuredin the greenhouse translate into performance in the field Thespecies that performed best in the greenhouse are most wide-spread geographically Overall the eight species ranked in vir-tually the same order as in their distributional ranking (thatis the number of southern California wetlands in which theyoccur Table 1) The top three performers in the greenhousewere Sv Jc and Fs and they all occur in 22 or 23 wetlandsLc ranked fourth in greenhouse performance and fourth inregional distribution occurring in 13 wetlands in the historicalrecord and in 11 wetlands in 1998 The others ranked fifththrough eighth in performance they are the most restrictedin their distributions now occurring in only six to eight sitesJc Fs and Sv are all highly productive long-lived perennialsthat reproduce vegetatively and form monotypic patches(Zedler et al 1980 Ferren 1985) It is not surprising thatthey persist in wetlands that experience closure to tidal in-fluence
What functions were lost when short-lived species dropped out of Tijuana Estuary Sb and Se are short-lived halophytes that do not reproducevegetatively Their roles in the ecosystem are arguably uniquefrom these perspectives alone Their annual recruitment ofseedlings following winter rainfall adds seasonality to thecanopy and soil perhaps changing food availability bothabove and below ground Regrettably these functions have notbeen quantified
1012 BioScience December 2001 Vol 51 No 12
Features
Figure 7 Adaptive restoration exemplified by theexperimental planting of 87 2-by-2-meter plots with 0 13 and 6 species in a newly excavated marsh plain atTijuana Estuary Photograph taken in June 1997 2months after planting greenhouse-grown seedlings (90per plot)
As our field data docu-ment Sv Sb and Se are allgood recruiters capable offilling canopy openings withseedlings and rapidly reveg-etating bare sites Howeverthe 1998 recruitment data(Lindig-Cisneros 2001) showthat Sb and Se producedseedlings earlier than Sv Inearly April 1998 there were2199 Sb and 220 Se seedlingsbut only 89 Sv seedlings Inour experimental design wepulled seedlings of invadingspecies to retain assemblagecomposition in 1997ndash1998so there was ample oppor-tunity for Sv seedlings to re-cruit later In early May 1998there were 9261 Sv seedlingsbut only 842 Sb and 50 Se re-cruitment then declined forall species over the late Mayand July sampling dates despite the fact that seedlings werebeing pulled and space remained available for establishmentWe infer from these results that Sb and Se have important rolesin filling gaps that is space opened late in the year is filled bySb and Se the following winter and spring
Both Sb and Se are capable of rapid height growth asdemonstrated in the greenhouse Se might be the first speciesto provide perch sites for the endangered Beldingrsquos Savannahsparrow which defends its nesting territories by singing whileperching on the tallest plants (Powell 1993) The verticalstructure afforded by Se differs from that of Sv in having amore horizontal branching angle and more lay-ers attributes that may increase its attraction ofsmall birds It is not uncommon to see sparrowsperching on tall branches of Se in the fieldwhether the plants are alive or dead We concludethat the loss of Sb and Se could impair the com-munityrsquos ability to recover from disturbanceand to support an endangered bird These func-tions are not entirely eliminated by the demiseof Sb and Se but the ecosystemrsquos functional po-tential might not be realized in their absence
Is the loss of species reversibleTijuana Estuaryrsquos current salt marsh communityis certainly different from that prior to 1984when an open canopy supported thousands ofSb seedlings per square meter and Se was com-mon throughout the tidal marsh Today thesespecies persist in a tiny fraction of the area theyoccupied historicallyAs short-lived species bothSb (annual) and Se (lives only a few years)
depend on recruitment for their persistence while all of theother marsh plain species reproduce vegetatively Seedlingswere young when Tijuana Estuary became nontidal in April1984 and one likely cause of their mortality was the dryingof surface soil and rise in salinity to an average of greater than100 ppt (Figure 9) Moderate hypersalinity (40ndash45 ppt) is typ-ical for marsh plain soils but extreme hypersalinity is morecharacteristic of higher elevations with less frequent tidal in-undation (Noe and Zedler 2001) Sb and Se can germinate inseawater but higher salinities reduce germination rates (Zedlerand Beare 1986) Had their seed banks persisted through the
December 2001 Vol 51 No 12 BioScience 1013
Features
Figure 8 An 8-ha marsh plain excavation at Tijuana Estuary Because of the large size of thissite plantings emphasized species that would not likely recruit on their own
Figure 9 Soil salinity at Tijuana Estuary (data from the Pacific EstuarineResearch Lab) Means before A on the x-axis include Spartina marsh andthe marsh plain means after A are marsh plain only Means before B arefor interstitial soil water after B are for soil pastes (adding deionized waterunderestimates interstitial salinity)
period of tidal closure Sb and Se should have germinated in1985 they did not
A recent reintroduction experiment by Vivian-Smith andZedler (2001) indicated that neither species retains a marshplain seed bank at Tijuana Estuary Vivian-Smith added 2000seeds of each species in 1998 Sb produced 272 seedlings andSe produced 2 with seedlings establishing only where seedswere added Also the current Sv-dominated canopy reducedrecruitment with Sb seedling density twice as high where thecanopy was removed (Table 4) Hence we learned that a lackof seeds and shading by Sv restricted recovery of the Sb pop-ulation And because H N Morzaria-Luna (University ofWisconsinndashMadison unpublished data) could not locate ei-ther species during her search in 2001 we suspect that changesin both the physical and biological conditions of this marshfurther inhibit recovery We explore each of these in turn
Irreversible physical changes According to recent in-formation both elevation and soil salinities have changed inthe 16 years since tidal closure A study of marsh accretion rates(Weis et al forthcoming) using cesium-137 (137Cs a globallydispersed radioisotope) suggests that since 1984 (the yearwhen Sb and Se declined) the marsh plain elevation has risen13 cm based on two soil cores At three additional locationsbenchmarks placed on the marsh plain in the 1970s by the USArmy Corps of Engineers were relocated there we have mea-sured an average of 6 cm accretion since 1984 (Morzaria-Lunaunpublished data) Further sedimentation data from TijuanaEstuary corroborate this rapid rise in elevation Cahoon andcolleagues (1996) measured 20ndash85 (mean 59) cm accretionin the S foliosa marsh between 1992 and 1993 and in anearby tidal pond (lower elevation) Ward (2000) found4ndash127 (mean 88) cm of sediment accretion from 1997 to1998 The marsh plain elevation has risen because of sedi-mentation and the recent record of flooding provides anexplanation
Tijuana Estuary has a large watershed with highly erodablesoils and major slope failures that are visible both north andsouth of the USndashMexico border The 1993 floodwaters cut anentirely new river channel just inland of the estuary Thehuge volume of sediment that was mobilized could havecontributed to a rise in elevation on the marsh plain In-creased soil salinities (now averaging 60 ppt Figure 9) cor-
roborate a shift toward a higher and hence drier marshplain For intertidal species that are sensitive to 5ndash10-cm dif-ferences in elevation the accretion rates and associated salin-ities documented at Tijuana Estuary would seem to be fatalPersistent hypersaline conditions would reduce Sb and Se es-tablishment drier conditions would favor Sv and densercanopies of Sv would reduce the persistence of seedlings
If rising elevation and increasing salinity prevent reestab-lishment of Sb and Se then reversing these changes would re-quire a massive sediment removal effortmdashthe marsh plainwould have to be bulldozed to lower the elevation by 6ndash13 cmor more Such a disruption to the vegetation would not be al-lowed by resource managers because the plant canopy isused by light-footed clapper rails for cover and Beldingrsquos Sa-vannah sparrows for nesting
There is an alternative Although these recent physicalchanges are not reversible it is possible to excavate sedi-ments and restore areas where sediment loads have shiftedtopography well out of the tidal range Exactly this procedureis planned for up to 200 ha of former tidal wetlands in thesouthern arm of the estuary (Entrix et al 1991) The re-moval of sediments in the south arm can conceivably com-pensate for some of the accretion that is occurring in the es-tuary and in the region Attempts are also being made tocontrol the flow of sediments upstream
Irreversible biological changes When the full com-plement of species is present the community presumably hasmaximum biocomplexitymdashthat is a variety of interspecificinteractions and feedback mechanisms Its biodiversity wouldlead to biocomplexity It is possible that the marshrsquos bio-complexity helped sustain its biodiversity If some interactionsfacilitated rare species and others kept dominants from out-competing subordinates then more species would persist Inthat event losing some species would make the communityless able to support all the naturally occurring species bothplant and animalWhere dense canopy cover reduces seedlingrecruitment any species that reduces overall biomass above-ground could reduce shading and enhance recruitment andspecies richness The greenhouse experiments with repli-cated assemblages suggest such a species effect
We explored effects of selected species by comparing mi-crocosms with and without such a species We found threespecies (Sv Fs and Jc) that tended to increase whole-potbiomass (a typical effect of productive species) and one (Tc)that tended to decrease whole-pot biomass (an unusual effect)Assemblages with Tc had lower shoot biomass than assem-blages without it Tc produces little biomass when grownalone it is short with an unimpressive canopy Althoughlow in biomass Tc accumulates a disproportionate concen-tration of N twice that for Sv In the microcosms total ash-free biomass values for Tc and Sv were significantly negativelycorrelated (r = ndash043 p lt 01 n = 72 sextets) Despite itssmall size Tc seems capable of reducing the biomass of SvField data from Bahiacutea San Quintiacuten also suggest a negative re-lationship between Tc and Sv cover (r = ndash0133 p = 05 n =224) We conclude that Tc has some mechanism for reduc-
1014 BioScience December 2001 Vol 51 No 12
Features
Table 4 Experimental attempt to reestablish Sb and Seat Tijuana Estuary in 1998 (unpublished datacollected by Gabrielle Vivian-Smith Pacific EstuarineResearch Lab) Species codes are Sb Salicorniabigelovii Se Suaeda esteroa The ldquoseeds addedrdquotreatment had 2000 seeds added per speciesldquoRecruitmentrdquo is the total number of seedlings found
Treatment Recruitment
No seeds added 0 Sb 0 SeSeeds added canopy left intact 92 Sb 0 SeSeeds added canopy cleared 180 Sb 2 Se
ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features
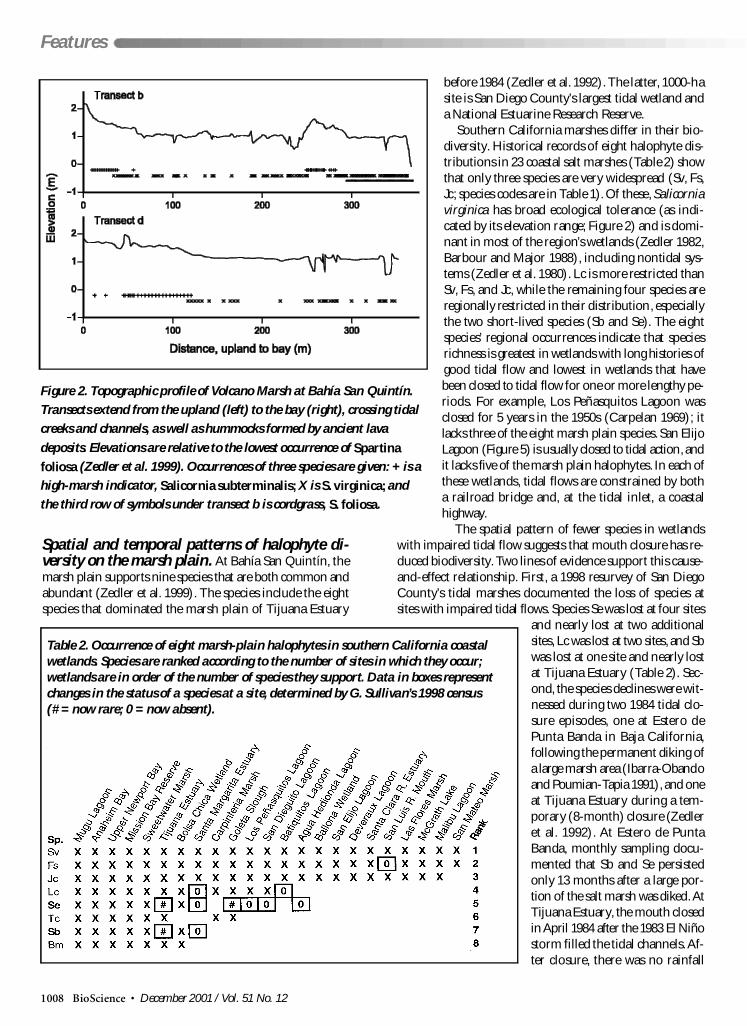
Spatial and temporal patterns of halophyte di-versity on the marsh plain At Bahiacutea San Quintiacuten themarsh plain supports nine species that are both common andabundant (Zedler et al 1999) The species include the eightspecies that dominated the marsh plain of Tijuana Estuary
before 1984 (Zedler et al 1992) The latter 1000-hasite is San Diego Countyrsquos largest tidal wetland anda National Estuarine Research Reserve
Southern California marshes differ in their bio-diversity Historical records of eight halophyte dis-tributions in 23 coastal salt marshes (Table 2) showthat only three species are very widespread (Sv FsJc species codes are in Table 1) Of these Salicorniavirginica has broad ecological tolerance (as indi-cated by its elevation range Figure 2) and is domi-nant in most of the regionrsquos wetlands (Zedler 1982Barbour and Major 1988) including nontidal sys-tems (Zedler et al 1980) Lc is more restricted thanSv Fs and Jc while the remaining four species areregionally restricted in their distribution especiallythe two short-lived species (Sb and Se) The eightspeciesrsquo regional occurrences indicate that speciesrichness is greatest in wetlands with long histories ofgood tidal flow and lowest in wetlands that havebeen closed to tidal flow for one or more lengthy pe-riods For example Los Pentildeasquitos Lagoon wasclosed for 5 years in the 1950s (Carpelan 1969) itlacks three of the eight marsh plain species San ElijoLagoon (Figure 5) is usually closed to tidal action andit lacks five of the marsh plain halophytes In each ofthese wetlands tidal flows are constrained by botha railroad bridge and at the tidal inlet a coastalhighway
The spatial pattern of fewer species in wetlandswith impaired tidal flow suggests that mouth closure has re-duced biodiversity Two lines of evidence support this cause-and-effect relationship First a 1998 resurvey of San DiegoCountyrsquos tidal marshes documented the loss of species atsites with impaired tidal flows Species Se was lost at four sites
and nearly lost at two additionalsites Lc was lost at two sites and Sbwas lost at one site and nearly lostat Tijuana Estuary (Table 2) Sec-ond the species declines were wit-nessed during two 1984 tidal clo-sure episodes one at Estero dePunta Banda in Baja Californiafollowing the permanent diking ofa large marsh area (Ibarra-Obandoand Poumian-Tapia 1991) and oneat Tijuana Estuary during a tem-porary (8-month) closure (Zedleret al 1992) At Estero de PuntaBanda monthly sampling docu-mented that Sb and Se persistedonly 13 months after a large por-tion of the salt marsh was diked AtTijuana Estuary the mouth closedin April 1984 after the 1983 El Nintildeostorm filled the tidal channels Af-ter closure there was no rainfall
1008 BioScience December 2001 Vol 51 No 12
Features
Table 2 Occurrence of eight marsh-plain halophytes in southern California coastalwetlands Species are ranked according to the number of sites in which they occurwetlands are in order of the number of species they support Data in boxes representchanges in the status of a species at a site determined by G Sullivanrsquos 1998 census ( = now rare 0 = now absent)
Figure 2 Topographic profile of Volcano Marsh at Bahiacutea San Quintiacuten
Transects extend from the upland (left) to the bay (right) crossing tidal
creeks and channels as well as hummocks formed by ancient lava
deposits Elevations are relative to the lowest occurrence of Spartina
foliosa (Zedler et al 1999) Occurrences of three species are given + is a
high-marsh indicator Salicornia subterminalis X is S virginica and
the third row of symbols under transect b is cordgrass S foliosa
in an unusually dry growing season Channel salinities roseto about 60 parts per thousand (ppt) and soil salinities roseto an average of 104 ppt (Zedler et al 1992) Species Sb andSe were virtually extirpated and did not recover from the 8-month nontidal catastrophe (Table 2)
The pattern of Sb and Se loss with reduced tidal influenceis consistent across the region in both space and time and atboth Estero de Punta Banda and Tijuana Estuary The patternis also obvious from monitoring small-scale (025 m2) quadrats(Figure 6) Prior to tidal closure the marsh plain of TijuanaEstuary supported a mean of 45 species during closure themean dropped below 20 Little recovery was evident in sub-sequent years despite the return of tidal influence Our May1999 data from Bahiacutea San Quintiacuten had a mean of 47 speciesper 025-m2 quadrat (n = 224) We conclude that biodiver-sity has declined at Punta Banda and Tijuana estuaries as aresult of interrupted tidal influence
Testing speciesrsquo roles in ecosystem function Given the 90 loss of wetland habitat in southern Californiarestoration is a high priority Early attempts sought only toprovide vegetation cover It was common to plant just one ortwo easily grown species (S foliosa in low elevations Sv on themarsh plain Zedler 2001) We hypothesize that ecosystemfunctioning might be impaired if too few species are re-stored No one knows how many are needed for the systemto develop at maximum speed or to achieve desired levels ofecosystem functions such as the development and mainte-nance of natural community composition the developmentof appropriate canopy structure and the accumulation of or-ganic matter and nitrogen These span the range of indica-tors that Noss (1990) recommends for monitoring biodi-versity The assessment of just one function such as netprimary productivity above ground (or its surrogate struc-tural attribute biomass at the end of the growing season) isnot likely to serve ecosystem managers or settle internationaldebates about the importance of diversity to ecosystem func-tioning Hence we tested the effects of species-rich plantingson the development of multiple functions that are critical inrestoration sites
In 1997 we established a large field experiment at TijuanaEstuary to test the effects of species richness on three specificresponse variables
1 Recruitment Which species need to be planted andwhich readily establish and spread
2 Canopy complexity Which grow taller and producemore layers as might be needed by insects and birds
3 Belowground biomass and N accumulation Whichplots store the most N in roots and rhizomes
Our experimental site was a graded marsh plain thathad room for 87 2 x 2-m plots distributed among fiveblocks along an excavated tidal channel (Figure 7) Our ex-perimental conditions included single-species plots of eachof the eight species of the nearby reference marsh plusrandomly chosen six- and three-species assemblages whichroughly correspond to small-scale richness levels beforeand after tidal closure at Tijuana Estuary (Figure 6) Sev-eral of our experimental assemblages occurred in TijuanaEstuary in 1974 and Bahiacutea San Quintiacuten in 1999 but ran-dom draws missed the most common trio (Bm Sv Sb)and sextet (Bm Sv Sb Fs Jc Se) from Tijuana Estuaryrsquos025-m2 plots Hence we added one plot with Bm Sv Sband one with Bm Sv Sb Fs Jc Se The 87 field experimentalplots were planted in April 1997 and replanted and weededas needed that summer to maintain planting treatments andcompensate for early mortality The assemblages planted re-sulted in similar numbers of plants introduced within theexperimental site Thus differential starting densities werenot of concern in comparing recruitment Assemblagecomposition was maintained during 1998 by removing
