Embed Size (px)
Citation preview

General and Comparative Endocrinology 158 (2008) 178–182
Contents lists available at ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier.com/ locate /ygcen
Effects of Castration castration and Androgen-Treatment androgen-treatment on the Expression expression of FSH- b FSH-b
and LH- b LH-b in the Threespine three-spine stickleback, Gasterosteus aculeatus – Feedback Dif ferences Mediating gasterosteus aculeatus—Feedback
differences mediating the Photoperiodic Maturation Response?photoperiodic maturation response?q
Anna Hellqvist a,*, Monika Schmitz b, Bertil Borg a
a Department of Zoology, Stockholm University, S-106 91 Stockholm, Swedenb Department of Biology, Karlstad University, S-65188 Karlstad, Sweden
a r t i c l e i n f o
Article history:
Received 3 March 2008
Revised 7 July 2008
Accepted 9 July 2008
Available online 18 July 2008
Keywords:
Stickleback
Castration
Androgen treatmentAndrogen-treatment
FSH
LH
mRNA expression
Photoperiod
0016-6480/$ - see front matter © 2008 Elsevier Inc. A
doi:10.1016/j.ygcen.2008.07.004
q Supported by the Swedish Natural Science Researc
Swedish Forestry and Agriculture Research Council (to
mous reviewers for valuable comments.
* Corresponding author. Fax: +46 08 164095.
E-mail address: [email protected] (A. Hell
a b s t r a c t
In many animals, including the threespine three-spine stickleback (Gasterosteus aculeatus), photoperiod strongly
influences reproduction. The aim of this study was to investigate if feedback mechanisms on the brain-pituitary-gonadal brain–
pituitary–gonadal axis play a role in mediating the photoperiodic response in the stickleback. To that
end, stickleback males, exposed to either non-stimulatory short photoperiod (Light:Dark (light/dark 8:16) or under
stimulatory long photoperiod (LD 16:8), were subjected to either sham-operation, castration, castration
combined with treatment with the androgens 11-ketoandrostenedione (11KA) and testosterone (T), and
the effects on levels of luteinizing hormone (LH) and follicle-stimulating hormone (FSH) b -mRNA (FSH)-b mRNA were
analyzed. During breeding season the kidney of the stickleback male hypertrophies and produces a glue
used for building nests. Kidney weight and expression of both LH- b LH-b and FSH- b FSH-b were higher in sham-
operated fish kept under long than under short photoperiod. Under both photoperiods, LH- b LH-b mRNA
levels were lower in castrated males compared to sham-operated males and treatment with 11KA and T
increased expression, indicating a positive feedback. A positive feedback was also found on FSH- b FSH-b expres-
sion under long photoperiod, where castration decreased, and androgen replacement restored FSH- b FSH-b
mRNA expression. On the contrary, castration under short photoperiod instead increased FSH- b FSH-b levels
whereas treatment with 11KA and T decreased FSH- b FSH-b expression, indicating a negative feedback on FSH- b FSH-
b under these conditions. The positive feedback on FSH- b FSH-b expression under stimulatory photoperiod
may accelerate maturation, whereas the negative feedback under inhibitory photoperiod may suppress
maturation. This could be part of the mechanisms by which photoperiod controls maturation.
© 2008 Elsevier Inc. All rights reserved.
This has been observed in several studies on birds, e.g. in Japanese
quail, Coturnix japonica, where both plasma LH and FSH levels were
higher in castrated birds kept under stimulatory long photoperi-
ods than under shorter photoperiods (Urbanski and Follett, 1982).
These two ways in which photoperiods may control the BPG axis BPG-axis
are not mutually exclusive, both mechanisms were found to be
present in tree sparrows, Spizella arborea (Wilson, 1985).
