Embed Size (px)
Citation preview

J. Mamm. Soc. Japan 18(1) : 19-32 June 1993
Gliding Flight in the Japanese Giant Flying Squirrel Petaurista leucogenys
Motokazu ANDO* and Satoshi SHIRAISHI**
* Environment Division, Shiga Prefectural Government, Shiga 520, Japan
* * Zoological Laboratory, Faculty of Agriculture, Kyushu University 46-06, Fukuoka 812, Japan
(To whom reprint requests should be addressed.) (Accepted 20 February 1993)
Abstract. Field observations of gliding and quadrupedal locomotion in the Japanese giant flying squirrel Petaurista leucogenys were made at five shrine groves with different topographies, tree densities and canopy heights. The main mode of locomotion was gliding, although arboreal quadrupedal locomo- tion was preferred to gliding flight in forests with a canopy height of less than 10 m, presumably because gliding was not energetically advantageous. The mean horizontal length of glides varied from 17.1-33.1 m among the study areas. Short glides of less than 10 m were infrequent. The longest glide recorded was of 115 m, but such long distance glides were rare and normally limited to circumstances where no intervening trees were available. Even when long distance glides were topographically possible, the animals tended to repeat glides of less than 50 m. Squirrels were found capable of gliding at a glide ratio (horizontal distance/vertical drop) of 3.5, but such extremes were not normal. More typically, glide ratios were found to be within the range of 1-3. Both glide ratio and speed tended to increase in longer glides, and horizontal gliding speeds ranged from 7.0-13.3 m/sec. The reasons why glides rarely approach the maximum capacity are discussed in relation to 1) wind velocity, 2) small home ranges, and 3) necessity of buffering landing impacts.
Key words : Petaurista leucogenys ; Gliding flight ; Gliding distance ; Glide ratio ; Aerodynamics.
The Japanese giant flying squirrel Petaurista leucogenys is almost exclusively arboreal, scarcely coming to the ground at any stage of its life cycle. In the Honshu, Kyushu and Shikoku islands of Japan, they are common in low- montane forests and even in small groves surrounding village shrines where other sciurid species are absent. Shrine groves in particular provide flying squirrels with suitable sheltering places and food (AndG & Shiraishi, 1983 ; Andb et al., 1985). The broad pattern of habitat utilization exhibited by P. leucogenys is a direct consequence of its excellent gliding capabilities.
The gliding adaptations of P. leucogenys have been discussed with respect to various aspects of its habitat utilization (Ando & Imaizumi, 1982 ; Baba et al., 1982), relative growth (Andb & Shiraishi, 1984) and quadrupedal locomotion

20 Anda & Shiraishi
(And6 & Shiraishi, 1991). Imaizumi (1983, 1988) has described gliding perfor- mance in extreme habitats where flying squirrels were obliged to exhibit their maximum gliding capability because of the topography. There have been however, no detailed field observations of its gliding performance in more typical habitats.
Although there have been morphological studies discussing the evolution of other mammalian gliders through comparative myology (Johnson-Murray, 1977, 1987) and interspecific allometry (Thorington & Heaney, 1981 ; Scheibe et al., 1990), surprisingly little is known about gliding performance in the field however, and there have been no systematic studies except an aerodynamic analysis of Petaurus breviceps papuanus in the field (Nachtigall et al., 1974; Nachtigall, 1979).
In discussing the gliding performance of such mammals, topography and habitat cannot be ignored because gliding only becomes efficient when there is sufficient vertical variation in the habitat. Also, gliding is sensitive to environ- mental conditions such as wind velocity. Thus, we have made extensive field observations of gliding performance in P. leucogenys at five study areas with different topographies and habitat structures in an effort to elucidate the gliding ecology of this species.
Study Areas
Field observations were carried out a t five shrine groves in Kyushu and in the Kanto district of Honshu, Japan: Yakuoin Temple (YT) on Mt. Takao in Hachioji, on the outskirts of Tokyo; Suiyo Shrine (SU) in Amagi, Fukuoka Prefecture; Tenjin Shrine (TE) in Yamaguni, Oita Prefecture; Takachiho Shrine (TA) at Takachiho, Miyazaki Prefecture; and Yawata Shrine (YS) in Ogawa, Saitama Prefecture.