December 2001 Vol 51 No 12 BioScience 1009
Features
Figure 3 A remotely sensed image of Volcano Marshshowing tidal channels Mosaic compiled by BruceNyden Pacific Estuarine Research Lab San Diego StateUniversity
seedlings of other species thereafter recruits were allowedto persist
Seedlings were censused by Roberto Lindig-Cisneros(2001) canopy architecture was documented by Keer andZedler (forthcoming) and biomass and N accumulationwere assessed by J Callaway Because we were most con-cerned about the increasing ability of the site to retain Nwe assessed biomass and N accumulation in midwinter af-ter most plants had slowed growth Furthermore becausewe wanted to minimize destructive sampling we removedsmall root cores in year 2 and root and shoot samples at theend of year 3 Both biomass components were analyzed forabove- and belowground N content
We complemented the field study with an experiment inthe greenhouse where we were able to compare individual as-semblages We replicated all 15 randomly drawn trios and sex-
tets 8-fold all solos 16-fold and the common trioand sextet 16-fold for a total of 400 microcosmsPlanting treatments were maintained in the potsthrough vegetative growth and seedling recruit-ment but assemblage composition did not changeAll but 80 pots were grown for 2 years and as-sessed for canopy attributes and harvested for bio-mass and N content above and below ground
The importance of diversity to restorationResults from the field and greenhouse experimentssupport the general hypothesis that diversity mat-ters in restoration sites Our plantings with sixspecies enhanced recruitment made canopiesmore complex increased biomass and increasedN accumulation
Recruitment Only three species (Sv Sb andSe) recruited readily (Table 3) Of these only one(Sv) is widely distributed (Table 2) and available todisperse into restoration sites It germinates in sea-water and produces abundant seedlings in bothnatural and restored wetlands including our un-planted plots it does not need to be planted torestoration sites if a seed source exists nearby Theother two abundant recruiters Sb and Se are themarsh plainrsquos only short-lived species Personalobservation revealed that Se recruitment potentialwas much lower than that of Sb reflecting its lowerdensity in regional wetlands and differences inseedling mortality within this experiment Thefact that both recruited readily in the bare restora-tion site and poorly in the salt marsh where it oc-curred historically is evidence that the Tijuana Es-tuary marsh plain no longer provides suitablehabitat for these species The remaining five speciesdid not readily colonize either our planted or un-planted plots Their inability to recruit may be at-
1010 BioScience December 2001 Vol 51 No 12
Features
Table 3 Seedling recruitment of eight marsh-plainspecies into experimental plots Data are for the 1998growing season Note that the number of plantsintroduced to the site as seedlings in April 1997(column 2) was similar but only three speciesrecruited extensively Species codes are in Table 1
Code Number planted Number of recruits
Sv 840 17703Sb 735 15978Se 795 1668Tc 810 73Jc 840 33Bm 720 26Lc 795 24Fs 765 2
Figure 4 Coastal wetlands of southern California 1Tijuana Estuary 2Sweetwater Marsh 3 Mission Bay Reserve 4 Los Pentildeasquitos Lagoon 5San Dieguito Lagoon 6 San Elijo Lagoon 7 Batiquitos Lagoon 8 AguaHedionda Lagoon 9 Buena Vista Lagoon (brackish wetland) 10 SanLuis Rey River Mouth 11 Santa Margarita Estuary 12 Las FloresMarsh 13 San Mateo Marsh 14 Upper Newport Bay 15 Bolsa ChicaWetland 16 Anaheim Bay 17 Los Cerritos Marsh (mostly filled anddeveloped) 18 Ballona Wetland 19 Malibu Lagoon 20 Mugu Lagoon21 McGrath Lake 22 Santa Clara River Estuary 23 Ventura River(brackish) 24 Carpinteria Marsh 25 Goleta Slough 26 DeverauxLagoon
tributable to small seed sup-ply poor germination or un-suitable conditions for es-tablishment
Canopy architectureKeer and Zedler (forthcom-ing) found that all plotsrapidly achieved high coverbut that canopy height andlayering were slow to de-velop Cover is a poor mea-sure of canopy complexity itdoes not differentiate speciesor species-richness treat-ments Maximum heightswere greatest for trios andsextets reaching about 60cm by year 3 Sv grew thetallest in the first 18 months of study followed by Sb and SeLayering (the number of times a vertical rod encountersplant parts as it is lowered through the canopy) was greaterfor trios and sextets both of which achieved three layerswithin 18 months The effects of individual species werelikely to be seen early in the development of the plots Fs andSe had six and five layers respectively with Jc at four layersLayering is hypothesized to enhance habitat for both arthro-pods and birds These canopies are far from fully developedreference marshes with monotypic Sv averaged 45 cm tall withseven layers and areas with multiple species were 55 cm talland had 13 layers (Keer and Zedler forthcoming)
Biomass and N accumulation In the field plots above-ground biomass averaged under 300 g per m2 for solos andover 400 g per m2 for sextets Belowground biomass also in-creased with species richness and over timemdashmeans doubledbetween years 2 and 3 (to 500g per m2 for sextets) The rapidincrease in belowground biomass is highly desirable for barerestoration sites To determine whether N crops increased wemeasured biomass and tissue N concentrations The result-ing estimates of accumulated N showed a higher crop for sex-tets than for unplanted and solo plots The rapid accumula-tion of N is beneficial for restoration sites that have coarse soiland where N is the limiting factor (a common problem in cre-ated and restored wetlands Langis et al 1991 Boyer andZedler 1996 1998 Zedler and Callaway 1999 2000 Boyer etal 2000) Our greenhouse experiment revealed similar pat-terns in a more benign environment (that is one with warmertemperatures) Total biomass was higher for sextets becausebelowground biomass increased with species richness As inthe field N crops tracked biomass with greater total N ac-cumulation and greater belowground accumulation at year2A species composition effectmdashnamely the chance inclusionof high biomass producers in the randomly drawn assem-blagemdashis indicated because Jc Fs and Lc solos all accumu-lated more N than the mean for sextets and Sv and Bm
solos both accumulated as much as the triosAlthough Tc Seand Sb solos all had below-average N crops Sb and Tc accu-mulated the most N per unit biomass
These findings were used to design the planting of the 8-ha ldquoFriendship Marshrdquo a $3 million project that aims to sup-port rare species of plants and animals (Figure 8) Located inthe south arm of Tijuana Estuary this site was opened to tidalflushing in February 2000 after floodborne sediments (1ndash2 m)were excavated and the historical marsh plain uncoveredBecause Sv dominates the nearby wetlands and readily es-tablishes on its own it was not introduced as seeds or plantsInstead we planted five species for which we had abundantseedlings (Fs Jc Bm Lc and Se) to help provide the functionsmentioned abovemdashnamely recruitment complex canopiesand biomass and N accumulation The protocol of not plant-ing Sv (to slow time to dominance) and adding multispeciesclusters (to accelerate the development of species-rich
December 2001 Vol 51 No 12 BioScience 1011
Features
Figure 5 San Elijo Lagoon San Diego County California
Figure 6 Historical patterns of species richness atTijuana Estuary data (means and standard errors) arefrom 025-m2 circular plots
canopies) is new to the region Previous marsh plain restora-tion efforts have focused on planting Sv
Although the field and greenhouse experiments taught usthat species-rich plots promote recruitment develop morecomplex canopies and accumulate more biomass and Nthe performance of sextets could be attributable to the com-bined performance of the six species or to the chance inclu-sion of a single high-performance species (a species compo-sition effect) Data from the replicated assemblages in thegreenhouse allowed us to evaluate these alternatives In the mi-crocosms species composition was stable over time (whereasrecruitment increased richness of solo plots in the field) Wecompared the eight species and found that Jc produced themost shoot biomass Fs the most root biomass and Sv themost total biomass while Tc had the highest rootshoot bio-mass But their rankings changed when ash-free biomass
was considered because Sv had the most ash per plant andSb the most ash per unit biomass For ash-free biomass Jc washighest for shoots Fs for roots and Jc for total with Tc stillhaving the highest rootshoot ratio The N concentration ofshoots was highest for Sb for roots it was highest for Tc Jchad the highest crop of N in its shoots and Fs the highest rootN crop while Jc had the highest total N crop Height was great-est for Se and layering was greatest for Fs Interestingly theregionrsquos ldquosuperplantrdquo Sv achieved its first place overall rank-ing not by excelling in multiple attributes but by scoring sec-ond or third in most attributes It scored highest only in to-tal biomass but it achieved high biomass by accumulating alarge volume of salts in its tissues in addition to accumulat-ing considerable carbon
This meta-analysis supports both alternatives Speciescomposition effects (Symstad et al 1998) were indicated forevery attribute considered (there is at least one species thatcan match the performance of the average sextet) but it is notthe same species for all response variables (that is all speciesare needed to achieve high levels of all functions) Further-more each species excelled in at least one attribute Thus weconclude that diversity matters a great deal and the onlyway to ensure that multiple functions are provided is to plantand sustain all species Retaining diversity results in less riskthat functions will be eliminated The planting of species-richassemblages in restoration sites would help ensure that morefunctions are restored
There is support for the suggestion that attributes measuredin the greenhouse translate into performance in the field Thespecies that performed best in the greenhouse are most wide-spread geographically Overall the eight species ranked in vir-tually the same order as in their distributional ranking (thatis the number of southern California wetlands in which theyoccur Table 1) The top three performers in the greenhousewere Sv Jc and Fs and they all occur in 22 or 23 wetlandsLc ranked fourth in greenhouse performance and fourth inregional distribution occurring in 13 wetlands in the historicalrecord and in 11 wetlands in 1998 The others ranked fifththrough eighth in performance they are the most restrictedin their distributions now occurring in only six to eight sitesJc Fs and Sv are all highly productive long-lived perennialsthat reproduce vegetatively and form monotypic patches(Zedler et al 1980 Ferren 1985) It is not surprising thatthey persist in wetlands that experience closure to tidal in-fluence
What functions were lost when short-lived species dropped out of Tijuana Estuary Sb and Se are short-lived halophytes that do not reproducevegetatively Their roles in the ecosystem are arguably uniquefrom these perspectives alone Their annual recruitment ofseedlings following winter rainfall adds seasonality to thecanopy and soil perhaps changing food availability bothabove and below ground Regrettably these functions have notbeen quantified
1012 BioScience December 2001 Vol 51 No 12
Features
Figure 7 Adaptive restoration exemplified by theexperimental planting of 87 2-by-2-meter plots with 0 13 and 6 species in a newly excavated marsh plain atTijuana Estuary Photograph taken in June 1997 2months after planting greenhouse-grown seedlings (90per plot)
As our field data docu-ment Sv Sb and Se are allgood recruiters capable offilling canopy openings withseedlings and rapidly reveg-etating bare sites Howeverthe 1998 recruitment data(Lindig-Cisneros 2001) showthat Sb and Se producedseedlings earlier than Sv Inearly April 1998 there were2199 Sb and 220 Se seedlingsbut only 89 Sv seedlings Inour experimental design wepulled seedlings of invadingspecies to retain assemblagecomposition in 1997ndash1998so there was ample oppor-tunity for Sv seedlings to re-cruit later In early May 1998there were 9261 Sv seedlingsbut only 842 Sb and 50 Se re-cruitment then declined forall species over the late Mayand July sampling dates despite the fact that seedlings werebeing pulled and space remained available for establishmentWe infer from these results that Sb and Se have important rolesin filling gaps that is space opened late in the year is filled bySb and Se the following winter and spring
Both Sb and Se are capable of rapid height growth asdemonstrated in the greenhouse Se might be the first speciesto provide perch sites for the endangered Beldingrsquos Savannahsparrow which defends its nesting territories by singing whileperching on the tallest plants (Powell 1993) The verticalstructure afforded by Se differs from that of Sv in having amore horizontal branching angle and more lay-ers attributes that may increase its attraction ofsmall birds It is not uncommon to see sparrowsperching on tall branches of Se in the fieldwhether the plants are alive or dead We concludethat the loss of Sb and Se could impair the com-munityrsquos ability to recover from disturbanceand to support an endangered bird These func-tions are not entirely eliminated by the demiseof Sb and Se but the ecosystemrsquos functional po-tential might not be realized in their absence
Is the loss of species reversibleTijuana Estuaryrsquos current salt marsh communityis certainly different from that prior to 1984when an open canopy supported thousands ofSb seedlings per square meter and Se was com-mon throughout the tidal marsh Today thesespecies persist in a tiny fraction of the area theyoccupied historicallyAs short-lived species bothSb (annual) and Se (lives only a few years)
depend on recruitment for their persistence while all of theother marsh plain species reproduce vegetatively Seedlingswere young when Tijuana Estuary became nontidal in April1984 and one likely cause of their mortality was the dryingof surface soil and rise in salinity to an average of greater than100 ppt (Figure 9) Moderate hypersalinity (40ndash45 ppt) is typ-ical for marsh plain soils but extreme hypersalinity is morecharacteristic of higher elevations with less frequent tidal in-undation (Noe and Zedler 2001) Sb and Se can germinate inseawater but higher salinities reduce germination rates (Zedlerand Beare 1986) Had their seed banks persisted through the
December 2001 Vol 51 No 12 BioScience 1013
Features
Figure 8 An 8-ha marsh plain excavation at Tijuana Estuary Because of the large size of thissite plantings emphasized species that would not likely recruit on their own
Figure 9 Soil salinity at Tijuana Estuary (data from the Pacific EstuarineResearch Lab) Means before A on the x-axis include Spartina marsh andthe marsh plain means after A are marsh plain only Means before B arefor interstitial soil water after B are for soil pastes (adding deionized waterunderestimates interstitial salinity)
period of tidal closure Sb and Se should have germinated in1985 they did not
A recent reintroduction experiment by Vivian-Smith andZedler (2001) indicated that neither species retains a marshplain seed bank at Tijuana Estuary Vivian-Smith added 2000seeds of each species in 1998 Sb produced 272 seedlings andSe produced 2 with seedlings establishing only where seedswere added Also the current Sv-dominated canopy reducedrecruitment with Sb seedling density twice as high where thecanopy was removed (Table 4) Hence we learned that a lackof seeds and shading by Sv restricted recovery of the Sb pop-ulation And because H N Morzaria-Luna (University ofWisconsinndashMadison unpublished data) could not locate ei-ther species during her search in 2001 we suspect that changesin both the physical and biological conditions of this marshfurther inhibit recovery We explore each of these in turn
Irreversible physical changes According to recent in-formation both elevation and soil salinities have changed inthe 16 years since tidal closure A study of marsh accretion rates(Weis et al forthcoming) using cesium-137 (137Cs a globallydispersed radioisotope) suggests that since 1984 (the yearwhen Sb and Se declined) the marsh plain elevation has risen13 cm based on two soil cores At three additional locationsbenchmarks placed on the marsh plain in the 1970s by the USArmy Corps of Engineers were relocated there we have mea-sured an average of 6 cm accretion since 1984 (Morzaria-Lunaunpublished data) Further sedimentation data from TijuanaEstuary corroborate this rapid rise in elevation Cahoon andcolleagues (1996) measured 20ndash85 (mean 59) cm accretionin the S foliosa marsh between 1992 and 1993 and in anearby tidal pond (lower elevation) Ward (2000) found4ndash127 (mean 88) cm of sediment accretion from 1997 to1998 The marsh plain elevation has risen because of sedi-mentation and the recent record of flooding provides anexplanation
Tijuana Estuary has a large watershed with highly erodablesoils and major slope failures that are visible both north andsouth of the USndashMexico border The 1993 floodwaters cut anentirely new river channel just inland of the estuary Thehuge volume of sediment that was mobilized could havecontributed to a rise in elevation on the marsh plain In-creased soil salinities (now averaging 60 ppt Figure 9) cor-
roborate a shift toward a higher and hence drier marshplain For intertidal species that are sensitive to 5ndash10-cm dif-ferences in elevation the accretion rates and associated salin-ities documented at Tijuana Estuary would seem to be fatalPersistent hypersaline conditions would reduce Sb and Se es-tablishment drier conditions would favor Sv and densercanopies of Sv would reduce the persistence of seedlings
If rising elevation and increasing salinity prevent reestab-lishment of Sb and Se then reversing these changes would re-quire a massive sediment removal effortmdashthe marsh plainwould have to be bulldozed to lower the elevation by 6ndash13 cmor more Such a disruption to the vegetation would not be al-lowed by resource managers because the plant canopy isused by light-footed clapper rails for cover and Beldingrsquos Sa-vannah sparrows for nesting