Feedback effects on the BPG-axis play an important role in con-
trolling reproduction in teleost fishes. Both positive and negative
feedback effects on the BPG-axis have been reported (Crim and
Evans, 1979; Billard et al., 1977). Aromatization, the conversion of
androgens to estrogens, has been found to be involved in both the
positive and negative feedback effects in fishes (Crim et al., 1981;
de Leeuw et al., 1986). It is, however, not known whether changes
1. Introduction
The photoperiod is an important factor in the control of sea-
sonal reproduction. Under stimulatory photoperiods, an increased
secretion of gonadotropins (GTHs) from the pituitary activate ver-
tebrate gonads. Previous studies have shown that photoperiod can
alter the sensitivity of steroid feedback on the secretion of GTHs
in the brain-pituitary-gonadal brain–pituitary–gonadal (BPG) axis in mammals (Turek,
1977; Rosa and Bryant, 2003). For instance, higher doses of testos-
terone (T) were needed to suppress LH and FSH plasma levels in
castrated golden hamsters, Mesocricetus auratus, under a stimula-
tory long photoperiod than under a non-stimulatory short photo-
period (Turek, 1977). However, GTH secretion patterns may also
change independently of changes in feedbacks on the BPG-axis.
ll rights reserved.
in feedback mechanisms play a role in the photoperiodic control of
reproduction in fish.
The threespine three-spine stickleback (Gasterosteus aculeatus) has a
marked seasonal reproductive cycle in which spermatogenesis
and the development of androgen-dependent secondary sexual
characters in the male are separated in time (e.g. Borg, 1982).
h Council (to BB) B.B.) and by the
MS). M.S.). We thank two anony-
qvist).

A. Hellqvist et al. / General and Comparative Endocrinology 158 (2008) 178–182 179
During the breeding season, the stickleback displays distinctive
secondary sexual characters such as the red breeding colours and
a hypertrophied kidney that secrets a glue, consisting of the andro-
gen induced protein spiggin (Jakobsson et al., 1999; Jones et al.,
2001), used for nest building. Sexual maturation in the stickleback
is strongly dependent on photoperiod. In winter, long photoperi-
ods induce maturation, while shorter photoperiods do not, even if
temperatures are high (e.g. Baggerman, 1957; Borg et al., 1987).
The aim of the present study was to investigate whether feed-
back mechanisms play a role in mediating the photoperiodic
response in the stickleback. To that end, males exposed to either
short or long photoperiod were subjected to either sham-operation
castration, castration combined with androgen replacement with
the non-aromatisable androgen 11-ketoandrostenedione (11KA)
or the aromatisable androgen testosterone (T), and the effects on
expression of LH- b LH-b and FSH- b FSH-b mRNA were analyzed.
2. Material & MethodsMaterials and methods
2.1. The animals and experimental protocol
The study was performed with the permission of the Stockholm
Northern Animal Experiment Ethical Board (no. (No. N184/00). Adult
non-breeding sticklebacks were caught in the Öresund on the 15-16 15–
16 December 2001. The fish were kept at 7 °C and under short day
photoperiod (LD 8:16) until the onset of the experiment.
The castration/implant experiment was started on the 4-6 4–6 Janu-
ary 2002. The fish were anaesthetized with 0.1% 2-phenoxyethanol,
c. ca. 1.5 mm long incisions were made into the abdominal cavity on
each side and the testes were excised using fine forceps. This type
of surgery drastically reduces circulating levels of T and 11-ketotes-
tosterone (11KT) in breeding sticklebacks (Mayer et al., 1990).
Sham-operated fish were treated similarly except that the testes
were mot removed. The castrated control and sham-operated fish
were implanted with empty Silastic capsules (length 5 mm, 1.2 mm
outer diameter) sealed with silicone glue. The other two castrated
groups were implanted with capsules, described as above, contain-
ing either crystalline T (KEBO) or 11KA (Sigma), which is converted
to 11KT in blood cells of sticklebacks (Mayer et al., 1990). The fish
were divided between two 700 l aquaria kept under long photope-
riod (16:8 Light:Dark) light/dark) or under short photoperiod (8:16 LD). In the
castration/replacement experiment, 16 sham-operated, 18 control
castrated, 17 11KA treated and 17 T treated fish were placed under
each regime.
Temperature was raised gradually to 20 °C over the first week to
allow adaptation. All aquaria contained brackish water (0.5% salin-
ity), which was constantly filtered and aerated, and the bottoms
were covered with sand. The fish were fed daily with frozen red
midge larvae. The fish in the castration/replacement experiment
were sampled on the 14-15 14–15 February 2002. The fish were anaesthe-
tized and weighed. Pituitaries were collected and frozen in liquid
nitrogen, and kidneys were excised and weighed (± 1(±1 mg). Visual
inspection under a dissecting microscope confirmed that the pres-
ence of capsules and the completeness of castration (absence of
testes remnants) in all fish.