The YT study area is in the temple precinct covering an area of more than 10 ha (Fig. lA), and is located on a steep mountainside with a maximum
Fig. 1. Views of study areas. A, Yakuoin Temple (YT) ; B, Suiyo Shrine (SU) ; C, Tenjin Shrine (TE).

Gliding Flight of Petaurista leucogenys 21
inclination of 35". Cryptomeria japonica trees, hundreds of years old and more than 30 m high, are abundant, although many of them stand too far apart to allow branch-to-branch quadrupedal locomotion by flying squirrels, forcing them to glide frequently and making them easily observed. Study site SU is situated a t the foot of a gentle hill (Fig. lB), and includes the 2 ha precinct of a small shrine. Acer palmatum trees, 8-12 m tall, line the approach to the shrine, while outside the shrine precinct there is a 25 years old plantation of densely grown (500/ha) 10- 15 m high C. japonica. Because of the density of the plantation, the flying squirrels here can reach most of the trees using quadrupedal locomotion via overlapping branches.
Area TE is in a small (0.7 ha) shrine grove on' level ground at the foot of a hill, and 40 m away from a neighboring hill plantation (Fig. 1C). About one- third of the area is covered with large C. japonica and Quercus trees, about 70% of which can be reached without resorting to gliding. Usually three or four P. leucogenys live in the shrine grove, with their home ranges extending out to the neighboring hills.
Study site T A is in the 4 ha shrine precinct densely covered with giant C. japonica and Quercus spp. up to 30 m iall. Quadrupedal locomotion (hereafter referred to as non-gliding) is possible to almost all the trees in the area. The range of the squirrels is, however, restricted to within the shrine precinct as the region is completely surrounded by the residential area of a local town. Animal density in T A (more than 8/ha) was the highest among the five study areas.
Area YS covers a small (0.3 ha) shrine grove on level land surrounded by paddy fields and a local village. There are only 15 large trees over 20 m high, and only one individual lived, with a home range of about 0.7 ha, used the grove.
Methods
Field observations of gliding and non-gliding locomotion of adult P. leucogenys, were made over a total of 621 hr from January 1975 to June 1989. The horizontal distance of each glide was measured by plotting starting and landing points on field maps. Vertical drop, gliding speed, and gliding posture were determined whenever possible. The heights of tall trees in the field were measured, and the height of starting and landing points were determined photographically by comparing trees of known height with the proportional heights of photographed points.
Video, 16-mm motion picture photography (18 frames/sec) and continuous still photography (5 frameslsec) were employed to analyze glide postures and speeds. Gliding speeds were calculated by dividing the horizontal gliding distance by the flight duration determined from the number of film frames. Gliding speeds several meters before landing were also measured by 16-mm motion pictures taken perpendicular to the glide path. Horizontal distances associated with arboreal quadrupedal locomotion were measured after record-
ing the routes traveled on field maps. Single non-gliding journey was defined as the distance from a departure point in one tree to another point a t which the individual started feeding, resting or gliding.
Results
1. The Ratio of Gliding to Non-gliding Distances Traveled The transition from simple jumping to full gliding flight was progressive.
