Embed Size (px)
Citation preview

Universidad de La Salle Universidad de La Salle
Ciencia Unisalle Ciencia Unisalle
Maestría en Ciencias Veterinarias Facultad de Ciencias Agropecuarias
1-1-2016
Efecto in vitro de los taninos condensados de las plantas Efecto in vitro de los taninos condensados de las plantas
leucaena leucocephala, calliandra calothyrsus y flemingia leucaena leucocephala, calliandra calothyrsus y flemingia
macrophylla sobre huevos y larvas l3 de nematodos macrophylla sobre huevos y larvas l3 de nematodos
gastrointestinales de ovinos gastrointestinales de ovinos
Adriana del Socorro Bernal Peralta Universidad de La Salle, Bogotá
Ángela Lucia Camargo Silva Universidad de La Salle, Bogotá
Follow this and additional works at: https://ciencia.lasalle.edu.co/maest_ciencias_veterinarias
Citación recomendada Citación recomendada Bernal Peralta, A. d., & Camargo Silva, Á. L. (2016). Efecto in vitro de los taninos condensados de las plantas leucaena leucocephala, calliandra calothyrsus y flemingia macrophylla sobre huevos y larvas l3 de nematodos gastrointestinales de ovinos. Retrieved from https://ciencia.lasalle.edu.co/maest_ciencias_veterinarias/56
This Tesis de maestría is brought to you for free and open access by the Facultad de Ciencias Agropecuarias at Ciencia Unisalle. It has been accepted for inclusion in Maestría en Ciencias Veterinarias by an authorized administrator of Ciencia Unisalle. For more information, please contact [email protected].

UNIVERSIDAD DE LA SALLE
FACULTAD DE CIENCIAS AGROPECUARIAS
MAESTRÍA EN CIENCIAS VETERINARIAS
EFECTO IN VITRO DE LOS TANINOS CONDENSADOS DE LAS PLANTAS
LEUCAENA LEUCOCEPHALA, CALLIANDRA CALOTHYRSUS Y FLEMINGIA
MACROPHYLLA SOBRE HUEVOS Y LARVAS (L3) DE NEMATODOS
GASTROINTESTINALES DE OVINOS.
Preparado por:
ADRIANA DEL SOCORRO BERNAL PERALTA
ÁNGELA LUCÍA CAMARGO SILVA
Trabajo de grado como requisito para optar por el título de
Magister en Ciencias Veterinarias
Bogotá
2016

ii
UNIVERSIDAD DE LA SALLE
FACULTAD DE CIENCIAS AGROPECUARIAS
MAESTRÍA EN CIENCIAS VETERINARIAS
EFECTO IN VITRO DE LOS TANINOS CONDENSADOS DE LAS PLANTAS
LEUCAENA LEUCOCEPHALA, CALLIANDRA CALOTHYRSUS Y FLEMINGIA
MACROPHYLLA SOBRE HUEVOS Y LARVAS (L3) DE NEMATODOS
GASTROINTESTINALES DE OVINOS.
Preparado por:
ADRIANA DEL SOCORRO BERNAL PERALTA
76131222
ÁNGELA LUCÍA CAMARGO SILVA
Código: 76131215
Director
DILDO MÁRQUEZ LARA, M.V., Esp., M.Sc.
Bogotá
2016

iii
APROBACIÓN
DIRECTOR
________________________________ Dildo Márquez Lara JURADO
________________________________ Felipe Gamboa Ruiz JURADO
________________________________ Julio Enrique Gómez JURADO
________________________________ Martin Orlando Pulido

iv
DIRECTIVOS
Rector Hermano Carlos Gabriel Gómez Restrepo Vicerrector Académico Hermano Carlos Carvajal Costa Vicerrector de Promoción Hermano Frank Leonardo Ramos Baquero y Desarrollo Humano Vicerrector Administrativo Dr. Eduardo Ángel Reyes Vicerrector de Investigación Dr. Luis Eduardo Ramírez Hernández y Transferencia Secretaria General Dra. Patricia Ortiz Valencia Decana Facultad de Dra. Claudia Aixa Mutis Barreto Ciencias Agropecuarias Secretario Académico Dr. Alejandro Tobón González Director de Postgrados en Dr. Ernesto Andrés Dalmau Barros Ciencias Veterinarias

v
COMPROMISO
El presente trabajo de grado no contiene ideas contrarias a la doctrina, moral y dogma
de la Iglesia Católica.
Todo lo escrito y expresado en este trabajo se encuentra dentro de los lineamientos
de honestidad, autenticidad y responsabilidad, y los contenidos que aquí se presentan
son de carácter estrictamente educativo, científico e informativo y no son
responsabilidad ni del director del proyecto ni de los jurados de la Universidad de La
Salle.

vi
AGRADECIMIENTOS
A Dios por darme la fortaleza y perseverancia durante este camino, a mi padre Raul
Camargo por su apoyo incondicional, a mi madre Nubia Silva por darme la vida, a mi
compañera de trabajo por su dedicación y paciencia y a todas las personas que de
una forma u otra se interesaron por mi trabajo siendo apoyo cada momento para
continuar.
Ángela.
Al Todopoderoso por permitirnos salir adelante en este proceso, a mi esposo Jaime
y a mi hijo Andrés por ser mis pilares y no dejarme decaer, a Ángela por ser mi guía y
punto de apoyo en todos los momentos vividos con ésta investigación, a Klarena por
toda su ayuda y consejos y a mi familia que siempre me han brindado su confianza.
Adriana
Nuestros más sinceros agradecimientos al Dr. Dildo marquez por acogernos y dirigir
nuestro trabajo de investigación, al Dr.Jaime Andrés Cubides por su apoyo, compartir
sus conocimientos y por el tiempo dedicado durante todo nuestro trabajo, A la doctora
Leyla y al Dr. César Díaz por sus valiosos aportes, a Lucio y a Estelita por su ayuda
incondicional para la realización de nuestro trabajo.

vii
RESUMEN
La ganadería ovina en Colombia ha ganado un importante terreno durante los últimos años. El hato ovino Colombiano asciende a 2 millones de cabezas en todo el país. Las infecciones por nematodos gastrointestinales se han relacionado con grandes pérdidas económicas para este sector en regiones tropicales. La creciente resistencia a medicamentos antihelmínticos tradicionales de estos nematodos es el resultado del inadecuado manejo de los productos farmacéuticos, la falta de tecnificación, entre otros factores; por lo cual se deben implementar alternativas terapéuticas que sean sostenibles y eficaces desde la perspectiva ambiental; es así, que el uso de plantas medicinales con actividad antihelmíntica como la Leucaena leucocephala, Calliandra calothyrsus y Flemingia macrophylla, podrían plantearse como una solución alternativa al problema de resistencia de éstos parásitos a productos alopáticos.En ésta investigación se evaluó el efecto in vitro de los taninos condensados de extractos de las plantas L. leucocephala, C. calothyrsus y F. macrophylla sobre huevos y larvas L3
de nematodos gastrointestinales de ovinos. Metodología: las fases de entrenamiento y pruebas finales se desarrollaron gracias a la facilitación del laboratorio de helmintología en la corporación Colombiana de investigación agropecuaria (CORPOICA) de Tibaitatá; el proceso de obtención de taninos condensados fue realizado por el laboratorio de química en la Universidad del Tolima (Ibagué); Se hicieron diferentes concentraciones de dichos taninos a partir de las cuales se llevaron a cabo las pruebas de inhibición de la eclosión y eliminación de la cutícula; previa obtención de huevos y larvas en estadio infectante L3, por medio de incubación y cultivo de los mismos a partir de un ovino infestado naturalmente. Se realizó la identificación morfométrica de larvas L3 determinando en porcentaje la participación de cada género y especie. Resultados: En la identificación morfométrica el porcentaje más representativo fue de un 58% correspondiente a Haemonchus contortus seguido de un 26% de Trichostrongylus falculatus, en contraste Cooperia curticei registró el 1%. Los extractos de taninos condensados de Calliandra calothyrsus presentaron mayor eficacia en la inhibición de la eclosión de huevos, tanto en la D50 1075,25 μg/ml como la D99 3601,23 μg/ml, los de Flemingia macrophylla y Leucaena leucocephala, inhibieron el proceso de eclosión de huevos de manera significativa ya que a mayor concentración de la dosis se presentó una mejor respuesta, a diferencia de C. calothyrsus, donde no fue evidente un efecto dosis dependiente. Los resultados obtenidos por el análisis Logyt Probit en la eliminación de la cutícula se realizaron con base a los valores promedios en cada prueba, la D50 que mejor eficacia mostró fue la del extracto de tanino condensado de C. calothyrsus 1,915 μg/ml, con respecto a los extractos de las otras dos plantas, mientras que F. macrophylla presentó D99 más eficaz (326 μg/ml). Todos los extractos inhibieron la eliminación de la cutícula, desde la concentración más baja (31,25 μg/ml) y el efecto fue dosis dependiente; los porcentajes de inhibición con la concentración más alta (1000 μg/ml) fueron respectivamente del 99%, 98% y 92% para Calliandra, Leucaena y Flemingia.
Palabras Clave: Ovinos, nematodos gastrointestinales, taninos condensados, Leucaena leucocephala, Calliandra calothyrsus, Flemingia macrophylla.

viii
ABSTRACT
Sheep farming in Colombia has gained significant ground in recent years. Colombian sheep herd amounts to 2 million head across the country. The gastrointestinal nematode infections have been linked to major economic losses for this sector in tropical regions. The growing resistance to anthelmintics traditional medicines of these nematodes is the result of improper handling of pharmaceutical products, lack of mechanization, among other factors; therefore be implemented therapeutic alternatives that are sustainable and effective from an environmental perspective; so that the use of medicinal plants with anthelmintic activity as Leucaena leucocephala, Calliandra calothyrsus and Flemingia macrophylla could pose as an alternative solution to the problem of resistance of these parasites to research products alopáticos. In this investigation in vitro effect was evaluated condensed tannins from plant extracts L. leucocephala, C. calothyrsus and F. macrophylla on eggs and larvae L3 of gastrointestinal nematodes of sheep. Methodology: training phases and final test were developed thanks to the facilitacion helminthology laboratory in the Colombian Agricultural Research Corporation (CORPOICA) of Tibaitata; the process of obtaining condensed tannins was carried out by the laboratory of chemistry at the University of Tolima (Ibague); different concentrations of these tannins from which testing inhibition of hatching and removal of the cuticle were conducted, prefetching eggs and larvae infective L3 stage, through incubation and cultivation thereof from a naturally infested sheep. Morphometric identification of L3 larvae determining percentage participation of each genus and species was performed. Results: In the morphometric identifying the most representative percentage was 58% for Haemonchus contortus followed by 26% of Trichostrongylus falculatus, by contrast Cooperia curticei recorded 1%. Extracts of Calliandra calothyrsus condensed tannins had higher efficacy in inhibiting egg hatching, both the D50 1075.25 ug / ml as the D99 3601.23 ug / ml, the Flemingia macrophylla and Leucaena leucocephala, inhibited egg hatching process significantly since the higher dosage strength showed a better response, unlike C. calothyrsus, which was not evident a dose dependent effect. The results obtained by analysis Logyt Probit in removing the cuticle were performed based on the average values in each test, the D50 better efficacy showed was the extract condensed tannin C.calothyrsus 1.915 ug / ml, with respect to extracts from the other two plants, while D99 F.macrophylla showed more effective (326 ug / ml). All extracts inhibited the elimination of cuticle from the lowest concentration (31.25 ug / ml) and the effect was dose dependent; percent inhibition with the higuest concentration (1000 mg/ml) were respectively 99%, 98% and 92% for Calliandra, Leucaena and Flemingia. Keywords: Sheep, gastrointestinal nematodes, condensed tannins, Leucaena leucocephala, Calliandra calothyrsus, Flemingia macrophylla.

ix
TABLA DE CONTENIDO
1. INTRODUCCIÒN………………………………………….....…………………………..17
2. MARCO TEÓRICO…………………………………………………………………........21
2.1. Nematodos gastrointestinales…….……………………………………………….... 21
2.2. Patogenia y signos clínicos………………………………………………………….. 25
2.3. Métodos de control de nematodos gastrointestinales………………………………27
2.3.1. Antihelmínticos convencionales………………………………………………….28
2.3.2. Métodos no convencionales……………………………………………………...31
2.3.2.1. Herbolaria………………………………………………………………………...31
2.3.2.2. Control biológico…………………………………………………………………32
2.3.2.3. Inmunización con vacunas……………………………………………………..32
2.3.2.4. Agujas de cobre………………………………………………………………....33
2.4. Resistencia antihelmíntica…………………………………………………………….33
2.5. Diagnóstico de nematodos gastrointestinales en ovinos………………………….34
2.6. Fitoterapia……………………………………………………………………………....40
2.6.1. Características de la fitoterapia…………………………………………………..41
2.6.2. Historia de la fitoterapia…………………………………………………………...42
2.6.3. Futuro de la fitoterapia…………………………………………………………….44
2.7. Metabolitos secundarios de las plantas………………………………………………45
2.8. Taninos………………………………………………………………………………….47
2.8.1. Estructura química de taninos……………………………………………………47
2.8.2. Funciones de los taninos………………………………………………………....50
2.8.3. Ventajas y desventajas……………………………………………………………51

x
2.8.3.1. Efectos de los taninos sobre el consumo voluntario de alimento………….51
2.8.3.2. Efectos sobre la digestibilidad de los alimentos……………………………..54
2.8.3.2.1 Efectos en la fermentación ruminal………………………………………….56
2.8.3.3 .Efectos sobre la producción…………………………………………………...58
2.8.3.4. Efectos sobre la reproducción…………………………………………………59
2.8.3.5. Efectos sobre la salud animal………………………………………………….60
2.8.3.6 Propiedades antiparasitarias……………………………………………………61
2.8.4 Toxicidad de los taninos.…………...………………………………………….........64
2.9 Leucaena leucocephala….…………………………………………………………….65
2.10 Calliandra calothyrsus…….…………………………………………………………..66
2.11 Flemingia macrophylla………………………………………...................................67
3. METODOLOGÍA………………………………………………………………………….69
3.1 Localización………………………………………………..........................................69
3.2 Material orgánico………………………………………………………………………..69
3.2.1. Técnica de obtención de huevos…………………………………………………70
3.2.2. Técnica coprocultivo……………………………………………………………….71
3.2.3. Técnica lavado de larvas………………………………………………………….72
3.2.4. Conteo larvas L3………………………………………………………………………………………………………72
3.2.5. Determinación de concentración de hipoclorito de sodio………………………73
3.2.6. Calibración de ocular para identificación morfométrica………………………..74
3.3. Extractos de plantas……………………………………………………………………74
3.4. Identificación morfométrica……………………………………………………………78
3.5. Pruebas In vitro…………………………………………………………………………75
3.5.1. Prueba de inhibición de la eclosión………………………………………………75

xi
3.5.2. Prueba de eliminación de la cutícula……………………………………………76
3.6. Diseño estadístico……………………………………………………………………..77
4. Resultados………………………………………………………………………………..78
4.1. Identificación morfométrica de larvas en estadio infectante (L3) de nematodos
gastrointestinales de ovinos…………………………………………………………….....78
4.2. Resultados prueba de sensibilidad de la cutícula con el hipoclorito…………......80
4.3. Prueba de eliminación de la cutícula…………………………………………………81
4.4. Prueba de inhibición de la eclosión ……………………………………………….....86
5. Discusión……………………………………………………………………………….....91
6. Conclusiones……………………………………………………………………………...98
7. Lista de referencias………………………………………………………………………99
8. Anexos…………………………………………………………………………………...116

xii
LISTA DE TABLAS
Tabla 1. Antihelmínticos convencionales, mecanismos de acción y algunos productos
comerciales…………………………………………………………………………………..28
Tabla 2. Clasificación de las especies forrajeras de clima templado según la
concentración de taninos presentes en su composición………………………………..50
Tabla 3. Promedio de medidas tomadas en la identificación morfométrica de 100 L3 de
nematodos gastrointestinales……………………………………………………………...79
Tabla 4. Resultados prueba de sensibilidad con hipoclorito de sodio…………………80
Tabla 5. Valores promedio del porcentaje de inhibición de la eliminación de la cutícula
de las observaciones repetidas seleccionadas por planta y concentración…………..82
Tabla 6. Concentración del extracto de taninos condensados Leucaena leucocephala
que se requiere para inhibir el 50 % y el 99% de la eliminación de la cutícula de larvas
L3 de nematodos gastrointestinales de ovinos…………………………………………..83
Tabla 7. Concentración del extracto de taninos condensados Calliandra calothyrsus
que se requiere para inhibir el 50 % y el 99% de la eliminación de la cutícula de larvas
L3 de nematodos gastrointestinales de ovinos…………………………………………..83
Tabla 8. Concentración del extracto de taninos condensados Flemingia macrophylla
que se requiere para inhibir el 50 % y el 99% de la eliminación de la cutícula de larvas
L3 de nematodos gastrointestinales de ovinos…………………………………………..83
Tabla 9. Valores promedio y desviación estándar del porcentaje de inhibición de la
eclosión por planta y concentración……………………………………………………….87

xiii
Tabla 10. Concentración del extracto de taninos condensados Calliandra calothyrsus
que se requiere para inhibir el 50 % y el 99% de la eclosión de huevos de nematodos
gastrointestinales de ovinos………………………………………………………………..88
Tabla 11. Concentración del extracto de taninos condensados Leucaena leucocephala
que se requiere para inhibir el 50 % y el 99% de la eclosión de huevos de nematodos
gastrointestinales de ovinos……………………………………………...........................88
Tabla 12. Concentración del extracto de taninos condensados Flemingia macrophylla
que se requiere para inhibir el 50 % y el 99% de la eclosión de huevos de nematodos
gastrointestinales de ovinos……………………………………………...........................88

xiv
LISTA DE FIGURAS
Figura 1. Representación del ciclo de vida de nematodos gastrointestinales en
pequeños rumiantes………………………………………………………………………...25
Figura 2. Puntuación de tarjetas utilizadas para evaluación de los cinco puntos…….36
Figura 3. Larvas (L3) de especies representativas de nematodos gastrointestinales en
ovejas, encontradas tras cultivo larvario……………………….…………………………38
Figura 4. Tanino hidrolizable (Gallotannin) compuesto por ésteres de ácido gálico,
vinculados a un núcleo de azúcar………………………………………………………....49
Figura 5. Estructura básica de un tanino condensado, mostrando estereoquímica,
vínculos interflavánicos e hidroxilación del anillo beta…………………………………..49
Figura 6. Larvas (L3), Oesophagostonum columbianum, Trichostrongylus falculatus y
Haemonchus contortus………………………………………………….……...……….....78
Figura 7. Inhibición de la eliminación de la cutícula del extracto de TC de cada planta
por concentración……………………………………......................................................84
Figura 8. Efecto inhibición de la eliminación de la cutícula del extracto de TC de cada
planta al minuto 60………………………………………………………………………….85
Figura 9. Cambios físicos y alteraciones de la cutícula………………………………...91
Figura 10. Inhibición de la eclosión del extracto de TC de cada planta por
concentración…………………………………………………….………………………….89
Figura 11. Efectos del extracto de TC de cada planta al aumentar la concentración en
la inhibición de la eclosión………………………………………………………………….90

xv
LISTA DE ANEXOS
Anexo 1. Imágenes de Recuento de Huevos por gramo de materia fecal………….117
Anexo 2. Imágenes Técnica de Coprocultivo y Obtención de larvas………………..118
Anexo 3. Imágenes del Lavado de larvas……………………………………………...120
Anexo 4. Imágenes del procedimiento para la determinación de la concentración
adecuada de hipoclorito de sodio para la prueba de eliminación de la cutícula……121
Anexo 5. Imágenes identificación morfométrica de Larvas L3………………………………...122
Anexo 6. Medidas de referencia para la Identificación morfométrica de larvas L3 de
nematodos gastrointestinales más comunes de pequeños rumiantes………………124
Anexo 7. Identificación morfométrica de 100 larvas L3 de nematodos gastrointestinales
ovinos……………………………………………………………………………………….125
Anexo 8. Análisis Logit Probit de la prueba de inhibición de la eclosión Leucaena
leucocephala……………………………………………………………………………….129
Anexo 9. Efecto de las diferentes concentraciones del extracto de Leucaena
leucocephala en la prueba in vitro de inhibición de la eclosión de huevos de nematodos
gastrointestinales de ovinos………………………………………………………………130
Anexo 10. Análisis Logit Probit de la prueba de inhibición de la eclosión Calliandra
calothyrsus………………………………………………………………………………….131
Anexo 11. Efecto de las diferentes concentraciones del extracto de Calliandra
calothyrsus en la prueba in vitro de inhibición de la eclosión de huevos de nematodos
gastrointestinales de ovinos………………………………………………………….…..132

xvi
Anexo 12. Análisis Logit Probit de la prueba de inhibición de la eclosión Flemingia
macrophylla………………………………………………………………………………...133
Anexo 13. Efecto de las diferentes concentraciones del extracto de Flemingia
macrophylla en la prueba in vitro de inhibición de la eclosión de huevos de nematodos
gastrointestinales de ovinos…………………………………………..…………………..134
Anexo 14. Análisis Logit Probit de la prueba de eliminación de la cutícula Leucaena
leucocephala……………………………………………………………………………….135
Anexo 15. Efecto de las diferentes concentraciones del extracto de Leucaena
leucocephala en la prueba in vitro de inhibición de la eliminación de la cutícula de
nematodos gastrointestinales de ovinos………………………………………………..136
Anexo 16. Análisis Logit Probit de la prueba de eliminación de la cutícula Calliandra
calothyrsus………………………………………………………………………………….137
Anexo 17. Efecto de las diferentes concentraciones del extracto de Calliandra
calothyrsus en la prueba in vitro de inhibición de la eliminación de la cutícula de
nematodos gastrointestinales de ovinos………………………………………………...138
Anexo 18. Análisis Logit Probit de la prueba de eliminación de la cutícula Flemingia
macrophylla………………………………………………………………………………...139
Anexo 19. Efecto de las diferentes concentraciones del extracto de Flemingia
macrophylla en la prueba in vitro de inhibición de la eliminación de la cutícula de
nematodos gastrointestinales de ovinos………………………………………………...140

17
1. INTRODUCCIÓN
El inventario ovino-caprino mundial según la FAO (2006) ubica a Colombia en el
puesto 63 de los productores ovinos, después de países como China; a nivel
nacional esta actividad productiva se encuentra de forma más intensiva en los
departamentos de Guajira, Magdalena, Atlántico, Bolívar, Sucre y Córdoba; así
mismo Santander, Nariño, Putumayo, Cesar y el altiplano Cundiboyacense
(Ministerio de Agricultura y Desarrollo Rural, 2006).
En los últimos años, el desarrollo de las producciones y la agroindustria en el
sector ovino ha impulsado la importación de animales, la incorporación de
tecnología, la adquisición de equipos e implementos, la capacitación técnica, entre
otros; permitiendo satisfacer, en parte, la demanda incipiente de productos que se
presenta tanto en Colombia, como en algunos mercados en países vecinos. Las
ovejas son comunes en varias zonas geográficas del país, adaptándose con
facilidad a los diferentes climas. De este rumiante menor se obtiene un triple
propósito carne, lana y leche por lo cual se han venido desarrollando nuevas formas
de obtención y comercialización de animales, realizando importaciones de
reproductores de México, Chile y Uruguay, fomentando la apertura de mercados
internacionales que demandan la carne ovina del país como en las Antillas
(Asoovinos, 2010-2018).
Según los datos reportados por la FAO (2003), Colombia, históricamente ha
tenido un consumo per cápita de carne ovina y caprina de alrededor de los 500 gr;

18
con base en la información obtenida por visitas regionales se han podido detectar
niveles de consumo significativos en las regiones productoras. En el año 2013,
según el Instituto Colombiano Agropecuario la población de ovinos es 1.177.944
revistiendo gran importancia sanitaria ya que la especie ovina es susceptible a
enfermedades epidemiológicamente importantes y cumple un papel decisivo en la
supervivencia y diseminación de agentes virales, parasitarios o bacterianos.
La mayoría de los ovinos tienen determinada predisposición a infecciones
parasitarias, esto debido a que el proceso de reproducción y crianza se desarrolla
de forma colectiva. Los nematodos gastrointestinales (NGI) de la familia
Trichostrongylidae son los más frecuentes en las explotaciones ovinas (Herrera,
Ríos y Zapata, 2013). A nivel mundial, las infecciones parasitarias se asocian a
importantes pérdidas económicas a la industria ganadera (Vercruysse y Claerebout,
2001), esto debido a que los NGI tienen un impacto negativo en la eficiencia
nutricional y en la productividad de los ovinos; esta especie es generalmente más
propensa a sufrir parasitismo gastrointestinal, ya que la mayoría de tiempo
permanecen en pasturas que tienden a estar contaminadas con nematodos en
estadio infectante (L3) (Papadopoulos et al., 2003). Estos NGI atacan a rumiantes
de diferentes edades en zonas tropicales, subtropicales y templadas del mundo.
En Colombia la incidencia de las parasitosis gastrointestinales ocupa un alto
nivel de presentación dentro de las 5 principales enfermedades de los sistemas de
producción en rumiantes para el año 2010 (Díaz et al., 2012), reportándose en el
departamento de Córdoba una prevalecía del 97.70% de Tricostrongilidios
principalmente Haemonchus contortus (Ensuncho-Hoyos et al., 2014); en Antioquia
un estudio realizado en el año 2013 se encontró una frecuencia de infección por

19
NGI del 86.6% con una prevalencia del 66.3% de H. contortus seguido por
Oesophagostomum spp. 38.9%, Trichostrongylus spp. 34.7% y Ostertagia spp.
24.2% (Herrera O, L., Ríos O, L., y Zapata S, R. 2013)
La infección por parásitos de los géneros: Haemonchus spp, Trichostrongylus
spp, Teladorsagia spp, Cooperia spp, Nematodirus spp, Bunostomum spp y
Gaigeria spp, constituyen uno de los principales problemas de las explotaciones
ovinas de los países del trópico, ya que generan grandes pérdidas económicas
debidas a la disminución en la ganancia de peso, inmunosupresión y gastroenteritis
asociada a síndrome de mala absorción, lo que se traduce en la deficiencia en la
producción y reproducción alterando de este modo el desarrollo anormal de las
actividades fisiológicas (Drudge, Szanto, Wyant y Elam, 1964).
El control de las parasitosis internas en animales a nivel mundial se ha venido
realizando a lo largo del tiempo con el uso de agentes químicos, pero debido a su
manejo inadecuado, estos organismos han sufrido un proceso de selección, que ha
generado resistencia a la mayoría de estos productos (Brasil et al., 2012).
De acuerdo con lo anterior, la Organización de las Naciones Unidas para la
Alimentación y la Agricultura en el año 2004 sugiere que se desarrollen y apliquen
estrategias que no afecten negativamente al medio ambiente y que además sean
efectivas desde el punto de vista de la salud animal. Por lo anterior, el uso de
extractos de plantas, específicamente, de taninos condensados para el control de
parásitos ha demostrado tener buenos resultados en el manejo integrado de NGI
(Hoste, Jackson, Athanasiadou, Thamsborg y Hoskin, 2006), además, por su corto
tiempo de biodegradación en el suelo contribuyen de forma positiva a la
sostenibilidad ambiental (Domingues et al., 2013)

20
El objetivo general de esta investigación consistió en evaluar el efecto in vitro de
los extractos de taninos condensados de las plantas: Leucaena leucocephala,
Calliandra calothyrsus y Flemingia macrophylla sobre huevos y larvas L3 de NGI de
ovinos.En cuanto a los objetivos específicos se realizó la identificación morfométrica
de larvas L3 de nematodos gastrointestinales de ovinos provenientes de infección
natural y posteriormente se evaluó en condiciones in vitro, el efecto de los taninos
condensados de las plantas anteriormente mencionadas, por medio de las pruebas
de inhibición de la eclosión y eliminación de la cutícula de larvas L3.

21
2. MARCO TEORICO
2.1. Nematodos gastrointestinales.
La familia de los NGI en los ovinos está constituida por parásitos de los géneros:
Haemonchus spp, Trichostrongylus spp, Teladorsagia spp, Cooperia spp, Gaigeria
spp, Nematodirus spp y Bunostomum spp, todos tienen la capacidad de generar,
entre otras alteraciones, gastroenteritis, anemia, debilidad, disminución del diámetro
de la fibra, pérdida de lana y pérdida de peso (Craig, 2009).
Las especies anteriormente mencionadas fueron introducidas probablemente al
neotrópico con los viajes de los conquistadores y el traslado respectivo del ganado
(Giudici, 1999; Hoberg, Lichtentels y Gibbons, 2004).
Los nematodos de la familia Trichostrongylidae son los más frecuentes en las
explotaciones ovinas (Herrera, Ríos y Zapata, 2013; Berghen, Hilderson, Dorny y
Veracruysse, 1993; Zajac, 2006). Por su parte H. contortus y Trichostrongylus son
considerados como los parásitos más importantes en pequeños rumiantes en
regiones cálidas y húmedas de todo el mundo. En climas fríos y húmedos
Teladorsagia spp., es estimado como agente causal de pérdidas económicas en
dichas explotaciones; una característica importante, es que tiende a presentarse
con frecuencia como una infección mixta junto con Trichostrongylus (Craig, 2009).
Cooperia spp., se considera como un patógeno leve que no genera alteraciones
graves pero también podría presentarse en infecciones mixtas agudas y en dicho
caso si podría contribuir a exacerbar el cuadro clínico (Craig, 2009).

22
H. contortus se produce en el abomaso de los rumiantes, dentro de sus
características principales se encuentra su longitud que oscila entre los 10 a 30 mm.
Debido a la capacidad hematófaga de este parásito se puede observar su
intestino con una coloración roja, además, el útero y los ovarios se ubican en forma
de envoltura alrededor del intestino dando una apariencia de bastón. Las hembras
adultas producen de 5000 a 6000 huevos al día que son expulsados dentro de las
heces al exterior de huésped, deben transcurrir mínimo de 5 a 10 días a partir de
la salida de los huevos para que se complete el desarrollo de la fase L3 infectante;
estas larvas cuentan con una cutícula que las protege de las condiciones adversas
del medio ambiente para que luego pueda migrar a la parte superior de los pastos,
la oveja que se encuentra en pastoreo ingiere dichas larvas infectantes, estas se
mueven del rumen hacia el abomaso, entrando a las fosas gástricas y allí ocurre la
muda a L4. Las L4 se alimentan de sangre y mudan al estadio adulto en donde se
da el acoplamiento y la posterior expulsión de huevos. Como característica adicional
las L4 en sus etapas tempranas tienen la capacidad de hacer hipobiosis, inactivando
su desarrollo y evadiendo la respuesta inmune del huésped hasta que las
condiciones al interior y exterior sean las adecuadas para reanudar su desarrollo
incrementando la producción de huevos, este es un fenómeno muy común en ovinos
durante el periodo de periparto (Craig, 2009).
H. contortus, es considerado como el parásito que genera mayores pérdidas
económicas en las explotaciones ovinas ya que es capaz de producir lesiones en
las glándulas gástricas parasitadas como resultado de la penetración y crecimiento
de las larvas en su interior, dando lugar a una gran dilatación y marcada protrusión
sobre la superficie de la mucosa. Las células de las glándulas parasitadas son

23
reemplazadas por otras células no diferenciadas. Al salir las primeras larvas de la
mucosa, se aprecian alteraciones en las glándulas circundantes a las parasitadas,
la salida del parásito produce lisis en las células epiteliales del borde superior de las
glándulas, estimulando la rápida división celular y originando una marcada
hiperplasia con engrosamiento de la mucosa, edema submucoso y aumento de
células plasmáticas. En las infecciones por H. contortus, la hematofagia llevada a
cabo por las larvas a medida que van emergiendo de las glándulas es considerado
como el daño más grave. A los 35 días se ven claramente pequeñas úlceras con
hemorragias capilares. La parasitación del abomaso da lugar a la disminución de la
secreción de ácido clorhídrico, que facilita el incremento del pH gástrico y esta
modificación altera la digestión proteica ya que el pepsinógeno no se transforma en
pepsina, afectando el proceso digestivo perdiendo el efecto bacteriostático del pH
bajo, lo que aumenta el número de bacterias y facilita la aparición de diarreas
(Caracostantogolo, Martínez y Ribicich, 2007).
La especie T. axei, reside en el abomaso. Este es uno de los pocos nematodos
compartidos normalmente con el ganado. El ciclo de vida de estos es directo con
huevos que pasan en heces; de manera similar a H. contortus los ovinos cuando
están en pastoreo ingieren larvas infectantes que se desarrollan en las criptas de la
mucosa del abomaso emergiendo aproximadamente 15 días después de la
infección, larvas adultas L4.
Teladorsagia spp., se presenta a menudo como una infección mixta con
Trichostrongylus, afectando el abomaso e intestino delgado, causando enfermedad.