2.2. Dot-blot analysis of LH¡ b LH-b and FSH- b FSH-b mRNA expression
Total RNA was extracted from single pituitaries using Trizol
reagent (Invitrogen) following the manufacture’s instructions, and
was then denatured and blotted on Hybond-N+, Nylon membrane
(Amersham Pharmacia Biotech) through a Hybri-blot manifold
(Life Technologies). The membranes were allowed to dry and were
then UV- crosslinked.UV-crosslinked.
The membranes were hybridized using a cRNA probe. The ca.
300 bp long antisense probes, based on stickleback LH- b LH-b and FSH- b FSH-
b (Hellqvist et al., 2004, accession nr. Accession Nos. AJ534969 and AJ534871,
respectively) were synthesized by in vitro transcription using SP6
or T7 RNA polymerase, respectively, and labelled with (a-32P) UTP
according to the Strip-EZ
TM Strip-EZ™ RNA kit (Ambion). The blots were
prehybridized for 1hour h at 65 °C, with ULTRAhyb hybridization solu-
tion (Ambion) and then incubated over night with the radioactive
probe (1x10(1 £ 10 6 cpm/ml hybridisation solution). The blots were
washed twice in 1 x 1£ SSC, 0.1% SDS at 65 °C for 15minutes min each and
then once for 15 min at 65 °C and once for 30 min at 68 °C in 0.1 x 0.1£
SSC, 0.1% SDS. The hybridization signals of the blots were counted
and analysed analyzed using a biophosphoimager (BioRad). The membranes
were stripped according to the Strip-EZ
TM Strip-EZ™ protocols and subse-
quently hybridized with a 32P-labeled stickleback 18S rRNA probe
to correct for unequal RNA loadings.
2.3. Statistics
The data were analyzed with STATISTICA ’99 edition (StatSoft
Inc.), using two-way ANOVA for multiple comparisons and Tukey
honest significant difference (post hoc) test for comparisons
between pair of groups. Prior to analysis, the data were log trans-
formed.
3. Results
3.1. Kidney somatic index
Sham-operated males displayed significantly higher KSI under
long photoperiod than under short photoperiod (p(p < 0.001, Table
1). Under long photoperiod, castrated males had significantly lower
kidney somatic index (KSI) compared to sham-operated males
(p(p < 0.001), whereas under short photoperiod, where KSI was low
in both sham-operated and castrated males, there was no signifi-
cant difference in KSI. Treatment with 11KA significantly increased
KSI in castrated males under both photoperiods (p(p < 0.001 in each
comparison), whereas treatment with T did not. There was no
effect of photoperiod on KSI in the groups treated with 11KA, T or
in castrated controls.
3.2. FSH- b FSH-b and LH- b LH-b mRNA expression
In both experiments, expressions of both FSH- b FSH-b and LH- b LH-b in
sham-operated males were significantly higher under long photo-
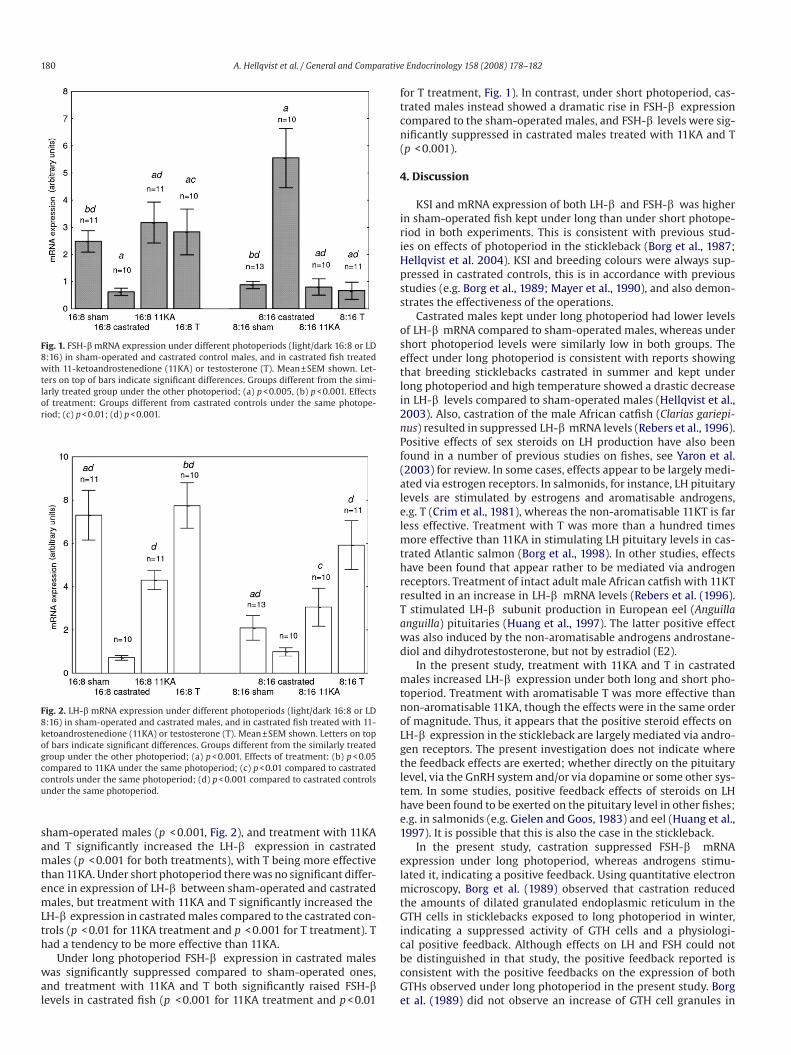
