-
저작자표시-비영리-변경금지 2.0 대한민국
이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게
l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다.
다음과 같은 조건을 따라야 합니다:
l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건을 명확하게 나타내어야
합니다.
l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다.
저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다.
이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다.
Disclaimer
저작자표시. 귀하는 원저작자를 표시하여야 합니다.
비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다.
변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
http://creativecommons.org/licenses/by-nc-nd/2.0/kr/legalcodehttp://creativecommons.org/licenses/by-nc-nd/2.0/kr/
-
지리학석사 학위논문
A Diatom-Based Reconstruction of
the Paleoenvironmental Changes
during the Last Deglaciation
in Jeju Island, Korea
규조분석을 통한 마지막 해빙기 동안의
제주도 하논 마르형 호수 고환경 복원
2015년 8월
서울대학교 대학원
지리학과
한 지 우
-
A Diatom-Based Reconstruction of the
Paleoenvironmental Changes during the Last Deglaciation
in Jeju Island, Korea
지도교수 박 정 재
이 논문을 지리학석사 학위논문으로 제출함
2015년 4월
서울대학교 대학원
지리학과
한지우
한지우의 지리학석사 학위논문을 인준함
2015년 6월
위 원 장 (인)
부위원장 (인)
위 원 (인)
-
i
Abstract
A Diatom-Based Reconstruction of the
Paleoenvironmental Changes during the Last Deglaciation in Jeju
Island, Korea
Jiwoo Han
Department of Geography
The Graduate School
Seoul National University
Reconstructing the paleoclimate/paleoenvironment has become
more
important as the prediction of future climate change becomes a
more pressing
issue. Future climate change can be predicted by reconstructing
the
paleoclimate of the past, as it reoccurs in a repeated cyclical
fashion. In
particular, the last deglaciation is the focus of much research
these days
because it consists of various climate shifts which may be
similar to the future
climate change triggered by global warming.
Hanon maar paleolake, in the southern part of Jeju Island, is
located
in a geographically significant place that can provide the
missing link to the
paleoenvironment between Japan and China because the southern
part of Jeju
Island is influenced by the East Asia monsoon and the Kuroshio
Current.
However, only the morphology and terrestrial environment of
Hanon maar has
been researched so far, so it is necessary to investigate Hanon
maar
paleoenvironment using another type of proxy data to observe it
from a
different angle. Because Hanon maar had been a paleolake until
500 years
ago, diatom analysis is an appropriate methodology to
reconstruct the
paleoenvironment around Hanon paleolake; it provides information
on
-
ii
lacustrine environmental changes.
The aquatic environment during the last deglaciation of Hanon
maar
paleolake in Jeju Island, Korea, has been reconstructed through
diatom
analysis. Diatom analysis is a methodology investigating diatom
microfossils in
sediments, which are phytoplankton with silicic valves. Diatoms
are a good
indicator of environmental changes; they provide various
environmental
information such as salinity, water depth, acidification,
trophic status, water
temperature, and so on. Among them, water depth, trophic status,
saprobity,
water temperature and the acidification of Hanon paleolake
during the last
deglaciation has been reconstructed in detail based on the
information derived
from diatom analysis.
The sediment core(HN-1) had been extracted and analyzed
using
diatom analysis. This study covers from 90 to 250cm in the 10
meter long core,
which includes the last deglaciation(ca. 15,500 – 8,000 cal. yr
BP). After
identifying diatoms by microscope analysis, diatom diagrams were
constructed.
The zones in the diagram were determined based on constrained
incremental
sum of squares cluster analysis, and climate events during the
last deglaciation
in Hanon paleolake have been zoned: Oldest Dryas for 15,440 –
14,670 cal. yr
BP, the beginning of Bølling-Allerød for 14,670 – 14,180 cal. yr
BP, ongoing
Bølling-Allerød for 14,180 cal. yr BP – 12,810 cal. yr BP,
Younger Dryas for
12,810 – 12,150 cal. yr BP, Preboreal for 12,150 – 10,440 cal.
yr BP, and Boreal
for 10,440 – 7,980 cal. yr BP. The time table of the climate
shifts in Hanon
maar which is reconstructed in this study corresponds with other
studies of
Hanon maar paleoclimate.
The result of the diatom diagram was schematized to reconstruct
the
water depth, trophic status, saprobity, water temperature and
acidification of
the paleolake based on changes in the diatom assemblage and
limnological
processes. The reconstructed aquatic environment has also been
drawn in the
-
iii
graph which outlined relative phase-dependent environment
changes in the
Hanon paleolake. Afterwards, the reconstructed environment based
on the
diatom diagram has been verified by the results of PCA and P:B
ratio. The
components of Axis 1, 2 and 3 and the value of P:B ratio were
made into several
graphs, and they were compared to each other. Based on the
meaning of each
value such as trophic status, water depth and pH, the
verification made the
previously reconstructed lacustrine environment revised.
Overall, Hanon maar
paleoenvironment during the last deglaciation has changed as
follows: cold and
dry for the Oldest Dryas, increasing temperatures and moisture
for the Bølling-
Allerød, cold and wet-dry for the Younger Dryas, an increase in
temperatures
and temporarily drier for the Preboreal, and warm and dry/wet
for the Boreal.
This was the first time a paleoenvironment of Korean freshwater
zone
was constructed using diatom analysis; therefore, this study
itself is meaningful.
Furthermore, this study reconstructed trophic status, water
depth, saprobity,
water temperature and acidification of Hanon paleolake by diatom
analysis. In
conclusion, it was possible to reconstruct the paleo-lacustrine
environment of
Hanon maar paleolake during the last deglaciation using diatom
analysis, and
it provided a new proxy data for the
paleoenvironment/paleoclimate during the
last deglaciation on the Korean Peninsula.
Keyword : diatom analysis, Jeju Island, Hanon maar paleolake,
reconstruction of paleoenvironment, paleoclimate, the last
deglaciation Student Number : 2013-20114
-
iv
Table of Contents
Abstract
..............................................................................
i
Table of Contents
...............................................................
iv
List of Figures
....................................................................
vii
List of Tables
......................................................................
ix
Chapter 1. Introduction
...................................................... 1
1.1. Study Backgrounds
.......................................................... 1
1.2. Regional and Temporal Settings: Hanon Paleo-Maar Lake in
Jeju
Island
.........................................................................
3
1.3. Research Purpose and Structure
......................................... 7
Chapter 2. Literature Review
............................................. 10
2.1. Studies on Hanon Paleo-Maar Lake
................................... 10
2.2. Introduction to Diatoms
.................................................. 14
2.3. Studies on Diatom Analysis for Reconstruction of
Paleoenvironment in Korea and Abroad ..........................
15
Chapter 3. Methodology
..................................................... 18
3.1. Preparation of Diatom Slides
............................................ 18
3.2. Microscope Examination
.................................................. 20
3.3. Diatom Diagram and Diatom Concentration
....................... 21
3.4. Principal Component Analysis
.......................................... 25
3.5. The Ratio of Planktonic to Benthic Diatom Species
.............. 26
Chapter 4. Research Results and Analysis
.......................... 28
4.1. Principal Component Analysis
.......................................... 28
4.2. Diatom Flora in the Diagram
............................................ 40
-
v
4.1.1. Zone 1: 15,440 cal. yr BP – 14,670 cal. yr BP (Oldest
Dryas)
..................................................................................
43
4.1.2. Zone 2-a: 14,670 cal. yr BP – 14,180 cal. yr BP (The
beginning of Bølling-Allerød)
.......................................... 43
4.1.3. Zone 2-b: 14,180 cal. yr BP – 12,810 cal. yr BP
(Bølling-
Allerød)
.......................................................................
44
4.1.4. Zone 3: 12,810 cal. yr BP – 12,150 cal. yr BP (Younger
Dryas)
..................................................................................
44
4.1.5. Zone 4: 12,150 cal. yr BP – 10,440 cal. yr BP
(Pre-Boreal: the
beginning of Holocene)
.................................................. 45
4.1.6. Zone 5: 10,440 cal. yr BP – 7,980 cal. yr BP (Boreal)
......... 45
4.3. The Ratio of Planktonic to Benthic Diatom Species
.............. 46
Chapter 5. Discussion
........................................................ 51
5.1. Reconstructing the Paleoenvironment of Hanon Maar based
on
the Diatom Diagram and Schematization ........................
52
5.1.1. Reconstructing the Paleoenvironment of Hanon Maar
through
schematization of each zone based on Diatom Diagram ......
52
5.1.2. Summary of the Paleoenvironment of Hanon Maar based
on
the Schematization
....................................................... 63
5.2. Verification of the Reconstructed Paleoenvironment of
Hanon
Maar Based on Axis 1, 2, 3, and the Values of P:B Ratio ...
65
5.3. Comparison and Analysis between Diatom Analysis and
Other
Multi-Proxy Data from Another Research on the
Paleoenvironment of Hanon Maar
.................................. 74
-
vi
Chapter 6. Conclusions
...................................................... 81
Bibliography
......................................................................
84
APPENDIX
.........................................................................
94
Appendix I. A diagram including all diatom species
.................... 94
Appendix II. A count sheet of 17 major diatom species
.............. 96
Appendix III. The component scores of Axis 1, 2, 3 and 4 by
depth
..............................................................................
105
Appendix IV. The component scores of major species at Axis 1, 2,
3
and 4 from PCA
........................................................ 108
Appendix V. Images of diatom species in Hanon maar paleolake
109
국 문 초 록
......................................................................
