Embed Size (px)
Citation preview

DNK detektivi
(seminar)
Martin Gorjan
mentor: dr. Ales Omerzu∗
somentor: doc. dr. Vita Dolzan†
10. november 2005
Povzetek
Skoraj vsaka celica v cloveskem telesu vsebuje identicno molekulo DNK, te pa se medposamezniki na nekaterih delih (priblizno 1% celotnega zapisa) razlikujejo. S pomocjoanalize kratkih odsekov DNK (fragmentov STR), ki se med posamezniki razlikujejo podolzini, je mozna zelo natancna identifikacija posamezne osebe. Uporaba pomnozevalnereakcije PCR omogoca analizo izjemno majhne kolicine vzorca tkiva, ki poteka s pomocjokapilarne gel elektroforeze. Opisane tehnologije pa niso pomembne samo za forenzicnoanalizo, pac pa so temeljnega pomena za molekularno biologijo in vse genske tehnologije.
∗Institut Jozef Stefan†Medicinska Fakulteta

Kazalo
1 Uvod 3
2 Zajem in ekstrakcija DNK 32.1 Zajem vzorca tkiva . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.2 Ekstrakcija DNK . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
3 Analiza DNK 53.1 Struktura DNK . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53.2 Kodiranje informacij v DNK (STR) . . . . . . . . . . . . . . . . . . . . . . . 63.3 Polimerazna verizna reakcija (PCR) . . . . . . . . . . . . . . . . . . . . . . . 8
3.3.1 Biokemicne reakcije . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83.3.2 Potek polimerazne verizne reakcije . . . . . . . . . . . . . . . . . . . . 9
3.4 Kapilarna gel elektroforeza . . . . . . . . . . . . . . . . . . . . . . . . . . . . 113.4.1 Teorija elektroforeze . . . . . . . . . . . . . . . . . . . . . . . . . . . . 113.4.2 Teorija gel elektroforeze polielektrolitov . . . . . . . . . . . . . . . . . 13
4 Identifikacija 15
2

1 Uvod
Petdeset let je odkar sta Watson in Crick razvozlala 3D strukturo dvojne vijacnice deoksiri-bonukleinske kisline (DNK) [1], ki je nosilec genetskega materiala vseh znanih zivih bitij [2].S tem so prej teoreticni geni dobili tudi fizicno obliko in sledila so leta intenzivnega razisko-vanja te molekule in njene biokemije. Z odkritjem in uporabo encimov restrikcijskih nukleaz[3], ki selektivno rezejo dolgo molekulo DNK na manjse dele ter kmalu nato se odkritjempolimerazne verizne reakcije (PCR) [4], se je v osemdesetih letih zacel pravi razcvet genetskihtehnologij. Leta 1986 so v Angliji ze obsodili prvega kriminalca na podlagi identifikacije zgenetsko analizo [5], zadnjih deset let pa je postala uporaba DNK v forenzicne namene dokajrutinska in zelo uspesna.
Celoten postopek sestavlja vec korakov, in danes uporabljani postopki se nekoliko ra-zlikujejo od prvih izpred dvajsetih let. V nadaljevanju je opisan celotni postopek sodobneforenzicne DNK analize (strokovno imenovane PCR/STR): od zajema vzorca tkiva in ekstrak-cije molekule DNK iz tkiva, prek opisa kodiranja za forenzicno analizo pomembnih informacijv molekuli DNK, pomnozevanje STR fragmentov DNK, ugotavljanje velikosti teh fragmen-tov s pomocjo kapilarne gel elektroforeze in koncno ovrednotenje rezultatov ter (morebitna)identifikacija.
Kljub veliki komercializaciji in avtomatizaciji je postopek se vedno precej drag (cena eneanalize je reda velikosti 100.000 SIT), v veliki meri tudi na racun dragih patentiranih kemikalij,predvsem barvil in fotoaktivnih snovi, potrebnih za zaznavo DNK pri koncni analizi. Ravnona tem podrocju je tudi za fizike se dovolj manevrskega prostora za odkritja novih postopkovzaznavanja DNK fragmentov in s tem znizanja cene ter posledicno povecanja dostopnostiforenzicne DNK analize.
2 Zajem in ekstrakcija DNK
2.1 Zajem vzorca tkiva
Molekula DNK se nahaja v celicnem jedru vecine celic, ki sestavljajo clovesko telo, polegtega so krajse krozne molekule DNK tudi v celicnem organelu mitohondriju (mtDNK) [2].Obe sta primerni za DNK analize, vendar s pomembno razliko: le jedrna DNK omogocaidentifikacijo posameznika, saj ima vsaka oseba (z izjemo enojajcnih dvojckov) edinstvenojedrno DNK, mitohondrijska DNK pa se podeduje le od matere in je zato identicna medmatero in otroci. Kot taka je primerna le za ugotavljanje sorodstvenih razmerij, odlikuje pajo tudi manjsa verjetnost degradacije. mtDNK se uporablja predvsem za identifikacijo drugacenerazpoznavnih trupel naravnih in drugih nesrec. Obe vrsti DNK pa se ne spreminjata medcelicno delitvijo (mitozo), kar pomeni, da imajo vse telesne celice posameznika (z izjemorelativno majhnega stevila mutacij, ki pa ne motijo te vrste analize) identicno DNK. Zapridobitev DNK so torej primerne skoraj vse telesne celice [6].
Za sodobni postopek analize zadostuje ze izredno majhna kolicina DNK: zadostuje zezajem nekaj deset celic z jedrom [7]. V Tabeli 1 je predstavljenih nekaj najpogostejsih tipovcelic, puscenih na kraju zlocina in pogosta mesta njih nahajanja [8]. Povrsino, kjer se nahajajote celice, drgnejo z bombazno krpico, s katere jih nato sperejo z destilirano vodo. Dobljenecelice shranijo v posebne papirne vrecke, kjer se posusijo in pri sobni temperaturi zdrzijodo dveh let. Ce nameravajo hraniti vzorce vec kot dve leti, jih je priporocljivo zamrznitina −20o [9]. DNK je precej stabilna molekula; k nje degradaciji (DNK razpada na krajse
3

tip celic mesto nahajanjakozna povrhnjica kapa, ocala, orozjeslina uporabljena cigareta, pisemska ovojnicabele krvnicke orozje, krogle, umazano perilosperma odeja, uporabljen kondom, umazano perilolasna korenina kapa, odeja, obleka
Tabela 1: Najpogostejsi tipi celic za DNK analizo in njih nahajalisca.
dele) najbolj pripomorejo vrocina in vlaga (pospesujeta rast mikrobov, ki razgrajujejo DNK,visja temperatura pa sama po sebi povzroca vecjo molekularno ”zivahnost”in s tem vecjoverjetnost za degenerativne kemicne reakcije na DNK, kot je npr. hidroliza - odcep vode inzlom fosfatnega ogrodja (glej 3.1)) ter soncni zarki (vidna svetloba in UV). Zelo pomembnodejstvo pa je, da degradacija DNK zgolj zmanjsuje kolicino dosegljive informacije o identiteti,ne povzroca pa spremembe te informacije [6]. Po zajemu (domnevnega) vzorca tkiva, najprejpreverijo, ce je to res bioloski vzorec (in ne denimo packa kecapa ali loscila za cevlje) inali vsebuje dovoljsnjo kolicino dovolj kvalitetne DNK (dovolj veliki fragmenti) za nadaljnjoanalizo. Z uvedbo sodobne PCR/STR analize, ki je opisana v nadaljevanju, sta se obe zahteviv primerjavi s predhodnimi metodami znizali za priblizno dva velikostna reda, tako da vecinalaboratorijev niti ne izvaja vec testov o kolicini in kvaliteti DNK [6].