period than under short photoperiod (P(p < 0.001, Figs. 1 and 2).
There was no difference in LH- b LH-b levels between the two pho-
toperiods in castrated control males and in castrated males treated
with either 11KA or T (Fig. 2). Under long photoperiod, LH- b LH-b mRNA
levels were significantly reduced in castrated males compared to
Table 1
Kidneysomatic indices (KSI) in sham-operated males, castrated controls and
castrated males treated with 11-ketoandrostenedion (11KA) or testosterone (T)
implants and kept under different photoperiods (16 h light/8 h dark, 8 h light/16 h
dark)*
Treatment KSI
16:8 sham 2.67 ± 0.17b
16:8 castrated 0.78 ± 0.06
16:8 11KA 2.72 ± 0.25b
16:8 T 1.21 ± 0.15
8:16 sham 0.94 ± 0.17a
8:16 castrated 0.74 ± 0.09
8:16 11KA 2.65 ± 0.18b
8:16 T 1.17 ± 0.19
* Values are mean ± SEM, n = 10–13 fish per group. a p < 0.001 compared to sham-operated fish under LD 16:8. b p < 0.001 compared to castrated to castrated controls under the same photope-
riod.
180 A. Hellqvist et al. / General and Comparative Endocrinology 158 (2008) 178–182
sham-operated males (p(p < 0.001, Fig. 2), and treatment with 11KA
and T significantly increased the LH- b LH-b expression in castrated
males (P(p < 0.001 for both treatments), with T being more effective
than 11KA. Under short photoperiod there was no significant differ-
ence in expression of LH- b LH-b between sham-operated and castrated
males, but treatment with 11KA and T significantly increased the LH- b
LH-b expression in castrated males compared to the castrated con-
trols (P(p < 0.01 for 11KA treatment and Pp < 0.001 for T treatment). T
had a tendency to be more effective than 11KA.
Under long photoperiod FSH- b FSH-b expression in castrated males
was significantly suppressed compared to sham-operated ones,
and treatment with 11KA and T both significantly raised FSH- b FSH-b
levels in castrated fish (P(p < 0.001 for 11KA treatment and p < 0.01
for T treatment, Fig. 1). In contrast, under short photoperiod, cas-
trated males instead showed a dramatic rise in FSH- b FSH-b expression
compared to the sham-operated males, and FSH- b FSH-b levels were sig-
nificantly suppressed in castrated males treated with 11KA and T
(P(p < 0.001).
4. Discussion
KSI and mRNA expression of both LH- b LH-b and FSH- b FSH-b was higher
in sham-operated fish kept under long than under short photope-
riod in both experiments. This is consistent with previous stud-
ies on effects of photoperiod in the stickleback (Borg et al., 1987;
Hellqvist et al. 2004). KSI and breeding colours were always sup-
pressed in castrated controls, this is in accordance with previous
studies (e.g. Borg et al., 1989; Mayer et al., 1990), and also demon-
strates the effectiveness of the operations.