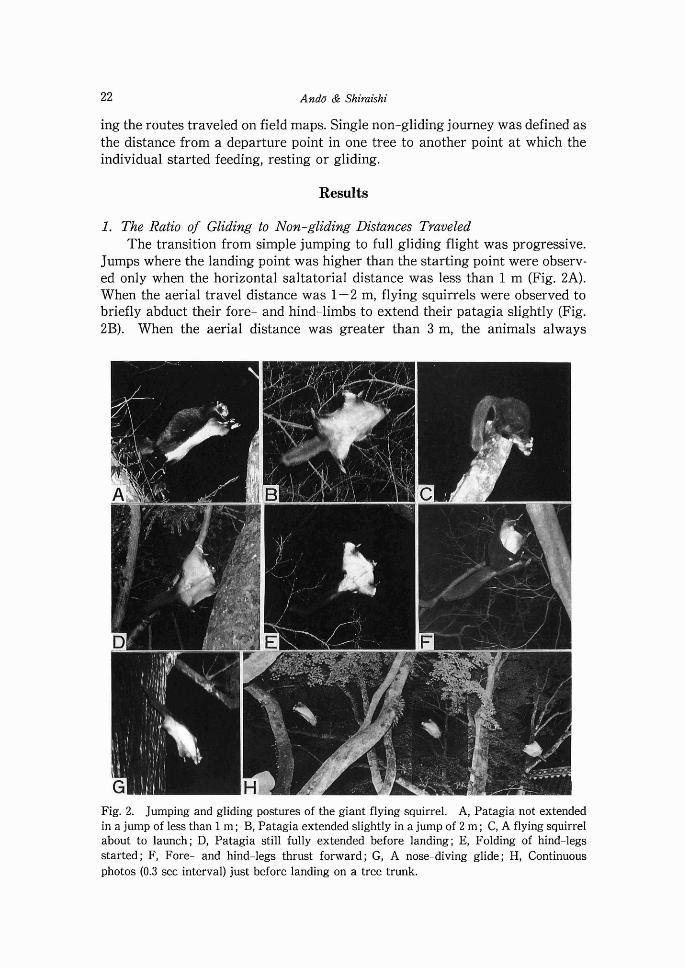
Jumps where the landing point was higher than the starting point were observ- ed only when the horizontal saltatorial distance was less than 1 m (Fig. 2A). When the aerial travel distance was 1-2 m, flying squirrels were observed to briefly abduct their fore- and hind-limbs to extend their patagia slightly (Fig. 2B). When the aerial distance was greater than 3 m, the animals always
Fig. 2. Jumping and gliding postures of the giant flying squirrel. A, Patagia not extended in a jump of less than 1 m ; B, Patagia extended slightly in a jump of 2 m ; C, A flying squirrel about to launch; D, Patagia still fully extended before landing; E, Folding of hind-legs started; F, Fore- and hind-legs thrust forward; G, A nose-diving glide; H, Continuous photos (0.3 sec interval) just before landing on a tree trunk.
Gliding Flight of Petaurista leucogenys 23
assumed the full gliding posture, with fully extended elbow and knee joints and styliform cartilage. Therefore, we defined glides to be those locomotory events covering a horizontal distance of greater than 3 m.
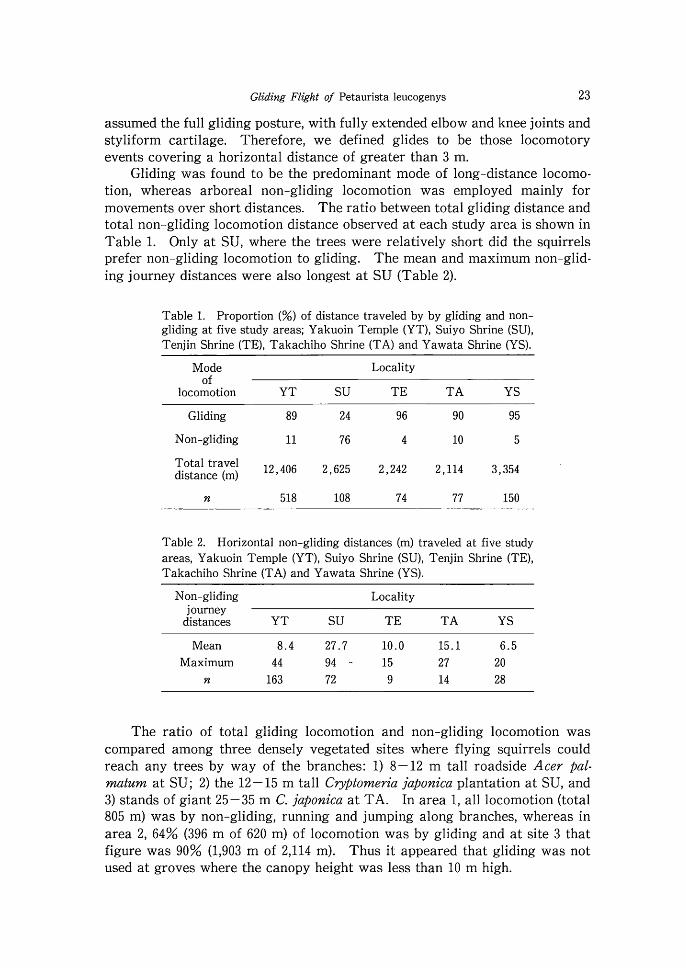
Gliding was found to be the predominant mode of long-distance locomo- tion, whereas arboreal non-gliding locomotion was employed mainly for movements over short distances. The ratio between total gliding distance and total non-gliding locomotion distance observed a t each study area is shown in Table 1. Only at SU, where the trees were relatively short did the squirrels prefer non-gliding locomotion to gliding. The mean and maximum non-glid- ing journey distances were also longest a t SU (Table 2).
Table 1. Proportion (%) of distance traveled by by gliding and non- gliding at five study areas; Yakuoin Temple (YT), Suiyo Shrine (SU), Tenjin Shrine (TE), Takachiho Shrine (TA) and Yawata Shrine (YS).
Mode Locality of
locomotion YT SU T E T A YS
Gliding 89 24 96 90 95
Non-gliding 11 76 4 10 5
12,406 2,625 2,242 2,114 3,354 distance (m)
n 518 108 74 77 150
Table 2. Horizontal non-gliding distances (m) traveled at five study areas, Yakuoin Temple (YT), Suiyo Shrine (SU), Tenjin Shrine (TE), Takachiho Shrine (TA) and Yawata Shrine (YS).