24
Las infecciones mixtas, con la participación de múltiples géneros y especies por
lo general tienen un mayor impacto en el hospedero que las infecciones mono
específicas (Wimmer, Craig, Pilkington y Pemberton, 2004), este parásito produce
huevos segmentados de concha fina que miden 90 × 50 micras y su ciclo de vida
es similar al de otros nematodos gastrointestinales mencionados con anterioridad;
es importante destacar que el ciclo de vida de los nematodos gastrointestinales más
importantes sigue un patrón similar (Levine, 1968).
Nematodirus spp., por su parte, se ubica en el íleon, presentando una forma
trenzada y estrías transversales. Los huevos miden 195 × 95 micras y son de color
café con lados paralelos. La oveja es considerada como huésped normal. Este
parásito estimula una respuesta inmune protectora temprana, y tiene la capacidad
de someterse a hipobiosis durante condiciones desfavorables (Craig, 2009); su ciclo
de vida es similar al de los parásitos ya mencionados, pero a diferencia de estos el
estadio L3 infectante se desarrolla dentro del huevo (Roeber, Jex y Gasser, 2013).

25
Figura 1. Representación del Ciclo de vida de NGI en pequeños rumiantes (Tomado y modificado de: Roeber, Jex y Gasser, 2013)
2.2. Patogenia y signos clínicos.
Existen tres grupos de ovinos que se pueden considerar más propensos a tener
altas cargas parasitarias: los jóvenes, por su escaso desarrollo inmunitario; los
adultos, ya que son animales inmunosuprimidos y los animales expuestos a altas
cargas parasitarias (Zajac, 2006). La gravedad de la enfermedad parasitaria está
directamente relacionada con factores como: la carga parasitaria, el estado general

26
e inmunológico del animal, estrés, el clima, el tipo de pasto carga de animales y la
dieta (Kassai, 1999).
La patogénesis de la enfermedad por NGI está ligada a una depresión en el
consumo de alimento voluntario (Fox, 1997), en estudios realizados por el mismo
autor se determina la posibilidad de que el incremento de gastrina a nivel sanguíneo,
puede contribuir a la disminución del apetito de los animales afectados por estos
parásitos.
Los principales efectos patógenos de dichos parásitos son causados por los
estadios adultos L4, que causan anemia grave ya que se alimentan de sangre, dicha
anemia suele hacerse evidente aproximadamente dos semanas después de la
infección (Baker et al., 1959). La enfermedad aguda es generalmente dependiente
de la intensidad y se asocia con signos de anemia hemorrágica, heces de color
oscuro, edema, debilidad, entre otros signos que ya han sido mencionados
anteriormente.
A diferencia de muchos otros parásitos gastrointestinales, H. contortus no es una
causa principal de diarrea, y sus efectos no son fácilmente detectables mediante la
observación de rutina (Zajac, 2006). Los signos primarios de la haemoncosis son
anemia e hipoproteinemia. El inicio de la enfermedad puede ser repentino, con
animales aparentemente sanos. En los casos crónicos puede haber pérdida de lana,
disminución de diámetro de la fibra, y pérdida de peso, las cavidades del cuerpo
pueden estar llenas de líquido, principalmente edema intermandibular, se presenta
también la palidez de las membranas mucosas y la materia fecal es de consistencia
variable (Craig, 2009).

27
Las infecciones con Trichostrongylus spp., causan graves daños a la mucosa
duodenal generando signos característicos de enteritis generalizada como edema y
pérdida de proteínas en el plasma con posterior hipoalbuminemia e hipoproteinemia
y hemorragia en el lumen intestinal (Taylor et al., 2007). T. axei altera la mucosa
gástrica, mientras que T. colubriformis, T. vitrinus, y T. rugatus causa atrofia de las
vellosidades en el intestino delgado anterior, por tal razón se genera pérdida de
proteínas a través del lumen del intestino y se altera la absorción de nutrientes
específicos, lo cual da lugar a la pérdida de masa corporal en corderos. En altas
cargas parasitarias se puede presentar diarrea acuosa prolongada, manchando la
lana de los miembros posteriores (Levine, 1968).
Teladorsagia spp., genera daño al huésped por la disminución de la producción
de HCL, la formación de edema de los pliegues abomasales así como descamación
de la mucosa, dando lugar a la hipoalbuminemia y niveles séricos elevados de
pepsinógeno (Fox, 1997).
2.3. Métodos de control de nematodos gastrointestinales.
El conocimiento de factores epidemiológicos tiene importantes implicaciones para
el control del parasitismo y el desarrollo de mejores regímenes de tratamientos
antihelmínticos estratégicos (Barger, 1999). En general, la medición de los efectos
de las infecciones parasitarias subclínicas, se realiza de forma rutinaria a partir de
parámetros de rendimiento, tales como aumento de peso, conversión alimenticia,
utilización de forraje, tasa de concepción, intervalo entre partos, producción de leche
y la resistencia a enfermedades (Corwin, 1997).

28
Según Hoste y Torres-Acosta (2011) existen tres principios fundamentales para
el control de parásitos gastrointestinales de rumiantes, como primera medida se
encuentra la reducción de la exposición a larvas en estadío infectante (L3), esto,
mediante pastoreo rotativo y disminución de la carga de animales; en segundo lugar
el desarrollo de respuestas más favorables por medio del hospedero, que se
consigue mediante la vacunación, la nutrición adecuada, selección genética y
mejoramiento de resistencia de los rebaños; el tercer principio va dirigido a la
reducción de las cargas parasitarias mediante la administración de antihelmínticos
tradicionales (sintéticos) o no tradicionales (vegetales o minerales).
2.3.1 Antihelmínticos convencionales.
Los antihelmínticos constituyen actualmente el principal método de control de los
nematodos de rumiantes en el mundo (Láñese, Gascon y Prichard, 1993).
Tabla 1. Antihelmínticos convencionales, mecanismo de acción y productos
comerciales.
GRUPO
FARMACOLÓGICO
PRINCIPIO
ACTIVO
MECANISMO DE ACCIÓN
PRODUCTO COMERCIAL
Imidazoles
Levamisol
Tetramisol
Parálisis espástica del
parasito por la liberación de
acetilcolina con efectos
muscarínicos y nicotínicos
Levamisol 15% VECOL®
Levamisol 3.5% Colvet®
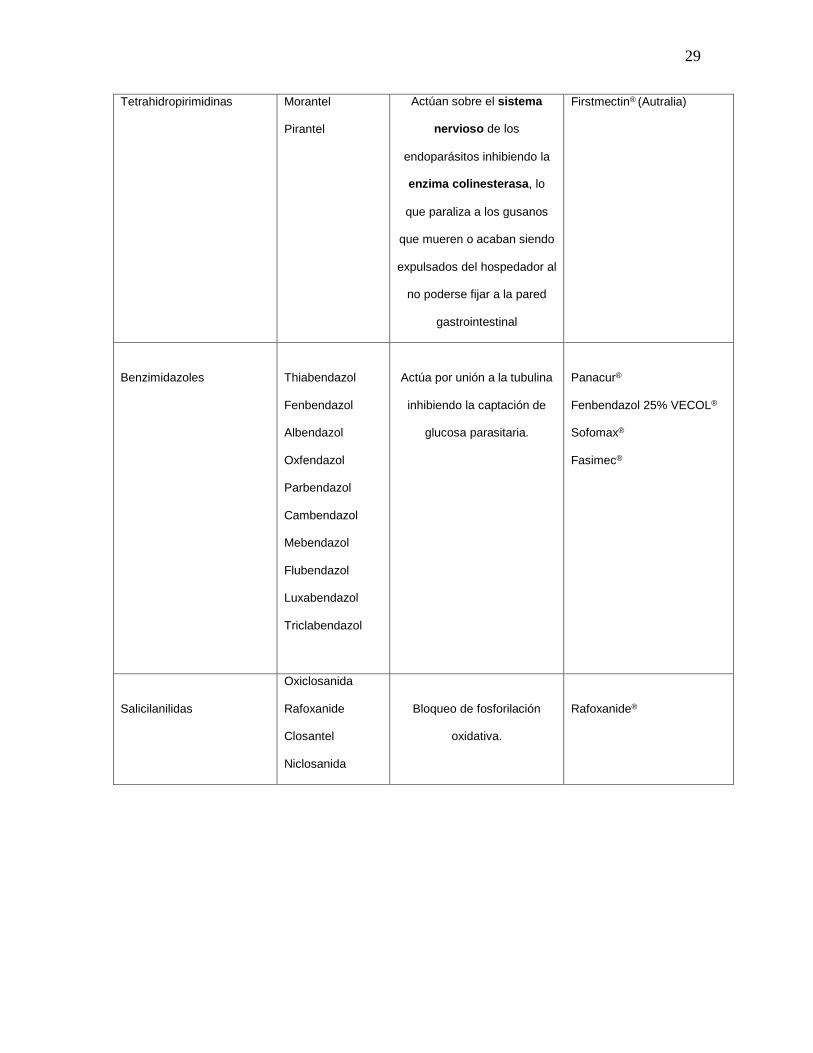
29
Tetrahidropirimidinas Morantel
Pirantel
Actúan sobre el sistema
nervioso de los
endoparásitos inhibiendo la
enzima colinesterasa, lo
que paraliza a los gusanos
que mueren o acaban siendo
expulsados del hospedador al
no poderse fijar a la pared
gastrointestinal
Firstmectin® (Autralia)
Benzimidazoles
Thiabendazol
Fenbendazol
Albendazol
Oxfendazol
Parbendazol
Cambendazol
Mebendazol
Flubendazol
Luxabendazol
Triclabendazol
Actúa por unión a la tubulina
inhibiendo la captación de
glucosa parasitaria.
Panacur®
Fenbendazol 25% VECOL®
Sofomax®
Fasimec®
Salicilanilidas
Oxiclosanida
Rafoxanide
Closantel
Niclosanida
Bloqueo de fosforilación
oxidativa.
Rafoxanide®

30
Ivermectinas/ Avermectinas
Abamectina
Doramectina
Moxidectina
Estimula la liberación del
GABA, agente
neurotransmisor inhibitorio
que actúa a nivel de las
neuronas presinápticas
parasitarias, produciendo la
muerte por parálisis flácida.
Abamectina 1% Bayer®
Doramec® L.A
Equest®
Ivermectinas/ milbemicynas
Ivermectina
Generan un incremento en la
permeabilidad de la
membrana celular a los iones
de cloro con la consecuente
hiperpolarización de la célula
nerviosa, resultando en una
parálisis y muerte del parásito
Ivomec®
Ivertin®
Virbamec®
A partir de 1980 se han realizado trabajos que evidencian los efectos negativos
que tienen en el ambiente algunos antihelmínticos como la fenotiazina que, aunque
en desuso, ejercía efectos adversos sobre el crecimiento del trébol, lo que se
traducía en una reducción del forraje y, por tanto, en una disminución de la
producción (Waller, 1999). En relación con los benzimidazoles, se ha sugerido que
éstos no son totalmente inocuos, habida cuenta del efecto residual indeseable que
tienen sobre algunos hongos saprofíticos que invaden las heces, especialmente el
fenbendazol y el oxfendazol, debido a que los mismos son excretados en gran
cantidad en las heces de los rumiantes (Waller, 1999). De los antihelmínticos que
se utilizan actualmente por los productores, son las ivermectinas/milbemycinas las

31
que probablemente ejercen más efectos negativos sobre el medio ambiente, en
poblaciones de insectos benéficos asociados al estiércol, principalmente en sus
formas larvarias (Papadopoulos, 2008).
El descubrimiento de nuevos compuestos químicos para el control de helmintos
en animales ha sido particularmente intenso con la introducción del thiabendazol
(Márquez, 2003), situación que produjo cambios radicales y profundo impacto en el
control del parasitismo gastrointestinal, dado el alto nivel de eficacia de estos
compuestos (Waller, 1999). Sin embargo, aunque el control de todos los nematodos
gastrointestinales se ha basado en el uso de antihelmínticos desde hace muchos
años (Wostenholme et al., 2004), se ha favorecido en gran medida la aparición y
desarrollo de la resistencia a éstos productos (Papadopoulos, 2008).
2.3.2. Métodos no convencionales.
2.3.2.1. Herbolaria.
Se ha establecido el uso de plantas bioactivas, que han demostrado afectar de
forma efectiva a gran mayoría de la población de parásitos en diversas etapas de
su ciclo de vida. Dichas propiedades bioactivas se atribuyen a los metabolitos
secundarios de las plantas, motivo por el cual estudios recientes han sugerido que
su utilización como nutracéuticos con propiedades antihelmínticas (Hoste et al.,
2006). Se han realizado hasta el momento estudios in vitro e in vivo en los cuales
se evaluó la actividad antihelmíntica de los taninos condensados evidenciando la
la alteración de la cutícula tras la exposición a dicho metabolito, además de retardar
el desarrollo larvario (Hoste et al., 2006).

32
2.3.2.2. Control biológico.
Actualmente se vienen realizando estudios de control biológico con hongos
nematófagos dentro de los cuales se encuentran Duddingtonia flagrans ofreciendo
buenas expectativas para su utilización, así como su posible introducción en
bloques minerales que contengan clamidosporas. Existen nematodos predadores
en el suelo que se alimentan de larvas infectantes de nematodos gastrointestinales;
por lo que se ha propuesto su empleo a fin de disminuir grandes poblaciones de
dichos parásitos que se encuentran en los potreros infestando los pastos (Quiroz et
al., 2011).
Se ha mencionado la actividad ovicida y/o sobre larvas en estadio temprano, de
bacterias prebióticas provenientes de cabras tales como Bifidobacterium DDBA,
Lactobacillus DDL19, Lactobacillus DDL48; pero su actividad se limita a infecciones
parasitarias moderadas (Quiroz et al., 2011).
2.3.2.3. Inmunización con vacunas.
La elaboración de vacunas con DNA que contienen específicamente antígeno H-
11 contra H. contortus. (Medina et al., 2014), ha sido tema de estudio y se han
obtenido resultados favorables en cuanto a su efectividad contra nematodos
gastrointestinales; dicha vacuna genera una respuesta inmune de protección sin
causar ningún daño al hospedero. Sin embargo es un área que requiere más
estudios a fin de determinar aspectos de bioseguridad animal, como de amplio
espectro de efectividad, evaluación de diferentes vías de administración, etc.
(Quiroz et al., 2011).

33
2.3.2.4. Agujas de cobre.
Se ha reportado la disminución significativa de cargas de H. contortus entre un 75 y
90% mediante el uso de capsulas (Martínez, 2010) y agujas (Aguilar et al., 2011) de
óxido de cobre, que liberan iones de cobre que ejercen un efecto antihelmíntico;
pero no se evidencia mejoría en la ganancia de peso, además, se ha demostrado la
acumulación de este mineral en el hígado (Aguilar et al., 2011 citado en Medina et
al., 2014).
2.4. Resistencia antihelmíntica.
El uso de antiparasitarios de relativo bajo costo, efectivos y de fácil aplicación ha
hecho posible el control de las plagas que afectan las especies productivas del
mundo entero, de esta forma el uso sistemático de químicos por los productores ha
conducido al desarrollo de la resistencia parasitaria (Nari, Eddi, Martins y Benavides,
2003).
La resistencia se ha definido cómo la disminución de la eficacia de un compuesto
químico contra una población de parásitos que, por lo general, es sensible a ese
medicamento. Es de naturaleza genética (Lara, 2007).
La resistencia puede ser intrínseca o adquirida (Mottier, Alvarez, Fairweather y
Lanusse, 2006). En la resistencia intrínseca, un parásito es naturalmente insensible
a un medicamento, debido a la ausencia de receptores o a la imposibilidad del
fármaco para entrar al sitio de acción de la misma, como ocurre en la resistencia de
los trematodos y cestodos a los endectocidas. La resistencia adquirida se presenta
en los parásitos que inicialmente son susceptibles a la acción terapéutica de un

34
fármaco, y posteriormente dejan de serlo luego de la ocurrencia de modificaciones
genéticas que son heredables de generación en generación (Márquez, 2003).
Larsen, Anderson, Ware y Fegely, (2006) demostraron que, a pesar de los
problemas de resistencia a los medicamentos, la productividad agrícola se puede
mantener o aumentar si se ajustan las técnicas de manejo de dichos productos,
pero además hacen hincapié en la necesidad desarrollar prácticas enfocadas al
manejo integrado de nematodos gastrointestinales.
Se han desarrollado líneas de investigación relacionadas con vías alternativas o
naturales para el control de parásitos gastrointestinales en rumiantes. Algunos de
estos métodos alternativos propuestos son de largo plazo, como la mejora genética
para la obtención de animales resistentes a parásitos gastrointestinales (Morris et
al., 2000) y el desarrollo de vacunas (Knox et al., 2003; Smith y Zarlenga, 2006).
2.5. Diagnóstico de nematodos gastrointestinales en ovinos.
El diagnóstico específico y sensible de infecciones por nematodos gastrointestinales
apunta al control efectivo de la enfermedad, que ahora es especialmente importante
debido a los problemas asociados con resistencia antihelmíntica en las poblaciones
de parásitos (Roeber, Jex y Gasser, 2013).
Las diferentes especies de nematodos gastrointestinales pueden ser variables
en su patogenicidad, distribución y susceptibilidad a diferentes antihelmínticos, por
lo que el diagnóstico preciso de infecciones parasitarias son de gran importancia,
ya que esta información apoya directamente a las estrategias de control de parásitos
(Roeber, Jex y Gasser, 2013).

35
Diversos métodos se emplean para el diagnóstico de infecciones por nematodos
gastrointestinales, siendo más comúnmente empleados: la observación de signos
clínicos indicativos de la enfermedad, el examen microscópico de las heces de los
huéspedes infectados y los enfoques de diagnóstico bioquímicos y / o serológicos;
se podría decir que estas técnicas son poco detalladas y por lo tanto carecen de
sensibilidad y especificidad (Kassai, 1999).
La enfermedad causada por nematodos gastrointestinales se manifiesta en una
variedad de signos clínicos que incluyen: desgrasado, anemia, pérdida de la
condición corporal y en casos graves la muerte. El tipo y el grado de manifestación
clínica también están influenciados por factores, como la especie y la carga de
nematodos que participan, el estado nutricional, inmunológico y reproductivo en el
cual se encuentre el huésped (Hungerford, 1990).
Para ayudar al diagnóstico de las infecciones por nematodos gastrointestinales,
se han empleado una serie de enfoques para la interpretación de los signos clínicos
vinculados a este tipo de parasitosis. Dentro de estos enfoques encontramos la
condición corporal del animal (Russel, Doney y Gunn, 1969), 'dag' (Larsen et al.,
1994) o la puntuación del grado de anemia FAMACHA (Van Wyk y Bath, 2002).
Estos métodos, aunque son útiles y con aplicabilidad a nivel de granja, son enfoques
muy subjetivos y carecen de especificidad, ya que los signos clínicos pueden
relacionarse con una muy amplia gama de enfermedades y problemas que afectan
al animal (Van Wyk y Bath, 2002).
Recientemente se ha establecido un método de evaluación en campo que se ha
considerado sencillo, económico y eficaz, en donde se realiza la evaluación de cinco
puntos específicos en el animal (Five Point Check©). El primer elemento a evaluar

36
es la nariz, ya que una secreción clara o incluso purulenta puede ser indicativo de
presencia de gusanos nasales; el segundo son los ojos, este punto hace referencia
al método FAMACHA mencionado anteriormente, su limitación se presenta al no ser
aplicable en detección de parásitos no hematófagos; el tercero es la mandíbula, la
formación de edema frío intermandibular es un firme indicativo de hipoproteinemia
causada por parásitos hematófagos; el cuarto es la condición corporal que pudiera
relacionarse principalmente con parásitos como Teladorsagia spp., Trichostrongylus
spp., Nematodirus spp, y Oesophagostomum columbianum; el quinto elemento a
evaluar es la cola, ya que el parasitismo es una de las principales causas de diarrea,
lo que puede ser de ayuda para determinar los animales a tratar de forma selectiva
(Bath y Van Wyk, 2009).
Figura 2. Puntuación de tarjetas utilizadas para evaluación de los cinco puntos. (Tomado y modificado de: Bath y Van Wyk, 2009).

37
El conteo de huevos en las heces es el método más común para el diagnóstico
de infecciones por nematodos gastrointestinales y se emplea de forma rutinaria en
parasitología; dentro de los métodos empleados para dicho fin se encuentran el
método de flotación por medio de centrifugación (Lane, 1922), la técnica de dilución
Stoll (Stoll, 1923), el método de McMaster (Gordon y Whitlock, 1939) y el método
de flotación Wisconsin (Cox y Todd, 1962), de los cuales el método de McMaster es
el más ampliamente utilizado (Nicholls y Obendorf, 1994). Las aplicaciones de estos
estos métodos incluyen: la determinación del grado de contaminación con huevos
de helmintos (Gordon, 1967), la estimación de intensidad de la infección (McKenna,
1987), la evaluación de la eficacia de lo antihelmínticos (Waller, 1999) y como un
medio de orientación para las decisiones de control y tratamiento (Brightling, 1988).
En los casos de infecciones mixtas se han empleado los métodos de cultivos
fecales con el fin de identificar los diferentes géneros de nematodos presentes en
dichas infecciones, estos métodos se han desarrollado para identificar los estadios
tempranos L1 o L3 infectivas de nematodos estrongilidos, los más utilizados implican
la incubación de material fecal para la obtención de L3 y su posterior diferenciación
morfológica (Roeber, Jex y Gasser, 2013).

38
Figura 3. Larvas (L3) de especies representativas de nematodos gastrointestinales en ovejas, encontradas tras el cultivo larvario (LC). (Roeber, Jex y Gasser, 2013) Otras alternativas diagnosticas tienen enfoques bioquímicos e inmunológicos
desarrollados con el fin de proporcionar un diagnóstico específico de infección.
Estos métodos se basan principalmente en la medición de parámetros como
detección de pepsinógeno sérico elevado, ya que las glándulas de la mucosa
gástrica al ser destruidas por el parásito, disminuyen la producción de ácido
clorhídrico por parte de las células parietales, causando un incremento en el pH del
abomaso resultando en la alteración de la transformación de pepsinógeno a pepsina
activa (Levine, 1968). Por lo tanto, un aumento en la concentración de pepsinógeno
sérico podrá relacionarse con daños en la mucosa. Sin embargo la existencia de
otras enfermedades parasitarias o no parasitarias pueden ser las responsables de
un aumento moderado de las concentraciones de pepsinógeno en la sangre, lo que
limita la especificidad de esta prueba. Los niveles de gastrina o anticuerpos

39
circulantes podrían ser indicativos de infecciones parasitarias ya que los nematodos
estrongilidios pueden estimular las células T, aumentando como consecuencia la
producción de gastrina, pero al igual que la prueba anteriormente mencionada, tiene
poca especificidad ya que factores tales como la dieta, la lactancia o lesiones
abomasales, también pueden afectar los niveles de gastrina. (Berghen et al., 1993).
Se han establecido también métodos diagnósticos de medición inmunológica
que se basan en la detección de la respuesta inmune de los animales afectados y
los antígenos del parasito (Ogunremi et al., 2008). Este método diagnóstico puede
clasificarse como directo o indirecto.
Los métodos inmunológicos directos proporcionan evidencia de una infección y
pueden estar basados en la detección de antígenos de parásitos que se encuentran
presentes en la circulación y en los excrementos de los huéspedes infectados
(Cohen y Sadun, 1976). Los métodos inmunológicos indirectos están basados en la
detección de anticuerpos anti-parásitos o respuestas inmunes mediadas por células
del huésped infectado. Dentro de estas pruebas se encuentran: la prueba de fijación
del complemento, inmunofluorescencia indirecta, hemaglutinación indirecta y ELISA
siendo este último, el más utilizado (Doenhoff, Chiodini y Hamilton., 2004).
Existen técnicas moleculares que son consideradas como alternativas viables
para la identificación de especies, así como las diferencias a nivel molecular dentro
de una especie (cepas) de parásitos. Dichas técnicas se basan en las diferencias
en DNA determinadas mediante técnicas moleculares como PCR, AFLP, RAPD,
RFLP, PCR-SSCP, PCR en tiempo real. Estas técnicas también se han empleado
como medio para diferenciar y evaluar la diversidad genética entre población de
parásitos (Ahmed et al., 2011).

40
2.6. Fitoterapia.
La fitoterapia es un neologismo empleado por Henri Leclerc, médico francés (1870-
1955), en los comienzos de siglo, desde entonces la palabra fitoterapia es utilizada
para designar la utilización de las plantas medicinales con fines terapéuticos, que
serviría más tarde para diferenciarla de la forma de curar actual; la medicina
sintética o convencional (Avello y Cisternas, 2010).
La Fitoterapia consiste en el empleo de plantas medicinales para fines
terapéuticos. Según la OMS (1978 citado en Palomo y Revuelta, 2012) son: “todo
aquel vegetal que contiene uno o varios principios activos que pueden ser utilizados
en el tratamiento curativo o paliativo de determinadas enfermedades” basada en
que su capacidad terapéutica depende de varios factores como los principios
activos, su distribución en todas las plantas, la existencia de plantas que combinen
estos principios y además el conocimiento de las propiedades terapéuticas de cada
uno de ellos y en qué especie se encuentran (Palomo y Revuelta, 2012).
Para situar los límites de la fitoterapia en la Terapéutica actual, y por lo tanto el
concepto de la misma, se debe partir de tres premisas:
- Si bien los productos fitoterápicos suelen tener márgenes terapéuticos más
amplios y suelen tener menos efectos adversos que los fármacos sintéticos, natural
no es sinónimo de inocuo.
- Actualmente existe una base científica que sustenta la eficacia de muchos
productos fitoterápicos para determinadas indicaciones que constituyen alternativas

41
fuertemente deseables para muchas patologías menores, enfermedades crónicas y
prácticas profilácticas.
- La eficacia se consigue solamente con el uso adecuado de los preparados
fitoterápicos, tanto en lo que tiene que ver con las indicaciones, como con la forma
de administración y la dosificación. La responsabilidad del profesional sanitario en
este aspecto, y por ende de su preparación curricular, es insoslayable. También se
requiere una legislación adecuada. Por lo tanto no se debe maximizar ni minimizar
las posibilidades de la fitoterapia, sino tener en cuenta que el lugar que debe ocupar
en la terapéutica es ni más ni menos que aquel para el cual ha demostrado su
utilidad (Palomo y Revuelta, 2012).
2.6.1 Características de la fitoterapia.
La base de los medicamentos fitoterápicos son las drogas vegetales y los diferentes
tipos de productos que de ellas se obtienen. El término droga vegetal no debe
confundirse con el de planta medicinal. La OMS definió en (1978 citado en
Cañigueral, Dellacassa y Bandoni, 2003), estos conceptos como se indica a
continuación:
-Planta medicinal es cualquier planta que en uno o más de sus órganos contiene
sustancias que pueden ser usadas con finalidad terapéutica o que son precursores
de la semisíntesis químico-farmacéutica.
- Droga vegetal es la parte de la planta medicinal utilizada en terapéutica
- Principios activos son las sustancias responsables de la acción farmacológica
(Cañigueral, Dellacassa y Bandoni, 2003).

42
La fitoterapia utiliza matrices vegetales complejas. Estas matrices las constituyen
plantas enteras, partes de ellas (hojas, tallos, raíces, etc.) y también productos de
éstas, resultados de tratamientos directos con algún disolvente o medio que concen-
tre los compuestos afines y facilite su administración, son los llamados extractos.
En cualquier caso en esta matriz compleja nos encontramos con un sin número de
compuestos de diferente naturaleza química, a esta mezcla se la llama fitocomplejo
(Hoffmann, 1992, citado en Avello y Cisternas 2010).
El fitocomplejo es la mezcla de sustancias activas y otras acompañantes que
actúan en conjunto para lograr un mismo fin terapéutico, que no sería el mismo si
se administraran por separado, o sea como monosustancias (Muñoz, Montes y
Wilkomirsky, 1999 citado en Avello y Cisternas 2010).
2.6.2 Historia de la fitoterapia.
El Dr. Auguste Soins en el año de 1865 define con el término fitoterapia la terapia
que utiliza las plantas medicinales y Dr. Henri Lecler francés fue quien la popularizó
entre la clase médica y científica a principios del siglo XX. Fitoterapia deriva del
griego “Phuton”, planta y de “Therapeia”, tratamiento, e incluyen otras terapias
especiales como la aromaterapia (con aceites esenciales) y la gemoterapia (con
yemas de tejidos vegetales). La historia propiamente dicha de las plantas se inicia
con la realización de los primeros herbarios y su constatación por escrito; según
Campbell datarían de los asirios (Berdonces, 2003).
El primer texto escrito sobre la terapia con plantas se encuentran en unas
tabletas de arcilla grabadas en escritura cuneiforme de la época de los sumerios,
datadas aproximadamente en el año 3000 a. C., donde se citan las propiedades

43
calmantes de la adormidera (Papaver somniferum). Los egipcios iniciaron la
fitoterapia más “racional” conllevando a una sistematización de los diferentes
remedios vegetales, el papiro de Ebbers (1500 a. C) y el de Edwin Smith (1600 a.
C.) son quizás los más conocidos que hacen referencia al uso de las plantas
medicinales. El más antiguo conocido es el papiro Kahun (1900 a.C.) y relata
remedios en relación con las enfermedades femeninas y como asistir un parto.
Hipócrates atribuye el desarrollo de los medicamentos a observación, molienda,
mezcla, cocción, combinando los remedios débiles para adaptarlos a la constitución
y fuerza del individuo. De las 257 plantas mencionadas en los trabajos hipocráticos,
tan sólo 27 no se siguen utilizando como tales hoy en día. En la edad media se
siguió el saber de los antiguos como Galeno, Cratevas, Dioscórides o Plinio, en el
siglo IV se escribió el Pseudo-apuleyo (Herbarius apuleius) uno de los más usados
en Europa Central que contiene las descripciones y recetas con más de 100 plantas
medicinales. La llegada de la imprenta en el renacimiento presentó un desarrollo de
los textos de fitoterapia, muchos de ellos compilaciones exhaustivas de textos
manuscritos más antiguos.
Una gran cantidad de autores contribuyeron al inicio de la botánica sistemática
intrínsecamente ligada a la sistematización de todas las especies de seres vivos.
Destacando por encima de todos ellos a Linneo, aceptado como el padre de ésta
clasificación, otros autores contribuyeron al desarrollo de esta sistematización y en
el campo de la botánica se distinguieron los franceses Lamarck y De Candolle.
Los autores españoles también contribuyeron con sus escritos, al desarrollo de
la fitoterapia entre estos encontramos a Nicolás Monardes, Andrés de Laguna y
Francisco Hernández (Berdonces, 2003).

44
En la actualidad, la utilización de plantas medicinales, ha alcanzado un gran auge
como recurso terapéutico alterno para afrontar las carencias de la medicina química,
debido a que la mayoría de ellas, tienen propiedades terapéuticas; sin embargo,
otras tienen algún factor que les confiere propiedades tóxicas, cuando son utilizadas
de manera inadecuada, produciendo efectos y/o reacciones adversas (Osorio, 2010
citado en Martínez, Meza y Guevara 2013).
2.6.3 Futuro de la fitoterapia.
Una de las ventajas de la fitoterapia es que puede aportar un elevado número de
drogas vegetales y preparaciones que se han empleado de forma empírica durante
siglos. Los avances en la tecnología actual permite a los investigadores optimizar la
eficacia, la normalización y la valoración clínica de todos estos medicamentos de
uso tradicional, para cumplir con los requisitos exigidos por los estándares
internacionales actuales; es por eso que se deben dirigir todos los esfuerzos a la
investigación del perfil químico-farmacológico de extractos y combinaciones para
racionalizar su aplicación terapéutica. Cuatro áreas de investigación determinarán
el próximo futuro de la fitoterapia: los métodos de alta tecnología para realizar los
análisis químicos y la estandarización de los extractos vegetales; la búsqueda de
otros compuestos bioactivos para el desarrollo de medicamentos; la integración de
los nuevos métodos de biología molecular para el tamizado farmacológico de las
plantas y sus constituyentes y el desarrollo de estudios clínicos protocolizados y de
ensayos de biodisponibilidad de los extractos vegetales estandarizados (Wagner,
2006).