Castrated males kept under long photoperiod had lower levels
of LH- b LH-b mRNA compared to sham-operated males, whereas under
short photoperiod levels were similarly low in both groups. The
effect under long photoperiod is consistent with reports showing
that breeding sticklebacks castrated in summer and kept under
long photoperiod and high temperature showed a drastic decrease
in LH- b LH-b levels compared to sham-operated males (Hellqvist et al.,
2003). Also, castration of the male African catfish (Clarias gariepi-
nus) resulted in suppressed LH- b LH-b mRNA levels (Rebers et al., 1996).
Positive effects of sex steroids on LH production have also been
found in a number of previous studies on fishes, see Yaron et al.
(2003) for review. In some cases, effects appear to be largely medi-
ated via estrogen receptors. In salmonids, for instance, LH pituitary
levels are stimulated by estrogens and aromatisable androgens,
e.g. T (Crim et al., 1981), whereas the non-aromatisable 11KT is far
less effective. Treatment with T was more than a hundred times
more effective than 11KA in stimulating LH pituitary levels in cas-
trated Atlantic salmon (Borg et al., 1998). In other studies, effects
have been found that appear rather to be mediated via androgen
receptors. Treatment of intact adult male African catfish with 11KT
resulted in an increase in LH- b LH-b mRNA levels (Rebers et al. (1996).
T stimulated LH- b LH-b subunit production in European eel (Anguilla
anguilla) pituitaries (Huang et al., 1997). The latter positive effect
was also induced by the non-aromatisable androgens androstane-
diol and dihydrotestosterone, but not by estradiol (E2).
In the present study, treatment with 11KA and T in castrated
males increased LH- b LH-b expression under both long and short pho-
toperiod. Treatment with aromatisable T was more effective than
non-aromatisable 11KA, though the effects were in the same order
of magnitude. Thus, it appears that the positive steroid effects on LH- b
LH-b expression in the stickleback are largely mediated via andro-
gen receptors. The present investigation does not indicate where
the feedback effects are exerted; whether directly on the pituitary
level, via the GnRH system and/or via dopamine or some other sys-
tem. In some studies, positive feedback effects of steroids on LH
have been found to be exerted on the pituitary level in other fishes;
e.g. in salmonids (e.g. Gielen and Goos, 1983) and eel (Huang et al.,
1997). It is possible that this is also the case in the stickleback.
In the present study, castration suppressed FSH- b FSH-b mRNA
expression under long photoperiod, whereas androgens stimu-
lated it, indicating a positive feedback. Using quantitative electron
microscopy, Borg et al. (1989) observed that castration reduced
the amounts of dilated granulated endoplasmic reticulum in the
GTH cells in sticklebacks exposed to long photoperiod in winter,
indicating a suppressed activity of GTH cells and a physiologi-
cal positive feedback. Although effects on LH and FSH could not
be distinguished in that study, the positive feedback reported is
consistent with the positive feedbacks on the expression of both
GTHs observed under long photoperiod in the present study. Borg
et al. (1989) did not observe an increase of GTH cell granules in
Fig. 1. FSH-b mRNA expression under different photoperiods (light/dark 16:8 or LD
8:16) in sham-operated and castrated control males, and in castrated fish treated
with 11-ketoandrostenedione (11KA) or testosterone (T). Mean ± SEM shown. Let-
ters on top of bars indicate significant differences. Groups different from the simi-
larly treated group under the other photoperiod; (a) p < 0.005, (b) p < 0.001. Effects
of treatment: Groups different from castrated controls under the same photope-
riod; (c) p < 0.01; (d) p < 0.001.
Fig. 2. LH-b mRNA expression under different photoperiods (light/dark 16:8 or LD
8:16) in sham-operated and castrated males, and in castrated fish treated with 11-
ketoandrostenedione (11KA) or testosterone (T). Mean ± SEM shown. Letters on top
of bars indicate significant differences. Groups different from the similarly treated
group under the other photoperiod; (a) p < 0.001. Effects of treatment: (b) p < 0.05
compared to 11KA under the same photoperiod; (c) p < 0.01 compared to castrated
controls under the same photoperiod; (d) p < 0.001 compared to castrated controls
under the same photoperiod.