Non-gliding Locality journey
distances YT SU T E TA YS
Mean 8 .4 27.7 10.0 15.1 6 .5 Maximum 44 94 - 15 27 20
n 163 72 9 14 28
The ratio of total gliding locomotion and non-gliding locomotion was compared among three densely vegetated sites where flying squirrels could reach any trees by way of the branches: 1) 8-12 m tall roadside Acer pal- matum at SU; 2) the 12-15 m tall CryPtomeria japonica plantation at SU, and 3) stands of giant 25-35 m C. japonica a t TA. In area 1, all locomotion (total 805 m) was by non-gliding, running and jumping along branches, whereas in area 2, 64% (396 m of 620 m) of locomotion was by gliding and a t site 3 that figure was 90% (1,903 m of 2,114 m). Thus it appeared that gliding was not used a t groves where the canopy height was less than 10 m high.
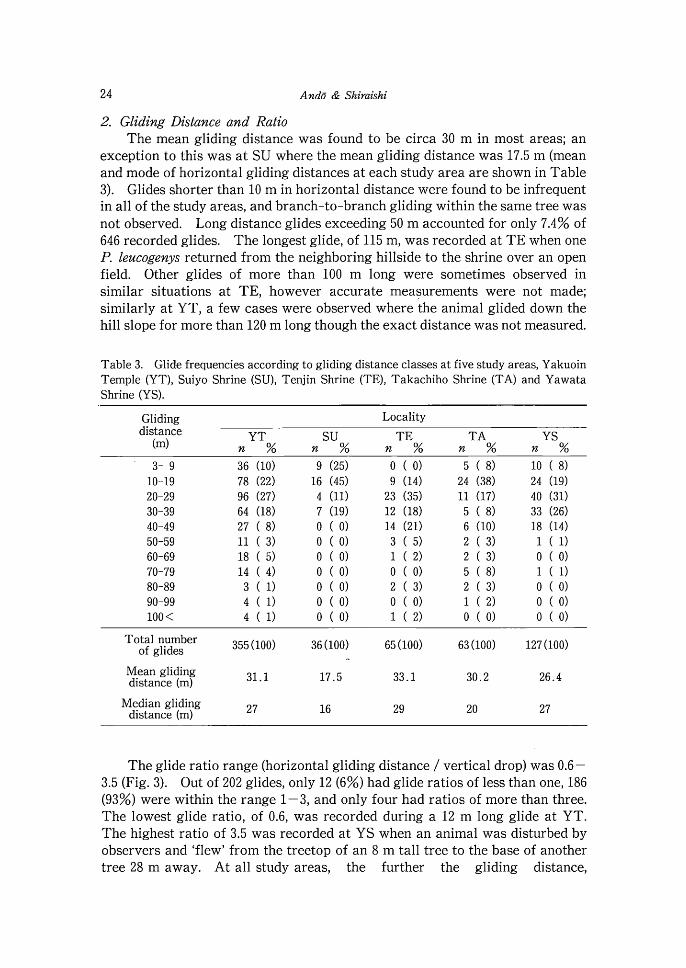
2. Gliding Distance and Ratio The mean gliding distance was found to be circa 30 m in most areas; an
exception to this was a t SU where the mean gliding distance was 17.5 m (mean and mode of horizontal gliding distances a t each study area are shown in Table 3). Glides shorter than 10 m in horizontal distance were found to be infrequent in all of the study areas, and branch-to-branch gliding within the same tree was not observed. Long distance glides exceeding 50 m accounted for only 7.4% of 646 recorded glides. The longest glide, of 115 m, was recorded at T E when one P. leucogenys returned from the neighboring hillside to the shrine over an open field. Other glides of more than 100 m long were sometimes observed in similar situations at TE, however accurate measurements were not made; similarly a t YT, a few cases were observed where the animal glided down the hill slope for more than 120 m long though the exact distance was not measured.
Table 3. Glide frequencies according to gliding distance classes at five study areas, Yakuoin Temple (YT), Suiyo Shrine (SU), Tenjin Shrine (TE), Takachiho Shrine (TA) and Yawata Shrine (YS).
Gliding Locality
Total number 355 of glides 36 (100) 65 (100) 63 (100) 127 (100)
