Embed Size (px)
Citation preview

BRAIN RESEARCH
E L S E V I E R Brain Research 690 (1995) 225-230
Short communication
Enkephalin-immunoreactive fastigial neurons in the rat cerebellum project to upper cervical cord segments
M i c h i k o Ikeda a, T a k e s h i H o u t a n i a, H i r o n o b u N a k a g a w a a, K a z u y a s u B a b a a, Ak i to sh i K o n d o h ~,
T e i z o U e y a m a a T o m o k o Y a m a m o t o b, H i sae G e m b a b, T e t s u o S u g i m o t o a,*
Department of Anatomy, Kansai Medical University, Moriguchi, Osaka 570, Japan b Department of Physiology, Kansai Medical Universi~, Moriguchi, Osaka 570, Japan
Abstract
By using enkephalin immunohistochemistry combined with retrograde fluorescent labelling, a great majority of neurons in the rat cerebellum sending their axons to the spinal cord were shown to contain enkephalin immunoreactivity. These neurons were numerous and clustered in the fastigial nucleus but far less abundant in other cerebellar nuclei. Enkephalin-immunoreactive fibers present in the ventral horn and the central cervical nucleus of upper cervical cord segments almost completely disappeared contralaterally following kainic acid-induced cell loss in the fastigial nucleus. The results indicate that fastigial and some other cerebellar nucleus neurons provide enkephalin-containing projections toward these spinal sites.
Keywords: Enkephalin; Cerebellar nucleus; Cervical cord; Cerebellospinal projection; Rat; Immunohistochemistry; Double labelling
Previous anatomical and electrophysiological studies have disclosed direct spinal projections arising from cere- bellar nucleus neurons in a variety of animal species including rats [1,3,6,10,15,16,20-24] (for further refs., see [7,20]). The projections originate mainly from the fastigial nucleus, and additionally from some regions of the cerebel- lar nucleus [7,10,15,16,20], with the principal termination sites being located contralateral at the upper cervical spinal cord [3,6,21,22,24]. According to Wilson and colleagues [23,24], fastigiospinal neurons can be activated by a num- ber of peripheral inputs and by stimulation of the labyrinth, and the axons exert a monosynaptic excitatory action on some upper cervical motoneurons. On the other hand, authors in anatomical studies claim several termination sites for fastigio- or interposito-spinal projections such as ventral horn cells [6], interneurons in the intermediate zone [3] and both ventral horn and intermediate gray matters [21,22]. At present, we have little information of cell profiles, chemical features and the extent of terminal sites of cerebellospinal projections.
Enkephalin is well-known to be expressed by a subpop- ulation of cerebellar neurons [11,18]. We noticed its pref- erential localization to neurons in the rat fastigial nucleus.
* Corresponding author. Fax: (81) (6) 992-0609.
0006-8993/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved SSDI 0 0 0 6 - 8 9 q 3 ( 9 5 ) 0 0 5 5 7 - 9
We were also aware of considerable reductions of descend- ing enkephalinergic fibers following lesioning at supraspinal levels. In this communication we report evi- dence for enkephalin in spinal projections from the rat cerebellum.
Male Sprague-Dawley rats (b.wt. 180-250 g) were maintained on a 12 h light-dark cycle with free access to rat chow and tap water. Surgery and vascular perfusion were done under intraperitoneal pentobarbital anesthesia (40 mg/kg b.wt.). Some rats received colchicine (Sigma; 200 /zg in 20 /zl 0.9% saline) in the lateral cerebral ventricle 2 days before perfusion. Kainic acid (Sigma; 2 /xg in 1 /zl 0.1 M Na-phosphate buffer, pH 7.3) was administered stereotaxically through the glass micropipette into the left fastigial nucleus of untreated rats; the con- tralateral fastigial nucleus was given vehicle. These rats were allowed to survive for 5 days before peffusion. For a double-label study of fastigiospinal neurons, a retrograde fluorescent tracer True Blue (Sigma; a total amount of 100 /~g in 5 /xl saline) was injected through a Hamilton microsyringe into the C1-C5 spinal cord segments with multiple needle penetrations 5 days before colchicine ad- ministration. All the animals were reanesthetized and per- fused intracardially with a fixative (2 liters/kg) consisted of 4% paraformaldehyde and 0.1 M Na-phosphate (pH 7.3). Brains and spinal cords were removed, saturated in

226 M. lkeda et al. /Brain Research 690 (1995) 225-230
25% sucrose for 2 days at 4°C, frozen and cut into transverse sections of 40 /zm or 10 /xm (for double-label study) thickness. Enkephalin was immunostained as de- scribed previously [19]. For the immunostaining of True Blue-labelled neurons, tissue sections were first photo- graphed with a fluorescence microscope, then reacted for enkephalin and observed for neuronal immunolabel. All the immunostained sections were examined under bright- field illumination. Some adjacent sections were stained with 0.1% Cresyl violet to define cytoarchitectural bound- aries of the nucleus and extents of injected chemicals. The nomenclature of the cerebellar nucleus in [8,13] was used in this study.