A. Hellqvist et al. / General and Comparative Endocrinology 158 (2008) 178–182 181
sham-operated fish, but instead a decrease compared to castrated
fish. This indicates that the increased amounts of GTHs that the
former produced were released, rather than stored. Unfortunately,
it is at present not possible to measure circulating levels of LH and
FSH in the stickleback.
Contrarily, castrated males under short photoperiod showed
a dramatic rise in FSH- b FSH-b expression compared to the sham-oper-
ated males, indicating a negative feedback instead under this con-
dition. This is also consistent with androgens suppressing FSH- b FSH-b
mRNA expression under the short photoperiod. It cannot be said
at which level(s) the feedback effects on FSH- b FSH-b are exerted. The
complexity of the response could be consistent with more than
one level being involved.
Castration of sticklebacks kept under long photoperiod and
high temperature in summer had no effect on FSH- b FSH-b expression
(Hellqvist et al., 2003). There are few other studies on the effects of
castration on expression of FSH- b FSH-b mRNA in fishes. In coho salmon
(Oncorhynchus kisutch), castration resulted in elevated plasma
levels of FSH (Larsen and Swanson, 1997), and castration of Atlan-
tic salmon could have both positive or negative effects on plasma
and pituitary FSH levels depending on season (Borg et al., 1998;
Antonopoulou et al., 1999a, b). In the present study, FSH- b FSH-b
mRNA levels under long photoperiod were restored with 11KA
and T treatment which increased the expression in castrated
fish. In contrast, under short photoperiod, FSH- b FSH-b levels were
suppressed in castrated males treated with 11KA and T. Since
both the non-aromatisable 11KA and the aromatisable T lowered
the FSH- b FSH-b expression under short photoperiod and increased
it under long photoperiod, both the inhibitory and the stimula-
tory effects are probably exerted via an androgen receptor. In
contrast, treatment of intact coho salmon with the aromatisable
androgen T or E resulted in a reduction of transcript levels of
FSH- b FSH-b (Dickey and Swanson, 1998) indicating an effect via an
estrogen receptor. In Atlantic salmon, T had both negative and
positive effects on pituitary FSH in salmon, depending on dose
and season (Antonopoulou et al., 1999b). Atlantic salmon treated
with 11KA showed increased plasma and pituitary FSH levels
(Borg et al., 1998). Unlike the feedback control of LH which is
relatively consistent, the feedback control of FSH is complex and
changes markedly between seasons/photoperiods in both the
stickleback and salmonids. There are very few previous studies
on feedback effects under different photoperiods in fishes. Early
maturing male masu salmon, Oncorhynchus masou, reached full
maturity (spermiation) earlier under LD 8:16 than under LD 16:8
in autumn (Amano et al. 2001). Castration had similar effects on
LH (GTH II) pituitary content under both photoperiods; levels
were much lower than in controls. Castration increased pituitary
FSH (GTH I) content under the non-stimulatory long, but not
under stimulatory short photoperiod after 30 days (Amano et al.
2001). It is difficult to interpret what role, if any, this could have
in the control of maturation as there was no significant effect
of photoperiod on FSH pituitary levels in controls. T implants
increased LH pituitary levels similarly in immature salmon kept
under LD 8:16 and 16:8, but had no effect on FSH levels (Amano
et al. 2001).
In the present study, there are no indications for a feedback-
independent increase of GTH secretion by the stimulatory
photoperiods as found in birds (Urbanski & Follett). The expres-
sion of LH- b LH-b was not influenced by photoperiod in the castrated
groups, whereas the expression of FSH- b FSH-b in castrated controls was
much higher under LD 8:16 than under LD 16:8, which is the oppo-
site to what one should expect with a feedback -independent feedback-independent stim-
ulation. The possible biological significance of this is unknown.
Feedback effects on LH- b LH-b were similar under both photoperiods.
A positive feedback was also found on FSH- b FSH-b mRNA expression
under long photoperiod, where castration drastically decreased,
and androgen replacement restored, expression. In contrast, cas-
tration under short photoperiod increased FSH- b FSH-b transcript levels
drastically and treatment with 11KA and T decreased FSH- b FSH-b expres-
sion, revealing a negative feedback on FSH- b. FSH-b. Thus, the feedback
effects on FSH are very different under long and short photoperiod.