There is an alternative Although these recent physicalchanges are not reversible it is possible to excavate sedi-ments and restore areas where sediment loads have shiftedtopography well out of the tidal range Exactly this procedureis planned for up to 200 ha of former tidal wetlands in thesouthern arm of the estuary (Entrix et al 1991) The re-moval of sediments in the south arm can conceivably com-pensate for some of the accretion that is occurring in the es-tuary and in the region Attempts are also being made tocontrol the flow of sediments upstream
Irreversible biological changes When the full com-plement of species is present the community presumably hasmaximum biocomplexitymdashthat is a variety of interspecificinteractions and feedback mechanisms Its biodiversity wouldlead to biocomplexity It is possible that the marshrsquos bio-complexity helped sustain its biodiversity If some interactionsfacilitated rare species and others kept dominants from out-competing subordinates then more species would persist Inthat event losing some species would make the communityless able to support all the naturally occurring species bothplant and animalWhere dense canopy cover reduces seedlingrecruitment any species that reduces overall biomass above-ground could reduce shading and enhance recruitment andspecies richness The greenhouse experiments with repli-cated assemblages suggest such a species effect
We explored effects of selected species by comparing mi-crocosms with and without such a species We found threespecies (Sv Fs and Jc) that tended to increase whole-potbiomass (a typical effect of productive species) and one (Tc)that tended to decrease whole-pot biomass (an unusual effect)Assemblages with Tc had lower shoot biomass than assem-blages without it Tc produces little biomass when grownalone it is short with an unimpressive canopy Althoughlow in biomass Tc accumulates a disproportionate concen-tration of N twice that for Sv In the microcosms total ash-free biomass values for Tc and Sv were significantly negativelycorrelated (r = ndash043 p lt 01 n = 72 sextets) Despite itssmall size Tc seems capable of reducing the biomass of SvField data from Bahiacutea San Quintiacuten also suggest a negative re-lationship between Tc and Sv cover (r = ndash0133 p = 05 n =224) We conclude that Tc has some mechanism for reduc-
1014 BioScience December 2001 Vol 51 No 12
Features
Table 4 Experimental attempt to reestablish Sb and Seat Tijuana Estuary in 1998 (unpublished datacollected by Gabrielle Vivian-Smith Pacific EstuarineResearch Lab) Species codes are Sb Salicorniabigelovii Se Suaeda esteroa The ldquoseeds addedrdquotreatment had 2000 seeds added per speciesldquoRecruitmentrdquo is the total number of seedlings found
Treatment Recruitment
No seeds added 0 Sb 0 SeSeeds added canopy left intact 92 Sb 0 SeSeeds added canopy cleared 180 Sb 2 Se
ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features

in an unusually dry growing season Channel salinities roseto about 60 parts per thousand (ppt) and soil salinities roseto an average of 104 ppt (Zedler et al 1992) Species Sb andSe were virtually extirpated and did not recover from the 8-month nontidal catastrophe (Table 2)
The pattern of Sb and Se loss with reduced tidal influenceis consistent across the region in both space and time and atboth Estero de Punta Banda and Tijuana Estuary The patternis also obvious from monitoring small-scale (025 m2) quadrats(Figure 6) Prior to tidal closure the marsh plain of TijuanaEstuary supported a mean of 45 species during closure themean dropped below 20 Little recovery was evident in sub-sequent years despite the return of tidal influence Our May1999 data from Bahiacutea San Quintiacuten had a mean of 47 speciesper 025-m2 quadrat (n = 224) We conclude that biodiver-sity has declined at Punta Banda and Tijuana estuaries as aresult of interrupted tidal influence
Testing speciesrsquo roles in ecosystem function Given the 90 loss of wetland habitat in southern Californiarestoration is a high priority Early attempts sought only toprovide vegetation cover It was common to plant just one ortwo easily grown species (S foliosa in low elevations Sv on themarsh plain Zedler 2001) We hypothesize that ecosystemfunctioning might be impaired if too few species are re-stored No one knows how many are needed for the systemto develop at maximum speed or to achieve desired levels ofecosystem functions such as the development and mainte-nance of natural community composition the developmentof appropriate canopy structure and the accumulation of or-ganic matter and nitrogen These span the range of indica-tors that Noss (1990) recommends for monitoring biodi-versity The assessment of just one function such as netprimary productivity above ground (or its surrogate struc-tural attribute biomass at the end of the growing season) isnot likely to serve ecosystem managers or settle internationaldebates about the importance of diversity to ecosystem func-tioning Hence we tested the effects of species-rich plantingson the development of multiple functions that are critical inrestoration sites
In 1997 we established a large field experiment at TijuanaEstuary to test the effects of species richness on three specificresponse variables
1 Recruitment Which species need to be planted andwhich readily establish and spread
2 Canopy complexity Which grow taller and producemore layers as might be needed by insects and birds
3 Belowground biomass and N accumulation Whichplots store the most N in roots and rhizomes
Our experimental site was a graded marsh plain thathad room for 87 2 x 2-m plots distributed among fiveblocks along an excavated tidal channel (Figure 7) Our ex-perimental conditions included single-species plots of eachof the eight species of the nearby reference marsh plusrandomly chosen six- and three-species assemblages whichroughly correspond to small-scale richness levels beforeand after tidal closure at Tijuana Estuary (Figure 6) Sev-eral of our experimental assemblages occurred in TijuanaEstuary in 1974 and Bahiacutea San Quintiacuten in 1999 but ran-dom draws missed the most common trio (Bm Sv Sb)and sextet (Bm Sv Sb Fs Jc Se) from Tijuana Estuaryrsquos025-m2 plots Hence we added one plot with Bm Sv Sband one with Bm Sv Sb Fs Jc Se The 87 field experimentalplots were planted in April 1997 and replanted and weededas needed that summer to maintain planting treatments andcompensate for early mortality The assemblages planted re-sulted in similar numbers of plants introduced within theexperimental site Thus differential starting densities werenot of concern in comparing recruitment Assemblagecomposition was maintained during 1998 by removing
December 2001 Vol 51 No 12 BioScience 1009
Features
Figure 3 A remotely sensed image of Volcano Marshshowing tidal channels Mosaic compiled by BruceNyden Pacific Estuarine Research Lab San Diego StateUniversity
seedlings of other species thereafter recruits were allowedto persist
Seedlings were censused by Roberto Lindig-Cisneros(2001) canopy architecture was documented by Keer andZedler (forthcoming) and biomass and N accumulationwere assessed by J Callaway Because we were most con-cerned about the increasing ability of the site to retain Nwe assessed biomass and N accumulation in midwinter af-ter most plants had slowed growth Furthermore becausewe wanted to minimize destructive sampling we removedsmall root cores in year 2 and root and shoot samples at theend of year 3 Both biomass components were analyzed forabove- and belowground N content
We complemented the field study with an experiment inthe greenhouse where we were able to compare individual as-semblages We replicated all 15 randomly drawn trios and sex-
tets 8-fold all solos 16-fold and the common trioand sextet 16-fold for a total of 400 microcosmsPlanting treatments were maintained in the potsthrough vegetative growth and seedling recruit-ment but assemblage composition did not changeAll but 80 pots were grown for 2 years and as-sessed for canopy attributes and harvested for bio-mass and N content above and below ground
The importance of diversity to restorationResults from the field and greenhouse experimentssupport the general hypothesis that diversity mat-ters in restoration sites Our plantings with sixspecies enhanced recruitment made canopiesmore complex increased biomass and increasedN accumulation
Recruitment Only three species (Sv Sb andSe) recruited readily (Table 3) Of these only one(Sv) is widely distributed (Table 2) and available todisperse into restoration sites It germinates in sea-water and produces abundant seedlings in bothnatural and restored wetlands including our un-planted plots it does not need to be planted torestoration sites if a seed source exists nearby Theother two abundant recruiters Sb and Se are themarsh plainrsquos only short-lived species Personalobservation revealed that Se recruitment potentialwas much lower than that of Sb reflecting its lowerdensity in regional wetlands and differences inseedling mortality within this experiment Thefact that both recruited readily in the bare restora-tion site and poorly in the salt marsh where it oc-curred historically is evidence that the Tijuana Es-tuary marsh plain no longer provides suitablehabitat for these species The remaining five speciesdid not readily colonize either our planted or un-planted plots Their inability to recruit may be at-
1010 BioScience December 2001 Vol 51 No 12
Features
Table 3 Seedling recruitment of eight marsh-plainspecies into experimental plots Data are for the 1998growing season Note that the number of plantsintroduced to the site as seedlings in April 1997(column 2) was similar but only three speciesrecruited extensively Species codes are in Table 1
Code Number planted Number of recruits
Sv 840 17703Sb 735 15978Se 795 1668Tc 810 73Jc 840 33Bm 720 26Lc 795 24Fs 765 2
Figure 4 Coastal wetlands of southern California 1Tijuana Estuary 2Sweetwater Marsh 3 Mission Bay Reserve 4 Los Pentildeasquitos Lagoon 5San Dieguito Lagoon 6 San Elijo Lagoon 7 Batiquitos Lagoon 8 AguaHedionda Lagoon 9 Buena Vista Lagoon (brackish wetland) 10 SanLuis Rey River Mouth 11 Santa Margarita Estuary 12 Las FloresMarsh 13 San Mateo Marsh 14 Upper Newport Bay 15 Bolsa ChicaWetland 16 Anaheim Bay 17 Los Cerritos Marsh (mostly filled anddeveloped) 18 Ballona Wetland 19 Malibu Lagoon 20 Mugu Lagoon21 McGrath Lake 22 Santa Clara River Estuary 23 Ventura River(brackish) 24 Carpinteria Marsh 25 Goleta Slough 26 DeverauxLagoon
tributable to small seed sup-ply poor germination or un-suitable conditions for es-tablishment
Canopy architectureKeer and Zedler (forthcom-ing) found that all plotsrapidly achieved high coverbut that canopy height andlayering were slow to de-velop Cover is a poor mea-sure of canopy complexity itdoes not differentiate speciesor species-richness treat-ments Maximum heightswere greatest for trios andsextets reaching about 60cm by year 3 Sv grew thetallest in the first 18 months of study followed by Sb and SeLayering (the number of times a vertical rod encountersplant parts as it is lowered through the canopy) was greaterfor trios and sextets both of which achieved three layerswithin 18 months The effects of individual species werelikely to be seen early in the development of the plots Fs andSe had six and five layers respectively with Jc at four layersLayering is hypothesized to enhance habitat for both arthro-pods and birds These canopies are far from fully developedreference marshes with monotypic Sv averaged 45 cm tall withseven layers and areas with multiple species were 55 cm talland had 13 layers (Keer and Zedler forthcoming)
Biomass and N accumulation In the field plots above-ground biomass averaged under 300 g per m2 for solos andover 400 g per m2 for sextets Belowground biomass also in-creased with species richness and over timemdashmeans doubledbetween years 2 and 3 (to 500g per m2 for sextets) The rapidincrease in belowground biomass is highly desirable for barerestoration sites To determine whether N crops increased wemeasured biomass and tissue N concentrations The result-ing estimates of accumulated N showed a higher crop for sex-tets than for unplanted and solo plots The rapid accumula-tion of N is beneficial for restoration sites that have coarse soiland where N is the limiting factor (a common problem in cre-ated and restored wetlands Langis et al 1991 Boyer andZedler 1996 1998 Zedler and Callaway 1999 2000 Boyer etal 2000) Our greenhouse experiment revealed similar pat-terns in a more benign environment (that is one with warmertemperatures) Total biomass was higher for sextets becausebelowground biomass increased with species richness As inthe field N crops tracked biomass with greater total N ac-cumulation and greater belowground accumulation at year2A species composition effectmdashnamely the chance inclusionof high biomass producers in the randomly drawn assem-blagemdashis indicated because Jc Fs and Lc solos all accumu-lated more N than the mean for sextets and Sv and Bm
solos both accumulated as much as the triosAlthough Tc Seand Sb solos all had below-average N crops Sb and Tc accu-mulated the most N per unit biomass
These findings were used to design the planting of the 8-ha ldquoFriendship Marshrdquo a $3 million project that aims to sup-port rare species of plants and animals (Figure 8) Located inthe south arm of Tijuana Estuary this site was opened to tidalflushing in February 2000 after floodborne sediments (1ndash2 m)were excavated and the historical marsh plain uncoveredBecause Sv dominates the nearby wetlands and readily es-tablishes on its own it was not introduced as seeds or plantsInstead we planted five species for which we had abundantseedlings (Fs Jc Bm Lc and Se) to help provide the functionsmentioned abovemdashnamely recruitment complex canopiesand biomass and N accumulation The protocol of not plant-ing Sv (to slow time to dominance) and adding multispeciesclusters (to accelerate the development of species-rich
December 2001 Vol 51 No 12 BioScience 1011
Features
Figure 5 San Elijo Lagoon San Diego County California
Figure 6 Historical patterns of species richness atTijuana Estuary data (means and standard errors) arefrom 025-m2 circular plots
canopies) is new to the region Previous marsh plain restora-tion efforts have focused on planting Sv
Although the field and greenhouse experiments taught usthat species-rich plots promote recruitment develop morecomplex canopies and accumulate more biomass and Nthe performance of sextets could be attributable to the com-bined performance of the six species or to the chance inclu-sion of a single high-performance species (a species compo-sition effect) Data from the replicated assemblages in thegreenhouse allowed us to evaluate these alternatives In the mi-crocosms species composition was stable over time (whereasrecruitment increased richness of solo plots in the field) Wecompared the eight species and found that Jc produced themost shoot biomass Fs the most root biomass and Sv themost total biomass while Tc had the highest rootshoot bio-mass But their rankings changed when ash-free biomass
was considered because Sv had the most ash per plant andSb the most ash per unit biomass For ash-free biomass Jc washighest for shoots Fs for roots and Jc for total with Tc stillhaving the highest rootshoot ratio The N concentration ofshoots was highest for Sb for roots it was highest for Tc Jchad the highest crop of N in its shoots and Fs the highest rootN crop while Jc had the highest total N crop Height was great-est for Se and layering was greatest for Fs Interestingly theregionrsquos ldquosuperplantrdquo Sv achieved its first place overall rank-ing not by excelling in multiple attributes but by scoring sec-ond or third in most attributes It scored highest only in to-tal biomass but it achieved high biomass by accumulating alarge volume of salts in its tissues in addition to accumulat-ing considerable carbon
This meta-analysis supports both alternatives Speciescomposition effects (Symstad et al 1998) were indicated forevery attribute considered (there is at least one species thatcan match the performance of the average sextet) but it is notthe same species for all response variables (that is all speciesare needed to achieve high levels of all functions) Further-more each species excelled in at least one attribute Thus weconclude that diversity matters a great deal and the onlyway to ensure that multiple functions are provided is to plantand sustain all species Retaining diversity results in less riskthat functions will be eliminated The planting of species-richassemblages in restoration sites would help ensure that morefunctions are restored
There is support for the suggestion that attributes measuredin the greenhouse translate into performance in the field Thespecies that performed best in the greenhouse are most wide-spread geographically Overall the eight species ranked in vir-tually the same order as in their distributional ranking (thatis the number of southern California wetlands in which theyoccur Table 1) The top three performers in the greenhousewere Sv Jc and Fs and they all occur in 22 or 23 wetlandsLc ranked fourth in greenhouse performance and fourth inregional distribution occurring in 13 wetlands in the historicalrecord and in 11 wetlands in 1998 The others ranked fifththrough eighth in performance they are the most restrictedin their distributions now occurring in only six to eight sitesJc Fs and Sv are all highly productive long-lived perennialsthat reproduce vegetatively and form monotypic patches(Zedler et al 1980 Ferren 1985) It is not surprising thatthey persist in wetlands that experience closure to tidal in-fluence
What functions were lost when short-lived species dropped out of Tijuana Estuary Sb and Se are short-lived halophytes that do not reproducevegetatively Their roles in the ecosystem are arguably uniquefrom these perspectives alone Their annual recruitment ofseedlings following winter rainfall adds seasonality to thecanopy and soil perhaps changing food availability bothabove and below ground Regrettably these functions have notbeen quantified
1012 BioScience December 2001 Vol 51 No 12
Features
Figure 7 Adaptive restoration exemplified by theexperimental planting of 87 2-by-2-meter plots with 0 13 and 6 species in a newly excavated marsh plain atTijuana Estuary Photograph taken in June 1997 2months after planting greenhouse-grown seedlings (90per plot)