112
-
vii
List of Figures
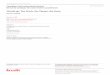
Figure 1. Map of Hanon maar and coring site(yellow arrow)
............... 4
Figure 2. Climate data of Seogwipo City, Jeju Island (1981-2010)
....... 5
Figure 3. Research flow chart
......................................................... 9
Figure 4. The time table of climate events in Hanon maar
paleolake
(Chung, 2007; Park et al., 2014a; Park et al., 2014b)........
12
Figure 5. The morphology of a diatom
........................................... 14
Figure 6. PCA graphs with major diatom taxa (A) excluding “spp.”
and
(B) including “spp.”
....................................................... 28
Figure 7. Principal component analysis of the HN-1 diatom data
....... 32
Figure 8. Diagram for examination of Axis 1, 2 and 3
...................... 34
Figure 9. The graphs of component scores on the PCA Axis 1, 2, 3
.... 36
Figure 10. The graph of component scores on the PCA Axis 1
........... 37
Figure 11. The graph of component scores on the PCA Axis 2
........... 38
Figure 12. The graph of component scores on the PCA Axis 3
........... 39
Figure 13. Diatom diagram including “spp.”
.................................... 42
Figure 14. The respective changes of planktonic and benthic
species . 49
Figure 15. P:B ratio
.....................................................................
50
Figure 16. Diatom diagram excluding “spp.”
................................... 54
Figure 17. Schematization of diatom assemblage changes – zone 1
.. 57
Figure 18. Schematization of diatom assemblage changes – zone
2-a 57
Figure 19. Schematization of diatom assemblage changes – zone
2-b59
Figure 20. Schematization of diatom assemblage changes – zone 3
.. 59
Figure 21. Schematization of diatom assemblage changes – zone 4
.. 61
Figure 22. Schematization of diatom assemblage changes – zone 5
.. 62
Figure 23. Schematization of relative phase-dependent
reconstruction
of aquatic environmental changes
................................. 64
Figure 24. P:B ratio and the component scores of Axis 2
................. 67
Figure 25. The component scores of Axis 1 and 2
........................... 70
Figure 26. The component scores of Axis 1 and 3
........................... 71
-
viii
Figure 27. The component scores of Axis 2 and 3
........................... 72
Figure 28. The revised version of the paleoenvironment in Hanon
..... 73
Figure 29. Climate shifts in Hanon maar by adding the one
reconstructed by diatom proxy data of this study ..........
75
Figure 30. Diagram for comparisons between the changes of
Botryococcus & Celtis and D. confervacea & PC 1, 2, 3 ..
77
-
ix
List of Tables
Table 1. Eigenvalues and variance explained by PCA of the
diatom
species from core HN-1
................................................... 29
Table 2. Saprobity index – the classes of water quality (Kelly
et al.,
2005)
............................................................................
55
Table 3. Habitat environments of major diatom species in Hanon
.... 56
Table 4. Table of relative phase-dependent reconstruction of
the
aquatic environmental changes in Hanon maar paleolake .....
63
-
1
Chapter 1. Introduction
1.1. Study Backgrounds
Global warming is a very familiar and even a clichéd topic
these
days. However, it poses an important question called ‘abrupt
climate
change’ impact that could possibly happen in the future. The
hygienic,
economic and cultural impact cannot be imagined easily in the
aftermath of
climate change triggered by global warming. However, the matter
of
whether climate change can be predicted is important because it
is directly
related to our lives rather than those reasons. Because climate
is repetitive
according to the Milankovitch Theory (Roberts, 1998), building a
predictive
model based on fluctuations of past climate is a good way to
prepare for
climate shifts in the future.
There are several ways to build a predictive model for
future
climate; however, analyzing proxy data is the most common way.
Proxy
data is a substitute literally for real climatic/environmental
data; therefore,
it cannot show us the exact same environment at that time due
to
limitations in the data. For example, the figures of dinosaurs
people think
could be wrong because they are restored from their proxy data;
fossilized
bones, imprints, et cetera (Conway et al., 2012). Therefore,
researchers
should use and gather as much proxy data as possible to minimize
errors
that may occur. A predictive model for future climate cannot be
made
without proxy data. Models are reconstructions of climate shifts
that have
happened in the past. Therefore, proxy data is necessary to
build predictive
models of the future climate.
-
2
Diatoms are cosmopolitan primary producers. They are
everywhere
that contains moisture and water; therefore, it is one of the
easiest ways to
obtain climate data among many other proxy data (Round et al.,
1990).
That is, diatom is one of the crucial proxy data to
reconstruct
paleoenvironment. Through diatom analysis, aquatic environment
can be
reconstructed according to the habitats of each diatom species
such as
water depth, temperature, pH, salinity, and so on (Mackay et
al., 2005). The
aquatic environment is always related to the atmosphere and/or
other
environmental factors (Meyers et al., 1993; Kuwae et al., 2002;
Wang et al.,
2012). In summation, diatoms offer valuable information for
paleoenvironmental reconstructions (Wang et al., 2012; Chen et
al., 2014;
Katsuki et al., 2003; Ribeiro and Senna, 2010).
Jeju Island is located in a critical are affected by the East
Asia
monsoon and the Kuroshio Current. Also, Hanon maar lies between
Japan
and China, so the data from Hanon maar can bridge the gap
between those
two countries and play a key role for reconstructing East Asia
climate
(Chung, 2007; Park, 2015). Therefore, it is necessary to
compile
environmental and climatic data for reconstruction of
paleoenvironment
around Jeju Island. Above all, Hanon maar paleolake is a good
place to
study paleoenvironment because it contains over 10m long
sediment under
the ground that shows good preservation (Park et al., 2014a;
Park et al.,
2014b). However, Hanon paleolake has not been studied enough in
spite of
this in the field of paleoclimatology; what has been done only
focuses on the
terrestrial environment and climate (Chung, 2007; Park et al.,
2014a; Park
et al., 2014b) even though the place was a paleolake until
around 500 years
ago (Bowers et al., 2014). Consequently, reconstructing the
paleoenvironment in Hanon maar using diatom analysis is
necessary to
-
3
observe it within the paleolake, which is close to what really
happened.
1.2. Regional and Temporal Settings: Hanon Paleo-
Maar Lake in Jeju Island
Hanon paleo-maar Lake, which is located in 400m from the
coast
of southern part of Jeju Island, is the only maar lake on the
Korean
Peninsula (Bowers et al., 2014; Choi et al., 2006; Chung, 2007;
Park et al.,
2014a; Park et al., 2014b). It is located at 33°14’N, 126°32’E
and is 53m
above sea level. Hanon maar was formed during the late
Pleistocene, ca.
50,000 yr BP. The diameter is about 1km for the crater and 850m
for the
crater lake, and the floor area is about 216,000m2 (Bowers et
al., 2014). The
depth of water is estimated about 5m for average and 13m for
maximum
depth according to Choi et al. (2006).
Hanon maar paleolake is a critical place to reconstruct
paleoclimate
and paleoenvironment in East Asia, not only because of its
location, but
also because of the particular physical environment. According
to Yoon et al.
(2006c) and Lee et al. (2008), Hanon crater lake developed in
the summit
of Hanon maar as a closed aquatic environment, which has low
inflow and
outflow(low energy environment). Therefore, the characteristics
of the
sediment in Hanon paleolake are mainly influenced by its own
ecology and
climate, so the sediment reflects the changes in the biological,
geological
and morphological environment within the lake. Thus, the
sediments of
Hanon paleolake are very important as an indicator of the
paleoenvironment considering its location and topography.
Especially,
-
4
diatoms living in the lake will be a good reference for
reconstructing
paleoenvironment.
Figure 1. Map of Hanon maar and coring site(yellow arrow)
The physical environment of Jeju Island where Hanon maar lies
is
a crucial place to observe the paleoclimate and paleoenvironment
around
East Asia because Jeju Island is affected by two important
environmental
factors concerning the environment of East Asia. First, the
Kuroshio
Current flows south of Jeju Island. It is a warm current
starting from the
east side of Taiwan and transfers heat energy from the tropics;
therefore,
-
5
the current definitely has an influence on the climate and
ecology of Jeju
Island, especially the southern part (Park, 2015; Chung,
2007).
The second factor is the East Asian monsoon(EAM). Hanon maar
paleolake is located in the EAM belt, and the climate of Jeju
Island is
normally hot and humid in summer and cold and dry in winter
(Anonymous, 2010). However, the characteristics of climate
around
Hanon maar during winter are lessened than in other parts of the
Korean
Peninsula and the northern part of Jeju Island due to the warm
Kuroshio
Current. According to the data from Domestic Climate Data at
Korea
Meteorological Administration, the lowest mean monthly
temperature is
6.8°C in January, the highest mean monthly temperature is 27.1°C
in
August, and the mean rainfall is 1,923mm in southern Jeju
Island
(Anonymous, 2010). It rains mainly between July and August
(Figure 2).
Figure 2. Climate data of Seogwipo City, Jeju Island
(1981-2010)
-
6
Accordingly, Hanon maar is a transitional zone between
climate
signals of North Atlantic(EAM – terrestrial deposits) and
western North
Pacific(Kuroshio Current – pelagic deposits) (Park and Park,
2015).