.!
d
at
Fig. 1. Enkephalin immunostaining of the fastigial nucleus. In this colchicine-treated rat, immunolabelled neurons are distributed in the fastigial, anterior and posterior interpositus and dentate nuclei, with highest accumulations in the fastigial medial subdivision (B), fastigial dorsolateral protuberance (A) and posterior interpositus nucleus. Scale bar = 100 p.m (A,B).
rostra l dmc
d dlp L i S
Mvl
f ' ,
a , LV' , c a u d a l e L , I
Fig. 2. Distribution of enkephalin-immunoreactive cerebellar nucleus neurons. Each dot represents 5-10 neurons which are seen in transverse sections arranged from caudal (a) to rostral (h). dlh, dorsolateral hump; dip, dorsolateral protuberance of the fastigial nucleus; dmc, dorsomedial crest region; IA, anterior interpositus nucleus; IP, posterior interpositus nucleus; L, dentate nucleus; LV, lateral vestibular nucleus; M, fastigial nucleus; Mm, medial subdivision of the fastigial nucleus; Mvl, ventrolat- eral subdivision of the fastigial nucleus.
In the cerebellar nucleus, enkephalin-immunoreactive fibers were more or less distributed in each nucleus and a small number of immunolabelled neurons were scattered within each cerebellar nucleus in good agreement with previous findings [11,18]. Following colchicine treatment, numerous immunolabelled neurons and their proximal den- drites became visible (Fig. 1). These neurons were either multipolar or fusiform, and usually contained small or medium-sized cell bodies ranging between 10-20 /xm in diameter. They were distributed predominantly to the fasti- gial nucleus but also to interpositus and dentate nuclei, with highest numbers being present in the fastigial medial subdivision, dorsolateral protuberance and the caudalmost portion of the posterior interpositus nucleus (Fig. 2).
In view of preferential neuronal expression of enkephalin in the fastigial nucleus as described above, we then tested if fastigial nucleus lesions may produce spe- cific reductions of enkephalin-immunoreactive fibers in the cervical spinal cord in which fastigial projections are reported to terminate [6,21,22]. Following kainic acid ad- ministration to the left fastigial nucleus, the majority of neuronal population were damaged from its main body (Fig. 3A) sparing a small portion at the rostral pole. The overlying vermal cortices and the medial part of the inter- positus nucleus were also partially affected by the lesion.
M. lkeda et al. / Brain Research 690 (1995) 225-230 227
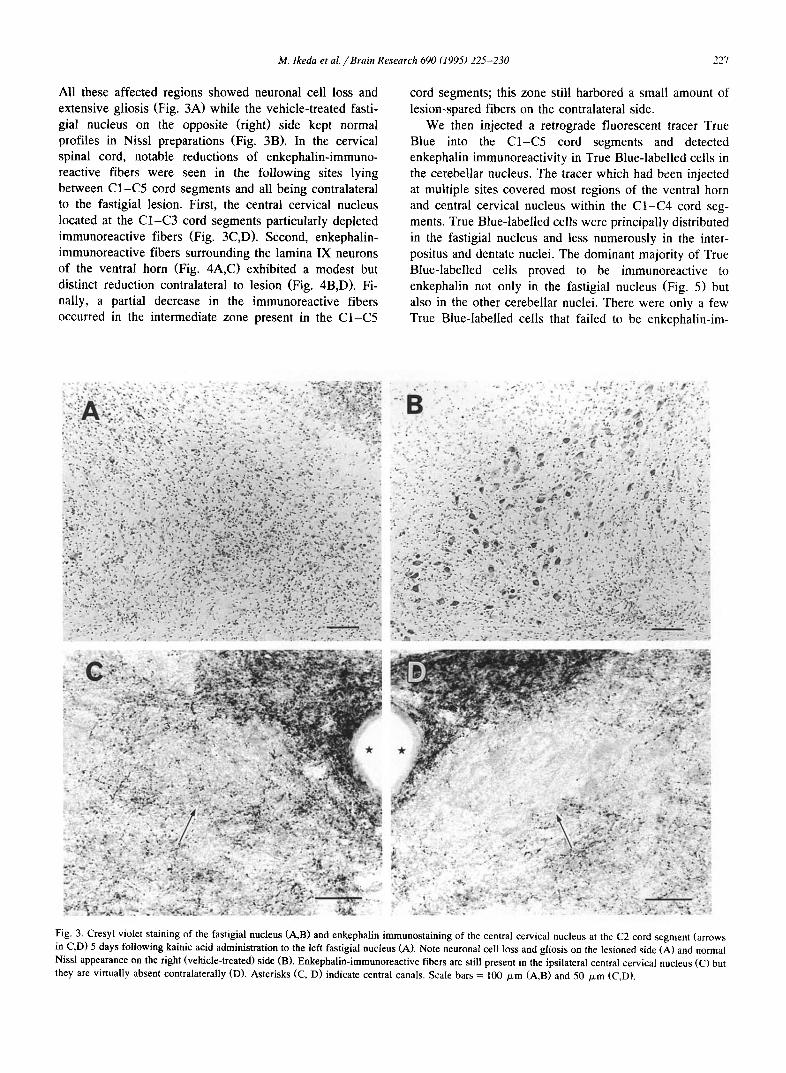
All these affected regions showed neuronal cell loss and extensive gliosis (Fig. 3A) while the vehicle-treated fasti- gial nucleus on the opposite (right) side kept normal profiles in Nissl preparations (Fig. 3B). In the cervical spinal cord, notable reductions of enkephalin-immuno- reactive fibers were seen in the following sites lying between C1-C5 cord segments and all being contralateral to the fastigial lesion• First, the central cervical nucleus located at the C1-C3 cord segments particularly depleted immunoreactive fibers (Fig. 3C,D). Second, enkephalin- immunoreactive fibers surrounding the lamina IX neurons of the ventral horn (Fig. 4A,C) exhibited a modest but distinct reduction contralateral to lesion (Fig. 4B,D). Fi- nally, a partial decrease in the immunoreactive fibers occurred in the intermediate zone present in the C1-C5