45
2.7. Metabolitos secundarios de las plantas.
Son compuestos vegetales que no son estrictamente esenciales para las funciones
principales de las plantas, tales como su crecimiento y reproducción, pero han sido
asociados con los mecanismos de defensa de las plantas contra los herbívoros
(Harborne 1999; Karban et al., 1999 citado en Athanasiadou y Kyriazakis, 2004).
Partes de plantas o extractos ricos en metabolitos secundarios se han utilizado
para combatir el parasitismo durante muchos siglos y en varias partes del mundo
este tipo de productos se siguen utilizando para este fin. Existe evidencia de los
efectos antiparasitarios de éstos metabolitos secundarios contra helmintos, como
nematodos y también contra protozoos y parásitos externos (Instituto Internacional
de Reconstrucción Rural, 1994 citado en Athanasiadou y Kyriazakis, 2004).
La variación en el contenido de metabolitos secundarios de plantas es el
resultado de muchos factores; como el fenotipo de la planta (fenotipo y variedad),
las diferentes características ambientales (radiación solar y disponibilidad de agua),
la velocidad de crecimiento, madurez, condición nutricional del suelo, depredación
y las enfermedades (Waterman y Mole, 1994), así mismo, la aparición de los
compuestos secundarios está relacionada con los mecanismos de defensa de la
planta y los efectos del suelo y del clima, puede haber un componente genético u
otras variaciones pero el genotipo puede ser modificado por una variedad de
características bióticas y abióticas, como alteraciones en la bioquímica causadas
por el cambio de patrones de asignación de recursos que reflejen las diferentes
demandas fisiológicas asociadas con el crecimiento, la defensa y la reproducción
(Bowers & Stamp, 1992 citado en Animan y Ebrahimzadeh, 2002).

46
No todos los metabolitos secundarios se encuentran en todos los grupos de
plantas, éstos son sintetizados en pequeñas cantidades y no de forma generalizada,
estando a menudo su producción, restringida a un determinado género de plantas,
a una o varias familias, o incluso a algunas especies, teniendo funciones ecológicas
específicas como atrayentes o repelentes de animales; muchos son pigmentos que
proporcionan color a flores y frutos, jugando un papel esencial en la reproducción
atrayendo a insectos polinizadores o atrayendo a diversos animales que van a
utilizar los frutos como fuente de alimento, contribuyendo a la dispersión de semillas.
Otros compuestos tienen función protectora frente a predadores, actuando como
repelentes, proporcionando a la planta sabores amargos, haciéndolas indigestas o
venenosas. Intervienen en los mecanismos de defensa de las plantas contra una
gran variedad de patógenos, actuando como pesticidas naturales. Se agrupan en 4
clases principales: Terpenos entre los que se encuentran las hormonas, pigmentos
o aceites esenciales; Compuestos fenólicos como cumarinas, flavonoides, lignina y
taninos; saponinas, glicósidos cardiacos, glicósidos cianogénicos y glucosinolatos;
y Alcaloides (García y Pérez, 2009).
2.8. Taninos.
Los taninos son compuestos fenólicos secundarios de elevado peso molecular (500
a > 20000) presentes en la naturaleza, que se encuentran frecuentemente en frutas,
árboles,en forrajeras templadas principalmente leguminosas, y otras especies como
sorgo y maíz utilizadas comúnmente en la alimentación del ganado (Otero y Hidalgo,
2004).

47
Los taninos son almacenados dentro de los tejidos epidérmicos y sub epidérmicos
de las hojas, pero se pueden encontrar dentro del pericarpo de los frutos y raíces.
A nivel celular, los taninos hidrosolubles, predominan dentro de la pared celular y
entre los espacios intracelulares. Los taninos condensados, son almacenados en la
vacuolas intracelulares en forma libre y en proporciones variables, vinculadas a las
fibras (lignina) o proteínas de las paredes celulares (Frutos et al., 2002).
2.8.1 Estructura química.
Los taninos se dividen en 2 clases: condensados e hidrolizables, sobre la base de
su estructura química (Hagerrman y Butler, 1989 citado en Hagerman et al., 1992).
Los taninos condensados son compuestos de polímeros de flavonoides, que
pueden ser oxidativamente degradados en ácido para producir antocianidinas. Los
taninos hidrolizables son polímeros de ácido gálico o de ácido hexahidroxidifenólico,
ésteres de glucosa y otros polifenoles.
Las principales diferencias estructurales existentes entre los taninos condensados
e hidrolizables afectarían la actividad de los taninos. Los principales efectos estarían
dados por la interacción con la proteína por parte de los taninos condensados, no
siendo así con los taninos hidrolizables, ya que estos últimos son rápidamente
degradados en grupos fenólicos más pequeños, incapaces de reaccionar con las
proteínas del medio (Hagerman et al., 1992).
Cada proteína tiene su pH distintivo y óptimo para unirse a los taninos
condensados, por ejemplo la albúmina del suero bovino puede unirse en un pH de
4.1 a 6.1 y disociarse cuando el pH es menor a 3.5 (Jones y Mangan, 1997, citado
en Márquez y Suárez, 2008).

48
En general, en un rango de pH de entre 5 y 7,5 en el rumen e intestino delgado,
la proteína permanece unida a los taninos, pero a pH bajos (pH < 3,5) la proteína
ligada por los taninos condensados es liberada (Hagerman et al., 1992).
Los taninos hidrolizables pueden ser hidrolizados por calentamiento con ácido
débil (Figura 4). En contraste, los taninos condensados pueden ser degradados por
oxidación solamente por el ácido mineral caliente (Figura 5). Los factores que
favorecen la formación de complejos incluyen su peso molecular relativamente alto
y su gran flexibilidad estructural. Las proteínas que muestran la mayoría de afinidad
por los taninos son relativamente grandes e hidrófobas, además de poseer una
estructura abierta, flexible y son ricas en prolina (Bunglavan y Dutta, 2013).
Los taninos hidrolizables tienen en su estructura química un carbohidrato en
posición central, generalmente d-Glucosa y presentan bajo peso molecular (500-
3,000 Daltons), haciéndolos más solubles en agua y de fácil absorción en el intestino
delgado, convirtiéndose en potencialmente tóxicos para los rumiantes cuando son
suministrados en grandes cantidades sin el suficiente tiempo de adaptación de la
flora microbial (Min et al., 2003, citado por Márquez y Suárez, 2008).

49
Figura 4. Tanino hidrolizable (gallotannin) compuesto por ésteres de ácido gálico
vinculados a un núcleo de azúcar (Naumann et al., 2013).
Figura 5. Estructura básica de un tanino condensado mostrando estereoquímica,
vínculos interflavánicos e hidroxilación del anillo B (Naumann et al., 2013).
En general, la concentración de los taninos presentes en la naturaleza muestra
una gran variabilidad. Los taninos que se producen en forrajes templados tienen una
masa molecular relativa de 2000 a 4000 que comprende de 10 a 12 oligómeros de
taninos condensados (Foo et al., 1986, citado en Min, Miller y Loetz, 2003)
50
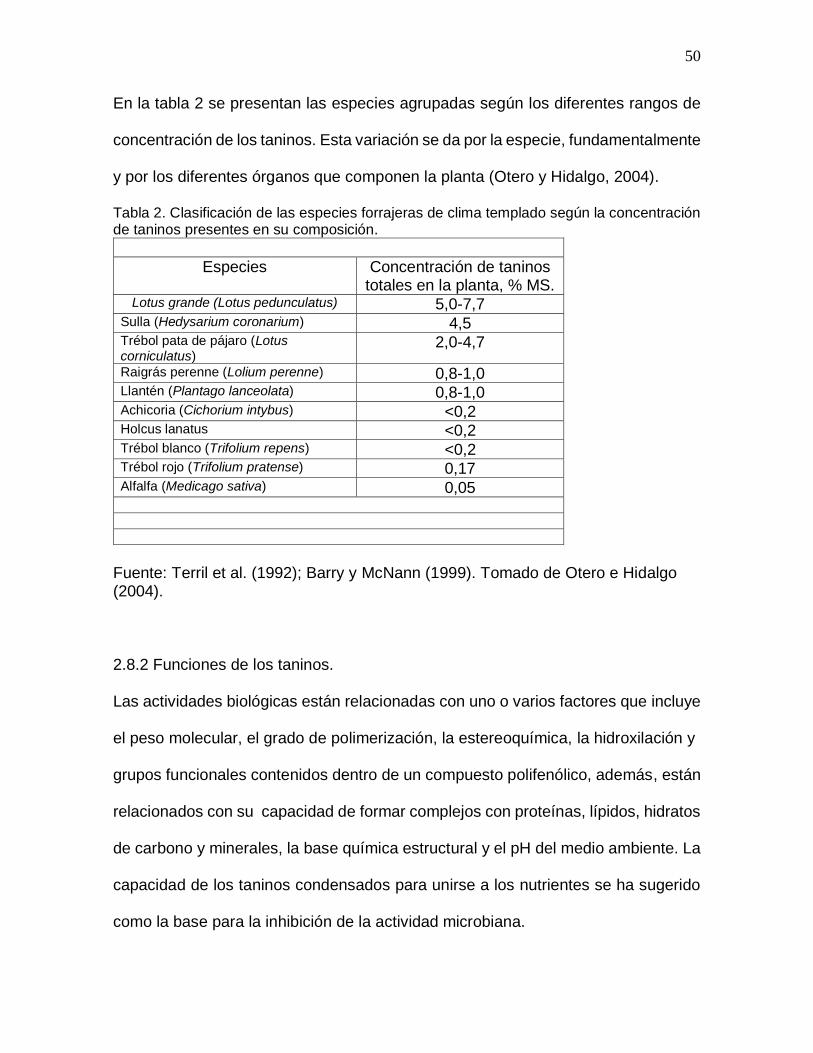
En la tabla 2 se presentan las especies agrupadas según los diferentes rangos de
concentración de los taninos. Esta variación se da por la especie, fundamentalmente
y por los diferentes órganos que componen la planta (Otero y Hidalgo, 2004).
Tabla 2. Clasificación de las especies forrajeras de clima templado según la concentración de taninos presentes en su composición.
Especies Concentración de taninos totales en la planta, % MS.
Lotus grande (Lotus pedunculatus) 5,0-7,7 Sulla (Hedysarium coronarium) 4,5 Trébol pata de pájaro (Lotus corniculatus)
2,0-4,7
Raigrás perenne (Lolium perenne) 0,8-1,0 Llantén (Plantago lanceolata) 0,8-1,0 Achicoria (Cichorium intybus) <0,2 Holcus lanatus <0,2 Trébol blanco (Trifolium repens) <0,2 Trébol rojo (Trifolium pratense) 0,17 Alfalfa (Medicago sativa) 0,05
Fuente: Terril et al. (1992); Barry y McNann (1999). Tomado de Otero e Hidalgo (2004). 2.8.2 Funciones de los taninos.
Las actividades biológicas están relacionadas con uno o varios factores que incluye
el peso molecular, el grado de polimerización, la estereoquímica, la hidroxilación y
grupos funcionales contenidos dentro de un compuesto polifenólico, además, están
relacionados con su capacidad de formar complejos con proteínas, lípidos, hidratos
de carbono y minerales, la base química estructural y el pH del medio ambiente. La
capacidad de los taninos condensados para unirse a los nutrientes se ha sugerido
como la base para la inhibición de la actividad microbiana.

51
La inhibición es el resultado de la interacción directa de los taninos condensados
con constituyentes de la pared celular microbiana y de la interacción indirecta con
nutrientes haciendo que no se encuentren disponibles para los microorganismos
(Smith, Zoetendal y Mackie., 2005).
Efectos de las actividades de los taninos condensados se han observado en la
digestión de rumiantes, el ciclo de nutrientes en el medio ambiente y como un
nutraceútico componente de un alimento que proporciona beneficios para la salud
relacionados con el tratamiento y la prevención de la enfermedad en los seres
humanos y el ganado (Naumann et al., 2013).
2.8.3 Ventajas y desventajas.
Los taninos pueden comportarse como beneficos o perjudiciales para los rumiantes
dependiendo de cuáles y cuanta cantidad se ingiere, su estructura y peso molecular
y la fisiología de las especies que lo consumen (Hagerman y Butler, 1991 citado en
Frutos, Hervás, Giráldez y Mantecón., 2004).
2.8.3.1 Efecto de los taninos sobre el consumo voluntario del alimento
Los taninos pueden tener efectos positivos o negativos sobre el valor nutritivo de los
forrajes según la concentración en la que se encuentren. Si se presenta en altas
concentraciones deprimen el consumo voluntario en los animales, la palatabilidad y
la digestibilidad. En cambio cuando la proporción de taninos es moderada o baja (2
a 4% de la MS), su efecto es beneficioso, previniendo infecciones y aumentando la
distribución de Nitrógeno no amónico y de los aminoácidos esenciales desde el
rumen (Scribano, 2012).

52
El consumo de altas concentraciones de taninos condensados (> 7% MS) ha
sido asociado con un número de efectos adversos en rumiantes, como la reducción
de la ingestión del alimento, la inhibición del crecimiento y la interferencia con la
morfología y la actividad proteolítica de los microbios en el rumen, mientras bajas o
moderadas concentraciones (< 6% MS) han demostrado efectos positivos en los
herbívoros, por ejemplo: incremento del peso y la producción de leche ( Min et al.,
2003; Waghorn y Mc Nave, 2003 citado en Hoste et al., 2006).
Frutos et al. (2004) no encontró ninguna reducción en el consumo de alimento
voluntario entre ovejas a las que se les proporcionó una alimentación que contenía
Harina de soja tratada con HT (20,8 g HT kg-1 MS de alimentación). Sin embargo,
en un experimento con ovejas alimentadas con 8 g de ácido tánico por Kg de peso
vivo, el consumo voluntario cayó drásticamente después de 24 h (desde 18 a 2,5 g
MS Kg de peso vivo) (Zhu, Filippich y Alsalam., 1992).
Tres mecanismos principales se han sugerido para explicar los efectos negativos
de las altas concentraciones de taninos en el consumo voluntario, una reducción de
la palatabilidad de la alimentación, el enlentecimiento de la digestión y el desarrollo
de aversiones condicionadas (Frutos et al., 2004).
- La reducción en la palatabilidad de los vegetales que contienen dichos
compuestos que podría deberse a una reacción de los taninos con las
proteínas de la saliva y las de la membrana mucosa de la boca durante la
masticación o directamente con los receptores gustativos provocando una
sensación de astringencia que aparece por la formación de complejos entre
los taninos y las glicoproteínas de la saliva (Mcleod, 1974).

53
Numerosas especies de herbívoros basan su alimentación en especies
vegetales con un alto contenido de taninos, habiéndose observado en su
saliva la presencia de proteínas ricas en el aminoácido prolina (Foley, Iason
y MacArthur., 1999; McMahon et al., 2000), con una elevada capacidad de
unirse a los taninos, los complejos taninos-proteínas ricas en prolina, al
contrario de los demás complejos tanino-proteína, son estables en todo el
rango de PH del tracto digestivo, pudiendo anular su efecto negativo sobre la
palatabilidad y por lo tanto, sobre la ingestión de alimento, minimizando las
pérdidas de nitrógeno fecal (McMahon et al., 2000).
Se cree que los herbívoros “ramoneadores” secretan proteínas ricas en
prolina de forma permanente, mientras que en el ganado ovino por ejemplo,
únicamente podría inducirse su producción en respuesta al consumo de
vegetales con alto contenido de taninos (Cheeke y Palo, 1995 citado en
Ramos, et al., 1998; Makkar y Becker, 1998).
- Un enlentecimiento de la digestión de la materia seca en el rumen, éste
perjudicaría el vaciado del tracto digestivo, generando signos de repleción y
actuando a modo de retro señal sobre los centros nerviosos involucrados en
el control de la ingestión (Waghorn, Shelton y McNabb., 1994).
Narjisse, Elhonsali y Olse. (1995) Infundieron taninos extraídos de hojas
de roble (Puercos ilex) directamente en el rumen de ovejas y cabras para
determinar si los factores independientes de la palatabilidad fueron
encargados de la reducción voluntaria en el consumo de alimento. La
desaceleración de la digestión de la materia seca en el rumen perjudica el
vaciado del tracto digestivo, generando señales de que el animal está lleno

54
y proporcionando una retroalimentación a los centros nerviosos implicados en
el control de la ingesta.
- El desarrollo de aversiones condicionadas, el tercer mecanismo mediante el
cual los animales reducen el consumo de alimentos que contienen taninos u
otros compuestos que puedan resultar tóxicos, se basa en la identificación de
las consecuencias negativas post-prandiales de su ingestión y el desarrollo
subsiguiente de aversiones condicionadas (Waghorn, 1996; Launchbaugh,
Provenza y Piste., 2001), por lo que parece probable que las consecuencias
Post-prandiales de la ingestión de alimentos ricos en taninos están mediados
por factores relacionados con fermentación microbiana (Waghorn, 1996).
2.8.3.2 Efectos sobre la digestibilidad de los alimentos
Los taninos reducen la digestibilidad de la dieta afectando las proteínas y también
a otros componentes de los piensos en diferentes grados (Kumar y Singh, 1984
citado en Frutos et al., 2004).
Su principal efecto sobre las proteínas se basa en su capacidad para formar
enlaces de hidrógeno que son estables entre un pH de 3,5 y 8 (aproximadamente).
Estos complejos se disocian cuando el pH cae por debajo de 3,5 (como en el
abomaso, pH de 2,5 – 3) o es mayor de 8 por ejemplo en el duodeno, lo que explica
la actividad de los taninos en las vías digestivas (McLeod, 1974; Hagerman et al.,
1992).

55
Las modificaciones de la digestibilidad como consecuencia de los taninos, se
asocia fundamentalmente con cambios en el patrón de fermentación ruminal pero
también con cambios en la digestibilidad intestinal posterior (Waghorn, 1996).
Otro efecto de la unión de los taninos condensados a las proteínas en la digestión
de los rumiantes es el cambio en la excreción de nitrógeno de amoniaco urinario
(NH3-N) a nitrógeno fecal. Pérez – Maldonado y Norton (1996) Demostraron que el
nitrógeno fecal aumenta 14% cuando las ovejas y cabras son alimentadas con
dietas que contienen D. intortum y C. calothyrsus a una concentración de 300 g/kg
de materia seca. El aumento en la excreción fecal de nitrógeno podría ser el
resultado de una disminución de la absorción de nitrógeno por los rumiantes que lo
consumen; sin embargo estos autores observaron el efecto contrario cuando las
ovejas y cabras fueron alimentadas con dietas que contienen taninos condensados
que absorben el 19% más nitrógeno que aquellas que consumen una dieta libre de
taninos condensados.
Los taninos condensados reducen la producción de CH4 (metano) por los
rumiantes (Tedeschi et al., 2011 citado en Naumann et al., 2013).
Los pesos moleculares pueden ser un factor en la producción de CH4 (metano),
Huang et al. (2011) compararon la producción in vitro de metano del ganado
alimentado con especies de Leucaena con taninos condensados de varios pesos
moleculares que variaron desde 442 hasta 1566 Daltons. El tanino condensado de
mayor peso molecular produjo cantidades más pequeñas de gas y metano, lo que
sugiere que la digestión de la materia orgánica en general se redujo y que el tipo de
tanino condensado presente afectó indirectamente las poblaciones microbianas
metanogénico en el rumen.

56
2.8.3.2.1 Efectos en la fermentación ruminal
El rumen se caracteriza por poseer un pH 6 - 7. A esta gama de pH, los complejos
que se forman entre los taninos condensados y las proteínas son estables (Butter
et al., 1999 citado en Abdu et al., 2012) y contribuyen a proteger las proteínas de la
degradación en el rumen. La formación de complejos entre los taninos condensados
y las proteínas alimentarias o la fijación de los mismos a las enzimas bacterianas
reducen globalmente la proteólisis ruminal (Min et al., 2003).
En general esta reducción en la degradación de la proteína está asociada con
una menor producción de nitrógeno amoniacal y un mayor flujo de nitrógeno sin
amoníaco al duodeno (Waghorn, 1996).
El efecto de los taninos, tanto condensados como hidrolizables sobre la
degradación de la proteína consiste en una reducción de la fracción inmediatamente
degradable y, básicamente en una reducción del ritmo fraccional de degradación
(Reed, 1995).
Los mecanismos por los cuales los taninos reducen la degradación ruminal
de los diferentes componentes de la dieta no son del todo conocidos. Entre los más
aceptados están la privación del sustrato (Scalbert, 1991; McAllister et al, 1994b;
McMahon et al., 2000, citados en Frutos et al., 2004), inhibición de la enzima (Barry
y Manley, 1984; Bae et al., 1993; Jones et al., 1994 citados en Frutos et al., 2004) y
acción directa sobre microorganismos ruminales (Leinmüller et al., 1991; Scalbert,
1991 citados en Frutos et al., 2004).

57
- Privación del Sustrato: La presencia de taninos condensados puede provocar
una precipitación de las enzimas extracelulares de los microorganismos ruminales,
impidiendo así la adhesión a los alimentos (Barrey y Manley, 1984).
Los taninos condensados son agentes quelantes por lo que podrían llegar a
reducir la disponibilidad de determinados iones metálicos que son necesarios para
el metabolismo de los microorganismos ruminales (McMahon et al., 2000).En este
sentido, Scalbert (1991) observó cómo la presencia de taninos oxidados tenía un
claro efecto tóxico sobre las bacterias metanogénicas.
- Inhibición enzimática: La capacidad de los taninos condensados para unirse a
los nutrientes se ha sugerido como la base para la inhibición de la actividad
microbiana. La inhibición es el resultado de la interacción directa de los taninos
condensados con constituyentes de la pared celular microbiana, y la interacción
indirecta de los mismos con nutrientes haciendo que no estén disponibles para los
microorganismos (Smith et al., 2005).
Nelson et al. (1997) señalaron que los distintos mecanismos de respuesta a la
presencia de taninos condensados dependen de cada microorganismo ruminal, lo
que explicaría la distinta susceptibilidad entre cepas bacterianas. La unión de los
taninos con enzimas sean bacterianas o endógenas, no implica necesariamente una
disminución de su actividad (Makkar, Singh y Dowra., 1988).
Resulta más fácil que los taninos condensados inhiban la actividad de las
enzimas hemicelulolíticas que de las celolulíticas (Waghorn, 1996), debido a que
estas últimas se encuentran unidas a las bacterias y los complejos enzimáticos
hemicelulolíticos son extracelulares. Por ello en presencia de taninos condensados,

58
la mayoría de los investigadores han podido observar una menor digestibilidad de
la hemicelulosa que de la celulosa (Barrey y Manley, 1984).
- Acción directa sobre los microorganismos del rumen: Parece probable que los
taninos actuen sobre los microorganismos ruminales alterando la permeabilidad de
sus membranas (Leinmüller, Steingass y Menke., 1991; Scalbert, 1991).
El grado de tolerancia de los microorganismos del rumen es específico de cada
microorganismo en cuestión, explicando la diferencia en la susceptibilidad de las
cepas bacterianas, dependiendo del tipo tanino, y las diferencias entre taninos
hidrolizados y taninos condensados; aunque pocos microorganismos tolerantes del
rumen han sido descritos, es muy factible que su verdadera diversidad sea mayor
que lo actualmente conocido ((McSweeney, Palmer, Bunch y Krause., 2001).
2.8.3.3 Efectos sobre la Producción
El consumo de taninos puede afectar la ingestión voluntaria de alimento y su utilidad
digestiva por lo que se puede esperar consecuencias sobre la productividad de los
animales que las consumen. La disponibilidad de nutrientes se reduce debido a los
complejos formados entre los taninos y varios tipos de macromoléculas, Barry
(1985) observó una reducción significativa en la ganancia de peso vivo en corderos
alimentados con L. pedunculatus (con un alto contenido de taninos condensados
76-90 g/kg de materia seca. Sin embargo, existen autores que sugieren que la
ingestión continua de taninos, podría dar lugar a una adaptación parcial a los
mismos, con la desaparición o al menos la atenuación de sus efectos nocivos (Barry,
1985; Silanikove 2000).

59
Se ha demostrado que el consumo de plantas que contienen taninos
condensados (L. corniculatus) en cantidades moderadas afecta el crecimiento de
ovinos en desarrollo (Ramírez-Restrepo y Barry, 2005). Por otra parte, la ingestión
de niveles bajos de taninos influye también en los niveles de producción y la calidad
de la leche (Min et al., 2003; Molle, Decidía, Cabiddu y Sitzia., 2003; Rochfort,
Parker y Dunshea., 2008). El consumo de lotos o de sulla (Hedysarum coronarium)
se ha asociado a un incremento de la producción de leche en ovinos y bovinos (Min
et al., 2003).
Se ha demostrado un aumento de la tasa de proteínas en la leche en vacas de
un 10% y un 12% en ovejas que consumen taninos condensados, la tasa de lactosa
se incrementó a un 14% en ovejas al ser comparado con animales que no recibieron
esta suplementación con plantas ricas en taninos condensados (Min et al., 2003;
Rochfort et al., 2008). Finalmente, una ingestión moderada de taninos condensados
(de 2 a 4% del total de la ración) se asocia a un aumento de la producción de lana.
Por ejemplo, el consumo de L. corniculatus indujo a un aumento del 11% de la
producción de lana y hasta el14% en la tasa de ovulación de ovinos sin afectar el
consumo voluntario (Luque et al., 2000), que está asociado con un incremento de
la absorción en particular de cisteína, el cual es un aminoácido esencial para la
producción de la lana (Mc Nabb, Waghorn, Barry y Shelton., 1993).
2.8.3.4 Efectos sobre la reproducción
En relación al efecto de los taninos condensados sobre la eficiencia reproductiva de
los animales Min et al. (1999) observaron que en ovejas en pastoreo alimentadas
con Lotus corniculatus (17 g de taninos condensados/kg de materia seca)

60
aumentaba un 25% la producción de corderos lo cual se debía a un aumento, en la
tasa de ovulación y como consecuencia la prolificidad de los animales.
2.8.3.5 Efectos sobre la salud animal
- Prevención de la timpanización espumosa: La hinchazón se produce cuando en el
pastoreo los rumiantes consumen grandes cantidades de plantas leguminosas
como alfalfa o trébol, los gases producidos en el rumen durante la fermentación no
pueden ser lanzados en la forma normal ya que se encuentran atrapados en una
espuma persistente causada por la rápida liberación de proteínas solubles
durante la masticación y la degradación ruminal. Se ha comprobado que los
animales que consumen leguminosas que contienen altos niveles de taninos
condensados reducen notablemente la producción de gas mediante la precipitación
de la espuma de las proteínas de las plantas, previniendo así, la aparición de
timpanismo por la acción de los taninos condensados (Min et al., 2005). Un ejemplo
de esta situación se da en el caso de animales que pastan con Onobrychis viciifolia
que debido a su alto contenido de taninos condensados no produce timpanismo
(Barry y McNabb, 1999; McMahon et al., 2000). La sustitución de una pequeña
cantidad aproximadamente 10% de materia seca de alfalfa por Onobrychis viciifolia
ofrece indudables beneficios en la prevención de la hinchazón (McMahon et al.,
2000).
-Prevención de episodios diarreicos: El consumo de forrajes ricos en taninos
condensados se asocia con un aumento de la materia seca de las heces lo que
contribuye a evitar los episodios diarreicos y a la producción de lana limpia (Min et
al., 2001). Además, como consecuencia disminuyen los riesgos de miasis cutáneas

61
debido a la disminución de las suciedades de las zonas del periné como las diarreas
(Larsen et al., 1994).
2.8.3.6 Propiedades antiparasitarias
El papel antiparasitario de los taninos condensados ha sido confirmado por los
resultados de varios bioensayos in vitro que evaluaban los efectos de plantas o
específicamente extractos crudos de taninos condensados de diversos orígenes,
sobre los diferentes estadios de NGI (Athanasiadou et al., 2001; Barrau et al., 2005
citado en Hoste et al., 2006).
Investigaciones previas realizadas por Molan et al. (2000a, 2000b, citado en
Molan, 2014) mostraron por primera vez que los taninos condensados extraídos de
una amplia gama de forrajes tienen efectos inhibidores directos contra las larvas de
gusanos pulmonares en ovejas y ciervos in vitro, así como la capacidad de
desacelerar el proceso de eclosión y prevenir la eclosión de las larvas para alcanzar
el pleno desarrollo a larvas infectivas (Molan et al., 2002; Schreus et al., 2002; Molan
y Farag, 2010 citado en Molan, 2014).
Los taninos condensados afectan las poblaciones de gusanos adultos en
cabras, estos efectos en la biología del gusano dependerá del tiempo de
administración en relación con el ciclo parasitario por ejemplo: la administración de
extracto de taninos condensados de quebracho tiene un impacto en el
establecimiento temprano de las larvas del tercer estadio en cabras así como un
efecto persistente en la reducción de excreción de huevos que podría ser una
consecuencia de una disminución en la carga de gusanos o una reducción en la

62
fecundidad de gusanos hembras, en comparación con las ovejas, una disminución
de la carga parasitaria en el abomaso se ha observado con independencia del
momento de la administración del tanino (Paolini et al., 2003).
Aunque el mecanismo por el cual los taninos afectan los nematodos aún es
indefinido, es posible que los taninos formen complejos con las proteínas de la
superficie del parásito y causen disturbios en su metabolismo por lo cual la
respuesta metabólica de los nematodos a los taninos sería desviar los nutrientes
utilizados para la reproducción disminuyendo así la fecundidad (Alhag, Abdelrahim
y Verla, 2014).
En un estudio realizado por Martínez et al. (2013) en condiciones tanto in vitro
como in vivo usando microscopía electrónica de barrido (SEM) se evaluaron los
cambios estructurales de adultos de H. contortus obtenidos del abomaso de cabras
donantes infectadas, los cuales fueron mantenidos durante 24 horas en contacto
con plantas ricas en taninos una de clima tropical Tzalam (L.latisiliquum) y otra de
clima templado Esparceta (Onobrychis viciifolia) y comparado con los gusanos
mantenidos en PBS (control) para los ensayos in vitro. Para los estudios in vivo las
cabras fueron alimentadas con una dieta libre de taninos hasta el día 27 post
infección cuando la infección era latente, a partir del día 28 algunas cabras fueron
alimentadas con forrajes de plantas ricas en taninos por 7 días consecutivos. Por
microscopía se observaron cambios estructurales en la cutícula de los parásitos
correspondientes a pliegues longitudinales y transversales y crestas cuticulares más
gruesas, también agregados materiales alrededor de la cápsula bucal y/o en la vulva
y el ano, cambios en el sistema reproductivo de las hembras fueron encontrados in
vitro pero no in vivo; se cree que estas lesiones estructurales encontradas en los

63
gusanos podrían afectar su nutrición y motilidad con las posibles consecuencias en
su reproducción.
Ponè et al. (2011) evaluaron los efectos ovicidas de los extractos acetónicos de
tres plantas leguminosas forrajeras: Calliandra calotyrsus, Gliricidia sepium y
Leucaena diversifolia a concentraciones de 0.075, 0.15, 0.3, 0.6 y1.25 mg/ml, in
vitro encontrando efectos negativos sobre huevos y larvas de pequeños rumiantes.
A la mayor concentración los extractos inhibieron el desarrollo de L1 y L2, la actividad
larvicida observada fue diferente entre extractos, lo que se puede atribuir a los
compuestos fenólicos y en particular a los taninos; estas sustancias son capaces
de penetrar la cutícula del nematodo y prevenir la absorción de glucosa o bloquear
los receptores post sinápticos y en consecuencia paralizar la larva.
Von Son et al. (2012) investigó la actividad antihelmíntica in vitro de 5 plantas
leguminosas tropicales Arachis pintoi CIAT 22160 (Ap 22160), Gliricidia sepium,
Cratylia argentea (Ca yacapani), C. argentea CIAT 22386 (Ca 22386), C. argentea
veranera contra larvas infectivas L3 de H.contortus y el papel de los taninos en el
efecto antihelmíntico. Los resultados de este estudio mostraron que el extracto C.A.
22386 tuvo un efecto dependiente de la dosis en la inhibición de la migración larval,
esto indicaba que dosis crecientes del extracto produjo mayor inhibición de la
migración de larvas. A. pintoi 22160, C.a. yacapani, C.a. veranera y C.a. 22386
inhibieron el proceso de eliminación de la cutícula de las larvas y solo C.a. 22386,
C.a. Veranera y G. sepium disminuyeron su motilidad.
Varios estudios en ovejas han demostrado que el mejoramiento de la nutrición
proteica disminuye la infestación de parásitos. Esto se supone que es mediado por
un aumento de la inmunidad del huésped, que puede ser especialmente importante

64
con la selección de su inmunidad frente a parásitos gastrointestinales debido a que
los taninos condensados presentes en los forrajes tienen el potencial para ayudar
en su control (Min, Miller y Loetz, 2003).
2.8.4 Toxicidad de taninos condensados.
Las intoxicaciones severas de diversas especies de animales después de la
ingestión excesiva de plantas ricas en taninos hidrolizables ha sido atribuida, sobre
todo, a la presencia de gallotaninos (Min et al., 2003; Mueller-Harvey, 2006). En los
rumiantes, los taninos hidrolizables son hidrolizados por las bacterias ruminales
provocando la liberación del ácido gálico, que es rápidamente descarboxilado en
pirogalol y convertido inmediatamente en resorcinol (Min et al., 2003).
Estos metabolitos resultados de la descomposición del ácido gálico (resorcinol y
punicalagin) son hepato y nefro-tóxicos y son rápidamente absorbidos por la sangre
(Mueller-Harvey, 2006). Sin embargo, se ha demostrado que la toxicidad de los
taninos hidrolizables depende de su calidad (estructura química, peso metabólico)
y de la cantidad (dosis) ingerida (Zhu et al., 1992).
Los taninos condensados son raramente asociados con una toxicidad aguda en
los rumiantes (Butter et al., 1999 citado en Abdu et al., 2012). Un nivel de consumo
bajo o moderado de taninos condensados está asociado con efectos favorables,
mientras que la ingestión de elevados niveles resulta en aspectos negativos en
parámetros de salud y sobre la fisiología digestiva de los animales (Min et al., 2003).

