Embed Size (px)
Citation preview

Dinámica de ecosistemas dominados por especies germinadoras obligadas en el oeste de la cuenca mediterránea: respuesta sucesional a incendios recurrentes
Victor Manuel Santana Pastor

Dinámica de ecosistemas dominados por especies germinadoras obligadas en el oeste de la cuenca med iterránea:
respuesta sucesional a incendios recurrentes.
___________________
Dynamics of ecosystems dominated by obligate seeder s in the western Mediterranean Basin: successional response to
recurrent fires.
Memoria presentada por:
Victor Manuel Santana Pastor para optar al grado de doctor en Ciencias Biológicas
Alicante, Marzo de 2011.



Manuel Jaime Baeza Berná, Investigador Senior de la Fundación CEAM y Profesor Asociado de la Universidad de Alicante. HACE CONSTAR: Que el trabajo descrito en la presente memoria, titulado: “Dinámica de ecosistemas dominados por especies germinadoras obligadas en el oeste de la cuenca mediterránea: respuesta sucesional a incendios recurrentes” ha sido reallizado bajo su dirección por D. Victor Manuel Santana Pastor en la Fundación de la Generalitat Valenciana Centro de Estudios Ambientales del Mediterráneo (CEAM), y reúne todos los requisitos necesarios para su aprobación como Tesis Doctoral.
Alicante, 25 de Enero de 2011
Dr. M. Jaime Baeza Berná

AGRADECIMIENTOS
Esta Tesis doctoral ha sido posible gracias a la beca de Formación de Personal
Universitario (FPU) otorgada por el Ministerio de Educación. La financiación de
la investigación ha sido posible gracias a los proyectos FIREMED (AGL200/8-
04522/FOR) y Consolider-Ingenio 2010 (GRACCIE CSD2007-00067). El CEAM
está financiado por la Generalitat Valenciana y la Fundación Bancaja.
En primer lugar, quisiera agradecer a mi director de Tesis, Jaime Baeza,
la oportunidad de realizar esta tesis doctoral dentro de un campo de la ecología
que me encanta como son los incendios forestales. Ha sido un director
excelente además de un amigo. También los compañeros del CEAM, Alejandro,
David, Joanet, Alberto, Vanesa, Jaime, Esteban, Karim y Ramón han sido un
gran apoyo moral y logístico a la hora de realizar esta tesis doctoral y, sin el
cual, el trabajo hubiera sido mucho más difícil. También agradecer el apoyo de
la gente del CEAM de Valencia, Mª Carmen, Emilio y Cristina, que se han
encargado del papeleo infame de la beca. José Antonio Valiente colaboró
desinteresadamente en la puesta en marcha del complejo mecanismo del data
logger.
Los propietarios de las parcelas de las quemas experimentales, Cristóbal
Miró, Ramón Gisbert y Victoriano Fuentes amablemente dieron su permiso para
poder realizar este trabajo.
Agradecer el recibimiento prestado por los anfitriones en las estancias
cortas en el extranjero que he realizado durante esta Tesis: Rob Marrs del
Applied Vegetation Dynamics Lab de la Universidad de Liverpool (Reino Unido),
Ross Bradstock del Centre for Environmental Risk Management of Bushfires de
la Universidad de Wollongong (Australia) y Mike Palmer del Department of
Botany de la Oklahoma State University (Estados Unidos). Además de
contribuir con su experiencia y críticas constructivas al desarrollo de esta tesis,
me han enseñado a ver la ecología desde nuevas perspectivas. Fernando
Maestre de la Universidad Rey Juan Carlos de Madrid también ha contribuido
decisivamente en uno de los capítulos de esta tesis.
No quisiera olvidarme de la gente de la Universidad de Alicante. Román,
Marian, Tadas, Noelia y Jordi me introdujeron en el mundo de la investigación
gracias a la beca de colaboración en el departamento de Ecología (hace ya

unos cuantos años). Susana y Andreu han colaborado con el diseño y
preparación de las parcelas de estudio. Mención especial para José Huesca
que me ha ayudado amablemente en las tareas de laboratorio. A todos los
becarios, laborantes, contratados y amigos de la UA, Karen, Rosario, Anna,
Ángeles, Soraya, Estrella, Adela, Bea, Juanjo, Luna, Diana, Núria, Olga,
imprescindibles en esas largas cenas de departamento.
El duro trabajo de campo no podría haberse realizado sin la ayuda de la
gente en prácticas. Oriol, David, Benjamín, Isabel, María, Silvia, Thanos,
Christos, Raquel & Raquel. Espero que no me guarden rencor por haberlos
llevado a trabajar bajo los terribles dominios del Ulex. A todos ellos muchas
gracias.
Un record especial per a la manada d'antics companys de llicenciatura,
Vicen, Santi, Soraya, Quique, Sofia, Isa, Susi, Cox, Peruan i Juanjo, que poc a
poc anem alcançant metes més llunyanes en açò de la Biologia. També, com
no, als meus amics de tota la vida i de la filà Kabilenyos de Sant Vicent. Espere
que després de llegir esta tesis doctoral (vos obligaré, no ho dubteu)
s'assabenten d'una vegada a què dedique la meua vida.
Un agraïment especial a la meua família, ma mare Victoria, mon pare
Juan Luis i el meu germà Abel. Pel seu recolzament incondicional al llarg de la
meua vida acadèmica (i no acadèmica també), i que han fet que siga la
persona que sóc hui en dia.
Finalment, el millor dels agraïments vull donar-lo a Mamen, per estar
sempre ahí, tant en el moments bons com dolents, i per fer-me creure que en
esta vida tot és possible.
A tots, moltes gràcies.

ÍNDICE
Síntesis 1
1. Introducción general 5
2. Área de estudio 29
3. Características estructurales del combustible modulando las
temperaturas del suelo en parches de diferentes especies de matorral de
la cuenca mediterránea 37
4. Establecimiento sucesional de plántulas en matorrales mediterráneos
dominados por germinadoras obligadas 55
5. La recurrencia de incendios y el tiempo desde el incendio como
conductores de la inflamabilidad en matorrales mediterráneos 75
6. Sucesión secundaria en campos de cultivo abandonados del sureste de
España: ¿puede el fuego desviarla? 93
7. Efecto del régimen de temperatura después del fuego en la dormancia y
germinación de semillas de seis especies de Fabaceae australianas 113
8. Discusión general 127
9. Conclusiones 137


1
SÍNTESIS
El abandono masivo de campos de cultivo a mediados del siglo XX en la
Comunitat Valenciana ha conllevado un aumento de ecosistemas en estados
iniciales de la sucesión dominados por especies germinadoras obligadas.
Debido a que este tipo de especies acumulan en su estructura gran cantidad de
combustible muerto y que a consecuencia de su desarrollo la conectividad
entre áreas forestales es mayor, la frecuencia y extensión de incendios
forestales se ha incrementado en las últimas décadas. Este hecho supone un
riesgo tanto para la fucionalidad del ecosistema como para la seguridad
humana. Por lo tanto, el objetivo general de esta tesis es indagar en los
procesos sucesionales que rigen los ecosistemas dominados por especies
germinadoras obligadas en el oeste de la cuenca mediterránea. Especialmente
en: (1) los mecanismos que controlan el establecimiento de individuos a lo
largo de la sucesión, (2) los patrones de abundancia de las especies
dominantes en función del tiempo desde el incendio y de la recurrencia de
incendio, (3) la dinámica de los diferentes tipos de combustible asociada a los
cambios sucesionales, (4) la variabilidad en los efectos del fuego en el
ecosistema dependiendo de la especie dominante en el dosel y que
característica estructural del combustible sería la más influyente sobre estos
mismos efectos. El conocimiento de estos procesos contribuirá a la toma de
decisiones en la gestión de estos ecosistemas altamente propensos al fuego.
Para ello se han realizado diferentes trabajos de campo en el interior de la
Comunitat Valenciana. Como conclusiones más importantes se destaca que: 1)
existen diferencias en el nicho de regeneración de las especies germinadoras
obligadas que hace que unas especies sean más competitivas que otras a lo
largo del gradiente sucesional o en diferentes regímenes de incendio. Aunque,
la mayor parte de los individuos de las especies estudiadas se establecen en
etapas inmediatamente post-fuego, Cistus albidus se regenera prioritariamente
en ambientes recientemente perturbados, mientras que Ulex parviflorus y
Rosmarinus officinalis pueden establecerse en periodos entre incendios. 2)
Existen procesos de sustitución especies a lo largo de la sucesión en los
matorrales dominados por especies germinadoras. Estos ecosistemas se rigen
por el mecanismo sucesional de tolerancia, y tras una primera etapa dominada

2
por Ulex parviflorus y Cistus albidus la comunidad pasa a estar dominada por
Rosmarinus officinalis. Un fuego recurrente no afecta los patrones de
sustitución entre especies, pero si que afecta a la abundancia y retrasa en el
tiempo el punto donde las especies alcanzan su óptimo. 3) Los campos de
cultivo abandonados pueden establecer diferentes trayectorias sucesionales
dependiendo del régimen de recurrencia de incendio. En ausencia de fuego,
son dominados en una primera etapa por Pinus halepensis, que con el paso del
tiempo se convierten en una formación mixta de pinar con especies
rebrotadoras y de hoja ancha como Quercus ilex y Q. coccifera. Un solo
incendio puede sustituir el pinar por un matorral de R. officinalis, donde el
establecimiento de especies germinadoras obligadas y de especies
rebrotadoras de etapas sucesionales posteriores pueden estar impedidas. Una
alta recurrencia de incendio en intervalos cortos de tiempo desvían el
ecosistema hacia una comunidad dominada por terófitos o herbáceas como
Brachypodium retusum. 4) La capacidad de retener combustible muerto y su
disposición en la estructura de la planta es determinante en los efectos del
fuego en el ecosistema. Bajo parches de la especie que más acumula
combustible muerto, U. parviflorus, se experimentan las mayores tasas de
consumo de biomasa y de temperatura de suelo. En contra, bajo los parches
de R. officinalis, la especie con menor acumulación de combustible muerto, se
encuentran los efectos opuestos. 5) Los procesos sucesionales de sustitución
de especies en ecosistemas de matorral llevan asociada una función de
inflamabilidad basada en la cantidad de combustible muerto acumulado. Una
primera etapa de la sucesión dominada por U. parviflorus, especie que mayor
cantidad de combustible muerto acumula, seguida por una dominancia de R.
officinalis, especie que acumula menor cantidad, conlleva a una función de
inflamabilidad de forma jorobada; es decir, tras un incremento inicial del
combustible muerto acumulado, éste disminuye con el transcurso de la
sucesión. 6) Tras un fuego recurrente no existe un incremento en la cantidad
de combustible muerto acumulado a nivel de comunidad. Por lo tanto, se
sugiere la ausencia de un bucle de retroalimentación positivo entre las especies
que acumulan mayor cantidad de combustible muerto y una regeneración
estimulada por el fuego. 7) Un efecto indirecto del fuego, como el aumento del
régimen de temperaturas diarias del suelo, puede ser un desencadenante de la

3
ruptura física de la dormancia en semillas de leguminosas de sureste de
Australia. Este efecto difiere entre las diferentes especies y, además, esta
modulado por el rango de temperaturas y el tiempo de exposición

4

5
CAPÍTULO 1-
INTRODUCCIÓN GENERAL

6

7
CAPÍTULO 1- INTRODUCCIÓN GENERAL
- Los ecosistemas mediterráneos y su relación con el fuego
La explotación de recursos naturales mediante la acción antrópica ha
modificado gran parte de los sistemas naturales durante milenios (Vitousek et
al. 1997). Sin embargo, debido a los cambios socio-económicos ocurridos a
nivel global desde el siglo pasado, la actividad en muchos de estos
ecosistemas ha cesado, mientras que en otros se ha iniciado o incluso se ha
incrementado (Cramer et al. 2008). Como consecuencia, están surgiendo en
diferentes biomas de todo el mundo ecosistemas emergentes (también
denominados ecosistemas noveles) con nuevas combinaciones de especies y
abundancias relativas que no se observaban previamente (Hobbs et al. 2006).
En algunos casos, estos ecosistemas pueden prevenir la instalación y
regeneración de las especies previas a la explotación, ya sea por competencia
o por modificar los mecanismos que controlan el funcionamiento del
ecosistema, incluyendo cambios en el régimen de perturbaciones (Suding et al.
2004, Hobbs et al. 2006).
La cuenca mediterránea ha sufrido durante milenios una alta antropización
de sus sistemas naturales. La agricultura, ganadería o explotación de otros
recursos naturales han sido una constante a lo largo de su territorio desde la
época neolítica, hace aproximadamente unos 10.000 años (Blondel y Aronson
1999, Blondel 2006). Sin embargo, la intensa industrialización de esta región
desde mediados del siglo pasado ha conducido al abandono de la actividad
rural y, en consecuencia, a un abandono generalizado de vastas extensiones
de terreno dedicadas al cultivo o al pastoreo (Le Houérou 1993). Actualmente,
estos sistemas se encuentran en estados iniciales de la sucesión, dominados
por especies de crecimiento rápido y alta capacidad de colonización de
espacios abiertos (Cramer et al. 2008).
Paralelamente al factor humano, el fuego es considerado una de las fuerzas
moduladoras más importantes del paisaje mediterráneo (muchas veces
provocado por la propia actividad humana). Tradicionalmente, se ha
considerado que estos ecosistemas poseen alta capacidad de regeneración y
que, con el paso del tiempo, recuperan su composición y estructura (Hanes

8
1971, Trabaud y Lepart 1980). La capacidad de regeneración viene
determinada por una serie de estrategias de supervivencia intrínsecas a las
especies que permiten su persistencia tras el fuego. Muchas especies
sobreviven a nivel individual gracias a estructuras enterrados bajo el suelo
(como por ejemplo tubérculos, rizomas o lignotubérculos) que las protegen de
las altas temperaturas, permitiéndoles rebrotar posteriormente (especies
rebrotadoras). Algunas especies de pinos mediterráneos o el alcornoque
presentan gruesas cortezas que protegen los tallos. Otro tipo especies que no
sobreviven al efecto del fuego, persisten a la perturbación mediante
poblaciones de semillas enterradas en el suelo (especies germinadoras
obligadas) (Keeley 1986, Pausas et al. 2004). Estas especies disponen a
menudo de un banco de semillas persistente (en el suelo o de copa), y el
reclutamiento de nuevos individuos viene determinado por las condiciones
generadas durante y posteriormente al fuego; por ejemplo, rompiendo el estado
de dormancia de las semillas o abriendo los conos de las especies con frutos
serótinos (Bond y van Wilgen 1996). Algunas especies muestran ambas
estrategias (especies facultativas), mientras que otras no pueden regenerarse
después del incendio mediante estas estrategias y su persistencia depende de
la capacidad de colonización desde áreas no quemadas.
A pesar de que estos ecosistemas se consideran resilientes, la
presencia y/o abundancia relativa, tanto de las especies como de los grupos
funcionales dominantes, puede estar determinada por el régimen de incendios
(intensidad, severidad, recurrencia, frecuencia, extensión y estacionalidad del
fuego; ver Fox y Fox (1987) para una descripción detallada de los
componentes del régimen de incendio). Por ejemplo, altas severidades o
recurrencias de incendio pueden reducir la capacidad de regeneración de las
especies, ya sea por eliminar directamente sus estructuras de persistencia
(Lloret y López-Soria 1993, Herranz et al. 1999) o por impedir que en cortos
periodos entre incendios, éstas repongan sus estructuras de reserva o bancos
de semillas (Zedler et al. 1983). Por el contrario, largos periodos en ausencia
de incendios puede ir en detrimento de especies de vida corta cuya
regeneración es dependiente del fuego (Keeley 1986). La extensión del
incendio puede limitar la reentrada de especies extintas como consecuencia del
fuego (Rodrigo et al. 2004). La estacionalidad puede afectar la regeneración de

9
especies cuya acumulación de recursos es baja o el banco de semillas no ha
sido completamente repuesto (Cruz y Moreno 2001, Domínguez et al. 2004).
No obstante, dentro del contexto de la presente tesis doctoral, cabe aclarar
algunas ambigüedades comunes entre dos componentes del régimen de
incendios como son la recurrencia y la frecuencia de incendios. La recurrencia
de incendios se considera como el número de incendios que ocurren en un
punto concreto y en un determinado periodo de tiempo, mientras que la
frecuencia de incendio se refiere al promedio de incendios ocurridos a lo largo
de un cierto periodo de tiempo y sobre un área concreta (sensu Johnson 1992).
En la actualidad, los incendios forestales en la cuenca mediterránea es uno
de los objetivos prioritarios dentro de la investigación forestal a nivel europeo,
ya que una media de 500.000 ha se queman anualmente en Europa. Los
cambios socioeconómicos ocurridos desde el siglo pasado han conllevado una
variación en el régimen de incendios hacia una mayor recurrencia y superficie
quemada. Los procesos que controlan la colonización y sucesión de las áreas
de cultivo abandonadas han supuesto un incremento en la cantidad de
combustible acumulado y una mayor conectividad entre los sistemas forestales.
Evidencias de algunas investigaciones han demostrado que el cambio en el
régimen de incendios podría estar potenciado como resultado del proceso de
cambio climático actual (Pausas 2004, Mouillot y Field 2005). Como
consecuencia directa, la resiliencia de estos ecosistemas se ha visto
comprometida en algunos casos, induciendo en el ecosistema procesos de
degradación a nivel de vegetación y de suelo (Lloret 2004). De hecho, en los
últimos años se ha sugerido una tendencia al reemplazo de los sistemas
forestales por matorrales, donde podría existir una pérdida en la calidad de los
ecosistemas (Lloret et al. 2002, Valladares et al. 2004). Además, cabe destacar
que la regeneración de los ecosistemas mediterráneos es altamente
dependiente de la disponibilidad hídrica (Lloret et al. 2004, Lloret et al. 2009) y
que, por lo tanto, estos procesos de degradación se podrían ver potenciados en
el marco de cambio climático actual (De Luis et al. 2001, Pausas 2004). Pero
sin duda, a parte del efecto en el funcionamiento del ecosistema, el efecto más
traumático del fuego es la pérdida de vidas humanas y de propiedades, así
como también de los elevados costes de su extinción.

10
- Los ecosistemas dominados por especies germinadoras obligadas en el oeste
de la cuenca mediterránea
El término vegetación natural potencial que define los estados sucesionales
maduros de la vegetación ha sido objeto de amplias controversias y críticas en
los últimos años debido a problemas asociados con su definición y con la
dinámica del ecosistema (ver Chiarucci et al. 2010 para una amplia discusión
del tema). Considerando esta controversia, se podría decir en sentido amplio y
solamente a modo de referencia que, los estados sucesionales más maduros
del oeste de la cuenca mediterránea (zonas de ombroclima seco de la
Comunitat Valenciana) deberían estar compuestos por bosques esclerófilos de
hoja ancha dominados por Quercus ilex (Barberó et al. 1992, Zavala et al 2000,
Zavala 2003, Quézel 2004). Sin embargo, este tipo de vegetación es muy
escasa en la actualidad debido a la explotación experimentada en esta zona
durante milenios y, solamente, aparecen pequeños rodales aislados y
dispersos a lo largo del paisaje. En su lugar, encontramos estados degradados
de esta vegetación cuya composición de especies varía en función del
substrato litológico. Sobre substratos calizos predominan comunidades
constituidas por especies rebrotadoras, con arbustos de enraizamiento
profundo que aprovechan las fisuras verticales de las rocas. Muchas de estas
plataformas calizas están ocupadas por coscojares (Quercus coccifera), con
pies arbustivos de carrasca (Q. ilex) más o menos disperso (Abad et al. 1996).
En las partes bajas de las vertientes donde se acumula más suelo,
especialmente sobre substrato margoso no consolidado, son frecuentes los
abancalamientos abandonados que atestiguan antiguos cultivos. La
combinación entre las propiedades intrínsecas de estos suelos y los efectos del
laboreo aplicado en el pasado, resultan en comunidades vegetales arbustivas y
arbóreas dominadas por especies germinadoras obligadas. Estos sistemas se
encuentran dominados por especies con una gran capacidad de colonización
típicas de estados sucesionales iniciales, tanto de porte arbóreo (Pinus
halepensis) como arbustivo (Ulex parviflorus, Cistus albidus y Rosmarinus
officinalis) (Abad et al. 1996, Baeza et al. 2007).
Los procesos de regeneración sobre antiguos campos de cultivos asociados
a especies germinadoras obligadas han supuesto un problema emergente en

11
las últimas décadas. En algunos casos, estas especies conforman matorrales
con una alta continuidad horizontal y vertical del combustible. Un caso bien
conocido son los aulagares dominados por U. parviflorus, que suponen uno de
las matorrales con mayor riesgo de incendio de la Comunitat Valenciana debido
a su alta capacidad de acumular combustible fino y muerto (Baeza et al. 2002,
De Luis et al. 2004, Duguy et al. 2007). Estos ecosistemas, además de suponer
un problema por el incremento en biomasa altamente inflamable en el paisaje,
han supuesto un aumento en la conectividad de las masas forestales que
anteriormente se encontraban separados por cultivo. Como consecuencia
directa, en las últimas décadas el régimen de incendios ha variado hacia una
mayor recurrencia de incendios y mayor superficie quemada en la Comunitat
Valenciana (Pausas 2004). Por esta razón, en los últimos años se han
realizado numerosos esfuerzos en conocer las bases ecológicas que rigen
estos ecosistemas (Baeza 2001, Baeza y Vallejo 2006, Verdú y Pausas 2007,
De Luis et al. 2008, Raventós et al. 2010) y el desarrollo de estrategias de
manejo (quemas controladas o desbroce) (Baeza 2001, Baeza et al. 2002,
Baeza et al. 2003, De Luis et al. 2005, Duguy et al. 2007, Baeza y Roy 2008).
Tradicionalmente, las especies germinadoras obligadas han sido clasificadas
dentro de un grupo homogéneo donde comparten una serie de atributos
comunes y que, en gran medida, contrastan con especies de diferente
estrategia regenerativa, como las especies rebrotadoras (Verdú 2000, Pausas
et al. 2004). Estas especies confían su regeneración post-fuego a partir de
semillas enterradas en el banco de semillas del suelo, generalmente, con
cubiertas duras que las hacen persistentes a las altas temperaturas (Paula y
Pausas 2008). Además, su germinación y establecimiento suele estar
favorecido por efectos directos (altas temperaturas, humo, fertilización por
cenizas; Baeza y Vallejo 2006, Moreira et al. 2010) e indirectos del fuego
(liberación de espacio, incremento de recursos del suelo, modificación del
espectro de luz, alternancia de temperaturas diaria en el suelo; Thanos y
Rundel 1995, DeBano et al. 1998, Baeza y Roy 2008) que favorecen su
emergencia y supervivencia. Tras el establecimiento, estas especies presentan
altas tasas de crecimiento de sus partes aéreas que les confiere un bajo ratio
raíz:tallo (Verdú 2000, Hernandez et al. 2010, Paula y Pausas en prensa).
Además, durante el crecimiento asignan gran parte de los nutrientes

12
disponibles a estructuras u órganos con alta tasa de renovación, como las
hojas (Carreira y Niell 1992, Saura-Mas y Lloret 2009). Habitualmente, las
hojas de las especies germinadoras no son esclerófilas, pero sus
características intrínsecas, junto con la estructura de su sistema radical (baja
proporción de biomasa repartida en raíces largas, ramificadas y finas que las
convierte en eficientes captadoras de recursos), confieren a este tipo de
especies alta capacidad de resistir el déficit hídrico estival (Paula y Pausas
2006, Saura-Mas y Lloret 2007, Paula y Pausas en prensa). Incluso, algunas
especies como U. parviflorus carecen de hojas reales y poseen tallos
fotosintéticos y espinosos altamente competitivas en zonas de alta radiación
(Valladares et al. 2003). Si bien es cierto, que los atributos morfológicos y
propiedades químicas de este tipo de especies les confieren una fácil ignición y
combustión de su biomasa. Las especies germinadoras obligadas son
consideradas altamente inflamables como consecuencia de su alta relación
muerto:vivo y fino:grueso en la estructura aérea de su combustible (Papió y
Trabaud 1991, Pereira et al. 1995, Dimitrakopoulos y Panov 2001,
Dimitrakopoulos 2001, Baeza et al. 2006, Saura-Mas et al. 2010, Baeza et al.
en prensa). Además, estas propiedades son variables a lo largo de su
ontogenia (Baeza et al. 2006, Baeza et al. en prensa) y se ha sugerido que la
retención de estos rasgos altamente inflamables podría ser el resultado de
presiones evolutivas asociadas al clima y el fuego (Saura-Mas et al. 2010).
- Avances en el conocimiento en los ecosistemas dominados por germinadoras
obligadas y propuestas de investigación.
A pesar de los recientes esfuerzos realizados en el estudio de los ecosistemas
de la cuenca mediterránea dominados por especies germinadoras obligadas,
todavía existe una falta de información de su funcionamiento en relación con la
actual tendencia de incremento de la recurrencia de incendios. Se ha sugerido
que, en ecosistemas propensos al fuego, las especies altamente inflamables
con una regeneración facilitada por el fuego podrían establecer bucles de
retroalimentación positivos que promovieran la expansión de las especies más
inflamables en detrimento de las menos inflamables (Wilson y Agnew 1992);
incluso, podrían establecer estados alternativos a los esperados en ausencia

13
de fuego (Bond et al. 2005). Dentro de este marco se podrían establecer los
ecosistemas de matorral dominados por especies germinadoras obligadas
(dominados por U. parviflorus, C. albidus y R. officinalis) resultantes del
abandono de cultivos o de la escasa regeneración post-fuego de formaciones
de pinar (Baeza 2001). Además, se ha sugerido que en este tipo de matorral
mediterráneo, con alta densidad de especies pioneras, se podrían establecer
mecanismos de inhibición de especies típicas de estados más avanzados,
arrestando el proceso natural de sucesión (Acacio et al. 2007, Siles et al.
2008). De hecho, especies altamente inflamables, propensas a su auto-
inmolación, y que inhiben la entrada de especies de estados más maduros
podrían ser consideradas un caso especial de reemplazo direccional entre
especies, en el cual, la sustitución de las especies de estados iniciales podría
estar impedido (Platt y Connell 2003). Por lo tanto, son necesarias nuevas
aproximaciones que determinen las trayectorias sucesionales de este tipo de
ecosistemas bajo escenarios de diferente recurrencia de incendio y que,
además, establezcan la posible existencia de procesos de retroalimentación
positivos entre las especies más inflamables de estados iniciales de la sucesión
y el fuego, en detrimento de aquellas pertenecientes a estados más maduros.
Este hecho puede alcanzar especial relevancia dentro del marco ecológico de
la restauración de este tipo de ecosistemas en la Comunitat Valenciana, donde
se pretenden promocionar trayectorias sucesionales que conduzcan hacia
estados dominados por especies rebrotadoras (Valdecantos et al. 2009), con
una mayor resiliencia al fuego y una menor acumulación de combustible fino
muerto (Baeza et al. en prensa).
Gran parte de las aproximaciones realizadas en estos ecosistemas en
relación con la recurrencia de incendios se han llevado a cabo a nivel
individual, usando modelos, u obviando el componente dinámico de estos
ecosistemas en el gradiente sucesional (Pausas 1999, Lloret et al. 2003, De
Luis et al. 2006). Trabajos previos han observado que tanto la recurrencia de
incendios como el intervalo de tiempo entre fuegos pueden determinar cambios
en la abundancia y composición de especies (Lloret et al. 2002, Delitti et al.
2005, Eugenio y Lloret 2006, Baeza et al. 2007, Vilà-Cabrera et al. 2008). Sin
embargo, es necesario integrar esta respuesta dentro de un concepto
sucesional. Aproximaciones comparativas que analizan un momento puntual

14
tras el incendio pueden obviar la variabilidad debida a procesos sucesionales
de sustitución entre especies (Connell y Slatyer 1977, Huston y Smith 1987).
Por lo tanto, son necesarios estudios que integren la variabilidad temporal de
las especies en la respuesta a la recurrencia de incendio; es decir, además de
los posibles cambios de abundancia, la edad en que las especies alcanzarían
su óptimo y los cambios en la amplitud de su nicho a lo largo del tiempo.
Generalmente, las especies germinadoras obligadas presentan atributos
funcionales pertenecientes a estados iniciales de la sucesión (Verdú 2000). Sin
embargo, existe una falta en el conocimiento de los posibles procesos
sucesionales que rigen estos ecosistemas y, por lo tanto, de la existencia de
posibles implicaciones en la asignación de recursos o historias de vida que
hagan unas especies más competitivas frente a otras en determinados
ambientes del gradiente sucesional. Un componente imprescindible para la
determinación de la dinámica sucesional es conocer el nicho de regeneración
de las especies que forman las comunidades (Grubb 1977); la dinámica de
establecimiento de plántulas revela el reemplazo potencial de los individuos
adultos en posteriores etapas (Harper 1977). Sin embargo, a pesar de que en
ecosistemas de la cuenca mediterránea se han observado patrones de
establecimiento de plántulas en etapas sucesionales entre fuegos (Clemente et
al. 1996, Lloret 1998, Lloret et al. 2005), la mayoría de trabajos sobre
regeneración en estas especies se han centrado solamente en la etapa
inmediatamente posterior al fuego, ignorando las etapas más tardías de la
sucesión (Baeza 2001, Quintana et al. 2004, De Luis et al. 2008).
Los cambios sucesionales en la composición de especies suelen estar
asociados a una función de inflamabilidad del combustible que dibuja en el
tiempo el riesgo de incendio (McCarthy et al. 2001). Las propiedades
intrínsecas del combustible de las especies germinadoras obligadas les
confieren una alta inflamabilidad (Saura-Mas et al. 2010); sin embargo, esta
inflamabilidad varia tanto entre las especies como a lo largo de su ontogenia
(Baeza et al. 2006, Baeza et al. in press). Concretamente, especial relevancia
se ha atribuido a la capacidad de acumulación de combustible muerto fino, ya
que estos combustibles disminuyen los tiempos de ignición y facilitan la
combustión por su bajo contenido en humedad (Bond y van Wilgen 1996). Por
tanto, diferentes funciones de inflamabilidad podrían establecerse dependiendo

15
de las propiedades intrínsecas de cada una de las especies, de la composición
específica de la vegetación y del orden de reemplazo en el tiempo. McCarthy et
al. (2001) definieron una serie de funciones donde, por ejemplo, la
inflamabilidad permanecía invariable a lo largo del tiempo, crecía de forma
constante, crecía hasta alcanzar una asíntota, o bien, tras un incremento inicial,
ésta decrecía con el transcurso de la sucesión. Sin embargo, a pesar de la
importancia que la dinámica temporal del combustible puede tener en la
frecuencia de incendios (como en los ecosistemas mediterráneos de California;
Minnich y Chou 1997), esta función de inflamabilidad permanece sin ser
estimada en los ecosistemas dominados por especies germinadoras del oeste
de la cuenca mediterránea. Además, aparte del efecto que puede tener la
dinámica temporal del combustible en la frecuencia de incendios, el reemplazo
de especies con diferente grado de inflamabilidad podría conllevar cambios en
la severidad del fuego y/o efectos en el ecosistema (Keeley 2009). Los efectos
del fuego en el ecosistema suelen estar ligados a un calentamiento significativo
del suelo (en superficie y profundidad) que afecta a las estructuras de
persistencia de las plantas o a las propiedades del suelo (Bradstock y Auld
1995, Certini 2005, De Luis et al. 2005, Baeza y Vallejo 2006). Por lo tanto, son
necesarios nuevos estudios que clarifiquen estos efectos dependiendo de la
especie dominante en el dosel. Además, cabría determinar que característica
morfológica y/o estructural del combustible es la más relevante en generar
estos efectos; ya que, estudios previos han observado que estos efectos
pueden estar vinculados, simplemente, a características individuales del
combustible como la arquitectura de las ramas muertas de los estratos
inferiores de la planta (Schwilk 2003) o a combustibles finos acumulados sobre
la superficie del suelo (Bradstock y Auld 1995).
No obstante, ecosistemas propensos al fuego y con una importante
presencia de especies germinadoras obligadas no son exclusivos de la cuenca
mediterránea. Estas especies pueden alcanzar una cierta relevancia en
ecosistemas de otras regiones de California, Sudáfrica o Australia (Bell et al.
1993, Keeley 1995), o incluso, en ecosistemas con clima no mediterráneo (Auld
y O’Connell 1991). Por lo tanto, el estudio de estos ecosistemas en la cuenca
mediterránea puede contribuir a determinar la ecología de este grupo funcional
de una forma más transversal y global. Sin embargo, nuevos estudios

16
centrados en este tipo de especies de diferentes regiones geográficas serían
necesarios para la realización de estudios comparativos entre regiones.
Por lo tanto, el objetivo general de esta tesis es indagar en los procesos
sucesionales que rigen los ecosistemas dominados por especies germinadoras
obligadas en el oeste de la cuenca mediterránea. Especialmente en: (1) los
mecanismos que controlan el establecimiento de individuos a lo largo de la
sucesión, (2) los patrones de abundancia de las especies dominantes en
función del tiempo desde el incendio y de la recurrencia de incendio, (3) la
dinámica de los diferentes tipos de combustible asociada a los cambios
sucesionales, (4) la variabilidad en los efectos del fuego en el ecosistema
dependiendo de la especie dominante en el dosel y que característica
estructural del combustible sería la más influyente sobre estos mismos efectos
y, (5) determinar los efectos del régimen de temperatura después del fuego en
la dormancia y germinación de seis especies germinadoras obligadas del
sureste de Australia. Para ello se han realizado diferentes trabajos de campo
en el interior de la Comunitat Valenciana. Estas aproximaciones se detallan a
continuación donde, además, se define la estructura de la tesis por capítulos.
- Capítulo 3. Se evalúa el papel de las diferentes características estructurales
del combustible de las especies dominantes en la determinación de los
posibles efectos sobre el ecosistema. Concretamente, se compara si existen
diferencias entre especies en las características estructurales del
combustible como su distribución por tamaños, la retención de combustible
muerto, la altura del combustible o su densidad aparente. Se analiza si las
diferencias entre las características estructurales del combustible se
traducen en diferencias en el consumo de combustible o en las temperaturas
del suelo. Determina que rasgos estructurales del combustible son los más
importantes modulando las temperaturas del suelo.
- Capítulo 4 . Se profundiza en el nicho de regeneración de las principales
especies de matorral, determinando cual es el patrón de establecimiento a lo
largo de la sucesión y cuales son los factores (bióticos y abióticos) que
dirigen la disponibilidad de micro-hábitats adecuados para el establecimiento

17
de plántulas. La hipótesis inicial es que el fuego desencadena una alta
disponibilidad de micro-hábitats, estableciéndose la mayoría de individuos
en etapas inmediatamente post-fuego; en etapas más tardías, la
disponibilidad de micro-hábitats decrecerá y el establecimiento estará
altamente restringido.
- Capítulo 5. En este capítulo se modeliza la respuesta sucesional de las
principales especies y formas vitales. Se determina la función de
inflamabilidad de estos ecosistemas a lo largo del tiempo en función del
desarrollo del dosel leñoso, la acumulación de combustible muerto y la
presencia de herbáceas. Valora el papel de la recurrencia de incendios en la
respuesta de las especies dominantes y cómo esta afecta globalmente la
inflamabilidad del ecosistema. Específicamente trata de comprobar la
hipótesis de que las especies que acumulan mayor cantidad de combustible
muerto y que tienen una germinación estimulada por el fuego se verán
favorecidas por el incremento de la recurrencia de incendio. Se discute la
posible existencia de un bucle de retroalimentación positivo entre fuego e
inflamabilidad del ecosistema.
- Capítulo 6. Se determinan las posibles trayectorias sucesionales en campos
de cultivo abandonados en ausencia de fuego y bajo diferente grado de
recurrencia de incendio. La hipótesis inicial es que, a largo plazo y en
ausencia de incendios, la sucesión se dirigirá hacia comunidades dominadas
por especies esclerófilas de hoja ancha. En cambio, si la trayectoria
sucesional se ve afectada por fuego esta tendencia se podría ver desviada
dependiendo del régimen de incendios. Bajo regímenes de baja recurrencia,
con un solo incendio, la vegetación será capaz de volver a su estado previo
sin desviaciones significativas de la trayectoria esperada en ausencia de
fuegos. Sin embargo, bajo regímenes de alta recurrencia de incendio la
sucesión de la vegetación será desviada hacia posibles estados alternativos,
dominados por especies que pueden establecer un bucle de
retroalimentación positivo con el fuego y donde la entrada de especies de
estados sucesionales más tardíos se puede ver limitada.

18
- Capítulo 7. Se indaga en los factores que controlan la germinación en
especies germinadoras obligadas pertenecientes a regiones geográficas
diferente a la cuenca mediterránea. Concretamente, se estudia si el régimen
de temperaturas diarias del suelo después del paso del fuego desempeña un
papel importante en la ruptura de la dormancia física de algunas especies de
la familia Fabaceae del sureste de Australia.
BIBLIOGRAFÍA
Abad N, Caturla RN, Baeza MJ, Bladé C, Vieira F, Carbó E, Valdecantos A,
Bonet A, Serrasolses I, Guàrdia R, Raventós J, Alloza JA, Escarré A,
Bellot J, Vallejo VR (1996) Regeneración de los montes quemados. En:
Vallejo VR (ed.) La restauración de la cubierta vegetal en la Comunidad
Valenciana. Fundación CEAM – Generalitat Valenciana, Valencia.
Acacio V, Holmgren M, Jansen P, Schrotter O (2007) Multiple recruitment
limitation causes arrested succession in Mediterranean oak systems.
Ecosystems 10: 1220-1230.
Auld TD, O’Connell MA (1991) Predicting patterns of post-fire germination in 35
eastern Australian Fabaceae. Australian Journal of Ecology 16: 53-70.
Baeza MJ (2001) Aspectos ecológicos y técnicas de control de combustible
(roza y quema controlada) en matorrales con alto riesgo de incendio
dominados por Ulex parviflorus (Pourr.). Tesis Doctoral, Departamento
de Ecología. Universidad de Alicante.
Baeza MJ, De Luís M, Raventós J, Escarré A (2002) Factors influencing fire
behaviour in shrublands of different stand ages and the implications for
using prescribed burning to reduce wildfire risk. Journal of
Environmental Management 65: 199-208.
Baeza MJ, Raventós J, Escarré A, Vallejo VR (2003) The effect of shrub
clearing on the control of the fire-prone species Ulex parviflorus. Forest
Ecology and Management 186: 47-59.
Baeza MJ, Vallejo VR (2006) Ecological mechanisms involved in dormancy
breakage in Ulex parviflorus seeds. Plant Ecology 183: 191-205.