As our field data docu-ment Sv Sb and Se are allgood recruiters capable offilling canopy openings withseedlings and rapidly reveg-etating bare sites Howeverthe 1998 recruitment data(Lindig-Cisneros 2001) showthat Sb and Se producedseedlings earlier than Sv Inearly April 1998 there were2199 Sb and 220 Se seedlingsbut only 89 Sv seedlings Inour experimental design wepulled seedlings of invadingspecies to retain assemblagecomposition in 1997ndash1998so there was ample oppor-tunity for Sv seedlings to re-cruit later In early May 1998there were 9261 Sv seedlingsbut only 842 Sb and 50 Se re-cruitment then declined forall species over the late Mayand July sampling dates despite the fact that seedlings werebeing pulled and space remained available for establishmentWe infer from these results that Sb and Se have important rolesin filling gaps that is space opened late in the year is filled bySb and Se the following winter and spring
Both Sb and Se are capable of rapid height growth asdemonstrated in the greenhouse Se might be the first speciesto provide perch sites for the endangered Beldingrsquos Savannahsparrow which defends its nesting territories by singing whileperching on the tallest plants (Powell 1993) The verticalstructure afforded by Se differs from that of Sv in having amore horizontal branching angle and more lay-ers attributes that may increase its attraction ofsmall birds It is not uncommon to see sparrowsperching on tall branches of Se in the fieldwhether the plants are alive or dead We concludethat the loss of Sb and Se could impair the com-munityrsquos ability to recover from disturbanceand to support an endangered bird These func-tions are not entirely eliminated by the demiseof Sb and Se but the ecosystemrsquos functional po-tential might not be realized in their absence
Is the loss of species reversibleTijuana Estuaryrsquos current salt marsh communityis certainly different from that prior to 1984when an open canopy supported thousands ofSb seedlings per square meter and Se was com-mon throughout the tidal marsh Today thesespecies persist in a tiny fraction of the area theyoccupied historicallyAs short-lived species bothSb (annual) and Se (lives only a few years)
depend on recruitment for their persistence while all of theother marsh plain species reproduce vegetatively Seedlingswere young when Tijuana Estuary became nontidal in April1984 and one likely cause of their mortality was the dryingof surface soil and rise in salinity to an average of greater than100 ppt (Figure 9) Moderate hypersalinity (40ndash45 ppt) is typ-ical for marsh plain soils but extreme hypersalinity is morecharacteristic of higher elevations with less frequent tidal in-undation (Noe and Zedler 2001) Sb and Se can germinate inseawater but higher salinities reduce germination rates (Zedlerand Beare 1986) Had their seed banks persisted through the
December 2001 Vol 51 No 12 BioScience 1013
Features
Figure 8 An 8-ha marsh plain excavation at Tijuana Estuary Because of the large size of thissite plantings emphasized species that would not likely recruit on their own
Figure 9 Soil salinity at Tijuana Estuary (data from the Pacific EstuarineResearch Lab) Means before A on the x-axis include Spartina marsh andthe marsh plain means after A are marsh plain only Means before B arefor interstitial soil water after B are for soil pastes (adding deionized waterunderestimates interstitial salinity)
period of tidal closure Sb and Se should have germinated in1985 they did not
A recent reintroduction experiment by Vivian-Smith andZedler (2001) indicated that neither species retains a marshplain seed bank at Tijuana Estuary Vivian-Smith added 2000seeds of each species in 1998 Sb produced 272 seedlings andSe produced 2 with seedlings establishing only where seedswere added Also the current Sv-dominated canopy reducedrecruitment with Sb seedling density twice as high where thecanopy was removed (Table 4) Hence we learned that a lackof seeds and shading by Sv restricted recovery of the Sb pop-ulation And because H N Morzaria-Luna (University ofWisconsinndashMadison unpublished data) could not locate ei-ther species during her search in 2001 we suspect that changesin both the physical and biological conditions of this marshfurther inhibit recovery We explore each of these in turn
Irreversible physical changes According to recent in-formation both elevation and soil salinities have changed inthe 16 years since tidal closure A study of marsh accretion rates(Weis et al forthcoming) using cesium-137 (137Cs a globallydispersed radioisotope) suggests that since 1984 (the yearwhen Sb and Se declined) the marsh plain elevation has risen13 cm based on two soil cores At three additional locationsbenchmarks placed on the marsh plain in the 1970s by the USArmy Corps of Engineers were relocated there we have mea-sured an average of 6 cm accretion since 1984 (Morzaria-Lunaunpublished data) Further sedimentation data from TijuanaEstuary corroborate this rapid rise in elevation Cahoon andcolleagues (1996) measured 20ndash85 (mean 59) cm accretionin the S foliosa marsh between 1992 and 1993 and in anearby tidal pond (lower elevation) Ward (2000) found4ndash127 (mean 88) cm of sediment accretion from 1997 to1998 The marsh plain elevation has risen because of sedi-mentation and the recent record of flooding provides anexplanation
Tijuana Estuary has a large watershed with highly erodablesoils and major slope failures that are visible both north andsouth of the USndashMexico border The 1993 floodwaters cut anentirely new river channel just inland of the estuary Thehuge volume of sediment that was mobilized could havecontributed to a rise in elevation on the marsh plain In-creased soil salinities (now averaging 60 ppt Figure 9) cor-
roborate a shift toward a higher and hence drier marshplain For intertidal species that are sensitive to 5ndash10-cm dif-ferences in elevation the accretion rates and associated salin-ities documented at Tijuana Estuary would seem to be fatalPersistent hypersaline conditions would reduce Sb and Se es-tablishment drier conditions would favor Sv and densercanopies of Sv would reduce the persistence of seedlings
If rising elevation and increasing salinity prevent reestab-lishment of Sb and Se then reversing these changes would re-quire a massive sediment removal effortmdashthe marsh plainwould have to be bulldozed to lower the elevation by 6ndash13 cmor more Such a disruption to the vegetation would not be al-lowed by resource managers because the plant canopy isused by light-footed clapper rails for cover and Beldingrsquos Sa-vannah sparrows for nesting
There is an alternative Although these recent physicalchanges are not reversible it is possible to excavate sedi-ments and restore areas where sediment loads have shiftedtopography well out of the tidal range Exactly this procedureis planned for up to 200 ha of former tidal wetlands in thesouthern arm of the estuary (Entrix et al 1991) The re-moval of sediments in the south arm can conceivably com-pensate for some of the accretion that is occurring in the es-tuary and in the region Attempts are also being made tocontrol the flow of sediments upstream
Irreversible biological changes When the full com-plement of species is present the community presumably hasmaximum biocomplexitymdashthat is a variety of interspecificinteractions and feedback mechanisms Its biodiversity wouldlead to biocomplexity It is possible that the marshrsquos bio-complexity helped sustain its biodiversity If some interactionsfacilitated rare species and others kept dominants from out-competing subordinates then more species would persist Inthat event losing some species would make the communityless able to support all the naturally occurring species bothplant and animalWhere dense canopy cover reduces seedlingrecruitment any species that reduces overall biomass above-ground could reduce shading and enhance recruitment andspecies richness The greenhouse experiments with repli-cated assemblages suggest such a species effect
We explored effects of selected species by comparing mi-crocosms with and without such a species We found threespecies (Sv Fs and Jc) that tended to increase whole-potbiomass (a typical effect of productive species) and one (Tc)that tended to decrease whole-pot biomass (an unusual effect)Assemblages with Tc had lower shoot biomass than assem-blages without it Tc produces little biomass when grownalone it is short with an unimpressive canopy Althoughlow in biomass Tc accumulates a disproportionate concen-tration of N twice that for Sv In the microcosms total ash-free biomass values for Tc and Sv were significantly negativelycorrelated (r = ndash043 p lt 01 n = 72 sextets) Despite itssmall size Tc seems capable of reducing the biomass of SvField data from Bahiacutea San Quintiacuten also suggest a negative re-lationship between Tc and Sv cover (r = ndash0133 p = 05 n =224) We conclude that Tc has some mechanism for reduc-
1014 BioScience December 2001 Vol 51 No 12
Features
Table 4 Experimental attempt to reestablish Sb and Seat Tijuana Estuary in 1998 (unpublished datacollected by Gabrielle Vivian-Smith Pacific EstuarineResearch Lab) Species codes are Sb Salicorniabigelovii Se Suaeda esteroa The ldquoseeds addedrdquotreatment had 2000 seeds added per speciesldquoRecruitmentrdquo is the total number of seedlings found
Treatment Recruitment
No seeds added 0 Sb 0 SeSeeds added canopy left intact 92 Sb 0 SeSeeds added canopy cleared 180 Sb 2 Se
ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features

seedlings of other species thereafter recruits were allowedto persist
Seedlings were censused by Roberto Lindig-Cisneros(2001) canopy architecture was documented by Keer andZedler (forthcoming) and biomass and N accumulationwere assessed by J Callaway Because we were most con-cerned about the increasing ability of the site to retain Nwe assessed biomass and N accumulation in midwinter af-ter most plants had slowed growth Furthermore becausewe wanted to minimize destructive sampling we removedsmall root cores in year 2 and root and shoot samples at theend of year 3 Both biomass components were analyzed forabove- and belowground N content
We complemented the field study with an experiment inthe greenhouse where we were able to compare individual as-semblages We replicated all 15 randomly drawn trios and sex-
tets 8-fold all solos 16-fold and the common trioand sextet 16-fold for a total of 400 microcosmsPlanting treatments were maintained in the potsthrough vegetative growth and seedling recruit-ment but assemblage composition did not changeAll but 80 pots were grown for 2 years and as-sessed for canopy attributes and harvested for bio-mass and N content above and below ground
The importance of diversity to restorationResults from the field and greenhouse experimentssupport the general hypothesis that diversity mat-ters in restoration sites Our plantings with sixspecies enhanced recruitment made canopiesmore complex increased biomass and increasedN accumulation
Recruitment Only three species (Sv Sb andSe) recruited readily (Table 3) Of these only one(Sv) is widely distributed (Table 2) and available todisperse into restoration sites It germinates in sea-water and produces abundant seedlings in bothnatural and restored wetlands including our un-planted plots it does not need to be planted torestoration sites if a seed source exists nearby Theother two abundant recruiters Sb and Se are themarsh plainrsquos only short-lived species Personalobservation revealed that Se recruitment potentialwas much lower than that of Sb reflecting its lowerdensity in regional wetlands and differences inseedling mortality within this experiment Thefact that both recruited readily in the bare restora-tion site and poorly in the salt marsh where it oc-curred historically is evidence that the Tijuana Es-tuary marsh plain no longer provides suitablehabitat for these species The remaining five speciesdid not readily colonize either our planted or un-planted plots Their inability to recruit may be at-
1010 BioScience December 2001 Vol 51 No 12
Features
Table 3 Seedling recruitment of eight marsh-plainspecies into experimental plots Data are for the 1998growing season Note that the number of plantsintroduced to the site as seedlings in April 1997(column 2) was similar but only three speciesrecruited extensively Species codes are in Table 1
Code Number planted Number of recruits
Sv 840 17703Sb 735 15978Se 795 1668Tc 810 73Jc 840 33Bm 720 26Lc 795 24Fs 765 2
Figure 4 Coastal wetlands of southern California 1Tijuana Estuary 2Sweetwater Marsh 3 Mission Bay Reserve 4 Los Pentildeasquitos Lagoon 5San Dieguito Lagoon 6 San Elijo Lagoon 7 Batiquitos Lagoon 8 AguaHedionda Lagoon 9 Buena Vista Lagoon (brackish wetland) 10 SanLuis Rey River Mouth 11 Santa Margarita Estuary 12 Las FloresMarsh 13 San Mateo Marsh 14 Upper Newport Bay 15 Bolsa ChicaWetland 16 Anaheim Bay 17 Los Cerritos Marsh (mostly filled anddeveloped) 18 Ballona Wetland 19 Malibu Lagoon 20 Mugu Lagoon21 McGrath Lake 22 Santa Clara River Estuary 23 Ventura River(brackish) 24 Carpinteria Marsh 25 Goleta Slough 26 DeverauxLagoon
tributable to small seed sup-ply poor germination or un-suitable conditions for es-tablishment
Canopy architectureKeer and Zedler (forthcom-ing) found that all plotsrapidly achieved high coverbut that canopy height andlayering were slow to de-velop Cover is a poor mea-sure of canopy complexity itdoes not differentiate speciesor species-richness treat-ments Maximum heightswere greatest for trios andsextets reaching about 60cm by year 3 Sv grew thetallest in the first 18 months of study followed by Sb and SeLayering (the number of times a vertical rod encountersplant parts as it is lowered through the canopy) was greaterfor trios and sextets both of which achieved three layerswithin 18 months The effects of individual species werelikely to be seen early in the development of the plots Fs andSe had six and five layers respectively with Jc at four layersLayering is hypothesized to enhance habitat for both arthro-pods and birds These canopies are far from fully developedreference marshes with monotypic Sv averaged 45 cm tall withseven layers and areas with multiple species were 55 cm talland had 13 layers (Keer and Zedler forthcoming)
Biomass and N accumulation In the field plots above-ground biomass averaged under 300 g per m2 for solos andover 400 g per m2 for sextets Belowground biomass also in-creased with species richness and over timemdashmeans doubledbetween years 2 and 3 (to 500g per m2 for sextets) The rapidincrease in belowground biomass is highly desirable for barerestoration sites To determine whether N crops increased wemeasured biomass and tissue N concentrations The result-ing estimates of accumulated N showed a higher crop for sex-tets than for unplanted and solo plots The rapid accumula-tion of N is beneficial for restoration sites that have coarse soiland where N is the limiting factor (a common problem in cre-ated and restored wetlands Langis et al 1991 Boyer andZedler 1996 1998 Zedler and Callaway 1999 2000 Boyer etal 2000) Our greenhouse experiment revealed similar pat-terns in a more benign environment (that is one with warmertemperatures) Total biomass was higher for sextets becausebelowground biomass increased with species richness As inthe field N crops tracked biomass with greater total N ac-cumulation and greater belowground accumulation at year2A species composition effectmdashnamely the chance inclusionof high biomass producers in the randomly drawn assem-blagemdashis indicated because Jc Fs and Lc solos all accumu-lated more N than the mean for sextets and Sv and Bm
solos both accumulated as much as the triosAlthough Tc Seand Sb solos all had below-average N crops Sb and Tc accu-mulated the most N per unit biomass
These findings were used to design the planting of the 8-ha ldquoFriendship Marshrdquo a $3 million project that aims to sup-port rare species of plants and animals (Figure 8) Located inthe south arm of Tijuana Estuary this site was opened to tidalflushing in February 2000 after floodborne sediments (1ndash2 m)were excavated and the historical marsh plain uncoveredBecause Sv dominates the nearby wetlands and readily es-tablishes on its own it was not introduced as seeds or plantsInstead we planted five species for which we had abundantseedlings (Fs Jc Bm Lc and Se) to help provide the functionsmentioned abovemdashnamely recruitment complex canopiesand biomass and N accumulation The protocol of not plant-ing Sv (to slow time to dominance) and adding multispeciesclusters (to accelerate the development of species-rich
December 2001 Vol 51 No 12 BioScience 1011
Features
Figure 5 San Elijo Lagoon San Diego County California
Figure 6 Historical patterns of species richness atTijuana Estuary data (means and standard errors) arefrom 025-m2 circular plots
canopies) is new to the region Previous marsh plain restora-tion efforts have focused on planting Sv
Although the field and greenhouse experiments taught usthat species-rich plots promote recruitment develop morecomplex canopies and accumulate more biomass and Nthe performance of sextets could be attributable to the com-bined performance of the six species or to the chance inclu-sion of a single high-performance species (a species compo-sition effect) Data from the replicated assemblages in thegreenhouse allowed us to evaluate these alternatives In the mi-crocosms species composition was stable over time (whereasrecruitment increased richness of solo plots in the field) Wecompared the eight species and found that Jc produced themost shoot biomass Fs the most root biomass and Sv themost total biomass while Tc had the highest rootshoot bio-mass But their rankings changed when ash-free biomass
was considered because Sv had the most ash per plant andSb the most ash per unit biomass For ash-free biomass Jc washighest for shoots Fs for roots and Jc for total with Tc stillhaving the highest rootshoot ratio The N concentration ofshoots was highest for Sb for roots it was highest for Tc Jchad the highest crop of N in its shoots and Fs the highest rootN crop while Jc had the highest total N crop Height was great-est for Se and layering was greatest for Fs Interestingly theregionrsquos ldquosuperplantrdquo Sv achieved its first place overall rank-ing not by excelling in multiple attributes but by scoring sec-ond or third in most attributes It scored highest only in to-tal biomass but it achieved high biomass by accumulating alarge volume of salts in its tissues in addition to accumulat-ing considerable carbon
This meta-analysis supports both alternatives Speciescomposition effects (Symstad et al 1998) were indicated forevery attribute considered (there is at least one species thatcan match the performance of the average sextet) but it is notthe same species for all response variables (that is all speciesare needed to achieve high levels of all functions) Further-more each species excelled in at least one attribute Thus weconclude that diversity matters a great deal and the onlyway to ensure that multiple functions are provided is to plantand sustain all species Retaining diversity results in less riskthat functions will be eliminated The planting of species-richassemblages in restoration sites would help ensure that morefunctions are restored