The shift between negative and positive feedback under different
photoperiods is not found in mammals, where there are instead
changes in effectiveness of negative feedbacks (Turek, 1977; Rosa
and Bryant, 2003).
FSH levels peak earlier in the season than LH levels in most
studied fishes (Suzuki et al. 1988). Also in the stickleback FSH- b FSH-b
expression reaches a peak earlier in the season than LH- b LH-b and FSH
is therefore more likely than LH to control the onset of maturation
(Hellqvist et al., 2006). The positive feedback under long photope-
riod is likely to accelerate maturation whereas the negative feed-
back under short photoperiod may suppress maturation, which
could be part of the mechanisms by which photoperiod controls
maturation in the stickleback.
Appendix A. Supplementary data
Supplementary data associated with this article can be found,
in the online version, at doi:10.1016/j.ygcen.2008.07.004.
References
Amano, M., Ikuta, K., Kitamura, S., Aida, K., 2001. Effects of photoperiod on pituitary gonadotropin levels in masu salmon. J. Exp. Zool. 289, 449–455.
Antonopoulou, E., Bornestaf, C., Swanson, P., Borg, B., 1999a. Feedback control of gonadotropins in Atlantic salmon, Salmo salar, male parr. I. Castration effects in rematuring and nonrematuring fish. Gen. Comp. Endocrinol. 114, 132–141.
Antonopoulou, E., Swanson, P., Mayer, I., Borg, B., 1999b. Feedback control of gonado-tropins in Atlantic salmon, Salmo salar, male parr. II. Aromatase inhibitor and androgen effects. Gen. Comp. Endocrinol. 114, 142–150.
Baggerman, B., 1957. An experimental study on the timing of breeding and migra-tion in the three-spined stickleback (Gasterosteus aculeatus). Arch. Néerl. Zool. 12, 105–318.
Billard, R., Richard, M., Breton, B., 1977. Stimulation of gonadotropin secretion after castration in rainbow trout. Gen. Comp. Endocrinol. 33, 163–165.
Borg, B., 1982. Seasonal effects of photoperiod and temperature on spermatogene-sis and secondary sexual characters in the three-spined stickleback, Gasteros-teus aculeatus L.. Can. J. Zool. 60, 3377–3386.
Borg, B., Andersson, E., Mayer, I., Zandbergen, M.A., Peute, J., 1989. Effects of castra-tion on pituitary gonadotropic cells of the male three-spined stickleback, Gaster-osteus aculeatus L., under long photoperiod in winter: Indications for a positive feedback. Gen. Comp. Endocrinol. 76, 12–18.
Borg, B., Antonopoulou, E., Mayer, I., Andersson, E., Berglund, I., Swanson, P., 1998. Effects of gonadectomy and androgen treatments on pituitary and plasma lev-els of gonadotropins in Atlantic salmon, Salmo salar, mature male parr—phys-iological positive feedback control of both gonadotropins. Biol. Reprod. 58, 814–820.
Borg, B., Peute, J., Reschke, M., van den Hurk, R., 1987. Effects of photoperiod and temperature on testes, retinal epithelium, and pituitary gonadotropic cells of the three-spined stickelback, Gasterosteus aculeatus L.. Can. J. Zool. 65, 14–19.
Crim, L.W., Evans, D.M., 1979. Stimulation of pituitary gonadotropin by testoster-one in juvenile rainbow trout (Salmo gairdneri). Gen. Comp. Endocrinol. 37, 192–196.
Crim, L.W., Peter, R.E., Billard, R., 1981. Onset of gonadotropic hormone accumula-tion in the immature trout pituitary gland in response to estrogen or aromatisa-ble androgen steroid hormones. Gen. Comp. Endocrinol. 44, 374–381.
de Leeuw, R., Wurth, Y.A., Zandbergen, M.A., Peute, J., Goos, H.J.Th., 1986. The effects of aromatisable androgens, non-aromatisable androgens, and estrogens on gon-adotropin release in castrated African catfish, Clarias gariepinus (Burchell). Cell Tissue Res. 243, 587–594.