19
Baeza MJ, Raventós J, Escarré A, Vallejo VR (2006) Fire risk and vegetation
structural dynamics in Mediterranean shrubland. Plant Ecology 187: 189-
201.
Baeza MJ, Valdecantos A, Alloza JA, Vallejo VR (2007) Human disturbance
and environmental factors as drivers of long-term post-fire regeneration
patterns in Mediterranean forests. Journal of Vegetation Science 18: 243-
252.
Baeza MJ, Roy J (2008) Germination of an obligate seeder (Ulex parviflorus)
and consequences for wildfire management. Forest Ecology and
Management 256: 685-693.
Baeza MJ, Santana VM, Pausas JG, Vallejo VR (en prensa) Successional
trends in standing dead biomass in Mediterranean Basin species. Journal
of Vegetation Science.
Barberó M, Loisel R, Quézel P (1992) Biogeography, ecology and history of
Mediterranean Quercus ilex ecosystems. Vegetatio 99-100: 19-34.
Bell DT, Plummer JA, Taylor SK (1993) Seed germination ecology in
southwestern western Australia. Botanical Review 59: 24-73.
Blondel J, Aronson J (1999) Biology and wildlife of the Mediterranean region.
Oxford University press, New York.
Blondel J (2006) The ‘design’ of Mediterranean landscapes: a millennial story of
humans and ecological systems during the historic period. Human
Ecology 34: 713-729.
Bond WJ, van Wilgen BW (1996) Fire and plants. Chapman & Hall, London.
Bond WJ, Woodward FI, Midgley GF (2005) The global distribution of
ecosystems in a world without fire. New Phytologist 165: 525-538.
Bradstock RA, Auld TD (1995) Soil temperatures during experimental bushfires
in relation to fire intensity: consequence for legume germination and fire
management in south-eastern Australia. Journal of Applied Ecology 32:
76-84.

20
Carreira JA, Niell FX (1992) Plant nutrient changes in a semiarid Mediterranean
shrubland after fire. Journal of Vegetation Science 3: 457-466.
Certini G (2005) Effects of fire on properties of forests soils: a review. Oecologia
143: 1-10.
Chiarucci A, Araújo MB, Decocq G, Beierkuhnlein C, Fernández-Palacios JM
(2010) The concept of potential natural vegetation: an epitaph? Journal
of Vegetation Science 21: 1172-1178.
Clemente AS, Rego FC, Correia OA (1996) Demographic patterns and
productivity of post-fire regeneration in Portuguese Mediterranean
Maquis. International Journal of Wildland Fire 6: 5-12.
Connell JH, Slatyer RO (1977) Mechanisms of succession in natural
communities and their role in community stability and organization.
American Naturalist 111: 1119-1144.
Cramer VA, Hobbs RJ, Standish RJ (2008) What’s new about old fields? Land
abandonment and ecosystem assembly. Trends in Ecology and
Evolution 23: 104-112.
Cruz A, Moreno JM (2001) Seasonal course of total non-structural
carbohydrates in the lignotuberous Mediterranean-type shrub Erica
australis. Oecologia 128: 343-350.
DeBano LF, Neary DG, Ffolliott PF (1998) Fire’s effects on Ecosystems. John
Wiley & Sons, New York.
Delitti W, Ferran A, Trabaud L, Vallejo VR (2005) Effects of fire recurrence in
Quercus coccifera L. shrublands of the Valencia Region (Spain): I. Plant
composition and productivity. Plant Ecology 177: 57-70.
De Luís M, García-Cano MF, Cortina J, Raventós J, González-Hidalgo JC,
Sánchez JR (2001) Climatic trends, disturbances and short-term
vegetation dynamics in a Mediterranean shrubland. Forest Ecology and
Management 147: 25-37.

21
De Luís M, Baeza MJ, Raventós J, González-Hidalgo JC (2004) Fuel
characteristics and fire behaviour in mature Mediterranean gorse
shrublands. International Journal of Wildland Fire 13: 79-87.
De Luís M, Raventós J, González-Hidalgo JC (2005) Factors controlling
seedling germination after fire in Mediterranean gorse shrublands.
Implications for fire prescription. Journal of Environmental Management
76: 159-166.
De Luís M, Raventós J, González-Hidalgo JC (2006) Post-fire vegetation
succession in Mediterranean gorse shrublands. Acta Oecologica 30: 54-
61.
De Luís M, Raventós J, Wiegand T, González-Hidalgo JC (2008) Temporal and
spatial differentiation in seedling emergence may promote species
coexistence in Mediterranean fire-prone ecosystems. Ecography 31: 620-
629.
Dimitrakopoulos AP (2001) A statistical classification of Mediterranean species
based on their flammability components. International Journal of Wildland
Fire 10: 113-118.
Dimitrakopoulos AP, Panov PI (2001) Pyric properties of some dominant
Mediterranean vegetation species. International Journal of Wildland Fire
10: 23-27.
Domínguez L, Calvo L, de Luis E (2002) The impact of wildfire season on
regeneration of Quercus pyrenaica forest and Pinus sp. stands. Journal
of Mediterranean Ecology 3: 47-54.
Duguy B, Alloza JA, Röder A, Vallejo VR, Pastor F (2007) Modelling the effects
of landscape fuel treatments on fire growth and behaviour in a
Mediterranean landscape (eastern Spain). International Journal of
Wildland Fire 16: 619-632.
Eugenio M, Lloret F (2006) Effects of repeated burning on Mediterranean
communities of the northeastern Iberian Peninsula. Journal of Vegetation
Science 17: 755-764.

22
Fox MD, Fox BJ (1987) The role of fire in scleromorphic forests and shrublands
of eastern Australia. En: Trabaud L, (ed.). The role of fire in ecological
systems. SPB, The Hague.
Grubb PJ (1977) The maintenance of species-richness in plant communities:
the importance of the regeneration niche. Biological Reviews 52: 107-
145.
Hanes TL (1971) Succession after fire in the chaparral of southern California.
Ecological Monographs 41: 27-52.
Harper JL (1977) Population biology of plants. Academic press, London.
Hernandez EI, Vilagrosa A, Pausas JG, Bellot J (2010) Morphological traits and
water use strategies in seedlings of Mediterranean coexisting species.
Plant Ecology 207: 233-244.
Herranz JM, Ferrandis P, Martínez-Sánchez JJ (1999) Influence of heat on
seed germination of nine woody Cistaceae species. International Journal
of Wildland Fire 9: 173-182.
Hobbs RJ, Arico S, Aronson J, Baron, Bridgewater P, Cramer VA, Epstein PR,
Ewel JJ, Klink CA, Lugo AE, Norton D, Ojima D, Richardson DM,
Sanderson EW, Valladares F, Vilà M, Zamora R, Zobel M (2006) Novel
ecosystems: a theorethical and management aspects of the new
ecological world order. Global Ecology and Biogeography 15: 1-7.
Huston M, Smith T (1987) Plant succession: life history and competition. The
American Naturalist 130: 168-198.
Johnson EA (1992) Fire and vegetation dynamics: studies from the North
American boreal forests. Cambridge University Press, Cambridge.
Keeley JE (1986) Resilience of Mediterranean shrub communities to fires. En:
Dell B, Hopkins AJM, Lamont BB (eds.) Resilience in Mediterranean-type
Ecosystems. Dr W. Junk Publishers, Dordrecht.
Keeley JE (1995) Seed-germination patterns in fire-prone Mediterranean-
climate regions. En: Arroyo MTK, Zedler PH, Fox MD (eds.) Ecology and
biogeography of Mediterranean ecosystems in Chile, California and
Australia. Springer-Verlag, New York.

23
Keeley JE (2009) Fire intensity, fire severity, and burn severity: a brief review
and suggested usage. International Journal of Wildland Fire 18: 116-126.
Le Houérou HN (1993) Land degradation in Mediterrranean Europe: can
agroforestry be a part of the solution? A prospective review. Agroforestry
Systems 21: 43-61.
Lloret F (1998) Fire, canopy cover and seedling dynamics in Mediterranean
shrubland of northeastern Spain. Journal of Vegetation Science 9: 417-
430.
Lloret F (2004) Régimen de incendios y regeneración. En: Valladares F (ed.)
Ecología del bosque mediterráneo en un mundo cambiante. Ministerio de
Medio Ambiente, Madrid.
Lloret F, López-Soria L (1993) Resprouting of Erica multiflora after experimental
fire treatments. Journal of Vegetation Science 4: 367-374.
Lloret F, Calvo E, Pons X, Díaz-Delgado R (2002) Wildfires and landscape
patterns in the eastern Iberian Peninsula. Landscape Ecology 17: 745-
759.
Lloret F, Pausas JG, Vilà M (2003) Responses of Mediterranean plant species
to different fire frequencies in Garraf Natural Park (Catalonia, Spain):
field observations and modelling predictions. Plant Ecology 167: 223-
235.
Lloret F, Peñuelas J, Estiarte M (2004) Experimental evidence of reduced
diversity of seedlings due to climate modification in a Mediterranean-type
community. Global Change Biology 10: 248-258.
Lloret F, Peñuelas J, Estiarte M (2005) Effects of vegetation canopy and climate
on seedling establishment in Mediterranean shrubland. Journal of
Vegetation Science 16: 67-76.
Lloret F, Peñuelas J, Prieto P, Llorens L, Estiarte M (2009) Plant community
changes induced by experimental climate change: seedling and adult
species composition. Perspectives in Plant Ecology, Evolution and
Systematics 11: 53-63.

24
McCarthy MA, Gill AM, Bradstock RA (2001) Theoretical fire interval
distribution. International Journal of Wildland Fire 10: 73-77.
Minnich RA, Chou YH (1997) Wildland fire patch dynamics in the chaparral
southern California and northern Baja California. International Journal of
Wildland Fire 7: 221-248.
Moreira B, Tormo J, Estrelles E, Pausas JG (2010) Disentangling the role of
heat and smoke as germination cues in Mediterranean Basin flora.
Annals of Botany 105: 627-635.
Mouillot F, Field CB (2005) Fire history and the global carbon budget: a 1º x 1º
fire history reconstruction for the 20th century. Global Change Biology
11: 398-420.
Papió C, Trabaud L (1991) Comparative study of the aerial structure of five
shrubs of Mediterranean shrublands. Forest Science 37: 146-159.
Paula S, Pausas JG (2006) Leaf traits and resprouting ability in the
Mediterranean basin. Functional Ecology 20: 941-947.
Paula S, Pausas JG (2008) Burning seeds: germinative response to heat
treatments in relation to resprouting ability. Journal of Ecology 96: 543-
552.
Paula S, Pausas JG (en prensa) Root traits explain different foraging strategies
between resprouting life histories. Oecologia
Pausas JG (1999) Response of plant functional types to changes in the fire
regime in Mediterranean ecosystems: a simulation approach. Journal of
Vegetation Science 10: 717-722.
Pausas JG (2004) Changes in fire and climate in the eastern Iberian Peninsula
(Mediterranean Basin). Climatic Change 63: 337-350.
Pausas JG, Bradstock RA, Keith DA, Keeley JE, Network GF (2004) Plant
functional traits in relation to fire in crown-fire ecosystems. Ecology 85:
1085-1100

25
Pereira JM, Sequeira NM, Carreiras JM (1995) Structural properties and
dimensional relations of some Mediterranean shrub fuels. International
Journal of Wildland Fire 5: 35-42.
Platt WJ, Connell JH (2003) Natural disturbances and directional replacement
of species. Ecological Monographs 73: 507-522.
Quézel P (2004) Large-scale post-glacial distribution of vegetation structure in
the Mediterranean region. En: Mazzoleni S, Di Pasquale G, Mulligan M,
Di Martino P, Rego F (eds.) Recent dynamics of the Mediterranean
vegetation and landscape. J. Wiley & Sons, Chichester.
Quintana JR, Cruz A, Fernández-González F, Moreno JM (2004) Time of
germination and establishment success after fire of three obligate
seeders in a Mediterranean shrubland of central Spain. Journal of
Biogeography 31: 241-249.
Raventós J, Wiegand T, De Luis M (2010) Evidence for the spatial segregation
hypothesis: a test with nine-year survivorship data in a Mediterranean
shrubland. Ecology 91: 2110-2120.
Rodrigo A, Retana J, Picó FX (2004) Direct regeneration is not the only
response of Mediterranean forests to large fires. Ecology 85: 716-729.
Saura-Mas S, Lloret F (2007) Leaf and shoot water content and leaf dry matter
content of Mediterranean woody species with different post-fire
regenerative strategies. Annals of Botany 99: 545-554.
Saura-Mas S, Lloret F (2009) Linking post-fire regenerative strategy and leaf
nutrient content in Mediterranean woody plants. Perspectives in Plant
Ecology, Evolution and Systematics 11: 219-229.
Saura-Mas S, Paula S, Pausas JG, Lloret F (2010) Fuel loading and
flammability in the Mediterranean Basin woody species with different
post-fire regenerative strategies. International Journal of Wildland Fire
19: 783-794.
Schwilk DW (2003) Flammability is a niche construction trait: canopy
architecture affects fire intensity. The American Naturalist 162: 725-733.

26
Siles G, Rey PJ, Alcántara JM, Ramírez JM (2008) Assessing the long-term
contribution of nurse plants to restoration of Mediterranean forests
through Markovian models. Journal of Applied Ecology 45: 1790-1798.
Suding KN, Gross KL, Houseman GR (2004) Alternative status and positive
feedbacks in restoration ecology. Trends in Ecology and Evolution 19:
46-53.
Thanos CA, Rundel PW (1995) Fire-followers in chaparral: nitrogenous
compounds trigger seed germination. Journal of Ecology 83: 207-216.
Trabaud L, Lepart J (1980) Diversity and stability in garrigue ecosystems after
fire. Vegetatio 43: 49-57.
Valdecantos A, Baeza MJ, Vallejo VR (2009) Vegetation management for
promoting ecosystem resilience in fire-prone Mediterranean shrublands.
Restoration Ecology 17: 414-421.
Valladares F, Hernandez LG, Dobarro I, García-Pérez C, Sanz R, Pugnaire FI
(2003) the ratio of leaf to total photosinthetic area influences shade
survival and plastic response to light of green-stemmed leguminous
shrub seedlings. Annals of Botany 91: 577-584.
Valladares F, Camarero JJ, Pulido F, Gil-Pelegrín E (2004) El bosque
mediterráneo, un sistema humanizado y dinámico. En: Valladares F (ed.)
Ecología del bosque mediterráneo en un mundo cambiante. Ministerio de
Medio Ambiente, Madrid.
Verdú M (2000) Ecological and evolutionary differences between Mediterranean
seeders and resprouters. Journal of Vegetation Science 11: 265-268.
Verdú M, Pausas JG (2007) Fire drives phylogenetic clustering in
Mediterranean Basin woody plant communities. Journal of Ecology 95:
1316-1323.
Vilà-Cabrera A, Saura-Mas S, Lloret F (2008) Effects of fire frequency on
species composition in a Mediterranean shrubland. Ecoscience 15: 519-
528.
Vitousek PM, Money HA, Lubchenco J, Melillo J (1997) Human domination of
Earth’s ecosystems. Science 277: 494-499.

27
Wilson JB, Agnew ADQ (1992) Positive-feedback switches in plant-
communities. Advances in Ecological Research 23: 263-336.
Zavala MA, Espelta JM, Retana J (2000) Constraints and trade-offs in
Mediterranean plant communities: the case of holm oak-Aleppo pine
forests. Botanical Review 66: 119-149.
Zavala MA (2003) Dinámica y sucesión en bosques mediterráneos: modelos
teóricos e implicaciones para la selvicultura. En: Rey-Benayas JM,
Espigares T, Nicolau JM (eds.) Restauración de ecosistemas
mediterráneos. Universidad de Alcalá, Madrid.
Zedler PH, Gautier CR, McMaster GS (1983) Vegetation change in response to
extreme events: the effect of a short interval between fires in California
chaparral and coastal scrub. Ecology 64: 809-818.

28

29
CAPÍTULO 2- ÁREA DE ESTUDIO

30

31
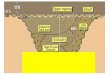
CAPÍTULO 2 - ÁREA DE ESTUDIO El área de estudio se encuentra en el interior de las provincias de Alicante y
Valencia, en el sureste de la Península Ibérica (Figura 1). Geológicamente, el
área corresponde a las últimas estribaciones de las cordilleras Béticas,
concretamente a la zona Prebética. En el área de estudio predominan los
materiales de origen sedimentario y de naturaleza caliza. Excluyendo los
depósitos aluviales y coluviales, los substratos calizos suponen
aproximadamente un 55% del territorio forestal y los margosos un 35%. Las
condiciones climáticas secas en el área de estudio determinan las
características de los suelos, y por ejemplo, las calizas presentan suelos rojos
fisurales (Leptosoles, Cambisoles y Luvisoles; FAO 1988) en diferente grado de
descarbonatación y, normalmente son pedregosos, bien estructurados y poco
erosionables. Estos suelos, no han sido cultivados intensamente en los medios
forestales y su capacidad de infiltración es muy elevada. Por otro lado, el
segundo gran tipo de substrato esta formado por margas y arcilla, que dan
Figura 1. Área de estudio. Los números corresponden con el identificador
de las diferentes parcelas.
1
2
3
4
5 6
7 8
9 10
12
11 13
14
Km

32
lugar a coluvios más o menos pedregosos, de profundidad media, de subsuelo
compacto y poco fisural. Los suelos suelen ser de tipo Regosol calcáreo (FAO
1989). En la Comunidad Valenciana, una gran parte de los suelos
desarrollados sobre substratos margosos no están consolidados y han sido
cultivados en algún momento (Abad et al. 1996).
En el área de estudio predomina el clima típicamente Mediterráneo, bajo
condiciones de ombroclima seco (precipitación media anual: 350-600 mm;
Pérez-Cueva 1994). El termoclima contempla un tipo ampliamente
representado en la Comunidad Valenciana: el mesomediterráneo (Tª media
anual 13-17ºC). Sin embargo, hay que exceptuar una zona de estudio
(Guadalest, Tabla 1, Figura 1) que presenta un termoclima termomediterráneo
(Tª media anual 17-19ºC). Investigaciones previas han mostrado que en las
últimas décadas se está observando una tendencia al descenso de la
precipitación estival y un claro patrón de incremento de la temperatura media
anual y estival (De Luis et al. 2001, Pausas 2004).
En general, el tipo de substrato es un determinante clave de la vegetación en el
área de estudio. Sobre substratos calizos predominan comunidades
constituidas por especies rebrotadoras, principalmente de Q. coccifera (Abad et
al. 1996). En substrato no consolidado y margoso, son frecuentes los
abancalamientos abandonados, donde la vegetación dominante esta
constituida por especies germinadoras. Estos sistemas se encuentran
dominados por especies tanto de porte arbóreo (Pinus halepensis Mill.) o
arbustivo (Ulex parviflorus Pourr., Cistus albidus L. y Rosmarinus officinalis L.)
con una gran capacidad de colonización, típicas de estados iniciales de la
sucesión (Abad et al. 1996, Baeza et al. 2007). No menos de un 30% de la
superficie forestal actual de la Comunidad Valenciana ocupa abancalamientos
que se hallan en un estado generalizado de desmantelamiento, especialmente
los muretes de contención de piedra seca. En casos extremos, los
abancalamientos han desaparecido por completo quedando el substrato
litológico descarnado.

33
Identificador Nombre nº fuegos Altura (m) Pendiente (º) Orientación Suelo 1er
Fuego 2o Fuego 3
er Fuego XUTM YUTM Capítulo
1 Ayora a 2 833 28 ENE Margo-calizo 1979 1985 - 682077 4339697 5
2
Ayora b
0 763 10 SSO Margo-calizo - - - 679798 4334808 6
1 1041 31 NNE Margas 1979 - - 676400 4332099 4, 5, 6
2 1041 23 NE Margas 1979 1996 - 676684 4332010 4, 5, 6
3 1041 23 NE Margas 1979 1996 2006 676812 4331950 3, 4, 6
3 Ayora c 2 831 6 O Margas 1979 1991 - 676537 4322175 5
4 Ayora d 2 735 14 E Calizas 1979 1984 - 687777 4308441 5
5
Fontanars
1 710 30 NO Margo-calizo 1978 - - 697715 4293747 5
2 718 25 O Calizas 1978 1984 - 696615 4294317 5
6 Banyeres 1 820 14 NO Margas 1991 - - 704587 4289588 5
7
Onil
0 940 47 ENE Margas - - - 703401 4280552 6
1 940 35 NO Margas 1984 - - 703392 4280698 4, 5, 6
2 940 35 NO Margas 1984 1994 - 703339 4280706 4, 5, 6
3 940 35 NO Margas 1984 1994 2006 703338 4280686 3, 4, 6
8 La Venteta 0 997 22 NE Margas - - - 709253 4281051 6
9
Pardines
0 900 4 N Margas - - - 711154 4283127 6
1 900 4 N Margas 1984 - - 711302 4283187 4, 5, 6
2 900 4 N Margas 1984 1994 - 711215 4283194 4, 5, 6
3 900 4 N Margas 1984 1994 2006 711202 4283207 4, 6, 3
10 Fontroja 0 834 32 NE Margas - - - 716895 4283094 6
11 Els Plans 0 920 11 NO Margas - - - 721637 4280334 6
12
La Torre
1 840 16 N Margas 1984 - - 725918 4276893 5
2 840 16 N Margas 1984 1994 - 725936 4276847 5
13 Confrides 1 789 15 O Margas 1991 - - 738829 4285347 5
14 Guadalest 1 417 3 S Margas 1991 - - 744675 4283954 5
Tabla 1. Descripción de las zonas de estudio utilizadas en la tesis doctoral. El número identificador corresponde con la
ubicación en la figura 1.

34
Dentro del área de estudio se seleccionaron 14 zonas ubicadas sobre antiguos
campos de cultivo abandonados a lo largo del siglo XX (Tabla 1). Estas zonas
han sido colonizadas por especies germinadoras, formando inicialmente
bosques de P. halepensis (excepto la zona de estudio de Ayora, que
corresponde a una formación mixta con Pinus pinaster Ait.). Sin embargo, parte
de estas zonas han sido objeto de diferentes incendios forestales desde 1978
(Tabla 1). Además, en años posteriores han ocurrido fuegos recurrentes dentro
de las áreas previamente quemadas y, en la actualidad, las zonas
seleccionadas conforman un mosaico de sistemas entre bosque y matorral en
diferente grado de desarrollo. Por lo tanto, se dispone de parcelas con un
diferente número de recurrencia de incendio y con características climáticas,
ambientales y de substrato similares entre ellas (Tabla 1). Por ejemplo, en
Ayora b, Onil y Pardines se dispone de cuatro parcelas con 0, 1, 2 y 3
incendios cada una. El tercer fuego sobre estas parcelas fue una quema
experimental realizada expresamente para esta tesis doctoral (ver capitulo 6).
La zona de Ayora corresponde a uno de los incendios más catastróficos de la
cuenca mediterránea, que en 1979 arrasó aproximadamente unas 30.000
hectáreas. Su gran extensión nos ha permitido establecer en su interior 4 zonas
de estudio diferentes con fuegos recurrentes independientes (Ayora a, Ayora b,
Ayora c y Ayora d; ver Tabla 1 para más detalles). Parte de las zonas utilizados
en esta tesis han formado parte de estudios previos dentro de proyectos de
investigación desarrollados por el Departamento de Restauración Forestal del
CEAM, donde se incluyen tanto estudios de dinámica de la vegetación como
de manejo de la vegetación para reducir el riesgo de incendio (Baeza 2001,
Baeza et al. 2002, Baeza et al. 2007). La altura sobre el nivel del mar de estas
zonas abarca entre 710-1041 m y se seleccionaron evitando exposiciones sur
para reducir la variabilidad ambiental. Sin embargo, hay que resaltar que la
zona de Guadalest es una excepción ya que se encuentra a 417 m de altura y
exposición sur.

35
BIBLIOGRAFÍA
Abad N, Caturla RN, Baeza MJ, Bladé C, Vieira F, Carbó E, Valdecantos A,
Bonet A, Serrasolses I, Guàrdia R, Raventós J, Alloza JA, Escarré A,
Bellot J, Vallejo VR (1996) Regeneración de los montes quemados. En:
Vallejo VR (ed.) La restauración de la cubierta vegetal en la Comunidad
Valenciana. Fundación CEAM – Generalitat Valenciana, Valencia.
Baeza MJ (2001) Aspectos ecológicos y técnicas de control de combustible
(roza y quema controlada) en matorrales con alto riesgo de incendio
dominados por Ulex parviflorus (Pourr.). Tesis Doctoral, Departamento
de Ecología. Universidad de Alicante.
Baeza MJ, De Luís M, Raventós J, Escarré A (2002) Factors influencing fire
behaviour in shrublands of different stand ages and the implications for
using prescribed burning to reduce wildfire risk. Journal of
Environmental Management 65: 199-208.
Baeza MJ, Valdecantos A, Alloza JA, Vallejo VR (2007) Human disturbance
and environmental factors as drivers of long-term post-fire regeneration
patterns in Mediterranean forests. Journal of Vegetation Science 18: 243-
252.
De Luís, M., García-Cano, M.F., Cortina, J., Raventós, J., González-Hidalgo,
J.C., Sánchez, J.R. 2001. Climatic trenes, disturbances and short-term
vegetation dynamics ina Mediterranean shrubland. Forest Ecology and
Management 147: 25-37.
FAO 1988. Soil map of the world. Revised legend. World soil resources report,
60. 99 pp. Rome.
Pérez-Cueva A (1994) Atlas climàtic de la Comunitat Valenciana. Direcció
General d’Ubanisme i Ordenació del Territori, Generalitat Valenciana.
Col·lecció Territori, Nº4.
Pausas, J.G. 2004. Changes in fire and climate in the eastern Iberian Peninsula
(Mediterranean Basin). Climatic Change 63: 337-350.

36

37
CAPÍTULO 3-
CARACTERÍSTICAS ESTRUCTURALES DEL COMBUSTIBLE
MODULANDO LAS TEMPERATURAS DEL SUELO EN PARCHES
DE DIFERENTES ESPECIES DE MATORRAL DE LA CUENCA
MEDITERRÁNEA
RESUMEN: La habilidad de un combustible para arder está determinada por su inflamabilidad, una propiedad que varía entre especies y que está parcialmente determinada por diferentes características estructurales del combustible como el ratio superficie: volumen de ramas y hojas, la retención de de ramillas muertas, la arquitectura de su copa y la densidad aparente. Nuestro objetivo fue valorar el rol de estas características estructurales en la modulación de temperaturas bajo diferentes parches de vegetación. Los resultados mostraron que hubieron diferencias contrastadas el la estructura del combustible entre los parches de diferentes especies de matorral mediterráneo. Las diferencias en la estructura del combustible fueron importantes en la modulación de las temperaturas ya que, sin tener en cuenta el combustible total, las temperaturas más altas fueron encontradas con la mayor cantidad y densidad de combustible muerto. Sorprendentemente, las temperaturas bajo especies herbáceas fueron altas en relación a las especies arbustivas; el tiempo de residencia de sus temperaturas fue más corto. Por lo tanto, las estrategias de manejo que promuevan especies que acumulan menor cantidad de combustible fino muerto podría ser crucial para conseguir fuegos menos severos con un impacto reducido en el ecosistema.
Este capítulo reproduce el siguiente manuscrito: Santana VM, Baeza MJ, Vallejo VR (en prensa) Fuel structural traits modulating soil temperatures in different species-patches of Mediterranean Basin shrublands. International Journal of Wildland Fire

38

39
Fuel structural traits modulating soil temperatures in different
species-patches of Mediterranean Basin shrublands
Victor M. SantanaA, M. Jaime BaezaA, B, V. Ramón VallejoA
AFundación de la Generalitat Valenciana Centro de Estudios Ambientales del
Mediterráneo (CEAM). Parque Tecnológico Paterna. C/Charles Darwin, 14.
46980 Paterna, Valencia. Spain BDepartamento de Ecología, Universidad de Alicante. Ap. 99. 03080 Alicante.
Spain.
Abstract The ability of a fuel to burn is determined by its flammability, a property which varies from one species to another and is partially determined by different fuel structural traits such as surface-to-volume ratio of twigs and leaves, retention of standing dead twigs, canopy architecture and bulk density. Our aim was to assess the role of these fuel structural traits in modulating soil temperatures under different species patches. The results showed that there were contrasted differences in the fuel-structure complex among different species-patches in Mediterranean Basin shrublands. The differences in the fuel structure were important in modulating soil temperature since, regardless of the total fuel load, the highest temperatures were found under the species with the highest loads and densities of dead fine fuel. Surprisingly, temperatures under herbaceous species were high in relation to shrubby species; however, temperature-residence-times were shorter. Therefore, management strategies which promote species that accumulate low contents of fine dead fuel could be crucial for attaining less severe fires with a reduced impact on ecosystem functioning. Key words: Dead fuel, Fire behaviour, Fire intensity, Fire severity, Flammability.

40
1. Introduction
Management of high fire-risk vegetation is a
challenge for fire fighters and land managers,
especially in the next decades when forecasts
predict shifts in the fire regime as a result of
climate change (Pausas 2004; Mouillot and Field
2005; Westerling et al. 2006). Designing new fuel
treatment strategies to reduce fire hazard requires
further understanding of the functional
relationships between fuel characteristics and the
processes associated with fire, so that when a
wildfire ignites in a treated landscape, it spreads
more slowly, burns with less intensity and causes
fewer negative effects on the ecosystem
functioning (Conard et al. 2001; Fernandes and
Botelho 2003).
The ability of a fuel to burn is determined by its
flammability, a property that indicates how easily it
will ignite (ignitability), how quick the flames will
spread (combustibility) and how stable its burning
rate will be (sustainability) (Troumbis and Trabaud
1989). This property varies substantially from one
species to another, and it is determined by the
interaction of many different fuel structural traits
such as surface-to-volume ratio of twigs and
leaves, retention of standing dead twigs, canopy
architecture and bulk density (Cornelissen et al.
2003). Moreover, flammability can also be partially
modulated by the content of minerals, volatile oils,
waxes and resins in fuels (Philpot 1970;
Dimitrakopoulos and Panov 2001; Alessio et al.
2008). Fuel structural traits have been widely
analysed in laboratory studies aimed at classifying
species into possible fire-risk levels (Rothermel
1972; Papió and Trabaud 1991; Pereira et al.
1995; Dimitrakopoulos and Panov 2001;
Dimitrakopoulos 2001; Baeza et al. 2006; Saura-
Mas et al. 2010). However, there is still a lack of
knowledge linking the role of these species-
specific traits to fuel consumption processes in
real fires and their possible effects on ecosystem
functioning (Pérez and Moreno 1998; Molina and
Llinares 2001; Schwilk 2003).
Fire effects on ecosystem functioning are
mainly determined by ground and subsurface
heating which affects plant fire-persistence
structures, like seedbanks, rhizomes, buds and
bulbs, and alters soil properties and erosion
processes (Beadle 1940; Bradstock and Auld
1995; Brooks 2002; Certini 2005). Readily
measurable parameters such as the amount of
heat released by the physical combustion of fuel
(fire intensity sensu Keeley 2009) or the losses in
organic matter above and belowground (fire
severity sensu Keeley 2009) have often been
linked to fire effects on the ecosystem (Moreno
and Oechel 1991; Schimmel and Granström 1996;
Keeley et al. 2005); however, they are not always
good descriptors of fire impacts on the
ecosystems (Hartford and Frandsen 1992;
Bradstock and Auld 1995; Keeley and McGinnis
2007). The failure of these relationships has been
attributed to the very little radiant or convected
heat that is transferred from the combustion of
aerial fuels to the soil or, simply, to the fact that
soil temperatures are more dependent on a single
fuel structural trait, such as the architecture of the
dead branches in the lower strata of plants
(Schwilk 2003) or the fine fuels lying on the soil
surface (Bradstock and Auld 1995).
The aim of this work is to assess the role of
species-specific fuel structural traits in determining
soil temperatures in Mediterranean Basin
shrublands dominated by obligate seeders. As this
vegetation is characterised by very flammable
foliage with abundant dead woody material that
favours the rapid spread of fires, it has become
one of the most problematic vegetation types in
this area for fire managers (Duguy et al. 2007;
Saura-Mas et al. 2010). The existence of
individuals of the same species clumped together
on our study sites allowed us to establish patches
composed almost exclusively of a single species.
On these patches we also assessed the fuel traits
of the dominant species before setting three
experimental fires. Specifically, we assessed: 1)
whether the different species-patches show
differences in fuel structural traits such as fuel
distribution by size classes, standing dead fuel,
fuel height and fuel bulk density; 2) whether the

41
differences in fuel structural traits between species
produce changes in fire effects, such as fuel
consumption and soil temperatures; and 3) which
fuel structural traits are most important for
modulating soil temperatures.
2. Material and methods
2.1 Experimental fires
We selected three sites in the Valencia Region
(Spain) to apply different experimental fires (Table
1). These sites have been subjected to frequent
fire episodes, both natural and human-caused,
with the most recent ones being experimental fires
for fuel management studies (Baeza et al. 2002b).
At the time of our experimental fires, the
vegetation was a shrubland dominated by several
obligate-seeding species. The woody stratum was
mainly composed of the shrubs Cistus albidus L.
(Cistus hereafter), Rosmarinus officinalis L.
(Rosmarinus hereafter) and Ulex parviflorus Pourr.
(Ulex hereafter), whereas the herbaceous stratum
consisted of perennial resprouting species, mainly
the grass Brachypodium retusum (Pers.) Beauv.
(Brachypodium hereafter). There were a few small
isolated individuals of woody resprouting species
like Quercus coccifera L. and Juniperus oxycedrus
L. At each site, we selected one plot of
approximately 30 x 20 m where we set an
experimental fire. The areas to be burned were
previously delimited by a 4 m-wide fire break in
which the vegetation was eliminated through
mechanical brushing. All three sites were burned
in June 2006, and there was a one-week
separation between each experimental fire. Fires
were ignited as a line encompassing the entire
upwind flank of the experimental plot (headfires).
As a safety measure, both fire-fighters and forest
rangers were present for each experimental fire.
2.2 Fuel structure of species-patches
Prior to the experimental fires, the existence of
individuals of the same species clumped together
on each site allowed us to distinguish patches
composed almost exclusively of a single species.
Table 1. Description of the experimental plots, including weather conditions and fire behaviour variables for the three different sites during the experimental fires.
Onil Pardines Ayora
Latitude 38º39'N 38º40'N 39º07'N
Longitude 0º39'W 0º39'W 0º57'W
Slope (º) 35 4 23
Aspect NW N N
Previous fire events 1986, 1994 1986, 1994 1979, 1996
Fuel load (g·m-2)
(n=6)
B. retusum 68±51 35±41 112±66
C.albidus 427±419 385±476 411±264
R. officinalis 130±201 61±144 33±81
U. parviflorus 235±168 636±601 194±166
Other species 38±56 347±540 186±180
Litter 370±209 334±362 344±250
Total 1274±705 1814±648 1285±468
Fuel moisture (%)
(n=5)
B. retusum 20±2,7 20,1±6,4 32,3±1,7
C.albidus 46,6±1,9 49,2±2,8 53,4±2,8
R. officinalis 54,1±3,1 52,1±1,4 59,8±3,4
U. parviflorus 44,1±1,3 49,9±2,6 50,2±3,1
U. parviflorus dead 15,1±9,2 13,8±4,7 15,7±3
Soil 12±2,8 7,8±2,2 20±9,5
Litter 6,9±0,7 8,4±1,6 12±4,5
Air temperature (ºC) 21,3 25,1 24,1
Air HR (%) 42 53,1 57,3
Wind speed (m/s) 4,9 1,6 3,7
Rate of spread (m.s-1)
(n=3) 0.21±0.04 0.11±0.04 0.25±0.02
Fire-line intensity
(Kw m-1) 4060 3288 4185
Tmax (ºC)
(0 cm depth, n=3) 283±175 445±147 278±91
We selected five different patches for each
dominant species (Brachypodium, Cistus,
Rosmarinus and Ulex) within each experimental
fire. These species have contrasting differences in
morphological and fuel traits (Table 2). Thus, to
assess the differences in fuel structural
characteristics between patches, we marked off
one 0.5 x 0.5 m square in each patch and
characterised all the vegetation present (Figure 1).
For shrubs, we assessed the total fuel load and its

42
Table 2. Traits description for the dominant species.
Trait Species Reference
Brachypodium Cistus Rosmarinus Ulex
Vital form perennial graminoid shrub shrub shrub A
Leaf phenology evergreen drought semi-deciduous evergreen evergreen A
Leaf shape linear broad linear spines A
Surface to volume ratio (m2 m-3) 20 x 10-3 3,64 x 10-3 3,54 x 10-3 5,00 x 10-3 B, C
Mass to volume ratio (Kg m-3) 442 280 410 614 B, C
Heat of combustion (KJ Kg-1) 17638 19520 22296 21077 B, C
References: A= Paula et al. 2009; B= Cohen et al. 2003; C= Elvira and Hernando 1989
size class distribution on the basis of the
allometric relationships estimated for these
species in this area (for Ulex see Baeza et al.
2006; Cistus and Rosmarinus relations from M. J.
Baeza unpublished data). Allometric relationships
were applied to the basal stem diameters of the
individuals rooted inside each square. We
assumed that all the fuel of the plants rooted in
each 0.5 x 0.5 m square was contained within the
volume located over each square (Pérez and
Moreno 1998). We distinguished between four
different fuel fractions: on the one hand, fine live
and dead fuel (Ø <0.6 cm) and on the other hand,
coarse live and dead fuel (Ø >0. 6 cm). Leaves
and spines were included within the fine fuels.
Fuels larger than 2.5 cm were very scarce, and
they were included within the Ø >0.6 cm class. In
order to analyze the architecture of the fuel, we
assessed different fuel distribution parameters.
We measured total height as the visually averaged
top of the canopy of the individuals rooted inside
each square, disregarding occasional taller stems
(Figure 1). Fine dead fuel is usually concentrated
in the lower parts of plants, accumulating as the
plant grows and produces new live shoots in the
upper part of stems. This fact enabled us to
differentiate two strata in the vertical distribution of
the fuel-structure complex, the fine dead and the
fine live fuel stratas (Baeza et al. 2006). Thus, we
visually averaged the height of the fine dead fuel
layer from the ground level to the first branches
with live leaves (hereafter first live branches
height) (Figure 1). We assumed the total fuel bulk
density (FBD hereafter) to be the fuel inside the
volume comprised by the 0.5 x 0.5 m square
surface and the total height of individuals
(Rothermel 1972; Fernandes 2001). In addition,
we assumed the fine dead and green FBD to be
the fuel of these fractions comprised within the
lower and higher strata respectively delimited by
the height of the first live branches (Figure 1).
Spaces between patches of woody-shrubs
were dominated mainly by Brachypodium.
Because allometric relationships could not be
applied in this herbaceous species, we calculated
its fuel components by means of a multiple linear
regression. Outside the three experimental plots,
we measured percent cover and average height
(four points in each square) in ten 0.5 x 0.5 m
squares dominated by this species. Then, we
extracted, dried and weighed the fuel in the
laboratory. The resultant regression (Brachfuel =
4,28 Brachheight + 0,59 Brachcover - 18,69; r2=0,88;
p<0,001; n=30) was used to estimate the total fuel
in the Brachypodium patches of the experimental
plots. To calculate the fraction of dead and live
fuel, we used the proportion estimated for this
species in this region in summer periods, where
approximately 80% of its fuel is dead (Caturla
2002). Because dead and live fuels are mixed in
the same space for this species, we calculated
only the total FBD.
The proportion of biomass corresponding to
litter accumulated on the soil was insignificant
within the fuel-complex structure. Most squares
were bare soils with sparse litter particles, and
litter accumulated only in a few Rosmarinus and
Cistus patches. Although these species can