Therefore, it is possible to say that Hanon paleolake shows the
climate
fluctuations around East Asia. Researchers in China and Japan
have
actively studied paleoclimate and paleoenvironment changes in
East Asia,
and Hanon maar paleolake is a central place to connect studies
from China
and Japan. In other words, in the context of that Hanon maar is
located in a
transitional zone between terrestrial and marine environment and
there is
not enough data since the last glacial period, Hanon maar has an
important
meaning in the matter of climate change in East Asia (Park,
2015).
The coring site of HN-11 for this study is pointed in Figure 1.
The
core was taken using a hydraulic piston corer in November, 2012.
Its total
length is about 10m, and each is sampled by 1cm thickness. For
this study,
samples between 250 and 90cm were analyzed every 2cm, which
covers
from 15,500 to 8,000 cal. yr BP. Samples from the core were
stored in
refrigerator before using, so the samples for this study were in
wet
condition, not dry.
Reconstructing the environment during the last deglaciation is
one
of the main goals in this study. Because the last deglaciation
is a transition
period which consists of centennial- to millennial-scale climate
changes, it
is necessary to observe exactly how climate had shifted to
improve the
predictability of future climate change (Park, 2015). During the
period
being studied, the Bølling-Allerød(BA), Younger Dryas(YD),
Preboreal,
8.2ka event and Holocene Climate Optimum(HCO) for
centennial-scale
1 The same core(HN-1) had been used for this study with Park et
al., 2014a and
2014b.
-
7
occurred. In the northern hemisphere, the BA, Preboreal and HCO
were
warm periods(interglacial and interstadial), while the Younger
Dryas and
the 8.2ka events were cold periods(stadial). The cause of both
stadial
events has not been revealed clearly, yet. However, it has
become common
knowledge that the YD and the 8.2ka happened because freshwater
from
melted glacier flowed into the oceans and cut the thermohaline
circulation,
which is called Atlantic Meridional Overturning Circulation
nowadays (Park,
2015). Researchers are very interested in both climate events
because there
is a possibility that the abrupt decline in temperature could
happen again in
the future due to global warming; glacier melting will cause
unpredictable
environmental changes as global warming proceeds continuously or
even is
accelerated in the future. That is, the unpredictable
environmental change
can be predicted through studying in climate change during the
YD and the
8.2ka event. Therefore, researching in the last deglaciation is
important for
our future. In particular, the transition between BA and YD was
the period
when climate had been changed abruptly, so it is necessary to
be
reconstructed in detail; this is why the last deglaciation has
been chosen for
this study.
1.3. Research Purpose and Structure
In this study, the paleoenvironment during the last
deglaciation(ca.
15,500 - 8,000 cal. yr BP) in Hanon maar will be reconstructed
by using
diatom analysis. Through diatom analysis, the paleoenvironment
in Hanon
maar can be reconstructed in different aspects showing the
environmental
-
8
changes within the lake. It is possible because diatoms live in
the lake, so
they can reflect environmental changes of the paleolake much
more
sensitively and closely as the climate change around Hanon
maar.
Furthermore, the reconstructed environment during the last
deglaciation
will provide proxy data that would be useful for predicting
future climate
change on the Korean Peninsula. This study could be helpful to
bridge the
gap between China and Japan which has been missed out frequently
so that
the change of paleoenvironment in East Asia can be
reconstructed.
To sum up, the purposes of this research are to:
1. Reconstructing the paleoenvironment in Hanon maar during
the last deglaciation using diatom analysis, which indicates
various environmental changes in the paleolake.
2. Providing another proxy data for the reconstruction of
paleoenvironment on the Korean Peninsula during the last
deglaciation.
The literature review related to this study is explained in
Chapter 2,
including the introduction of diatoms. The methodology, diatom
analysis, is
introduced in Chapter 3: how to prepare diatom microscope
slides, how to
count diatoms for microscope analysis, how to draw diatom
diagram, and
applications of PCA and P:B ratio. In Chapter 4, the results of
diatom
diagram, PCA and P:B ratio are described. In Chapter 5, the
results from
the previous chapter are interpreted, and the verification of
the previous
interpretation of this study and comparison study to other proxy
data of
other studies are discussed.
-
9
Figure 3. Research flow chart
-
10
Chapter 2. Literature Review
2.1. Studies on Hanon Paleo-Maar Lake
Hanon maar is a significant place for reconstructing the
paleoenvironment of the Korean Peninsula and even East Asia due
to some
reasons explained in Chapter 1.2. There are several studies
dealing with
Hanon maar paleolake2 in various fields.
First, there have been done several studies to reconstruct
paleoclimate of Hanon. Among of them, Chung (2007) and Park et
al.
(2014a; 2014b) especially used microfossil analysis – pollen –
in the
sediment of Hanon for reconstructing. Chung (2007) analyzed the
pollen
record of Hanon maar, which was taken at the center of the lake,
and
showed how it provided a vegetation history of Jeju Island
during the last
deglaciation. Three zones were established: zone 1(21,800-14,400
cal. yr BP)
dominated by Artemisia and Gramineae shows much a colder and
drier
climate than the present one on Jeju Island, which means the
period is
related to the Last Glacial Maximum(LGM). Zone 2(14,400-11,800
cal. yr
BP) shows a sudden increased in Polypodiaceae ferns and an
abrupt
decline in herbaceous taxa indicates the transitional period
from glacial to
interglacial while suggesting warmer climate than before. Zone
3(11,800-
9,900 cal. yr BP) showing the retreat of grassland vegetation
and further
expansion of temperate deciduous broadleaved forests displays
similar
climate, warm and humid, to the modern climate in Jeju Island.
He asserts
that it is related to stronger influence of the East Asian
summer monsoon.
2 “Hanon maar paleolake” is a correct term; however, it is going
to be written in
mostly “Hanon” after then for the simplicity.
-
11
Park et al. (2014a; 2014b) also studied the area of Hanon maar
lake
using pollen analysis and many geochemical proxies such as
magnetic
susceptibility, grain size, δ13C, δ15N, total organic carbon,
carbon-nitrogen
ratio, algae, etc. Park et al. (2014a; 2014b) recovered an
approximately
10m-long core, and they published two papers covering different
time
scopes – the one between 32.5 and 6.9k cal. yr BP and another
one focusing
on the last deglaciation. In particular, Park et al. (2014b)
divided into 6
zones to reconstruct the climate change and its effects on the
vegetation
around Hanon in the past. It contends that there were Oldest
Dryas(15,450-
14,650 cal. yr BP), Bølling-Allerød(14,650-12,900 cal. yr BP),
Younger
Dryas(12,900-11,900 cal. yr BP), Pre-Boreal(11,900-10,300 cal.
yr BP),
Boreal(10,300-7,800 cal. yr BP), and Holocene Climate
Optimum(7,800-
7,300 cal. yr BP) occurred during this time frame. The paper
interpreted
proxy data conjunctly so that they could reconstruct
environmental change,
particularly in vegetation and climate, accurately and with
high-resolution.
The Younger Dryas on the Korean Peninsula is detected for the
first time in
this paper. The timelines for Park et al. (2014a) and Park et
al. (2014b) are
slightly different because the samples were analyzed at 8cm
intervals in
Park et al. (2014a) – pre LGM(32,500-25,200 cal a BP3), the
earlier part of
LGM(25,200-21,500 cal a BP), the later part of LGM(21,500-17,600
cal a
BP), early deglacial period(17,600-14,700 cal a BP), late
deglacial
period(14,700-10,700 cal a BP), and early Holocene(10,700-6,900
cal a BP).
This paper focuses much on the climate changes in orbital- and
millennial-
scale.
3 All dates in Park et al. (2014a) is written in cal a BP which
is calibrated with the
intcal09 data set (Park et al., 2014a; Reimer et al., 2009).
-
12
Figure 4. The time table of climate events in Hanon maar
paleolake (Chung, 2007; Park et al., 2014a; Park et al., 2014b)
Those results from both Chung (2007) and Park et al. (2014a;
2014b) were slightly different because the intervals of analyzed
samples
were different for each study; Chung (2007) did 10cm interval,
Park et al.
(2014a) did 8cm interval, and Park et al. (2014b) did 2cm
interval.
Therefore, the distinction in resolution delineates slightly
different stories
in each scale. Timeline of Hanon maar lake from the three
studies have
been schematized in Figure 4. Yoon et al. (2006a; 2006b; 2006c)
and Lee
et al. (2008) also reconstructed the paleoclimate and
paleoenvironment of
Hanon and Jeju Island using by sedimentological analyses.
Second, some researchers tried to reconstruct the
geomorphology
of Hanon such as Yoon et al. (2006a) and Choi et al. (2006).
Yoon et al.
(2006a) recovered the morphology and geological process in Hanon
crater
using resistivity survey and boring. Choi et al. (2006)
reconstructed the
volcanic lake in Hanon crater by applying the spatial
statistical techniques
based on the depth information from the seismic survey and known
data.
Overall, there were some attempts to recover Hanon morphology.
Third,
-
13
Lee and Ahn (2005) did a simple fundamental study in flora and
fauna of
Hanon to reconstruct what it looked like before people converted
it to
farmland.
In conclusion, considering the value of Hanon in an aspect
of
reconstructing paleoenvironment and paleoclimate, there have
been fewer
researches done until now than expected. Although there are some
studies
have been carried out to reconstruct paleoclimate using pollen
analyses and
geochemical analyses (Park et al, 2014a; Park et al., 2014b;
Chung, 2007;
Lee et al., 2008; Bowers et al., 2014), it is not still enough
for Hanon
reconstruction. It is necessary to have much closer look of the
real past
environment around Hanon.