Dickey, J-T., Swanson, P., 1998. Effects of sex steroids on gonadotropin (FSH and LH) regulation in coho salmon (Oncorhynchus kisutch). J. Mol. Endocrinol. 21, 291–306.
Gielen, J.Th., Goos, H.J.Th., 1983. The brain–pituitary axis in the rainbow trout, Salmo gairdneri. II. Direct effect of gonadal steroids on the gonadotropic cells. Cell Tissue Res. 233, 377–388.
Hellqvist, A., Schmitz, M., Lindberg, C., Olsson, P-E., Borg, B., 2003. LH-b and FSH-b mRNA expression in nesting and post-breeding three-spined stickleback, Gast-erosteus aculeatus, and effects of castration on expression of LH-b, FSH-b and spiggin mRNA. Fish Physiol. Biochem. 25, 311–317.
Hellqvist, A., Bornestaf, C., Borg, B., Schmitz, M., 2004. Cloning and sequencing of the FSH-b and LH-b-subunit in the three-spined stickleback, Gasterosteus aculeatus,

182 A. Hellqvist et al. / General and Comparative Endocrinology 158 (2008) 178–182
and effects of photoperiod and temperature on LH-b and FSH-b mRNA expres-sion. Gen. Comp. Endocrinol. 135, 167–174.
Hellqvist, A., Schmitz, M., Mayer, I., Borg, B., 2006. Seasonal changes in expression of LH-b and FSH-b in male and female three-spined stickleback, Gasterosteus aculeatus. Gen. Comp. Endocrinol. 145, 263–269.
Huang, Y.S., Schmitz, M., Le Belle, N., Chang, C.F., Quérat, B., Dufour, S., 1997. Andro-gens stimulate gonadotropin-II b-subunit in eel pituitary cells in vitro. Mol. Cell. Endocrinol. 131, 157–166.
Jakobsson, S., Borg, B., Haux, C., Hyllner, S.J., 1999. An 11-ketotestosterone induced kidney-secreted protein: the nest building glue from male three-spined stickleback, Gasterosteus aculeatus. Fish Physiol. Biochem. 20, 79–85.
Jones, I., Lindberg, C., Jakobsson, S., Hellqvist, A., Hellman, U., Borg, B., Olsson, P.O., 2001. Molecular cloning and characterization of spiggin. J. Biol. Chem. 21, 17857–17863.
Larsen, D.A., Swanson, P., 1997. Effects of gonadectomy on plasma gonadotropins I and II in coho salmon, Oncorhynchus kisutch. Gen. Comp. Endocrinol. 108, 152–160.
Mayer, I., Borg, B., Schulz, R., 1990. Seasonal changes in and effects of castration/androgen replacement on the plasma levels of five androgens in the male
three-spined stickleback, Gasterosteus aculeatus L.. Gen. Comp. Endocrinol. 79, 23–30.
Rebers, F.E.M., Tensen, C.P., Schulz, R.W., Goos, H.J.T., Bogerd, J., 1996. Modulation of glycoprotein hormone a- and gonadotropin II b-subunit mRNA levels in the pituitary gland of mature male African catfish, Clarias gariepinus. Fish Physiol. Biochem. 17, 99–108.
Rosa, H.J.D., Bryant, M.J., 2003. Seasonality of reproduction in sheep. Review. Small Ruminant Res. 48, 155–171.
Suzuki, K., Kanamori, A., Nagahama, Y., Kawauchi, H., 1988. Development of salmon GTH I and GTH II radioimmunoassays. Gen. Comp. Endocrinol. 71, 459–467.
Turek, F., 1977. The interaction of photoperiod and testosterone in regulating serum gonadotropin levels in castrated male hamsters. Endocrinology 101, 1210–1215.
Urbanski, H.F., Follett, B.K., 1982. Photoperiodic modulation of gonadotrophin secre-tion in castrated Japanese quail. J. Endocrinol. 92, 73–83.
Wilson, F.E., 1985. Androgen feedback-dependent and -independent control of pho-toinduced LH secretion in male tree sparrows (Spizella arborea). J. Endocrinol. 105, 141–152.
Yaron, Z., Gur, G., Melamed, P., Rosenfield, H., Elizur, A., Levavi-Sivan, B., 2003. Regu-lation of fish gonadotropins. Int. Rev. Cytol. 225, 131–185.