43
Figure 1. Fuel structural traits measured for shrubby species. FBD = fuel bulk density.
accumulate litter as a consequence of the annual
leaf fall, this litter layer was distributed
heterogeneously in small and isolated spots under
the canopy and did not reach thicknesses higher
than 2.5 cm in any case. Therefore, we assumed
that the influence of the litter layer on fire effects
was minimum, and it was not measured
quantitatively because of the difficulties entailed in
measuring it without using destructive methods
that alter the fuel structure system.
2.3 Species-patches fire effects
Variability in fire effects under the different species
patches was measured using two proxies. First,
we calculated the percentage of the total fuel
consumed for each square. We calculated the
difference between the pre-fire fuel load and the
fuel remaining after fire. For this, we clipped all the
unburned fuel remaining within each square; then
we took it to the laboratory where it was oven
dried at 80ºC for 24 h and weighed. Secondly, we
estimated the possible fire effects on ecosystem
functioning on the basis of the maximum
temperatures (Tmax hereafter) and duration of
heating over threshold temperatures (temperature-
residence-time) in the soil profile. In these
ecosystems dominated by obligate seeders, the
largest soil temperature increases due to fire have
been found in the first 1 cm of the profile (Baeza et
al. 2002a; De Luis et al. 2004) where, in addition,
the maximum density of seeds stored in the
seedbank has also been found (Baeza 2001;
Clemente et al. 2007). Thus, we measured the
temperature at the 1 cm depth under the different
vegetation patches as a possible proxy of the fire
effects on ecosystem functioning. The availability
of only 17 insulated chrome-alumel thermocouples
(K-type) per fire limited the equal distribution of
thermocouples among species. The sensors of the
thermocouples were placed in the centre of the
squares and distributed as follows: 5 for Cistus, 4
for Ulex, 4 for Rosmarinus and 4 for
Brachypodium. A narrow hole (3 cm wide, 2 cm
deep approximately) was excavated and the
sensors were inserted horizontally at the desired
depth. Then, the soil was restored as closely as
possible to the original state. If there was any litter
present, it was removed and restored after setting
up the sensors. After each fire, the buried sensors
were excavated and their exact depth was
checked. Thermocouples displaced from their
original depth during the fires were discarded in
the data analysis. A total of one thermocouple for
Brachypodium, three for Cistus, two for
Rosmarinus and one for Ulex were discarded in all
the experimental fires. The thermocouples were
protected from fire by a stainless steel sheath, and
they were connected to a datalogger (CR1000;
Campbell Scientific, North Logan, Utah, USA)
located outside the fire perimeter. The response
time of the thermocouples to temperature
fluctuations was 1 second, and the soil

44
temperature was registered every 10 seconds.
The register began 20 min before the
experimental fire was initiated and ended 2 hours
after the fire had been extinguished. The data
logger was also connected to a meteorological
station installed contiguous to the experimental
plot which registered wind speed, air temperature
and air relative humidity (Table 1).
2.4 Data analysis
We tested differences for each fuel structural trait
among the different species patches by means of
two-factor ANOVA. We used species-patches as a
fixed factor and site as random. Variables were
transformed to homogenise variances when
necessary. When significant differences were
found, Tukey’s HSD test for multiple pair-wise
comparisons of means was performed.
Differences in coarse dead and live fuel load, first-
live-branches height and fine live and fine dead
FBD could be tested only for shrubby species.
The assessment of which fuel component was
the most important one for determining Tmax and
temperature-residence-times was difficult because
the values of the fuel structural traits are
correlated with each other. However, performing a
separate correlation analysis of each fuel structure
component with the soil temperature parameters
could provide information about this. We assumed
that the most influential trait would be determined
by the strongest correlation coefficient with
temperature values. Correlation analyses were
performed separately for each species, and then,
for all shrubby species together. All statistical
analyses were performed using the SPSS v. 15
package (SPSS Inc., Chicago, IL).
3. Results
3.1 Fuel structural traits
Total fuel load within Brachypodium patches
(Mean ± SD; 216 ± 87 g m -2) was drastically lower
than in the shrub species patches, whilst there
were no significant differences found between
shrub species (3401 ± 1219 g m-2 for Ulex; 2926
± 1279 g m-2 for Rosmarinus; and 2122 ± 714 g m-
2 for Cistus; Table 3). However, the distribution of
fuel by size classes was markedly different among
shrubs. Fine dead fuel for Ulex represented 34%
of total fuel and was significantly higher than
Cistus and Rosmarinus (Figure 2A, Table 3),
which only reached 16% and 7% respectively. The
live fuel fraction was the most abundant one for all
Brachypodium Cistus Rosmarinus UlexF
uel (
g m
-2)
0
500
1000
1500
2000
Fine deadCoarse deadFine liveCoarse live
Brachypodium Cistus Rosmarinus Ulex
Hei
ght (
cm)
0
20
40
60
80
100
Total height First live branches
Brachypodium Cistus Rosmarinus Ulex
Bul
k de
nsity
(g
m-3
)
0
2000
4000
6000
8000
10000 Total Fine deadFine live
a
bbcc
a'
b' b'
c'
a
abb
c
a'a'
b'
A
B
C
Figure 2 . Fuel structural traits for the different species
patches. A) Fuel distribution by size classes, B) Total
and first-live-branches-height average, C) Bulk density
for total and fine fuel fractions. Error bars represent the
standard error. Different letters indicate significant (p
<0.05) differences between species patches (HSD
Tukey’s test).
shrubby species. Fine live fuel was the most
abundant fraction for Rosmarinus and it was
significantly higher than Cistus and Ulex (Figure
2A, Table 3). Coarse live fuel was similar for these
three species, representing 38-49% of the total
fuel load. Coarse dead fuel was scarce for all

45
Table 3. Results of the two-factor ANOVAs comparing fuel structural traits between the different species patches (p<0,05 *; p<0,01 **; p<0,001 ***). FBD = fuel bulk density. Fuel structure trait df F p Fuel structure trait df F p
Total fuel load Total height
Species patch 3 49,17 <0,001*** Species patch 3 122,91 <0,001***
Plot 2 0,228 0,803 Plot 2 1,29 0,342
Species patch*Plot 6 3,158 0,011* Species patch*Plot 6 2,22 0,057
Fine dead fuel load Live branches height
Species patch 3 9,21 0,012* Species patch 2 39,28 0,002**
Plot 2 1,45 0,306 Plot 2 0,68 0,558
Species patch*Plot 6 2,43 0,039* Species patch*Plot 4 1,13 0,359
Fine live fuel load Total FBD
Species patch 3 114,4 <0,001*** Species patch 3 3,76 0,079
Plot 2 0,11 0,898 Plot 2 0,15 0,856
Species patch*Plot 6 1,937 0,094 Species patch*Plot 6 4,77 0,001**
Coarse dead fuel load Fine dead FBD
Species patch 2 2,85 0,17 Species patch 2 6,25 0,059
Plot 2 0,09 0,913 Plot 2 1,99 0,251
Species patch*Plot 4 1,36 0,266 Species patch*Plot 4 2,15 0,094
Coarse live fuel load Fine live FBD
Species patch 2 1,5 0,327 Species patch 2 3,63 0,126
Plot 2 0,68 0,555 Plot 2 0,4 0,694
Species patch*Plot 4 2,53 0,057 Species patch*Plot 4 1,21 0,324
species and did not exceed 6% in any species
(Figure 2A, Table 3). There were also differences
among the heights of shrubs. Rosmarinus patches
were significantly higher than Ulex in total height,
whereas Cistus had intermediate values (Table 3,
Figure 2B). The average height of the first live
branches was significantly closer to the soil in
Rosmarinus patches than in Cistus and Ulex
patches (Table 3, Figure 2B). Differences close to
significance were found in total and fine dead fuel
FBD (Table 3). Total FBD and fine dead FBD were
slightly higher in Ulex patches than in the other
shrub patches (Figure 2C). In contrast, the fuel-
structure complex showed marked differences
between the Brachypodium herbaceous patches
and the shrub patches. Not only did the grass
patches have a lower total fuel load, but they also
contained only fine fuel, most of which was dead
(Figure 2A). Total height was also lower and did
not exceed 13 cm. However, because all its fuel
was concentrated at low heights above the soil
surface, its total FBD was higher than Ulex
patches and more than twice the values of Cistus
and Rosmarinus (Figure 2C).
3.2 Species-specific fire effects
Among shrubs, fuel consumption was highest in
Ulex patches (83.7%) (Figure 3A), whereas Cistus
and Rosmarinus consumption showed similar
values (70.1 and 68% respectively). Practically all
the fuel in Brachypodium patches was consumed
(Figure 3A). Maximum temperatures reached at
the 1 cm-depth were highly variable. They ranged
from 24ºC to 99ºC. The highest temperature
values were registered under Ulex patches,
though variabilty was high (Figure 3B), followed by
Brachypodium, Cistus and Rosmarinus patches
respectively (Figure 3B). Temperature-residence-
time above 40ºC was also variable, ranging
between 30.1 and 0.6 min in the different species
patches. Brachypodium residence-times were
approximately two times those of Ulex, whereas

46
Cistus and Rosmarinus experienced very short
times (Figure 3C). Temperatures above 50ºC were
of relatively short duration (less than 5 min) for all
species patches, but in this case longer durations
were experienced under Ulex patches.
Temperatures above 60ºC were found only under
Ulex patches and lasted no longer than 2 min
(Figure 3C).
3.3 Fuel structure traits that modulate soil
temperatures
Pearson correlation coefficients showed that Tmax
was better correlated with fuel structure traits than
BrachypodiumCistus Rosmarinus Ulex
Max
imum
tem
pera
ture
(ºC
)
20
30
40
50
60
70
80
90B
BrachypodiumCistus Rosmarinus Ulex
Fue
l con
sum
ptio
n (%
)
0
20
40
60
80
100 A
BrachypodiumCistus Rosmarinus Ulex
Res
iden
ce ti
me
(min
)
0
10
20
30
40 >40ºC >50ºC >60ºC
C
Figure 3. Fire effects in the different species-patches. A)
Consumed fuel; B) Maximum temperatures at 1 cm
depth. Boxes show the first and third quartiles and the
dividing line is the median; C) Soil temperature-
residence-times at 1cm depth. Error bars represent the
standard error.
with temperature-residence-times (Table 4). For
the whole set of shrub patches, fine dead fuel was
the most important fraction determining Tmax,
moreso than the rest of fractions and the total fuel
load. In addition, the spatial distribution of this
fraction was also important since the highest
correlation coefficient was found with fine dead
FBD. By species, Ulex patches showed the
strongest coefficients with Tmax, especially the
fine dead FBD (Table 4), whilst Cistus patches
were negatively correlated with the total FBD
(Table 4). The weakest coefficients were found in
Rosmarinus patches, which correlated negatively
with the coarse dead fuel load. For the grass
Brachypodium, Tmax was positively correlated
with the total fuel load.
Correlation analyses for temperature-
residence-times were performed only for
temperatures >40ºC, because patches showing
higher temperature thresholds were scarce. For
the collective set of shrub patches, the fine dead
fuel was also the most important fuel fraction in
determining residence-times. However, the spatial
distribution of the fuel fractions in the canopy was
also important in determining residence-times,
since the highest correlation coefficient was found
with total FBD. By species, Ulex had the
strongest correlation with total FBD (Table 4). No
significant correlations were found in Cistus and
Rosmarinus patches (Table 4). Patches of
Brachypodium had the strongest correlation with
the total height.
4. Discussion
The intrinsic differences in fuel structural traits
among species may entail variability in soil
temperatures in ecosystems dominated by
obligate seeders in Mediterranean Basin
shrublands. Subtle differences in the fuel-complex
structure could be crucial in modulating fire effects
at the micro-scale level, both for fuel consumption
and for soil temperatures. These results agree
with studies in other fire-prone vegetation types
where fire parameter variability is controlled by the
fuel array provided by the dominant species with

47
Table 4. Pearson r correlations between fuel structure traits and soil temperatures for each species-patch. The strongest correlations are marked in bold letters (p<0,05*; p<0,01 **; p<0,001 ***).
Fuel structure trait
Fuel load Height Fuel bulk density
Variable Total
Fine
live
Fine
dead
Coarse
live
Coarse
dead Total
Live
branch Total
Fine
live
Fine
dead
Tmax (1cm depth)
Brachypodium (n=11) 0,596* - - - - -0,253 - 0,497 - -
Cistus (n=12) -0,352 -0,479 -0,238 -0,203 -0,184 0,603* 0,552* -0,693** -0,608* -0,421
Rosmarinus (n=10) -0,198 -0,291 -0,226 -0,008 -0,578* 0,02 -0,121 -0,329 -0,46 0,045
Ulex (n=11) 0,652* -0,044 0,775** 0,696** 0,33 -0,485 -0,351 0,842** 0,18 0,871***
All shrubs (n=33) 0,416* -0,227 0,646*** 0,401* 0,107 -0,252 0,068 0,623*** 0,054 0,665***
Residence time >40ºC (min)
Brachypodium (n=11) -0,337 - - - - 0,711** - -0,516 - -
Cistus (n=12) 0,403 0,375 0,115 0,432 0,141 0,298 0,081 0,165 -0,02 0,077
Rosmarinus (n=10) -0,112 -0,272 0,179 -0,089 -0,152 0,297 0,433 -0,257 -0,332 -0,026
Ulex (n=11) 0,351 0,069 0,383 0,324 0,329 -0,379 -0,184 0,500* 0,269 0,425
All shrubs (n=33) 0,484** -0,074 0,652*** 0,400* 0,275 -0,325 0,014 0,709*** 0,267 0,629***
respect to soil temperatures (Bradstock et al.
1992; Savadogo et al. 2007; Wright and Clarke
2008), flame length (Bradstock and Gill 1993;
Morvan and Dupuy 2004), fire severity (De Luis et
al. 2005) and the rate of spread (van Wilgen et al.
1990; Fernandes 2001; Fernandes 2009).
The accumulated fine dead fuel load and its
spatial distribution in the plant architecture
together probably represent one of the most
important fuel structure traits determining fuel
consumption and soil temperatures in our
experimental fires. The low moisture content of
dead fuel increases both the risk of fire ignition
and the capacity of a fire to grow after ignition.
The dead fuel turns into the first heat source in the
initial steps of fire and its combustion contributes
to the loss of moisture in green fuel, which acts as
a heat sink until it, in turn, catches fire and
releases heat (Johnson 1992; Bond and van
Wilgen 1996; Sun et al. 2006). In Ulex patches,
where the proportion of accumulated fine dead
fuel was higher than in the other shrubby species,
the total amount of fuel consumed reached greater
values. The heat released by the consumption of
dead fuels in these patches could favour the
combustion of other fuel fractions such as fine and
coarse live fuels. In contrast, in Rosmarinus
patches, where the lowest accumulation of fine
dead fuel was registered, the heat released would
not be high enough to ignite green fuels, resulting
in a lower consumption of other fractions.
The spatial distribution of these fine dead fuels
could be a crucial factor in determining soil
temperatures. The highest temperature values
were observed under Ulex patches, and they were
also better correlated to the bulk density of fine
dead fuels. The considerable loads of fine dead
fuel in the lower strata lead to high densities in an
aerated state close to the soil; as a result, when
these fuel loads burn they can be consumed
easily, heating the soil profile. The burned fuel
could even collapse over the soil surface and
increase the heating efficiency. In this sense,
Schwilk (2003) demonstrated that both fuel
flammability and soil temperatures under
Adenostoma fasciculatum plants depended on the
architecture of the dead fuel accumulated. The
retention of dead branches in an aerated state in
the canopy close to the soil was likely to increase
flammability more than the same amount of
branches lying on the soil surface. In our study,
Rosmarinus patches registered the lowest soil
temperatures. This may be explained firstly, by its
lower fuel consumption and release of heat, and

48
secondly, by the low bulk density of its fuels,
which were more dispersed in the vertical canopy
structure and which, on consumption, could not
concentrate enough heat in the lower strata to
significantly heat up the soil profile. In fact, as has
been widely documented in other fire-prone
shrublands, under fuel beds with the same fuel
load, maximum soil temperatures and longer
residence-times were significantly higher for the
fuels with a lower height and a higher bulk density
(Bradstock et al. 1992; Molina and Llinares 2001;
Wright and Clarke 2008).
It has been demonstrated that the litter layer
can affect soil temperatures by buffering soil
heating in the case of low intensity fires with a
high rate of spread (Hartford and Frandsen 1992;
Valette et al. 1994). Despite the fact that litter
accumulation was low in our study, it could have
some effect in Rosmarinus and Cistus patches, as
these species accumulate litter under their
canopies due to an annual leaf release. This could
explain the negative correlation between their bulk
fuel densities and soil temperatures. High stem
densities could result in an increase in leaf release
and accumulation. Therefore, as our fires showed
high rates of spread and the fuel consumption of
these species was not severe, the litter layer could
have buffered soil heating. In fact, it has been
proposed that litter layers formed by obligate
seeders have low flammability; the high packing
ratio resulting from their small leaf sizes inhibits
the aeration of the fuel bed, thus hindering its
ignition and consumption (Scarff and Westoby
2006).
The Brachypodium herbaceous patches
behaved differently from the shrub patches. The
high proportion of fine dead fuel in these grass
patches was completely consumed by fire.
Surprisingly, we observed high soil temperature
values under these patches despite their low fuel
load. The high bulk densities of fine fuels in a few
centimetres near the soil surface were able to heat
the soil profile efficiently when they were burnt.
However, despite the fact that Brachypodium and
Ulex reached similar Tmax values, Brachypodium
temperature-residence-times above 50ºC or 60ºC
were lower in comparison. Because the low fine-
fuel load of this grass is consumed rapidly, the
heat released during the burn would not last long
enough to reach long residence-times. In contrast,
Ulex patches, composed of greater loads and
coarser fuels, would sustain a more prolonged
consumption. In fact, deeper penetrations and
longer temperature durations have been atributed
to the combustion of woody fuel collapsed on the
soil surface (Odion and Davis 2000).
At community level, the critical role of fine
dead fuels in determining the probability of fire
ocurrence through their high ignitability (Bond and
van Wilgen 1996), and in driving fire behaviour
through their high combustion rate (Rothermel
1972; Fernandes 2001; Anderson and Anderson
2009; Davis et al. 2009, 2010), has been widely
recognized and modelled in different fire-affected
shrublands. In fact, U. parviflorus-dominated
shrublands are regarded by fire managers as one
of the most problematic vegetation types on our
study site (eastern Iberian peninsula) because of
the high amounts and high bulk densities of their
fine dead fuel, which result in fuel beds of high
density and continuity (Baeza 2001; Baeza et al.
2002b; De Luis et al. 2004; De Luis et al. 2005;
Duguy et al. 2007). Species composition in
eastern Iberian peninsula ecosystems is
determined in part by the fire history; shifts in the
dominance of one species with respect to others
are driven by the effect of recurrent fires or
successional processes (Baeza et al. 2007).
Therefore, management strategies to drive
vegetation types within a desired successional
trajectory or a new preferable state dominated by
species with lower dead-fuel contents are highly
desirable. The promotion of mature and stable
stages of these shrublands dominated by R.
officinalis (Santana et al. 2010), could be a crucial
step to reduce fire-ignition risk, fire severity and
possible impacts on the ecosystem.
Acknowledgements

49
We thank J. Scheiding for her assistance with the
English revision of the text and the Fontroja
Natura-UA Scientific Station, the Conselleria de
Territori i Habitatge and the Consorcio de
Bomberos de Alicante and Valencia for their
fieldwork support. V.M. Santana was supported by
a FPU grant awarded by the Spanish Ministry of
Education and Science. We also thank to F.
Moreira for his suggestions in the early draft of this
study. This research was carried out within the
FIREMED (AGL200/8-04522/FOR) and
Consolider-Ingenio 2010 (GRACCIE CSD2007-
00067) projects. CEAM is supported by the
Generalitat Valenciana and Fundación Bancaja.
References
Alessio GA, Peñuelas J, De Lillis M, Llusià J
(2008) Implications of foliar terpene content
and hydration on leaf flammability of Quercus
ilex and Pinus halepensis. Plant Biology 10,
123-128.
Anderson SAJ, Anderson WR (2009) Predicting
the elevated dead fine fuel moisture content in
gorse (Ulex europaeus L.) shrub fuels.
Canadian Journal of Forest Research 39,
2355-2368.
Baeza MJ (2001) Aspectos ecológicos y técnicas
de control de combustible (roza y quema
controlada) en matorrales con alto riesgo de
incendio dominados por Ulex parviflorus
(Pourr.). Ph.D thesis, Departamento de
Ecología. Universidad de Alicante.
Baeza MJ, Raventós J, Escarré A (2002a) Ulex
parviflorus germination after experimental
burning: effects of temperature and soil depth.
In ‘Fire and Biological Processes’ (Eds L
Trabaud, R Prodon) pp. 83-91. (Backhuys
Publisher: Leiden, The Netherlands).
Baeza MJ, de Luis M, Raventós J, Escarré A
(2002b) Factors influencing fire behaviour in
shrublands of different stand ages and the
implications for using prescribed burning to
reduce wildfire risk. Journal of Environmental
Management 65, 199-208.
Baeza MJ, Raventos J, Escarré A, Vallejo VR
(2006) Fire risk and vegetation structural
dynamics in Mediterranean shrubland. Plant
Ecology 187, 189-201.
Baeza MJ, Valdecantos A, Alloza JA, Vallejo VR
(2007) Human disturbance and environmental
factors as drivers of long-term post-fire
regeneration patterns in Mediterranean
forests. Journal of Vegetation Science 18, 243-
252.
Beadle NCW (1940) Soil temperatures during
forest fires and their effect on the survival of
vegetation. The Journal of Ecology 28, 180-
192.
Bond WJ, van Wilgen BW (1996) Fire and plants.
(Chapman & Hall: London)
Bradstock RA, Auld TD, Ellis ME, Cohn JS (1992)
Soil temperatures during bushfires in semi-
arid, mallee shrublands. Australian Journal of
Ecology 17, 433-440.
Bradstock RA, Gill AM (1993) Fire in semi-arid,
Mallee shrublands: size of flames from
discrete fuel arrays and their role in the spread
of fire. International Journal of Wildland Fire 3,
3-12.
Bradstock RA, Auld TD (1995) Soil temperatures
during experimental bushfires in relation to fire
intensity: Consequence for legume
germination and fire management in south-
eastern Australia. Journal of Applied Ecology
32, 76-84.
Brooks ML (2002) Peak fire temperatures and
effects on annual plants in the Mojave desert.
Ecological Applications 12, 1088-1102.
Caturla RN (2002) Efecte del foc i la recurrència
d’incendis en camps abandonats dominats per
Brachypodium retusun (Pers.) Beauv. a la
Comunitat Valenciana. PhD thesis,

50
Departamento de Ecología. Universidad de
Alicante.
Certini G (2005) Effects of fire on properties of
forest soils: a review. Oecologia 143, 1-10.
Clemente AS, Rego FC, Correia O (2007) Seed
bank dynamics of two obligate seeders, Cistus
monspeliensis and Rosmarinus officinalis, in
relation to time since fire. Plant Ecology 190,
175-188.
Cohen M, Cuiñas P, Diez C, Fernandes P,
Guijarro M, Moro C (2003) Wildland fuel
particles characterisation: database content
(Deliverable D6-03-A1). Fire Star: a decision
support system for fuel management and fire
hazard reduction in Mediterranean wildland -
urban interfaces, Project no. EVG1-CT-2001-
00041, European Commission. Accessed
September 15, 2009 at
http://www.eufirestarpro.org
Conard SG, Hartzell T, Hilbruner MW, Zimmerman
GT (2001) Changing fuel management
strategies - The challenge of meeting new
information and analysis needs. International
Journal of Wildland Fire 10, 267-275.
Cornelissen JHC, Lavorel S, Garnier E, az S,
Buchmann N, Gurvich DE, Reich PB, Steege
Ht, Morgan HD, Heijden MGA, Pausas JG,
Poorter H (2003) A handbook of protocols for
standardised and easy measurement of plant
functional traits worldwide. Australian Journal
of Botany 51, 335-380.
Davies GM, Legg CJ, Smith AA, MacDonald AJ
(2009) Fire rate of spread in Calluna vulgaris-
dominated moorlands. Journal of Applied
Ecology 46, 1054-1063.
Davies GM, Smith AA, MacDonald AJ, Bakker D,
Legg CJ (2010) Fire intensity, fire severity and
ecosystem response in heathlands: factors
affecting the regeneration of Calluna vulgaris.
Journal of Applied Ecology 47, 356-365.
De Luis M, Baeza MJ, Raventós J, González-
Hidalgo JC (2004) Fuel characteristics and fire
behaviour in mature Mediterranean gorse
shrublands. International Journal of Wildland
Fire 13, 79-87.
De Luis M, Raventós J, González-Hidalgo JC
(2005) Factors controlling seedling
germination after fire in Mediterranean gorse
shrublands. Implications for fire prescription.
Journal of Environmental Management 76,
159-166.
Dimitrakopoulos AP (2001) A statistical
classification of Mediterranean species based
on their flammability components. International
Journal of Wildland Fire 10, 113-118.
Dimitrakopoulos AP, Panov PI (2001) Pyric
properties of some dominant Mediterranean
vegetation species. International Journal of
Wildland Fire 10, 23-27.
Duguy B, Alloza JA, Röder A, Vallejo R, Pastor F
(2007) Modelling the effects of landscape fuel
treatments on fire growth and behaviour in a
Mediterranean landscape (eastern Spain).
International Journal of Wildland Fire 16, 619-
632.
Elvira LM, Hernando C (1989) Inflamabilidad y
energía de las especies de sotobosque.
Colección Monografías INIA, nº 68. Madrid.
Fernandes PA (2001) Fire spread prediction in
shrub fuels in Portugal. Forest Ecology and
Management 144, 67-74.
Fernandes PA, Botelho H (2003) A review of
prescribed burning effectiveness in fire hazard
reduction. International Journal of Wildland
Fire 12, 117-128.
Fernandes P (2009) Combining forest structure
data and fuel modelling to classify fire hazard
in Portugal. Annals of Forest Science 66, 415
Hartford RA, Frandsen WH (1992) When it's hot,
it's hot... or maybe it's not! (surface flaming
may not portend extensive soil heating).
International Journal of Wildland Fire 2, 139-
144.

51
Johnson EA (1992) Fire and vegetation dynamics:
studies from the North American boreal forest.
(Cambridge University Press: Cambridge)
Keeley J.E. (2009) Fire intensity, fire severity and
burn severity: a brief review and suggested
usage. International Journal of Wildland Fire
18, 116-126.
Keeley JE, Fotheringham CJ, Baer-Keeley M
(2005) Determinants of post-fire recovery and
succession in Mediterranean-climate
shrublands of California. Ecological
Applications 15, 1515-1534.
Keeley JE, McGinnis TW (2007) Impact of
prescribed fire and other factors on cheatgrass
persistence in a Sierra Nevada ponderosa
pine forest. International Journal of Wildland
Fire 16, 96-106.
Molina MJ, Llinares JV (2001) Temperature-time
curves at the soil surface in maquis summer
fires. International Journal of Wildland Fire 10,
45-52.
Moreno JM, Oechel WC (1991) Fire intensity
effects on germination of shrubs and herbs in
southern California chaparral. Ecology 72,
1993-2004.
Morvan D, Dupuy JL (2004) Modeling the
propagation of a wildfire through a
Mediterranean shrub using a multiphase
formulation. Combustion and Flame 138, 199-
210.
Mouillot F, Field CB (2005) Fire history and the
global carbon budget: a 1º x 1º fire history
reconstruction for the 20th century. Global
Change Biology 11, 398-420.
Odion DC, Davis FW (2000) Fire, soil heating, and
the formation of vegetation patterns in
Chaparral. Ecological Monographs 70, 149-
169.
Paula S, Arianoutsou M, Kazanis D, Tavsanoglu,
Lloret F, Buhk C, Ojeda F, Luna B, Moreno
JM, Rodrigo A, Espelta JM, Palacio S,
Fernández-Santos B, Fernandes PM, Pausas
JG, Michener WK (2009) Fire-related traits for
plant species of the Mediterranean Basin.
Ecology 90, 1420.
Papió C, Trabaud L (1991) Comparative study of
the aerial structure of five shrubs of
Mediterranean shrublands. Forest Science 37,
146-159.
Pausas JG (2004) Changes in fire and climate in
the eastern Iberian peninsula (Mediterranean
Basin). Climatic Change 63, 337-350.
Pereira JM, Sequeira NM, Carreiras JM (1995)
Structural properties and dimensional relations
of some Mediterranean shrub fuels.
International Journal of Wildland Fire 5, 35-42.
Pérez B, Moreno JM (1998) Methods for
quantifying fire severity in shrubland-fires.
Plant Ecology 139, 91-101.
Philpot CW (1970) Influence of mineral content on
the pyrolysis of plant materials. Forest Science
16, 461-471.
Rothermel RC (1972) A mathematical model for
predicting fire-spread in wildland fuels. United
States Department of Agriculture, Forest
Service, Intermountain Forest and Range
Experiment Station. Research paper INT-115.
(Ogden, Utah)
Santana VM, Baeza MJ, Marrs RH, Vallejo VR
(2010). Old-field secondary succession in SE
Spain: can fire divert it? Plant Ecology 211,
337-349.
Savadogo P, Zida D, Sawadogo L, Tiveau D,
Tigabu M, Odén PC (2007) Fuel and fire
characteristics in savanna-woodland of West
Africa in relation to grazing and dominant
grass type. International Journal of Wildland
Fire 16, 531-539.
Saura-Mas S, Paula S, Pausas JG, Lloret F
(2010) Fuel loading and flammability in the
Mediterranean Basin woody species with
different post-fire regenerative strategies.

52
International Journal of Wildland Fire 19, 783-
794.
Scarff FR, Westoby M (2006) Leaf litter
flammability in some semi-arid Australian
woodlands. Functional Ecology 20, 745-752.
Schimmel J, Granström A (1996) Fire severity and
vegetation response in the boreal Swedish
forest. Ecology 77, 1436-1450.
Schwilk DW (2003) Flammability is a niche
construction trait: canopy architecture affects
fire intensity. The American Naturalist 162,
725-733.
Sun L, Zhou X, Mahalingam S, Weise DR (2006)
Comparison of burning characteristics of live
and dead chaparral fuels. Combustion and
flame 144, 349-359.
Troumbis AS, Trabaud L (1989) Some questions
about flammability in fire ecology. Acta
Oecologica-Oecologia Plantarum 10, 167-175.
Valette JC, Gomendy V, Maréchal J, Houssard C,
Gillon D (1994) Heat transfer in the soil during
very low-intensity experimental fires: the role
of duff and soil moisture content. International
Journal of Wildland Fire 4, 225-237.
van Wilgen BW, Higgins K, Bellstedt D (1990) The
role of vegetation structure and fuel chemistry
in excluding fire from forest patches in the fire-
prone fynbos shrublands of South Africa.
Journal of Ecology 78, 210-222.
Westerling AL, Hidalgo HG, Cayan DR, Swetnam
TW (2006) Warming and Earlier Spring
Increase Western U.S. Forest Wildfire Activity.
Science 313, 940-943.
Wright BR, Clarke PJ (2008) Relationships
between soil temperatures and properties of
fire in feathertop spinifex (Triodia schinzii
(Henard) Lazarides) sandridge desert in
central Australia. The Rangeland Journal 30,
317-325.

53
Apéndice fotográfico
Foto 1. Data logger para el registro de temperaturas Foto 2. Quema experimental de Onil
Foto 3. Quema experimental de Pardines Foto 4. Quema experimental de Ayora
Foto 5. Detalle de una quema experimental Foto 6. Parcela de estudio después de la quema
Foto 7. Parcela de estudio anterior a la quema Foto 8. Estructura del combustible en Ulex parviflorus

54

55
CAPÍTULO 4-
ESTABLECIMIENTO SUCESIONAL DE PLÁNTULAS EN
MATORRALES MEDITERRÁNEOS DOMINADOS POR
GERMINADORAS OBLIGADAS
RESUMEN: Las plántulas son particularmente sensibles a las condiciones ambientales en el momento de su establecimiento. Dentro de la sucesión, las condiciones ambientales varían en el tiempo y las especies tendrán una mejor posibilidad de regenerar en un punto concreto dependiendo de las características intrínsecas que determinan su nicho de regeneración. En un matorral mediterráneo, determinamos el nicho de regeneración y el patrón de establecimiento de plántulas a lo largo de la sucesión en las principales especies (Cistus albidus, Rosmarinus officinalis y Ulex parviflorus). El establecimiento de plántulas fue seguido durante tres años utilizando cohortes estacionales y en tres diferentes etapas a lo largo del gradiente sucesional (post-fuego, joven y maduro). Hubo un incremento del establecimiento inmediatamente después del fuego en todas las especies. Posteriormente, las especies experimentaron un declive en su establecimiento según la sucesión progresa, hasta que prácticamente no se encontró establecimiento en las etapas maduras. La gruesa capa de restos orgánicos y la probable competencia con individuos adultos impidió el establecimiento de plántulas. El establecimiento de Cistus estuvo muy ligado a ambientes post-fuego, mientras que Rosmarinus y Ulex también se establecieron en etapas jóvenes. En contraste a los estudios de otros matorrales mediterráneos, el establecimiento en nuestra área no está restringida solamente a etapas post-fuego, y las especies de matorral también se establecieron como oportunistas cuando hubo espacios abiertos a lo largo de la sucesión. Las diferencias en los micro-sitios preferidos para el establecimiento sugiere una diferenciación en los nichos de regeneración y unas condiciones ambientales particulares donde las diferentes especies son particularmente más competitivas a lo largo de la sucesión. Cistus estaría altamente ligada a ambientes perturbados, mientras Ulex y Rosmarinus podrían verse beneficiados por periodos entre fuegos más largos. Este capítulo reproduce el siguiente manuscrito: Santana VM, Baeza MJ, Maestre FT (enviado) Successional seedling establishment in Mediterranean shrublands dominated by obligate seeders.

56

57
Successional seedling establishment in Mediterranea n
shrublands dominated by obligate seeders
Victor M. SantanaA , M. Jaime BaezaA, B , Fernando T. Maestre C
AFundación de la Generalitat Valenciana Centro de Estudios Ambientales del
Mediterráneo (CEAM). Parque Tecnológico Paterna. C/Charles Darwin, 14.
46.980 Valencia. Spain. BDepartamento de Ecología, Universidad de Alicante. Ap. 99. 03080 Alicante.
Spain. CDepartamento de Biología y Geología, Área de Biodiversidad y Conservación,
Escuela Superior de Ciencias Experimentales y Tecnología, Universidad Rey
Juan Carlos, C/ Tulipán s/n, E-28933 Móstoles, Spain.
Abstract Seedlings are expected to be particularly sensitive to the environmental conditions at the time of establishment. Within succession, environmental conditions vary over time and a species will have a better chance to regenerate on a particular place depending on the intrinsic traits that determine its regeneration niche. In a Mediterranean shrubland, we determined the regeneration niche and the pattern of seedling establishment along succession of the main species (Cistus albidus, Rosmarinus officinalis and Ulex parviflorus). The establishment of species was monitored for three years using seasonal cohorts in three different stages along a successional gradient (post-fire, building, mature). There was a flush of establishment immediately after fire in all species. Then, the species experienced a declining establishment phase as succession progresses, until practically no seedling establishment was found at mature stages. Both the thick litter layer and the likely competition with adult plants precluded seedling establishment. The establishment of Cistus was very closely tied to the post-fire environment, while recruitment of Rosmarinus and Ulex also occurred in the building stage. In contrast to what has been reported in other Mediterranean shrublands, recruitment at our study area was not restricted solely to post-fire stages, and shrubs also recruited as opportunistic species when open gaps in the canopy were available along succession. The differences in preferred micro-sites for the establishment suggest a differentiation in regeneration niches and a particular set of environmental conditions where the different species would be particularly competitive through succession. Cistus would be tied to highly perturbed environments, whereas Ulex and Rosmarinus could be benefited in longer inter-fire periods.
Keywords: Competition; fire severity; open gaps; population dynamics; regeneration niche.