Because Hanon was a (maar) lake, the proper way of
reconstructing
paleo-environment would be reconstructing the “lake” environment
of
Hanon in the past; Hanon was a lake basically, so reconstructing
aquatic
environment of the past would be a key to investigate
paleoenvironment
and paleoclimate around Hanon. Diatoms live in a lake, and thus
they show
the status of lake at that time closely. However, there is no
research using
diatom analysis that has been done around Hanon. This is why
diatom
analysis should be performed in this area. Without
reconstructing
lacustrine environment in Hanon, the reconstructed
paleoenvironment
around Hanon would just show the half of what happened in the
past
actually. Therefore, diatom analysis is going to be carried out
to
reconstruct Hanon paleo-maar lacustrine environment in the past,
where is
a significant place to study for paleoenvironment reconstruction
in East
Asia.
-
14
2.2. Introduction to Diatoms
The first observation of diatom was recorded in 1703 by an
English
gentleman. The recorded diatom adhered to the roots of the
Lemna, which
are pond-weed herbs (Round et al., 1990). After then, studies on
diatoms
have begun and being done actively until now.
Figure 5. The morphology of a diatom – two valves(epitheca and
hypotheca) are tied up together by girdle bands (Kelly et al.,
2005)
Diatoms are unicellular algae and phytoplankton. The name
diatom
comes from Greek, which means “cut into two.” It is because a
diatom
consists of two valves with girdle bands (Figure 5). When a
diatom dies,
-
15
their silicic valves become separated and deposited on the
floor, and those
valves are counted by diatomists. As diatoms are
phytoplankton
aforementioned, most of them are photosynthetic, so their
habitats are
usually in/on sunlit places. They occur in environment
containing
water/moisture such as streams, lakes, oceans, and even in
wetlands. That
is, diatoms are in everywhere. Also, diatoms are primary
producers – they
play an important role in nutrient supply along with bacteria
and other
kinds of plankton (Sigman and Hain, 2012) in aquatic
environment.
Diatoms are ecological indicators because each diatom species
lives
differently, conditions by conditions such as pH, salinity,
water
temperature, water depth, trophic level, thermal stratification,
and so on.
For these reasons, they are being used as an indicator of
pollution these
days. What is better, their frustules(valves) are made from
silica, SiO2. It is
because silica is insoluble, they are fossilized when they die
and sink into
the sediment. Also, their complexly shaped valves seem different
according
to each species. Therefore, diatoms can be used as a proxy data
to unearth
the paleoenvironment. In other words, diatoms can be good
indicators of
past environment (Mackay et al., 2005).
2.3. Studies on Diatom Analysis for Reconstruction
of Paleoenvironment in Korea and Abroad
There have been a lot of works related to diatom analysis. Due
to its
useful roles as proxy data, diatom analysis is a popular method
in many
other countries, especially in United Kingdom, Germany, and
Japan.
-
16
Diatom analysts can be divided into two groups largely: people
who classify
diatoms taxonomically and people who analyze the change of
diatom
assemblages to reconstruct an environment of the past or
present. In the
latter case, it is necessary to have adequate information to
interpret diatoms
in the environment; which environment(pH, salinity, temperature,
organic
pollution, etc) is the best for which diatom species. Chapter
2.3. mainly
focuses on studies on diatom analysis for reconstructing the
paleoenvironment – the latter usage.
There are many paleoclimate/paleoenvironment studies
applying
diatom analysis in Japan. Japanese diatomists have worked on
reconstructing the paleoenvironment using diatoms such as in
Lake Biwa
and Suigetsu (Meyers et al., 1993; Kuwae et al., 2002; Kosseler
et al., 2011).
Beyond reconstruction of lacustrine environmental changes in the
past,
many diatom researchers study on various topics using diatom
analysis
such as reconstructing productivity changes in the ocean and
lagoon
(Katsuki et al., 2003; Katsuki et al., 2012). Also, there have
been many
studies on paleoclimate and paleoenvironment using diatoms
rousingly in
China. Especially, many paleoclimate studies have being
performed in the
Long Gang Volcanic Field (LGVF) region recently – Lake
Xiaolongwan,
Sihailongwan, and Erlongwan – in Jilin Province, northeast China
(Wang et
al., 2012).
In contrast with the situation in other countries in the
world,
especially comparing to Japan and China, studies on diatom
analysis in
Korea are very calm. Most diatom analyses in South Korea have
been
carried out in coastal regions or ocean – brackish water and
seawater.
There are a few of researchers performing in lagoon and/or
ocean: Go et al.
(2013) in Hwajinpo on the eastern coast of Korea, Bak et al.
(2006; 2009;
-
17
2010) in Seokrim-dong in Seosan City and Uleung Basin in East
Sea, and
Yoon et al. (2004) in the eastern part of Jeju Island, etc.
Research
performed in coastal regions is generally to determine sea level
fluctuation
in the past.
Reconstructing sea level changes is important; however,
reconstructing inland paleoenvironmental change is also
important,
because the environments of brackish and salt water contain
different
stories from that of freshwater. That is, investigating
freshwater diatoms in
the past would let us see things that cannot be seen in lagoons
or oceans
such as how strong wind blew or how much it rained or how strong
sunlight
was, etc. Therefore, to reconstruct regions in freshwater
environments is to
reconstruct the major part of the past environment on the
Korean
Peninsula. Even though there are some studies which have done
in
freshwater regions in Korea, the past environment has not been
studied.
Instead, it was done to investigate the habitats of diatoms and
the present
environment to observe how much the place is polluted. Although
it seems
that using diatom analysis in the field of paleoenvironment
reconstruction
is not popular yet, collecting and recording diatoms in Korea
have been
more actively pursued recently4. There may be some ongoing
studies that
are reconstructing the paleoenvironment right now, but more are
needed.
Because diatom analysis is a good indicator of ecological and
environmental
change, it is a good tool to reconstruct paleoenvironment. There
ought to be
more studies on this field on the Korean Peninsula.
4 By leading of National Institute of Biological Resources,
Korean diatoms in
streams, lakes, and oceans are collected and recorded. The
project has started since 2010s. They published illustrated books
of diatoms and provide the information on their website.
-
18
Chapter 3. Methodology
3.1. Preparation of Diatom Slides
For diatom analysis, sample preparation is needed in some ways
to
remove materials such as calcareous and organic matters and to
improve
visibility of diatoms under the microscope. According to Round
et al. (1990),
“there is no universally best method and every diatomist has
some
preferred recipe.” There are various methods for diatom
preparation, and
the method can be modified properly depends on soil condition in
each
environment. In this study, a modified version of Katsuki et al.
(2003)
was chosen for sample preparation. The samples are dried at
room
temperature5 and treated as below:
a. About 0.1g of sample is treated by H2O2 to remove organic
matter
and distilled water in a 100mL beaker6.
b. The sample is boiled for 1-2 hours at 120°C until the foam is
gone.
c. A minute amount of sodium hexametaphosphate(SHMP) is
mixed
and left for 15-30 minutes.
d. The pH of the solution is checked, and the surface solution
is
thrown away when pH is less than 7 (In case the pH shows
more
than 7, mix it with distilled water and check the pH. Repeat
the
same step until it shows less than 7).
5 Samples for diatom analysis should be dried naturally because
silicic valves are
easily broken by heat. 6 The sediments in Hanon barely contain
calcareous matters, so using
hydrochloride (HCl) could be omitted for this experiment.
-
19
e. The beaker is filled with distilled water to the top and left
for 5
hours.
f. The pH is checked again, which shows less than 7, and then
its
surface solution thrown away.
g. The beaker is filled with distilled water until the water
level is at
25mL and shaken thoroughly.
h. The sample is taken by using micropipette (Make sure the
sample is
shaken thoroughly and taken from the middle of the beaker).
The
amount of sample can be decided according to each sample’s
abundance of diatoms.
i. The sample is put on a heated cover slide with low
temperature.
j. When the sample is dried, minute mountmedia (i.e. Naphrax) is
put
to mount samples on the cover slide.
k. The mountmedia is melted at 120°C until its bubbles are
gone.
l. The cover slide is put onto a microscope slide.
In step g, each 25mL water volume of beakers should be measured
by
cylinder accurately. The values are used to calculate diatom
concentration
afterward. Three samples at 100, 164, 250cm had insufficient
amount. The
remained samples of 100 and 250 were almost half of normal, so a
lesser
amount of distilled water was used in step g. In the similar
way, only 1/10
amount remained for sample 164, so the sample was treated by
less H2O2
and distilled water in step a and g.
-
20
3.2. Microscope Examination
Among of 10m core of HN-1 from Hanon in Jeju Island,
sediment
from 90cm to 250cm has been examined for this study. Samples for
diatom
analysis are taken every 2cm interval, and the thickness of each
sample is
1cm; 81 slides have been examined. Diatoms in the slides are
counted on an
optical microscope, Leica ICC50 microscope, at a magnification
of ×1000
with an oil immersion objective (index of refraction = 1.515).