65
2.9. Leucaena leucocephala.
La Leucaena es una leguminosa arbórea que puede alcanzar alturas comprendidas
entre los 2 y 20 mts, es muy resistente a la sequía ya que posee un sistema radical
profundo el cual se puede extender hasta 5 mts para obtener el agua subterránea.
Tiene la capacidad de desarrollarse en zonas con precipitaciones entre los 300 y
3000mm/año, no tolera suelos con mal drenaje interno, pudiendo crecer en suelos
de baja fertilidad natural pero con un buen drenaje y sin problemas de aluminio
intercambiable (Espinoza, 1996 citado en Espinoza, Torres y Chacón, 2007).
Es una planta que contiene proteína sobrepasante, razón por la cual las
respuestas animales son satisfactorias, los valores de taninos condensados (4.7%)
obtenidos por Espinoza et al. (2003) en el estado de Cojedes Venezuela, indican
que los materiales evaluados en este estudio pueden incrementar la cantidad de
aminoácidos disponibles en el tracto duodenal debido a la protección que ejercen
estos taninos a la proteína cruda por lo cual escapan de las bacterias del rumen
para ser utilizadas y sintetizadas en el tracto posterior del animal.
L. leucocephala es muy utilizada en Cuba debido a su alto contenido en
proteínas, minerales y vitaminas; trabajos recientes desarrollados en el Instituto de
Ciencia Animal destacan el contenido de polifenoles totales en un 5.84% y taninos
condensados totales entre el 1.8 y 4%; además de la presencia de saponinas, se
ha demostrado su potencialidad para reducir la metanogénesis en condiciones in
vitro (Galindo et al., 2009).

66
Agregando ventajas a su uso como forrajera se menciona el efecto en la
disminución de parásitos internos; se ha comprobado que leguminosas no sólo las
arbóreas, sino algunas forrajeras tradicionales presentan altos niveles de taninos
condensados, éstas sustancias aparecen con un efecto negativo sobre los
nematodos al limitar el establecimiento de los mismos en el aparato digestivo o
interfiriendo con su longevidad o reproducción. Pertenece a la familia Fabaceae,
Subfamilia mimoseae, Tribu mimoseae, Género Leucaena especie Leucocephala.
(Scribano, 2012).
2.10. Calliandra calothyrsus
Es un árbol de la agroforestería y sus hojas se utilizan para alimentación animal.
La madera es apta para leña y ha demostrado ser una alternativa útil a las especies
de Leucaena y eucalipto. Puede ser aceptada por los habitantes del bosque o de
las comunidades agrícolas. La especie pertenece a la familia de las leguminosas y
la subfamilia Mimosoideae (Powell, 1997 citado en Sebuliba et al., 2012).
La especie C. calothyrsus, conocida como “pelo de ángel”, es un árbol frondoso
originario de Guatemala, pero distribuido por todo el mundo por sus múltiples usos.
Esta planta tiene la facilidad de crecer en suelos empinados, secos, en tierras
afectadas por erosión o fuego, en zonas dominadas por malezas e incluso en tierras
ácidas (a pH 5.0); puede crecer en altitudes mayores a 1500 msnm, y la cantidad
óptima de lluvia para su desarrollo está entre 2000 mm y 4000 mm anuales, pero
se puede desarrollar en áreas de menor pluviosidad, con 2-4 meses de estación
seca. Desde 1936 esta especie se utiliza para la producción de pulpa de madera
debido a que posee un sólido sistema de defensa, compuesto por gran cantidad de

67
taninos inhibidores de crecimiento de larvas que hacen muy fuerte su tallo. C.
calothyrsus es una leguminosa arbustiva potencialmente valiosa como suplemento
forrajero en la producción del ganado bovino; sin embargo, el uso de algunas
accesiones se encuentra limitado por presentar diversos factores denominados
como antinutricionales y diferentes grados de digestibilidad, lo que permite sugerir
y estimular el estudio químico de estas accesiones (Lascano, 1995; Jackson, 1996
citados en Murillo et al., 2008).
Se infiere comúnmente que cualquier restricción nutricional es debido a su
contenido de taninos, aunque todavía no se han identificado sustancias tóxicas en
Calliandra, sí se han reportado altas concentraciones de taninos condensados
(hasta 11%). Existen informes contradictorios de si las altas concentraciones de
taninos condensados pueden ser responsables de la reducción de la palatabilidad
y digestibilidad en los animales, sin embargo, parece que la alimentación como un
suplemento en lugar de una alimentación única sería una forma de evitar estos
problemas (Palmer, Macqueen y Gutteridge, 1994).
2.11. Flemingia macrophylla.
Flemingia es un arbusto leñoso de origen asiático, con hojas trifoliadas. El hábitat
de esta planta ha sido estudiado en Indonesia, y se ha informado de que flemingia
es un arbusto resistente que puede resistir bien largos períodos de sequía o ligeras
inundaciones. Por otra parte, se ha afirmado que pueden tolerar suelos ácidos
infértiles ricos en aluminio soluble (Budelman, 1989; citado en Sankol y Ly, 2001).
Presenta poca susceptibilidad al ataque de enfermedades. Un dato importante
es que puede ser usada como un método de reducción de las poblaciones de

68
nematodos como Paratylenchus sp., Helicotylenchus sp. y Rotylenchus sp., pero no
de la especie Meloidogyne sp. (Yamoah et al., 1986 citado en Ayala y Pérez, 2006).
En algunas partes, es utilizada como forraje para la alimentación animal, no
obstante hay autores que mencionan el hecho de que el alto contenido de fibra y
taninos hace de la materia verde algo muy difícil de consumir para los animales. A
pesar de esto se ha reportado su uso como forraje, especialmente para especies
ovinas y caprinas (Ansah-Adjaye 1977 citado en Ayala y Pérez, 2006).
Fäsler (1993 citado en Rosales, 1999), midió el consumo, la digestibilidad y la
retención de nitrógeno en ovejas alimentadas con sólo pasto de baja calidad
(Brachiaria didtyoneura) en un 60%, suplementado con Cratylia argentea sola (40%)
o suministrado en una mezcla con Flemingia macrophylla a dos niveles; los
resultados demostraron que a medida que la proporción de F. macrophylla se
incrementaba en la mezcla, hubo una excreción más grande de nitrógeno fecal y
una reducción en la digestibilidad de la materia seca y la fibra, lo cual confirmó según
un estudio de Lascano y Palacios (1993) que el contenido de taninos en las plantas
se puede utilizar para reducir la magnitud de la degradación de la proteína soluble
en el rumen y de ésta forma incrementar la cantidad del flujo de nitrógeno no
amoniacal hacia el intestino delgado.
Estudios realizados por Cano, Carulla y Lascano (1994) describieron que en
plantas de trópico alto como F. macrophylla, el 79% de los taninos fueron extractos
solubles y sólo el 14%, estuvieron ligados a proteínas y el 7% a la fibra.

69
3. METODOLOGÍA
Este proyecto de investigación se aprobó por el Comité de Ética de la Universidad
de la Salle con fecha del 14 de Agosto del 2015 con el consecutivo 139.
3.1 Localización
La presente investigación se desarrolló en dos fases una de entrenamiento y otra
correspondiente a las pruebas finales en el laboratorio de helmintología veterinaria
de la Corporación Colombiana de investigación agropecuaria (CORPOICA) de
Tibaitatá en el kilómetro 14 vía Bogotá- Mosquera; a una altura de 2630 msnm con
una humedad relativa de 72% y temperatura entre 6 y 22 ºC.
3.2 Material orgánico
Se colectaron muestras de materia fecal de un ovino raza Hampshire de 8 meses
de edad con una condición corporal de 3, adquirido como parte del proyecto de
investigación, los muestreos se llevaron a cabo en predios de la Corporación
Colombiana de investigación agropecuaria (CORPOICA) de Tibaitatá en Mosquera
Cundinamarca, donde se determinó la carga parasitaria (huevos por gramo de
materia fecal) empleando el método de McMaster (Gordon y Whitlock, 1939). La
carga parasitaria del animal en el presente trabajo se mantuvo en un rango entre
2000 y 2500 Hpg, se verificaron también los cinco puntos de chequeo para valorar
el estado del paciente durante el experimento garantizando su bienestar.
Posteriormente se realizó la obtención de huevos de nematodos, así como de
larvas L3, efectuando la identificación morfométrica de las mismas en porcentaje

70
por género y especie de acuerdo con la metodología propuesta por Van Wyk, J.A.
y Mayhew, E. (2013).
3.2.1. Técnica de Recuperación de huevos
20 g de heces fueron tomadas desde la ampolla rectal del ovino empleado en el
estudio. Los huevos se colectaron mediante el uso de diferentes tamices de
acuerdo al método publicado por Bizimenyera et al. (2006) basado en el método
original reportado por Coles et al. (1992). Dichas muestras se suspendieron en agua
tibia (37°C) y luego fueron filtradas a través de mallas de 1 mm, 106 μm, 53 μm and
25 μm; los huevos retenidos en el último tamiz, fueron removidos y lavados con
agua destilada, el resultado de este lavado fue almacenado en tubos Falcon de 15
mL para ser centrifugados a 3000 rpm durante 5 minutos. Luego el sobrenadante
fue desechado, y se adicion sacarosa hasta completar nuevamente un volumen de
15 ml y se llevó nuevamente a centrifuga durante el mismo periódo de tiempo y rpm
anteriormente mencionadas. De manera cuidadosa, se extrajo el sobrenadante que
contiene la mayoría de huevos de nematodos gastrointestinales; este sobrenadante
se pasó una vez más por el tamiz de 25 μm, enseguida se realizó el lavado de este
tamiz, utilizando agua destilada y depositando el resultado de este lavado en tubos
Falcon de 50ml, estos tubos se dejaron decantando durante 15 minutos a 22OC;
finalmente se desechó el sobrenadante y se depositaron en un solo tubo, todos los
sedimentos. Previamente a la realización de los montajes se llevó a cabo el conteo
de los huevos, con el fin de garantizar que existían aproximadamente 100 huevos
en 100μl.

71
3.2.2. Técnica de Coprocultivo y Obtención de Larvas
Se recogieron las heces del ovino mediante un sistema de pañal o envase que
permitió mantener a las mismas dentro del recipiente para ser recolectadas sin
caerse al suelo, se mantuvieron en refrigeración, luego se colocaron en una bandeja
plástica mezcladas con una solución de agua destilada y corriente en proporción
50:50 para ser ablandadas hasta que su consistencia fue la de una papilla; una vez
en este punto se adicionó un poco de aserrin para formar bolitas que se depositaron
en frascos de vidrio de boca ancha , los cuales se taparon con papel al que se le
hicieron unos huecos pequeños para la entrada de aire y se sellaron con un caucho.
Estos montajes se incubaron a 27o C durante 7 días según la metodología propuesta
por el Laboratorio Central Veterinario de Weybridge (1986) y Roeber, Jex y Gasser
(2013), durante los cuales día de por medio se sacaron y destaparon para aireación
y verificación de humedad hecho fundamental, principalmente para el proceso de
identificacion morfométrica, pues de acuerdo a lo discutido por Van Wyk et al. (2004)
y Rossanigo y Gruner (1996) puede variar tanto la longitud total de la larva asi como
la longitud de la cola de la vaina en relacion con la humedad del coprocultivo. A los
8 días se retiró la cubierta y se les adicionó una solución 50:50 de agua destilada y
corriente, se voltearon y se dejaron 3 horas para que las larvas migraran hacia abajo
a una zona de menor densidad y luz; luego se tomaron muestras de cada frasco
con pipeta y se depositó una gota en la placa para observar al microscopio si había
suficientes larvas, éste proceso se repitió varias veces hasta conseguir la mayor
cantidad de larvas, las cuales fueron refrigeradas para su posterior concentración.

72
3.2.3. Lavado de Larvas
En una vénula de 20 ml se adicionó 15 ml de solución de sacarosa fría al 40 % a
la que se agregó una solución de 5 ml de larvas L3 conservando una relación de
7.5: 2.5, dejándola caer despacio por las paredes del tubo. El mismo procedimiento
se realizó con 3 vénulas más. Después el total de vénulas (4) se pesaron en una
báscula de manera que quedaran calibradas para poderlas colocar dentro de la
centrífuga de refrigeración a 3500 rpm por 10 minutos a una temperatura de 4°C.
Una vez cumplido el tiempo se procedió a sacarlas revisando la formación de
un anillo blanco que es el sitio donde están concentradas las larvas, con una pipeta
se extrajeron las larvas de éste anillo, teniendo la precaución de no sacar residuos
de sacarosa y detritus para ser depositadas en tubos de ensayo en un volumen de
2 ml, se adicionó agua destilada hasta un volumen de 8 ml para centrifugarlos,
luego se sacó el sobrenadante y se procedió a realizar 2 lavados más de manera
que se eliminaran los residuos de sacarosa quedando las larvas L3 lavadas, siendo
almacenadas y conservadas en una caja de manera horizontal.
3.2.4. Conteo de Larvas
Con una pipeta de 50 μl se extrajo de un tubo de ensayo con un volumen específico
de larvas una muestra para ser depositada en forma de 10 gotas en una lámina y
se procedió a contar cuántas larvas había en cada gota para determinar el número
total en la misma, este procedimiento se realizó de igual forma en otras 4 láminas.

73
Al final se sacó el número promedio de larvas de las 5 láminas y se relacionó con
el volumen inicial para obtener el número total de larvas. Este procedimiento se
realizó previo a cada montaje para garantizar que se tuviera el número suficiente de
larvas en la ejecución de cada prueba.
3.2.5 Determinación de la concentración adecuada de Hipoclorito de Sodio para la
prueba de eliminación de la cutícula
El hipoclorito de sodio hace la acción del ácido clorhídrico llegando al estómago de
la larva para soltar la cutícula. La determinación de la solución más adecuada del
mismo que permitió una mejor eliminación de la cutícula se realizó de la siguiente
manera: De una solución de hipoclorito al 15% se prepararon 2 diluciones al 7.5%
y 3 más al 3.75% en tubos falcon de 15ml, luego en 5 tubos de ensayo, se adicionó
un volumen 15,625 μl de agua destilada en cada uno, después en uno de los tubos
se colocó un volumen de 125 μl y en otro 150 μl de la dilución al 7.5%, los mismos
volúmenes fueron adicionados en los otros 2 tubos y en el 3 tubo se adicionó un
volumen de 137.5 μl pero con la dilución al 3.75% . En una placa de 24 pozos
previamente limpiada con alcohol, se agregó en cada pozo 6 por fila un volumen
total 100 L3 más 1400 μl de cada dilución hasta completar la placa. Luego se inició
el cronometraje de 10 minutos para adicionar lugol en cada uno de los pozos
pertenecientes a las dos primeras concentraciones, después de 10 minutos se
colocó en otros 2 pozos de la misma manera hasta llenar toda la placa para un total
de 60 minutos. Una vez terminado el proceso se dio inició a la lectura de cada pozo,
cuantificando la cantidad de L3 con y sin cutícula. De acuerdo a los resultados
obtenidos se determinó que la dilución de Hipoclorito de sodio más adecuada para

74
la eliminación de la cutícula fue la de 3.75% con 150 μl que mostró una eliminación
gradual de la cutícula desde los 10 a los 60 minutos en los 6 pozos.
3.2.6. Calibración del Ocular para la identificación morfométrica de las larvas L3
La calibración del microscopio se realizó utilizando un micrómetro de platina el cual
consta de un disco de vidrio que contiene una línea de 1 mm de longitud con una
escala calibrada de 0 a 100 micras divisiones que equivalen a 100 micras y cada
división a 10 micras y un micrómetro ocular el cual posee una reglilla de 100
divisiones pero sin medida de precisión. que se determinó de la siguiente forma: Se
insertó el micrómetro ocular en el ocular del microscopio, luego se colocó el
micrómetro de platina como se ubica una lámina portaobjeto en el microscopio
enfocando con el objetivo de 10 X la escala del micrómetro de platina, alineando al
cero del micrómetro de platina con el cero del micrómetro ocular que se superpuso
con una línea del micrómetro de la platina, determinando en el micrómetro ocular el
número de divisiones que se encontraron entre el punto cero y la línea de
superposición al igual que en el micrómetro de platina; cada división tuvo un valor
diferente según el ocular con que se observó, es decir para el objetivo de 40X fue
2.5 micras y para el objetivo de 10X fue 10 micras.
3.3 Extractos de plantas
El proceso de extracción de taninos condensados liofilizados de las plantas
Leucaena leucocephala, Calliandra calothyrsus y Flemingia macrophylla fue
realizado en el laboratorio de química de la Universidad del Tolima en la ciudad de
Ibagué.

75
3.4 Identificación morfométrica de larvas en estadio infectante (L3) de nematodos
gastrointestinales de ovinos
Se extrajo con la pipeta una muestra de solución con larvas y se depositó en una
lámina, luego se procedió a exponer la lámina con las larvas al calor por breves
segundos momento en el cual se retiró y se adicionó una gota de lugol a la lámina,
colocando un cubreobjetos; la coloración de contraste que las larvas adoptaron
facilitó la observación de los distintos detalles morfológicos y permitió diferenciar a
las larvas infectantes de las de vida libre , el mismo procedimiento se repitió varias
veces hasta completar 100 larvas L3. El análisis morfométrico se realizó basados
en parámetros como longitud total de la larva, ancho de la larva, longitud de la vaina
de la cola y extensión y longitud total del cuerpo (Van Wyk et al., 2004),
determinando de forma porcentual la participación de cada género y especie de
nematodos gastrointestinales.
3.5 Pruebas in vitro
3.5.1 Prueba de inhibición de la eclosión
De acuerdo a lo planteado por De Souza (2012). Los huevos fueron seleccionados
al azar para organizar los niveles de tratamiento; donde en cada nivel existian 100
huevos suspendidos en 100 μL; 6 niveles de tratamiento tuvieron 250 μL de cada
una de las diferentes concentraciones de taninos condensados de las plantas
empleadas en el estudio (1000, 500, 250, 125, 62.5 y 31.25 μg/ml), un control

76
positivo compuesto por Fenbendazol (25 mcg/mL) y un control negativo integrado
por tween al 0.4%; cada nivel de tratamiento fue completado hasta un volumen de
2000 μL. Posteriormente, cada uno de los montajes fue llevado a incubadora a
27°C durante 4 horas; terminado este proceso se añadió una gota de solución de
Lugol para frenar el proceso de eclosión y las larvas (L1) y huevos se contaron
utilizando un microscopio invertido de acuerdo a los lineamientos propuestos por la
WAAVP (World Association for the Advancement Veterinary Parasitology) descritos
por (Powers et al., 1982). A través de la siguiente formula se obtuvo el porcentaje
de inhibición de la eclosión (IE) para cada nivel de tratamiento:
IE (%) = 100 (P test / P total), en donde, “P test” es el número de huevos y “P total”
es el número de huevos + larvas L1.
3.5.2 Prueba de eliminación de la cutícula (Exsheathment)
Esta prueba se llevó a cabo según lo propuesto por De Souza (2012). Se pesó la
cantidad total del extracto de taninos condensados purificados de cada planta
incluida en el estudio, en una báscula electrónica y luego se tomó una muestra de
10 μl para hacer las diluciones (1000 μl/ml, 500 μl/ml, 250 μl/ml 125 μl/ml, 62.5
μl/ml, 31,25 μl/ml y control). Se realizaron todos los cálculos pertinentes y se
adicionó a cada uno de los tubos de ensayo la solución de tween, la concentración
de taninos condensados,100 L3 y el volumen restante con agua destilada auto
clavada. Estos montajes se colocaron en una incubadora a 22°C por 3 horas; una
vez cumplido el tiempo se llevaron a una centrífuga a 6000 rpm por 2 minutos, se

77
extrajo el sobrenadante, se sacó un volumen de 8 ml y se dejaron 2ml, se hicieron
2 lavados.
Para la ejecución de los montajes, fueron utilizadas unas placas de 24 pozos en
las cuales se hicieron 12 observaciones repetidas por concentración, preparadas
de la siguiente manera: Se adicionaron 1400 μl de Hipoclorito de sodio de la
concentración que mejor resultado arrojo, que en este caso para la investigación fue
la de 3.75% con 150 μl ya que permitió la liberación gradual de la cutícula, luego se
colocó un volumen de 100 μl de la solución de cada concentración conteniendo
aproximadamente 100 L3; tan pronto se terminó de llenar el último pozo, se contaron
10 minutos después de los cuales se inició la adición del lugol en cada hilera de
forma vertical, cada 10 minutos hasta terminar la placa. Una vez culminado el
proceso, se realizó la cuantificación de las larvas con y sin cutícula.
3.6 Diseño estadístico
A los resultados de las réplicas de las pruebas in vitro (% de inhibición de la
eclosión y % de inhibición de eliminación de la cutícula) se les realizó una regresión
Logyt Probit, con un rango de confiabilidad del 95% (p<0,05).
Para la comparación de efectividad entre los extractos de taninos condensados
de las diferentes plantas por concentración a utilizar en la investigación se efectuó
un ANOVA (DCA) y se realizó una prueba de medias de Tukey-HSD para establecer
el mejor tratamiento para cada prueba. El paquete estadístico utilizado para los
análisis de los datos fue el SPSS 20.

78
4. RESULTADOS
4.1 Identificación morfométrica de larvas en estadio infectante (L3) de nematodos
gastrointestinales de ovinos.
El objetivo principal de la identificación morfométrica fué determinar el porcentaje
de participación de cada especie del gran grupo de nematodos gastrointestinales
de los pequeños rumiantes en nuestro estudio, en donde el porcentaje que fue más
representativo de un 58% correspondió al género Haemonchus especie contortus
seguido de un 26% del género y especie Trichostrongylus falculatus; en contraste
el género que menor porcentaje registró del 1% fue el de Cooperia especie curticei
(Tabla 3). Aunque en el análisis morfométrico se tuvieron en cuenta los parámetros
como longitud total de la larva, ancho, longitud de la vaina de la cola y extensión y
longitud total del cuerpo de la larva (Van Wyk et al., 2004). Se determinó realizar la
clasificación en la presente investigación de acuerdo a la medida de la extensión de
la cola de la vaina (Anexo 7) propuesto por Van Wyk, J.A. y Mayhew, E. (2013).
Figura 6. Larvas (L3) Oesophagostomum columbianum, Trichostrongylus falculatus y Haemonchus contortus.
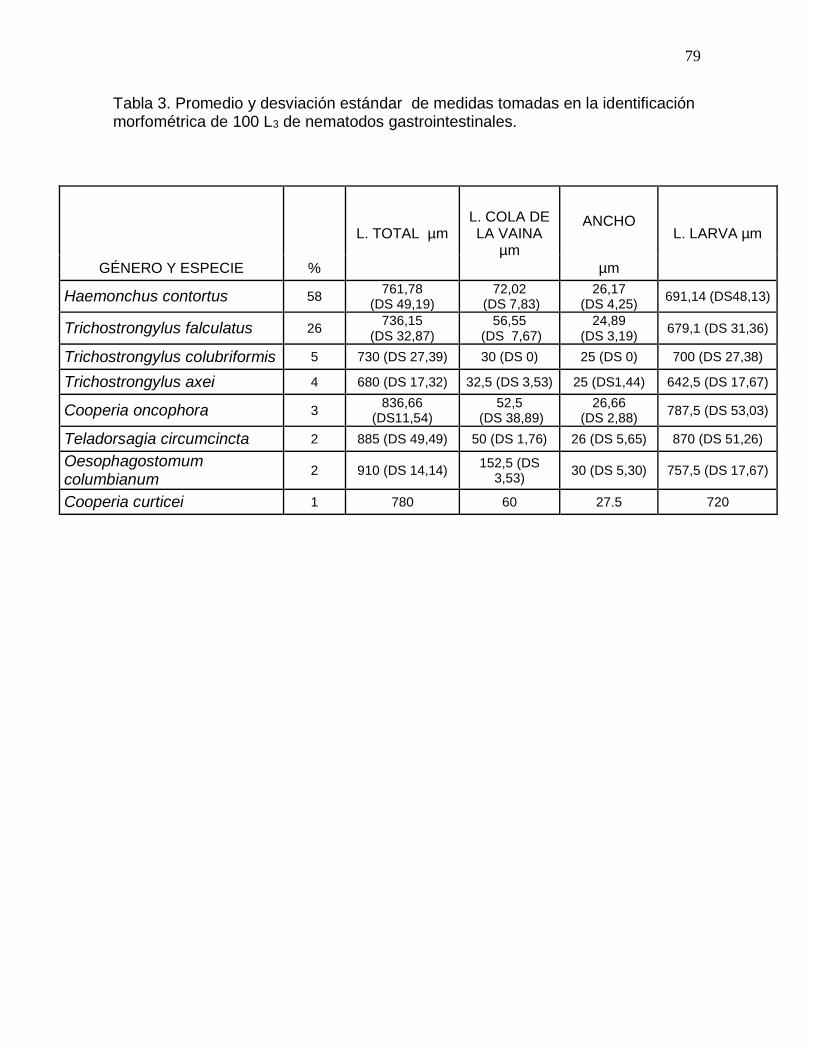
79
Tabla 3. Promedio y desviación estándar de medidas tomadas en la identificación morfométrica de 100 L3 de nematodos gastrointestinales.
L. TOTAL µm
L. COLA DE LA VAINA
µm
ANCHO L. LARVA µm
GÉNERO Y ESPECIE % µm
Haemonchus contortus 58 761,78
(DS 49,19) 72,02
(DS 7,83) 26,17
(DS 4,25) 691,14 (DS48,13)
Trichostrongylus falculatus 26 736,15
(DS 32,87) 56,55
(DS 7,67) 24,89
(DS 3,19) 679,1 (DS 31,36)
Trichostrongylus colubriformis 5 730 (DS 27,39) 30 (DS 0) 25 (DS 0) 700 (DS 27,38)
Trichostrongylus axei 4 680 (DS 17,32) 32,5 (DS 3,53) 25 (DS1,44) 642,5 (DS 17,67)
Cooperia oncophora 3 836,66
(DS11,54) 52,5
(DS 38,89) 26,66
(DS 2,88) 787,5 (DS 53,03)
Teladorsagia circumcincta 2 885 (DS 49,49) 50 (DS 1,76) 26 (DS 5,65) 870 (DS 51,26)
Oesophagostomum columbianum
2 910 (DS 14,14) 152,5 (DS
3,53) 30 (DS 5,30) 757,5 (DS 17,67)
Cooperia curticei 1 780 60 27.5 720
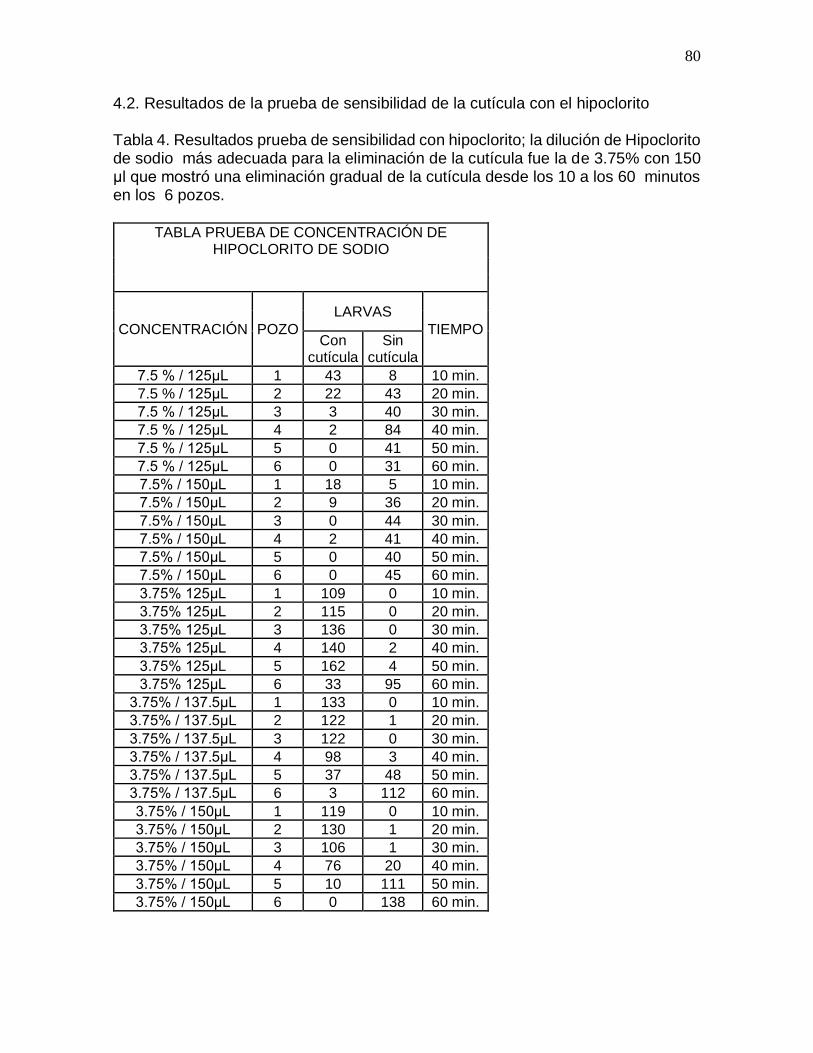
80
4.2. Resultados de la prueba de sensibilidad de la cutícula con el hipoclorito
Tabla 4. Resultados prueba de sensibilidad con hipoclorito; la dilución de Hipoclorito de sodio más adecuada para la eliminación de la cutícula fue la de 3.75% con 150 μl que mostró una eliminación gradual de la cutícula desde los 10 a los 60 minutos en los 6 pozos.
TABLA PRUEBA DE CONCENTRACIÓN DE HIPOCLORITO DE SODIO
CONCENTRACIÓN POZO
LARVAS
TIEMPO Con
cutícula Sin
cutícula
7.5 % / 125μL 1 43 8 10 min.
7.5 % / 125μL 2 22 43 20 min.
7.5 % / 125μL 3 3 40 30 min.
7.5 % / 125μL 4 2 84 40 min.
7.5 % / 125μL 5 0 41 50 min.
7.5 % / 125μL 6 0 31 60 min.
7.5% / 150μL 1 18 5 10 min.
7.5% / 150μL 2 9 36 20 min.
7.5% / 150μL 3 0 44 30 min.
7.5% / 150μL 4 2 41 40 min.
7.5% / 150μL 5 0 40 50 min.
7.5% / 150μL 6 0 45 60 min.
3.75% 125μL 1 109 0 10 min.
3.75% 125μL 2 115 0 20 min.
3.75% 125μL 3 136 0 30 min.
3.75% 125μL 4 140 2 40 min.
3.75% 125μL 5 162 4 50 min.
3.75% 125μL 6 33 95 60 min.
3.75% / 137.5μL 1 133 0 10 min.
3.75% / 137.5μL 2 122 1 20 min.
3.75% / 137.5μL 3 122 0 30 min.
3.75% / 137.5μL 4 98 3 40 min.
3.75% / 137.5μL 5 37 48 50 min.
3.75% / 137.5μL 6 3 112 60 min.
3.75% / 150μL 1 119 0 10 min.
3.75% / 150μL 2 130 1 20 min.
3.75% / 150μL 3 106 1 30 min.
3.75% / 150μL 4 76 20 40 min.
3.75% / 150μL 5 10 111 50 min.
3.75% / 150μL 6 0 138 60 min.