58
1. Introduction
In plant community dynamics, succession is
basically governed by adult decline and seedling
establishment (Harper, 1977). Seedlings are
expected to be particularly sensitive to the
environmental conditions at the time of
establishment, when they are most vulnerable
(Meyer, 1986; Kitajima and Fenner, 2000), and
their dynamics drive the potential replacement of
adults in future stages (Steven, 1991; Kellman and
Kading, 1992). Within successional gradients,
environmental conditions vary over time and,
consequently, a species will have a better chance
to regenerate on a particular place depending on
the intrinsic traits that determine its regeneration
niche (Grubb, 1977; Pickett and Bazazz, 1978;
Bazzaz, 1979; Fowler, 1988). Therefore,
approaches covering a broad range of
environmental conditions within the successional
gradient are basic for understanding both the
requirements for establishment of a species and
the role of seedling dynamics in driving population
patterns.
The occurrence of fires is a key factor driving
plant population patterns in Mediterranean
shrublands (Pierce and Cowling, 1991; Roy and
Sonié, 1992; Pausas, 1999; DeSimone and
Zedler, 1999; Keeley et al., 2006). In fact, the
persistence of obligate seeders relies completely
on seeds stored in the seed bank, as these
species are very sensitive and most individuals die
from the effects of fire (Pausas et al., 2004). In
these shrublands, the regeneration of species with
soil seed banks is almost completely tied to the
flush of seedling emergence and establishment
promoted by direct (e.g., heat, smoke, charred
wood) and indirect fire effects, which change the
environmental conditions and trigger the
availability of resources (e.g., increases in nutrient
levels, increases in light, decreases in
competition, shifts in daily soil temperature
regime) during the first stages after the
perturbation (Keeley, 1991; Bell et al., 1993;
Thanos and Rundel, 1995; DeBano et al., 1998;
Moreira et al., 2010; Santana et al., 2010a). These
fire effects break seed dormancy and enhance
seedling survival and, as a result, the regenerated
population is almost totally composed of even-
aged individuals (Keeley, 1992). Later
successional stages without fire provide
opportunities to regenerate the populations of
species that do not depend exclusively on fire to
become established; however, these regeneration
processes are very scarce and are mainly
attributed to bird-dispersed resprouting species
(Keeley, 1992; Keeley, 1995; Siles et al., 2008).
Probably for this reason, most studies on seedling
dynamics in Mediterranean shrublands have been
centred on early post-fire successional stages,
neglecting later stages (Moreno and Oechel,
1992; Quintana et al., 2004; De Luis et al., 2008).
Nevertheless, it has been suggested that the
establishment of seeder species in the
Mediterranean Basin may not be so closely tied to
fire events and that, in some cases, new
individuals could become established during inter-
fire periods (Clemente et al., 1996; Lloret, 1998;
Lloret et al. 2005). Therefore, accurate knowledge
on what are suitable micro-habitats for the
recruitment of these species and whether there is
a particular set of environmental conditions in
which the different species would be more
competitive during succession, remains to be
specifically assessed in Mediterranean
shrublands.
In the Mediterranean Basin, ecosystems
dominated by obligate seeders are mainly linked
to landscapes with long histories of perturbations
caused by human exploitation (old-fields and
livestock) and/or fire recurrence (Baeza et al.,
2007). These ecosystems stay in early
successional stages and are dominated by
pioneering species with persistent soil seed-banks
(Verdú, 2000; Pausas et al., 2004). In addition,
these species accumulate large amounts of
standing dead fine fuels on the plant structure,
making them one of the most problematic
vegetation types in terms of fire-risk (Duguy et al.,
2007; Saura-Mas et al., 2010). Species
composition in these ecosystems is highly

59
influenced by the fire regime; the fire-return
interval can, in fact, drive shifts in species
dominance (Pausas, 1999; Eugenio and Lloret,
2006; Baeza et al., 2007). Thus, the determination
of the ecological factors driving successional
seedling dynamics could be a key step in
designing new fuel-management strategies that
direct vegetation dynamics towards a desired
state or successional trajectory with lower
accumulations of dead fuel. Moreover, this issue
also has a special relevance in the light of climate
change, where shifts in fire regimes are predicted
(Pausas, 2004), and the presence of these
species could be enhanced (Verdú and Pausas,
2007).
The aim of this paper was to determine
seedling dynamics in Mediterranean Basin
shrublands dominated by obligate seeders at
different successional stages after fire. In addition,
we identified a series of biotic and abiotic factors
that could potentially drive the availability of
suitable micro-habitats for seedling regeneration.
Our initial hypothesis was that fire triggers the
availability of micro-habitats needed for seedling
establishment, and that this occurs mainly at the
immediate post-fire stage; at later successional
stages, micro-habitat availability decreases and
seedling establishment is restricted. To test this
hypothesis, we used shrublands with three
markedly different stages within the successional
gradient: an immediate post-fire stage, a building
stage where the community was composed of
individuals in a growth phase (10-12 years after
fire), and a mature stage where the community
was composed of individuals at a mature or
senescent phase (22-27 years after fire). Then,
during a three-year period, we monitored seedling
emergence and survival of the three dominant
obligate seeders (Cistus albidus L., Rosmarinus
officinalis L. and Ulex parviflorus Pourr.) at these
successional stages using seasonal cohorts.
2. Methods
2.1. Study area and site selection
The study was carried out in the interior of the
Valencia region (south-east Spain) on three sites:
Onil (38º39’N-0º39’W), Pardines (38º40’N-0º39’W)
and Ayora (39º07’N-0º57’W). The study sites were
in all cases old-field terraces that had been
abandoned ca. 50-60 years ago and had a well-
documented history of exploitation and fire
occurrence (Santana et al. 2010b). Their altitude
range between 900 and 1050 m.a.s.l. and their
climate is typically Mediterranean. Mean annual
rainfall ranges between 466 mm (Onil) to 537 mm
(Ayora). There is a pronounced summer drought
from June to August, with no more than 65 mm of
rain at any site. The mean annual temperature is
approximately 14ºC, and the mean maximum
temperature for the hottest month (July) is 30ºC.
To minimise environmental variability between
sites, all sites were oriented north, located on
marls and their soils were Regosols (FAO,
1988). At the onset of the study, the vegetation
consisted of shrublands (ca. 1-1.5 m in height)
dominated by nanophanerophytes at different
successional stages. The obligate seeders
Cistus, Rosmarinus and Ulex dominated the
vegetation, whereas resprouting shrubs such as
Quercus coccifera L. and Juniperus oxycedrus L.
were scarce. The grass Brachypodium retusum
(Pers.) Beauv. was the main herbaceous species.
Each site consisted of three sub-areas that
had been burned in different years and that we
assumed to be at different successional stages
after fire. In 1984, two different wildfires burned
the Onil and Pardines sites; then, in 1994, an
experimental fire was performed at each site over
part of the area burned before (see Baeza et al.,
2002 for details). The Ayora site was affected
by a wildfire in 1979, and, subsequently, part
of this area was burned again by another
wildfire in 1996. Finally, in June of 2006, an
experimental fire treatment was applied to
reburn part of the areas that had already
been burned twice. This fire history allowed us
to define three different stages within the
successional gradient at each site: first an

60
Axis 1 (48.8%)-30 -20 -10 0 10 20 30
Axi
s 2
(19.
9%)
-30
-20
-10
0
10
20
30
Post-fire
Building
Mature
Shrub and litter cover
Rosmarinus proximity C
istu
s an
d U
lex
prox
imity
Figure 1. Principal-coordinate analysis (PCO) of the
environmental conditions characterising the different
successional stages. Data represent means ± SE
(n=60).
immediately post-fire stage resulting from the
experimental burnings in 2006 (post-fire,
hereafter); second, a 10-12 year-old building stage
resulting from the 1994-1996 fires, in which the
vegetation consisted of individuals in the growth
phase (building, hereafter); and third, a 22-27
year-old mature stage resulting from the first fires
in 1979 and 1984, where the vegetation was
mainly composed of individuals in the mature or
senescent phase (mature, hereafter). On each
site, the three different successional stages were
located no farther than 500 m from each other.
We acknowledge that the recurrent fires at
short intervals experienced by the vegetation on
our study sites could have led to alterations in
seed bank composition and species abundance
during their post-fire and building stages (Santana
et al., 2010b). However, we accepted this
drawback because our aim was to determine
seedling dynamics with respect to three
contrasted environments within the succession,
and not to compare species abundance.
2.2. Characterisation of the successional stages
We studied the seedling dynamics of the three
dominant species (Cistus, Rosmarinus, and Ulex).
Together, these species had a relative cover
greater than 75% in the shrub layer at every
successional stage. To assess the factors
influencing seedling emergence at each
successional stage, we considered a series of
biotic and abiotic factors that characterise suitable
micro-habitats for this emergence. For doing this,
a 10 x 10 m plot was laid out within each
successional stage, and 20 points within this plot
were randomly chosen from a 1 x 1 m grid. At
each point, we set twenty 0.5 x 0.5 m quadrats for
monitoring seedling dynamics. In June 2006, we
visually estimated at each quadrat the percentage
cover for shrubs, herbs, stones (larger than 2 cm
in diameter) and litter (> 2 cm deep). Stones larger
than 2 cm in diameter and litter accumulations
with depth greater than 2 cm were considered
unsuitable microhabitats for seedling emergence
because the mean height of seedlings of our study
species was less than 2 cm (Lloret, 1998).
Proximity to the nearest adult plant of Cistus,
Rosmarinus and Ulex was also measured for each
quadrat as a proxy of seed availability. In the post-
fire stage, an additional index of fire severity,
based on Ryan and Noste (1985) was estimated.
This index was intended to infer soil temperatures
reached during the passage of fire, and it ranged
from 0 (low) to 5 (high). Concretely, the levels of
severity were estimated as follows: 0) unburned-
plant parts green and unaltered, no direct effect
for heat; 1) very low- more than 50% of plants and
litter remained unburned; 2) low- between 10 and
50% of plants and litter remained unburned, 3)
moderate- less than 10% remained unburned; 4)
high- only stems >5 mm in diameter remained,
black ash deposition; 5) very high- all plants and
litter practically consumed, white ash deposition.
Finally, changes in shrub composition during
succession were evaluated at each stage with
three perpendicular 20 m long transects spaced 7
m apart across the maximum slope. A metal rod
was used to record contacts with individual
species, and measurements were taken

61
Table 1. Micro-plot characteristics and correlation matrix with the first two axes of the principal coordinate analysis (PCO). Data represent means ± SE (n = 60). The highest correlation coefficients (ρ ≥ 0.6) and their P values are shown in bold type.
Micro-plot characteristics Successional stage Axis 1 Axis2
Post-fire Building Mature Spearman's ρ P Spearman's ρ P
Litter (%) 37.8 ± 3.2 46.3 ± 4.4 90.1 ± 2 -0.756 <0.001 -0.166 0.026
Stones (%) 12.7 ± 1.7 11.3± 1.5 3.7 ± 1.7 0.507 <0.001 0.178 0.017
Herbs (%) 1.3 ± 0.2 31.3± 3.1 13.2 ± 2.7 -0.237 0.001 0.328 <0.001
Shrubs (%) 10.4 ± 1.4 59.5± 4.1 76± 3.8 -0.772 <0.001 0.202 0.007 Cistus albidus proximity (cm) 32.8 ± 4.9 30.5± 2.9 144 ± 15.4 -0.406 <0.001 -0.622 <0.001 Rosmarinus officinalis proximity (cm) 177 ± 13.1 125.8 ± 12.2 36.5± 5.6 0.898 <0.001 -0.17 0.023 Ulex parviflorus proximity (cm) 54.3 ± 6.27 32.7± 3.5 104,3 ± 9.7 -0.319 <0.001 -0.665 <0.001
Severity (1-5) 2.5 ± 0.2 - - - - - -
every 20 cm along the transects (100 points per
transect).
2.3. Seedling monitoring
Seedling monitoring started in October 2006.
Seedling emergence and survival was monitored
in the quadrats previously described every two
months until June 2009. To facilitate their
identification during subsequent samplings,
seedlings were tagged with colour-coded rings
indicating the time of germination, and their
coordinates within the quadrat were recorded. The
fate of each seedling was tracked over the whole
study period. Germination of Mediterranean Basin
shrubs occurs preferentially in the wet seasons of
autumn and spring, whereas it is negligible in
summer (Lloret, 1998; Quintana et al., 2004; De
Luis et al., 2008). Therefore, and to simplify the
analyses of our data, we pooled the emergent
seedlings into 2 annual cohorts according to their
time of emergence. Seedlings emerging in the
October and December samplings were
considered autumn cohorts, whereas those
emerging in February, April and June were pooled
in the spring cohort of the respective years. The
last monitoring for the survival of previously
emerged seedlings was performed in September
2009 (after the last summer drought period).
2.4. Statistical analyses
To evaluate whether environmental conditions
(cover of shrubs, herbs, litter and stones, and
proximity to adult individuals) differed globally
between successional stages (post-fire, building
and mature), the semi-parametric multivariate
analysis of variance (PERMANOVA) developed by
Anderson (2001) was performed. For this analysis,
we used site as a random variable and
successional stage as a nested variable within
site. In addition, we conducted a principal
coordinates analysis (PCO) to identify the
particular environmental variables responsible for
the multivariate patterns observed. The first two
axes were correlated with the environmental
conditions measured using the Spearman’s
correlation coefficient. PERMANOVA and PCO
were performed using the programs
PERMANOVA 1.6 (Anderson 2005) and CAP
(Anderson 2004), respectively (both can be freely
downloaded from http://www.stat.auckland.ac.nz/
~mja/Programs.htm). For these analyses, we used
Bray-Curtis distance (appropriate for the dataset
containing a miscellaneous mixture of variables
and numerous zeros; Quinn and Keough, 2002)
and 4999 permutations (permutation of raw data;
Anderson and Ter Braak, 2003).
Differences in the number of emerged
seedlings between cohorts of the same
successional stage were checked with one-way
ANOVA. Post-hoc HSD Tukey tests were
performed when differences were observed

62
between the different cohorts. The data were log-
transformed when necessary to achieve a normal
distribution of the residuals and ensure
homoscedasticity.
Redundancy Analysis (RDA) was used to
explore the relationships between emerged
seedlings and micro-site characteristics for each
successional stage. We performed this analysis
with the autumn 2006 cohort, which was the most
representative of the flush of emergence in the
post-fire stage and the closest in time to the
sampling of micro-site characteristics.
Redundancy Analysis was performed using the
Vegan 1.9 package (Oksanen et al., 2007) in the
R software environment (version 2.6.1; R
Development Core Team, Vienna, Austria,
http://www.r-project.org/). The number of
emerging seedlings in each quadrat for each
species was used as dependent variables in the
ordination. These data were loge(x+1)
transformed. The forward selection procedure in
the Vegan package using the AIC statistic
(Oksanen et al., 2007) was used to select the
model with the most significant explanatory
variables determining seedling emergence.
Concretely, the explanatory variables used were:
shrub, herb, litter and stone cover, proximity to
adult individuals and fire severity (in the case of
the post-fire stage only). The significance of both
the model selected and the explanatory variables
axes was tested using a permutation test
(n=1000). We used site as a conditional variable
(covariable) in the analyses to remove its possible
effect.
Seedling survival at the different successional
stages for each species was analysed with the
Survival package (Crawley, 2007) in the R
software environment. We estimated the survival
curves of each cohort with the non-parametric
Kaplan-Meier analysis. We regarded as censored
data the seedlings alive at the end of the study
and those checked as alive at intermediate
samplings but not detected in later samplings.
Then, the shape differences in survival curves
between successional stages were tested by log-
rank tests (Pyke and Thompson, 1986). Since all
cohorts were not equally abundant, the statistical
analyses were carried out only when the density of
the respective cohort was at least 1 individual m-2
(Quintana et al., 2004). Therefore, for this
analysis, seedlings for the three sites in the same
census were pooled. The analysis could only be
performed in autumn cohorts, which were the
most abundant. However, even in some autumn
cohorts of Rosmarinus and Ulex, the analysis
could not be carried out due to their low individual
densities at some successional stages.
3. Results
3.1. Successional gradient after fire
Environmental conditions differed between
successional stages (PERMANOVA, d.f = 6, F =
21.49, P < 0.001). Moreover, there were
significant differences among sites
(PERMANOVA, d.f. = 2, F = 5.59, P < 0.001). The
first two PCO axes explained approximately 68%
of the variation observed in the data, and clearly
separated the three successional stages along
axis 1 (Figure 1). All the environmental conditions
measured were significantly correlated with the
first two axes, but we emphasize only those
variables with correlation coefficients (ρ) ±≥ 0.6.
Litter and shrub cover were the most important
variables changing environmental conditions along
the successional gradient and they were
negatively correlated with the first ordination axis
(Figure 1, Table 1). The proximity of Rosmarinus
to the seedling monitoring quadrats was positively
correlated with the first axis of the PCO, indicating
a greater proximity in mature stages (Figure 1,
Table 1). Those of Cistus and Ulex were
negatively correlated with axis 2, indicating a
slighter proximity in building stages (Figure 1,
Table 1). The post-fire stage experienced
intermediate values of severity according to our
index (Table 1). The shrub stratum was clearly
dominated by our three study species in all
successional stages, although its abundance was
variable. At the post-fire stage, vegetation cover
was low and it was dominated mainly by dead

63
individuals of Ulex and Cistus as a consequence
of fire (Table 2). At the building stage, Cistus and
Ulex were co-dominant, whereas Rosmarinus was
scarce. However, at mature stages Rosmarinus
became dominant and Cistus and Ulex
experienced a senescent phase, since most of
their individuals were found dead (Table 2).
Table 2. Shrub cover of stands at different successional stages. Data represent means ± SE (n = 3).
Successional stage
Shrub cover (%) Post-fire Building Mature
C. albidus 0.2 ± 0.2 30.1 ± 2.3 2.7 ± 1.9
R. officinalis 0.1 ± 0.1 2.2 ± 0.5 55.3 ± 4.7
U. parviflorus 0 21 ± 5.8 5.1± 1.1
C. albidus dead 7.3 ± 1.7 3.2 ± 1.9 0.2 ± 0.2
U. parviflorus dead 5.2 ± 0.9 2.7 ± 1.2 42 ± 4.3
Other shrubs 3.2 ± 1.3 20.8 ± 5.5 16.7 ± 4
3.2. Suitable micro-habitats for seedling
emergence
During the three years of the study, a total of 3664
seedling emergences were recorded. The most
abundant was Cistus (3003), followed by Ulex
(471) and Rosmarinus (190). Seedling emergence
for the three species was generally higher in
autumn than in spring cohorts (Table 3).
In the post-fire stage, the three species
showed a flush of germination in the first wet
season after fire (autumn 2006), with values
significantly higher than the subsequent cohorts
(approximately 112, 2 and 7 individuals·m-2 for
Cistus, Rosmarinus and Ulex, respectively; Table
3). After this, germination decreased in time until it
became practically inexistent three years after fire.
Cistus germination was approximately one order
of magnitude greater in number than the other
species in all the cohorts sampled (Table 3).
Seedling emergence was slightly correlated with
high-severity micro-habitats for Cistus and Ulex,
although the effect of this variable was not
significant within the model selected (Figure 2A).
In contrast, Rosmarinus was negatively affected
by fire severity, and was positively correlated with
the proximity of adult individuals of the same
species (Figure 2A).
In the building stage, the number of emerged
seedlings was closely similar among cohorts of
the same season. The most numerous cohorts
(autumn) were considerably fewer than the post-
fire flush of emergence in both Cistus (19-11
individuals·m-2) and Rosmarinus (1 individual·m-2;
Table 3). Ulex experienced a maximum of
emergence in the autumn 2008 cohort (10-11
individuals·m-2; Table 3), which was even higher
than the post-fire flush. In the other autumn
cohorts, emergence ranged between 3 and 4
individuals·m-2. Cistus and Ulex preferably emerg-
ed on open microsites since they were negatively
Table 3. Mean of emerged seedling cohorts (ind m-2; SE in brackets, n=3) throughout the three years of study. Means that are in the same row but are significantly different (one-way ANOVA, P<0.05, HSD Tukey test) are indicated with different letters.
Cohort Species and successional
stage Autumn 06 Spring 06 Autumn 07 Spring 08 Autumn 08 Spring 09 F P
Cistus albidus
Post-fire 112.2 ± 26.8 a 4.8 ± 3.5 bc 16.8 ± 3 ab 1.6 ± 0.4 cd 12.4 ± 3.1 b 0.3 ± 0.2 d 23.32 <0.001
Building 19.3 ± 10.2 a 1.6 ± 0.7 abc 10.7 ± 3.9 a 0.8 ± 0.4 bc 11.1 ± 4.9 ab 0.4 ± 0.1 c 7.49 0.002
Mature 2.1 ± 1.1 0.4 ± 0.2 2.8 ± 1.8 0.4 ± 0.3 2.1 ± 1.1 0.1 ± 0.1 2.36 0.147
Rosmarinus officinalis
Post-fire 1.9 ± 0.5 a 1.3 ± 0.3 ab 0.1 ± 0.1 b 0.2 ± 0.2 b 0.2 ± 0.2 b 0.1 ± 0.1 b 6.57 0.004
Building 1 ± 0.4 0.2 1.5 ± 0.9 0.5 ± 0.3 0.7 ± 0.6 0.2 ± 0.1 1.08 0.417
Mature 1.3 ± 0.3 0.3 ± 0.2 2.5 ± 2.1 0.3 ± 0.2 0.4 ± 0.2 0.3 ± 0.2 0.63 0.681
Ulex parviflorus
Post-fire 6.6 ± 4 a 0.8 ± 0.2 ab 1.1 ± 0.1 ab 0.3 ± 0.2 b 0.5 ± 0.2 b 0.1 ± 0.1 b 7.03 0.005
Building 3.5 ± 2.2 0.5 ± 0.3 3.5 ± 2.2 0.4 ± 0.3 10 .6 ± 5 0.6 ± 0.3 2.68 0.075
Mature 1.6 ± 0.3 a 0.13 ± 0.1 b 0.8 ± 0.3 ab 0.1 ± 0.1 b 0.5 ± 0.1 b 0.2 ± 0.1 b 7.56 0.002

64
influenced by shrub and herb cover (Figure 2B). In
contrast, Rosmarinus was not so linked to open
spaces, and preferred micro-sites near adult
plants (Figure 2B).
Germination in the mature stages was the
lowest for all successional stages and species, as
it varied between 1 and 3 individuals m-2 (Table 3).
Seedling emergence for all three species was
negatively correlated with litter cover >2 cm deep
(Figure 2C). The germination of Cistus was slightly
influenced by the proximity of adult individuals
(Figure 2C).
3.3. Seedling survival and establishment
In all three species, mortality was generally
highest shortly after germination, and clearly so
during the very early establishing phase (winter-
spring). After that, mortality was more sustained,
showing some peaks in summer periods (Figure
3).
The percentages of survival in Cistus were the
lowest of the three species studied, ranging from
5-15% in all successional stages at the end of our
study. However, there was one exception, the first
post-fire cohort (autumn 2006), which reached
26% survival (Table 4). The Log-Rank
comparisons of survival curves for the autumn
2006 cohort showed that survival in the post-fire
stage was significantly different from that in the
other successional stages (Table 4). However, this
difference disappeared in subsequent cohorts,
and only subtle differences were found between
the post-fire and the building stages in the autumn
2008 cohort (Table 4). Consequently, Cistus
establishment was practically confined to the initial
post-fire cohort of 29 individuals·m-2, since neither
in subsequent post-fire cohorts nor in other
successional stages did Cistus establishment
reach more than 1 individual·m-2.
Rosmarinus seedlings had percentages of
survival ranging from 33 to 61% in the post-fire
and building stages, respectively. In this species,
no significant differences in the survival curves
were found between these two stages (Figure 3,
Table 4). However, the survival curves in the
Figure 2 . Redundancy analysis (RDA) showing the relationships between emerging seedlings and micro-plot characteristics for the Autumn 2006 cohort. A) the model selected for the Post-fire stage (F= 0.136, P= 0.032) explained 29% of the total variance, axes 1 and 2 explained 5% and 2% and the conditional variable explained 22% of the variance; B) the model for the Building stage (F= 0.643, P< 0.005) explained 49% of the total variance, axes 1 and 2 explained 31% and 2% and the conditional variable explained 16%; C) Model for Mature stage (F= 0.403, P< 0.005) explained 22% of the total variance, axes 1 and 2 explained 21% and 1% and the conditional variable had no effect. Grey arrows are predictor variables, whilst emergent seedlings of the different species are shown in black. The numbers in brackets are P-values (permutation test with 1000 iterations).

65
Table 4. Number of emerged seedlings, percentage of survival and number of seedlings established at the end of the study for Autumn cohorts. The effect of the successional stage on the survivorship of seedlings of each species was compared by Log-Rank tests (P<0.05 are shown in bold).
Cohort Emerged Survival Established Log-rank
comparison
(individuals) (%) (individuals m-2) Post-fire Building
Cistus albidus
Autumn 2006
Post-fire 1665 26.3 29.2
Building 290 4.5 0.9 <0.001
Mature 32 6.2 0.1 0.008 0.395
Autumn 2007
Post-fire 252 11.3 1.9
Building 161 8.7 0.9 0.683
Mature 42 14.3 0.4 0.444 0.309
Autumn 2008
Post-fire 188 7.4 0.9
Building 166 4.8 0.5 0.004
Mature 32 15.6 0.3 0.742 0.221
Rosmarinus officinalis
Autumn 2006
Post-fire 28 35.7 0.7
Building 15 33.3 0.3 0.939
Mature 19 5.3 0.1 0.023 0.049
Autumn 2007
Post-fire 0 - -
Building 23 60.9 0.9 -
Mature 38 34.2 0.9 - 0.041
Autumn 2008
Post-fire 3 - -
Building 11 - - -
Mature 6 - - - -
Ulex parviflorus
Autumn 2006
Post-fire 99 36.4 2.4
Building 52 26.9 0.9 0.489
Mature 24 25 0.4 0.307 0.551
Autumn 2007
Post-fire 17 47.1 0.5
Building 52 34.6 1.2 0.706
Mature 12 - - - -
Autumn 2008
Post-fire 7 - -
Building 158 40.5 4.3 -
Mature 7 - - - -
mature stages were significantly different from
those in both the post-fire and the building stages
in all the cohorts studied (Figure 3, Table 4); in the
autumn 2006 cohort, mature-stage survival was
highly reduced to 5%, and in the autumn 2007
cohort it was reduced to 34% (note that for the
building stage it was 61%). The cohort from
autumn 2008 was very scarce, making it
impossible to perform any analysis.
The percentage of survival for Ulex seedlings
ranged from 25% to 47% for all cohorts and
successional stages. Environmental conditions at
the different successional stages did not affect
seedling survival in this species, as no differences

66
Autumn 2006
Autumn 2007
Season
Autumn 2006
Autumn 2007
Autumn 2008
Season
Autumn 2006
0,0
0,2
0,4
0,6
0,8
1,0
MatureBuildingPost-fire
Autumn 2007
See
dlin
g su
rviv
al
0,0
0,2
0,4
0,6
0,8
1,0
Autumn 2008
Season
0,0
0,2
0,4
0,6
0,8
1,0
Cistus Rosmarinus Ulex
AU
T06
SP
R07
AU
T07
SP
R08
AU
T08
SP
R09
Sep
09
AU
T06
SP
R07
AU
T07
SP
R08
AU
T08
SP
R09
Sep
09A
UT
06
SP
R07
AU
T07
SP
R08
AU
T08
SP
R09
Sep
09
Figure 3 . Kaplan-Meier survival curves for the cohorts with the most abundant seedlings. AUT = Autumn, SPR = Spring, and Sep = September. in survival curves were found between any
successional stage in any cohort (Figure 3, Table
4). There was a flush of establishment in the first
post-fire cohort (autumn 2006) of approximately 2
individuals·m-2, whilst establishment in the
subsequent cohorts was negligible. However, we
should highlight that, in the building stage,
notorious establishments between 1-4
individuals·m-2 per year were found. Establishment
in the mature stages was virtually nil.
4. Discussion
Obligate seeder shrubs experienced a declining
pattern of emergence and establishment as
succession progressed; this suggests a reduction
in the availability of suitable micro-habitats for their
regeneration through time. In the three species
studied there was a flush of germination in the
immediate post-fire cohort, which, in addition, had
the highest levels of survival and establishment.
The regeneration that occurs at this first stage
may be the main determinant of community
composition through time, until senescence
processes take place in adult individuals. This kind
of regeneration pattern may be in agreement with
the tolerance mechanism proposed by Connell
and Slatyer (1977). However, some shifts in
species composition with respect to the initial
regenerated community can occur: in our case,
establishment was not restricted to the immediate
post-fire stages and was found in some species
(i.e., Ulex and Rosmarinus) at least until the
building stages (10-12 years after fire). In fact,
seedling recruitment processes of seeder species
in inter-fire periods have previously been observed
in Mediterranean Basin shrublands; Lloret (1998)
found patterns of seedling establishment in a
similar shrubland in NE Spain 10 years after fire,
and Clemente et al. (1996) observed significant
recruitment of new individuals in fire-free periods

67
in Rosmarinus shrublands in Portugal.
Nevertheless, our results suggest that, over the
course of time, this recruitment will decrease with
the decline in micro-habitats suitable for seedling
establishment, until it practically ends in mature
stages (22-27 years after fire). In a similar
direction, Siles et al. (2008) found in shrublands in
S Spain that the time required to reach a steady
community dominated by Rosmarinus was at least
18 years. The ability of our study shrubs to recruit
in inter-fire periods agrees with results reported for
some suffrutescent species from the chaparral
(Keeley et al., 2006) and subshrubs in the
Californian coastal sage scrub (DeSimone and
Zedler, 1999), which are also able to establish
uneven-aged populations after fire with
recruitment linked to open gaps. These species
may be classified as ‘opportunistic’ species that
germinate and establish on open sites or disturbed
soil following fire and/or non-fire disturbance
(Ackerly, 2004). However, it is worth noting that
the recruitment patterns detected in our study
contrast with those described for most obligate
seeder shrubs in Mediterranean regions, where
germination and recruitment depend entirely on
fire cues and are found almost exclusively during
the first year post-fire (Pierce and Cowling, 1991;
Bell et al., 1993; Keeley, 1992, 1995).
The subtle differences detected in the
preferential micro-habitats for germination and
establishment of each species suggested a
differentiation in their regeneration niche (Grubb,
1977). These differences may imply that the
chance of establishment and coexistence of
species varies over time because of the changing
environmental conditions within succession
(Bazzaz, 1979). In this context, we propose a
conceptual scheme that shows how the
establishment patterns vary during succession for
each species and in relation to canopy
development (Figure 4). Immediately after fire, the
availability of resources and/or factors needed for
the regeneration of species reaches its optimum.
Direct fire-effects (i.e., soil temperatures and/or
smoke) can enhance germination in soil-stored
seeds (Bell et al., 1993; Keeley, 1995; Moreira et
al., 2010), and our study species probably
responded to these effects. However, the
responses varied between species. The
germination of species with hard-coated seeds
broken by heat, such as Cistus and Ulex (Roy and
Sonié, 1992; Baeza and Roy, 2008), was
positively correlated with high fire severity micro-
sites. Note that the severity of our fires was
generally intermediate, and the micro-sites with
higher-severity fires probably experienced
optimum temperatures for breaking the seed coat
without reaching deleterious temperatures.
Rosmarinus, in contrast, is a soft-seeded species
that shows some sensitivity to high temperatures
(Trabaud and Casal, 1989; Moreira et al., 2010)
and had a negative correlation with fire severity;
however, the fact that its germination is stimulated
by smoke (Moreira et al., 2010) could partly
explain its enhanced germination after fire. In
addition, other indirect fire-effects such as the
consumption of vegetation and the opening of
large gaps in the canopy can also promote
species regeneration. The incidence of solar
radiation can lead to higher daily fluctuations in
soil temperature and contribute to rendering hard-
seeded species permeable. Furthermore, the high
red to far-red ratio in the light spectrum reaching
the soil surface can also enhance germination. In
fact, these two germination cues have previously
been observed for Ulex, Cistus and other fire-
prone species (Roy and Arianoutsou-Faraggitaki,
1985; Roy and Sonié, 1992; Baeza and Roy,
2008; Santana et al., 2010a). In post-fire
environments, seedling survival is expected to be
enhanced both by increasing the availability of soil
nutrients through ash deposition and by increasing
soil water availability due to a lack of competition
(Thanos and Rundel, 1995; DeBano et al., 1998).
Although these soil resources were not directly
measured, we can infer that this effect took place
only in the first post-fire cohort of Cistus. The other
species did not need these resources to
experience higher survival rates in the post-fire
stage than in the later successional stages.

68
Figure 4 . Conceptual scheme showing A) canopy development along succession for the three main species, and B) patterns of seedling establishment.
In the subsequent building stage, the
availability of suitable micro-habitats for species
regeneration declines as the direct and indirect
fire-effects decrease through time. Open gaps in
the canopy left among the building individuals, and
the absence of a thick litter layer, are the main
requirements for achieving germination in hard-
coated seeds (i.e., Cistus and Ulex). The seeds
that remain ungerminated in the soil after fire or
those produced by the new individuals can be
stimulated by the high levels of light and radiation
affecting the soil in open gaps. However, the
existence of seedling emergence did not translate
into establishment for all species; emerging
seedlings of Cistus had high mortality rates and
there was practically no Cistus establishment in
the building stages. This suggests that the
regeneration of Cistus could be tied to
environments immediately post-disturbance,
where the high availability of soil resources is the
only way to counterbalance its high mortality rate.
In fact, this high mortality rate has also been found
for C. salvifolius and C. villosus (ca. 90%) in
unburned stands of the Greek maquis (Troumbis
and Trabaud, 1987). Ulex was the only species
with a significant recruitment in open gaps in our
study. In contrast, the germination of Rosmarinus
differed from that of the hard-seeded species in
that it was more related to the proximity of adult
individuals than to canopy gaps. This may have
several explanations. For example, the fact that
the abundance of Rosmarinus individuals was
scarce (low seed availability) could mean that this
species has a low dispersion ability. It is also
possible that Rosmarinus germination and
establishment are not so linked to open gaps as to
safer sites next to parent plants (Ellner and
Shmida, 1981; Escudero et al., 1999). Rosmarinus
had a low number of established individuals in the
building stages, but this was probably due to its
low number of emergent seedlings since it showed
high survival rates. Our findings contrast with
Lloret (1998), who did not find any relationship
between seedling germination and open gaps; this
could be the result of the analyses conducted by
this author, who analysed all the species together.
As the succession progresses towards mature
stages (22-27 year after fire), the established
individuals grow and the availability of resources
needed for germination, such as light and space,
declines. The annual leaf release and the
individuals of short-lived species in senescent
phases can increase the cover and thickness of
the litter layer, impeding seedling emergence. In

69
fact, litter depth can influence recruitment by
decreasing the surface suitable for germination
(Facelli and Pickett, 1991; Lloret, 1998; Baeza and
Roy, 2008). In addition, a high density of adults
may have a negative effect on seedling
establishment of these pioneering species
because of competition for light and soil resources
(Aguiar et al., 1992; Veenendaal et al., 1995). This
could explain the high mortality rates experienced
in Rosmarinus seedlings at mature stages in
comparison with previous stages. Thus, the
combination of all these factors can eliminate the
availability of suitable micro-habitats for
regeneration in our study species at mature
stages.
Traditionally, obligate seeder species in the
Mediterranean basin have been globally regarded
as pioneer species of early successional stages
due to their high production of small seeds, their
high dispersion ability and their high growth rates
(Verdú, 2000; Pausas et al., 2004). Nevertheless,
our findings suggest that the differences in the
regeneration niches of these species may point to
the existence of tradeoffs that make them more
competitive in a particular set of environmental
conditions during succession (Parrish and Bazzaz,
1982; Silvertown, 2004). Some species generate a
large number of seeds whose establishment is
very dependent on post-fire conditions (i. e., heat,
open gaps, high soil resource levels). This seems
to be the case of Cistus and other short-lived
subshrubs that generate a large offspring after fire
and become dominant in recurrently perturbed
environments (Roy and Sonié, 1992; Pausas,
1999; Baeza et al., 2007; Santana et al., 2010b).
In contrast, species like Ulex, whose recruitment is
not so linked to fire, may take advantage of
building stages to colonise open gaps and expand
their abundance through time. Therefore, although
the response of a species to post-fire
environmental conditions (i.e., sensitivity to fire
severity, dispersion and distribution of the
seedbank, survival ability) would be the main
factor determining species establishment and
possible spatial coexistence (Moreno and Oechel,
1992; Quintana et al., 2004; De Luis et al., 2008),
the ability of a species to establish in inter-fire
periods may also be an important factor driving
seeder-species coexistence through the
successional process in the western
Mediterranean Basin.
Acknowledgements
We thank J. Scheiding for the revision of the
English text and the Font Roja Natura-UA
Scientific Station for fieldwork support. V.M.
Santana was supported by a FPU grant awarded
by the Spanish Ministry of Education and Science.
F. T. Maestre is supported by the European
Research Council under the European
Community's Seventh Framework Programme
(FP7/2007-2013)/ERC Grant agreement n°
242658 (BIOCOM). This research was carried out
within the FIREMED (AGL200/8-04522/FOR) and
Consolider-Ingenio 2010 (GRACCIE CSD2007-
00067) projects. CEAM is supported by the
Generalitat Valenciana and Fundación Bancaja.
References
Ackerly, D. 2004. Functional strategies of
chaparral shrubs in relation to seasonal water
deficit and disturbance. Ecological
Monographs 74, 25-44.
Aguiar, M.R., Soriano, A., Sala, O.E. 1992.
Competiton and facilitation in the recruitment
of seedlings in Patagonian steppe. Functional
Ecology 6, 66-70.
Anderson, M.J. 2001. A new method for non-
parametric multivariate analysis of variance.
Austral Ecology 26, 32-46.
Anderson, M.J. 2004. CAP: a FORTRAN
computer program for canonical analysis of
principal coordinates. Department of Statistics,
University of Auckland.
Anderson, M.J. 2005. PERMANOVA: a FORTRAN
computer program for permutational analysis

70
of variance. Department of Statistics,
University of Auckland.
Anderson, M.J., Ter Braak, C.J.F. 2003.
Permutation tests for multi-factorial analysis of
variance. Journal of Statistical Computation
and Simulation 73, 85-113.
Baeza, M.J., De Luis, M., Raventós, J., Escarré,
A. 2002. Factors influencing fire behaviour in
shrublands of different stand ages and the
implications for using prescribed burning to
reduce wildfire risk. Journal of Environmental
Management 65, 199-208.
Baeza, M.J., Valdecantos, A., Alloza, J.A., Vallejo,
V.R. 2007. Human disturbance and
environmental factors as drivers of long-term
post-fire regeneration patterns in
Mediterranean forests. Journal of Vegetation
Science 18, 243-252.
Baeza, M.J., Roy, J. 2008. Germination of an
obligate seeder (Ulex parviflorus) and
consequences for wildfire management. Forest
Ecology and Management 256, 685-693.
Bazzaz, F.A. 1979. The physiological ecology of
plant succession. Annual Review of Ecology
and Systematics 10, 351-371.
Bell, D.T., Plummer, J.A. & Taylor, S. K. 1993.
Seed germination ecology in southwestern
western Australia. Botanical Review 59, 24-73.
Clemente, A.S., Rego, F.C., Correia, O. 1996.
Demographic patterns and productivity of post-
fire regeneration in Portuguese Mediterranean
maquis. International Journal of Wildland Fire
6, 5-12.
Crawley, M.J. 2007. The R book. John Wiley &
Sons, Ltd, Chichester.
Connell, J.H., Slatyer, R.O. 1977. Mechanisms of
succession in natural communities, and their
role in the community stability and
organization. The American Naturalist 111,
1119-1144.
De Luis, M., Raventos, J., Wiegand, T., González-
Hidalgo, J.C. 2008. Temporal and spatial
differentiation in seedling emergence may
promote species coexistence in Mediterranean
fire-prone ecosystems. Ecography 31, 620-
629.
DeBano, L.F., Neary, D.G., Ffolliott, P.F. 1998.
Fire’s effects on Ecosystems. John Wiley &
Sons, New York.
DeSimone, S.A., Zedler, P.H. 1999. Shrub
seedling recruitment in unburned Californian
coastal sage scrub and adjacent grassland.
Ecology 80, 2018-2032.
Duguy, B., Alloza, J.A., Röder, A., Vallejo, R.,
Pastor, F. 2007. Modelling the effects of
landscape fuel treatments on fire growth and
behaviour in a Mediterranean landscape
(eastern Spain). International Journal of
Wildland Fire 16, 619-632.
Ellner, S., Shmida, A. 1981. Why are adaptations
for long-range seed dispersal rare in desert
plants? Oecologia 51, 133-144.
Escudero, A., Somolinos, R.C., Olano, J.M.,
Rubio, A. 1999. Factors controlling the
establishment of Helianthemum squamatum,
an endemic gypsophile of semi-arid Spain.
Journal of Ecology 87, 290-302.
Eugenio, M., Lloret, F. 2006. Effects of repeated
burning on Mediterranean communities of the
northeastern Iberian Peninsula. Journal of
Vegetation Science 17, 755-764.
Facelli, J.M., Pickett, S.T.A. 1991. Plant litter: its
dynamics and effects on plant community
structure. The Botanical Review 57, 1-32.
FAO 1988. Soil map of the world. Revised legend.
World soil resources report, 60. 99 pp. Rome.
Fowler, N.L. 1988. What is a safe site?:
neighbour, litter, germination date, and patch
effects. Ecology 69, 947-961.
Grubb, P.J. 1977. The maintenance of species-
richness in plant communities: the importance

71
of the regeneration niche. Biological Reviews
52, 107-145.
Harper, J.L. 1977. Population biology of plants.
Academic press, London.
Keeley, J.E. 1991. Seed germination and life
history syndromes in the California chaparral.
The Botanical Review 57, 81-116.
Keeley J.E. 1992. Recruitment of seedlings and
vegetative sprouts in unburned chaparral.
Ecology 73, 1194-1208.
Keeley J.E. 1995. Seed-germination patterns in
fire-prone Mediterranean-climate regions. In:
Arroyo, M.T.K., Zedler, P.H., Fox, M.D. (Eds.)
Ecology and biogeography of Mediterranean
ecosystems in Chile, California and Australia.
Springer-Verlag, New York. pp. 239-273.
Keeley, J.E., Fotheringham, C. J., Baer-Keeley, M.
2006. Demographic patterns of postfire
regeneration in Mediterranean-climate
shrublands of California. Ecological
Monographs 76, 235-255.
Kellman, M., Kading, M. 1992. Facilitation of tree
seedling establishment in a sand dune
succession. Journal of Vegetation Science 3,
679-688.
Kitajima, K., Fenner, M. 2000. Ecology of seedling
regeneration. In: Fenner, F. (Ed.) Seeds: the
ecology of regeneration in plant communities.
CAB international, Wallingford. pp. 331-359.
Lloret, F. 1998. Fire, canopy cover and seedling
dynamics in Mediterranean shrubland of
northeastern Spain. Journal of Vegetation
Science 9, 417-430.
Lloret, F., Peñuelas, J., Estiarte, M. 2005. Effects
of vegetation canopy and climate on seedling
establishment in Mediterranean shrubland.
Journal of Vegetation Science 16, 67-76.
Meyer, S.E. 1986. The ecology of gypsophile
endemism in the eastern Mojave desert.
Ecology 67, 1303-1313.
Moreira, B., Tormo, J., Estrelles, E., Pausas, J. G.
2010. Disentangling the role of heat and
smoke as germination cues in Mediterranean
Basin flora. Annals of Botany 105, 627-635.
Moreno, J.M., Oechel, W.C. 1992. Factors
controlling postfire seedling establishment in
southern California chaparral. Oecologia 90,
50-60.
Oksanen, J., Kindt, R., Legendre, P., O’Hara, B.,
Henry, M., Stevens, H. 2007. Vegan:
Community Ecology Package. R package
version 1.8-7. http://cran.r-project.org,
http://cc.oulu.fi/fi/~jarioksa/. Accessed 12
September 2007.
Parrish, J.A.D., Bazzazz, F.A. 1982. Responses of
plants from three successional communities to
a nutrient gradient. Journal of Ecology 70, 233-
248.
Pausas, J.G. 1999. Response of plant functional
types to changes in the fire regime in
Mediterranean ecosystems: A simulation
approach. Journal of Vegetation Science 10,
717-722.
Pausas, J.G. 2004. Changes in fire and climate in
the eastern Iberian peninsula (Mediterranean
Basin). Climatic Change 63, 350.
Pausas, J.G., Bradstock, R.A., Keith, D.A.,
Keeley, J.E., GCTE. 2004. Plant functional
traits in relation to fire in crown-fire
ecosystems. Ecology 85, 1085-1100
Pickett, S.T.A., Bazzaz, F.A. 1978. Organization of
an assemblage of early successional species
on a soil moisture gradient. Ecology 59, 1248-
1255.
Pierce, S.M., Cowling, R.M. 1991. Dynamics of
soil-stored seed banks of six shrubs in fire-
prone dune fynbos. Journal of Ecology 79,
731-747.
Pyke, D.A., Thompson, J.N. 1986. Statistical
analysis of survival and removal rate
experiments. Ecology 67, 240-245.