cord segments; this zone still harbored a small amount of lesion-spared fibers on the contralateral side.
We then injected a retrograde fluorescent tracer True Blue into the C1-C5 cord segments and detected enkephalin immunoreactivity in True Blue-labelled cells in the cerebellar nucleus. The tracer which had been injected at multiple sites covered most regions of the ventral horn and central cervical nucleus within the C1-C4 cord seg- ments. True Blue-labelled cells were principally distributed in the fastigial nucleus and less numerously in the inter- positus and dentate nuclei. The dominant majority of True Blue-labelled cells proved to be immunoreactive to enkephalin not only in the fastigial nucleus (Fig. 5) but also in the other cerebellar nuclei. There were only a few True Blue-labelled cells that failed to be enkephalin-im-
. " " .: . ..7 ~ ' ~ . , " " . ~.:" t . ~ " '" ' : "
• ~ " . . " : -'; ;*~t: :~, ' ; 'a ' ,~ !'. 3 . -,~, . . - ~: , ,
• - ' . . . . . ,:~c~,~'-;,q . ? ~ - , , ~ , O ~ ~ 2. ' , .",'*" ; . ' - ' . . , * ; ~ ' "~i';', : ,,~'" "
~ . '~ ~ . . ~ , , .,~ ,, ~,, :,-'~ . . . . . , . . . . • , ; 7 , ~ : ~ ,~"
Fig. 3. Cresyl violet staining of the fastigial nucleus (A,B) and enkephalin immunostaining of the central cervical nucleus at the C2 cord segment (arrows in C,D) 5 days following kainic acid administration to the left fastigial nucleus (A). Note neuronal cell loss and gliosis on the lesioned side (A) and normal Nissl appearance on the right (vehicle-treated) side (B). Enkephalin-immunoreactive fibers are still present in the ipsilateral central cervical nucleus (C) but they are virtually absent contralaterally (D). Asterisks (C, D) indicate central canals. Scale bars = 100 ~ m (A,B) and 50 /zm (C,D).

228 M. Ikeda et al. / Brain Research 690 (1995) 225-230
munopositive. Thus, a great majority of "spinal projecting neurons were indicated by this double-label study to con- tain enkephalin immunoreactivity. On the other hand, there were also a considerable number of enkephalin-immuno- reactive neurons that failed to be retrogradely labelled (for instance, see unarrowed neurons in Fig. 5B,D). Such neu- rons were found in each cerebellar nucleus, and seemed to be distributed randomly but not restricted to the regions where enkephalin-immunoreactive neurons were clustered•
In the results we have shown enkephalin immuno- reactivity contained in cerebellospinal projection neurons and distinct terminal sites. We found that double-labelled cells are predominant in the fastigial nucleus and that the majority of True Blue-labelled cells contained enkephalin immunolabel. These findings not only support results from previous tracer studies which indicate an accentuated dis- tribution pattern of cerebellospinal neurons [7,15,16,20], but also provide insights into plausible roles of the projec-
tion system implicated in opioid-mediated modulation of upper cervical spinal cord by cerebellar output neurons.