Mean gliding distance (m) 31.1 17.5 33.1 30.2 26.4
Median gliding distance (m) 27 16 29 20 27
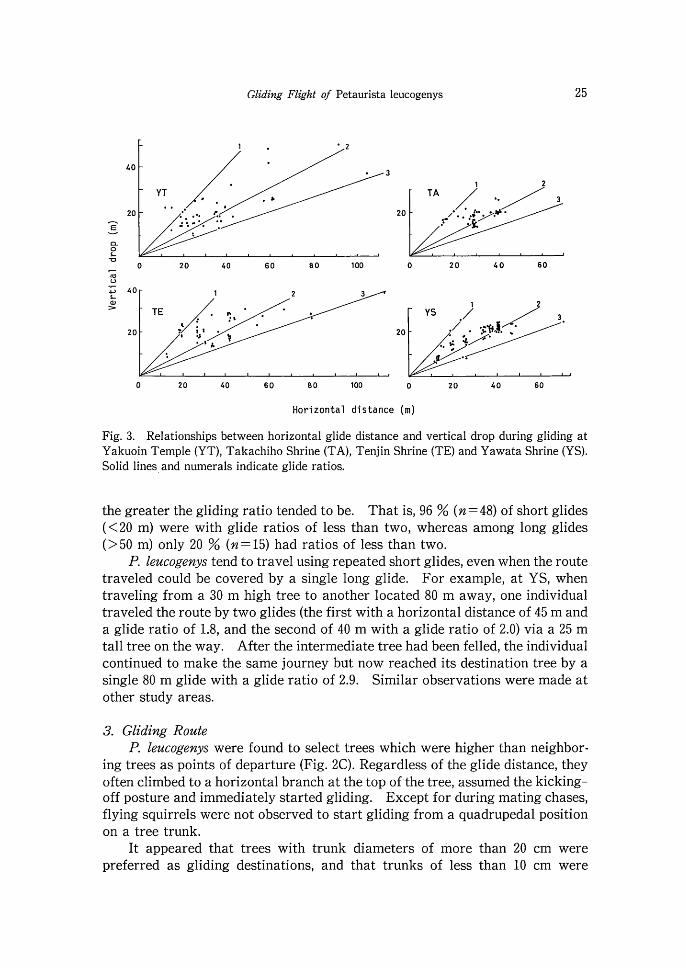
The glide ratio range (horizontal gliding distance / vertical drop) was 0.6- 3.5 (Fig. 3). Out of 202 glides, only 12 (6%) had glide ratios of less than one, 186 (93%) were within the range 1-3, and only four had ratios of more than three. The lowest glide ratio, of 0.6, was recorded during a 12 m long glide at YT. The highest ratio of 3.5 was recorded a t YS when an animal was disturbed by observers and 'flew' from the treetop of an 8 m tall tree to the base of another tree 28 m away. At all study areas, the further the gliding distance,
Gliding Flight of Petaurista leucogenys
Horizontal distance (m)
Fig. 3. Relationships between horizontal glide distance and vertical drop during gliding a t Yakuoin Temple (YT), Takachiho Shrine (TA), Tenjin Shrine (TE) and Yawata Shrine (YS). Solid lines and numerals indicate glide ratios.
the greater the gliding ratio tended to be. That is, 96 % (n=48) of short glides (<20 m) were with glide ratios of less than two, whereas among long glides (>50 m) only 20 % ( n= 15) had ratios of less than two.
P. leucogenys tend to travel using repeated short glides, even when the route traveled could be covered by a single long glide. For example, at YS, when traveling from a 30 m high tree to another located 80 m away, one individual traveled the route by two glides (the first with a horizontal distance of 45 m and a glide ratio of 1.8, and the second of 40 m with a glide ratio of 2.0) via a 25 m tall tree on the way. After the intermediate tree had been felled, the individual continued to make the same journey bat now reached its destination tree by a single 80 m glide with a glide ratio of 2.9. Similar observations were made a t other study areas.
3. Gliding Route P. leucogenys were found to select trees which were higher than neighbor-
ing trees as points of departure (Fig. 2C). Regardless of the glide distance, they often climbed to a horizontal branch at the top of the tree, assumed the kicking- off posture and immediately started gliding. Except for during mating chases, flying squirrels were not observed to start gliding from a quadrupedal position on a tree trunk.