72
Quinn , G.P., Keough, M.J. 2002. Experimental
design and data analysis for biologists.
Cambridge University Press, Cambridge.
Quintana, J.R., Cruz, A., Fernandez-Gonzalez, F.,
Moreno, J.M. 2004. Time of germination and
establishment success after fire of three
obligate seeders in a Mediterranean shrubland
of central Spain. Journal of Biogeography 31,
241-249.
Roy, J., Arianoutsou-Faraggitaki, M. 1985. Light
quality as the environment trigger for the
germination of the fire-promoted species
Sarcopterium spinosum L. Flora 177, 345-349.
Roy, J., Sonié, L. 1992. Germination and
population dynamics of Cistus species in
relation to fire. Journal of Applied Ecology 29,
647-655.
Ryan, K.C., Noste, N.V. 1985. Evaluating
prescribed fires. In: Lotan, J.E., Kilgore, B.M.,
Fischer, W.C., Mutch, R.W. (Eds.)
“Proceedings, symposium and workshop on
wilderness fire”15-18 November 1983,
Missoula, MT. USDA Forest Service,
Intermountain forest range experimental
station, General technical report INT-182.
Missoula. pp. 230-238.
Santana, V.M., Bradstock, R.A., Ooi, M.K.J.,
Denham, A.J., Auld, T.D., Baeza, M.J. 2010a.
Effects of soil temperature regimes after fire on
seed dormancy and germination in six
Australian Fabaceae species. Australian
Journal of Botany 58, 539-545.
Santana, V.M., Baeza, M.J., Marrs, R.H., Vallejo,
V.R. 2010b. Old-field secondary succession in
SE Spain: can fire divert it? Plant Ecology 211,
337-349.
Santana, V.M., Baeza, M.J., Vallejo, V.R. in press.
Fuel structural traits modulating soil
temperatures in different species-patches of
Mediterranean Basis shrublands. International
Journal of Wildland Fire
Saura-Mas, S., Paula, S., Pausas, J.G., Lloret, F.
2010. Fuel loading and flammability in the
Mediterranean Basin woody species with
different post-fire regenerative strategies.
International Journal of Wildland Fire 19, 783-
794.
Siles, G., Rey, P.J., Alcántara, J.M., Ramírez, J.M.
2008. Assessing the long-term contribution of
nurse plants to restoration of Mediterranean
forests through Markovian models. Journal of
Applied Ecology 45, 1790-1798.
Silvertown, J. 2004. Plant coexistence and the
niche. Trends in Ecology and Evolution 19,
605-611.
Steven, D.D. 1991. Experiments on mechanisms
of tree establishment in old-field succession:
seedling survival and growth. Ecology 72,
1076-1088.
Thanos, C.A., Rundel, P.W. 1995. Fire-followers in
chaparral: nitrogenous compounds trigger
seed germination. Journal of Ecology 83, 207-
216.
Trabaud, L., Casal, M. 1989. Réponses des
semences de Rosmarinus officinalis à
différents traitments simulant une action de
feu. Acta Oecologica 10, 355-363.
Troumbis, A., Trabaud, L. 1987. Dynamique de la
banque de graines de deux espèces de Cistes
dans les maquis grecs. Acta Oecologica 8,
167-179.
Veenendaal, E. M., Swaine, M. D., Agyeman, V.
K., Blay, D., Abebrese, I. K., Mullins, C. E.
1995. Differences in plant and soil water
relations in and around a forest gap in west
Africa during the wry season may influence
seedling establishment and survival. Journal of
Ecology 84, 83-90.
Verdú, M. 2000. Ecological and evolutionary
differences between Mediterranean seeders
and resprouters. Journal of Vegetation
Science 11, 265-268.

73
Verdú, M., Pausas, J.G. 2007. Fire drives
phylogenetic clustering in Mediterranean Basin
woody plant communities. Journal of Ecology
95, 1316-1323

74
Apéndice fotográfico
Foto 1. Cuadro de muestreo Foto 2. Plántulas de Cistus albidus
Foto 3. Plántulas de Rosmarinus officinalis Foto 4. Plántula de Ulex parviflorus
Foto 5. Etapa post-incendio Foto 6. Matorral joven
Foto 7. Matorral en una etapa sucesional madura

75
CAPÍTULO 5- LA RECURRENCIA DE INCENDIOS Y EL TIEMPO DESDE EL INCENDIO COMO CONDUCTORES DE LA INFLAMABILIDAD EN MATORRALES MEDITERRÁNEOS RESUMEN: Diferencias en la inflamabilidad de las especies y su habilidad regenerativa post-fuego pueden ser un factor clave en el estableciendo vínculos entre el régimen de incendios y la dinámica de la vegetación. Nosotros formulamos la hipótesis de que las especies leñosas que acumulan una mayor cantidad de de combustible muerto y tienen un reclutamiento estimulado por el fuego se verían beneficiadas por un incremento de la recurrencia de incendios y, por lo tanto, establecerían un bucle de de retroalimentación positiva con el fuego. Para ello, comparamos dos series de matorrales quemados una y dos veces. Además, valoramos el cambio en la estructura del ecosistema durante la sucesión post-fuego (25 años) determinando el cambio en formas vitales, especies, combustible muerto y presencia de herbáceas. Encontramos que, a nivel de comunidad, las especies de matorral de las etapas iniciales de la sucesión son las más inflamables como consecuencia de su mayor crecimiento y acumulación de combustible muerto. Como consecuencia, la función de riesgo de incendio a lo largo del tiempo basada en el combustible leñoso tiene forma jorobada, debido a la sustitución sucesional por especies menos inflamables. Sin embargo, la acumulación de combustible muerto no se encuentra bajo una selección positiva a causa de fuegos recurrentes, ya que un segundo fuego en el periodo de máximo riesgo de incendio promovió una comunidad con menor cantidad de combustible muerto. Este hecho sugiere la falta de una retroalimentación positiva fuego-combustible muerto acumulado. En contra, bajo el régimen de incendio estudiado, el sistema se ve desplazado hacia comunidades de bajo porte y herbáceas. Este capítulo reproduce el siguiente manuscrito: Santana VM, Baeza MJ, Marrs RH (en preparación) Fire recurrence and time-since-fire as flammability drivers in Mediterranean Basin shrublands.

76

77
Fire recurrence and time-since-fire as flammability drivers in
Mediterranean Basin shrublands
Victor M. SantanaA, M. Jaime BaezaA,B, Rob H. MarrsC
AFundación de la Generalitat Valenciana Centro de Estudios Ambientales del
Mediterráneo (CEAM). Parque Tecnológico Paterna. C/Charles Darwin, 14.
46.980 Valencia. Spain
BDepartamento de Ecología, Universidad de Alicante. Ap. 99. 03080 Alicante.
Spain
CApplied Vegetation Dynamics Laboratory, School of Environmental Sciences,
University of Liverpool, Liverpool, L69 3GP, UK.
Abstract Differences in both the flammability of species and their post-fire regenerative abilities can be the key for establishing a mutual link between fire regime and vegetation dynamics. We hypothesised that woody species that accumulate the greatest amount of dead fuel and have fire-stimulated recruitment would be benefited by increasing fire recurrence and, thus, establish a positive fire-vegetation’s flammability feedback. For this, we compared two seres of shrublands burned once and twice. In addition, we assessed the change in ecosystem structure during the post-fire succession (25 years) by assessing the change in life-forms, species, dead fuel load and the presence of herbaceous species. We found that, at community level, early successional shrubs are the most flammable species as consequence of their higher growth and accumulation of dead fuel. As consequence, the fire-risk function through time of woody combustible is hump-shaped due to the successional replacement with less flammable species. However, the accumulation of dead fuel is not under positive selection by recurrent fires, since a second fire in the period of maximum fire risk promote a community with less amount of dead fuel. This fact suggest the lack of positive fire-dead fuel accumulation feedback. In contrast, under the studied fire regime, the systems are displaced to grass-scrublands. Keywords: Dead fuel, feedback, HOF models, succession, vegetation structure

78
1. Introduction
Differences in both the flammability of species and
their post-fire regenerative abilities can be the key
for establishing a mutual link between fire regime
and vegetation dynamics; for example, a positive
fire-feedback may occur if the more flammable
species also show more successful regenerative
trends after fire (Wilson and Agnew 1992). In this
sense, traits that enhances species flammability,
for example the accumulation of dead fuel within
the vegetation structure, have been proposed as
niche construction traits in fire-prone
environments; i.e, they modify the environment,
either increasing fire recurrence and/or fire
severity, and provide fire-cleared gaps that benefit
the more flammable species (Odling-Smee et al.
1996; Schwilk 2003). Moreover, it have been
suggested that traits enhancing the flammability of
plants may be subjected to positive selective
pressures in fire-prone environments (Mutch 1970;
Bond and Midgley 1995; Kerr et al. 1999; Gagnon
et al. 2010) and they can exhibit an evolutionary
correlation with fire-dependent seedling
recruitment (Schwilk and Ackerly 2001). This may
be the case for pyrogenic species where seed
germination is stimulated by fire cues, or where
seed is released from serotinous cones after heat
exposure (Schwilk and Ackerly 2001; Schwilk
2003). Resprouting species, such as herbs and
grasses, can also expand under recurrent fire
regimes as consequence of their high flammability
(e.g. fine texture and very low moisture in
summer) and high regeneration ability (Menaut et
al. 1990; Vilà et al. 2001). In fact, it has been
suggested that many ecosystems around the
world that could develop into forest are instead
occupied by stable pyrogenic vegetation
maintained by fire, for example savanna
grasslands (Bond et al. 2005; Grigulis et al. 2005;
Hoffman et al. 2009) and shrublands (Mermoz et
al. 2005; Bowman et al. 2007; Warman and Moles
2009; Odion et al. 2010).
Among the major factors that have been
reported to determine the flammability of
vegetation are the amount of dead fuel held in the
vegetation and the quantity of herbaceous species
(Bradstock and Gill 1993; Baeza et al. 2002).
Dead wood, especially if it has a very fine
structure, confers high ignitability to fuels as it
usually has a very low moisture content (Bond and
van Wilgen 1996). Once this dead wood catches
fire, it acts as a primary first heat source in the
initial stages of the fire and contributes to the
secondary combustion of green fuel, thus helping
to propagate the fire and enhancing fire severity
(Johnson 1992; Sun et al. 2006; Schwilk 2003;
Santana et al. in press). Herbaceous species, on
the other hand, can produce a continuous
horizontal and vertical structure of fine fuel
(Bradstock and Gill 1993; Vilà et al. 2001), which
combined with the fact that they have a very fine
structure and they become very dry in
Mediterranean summer conditions may also result
in increasing the ignitability and spread of fires
(Cheney et al. 1993; Vilà et al. 2001). In fact, the
dead-to-live fine fuel ratio has been suggested as
an effective indicator of fire-risk in fire-prone
ecosystems (Bond and van Wilgen 1996; De Luis
et al. 2004), and its assessment through time after
fire may be crucial in helping to forecast future fire
occurrence (Minnich and Chou 1997).
In the western Mediterranean Basin, previous
studies have showed that seeding species with
fire-stimulated recruitment (i.e., soil stored seed
with dormancy broken by heat and/or smoke;
Moreira et al. 2010) exhibit traits that heighten
flammability; for example they have a high fine
fuel proportion, a high dead-to-live fuel ratio and
low ignition temperatures (Saura-Mas et al. 2010).
These observations suggest a link between post-
fire regenerative and flammability traits, resulting
in a possible fitness advantage for more
flammable species. In this case, an understanding
of factors that affect fire and any potential positive
feedbacks will be a key step in understanding
ecosystem function where there is recurrent fires.
It will clearly have relevance in the future, where
as consequence of projected climate change (with
longer, hotter and drier summers), an increased
fire occurrence in this region have been predicted

79
(Pausas 2004). In south-east Spain large areas of
former agricultural land was abandoned in the last
half of the twentieth century, and these areas have
been colonized almost exclusively by species with
these traits (Lloret et al. 2002, Baeza et al. 2007).
Consequently, the presence of these new fire-
adapted ecosystems increase the likelihood of fire
occurrence within the area, link forested areas and
increase both the size and frequency of fires
(Pausas 2004).
It has been hypothesised that the
accumulation of dead fuel in the canopy structure
would be driven by biological mechanisms linked
to plant succession (Odum 1969). Dead fuel
accumulation would be the consequence of
ontogenetic shifts in the flammability properties of
species and the turnover of species with different
flammability traits (Baeza et al. in press). In this
sense, the response of species and fuel types with
respect to time-since-fire may lead to different
functions of flammability depending on these
processes (McCarthy et al. 2001), for example:
flammability may (a) remain unchanged through
time, (b) increase constantly, (c) increase to an
asymptote, and (d) exhibit a humped-shaped
response where intermediate time periods
produce the greatest probability of fire and long
intervals a reduced probability (see Rothermel and
Philpot 1973; Bond and van Wilgen 1996; Díaz-
Delgado et al. 2004; Keane at al. 2004; Mermoz et
al. 2005; Baeza et al. 2006; Odion et al 2010;
Baeza et al. in press for some examples).
However, in spite of the importance in modelling
all of these factors to predict fire-risk, there is no
information available on the changing response of
species and fuel load in shrublands dominated by
seeding species in the western Mediterranean
Basin.
In this paper therefore, we report a study of the
successional response of the main species and
vital forms in terms of flammability in shrublands
dominated by seeding species within the western
Mediterranean Basin. We addressed three
questions: (1) How do the ecosystem structure
change during the post-fire succession? This
question was answered thought assessing the
change in life-forms, species, dead fuel load and
the presence of herbaceous species during a 25
year period. (2) How does recurrent fires alter
ecosystem structure? This question was answered
by comparing a post-fire succession after one fire
to a post-fire succession where there were two
fires. (3) Would those woody species that
accumulate the greatest amount of dead fuel be
benefited by increasing fire recurrence?, i.e. a
positive fire-vegetation’s flammability feedback
effect. This question was answered by comparing
the amount of dead fuel accumulated in vegetation
structure after one and two fires.
2. Methods
2.1 Study area
The study was carried out in the interior of the
Valencia region in the south-east of Spain (UTM
coordinates: 676400E - 4332099N, upper left
corner; 744675E - 4283954N, lower right corner)
over an area of approximately 4000 km2. The
region has a dry Mediterranean climate with an
annual rainfall range of 450-600 mm and a mean
annual temperature range of 13-16ºC. The study
plots were all located between 800 to 1050
m.a.s.l., and the bedrock were either marls or
marls mixed with limestone. The landscape is a
mountainous mosaic composed of Pinus
halepensis forests, agricultural land and
abandoned fields where shrubland and woodland
have regenerated naturally (nomenclature follows
Bolòs 1993). The potential vegetation of the area
would be a broad-leaved sclerophyllous forest of
Quercus ilex; this community is, however
extremely scarce at present due to past
deforestation and other exploitation, and is
confined to valley bottoms and other very limited
areas. Fire is an important factor shaping this
landscape and parts of this region are burned
almost every summer.
Eight sites were used in this study; they were
all P. halepensis forests that had colonized crop-
fields abandoned in the 1940s (Santana et al.

80
Table 1. Fire history description of the two seres at eight study sites. Sere
# Site nº fires 1st Fire 2nd Fire X UTM Y UTM Sampling date
1 Guadalest 1 1991 - 744675 4283954 1994, 1998, 2003 Banyeres 1 1991 - 704587 4289588 1994, 1998, 2003 Confrides 1 1991 - 738829 4285347 1994, 1998, 2003 Onil 1 1984 - 703392 4280698 1994, 1998, 2003, 2006, 2009 Pardines 1 1984 - 711302 4283187 1994, 1998, 2003, 2006, 2009 Ayora 1 1979 - 676400 4332099 2002, 2006, 2009 La Torre 1 1984 - 725918 4276893 1994, 1998, 2003 Fontanars 1 1978 - 697715 4293747 2009
2 Onil 2 1984 1994* 703339 4280706 1997, 1998, 2003, 2006, 2009 Pardines 2 1984 1994* 711215 4283194 1997, 1998, 2003, 2006, 2009 Ayora 2a 1979 1996 676684 4332010 2002, 2006, 2009 Ayora 2b 1979 1991 676537 4322175 2002, 2009 Ayora 2c 1979 1985 682077 4339697 2009 Ayora 2d 1979 1984 687777 4308441 2002, 2009 La Torre 2 1984 1994* 725936 4276847 1997, 1998, 2003 Fontanars 2 1978 1984 696615 4294317 2009
*= an experimental fire; Different letters in fire frequency means independent fires within the first fire. 2010), and had been subjected to a different fires
histories over the last 30 years (Table 1). At
present, their vegetation is dominated by shrub
species, which are mainly obligate seeders with
soil seedbanks and fire-enhanced recruitment
(Baeza and Vallejo 2006; Moreira et al. 2010),
such as Ulex parviforus, Cistus albidus and
Rosmarinus officinalis. There are also sparse
individuals of P. halepensis, a tree with serotinous
cones and a canopy seedbank. The resprouting
grass Brachypodium retusum dominates the
herbaceous strata. These sites were selected with
the aim of reducing environmental variability to a
minimum in that they experienced similar
morphological characteristics and land-use history
(Table 1). The only exception was the Guadalest
site, which unlike the others, had a southerly
aspect.
2.2 Design of the study
We studied two different seres of post-fire
succession covering a span of approximately 25
years. This was done using vegetation
assessments carried out between 1994 to 2009 to
build a sequence of vegetation dynamics for
vegetation either burned once (Sere 1) or twice
(Sere 2) (detailed in Baeza 2001; Baeza et al.
2007; Santana et al. 2010). Sere 1 was an
assessment of the response of the vegetation
after a single wildfire; one large patch was
available at each of the eight sites, and they were
sampled at 3-5 year intervals. The complete
history of fire occurrence and vegetation sampling
of this sere is summarised in Table 1. Sere 2 was
composed of sub-patches within the original
burned areas at each site (Sere 1) where a
secondary burn had occurred. The secondary
burn was either a second wildfire or an
experimental burn (Table 1, Baeza et al. 2002);
most sites had just one secondary burn but the
Ayora site had four independent fires within the
area burned once. These recurrent fires occurred
at time intervals ranging for 5 to 17 years after the
first fire (Table 1).
All together there were 16 sampling plots
(Table 1) and this design allowed us to depict a
period of successional time period of between 3
and 31 years for plots burned once, and a period
from 3 to 25 to plots burned twice. However, in
order to make comparisons between the two fire
recurrences, we only considered the first 25 years
of Sere 1.
2.3. Vegetation sampling
The vegetation was sampled in a standard
manner throughout. In each plot, between three

81
and five transects (3-7 m apart) were laid out and
the cover of all vascular species assessed using
the point-intersect method (Greig-Smith 1983).
Because the data were obtained from different
studies, the length of transects were variable
between 10-20 m, but the total sum of all transects
was always 50 m. A metal rod was used to record
contact with individual species, and
measurements were taken every 20 cm along the
transect (250 points per site). In addition, contacts
with dead woody fuel standing in vegetation
structure were also recorded.
2.3. Statistical analysis
Huisman-Olff-Fresco (HOF) models (Huisman et
al. 1993) were used to describe the response of
individual species to time-since-fire and fire
recurrence. HOF models are a means of
describing species response to environmental
gradients (here time) and intra- and inter-specific
interactions (Lawesson and Oksanen 2002). An
hierarchical series of five response models are
fitted, ranked by their increasing complexity
(Model I, no species trend; Model II, increasing or
decreasing trend; Model III, increasing or
decreasing trend below maximum attainable
response; Model IV, symmetrical response curve;
Model V, skewed response curve; Huisman et al
1993). In this study a Poisson error distribution
was used and the resultant equations are
presented in S1 in the Electronic Supplementary
Material). The AIC statistic (Akaike 1973) was
used to select the most parsimonious model for
species in each sere. For those species with
unimodal responses, the HOF procedure
estimated (1) the location of the maximum cover
reached during the succession (top), (2) the point
in time where this maximum cover is reached
(optimum) and (3) the niche width (based on the t-
intervals, t=tolerance, Huisman et al. 1993). In the
case of symmetric unimodal response (Model IV),
the lower and upper t-intervals around the
optimum are identical, whereas with the skewed
model (Model V), the t-intervals are unequal.
These model parameters (top, optimum and niche
width) were used to help interpret the post-fire
species response.
In addition, the species were divided into two
functional groups depending on their fuel type: i.e.
woody species and herbaceous species; these
were further subdivided as follows: woody species
into scrub (< 50 cm tall), shrubs ( 50 cm < X > 150
cm tall) and trees (>150 cm tall), and herbaceous
species into grasses and forbs. Classification of
species was made following Paula et al. (2009)
and authors’ personal observations. HOF models
were derived for each functional type. The
parameters for the selected models for both vital
forms and species are showed in the table S2 of
the ESM.
The change in the dead woody fuel was also
modelled with respect to time-since-fire by fitting
second- and third-order polynomial functions; the
reduction in the AIC statistic was used to choose
the best fit curve (2nd order for both seres).
All statistical analyses were implemented in
the R software environment (version 2.10.1; R
Development Core Team, Vienna, Austria,
http://www.r-project.org/). The HOF models were
fitted using the GRAVY package (R package
version 0.0-21, http://cc.oulu.fi/~jarioksa/softhelp/
softalist.html).
3. Results
For Sere 1, the woody vegetation was dominated
by shrubs (Figure 1), which experienced a skewed
response (model V). They reached a cover
optimum approximately 7 years after fire and,
cover was maintained at relatively high level
(>60%) throughout the sere albeit decreasing
slowly with time (Figure 1, Table 2A). Trees and
forbs increased slightly towards the end of the
sere and grasses showed an unimodal responses
peaking at approximately 13 years.
In Sere 2, the cover of the shrubs, the main
woody species, was reduced in the early stages
(Figure 1). The response model changed from V
to III and their cover in the initial stages of
succession was less than in Sere 1; however, the
shrub cover was comparable to those found in

82
Shrubs
5 10 15 20 25
Scrub
5 10 15 20 25
Grasses
Years5 10 15 20 25
Res
pons
e
0,0
0,2
0,4
0,6
0,8
1,0
Forbs
Years5 10 15 20 25
Woody species
Herbaceous species
5 10 15 20 25
Res
pons
e
0,0
0,2
0,4
0,6
0,8
1,0
1 Fire2 Fires
Trees
Figure 1. HOF models for the vital forms in relation to time-since-fire in two seres, one subject to a single fire and the second subject to two fires.
Sere 1 approximately 20 years after the second
burn (Figure 1, Table 2A). The responses of the
grasses were very similar to Sere 1 but they
increased slightly in abundance and niche width
(Figure 1, Table 2A). Forbs and especially trees
remained very low throughout, and there was no
evidence of an increase in trees at the end of the
sere.
In both seres, the changes in responses of
the functional types were driven by changes in the
component species. However, there were
important differences between the two seres
(Figure 2, Table 2B). In Sere 1, U. parviflorus and
C. albidus, with unimodal responses were the
pioneer species peaking at 8.8 and 13.6 years,
followed by R. officinalis and P. halepensis still
both increasing at 25 years. R. officinalis was the
dominant species in the late stages of the
succession. In sere 2, a similar pattern was found
for the two pioneer species in that both U.
parviflorus and C. albidus had unimodal
responses, however, here the peaks were delayed
until 16.1 and 15.0 years. In addition, their
response in abundance was different between
them: U. parviflorus decreased its top (maximum
cover reached) from 0.69 to 0.28, whereas C.
albidus increased from 0.09 to 0.21. In the last
stages of the succession R. officinalis showed an
almost identical increasing pattern to sere 1 but
with a slight decrease in abundance. P.
halepensis was not found in sere 2.
For herbaceous species, the dominant
species in both seres was the grass B. retusum
which experienced an unimodal response in
relation to time-since-fire (model IV) (Figure 2,
Table 2B). This species experienced an increase
in abundance and niche width in Sere 2 (Figure 2,
Table 2B). In addition, other grasses as
Brachypodium phoenicoides also showed an
increasing trend after the second fire, changing
from a model I to a model V (Figure 2, Table 2B).
The polynomial function fitted to the
dead woody fuel in Sere 1 (second-order, p<
0.001, F2, 20= 35.86, R2= 0.78) showed a
gradual increase from 0 in the third year to
50% cover at 17 years and then a slight
decrease to 40% cover at 25 years (Figure 3).
In Sere 2, the amount of dead fuel was
always less than Sere 1 and it experienced
a gradual increase until approximately

83
Table 2. HOF model parameters in relation to fire recurrence for A) vital forms and B) the main species; the estimated values for the top (maximum response), the optimum (age at which the top value is estimated) and niche width are presented. The niche width is calculated for the unimodal Model IV as 2 x the tolerance value (2xt), and for the skewed model V as the sum of the left and right tolerance values (t+t). The % reduction in deviance of the selected model relative to the null model (Model I) is presented. A)
Sere
# Functional
group Vital form
HOF model
Top Optimum 2xt t+t Deviance reduction (%)
df
1 Trees II - - - - 22,1 21
Woody species
Shrubs V 0,83 6,97 - 3,96 + 52,95 56.9 19
Scrubs I - - - - - 22
Grasses IV 0,58 12,13 16,62 - 19,3 20
Herbaceous species
Forbs II - - - - 41,4 21
2 Trees II - - - - 22,1 20
Woody species
Shrubs III - - - - 85,1 19
Scrubs I - - - - - 21
Grasses IV 0,64 13,42 17,02 - 19,7 19
Herbaceous species
Forbs I - - - - - 21
B)
Sere #
Functional group
Species Vital form
HOF model
Top Optimum 2xt t+t Deviance reduction (%)
df
1 Pinus halepensis Tr II - - - - 21,5 21
Woody species
Cistus albidus Sh IV 0,09 13,59 12,56 - 17,5 20
Rosmarinus officinalis Sh II - - - - 60,2 21
Ulex parviflorus Sh IV 0,69 8,83 13,12 - 70,8 20
Brachypodium retusum Gr IV 0,56 12,25 15,82 - 22,2 20
Herbaceous species
Brachypodium phoenicoides Gr I - - - - - 22 2
Pinus halepensis Tr - - - - - - -
Woody species
Cistus albidus Sh IV 0,21 15,03 13,24 - 37,3 19
Rosmarinus officinalis Sh II - - - - 61,6 20
Ulex parviflorus Sh IV 0,28 16,07 11,04 - 51,7 19
Brachypodium retusum Gr IV 0,61 13,51 17,10 - 18,7 19
Herbaceous species
Brachypodium phoenicoides Gr V 0,08 5,74 - 0,88 + 4,01 34,7 18 Sh= shrub, Tr= tree, Sc= scrub, Gr= grass, F= forb. 37% cover at 25 years (second-order, p< 0.001,
F2, 19= 15.19, R2= 0.61; Figure 3).
4. Discussion
4.1 Vegetation structure along succession
This paper is the first to attempt to assess the
change in species and life-history attributes in
terms of flammability during the post-fire
succession in ecosystems dominated by seeding
species with the western Mediterranean Basin.
We answered three questions. First we showed
that over a 25 year period there was an unimodal
response of both grasses and shrubs after a
single fire, increasing to a peak and then reducing.
The only groups that showed a continuing small
increase with time since burning are the forbs and
the trees. This overall pattern obscured changes
in individual species with a clear replacement of
shrub species with time (U. parviforus > C.albidus
> R. officinalis).

84
P. halepensis (Tr)
Res
pons
e
0,0
0,2
0,4
0,6
0,8
1,0
1 Fire2 Fires
C. albidus (Sh) R. officinalis (Sh)
U. parviflorus (Sh)
5 10 15 20 25
Res
pons
e
0,0
0,2
0,4
0,6
0,8
1,0
B. retusum (Gr)
Years5 10 15 20 25
B. phoenicoides (Gr)
Years5 10 15 20 25
Figure 2. HOF models for the main species in relation to time since fire in two Seres, one subject to a single fire and the second subject to two fires. Sh= shrub, Tr= tree, , Gr= grass, F= forb.
Second, we showed that a second fire
changed the post-fire dynamics, the main results
being that grasses and shrubs peaked slightly
later than subject to a single fire. Moreover, there
were almost no forbs or trees after a second fire.
The order of shrub turnover remained similar to
the single-burn sere in shrubs, however, their
abundance was quite reduced. The response to a
recurrent fire was variable depending on the
species but their balance was negative. The
largest shrubs U. parviflorus declined with a
second fire but other species, such as C. albidus,
were enhanced.
Third we showed that dead wood was
accumulated immediately after fire but as
polynomial responses were detected for both
seres this implies a decline in the later stages of
the succession. After a single fire the dead wood
accumulation increased to 17 years and then here
was a reduction up to the 25 years sampled. After
a second fire, the response was delayed, and
although a polynomial was fitted the dead wood
was always inferior to the one single fire sere.
Therefore, for these systems, our initial
hypotheses that there was a positive fire-mediated
switch for the shrubby species with higher
accumulation of dead fuel and fire-enhanced
recruitment throughout the succession (sensu
Wilson and Agnew 1992) is rejected. This
suggests that these ecosystems are not a stable
self-maintaining community with respect to
successive fires within the fire-return interval
studied here, although more fire recurrences are
needed to test this hypothesis.
In addition to the change in woody fuels we
also found an increase in herbs (i.e., grasses and
forbs) and scrub during the succession. Although
these life-forms do not accumulate large amounts
of dead woody fuel they do impact on flammability
at community level. They are small and short-lived
species and produce a very fine, dry fuel load that
can produce a continuous horizontal fuel bed
throughout the community. Most of the species in
this strata have effective strategies for
regeneration after disturbance. Scrub species
ripen and produce seeds very early after fire (1-3
years) and some species regenerate by
resprouting (facultative species sensu Pausas et
al. 2004). Grasses also show high fire-tolerance
as they resprout quickly and can prevent the
establishment of shrub and tree seedlings
(Menaut et al. 1990; Berkowitz et al. 1995 Vilà et
al 2001; Grigulis et al. 2005). In our study area we
found an increase in B. retusum and scrub
species after three fires (Santana et al. 2010), and
similar patterns have been observed for the grass

85
Ampelodesmos mauritanica elsewhere in the NE
of Iberian Peninsula (Vilà et al 2001; Grigulis et al.
2005). In fact, positive feedback between grass
expansion and susceptibility to fire has been
previously described in many fire-prone
ecosystems (Hughes et al. 1991; D’Antonio and
Vitousek 1992; Bond et al. 2005; Hoffman et al.
2009), and in the case of the studied fire-return
interval studied, it may successfully establish a
positive feedback between their expansion and
susceptibility to fire.
4.1. Selection of plant traits in fire-prone
ecosystems
There is little information about the ecological
significance of retaining traits that enhance
flammability such as increasing the dead fuel load
within the vegetation. Mutch (1970) hypothesised
that these traits could be under positive selection
in fire-prone environments, i.e., fire-dependent
species burn more readily than non-fire dependent
plant ones because selection has favoured
characteristics that make them more flammable
(Bond and Midgley 1995; Zedler 1995; Kerr et al.
1999; Schwilk and Ackerly 2001). However, these
suggestions have been derived form studies of
individual-level selections, and they have been
criticized because putative flammability is an
emergent property of communites made up of
different species rather than form characteristics
of individual plants (Troumbis and Trabaud 1989;
Whelan 1995). Here, regardless of what happens
at the individual level, at the community level we
observed that the flammability enhancing trait
(retention of dead woody fuel) is not under positive
selection and hence providing a fitness
advantage. The regeneration after a recurrent fire
in approximately the period of the highest fire-risk
(5-17 years) led to a community with less amount
of dead woody fuel accummulated; the intervals of
time between fires may be too short for plants to
reach reproductive age and replenish successfully
their seedbank and, consequently, these species
can be rapidly excluded (Zedler et al. 1983;
Jacobsen et al. 2004; Eugenio and Lloret 2006;
Vilà-Cabrera et al. 2008). However, in longer
periods of time in absence of fire, when the
species may have replenished succesfully their
seedbank, the amount of dead fuel enhancing fire
probability is also reduced.
Dead wood
5 10 15 20 25
Cov
er (
%)
0
20
40
60
80
100
,3
Figure 3. Polynomial regressions for the evolution at community level of the dead woody fuels in relation to the time-since-fire. Filled dots and solid line correspond to one fire and empty dots and dashed line to two fires.
It is also possible that flammability-related
traits could be the result of physiological and
morphological responses to other selection
pressures found in Mediterranean ecosystems, for
example herbivory and seasonal changes in
temperature, drought and radiation (Snyder 1984,
Lavorel and Garnier 2002, Ackerly 2004).
Therefore, we hypothesise that the humped
function of flammability observed during the post-
fire succession, i.e., the woody fuel reached an
asymptote with time but the dead fuel
accumulation decreased after reaching a
maximum may be the result of intrinsic traits of
individual species as they occur within the
successional gradient of environmental resources.
Early-successional species from resource-rich
habitats after perturbation tend to be inherently
short-lived and fast growing with a high resource
capture and fast turnover of organs (leaves), while
the reverse would be for late-successional species
(Bazzaz 1979, Carreira and Niell 1992). Thus,
early-successional species might be expected to
have early and prolific branching which would
reduce light availability and, in certain cases,
increase the dead wood component of the lowest
branches (Schlessinger and Gill 1980, Larcher
1995). In addition, changes in resource allocation

86
pattern between shoots and roots has also been
suggested as a possible determinant of dead fuel
accumulation (Schwilk and Ackerly 2005, Cowan
and Ackerly 2010). Species with a high shoot: root
ratio would be expected to be less drought tolerant
in the summer and hence there might be an
increase dieback of part of the biomass in summer
droughts (Orsham 1963, Montserrat-Martí 2004).
Thus, an increased allocation to above-ground
biomass in early-successional species combined
with an increased shoot dieback over time may
drive differences in dead fuel retention. This
hypothesis is supported in our study as the early-
successional U. parviflorus allocated less biomass
to roots (lower root:shoot ratio) that the late-
successional R. officinalis (Hernandez et al.
2010). Nevertheless, little is known about the
physiological causes of dead fuel production, and
further investigations are needed to confirm these
speculations. The fire-risk function depicted by the
woody fuels through time is in accordance with the
results found on these systems by Baeza et al. (in
press) and other fire-prone systems, where long
intervals without fire lead to a decreased
probability of fire due to the successional
replacement with less-flammable species (Bond
and van Wilgen 1996, Mermoz et al. 2005, Odion
et al 2010).
Acknowledgements
V.M. Santana was supported by a FPU grant
awarded by the Spanish Ministry of Education and
Science. This research was carried out within the
FIREMED (AGL200/8-04522/FOR) and
Consolider-Ingenio 2010 (GRACCIE CSD2007-
00067) projects. CEAM is supported by the
Generalitat Valenciana and Fundación Bancaja.
References
Ackerly DD (2004) Functional strategies of
chaparral shrubs in relation to seasonal water
deficit and disturbance. Ecol Monogr 74: 25-44
Akaike H (1973) Information theory and an
extension of the maximum likelihood principle.
In: Petrov BN, Csaki F (eds) 2nd international
symposium on information theory. Akademiai
Kiado, Budapest.
Baeza MJ (2001) Aspectos ecológicos y técnicas
de control de combustible (roza y quema
controlada) en matorrales con alto riesgo de
incendio dominados por Ulex parviflorus
(Pourr.). Ph.D thesis, Departamento de
Ecología. Universidad de Alicante.
Baeza MJ, De Luis M, Raventòs J, Escarré A
(2002) Factors influencing fire behaviour in
shrublands of different stand ages and the
implications for using prescribed burning to
reduce wildfire risk. J Environ Manag 65: 199-
208
Baeza MJ, Vallejo VR (2006) Ecological
mechanisms involved in dormancy breakage in
Ulex parviflorus seeds. Plant Ecol 183: 191-
295
Baeza MJ, Raventòs J, Escarré A, Vallejo VR
(2006) Fire risk and vegetation structural
dynamics in Mediterranean shrubland. Plant
Ecol 187: 189-201
Baeza MJ, Valdecantos A, Alloza JA, Vallejo VR
(2007) Human disturbance and environmental
factors as drivers of long-term post-fire
regeneration patterns in Mediterranean
forests. J Veg Sci 18: 243-252
Baeza MJ, Santana VM, Pausas JG, Vallejo VR
(in press) Successional trends in standing
dead biomass in Mediterranean Basin species.
J Veg Sci
Bazzaz FA (1979) The physiological ecology of
plant succession. Ann Rev Ecol Syst 10: 351-
371
Berkowitz AR, Canham CD, Kelly VR (1995)
Competition vs. facilitation of the tree seedling
growth and survival in early-successional
communities. Ecology 76: 1156-1168