81
4.3. Prueba de eliminación de la cutícula
Los resultados obtenidos por el análisis Logyt Probit se realizaron con base a los
valores promedios que se dieron en cada prueba de acuerdo al número de
observaciones repetidas seleccionadas (Tabla 5). La dosis efecto 50 que mejor
eficacia mostró en los resultados de la inhibición de la eliminación de la cutícula fue
la del extracto de taninos condensados de Calliandra calothyrsus 19,15 μg/ml (Tabla
7) con respecto a los extractos de Leucaena leucocephala (Tabla 6) y Flemingia
macrophylla (Tabla 8). que presentó la dosis efecto 99 más eficaz 326 μg/ml.
El análisis Logyt Probit también mostró que la pendiente de la regresión fue
positiva en todos los casos por lo que el incremento en la concentración del extracto
de taninos condensados incrementó la inhibición del desenvaine larval.
Los controles negativos no presentaron un efecto significativo en la inhibición;
sin embargo todos los extractos inhibieron la eliminación de la cutícula, la cual se
inició desde la menor concentración (31,25 μg/ml) y el efecto fue dosis dependiente;
los porcentajes de la inhibición con la concentración más alta (1000 μg/ml), fueron
respectivamente del 99%, 98% y 92% sin que se presenten diferencias significativas
(p>0,05) en todas las concentraciones entre los extractos de las plantas Calliandra,
Leucaena y Flemingia.
Con el extracto de C. calothyrsus se observaron inhibiciones altas que se
presentaron desde la concentración más baja (31,25 μg/ml), donde mostró un % de
inhibición del desenvaine larval del 88,46% al minuto 60, en contraste los % de
inhibición para los extractos de L. leucocephala (77,45%) y F. macrophylla (74,74%)
fueron menores, no obstante, casi el 100% de la inhibición en el mismo extracto fue
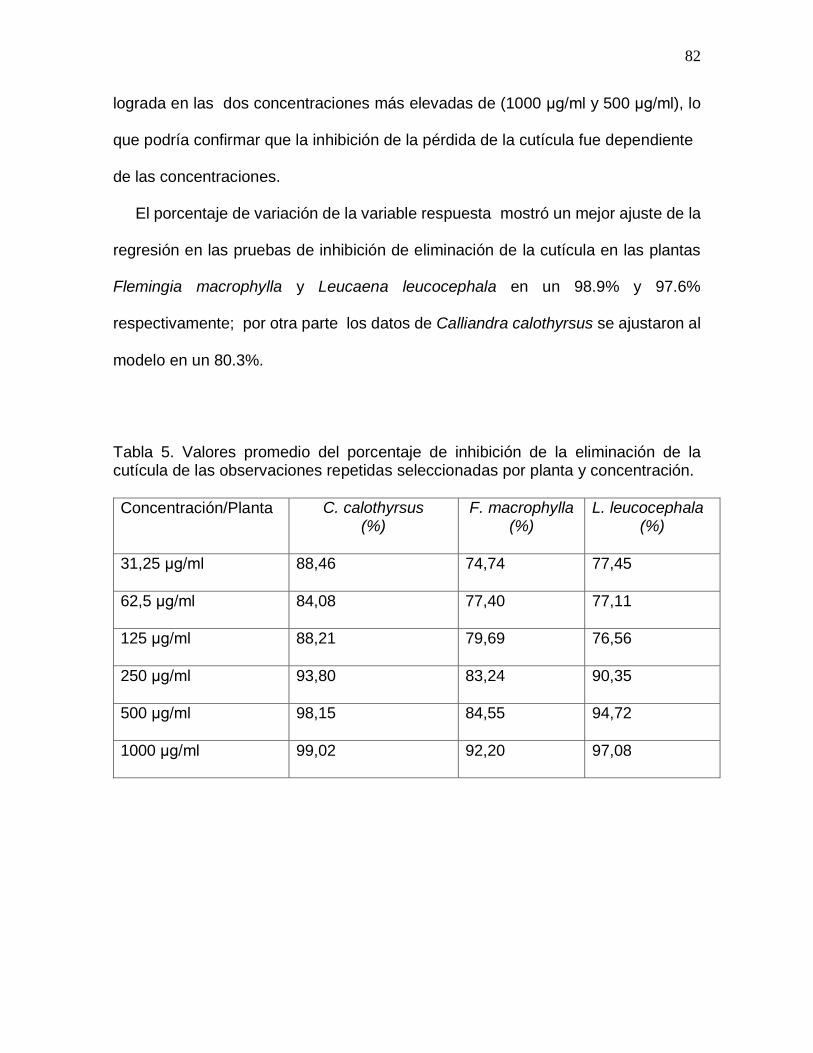
82
lograda en las dos concentraciones más elevadas de (1000 μg/ml y 500 μg/ml), lo
que podría confirmar que la inhibición de la pérdida de la cutícula fue dependiente
de las concentraciones.
El porcentaje de variación de la variable respuesta mostró un mejor ajuste de la
regresión en las pruebas de inhibición de eliminación de la cutícula en las plantas
Flemingia macrophylla y Leucaena leucocephala en un 98.9% y 97.6%
respectivamente; por otra parte los datos de Calliandra calothyrsus se ajustaron al
modelo en un 80.3%.
Tabla 5. Valores promedio del porcentaje de inhibición de la eliminación de la cutícula de las observaciones repetidas seleccionadas por planta y concentración.
Concentración/Planta C. calothyrsus (%)
F. macrophylla (%)
L. leucocephala (%)
31,25 μg/ml 88,46 74,74 77,45
62,5 μg/ml 84,08 77,40 77,11
125 μg/ml 88,21 79,69 76,56
250 μg/ml 93,80 83,24 90,35
500 μg/ml 98,15 84,55 94,72
1000 μg/ml 99,02 92,20 97,08
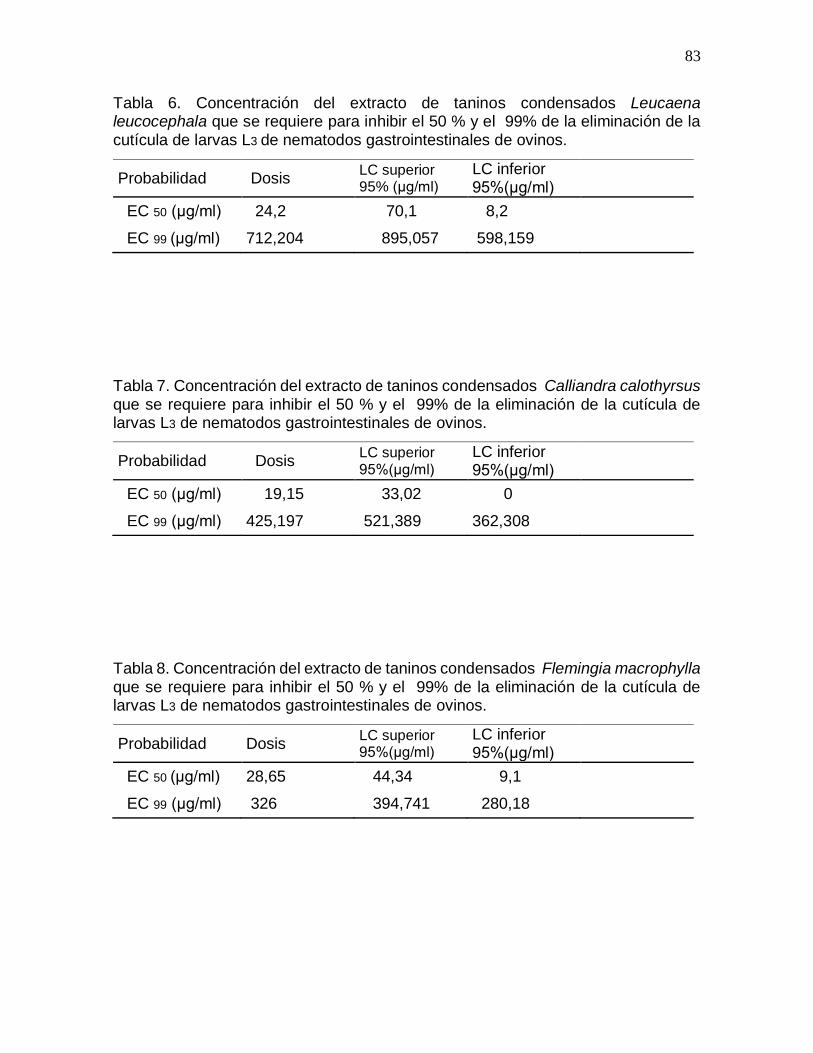
83
Tabla 6. Concentración del extracto de taninos condensados Leucaena leucocephala que se requiere para inhibir el 50 % y el 99% de la eliminación de la cutícula de larvas L3 de nematodos gastrointestinales de ovinos.
Probabilidad Dosis LC superior 95% (μg/ml)
LC inferior 95%(μg/ml)
EC 50 (μg/ml) 24,2 70,1 8,2
EC 99 (μg/ml) 712,204 895,057 598,159
Tabla 7. Concentración del extracto de taninos condensados Calliandra calothyrsus que se requiere para inhibir el 50 % y el 99% de la eliminación de la cutícula de larvas L3 de nematodos gastrointestinales de ovinos.
Probabilidad Dosis LC superior 95%(μg/ml)
LC inferior 95%(μg/ml)
EC 50 (μg/ml) 19,15 33,02 0
EC 99 (μg/ml) 425,197 521,389 362,308
Tabla 8. Concentración del extracto de taninos condensados Flemingia macrophylla que se requiere para inhibir el 50 % y el 99% de la eliminación de la cutícula de larvas L3 de nematodos gastrointestinales de ovinos.
Probabilidad Dosis LC superior 95%(μg/ml)
LC inferior 95%(μg/ml)
EC 50 (μg/ml) 28,65 44,34 9,1
EC 99 (μg/ml) 326 394,741 280,18
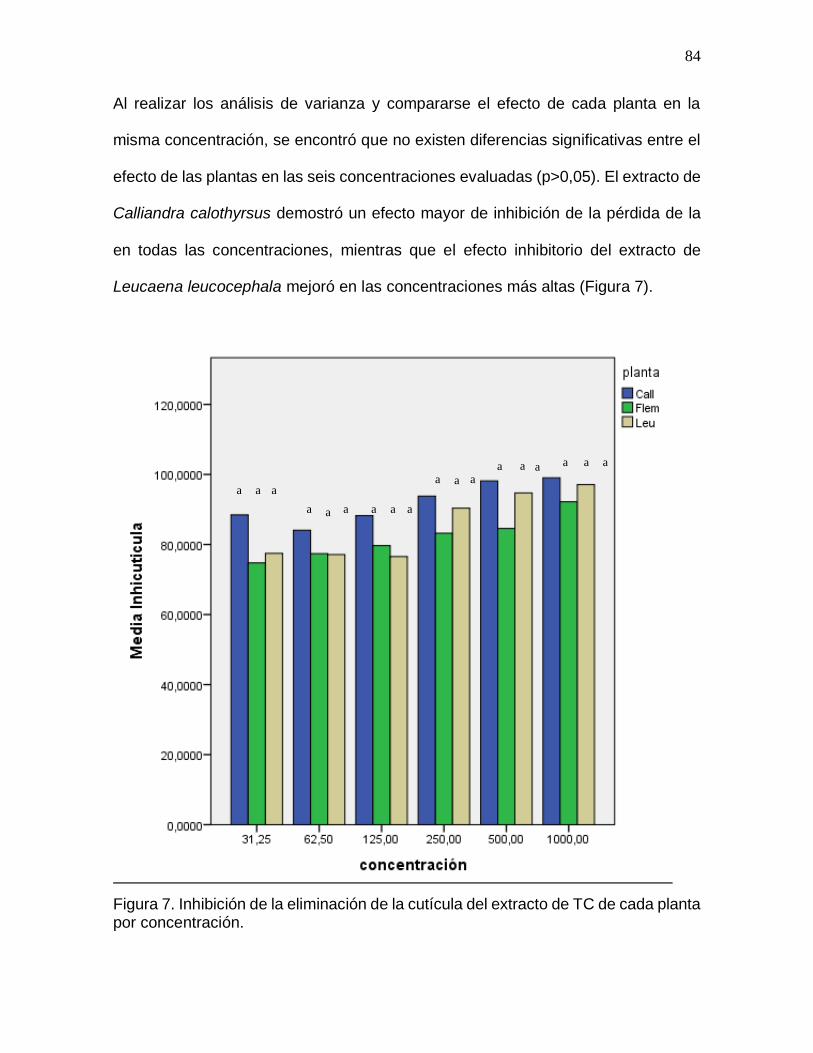
84
Al realizar los análisis de varianza y compararse el efecto de cada planta en la
misma concentración, se encontró que no existen diferencias significativas entre el
efecto de las plantas en las seis concentraciones evaluadas (p>0,05). El extracto de
Calliandra calothyrsus demostró un efecto mayor de inhibición de la pérdida de la
en todas las concentraciones, mientras que el efecto inhibitorio del extracto de
Leucaena leucocephala mejoró en las concentraciones más altas (Figura 7).
Figura 7. Inhibición de la eliminación de la cutícula del extracto de TC de cada planta por concentración.
a a
a a a a a
a
a
a a a a a a a a
a
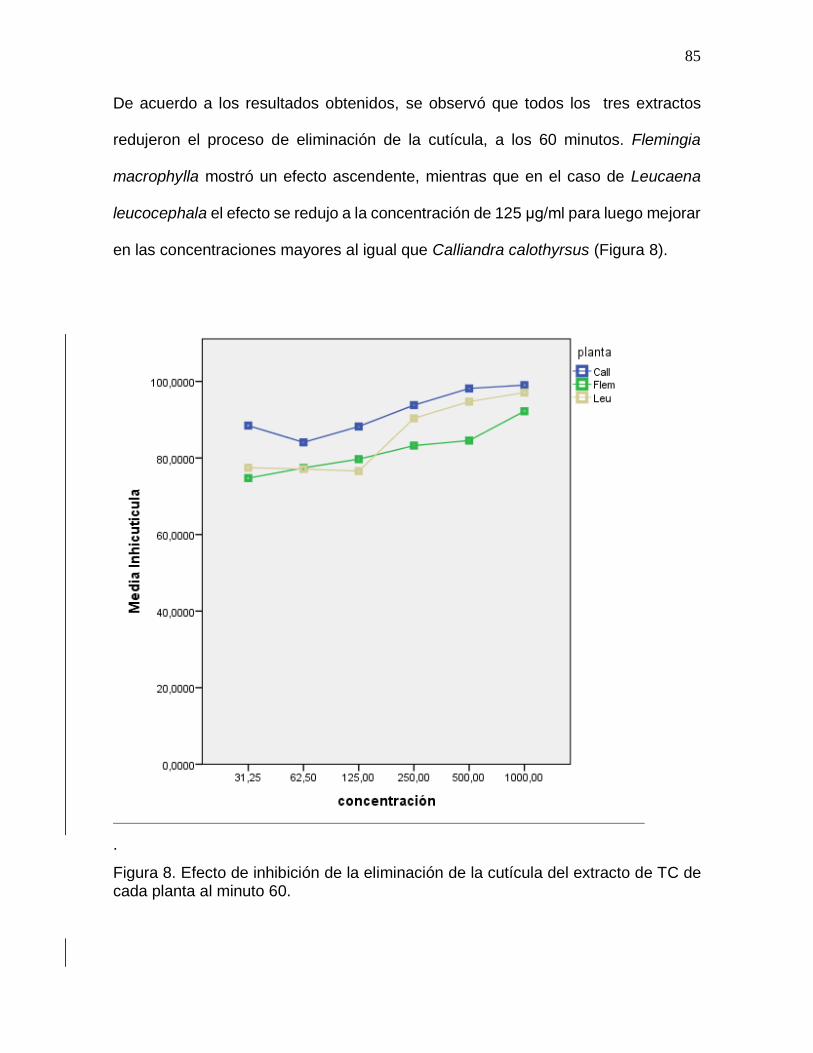
85
De acuerdo a los resultados obtenidos, se observó que todos los tres extractos
redujeron el proceso de eliminación de la cutícula, a los 60 minutos. Flemingia
macrophylla mostró un efecto ascendente, mientras que en el caso de Leucaena
leucocephala el efecto se redujo a la concentración de 125 μg/ml para luego mejorar
en las concentraciones mayores al igual que Calliandra calothyrsus (Figura 8).
.
Figura 8. Efecto de inhibición de la eliminación de la cutícula del extracto de TC de cada planta al minuto 60.

86
Figura 9. Cambios físicos y alteraciones de la cutícula; (A) L3 perdiendo la cutícula; (B) (c) Diferenciación entre L3 con y L3 sin cutícula; (D) Adhesión de taninos condensados a la cutícula de L3.
4.4. Prueba de inhibición de la eclosión
En el presente estudio los taninos condensados de la planta Calliandra calothyrsus
fueron los que presentaron mayor eficacia en la inhibición de la eclosión de huevos
de nematodos gastrointestinales, tanto en la dosis efectiva 50 como la 99, siendo
estas 1075,25 μg/ml y 3601,23 μg/ml respectivamente (Tabla 10).
Referente a los resultados obtenidos de los controles positivos y negativos, no
se encontró efecto significativo en la inhibición de la eclosión de huevos de NGI.
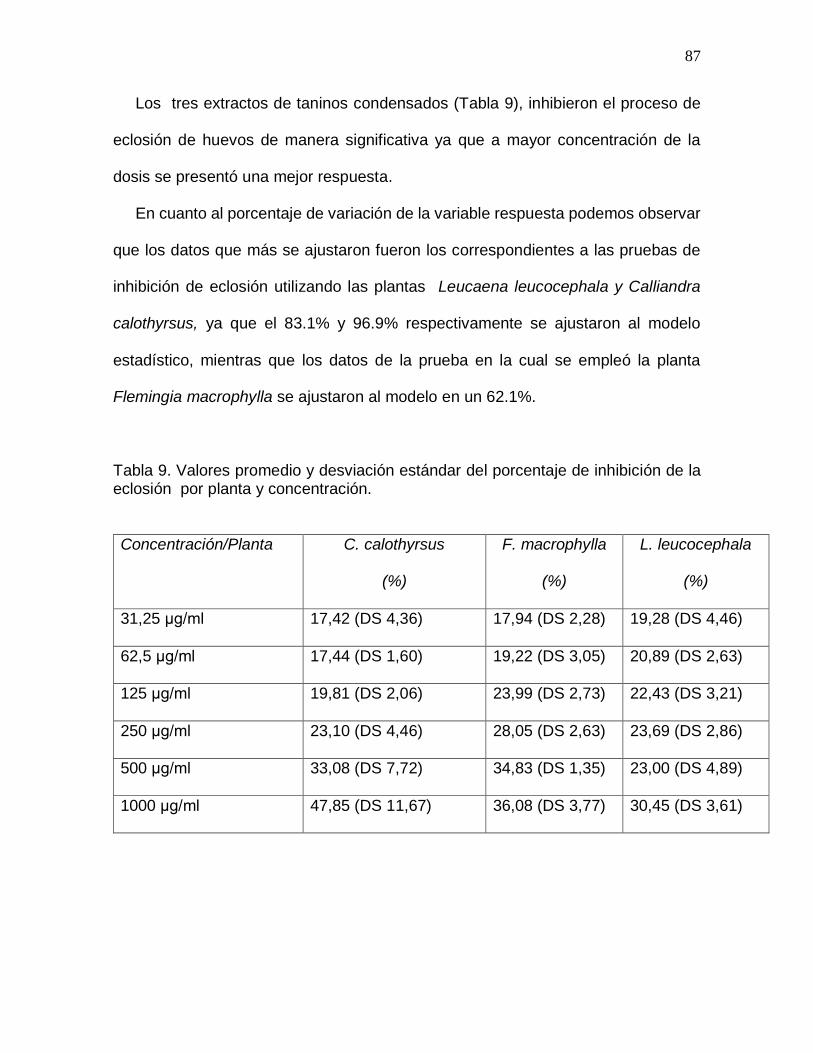
87
Los tres extractos de taninos condensados (Tabla 9), inhibieron el proceso de
eclosión de huevos de manera significativa ya que a mayor concentración de la
dosis se presentó una mejor respuesta.
En cuanto al porcentaje de variación de la variable respuesta podemos observar
que los datos que más se ajustaron fueron los correspondientes a las pruebas de
inhibición de eclosión utilizando las plantas Leucaena leucocephala y Calliandra
calothyrsus, ya que el 83.1% y 96.9% respectivamente se ajustaron al modelo
estadístico, mientras que los datos de la prueba en la cual se empleó la planta
Flemingia macrophylla se ajustaron al modelo en un 62.1%.
Tabla 9. Valores promedio y desviación estándar del porcentaje de inhibición de la eclosión por planta y concentración.
Concentración/Planta C. calothyrsus
(%)
F. macrophylla
(%)
L. leucocephala
(%)
31,25 μg/ml 17,42 (DS 4,36) 17,94 (DS 2,28) 19,28 (DS 4,46)
62,5 μg/ml 17,44 (DS 1,60) 19,22 (DS 3,05) 20,89 (DS 2,63)
125 μg/ml 19,81 (DS 2,06) 23,99 (DS 2,73) 22,43 (DS 3,21)
250 μg/ml 23,10 (DS 4,46) 28,05 (DS 2,63) 23,69 (DS 2,86)
500 μg/ml 33,08 (DS 7,72) 34,83 (DS 1,35) 23,00 (DS 4,89)
1000 μg/ml 47,85 (DS 11,67) 36,08 (DS 3,77) 30,45 (DS 3,61)

88
Tabla 10. Concentración del extracto de taninos condensadosCalliandra calothyrsus que se requiere para inhibir el 50 % y el 99% de la eclosión de huevos de nematodos gastrointestinales de ovinos.
Probabilidad Dosis LC superior 95%(μg/ml)
LC inferior 95%(μg/ml)
EC 50 (μg/ml) 1075,25 1675,58 810,063
EC 99 (μg/ml) 3601,23 10279,3 2619,83
Tabla 11. Concentración del extracto de taninos condensados Leucaena leucocephala que se requiere para inhibir el 50 % y el 99% de la eclosión de huevos de nematodos gastrointestinales de ovinos.
Probabilidad Dosis LC superior 95% (μg/ml)
LC inferior 95%(μg/ml)
EC 50 (μg/ml) 2309,7 6916,80 1472,5
EC 99 (μg/ml) 8411,9 27141,7 5083,0
Tabla 12. Concentración del extracto de taninos condensados Flemingia macrophylla que se requiere para inhibir el 50 % y el 99% de la eclosión de huevos de nematodos gastrointestinales de ovinos.
Probabilidad Dosis LC superior 95%(μg/ml)
LC inferior 95%(μg/ml)
EC 50 (μg/ml) 1458,5 1062,7 2561,0
EC 99 (μg/ml) 5494,3 10447,4 3793,4
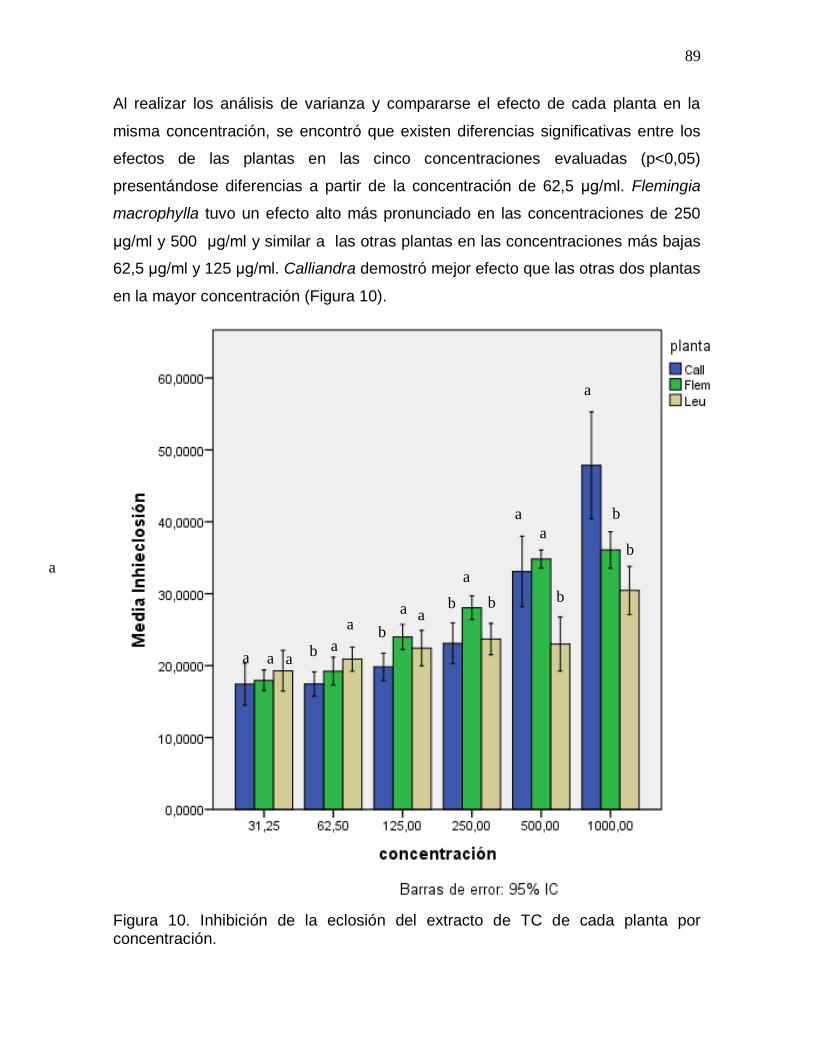
89
Al realizar los análisis de varianza y compararse el efecto de cada planta en la
misma concentración, se encontró que existen diferencias significativas entre los
efectos de las plantas en las cinco concentraciones evaluadas (p<0,05)
presentándose diferencias a partir de la concentración de 62,5 μg/ml. Flemingia
macrophylla tuvo un efecto alto más pronunciado en las concentraciones de 250
μg/ml y 500 μg/ml y similar a las otras plantas en las concentraciones más bajas
62,5 μg/ml y 125 μg/ml. Calliandra demostró mejor efecto que las otras dos plantas
en la mayor concentración (Figura 10).
Figura 10. Inhibición de la eclosión del extracto de TC de cada planta por concentración.
a
b
b
a
b
b
a
a a
b
a
a
b
a ab
a
b a
b
a
a a
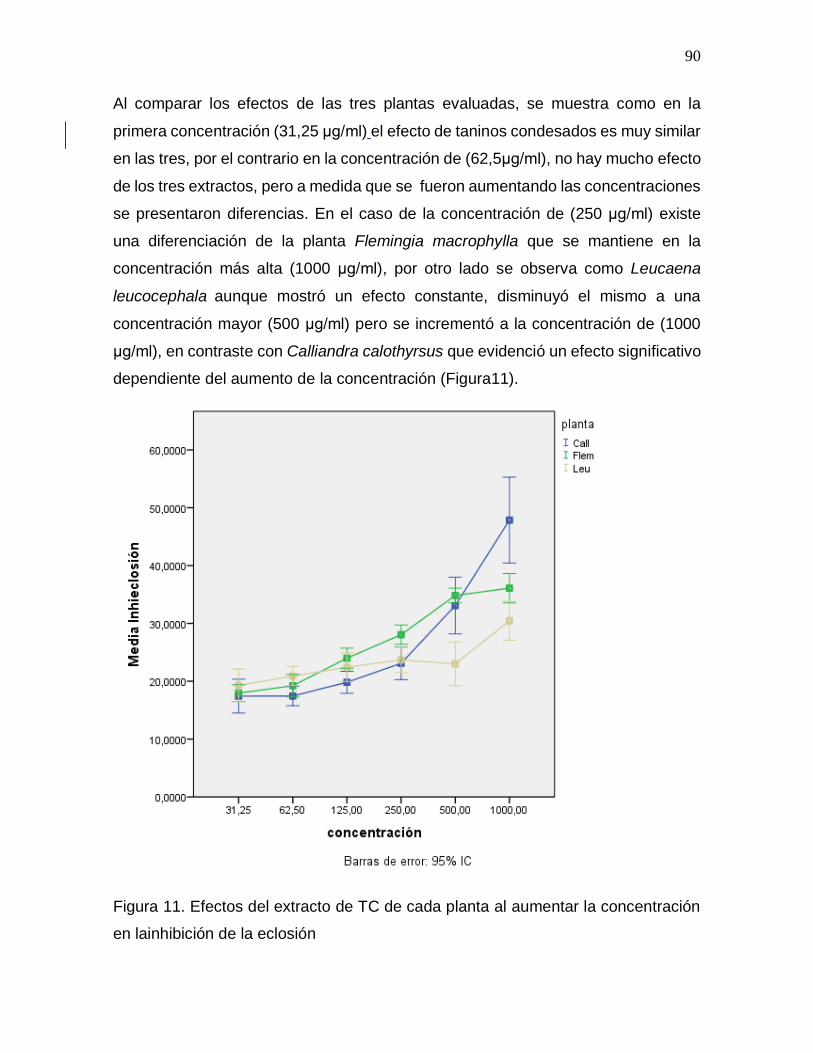
90
Al comparar los efectos de las tres plantas evaluadas, se muestra como en la
primera concentración (31,25 μg/ml) el efecto de taninos condesados es muy similar
en las tres, por el contrario en la concentración de (62,5μg/ml), no hay mucho efecto
de los tres extractos, pero a medida que se fueron aumentando las concentraciones
se presentaron diferencias. En el caso de la concentración de (250 μg/ml) existe
una diferenciación de la planta Flemingia macrophylla que se mantiene en la
concentración más alta (1000 μg/ml), por otro lado se observa como Leucaena
leucocephala aunque mostró un efecto constante, disminuyó el mismo a una
concentración mayor (500 μg/ml) pero se incrementó a la concentración de (1000
μg/ml), en contraste con Calliandra calothyrsus que evidenció un efecto significativo
dependiente del aumento de la concentración (Figura11).
Figura 11. Efectos del extracto de TC de cada planta al aumentar la concentración
en lainhibición de la eclosión