It appeared that trees with trunk diameters of more than 20 cm were preferred as gliding destinations, and that trunks of less than 10 cm were

seldom used as destinations. Usually vertical trunks were used as landing sites, but occasionally landings were made on leaning trunks. Sometimes, the squirrels glided toward slender horizontal branches. Although landings on such branches were usually observed after short glides, in one case one individ- ual landed on a branch after a long distance glide of 58 m. No cases of adults falling to the ground, through failure in landing, were observed.
P. leucogenys are clearly capable of gliding to targets not directly visible from their departure points, doing so by gliding in large curves. For example, when gliding between roadside Cryptomeria trees, individuals were observed gliding first toward open land and then gliding in a curve to land on a target tree. In some cases of long distance gliding, flying squirrels landed on the further side of the target tree, after a circuitous glide. When gliding squirrels were illuminated with flashlights by observers, it happened that they greatly altered their flight directions to land on a different target trees. Although P. leucogenys are able to finely adjust their gliding course in order to pass between branches, occasionally they were observed to contact twigs with their flying membranes. In one extreme one individual became suspended in the air when it became caught on a branch by its membrane while gliding.
4. Gliding Posture Regardless of glide distance and glide ratio, the sequence and timing of
postural changes just after launching from a branch and just before landing on a tree trunk remain essentially the same (Table 4). The flying membrane is kept fully extended until just before landing (Fig. 2D), however, when landing on horizontal branches, they typically grasp the branch with the fore-limbs then climb on to the branch with a kicking movement.