87
Bolòs O, Vigo J, Masalles RM, Ninot JM (1993)
Flora manual dels Països Catalans. Pòrtic,
Barcelona.
Bond WJ, Midgley JJ (1995) Kill thy neighbour: an
individualistic argument for the evolution of
flammability. Oikos 73: 79-85
Bond WJ, van Wilgen BW (1996) Fire and plants.
Chapman & Hall, London.
Bond WJ, Woodward FI, Midgley GF (2005) The
global distribution of ecosystems in a world
without fire. New Phytol 165: 525-538
Bowman DMJS, Boggs GS, Prior LD (2007) Fire
maintains an Acacia aneura shrubland- Triodia
grassland mosaic in central Australia. J Arid
Environ 72: 34-47
Bradstock RA, Gill AM (1993) Fire in semi-arid,
Mallee shrublands: size of flames from
discrete fuel arrays and their role in the spread
of fire. Int J Wildland Fire 3: 3-12
Carreira JA, Niell FX (1992) Plant nutrient
changes in a semiarid Mediterranean
shrubland after fire. J Veg Sci 3: 457-466.
Cheney NP, Gould JS, Catchpole WR (1993) The
influence of fuel, weather and fire shape
variables on fire-spread in grasslands. Int J
Wildland Fire 3: 31-44
Cowan PD, Ackerly DD (2010) Post-fire
regeneration strategies and flammability traits
of California chaparral shrubs. Int J Wildland
Fire 19: 984-989
D’Antonio CM, Vitousek PM (1992) Biological
invasions by exotic grasses, the grass-fire
cycle, and global change . Ann Rev Ecol Syst
23: 63-87
De Luis M, Baeza MJ, Raventós J, González-
Hidalgo JC (2004) Fuel characteristics and fire
behaviour in mature Mediterranean gorse
shrublands. Int J Wildland Fire 13: 79-87
Díaz-Delgado R, Lloret F, Pons X (2004)
Statistical analysis of fire frequency models for
Catalonia (NE Spain, 1975-1998) Based on
fire scar maps from Landsat MSS data. Int J
Wildland Fire 13: 89-99
Eugenio M, Lloret F (2006) Effects of repeated
burning on Mediterranean communities of the
northeastern Iberian Peninsula. J Veg Sci 7:
755-764
Gagnon PR, Passmore HA, Platt WJ, Myers JA,
Paine CET, Harms KE (2010) Does
pyrogenicity protect burning plants? Ecology
91: 3481-3486
Greig-Smith P (1983) Quantitative plant ecology.
Blackwell, Oxford.
Grigulis K, Lavorel S, Davies ID, Dossantos A,
Lloret F, Vilà M (2005) Landscape-scale
positive feedbacks between fire and expansion
of the large tussock grass, Ampelodesmos
mauritanica in Catalan shrublands. Global
Change Biol 11: 1042-1053.
Hernández E, Vilagrosa A, Pausas JG, Bellot J
(2010) Morphological traits and water use
strategies in seedlings of Mediterranean
coexisting species. Plant Ecol 207: 233-244
Hoffmann WA, Adasme R, Haridasan M, de
Carvalho MT, Geiger EL, Mireia MAB, Gotsch
SG, Franco AC (2009) Tree topkill, not
mortality, governs the dynamics of savanna-
forest boundaries under frequent fire in central
Brazil. Ecology 90: 1326-1337
Hughes F, Vitousek PM, Tunison T (1991) Alien
grass invasion and fire in the seasonal
submontane zone of Hawaii. Ecology 72: 743-
746
Huisman J, Olff H, Fresco LFM (1993) A
hierarchical set of models for species
response analysis. J Veg Sci 4: 37-46
Jacobsen AL, Davis SD, Babritius SL (2004) Fire
frequency impacts non-sprouting chaparral
shrubs in the Santa Monica Mountains of
southern California. In: Arianoutsou M,
Papanastasis VP (eds) Ecology, conservation

88
and management of Mediterranean climate
ecosystems. Millpress, Rotterdam.
Johnson EA (1992) Fire and vegetation dynamics:
studies from the North American boreal forest.
Cambridge University Press, Cambridge.
Keane RE, Cary GJ, Davies ID, Flannigan MD,
Gardner RH, Lavorel S, Lenihan JM, Li C,
Rupp TS (2004) A classification of landscape
fire succession models: spatial simulations of
fire and vegetation dynamics. Ecol Model 179:
3-27.
Kerr B, Schwilk DW, Bergman A, Feldman MW
(1999) Rekindling an old flame: a haploid
model for the evolution and impact of
flammability in resprouting plants. Evol Ecol
Res 1: 807-833
Larcher W (1995) Physiological plant ecology, 3rd
edn. Springer, Berlin.
Lavorel S, Garnier E (2002) Predicting changes in
community composition and ecosystem
functioning from plant traits: revisiting the Holy
Grail. Funct Ecol 16: 545-556
Lawesson JE, Oksanen J (2002) Niche
charateristics of Danish woody species as
derived from coenoclines. J Veg Sci 13: 279-
290
Lloret F, Calvo E, Pons X, Díaz-Delgado R (2002)
Wildfires and landscape patterns in the
eastern Iberian Peninsula. Landscape Ecol 17:
745-759
McCarthy MA, Gill AM, Bradstock RA (2001)
Theoretical fire interval distributions. Int J
Wildland Fire 10: 73-77
Menaut JC, Gignoux J, Prado C, Clobert J (1990)
Tree community dynamics in a humid savanna
of the Cote-d’Ivoire: modelling the effects of
fire and competition with grass and
neighbours. J. Biogeogr 17: 471-481
Mermoz M, Kitzberger T, Veblen T (2005)
Landscape influences on occurrence and
spread of wildfires in Patagonian forests and
shrublands. Ecology 86: 2705-2715
Minnich RA, Chou YH (1997) Wildland fire patch
dynamics in the chaparral of southern
California and northern Baja California. Int J
Wildland Fire 7: 221-248
Montserrat-Martí G, Palacio-Blasco S, Milla
Gutiérrez R (2004) Fenología y características
funcionales de las plantas leñosas
mediterráneas. In: Valladares F (ed) Ecología
del bosque mediterráneo en un mundo
cambiante. Ministerio de Medio Ambiente,
Madrid.
Moreira B, Tormo J, Estrelles E, Pausas JG
(2010) Disentangling the role of heat and
smoke as germination cues in Mediterranean
Basin flora. Ann Bot 105: 627-635
Mutch RW (1970) Wildland fires and ecosystems-
a hypothesis. Ecology 51: 1046-1051.
Odion DC, Moritz MA, DellaSala D (2010)
Alternative community states maintained by
fire in the Klamath Mountains, USA. J Ecol 98:
96-105
Odling-Smee FJ, Laland KN, Feldman MW (1996)
Niche construction. Am Nat 147: 641-648
Odum EP (1969) Strategy of ecosystem
development. Science 164: 262-270.
Orsham G (1963) Seasonal dimorphism of desert
and Mediterranean chamephytes and its
significance as a factor in their water economy.
In: Rutter AJ, Whitehead FH (eds) The water
relations of plants. Blackwell Scientific
Publications, London.
Paula S, Arianoutsou M, Kazanis D, Tavsanoglu
Ç, Lloret F, Buhk C, Ojeda F, Luna B, Moreno
JM, Rodrigo A, Espelta JM, Palacio S,
Fernández-Santos B, Fernandes PM, Pausas
JG (2009) Fire-related traits for plant species
of the Mediterranean Basin. Ecology 90: 1420

89
Pausas JG (2004) Changes in fire and climate in
the eastern Iberian Peninsula (Mediterranean
Basin). Climatic Change 63: 337-350
Pausas JG, Bradstock RA, Keith DA, Keeley JE,
Network GF (2004) Plant functional traits in
relation to fire in crown-fire ecosystems.
Ecology 85: 1085-1100
Rothermel RC, Philpot CW (1973) Predicting
changes in chaparral flammability. J Forest 71:
640-643.
Santana VM, Baeza MJ, Marrs RH, Vallejo VR
(2010) Old-field secondary succesión in SE
Spain: can fire divert it? Plant Ecol 211: 337-
349
Santana VM, Baeza MJ, Vallejo VR (in press) Fuel
structural traits modulating soil temperatures in
different species patches of Mediterranean
Basin shrublands. Int J Wildland Fire
Saura-Mas S, Paula S, Pausas JG, Lloret F
(2010) Fuel loading and flammability in the
Mediterranean Basin woody species with
different post-fire regenerative strategies. Int J
Wildland Fire 19: 783-794
Schlessinger WH, Gill DS (1980) Biomass,
production, and changes in the availability of
light, water, and nutrients during the
development of pure stands of the chaparral
shrub, Ceanothus megacarpus, after fire.
Ecology 64: 781-789
Schwilk DW (2003) Flammability is a niche
construction trait: canopy architecture affects
fire intensity. Am Nat 162: 725-733
Schwilk DW, Ackerly DD (2001) Flammability and
serotiny as strategies: correlated evolution in
pines. Oikos 94: 326-336
Schwilk DW, Ackerly DD (2005) Is there a cost of
resprouting? Seedling growth rate and
tolerance in sprouting and non-sprouting
Ceanothus (Rhamnaceae). Am J Bot 92: 404-
410
Snyder JR (1984) The role of fire: Mutch ado
about nothing. Oikos 43: 404-405
Sun L, Zhou X, Mahalingam S, Weise DR (2006)
Comparison of burning characteristics of live
and dead chaparral fuels. Combust Flame
144: 349-359
Troumbis AY, Trabaud L (1989) Some questions
about flammability in fire ecology. Acta
Oecologica-Oecologia Plantarum 10: 167-175
Vilà M, Lloret F, Ogheri E, Terradas J (2001)
Positive fire-grass feedback in Mediterranean
Basin woodlands. Forest Ecol Manag 147: 3-
14
Vilà-Cabrera A, Saura-Mas S, Lloret F (2008)
Effects of fire frequency on species
composition in a Mediterranean shrubland.
Ecoscience 15: 519-528
Warman L, Moles AT (2009) Alternative stable
states in Australia’s wet tropics: a theoretical
framework for the field data and field-case for
the theory. Landscape ecol 24: 1-13
Whelan RJ (1995) The ecology of fire. Cambridge
University Press. Cambridge.
Wilson JB, Agnew ADQ (1992) Positive-feedback
switches in plant-communities. Adv Ecol Res
23: 263-336
Zedler PH (1995) Are some plants born to burn?
TREE 10: 393-395
Zedler PH, Gautier CR, McMaster GS (1983)
Vegetation change in response to extreme
events: the effect of a short interval between
fires in California chaparral and coastal scrub.
Ecology 64: 809-818

90
ELECTRONIC SUPPLEMENTARY MATERIAL Table S1. The equations fitted to the 5 models in the HOF procedure; y = the species response variable; x = the gradient variable; a, b, c and d are parameters to be estimated; M = a constant and is the maximum value attainable (M = 250 in our case).
HOF Model Equation
I y = M * 1/(1+ea)
II y = M * 1/(1+ ea+bx)
III y = M * 1/(1+ ea+bx) * 1/(1+ec)
IV y = M * 1/(1+ ea+bx) * 1/(1+ec-bx)
V y = M * 1/(1+ ea+bx) * 1/(1+ec-dx) where b and d have opposite signs

91
Table S2. HOF model parameters for the complete set of A) vital forms and B) species fitted. A Poisson error distribution was used to compute these values using the GRAVY package (R package version 0.0-21, http://cc.oulu.fi/~jarioksa/softhelp/softalist.html). A) Sere Functional
Model parameters # group
Vital form HOF model
a b c d
1 Trees II 4,23 -2,18 - -
Woody species
Shrubs V -1,72 0,66 -0,37 28,72
Scrub I 2,10 - - -
Grasses IV -3,01 4,46 0,69 -
Herbaceous species
Forbs II 4,59 -2,39 - -
2 Trees II 7,13 -2,63 - -
Woody species
Shrubs III 1,53 -8,30 -0,95 -
Scrub I 1,92 - - -
Grasses IV -3,57 4,60 0,78 -
Herbaceous species
Forbs I 2,85 - - - B) Sere Functional
Model parameters # group
Species Vital form
HOF model
a b c d 1
Pinus halepensis Tr II 4,22 -2,20 - -
Woody species
Cistus albidus Sh IV -1,80 5,53 3,49 -
Rosmarinus officinalis Sh II 2,83 -3,00 - -
Ulex parviflorus Sh IV -3,30 6,29 0,05 -
Brachypodium retusum Gr IV -3,00 4,62 0,85 -
Herbaceous species
Brachypodium phoenicoides Gr I 3,76 - - - 2
Pinus halepensis Tr - - - -
Woody species
Cistus albidus Sh IV -2,50 4,87 2,82 -
Rosmarinus officinalis Sh II 4,15 -4,20 - -
Ulex parviflorus Sh IV -3,60 5,89 3,35 -
Brachypodium retusum Gr IV -3,40 4,43 0,85 -
Herbaceous species
Brachypodium phoenicoides Gr V 6,65 -80,00 2,07 -3,10 Sh= shrub, Tr= tree, Sc= scrub, Gr= grass, F= forb.

92
Apéndice fotográfico
Foto 1. Matorral de 3 años de edad después de un fuego Foto 2. Matorral de 9 años de edad después de 1 fuego
Foto 3. Matorral de 25 años con individuos senescentes
Foto 4. Individuos senescentes de Ulex parviflorus Foto 5. Matorral quemado 2 veces a los 10 años de edad

93
CAPÍTULO 6- SUCESIÓN SECUNDARIA EN CAMPOS DE CULTIVO ABANDONADOS DEL SURESTE DE ESPAÑA: ¿PUEDE EL FUEGO DESVIARLA? RESUMEN: En la cuenca mediterránea, grandes áreas cultivadas fueron abandonadas en el pasado siglo y hoy en día se encuentran en varias etapas de sucesión. El objetivo de este trabajo es analizar las trayectorias sucesionales de estos ecosistemas, y valorar las posibles desviaciones en estas trayectorias debido a la ocurrencia de incendios en altos o bajos niveles de recurrencia. Campos de cultivo abandonados aproximadamente 50 o 100 años atrás fueron seleccionados en el sureste de España. Dentro de los campos de cultivo abandonados hace 50 años, se establecieron parcelas que habían sido quemadas por 1, 2 y 3 incendios en los últimos 25 años. Los valores de cobertura para todas las especies vasculares fueron estimados y después analizados mediante análisis multivariante. Las distancias euclídeas entre las comunidades resultantes fueron usadas como un indicador de la posible desviación de la trayectoria sucesional en ausencia de fuegos. Nuestros resultados señalan la posibilidad de que existan diferentes trayectorias sucesionales dependiendo de la ocurrencia y recurrencia de incendios. En ausencia de fuego, la vegetación es dominada por especies pioneras, principalmente Pinus. Con el paso del tiempo esta vegetación pasa a estar dominada por especies arbóreas de etapas sucesionales tardías (Quercus). Sin embargo, cuando las etapas sucesionales tempranas son afectadas por el fuego, la sucesión puede ser desviada. Un simple incendio es suficiente para cambiar bosques de Pinus en estados estables alternativos de de matorral dominado por Rosmarinus officinalis, donde la colonización de especies en etapas sucesionales posteriores puede verse impedida. La desviación se incrementa bajo un régimen de alta recurrencia de incendio, donde la vegetación cambia hacia especies de bajo porte y herbáceas. Este capítulo reproduce el siguiente manuscrito: Santana VM, Baeza MJ, Marrs RH, Vallejo VR (2010) Old-field secondary succession in SE Spain: can fire divert it? Plant Ecology 211: 337-349

94

95
Old-field secondary succession in SE Spain: can fir e divert it?
Victor M. Santana A, M. Jaime BaezaA, B, Rob H. MarrsC V. Ramón VallejoA
AFundación de la Generalitat Valenciana Centro de Estudios Ambientales del
Mediterráneo (CEAM). Parque Tecnológico Paterna. C/Charles Darwin, 14.
46.980 Valencia. Spain
BDepartamento de Ecología, Universidad de Alicante. Ap. 99. 03080 Alicante.
Spain CApplied Vegetation Dynamics Laboratory, School of Environmental Science,
University of Liverpool, PO Box 147, Liverpool, L69 7ZB, UK
Abstract In the Mediterranean Basin, most cultivated areas were abandoned in the last century and are now in various stages of old-field succession. The aim of this work was to analyse the successional trajectories of these ecosystems, and to assess possible deviations in these pathways due to fire occurrence at high or low recurrence levels. Old-fields abandoned either about 50 or about 100 years ago were selected in SE Spain. Within the 50-year-old abandoned fields, plots were established which had been burned by 1, 2 and 3 fires in the last 25 years. Cover values of vascular species were sampled and then analysed by means of multivariate analysis. Euclidean distances between resulting communities were used as an indicator of the possible deviation from the unburned successional pathway. Our results pointed to the possibility that different successional pathways may exist depending on fire occurrence and recurrence. In the absence of fire, the vegetation is dominated by pioneer species, mainly Pinus. With the passage of time this vegetation will become dominated by later successional tree species (Quercus). However, when early-successional communities are affected by fire, the succession can be diverted. A single fire is enough to change Pinus forests into alternative stable-states dominated by Rosmarinus officinalis shrub communities, where the colonisation of species in later-successional stages is arrested. This deviation increases in high fire-recurrence regimes where the vegetation changes to dwarf shrubs and herbs. Key words: alternative stable-state; arrested succession; CCA; fire recurrence; Mediterranean vegetation; successional pathway.

96
1. Introduction
In the Mediterranean Basin, late-successional
forests are associated mainly with broad-leaved
sclerophyllous species. In fact, under mesic
conditions, Quercus ilex is often the dominant
species in eastern Spain (Barbero et al. 1992;
Zavala et al. 2000, Zavala 2003; Quézel 2004).
However, in the Mediterranean Basin, there is a
long history of exploitation and deforestation, and
most of the natural vegetation was removed
several centuries ago and the land converted to
agriculture (Blondel and Aronson 1999). More
recently, some of this agricultural land has been
taken out of production and natural regeneration
has occurred. As a result, this has produced a
series of old-field successional communities in this
region (Cramer et al. 2008). An inevitable
consequence of this successional development
has been an increased in vegetation biomass and
hence an added fire risk. This is a particular threat
when the successional patches link forested areas
together. These changes in land-use, together
with climate change (longer, hotter and drier
summers), are contributing factors to the
increased size and frequency of fires in recent
decades (Pausas 2004).
Although old-field successions in the
Mediterranean Basin have been widely studied
(Debussche et al. 1982; Tatoni and Roche 1991;
Debussche et al. 1996; Ne’eman and Izhaki 1996;
Verdú and García-Fayos 1998; Bonet and Pausas
2007), few of them cover long time periods. It is
widely accepted that when cultivation practices
have not exceeded the biotic and abiotic
degradation thresholds of the ecosystem, plant
communities assemble along a broadly repeatable
pathway to largely resemble the composition,
structure, and function of the late-successional
state that existed before clearing (Cramer et al.
2008). The transition to late stages of succession
is often slow because the propagule bank in the
soil necessary for species establishment has been
depleted or destroyed. Plant establishment is,
therefore, dependent on the dispersal and
subsequent germination of seeds from off-site
sources. This dispersal limitation is more likely to
impede colonization of the late-successional
species, which tend be slower-growing and are
dispersed by frugivores (Debussche et al. 1982,
Pons and Pausas 2007) than early–successional
species which are usually faster-growing and have
a high dispersal capacity (Grubb 1998; Bonet and
Pausas 2004).
Succession after field abandonment, where all
of the pre-existing vegetation has been removed,
differs from succession after fire, where many
propagules of the original species are left in situ.
Classically, Mediterranean systems have been
described as auto-successional after fire, i.e., the
fire-affected vegetation recovers its previous
composition with time (Hanes 1971). In this case,
fires have been viewed as processes that interrupt
succession, or delay it, by returning the system to
an earlier successional state. Nevertheless, it is
possible that disturbance can sometimes cause an
abrupt change in ecosystem function and/or
structure, diverting the succession to alternative
stable-states that differ substantially from the
original system (Friedel 1991; Laycock 1991;
Scheffer et al. 2001). In cases where the
alternative states are persistent and the
established vegetation prevents the development
of late-successional species, succession is
strongly delayed or practically stopped in an
arrested succession (Putz and Canham 1992;
Acacio et al. 2008). Factors such as the fire
disturbance regime (extent, frequency, intensity,
severity, seasonality and recurrence- see Fox and
Fox 1987), climatic conditions before, during and
after the disturbance, the previous species
composition and the previous land use can modify
the successional trajectory to follow multiple
pathways in a given environment (Picket et al.
1987; Keeley et al. 2005; Cramer et al. 2008). In
fire-prone ecosystems, deviations from the
expected successional pathways have been
proposed especially where there is a high fire
recurrence over relatively short time periods
(Noble and Slatyer 1980; Trabaud 1991; Eugenio
and Lloret 2004; Donato et al. 2008). As a

97
consequence vegetation can be diverted to
communities composed mainly of small-sized
species with high proportion of fine fuels, which
can provide a positive feedback on the likelihood
of fire recurrence (Zedler et al. 1983; Haidinger
and Keeley 1993; Lloret and Vilà 2003; Lloret et al.
2003; Eugenio and Lloret 2006; Baeza et al.
2007).
In this paper, our aim was to analyse
secondary succession in old-fields in eastern
Spain in the absence, and in the presence of fire.
Our starting hypothesis was that after
abandonment, vegetation would in the long-term
resemble a late-successional woodland
community composed of sclerophyllous broad-
leaved species. To test this hypothesis, we
compared the vegetation in unburned old-fields
that had been abandoned in the medium- and
distant-past (50 and 100 years ago respectively).
In addition, we assessed the impact of fire on old-
fields abandoned in the medium term (50 years
ago). For this we used plots affected by different
fire recurrences, and we tested two hypotheses,
that: (1) after a single fire, the vegetation would be
able to return to its pre-disturbance state, thus
supporting stand self-replacement as the most
common pathway in low fire recurrence regimes
where no significant deviations from the unburned
pathway would be expected; and (2) at high fire
recurrences the vegetation would be diverted to
possible alternative states, dominated by regime-
driven species.
2. Methods
2.1 Study area and experimental design
The study was conducted in the interior of Alicante
and Valencia provinces, eastern Spain, where the
sites experience a meso-Mediterranean climate
with an annual rainfall range between 450-600
mm and a mean annual temperature range
between 13-16ºC. The bedrock was marl and soils
were Regosols. The potential vegetation is mainly
a broad-leaved sclerophyllous forest of Q. ilex,
which, except for valley bottoms and other very
localised areas, is extremely scarce at present
due to past deforestation and exploitation. The
landscape is a mountainous mosaic with
agricultural lands and abandoned fields covered
mainly by regenerated forests and shrublands with
different degrees of development and
composition. The main species are the grass
Brachypodium retusum, the shrubs Rosmarinus
officinalis, Ulex parviflorus and Quercus coccifera,
and some pine forests dominated by Pinus
halepensis and P. pinaster.
Six sites containing old-field plant communities
on former arable land were selected on the basis
of their time since abandonment. The sites, which
had already been part of previous studies, were
selected to have similar soil and topographical
characteristics with the aim of reducing to the
minimum the effect of environmental variability on
vegetation regeneration (Baeza et al. 2002). Two
time ranges were available: (1) Long-term
abandonment (LT), with three sites abandoned
approximately 100 years ago, and (2) medium-
term abandonment (MT), with three sites
abandoned approximately 50 years ago. All of
these sites have been colonised by Pinus forest.
The time since abandonment was estimated
by means of interviews with landowners and
managers. To corroborate this information, we
inspected aerial photographs from 1956, 1978 and
2000. In the 1956 photographs the 100-year-old
sites had well-developed woodlands dominated by
Pinus sp., whereas the 50-year-old sites were
dominated by shrub and herb species and were
clearly at an earlier development stage. All the
selected sites were marginal fields located near
late-successional mature forests. Therefore, we
assumed that the propagule availability of late-
successional species dispersed by birds and
mammals was not limited.
In addition, within each of the three MT sites
there were unburned areas as well as areas which
had been subjected to different fire recurrences.
The first burn on each site was a wildfire that
occurred between 22 and 26 years ago
(abandonment and burning history summarised in
Table 1); this first fire covered an area of at least

98
Table 1 Site description and fire history. Experimental fires are marked with* in fire year. DIR = Direct Incident Radiation, HL = Heat Load.
Site name Latitude-Longitude Burning treatment
Time since abandonment
(years)
Fire year Altitude (m) Slope (º) Aspect DIR HL
La Venteta 38º39'N-0º35'W Unburned 100 No fire 1044 22 NE 0,74 0,71
Fontroja 38º40'N-0º30'W Unburned 100 No fire 834 32 NE 0,63 0,56
Els Plans 38º38'N-0º27'W Unburned 100 No fire 863 11 NW 0,88 0,94
38º39'N-0º39'W Unburned 50 No fire 940 47 ENE 0,51 0,34
1 fire 50 1984 940 35 NW 0,59 0,79
2 fires 50 1994* 940 35 NW 0,59 0,79
Onil
3 fires 50 2006* 940 35 NW 0,59 0,79
38º40'N-0º39'W Unburned 50 No fire 900 4 N 0,93 0,94
1 fire 50 1984 900 4 N 0,93 0,94
2 fires 50 1994* 900 4 N 0,93 0,94
Pardines
3 fires 50 2006* 900 4 N 0,93 0,94
39º07'N-0º57'W Unburned 50 No fire 763 10 SSW 1,02 1,03
1 fire 50 1979 1041 31 NNE 0,61 0,58
2 fires 50 1996 1041 23 NE 0,74 0,79
Ayora
3 fires 50 2006* 1041 23 NE 0,74 0,79
50 ha where, according to landowners and aerial
photographs, the sites were well-developed
mature Pinus forests at that time. Since then, sub-
areas of this burned area have been reburned,
either by subsequent experimental burns (Baeza
et al. 2002) or by other wildfires (e.g., the Ayora
site, Table 1). Within the area burned twice, a third
experimental burn was carried out in 2006.
Therefore, on each site we had at our disposal
four plots with similar characteristics but different
burning recurrences at distances of no more than
1km from each other. The medium-term
abandoned communities are denoted hereafter as
MTU (unburned), MT1 (burned once), MT2
(burned twice) and MT3 (burned thrice).
2.2 Vegetation sampling
The vegetation was sampled in 2006, except for
the sites burned thrice, which were sampled one
year after the fire in 2007. The cover for all
vascular species was estimated using the point-
intersect method (Greig-Smith 1983). Three
perpendicular 20 m long transects spaced 7 m
apart were evaluated on each plot across the
maximum slope. A metal rod was used to record
contact with individual species, and
measurements were taken every 20 cm along the
transect (100 points per transect). In addition, to
assess the success of late-successional species
establishment, the density of the main
sclerophyllous broad-leaved species (Q. ilex, Q.
coccifera, Olea europaea, Crataegus monogyna,
Juniperus oxycedrus and Rhamnus alaternus,
nomenclature follows Bolós et al. 1993) was
determined by counting the total number of
individuals within a 5m wide strip centred on the
transect line. We recorded all established
seedlings, saplings and trees as individuals.
At each plot, slope, aspect and elevation were
recorded, and the aspect values were used to
compute Direct Incident Radiation (DIR) and Heat
Load (HL) (McCune, 2007).
2.3 Data analysis
Species were classified according to their life-form
(Raunkiaer 1934). Because the assumption of
homogeneity of variances was not possible,
Kruskal-Wallis tests were used to test the
differences in relative cover of the dominant
species and life-forms between the different
treatments. We compared MTU cover against LT
cover to investigate species replacement during

99
the secondary succession, and also compared
MTU with the different burning histories to check
the effect of the different fire recurrences.
Similarly, one-way ANOVA was performed to find
differences in the total density of the main late-
successional species. When significant
differences were observed between communities
with different burning histories, post-hoc HSD
Tukey tests were performed (Zar 1996).
Ordination methods were then used to
evaluate compositional variations in relation to the
measured plot characteristics (fire recurrence
sequence, abandonment time, slope, elevation,
DIR, HL). Only species which were present in at
least two transects were included, i.e., 53 species
out of the 70 recorded. Species cover data were
transformed (loge(x+1)) and the downweighting
option for rare species was used. All multivariate
analyses were performed using the Vegan 1.9
package (Oksanen et al. 2007) in the R
environment (R Development Core Team 2007).
Initially, the vegetation data were analysed by
unconstrained ordination using Decorana (DCA),
and the significance of the relationship between
the first two axes and the plot characteristics
assessed using a permutation test (n=1000)
stratified by site. This analysis produced a
gradient length of 2.97 for the first axis, which
justified the use of the unimodal model (ter Braak
and Smilauer 1998). The relationship between
species composition and plot characteristics was
then investigated further using constrained
Canonical Correspondence Analysis (CCA). The
forward selection procedure in the Vegan package
(Oksanen et al. 2007) using the AIC statistic was
utilized to select the most significant
environmental variables. The significance of both
the model selected and the effects of the plot
characteristics axes was tested using a
permutation test (n=1000) with stratification by
site. The results for the DCA and the CCA were
similar, therefore only the CCA results are
reported here.
The relative impact of abandonment time and
fire recurrence in the vegetation dynamics was
assessed by plotting the relative positions of each
transect x fire recurrence/abandonment time
within the derived CCA model using two-
dimensional ellipses reflecting 95% confidence
intervals (Milligan et al. 2004). Thereafter, to
assess the relative impact of fire recurrence or
abandonment time in vegetation change at the
different sites, the following two-dimensional
Euclidean distances (Manly 1986; Mitchell et al.
2000) were calculated: (1) the distance between
the centroids of the MTU sites and the LT ones, to
assess the relative success in the replacement of
early-successional species by late-species in a
possible successional pathway; (2) the distance
between the centroids of the burned sites (MT1,
MT2 and MT3) in comparison with the MTU sites,
to assess the relative effect of the fire regime on
diverting the species composition from the
unburned community, and (3) the distance
between the centroids of MT2 and MT3 from
MT1, to assess the relative effects of increasing
fire recurrence.
3. Results
3.1 Species and life-form cover
Seventy species were detected, but the vegetation
was dominated by only eight species whose
combined cover exceeded 50% on all plots, and
80% on eight of the fifteen plots studied. Pinus
halepensis was the dominant Pinus species on all
sites except Ayora where P. pinaster was
dominant. The other dominant species were B.
retusum, Cistus albidus, Q. coccifera, Q. ilex, R.
officinalis and U. parviflorus. Due to their similar
life history traits, the two Pinus species were
regarded as the same taxon (Pinus sp.) in the
subsequent analysis in order to simplify the data
analysis and the interpretation of the results.
Macro-phanerophytes were dominant at both
abandonment times (MTU and LT). The cover of
R. officinalis decreased and the cover of both
Quercus sp. increased in the older (LT) fields.
Except for the rambling phanerophytes that
showed a slight increase with time, there was no
effect of time since abandonment on life-forms

100
Table 2 Relative cover of the seven most abundant species and life-forms on (a) old-fields at different abandonment times, and (b) areas of the medium-term abandoned old-fields which had been burned at different recurrences. Mean values (±SD, n=3) are presented; means in the same row that are significantly different (P<0,05 level, HSD Tukey test) are indicated with different letters.
A) Time since abandonment
50 years approx. 100 years approx.
Category
(MTU) (LT)
Significant response to time
since abandonment
Dominant species
Q. coccifera 3.7±2.9 b 11.9±9.2 a Increase
Q. ilex 0.1±0.2 b 17.6±2.4 a Increase
R. officinalis 13.4±10.8 a 0.8±1.5 b Decrease
B. retusum 18.8±14.4 15.6±4.8 None
C. albidus 0.1±0.2 0.9 ±0.8 None
Pinus sp. 47.4±11.9 34.5±5.6 None
U. parviflorus 0.9±0.9 0.8±1.3 None
Life-form
Rambling phanerophyte 0.3±0.6 b 5.5±3.4 a Increase
Macro-phanerophyte 55.4±19.1 66.5±12.1 None
Nano-phanerophyte 15.7±10.4 10±4.6 None
Chamaephyte 23.8±18.1 18±7 None
Hemi-cryptophyte 4.9±2 4.9±3.7 None
Terophyte 0.2±0.3 0 None
Geophyte 0 0.1±0.1 None
B) Burning treatment Significant response to
burning recurrence Unburned Burned x1 Burned x2 Burned x3
Category
(MTU) (MT1) (MT2) (MT3)
Dominant species
U. parviflorus 0.9±0.9 b 4.5±1.6 b 22 ±7.8 a 0.8±0.9 b Increase in x2
C. albidus 0.1±0.2 b 2.3±2.6 b 15.2±3.5 a 6±3.4 b Increase in x2
R. officinalis 13.4±10.8 b 48.6±7.8 a 3.6±3.6 c 0.4±0.7 c Increase in x1
Pinus sp. 47.4±11.9 a 4.2±6.3 b 0.4±0.7 b 0 b Decrease
B. retusum 18.8±14.4 19.9±11.1 33±11.3 47.2±.5 None
Q. coccifera 3.7±2.9 0.3±0.3 8±4.1 10±3.6 None
Q. ilex 0.1±0.2 0 0 0 None
Life-form
Nano-phanerophyte 19±7.8 b 60.9±10.4 a 52.7±10.6 a 17.6±9.9 b Increase in x1 and x 2
Hemi-cryptophyte 23.7±12.6 c 22.5±8 c 36.7±10.3 b 57.2±15.9 a Increase
Macro-phanerophyte 51.4±17.1 a 6.6±5.6 b 0.5±0.4 b 2.8±2.8 b Decrease
Chamaephyte 3.7±3.1 5.8±0.9 7.2±7.4 18.1±9.8 None
Terophyte 1.3±2.3 4±3.2 0.5±0.6 1.5±0.9 None
Geophyte 0 0 0.1±0.2 0.5±0.9 None
Rambling phanerophyte 0.3±0.6 0 0.1±0.2 0 None
(Table 2a). Burning had a much greater effect
(Table 2b). Pinus sp. had the greatest cover in the
unburned stands and they reduced by burning,
practically disappearing in the stands burned twice
and thrice. Moreover, in the stands burned once or
twice the vegetation became dominated by nano-
phanerophytes. In the once-burned treatment R.
officinalis was dominant, whereas in the twice-
burned treatment U. parviflorus and C. albidus
were dominant (Table 2b). Hemi-cryptophytes
increased in dominance with fire recurrence,
especially in thrice-burned stands. None of the
other treatments or life forms showed a significant
response to burning.

101
3.2 Late-successional species density
No significant differences in the total density of
late-successional species were found (p=0,579;
F=0,364, d.f =1) between MTU (Mean=40,5;
SD=28,3) and LT (Mean=40; SD=4,7) (Fig. 1).
However, some significant differences were found
between fire recurrence treatments. The density
was significantly higher in MTU (p=0,004;
F=10,309, d.f =3) compared to MT1 (Mean=8,8;
SD=5.7), MT2 (Mean=3,4; SD=1,4) and MT3
(Mean=5; SD=1,8). By species, Q. coccifera and
Q. ilex were the most abundant species in both
unburned communities. Rhamnus alaternus
showed a marked increase in the LT communities
compared to the MTU ones. In contrast, J.
oxycedrus decrease its density in LT compared to
MTU. All late-successional species strongly
decreased their density when they were affected
by fire. Density in MT1, MT2 and MT3
communities was markedly lower than in the MTU
ones (Fig. 1).
3.3 Multivariate analysis
The selection procedure within CCA included all 6
plot characteristics as significant (F=5,09;
p<0,005). The most important were fire recurrence
(F=11,11; p<0,001) and time since abandonment
(F=7,87; p<0,001), followed by DIR (F=3,73;
p<0,001), HL (F=3,01; p=0,002) elevation (F=2,93;
p=0,002) and slope (F=1,93; p=0,018). The
eigenvalues for the first four axes of the significant
CCA analysis were λ1=0,41, λ2=0,29, λ3=0,10 and
λ4=0,09. The total inertia was 2,31 and the first
two axes explained 70% of species-environment
variables.
The biplot relating species composition to the
environmental variables (Fig. 2a) showed that
late-successional species (e.g., Q. ilex, C.
monogyna and R. alaternus) and rambling
phanerophytes (e.g., Lonicera implexa and Rubia
peregrina) were associated positively with axis 1
and abandonment time. Pinus sp. was also
associated positively with axis 1 and age of
abandonment but less so than the
R. alaternus
MT3 MT2 MT1 MTU LT
Den
sity
(in
d. 1
00m
-2)
0
5
10
15
20
25
30
J. oxycedrus
MT3 MT2 MT1 MTU LT
Den
sity
(in
d. 1
00m
-2)
0
5
10
15
20
25
30
Q. ilex
MT3 MT2 MT1 MTU LT
Den
sity
(in
d. 1
00m
-2)
0
5
10
15
20
25
30
Q. coccifera
MT3 MT2 MT1 MTU LTD
ensi
ty (
ind.
100
m-2
)0
5
10
15
20
25
30
Others
MT3 MT2 MT1 MTU LT
Den
sity
(in
d. 1
00m
-2)
0
5
10
15
20
25
30
Figure. 1 Density of the main late-successional species in Long-term abandoned communities (LT), Medium-term abandoned unburned communities (MTU), Medium-term abandoned communities burned once (MT1), burned twice (MT2) and burned thrice (MT3). Mean values are presented. Error bars show standard error.