2.2 Ekstrakcija DNK
Telesne celice sesalcev so zelo kompleksne strukture: v najvecji meri jih sestavlja voda (70%celotne mase celice), sledijo proteini (20%), ostali gradniki kot so ioni, aminokisline (monomeriproteinov), sladkorji (monomeri polisaharidov), mascobne kisline in posamezni nukleotidi(monomeri DNK in RNK) prispevajo k masi celice relativno malo. Molekula DNK predstavljale okoli 0.25% celotne celicne mase [2], kar znasa okoli 6 pg [7]. Celice, ki v povprecju merijookoli 10 µm, obdaja plazemska membrana iz fosfolipidov, v notranjosti celice (citoplazma)pa plavajo mnogi celicni organeli ter celicno jedro, ki ga zopet obdaja membrana. Celicnojedro pri sesalcih zavzema okoli 10% prostornine celice, celotna dolzina molekule DNK pa jepri cloveku okoli 2 m, kar omogoca zelo spretno ”pakiranje” le-te. Clovek in druga diploidnabitja imajo dve razlicni molekuli DNK, eno od matere in eno od oceta. Vsaka je razdeljenana 23 delov imenovanih kromosomi. DNK, ki je negativo nabita, je ovita okoli pozitivnonabitih proteinov histonov, le-ti pa sestavljajo terciarno strukturo - kromatinsko vlakno, kiima premer okoli 30 nm. To vlakno je razpredeno znotraj celicnega jedra in celotno vlaknastostrukturo imenujemo kromatin. Ob celicni delitvi se kromatin s pomocjo tvorbe zankic sedodatno zgosti v kromosome, ki so vidni ze z opticnim mikroskopom. Poleg histonov obdajaDNK se kopica drugih proteinov, ki sluzijo kot prepisovalci informacij na RNK, aktivaciji alirepresiji genov, in se mnogo cem. Celotno DNK v celicnem jedru imenujemo genom. [2].
Za DNK analizo bi se radi znebili vseh teh proteinov, mascob, ionov itd. in dobili cimcistejso raztopino DNK (DNK je zelo dobro topna v vodi). Obstaja vec postopkov za ek-strakcijo DNK iz celic oz. tkiva, in njih izbira je odvisna od tipa celic in kolicine ter stanjavzorca. Vsi pa temelijo na razgradnji celicne stene (lizija), in izolaciji nezelenih komponent izraztopine, kar dosezejo s povecano temperaturo in dodatkom razlicnih kemikalij. Ena takihje detergent natrijev dodecil sulfat, ki razgradi celicno in jedrsko membrano ter denaturira
4
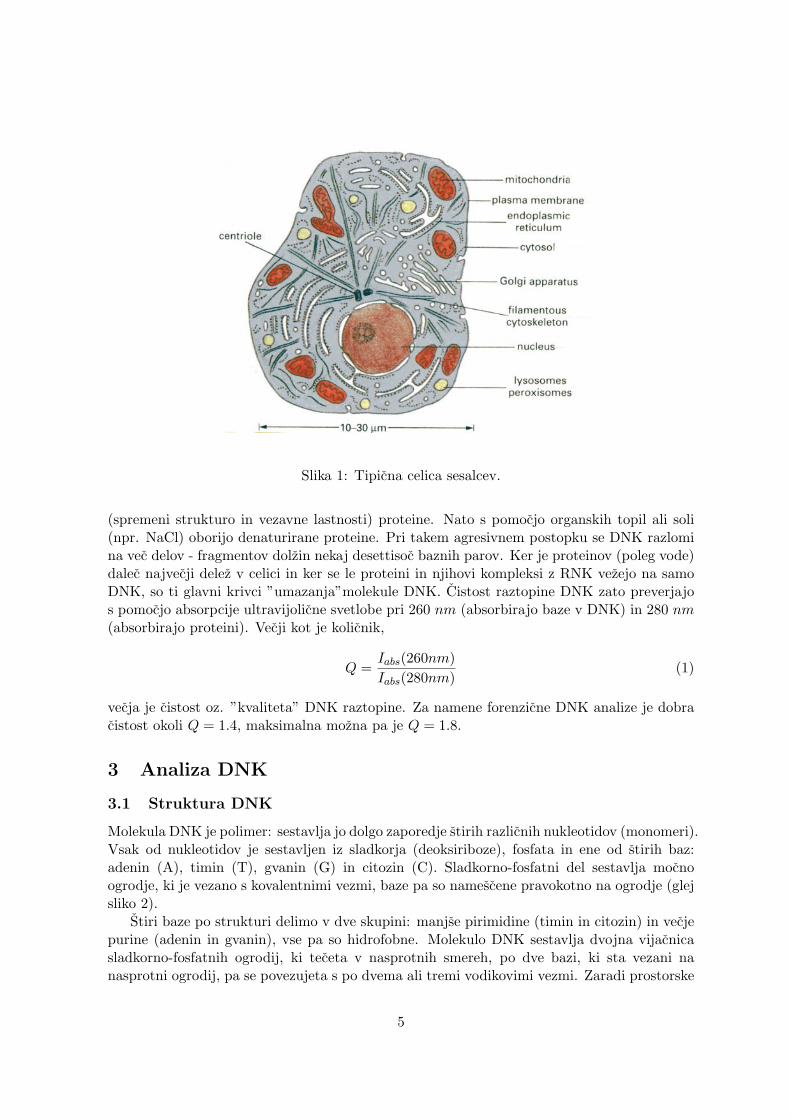
Slika 1: Tipicna celica sesalcev.