Because Leica
ICC50 has a built-in HD digital camera, pictures of diatoms are
taken for
reference in order to prevent/reduce mistakes and errors for the
first 1,500
valves. The identification of diatoms are referred to the
taxonomy studies
and illustrated books such as Algal Flora of Korea series (Joh,
2010; Joh et
al., 2010; Joh, 2011; Lee, 2011; Joh, 2012), Freshwater Algae of
North
America (Stoermer et al., 2003; Kingston, 2003; Kociolek and
Spaulding,
2003a; Kociolek and Spaulding, 2003b; Lowe, 2003), The Diatoms
(Round
et al., 1990), Bacillariophyceae 1, 2 (Krammer and
Lange-Bertalot, 1986;
Krammer and Lange-Bertalot, 1988), and Bibliotheca
Diatomologica
(Krammer and Lange-Bertalot, 1985; Lange-Bertalot and Krammer,
1987;
Lange-Bertalot and Krammer, 1989). Moreover, several websites
containing
illustrated guides are very helpful in identifying diatoms:
Common
Freshwater Diatoms of Britain and Ireland (Kelly et al., 2005),
Diatoms of
the United States (Spaulding et al., 2010), and Diatoms of the
Southern
California Bight (Kociolek, 2012). The classification system of
diatom
species in Hanon paleolake is based on Round et al. (1990).
Microscopic analysis in this study has been performed in
species
level. About 350 valves are counted for each slide on
average.
-
21
Unidentifiable diatoms are not counted; for example, there were
a few very
small and/or broken diatoms to identify with 1000× magnitude.
Some
broken diatoms which cannot be counted fully as one diatom
species are
not counted, either; centric diatoms without central area and/or
remained
less than 1/3 and pennate diatoms without raphe and/or both apex
and/or
remained less than a half are not counted. Those which are
unable to be
identified into species level are counted as genus level, and
they are written
in “xxx spp.”; that is, “xxx spp.” does not mean that it is a
sum of the genus.
The pictures of major diatom species from Hanon are in Appendix
V.
3.3. Diatom Diagram and Diatom Concentration
All prepared diatoms are counted by light microscopic
examination
as aforementioned – 350 diatom valves roughly for each sample.
Tilia
version 1.7.16 (Grimm, 1992) is used to construct stratigraphic
diatom
diagram with zonation based on constrained incremental sum of
squares
cluster analysis, CONISS in Tilia software: the zones are given
numbers for
identification such as zone 1, 2-a, 2-b, 3, 4 and 5. Totally,
six zones
including subzones are decided at depths of 120, 136, 156, 220
and 236cm
(Grimm, 1987). The zonation has been determined in consideration
of the
principles of CONISS and the pattern of change in diatom
assemblages
(Figure 10 and 12). Several clusters can be divided and merged
into slightly
differently according to the dendogram; however, six zones at
these depths
are established finally by considering the main changes of the
diatom
assemblages. Even though CONISS is a quantitative way to define
zones, it
-
22
is not always right (Bennett, 1996); clusters can be divided
into several
zones by cutting the dedrogram at various units of height(total
sum of
squares), so zones can be splitted and merged in various ways.
Therefore, it
is better to ponder the zonation based on CONISS while
considering the
changes in major diatom assemblages.
The data acquired from counting diatom valves is necessary to
be
converted into relative abundance with an equation below in
order to draw
a diatom diagram (Boden, 1991):
Relative abundance of a particular species in depth XX
=number of a particular species counted in depth XX
total number of diatom valves counted in depth XX× 100
The data with absolute values of counted diatom valves is not
appropriate
to compare the changes in each diatom assemblage because the
valves in
each depth are not counted in the exact same amount.
Therefore,
converting absolute values into relative values is required for
accurate
analysis. Finally, a diatom diagram can be constructed after
the
calculation and zonation. The diatom diagram shows the sum of
each
species through the depths, and it represents which species
are
dominant/rare, increase/decrease, appear/disappear, and so
forth.
Relevant species are arranged side by side in the diagram for
prehension
(Figure 13 and 16).
Hanon radiocarbon dates were not obtained individually in
this
study because the AMS radiocarbon dates were already measured
and the
age-depth model were already created by Park et al. (2014b),
which used
the exact same core(HN-1). They did not use the median ages.
They took the
-
23
minimum or maximum two-sigma ages to smoothen the interpolating
curve
(Park et al., 2014b). For more detailed information about the
radiocarbon
dating, refer to the Park et al. (2014a; 2014b).
Next, diatom concentration is calculated and described.
Calculating
diatom concentration is a way to estimate the actually existed
number of
population of diatom valves from the number of observed(counted)
diatom
valves (Moos et al., 2009; Boden, 1991; Scherer, 1994; Bak et
al., 2001; Bak
et al., 2002; Bak et al., 2010). The equation to calculate
diatom
concentration is as follow:
𝐷𝑖𝑎𝑡𝑜𝑚 𝑣𝑎𝑣𝑙𝑒𝑠 𝑝𝑒𝑟 1𝑔 𝑜𝑓 𝑑𝑟𝑦 𝑠𝑒𝑑𝑖𝑚𝑒𝑛𝑡 [𝑣𝑎𝑙𝑣𝑒𝑠
1𝑔] =
𝑐𝑜𝑢𝑛𝑡𝑒𝑑 𝑑𝑖𝑎𝑡𝑜𝑚 𝑣𝑎𝑙𝑣𝑒𝑠 [𝑣𝑎𝑙𝑣𝑒𝑠] ×𝑐𝑜𝑣𝑒𝑟 𝑠𝑙𝑖𝑑𝑒 𝑔𝑙𝑎𝑠𝑠 𝑎𝑟𝑒𝑎 [𝑚𝑚2]
𝑐𝑜𝑢𝑛𝑡𝑒𝑑 𝑎𝑟𝑒𝑎 [𝑚𝑚2]×
𝑏𝑒𝑎𝑘𝑒𝑟 𝑤𝑎𝑡𝑒𝑟 𝑣𝑜𝑙𝑢𝑚𝑒 [𝑚𝐿]
𝑝𝑖𝑝𝑒𝑡 𝑤𝑎𝑡𝑒𝑟 𝑣𝑜𝑙𝑢𝑚𝑒 [𝜇𝐿]× 1000 [𝑚𝐿 → 𝜇𝐿] ×
1
𝑑𝑟𝑦 𝑠𝑒𝑑𝑖𝑚𝑒𝑛𝑡 𝑣𝑜𝑙𝑢𝑚𝑒 [𝑔]
𝐷𝑟𝑦 𝑠𝑒𝑑𝑖𝑚𝑒𝑛𝑡7 𝑣𝑜𝑙𝑢𝑚𝑒 [𝑔]
= 𝑤𝑒𝑡 𝑠𝑒𝑑𝑖𝑚𝑒𝑛𝑡 𝑣𝑜𝑙𝑢𝑚𝑒 [𝑔] × (1 − 𝑤𝑎𝑡𝑒𝑟 𝑐𝑜𝑛𝑡𝑒𝑛𝑡 𝑟𝑎𝑡𝑒)
This equation is modified from the calculation method of Katsuki
et al.
(2003) based on the author’s advice. The graph of diatom
concentration is
inserted in the diatom diagram of Figure 13 and 16 to make it
easy to
compare the changes in diatom assemblage. The diatom
concentration is
also called “absolute number of diatom valves,” and its unit is
[valves/1g of
dry sediment].
7 Sediments for diatom analysis should be dried at room
temperature because
diatom valves, which consist of silica, are vulnerable to heat.
The sediments for this study were stored in refrigerator, so they
were in wet condition when they were prepared as sediment samples
for diatom analysis.
-
24
There are some factors which control the abundance of diatom
such
as diatom productivity, microbial decomposition, dilution by
clastic
sedimentation, dissolution of diatom valves, etc., and diatom
concentration
is ultimately related to the autochthonous water column
productivity and
paleoclimatic changes (Kuwae et al., 2002). There have been
several
studies in the relationship between diatom concentration and
environmental conditions: Meyers et al. (1993) maintains that
diatom
productivity increases as regional precipitation, soil erosion
and rock
weathering increase, and coarser detrital sediment particles are
usually
delivered with washed out nutrients during wetter intervals by
runoff.
Xiao et al. (1997) asserts that the values of higher biogenic
silica flux signify
warmer and wetter paleoclimatic conditions as well. That is,
water
temperature is closely associated with diatom productivity,
which is driven
by air temperature. Kuwae et al. (2002) also suggests that the
nutrients
washed into the lake result in enhanced diatom productivity, and
more
nutrient input into the lake happens during a warm period with
greater
levels of precipitation. Therefore, diatom concentration is
certainly
influenced by lake productivity(trophic status of the lake)
and
paleoclimate(water temperature in lake), which can be connected
to the
paleoclimate and paleoenvironment around the lake. The
interpretation of
diatom concentration is going to be explained in Chapter 5.
-
25
3.4. Principal Component Analysis (PCA)
In the field of community ecology, data are aligned and
organized
using ordination methods to find a relationship of species,
sites and
environmental variables. There are two ways of ordination
methods
whether or not it contains environmental variables – ordination
analysis
and canonical ordination analysis (Ko et al., 2015). Ordination
analysis is
used to investigate the relation between species and its
appearance by
deriving indirect environmental factors from species and
appearance data
which do not have environmental information. This is called
indirect
ordination analysis, and it includes Principal Component
Analysis(PCA),
Correspondence Analysis(CA), and so on (Ko et al., 2015).
PCA converts high-dimensional data into low-dimensional
data(dimension reduction) using orthogonal transformation to
extract
several principal components (Janžekovič and Novak, 2012).