102
A)
B)
Figure. 2 Biplots derived from the CCA analysis of burned and successional stages in SE Spain. (a) Species biplot with significant environmental variables; all species are shown, but the 23 most abundant species are denoted with the larger symbols and with species codes. (b) Sites biplot with significant environmental variables, ellipses show 95% confidence intervals of communities with different abandonment and burning history; Symbols: ● = Long-term abandoned communities (LT), □ = Medium-term abandoned communities (MTU), = Medium-term abandoned communities burned once (MT1), ■ =Medium-term abandoned communities burned twice (MT2), ◊ =Medium-term abandoned communities burned thrice (MT3). Species code: Ahu= Atractylis humilis, Bfr= Bupleurum fruticosum, Bph= Brachypodium phoenicoides, Bre= Brachypodium retusum, Cal= Cistus albidus, Ccl= Cistus clusii, Dgn=Daphne gnidium, Dpe= Dorycnium pentaphyllum, Ech= Euphorbia characeas, Gsc= Genista scorpius, Hma= Helianthemum marifolium, Hfi= Helictotrichion filifolium, Jox= Juniperus oxycedrus, Lim= Lonicera implexa, Psp= Pinus sp., Qco= Quercus coccifera, Qil= Quercus ilex, Ral= Rhamnus alaternus, Rof= Rosmarinus officinalis, Rpe= Rubia peregrina, Sdu= Stahelina dubia, Sof= Stipa offneri, Upa= Ulex parviflorus.
late-successional species group (Q. ilex, C.
monogyna and R. alaternus). Pinus sp. was
associated with species that appear in the MTU,
e.g., Stahelina dubia and Bupleurum fruticosum.
The species associated with a single burn 22-26
years ago (MT1) had a negative score on axis 1

103
and this group included R. officinalis, Cistus clusii,
Helictotrichion filifolium and J. oxycedrus. A group
of species was highly correlated with high fire
recurrence, and this species group included, the
chamaephytes and hemi-cryptophytes:
Helianthemum marifolium, Atractylis humilis,
Euphorbia characeas, Dorycnium pentaphyllum,
and the nano-phanerophytes: C. albidus, Genista
scorpius and U. parviflorus.
The plot positions reflected a change in
species dominance and composition in long-time
secondary succession with a clear separation
between MTU and LT along axis 1 (Fig. 2b). The
effect of fire had a major influence on site position
on axis 2. MT1 was relatively close to MTU, but
with increasing fire recurrence (MT2, MT3) were
placed towards the negative end of both axes.
When figure 2b is further decomposed to show
the relative impact of fire recurrence or
abandonment time on vegetation change on the
three medium-term abandoned sites, subtle
differences become apparent (Fig. 3). The Onil
and Ayora MTU are closer to the LT than the
Pardines site (Table 3). With respect to fire
recurrence, two important site-specific effects
were noted. The first is that at the Pardines site,
the community with a single burn (MT1) was the
least diverted on species composition, it was
relatively close to the MTU. In contrast, the others
MT1 sites, of Onil and Ayora, were more different
to MTU (Table 3). A possible explanation for this
might be that the Pardines MTU community is less
well developed than the other sites; however, field
observations and data inspection (12.6% for
Pardines site, and 1.3% and 0 for Onil and Ayora
sites respectively in percentage of Pinus sp.
cover) showed that vegetation development of
MT1 at Pardines is moving towards regeneration
of the Pinus community from seeds released by
burned cones, whereas at the Onil and Ayora sites
the vegetation has become dominated by R.
officinalis, C. clusii and H. filifolium. Second, with
increasing fire recurrence, the vegetation has
been diverted to more negative values on both
axes of the CCA plot (Fig. 3), into the lower-left
quadrant associated with C. albidus, U.
parviflorus, D. pentaphyllum, G. scorpius, H.
cinereum, A. humilis, E. characeas, Daphne
gnidium and B. retusum. MT2 and MT3
communities of the three sites moved away from
the MTU and MT1. The site most affected by
increasing fire recurrence was Pardines. In
contrast, the distances between MT2 and MT3
were short in all three sites (Fig. 3; Table 3).
4. Discussion
4.1 Succession in Mediterranean old-fields
If undisturbed, successions on abandoned old-
fields developed along a predictable pathway,
involving the change of the dominant species. The
communities abandoned ca. 50 years ago were
dominated by P. halepensis or P. pinaster, which
were substituted in dominance on the plots
abandoned ca. 100 years ago by sclerophyllous
broad-leaved species, e.g., Q. ilex, C. monogyna
and R. alaternus. This late-successional
community resembled the expected late-
successional forest typical of this region (Quézel
2004). Our results confirm models proposed for
other Mediterranean P. halepensis forests, where
Pinus forests are replaced by Q. ilex forests in
mesic conditions where there has been an
absence of perturbations (Zavala et al. 2000;
Zavala 2003; Capitanio and Carcaillet 2008).
In the case of secondary succession on
abandoned fields, few propagules of forest
species are left on the site. Thus, various
successional mechanisms must be responsible for
this change in species abundance through time. In
an initial phase, species establishment will depend
on the ability to input propagules from the
surroundings (Platt and Connell 2003). Pinus
halepensis and P. pinaster are wind-dispersed
species with a high capacity to colonise open
spaces (Nathan et al. 2000), whereas late-
successional species are usually bird-dispersed
and their colonisation would be expected to be
more gradual in time (Bonet and Pausas 2004).
However, we found no differences in the density of
the main late-successional species established at

104
the two abandonment times. Previous
observations in other Mediterranean old-fields
(Debussche et al. 1982; Ne’eman and Izhaki
1996; Verdú and García-Fayos 1998; Bonet and
Pausas 2007) indicated that the density of bird-
dispersed species was surprisingly important in
the first years of abandonment. Recently
abandoned fields are often used by the European
jay (Garrulus glandarius) to cache acorns in SE
Spain (Pons and Pausas 2007). Therefore, in our
study, it is possible that most of the late-
successional individuals colonized early in the
succession, when Pinus was also colonising. They
then increased in their importance as Pinus
declined, essentially following the tolerance model
of Connell and Slatyer (1977). Late-successional
species have been characterised by their lower
growth, longer life-span and shade-tolerance with
regard to early-successional species (Verdú 2000;
Zavala et al. 2000). However, it is worth noting
that the facilitation model (Connell and Slatyer
1977) might also operate under Pinus forests in
some Mediterranean ecosystems where there is a
greater colonization and establishment of late-
successional (especially Quercus) species. This
improved seedling establishment has been
attributed to modified shade, moisture and
temperature conditions (Lookingbill and Zavala
2000; Gómez 2004; Pons and Pausas 2006,
2007).
Therefore, in this study which only compares
two ages, we were able to show the existence of a
change to a late-successional community
dominated by broad-leaved species.
Nevertheless, in our study the colonisation of the
main late-successional species took place mainly
in the first phase of succession. To investigate
which successional mechanism is more important,
facilitation or tolerance (sensu Connell and Slatyer
1977), further investigations are needed.
4.2 Failure of stand self-replacement after fire
When the secondary succession in the
communities abandoned 50 years ago was burned
by a single wildfire about 25-30 years after
Figure. 3 Biplots derived from the CCA analysis of burned and successional stages in the Valencia Region, separated to show the distribution of the burning recurrences of medium-term abandoned communities at each site in relation to the long-term abandoned communities. Ellipses show 95% confidence intervals of communities with different abandonment and burning history. Symbols: ● = Long-term abandoned communities (LT), □ = Medium-term abandoned communities unburned (MTU), = Medium-term abandoned communities burned once (MT1), ■ = burned twice (MT2) and ◊ = burned thrice (MT3)
abandonment, there was little evidence of stand
self-replacement after a further 22-27 years. The
burned Pinus forest was replaced by shrub
communities dominated mainly by R. officinalis.
Thus, in spite of the fact that P. halepensis and P.
pinaster are considered good regenerators after

105
fire (Vega 2003), their regeneration on our study
sites was poor or nil. This suggests that other
factors were responsible for this low regeneration.
Perhaps this low regeneration was because the
fires on our study plots were exceptionally severe,
there were unusual climatic conditions for the
establishment of seedlings at the time after
burning or other pre- or post-fire plot
characteristics. The variability in Pinus
regeneration success in eastern Spain has been
linked to: post-fire micro-environmental conditions
created by the amount of unburned branches lying
on the soil, plot topographic characteristics like
aspect and slope, and pre-fire tree density
(Pausas et al. 2004). However, to our knowledge,
in the Mediterranean Basin there are no studies
that clearly demonstrate the biotic and/or abiotic
factors involved in the null or massive
regeneration of Pinus sp. after fire.
Table 3 Euclidean distances between the centroids of medium-term abandoned (MTU) and long-term abandoned (LT) communities, and between the centroids of medium-term abandoned communities with different fire recurrences (MT1, MT2 and MT3) and unburned communities.
Euclidean distance Onil Pardines Ayora
MTU and LT 1,99 2,52 1,96 MTU and MT1 1,16 0,77 1,59 MTU and MT2 1,42 2,49 1,67 MTU and MT3 2,19 3,58 2,44 MT1 and MT2 1,22 1,80 1,95 MT1 and MT3 2,23 2,90 2,94 MT2 and MT3 1,01 1,1 0,98
Multivariate analysis identified the possible
deviation of the post-fire communities from the
unburned secondary successional pathway, and
measurement of the Euclidean distances between
the unburned and burned communities provided a
comparative quantitative measure of this
deviance. Our results showed that a large change
in species composition was brought about by a
single fire at the Onil and Ayora sites, suggesting
a deviation towards possible alternative
communities. Only one site (Pardines) showed
signs of recovery, with a trajectory towards the
unburned communities and with some Pinus
regeneration.
In addition, the density of the main late-
successional species was reduced considerably
compared to the unburned community. There are
several possible causes for this. For example,
although these species have the ability to survive
after fire and resprout from buds (Verdú 2000), it
is possible that if the fire was very severe they
would be killed. It is also possible that their
colonization might be restricted because of the
development of an arrested succession; this
process has been demonstrated in similar
Mediterranean shrub communities dominated by
pioneer species (Acacio et al. 2007; Siles et al.
2009). This process involved in restricting
colonization might include an unattractiveness to
birds, and hence a reduced seed dispersal, or a
reduction in success of seedling establishment
because of competition from the established
vegetation (Gómez 2004; Pons and Pausas 2006,
2007).
Thus, even a single fire can impact on old-field
succession in the Mediterranean region. It is
possible that the communities will recover, but it is
also possible that the successional trajectory will
be modified and an alternative state, dominated by
R. officinalis will be produced. Where R. officinalis
shrub communities develop it is likely that they will
remain in this state for a long time, i.e, essentially
(at least in human terms) an alternative stable
state.
4.3 High fire recurrences divert succession
Despite the fact that the different ages of the
burned communities (22-27 years for MT1, 10-12
years for MT2 and 1 year for MT3) to some
degree affect species composition, especially in
short-lived species, the Euclidean distances
between the communities could suggest
deviations in the successional pathways. Where
fires occurred at relatively short intervals, the
deviation from the unburned successional
pathway could increase. The Euclidean distances
were greater after the second and third fire

106
compared to either the unburned or single fire
communities. In addition, the short Euclidean
distances found between communities burnt twice
or thrice could suggest the establishment of a new
community with an auto-successional character
promoted by these fire regimes. This change
would also be accompanied by a changed
vegetation structure. Low stature species,
including many dwarf shrub, herbs and forbs, and
particularly those with rapid maturation and
shorter lived tissues, responded positively to the
short interval fires. These communities were
dominated by nano-phanerophytes (mainly U.
parviflorus and C. albidus), hemi-cryptophytes
(mainly B. retusum) and chamaephytes. This
change in community composition was enhanced
by the local extinction of some species as a
consequence of the recurrent fires. It is possible
that the recurrent fires exhausted the seed bank
and/or bud-bank of propagules; the fire interval is
too short and the species have not had time to
replenish these propagule stores between fires
(Zedler et al. 1983). In our study, the extinction of
Pinus was most relevant; P. halepensis needs 7-
10 years to reach the reproductive stage, and
about 15 years to produce a consistent canopy
seed bank (Eugenio et al. 2006; Eugenio and
Lloret 2006).
The altered species composition after
recurrent fires, with different spectra of life history
types, may lead to distinct post-fire assemblages
and alternative successional pathways compared
to regimes with a lower fire recurrence or no fire
(Noble and Slatyer 1980; Johnston and Chapin
2006; Donato et al. 2008). Furthermore, the re-
establishment of locally lost species will depend
on the input of propagules from undisturbed
zones, and re-establishment success will depend
on the size and severity of the fire, along with the
capacity of these species to colonise (Platt and
Connell 2003).
Our results agree with the conceptual model
proposed by Baeza et al. (2007), which suggested
that stable communities composed of C. albidus
and U. parviflorus would become established in
old-fields of SE Spain when the fire intervals were
less than 10 years. Hence, our work evidences
that old-field successions could behave like other
Mediterranean ecosystems; where fire recurrence
is high the dominance of small-size shrubs and
herbs is promoted (Zedler et al. 1983; Haidinger
and Keeley 1993; Lloret and Vilà 2003; Lloret et
al. 2003; Eugenio and Lloret 2006; Baeza et al.
2007).
5. Conclusions
Different successional pathways can exist in
abandoned old-fields of SE Spain depending on
fire occurrence and recurrence. In the absence of
fire, the vegetation in early-stages of succession is
dominated by pioneer tree species, mainly Pinus.
Nevertheless, this vegetation with the passage of
time and if the input of propagules from external
vegetation is not limited, will become dominated
by species of later-successional forests, such as
Quercus sp.
However, if the vegetation is burned during
these early-stages of succession, the succession
could be diverted. This deviation is accompanied
by a change in species composition and structure.
A single fire is enough to change Pinus forests
into alternative stable-states dominated by R.
officinalis, where the colonisation of species of
later successional stages may be prevented. This
deviation could increase in regimes with high fire
recurrence, enhancing the change in species
composition to low-size shrubs and herbs even
more.
Acknowledgements
We thank J. Scheiding for revision of the English
text and the Font Roja Natura-UA Scientific
Station for fieldwork support. We also thank the
two referees whose suggestions greatly improved
the previous drafts of this study. V.M. Santana
was supported by a FPU grant awarded by the
Spanish Ministry of Education and Science. This
research was carried out within the FIREMED
(AGL200/8-04522/FOR) and Consolider-Ingenio
2010 (GRACCIE CSD2007-00067) projects.

107
CEAM is supported by the Generalitat Valenciana
and Bancaixa.
References
Acacio V, Holmgren M, Jansen P, Schrotter O
(2007) Multiple recruitment limitation causes
arrested succession in Mediterranean oak
systems. Ecosystems 10: 1220-1230.
Baeza MJ, De Luís M, Raventós J, Escarré A
(2002) Factors influencing fire behaviour in
shrublands of different stand ages and the
implications for using prescribed burning to
reduce wildfire risk. J Environ Manage 65:
199-208.
Baeza MJ, Valdecantos A, Alloza JA, Vallejo VR
(2007) Human disturbance and environmental
factors as drivers of long-term post-fire
regeneration patterns in Mediterranean
forests. J Veg Sci 18: 243-252.
Barberó M, Loisel R, Quézel P (1992)
Biogeography, ecology and history of
Mediterranean Quercus ilex ecosystems.
Vegetatio 99-100: 19-34.
Blondel J, Aronson J (1999) Biology and Wildlife
of the Mediterranean Region. Oxford
University Press, New York
Bolòs O, Vigo J, Masalles RM, Ninot JM (1993)
Flora Manual dels Països Catalans. Pòrtic,
Barcelona
Bonet A, Pausas JG (2004) Species richness and
cover along a 60-year chronosequence in old-
fields of southeastern Spain. Plant Ecol 174:
257-270.
Bonet A, Pausas JG (2007) Old field dynamics on
the dry side of the Mediterranean Basin:
patterns and processes in semiarid Southeast
Spain. In: Cramer VA, Hobbs RJ (eds) Old
fields: dynamics and restoration of
abandoned farmland, Island press,
Washington, pp 247-264
Capitanio R, Carcaillet C (2008) Post-Fire
Mediterranean vegetation dynamics and
diversity: A discussion of succession models.
For Ecol Manage 255: 431-439.
Connell JH, Slatyer RO (1977) Mechanisms of
succession in natural communities and their
role in community stability and organization.
Am Nat 111: 1119-1144.
Cramer VA, Hobbs RJ, Standish RJ (2008) What's
new about old fields? Land abandonment and
ecosystem assembly. Trends Ecol Evol 23:
104-112.
Debussche M, Escarré J, Lepart J (1982)
Ornitichory and plant succession in
Mediterranean orchards. Vegetatio 48: 255-
66.
Debussche M, Escarré J, Lepart J, Houssard C,
Lavorel S (1996) Changes in Mediterranean
plant succession: old-fields revisited. J Veg
Sci 7: 519-526.
Donato DC, Fontaine JB, Robinson WD, Kauffman
JB, Law BE (2009) Vegetation response to a
short interval between high-severity wildfires
in a mixed-evergreen forest. J Ecol 97: 142-
154.
Eugenio M, Lloret F (2004) Fire recurrence effects
on the structure and composition on
Mediterranean Pinus halepensis communities
in Catalonia (northeast Iberian Peninsula).
Ecoscience 11: 446-454.
Eugenio M, Lloret F (2006) Effects of repeated
burning on Mediterranean communities of the
northeastern Iberian Peninsula. J Veg Sci 7:
755-764.
Eugenio M, Verkaik I, Lloret F, Espelta JM (2006)
Recruitment and growth decline in Pinus
halepensis populations after recurrent
wildfires in Catalonia (NE Iberian Peninsula).
For Ecol Manage 231: 47-54.
Fox MD, Fox BJ (1987) The role of fire in
scleromorphic forests and shrublands of
Eastern Australia. In: Trabaud L (ed) The role

108
of fire in ecological systems, SPB, the Hague,
pp 23-48
Friedel MH (1991) Range condition assessment
and the concept of thresholds: a viewpoint. J
Range Manage 44: 422-426
Gómez JM (2004) Importance of microhabitat and
acorn burial on Quercus ilex early
recruitment: non-additive effects on multiple
demographic processes. Plant Ecol 172: 287-
297.
Greig-Smith P (1983) Quantitative Plant Ecology.
Blackwell, Oxford
Grubb PJ (1998) A reassessment of the strategies
of plants which cope with shortages of
resources. Perspect Plant Ecol Evol Syst 1: 3-
31.
Haidinger TL, Keeley JE (1993) Role of high fire
frequency in destruction of mixed chaparral.
Madroño 40: 141-147
Hanes TL (1971) Succession after fire in the
chaparral of Southern California. Ecol
Monogr. 41: 27-52.
Johnstone J, Chapin F (2006) Fire interval effects
on successional trajectory in boreal forests of
Northwest Canada. Ecosystems 9: 268-277.
Keeley JE, Fotheringham CJ, Baer-Keeley M
(2005) Determinants of postfire recovery and
succession in Mediterranean-climate
shrublands of California. Ecol Appl 15: 1515-
1534.
Laycock WA (1991) Stable states and thresholds
of range conditions on North American
rangelands: a view point. J Range Manage
44: 427-433
Lloret F, Vilà M (2003) Diversity patterns of plant
functional types in relation to fire regime and
previous land use in Mediterranean
woodlands. J Veg Sci 14 : 387-398.
Lloret F, Pausas JG, Vilà M (2003) Responses of
Mediterranean plant species to different fire
frequencies in Garraf Natural Park (Catalonia,
Spain): field observations and modelling
predictions. Plant Ecol 167: 213-225.
Lookingbill TR, Zavala MA (2000) Spatial pattern
of Quercus ilex and Quercus pubescens
recruitment in Pinus halepensis dominated
woodlands. J Veg Sci 11: 607-612.
Manly BFJ (1986) Multivariate Statistical Methods.
Chapman and Hall, London
McCune B (2007) Improved estimates of incident
radiation and heat load using non-parametric
regression against topographic variables. J
Veg Sci 18: 751-754.
Milligan AL, Putwain PD, Cox ES, Ghorbani J, Le
Duc MG, Marrs RH (2004) Developing an
integrated land management strategy for the
restoration of moorland vegetation on Molinia
caerulea-dominated vegetation for
conservation purposes in upland Britain. Biol
Conserv 119: 371-387.
Mitchell RJ, Auld MHD, Le Duc MG, Marrs RH
(2000) Ecosystem stability and resilience: a
review of their relevance for the conservation
management of lowland heaths. Perspec Plant
Ecol Evol Syst 3: 142-160.
Nathan R, Safriel UN, Noy-Meir I, Schiller G (2000)
Spatiotemporal variation in seed dispersal and
recruitment near and far from Pinus halepensis
trees. Ecology 81: 2156-2169.
Ne’eman G, Izhaki I (1996) Colonization in an
abandoned East-Mediterranean vineyard. J
Veg Sci 7: 465-472.
Noble I, Slatyer RO (1980) The use of vital
attributes to predict successional changes in
plant communities subject to recurrent
disturbances. Vegetatio 43: 5-21.
Oksanen J, Kindt R, Legendre P, O’Hara B, Henry
M, Stevens H (2007) Vegan: Community
Ecology Package. R package version 1.8-7.
http://cran.r-project.org, http://cc.oulu.fi/fi/
~jarioksa/. Accessed 12 September 2007

109
Picket STA, Collins SL, Armesto JJ (1987) A
hierarchical consideration of causes and
mechanisms of succession. Vegetatio 69: 109-
114.
Pausas JG (2004) Changes in fire and climate in
the eastern Iberian Peninsula (Mediterranean
Basin). Clim Change 63: 337-350.
Pausas JG, Ribeiro E, Vallejo VR (2004) Post-fire
regeneration variability of Pinus halepensis in
the eastern Iberian Peninsula. For Ecol
Manage 203: 251-259.
Pons X, Pausas JG (2006) Oak regeneration in
heterogeneous landscapes: The case of
fragmented Quercus suber forests in the
eastern Iberian peninsula. For Ecol Manage
231: 196-204.
Pons X, Pausas JG (2007) Acorn dispersal
estimated by radio-tracking. Oecologia 153:
903-911.
Platt WJ, Connell JH (2003) Natural disturbances
and directional replacement of species. Ecol.
Monogr 73: 507-522.
Putz FE, Canham CD (1992) Mechanisms of
arrested succession in shrublands: root and
shoot competition between shrubs and tree
seedlings. For Ecol Manage 49: 267-75
Quézel P (2004) Large-scale post-glacial
distribution of vegetation structure in the
Mediterranean region. In: Mazzoleni S, Di
Pasquale G, Mulligan M, Di Martino P, Rego F
(eds) Recent dynamics of the Mediterranean
vegetation and landscape, J. Wiley & Sons,
Chichester, pp 3-12
R Development Core Team (2007) R: A language
and environment for statistical computing. R
Foundation for Statistical Computing, Vienna,
Austria. ISBN 3-900051-07-0, URL
http://www.R-project.org. Accessed 12
September 2007
Raunkiaer C (1934) The life form of plants and
statistical plant geography. Clarendon Press,
Oxford
Scheffer M, Carpenter S, Foley JA, Folke C,
Walker B (2001) Catastrophic shifts in
ecosystems. Nature 413: 591-6.
Siles G, Rey PJ, Alcántara JM, Ramírez JM
(2008) Assessing the long-term contribution of
nurse plants to restoration of Mediterranean
forests through Markovian models. J Appl Ecol
45: 1790-1798.
Tatoni T, Roche P (1994) Comparison of old-field
and forest revegetation dynamics in Provence.
J Veg Sci 5: 295-302.
Ter Braak CJF, Smilauer P (1998) CANOCO
reference manual user’s guide to Canoco for
Windows: Software for Canonical community
ordination (version 4.5), Micro-computer
Power, Ithaca, New York
Trabaud L (1991) Fire regimes and phytomass
growth dynamics in a Quercus coccifera
garrigue. J Veg Sci 2: 307-314.
Vega JA (2003) Regeneración del género Pinus
tras incendios. Cuadernos Sociedad Española
de Ciencias Forestales 15: 59-68
Verdú M (2000) Ecological and evolutionary
differences between Mediterranean seeders
and resprouters. J Veg Sci 11: 265-268.
Verdú M, García-Fayos P (1998) Old-field
colonization by Daphne gnidium: seedling
distribution and spatial dependence at
different scales. J Veg Sci 9: 713-718.
Zar JH (1996) Biostatistical analysis. Prentice
May, Upper Saddle River, New Jersey
Zavala MA (2003) Dinámica y sucesión en
bosques mediterráneos: modelos teóricos e
implicaciones para la selvicultura. In: Rey-
Benayas JM, Espigares T, Nicolau JM (eds)
Restauración de ecosistemas mediterráneos,
Universidad de Alcalá, Madrid, pp 43-63

110
Zavala MA, Espelta JM, Retana J (2000)
Constraints and trade-offs in Mediterranean
plant communities: The case of holm oak-
Aleppo pine forests. Bot Rev 66: 119-149.
Zedler PH, Gautier CR, McMaster GS (1983)
Vegetation change in response to extreme
events: the effect of a short interval between
fires in California chaparral and coastal scrub.
Ecology 64: 809-818.

111
Apéndice fotográfico
Foto 1. Campo abandonado 50 años atrás Foto 2. Campo abandonado 100 años atrás
Foto 3. Zona con 1 incendio dominado por R. officinalis Foto 4. Zona con 2 incendios dominado por Cistus y Ulex
Foto 5. Disposición de las parcelas de estudio Foto 6. Parcela quemada 3 veces con 1 año de edad
Foto 7. Parcela quemada 3 veces Foto 8. Colonización de Quercus coccifera

112

113
CAPÍTULO 7- EFECTO DEL RÉGIMEN DE TEMPERATURA DESPUÉS DEL FUEGO EN LA DORMANCIA Y GERMINACIÓN DE SEMILLAS DE SEIS ESPECIES DE FABACEAE AUSTRALIANAS RESUMEN: Además de efectos directos del fuego como el calor, el humo y la madera carbonizada, el paso del fuego conlleva indirectamente cambios en las condiciones ambientales capaces de romper la dormancia física de las especies con semillas de cubierta dura. Después de un fuego, la apertura de la cubierta vegetal y el material quemado tendido sobre la superficie alteran las propiedades térmicas del suelo, creando elevadas temperaturas del suelo durante largos periodos de tiempo. Nosotros simulamos los regímenes de temperatura diarios experimentados a diferentes profundidades del suelo después de un fuego de verano. Nuestro objetivo fue determinar si estos regímenes de temperaturas junto con la duración de la exposición (5, 15 y 30 días) desempeñan un papel importante rompiendo lo dormancia física en seis leguminosas del sureste de Australia. Nuestros resultados mostraron que las temperaturas simuladas rompen la dormancia de las semillas. Este efecto es especialmente pronunciado en temperaturas que son esperadas que ocurran cerca de la superficie del suelo (de 0 a 2 cm. de profundidad). El tiempo de exposición interactúa con la temperatura para romper la dormancia, con los mayores valores de germinación alcanzados después de las exposiciones más largas y temperaturas más altas. Sin embargo, la respuesta varió entre especies. Por lo tanto, este efecto indirecto del fuego podría jugar un rol importante en la regeneración de las comunidades vegetales, ya que podría estimular la emergencia de plántulas independientemente de los efectos directos del fuego así como en interacción con ellos. Este capítulo reproduce el siguiente manuscrito: Santana VM, Bradstock RA, Ooi MKJ, Denham AJ, Auld TD,Baeza MJ (2010) Effects of soil temperature regimes after fire on seed dormancy and germination in six Australian Fabaceae species. Australian Journal of Botany 58: 539-545

114

115
Effects of soil temperature regimes after fire on s eed dormancy
and germination in six Australian Fabaceae species
Victor M. SantanaA,, Ross A. BradstockB, Mark K. J. OoiC,D, Andrew J.
DenhamC, Tony D. AuldC, M. Jaime BaezaA,E
AFundación de la Generalitat Valenciana Centro de Estudios Ambientales del
Mediterráneo (CEAM). Parque Tecnológico Paterna. C/ Charles Darwin, 14.
46980 Valencia. Spain. BCentre for Environmental Risk Management of Bushfires, Institute for
Conservation Biology and Environmental Management, University of
Wollongong, NSW 2522, Australia. CDepartment of Environment, Climate Change and Water NSW, PO Box 1967,
Hurstville NSW 2220, Australia. DDepartment of Animal & Plant Sciences, University of Sheffield, Sheffield S10
2TN, UK. EDepartamento de Ecología, Universidad de Alicante. Ap 99. 03080 Alicante.
Spain
Abstract In addition to direct fire cues such as heat, smoke and charred wood, the passage of fire leads indirectly to changes in environmental conditions which may be able to break physical dormancy in hard-coated seeds. After a fire, the open canopy and the burnt material lying on the surface alter the thermal properties of the soil, resulting in elevated soil temperatures for long periods of time. We simulated daily temperature regimes experienced at different depths of soil profile after a summer fire. Our aim was to determine whether these temperature regimes and the duration of exposure (5, 15 and 30 days) play an important role breaking physical seed dormancy in six legumes from south-eastern Australia. Our results showed that simulated temperature regimes break seed dormancy. This effect is specially pronounced at temperatures that are expected to occur near the soil surface (0 to 2 cm depth). The duration of exposure interacts with temperature to break dormancy, with the highest germination rates reached after the longest duration and highest temperatures. However, the germination response varied among species. Therefore, this indirect post-fire cue could play a role in the regeneration of plant communities, and could stimulate seedling emergence independent of direct fire cues as well as in interaction with direct cues.

116
1. Introduction
A flush of seedling emergence occurs immediately
after fire in many fire-prone environments around
the world (Kruger and Bigalke 1984; Auld 1986;
Keeley 1991; Trabaud 1994; Carrington and
Keeley 1999). Heat and smoke released during
the passage of fire are considered to be the most
important fire cues that break dormancy or
promote germination in soil stored seeds.
Temperatures reached during the passage of fire
can break physical dormancy of hard-coated
seeds, allowing subsequent water imbibition and
germination when environmental conditions are
suitable (Keeley 1991; Auld and O’Connell 1991;
Cocks and Stock 1997; Bell 1999). In addition,
smoke can also affect the physiology of seeds and
directly stimulate germination (Brown 1993; Dixon
et al. 1995; Keeley and Fotheringham 1998; Van
Staden et al. 2000; Moreira et al. 2010) or act in
combination with heat (Keeley 1991; Keith 1997;
Morris 2000; Thomas et al. 2003). Other direct
fire-cues, such as charred wood, can also act to
stimulate seed germination in some species
(Keeley 1987). These factors all play a key role in
determining vegetation recovery after fire,
especially in ecosystems dominated by obligate
seeders.
Most management strategies used to control
fuel load and/or maintain biodiversity in fire-prone
ecosystems throughout the world are dependent
on a good understanding of the relationship
between direct fire cues and germination
(Bradstock and Auld 1995; Baeza and Roy 2008),
and for this reason, they have been widely studied
both in field and laboratory experiments. In
particular, the relationship between high but short-
term temperature conditions experienced by
seeds during fire have been tested, identifying
optimal and lethal temperature thresholds for a
range of species (e.g. Keeley 1987; Auld and
O’Connell 1991; Baeza and Vallejo 2006; Paula
and Pausas 2008 and references therein).
However, the passage of fire also leads
indirectly to changes in environmental conditions,
particularly those experienced by seeds on or
within the soil. After fire, the layer of black ash and
the partially burnt organic material lying on the soil
surface can alter the thermal properties of the soil
(Walker et al. 1986), particularly where an opening
in the canopy has occurred and increased solar
radiation reaches the soil surface. As a
consequence, a shift in the range of daily soil
temperatures may occur (Sharrow and Wright
1977; Raison et al. 1986), in some cases
exceeding the thresholds for breaking physical
seed dormancy (Auld and Bradstock 1996).
This indirect fire cue may acquire special
relevance after summer fires, when daily soil
temperatures reach high levels and fluctuate most
widely. These high temperatures can be sustained
for significant lengths of time (i.e. up to several
hours a day) in comparison to the high
temperatures induced by the fire itself, which only
remain for a few minutes or hours (Bradstock and
Auld 1995). Furthermore, regimes of high daily
temperatures may continue for several weeks
post-fire. In Mediterranean ecosystems, this
indirect fire cue may be quite important, ensuring
that a flush of germination in some hard-seeded
species occurs in the wet season after summer,
irrespective of the season of fire. This germination
strategy has been proposed as an adaptive trait,
as it avoids germination and subsequent seedling
establishment failures during the dry period
(Baeza and Roy 2008).
The role that daily soil temperature regimes
play as an indirect fire cue for breaking seed
dormancy has been scarcely studied. However,
the implications for population dynamic processes
in fire-prone regions are potentially significant.
Additionally, soil temperatures after fire are
strongly correlated with air temperatures (Auld and
Bradstock 1996; Ooi et al. 2009) and climate
change forecasts predict significant increases in
air temperatures over the next few decades
throughout the world (IPCC 2007). To both inform
management and help to predict the long-term
consequences of climate change, it is necessary
to link future environmental changes to

117
mechanisms that can control population
processes.
The aim of our work is to therefore test
whether regimes of daily soil temperatures,
experienced by seeds after the passage of fire,
play an important role in breaking seed dormancy.
An understanding of this will provide insight into
the potential impact that changing climatic
conditions will have on germination patterns
promoted by this indirect-fire cue. We simulated
this indirect fire cue in the laboratory and
examined its effect on germination in six
Australian Fabaceae species commonly found in
fire-prone vegetation in south eastern Australia.
More specifically, we asked two questions: (i) Can
the daily variations in temperatures that occur
post-fire, at different depths in the soil, break
physical dormancy in six different Fabaceae
species? and (ii) Does the amount of time seeds
are exposed to such regimes of temperature (in
terms of days) affect seed dormancy?
2. Materials and methods
The six study species are typical shrubs or
subshrubs from the Fabaceae family, a significant
understorey component of sclerophyll vegetation
in the Sydney region (Australia). These species
are characterized by having soil stored seed
banks and by having seeds with physical
dormancy which is broken by heat (Auld and
O’Connell 1991; Ooi 2007). The study species
used were Acacia suaveolens (Sm.) Willd.,
Bossiaea obcordata (Vent.) Druce, Bossiaea
rhombifolia Sieber ex DC., Dillwynia retorta (J.C.
Wendl.) Druce, Gompholobium grandiflorum Sm.
and Pultenaea ferruginea Rudge.
Seeds of the six study species were collected
from the Blue Mountains National Park (33º48’S,
150º35’E) at some 200 m elevation, near the
western outskirts of Sydney. Vegetation ranges
from open heath to open forest, with the
overstorey dominated by Eucalyptus/Corymbia
species. Soils are derived from Hawkesbury
sandstone. Average annual rainfall for the nearby
Glenbrook Bowling Club Meteorological station is
approximately 971 mm distributed throughout the
year, with a peak in summer. Average summer
temperatures (max/min) for Springwood Bowling
Club Meteorological station (some 7 km W of
Glenbrook) are 29/17ºC and average winter
temperatures 16/6ºC. Field collections were made
in summer during December of 2007. Several
hundred ripe fruits were collected from at least 30
plants in each population. Seeds were stored in
paper bags at laboratory temperatures (approx.
22ºC) until they were processed for treatment
applications in August 2008.
The direct effects of fire in breaking physical
dormancy of most of our study species have been
previously studied in laboratory experiments by
Auld and O’Connell (1991). The most important
factor breaking dormancy was temperature,
whereas the time of exposure had variable effects
(1-120 min). All species experienced their
maximum of germination (ca. 90%) after
treatments of 80-100ºC. Further increases in
temperature had deleterious effects on seed
viability. However, the threshold of temperature for
enhanced germination (cf. untreated seeds)
differed between species. Seed dormancy was
broken in G. grandiflorum after seeds were
exposed to 40ºC, whereas for the rest of our study
species seed dormancy was largely unaffected at
this temperature. Bossiaea obcordata, B.
rhombifolia and D. retorta had seed dormancy
broken from 60ºC. One population of A.
suaveolens had seed dormancy broken at 60ºC,
while a second did not respond until 80ºC. The
species P. ferruginea was not studied; however,
the response of seven species of the same genus
was variable, with four species having seed
dormancy broken at 40ºC and three species at
60ºC.
Our experiment was designed to test the
effects of regimes of post-fire daily temperature in
the soil on physical dormancy. We used soil
temperatures measured in the Sydney region after
a summer fire (Auld and Bradstock 1996) to
determine the range of temperatures to be
applied. Auld and Bradstock (1996) found that the

118
soil temperatures exceeded 40ºC in burnt areas
over summer, down to a depth of 4.5 cm, with the
highest temperatures of above 60ºC recorded
near the soil surface, at 0.4 cm depth. Thus,
temperatures reached after the passage of a fire
may in some cases exceed the thresholds for
breaking physical seed dormancy of the study
species (Auld and O’Connell 1991). In contrast,
soil temperatures after a winter fire or in unburned
vegetation during summer did not rise above
40ºC. Thus, we simulated daily temperature
regimes using approximate summer conditions
from three different depths down the soil profile.
Two incubators and two ovens were set up to
apply dry heat at 12h/12h maximum/minimum
temperature cycles. Three temperature ranges
were chosen: 40/18ºC, 50/18ºC, 60/18ºC. In order
to test the effect of exposure time, temperature
treatments were factorially combined with three
durations of exposure: 5, 15 and 30 days. A fourth
temperature range of 28/18ºC, simulating
unburned vegetation conditions during summer,
was set up as a control (Auld and Bradstock
1996). Temperatures within each chamber were
measured with a thermocouple and recorded
every 15 minutes with a data logger. Although we
attempted to achieve temperature regimes of
50/18ºC and 60/18ºC, due to technical difficulties
the regimes we achieved were 47/16ºC and
61/16ºC. While the temperature regimes that were
applied do not exactly mimic actual temperature
fluctuations within a soil profile (i.e. in the field
fluctuations of temperatures may occur throughout
the day and the exposure to maximum
temperatures can be variable depending on the
depth of soil) the treatments can provide insight
about the additive effect of post-fire temperatures
regimes.
The control and the 30 day treatment started
first, the 15 day treatment started 15 days later
and the 5 day treatment started 25 days after the
first one. This was done so that germination could
start simultaneously for all treatments with
identical elapsed time from the end of the pre-
treatments.
For each species, 60 seeds were divided into
three replicates and used for each combination of
temperature and duration during the experiment.
Seeds were placed on one layer of filter paper in 9
cm Petri dishes. For the 15 and 5 day treatments,
dishes were placed into the control incubator until
their treatment started. For two species the
number of treatments was limited by seed
availability. For B. rhombifolia only the 30 day
treatments were possible while for B. obcordata,
the only treatment possible was 47/16ºC for 30
days.
In order to estimate the potential maximum
germination of seeds of each species, three
additional replicates were established with no heat
treated seeds individually scarified using a scalpel.
For germination assessment, all seeds were kept
in two germination chambers at 25ºC day
temperature and 18ºC night temperature in
darkness. Seeds were checked every four days
for the first month, then once a week in the second
month and only once in the third month. Petri
dishes were watered with distilled water when
required. A seed with a 1 mm long radicle was
scored as a germinant and removed.
Proportional data (number of germinants as a
fraction of the total number of viable seeds per
dish) were analysed using a one-way ANOVA. We
used the results of the scalpel treatment to
estimate the number of viable seeds per dish.
Duncan’s post-hoc tests were used to detect any
pair-wise differences among treatments (α = 0.05)
for each species. The data were checked for
normality using the Kolmogorov-Smirnov test and
for homogeneity of variances by the Levene’s test
and arcsine transformed when necessary. We
used a two-way ANOVA to determine the
significance of the two fixed factors (temperature
fluctuation ranges and exposure time) on
germination percentage for each species. This
analysis was only possible for the four species
with the complete set of treatments.
To examine the variation in germination
between temperature treatments with regard to
increasing time of exposure, we applied a

119
regression to the germination data for each
temperature treatment (40/18ºC, 47/16ºC and
61/16ºC) against the numbers of days of exposure
(5, 15 and 30 days) for each species. Then, we
compared the slopes of the regressions using an
F-test. Only regressions with slopes significantly
different from zero were compared. Germination
values from the Control treatment were used as
common starting point (0 days of exposure) in all
regressions.
3. Results
The mechanical scarification treatment showed
that viability and potential maximum germinability
of seeds used in the experiment was very high
(96.7% for A. suaveolens, 100% for G.
grandiflorum, and 98.3% for D. retorta and P.
ferruginea). Only B. obcordata and B. rhombifolia
had lower values (83.3 and 86.7% respectively).
Table 1. Results from the two-way ANOVAs for the four species with complete experimental design
Variable d.f Mean
Square F P
A.suaveolens
Time 2 0.008 0.964 0.4
Temperature 2 0.023 3 0.075
Time*Temperature 4 0.01 1.29 0.312
Residual 18 0.008
D. retorta
Time 2 0.155 8.92 0.002
Temperature 2 0.175 10.11 0.001
Time*Temperature 4 0.026 1.53 0.236
Residual 18 0.017
G. grandiflorum
Time 2 0.115 6.37 0.008
Temperature 2 0.209 11.63 0.001
Time*Temperature 4 0.047 2.63 0.068
Residual 18 0.018
P. ferruginea
Time 2 0.223 17.735 <0.001
Temperature 2 0.319 25.301 <0.001
Time*Temperature 4 0.065 5.18 0.006
Residual 18 0.013
The response to heat treatments differed
depending on the species. Daily temperature
regimes and exposure duration influenced
germination response in all species, except for A.
suaveolens (Table 1, Fig.1). Neither of these
factors significantly affected germination in A.
suaveolens (Table 1), where germination values
were low for all treatments. The control treatment
reached 5% germination, whilst the maximum in
any treatment was 22.4% (Fig. 1).
The output from the one-way ANOVA showed
that germination in D. retorta was significantly
greater than the control (11.9% germinated) at
61/16ºC after 15 and 30 days, reaching 37.3 and
62.7% respectively, but was not influenced by
lower temperatures or 5 days exposure at 61/16ºC
(Fig. 1). The slope of the relationship between
germination and exposure duration at 61/16ºC
temperature regime was considerably greater than
one, suggesting that further exposure may further
increase germination (Table 2).
A similar pattern was apparent for G.
grandiflorum, with significantly greater germination
than the control treatment (23.3% germinated) at
47/18ºC-30 days treatment (50% germinated), and
at 61/16ºC after 15 and 30 days of exposure, with
values of 53.3 and 80% respectively (Fig. 1). The
regression slope for the 47/18ºC treatment was
less than one, while at 61/16ºC with the regression
slope was almost 2 times higher and comparable
to D. retorta at this temperature regime (Table 2).
There was a significant interaction between
temperature and exposure duration for P.
ferruginea (Table 1). For all treatments,
germination increased with exposure duration
(Fig. 1), but the greatest effect was found at
61/16ºC, with a regression slope approximately 5
times higher than at 40/18ºC and 47/18ºC (Table
2). One-way ANOVA showed that germination
was significantly enhanced over the control
(10.2% germinated) at 61/16ºC after 15 and 30
days of exposure, reaching values of 37.3 and
88.1% respectively (Fig. 1).
Although all treatments were not possible for
B. obcordata and B. rhombifolia, both species also
showed a trend of enhanced germination in
relation to increasing daily temperatures (Fig. 1).
One-way ANOVA for B. rhombifolia showed
significant differences for all temperature