(spremeni strukturo in vezavne lastnosti) proteine. Nato s pomocjo organskih topil ali soli(npr. NaCl) oborijo denaturirane proteine. Pri takem agresivnem postopku se DNK razlomina vec delov - fragmentov dolzin nekaj desettisoc baznih parov. Ker je proteinov (poleg vode)dalec najvecji delez v celici in ker se le proteini in njihovi kompleksi z RNK vezejo na samoDNK, so ti glavni krivci ”umazanja”molekule DNK. Cistost raztopine DNK zato preverjajos pomocjo absorpcije ultravijolicne svetlobe pri 260 nm (absorbirajo baze v DNK) in 280 nm(absorbirajo proteini). Vecji kot je kolicnik,
Q =Iabs(260nm)Iabs(280nm)
(1)
vecja je cistost oz. ”kvaliteta” DNK raztopine. Za namene forenzicne DNK analize je dobracistost okoli Q = 1.4, maksimalna mozna pa je Q = 1.8.
3 Analiza DNK
3.1 Struktura DNK
Molekula DNK je polimer: sestavlja jo dolgo zaporedje stirih razlicnih nukleotidov (monomeri).Vsak od nukleotidov je sestavljen iz sladkorja (deoksiriboze), fosfata in ene od stirih baz:adenin (A), timin (T), gvanin (G) in citozin (C). Sladkorno-fosfatni del sestavlja mocnoogrodje, ki je vezano s kovalentnimi vezmi, baze pa so namescene pravokotno na ogrodje (glejsliko 2).
Stiri baze po strukturi delimo v dve skupini: manjse pirimidine (timin in citozin) in vecjepurine (adenin in gvanin), vse pa so hidrofobne. Molekulo DNK sestavlja dvojna vijacnicasladkorno-fosfatnih ogrodij, ki teceta v nasprotnih smereh, po dve bazi, ki sta vezani nanasprotni ogrodij, pa se povezujeta s po dvema ali tremi vodikovimi vezmi. Zaradi prostorske
5

Slika 2: Prostorska zgradba dvojne vijacnice DNK.
strukture baz sta mozna le para adenin - timin (dve vodikovi vezi) in citozin - gvanin (trivodikove vezi) [1]. Ravno ta lastnost omogoca kodiranje informacij v DNK, zaradi komple-mentarnosti pa omogoca tudi kopiranje teh informacij in tako pomnozevanje molekule (pricelicni delitvi, prepisovanju gena na RNK, PCR, itd.).
Celotna DNK pri cloveku vsebuje okoli tri miljarde baznih parov (ljudje pa imamo podve razlicni DNK). Le okoli 1% celotne DNK kodira gene (pri cloveku je teh okoli 60.000), kivsebujejo zapis za izdelavo proteinov in encimov (proteinski katalizatorji biokemicnih reakcij),preko teh pa izgradnjo in krmiljenje vecjega dela celicnih struktur in funkcij. Vseh funkcijostalega dela DNK, imenovanega tudi ”odvecna DNK”, se ne poznamo v celoti [2, 7, 9], meddrugim pomagajo pri oblikovanju sekundarne in terciarne strukture DNK (glej 2.2). Del DNK,ki kodira gene (kodirajoci del) je zelo obcutljiv na mutacije, ki so vecinoma destruktivne, inze sprememba enega samega baznega para lahko pripelje do neuspesnega razvoja nove celice.Posledica tega je, da je na kodirajocem delu zelo malo variacij med razlicnimi osebami. Celicaima posebne mehanizme (sklopi encimov), ki odpravljajo mutacije na DNK. Taka popravila somozna, ker je informacija zaradi dvojne vijacnice zapisana dvakrat. Ce se spremeni ena baza(kot posledica delovanja mutagenih kemikalij, UV svetlobe, itd.), obstaja velika verjetnost,da je njena parna baza ostala nespremenjena [2].
3.2 Kodiranje informacij v DNK (STR)
V nekodirajocem delu pa so prisotni tudi odseki DNK, na katerih se ponavlja enostavnasekvenca dolzine 2-6 baznih parov, celotna dolzina takih sekvenc pa je 50-1000 baznih parov.Ti odseki, imenujemo jih STR (Short Tandem Repeat), nimajo pomembne funkcije, in celicnimehanizmi za popravila DNK na njih ne odpravljajo mutacij, zato se na njih ohranjajo vveliko vecjem stevilu [2]. Doloceno lokacijo na DNK imenujemo lokus, dejanski zapis nalokusu pa imenujemo alel. Kadar sta v populaciji na enem lokusu prisotna vsaj dva alela in
6
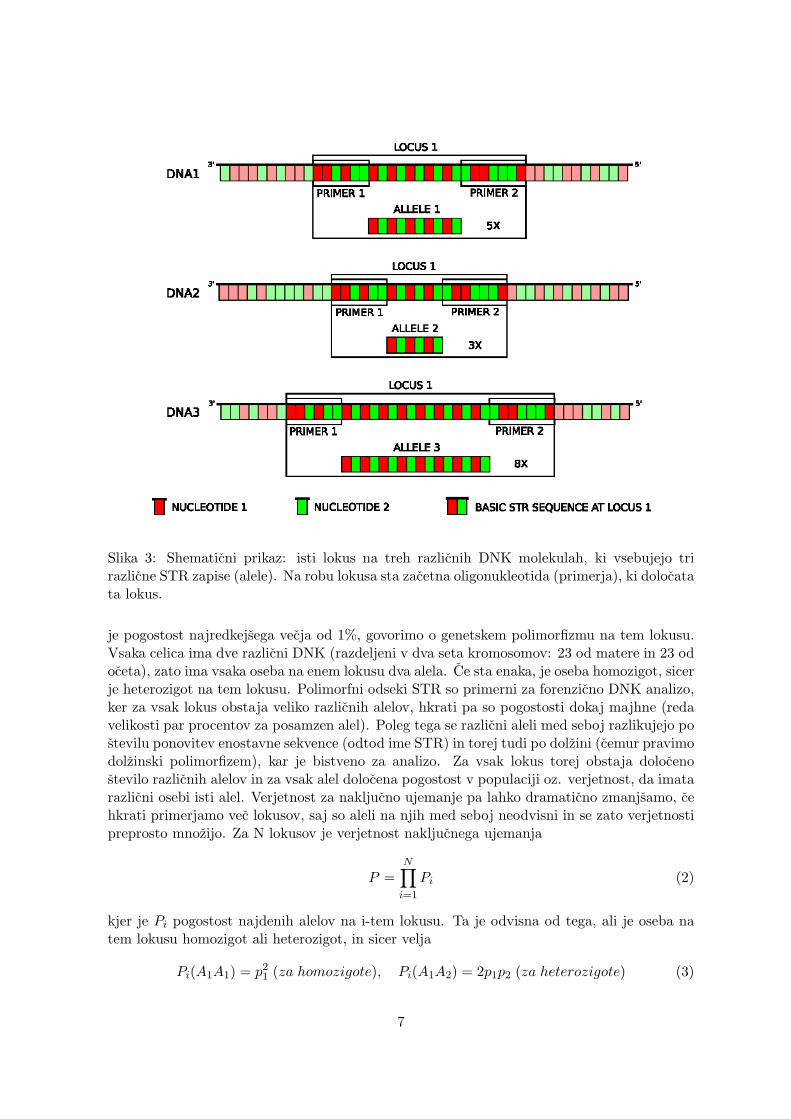
Slika 3: Shematicni prikaz: isti lokus na treh razlicnih DNK molekulah, ki vsebujejo trirazlicne STR zapise (alele). Na robu lokusa sta zacetna oligonukleotida (primerja), ki dolocatata lokus.