There is support for the suggestion that attributes measuredin the greenhouse translate into performance in the field Thespecies that performed best in the greenhouse are most wide-spread geographically Overall the eight species ranked in vir-tually the same order as in their distributional ranking (thatis the number of southern California wetlands in which theyoccur Table 1) The top three performers in the greenhousewere Sv Jc and Fs and they all occur in 22 or 23 wetlandsLc ranked fourth in greenhouse performance and fourth inregional distribution occurring in 13 wetlands in the historicalrecord and in 11 wetlands in 1998 The others ranked fifththrough eighth in performance they are the most restrictedin their distributions now occurring in only six to eight sitesJc Fs and Sv are all highly productive long-lived perennialsthat reproduce vegetatively and form monotypic patches(Zedler et al 1980 Ferren 1985) It is not surprising thatthey persist in wetlands that experience closure to tidal in-fluence
What functions were lost when short-lived species dropped out of Tijuana Estuary Sb and Se are short-lived halophytes that do not reproducevegetatively Their roles in the ecosystem are arguably uniquefrom these perspectives alone Their annual recruitment ofseedlings following winter rainfall adds seasonality to thecanopy and soil perhaps changing food availability bothabove and below ground Regrettably these functions have notbeen quantified
1012 BioScience December 2001 Vol 51 No 12
Features
Figure 7 Adaptive restoration exemplified by theexperimental planting of 87 2-by-2-meter plots with 0 13 and 6 species in a newly excavated marsh plain atTijuana Estuary Photograph taken in June 1997 2months after planting greenhouse-grown seedlings (90per plot)
As our field data docu-ment Sv Sb and Se are allgood recruiters capable offilling canopy openings withseedlings and rapidly reveg-etating bare sites Howeverthe 1998 recruitment data(Lindig-Cisneros 2001) showthat Sb and Se producedseedlings earlier than Sv Inearly April 1998 there were2199 Sb and 220 Se seedlingsbut only 89 Sv seedlings Inour experimental design wepulled seedlings of invadingspecies to retain assemblagecomposition in 1997ndash1998so there was ample oppor-tunity for Sv seedlings to re-cruit later In early May 1998there were 9261 Sv seedlingsbut only 842 Sb and 50 Se re-cruitment then declined forall species over the late Mayand July sampling dates despite the fact that seedlings werebeing pulled and space remained available for establishmentWe infer from these results that Sb and Se have important rolesin filling gaps that is space opened late in the year is filled bySb and Se the following winter and spring
Both Sb and Se are capable of rapid height growth asdemonstrated in the greenhouse Se might be the first speciesto provide perch sites for the endangered Beldingrsquos Savannahsparrow which defends its nesting territories by singing whileperching on the tallest plants (Powell 1993) The verticalstructure afforded by Se differs from that of Sv in having amore horizontal branching angle and more lay-ers attributes that may increase its attraction ofsmall birds It is not uncommon to see sparrowsperching on tall branches of Se in the fieldwhether the plants are alive or dead We concludethat the loss of Sb and Se could impair the com-munityrsquos ability to recover from disturbanceand to support an endangered bird These func-tions are not entirely eliminated by the demiseof Sb and Se but the ecosystemrsquos functional po-tential might not be realized in their absence
Is the loss of species reversibleTijuana Estuaryrsquos current salt marsh communityis certainly different from that prior to 1984when an open canopy supported thousands ofSb seedlings per square meter and Se was com-mon throughout the tidal marsh Today thesespecies persist in a tiny fraction of the area theyoccupied historicallyAs short-lived species bothSb (annual) and Se (lives only a few years)
depend on recruitment for their persistence while all of theother marsh plain species reproduce vegetatively Seedlingswere young when Tijuana Estuary became nontidal in April1984 and one likely cause of their mortality was the dryingof surface soil and rise in salinity to an average of greater than100 ppt (Figure 9) Moderate hypersalinity (40ndash45 ppt) is typ-ical for marsh plain soils but extreme hypersalinity is morecharacteristic of higher elevations with less frequent tidal in-undation (Noe and Zedler 2001) Sb and Se can germinate inseawater but higher salinities reduce germination rates (Zedlerand Beare 1986) Had their seed banks persisted through the
December 2001 Vol 51 No 12 BioScience 1013
Features
Figure 8 An 8-ha marsh plain excavation at Tijuana Estuary Because of the large size of thissite plantings emphasized species that would not likely recruit on their own
Figure 9 Soil salinity at Tijuana Estuary (data from the Pacific EstuarineResearch Lab) Means before A on the x-axis include Spartina marsh andthe marsh plain means after A are marsh plain only Means before B arefor interstitial soil water after B are for soil pastes (adding deionized waterunderestimates interstitial salinity)
period of tidal closure Sb and Se should have germinated in1985 they did not
A recent reintroduction experiment by Vivian-Smith andZedler (2001) indicated that neither species retains a marshplain seed bank at Tijuana Estuary Vivian-Smith added 2000seeds of each species in 1998 Sb produced 272 seedlings andSe produced 2 with seedlings establishing only where seedswere added Also the current Sv-dominated canopy reducedrecruitment with Sb seedling density twice as high where thecanopy was removed (Table 4) Hence we learned that a lackof seeds and shading by Sv restricted recovery of the Sb pop-ulation And because H N Morzaria-Luna (University ofWisconsinndashMadison unpublished data) could not locate ei-ther species during her search in 2001 we suspect that changesin both the physical and biological conditions of this marshfurther inhibit recovery We explore each of these in turn
Irreversible physical changes According to recent in-formation both elevation and soil salinities have changed inthe 16 years since tidal closure A study of marsh accretion rates(Weis et al forthcoming) using cesium-137 (137Cs a globallydispersed radioisotope) suggests that since 1984 (the yearwhen Sb and Se declined) the marsh plain elevation has risen13 cm based on two soil cores At three additional locationsbenchmarks placed on the marsh plain in the 1970s by the USArmy Corps of Engineers were relocated there we have mea-sured an average of 6 cm accretion since 1984 (Morzaria-Lunaunpublished data) Further sedimentation data from TijuanaEstuary corroborate this rapid rise in elevation Cahoon andcolleagues (1996) measured 20ndash85 (mean 59) cm accretionin the S foliosa marsh between 1992 and 1993 and in anearby tidal pond (lower elevation) Ward (2000) found4ndash127 (mean 88) cm of sediment accretion from 1997 to1998 The marsh plain elevation has risen because of sedi-mentation and the recent record of flooding provides anexplanation
Tijuana Estuary has a large watershed with highly erodablesoils and major slope failures that are visible both north andsouth of the USndashMexico border The 1993 floodwaters cut anentirely new river channel just inland of the estuary Thehuge volume of sediment that was mobilized could havecontributed to a rise in elevation on the marsh plain In-creased soil salinities (now averaging 60 ppt Figure 9) cor-
roborate a shift toward a higher and hence drier marshplain For intertidal species that are sensitive to 5ndash10-cm dif-ferences in elevation the accretion rates and associated salin-ities documented at Tijuana Estuary would seem to be fatalPersistent hypersaline conditions would reduce Sb and Se es-tablishment drier conditions would favor Sv and densercanopies of Sv would reduce the persistence of seedlings
If rising elevation and increasing salinity prevent reestab-lishment of Sb and Se then reversing these changes would re-quire a massive sediment removal effortmdashthe marsh plainwould have to be bulldozed to lower the elevation by 6ndash13 cmor more Such a disruption to the vegetation would not be al-lowed by resource managers because the plant canopy isused by light-footed clapper rails for cover and Beldingrsquos Sa-vannah sparrows for nesting
There is an alternative Although these recent physicalchanges are not reversible it is possible to excavate sedi-ments and restore areas where sediment loads have shiftedtopography well out of the tidal range Exactly this procedureis planned for up to 200 ha of former tidal wetlands in thesouthern arm of the estuary (Entrix et al 1991) The re-moval of sediments in the south arm can conceivably com-pensate for some of the accretion that is occurring in the es-tuary and in the region Attempts are also being made tocontrol the flow of sediments upstream
Irreversible biological changes When the full com-plement of species is present the community presumably hasmaximum biocomplexitymdashthat is a variety of interspecificinteractions and feedback mechanisms Its biodiversity wouldlead to biocomplexity It is possible that the marshrsquos bio-complexity helped sustain its biodiversity If some interactionsfacilitated rare species and others kept dominants from out-competing subordinates then more species would persist Inthat event losing some species would make the communityless able to support all the naturally occurring species bothplant and animalWhere dense canopy cover reduces seedlingrecruitment any species that reduces overall biomass above-ground could reduce shading and enhance recruitment andspecies richness The greenhouse experiments with repli-cated assemblages suggest such a species effect
We explored effects of selected species by comparing mi-crocosms with and without such a species We found threespecies (Sv Fs and Jc) that tended to increase whole-potbiomass (a typical effect of productive species) and one (Tc)that tended to decrease whole-pot biomass (an unusual effect)Assemblages with Tc had lower shoot biomass than assem-blages without it Tc produces little biomass when grownalone it is short with an unimpressive canopy Althoughlow in biomass Tc accumulates a disproportionate concen-tration of N twice that for Sv In the microcosms total ash-free biomass values for Tc and Sv were significantly negativelycorrelated (r = ndash043 p lt 01 n = 72 sextets) Despite itssmall size Tc seems capable of reducing the biomass of SvField data from Bahiacutea San Quintiacuten also suggest a negative re-lationship between Tc and Sv cover (r = ndash0133 p = 05 n =224) We conclude that Tc has some mechanism for reduc-
1014 BioScience December 2001 Vol 51 No 12
Features
Table 4 Experimental attempt to reestablish Sb and Seat Tijuana Estuary in 1998 (unpublished datacollected by Gabrielle Vivian-Smith Pacific EstuarineResearch Lab) Species codes are Sb Salicorniabigelovii Se Suaeda esteroa The ldquoseeds addedrdquotreatment had 2000 seeds added per speciesldquoRecruitmentrdquo is the total number of seedlings found
Treatment Recruitment
No seeds added 0 Sb 0 SeSeeds added canopy left intact 92 Sb 0 SeSeeds added canopy cleared 180 Sb 2 Se
ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features
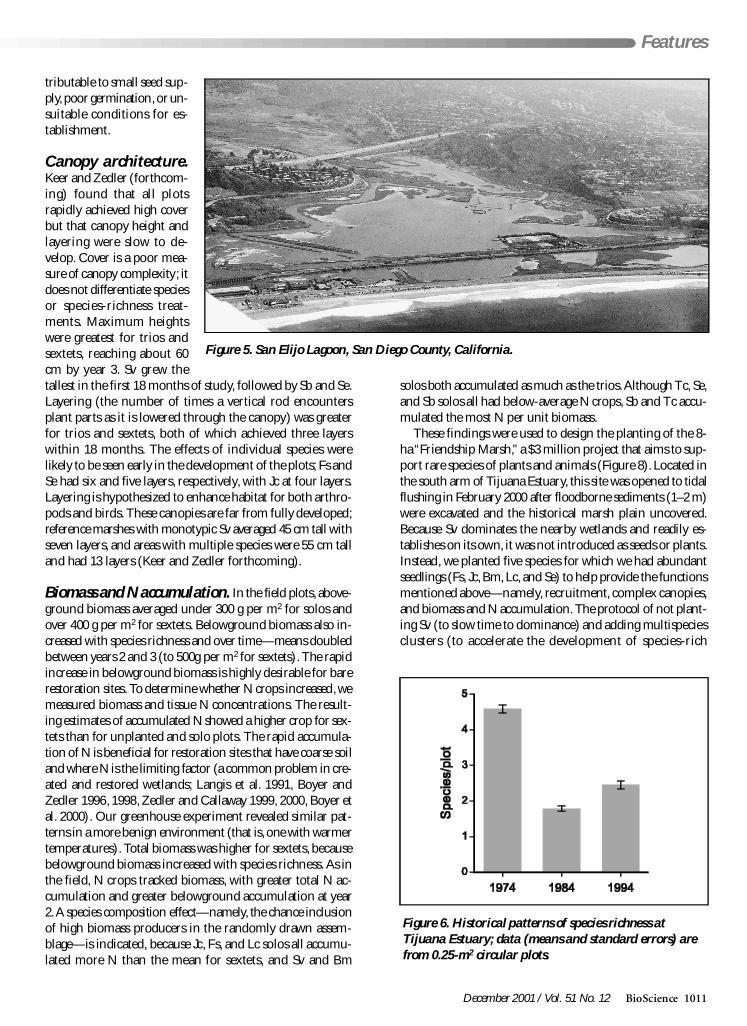
tributable to small seed sup-ply poor germination or un-suitable conditions for es-tablishment
Canopy architectureKeer and Zedler (forthcom-ing) found that all plotsrapidly achieved high coverbut that canopy height andlayering were slow to de-velop Cover is a poor mea-sure of canopy complexity itdoes not differentiate speciesor species-richness treat-ments Maximum heightswere greatest for trios andsextets reaching about 60cm by year 3 Sv grew thetallest in the first 18 months of study followed by Sb and SeLayering (the number of times a vertical rod encountersplant parts as it is lowered through the canopy) was greaterfor trios and sextets both of which achieved three layerswithin 18 months The effects of individual species werelikely to be seen early in the development of the plots Fs andSe had six and five layers respectively with Jc at four layersLayering is hypothesized to enhance habitat for both arthro-pods and birds These canopies are far from fully developedreference marshes with monotypic Sv averaged 45 cm tall withseven layers and areas with multiple species were 55 cm talland had 13 layers (Keer and Zedler forthcoming)
Biomass and N accumulation In the field plots above-ground biomass averaged under 300 g per m2 for solos andover 400 g per m2 for sextets Belowground biomass also in-creased with species richness and over timemdashmeans doubledbetween years 2 and 3 (to 500g per m2 for sextets) The rapidincrease in belowground biomass is highly desirable for barerestoration sites To determine whether N crops increased wemeasured biomass and tissue N concentrations The result-ing estimates of accumulated N showed a higher crop for sex-tets than for unplanted and solo plots The rapid accumula-tion of N is beneficial for restoration sites that have coarse soiland where N is the limiting factor (a common problem in cre-ated and restored wetlands Langis et al 1991 Boyer andZedler 1996 1998 Zedler and Callaway 1999 2000 Boyer etal 2000) Our greenhouse experiment revealed similar pat-terns in a more benign environment (that is one with warmertemperatures) Total biomass was higher for sextets becausebelowground biomass increased with species richness As inthe field N crops tracked biomass with greater total N ac-cumulation and greater belowground accumulation at year2A species composition effectmdashnamely the chance inclusionof high biomass producers in the randomly drawn assem-blagemdashis indicated because Jc Fs and Lc solos all accumu-lated more N than the mean for sextets and Sv and Bm
solos both accumulated as much as the triosAlthough Tc Seand Sb solos all had below-average N crops Sb and Tc accu-mulated the most N per unit biomass
These findings were used to design the planting of the 8-ha ldquoFriendship Marshrdquo a $3 million project that aims to sup-port rare species of plants and animals (Figure 8) Located inthe south arm of Tijuana Estuary this site was opened to tidalflushing in February 2000 after floodborne sediments (1ndash2 m)were excavated and the historical marsh plain uncoveredBecause Sv dominates the nearby wetlands and readily es-tablishes on its own it was not introduced as seeds or plantsInstead we planted five species for which we had abundantseedlings (Fs Jc Bm Lc and Se) to help provide the functionsmentioned abovemdashnamely recruitment complex canopiesand biomass and N accumulation The protocol of not plant-ing Sv (to slow time to dominance) and adding multispeciesclusters (to accelerate the development of species-rich
December 2001 Vol 51 No 12 BioScience 1011
Features
Figure 5 San Elijo Lagoon San Diego County California
Figure 6 Historical patterns of species richness atTijuana Estuary data (means and standard errors) arefrom 025-m2 circular plots
canopies) is new to the region Previous marsh plain restora-tion efforts have focused on planting Sv
Although the field and greenhouse experiments taught usthat species-rich plots promote recruitment develop morecomplex canopies and accumulate more biomass and Nthe performance of sextets could be attributable to the com-bined performance of the six species or to the chance inclu-sion of a single high-performance species (a species compo-sition effect) Data from the replicated assemblages in thegreenhouse allowed us to evaluate these alternatives In the mi-crocosms species composition was stable over time (whereasrecruitment increased richness of solo plots in the field) Wecompared the eight species and found that Jc produced themost shoot biomass Fs the most root biomass and Sv themost total biomass while Tc had the highest rootshoot bio-mass But their rankings changed when ash-free biomass
was considered because Sv had the most ash per plant andSb the most ash per unit biomass For ash-free biomass Jc washighest for shoots Fs for roots and Jc for total with Tc stillhaving the highest rootshoot ratio The N concentration ofshoots was highest for Sb for roots it was highest for Tc Jchad the highest crop of N in its shoots and Fs the highest rootN crop while Jc had the highest total N crop Height was great-est for Se and layering was greatest for Fs Interestingly theregionrsquos ldquosuperplantrdquo Sv achieved its first place overall rank-ing not by excelling in multiple attributes but by scoring sec-ond or third in most attributes It scored highest only in to-tal biomass but it achieved high biomass by accumulating alarge volume of salts in its tissues in addition to accumulat-ing considerable carbon
This meta-analysis supports both alternatives Speciescomposition effects (Symstad et al 1998) were indicated forevery attribute considered (there is at least one species thatcan match the performance of the average sextet) but it is notthe same species for all response variables (that is all speciesare needed to achieve high levels of all functions) Further-more each species excelled in at least one attribute Thus weconclude that diversity matters a great deal and the onlyway to ensure that multiple functions are provided is to plantand sustain all species Retaining diversity results in less riskthat functions will be eliminated The planting of species-richassemblages in restoration sites would help ensure that morefunctions are restored
There is support for the suggestion that attributes measuredin the greenhouse translate into performance in the field Thespecies that performed best in the greenhouse are most wide-spread geographically Overall the eight species ranked in vir-tually the same order as in their distributional ranking (thatis the number of southern California wetlands in which theyoccur Table 1) The top three performers in the greenhousewere Sv Jc and Fs and they all occur in 22 or 23 wetlandsLc ranked fourth in greenhouse performance and fourth inregional distribution occurring in 13 wetlands in the historicalrecord and in 11 wetlands in 1998 The others ranked fifththrough eighth in performance they are the most restrictedin their distributions now occurring in only six to eight sitesJc Fs and Sv are all highly productive long-lived perennialsthat reproduce vegetatively and form monotypic patches(Zedler et al 1980 Ferren 1985) It is not surprising thatthey persist in wetlands that experience closure to tidal in-fluence