Principal
components are the major components that can explain all the
variables in
the data, and each component is represented as uncorrelated axes
by
orthogonal transformation (Janžekovič and Novak, 2012). That is,
PCA
interprets and summarizes the major patterns of variation within
the data
(Väliranta and Weckstrom, 2007); therefore, it is helpful to
focus on the
main characteristics of the phenomenon (Janžekovič and Novak,
2012).
Even though PCA can indicate environmental niche due to its
process, it
cannot cover all dimensions of an environmental niche
(Janžekovič and
Novak, 2012).
PCA is carried out to see the tendency of diatom assemblages
in
this study. PCA graphs will be shown in Figure 7, and the graphs
drawn by
-
26
component scores of Axis 1, 2 and 3 will be shown in Figure 9 –
12 in
Chapter 4 and 5. Those are graphed by selecting species with
greater than 5%
frequency in at least one depth(sample) using CANOCO 5.0.2.0
(ter Braak
and Šmilauer, 2012). The variable loadings of major species are
going to be
interpreted to find out what each axis stands for in Chapter
4.1. It is because
the component scores reflect how significant a species is at
delineating the
variation within an assemblage (Allen et al., 2005).
Accordingly, it is
necessary to construe what each axis means. The analyses of the
axes are
going to be explained in Chapter 4.1 and the comparisons of
trend of
respective axes will be discussed in Chapter 5.2. The component
scores of
the major species are recorded in Appendix IV.
3.5. The Ratio of Planktonic to Benthic Diatom
Species (P:B ratio)
A graph describing P:B ratio has been constructed as well
(Figure
15). P:B ratio is a ratio of planktonic to benthic diatom
species. Planktonic
species are free floating species(not attached to a plant or
rock or bed);
therefore, they cannot usually thrive in shallow water comparing
to benthic
species. Actually, there are various conditions that planktonic
diatoms can
prosper besides water depth; for example, long ice-free season
is more
favorable to planktonic species than benthic species because ice
melts from
the littoral area(shallow water area) to the middle of the
water(deep water
area), so an ice-cover season is unfavorable to planktonic
diatoms (Wang et
al., 2012). However, the temperature in Seogwipo where Hanon is
located
-
27
does not fall less than 0°C (Figure 2), and thus
ice-free/ice-cover season is
not going to be considered significantly in this study.
Therefore, the P:B
ratio is likely to be related to the water depth of Hanon
paleolake; the graph
will be helpful to see the change of water depth according to
the change in
ratio between planktonic and benthic diatom species. The ratio
of
planktonic to benthic diatom species is calculated as below
(Wang et al.,
2013):
𝑃: 𝐵 𝑟𝑎𝑡𝑖𝑜 = ∑ 𝑝𝑙𝑎𝑛𝑘𝑡𝑜𝑛𝑖𝑐 𝑡𝑎𝑥𝑎
∑(𝑝𝑙𝑎𝑛𝑘𝑡𝑜𝑛𝑖𝑐 + 𝑏𝑒𝑛𝑡ℎ𝑖𝑐 𝑡𝑎𝑥𝑎)
A. ambigua, D. stelligera, and D. pseudostelligera are chosen
for
planktonic species. F. capucina var. mesolepta and Staurosirella
pinnata
are tychoplanktons; however, they are basically benthic species,
so these
species were not counted for planktonic taxa in the calculation
for the P:B
ratio. The result of P:B ratio is going to be show and explained
in Chapter
4.3, and it will be discussed in detail with the result of PCA
in Chapter 5.2.
-
28
Chapter 4. Research Results and Analysis
4.1. Principal Component Analysis (PCA)
Principal component analysis is carried out to condense all the
data
into two-dimensional presentation to show its major
representative
features by projection as explained previously (Janžekovič and
Novak,
2012). Above all, PCAs have been tried twice to see which one
would be
appropriate for analysis: one excluding “spp.” counting and the
other one
including “spp.” counting. The “spp.” counting in here means
counted
diatoms in genus level that were unable to be identified in
species level;
however, it does not mean that it is the sum of the genus level.
The reason
why this step is needed is to notice which one would be better
to interpret;
which one is easy to observe the meaning of principal
components.
Figure 6. PCA graphs with major diatom taxa (A) excluding “spp.”
and (B) including “spp.”
-
29
For principal component analyses, 17 and 26 diatom species
are
selected respectively in Figure 6: Figure 6-A excluding “spp.”
shows the
distribution of 17 species8 while Figure 6-B shows the
arrangement of 26
diatom species including “spp9.” Each axis in both graphs
represents the
same thing because both graphs are drawn from the same data.
Thus, it is
good to interpret only one of the graphs. Consequently, the
graph excluding
“spp.” is selected because it is plain enough to interpret the
meaning of axes
indicating information of environment/habitat/distribution in
species level.
Therefore, the simple version graph is better for interpretation
– easy to
find the tendency of the distribution of species.
From now on, the analysis on the results of PCA excluding “spp.”
is
going to be explained. The eigenvalues are essential to judge
how many axes
it should be considered for interpretation. As a result, four
axes are chosen
by referring to scree plot, and the eigenvalues are below (Table
1):
Table 1. Eigenvalues and variance explained by PCA of the diatom
species from core HN-1
8 Diadesmis Confervacea, Gomphonema parvulum, Eunotia incisa,
Planothidium
biporomum, Epithemia adnata, Cocconeis placentula, Cocconeis
placentula var. lineata, Aulacoseira ambigua, Fragilaria capucina,
Fragilaria capucina var. mesolepta, Discostella stelligera,
Staurosira construens var. venter, Staurosira construens,
Discostella pseudostelligera, Staurosirella leptostauron var.
dubia, Staurosirella leptostauron, and Staurosirella pinnata
(counterclockwise)
9 Pinnularia spp., Diadesmis confervacea, Eunotia spp., Eunotia
incisa, Gomphonema parvulum, Planothidium spp., Planothidium
biporomum, Epithemia adnata, Melosira spp., Epithemia spp.,
Cocconeis placentula, Gomphonema spp., Cocconeis placentula var.
lineata, Fragilaria spp., Aulacoseira ambigua, Fragilaria capucina
var. mesolepta, Navicula spp., Fragilaria capucina, Discostella
stelligera, Staurosira construens var. venter, Discostella
pseudostelligera, Staurosira construens, Staurosirella leptostauron
var. dubia, Staurosirella pinnata, Staurosirella leptostauron, and
Aulacoseira spp (counterclockwise)
-
30
The eigenvalues are 27.23% for Axis 1, 16.94% for Axis 2, 11.94%
for
Axis 3 and 9.64% for Axis 4, and their cumulative explained
variation is
65.74%. That is, the PCA explains 65.74% of the total variance
within the
first four axes, and the graphs of PCA are shown in Figure 710.
It is going to
be explained about what these axes represent respectively.
The graph of Axis 1 and 2 (Figure 7), S. construens var. venter
and
S. pinnata show the highest positive values, 0.7914 and 0.7952,
while C.
placentula and E. adnata show the highest negative values,
-0.6171 and
-0.654, along Axis 1. Considering S. construens var. venter and
S. pinnata
prefer mesotrophic (Fluin et al., 2010; Joh et al., 2010) while
C. placentula
and E. adnata prefer eutrophic (Joh, 2012; Kelly et al., 2005;
Round et al.,
1990; Krammer and Lange-Bertalot, 1988), Axis 1 can be
interpreted as a
representative of the nutrient status in the lake.
In the same way, D. confervacea and G. parvulum show highly
positive values of 0.794 and 0.5355, respectively, while A.
ambigua, F.
capucina var. mesolepta and C. placentula show highly negative
values
such as -0.6095, -0.6766 and -0.4939 along Axis 2, which is able
to observe
in the graph of Axis 2 and 3 in Figure 7. D. confervacea is a
benthic
species attached to plants (Jena et al., 2006; Torgan and
Santos, 2008). On
the other hand, C. placentula is also a benthic species but
attached to rocks
(Joh, 2012; Round et al., 1990; Kelly et al., 2005; Spaulding et
al., 2010), A.
ambigua is a planktonic species (Joh, 2010; Kelly et al., 2005;
Spaulding et
al., 2010), and F. capucina var. mesolepta is a tychoplanktonic
(Stoermer et
al., 1971). Therefore, Axis 2 seems to represent the water depth
in Hanon.
10 In Figure 7, the axis having smaller number is always
positioned on the
horizontal line; for example, Axis 1 is at horizontal line in
the graph of Axis 1 and 2.
-
31
E. incisa and F. capucina show the highest positive values such
as
0.7277 and 0.5623, while C. placentula var. lineata, E. adnata
and P.
biporomum show the highest negative values of -0.5039, -0.4923
and
-0.6173 along Axis 3, respectively (Figure 7 – Axis 1 and 3). E.
incisa prefers
an acidic condition (Oritz-Lerin and Cambra, 2007; Fukumoto et
al., 2012;
Flower et al., 1997), while the three diatom species with the
highest negative
values are alkaliphilous (Barbiero, 2000; Metcalfe et al., 1991;
Kelly et al.,
2005; Spaudling et al., 2010). Therefore, Axis 3 seems to be
related to the
pH of the lake.