je pogostost najredkejsega vecja od 1%, govorimo o genetskem polimorfizmu na tem lokusu.Vsaka celica ima dve razlicni DNK (razdeljeni v dva seta kromosomov: 23 od matere in 23 odoceta), zato ima vsaka oseba na enem lokusu dva alela. Ce sta enaka, je oseba homozigot, sicerje heterozigot na tem lokusu. Polimorfni odseki STR so primerni za forenzicno DNK analizo,ker za vsak lokus obstaja veliko razlicnih alelov, hkrati pa so pogostosti dokaj majhne (redavelikosti par procentov za posamzen alel). Poleg tega se razlicni aleli med seboj razlikujejo postevilu ponovitev enostavne sekvence (odtod ime STR) in torej tudi po dolzini (cemur pravimodolzinski polimorfizem), kar je bistveno za analizo. Za vsak lokus torej obstaja dolocenostevilo razlicnih alelov in za vsak alel dolocena pogostost v populaciji oz. verjetnost, da imatarazlicni osebi isti alel. Verjetnost za nakljucno ujemanje pa lahko dramaticno zmanjsamo, cehkrati primerjamo vec lokusov, saj so aleli na njih med seboj neodvisni in se zato verjetnostipreprosto mnozijo. Za N lokusov je verjetnost nakljucnega ujemanja
P =N∏
i=1
Pi (2)
kjer je Pi pogostost najdenih alelov na i-tem lokusu. Ta je odvisna od tega, ali je oseba natem lokusu homozigot ali heterozigot, in sicer velja
Pi(A1A1) = p21 (za homozigote), Pi(A1A2) = 2p1p2 (za heterozigote) (3)
7

A1 in A2 sta alela na opazovanem lokusu, p1 in p2 pa njuni pogostosti v populacij [5]. AmeriskiFBI (Federal Bureau of Investigation) npr. uporablja 13 lokusov, ki so porazdeljeni po skorajvseh kromosomih. Imenuje jih ”core” lokusi; verjetnost za nakljucno ujemanje je za ta izborlokusov manjsa kot 10−10 [10]. Britanska FSS (Forensic Science Service) uporablja se veclokusov in verjetnost za nakljucno ujemanje je okoli 10−15 [7].
3.3 Polimerazna verizna reakcija (PCR)
Za uspesno forenzicno analizo DNK potrebujemo le okoli 50 pg vzorca, za kar zadostujeze par celic [7], vendar je ta kolicina veliko premajhna za neposredno analizo fragmentovSTR. Najprej moramo pomnoziti stevilo molekul DNK, kar nam omogoca postopek imenovanpolimerazna verizna reakcija (Polymerase Chain Reaction ali PCR). Ta ponavadi poteka vmikro epruvetah (volumna nekaj µg) in je v bistvu ena od mnogih biokemijskih reakcij, ki seodvijajo v zivih celicah, od kjer so tudi izolirali encim polimeraze. Celotna biokemija DNKtemelji na lastnostih kemijskih vezi v vodi: mocne kovalentne vezi, ki so za vec redov velikostivecje od termicne energije kT pri sobni temperaturi, so nosilci informacije (veriga nukleotidovoz. enojna vijacnica1) in posledicno je informacija zelo stabilna. Ostale vezi, kot so ionske,vodikove in van der Waalsove, so reda velikosti termicne energije, in omogocajo dostop doinformacije (dve enojni vijacnici se povezujeta v dvojno vijacnico2 preko vodikovih vezi medbazami DNK). Termicna energija kT znasa pri sobni ali telesni temperaturi priblizno 2.5kJ/mol, energije ostalih vezi so v Tabeli 2 [2].
tip vezi E v vakuumu [kJ/mol] E v vodi [kJ/mol]kovalentna 375 375ionska 333 12.5vodikova 16.7 4.2van der Waalsova 0.4 0.4
Tabela 2: Energije kemijskih vezi v vodi in v vakuumu; kT = 2.5 kJ/mol.
3.3.1 Biokemicne reakcije
Encimi so biomakromolekule, ki zaradi svoje zgradbe selektivno katalizirajo dolocene biokemicnereakcije s sklapljanjem dveh ali vec reakcij, od katerih je katalizirana reakcija energijsko neu-godna (∆G1 > 0), druga pa energijsko bolj ugodna, kot je prva neugodna (∆G2 < −∆G1)[2]. Snov, ki je v celici (in pri PCR) nekaksen univerzalen nosilec proste energije, je adenozintrifosfat (ATP). S hidrolizo (odcepom H2O) kovalentne vezi na fosforju ATP, dobi sistemokoli 54 kJ/mol energije. Pri polimerizaciji DNK poteka se daljsa reakcijska pot, ki vodipreko adenozin monofostata (AMP) in pirofosfata (PPi) ter prinese okoli 108 kJ/mol en-ergije, ki se nato porabi za tvorbo kovalentne vezi med ze obstojecim delom verige ter novimnukleotidom; po tej poti dobimo v splosnem iz snovi A-H in B-OH snov A-B po naslednjih
1ang. single-stranded DNK ali ssDNK2ang. double-stranded DNK ali dsDNK
8

kemicnih reakcijah
B −OH + ATP → B −O − PO3 − PO3 + AMPA−H + B −O − PO3 − PO3 → A−B + PPi
PPi + H2O → 2Pi
(4)
Nukleotidi v posodi PCR nosijo s seboj potrebno energijo za njihovo polimerizacijo (cemurpravimo repna polimerizacija), saj so v obliki deoksinukleotidnega trifosfata (dNTP). Dejan-ska reakcija polimerizacije je
(dNMP )n + dNTP → (dNMP )n+1 + PPi (5)
Od deoksinukleotidnega trifosfata se odcepi pirofosfat, kar da energijo za vezavo nukleotidana ze obstojeco verigo deoksinukleotid monofosfatov (z drugimi besedami je to ssDNK) in stem podaljsanje le-te. Graficni prikaz opisanega dogajanja je na sliki 5.
Slika 4: Polimerizacija DNK.