What functions were lost when short-lived species dropped out of Tijuana Estuary Sb and Se are short-lived halophytes that do not reproducevegetatively Their roles in the ecosystem are arguably uniquefrom these perspectives alone Their annual recruitment ofseedlings following winter rainfall adds seasonality to thecanopy and soil perhaps changing food availability bothabove and below ground Regrettably these functions have notbeen quantified
1012 BioScience December 2001 Vol 51 No 12
Features
Figure 7 Adaptive restoration exemplified by theexperimental planting of 87 2-by-2-meter plots with 0 13 and 6 species in a newly excavated marsh plain atTijuana Estuary Photograph taken in June 1997 2months after planting greenhouse-grown seedlings (90per plot)
As our field data docu-ment Sv Sb and Se are allgood recruiters capable offilling canopy openings withseedlings and rapidly reveg-etating bare sites Howeverthe 1998 recruitment data(Lindig-Cisneros 2001) showthat Sb and Se producedseedlings earlier than Sv Inearly April 1998 there were2199 Sb and 220 Se seedlingsbut only 89 Sv seedlings Inour experimental design wepulled seedlings of invadingspecies to retain assemblagecomposition in 1997ndash1998so there was ample oppor-tunity for Sv seedlings to re-cruit later In early May 1998there were 9261 Sv seedlingsbut only 842 Sb and 50 Se re-cruitment then declined forall species over the late Mayand July sampling dates despite the fact that seedlings werebeing pulled and space remained available for establishmentWe infer from these results that Sb and Se have important rolesin filling gaps that is space opened late in the year is filled bySb and Se the following winter and spring
Both Sb and Se are capable of rapid height growth asdemonstrated in the greenhouse Se might be the first speciesto provide perch sites for the endangered Beldingrsquos Savannahsparrow which defends its nesting territories by singing whileperching on the tallest plants (Powell 1993) The verticalstructure afforded by Se differs from that of Sv in having amore horizontal branching angle and more lay-ers attributes that may increase its attraction ofsmall birds It is not uncommon to see sparrowsperching on tall branches of Se in the fieldwhether the plants are alive or dead We concludethat the loss of Sb and Se could impair the com-munityrsquos ability to recover from disturbanceand to support an endangered bird These func-tions are not entirely eliminated by the demiseof Sb and Se but the ecosystemrsquos functional po-tential might not be realized in their absence
Is the loss of species reversibleTijuana Estuaryrsquos current salt marsh communityis certainly different from that prior to 1984when an open canopy supported thousands ofSb seedlings per square meter and Se was com-mon throughout the tidal marsh Today thesespecies persist in a tiny fraction of the area theyoccupied historicallyAs short-lived species bothSb (annual) and Se (lives only a few years)
depend on recruitment for their persistence while all of theother marsh plain species reproduce vegetatively Seedlingswere young when Tijuana Estuary became nontidal in April1984 and one likely cause of their mortality was the dryingof surface soil and rise in salinity to an average of greater than100 ppt (Figure 9) Moderate hypersalinity (40ndash45 ppt) is typ-ical for marsh plain soils but extreme hypersalinity is morecharacteristic of higher elevations with less frequent tidal in-undation (Noe and Zedler 2001) Sb and Se can germinate inseawater but higher salinities reduce germination rates (Zedlerand Beare 1986) Had their seed banks persisted through the
December 2001 Vol 51 No 12 BioScience 1013
Features
Figure 8 An 8-ha marsh plain excavation at Tijuana Estuary Because of the large size of thissite plantings emphasized species that would not likely recruit on their own
Figure 9 Soil salinity at Tijuana Estuary (data from the Pacific EstuarineResearch Lab) Means before A on the x-axis include Spartina marsh andthe marsh plain means after A are marsh plain only Means before B arefor interstitial soil water after B are for soil pastes (adding deionized waterunderestimates interstitial salinity)
period of tidal closure Sb and Se should have germinated in1985 they did not
A recent reintroduction experiment by Vivian-Smith andZedler (2001) indicated that neither species retains a marshplain seed bank at Tijuana Estuary Vivian-Smith added 2000seeds of each species in 1998 Sb produced 272 seedlings andSe produced 2 with seedlings establishing only where seedswere added Also the current Sv-dominated canopy reducedrecruitment with Sb seedling density twice as high where thecanopy was removed (Table 4) Hence we learned that a lackof seeds and shading by Sv restricted recovery of the Sb pop-ulation And because H N Morzaria-Luna (University ofWisconsinndashMadison unpublished data) could not locate ei-ther species during her search in 2001 we suspect that changesin both the physical and biological conditions of this marshfurther inhibit recovery We explore each of these in turn
Irreversible physical changes According to recent in-formation both elevation and soil salinities have changed inthe 16 years since tidal closure A study of marsh accretion rates(Weis et al forthcoming) using cesium-137 (137Cs a globallydispersed radioisotope) suggests that since 1984 (the yearwhen Sb and Se declined) the marsh plain elevation has risen13 cm based on two soil cores At three additional locationsbenchmarks placed on the marsh plain in the 1970s by the USArmy Corps of Engineers were relocated there we have mea-sured an average of 6 cm accretion since 1984 (Morzaria-Lunaunpublished data) Further sedimentation data from TijuanaEstuary corroborate this rapid rise in elevation Cahoon andcolleagues (1996) measured 20ndash85 (mean 59) cm accretionin the S foliosa marsh between 1992 and 1993 and in anearby tidal pond (lower elevation) Ward (2000) found4ndash127 (mean 88) cm of sediment accretion from 1997 to1998 The marsh plain elevation has risen because of sedi-mentation and the recent record of flooding provides anexplanation
Tijuana Estuary has a large watershed with highly erodablesoils and major slope failures that are visible both north andsouth of the USndashMexico border The 1993 floodwaters cut anentirely new river channel just inland of the estuary Thehuge volume of sediment that was mobilized could havecontributed to a rise in elevation on the marsh plain In-creased soil salinities (now averaging 60 ppt Figure 9) cor-
roborate a shift toward a higher and hence drier marshplain For intertidal species that are sensitive to 5ndash10-cm dif-ferences in elevation the accretion rates and associated salin-ities documented at Tijuana Estuary would seem to be fatalPersistent hypersaline conditions would reduce Sb and Se es-tablishment drier conditions would favor Sv and densercanopies of Sv would reduce the persistence of seedlings
If rising elevation and increasing salinity prevent reestab-lishment of Sb and Se then reversing these changes would re-quire a massive sediment removal effortmdashthe marsh plainwould have to be bulldozed to lower the elevation by 6ndash13 cmor more Such a disruption to the vegetation would not be al-lowed by resource managers because the plant canopy isused by light-footed clapper rails for cover and Beldingrsquos Sa-vannah sparrows for nesting
There is an alternative Although these recent physicalchanges are not reversible it is possible to excavate sedi-ments and restore areas where sediment loads have shiftedtopography well out of the tidal range Exactly this procedureis planned for up to 200 ha of former tidal wetlands in thesouthern arm of the estuary (Entrix et al 1991) The re-moval of sediments in the south arm can conceivably com-pensate for some of the accretion that is occurring in the es-tuary and in the region Attempts are also being made tocontrol the flow of sediments upstream
Irreversible biological changes When the full com-plement of species is present the community presumably hasmaximum biocomplexitymdashthat is a variety of interspecificinteractions and feedback mechanisms Its biodiversity wouldlead to biocomplexity It is possible that the marshrsquos bio-complexity helped sustain its biodiversity If some interactionsfacilitated rare species and others kept dominants from out-competing subordinates then more species would persist Inthat event losing some species would make the communityless able to support all the naturally occurring species bothplant and animalWhere dense canopy cover reduces seedlingrecruitment any species that reduces overall biomass above-ground could reduce shading and enhance recruitment andspecies richness The greenhouse experiments with repli-cated assemblages suggest such a species effect
We explored effects of selected species by comparing mi-crocosms with and without such a species We found threespecies (Sv Fs and Jc) that tended to increase whole-potbiomass (a typical effect of productive species) and one (Tc)that tended to decrease whole-pot biomass (an unusual effect)Assemblages with Tc had lower shoot biomass than assem-blages without it Tc produces little biomass when grownalone it is short with an unimpressive canopy Althoughlow in biomass Tc accumulates a disproportionate concen-tration of N twice that for Sv In the microcosms total ash-free biomass values for Tc and Sv were significantly negativelycorrelated (r = ndash043 p lt 01 n = 72 sextets) Despite itssmall size Tc seems capable of reducing the biomass of SvField data from Bahiacutea San Quintiacuten also suggest a negative re-lationship between Tc and Sv cover (r = ndash0133 p = 05 n =224) We conclude that Tc has some mechanism for reduc-
1014 BioScience December 2001 Vol 51 No 12
Features
Table 4 Experimental attempt to reestablish Sb and Seat Tijuana Estuary in 1998 (unpublished datacollected by Gabrielle Vivian-Smith Pacific EstuarineResearch Lab) Species codes are Sb Salicorniabigelovii Se Suaeda esteroa The ldquoseeds addedrdquotreatment had 2000 seeds added per speciesldquoRecruitmentrdquo is the total number of seedlings found
Treatment Recruitment
No seeds added 0 Sb 0 SeSeeds added canopy left intact 92 Sb 0 SeSeeds added canopy cleared 180 Sb 2 Se
ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features

canopies) is new to the region Previous marsh plain restora-tion efforts have focused on planting Sv
Although the field and greenhouse experiments taught usthat species-rich plots promote recruitment develop morecomplex canopies and accumulate more biomass and Nthe performance of sextets could be attributable to the com-bined performance of the six species or to the chance inclu-sion of a single high-performance species (a species compo-sition effect) Data from the replicated assemblages in thegreenhouse allowed us to evaluate these alternatives In the mi-crocosms species composition was stable over time (whereasrecruitment increased richness of solo plots in the field) Wecompared the eight species and found that Jc produced themost shoot biomass Fs the most root biomass and Sv themost total biomass while Tc had the highest rootshoot bio-mass But their rankings changed when ash-free biomass
was considered because Sv had the most ash per plant andSb the most ash per unit biomass For ash-free biomass Jc washighest for shoots Fs for roots and Jc for total with Tc stillhaving the highest rootshoot ratio The N concentration ofshoots was highest for Sb for roots it was highest for Tc Jchad the highest crop of N in its shoots and Fs the highest rootN crop while Jc had the highest total N crop Height was great-est for Se and layering was greatest for Fs Interestingly theregionrsquos ldquosuperplantrdquo Sv achieved its first place overall rank-ing not by excelling in multiple attributes but by scoring sec-ond or third in most attributes It scored highest only in to-tal biomass but it achieved high biomass by accumulating alarge volume of salts in its tissues in addition to accumulat-ing considerable carbon
This meta-analysis supports both alternatives Speciescomposition effects (Symstad et al 1998) were indicated forevery attribute considered (there is at least one species thatcan match the performance of the average sextet) but it is notthe same species for all response variables (that is all speciesare needed to achieve high levels of all functions) Further-more each species excelled in at least one attribute Thus weconclude that diversity matters a great deal and the onlyway to ensure that multiple functions are provided is to plantand sustain all species Retaining diversity results in less riskthat functions will be eliminated The planting of species-richassemblages in restoration sites would help ensure that morefunctions are restored
There is support for the suggestion that attributes measuredin the greenhouse translate into performance in the field Thespecies that performed best in the greenhouse are most wide-spread geographically Overall the eight species ranked in vir-tually the same order as in their distributional ranking (thatis the number of southern California wetlands in which theyoccur Table 1) The top three performers in the greenhousewere Sv Jc and Fs and they all occur in 22 or 23 wetlandsLc ranked fourth in greenhouse performance and fourth inregional distribution occurring in 13 wetlands in the historicalrecord and in 11 wetlands in 1998 The others ranked fifththrough eighth in performance they are the most restrictedin their distributions now occurring in only six to eight sitesJc Fs and Sv are all highly productive long-lived perennialsthat reproduce vegetatively and form monotypic patches(Zedler et al 1980 Ferren 1985) It is not surprising thatthey persist in wetlands that experience closure to tidal in-fluence
What functions were lost when short-lived species dropped out of Tijuana Estuary Sb and Se are short-lived halophytes that do not reproducevegetatively Their roles in the ecosystem are arguably uniquefrom these perspectives alone Their annual recruitment ofseedlings following winter rainfall adds seasonality to thecanopy and soil perhaps changing food availability bothabove and below ground Regrettably these functions have notbeen quantified
1012 BioScience December 2001 Vol 51 No 12
Features
Figure 7 Adaptive restoration exemplified by theexperimental planting of 87 2-by-2-meter plots with 0 13 and 6 species in a newly excavated marsh plain atTijuana Estuary Photograph taken in June 1997 2months after planting greenhouse-grown seedlings (90per plot)
As our field data docu-ment Sv Sb and Se are allgood recruiters capable offilling canopy openings withseedlings and rapidly reveg-etating bare sites Howeverthe 1998 recruitment data(Lindig-Cisneros 2001) showthat Sb and Se producedseedlings earlier than Sv Inearly April 1998 there were2199 Sb and 220 Se seedlingsbut only 89 Sv seedlings Inour experimental design wepulled seedlings of invadingspecies to retain assemblagecomposition in 1997ndash1998so there was ample oppor-tunity for Sv seedlings to re-cruit later In early May 1998there were 9261 Sv seedlingsbut only 842 Sb and 50 Se re-cruitment then declined forall species over the late Mayand July sampling dates despite the fact that seedlings werebeing pulled and space remained available for establishmentWe infer from these results that Sb and Se have important rolesin filling gaps that is space opened late in the year is filled bySb and Se the following winter and spring
Both Sb and Se are capable of rapid height growth asdemonstrated in the greenhouse Se might be the first speciesto provide perch sites for the endangered Beldingrsquos Savannahsparrow which defends its nesting territories by singing whileperching on the tallest plants (Powell 1993) The verticalstructure afforded by Se differs from that of Sv in having amore horizontal branching angle and more lay-ers attributes that may increase its attraction ofsmall birds It is not uncommon to see sparrowsperching on tall branches of Se in the fieldwhether the plants are alive or dead We concludethat the loss of Sb and Se could impair the com-munityrsquos ability to recover from disturbanceand to support an endangered bird These func-tions are not entirely eliminated by the demiseof Sb and Se but the ecosystemrsquos functional po-tential might not be realized in their absence
Is the loss of species reversibleTijuana Estuaryrsquos current salt marsh communityis certainly different from that prior to 1984when an open canopy supported thousands ofSb seedlings per square meter and Se was com-mon throughout the tidal marsh Today thesespecies persist in a tiny fraction of the area theyoccupied historicallyAs short-lived species bothSb (annual) and Se (lives only a few years)
depend on recruitment for their persistence while all of theother marsh plain species reproduce vegetatively Seedlingswere young when Tijuana Estuary became nontidal in April1984 and one likely cause of their mortality was the dryingof surface soil and rise in salinity to an average of greater than100 ppt (Figure 9) Moderate hypersalinity (40ndash45 ppt) is typ-ical for marsh plain soils but extreme hypersalinity is morecharacteristic of higher elevations with less frequent tidal in-undation (Noe and Zedler 2001) Sb and Se can germinate inseawater but higher salinities reduce germination rates (Zedlerand Beare 1986) Had their seed banks persisted through the
December 2001 Vol 51 No 12 BioScience 1013
Features
Figure 8 An 8-ha marsh plain excavation at Tijuana Estuary Because of the large size of thissite plantings emphasized species that would not likely recruit on their own
Figure 9 Soil salinity at Tijuana Estuary (data from the Pacific EstuarineResearch Lab) Means before A on the x-axis include Spartina marsh andthe marsh plain means after A are marsh plain only Means before B arefor interstitial soil water after B are for soil pastes (adding deionized waterunderestimates interstitial salinity)
period of tidal closure Sb and Se should have germinated in1985 they did not