The enkephalin-immunopositive cerebellospinal neu- rons in our rats have medium-sized or small cell bodies. Judging from the cell size and the characteristic distribu- tion pattern, these neurons do not seem to be identical to previously reported GABA neurons (mostly acting as cere- bel lo-ol ivary neurons or nucleocort ical neurons) [2,4,5,9,14] or excitatory amino acid (glutamate and aspar- tate)-utilizing neurons [5,12,14,17]. It is generally agreed that most GABA neurons are small, that glutamate- and aspartate-utilizing neurons have cell bodies of variable sizes, and that all these neurons are scattered throughout most regions of the cerebellar nucleus [4,5,9,12,14,17]. Accordingly, partial coexistence may be possible between enkephalin and amino acid transmitters.
From the results of lesioning, we regarded the central cervical nucleus, the ventral horn regions and the interme-
= = : , ,
~ ~ , 4 ¸ ~ ~
Fig. 4. Enkephalin immunostaining of the lamina IX at the C2 (A,B) and C3 (C,D) cord segments, 5 days following kainic acid administration to the left fastigial nucleus (Fig. 3A,B). Enkephalin-immunoreactive fibers in the central (B) and medial (D) regions of the ventral horn are depleted contralaterally while identical regions on the ipsilateral side (A,C) harbor ample immunoreactive fibers. Scale bar = 50 ~m (A-D).

M. lkeda et al. / Brain Research 690 (1995) 225-230 229
, . . . .
i
Fig. 5. True Blue-labelled neurons in the medial subdivision of the fastigial nucleus (A) and at a further rostral level of the nucleus (C), and the corresponding areas immunostained for enkephalin (B,D). In this rat, True Blue is injected into the C1-C5 cord segments. Most neurons are double-labelled (corresponding arrows in panels A,B and C,D). Scale bar = 50 /.Lm (A-D).
diate zone as the termination sites of axons derived from the contralateral fastigial nucleus. Both ventral horn and intermediate gray matters have been listed as the terminal
sites of crossed fastigiospinal projections [6,21,22]. In these spinal regions, enkephalin may act directly on mo- toneurons or indirectly by the contact with interneurons of the intermediate zone and provide both direct and remote
opioid-induced effects upon upper cervical motoneurons. The central cervical nucleus is a so-far unreported site of termination of cerebellospinal projections. We demon- strated that the projections to this nucleus are contralateral. A role for enkephalin in reflex or coordinated activities of the cerebellospinal system is suggested from the present study since this upper cervical cord station represents an important link between cerebellar output neurons and pe- ripheral inputs.
Acknowledgements
The authors express their gratitude to Fumio Yamashita and Tetsuji Yamamoto for technical assistance, and to
Yuki Okada and Yuki Shiozaki for expert secretarial work. Supported in part by grants from the Ministry of Educa- tion, Science and Culture of Japan, the Ministry of Health
and Welfare of Japan, and the Science Research Promotion Fund of the Japan Private School Promotion Foundation.
References
[1] Achenbach, K.E. and Goodman, D.C., Cerebellar projections to pons, medulla and spinal cord in the albino rat, Brain Behat,. Et'ol., 1 (1968) 43-57.
[2] Angaut, P. and Sotelo, C., Synaptology of the cerebello-olivary pathway. Double labelling with anterograde axonal tracing and GABA immunocytochemistry in the rat, Brain Res., 479 (1989) 361-365.
[3] Asanuma, C., Thach, W.T. and Jones, E.G., Nucleus interpositus projection to spinal interneurons in monkeys, Brain Res., 191 (1980) 245-248.
[4] Batini, C., Buisseret-Delmas, C., Compoint, C. and Daniel, H., The GABAergic neurons of the cerebellar nuclei in the rat: projections to the cerebellar cortex, Neurosci. Lett., 99 (1989) 251-256.
[5] Batini, C., Compoint, C., Buisseret-Delmas, C., Daniel, H_ and Guegan, M., Cerebellar nuclei and the nucleocortical projections in

230 M. lkeda et al. /Brain Research 690 (1995) 225-230
the rat: retrograde tracing coupled to GABA and glutamate immuno- histochemistry, J. Comp. Neurol., 315 (1992) 74-84.