3.3.2 Potek polimerazne verizne reakcije
Polimeraza pa ne more zaceti sinteze nove ssDNK samo iz prostih nukleotidov (v obliki dNMP)in komplementarne podlage (na zacetku je to genomska ssDNK), pac pa rabi se nek kratek(nekaj baznih parov) zacek nove ssDNK verige vezan na podlago. Te kratke ssDNK molekule(imenujemo jih zacetni oligonukleotidi3) moramo dodati v epruveto pred zacetkom reakcije.
3gr. oligo pomeni malo
9
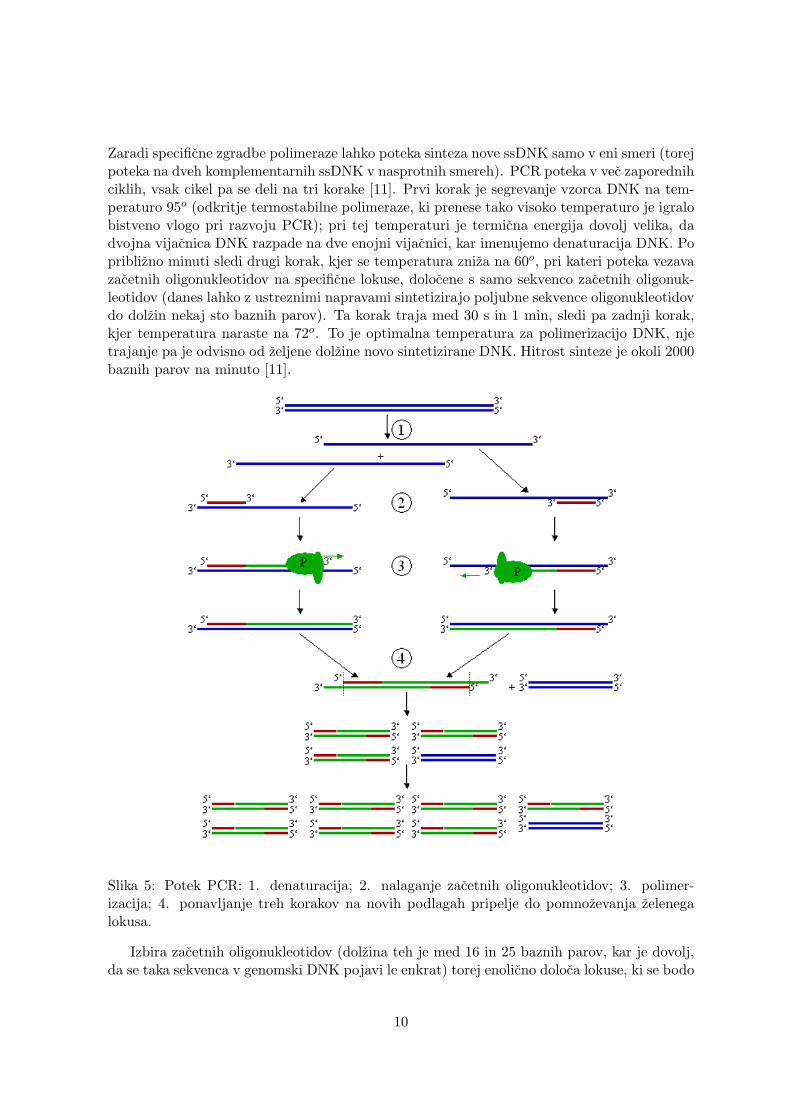
Zaradi specificne zgradbe polimeraze lahko poteka sinteza nove ssDNK samo v eni smeri (torejpoteka na dveh komplementarnih ssDNK v nasprotnih smereh). PCR poteka v vec zaporednihciklih, vsak cikel pa se deli na tri korake [11]. Prvi korak je segrevanje vzorca DNK na tem-peraturo 95o (odkritje termostabilne polimeraze, ki prenese tako visoko temperaturo je igralobistveno vlogo pri razvoju PCR); pri tej temperaturi je termicna energija dovolj velika, dadvojna vijacnica DNK razpade na dve enojni vijacnici, kar imenujemo denaturacija DNK. Popriblizno minuti sledi drugi korak, kjer se temperatura zniza na 60o, pri kateri poteka vezavazacetnih oligonukleotidov na specificne lokuse, dolocene s samo sekvenco zacetnih oligonuk-leotidov (danes lahko z ustreznimi napravami sintetizirajo poljubne sekvence oligonukleotidovdo dolzin nekaj sto baznih parov). Ta korak traja med 30 s in 1 min, sledi pa zadnji korak,kjer temperatura naraste na 72o. To je optimalna temperatura za polimerizacijo DNK, njetrajanje pa je odvisno od zeljene dolzine novo sintetizirane DNK. Hitrost sinteze je okoli 2000baznih parov na minuto [11].
Slika 5: Potek PCR: 1. denaturacija; 2. nalaganje zacetnih oligonukleotidov; 3. polimer-izacija; 4. ponavljanje treh korakov na novih podlagah pripelje do pomnozevanja zelenegalokusa.