For Axis 4 (Figure 7 – Axis 2 and 4), D. stelligera and D.
pseudostelligera show highly positive values of 0.6016 and
0.5853, while S.
construens and S. construens var. venter show highly negative
values of
-0.7213 and -0.3978. However, the Axis 4 was difficult to
interpret due to
the lack of habitat information. D. stelligera and D.
pseudostelligera have
very low sinking velocity, so they can persist throughout summer
thermal
stratification (Wang et al. 2012). S. construens and S.
construens var.
venter can increase in abundance as opportunistic diatoms
when
macrophytes are plentiful (Fluin et al., 2010). These species
are common
species all year around, especially from spring to summer (Kelly
et al., 2005;
Wang et al., 2012). Likewise, the information about these four
diatom
species does not correlate well, so it is hard to analyze what
Axis 4 reflects.
-
32
Axis 1 and 2 Axis 1 and 3
Axis 1 and 4 Axis 2 and 3
Axis 2 and 4 Axis 3 and 4
Figure 7. Principal component analysis of the HN-1 diatom
data
-
33
The interpretation of Axis 1, 2 and 3 is going to be examined
briefly
whether Axis 1, 2 and 3 indicates trophic status, water depth
and pH. The
principal components are compared to the change of several
diatom
assemblages. Several diatoms are plotted beside the diagrams of
each axis
(Figure 8), and the diatoms are compared to the axes based on
the ecology
of each diatom. In Figure 8, the diagrams of Axis 1, 2 and 3 are
10 times
enlarged to clarify their changes so as to compare other
variations precisely.
Also, the grey shadings in the diagram mean that the graph is
exaggerated
by three times.
First, C. placentula is very widely distributed except
oligotrophic
environment. Therefore, its fluctuation must be inversely
proportional to
the trend of Axis 1, and it seems they are in inverse proportion
to each other.
That is, it is right that Axis 1 indicates trophic status.
Second, Axis 2 is
compared to A. ambigua and D. confervacea. Because A. ambigua
is
planktonic while D. confervacea is epilithic/epiphytic, Axis 2
must be
proportional to D. confervacea and inversely proportional to A.
ambigua,
and so are they. Although it seems that the fluctuation of A.
ambigua does
not fit well to Axis 2, D. confervacea seems to match Axis 2
well. These
differences may have been caused to various environmental
factors such as
temperature, pH, trophic status, etc. Thus, it is possible to
say Axis 2
reflects water depth. Third, E. incisa and S. pinnata are
compared to
verify the interpretation of Axis 3 was right. Because E. incisa
reflects
acidification of lacustrine environment while S. pinnata is
alkaliphilous, E.
incisa must be proportional and S. pinnata must be inversely
proportional
to Axis 3. As a result, they are also harmonized with each
other. In
conclusion, the interpretations of Axis 1, 2 and 3 were right,
and there
would be no crucial problems/errors for further discussion.
-
34
Even though it was impossible to find out what Axis 4 means, it
is
still possible to reconstruct paleoenvironment of Hanon using
Axis 1, 2 and
3 by having interpretations so far. Additional graphs drawn by
components
of the first three axes are going to be shown in the next pages
(Figure 9 - 12).
These graphs are going to be discussed in detail to verify the
reconstructed
paleoenvironment by diatom diagram in Chapter 5.2.
Figure 8. Diagram for examination of Axis 1, 2 and 3 by
comparisons of several diatoms
Lastly, PCA graphs are used to rearrange the diatom species
in
diatom diagrams – gathering similar diatom assemblages together,
so this
-
35
analysis was also helpful to start solving a diatom diagram. The
component
scores of the Axis 1, 2, 3 and 4 are described in Appendix III,
and each
species score at Axis 1, 2, 3 and 4 is also reported in Appendix
IV.
-
36
Figure 9. The graphs of component scores on the PCA Axis 1, 2
and 3
-1.5
-1
-0.5
0
0.5
1
1.5
2
2.5
Axis 1
Axis 2
Axis 3
-
37
Figure 10. The graph of component scores on the PCA Axis 1
-1.5
-1
-0.5
0
0.5
1
1.5
2
2.5
Axis 1
-
38
Figure 11. The graph of component scores on the PCA Axis 2
-1.5
-1
-0.5
0
0.5
1
1.5
2
Axis 2
-
39
Figure 12. The graph of component scores on the PCA Axis 3
-1.5
-1
-0.5
0
0.5
1
1.5
2
Axis 3
-
40
4.2. Diatom Flora in the Diagram
In this chapter, diatom flora in the diagram is going to be
explained:
which species are dominant/rare, increase/decrease and
appear/disappear
(Figure 10). As explained in Chapter 3.3, the diatom assemblages
in the
stratigraphic diatom diagram are totally divided into five large
zones and
two subzones with zonation based on constrained incremental sum
of
squares cluster analysis, CONISS in Tilia software (Grimm, 1987)
(Figure 13
and 16). In the diagram, each diatom diagram is exaggerated by a
factor of
five with grey shadings, for a good grasp of each species’
fluctuations.
Original values are specified in black solid diagrams.
A total of 47 genera and 221 species have been identified in
81
samples with 2cm interval from Hanon paleo-maar lake sediment.
Diatoms
are present throughout the whole HN-1 core, and the composition
of
diatom species are getting very different from top to bottom in
the core.
As the depth is getting deeper, the sizes of diatoms are getting
smaller and
many broken valves found. Moreover, the diatom concentration at
the
bottom is very little degressively even though the concentration
is not low.
Also, genera Karayevia and Cavinula appear rarely in other
depths, but are
present in deeper parts of the core. On the other hand, the
sizes of diatoms
are getting larger and have distinguishable feature easy to
identify as the
depth is getting shallower. The degressive diatom concentration
is high
even though its tendency actually decreases as the depth gets
shallower; it
was hard to identify diatoms sometimes because they were
overlapped too
much due to its high concentration and density. It may be
connected to the
sedimentation rate in Hanon. In the upper depths, genera
Gomphonema,
-
41
Epithemia, Cocconeis and Diadesmis appear frequently. The upper
part of
the core is made up of a wide variety of diatoms, and they are
exhibited
evenly in general. The greater part of the species from Hanon
paleo-maar
lake consist of freshwater and fresh brackish species.
For the diagram in Chapter 4.2, the 26 species are selected
based
on their relative abundance which is larger than 5% frequency in
at least
one sample. Also, the “spp.” is included for Figure 13 because
it would be
good to try to examine the whole change of diatom assemblages
that were
counted originally. It is difficult to figure out the
paleoenvironmental
changes in detail based on the habitat information in genus
level because of
its ambiguity; therefore, the new diatom diagram which does not
contain
“spp.” assemblages will be discussed and interpreted in Chapter
5. Despite
of its difficulty, the diatom diagram including “spp.” is going
to be explained
from Chapter 4.1.1. to 4.1.6. for information for whom may be
interested. As
mentioned before, the order of arrangement of diatoms in the
diagrams was
referred to the result of PCA; PCA makes diatoms that have
common
environmental conditions gather. This makes it easy to
understand the
fluctuations in diatom assemblage over time and compare to the
other
changes of diatom assemblages changing in a similar or different
way. The
diagram showing the fluctuations of all species is in Appendix
I.
-
42
Figure 13. Diatom diagram including “spp.” – grey shading means
that it is enlarged by five times
-
43
4.1.1. Zone 1: 15,440 cal. yr BP – 14,670 cal. yr BP
(Oldest Dryas)
Zone 1 from 250cm to 236cm does contain diatoms, but quite a
few
diatoms in this zone are small and broken as mentioned before,
so it is hard
to identify them into species level. New species that do not
appear in upper
sediments are also observable. In particular, the sample at
250cm is very
difficult to identify diatoms due to the degressive low diatom
concentration
comparing to other depths.
As it is detectable in Figure 13, there are two main
dominant
species; Staurosira construens var. venter and Staurosirella
pinnata. They
reach approximately 50% together. Besides, Staurosira
construens,
Staurosirella leptostauron, Staurosirella leptostauron var.
dubia,
Discostella stelligera, Discostella pseudostelligera,
Aulacoseira spp., and
Navicula spp. are common in zone 1. The other species rarely
show up or
have very low relative abundance whose power of explanation
would be
poor for representing the change of environment in the past
around Hanon.
4.1.2. Zone 2-a: 14,670 cal. yr BP – 14,180 cal. yr BP
(The beginning of Bø lling-Allerø d)
Zone 2 between 236 and 220cm still consists of small
diatoms,
however, it is much better to identify than zone 1. Staurosira
construens
var. venter, the predominant species in zone 1, is still
abundant in zone 2-a.
Also, Staurosira construens, Aulacoseira ambigua, and
Cocconeis
placentula are dominant with about 70% in total abundance in
this zone.
-
44
Especially, C. placentula has begun to appear and increase
abruptly,
whereas S. pinnata decrease gradually.
4.1.3. Zone 2-b: 14,180 cal. yr BP – 12,810 cal. yr BP
(Bø lling-Allerø d)
The prevailing species in zone 2-b between 220 and 156cm from
the
surface of the earth of the core sediment are Cocconeis
placentula and
Eunotia incisa. C. placentula shows the maximum relative
abundance at
ca. 13,500 cal. yr BP throughout the core, which almost reaches
to 40% by
itself. E. incisa is present newly and increase until the midst
of zone 2-b
mainly. It is also told that Melosira spp. is dominant, and
Staurosira
construens var. venter decreases notably. Also, Aulacoseira
ambigua
maintains its abundance while Staurosirella pinnata keeps
decreasing. Also,
Eunotia spp. just appears.