A recent reintroduction experiment by Vivian-Smith andZedler (2001) indicated that neither species retains a marshplain seed bank at Tijuana Estuary Vivian-Smith added 2000seeds of each species in 1998 Sb produced 272 seedlings andSe produced 2 with seedlings establishing only where seedswere added Also the current Sv-dominated canopy reducedrecruitment with Sb seedling density twice as high where thecanopy was removed (Table 4) Hence we learned that a lackof seeds and shading by Sv restricted recovery of the Sb pop-ulation And because H N Morzaria-Luna (University ofWisconsinndashMadison unpublished data) could not locate ei-ther species during her search in 2001 we suspect that changesin both the physical and biological conditions of this marshfurther inhibit recovery We explore each of these in turn
Irreversible physical changes According to recent in-formation both elevation and soil salinities have changed inthe 16 years since tidal closure A study of marsh accretion rates(Weis et al forthcoming) using cesium-137 (137Cs a globallydispersed radioisotope) suggests that since 1984 (the yearwhen Sb and Se declined) the marsh plain elevation has risen13 cm based on two soil cores At three additional locationsbenchmarks placed on the marsh plain in the 1970s by the USArmy Corps of Engineers were relocated there we have mea-sured an average of 6 cm accretion since 1984 (Morzaria-Lunaunpublished data) Further sedimentation data from TijuanaEstuary corroborate this rapid rise in elevation Cahoon andcolleagues (1996) measured 20ndash85 (mean 59) cm accretionin the S foliosa marsh between 1992 and 1993 and in anearby tidal pond (lower elevation) Ward (2000) found4ndash127 (mean 88) cm of sediment accretion from 1997 to1998 The marsh plain elevation has risen because of sedi-mentation and the recent record of flooding provides anexplanation
Tijuana Estuary has a large watershed with highly erodablesoils and major slope failures that are visible both north andsouth of the USndashMexico border The 1993 floodwaters cut anentirely new river channel just inland of the estuary Thehuge volume of sediment that was mobilized could havecontributed to a rise in elevation on the marsh plain In-creased soil salinities (now averaging 60 ppt Figure 9) cor-
roborate a shift toward a higher and hence drier marshplain For intertidal species that are sensitive to 5ndash10-cm dif-ferences in elevation the accretion rates and associated salin-ities documented at Tijuana Estuary would seem to be fatalPersistent hypersaline conditions would reduce Sb and Se es-tablishment drier conditions would favor Sv and densercanopies of Sv would reduce the persistence of seedlings
If rising elevation and increasing salinity prevent reestab-lishment of Sb and Se then reversing these changes would re-quire a massive sediment removal effortmdashthe marsh plainwould have to be bulldozed to lower the elevation by 6ndash13 cmor more Such a disruption to the vegetation would not be al-lowed by resource managers because the plant canopy isused by light-footed clapper rails for cover and Beldingrsquos Sa-vannah sparrows for nesting
There is an alternative Although these recent physicalchanges are not reversible it is possible to excavate sedi-ments and restore areas where sediment loads have shiftedtopography well out of the tidal range Exactly this procedureis planned for up to 200 ha of former tidal wetlands in thesouthern arm of the estuary (Entrix et al 1991) The re-moval of sediments in the south arm can conceivably com-pensate for some of the accretion that is occurring in the es-tuary and in the region Attempts are also being made tocontrol the flow of sediments upstream
Irreversible biological changes When the full com-plement of species is present the community presumably hasmaximum biocomplexitymdashthat is a variety of interspecificinteractions and feedback mechanisms Its biodiversity wouldlead to biocomplexity It is possible that the marshrsquos bio-complexity helped sustain its biodiversity If some interactionsfacilitated rare species and others kept dominants from out-competing subordinates then more species would persist Inthat event losing some species would make the communityless able to support all the naturally occurring species bothplant and animalWhere dense canopy cover reduces seedlingrecruitment any species that reduces overall biomass above-ground could reduce shading and enhance recruitment andspecies richness The greenhouse experiments with repli-cated assemblages suggest such a species effect
We explored effects of selected species by comparing mi-crocosms with and without such a species We found threespecies (Sv Fs and Jc) that tended to increase whole-potbiomass (a typical effect of productive species) and one (Tc)that tended to decrease whole-pot biomass (an unusual effect)Assemblages with Tc had lower shoot biomass than assem-blages without it Tc produces little biomass when grownalone it is short with an unimpressive canopy Althoughlow in biomass Tc accumulates a disproportionate concen-tration of N twice that for Sv In the microcosms total ash-free biomass values for Tc and Sv were significantly negativelycorrelated (r = ndash043 p lt 01 n = 72 sextets) Despite itssmall size Tc seems capable of reducing the biomass of SvField data from Bahiacutea San Quintiacuten also suggest a negative re-lationship between Tc and Sv cover (r = ndash0133 p = 05 n =224) We conclude that Tc has some mechanism for reduc-
1014 BioScience December 2001 Vol 51 No 12
Features
Table 4 Experimental attempt to reestablish Sb and Seat Tijuana Estuary in 1998 (unpublished datacollected by Gabrielle Vivian-Smith Pacific EstuarineResearch Lab) Species codes are Sb Salicorniabigelovii Se Suaeda esteroa The ldquoseeds addedrdquotreatment had 2000 seeds added per speciesldquoRecruitmentrdquo is the total number of seedlings found
Treatment Recruitment
No seeds added 0 Sb 0 SeSeeds added canopy left intact 92 Sb 0 SeSeeds added canopy cleared 180 Sb 2 Se
ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features
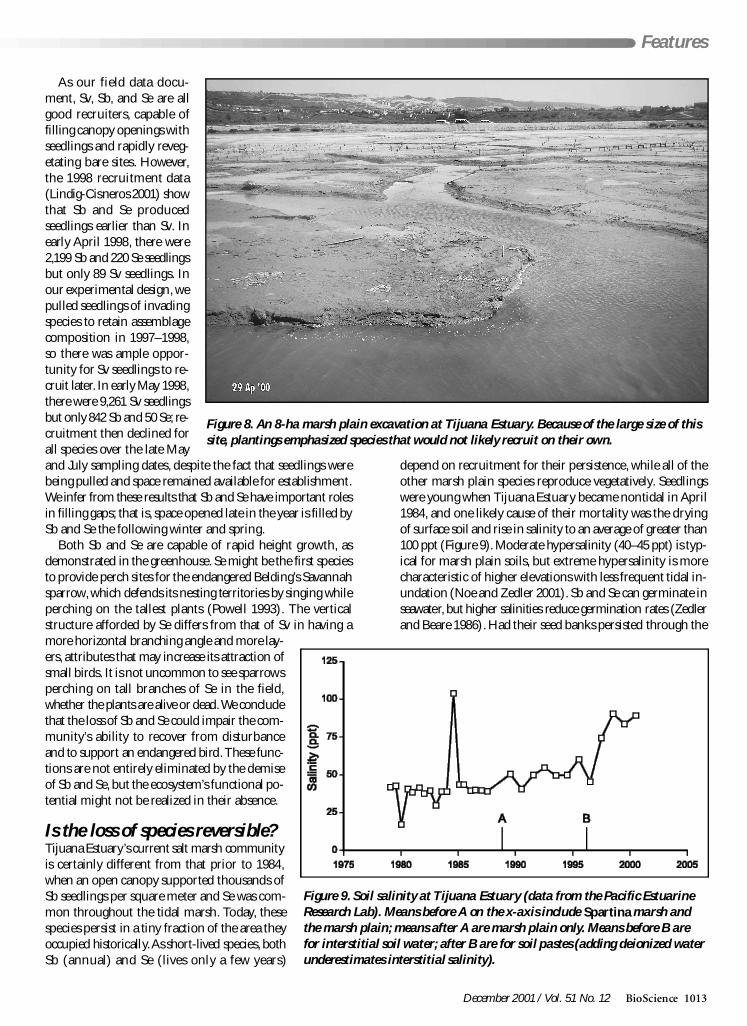
As our field data docu-ment Sv Sb and Se are allgood recruiters capable offilling canopy openings withseedlings and rapidly reveg-etating bare sites Howeverthe 1998 recruitment data(Lindig-Cisneros 2001) showthat Sb and Se producedseedlings earlier than Sv Inearly April 1998 there were2199 Sb and 220 Se seedlingsbut only 89 Sv seedlings Inour experimental design wepulled seedlings of invadingspecies to retain assemblagecomposition in 1997ndash1998so there was ample oppor-tunity for Sv seedlings to re-cruit later In early May 1998there were 9261 Sv seedlingsbut only 842 Sb and 50 Se re-cruitment then declined forall species over the late Mayand July sampling dates despite the fact that seedlings werebeing pulled and space remained available for establishmentWe infer from these results that Sb and Se have important rolesin filling gaps that is space opened late in the year is filled bySb and Se the following winter and spring
Both Sb and Se are capable of rapid height growth asdemonstrated in the greenhouse Se might be the first speciesto provide perch sites for the endangered Beldingrsquos Savannahsparrow which defends its nesting territories by singing whileperching on the tallest plants (Powell 1993) The verticalstructure afforded by Se differs from that of Sv in having amore horizontal branching angle and more lay-ers attributes that may increase its attraction ofsmall birds It is not uncommon to see sparrowsperching on tall branches of Se in the fieldwhether the plants are alive or dead We concludethat the loss of Sb and Se could impair the com-munityrsquos ability to recover from disturbanceand to support an endangered bird These func-tions are not entirely eliminated by the demiseof Sb and Se but the ecosystemrsquos functional po-tential might not be realized in their absence
Is the loss of species reversibleTijuana Estuaryrsquos current salt marsh communityis certainly different from that prior to 1984when an open canopy supported thousands ofSb seedlings per square meter and Se was com-mon throughout the tidal marsh Today thesespecies persist in a tiny fraction of the area theyoccupied historicallyAs short-lived species bothSb (annual) and Se (lives only a few years)
depend on recruitment for their persistence while all of theother marsh plain species reproduce vegetatively Seedlingswere young when Tijuana Estuary became nontidal in April1984 and one likely cause of their mortality was the dryingof surface soil and rise in salinity to an average of greater than100 ppt (Figure 9) Moderate hypersalinity (40ndash45 ppt) is typ-ical for marsh plain soils but extreme hypersalinity is morecharacteristic of higher elevations with less frequent tidal in-undation (Noe and Zedler 2001) Sb and Se can germinate inseawater but higher salinities reduce germination rates (Zedlerand Beare 1986) Had their seed banks persisted through the
December 2001 Vol 51 No 12 BioScience 1013
Features
Figure 8 An 8-ha marsh plain excavation at Tijuana Estuary Because of the large size of thissite plantings emphasized species that would not likely recruit on their own
Figure 9 Soil salinity at Tijuana Estuary (data from the Pacific EstuarineResearch Lab) Means before A on the x-axis include Spartina marsh andthe marsh plain means after A are marsh plain only Means before B arefor interstitial soil water after B are for soil pastes (adding deionized waterunderestimates interstitial salinity)
period of tidal closure Sb and Se should have germinated in1985 they did not
A recent reintroduction experiment by Vivian-Smith andZedler (2001) indicated that neither species retains a marshplain seed bank at Tijuana Estuary Vivian-Smith added 2000seeds of each species in 1998 Sb produced 272 seedlings andSe produced 2 with seedlings establishing only where seedswere added Also the current Sv-dominated canopy reducedrecruitment with Sb seedling density twice as high where thecanopy was removed (Table 4) Hence we learned that a lackof seeds and shading by Sv restricted recovery of the Sb pop-ulation And because H N Morzaria-Luna (University ofWisconsinndashMadison unpublished data) could not locate ei-ther species during her search in 2001 we suspect that changesin both the physical and biological conditions of this marshfurther inhibit recovery We explore each of these in turn
Irreversible physical changes According to recent in-formation both elevation and soil salinities have changed inthe 16 years since tidal closure A study of marsh accretion rates(Weis et al forthcoming) using cesium-137 (137Cs a globallydispersed radioisotope) suggests that since 1984 (the yearwhen Sb and Se declined) the marsh plain elevation has risen13 cm based on two soil cores At three additional locationsbenchmarks placed on the marsh plain in the 1970s by the USArmy Corps of Engineers were relocated there we have mea-sured an average of 6 cm accretion since 1984 (Morzaria-Lunaunpublished data) Further sedimentation data from TijuanaEstuary corroborate this rapid rise in elevation Cahoon andcolleagues (1996) measured 20ndash85 (mean 59) cm accretionin the S foliosa marsh between 1992 and 1993 and in anearby tidal pond (lower elevation) Ward (2000) found4ndash127 (mean 88) cm of sediment accretion from 1997 to1998 The marsh plain elevation has risen because of sedi-mentation and the recent record of flooding provides anexplanation
Tijuana Estuary has a large watershed with highly erodablesoils and major slope failures that are visible both north andsouth of the USndashMexico border The 1993 floodwaters cut anentirely new river channel just inland of the estuary Thehuge volume of sediment that was mobilized could havecontributed to a rise in elevation on the marsh plain In-creased soil salinities (now averaging 60 ppt Figure 9) cor-
roborate a shift toward a higher and hence drier marshplain For intertidal species that are sensitive to 5ndash10-cm dif-ferences in elevation the accretion rates and associated salin-ities documented at Tijuana Estuary would seem to be fatalPersistent hypersaline conditions would reduce Sb and Se es-tablishment drier conditions would favor Sv and densercanopies of Sv would reduce the persistence of seedlings
If rising elevation and increasing salinity prevent reestab-lishment of Sb and Se then reversing these changes would re-quire a massive sediment removal effortmdashthe marsh plainwould have to be bulldozed to lower the elevation by 6ndash13 cmor more Such a disruption to the vegetation would not be al-lowed by resource managers because the plant canopy isused by light-footed clapper rails for cover and Beldingrsquos Sa-vannah sparrows for nesting
There is an alternative Although these recent physicalchanges are not reversible it is possible to excavate sedi-ments and restore areas where sediment loads have shiftedtopography well out of the tidal range Exactly this procedureis planned for up to 200 ha of former tidal wetlands in thesouthern arm of the estuary (Entrix et al 1991) The re-moval of sediments in the south arm can conceivably com-pensate for some of the accretion that is occurring in the es-tuary and in the region Attempts are also being made tocontrol the flow of sediments upstream
Irreversible biological changes When the full com-plement of species is present the community presumably hasmaximum biocomplexitymdashthat is a variety of interspecificinteractions and feedback mechanisms Its biodiversity wouldlead to biocomplexity It is possible that the marshrsquos bio-complexity helped sustain its biodiversity If some interactionsfacilitated rare species and others kept dominants from out-competing subordinates then more species would persist Inthat event losing some species would make the communityless able to support all the naturally occurring species bothplant and animalWhere dense canopy cover reduces seedlingrecruitment any species that reduces overall biomass above-ground could reduce shading and enhance recruitment andspecies richness The greenhouse experiments with repli-cated assemblages suggest such a species effect
We explored effects of selected species by comparing mi-crocosms with and without such a species We found threespecies (Sv Fs and Jc) that tended to increase whole-potbiomass (a typical effect of productive species) and one (Tc)that tended to decrease whole-pot biomass (an unusual effect)Assemblages with Tc had lower shoot biomass than assem-blages without it Tc produces little biomass when grownalone it is short with an unimpressive canopy Althoughlow in biomass Tc accumulates a disproportionate concen-tration of N twice that for Sv In the microcosms total ash-free biomass values for Tc and Sv were significantly negativelycorrelated (r = ndash043 p lt 01 n = 72 sextets) Despite itssmall size Tc seems capable of reducing the biomass of SvField data from Bahiacutea San Quintiacuten also suggest a negative re-lationship between Tc and Sv cover (r = ndash0133 p = 05 n =224) We conclude that Tc has some mechanism for reduc-
1014 BioScience December 2001 Vol 51 No 12
Features
Table 4 Experimental attempt to reestablish Sb and Seat Tijuana Estuary in 1998 (unpublished datacollected by Gabrielle Vivian-Smith Pacific EstuarineResearch Lab) Species codes are Sb Salicorniabigelovii Se Suaeda esteroa The ldquoseeds addedrdquotreatment had 2000 seeds added per speciesldquoRecruitmentrdquo is the total number of seedlings found
Treatment Recruitment
No seeds added 0 Sb 0 SeSeeds added canopy left intact 92 Sb 0 SeSeeds added canopy cleared 180 Sb 2 Se
ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features

period of tidal closure Sb and Se should have germinated in1985 they did not
A recent reintroduction experiment by Vivian-Smith andZedler (2001) indicated that neither species retains a marshplain seed bank at Tijuana Estuary Vivian-Smith added 2000seeds of each species in 1998 Sb produced 272 seedlings andSe produced 2 with seedlings establishing only where seedswere added Also the current Sv-dominated canopy reducedrecruitment with Sb seedling density twice as high where thecanopy was removed (Table 4) Hence we learned that a lackof seeds and shading by Sv restricted recovery of the Sb pop-ulation And because H N Morzaria-Luna (University ofWisconsinndashMadison unpublished data) could not locate ei-ther species during her search in 2001 we suspect that changesin both the physical and biological conditions of this marshfurther inhibit recovery We explore each of these in turn
Irreversible physical changes According to recent in-formation both elevation and soil salinities have changed inthe 16 years since tidal closure A study of marsh accretion rates(Weis et al forthcoming) using cesium-137 (137Cs a globallydispersed radioisotope) suggests that since 1984 (the yearwhen Sb and Se declined) the marsh plain elevation has risen13 cm based on two soil cores At three additional locationsbenchmarks placed on the marsh plain in the 1970s by the USArmy Corps of Engineers were relocated there we have mea-sured an average of 6 cm accretion since 1984 (Morzaria-Lunaunpublished data) Further sedimentation data from TijuanaEstuary corroborate this rapid rise in elevation Cahoon andcolleagues (1996) measured 20ndash85 (mean 59) cm accretionin the S foliosa marsh between 1992 and 1993 and in anearby tidal pond (lower elevation) Ward (2000) found4ndash127 (mean 88) cm of sediment accretion from 1997 to1998 The marsh plain elevation has risen because of sedi-mentation and the recent record of flooding provides anexplanation
Tijuana Estuary has a large watershed with highly erodablesoils and major slope failures that are visible both north andsouth of the USndashMexico border The 1993 floodwaters cut anentirely new river channel just inland of the estuary Thehuge volume of sediment that was mobilized could havecontributed to a rise in elevation on the marsh plain In-creased soil salinities (now averaging 60 ppt Figure 9) cor-
roborate a shift toward a higher and hence drier marshplain For intertidal species that are sensitive to 5ndash10-cm dif-ferences in elevation the accretion rates and associated salin-ities documented at Tijuana Estuary would seem to be fatalPersistent hypersaline conditions would reduce Sb and Se es-tablishment drier conditions would favor Sv and densercanopies of Sv would reduce the persistence of seedlings
If rising elevation and increasing salinity prevent reestab-lishment of Sb and Se then reversing these changes would re-quire a massive sediment removal effortmdashthe marsh plainwould have to be bulldozed to lower the elevation by 6ndash13 cmor more Such a disruption to the vegetation would not be al-lowed by resource managers because the plant canopy isused by light-footed clapper rails for cover and Beldingrsquos Sa-vannah sparrows for nesting