Izbira zacetnih oligonukleotidov (dolzina teh je med 16 in 25 baznih parov, kar je dovolj,da se taka sekvenca v genomski DNK pojavi le enkrat) torej enolicno doloca lokuse, ki se bodo
10

pri PCR pomnozili. Po koncanem prvem ciklu se kolicina DNK na izbranih lokusih podvoji,dejanski novo sintetizirani fragmenti (za vsak lokus po en komplementarni par) pa so daljsiod samega STR dela (dolzino doloca cas pomnozevanja). Pri drugem ciklu predstavljatapodlago genomska DNK in novo sintetizirani fragmenti. Iz genomske podlage dobimo zopetenake fragmente kot pri prvem ciklu, iz fragmentov s prvega cikla pa dobimo, zaradi dolocenesmeri sinteze, fragmente, ki vsebujejo samo STR in zacetno sekvenco oligonukleotidov naobeh straneh. Kolicina predolgih STR fragmentov, ki niso dobri za koncno analizo rastelinearno s stevilom ciklov, kolicina pravih STR fragmentov pa raste (na zacetku) eksponentnokot 2N , ce je N stevilo koncanih ciklov. PCR pa ne more potekati v nedogled. Vse tereakcije so ravnotezne reakcije in zato odvisne tako od kolicine reaktantov kot produktov.Z manjsanjem stevila reaktantov se z vsakim ciklom zmanjsuje tudi ucinkovitost PCR. Kzmanjsanju ucinkovitosti prispeva tudi rekombinacija produktnih fragmentov in razpadanjeencima polimeraze, ki ima razpolovno dobo priblizno 30 min na 95o. V praksi to pomenizgornjo mejo nekje med 30 in 40 cikli, kar se vedno prinese vecmiljonsko pomnozitev izbranihSTR lokusov. [11]
3.4 Kapilarna gel elektroforeza
Pri koncni analizi ugotavljamo velikost s PCR pomnozenih fragmentov STR. Danes najboljsain najbolj razsirjena metoda je kapilarna gel elektroforeza (gel se nahaja v kapilari), ki potre-buje relativno malo vzorca (nekaj pikolitrov) in je lahko skoraj popolnoma avtomatizirana.Osnova kapilarne gel elektroforeze je gel elektroforeza, kjer vzorec (v nasem primeru fragmentissDNK, uporablja pa se jo se za druge polielektrolite npr. proteine) pod vplivom elektricnesile potujejo po gelu. Geli so porozne snovi: dolga vlakna gela so skoraj nakljucno razpore-jena po prostoru in tvorijo ti. matrico, med luknjami, ki jih tvorijo prekrizana vlakna, pase nahaja topilo (vodna raztopina pufrov in drugih snovi) [12]. Za analizo fragmentov STRje najbolj primeren poliakrilamidni gel, uporablja pa se tudi agar (predvsem za daljse frag-mente) [2, 6, 13]. Ker je molekula (v bistvu makroion) DNK negativno nabita (naboj 1e nanukleotid se nahaja na enem od kisikov na fosfatu [2]), jo elektricno polje vlece proti anodi,gel in topilo pa predstavljata upor, ki je odvisen od velikosti fragmenta. Posledica je razlicnagibljivost fragmentov DNK, ki je funkcija njihove dolzine. Ce damo torej raztopino vzorcafragmentov DNK na eno stran gela in nanj prikljucimo elektrodi pod elektricno napetostjo(nekaj kV), se bodo scasoma fragmenti DNK razporedili po velikosti.
3.4.1 Teorija elektroforeze
Nabiti delci (fragmenti DNK) v mediju z ioni (pufrska vodna raztopina) zmotijo porazdelitevionov medija. Nova porazdelitev je dolocena z ravnovesjem elektrostaticnih in Brownovih sil.Medij razdelimo na dve obmocji: bliznje (izhodisce je nabit delec), kjer so ioni medija stalno”absorbirani”na delec in so nemobilni, ter na ostalo obmocje z mobilnimi ioni. Obmocje znemobilnimi ioni se imenuje Sternova plast, obmocje z mobilnimi difuzna plast, obe skupajpa elektricna dvojna plast4 [13]. Meja med tema plastema je nekako slabo dolocena in jeodvisna tudi od casovne skale. Priblizno jo podaja Bjerrunova enacba [13]
lB =e2
4πεmε0kT(6)
4ang. electric double layer
11

Slika 6: Shema kapilarne gel elektroforeze.
kjer je εm dielektricna konstanta medija. lB je reda velikosti nm, v vodi je pri sobni temper-aturi 0.7nm. V teoriji povprecnega polja je ravnovesna porazdelitev mobilnih ionov podanas Poisson-Boltzmanovo enacbo
εmε0∇2Vp = −e∑k
ZkCkexp
(−ezkVp
kT
)(7)
qzk in Ck sta nabojna in volumska koncentracija ionov vrste k. Z linearizacijo enacbe (7)dobimo Debye-Hucklovo teorijo, ki velja za difuzno plast. Po tej teoriji je zunanje elektricnopolje v elektrolitu senceno, potencial Vp pada eksponentno z razdaljo od vrednosti na mejiplasti Vs
Vp(x) = Vse−κx (8)
Debyeva dolzina sencenja κ−1 je podana z
κ−1 =
(εmε0kT
e2∑
i z2i Ci
) 12
(9)
Splosna resitev Poisson-Boltzmanove enacbe (7) se imenuje Gouy-Chapmanov model [13].Primerjava napovedi te z natancnimi numericnimi izracuni in rezultati Monte-Carlo simulacijjo potrjuje kot zelo natancno, vendar za ta primer zadostuje enostavnejsi Debye-Huckelovmodel [13].
Zunanja elektricna sila deluje na oba, nabit delec in oblak debeline κ−1 s presezkomnasprotno nabitih ionov, ki obdaja delec; ta oblak ionov poleg tega se hidrodinamsko intera-gira z molekulami medija (viskozni upor). Natancen racun je prekompliciran za to obravnavo,preproste resitve pa obstajajo za limitna primera: za debelo Debyevo plast (κ−1 vecji od di-menzije delca R) lahko loceno izracunamo elektricno vlecno silo in viskozno silo. Hitrost delcaje v tem primeru sorazmerena s QE, kjer je Q celotni naboj na delcu in E jakost zunanjegaelektricnega polja. Gibljivost je odvisna tudi od oblike, za kroglo dobimo gibljivost
µ =v
E=
Q
6πηR(10)
12

Slika 7: Delec in dve obmocij elektricne dvojne plasti: (a) nabit delec dimenzije R, (b)Sternova plast debeline κ−1 z nemobilnimi ioni in (c) difuzna plast z mobilnimi ioni.
η je viskoznost medija, R pa radij krogle. V nasprotnem primeru, ko je Debyeva plast majhnav primerjavi z dimenzijami delca, je strig omejen na plast κ−1 okoli delca. Treba je resitiNavier-Stokesovo enacbo
η∇2Vp = −ρE (11)
kjer je ρ resitev Gouy-Chapmanovega (ali pa Debye-Huckelovega) modela znotraj Debyeveplasti ter nic zunaj. Hitrost v tem primeru postane neodvisna od dimenzije in oblike delca,kar popisuje enacba Smoluchowskega [13]
µ =εmε0ζ
η(12)
Zeta potencial ζ je potencial na povrsini delca, pri katerem se pojavi strig v kapljevini medija.V praksi ga je za biomolekule zelo tezko izracunati, zato se ga raje dobi eksperimentalno spomocjo enacbe Smoluchowskega (12).
S pojavom elektroforeze je povezan tudi pojav elektroosmoze: na steni kvarcne kapliarese nahajajo negativno nabite silanolne skupine, zato se tudi ob steni pojavi elektricna dvojnaplast s pozitivnimi ioni. Tudi te vlece elektricna sila in preko striga v kapljevini medijapovzrocijo gibanje le-tega. Dejanska elektroforeticna mobilnost je zato vsota
µ = µ0 + µeo (13)
kjer je µ0 mobilnost delca (10, 12) in µeo mobilnost, ki jo povzroca pojav elektroosmoze.Debyeva plast (κ−1) je v pufrskih raztopinah majhna, zato velja rezim Smoluchowskega [13].Ioni, ki obdajajo polielektroli, sencijo hidodinamske interakcije cez razdalje vecje od κ−1,upor se pojavi le v obmocju d + 2κ−1, kjer je d debelina polielektrolita. Sledi znano inneugodno dejstvo: mobilnost dolgega in gibkega polielektrolia v kapljevini je neodvisna odnjegove velikosti oz. dolzine [13]. Za locevanje STR fragmentov DNK po velikosti potrebujemododaten dejavnik - tega prispevajo vlakna gela v kapilari.