120
D. retorta
0
20
40
60
80
100
P. ferruginea
0
20
40
60
80
100
B. obcordata
0
20
40
60
80
100
a
b
a
Control47/16ºC 28/18ºC
30
bc
b
bc
c c cbc
bc
bc
bc
c
bc bc
c
c cc
Control40/18ºC 47/16ºC 61/16ºC28/18ºC
5 15 30 5 15 30 5 15 30
Control40/18ºC 47/16ºC 61/16ºC28/18ºC
5 15 30 5 15 30 5 15 30
B. rhombifolia
Ger
min
atio
n (%
)
0
20
40
60
80
100
a
b
c
b
Control40/18ºC 47/16ºC 61/16ºC28/18ºC
303030
A. suaveolens
Ger
min
atio
n (%
)
0
20
40
60
80
100
Control40/18ºC 47/16ºC 61/16ºC28/18ºC
5 15 30 5 15 30 5 15 30
G. grandiflorum
Ger
min
atio
n (%
)
0
20
40
60
80
100a
d dd
bcb
bcdbcdcd bcd
Control40/18ºC 47/16ºC 61/16ºC28/18ºC
5 15 30 5 15 30 5 15 30
Figure 1. Effect of treatments simulating daily soil temperature regimes in summer burned stands upon the germination of some Australian legumes. Different lower case letters above columns indicate significant differences between treatments (Duncan post-hoc test, p<0.05). Error bars indicate standard error. Control= 28/18ºC treatment with 30 days of exposure, simulating soil temperature regime under unburnt vegetation. The numbers 5, 15 and 30 beneath columns indicate the different heat exposure periods (in days) used in the experiment.
treatments compared to the control, reaching
germination of 69.2% for the 61/16ºC-30 day
treatment, while B. obcordata showed no
significant increase in germination after exposure
to 47/16 ºC for 30 days.
4. Discussion
Soil temperature regimes after summer fires could
play a key role in breaking physical seed
dormancy, independently of temperatures
experienced during fire. We observed a significant
increase in germination for several legume
species after treatment at a range of temperatures
representative of soil conditions in open post-fire

121
Table 2. Results of linear regression approach between percent germination and duration of heat treatments (5, 15 and 30 days) for the different species and daily temperature regimes
Species Treatment Slope Intercept r2 F P n A. suaveolens
40/18ºC 0.01 7.55 0.01 0.006 0.940 12
47/16ºC -0.15 12.19 0.03 0.350 0.567 12
61/16ºC 0.31 11.61 0.14 1.672 0.225 12 D. retorta
40/18ºC 0.23 8.19 0.06 0.641 0.442 12
47/16ºC 0.7 8.59 0.22 2.81 0.125 12
61/16ºC 1.77 9.68 0.83 47.665 <0.001 12 G. grandiflorum
40/18ºC 0.27 29.13 0.08 0.875 0.372 12
47/16ºC 0.92 16.83 0.39 6.413 0.03 12
61/16ºC 1.79 26.75 0.86 61.449 <0.001 12 P. ferruginea
40/18ºC 0.46 8.55 0.37 5.982 0.035 12
47/16ºC 0.57 9.36 0.4 6.614 0.028 12
61/16ºC 2.51 8.87 0.84 53.98 <0.001 12
areas. This effect would be especially pronounced
on seeds present in shallow or sandy soil profiles,
where temperatures reach their widest ranges
(Auld and Bradstock 1996).
Although few studies have investigated the
effect of post-fire soil temperature regimes on
native Australian species, there is evidence from
other regions that have shown similar responses
by members of the Fabaceae. In European heath
in the Mediterranean, daily temperature cycles
occurring in vegetation gaps promoted
germination in the gorse U. parviflorus (Baeza and
Roy 2008). In temperate European ecosystems,
Van Assche et al. (2003) found that slight
seasonal changes in daily temperature
fluctuations were key to breaking physical
dormancy of many herbaceous legumes. Other
evidences have been highlighted from studies on
invasive species, such as the gap recruitment
displayed by the tropical shrub Mimosa pigra
(Lonsdale 1993) and the European gorse, U.
europaeus in New Zealand (Ivens 1983). Several
studies in agricultural systems found that the hard
seeds of clover, Trifolium subterraneum, softened
in response to daily temperature regimes between
30ºC and 60ºC, if treated for several weeks or
months (Hagon 1971; Taylor 1981).
Auld and O’Connell (1991) observed that
many leguminous species from south-eastern
Australia had their physical dormancy broken to
varying degrees by temperatures experienced
during fire. The most important factor breaking
dormancy was temperature, with a few species
reaching significant germination levels after
treatment at 40ºC and 60ºC, but most reaching
their maximum germination after treatment at 80-
100ºC. The duration of exposure did not
significantly change the effect on dormancy,
however, it should be noted that temperatures
maintained in the soil during fire are short and
exposure duration was tested over a scale of only
minutes (Bradstock and Auld 1995). In contrast,
daily temperature regimes over the threshold for
breaking dormancy can remain after a summer fire
for weeks or months (Raison 1986; Auld and
Bradstock 1996). Our work has shown that
duration of treatment (5 to 30 days) interacts with
temperature to break physical seed dormancy in
some species, with the highest germination levels
reached after the longest treatment durations in
some cases. The strength of this interaction
increased with increasing temperature ranges. Not
surprisingly, the germination response to heat
treatments varied between species. For example,
A. suaveolens, a species whose physical

122
dormancy is broken from 60-80ºC (Auld and
O’Connell 1991), was not influenced by any of the
temperature ranges or duration tested; it is
unlikely that seeds would experience longer
durations of exposure to the treatment
temperatures in a natural setting, so germination
and recruitment of this species may be more
tightly bound to direct fire cues. In contrast, for
other species with a lower threshold for breaking
physical dormancy (40-60ºC), such as G.
grandiflorum, D. retorta, P. ferruginea, B.
obcordata and B. rhombifolia (Auld and O’Connell
1991), germination has the potential to be
determined by both direct and indirect cues.
Our results suggest that indirect fire cues
could have more influence than expected on the
germination response of some physically dormant
species, especially after summer fires.
Additionally, the influence of post-fire temperature
regimes within the soil may behave in an additive
and/or synergistic way with the direct fire cues
heat and smoke in overcoming seed dormancy.
For example, low intensity fires may not provide
adequate heat to break dormancy in seeds, with
temperatures greater than 40ºC reached only in
the upper 2 cm of the soil profile, and
temperatures of 60-70ºC occurring for only a few
minutes at 1 cm depth (e.g., Auld 1986; Bradstock
and Auld 1995). However, significant post-fire
germination levels could still be reached if the litter
layer was consumed and daily soil temperature
regimes were enhanced. Other indirect fire cues
such as the removal of canopy vegetation could
increase both soil temperatures and the red:far-
red light ratio, which can also promote germination
in leguminous species (Baeza and Roy 2008). The
combination of these factors may explain the
higher than expected emergence of Acacia
seedlings observed after fires studied in south-
eastern Australia (Monk et al. 1981; Auld 1986;
Bradstock and Auld 1995). It is nevertheless true
that high intensity fires and the opening of litter
and canopy gaps are highly corrrelated (Bradstock
and Auld 1995; Whight and Bradstock 1999).
Thus, the rupture of physical dormancy both via
fire temperatures or via daily temperature regimes
after fires are probably conflated and further field
studies taking into consideration both effects are
needed to put our experimental findings into
context. This mechanism, in addition, could play a
key role in inter-fire recruitment, promoting shrub
regeneration in gaps opened in the canopy
vegetation. In fact, other studies in fire prone-
ecosystems have contrasted these cues on
seedling establishment by comparing cleared with
burned plots, and observed, for example, that in
California chaparral germination was more tied to
direct effects of fire (Tyler 1995) whereas in other
Mediterranean Basin shrublands indirect effects
may increase their significance (Baeza and Roy
2008; Santana, unpublished data).
Enhanced germination resulting from summer
daily temperature regimes could be considered
adaptive for many physically dormant species in
Mediterranean fire-prone vegetation. This may
ensure that a flush of germination occurs
predominantly in autumn, independently of fire
season, avoiding germination during the summer
drought (Trabaud 1994; Bell 1999; Baeza and Roy
2009). While a strong seasonal pattern of rainfall
does not occur in south-eastern Australia, time
periods with adequate soil moisture to allow
seedling germination and emergence are much
more common in the cooler seasons (Bradstock
and Bedward 1992). Seeds with released physical
dormancy germinate, independently of season, as
soon as moisture conditions are suitable
(Hodgkinson 1991; Bell 1999). Therefore, there is
also the potential for an adaptive advantage in
these non-seasonal rainfall habitats. Probably,
advantages of this mechanism on these habitats
could be determined by the spreading germination
over time. Rupture of dormancy several weeks or
months after fire could be an advantage avoiding
unsuitable conditions in the immediate post-fire
period which could limit the success of seedling
establishment or survival (Frazer and Davis 1988;
Carrington 1999; De Luis et al. 2005). Hodgkinson
(1991) found in semiarid woodland with no
seasonal rainfall pattern in inner south-eastern

123
Australia higher germination and survival rates for
leguminous species regenerated after spring and
summer fires than in winter fires.
The rupture of seed dormancy by the soil
temperature regime could have implications on
seeds and seed bank dynamics within the
framework of predicted impacts of climate change,
where significant increases in mean air
temperature are forecast for the latter half of the
21st century (IPCC 2007; CSIRO 2007). In south-
eastern Australia, Auld and Bradstock (1996)
found that daily soil temperatures were
significantly related to air temperature at all soil
depths tested after a summer fire. In addition, Ooi
et al. (2009) found a relationship between
maximum air temperature and soil temperature in
bare soils in arid environments, where an air
temperature increase of 4ºC resulted in an
increase of approximately 10ºC in soil
temperature. Predicted increases in temperatures
may therefore promote germination in soil seed
banks that otherwise would persist ungerminated
after fire. Persistent seed banks play a
fundamental role minimising the risk of decline or
local extinction in plants for the cases where the
fire-free intervals are less than the primary juvenile
periods of the species (Auld and Denham 2006).
Acknowledgements
We thank Fiona Thomson for providing seeds for
this experiment. V.M. Santana was supported by a
FPU grant awarded by the Spanish Ministry of
Education and Science and by the Consolider-
Ingenio 2010 (GRACCIE CSD2007-00067) and
FIREMED (AGL2008-04522/FOR) projects. CEAM
is supported by the Generalitat Valenciana and
Fundación Bancaja.
References
Auld TD, O’Connell MA (1991) Predicting patterns
of post-fire germination in 35 eastern
Australian Fabaceae. Australian Journal of
Ecology 16, 53-70
Auld TD, Bradstock RA (1996) Soil temperatures
after the passage of a fire: Do they
influence the germination of buried
seeds? Australian Journal of Ecology 21, 106-
109
Auld TD (1986) Population dynamics of the shrub
Acacia suaveolens (Sm.) Willd.: fire and the
transition to seedlings. Australian Journal of
Ecology 11, 373-385
Auld TD, Denham AJ (2006) How much seed
remains in the soil after a fire? Plant Ecology
187, 15-24
Baeza MJ, Vallejo VR (2006). Ecological
mechanisms involved in dormancy breakage
in Ulex parviflorus seeds. Plant Ecology 183,
191-205
Baeza MJ, Roy J (2008) Germination of an
obligate seeder (Ulex parviflorus) and
consequences for wildfire management.
Forest Ecology and Management 256, 685-
693
Bell DT (1999) Turner Review No. 1. The process
of germination in Australian species.
Australian Journal of Botany 47, 475-517
Bradstock RA, Auld TD (1995) Soil temperatures
during experimental bushfires in relation to
fire intensity: consequences for legume
germination and fire management in south-
eastern Australia. Journal of Applied Ecology
32, 76-84.
Bradstock RA, Bedward M (1992) Simulation of
the effect of season of fire on post-fire
seedling emergence of two Banksia species
based on long-term rainfall records. Australian
Journal of Botany 40, 75-88
Brown NAC (1993) Promotion of germination of
fynbos seeds by plant-derived smoke. New
Phytologist 123, 575-583
Carrington ME (1999) Post-fire seedling
establishment in Florida sand pine scrub.
Journal of Vegetation Science 10, 403-412
Carrington ME, Keeley JE (1999) Comparison of
post-fire seedling establishment between

124
scrub communities in mediterranean and non-
mediterranean climate ecosystems. Journal of
Ecology 87, 1025-1036
Cocks MP, Stock WD (1997) Heat stimulated
germination in relation to seed characteristics
in fynbos legumes of the Western Cape
Province, South Africa. South African Journal
of Botany 63, 129-132.
CSIRO (2007) Climate change in Australia.:
Technical Report 2007. CSIRO, Australia
De Luis M, Raventos J, Gonzalez-Hidalgo JC
(2005) Fire and torrential rainfall: effects on
seedling establishment in Mediterranean
gorse shrublands. International Journal of
Wildland Fire 14, 413-422
Dixon KW, Roche S, Pate JS (1995) The
promotive effect of smoke derived from burnt
native vegetation on seed germination of
Western Australian plants. Oecologia 101,
185-192
Frazer JM, Davis SD (1988) Differential survival of
chaparral seedlings during the first summer
drought after wildfire. Oecologia 76, 215-21
Hagon MW (1971) The action of temperature
fluctuations on hard seeds of subterranean
clover. Australian Journal of Experimental
Agriculture 11, 440-443
Hodgkinson K.C. (1991) Shrub recruitment
response to intensity and season of fire in a
semi-arid woodland. Journal of Applied
Ecology 28, 60-70.
IPCC (2007) Climate Change 2007 Synthesis
Report. An assessment of the
Intergovernmental Panel on Climate Change,
Geneva.
Ivens GW (1983) The influence of temperature on
germination of gorse (Ulex europaeus L.).
Weed Research 23, 207-216
Keeley JE (1991) Seed germination and life
history syndromes in the California chaparral.
The Botanical Review 57, 81-116
Keeley JE (1987) Role of fire in seed germination
of woody taxa in California chaparral. Ecology
68, 434-443
Keeley JE, Fotheringham CJ (1998) Smoke-
induced seed germination in California
chaparral. Ecology 79, 2320-2336.
Keith DA (1997) Combined effects of heat shock,
smoke and darkness on germination of
Epacris stuartii Stapf., an endangered fire-
prone Australian shrub. Oecologia 112, 340-
344
Kruger FJ, Bigalke RC (1984) Fire in fynbos. In
“Ecological effects of fire in south African
ecosystems” (Eds P de Van Booysen, NM
Tainton) pp. 69-94 (Springer-Verlag, Berlin)
Lonsdale WM (1993) Losses from the seed bank
of Mimosa pigra: soil micro-organisms vs.
temperature fluctuations. Journal of Applied
Ecology 30, 654-660.
Monk D, Pate JS, Loneragan WA (1981) Biology
of Acacia pulchella R.Br. with special
reference to symbiotic nitrogen fixation.
Australian Journal of Botany 29, 579-592
Moreira B, Tormo J, Estrelles E, Pausas JG
(2010). Disentangling the role of heat and
smoke as germination cues in Mediterranean
Basin flora. Annals of Botany 105, 627-635
Morris EC (2000) Germination response of seven
east Australian Grevillea species
(Proteaceae) to smoke, heat exposure and
scarification. Australian Journal of Botany 48,
179-189
Ooi MKJ (2007) Dormancy classification and
potential dormancy-breaking cues for shrub
species from fire-prone south-eastern
Australia. In “Seeds: Biology, Development
and Ecology” (Eds SW Adkins, S Ashmore,
SC Navie) pp. 205-216 (C.A.B. International,
Wallingford, UK)
Ooi MKJ, Auld TD, Denham AJ (2009) Climate
change and bet-hedging: interactions

125
between increased soil temperatures and
seed bank persistence. Global Change
Biology 15, 2375-2386
Paula S, Pausas JG (2008) Burning seeds:
Germinative response to heat treatments in
relation to resprouting ability. Journal of
Ecology 96, 543 - 552
Raison RJ, Woods PV, Jakobsen BF, Bary GAV
(1986) Soil temperatures during and following
low-intensity prescribed burning in a
Eucalyptus pauciflora forest. Australian
Journal of Soil Research 24, 33-47
Sharrow SH, Wright HA (1977) Effects of fire, ash,
and litter on soil nitrate, temperature, moisture
and tobosagrass production in the Rolling
Plains. Journal of Range Management 30,
266-270.
Taylor GB (1981) Effect of constant temperature
treatments followed by fluctuating
temperatures on the softening of hard seeds
of Trifolium subterraneum L. Australian
Journal of Plant Physiology 8, 547-558
Thomas PB, Morris EC, Auld TD (2003)
Interactive effects of heat shock and smoke
on germination of nine species forming soil
seed banks within the Sydney region. Austral
Ecology 28, 674-683
Trabaud L (1994) Postfire plant community
dynamics in the Mediterranean basin. In “The
Role of Fire in Mediterranean-type
Ecosystems” (Eds JM Moreno, WC Oechel)
pp. 1-15 (Springer-Verlag, New York, USA).
Tyler CM (1995) Factors contributing to postfire
seedling establishment in chaparral: direct
and indirect effects of fire. Journal of Ecology
83, 1009-1020.
Van Assche JA, Debucquoy KLA, Rommens WAF
(2003) Seasonal cycles in the germination
capacity of buried seeds of some
Leguminoseae (Fabaceae). New Phytologist
158, 315-323
Van Staden J, Brown NAC, Jäger AK, Jonson TA
(2000) Smoke as germination cue. Plant
Species Biology 15, 167-178
Walker J, Raison RJ, Khanna PK (1986) Fire. In
“Australian soils: the human impact” (Eds JS
Russell, RF Isbell) pp. 185-216 (University of
Queensland Press, Brisbane)
Whight S, Bradstock RA (1999) Indices of fire
characteristics in sandstone heath near
Sydney, Australia. International Journal of
Wildland Fire 9, 145-153

126
Apéndice fotográfico
Foto 1. Semillas de Acacia suaveolens Foto 2. Semillas de Dillwynia retorta
Foto 3. Semillas de Gompholobium gradiflorum Foto 4. Semillas de Pultenaea ferruginea
Foto 5. Semillas de Bossiaea rhombifolia Foto 6. Semillas de Bossiaea obcordata
Foto 7. Semillas germinadas Foto 8. Especies leguminosas en su hábitat

127
CAPÍTULO 8- DISCUSIÓN GENERAL

128

129
CAPÍTULO 8 - DISCUSIÓN GENERAL
- Regeneración de especies germinadoras obligadas
Sin duda, el paso del fuego promueve la germinación y establecimiento de las
especies germinadoras obligadas estudiadas. Las altas temperaturas, el humo
o los nutrientes liberados podrían estimular la geminación y supervivencia de
nuevas plántulas (Capítulo 4). Sin embargo, este alto establecimiento podría no
ser consecuencia exclusiva de los efectos directos del fuego y otros efectos
indirectos, como el incremento de la fluctuación diaria del régimen de
temperaturas del suelo, pueden ser determinantes en la ruptura de la
dormancia física de las semillas. De hecho, se ha observado que, en especies
de leguminosas australianas, el régimen de temperaturas del suelo después de
un fuego de verano puede ser suficiente para romper la dormancia física de
semillas con cubiertas duras (Capítulo 7). Por lo tanto, la determinación de la
importancia relativa de cada uno de estos efectos (directos o indirectos) en la
ruptura de la dormancia puede ser clave a la hora de clarificar los procesos de
regeneración de estas especies. Sin embargo, destacar la dificultad de
determinar estos efectos, ya que pueden estar altamente correlacionados entre
si (por ejemplo las altas temperaturas del fuego y el grado de consumo de
biomasa aérea que determinará la fluctuación de temperaturas del suelo). En el
marco de la presente tesis se ha realizado un estudio comparativo entre estos
efectos en las especies principales de estudio (Cistus albidus, Rosmarinus
officinalis, Ulex parviflorus), sin embargo, no ha sido incluido en la memoria
final. A grandes rasgos, cabría resaltar que se ha encontrado un peso
importante de los efectos indirectos.
No obstante, el reclutamiento de estas especies no está restringido a
situaciones post-incendio, poniendo en entredicho la necesidad de los efectos
directos del fuego en la regeneración de estas especies. Ulex parviflorus o R.
officinalis, son capaces de establecerse con cierto éxito en periodos
sucesionales entre fuegos, especialmente cuando existen espacios abierto de
suelo y vegetación y/o la densidad de individuos adultos no es muy alta
(Capítulo 4). El incremento de la fluctuación diaria de las temperaturas del
suelo a consecuencia de la radiación incidente puede ser un factor ambiental

130
determinante en la ruptura física de la dormancia de especies con cubierta
dura. Este reclutamiento se ve restringido en etapas sucesionales maduras,
cuando los espacios abiertos son cerrados por la vegetación y la materia
orgánica acumulada en el suelo. Probablemente, los individuos adultos pueden
también excluir competitivamente el establecimiento de las plántulas. Por lo
tanto, los resultados sugieren que las especies germinadoras obligadas
estudiadas utilizan una estrategia oportunista; es decir, podrían germinan y
establecerse en sitios abiertos o suelos perturbados después de un fuego o de
cualquier otro tipo de perturbación (Ackerly 2004). Esta estrategia contrasta con
la mayoría de las especies de germinadoras obligadas de otras regiones
mediterráneas, donde la germinación y el establecimiento depende
completamente de de los efectos directos del fuego y se produce casi
exclusivamente en el primer año post-incendio (Pierce y Cowling 1991, Bell et
al. 1993, Keeley 1992, 1995).
Por otro lado, existen diferencias en los micro-hábitats preferenciales
para la germinación y establecimiento de las especies estudiadas, sugiriendo
diferencias en los nichos de regeneración (Capítulo 4). Destacar las diferencias
entre especies con diferentes características funcionales en sus semillas
(cubierta dura y blanda). Por ejemplo, las especies con cubiertas duras (C.
albidus y U. parviflorus) ven estimulada su germinación por valores intermedios
de severidad y, además, germinan en espacios abiertos de la cubierta vegetal.
En contra, R. officinalis con cubierta blanda ve limitado su germinación en
valores altos de severidad y, en periodos entre-fuegos, su germinación esta
ligada a micro-hábitats cercanos a individuos adultos. Por lo tanto, son
necesarios nuevos estudios que establezcan los patrones de coexistencia entre
especies y los factores que la determinan. De hecho, estas diferencias implican
que la oportunidad de establecimiento y coexistencia de las especies puede
variar en el tiempo a causa de las cambiantes condiciones ambientales a lo
largo de la sucesión.
- Trayectorias sucesionales en relación al régimen de recurrencia de incendios
Los ecosistemas dominados por especies germinadoras obligadas no son
estables dentro del régimen de alta recurrencia de incendio estudiado; es decir,

131
con el paso del tiempo no recuperan la composición y estructura previa al fuego
(Hanes 1971, Trabaud y Lepart 1980). Esta baja estabilidad se establece tanto
para los sistemas de pinar (dominados por Pinus halepensis) como de matorral
(dominados por C. albidus, R. officinalis y U. parviflorus) (Capítulo 5 y Capítulo
6). Incluso, la regeneración de los pinares puede verse comprometida por un
simple incendio (Capítulo 6), a pesar de que P. halepensis ha sido considerada
tradicionalmente como una especie altamente resiliente al fuego (Vega 2003).
Por lo tanto, son necesarios nuevos estudios que determinen las razones de la
baja regeneración obtenida en nuestra zona de estudio, ya que esta
regeneración puede ser determinante en la trayectoria sucesional que siga el
ecosistema.
Los campos de cultivo abandonado pueden albergar diferentes
trayectorias sucesionales dependiendo del régimen de recurrencia. En
ausencia de fuego, son dominados por sistemas de pinar (P. halepensis o P.
pinaster) en una primera etapa sucesional. Sin embargo, con el paso del
tiempo pueden evolucionar hacia un bosque mixto con presencia de especies
rebrotadoras (Quercus ilex). En contra, tras un solo incendio los sistemas de
pinar pueden pasar a estar dominados por especies de matorral. Tras una
primera etapa sucesional dominada por U. parviflorus y C. albidus, el matorral
podría alcanzar estados estables dominados por R. officinalis (Capítulo 6). De
hecho, en esta etapa sucesional más tardía se ha observado una
establecimiento prácticamente nulo tanto de las especies germinadoras
obligadas (C. albidus, R. officinalis, U. parviflorus; Capítulo 4) como de las
especies rebrotadoras (Capítulo 6). Los resultados sugieren que las diferentes
especies germinadoras obligadas tienen características en sus atributos vitales
las hacen más competitivas que otras a lo largo del gradiente sucesional. La
regeneración de C. albidus está prácticamente restringida a periodos post-
incendio, mientras que R. officinalis y U. parviflorus pueden establecerse en
períodos entre fuegos (Capítulo 4). Además, diferencias en el crecimiento
(tanto de la parte aérea como de la subterránea) y la longevidad de vida
podrían ser claves en los procesos de sustitución de las especies de matorral a
lo largo de la sucesión. Cabe destacar que un detallado estudio de estos
factores (longevidad de vida, crecimiento, producción de combustible muerto,
producción de semillas) se ha realizado para estas tres especies a lo largo de

132
una cronosecuencia de edades dentro del marco de la presente tesis; sin
embargo, este estudio no ha sido incluido en la memoria final. En general, se
podría sugerir que el mecanismo que rige los procesos sucesionales en
matorrales dominados por especies germinadoras obligadas es el de tolerancia
(sensu Connell y Slatyer 1977; Capítulo 4), y que además, el desarrollo de
estos matorrales podrían inhibir la entrada de especies de etapas sucesionales
más tardías como rebrotadoras (Capítulo 6). Sin embargo, Siles et al. (2008)
observaron que mientras bajo U. parviflorus la entrada de especies
rebrotadoras se veía inhibida, bajo R. officinalis se veía facilitada. Por lo tanto,
nuevos estudios que determinen si la sustitución de especies a lo largo de la
sucesión (U. parviflorus > R. officinalis) puede establecer cambios en la
probabilidad de de establecimiento de las especies rebrotadoras son
necesarios. Este hecho puede alcanzar especial relevancia en la restauración
de este tipo de ecosistemas en la Comunitat Valenciana, donde se pretenden
promocionar trayectorias sucesionales que conduzcan hacia estados
dominados por especies rebrotadoras (Valdecantos et al. 2009), con una mayor
resiliencia al fuego y una menor acumulación de combustible fino muerto
(Baeza et al. en prensa). Por otro lado, el escaso establecimiento de nuevos
individuos de especies rebrotadoras en los sistemas de romeral (R. officinalis),
así como de una propia autoregeneración, podría sugerir un colapso del
ecosistema en futuras etapas sucesionales. Por lo tanto, serían interesantes
nuevos estudios que indaguen en esta posibilidad, que podría llevar asociados
ecosistemas senescentes con procesos de sucesión regresiva (sensu Walker y
Reddell 2007).
Bajo una alta recurrencia de incendio (2-3 fuegos), los ecosistemas de
matorral sufren modificaciones en los patrones de abundancia de las especies
que los componen. La especie dominante en la primera etapa sucesional, U.
parviflorus, no sólo ve reducida su abundancia, sino que también ve retrasado
su óptimo en el tiempo. Rosmarinus officinalis sufre un ligero descenso en su
abundancia, mientras que P. halepensis ve prácticamente eliminada su
presencia (Capítulo 5). Probablemente, el intervalo de tiempo entre incendios
es demasiado corto para que las especies alcancen su madurez reproductiva
y/o rellenen satisfactoriamente sus bancos de semillas (Capítulo 5 y Capítulo
6). De hecho, dos fuegos consecutivos en periodos de tiempo menores a 10-15

133
años son suficientes para eliminar a P. halepensis, ya que este intervalo de
tiempo es demasiado corto para que la especie alcance su madurez
reproductiva (Eugenio y Lloret 2006). Por el contrario, especies de tipo
caméfitos o hemicriptófitos, de pequeño tamaño, con una rápida maduración
y/o capacidad de rebrote se pueden ver favorecidos por el régimen de alta
recurrencia de incendios. Se ha observado que especies de matorral que
producen un gran cantidad de semillas, como C. albidus, necesitan ambientes
perturbados para regenerarse satisfactoriamente y, además, en ambientes de
alta recurrencia aumenta su abundancia (Capítulo 4 y Capítulo 5). Los
resultados sugieren una transición hacia sistemas con una mayor presencia de
especies de matorral de bajo porte, caméfitos y herbáceas, especialmente de
Brachypodium retusum, que pueden establecer un vínculo con la ocurrencia de
incendio. En este tipo de ecosistemas, dominado por especies germinadoras
obligadas, se antoja imprescindible el estudio de la evolución del banco de
semillas de suelo de las diferentes especies y bajo diferente régimen de
recurrencia de incendios. Este estudio, a pesar de que se ha realizado a través
de la extracción de muestras de suelo y su posterior determinación de forma
indirecta (germinación en invernadero), no ha sido incluido en la presente tesis
doctoral.
- Dinámica y efectos de los combustibles en el ecosistema
A nivel de combustible, se ha observado una especial relevancia de la cantidad
de biomasa muerta acumulada en la estructura de los individuos a la hora de
determinar los efectos en el ecosistema (combustión de biomasa y
temperaturas del suelo; Capítulo 3). La especie que mas acumuló este tipo de
combustible, U. parviflorus, experimentó los mayores valores de combustión y
temperatura del suelo, mientras que la especie con menor combustible muerto,
R. officinalis, obtuvo los valores más bajos (Capítulo 3). Este hecho se ve
reflejado en la función de inflamabilidad descrita para este tipo de matorrales
donde la sustitución de U. parviflorus por R. officinalis a lo largo de la sucesión
conlleva una un función de forma jorobada; es decir, tras un incremento inicial,
con el paso del tiempo, la cantidad de combustible muerto se ve reducida
(Capítulo 5). Por lo tanto, estos resultados sugieren una varibilidad en los

134
posibles efectos del fuego sobre el ecosistema dependiendo de la etapa
sucesional. Sin embargo, nuevos estudios que demuestren esta hipótesis
empíricamente son necesarios. A nivel de recurrencia de incendios, se he
rechazado la hipótesis inicial sobre la posibilidad de un bucle de
retroalimentación positiva (sensu Wilson y Agnew 1992) entre las especies que
acumulan mayor cantidad de combustible muerto y tienen una germinación
estimulada por el fuego (Capítulo 5). Dentro del régimen de recurrencia de
incendio estudiado, la cantidad de combustible muerto acumulado se ve
reducida por un segundo incendio. Por lo tanto, a nivel de comunidad, se pone
en entredicho las ventajas de la retención de este tipo de combustibles en la
estructura por las diferentes especies. Dentro del contexto de la tesis doctoral,
se propone la hipótesis de que la acumulación de combustible muerto puede
ser debido a la respuesta intrínseca de las diferentes especies al gradiente de
recursos sucesional (Capítulo 5), como la disponibilidad de luz o de agua. No
obstante, todavía se conoce muy poco sobre las causas fisiológicas de la
producción de biomasa muerta y, por lo tanto, nuevos estudios que indaguen
en estos mecanismos son necesarios.
BIBLIOGRAFÍA
Ackerly D (2004) Functional strategies of chaparral shrubs in relation to
seasonal water deficit and disturbance. Ecological Monographs 74: 25-
44.
Baeza MJ, Santana VM, Pausas JG, Vallejo VR (en prensa) Successional
trends in standing dead biomass in Mediterranean Basin species. Journal
of Vegetation Science.
Bell DT, Plummer JA, Taylor SK (1993) Seed germination ecology in
southwestern western Australia. Botanical Review 59: 24-73.
Connell JH, Slatyer RO (1977) Mechanisms of succession in natural
communities and their role in community stability and organization.
American Naturalist 111: 1119-1144.

135
Eugenio M, Lloret F (2006) Effects of repeated burning on Mediterranean
communities of the northeastern Iberian Peninsula. Journal of Vegetation
Science 17: 755-764.
Hanes TL (1971) Succession after fire in the chaparral of southern California.
Ecological Monographs 41: 27-52.
Keeley JE (1992) Recruitment of seedlings and vegetative sprouts in unburned
chaparral. Ecology 73: 1194-1208.
Keeley JE (1995) Seed-germination patterns in fire-prone Mediterranean-
climate regions. En: Arroyo, M.T.K., Zedler, P.H., Fox, M.D. (Eds.)
Ecology and biogeography of Mediterranean ecosystems in Chile,
California and Australia. Springer-Verlag, New York.
Pierce SM, Cowling RM (1991) Dynamics of soil-stored seed banks of six
shrubs in fire-prone dune fynbos. Journal of Ecology 79: 731-747.
Siles G, Rey PJ, Alcántara JM, Ramírez JM (2008) Assessing the long-term
contribution of nurse plants to restoration of Mediterranean forests
through Markovian models. Journal of Applied Ecology 45: 1790-1798.
Trabaud L, Lepart J (1980) Diversity and stability in garrigue ecosystems after
fire. Vegetatio 43: 49-57.
Valdecantos A, Baeza MJ, Vallejo VR (2009) Vegetation management for
promoting ecosystem resilience in fire-prone Mediterranean shrublands.
Restoration Ecology 17: 414-421.
Vega JA (2003) Regeneración del género Pinus tras incendios. Cuadernos
Sociedad Española de Ciencias Forestales 15: 59-68
Walker J, Reddell P (2007) Retrogressive succession and restoration on old
landscapes. En: Walker LR, Walker J, Hobbs RJ (Eds) Linking
restoration and ecological succession. Springer, New York.
Wilson JB, Agnew ADQ (1992) Positive-feedback switches in plant-
communities. Advances in Ecological Research 23: 263-336.

136

137
CAPÍTULO 9-
CONCLUSIONES

138

139
CAPÍTULO 9 - CONCLUSIONES
1. Existen diferencias en el nicho de regeneración de las especies
germinadoras obligadas. Esto hace que unas especies sean más
competitivas que otras a lo largo del gradiente sucesional o en diferentes
regimenes de incendio. Aunque, la mayor parte de los individuos de las
especies estudiadas se establecen en etapas inmediatamente post-
fuego, Cistus albidus se regenera prioritariamente en ambientes
recientemente perturbados, mientras que Ulex parviflorus y Rosmarinus
officinalis pueden establecerse en periodos entre incendios.
2. Existen procesos de sustitución especies a lo largo de la sucesión en los
matorrales dominados por especies germinadoras. Estos ecosistemas
se rigen por el mecanismo sucesional de tolerancia, y tras una primera
etapa dominada por U. parviflorus y C. albidus la comunidad pasa a
estar dominada por R. officinalis. Un fuego recurrente no afecta los
patrones de sustitución entre especies, pero si que afecta a la
abundancia y retrasa en el tiempo el punto donde las especies alcanzan
su óptimo.
3. Los campos de cultivo abandonados pueden establecer diferentes
trayectorias sucesionales dependiendo del régimen de recurrencia de
incendio. En ausencia de fuego, son dominados en una primera etapa
por Pinus halepensis, que con el paso del tiempo se convierten en una
formación mixta de pinar con especies rebrotadoras y de hoja ancha
como Quercus ilex y Q. coccifera. Un solo incendio puede sustituir el
pinar por una matorral de R. officinalis, donde el establecimiento de
especies germinadoras obligadas y de especies rebrotadoras de etapas
sucesionales posteriores pueden estar impedidas. Una alta recurrencia
de incendio en intervalos cortos de tiempo desvían el ecosistema hacia
una comunidad dominada por terófitos o herbáceas como Brachypodium
retusum.

140
4. La capacidad de retener combustible muerto y su disposición en la
estructura de la planta es determinante en los efectos del fuego en el
ecosistema. Bajo parches de la especie que más acumula combustible
muerto, U. parviflorus, se experimentan las mayores tasas de consumo
de biomasa y de temperatura de suelo. En contra, bajo los parches de R.
officinalis, la especie con menor acumulación de combustible muerto, se
encuentran los efectos opuestos.
5. Los procesos sucesionales de sustitución de especies en ecosistemas
de matorral llevan asociada una función de inflamabilidad basada en la
cantidad de combustible muerto acumulado. Una primera etapa de la
sucesión dominada por U. parviflorus, especie que mayor cantidad de
combustible muerto acumula, seguida por una dominancia de R.
officinalis, especie que acumula menor cantidad, conlleva a una función
de inflamabilidad de forma jorobada; es decir, tras un incremento inicial
del combustible muerto acumulado, éste disminuye con el transcurso de
la sucesión.
6. Tras un fuego recurrente no existe un incremento en la cantidad de
combustible muerto acumulado a nivel de comunidad. Por lo tanto, se
sugiere la ausencia de un bucle de retroalimentación positivo entre las
especies que acumulan mayor cantidad de combustible muerto y una
regeneración estimulada por el fuego con el régimen de recurrencia de
incendios estudiado.
7. Un efecto indirecto del fuego, como el aumento del régimen de
temperaturas diarias del suelo, puede ser un desencadenante de la
ruptura física de la dormancia en semillas de leguminosas de sureste de
Australia. Este efecto difiere entre las diferentes especies y, además,
esta modulado por el rango de temperaturas y el tiempo de exposición.

141

142
La defensa de la tesi doctoral feta per s’ha desenvolupat en les llengües següents: i , fet que, unit al compliment de la resta de requisits establits en la normativa pròpia de la UA, li atorga la menció de Doctor Europeu. Alacant, de de EL SECRETARI EL PRESIDENT







![Rosmarinus officinalis ... file1396 !(1)5 : ˘ˇ ˆ ˙ ˝ ˛ ˘ˇ ˆ ˛ [88] 1,&., 6˝ -˝ ˘ˇˆ ˙˝ y< ( ˛˚˜ )1](https://img.pdfslide.tips/doc/110x75/5d3df78588c993c11a8d1cb4/rosmarinus-officinalis-15-88-1-6-.jpg)