There is an alternative Although these recent physicalchanges are not reversible it is possible to excavate sedi-ments and restore areas where sediment loads have shiftedtopography well out of the tidal range Exactly this procedureis planned for up to 200 ha of former tidal wetlands in thesouthern arm of the estuary (Entrix et al 1991) The re-moval of sediments in the south arm can conceivably com-pensate for some of the accretion that is occurring in the es-tuary and in the region Attempts are also being made tocontrol the flow of sediments upstream
Irreversible biological changes When the full com-plement of species is present the community presumably hasmaximum biocomplexitymdashthat is a variety of interspecificinteractions and feedback mechanisms Its biodiversity wouldlead to biocomplexity It is possible that the marshrsquos bio-complexity helped sustain its biodiversity If some interactionsfacilitated rare species and others kept dominants from out-competing subordinates then more species would persist Inthat event losing some species would make the communityless able to support all the naturally occurring species bothplant and animalWhere dense canopy cover reduces seedlingrecruitment any species that reduces overall biomass above-ground could reduce shading and enhance recruitment andspecies richness The greenhouse experiments with repli-cated assemblages suggest such a species effect
We explored effects of selected species by comparing mi-crocosms with and without such a species We found threespecies (Sv Fs and Jc) that tended to increase whole-potbiomass (a typical effect of productive species) and one (Tc)that tended to decrease whole-pot biomass (an unusual effect)Assemblages with Tc had lower shoot biomass than assem-blages without it Tc produces little biomass when grownalone it is short with an unimpressive canopy Althoughlow in biomass Tc accumulates a disproportionate concen-tration of N twice that for Sv In the microcosms total ash-free biomass values for Tc and Sv were significantly negativelycorrelated (r = ndash043 p lt 01 n = 72 sextets) Despite itssmall size Tc seems capable of reducing the biomass of SvField data from Bahiacutea San Quintiacuten also suggest a negative re-lationship between Tc and Sv cover (r = ndash0133 p = 05 n =224) We conclude that Tc has some mechanism for reduc-
1014 BioScience December 2001 Vol 51 No 12
Features
Table 4 Experimental attempt to reestablish Sb and Seat Tijuana Estuary in 1998 (unpublished datacollected by Gabrielle Vivian-Smith Pacific EstuarineResearch Lab) Species codes are Sb Salicorniabigelovii Se Suaeda esteroa The ldquoseeds addedrdquotreatment had 2000 seeds added per speciesldquoRecruitmentrdquo is the total number of seedlings found
Treatment Recruitment
No seeds added 0 Sb 0 SeSeeds added canopy left intact 92 Sb 0 SeSeeds added canopy cleared 180 Sb 2 Se
ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features

ing the aboveground biomass ofother species and where shootbiomass is reduced canopies aremore open and recruitmentshould be facilitated
Any of three mechanismscould lead to reduction ofwhole-pot biomass by Tc (1)direct competition for N whichreduces other speciesrsquo growth(2) allellopathy (negative chem-ical effects) or (3) indirect in-teractions that favor otherspecies which in turn reduce Svbiomass In support of directcompetition Tc sequesters a largemass of N because it has thehighest concentration of N in itsroots In the field long-livedclones of Tc could tie up sub-stantial stores of N In support ofindirect interaction Tcrsquos phe-nology and life history are highlycomplementary to that of Sb Tchas an open canopy (the fewest layers of all species) whichmost likely facilitates Sb germination Tcrsquos canopy declines inMay when Sb grows rapidly Sb reaches peak biomass fruitsand dies in fall The N in Sb tissues would then be releasedabout the time that Tc resumes aboveground growth The po-tential exists for complementary N use
The post hoc evaluation of assemblages with Tc suggesteda biocomplexity model in which Tc facilitates the co-occur-rence of Sb (Table 5) Like a keystone species Tc might be pre-venting overdominance by Sv Fs or Jc It could thus be a keyto biodiversity support in the Californian salt marsh (Table5) It is widespread at San Quintiacuten Bay (greater than 70 fre-quency in 025-m2 quadrats) Its absence in many Californiasalt marshes could be part of the reason Sv is so often dom-inant Ibarra-Obando and Poumian-Tapia (unpublisheddata) recorded large differences in Tc cover where tidal flush-ing was eliminated at Estero de Punta Banda (46 versus lessthan 10 cover in 1984 and 71 versus less than 3 in1985 for areas with and without tidal flushing respectivelyand occurrences of 73 versus 15 plusmn tidal flushing) The po-tential for low-biomass species to open Sv canopies is not un-precedented Cuscuta salina sometimes facilitates recruit-ment on the marsh plain (Pennings and Callaway 1996) butthis obligate parasite is not amenable to experimental plant-ings We conclude that Tc Sb and Se are all highly sensitiveto tidal interruption and suggest that establishing Tc inrestoration sites could promote establishment and persis-tence of Sb and Se Had it not been for the random drawingof species combinations we would not have tested assemblageswith and without Tc because this small and often incon-spicuous species is a winterndashspring ephemeral that displayslittle evidence of its rich store of N Hence we disagree with
Huston and colleagues (2000) who argue that randomlydrawn assemblages lack relevance
The biodiversity debate revisitedBiodiversity and function are positively related in the saltmarsh that is all of the species appear to be needed to pro-vide all of the functions Our comparison of species richnesslevels and single-species treatments contributes quantitativedata on multiple functions showing that different species per-form different functions best (Sullivan and Zedler 1999) Fur-thermore our biocomplexity model suggests that a small butN-rich species might keep a highly productive species incheck thereby allowing other species to coexist Ecosystemdisruptions such as tidal closure can be catastrophic byeliminating species but a more elusive response might alsooccur if the loss of a keystone species simultaneously removesthe mechanism for sustaining diversity The loss of biodi-versity might mean the loss of biocomplexity which in turnmeans loss of the ability to sustain biodiversity If Tc has akeystone role in biodiversity maintenance then its loss dur-ing tidal closure would have lasting effects on the entire as-semblage These biologically complex interactions are nowunder experimental study and long-term follow-up of theimmature vegetation in our experimental plots will helptest this model
ConclusionsThe Californian salt marsh plain is a model system for ex-ploring the importance of diversity to restoration Species-richplantings enhanced the restoration site by increasing re-cruitment of native species canopy complexity biomassand N accumulation
December 2001 Vol 51 No 12 BioScience 1015
Features
Table 5 Schematic based on greenhouse experimentation showing low species richnesswhere Triglochin concinna (Tc) is absent (and the presence of only the more productiveand aggressive competitors (Fs Jc) versus a more species-rich system where Tc is presentTc might reduce aboveground biomass and cover of the perennial Salicornia virginica (Sv)in the field thereby allowing some of the less competitive species to coexist with Sv Fs andJc Reduced biomass would make more light available for the annual S bigelovii (Sb) torecruit from seed Plants illustrated by Donovan McIntire
Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features

Once lost diversity might not be easily restored because ofchanges in the environment or disruption of critical speciesinteractions The hypothesis emerged that biocomplexity (inthis case species interaction) sustains biodiversity (persistenceof all species) when experiments showed that a low biomassspecies (Tc) reduces the growth of high-biomass species (es-pecially Sv) Mechanisms that might explain this surprisingoutcome include N sequestering allelopathy and phenologythat facilitates growth of an annual plant (Sb)
Experiments with young vegetation and randomly drawnassemblages are highly relevant in the restoration contextRestoration sites offer outstanding opportunities to ad-vance both the science and practice of restoration
Recommendations for restorationIn general
Do not ignore uncommon inconspicuous species inplanning the restoration of vegetation Even if domi-nant species are easy to propagate they might not ful-fill all essential functions or achieve any one desiredgoal They might also impair the establishment ofdesirable species that contribute functions that areless conspicuous
Plant species that recruit poorly
Employ measures of canopies that evaluate complexity(such as height and layering) not just plant cover
Incorporate more physical complexity into restorationsites and foster biological interactions (via mixed-species plantings) to sustain biological diversity
Use restoration sites to test further the ability oftopographic variability and interspecific interactions to enhance the persistence of species-rich assem-blages The design of restoration sites as experimentscan simultaneously vegetate the site and improveknowledge
For Californian salt marshes
Sow seeds of Sb and Se and plant seedlings of Bm TcLc Fs and Jc Allow Sv to recruit from seed on itsown so it will not immediately dominate the site
Introduce plants as multispecies assemblages to accelerate development of biomass (Fs Jc) N accumulation (Tc) height (Se) layering (Fs Se)shade (Lc) seedling density (Sb) and vegetativespread (Bm) Test the effect of planting Tc firstand then introducing short-lived species (Sb Se)
Track the outcomes by monitoring plantings in latespring (to evaluate Tc establishment and growth) andlate fall (to assess other speciesrsquo performance) and bymonitoring recruitment by volunteers (Sv) as well asplanted species
AcknowledgmentsWe thank the National Science Foundation (DEB 96-19875)and Earth Island Institute for support of this researchGeorgeann Keer and Roberto Lindig-Cisneros conductedstudies of canopy architecture and seedling recruitment re-spectively providing results that greatly improved our un-derstanding of species richness and species composition ef-fects The research could not have been completed withouthelp from our colleagues and staff at the Pacific Estuarine Re-search Laboratory at San Diego State University who helpedgrow transplant and maintain thousands of seedlings for thefield and greenhouse experiments
References citedBarbour MG Major J 1988 Terrestrial Vegetation of California Davis (CA)
California Native Plant SocietyBoyer KE Zedler JB 1996 Damage to cordgrass by scale insects in a con-
structed salt marsh Effects of nitrogen additions Estuaries 19 1ndash12mdashmdashmdash 1998 Effects of nitrogen additions on the vertical structure of a con-
structed cordgrass marsh Ecological Applications 8 692ndash705Boyer KE Callaway JC Zedler JB 2000 Evaluating the progress of restored
cordgrass (Spartina foliosa) marshes Belowground biomass and tissueN Estuaries 23 711ndash721
Cahoon DR Lynch JC Powell AN 1996 Marsh vertical accretion in a south-ern California estuary USA Estuarine Coastal and Shelf Science 4319ndash32
Carpelan LH 1969 Physical characteristics of southern California coastal la-goons Pages 319ndash344 in Castanares AA Phleger FB eds LagunasCosteras Un Simposio Mexico City (DF) Universidad Nacional Au-tonoma de Mexico
Chapin FS Schulze ED Mooney HA 1992 Biodiversity and ecosystemprocesses Trends in Ecology amp Evolution 7 107ndash108
Crocker RL Major J 1955 Soil development in relation to vegetation andsurface age at Glacier Bay Alaska Journal of Ecology 43 427ndash448
Entrix Inc PERL PWA Ltd 1991 Draft Environmental Impact ReportEnvironmental Impact Statement Tijuana Estuary Tidal Restoration Pro-gram Oakland (CA) California Coastal Conservancy and US Fish andWildlife Service
Ewel JJ Mazzarino MJ Berish CW 1991 Tropical soil fertility changes un-der monocultures and successional communities of different structureEcological Applications 1 289ndash302
Ferren WR Jr 1985 Environment History and Botanical Resources of aSouthern California Estuary Santa Barbara (CA) Department of Bio-logical Sciences University of California Publication No 4
Hector A et al 1999 Plant diversity and productivity experiments in Euro-pean grasslands Science 286 1123ndash1127
Hooper DU Vitousek PM 1997 The effects of plant composition and di-versity on ecosystem processes Science 277 1302ndash1305
Huston MA et al 2000 No consistent effect of plant diversity on produc-tivity Science 289 1255a
Ibarra-Obanda SE Poumian-Tapia M 1991 The effect of tidal exclusion onsalt marsh vegetation in Baja California Mexico Wetlands Ecology andManagement 1 131ndash148
Johnson KHVogt KA Clark HJ Schmitz OJ Vogt DJ 1996 Biodiversity andthe productivity and stability of ecosystems Trends in Ecology amp Evo-lution 11 372ndash377
Keer G Zedler JB Salt marsh canopy architecture differs with the numberand composition of species Ecological Applications Forthcoming
Knops JMH et al 1999 Effects of plant species richness on invasion dynamicsdisease outbreaks insect abundances and diversity Ecology Letters 2286ndash293
1016 BioScience December 2001 Vol 51 No 12
Features
Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features

Langis R Zalejko M Zedler JB 1991 Nitrogen assessment in a constructedand a natural salt marsh of San Diego Bay Ecological Applications 140ndash51
Lindig-Cisneros R 2001 Interactions among species richness canopy struc-ture and seedling recruitment PhD dissertation University of Wis-consin Madison
Naeem S Li SB 1997 Biodiversity enhances ecosystem reliability Nature 390507ndash509
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1994 Decliningbiodiversity can alter the performance of ecosystems Nature 368734ndash737
Naeem S Thompson LJ Lawler SP Lawton JH Woodfin RM 1995 Empir-ical evidence that declining species diversity may alter the performanceof terrestrial ecosystems Philosophical Transactions of the Royal Soci-ety of London Series B Biological Sciences 347 249ndash262
Naeem S Hakansson K Lawton JH Crawley MJ Thompson LJ 1996 Bio-diversity and plant productivity in a model assemblage of plant speciesOikos 76 259ndash264
Noe GB Zedler JB 2001 Spatio-temporal variation of salt marsh seedlingestablishment in relation to the abiotic and biotic environment Journalof Vegetation Science 12 61ndash74
Noss RF 1990 Indicators for monitoring biodiversity A hierarchical approachConservation Biology 4 355ndash364
Onuf CP 1987 The ecology of Mugu Lagoon California An estuarine pro-file Washington (DC) US Fish and Wildlife Service Biological Report85 (715)
Pennings SC Callaway RM 1996 Impact of a parasitic plant on the struc-ture and dynamics of salt marsh vegetation Ecology 77 1410ndash1419
Powell AN 1993 Nesting habitat of Belding Savannah sparrows in coastalsalt marshes Wetlands 13 219ndash223
Schwartz MW Brigham CA Hoeksema JD Lyons KG Mills MH van Mant-gem PJ 2000 Linking biodiversity to ecosystem function Implicationsfor conservation ecology Oecologia 122 297ndash305
Siemann E Tilman D Haarstad J Ritchie M 1998 Experimental tests of thedependence of arthropod diversity on plant diversity American Natu-ralist 152 738ndash750
Silver WL Brown S Lugo AE 1996 Effects of changes in biodiversity onecosystem function in tropical forests Conservation Biology 10 17ndash24
Sullivan G Zedler JB 1999 Functional redundancy among tidal marshhalophytes A test Oikos 84 246ndash260
Symstad AJ 2000 A test of the effects of functional group richness andcomposition on grassland invasibility Ecology 81 99ndash109
Symstad AJ Tilman D Willson J Knops JMH 1998 Species loss and ecosys-tem functioning Effects of species identity and community compositionOikos 81 389ndash397
Tilman D 1996 Biodiversity Population versus ecosystem stability Ecology77 350ndash363
mdashmdashmdash 1997 Community invasibility recruitment limitation and grasslandbiodiversity Ecology 78 81ndash92
Tilman D Downing JA 1994 Biodiversity and stability in grasslands Nature367 363ndash365
van der Heijden MGA Klironomos JN Ursic M Moutoglis P Streitwolf-Engel R Boller T Wiemken A Sanders IR 1998 Mycorrhizal fungal di-versity determines plant biodiversity ecosystem variability and pro-ductivity Nature 396 69ndash72
Vitousek PM Hooper DU 1993 Biological diversity and terrestrial ecosys-tem biogeochemistry Pages 3ndash14 in Schulze ED Mooney HA eds Bio-diversity and Ecosystem Function Berlin Springer-Verlag
Vitousek PM Reiners WA 1975 Ecological succession and nutrient reten-tion A hypothesis BioScience 25 376ndash381
Vivian-Smith G Zedler JB 2001 Enhancing rare plant populations at TijuanaEstuary Pages 82ndash38 in Zedler JB ed Handbook for Restoring Tidal Wet-lands Boca Raton (FL) CRC Press
Ward K 2000 Episodic colonization of an intertidal mudflat by cordgrass(Spartina foliosa) at Tijuana Estuary Masterrsquos thesis San Diego State Uni-versity San Diego
Weis DA Callaway JC Gersberg RM Vertical accretion rates and heavy
metal chronologies in wetland sediments of the Tijuana Estuary Estu-
aries Forthcoming
Winfield TP 1980 Dynamics of carbon and nitrogen in a southern California
salt marsh PhD dissertation University of CaliforniandashRiverside and
San Diego State University
Zedler JB 1982 The ecology of southern California coastal salt marshes A
community profile Washington (DC) US Fish and Wildlife Service Bi-
ological Report FWSOBS-3154
mdashmdashmdash ed 2001 Handbook for Restoring Tidal Wetlands Boca Raton
(FL) CRC Press
Zedler JB Beare PA 1986 Temporal variability of salt marsh vegetation The
role of low-salinity gaps and environmental stress Pages 295ndash306 in Wolfe
DA ed Estuarine Variability San Diego Academic Press
Zedler JB Callaway JC 1999 Tracking wetland restoration Do mitigation
sites follow desired trajectories Restoration Ecology 7 69ndash73
mdashmdashmdash 2000 Evaluating the progress of engineered tidal wetlands Ecological
Engineering 15 211ndash225
Zedler JB Callaway JC Desmond J Vivian-Smith G Williams G Sullivan G
Brewster ABradshaw B1999 Californian salt marsh vegetation An im-
proved model of spatial pattern Ecosystems 2 19ndash35
Zedler JB Nordby CS Kus BE 1992 The ecology of Tijuana Estuary Cali-
fornia A National Estuarine Research Reserve Washington (DC) NOAA
Office of Coastal Resource Management Sanctuaries and Reserves Di-
vision
Zedler JB Winfield T Williams P 1980 Salt marsh productivity with natural
and altered tidal circulation Oecologia 44 236ndash240
December 2001 Vol 51 No 12 BioScience 1017
Features





