91
5. DISCUSIÓN
Dentro de la familia Trychostrongylidae, los nematodos gastrointestinales de interés
son los del género Haemonchus y Trichostrongylus ya que son los más comunes y
los más diseminados pero también los más patógenos. Otros géneros están
también presentes pero en menor cantidad como Cooperia y Ostertagia (González
et al., 2003). El alto porcentaje de participación de las especies del género H. spp
y en especial el de H. contortus (58%) puede explicarse, debido a su gran potencial
reproductivo, característica particular de esta especie ya que las hembras adultas
tienen la capacidad de producir entre 5000 a 6000 huevos diarios lo que facilita
alcanzar altas cargas parasitarias de forma rápida. H. contortus y H. placei no eran
reconocidos como especies separadas según (Gibbons, 1979), pero más adelante
Le Jambre (1979) no pudo realizar cruces fértiles entre estas dos especies, por lo
cual se consideraron como especies separadas. La diferenciación entre H. contortus
y H. placei según Van wyk et al. (2004) es muy compleja y se podrían cometer
errores por su similitud, aun así con un análisis más preciso puede determinarse
mediante la determinación del porcentaje de la prolongación filamentosa de la cola
de la vaina larval, siendo del 10% al 15% para H. contortus y del 20% para H. placei.
En cuanto a las especies de los géneros Trichostrongylus spp., y Teladorsagia
spp., obtuvieron una participación correspondiente al 35% y 2 % respectivamente;
teniendo en cuenta que la identificación entre este dos géneros no puede llevarse a

92
cabo únicamente por medición ya que la longitud total entre los mismos, es muy
similar, lo que hace necesario analizar y detallar la morfología de su cola, ya que
representa un punto clave para la identificación de estas especies. De acuerdo a lo
anterior las L3 de T. axei y Te. spp., aunque son parecidas por la terminación de la
lisa de la cola, pueden diferenciarse por su longitud total de 720 µm para la primera
y 820 µm para Te. Spp; en contraste con otras especies del genero T. spp., en las
cuales las colas terminan de manera irregular y con protuberancias (Van Wyk, J.A.
y Mayhew, E., 2013). La cola de la vaina de T. falculatus mide aproximadamente 50
µm, siendo más larga que las demás especies de este género, lo que facilita su
identificación (Van Wyk et al., 2004). La identificacion de Cooperia spp. por su parte,
tiene una característica que facilita dicho proceso, ya que posee una cinta fibrilosa
que a la observación microscópica se ve como dos puntos refringentes en el sitio
donde empieza el esófago.
Ostertagia spp, presenta una particularidad para su diferenciación y es el hecho
de que dobla en longitud a Trichostongylus spp, contando además con un filamento
corto diferenciandola de Te. circumcincta y T. spp.,mientras que O.ostertagi termina
en una punta mas contundente que H. placei.
El uso indiscriminado de los antihelmínticos para el control de los parásitos
gastrointestinales, puede generar efectos secundarios como resistencia, toxicidad y
reacciones adversas. Un efecto indirecto es la amenaza a la integridad ambiental;
hay antecedentes que confirman que la administración de antiparasitarios ocasiona
cambios en los organismos colonizadores de la materia fecal de animales tratados

93
con estos productos (Iglesias et al., 2005), lo que ha venido incrementando la
utilización de extractos de plantas o nutracéuticos como alternativa o complemento
a las drogas sintéticas en el control de las parasitosis (Sandoval et al., 2012).
Entre las estrategias exploradas se encuentra el uso de plantas con metabolitos
secundarios, en especial las plantas ricas en compuestos polifenólicos como los
taninos condensados (TC) (Hoste et al., 2006; Brunet et al., 2008; Sandoval-Castro
et al., 2012) con fines antihelmínticos.
El posible papel antiparasitario de los taninos condensados es apoyado por una
gran mayoría de estudios in vivo, lo que ha sido confirmado por los resultados de
bioensayos in vitro que midieron los efectos de extractos de taninos condensados
en las diferentes etapas de los nematodos (Athanasiadou et al., 2001). Además del
contenido de taninos en la dieta, los experimentos in vitro indican que su estructura
química característica también podría ser importante en el proceso de modulación
de la actividad parasitaria (Hoste et al., 2006), los resultados obtenidos con las
unidades monoméricas (flavan-3-oles), sugieren que la naturaleza de los taninos
condensados, en particular la relativa proporción de prodelfinidinas a procianidinas
en plantas podría influir en su eficacia contra los nematodos; ésta hipotésis se apoya
en el hecho de que los derivados galoil de flavan 3 base de las prodelfinidinas, son
más eficaces contra los diferentes estadios parasitarios que las unidades básicas
de procianidinas (Molan et al., 2005).
Makkar et al., (1995) mencionan el efecto directo e indirecto de los taninos
condensados sobre los parásitos gastrointestinales, demostrando que estos TC
empleados en la dieta de ovinos y cabras producen una disminución significativa
del recuento de huevos por gramo de materia fecal; además de la inhibición del

94
desarrollo de larvas en las heces. Por su parte Niezen et al. (1995,1998) y Molan et
al. (2002) confirman que los efectos directos de los TC pueden ser mediados por
interacciones entre nematodos y TC que afectan directamente el funcionamiento
fisiológico del parásito gastrointestinal.
Ademola, Idowu y Fagbemi (2004) atribuyeron la actividad antihelmíntica sobre
huevos y larvas a la presencia de taninos condensados en los extractos utilizados.
Los extractos de taninos condensados de las tres plantas Calliandra calothyrsus,
Flemingia macrophylla, y Leucaena leucocephala, inhibieron el proceso de eclosión
de huevos de manera significativa en la mayoría de las concentraciones, indicando
un posible efecto dosis dependiente, que concuerda con un estudio realizado por
Alonso et al., (2008 a, b) en el cual se demostró éste efecto dependiente de la dosis
de los extractos de diferentes plantas sobre H. contortus y T. colubriformis. Estudios
previos han reportado la validez y efectividad in vitro en la inhibición de la eclosión
de huevos de NGI, utilizando taninos condensados (Furtado, K. et al., 2005). Los
ensayos in vitro realizados por Iqbal et al (2007) también mostraron una inhibición
de la eclosión de huevos de los nematodos dependiente de la dosis mientras que
no hubo efectos de taninos condensados en el adulto de Haemonchus contortus
concluyendo que los efectos antihelmínticos directos de los taninos condensados
parece estar limitado a las etapas de desarrollo.
Un reporte hecho por molan et al (2014) llevado a cabo utilizando taninos
condensados sobre los diferentes estadios de larvas y huevos de nematodos
gastrointestinales, demostró el efecto inhibidor directo del proceso de eclosión in
vitro utilizando diferentes concentraciones. De manera similar un estudio realizado
con el fin de determinar el efecto ovicida de tres extractos acetónicos de las plantas

95
Calliandra calotyrsus, Gliricidia sepium y Leucaena diversifolia en H. contortus a
diferentes concentraciones (0,075; 0.15; 0.3; 0.6 y 1.25 mg/ml), evidenció efectos
negativos con resultados variables de acuerdo a cada concentración, empleando
extractos que incluían todo un grupo de metabolitos secundarios; ésta diversidad de
resultados entre cada extracto fue relacionada directamente con la variabilidad en
la concentración, principalmente de los taninos condensados en cada planta (Ponè
et al. 2011). Una investigación reciente realizada por Engström et al. (2016) en la
cual se emplearon taninos condensados e hidrolizables, mostró variabilidad en los
resultados de la inhibición de la eclosión entre un 50%(TC) y 20% (TH), y en la dosis
efectiva 50, presentando una mejor actividad en concentraciones más bajas pero no
así en las concentraciones máximas.
Si bien en la presente investigación existió un efecto inhibitorio sobre huevos de
nematodos gastrointestinales, dicha inhibición no fue del 100%, resultados que se
pueden atribuir a la ausencia de otros metabolitos secundarios a los cuales se les
atribuye la actividad ovicida tal como lo reportado por Athanasiadou et al. (2001),
Molan et al. (2003) y Monteiro et al. (2011).
Teniendo en cuenta que las larvas de la familia Trichostrongylidae no tienen la
habilidad de infectar al hospedero si no pierden su cutícula (Hertzberg et al., 2002),
se determinó realizar en este proyecto la evaluación del efecto antihelmíntico in vitro
de extractos de taninos condensados correspondientes a Leucaena leucocephala,
Calliandra calothyrsus y Flemingia macrophylla sobre larvas L3 de nematodos
gastrointestinales de ovinos, mediante la prueba de la eliminación de la cutícula que
ha sido desarrollada para medir la capacidad de la planta de poder retrasar o inhibir
artificialmente el desenvaine larval (Bahuaud et al., 2006), siendo considerada como

96
una prueba sensible, reproducible y relevante para los procesos que se producen in
vivo (Díaz et al., 2011).
Aunque todos los extractos inhibieron el proceso de eliminación de la cutícula,
fueron C. calothyrsus y L. leucocephala los que mejores porcentajes de inhibición
presentaron frente a los porcentajes de F. macrophylla; esto concuerda con un
estudio realizado por (Bahuaud et al., 2006) en donde el mecanismo de acción de
los taninos condensados sobre el desenvaine de las larvas L3 de H. contortus y
T.colubriformis mostraron un retraso o inhibición total de la pérdida de la cutícula en
función de la planta aplicada. Brune y Hoste (2006) confirmaron este efecto pero
determinaron que monómeros (flavan-3-ols) de los taninos condensados afectaban
esta no pérdida de la cutícula estableciendo diferencias de susceptibilidad a estos
taninos por parte de la especie parasitaria (H.contortus y T.colubriformis), debido
probablemente a las características y composición de la superficie externa de cada
parásito, indicando un posible efecto de la interacción de la proteína parasitaria y el
tanino condensado. Un mecanismo de acción directa de los taninos sobre el control
de nematodos propuesto por Iqbal et al. (2007) es el de la unión de los mismos a la
cutícula de la larva, la cual está compuesta por una alta cantidad de glicoproteínas
que al ser destruidas y disminuidas producen la muerte del parásito.
Los taninos condensados son capaces de interrumpir el ciclo de vida de los
nematodos y sus efectos varian de acuerdo con la especie de parásito y etapa,
pudiendo prevenir la eclosión de las larvas para alcanzar el pleno desarrollo a larvas
infectivas (Molan et al., 2002 citado en Molan, 2014).
El extracto de Calliandra que evidenció altos porcentajes de inhibición en todas
las concentraciones, además de presentar las mejores dosis efecto 50 en las dos

97
pruebas y la dosis efecto 99 más eficaz en la inhibición de la eclosión, podría estar
relacionado con las altos contenidos de taninos condensados que se reportan en
esta planta de hasta 11% (Palmer, Macqueen y Gutteridae, 1994).
El extracto de Leucaena presentó también un efecto significativo dependiente de
la dosis de manera similar a otra investigación donde se mostró igual efecto desde
una dosis de 75 hasta 1200 μg/ml (Díaz et al., 2011).
La mayoría de estudios sobre las posibles propiedades antihelmínticas de los
extractos de plantas sobre las larvas L3 de los nematodos gastrointestinales, han
relacionado los efectos antiparasitarios observados con la concentración (Molan et
al., 2000; Paolini et al., 2003).
Fernex et al., (2012) utilizaron 4 extractos de plantas con alto contenido de
metabolitos secundarios en larvas L3 de H. contortus, y encontraron una inhibición
completa de su desarrollo, atribuyendo este efecto antihelmíntico, a sus compuestos
polifenólicos como un conjunto.
Poné et al., (2011) adjudicó la actividad antiparasitaria de los extractos de las
leguminosas forrajeras, principalmente, a la presencia de compuestos fenólicos y
particularmente, a los taninos que se encuentran en las hojas; éstas sustancias son
capaces de penetrar la cutícula del nematodo y prevenir la absorción de glucosa o
bloquear los receptores post-sinápticos y en consecuencia paralizar la larva.
Joji y Beena (2010) hacen mención de que los metabolitos secundarios como:
rotenoides, cumarinas, taninos condensados, flavonoides, triterpenos, entre otros,
tienen la capacidad de alterar la biología de los insectos.

98
6. CONCLUSIONES
Se demostró que los extractos de taninos condensados de las especies de plantas
evaluadas en ésta investigación presentaron propiedades antihelmínticas, por lo
que se podría sugerir su uso como alternativa de control de nematodos
gastrointestinales en pequeños rumiantes.
El empleo de este método de control alternativo a la utilización de
antiparasitarios, muestra una serie de beneficios ambientales que la convierten en
una tecnología limpia.
El efecto antihelmíntico atribuido a los extractos estudiados, 8podría estar
directamente relacionado con la presencia de los taninos condensados puros, en
ausencia de otros tipos de metabolitos secundarios de las plantas entre ellos,
terpenoides, fenilpropanoides, saponinas, y antraquinonas que han evidenciado
también actividad antihelmíntica.
En las tres plantas evaluadas aunque todas presentaron efectos inhibitorios en
las dos pruebas, la de mejor eficacia fue Calliandra calothyrsus, razón por la cual
se recomienda la realización de más estudios, donde se determine el efecto de las
mismas in vivo, garantizando el uso de este tipo de controles a los nematodos
gastrointestinales pero de forma segura.

99
7. LISTA DE REFERENCIAS
- Abdu, S.B., Ehoche, O.W., Adamu, A.M., Bawa, G.S., Hassan, M.R., Yashim,
S.M., y Adamu, H.Y. (2012). Effect of Varying Levels of Zizyphus (Zizyphus
mauritiana) Leaf Meal Inclusion in Concentrate Diet on Performance of
Growing Yankasa Ram Lambs Fed Maize Stover Basal Diet. Iranian Journal
of Applied Animal Science, Vol. 2 Nro.4, Pag: 323-330.
- Ademola, I., Fagbemi, B., Idowu, S. (2004). Evaluation of the anthelmintic activity of Khaya senegalensis extract against gastrointestinal nematodes of sheep: in vitro and in vivo studies. Veterinary Parasitology 122: 151–164
- Aguilar, A. J., Cámara, R., Torres, J. F. y Sandoval, C. (2011). El control de
los nematodos gastrointestinales de los caprinos ¿Dónde estamos?
Bioagrociencias. 4 (2):10-16.
- Alhag, A. M., Abdelrahim, B. N., y Verla, N. I. (2014). Wattle tannins as control
strategy for gastrointestinal nematodes in sheep. African Journal of Agricultural Research, 9(28), 2185-2189.
- Alonso-Díaz M.A., Torres-Acosta F.J., Sandoval-Castro C., Brunet S. and Hoste H., 2008a. The effects of four tropical taniniferous plants on the In vitro larval migration and kinetics of exsheathment of Trichostrongylus colubriformis stages. Vet Parasitology, 153, p. 187-192.
- Alonso-Díaz M.A., Torres-Acosta F.J., Sandoval-Castro C., Aguilar Caballero A.J. and Hoste H. 2008b. In vitro larval migration and kinetics of exsheathment of Haemonchus contortus exposed to four tropical taniniferous plants. Vet Parasitology, 153, p. 313-319.
- Alonso-Díaz, M. A., Torres-Acosta, J. F. J., Sandoval-Castro, C. A., y Hoste, H. (2010). Tannins in tropical tree fodders fed to small ruminants: A friendly foe? Small Ruminant Research, 89(2–3), 164-173.
- Ahmed, M., Singh, M., Bera, A., Bandyopadhyay, S., y Bhattacharya, D. (2011). Molecular basis for identification of species/isolates of gastrointestinal nematode parasites. Asian Pacific Journal of Tropical Medicine, 4(8), 589-593.
- Azaizeh, H., Halahleh, F., Abbas, N., Markovics, A., Muklada, H., Ungar, E. D. y Landau, S. Y. (2013). Polyphenols from Pistacia lentiscus and Phillyrea

100
latifolia impair the exsheathment of gastro-intestinal nematode larvae. Veterinary Parasitology, 191(1–2), 44-50.
- Argel, P.J., Lascano, C.E. (1998). Cratylia argentea: Una nueva leguminosa
arbustiva para suelos ácidos en zonas subhúmedas tropicales. Agroforestería para la Producción Animal en Latinoamérica. CIAT.
- Asovinos. Obtenido el 2 de Mayo, 2015 desde Http://www.asovinos.org/e
- Athanasiadou, S., y Kyriazakis, I. (2004). Plant secondary metabolites: antiparasitic effects and their role in ruminant production systems. Proceedings of the Nutrition Society, 63(04), 631–639.
- Athanasiadou, S., Kyriazakis, I., Jackson, F., Coop, R.L. (2001). Direct anthelmintic effects of condensed tannins towards different gastrointestinal nematodes of sheep: in vitro and in vivo studies. Veterinary Parasitology. 99, 205- 219.
- Avello L, M., y Cisternas F, I. (2010). Fitoterapia, sus orígenes, características y situación en Chile. Revista médica de Chile, 138(10), 1288-1293.
- Ayala Paz, D.E., Pérez Guzmán, J.M. 2006 Estudio fisiológico de Flemingia macrophylla (willd) y comportamiento de la planta en la universidad earth. Trabajo de grado. U. Earth. Guácimo, Costa Rica.
- Bahuaud, D., Martínez- Ortíz de Montellano, C., Chaveau, S., Prevot, F., Torres Acosta, J.F.J., Fouraste, I., Hoste, H. (2006). Effects of four tanniferous plant extracts on the in vitro exsheatment third – stage larvae of parasitic nematodes. Parasitology 132, 545-554.
- Barger, I. A. (1999). The role of epidemiological knowledge and grazing management for helminth control in small ruminants. International Journal for Parasitology, 29(1), 41-47; discussion 49-50.
- Barry, T. N., y Manley, T. R. (1984). The role of condensed tannins in the nutritional value of Lotus pedunculatus for sheep. 2. Quantitative digestion of carbohydrates and proteins. The British Journal of Nutrition, 51(3), 493-504.
- Barry, T. N. (1985). The role of condensed tannins in the nutritional value of Lotus pedunculatus for sheep. Rates of body and wool growth. The British Journal of Nutrition, 54(1), 211-217.
- Barry, T. N., y McNabb, W. C. (1999). The implications of condensed tannins
on the nutritive value of temperate forages fed to ruminants. The British Journal of Nutrition, 81(4), 263-272

101
- Bath, G. F. y van Wyk, J. A. (2009). The Five Point Check© for targeted
selective treatment of internal parasites in small ruminants. Small Ruminant Research, 86(1–3), 6-13.
- Baker, N. F., Cook, E. F., Douglas, J. R., y Cornelius, C. E. (1959). The pathogenesis of trichostrongyloid parasites. III. Some physiological observations in lambs suffering from acute parasitic gastroenteritis. The Journal of Parasitology, 45, 643-651.
- Becker, A., Reith, A., Napiwotzki, J., y Kadenbach, B. (1996). A quantitative
method of determining initial amounts of DNA by polymerase chain reaction cycle titration using digital imaging and a novel DNA stain. Analytical Biochemistry, 237(2), 204-207.
- Berghen, P., Hilderson, H., Vercruysse, J. y Dorny, P. (1993). Evaluation of
pepsinogen, gastrin and antibody response in diagnosing ostertagiasis. Veterinary Parasitology, 46(1-4), 175-195.
- Berdonces, J.L. 2003. Historia de la Fitoterapia. Natura Medicatrix 21(3): 142
– 152.
- Bizimenyera, E.S., Githiori, J.B., Eloff, J.N. y Swan, G.E. (2006). In vitro activity of Peltophorum africanum Sond. (Fabaceae) extracts on the egg hatching and larval development of the parasitic nematode Trichostrongylus colubriformis. Veterinary Parasitology. 142, 336–343.
- Brasil, B. S. A. F., Nunes, R. L., Bastianetto, E., Drummond, M. G., Carvalho,
D. C., Leite, R. C. y Oliveira, D. A. A. (2012). Genetic diversity patterns of Haemonchus placei and Haemonchus contortus populations isolated from domestic ruminants in Brazil. International Journal for Parasitology, 42(5), 469-479.
- Brightling, A., (1988). Sheep Diseases. Inkata Press, Melbourne, Australia pp. 3–10
- Brunet, S., Martínez-Ortiz-de-Montellano, C., Torres Acosta, J.F.J. Sandoval-Castro, C.A., Aguilar-Caballero, A.J., Capetillo-Leal, C., Hoste, H., 2008. Effect of the consumption of Lysiloma latisiliquum on the larval establishment of gastrointestinal nematodes in goats. Veterinary Parasitology. 157, 81-88.
- Bunglavan, S.J. y Dutta, N., J. (2013). Use of Tannins as Organic Protectants of Proteins in Digestion of Ruminants. Journal of Livestock Science. 4: 67-77.

102
- Cano, R., J., Carulla, E. y Lascano, C.E. (1994). Métodos de conservación de muestras de forraje de leguminosas tropicales y su efecto en el nivel y la actividad biológica de los taninos. Pasturas Tropicales. 16 (1): 2-7
- Cañigueral S., Dellacassa E. y Bandoni AL 2003. Plantas Medicinales y
Fitoterapia: ¿indicadores de dependencia o factores de desarrollo? Acta Farm Bonaerense 22: 265-278.
- Caracostantogolo J., Martínez L.M. y Ribicich M. (2007). Tesis: Caracterización de proteínas de membrana de intestino de Haemonchus contortus. Área de Parasitología, Instituto de Patobiología. Universidad de Buenos Aires. 12-13.
- CIAT. Obtenido 2 de mayo de 2015. Desde http://ciat.cgiar.org/es.
- Craig, T. M. (2009). CHAPTER 22 - Helminth Parasites of the Ruminant Gastrointestinal Tract. En D. E. A. M. Rings (Ed.), Food Animal Practice (Fifth Edition) (pp. 78-91). Saint Louis: W.B. Saunders.
- Cohen, S., Sadun, E.H., (1976). Immunology of Parasitic Infections. Blackwell Scientific Publications, Oxford, UK.
- Coles, G.C., Bauer, C., Borgsteede, F.H.M., Geerts, S., Klei, T.R., Taylor, M.A. y Waller, P.J. (1992). World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P.) methods for the detection of anthelmintic resistance in nematodes of veterinary.
- Corwin, R. M. (1997). Economics of gastrointestinal parasitism of cattle. Veterinary Parasitology, 72(3-4), 451-457; discussion 457-460.
- Cox, D. D. y Todd, A. C. (1962). Survey of gastrointestinal parasitism in Wisconsin dairy cattle. Journal of the American Veterinary Medical Association, 141, 706-709.
- De Souza, A.C. Protocolo Laboratório Sanidae Animal (2012). Embrapa Pecuária Sudeste.
- Díaz, O., Ortiz, J., Reina, J., Patiño, A., Linares, C. y González, P. (2012).
Colombia sanidad animal 2010. Instituto Colombiano Agropecuario. 8-72.
- Díaz, A., Torres, J., Sandoval, C., Aguilar, A., Hoste, H. (2008a). In vitro larval migration and kinetics of exsheathment of Haemonchus contortus exposed to four tropical tanniniferous plants. Vet. Parasitol. 153, 313–319.
- Díaz, A., Torres, J., Sandoval, C., Capetillo, C., Brunet, S., Hoste, H. (2008b). Effects of four tropical tanniniferous plants on the inhibition of larval migration

103
and the exsheathment process of Trichostrongylus colubriformis infective stage. Vet. Parasitology. 153, 187–192.
- Díaz, A., Torres- Acosta J.F., Sandoval-Castro C.A., Hoste, H. (2011). Comparing the sensitivity of two in vitro assays to evaluate the anthelmintic activity of tropical tannin rich plant extracts against Haemonchus contortus.Vet. Parasitology. 181(2-4):360-4.
- Drudge, J. H., Szanto, J., Wyant, Z. N., y Elam, G. (1964). Field Studies on Parasite Control in Sheep: Comparison of Thiabensazole, Ruelene, and Phenothiazine. American Journal of Veterinary Research, 25, 1512-1518.
- Doenhoff, M. J., Chiodini, P. L., y Hamilton, J. V. (2004). Specific and sensitive diagnosis of schist some infection: can it be done with antibodies? Trends in Parasitology, 20(1), 35-39.
- Domingues, L. F., Giglioti, R., Feitosa, K. A., Fantatto, R. R., Rabelo, M. D.,
de Sena Oliveira, M. C., y De Souza Chagas, A. C. (2013). In vitro and in vivo evaluation of the activity of pineapple (Ananas comosus) on Haemonchus contortus in Santa Inês sheep. Veterinary Parasitology, 197(1-2), 263-270.
- Eddi, C., Nari, A. y Amanfu, W. (2003). Taenia solium cysticercosis/taeniosis:
potential linkage with FAO activities; FAO support possibilities. Acta Tropica, 87(1), 145-148.
- Engström, M. T., Karonen, M., Ahern, J. R., Baert, N., Payré, B., Hoste, H., y Salminen, J.-P. (2016). Chemical Structures of Plant Hydrolyzable Tannins Reveal Their in Vitro Activity against Egg Hatching and Motility of Haemonchus contortus Nematodes. Journal of Agricultural and Food Chemistry, 64(4), 840-851.
- Ensuncho-Hoyos, C., Castellano-Coronado, A., Maza-Ángulo, L., Bustamante-Yánez, M. y Vergara-Garay, O. (2014). Prevalencia y grado de infección de nematodos gastrointestinales en ovinos de pelo en pastoreo de cuatro municipios de Córdoba, Colombia.
- Espinoza, F., Torres, A. y Chacón, E. (2007). Leucaena (Leucaena leucocephala) y Cují (Acacia macracantha y Mimosa tenuiflora) como aporte de proteína económica en los sistemas doble propósito. Memorias I simposio tecnología para la ganadería de los llanos de Venezuela. Pp. 47-70
- Fernex, E., Alonso, M., Valles, B. y Capetillo, C. (2012). In vitro anthelmintic activity of the tropical legumes on the esxsheathment and motility of Haemonchus contortus infective larvae. Exp. Parasitology 131(4), 413-8.

104
- Foley, W.J., Iason, G.R. y MacArthur, C. (1999). Role of secondary metabolites in the nutritional ecology of mammalian herbivores: How far have we come in 25 years? In nutritional ecology of herbivores. H.J. Jung y G.C. Fahey, J.I. (Eds). American society of animal science. pp. 130-209.
- Fox, M. T. (1997). Pathophysiology of infection with gastrointestinal nematodes in domestic ruminants: recent developments. Veterinary Parasitology, 72(3-4), 285-297; discussion 297-308.
- Furtado, K., Negrelle, R., Miguel, O., Zaniolo, S., Kapronezai, J., Ramos, S. y Sotello, A. (2005). The effect of Carica papaya L. (caricaceae) and Musa paradisiaca Linn. (Musaceae) on the development of gastrointestinal nematode eggs from ovine. Arquivos do Instituto Biológico São Paulo 72, 191–197.
- Frutos, P., Hervás, G., F. J. Giráldez, F. J. y Mantecón, A.R. (2004). Review.
Tannins and ruminant nutrition. Spanish Journal of Agricultural Research 2 (2), 191-202.
- Frutos, P., Hervàs, G., Ramos, G., Giráldez, F.J. y Mantecón, A.R. (2002).
Condensed tannin content of several shrub species from a mountain area in northern Spain, and its relationship to various indicators of nutrient value. Animal Feed Sci Tech. 95; 215-226.
- Galindo, J., González, N., Delgado, D., Sosa, A., Marrero, Y., González, R., Aldana, A. y Moreira, O. (2008). Efecto modulador de Leucaena leucocephala sobre la microbiota ruminal. Zootecnia tropical 26(3): 249-252. Instituto de Ciencia Animal la Habana Cuba.
- García, A. y Pérez- Urria Carril, E. (2009). Metabolitos Secundarios de
Plantas. Reduca (Biología). Serie Fisiología Vegetal. 2 (3): 119-145.
- Giudici, G. A. (1999). Changes in gastro-intestinal helminth species diversity in lambs under mixed grazing on irrigated pastures in the tropics (French West Indies). Veterinary Research, 30(6), 573-81.
- Gibbons, L.M. (1979). Revision of the genus Haemonchus Cobb, 1898: (Nematoda Trichostrongylidae). Systematic Parasitology. V. 1 n.1 p. 3-24
- Gomes, E. F. (2013). Inhibition of infective larvae exsheathment and egg hatching of the nematode Haemonchus contortus with extracts of tannin-rich plants (text). Universidad de São Paulo.
- Gordon, H.M. y Whitlock, H.V. (1939). A new technique for counting
nematode eggs in sheep faeces. J. Counc. Sci. Ind. Res. 12, 50–52.
- Gordon, H.M. (1967). Some aspects of the control of helminthosis in sheep. Veterinary Inspector N.S.W. 31, 88–99.

105
- Hagerman, A. E., Robbins, C.T., Weerasuriya, Y., Wilson, T.C. y MacArthur,
C. (1992). Tannin chemistry in relation to digestion. Journal Range Manage. 45:57-62.
- Hagerman, Ricco, R., Sena, G., Vai, V., Wagner, M. y Gurni, A. (1992)
Determinación de la Presencia de Taninos Condensados en Ephedra americana Humb. et Bonpl. ex Wild. (Ephedraceae) 23 (1): 11-5
- Harbone, J.B. (1993). Feeding preferences of vertebrates, including man. In:
Introduction to ecological biochemistry (4th Edn). J.B. Harbone (Ed), pp. 162-210. Academic Press (Reino Unido).
- Herrera O, L., Ríos O, L., y Zapata S, R. (2013). Frecuencia de la infección por nematodos gastrointestinales en ovinos y caprinos de cinco municipios de Antioquia. Revista MVZ Córdoba, 18(3), 3851-3860.
- Hervás, G., Frutos, P., Ramos, G., Giráldez, F.J., Álvarez del Pino, M.C. y Mantecón, A.R. (2000). Gas Production: Fermentation Kinetics for Feed Evaluation and to Assess Microbial Activity. Proceedings of an EAAP Satellite Symposium on Gas Production, pp. 76-77.
- Herazo, Meza, H. (2006). Revisión general de los aspectos fundamentales de la leguminosa Cratylia argentea (veranera). Línea Continental Tropical. Universidad de Sucre. Programa de Zootecnia. Obtenido el 18 de Noviembre, 2015 desde htpp://repositorio.unisucre.edu.co/handle/001/484.
- Hertzberg, H., Huwyler, U., Kohler, L., Rehbein, S., Wanner, M. (2002). Kinetics of exsheathment of infective ovine and bovine strongylid larvae in vivo and in vitro. Parasitology 125, 65-70
- Hoste, H., Jackson, F., Athanasiadou, S., Thamsborg, S. M. y Hoskin, S. O.
(2006). The effects of tannin-rich plants on parasitic nematodes in ruminants. Trends in Parasitology, 22(6), 253-261.
- Hoste, H., Torres-Acosta, J.F. (2011). Non chemical control of helminths in
ruminants: adapting solutions for changing worms in a changing world. Veterinary Parasitology. 180, 144–154.
- Hoberg, E. R., Lichtenfels, J. R., y Gibbons, L. (2004). Phylogeny for species
of Haemonchus (Nematoda: Trichostrongyloidea): considerations of their evolutionary history and global biogeography among Camelidae and Pecora (Artiodactyla). The Journal of Parasitology, 90(5), 1085-1102.
- Huang, X.D., Liang, J.B., Tan, H.Y., Yahya, R., Khamseekhiew, B. y Ho, Y.W. (2011). Effects of Leucaena condensed tannins of differing molecular weights

106
on in vitro CH4 production. Animal Feed Science. Technol. Vol.166 -167. 373-376.
- Hungerford, T.G., (1990). Diseases of Livestock, ninth ed. McGraw-Hill.
Medical, Sydney, Australia. - Iqbal, Z., Sarwar, M., Tabbar, A., Ahmed, S., Nisa, M., Sajid, M.S., Khan,
M.N., Muft, K.A., Yaseen, M. (2007). Direct and indirect anthelmintic effects of condensed tannins in sheep. Veterinary Parasitology 144: 125-131.
- Iglesias, L.E., Saumell, C.A., Fusé, L.A., Lifschitz, A.L., Rodríguez, E.M., Steffan, P.E., Fiel, C.A. (2005). Impacto ambiental de la ivermectina eliminada por bovinos tratados en otoño, sobre la coprofauna y la degradación de la materia fecal en pasturas (Tandil, Argentina). RIA 34 (3): 83-103, Inta Argentina.
- Joji, R.l. y Beena, J. 2010. Chemical Composition and Antibacterial Activity of the Volatile Oil from the Bark of Gliricidia Sepium. International Journal of Pharmacy and Pharmaceutical Sciences. Vol 2, Issue 3.
- Kassai, T., (1999). Veterinary Helminthology. Butterworth Heinemann,
Oxford, UK.
- Knox, D. P., Redmond, D. L., Newlands, G. F., Skuce, P. J., Pettit, D. y Smith, W. D. (2003). The nature and prospects for gut membrane proteins as vaccine candidates for Haemonchus contortus and other ruminant trichostrongyloids. International Journal for Parasitology, 33(11), 1129-1137.
- Lane, C. (1922). The mass diagnosis of ankylostome infestation (Part I).
Trans. R. Soc. Trop. Med. Hyg. 16, 274–315.
- Lanusse, C. E., Gascon, L. H. y Prichard, R. K. (1993). Gastrointestinal distribution of albendazole metabolites following netobimin administration to cattle: relationship with plasma disposition kinetics. Journal of Veterinary Pharmacology and Therapeutics, 16(1), 38-47.
- Larsen, J.W.A., Anderson, N., Vizard, A.L., Anderson, G.A. y Hoste, H.,
(1994). Diarrhea in Merino ewes during winter: association with trichostrongyloid larvae. Australian. Veterinary. Journal. 71, 365–372.
- Lara, D. M. (2007). Resistencia a Los Antihelmínticos en Nematodos de
Rumiantes y Estrategias Para Su Control. Corpoica.
- Larsen, J.W.A., Anderson, N., Ware, J.W. y Fegely, C.D., (2006). The productivity of Merino flocks in South-Eastern Australia in the presence of anthelmintic resistance. Small Ruminal. Res. 62, 87–93.