Table 4. The sequence and timing of postural changes during gliding
Time (sec) Postural changes -
After starting 0.15
Before landing 0.30
Starting horizontal extension of fore- and hind-legs simultaneously.
Assuming a typical gliding posture, with elbow, knee and styliform cartilage joints fully extended.
Folding of hind-legs started, but fore-legs fully extended (Fig. 2E). Notable stooping of the backbone to form deep arc.
Increased stooping. Hind-legs thrown toward the target tree trunk.
Folding of fore-legs started.
With deep bending of the backbone, both fore-and hind-legs pushed toward the tree trunk to grasp it with all four limbs simultaneously (Fig. 2F).
Long glides sometimes began with a steep descent (Fig. 2G). As the glide speed increased the angle of the glide decreased, until a steady glide was

Gliding Flight of Petaurista leucogenys 2 7
achieved in which the speed and glide angle were constant. On approaching the landing target, the body was raised and the glide angle decreased until the target was approached almost perpendicular towards the target (Fig. 2H). The angle of the body axis during flight varied significantly according to the conditions affecting gliding. Table 5 shows an example of body axis change when an individual glided a distance of 33 m with a ratio of 2.0 and landed on a rock at the base of a tree. During the 3.67 sec glide, constant speed and glide angle were maintained for only about 1.1 sec. Thus, considerable time in the air was spent accelerating and decelerating.
Table 5. The timing of body axis changes in an example of gliding flight (horizontal distance 3'3 m, glide ratio 2)
Timing (sec) Body axis
Began from a horizontal branch. Body axis showed depression of 45". Depression angle decreased to 15". Start of constant gliding with elevation angle of 0-5".
Slight increase in the elevation angle, with the tail being pushed slightly ventrally. Elevation angle increased to 45". Elevation angle increased to 80-90".
Landed on a rock.
5. Gliding Speed The speed of short jumps (1 -2 m) with only partial extension of the gliding
membrane, was about 3-4.5 m/sec (n=6), whereas the horizontal speed of gliding was much faster, in the range 7.0 - 13.3 m/sec (n = 15). The greater the glide ratio, the faster the glide tended to be (see Fig. 4). The slowest speed of 7.0 m/sec was recorded for a short, steep glide. When gliding was over distances of 40-50 m, with a glide ratio of about two, gliding speed was in the range of 8.5-11 m/sec. In glides longer than 80 m, with a glide ratio of about three, the range was 9.0-13.3 m/sec. Gliding speeds a t points several meters prior to landing were found to be slower than the mean speeds. It seems therefore that increasing the angle of the body axis just prior to landing is effective in decelerating as well as providing additional lift.
6. The Effect of Weather Conditions on Activity P. leucogenys were observed gliding during light rain and snow, and even in