3.4.2 Teorija gel elektroforeze polielektrolitov
Dobra teorija gibanja kratkih DNK fragmentov po gelu je sele v nastajanju, imamo pa vecbolj ali manj dobrih teoreticnih modelov na to temo [13]. Teoretski model, ki dobro opisuje
13

gibanje kratkih (do 1000 baznih parov) in srednjih (do 20000 baznih parov) fragmentov DNK,se imenuje model ”vrata-hodnik”5 [12]. Ta zavraca obstoj stalnih lukenj med vlakni gela,pac pa predpostavlja obstoj podrocij majhne in velike gibljivosti. Podrocja velike gibljivosti(vrata) so tam, kjer je vlakno gela samostojno, podrocja nizke gibljivosti (hodniki) pa tam,kjer se vlakna gela stikajo in tvorijo krizisca. Ko se molekula giblje skozi gel, si sama utirapot in oblikuje luknje. Gibanje poteka diskretno po plasteh, debelina plasti pa je krajsa odznacilne dolzine molekul vzorca, torej na gibajoco se molekulo deluje vec plasti naenkrat. Cemolekula naleti na vrata, zaradi velike gibljivosti, premakne vlakna gela le v trenutni plasti.Ce pa naleti na hodnik, zaradi majhne gibljivosti, premik vlaken gela v trenutni plasti povzrocitudi premike v sosednjih plasteh in odpre se ”hodnik”. Vec hodnikov se lahko zlije v daljsihodnik, ki sega cez nekaj plasti. O tem, ali bo molekula odpirala vec vrat ali vec hodnikovodloca ravnovesje dveh sil: elektrokineticne, ki jo povzroca gibajoca se molekula in upornesile, ki jo predstavljajo vlakna gela. Ko je razmerje uporne in elektrokineticne sile veliko,nastajajo predvsem hodniki, ko je majhno, predvem vrata. Ce sta enaki, se vrata in hodnikiodpirajo z enako verjetnostjo in takrat je tudi najvecja locljivost. Zveza med mobilnostjoDNK in obema silama je [12]
µ = µ0e−Fr/Fe (14)
µ0 je mobilnost fragmentov DNK v mediju (brez gela), ki jo podaja enacba (12), Fr je upornasila gela, Fe pa elektrokineticna sila molekule. Enacba splosne narave (14) se za primer DNKprevede na enacbo, ki direktno povezuje mobilnost in velikost fragmentov DNK
µ =µ0
Nµ0ks + 1(15)
kjer je N velikost fragmenta (stevilo baznih parov) in ks naklon premice v grafu velikostiDNK proti reciprocni gibljivosti, ki jo dobimo eksperimentalno. Enacbi (14) in (15) stapomembni za dolocanje koncentracije gela pri optimizaciji locljivosti za dano obmocje velikostimolekul v vzorcu. Vecanje koncentracije povecuje locljivost pri velikih molekulah, manjsanjekoncentracije pa povecuje locljivost pri manjsih molekulah. Vsak gel je karakteriziran s kdc,ki je produkt µ0 in ks in je neodvisna od sprememb temperature in elektricnega polja. Hitrostgibanja fragmenta DNK v odvisnoti od njegove velikosti je sorazmerna mobilnosti, in ker jehitrost v povprecju stalna, sledi za prepotovano dolzino
l = vt ∝ µt =µ0t
Nµ0ks + 1(16)
Kapilarna gel elektroforeza je boljsa od navadne gel elektroforeze zaradi vec stvari: strukturagela je bolj stabilizirana (znacilni premeri kapilar so 50-100 µm) in daje zato boljse rezultate,zaradi vecjega razmerja povrsina/prostornina se gel manj segreva, kar zopet poveca locljivost.Zaradi majhne debeline kapilare pa je mozna tudi avtomatska opticna detekcija.
Dejansko merijo cas, ki ga porabijo fragmenti z dolocenega lokusa DNK, da prepotujejocelotno dolzino kapilare. Vsi fragmenti v raztopini vzorca zacnejo potovati po kapilari ob istemcasu, a imajo glede na svojo dolzino razlicno hitrost. Na drugem koncu kapilare s pomocjofluorescence opazujemo njihov prihod. Dolocen kratek odsek na koncu kapilare osvetljujejoz lasersko svetlobo in opazujejo izsevano svetlobo iz tega odseka. Molekula DNK je le malofluorescentna, zato ze na zacetne oligonukleotide vezejo fluorescentne molekule (markerje), insicer razlicne (ki fluorescirajo pri razlicnih valovnih dolzinah) za vsak zacetni oligonukleotid,
5ang. door-corridor model
14
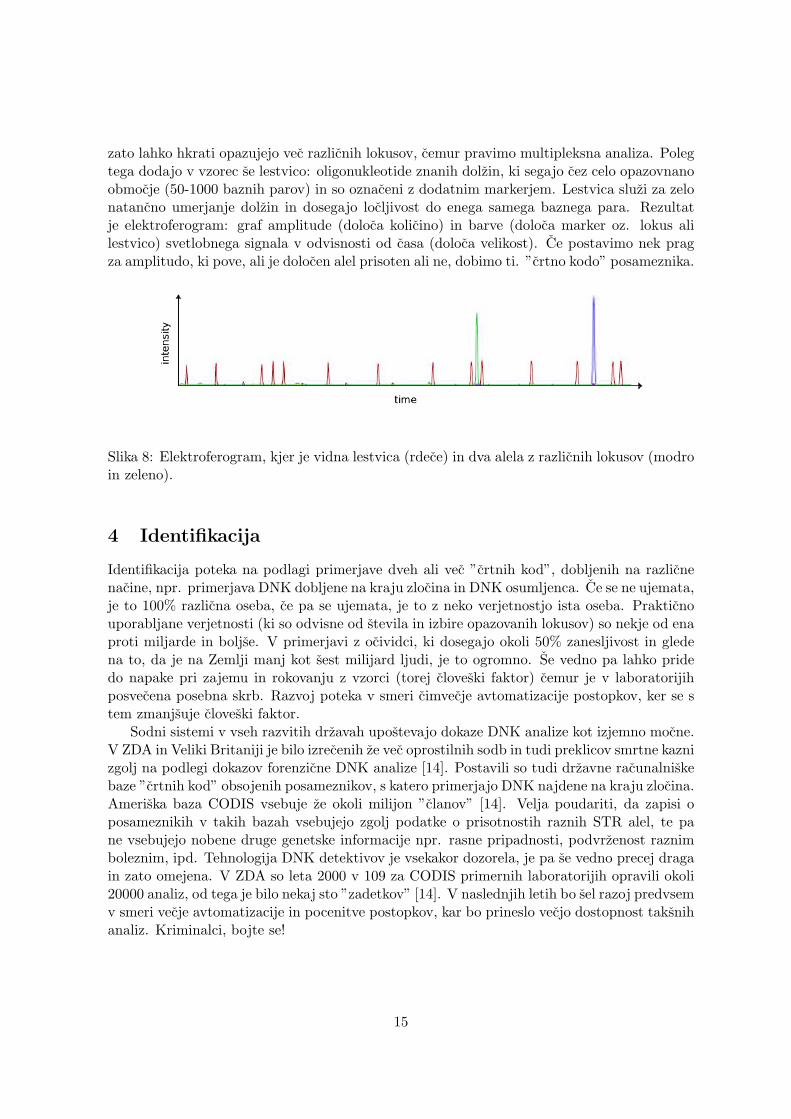
zato lahko hkrati opazujejo vec razlicnih lokusov, cemur pravimo multipleksna analiza. Polegtega dodajo v vzorec se lestvico: oligonukleotide znanih dolzin, ki segajo cez celo opazovnanoobmocje (50-1000 baznih parov) in so oznaceni z dodatnim markerjem. Lestvica sluzi za zelonatancno umerjanje dolzin in dosegajo locljivost do enega samega baznega para. Rezultatje elektroferogram: graf amplitude (doloca kolicino) in barve (doloca marker oz. lokus alilestvico) svetlobnega signala v odvisnosti od casa (doloca velikost). Ce postavimo nek pragza amplitudo, ki pove, ali je dolocen alel prisoten ali ne, dobimo ti. ”crtno kodo” posameznika.