107
- Lascano, C. y Palacios, E. (1993). Intake and digestibility by sheep fed mature grass alone and in combination with two tropical legumes. Tropical Agriculture (Trinidad). 70:365–358.
- Lascano C, Rincón A, Plazas CH, Bueno G, Argel P. Cultivar veranera Cratylia argentea (desvaux) o. Kuntze. Leguminosa arbustiva de usos múltiples para zonas con períodos prolongados de sequía en Colombia. Cali, Colombia: Corpoica, Minagricultura Y Ciat; 2002.
- Launchbaugh, K.L. y Provenza, F.D. (1994). The effect of flavor concentration and toxin doses on the formation and generalization of flavor aversions in lambs. Journal of Animal Science. 72, 10-13.
- Launchbaugh, K.L., Provenza, F.D. y Pfister, J.A. (2001). Herbivore response to anti-quality factors in forages. Journal of Range Management. 54, 431-440
- Leinmüller, E., Steingass, H. y Menke, K.H. (1991). Tannins in ruminants feeds tuffs. Biannual collection of recent German contributions concerning development TH Rough Animal Research. 33, 9-62
- Levine, N.D., (1968). Nematode Parasites of Domestic Animals and of Man.
Burgess Publishing Company, Minneapolis, USA
- Le Jambre, L.F. (1979). Hybridization studies of Haemonchus contortus. International Journal for Parasitology. 9: 455-463
- Luque, A., Barry, T.N., Mcnabb, W.C., Kemp, P.D. y McDonald, M.F. (2000). The effect of grazing Lotus corniculatus during summer-autumn on reproductive efficiency and wool production in ewes. Australian Journal of Agricultural Research, 51: 385 -391.
- MAFF, (1986). Manual of Veterinary Parasitological Laboratory Techniques. Her Majesty’s Stationary Office, London, UK pp. 20–27.
- Makkar, H.P.S., Singh, B. y Dowra, R.K. (1988). Effects of tannin-rich of Oak (Quercys incana) on various microbial enzyme activities of the bovine rumen. British Journal of nutrition. 60, 287-296.
- Makkar, H. P. S., Becker, K., Abel, H., y Szegletti, C. (1995). Degradation of condensed tannins by rumen microbes exposed to quebracho tannins (QT) in rumen simulation technique (RUSITEC) and effects of QT on fermentative processes in the RUSITEC. Journal of the Science of Food and Agriculture, 69(4), 495-500

108
- Makkar, H.P.S. y Becker, K. (1998). Do tannins in leaves of trees and shrubs from African and Himalayan regions differ in level and activity? Agroforestry System, 40:59–68
- Márquez, D. (2003). Nuevas tendencias para el control de parásitos de
bovinos en Colombia. Corpoica. Pág. 1-52. Vol 4 No.1
- Márquez Lara, D. y Suárez Londoño, A. (2008). El uso de taninos condensados como alternativa nutricional y sanitaria en rumiantes. Revista de Medicina Veterinaria Nro. 16 / julio – diciembre.
- Márquez, D. (2014). Control sostenible de los nematodos gastrointestinales en rumiantes. Corpoica Pág. 230- 254.
- Martínez, N., Meza D. y Guevara, M. (2013). Ampliación y profundización de
estudios en Fitoterapia. Encuentro de saberes desde la universidad de Carabobo, Venezuela. Vol. 11, Nº 2.
- Martínez-Ortiz-de-Montellano, C., Arroyo-López, C., Fourquaux, I., Torres-
Acosta., J., Sandoval-Castro, C.A. y Hoste, H. (2013). Scanning electron microscopy of Haemonchus contortus exposed to tannin-rich plants under in vivo and in vitro conditions. Experimental Parasitology 133, 281–286.
- Martínez, O. M. C. (2010). Mecanismo de acción de las plantas ricas en taninos sobre la población adulta de nematodos gastrointestinales de pequeños rumiantes. Tesis en cotutela presentada para obtener el grado de Doctor en Ciencias Agropecuarias. Francia: Université de Toulouse.
- Medina, P., Guevara, F., La O, M., Ojeda, N. y Reyes, E. (2014). Resistencia antihelmíntica en ovinos: una revisión de informes del sureste de México y alternativas disponibles para el control de nematodos gastrointestinales. Pastos y Forrajes, 37(3), 257-263.
- McLeod, M.N. (1974). Plant tannins their role in forage quality. Nutrition abstracts and reviews. 44, 803-812.
- McMahon, L.R., McAllister, T.A., Berg, B.P., Majak, W., Acharya, S.N., Popp, J.D., Coulman, B.E., et al. (2000). A review of the effects of forage condensed tannins on ruminal fermentation. Canadian Journal of plant science. 80(3): 469-485.
- McNabb, W. C., Waghorn, G. C., Barry, T. N. y Shelton, I. D. (1993). The effect of condensed tannins in Lotus pedunculatus on the digestion and metabolism of methionine, cysteine and inorganic sulphur in sheep. The British Journal of Nutrition, 70(2), 647-661.

109
- Min, B., Miller, P. y Loetz, M. (2003). Tannins for suppression of internal parasites. Journal of Animal Science. 81: E 102- E109.
- Ministerio de Agricultura y Desarrollo Rural. Observatorio Agrocadenas
Colombia. La cadena de ovinos y caprinos en Colombia. Bogotá: Ministerio de Agricultura y Desarrollo Rural; 2006.
- Monteiro, M., Bevilaqua, C., Morais, S., Machado, L., Camurc¸ a-Vasconcelos, A., Campello, C., Ribeiro, W. y Mesquita, M. (2011). Anthelmintic activity of Jatropha curcas L. seeds on Haemonchus contortus. Vet. Parasitology. 182, 259–263.
- McKenna, P.B., (1987). The estimation of gastrointestinal strongyle worm
burdens in Young sheep flocks: a new approach to the interpretation of faecal egg counts I. Development. NZ Veterinary Journal. 35, 94–97.
- McSweeney, C.S., Palmer, B., Bunch, R. y Krause, D.D. (2001). Effect of the tropical forage Calliandra on microbial protein synthesis and ecology in the rumen. Journal of applied microbiology. 90, 78-88.
- Min, B.R., McNabb, W.C., Barry, T.N., Kemp, P.D., Waghorn, G.C. y
McDonald, M.F. (1999). The effects of condensed tannins in Lotus corniculatus upon reproductive efficiency and wool production in sheep during late summer and autumn. Journal of Agricultural Science, Cambridge. 132, 323-334.
- Min, B.R., Fernández, J.M., Barry, T.N., Mcnabb, W.C. y Kemp, P.D. (2001).
The effect of condensed tannins in Lotus corniculatus upon reproductive efficiency and wool production in ewes during autumn. Animal Feed Science and Technology, 92: 185-202.
- Min, B. R., Barry, T. N., Attwood, G. T., y McNabb, W. C. (2003). The effect
of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: a review. Animal Feed Science and Technology, 106(1–4), 3-19.
- Min, B.R., Hart, S.P., Miller, D., Tomita, G.M., Loetz, E., y Sahlu, T. (2005).
The effect of grazing forage containing condensed tannins on gastro intestinal parasite infection and milk composition in Angora does. Veterinary Parasitology. 130: 105-113.
- Molle, G., Decandia, M., Cabiddu, A. y Sitzia, M. (2003). The performance of Mediterranean dairy sheep given access to Sulla (Hedysarum coronarium l.) and annual ryegrass (Lolium rigidum gaudin) pastures in different time proportions. Small Ruminant Research. 49: 319-328.

110
- Molan, A.L., (2014). Effect of purified condensed tannins from pine bark on larval motility, egg hatching and larval development of Teladorsagia circumcincta and Trichostrongylus colubriformis (Nematoda: Trichostrongylidae). Folia Parasitologica 61 [4]: 371–376.
- Molan, A.L. (2005). Effect of flavan-3-ols on in vitro egg hatching larval development and viability of infective larvae of Trichonstrongylus colubriformis. Int. J. Parasitology. 33, 1697-1698.
- Molan, A. L., Waghorn, G. C., y McNabb, W. C. (2002). Effect of condensed tannins on egg hatching and larval development of Trichostrongylus colubriformis in vitro. The Veterinary Record, 150(3), 65-69.
- Moilanen, J., y Salminen, J.-P. (2007). Ecologically neglected tannins and their biologically relevant activity: chemical structures of plant ellagitannins reveal their in vitro oxidative activity at high ph. Chemo ecology, 18(2), 73-83.
- Moreno-Murillo, B., Sánchez, A., Quevedo, R., Pabón, M., Juan E. Carulla F, J.E. (2008). 3-o-a-l-Ramnopiranosilflavonoides y otros derivados fenólicos de hojas de Calliandra calothyrsus. Meissner (Mimosaceae). Revista Colombiana de Química. vol.37 no.3 Bogotá Sept. /Dec.
- Mottier, L., Alvarez, L., Fairweather, I., y Lanusse, C. (2006). Resistance-
induced changes in triclabendazole transport in Fasciola hepatica: ivermectin reversal effect. Journal of Parasitology, 92(6), 1355–1360.
- Morris, C. A., Vlassoff, A., Bisset, S. A., Baker, R. L., Watson, H. T. G., West,
C. J., y Wheeler, M. (2000). Continued selection of Romney sheep for resistance or susceptibility to nematode infection: Estimates of direct and correlated responses.
- Mueller- Harvey, I. (2006). Unravelling the conundrum of tannins in animal
nutrition and health. Journal of the Science of Food and Agriculture. 86: 2010-2037.
- Nari, A., Eddi, C., Martins, J. y Benavides, E. (2003) Resistencia a los antiparasitarios, estado actual con énfasis en América Latina. Estudio FAO producción y sanidad animal. Organización de las Naciones Unidas para la Agricultura y la Alimentación. 8-32.
- Narjisse, H., Elhonsali, M.A., y Olsen, J.D. (1995). Effects of Oak (Quercus ilex) tannins on digestion and nitrogen balance in sheep and goats. Small Ruminant Research. 18, 201-206.

111
- Naumann, H.D., Muira, J.P., Lambert, B.D., Tedeschi, L.O. y Kothmanne M.M. (2013). Condensed Tannins in the Ruminant Environment: A Perspective on Biological Activity. Journal of Agricultural Sciences Vol. 1(1), pp. 8-20.
- Nicholls, J. y Obendorf, D.L. (1994). Application of a composite faecal egg
count procedure in diagnostic parasitology. Veterinary. Parasitology. 52, 337–342.
- Nikman, V., Ebrahimzadeh, H., (2002). Phenolics content in astragalus species. Pak. J. Bot., 34(3): 283-289
- Niezen, J. H., Charleston, W. A., Hodgson, J., Miller, C. M., Waghorn, T. S., y Robertson, H. A. (1998). Effect of plant species on the larvae of gastrointestinal nematodes which parasitise sheep. International Journal for Parasitology, 28(5), 791-803.
- Niezen, J. H., Waghorn, T. S., Charleston, W lucerne (Medicago sativa) or Sulla (Hedysarum coronarium) which contains condensed tan. A. G., y Waghorn, G. C. (1995). Growth and gastrointestinal nematode parasitism in lambs grazing either Nins. The Journal of Agricultural Science, 125(2), 281–289.
- Nelson, K.E., Pell, A.N., Duane, P.H., Giner- Chavez, B.I. y Schofield, P. (1997). Chemical and biological assays to evaluate bacterial inhibition by tannins. Journal of Chemical Ecology. 23, 1175-1194.
- Ogunremi, O., Halbert, G., Mainar Jaime, R., Benjamin, J. y Pfister, K. (2008). Accuracy of an indirect fluorescent-antibody test and of a complement-fixation test for the diagnosis of Babesia caballi in field samples from horses. Prev. Veterinary. Medicine. 83, 41–51.
- Otero, M. y Hidalgo, L. (2004). Condensed tannins in temperate forages
species: effects on the productivity of ruminants infected with internal parasites (a review). Livestock Research for Rural Development 16(2): 18-36.
- Oliveira, J.T.A., Rios, F.J.B., Vasconcelos, I.M., Ferreira, F.V.A., Nojosa, G.B.A., y Medeiros, D.A. (2004). Cratylia argentea seed lectin, a possible protein against plant-eating organisms: effects on rat metabolism and gut histology. Food and Chemical Toxicology. Nov.42 (11):1737 – 47.
- Palmer, B., Macqueen, D.J., y Gutteridge, R.C. (1994). Calliandra calothyrsus
- a multipurpose tree legume for humid locations. In: R.C. Gutteridge and H.M.

112
Shelton (Eds), Forage tree legumes in tropical agriculture. CAB International, UK. p. 65-74.
- Papadopoulos, E., Arsenos, G., Sotiraki, S., Deligiannis, C., Lainas, T. y Zygoyiannis, D. (2003). The epizootiology of gastrointestinal nematode parasites in Greek dairy breeds of sheep and goats. Small Ruminant Research, 47(3), 193-202.
- Papadopoulos, E. (2008). Anthelmintic resistance in sheep nematodes. Small Ruminant Research, 76(1–2), 99-103.
- Palomo López, P. y Revuelta García, E. (2012). Fitoterapia básica y podológica. Revista Internacional de Ciencias Podológicas. Vol. 6, Núm. 1, 39-50.
- Paolini, V., Frayssines, A., De La Farge, F., Dorchies, P. y Hoste, H. (2003).
Effects of condensed tannins on established populations and on incoming larvae of Trichostrongylus colubriformis and Teladorsagia circumcincta in goats. Veterinary. Research. 34 (2003) 331–339.
- Perez-Maldonado, R.A. y Norton, B.W. (1996ª). Digestion of C-labelled
condensed tannins from Desmodium intortum in sheep and goats. British of Journal of nutrition. 76, 501-513.
- Perez-Maldonado, R.A. y Norton, B.W. (1996b). The effects of condensed
tannins from Desmodium intortum and Calliandra Calothyrsus on protein and carbohydrate digestion in sheep and goats. British of Journal of nutrition. 76, 515-533.
- Poné, W., Tameli, K., Mbida, M., Tedonkeng, P. y Bilong B., (2011). In vitro
activities of acetonic extracts from leaves of three forage legumes (Calliandra calotyrsus, Gliricidia sepium and Leucaena diversifolia) on Haemonchus contortus. Asian Pacific Journal of Tropical Medicine 125-128.
- Powers, K.G., Wood, I.B., Eckert, J., Gibson, T. y Smith, H.J. (1982). World associations of the advancement of veterinary parasitology (W.A.A.V.P.) guidelines for evaluating the efficacy of anthelmintics in ruminants (bovine and ovine). Veterinary Parasitology 10, 265–284.
- Quiroz Romero H., Figueroa Castillo, J.A., Ibarra Veralde, F., Lopez Arellano, M.E. (2011). Capítulo 16. Ecología de larvas de nematodos gastrointestinales de bovinos, ovinos y caprinos. En epidemiologia de enfermedades parasitarias en animales domésticos (primera edición, pp. 254-288). México.
- Ramírez-Restrepo, C.A. y Barry, T.N. (2005). Alternative temperate forages
containing secondary compounds for improving sustainable productivity in grazing ruminants. Animal Feed Science and Technology, 120: 179-201.

113
- Ramos, G., Frutos, P., Giráldez, F.J. y Mantecón, A.R. (1998). Los compuestos secundarios de las plantas en la nutrición de los herbívoros. Arch. Zootecnia. 47: 597-620. Estación Agrícola Experimental.
- Reed, J.D. (1995). Nutritional toxicology of tannins and related polyphenols in forage legumes. Journal of Animal Science. 73, 1515-1528.
- Rochfort, S., Parker, A.J. y Dunshea, F.R. (2008). Plant bioactives for ruminant health and productivity. Phytochemistry, 69: 299-322.
- Roeber, F., Jex, A. R., y Gasser, R. B. (2013). Chapter Four - Next-
Generation Molecular-Diagnostic Tools for Gastrointestinal Nematodes of Livestock, with an Emphasis on Small Ruminants: A Turning Point? En D. Rollinson (Ed.), Advances in Parasitology (Vol. 83, pp. 267-333). Academic Press.
- Romero, H., Figueroa, J.A., Ibarra, F. y López, M.E. (2011). Capítulo 16. Ecología de larvas de nematodos gastrointestinales de bovinos, ovinos y caprinos. en Epidemiologia de enfermedades parasitarias en animales domésticos (primera edición, pp. 254-288). México.
- Rosales, Méndez, M. (1999). Mezclas de forrajes: Uso de la diversidad forrajera tropical en sistemas agroforestales. Agroforestería para la producción animal en américa latina. Roma (Italia). P. 201-230.
- Russel, A.J.F., Doney, J.M. y Gunn, R.G., (1969). Subjective assessment of body fat in live sheep. Journal. Agric. Science. 72, 451–454
- Sandoval-Castro, C.A., Torres-Acosta, J.F.J., Hoste, H., Salem, A.Z.M., Chan-Pérez, J.I. 2012. Using plant bioactive materials to control gastrointestinal tract helminths in livestock. Animal Feed Science and Technology. 176, 192-201 Torres
- Sankol, P., y Ly, J. (2001). Nutritive evaluation of tropical tree leaves for pigs. (Flemingia macrophylla). Livestock Research for Rural Development 13(5).
- Scalbert, A. (1991). Antimicrobial properties of tannins. Phytochemistry. 30, 3875-3883.
- Scribano, V. (2012). Pastoreo de Leucaena en el control de parásitos.
Producir XXI, Bs. As., 20(252):12-18.
- Sebuliba, E., Nyeko, P., M., Eilu, G., Luswata Kizza, C., y Ekwamu, A. (2012). Enhanced Growth of Multipurpose Calliandra (Calliandra calothyrsus) Using
Arbuscular Mycorrhiza Fungi in Uganda. The Scientific WorldJournal.
Volume 2012, Article ID 830357, 6 pages

114
- Silanikove, N. (2000). The physiological basis of adaptation in goats to hars
environments. Small Ruminant Research. 35, 181-193
- Smith, A.H., Zoetendal, E. y Mackie, R.I. (2005). Bacterial mechanisms to overcome inhibitory effects of dietary tannins. Microbial Ecology. 50: 197-205
- Smith, W. D. y Zarlenga, D. S. (2006). Developments and hurdles in
generating vaccines for controlling helminth parasites of grazing ruminants. Veterinary Parasitology, 139(4), 347-359
- Stoll, N.R. (1923). Investigations on the control of hookworm disease. XV. An effective method of counting hookworm eggs in faeces. American Journal Epidemiology. 3, 59–70.
- Taylor, M.A., Coop, R.L. y Wall, R.L. (2007). Veterinary Parasitology, third ed. Blackwell Publishing, Oxford, UK.
- Van Wyk, J.A. & Mayhew, E. (2013). ‘Morphological identification of parasitic nematode infective larvae of small ruminants and cattle: A practical lab guide’, Onderstepoort Journal of Veterinary Research 80(1), Art. #539, 14 pages.
- Van Wyk, J. A., Cabaret, J., y Michael, L. M. (2004). Morphological identification of nematode larvae of small ruminants and cattle simplified. Veterinary Parasitology, 119(4), 277-306.
- Van Wyk, J.A. y Bath, G.F. (2002). The FAMACHA system for managing haemonchosis in sheep and goats by clinically identifying individual animals for treatment. Veterinary Research. 33, 509–529
- Vercruysse, J., y Claerebout, E. (2001). Treatment vs non-treatment of helminth infections in cattle: defining the threshold. Veterinary Parasitology, 98(1-3), 195-214.
- Von Son –De Fernex, E., Alonso – Díaz, M.A., Valles de la Mora, B. y Capetillo-Leal, C.M. (2012). In vitro antihelmintic activity of five tropical legumes on the exsheathment and motility of Haemonchus contortus infective larvae. Exp. Parasitology. Aug: 31 (4): 413-8.
- Wagner, H. (2006). Futuro en la Investigación en Fitoterapia: Tendencias y Retos. Revista de Fitoterapia. 6 (2) 101-107.
- Waghorn, G. (1996). Condensed tannins and nutrient absorption from the small intestine in: proceedings of the Canadian society of animal science. 175-194.

115
- Wang, Y., Douglas, G. B., Waghorn, G. C., Barry, T. N., y Foote, A. G. (1996).
Effect of condensed tannins in Lotus corniculatus upon lactation performance in ewes. The Journal of Agricultural Science, 126(3), 353–362.
- Waller, P. (1999). International approaches to the concept of integrated control on nematode parasites of livestock. Int. J. for Parasitology. 29:155/164.
- Waterman, P.G. y Mole, S. (1994). Method in ecology. Analysis of phenolic plant metabolites. Blackwell Sci. Publ. London: 66 -103
- Wimmer, B., Craig, B. H., Pilkington, J. G., y Pemberton, J. M. (2004). Non-invasive assessment of parasitic nematode species diversity in wild Soay sheep using molecular markers. International Journal for Parasitology, 34(5), 625-631.
- Wolstenholme, A. J., Fairweather, I., Prichard, R., von Samson-Himmelstjerna, G., y Sangster, N. C. (2004). Drug resistance in veterinary helminths. Trends in Parasitology, 20(10), 469-476
- Zajac, A.M. (2006). Gastrointestinal nematodes of small ruminants: life cycle, anthelmintics, and diagnosis. Vet. Clin. North Am. Food Anim. Pract. 22, 529–541.
- Zhu, J., Filippich, L. J. y Alsalam, M. T. (1992). Tannic acid intoxication in sheep and mice. Research in Veterinary Science, 53(3), 280-292.

116
8. ANEXOS
Anexo1. Imágenes de Recuento de Huevos por gramo de materia fecal 1. Heces tomadas directamente 2. Pesaje de la muestra 3gr de la ampolla rectal del animal.
3. Adicion de 42 ml de una solución 4. Tamizaje y filtrado
de agua destilada con sal para mezcla
5. Lectura y conteo de huevos en la cámara de Mc Master
Fuente (Bernal y Camargo, 2016)

117
Anexo 2. Imágenes Técnica de Coprocultivo y Obtención de larvas 1. Recolección de heces del ovino 2. Mezcla con agua destilada y corriente
en proporción 50:50 para ablandar
3. Heces mezcladas con aserrín 4. Se vierte en frasco de boca ancha,
para formar una masa se tapa y deja con aireación
5. A incubadora 8 días/27°C 6. Volteo (3 horas) aireación día de por medio para migración larval

118
7. Almacenamiento de larvas contenidas en una mezcla de agua destilada y agua
corriente en proporción 50:50 y mantenidas en refrigeración.
Fuente (Bernal y Camargo, 2016)

119
Anexo 3. Imágenes del Lavado de larvas 1. Vénulas con 15 ml sln sacarosa 40% 2. Pesaje y calibración de vénulas + 5ml de larvas L3 proporción 7:5 -2.5
3. Colocadas en centrífuga de refrigeración 4. Separación de los diferentes a 3500 rpm por 5’ a 4°C elementos
5. Extracción y lavado (2) 6. Almacenamiento caja horizontal
Fuente (Bernal y Camargo, 2016)
* Anillo
con larvas
*Sacarosa
*Detritus

120
Anexo 4. Imágenes del procedimiento para la determinación de la concentración adecuada de Hipoclorito de Sodio para la prueba de eliminación de la cutícula.
1. Concentraciones del 2. Preparación del montaje Hipoclorito de Sodio (7.5% - 3.75%)
3. Adición del lugol a los pozos 4. Placa completa
5. Cuantificación de larvas con cutícula y sin cutícula
Fuente (Bernal y Camargo, 2016)

121
Anexo 5. Imágenes Identificación Morfométrica de Larvas L3
1. Extracción de una muestra con una solución de larvas
2. Exposición de lámina al calor 3. Se adicionó una gota de lugol
por breves segundos y se colocó un cubreobjetos
4. Observación al microscopio para identificación morfométrica total de 100 larvas
Fuente (Bernal y Camargo, 2016)