[6] Batton III, R.R., Jayaraman, A., Ruggiero, D. and Carpenter, M.B., Fastigial efferent projections in the monkey: an autoradiographic study, J. Comp. Neurol., 174 (1977) 281-306.
[7] Bentivoglio, M., The organization of the direct cerebellospinal pro- jections, Prog. Brain Res., 57 (1982) 279-290.
[8] Buisseret-Delmas, C. and Angaut, P., The cerebellar olivo- corticonuclear connections in the rat, Prog. Neurobiol., 40 (1993) 63-87.
[9] Fredette, B.J. and Mugnaini, E., The GABAergic cerebello-olivary projection in the rat, Anat. Embryol., 184 (1991) 225-243.
[10] Fukushima, K., Peterson, B.W., Uchino, Y., Coulter, J.D. and Wilson, V.J., Direct fastigiospinal fibers in the cat, Brain Res., 126 (1977) 538-542.
[11] Harlan, R.E., Shivers, B.D., Romano, G.J., Howells, R.D. and Pfaff, D.W., Localization of preproenkephalin mRNA in the rat brain and spinal cord by in situ hybridization, J. Comp. Neurol., 258 (1987) 159-184.
[12] Kaneko, T., Itoh, K., Shigemoto, R. and Mizuno, N., Glutaminase- like immunoreactivity in the lower brainstem and cerebellum of the adult rat, Neuroscience, 32 (1989) 79-98.
[13] Korneliussen, H.K., On the morphology and subdivision of the cerebellar nuclei of the rat, J. Hirnforsch., 10 (1968) 109-122.
[14] Kumoi, K., Saito, N., Kuno, T. and Tanaka, C., Immunohistochemi- cal localization of y-aminobutyric acid- and aspartate-containing neurons in the rat deep cerebellar nuclei, Brain Res., 439 (1988) 302-310.
[15] Leong, S.K., Shieh, J.Y. and Wong, W.C., Localizing spinal-cord- projecting neurons in adult albino rats, J. Comp. Neurol., 228 (1984) 1-17.
[16] Matsushita, M. and Hosoya, Y., The location of spinal projection neurons in the cerebeUar nuclei (cerebellospinal tract neurons) of the
cat. A study with the horseradish peroxidase technique, Brain Res.. 142 (1978) 237-248.
[17] Monaghan, P.L., Beitz, A.J., Larson, A.A., Altschuler, R.A., Madl, J.E. and Mullett, M.A., lmmunocytochemical localization of gluta- mate-, glutaminase- and aspartate aminotransferase-like immuno- reactivity in the rat deep cerebellar nuclei, Brain Res., 363 (1986) 364-370.
[18] Petrusz, P., Merchenthaler, I. and Maderdrut, J.L., Distribution of enkephalin-containing neurons in the central nervous system. In A. Bj/Srklund and T. H61delt (Eds.) Handbook of Chemical Neu- roanatomy, VoL 4: GABA and Neuropeptides in the CNS, Part 1, Elsevier, Amsterdam, 1985, pp. 273-334.
[19] Sugimoto, T. and Mizuno, N., Neurotensin in projection neurons of the striatum and nucleus accumbens, with reference to coexistence with enkephalin and GABA: an immunohistochemical study in the cat, J. Comp. Neurol., 257 (1987) 383-395.
[20] Takahashi, O., Satoda, T., Matsushima, R., Uemura-Sumi, M. and Mizuno, N., Distribution of cerebellar neurons projecting directly to the spinal cord: an HRP study in the Japanese monkey and the cat, J. Hirnforsch., 28 (1987) 105-113.
[21] Thomas, D.M., Kaufman, R.P., Sprague, J.M. and Chambers, W.W., Experimental studies of the vermal cerebellar projections in the brain stem of the cat (fastigiobulbar tract), J. Anat. (Lond.), 90 (1956) 371-385.
[22] Ware, C.B. and Mufson, E.J., Spinal cord projections from the medial cerebellar nucleus in tree shrew (Tupaia glis), Brain Res., 171 (1979) 383-400.
[23] Wilson, V.J., Uchino, Y., Maunz, R.A., Susswein, A. and Fukushima, K., Properties and connections of cat fastigiospinal neurons, Exp. Brain Res.. 32 (1978) 1-17.
[24] Wilson, V.J., Uchino, Y., Susswein, A. and Fukushima, K., Proper- ties of direct fastigiospinal fibers in the cat, Brain Res., 126 (1977) 543-546.





