Slika 8: Elektroferogram, kjer je vidna lestvica (rdece) in dva alela z razlicnih lokusov (modroin zeleno).
4 Identifikacija
Identifikacija poteka na podlagi primerjave dveh ali vec ”crtnih kod”, dobljenih na razlicnenacine, npr. primerjava DNK dobljene na kraju zlocina in DNK osumljenca. Ce se ne ujemata,je to 100% razlicna oseba, ce pa se ujemata, je to z neko verjetnostjo ista oseba. Prakticnouporabljane verjetnosti (ki so odvisne od stevila in izbire opazovanih lokusov) so nekje od enaproti miljarde in boljse. V primerjavi z ocividci, ki dosegajo okoli 50% zanesljivost in gledena to, da je na Zemlji manj kot sest milijard ljudi, je to ogromno. Se vedno pa lahko pridedo napake pri zajemu in rokovanju z vzorci (torej cloveski faktor) cemur je v laboratorijihposvecena posebna skrb. Razvoj poteka v smeri cimvecje avtomatizacije postopkov, ker se stem zmanjsuje cloveski faktor.
Sodni sistemi v vseh razvitih drzavah upostevajo dokaze DNK analize kot izjemno mocne.V ZDA in Veliki Britaniji je bilo izrecenih ze vec oprostilnih sodb in tudi preklicov smrtne kaznizgolj na podlegi dokazov forenzicne DNK analize [14]. Postavili so tudi drzavne racunalniskebaze ”crtnih kod” obsojenih posameznikov, s katero primerjajo DNK najdene na kraju zlocina.Ameriska baza CODIS vsebuje ze okoli milijon ”clanov” [14]. Velja poudariti, da zapisi oposameznikih v takih bazah vsebujejo zgolj podatke o prisotnostih raznih STR alel, te pane vsebujejo nobene druge genetske informacije npr. rasne pripadnosti, podvrzenost raznimboleznim, ipd. Tehnologija DNK detektivov je vsekakor dozorela, je pa se vedno precej dragain zato omejena. V ZDA so leta 2000 v 109 za CODIS primernih laboratorijih opravili okoli20000 analiz, od tega je bilo nekaj sto ”zadetkov” [14]. V naslednjih letih bo sel razoj predvsemv smeri vecje avtomatizacije in pocenitve postopkov, kar bo prineslo vecjo dostopnost taksnihanaliz. Kriminalci, bojte se!
15

Literatura
[1] James D. Watson and Francis H. Crick. Molecular structure of nucleic acids; a structurefor deoxyrobose nucleic acid. Nature, 171:737–738, 1953.
[2] Bruce Alberts, Dennis Bray, Julian Lewis, Martin Raff, Keith Roberts, and James D.Watson. Molecular Biology of the Cell. Garland Publishing, 3rd edition, 1994.
[3] D. Botstein, R. C. White, M. Skolnick, and R. W. Davis. Construction of a geneticlinkage map in man using restriction fragment length polymorphisms. Am. J. Hum.Genet., 32:314–331, 1980.
[4] R. K. Saiki, S. Scharf, F. Falowa, K. B. Mullis, G. T. Horn, H. A. Erlich, and N. Arnheim.Enzymatic amplification of beta-globin genomic sequences and restriction site analysisfor diagnosis of sickle cell anemia. Science, 230:1350–1354, 1985.
[5] Thomas Curran. Forensic DNA Analysis: Technology And Application. Svetovni splet,http://www.parl.gc.ca/information/library/PRBpubs/bp443-e.htm, 1997.
[6] Norah Rudin and Keith Inman. An Introduction to Forensic DNA Analysis. CRC Press,2nd edition, 2002.
[7] Mark Benecke. DNA typing in forensic medicine and in criminal investigations: a currentsurvey. Naturwissenscheften, 84:181–188, 1997.
[8] National Institute for Justice. What Every Law Enforcement Officer Should Know AboutDNA Evidence. Svetovni splet.
[9] Mark Benecke. Collection and handling of forensic DNA samples. Svetovni splet, http://www.benecke.com/dnacollection.pdf, 2005.
[10] Christian M. Ruitberg, Dennis J. Reeder, and John M. Butler. STRBase: A short tandemrepeat dna database for the human identity testing comunity. Nucleic Acids Research,29:320–322, 2001.
[11] Ed Rybicki. PCR Primer Design and Reaction Optimisation. Svetovni splet, http://www.mcb.uct.ac.za/pcroptim.htm, 2001.
[12] Branko Kozulic. Models of gel electrophoresis. Analitical Biochemistry, 231:1–12, 1995.
[13] Jean-Louis Viovy. Electrophoresis of dna and other polyelectrolytes: Physical mecha-nisms. Review of Modern Physics, 72:813–866, 2000.
[14] US Department of Justice. The FBI’s DNA and Databasing Initiatives. Svetovni splet.
16





