122
5. Análisis morfométrico basado en los siguientes parámetros
Fuente (Bernal y Camargo, 2016)
Longitud
total
Ancho
Longitud de la Vaina de
la cola
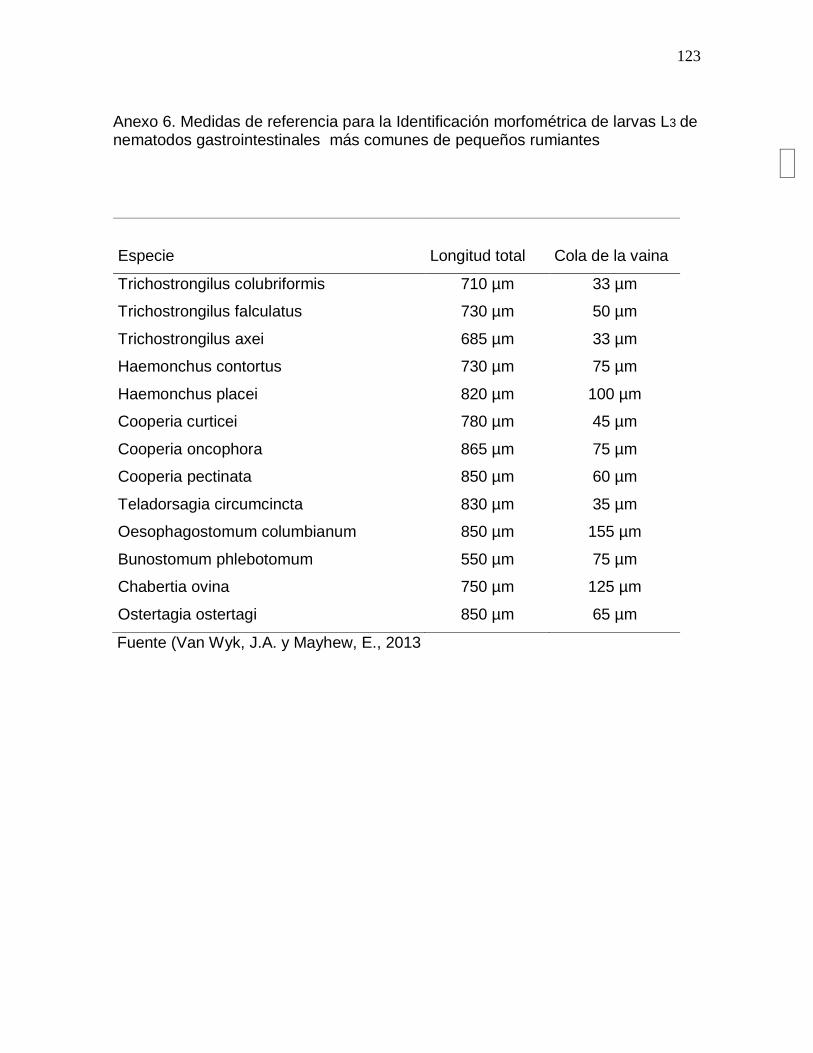
123
Anexo 6. Medidas de referencia para la Identificación morfométrica de larvas L3 de nematodos gastrointestinales más comunes de pequeños rumiantes
Fuente (Van Wyk, J.A. y Mayhew, E., 2013
Especie Longitud total Cola de la vaina
Trichostrongilus colubriformis 710 µm 33 µm
Trichostrongilus falculatus 730 µm 50 µm
Trichostrongilus axei 685 µm 33 µm
Haemonchus contortus 730 µm 75 µm
Haemonchus placei 820 µm 100 µm
Cooperia curticei 780 µm 45 µm
Cooperia oncophora 865 µm 75 µm
Cooperia pectinata 850 µm 60 µm
Teladorsagia circumcincta 830 µm 35 µm
Oesophagostomum columbianum 850 µm 155 µm
Bunostomum phlebotomum 550 µm 75 µm
Chabertia ovina 750 µm 125 µm
Ostertagia ostertagi 850 µm 65 µm
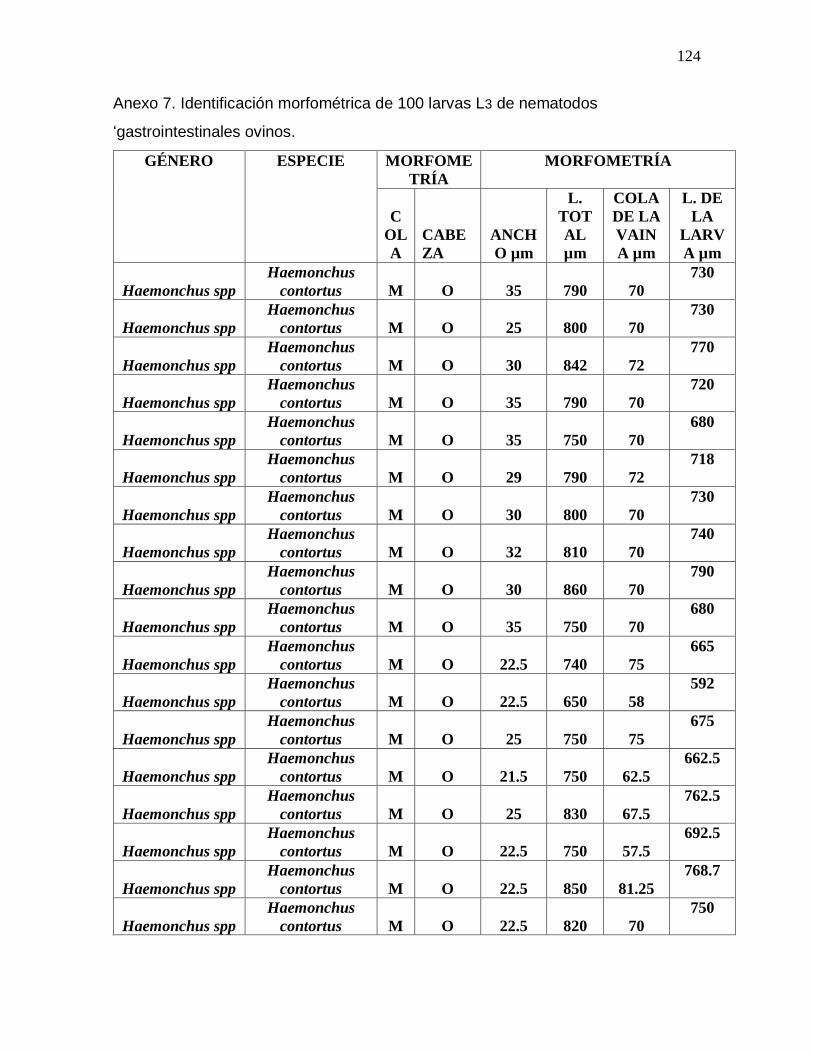
124
Anexo 7. Identificación morfométrica de 100 larvas L3 de nematodos
‘gastrointestinales ovinos.
GÉNERO ESPECIE MORFOME
TRÍA
MORFOMETRÍA
C
OL
A
CABE
ZA
ANCH
O µm
L.
TOT
AL
µm
COLA
DE LA
VAIN
A µm
L. DE
LA
LARV
A µm
Haemonchus spp
Haemonchus
contortus M O 35 790 70
730
Haemonchus spp
Haemonchus
contortus M O 25 800 70
730
Haemonchus spp
Haemonchus
contortus M O 30 842 72
770
Haemonchus spp
Haemonchus
contortus M O 35 790 70
720
Haemonchus spp
Haemonchus
contortus M O 35 750 70
680
Haemonchus spp
Haemonchus
contortus M O 29 790 72
718
Haemonchus spp
Haemonchus
contortus M O 30 800 70
730
Haemonchus spp
Haemonchus
contortus M O 32 810 70
740
Haemonchus spp
Haemonchus
contortus M O 30 860 70
790
Haemonchus spp
Haemonchus
contortus M O 35 750 70
680
Haemonchus spp
Haemonchus
contortus M O 22.5 740 75
665
Haemonchus spp
Haemonchus
contortus M O 22.5 650 58
592
Haemonchus spp
Haemonchus
contortus M O 25 750 75
675
Haemonchus spp
Haemonchus
contortus M O 21.5 750 62.5
662.5
Haemonchus spp
Haemonchus
contortus M O 25 830 67.5
762.5
Haemonchus spp
Haemonchus
contortus M O 22.5 750 57.5
692.5
Haemonchus spp
Haemonchus
contortus M O 22.5 850 81.25
768.7
Haemonchus spp
Haemonchus
contortus M O 22.5 820 70
750
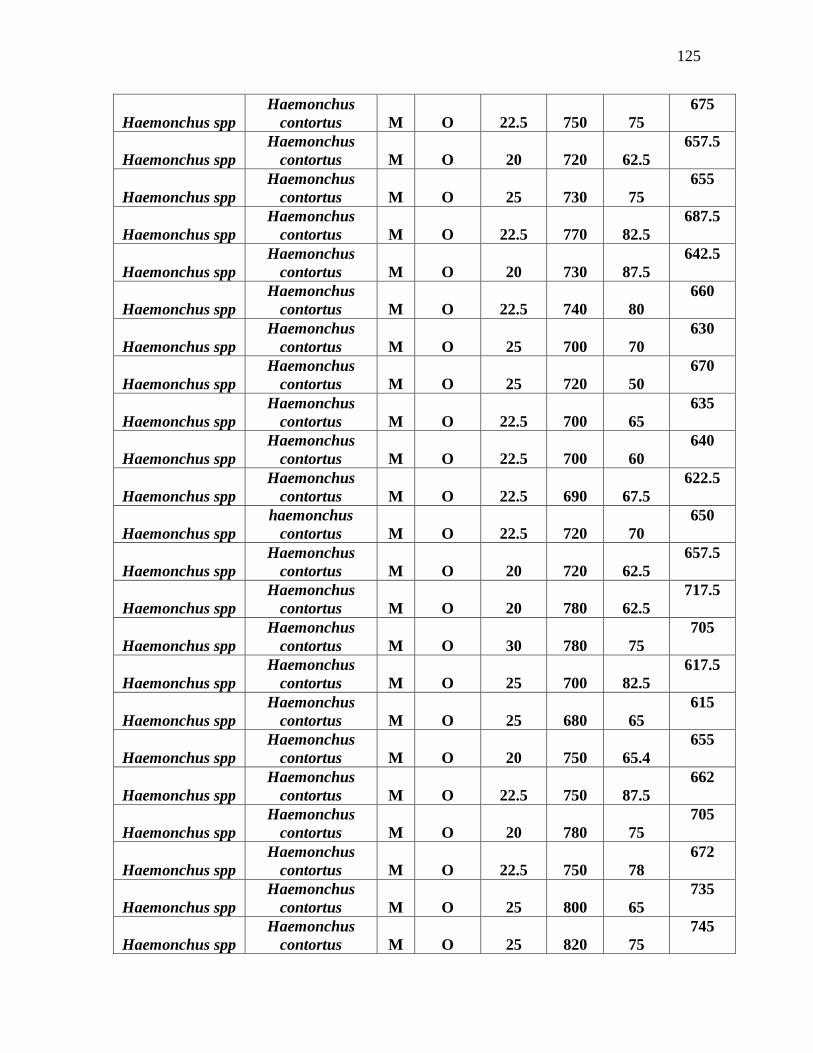
125
Haemonchus spp
Haemonchus
contortus M O 22.5 750 75
675
Haemonchus spp
Haemonchus
contortus M O 20 720 62.5
657.5
Haemonchus spp
Haemonchus
contortus M O 25 730 75
655
Haemonchus spp
Haemonchus
contortus M O 22.5 770 82.5
687.5
Haemonchus spp
Haemonchus
contortus M O 20 730 87.5
642.5
Haemonchus spp
Haemonchus
contortus M O 22.5 740 80
660
Haemonchus spp
Haemonchus
contortus M O 25 700 70
630
Haemonchus spp
Haemonchus
contortus M O 25 720 50
670
Haemonchus spp
Haemonchus
contortus M O 22.5 700 65
635
Haemonchus spp
Haemonchus
contortus M O 22.5 700 60
640
Haemonchus spp
Haemonchus
contortus M O 22.5 690 67.5
622.5
Haemonchus spp
haemonchus
contortus M O 22.5 720 70
650
Haemonchus spp
Haemonchus
contortus M O 20 720 62.5
657.5
Haemonchus spp
Haemonchus
contortus M O 20 780 62.5
717.5
Haemonchus spp
Haemonchus
contortus M O 30 780 75
705
Haemonchus spp
Haemonchus
contortus M O 25 700 82.5
617.5
Haemonchus spp
Haemonchus
contortus M O 25 680 65
615
Haemonchus spp
Haemonchus
contortus M O 20 750 65.4
655
Haemonchus spp
Haemonchus
contortus M O 22.5 750 87.5
662
Haemonchus spp
Haemonchus
contortus M O 20 780 75
705
Haemonchus spp
Haemonchus
contortus M O 22.5 750 78
672
Haemonchus spp
Haemonchus
contortus M O 25 800 65
735
Haemonchus spp
Haemonchus
contortus M O 25 820 75
745
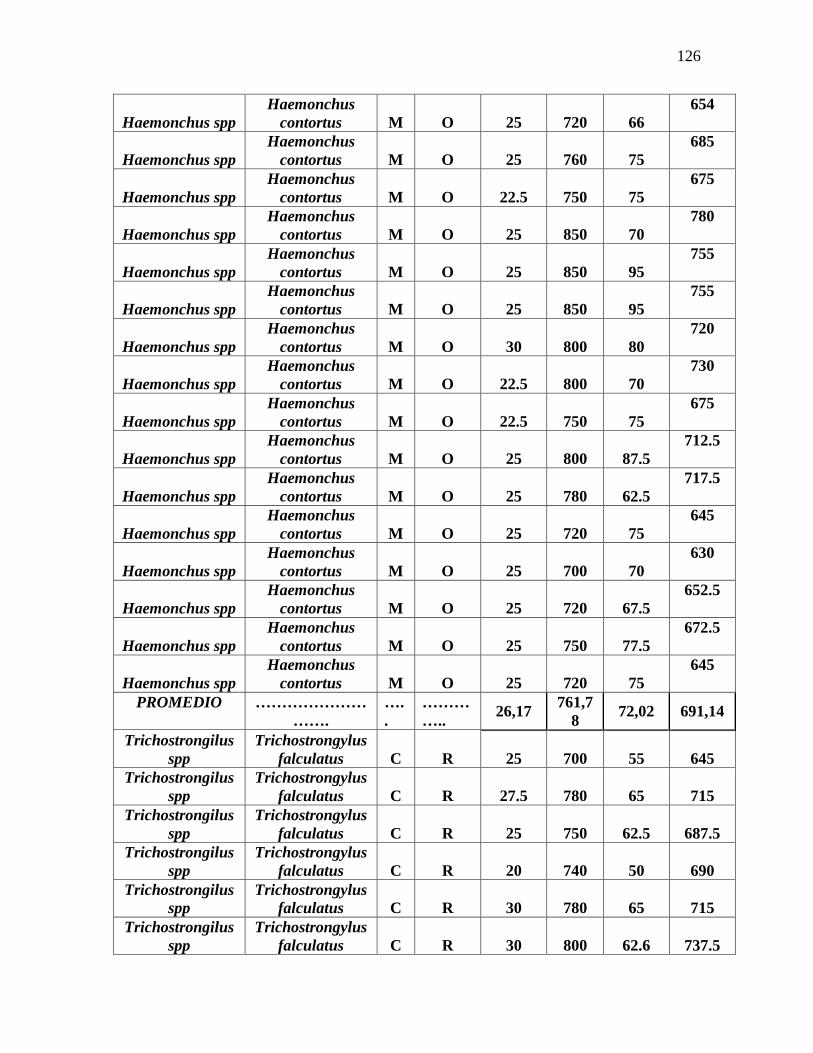
126
Haemonchus spp
Haemonchus
contortus M O 25 720 66
654
Haemonchus spp
Haemonchus
contortus M O 25 760 75
685
Haemonchus spp
Haemonchus
contortus M O 22.5 750 75
675
Haemonchus spp
Haemonchus
contortus M O 25 850 70
780
Haemonchus spp
Haemonchus
contortus M O 25 850 95
755
Haemonchus spp
Haemonchus
contortus M O 25 850 95
755
Haemonchus spp
Haemonchus
contortus M O 30 800 80
720
Haemonchus spp
Haemonchus
contortus M O 22.5 800 70
730
Haemonchus spp
Haemonchus
contortus M O 22.5 750 75
675
Haemonchus spp
Haemonchus
contortus M O 25 800 87.5
712.5
Haemonchus spp
Haemonchus
contortus M O 25 780 62.5
717.5
Haemonchus spp
Haemonchus
contortus M O 25 720 75
645
Haemonchus spp
Haemonchus
contortus M O 25 700 70
630
Haemonchus spp
Haemonchus
contortus M O 25 720 67.5
652.5
Haemonchus spp
Haemonchus
contortus M O 25 750 77.5
672.5
Haemonchus spp
Haemonchus
contortus M O 25 720 75
645
PROMEDIO …………………
…….
….
.
………
….. 26,17
761,7
8 72,02 691,14
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 700 55
645
Trichostrongilus
spp
Trichostrongylus
falculatus C R 27.5 780 65
715
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 750 62.5
687.5
Trichostrongilus
spp
Trichostrongylus
falculatus C R 20 740 50
690
Trichostrongilus
spp
Trichostrongylus
falculatus C R 30 780 65
715
Trichostrongilus
spp
Trichostrongylus
falculatus C R 30 800 62.6
737.5
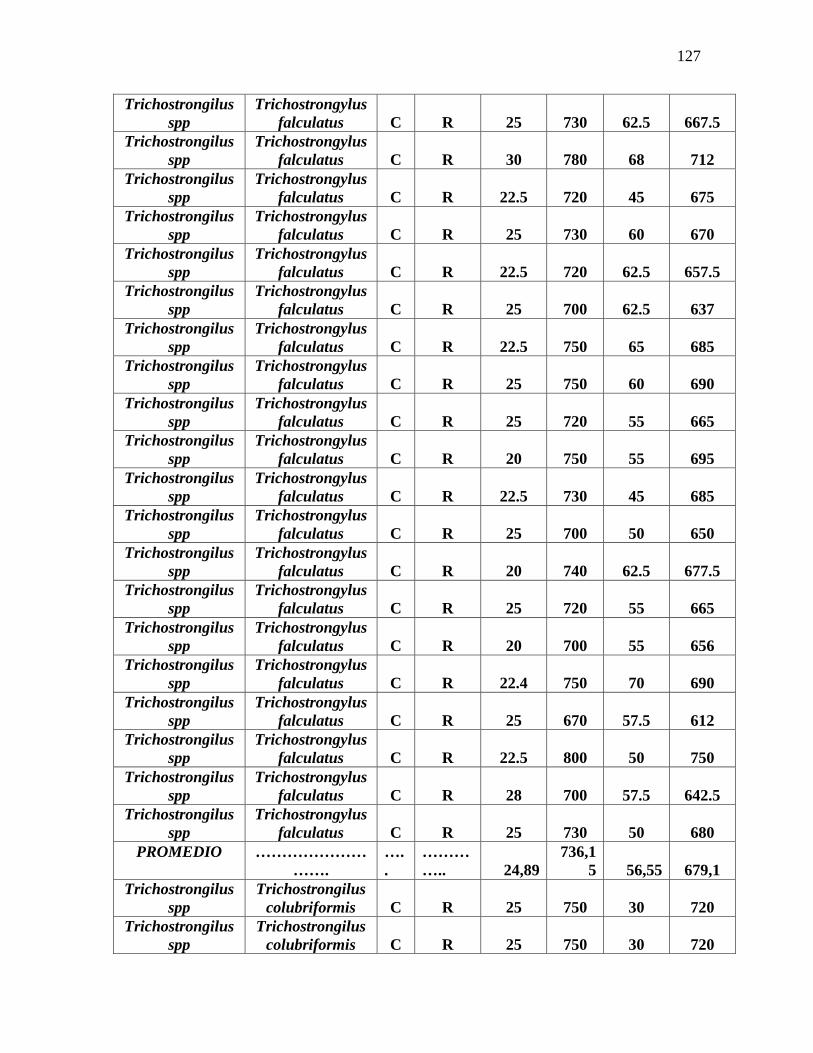
127
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 730 62.5
667.5
Trichostrongilus
spp
Trichostrongylus
falculatus C R 30 780 68
712
Trichostrongilus
spp
Trichostrongylus
falculatus C R 22.5 720 45
675
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 730 60
670
Trichostrongilus
spp
Trichostrongylus
falculatus C R 22.5 720 62.5
657.5
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 700 62.5
637
Trichostrongilus
spp
Trichostrongylus
falculatus C R 22.5 750 65
685
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 750 60
690
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 720 55
665
Trichostrongilus
spp
Trichostrongylus
falculatus C R 20 750 55
695
Trichostrongilus
spp
Trichostrongylus
falculatus C R 22.5 730 45
685
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 700 50
650
Trichostrongilus
spp
Trichostrongylus
falculatus C R 20 740 62.5
677.5
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 720 55
665
Trichostrongilus
spp
Trichostrongylus
falculatus C R 20 700 55
656
Trichostrongilus
spp
Trichostrongylus
falculatus C R 22.4 750 70
690
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 670 57.5
612
Trichostrongilus
spp
Trichostrongylus
falculatus C R 22.5 800 50
750
Trichostrongilus
spp
Trichostrongylus
falculatus C R 28 700 57.5
642.5
Trichostrongilus
spp
Trichostrongylus
falculatus C R 25 730 50
680
PROMEDIO …………………
…….
….
.
………
….. 24,89
736,1
5 56,55 679,1
Trichostrongilus
spp
Trichostrongilus
colubriformis C R 25 750 30
720
Trichostrongilus
spp
Trichostrongilus
colubriformis C R 25 750 30
720
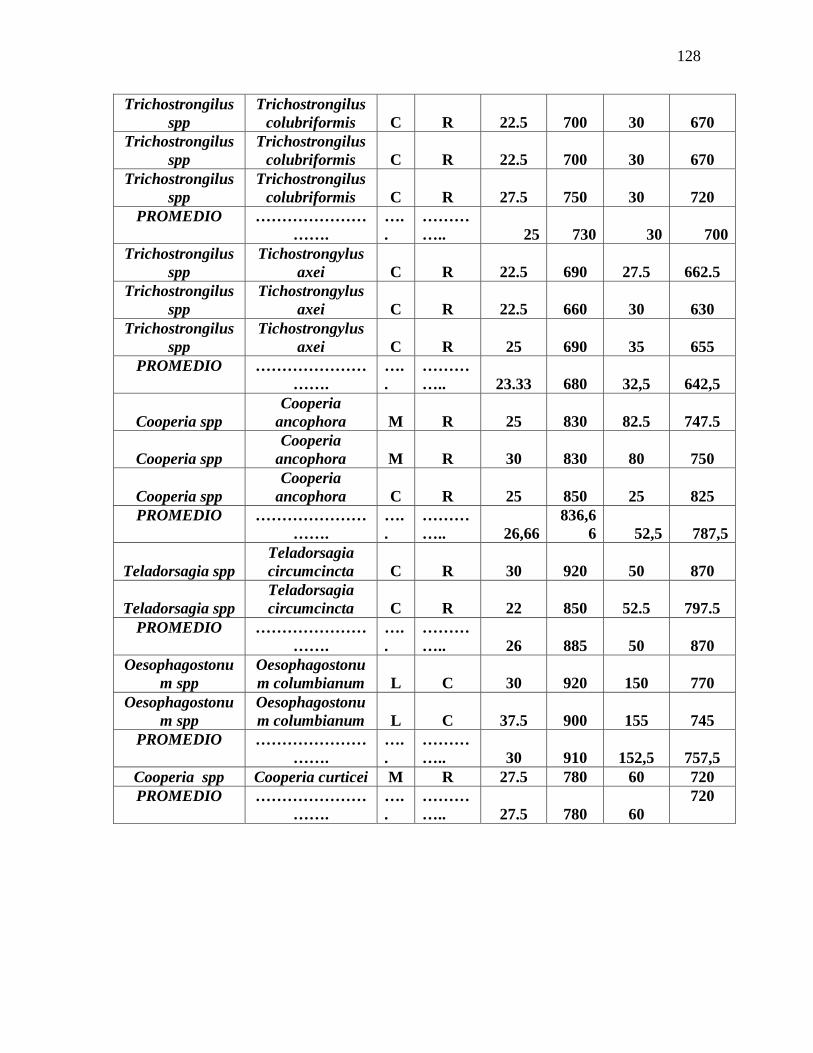
128
Trichostrongilus
spp
Trichostrongilus
colubriformis C R 22.5 700 30
670
Trichostrongilus
spp
Trichostrongilus
colubriformis C R 22.5 700 30
670
Trichostrongilus
spp
Trichostrongilus
colubriformis C R 27.5 750 30
720
PROMEDIO …………………
…….
….
.
………
….. 25 730 30 700
Trichostrongilus
spp
Tichostrongylus
axei C R 22.5 690 27.5
662.5
Trichostrongilus
spp
Tichostrongylus
axei C R 22.5 660 30
630
Trichostrongilus
spp
Tichostrongylus
axei C R 25 690 35
655
PROMEDIO …………………
…….
….
.
………
….. 23.33 680 32,5 642,5
Cooperia spp
Cooperia
ancophora M R 25 830 82.5
747.5
Cooperia spp
Cooperia
ancophora M R 30 830 80
750
Cooperia spp
Cooperia
ancophora C R 25 850 25
825
PROMEDIO …………………
…….
….
.
………
….. 26,66
836,6
6 52,5 787,5
Teladorsagia spp
Teladorsagia
circumcincta C R 30 920 50
870
Teladorsagia spp
Teladorsagia
circumcincta C R 22 850 52.5
797.5
PROMEDIO …………………
…….
….
.
………
….. 26 885 50 870
Oesophagostonu
m spp
Oesophagostonu
m columbianum L C 30 920 150
770
Oesophagostonu
m spp
Oesophagostonu
m columbianum L C 37.5 900 155
745
PROMEDIO …………………
…….
….
.
………
….. 30 910 152,5 757,5
Cooperia spp Cooperia curticei M R 27.5 780 60 720
PROMEDIO …………………
…….
….
.
………
….. 27.5 780 60
720

129
Anexo 8. Análisis Logit Probit de la prueba de inhibición de la eclosión Leucaena leucocephala

130
Anexo 9. Efecto de las diferentes concentraciones del extracto de Leucaena leucocephala en la prueba in vitro de inhibición de la eclosión de huevos de nematodos gastrointestinales de ovinos.

131
Anexo 10. Análisis Logit Probit de la prueba de inhibición de la eclosión Calliandra calothyrsus
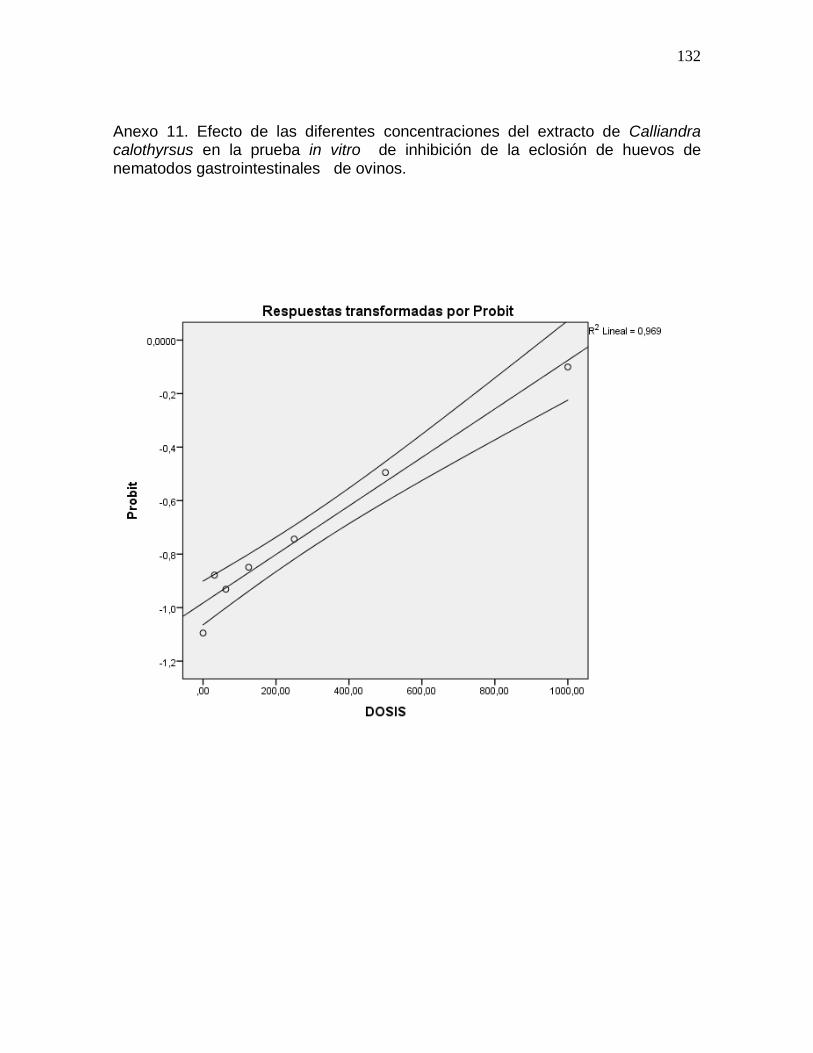
132
Anexo 11. Efecto de las diferentes concentraciones del extracto de Calliandra calothyrsus en la prueba in vitro de inhibición de la eclosión de huevos de nematodos gastrointestinales de ovinos.

133
Anexo 12. Análisis Logit Probit de la prueba de inhibición de la eclosión Flemingia macrophylla.
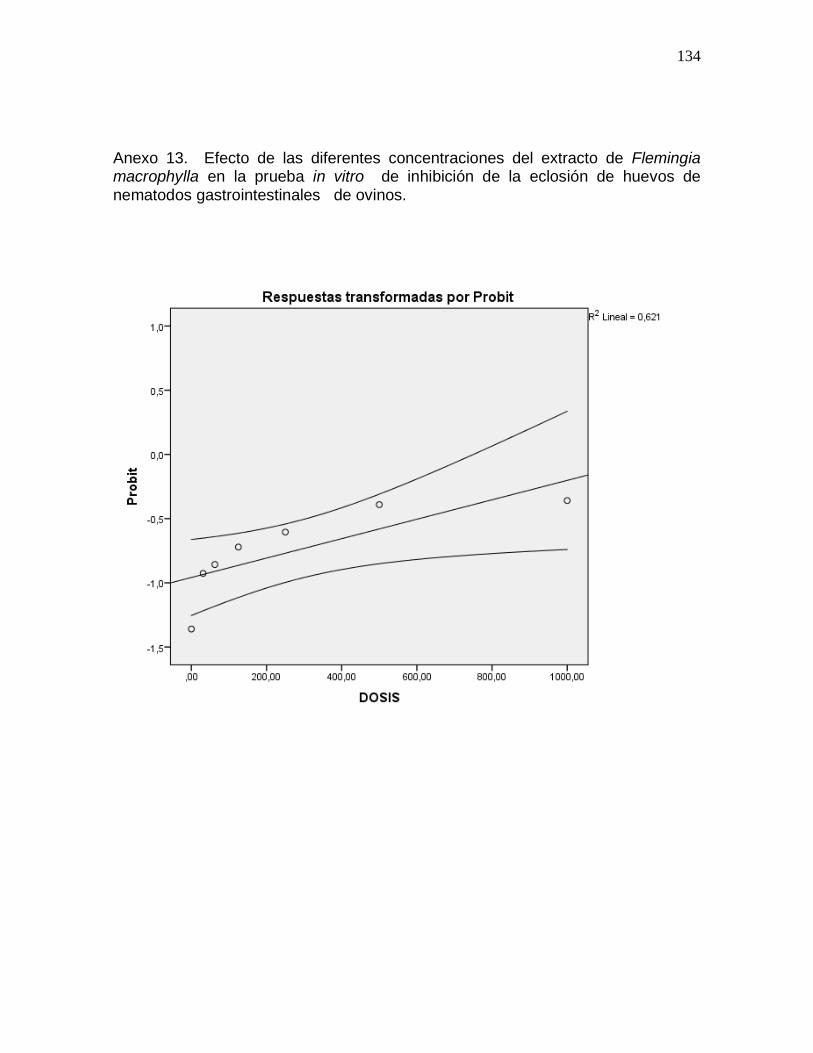
134
Anexo 13. Efecto de las diferentes concentraciones del extracto de Flemingia macrophylla en la prueba in vitro de inhibición de la eclosión de huevos de nematodos gastrointestinales de ovinos.

135
Anexo 14. Análisis Logit Probit de la prueba de eliminación de la cutícula Leucaena leucocephala
136
Anexo 15. Efecto de las diferentes concentraciones del extracto de Leucaena leucocephala en la prueba in vitro de inhibición de eliminación de la cutícula de larvas L3 de nematodos gastrointestinales de ovinos.

137
Anexo 16. Análisis Logit Probit de la prueba de eliminación de la cutícula Calliandra calothyrsus
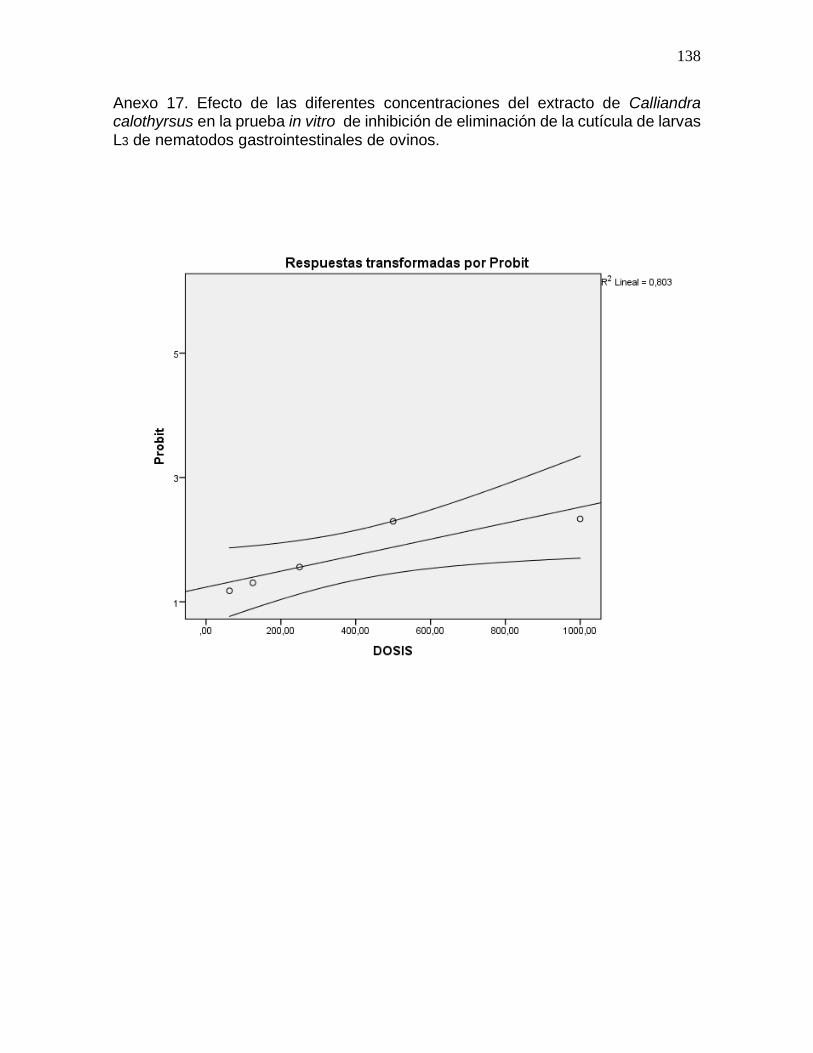
138
Anexo 17. Efecto de las diferentes concentraciones del extracto de Calliandra calothyrsus en la prueba in vitro de inhibición de eliminación de la cutícula de larvas L3 de nematodos gastrointestinales de ovinos.

139
Anexo 18. Análisis Logit Probit de la prueba de eliminación de la cutícula Flemingia macrophylla
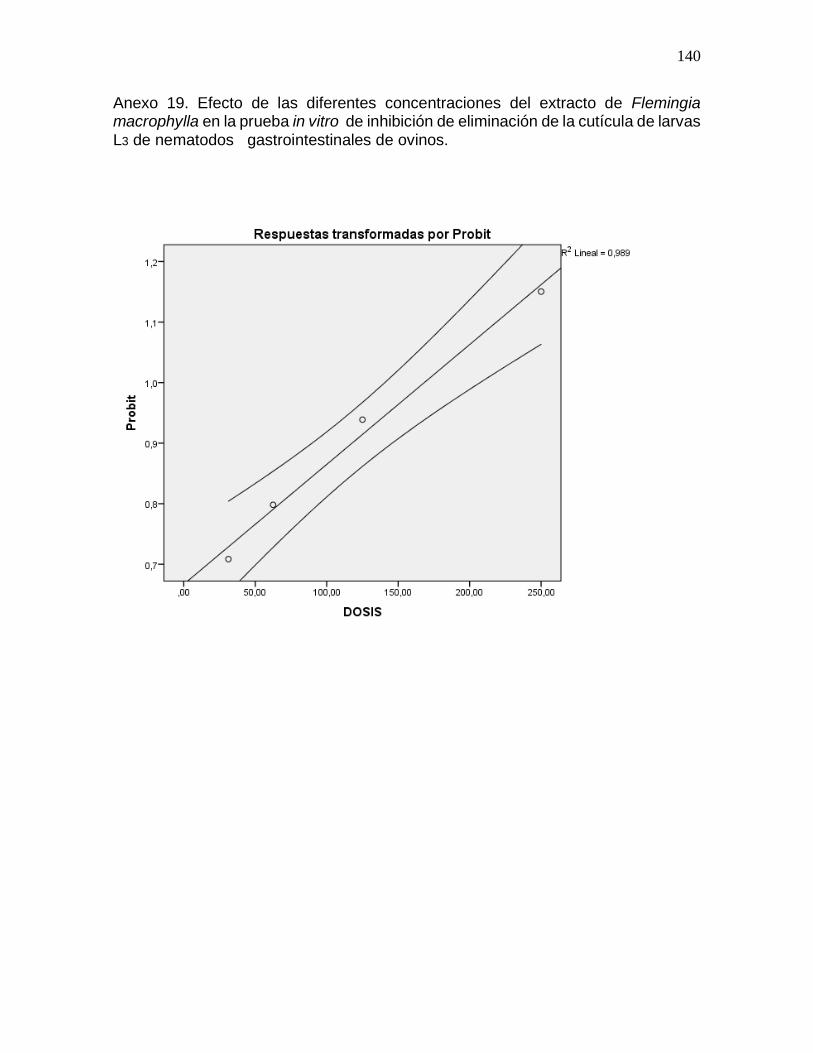
140
Anexo 19. Efecto de las diferentes concentraciones del extracto de Flemingia macrophylla en la prueba in vitro de inhibición de eliminación de la cutícula de larvas L3 de nematodos gastrointestinales de ovinos.





