4.1.4. Zone 3: 12,810 cal. yr BP – 12,150 cal. yr BP
(Younger Dryas)
First of all, Cocconeis placentula shows a declining trend,
albeit in
high relatively abundance. The prevalent species in zone 3
(156-136cm) are
Epithemia adnata, Planothidium biporomum, and Cocconeis
placentula
var. lineata by 15% of relative total abundance although their
percentage is
not that high. It is because the substantial shift in the diatom
species is
considered importantly. The actual dominant species with high
abundances
are Epithemia spp. and C. placentula. Melosira spp. and Eunotia
spp. are
-
45
still shown, but in decline. Navicula spp. keeps its relative
abundance
consistently throughout the whole core, HN-1. Pinnularia spp.
does similar
with Navicula spp., however, it will increase from zone 4 a
little bit.
4.1.5. Zone 4: 12,150 cal. yr BP – 10,440 cal. yr BP (Pre-
Boreal: the beginning of Holocene)
The predominant species in zone 4 (136-120cm) are Diadesmis
confervacea, Aulacoseira ambigua, Epithemia adnata, and
Planothidium
biporomum. Especially, D. confervacea appears for the first time
with high
relative abundance. Even though it shows a declining trend at
the end of
this zone but recovers shortly. Gomphonema parvulum and
Planothidium
spp. are common but quite dominant in this zone. Epithemia spp.
is also
prevailing by reaching its maximum relative abundance throughout
this
core, which is about 20%. Cocconeis placentula is also present
with high
abundance. There is an unusual abrupt increase and decrease
in
Staurosirella pinnata in the beginning of zone 4.
4.1.6. Zone 5: 10,440 cal. yr BP – 7,980 cal. yr BP
(Boreal)
The last zone, zone 5 covers sediments between 120 and 90cm. In
the
section, Diadesmis confervacea, Eunotia incisa, Staurosirella
pinnata, and
Staurosira construens is prevailing. Melosira spp. and
Pinnularia spp.
account for high relative abundance, too. It shows a pronounced
increase in
D. confervacea in zone 5. E. incisa increases after a while in
zone 5 begins
-
46
and maintains its richness until the end, which is the second
dominant
appearance for E. incisa throughout the core. Staurosira
construens var.
venter and Aulacoseira spp. are common species. There is two
striking
decreased species such as Cocconeis placentula and Epithemia
spp. Based
on the results of diatom diagram, the interpretation and
reconstruction of
paleoenvironment of Hanon in detail will be discussed in Chapter
5.1.
4.3. The Ratio of Planktonic to Benthic Diatom
Species (P:B ratio)
The graphs describing the changes of benthic and planktonic
diatoms have been drawn in Figure 14 and 15 based on the
calculation of
the equation explained in Chapter 3.5. Before starting looking
at the
fluctuation of P:B ratio in earnest, the graph in Figure 14 has
been drawn to
see the individual changes of planktonic and benthic species
during the last
deglaciation. There are remarkable changes in both diatom types:
abrupt
decrease in benthic diatoms and an increase in planktonic
diatoms
approximately at 13,730 and 13,780 cal. yr BP. Those are minimum
and
maximum points for each one in the graph. The slight age
discrepancy
between the two points is about 40 to 50 years. This could have
happened
because of the types of benthic diatom species such as
tychoplankton; all
the benthic species do not live in the same habitat. Some of
them live
attached to a rock or sand, whereas others live attached to
plant or lakebed.
All of these diatoms are called benthic species; however, they
do not
indicate directly shallow water depth even though they are
benthic. For
-
47
example, most benthic diatoms which is attached to a plant
inhabit shallow
water because plants usually inhabit littoral area for plant
photosynthesis
where light penetrates. On the other hand, another benthic
species attached
to a rock is able to inhabit much deeper water depth than the
one attached
to a plant because rocks run into a lakebed straightly caused by
runoff of
heavy rainfall – this can be related to precipitation.
Therefore, it is
necessary to search the habitat environments of the benthic
species for
more accurate interpretation. Overall, both benthic and
planktonic species
tend to decline slightly from 15,500 to 8,000 cal. yr BP.
Finally, the graph of the ratio change of planktonic to
benthic
species in Hanon maar paleolake during the last deglaciation has
been
constructed (Figure 15). Each change in planktonic and benthic
species is
combined by the equation mentioned in Chapter 3.5. The general
trend in
Figure 15 is similar to the one of planktonic species in Figure
14. As the
name of the graph is “planktonic to benthic,” it shows the ratio
change of
planktonic species comparing to benthic species (Figure 15).
Approximately
at 13,780 cal. yr BP, the graph is at the maximum, which means
the largest
abundance of planktonic species compared with benthic species in
Hanon
paleolake. The reason why planktonic diatoms thrived in this
period could
be connected to water depth, which is also related to the
strength of the
monsoon, sedimentation rate, and so on.
According to P:B ratio analysis, the diversity of planktonic
diatom
species in Hanon paleolake is much less than the one of benthic
species
during the last deglaciation. That is, it seems that the water
depth of Hanon
had been shallow in general from 15,500 to 8,000 cal. yr BP.
However, the
interpretation should be supplemented by considering
sedimentation rate,
strength of the monsoon, and other related environmental factors
to water
-
48
depth to reconstruct more accurate water depth. Last, the P:B
ratio graph
is also going to be interpreted in detail comparing to Axis 1,
2, 3
components and other proxy data of Hanon from Park et al.
(2014b) in
Chapter 5.3.
-
49
Figure 14. The respective changes of planktonic and benthic
species
-5
5
15
25
35
45
55
65
Planktonic
Benthic
-
50
Figure 15. P:B ratio in Hanon maar paleolake during the last
deglaciation
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
planktonic to benthic
-
51
Chapter 5. Discussion
The total length of core HN-1 is approximately 10m-long, and
the
range from 250 to 90cm of HN-1 is selected for this study to
observe the
environmental change during the last deglaciation, which covers
from
15,500 to 8,000 cal. yr BP – approximately 7,500 years among the
paleo-
maar lake history. The analysis has been performed every 2cm of
the
section. Totally, 47 genera and 221 species have been
enumerated
throughout the core. The AMS radiocarbon dates are adopted from
the
research by Park et al. (2014b), which had studied with the same
core
sediments.
In this chapter, the new version of diatom diagram is going to
be
interpreted. In Chapter 4, the diatom diagram included “spp.,”
but the one
in Figure 16 does not contain “spp.” As explained previous
chapter, it is
difficult to interpret and reconstruct paleoenvironment based on
the genus
level of diatoms because diatoms in genus level cover quite a
wide range of
inhabited environment. This is why most diatom analysis has been
being
performed in species level. Through the interpretation of the
diatom
diagram, the paleoenvironment in Hanon during the last
deglaciation
would be reconstructed in Chapter 5.1.1. Based on the
reconstructed
information, a schematization has been done in Chapter 5.1.2.
Also, there
will be a few more analyses(verification) using principal
components and
P:B ratio in Chapter 5.2. Last, comparative analysis with other
research in
Hanon, especially Park et al. (2014b), is going to be discussed
in Chapter
5.3. The diagram delineating all species is in Appendix I, and
counting data
of major species are in Appendix II.
-
52
5.1. Reconstructing the Paleoenvironment of
Hanon Maar based on the Diatom Diagram and
Schematization
5.1.1. Reconstructing the Paleoenvironment of Hanon
Maar through schematization of each zone based
on Diatom Diagram
To reconstruct the detailed environmental changes in Hanon, it
is
essential to interpret the diatom diagram. A new graph is drawn
to show the
transition of diatom assemblage lucidly according to temporal
trend (Figure
16). In Chapter 5.1.1., schematic diagrams are going to be drawn
(Chen et al.,
2014) and interpreted for a diatom-based reconstruction of
the
paleoenvironmental changes in Hanon. The reconstructed
lacustrine
environment based on schematic diagram will be summarized
and
schematized again for better vision.
The values of diatom concentration are shown in Figure 16.
The
total amount of diatoms seems to decrease gradually from the
bottom to the
top of the core according to the transition of absolute number
of diatom
valves per 1g of dry sample, which varies from 6.95×109 to
8.64×109
[valves]. The total amount of valves is quite a lot but normal.
According to
Katsuki et al. (2012), the absolute number of valves from Lagoon
Notoro-Ko
in northern Japan also had similar amount, which varies from
0.5×109 to
2.5×109; therefore, the absolute number of diatom valves in
Hanon are not
at abnormal levels. According to diatom concentration from Hanon
data,
the lake productivity had decreased during the last
deglaciation, and it
-
53
reached to the maximum at BA. It means that the climate during
BA was
wet and warm, and the trophic status was good. However, the
concentration
during Holocene maintained relatively low, and it means the
climate was
not warm and/or wet. These should be checked later with other
results of
this study. The diagram of diatom concentration is in Figure 13
and 16.
-
54
Figure 16. Diatom diagram excluding “spp.” – grey shading means
that it is enlarged by five times
-
55
Throughout the Chapter 5.1 from now on, there will be the
interpretation of diatom diagram with some schematizations for
better
understanding (Figure 17 – 22). The information is based on
lacustrine
environment concerning surrounding factors around a lake such
as
monsoon, vegetation, and so on. The schematic diagrams are
created by
referring to Chen et al. (2014). Before starting to look at the
schematic
diagrams, there will be an unfamiliar term – saprobity values.
The values
specify the levels of dissolved oxygen and Biological oxygen
deman