foggy conditions when target landing sites were barely visible to the naked eye of the observers. Wind strength however, clearly affected flying behavior. Wind conditions in the study areas were found to differ considerably according to the topography and vegetation. Even when winds of 5.5-8.0 m/sec (4 on the Beaufort scale) prevailed in open areas, the wind was often very light in the lee of the hills or of mature forest. While winds of 0-3.4 m/sec (Beaufort scales
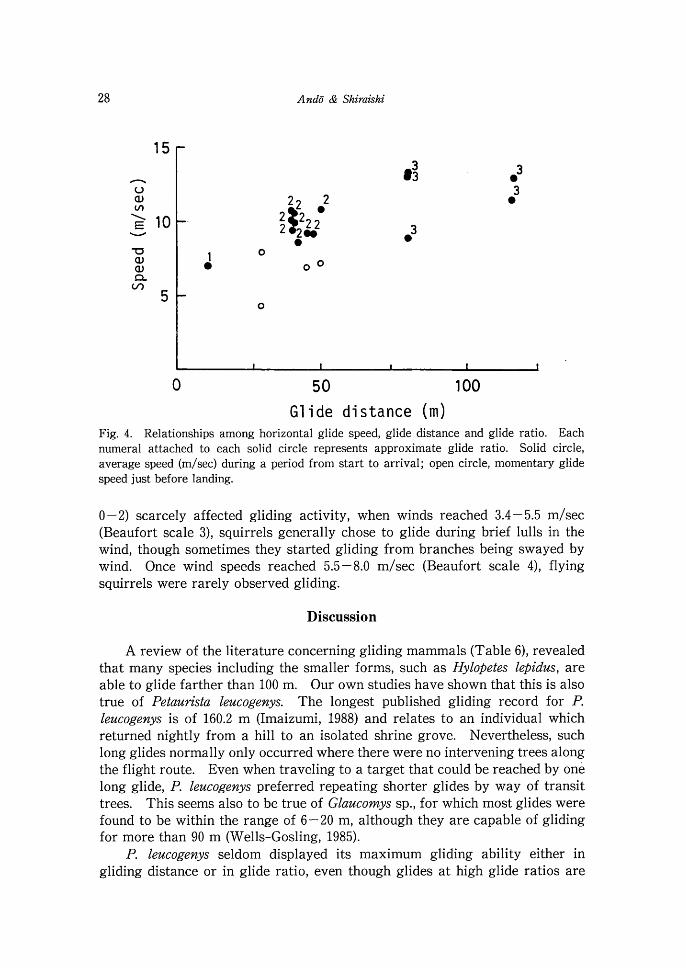
Gl i de d i stance (m) Fig. 4. Relationships among horizontal glide speed, glide distance and glide ratio. Each numeral attached to each solid circle represents approximate glide ratio. Solid circle, average speed (mlsec) during a period from start to arrival; open circle, momentary glide speed just before landing.
0 - 2) scarcely affected gliding activity, when winds reached 3.4 - 5.5 m/sec (Beaufort scale 3), squirrels generally chose to glide during brief lulls in the wind, though sometimes they started gliding from branches being swayed by wind. Once wind speeds reached 5.5-8.0 m/sec (Beaufort scale 4), flying squirrels were rarely observed gliding.
Discussion
A review of the literature concerning gliding mammals (Table 6), revealed that many species including the smaller forms, such as Hylopetes lepidus, are able to glide farther than 100 m. Our own studies have shown that this is also true of Petaurista leucogenys. The longest published gliding record for P. leucogenys is of 160.2 m (Imaizumi, 1988) and relates to an individual which returned nightly from a hill to an isolated shrine grove. Nevertheless, such long glides normally only occurred where there were no intervening trees along the flight route. Even when traveling to a target that could be reached by one long glide, P. leucogenys preferred repeating shorter glides by way of transit trees. This seems also to be true of Glaucomys sp., for which most glides were found to be within the range of 6-20 m, although they are capable of gliding for more than 90 m (Wells-Gosling, 1985).
P. leucogenys seldom displayed its maximum gliding ability either in gliding distance or in glide ratio, even though glides a t high glide ratios are

Gliding Flight of Petaurista leucogenys 29
Table 6. Gliding performances of mammals with flying membranes
Species Gliding distance(m) Source
Marsupialia Schoinobates volans Petaums breviceps ditto P. australis
Dermoptera Cynocephalus variegatus ditto
Rodentia Petaurista leucogenys P. petaurista P. sp. P. sp. P. sp. Glaucomys volans ditto G. sabrinus
Glaucomys sp. Hylopetes lepidus Anomalums derbianus
Grzimek, 1972 Nowak, 1991
1.7-2.7 Nachtigall, 1979 Nowak, 1991
11-13 Nowak, 1991 5 Imaizumi & Obara, 1966
Imaizumi, 1988 1 Thorington & Heaney, 1981 2.4 Thorington & Heaney, 1981 -
3 Davis, 1962 Nowak, 1991
2.8 Nowak, 1991 2-3 Thorington & Heaney, 1981
Wells-Gosling, 1985
Wells-Gosling, 1985
Nowak, 1991 Kingdon, 1974
energetically more efficient. Given a horizontal distance of about 30 m, squir- rels were capable of gliding at a glide ratio of 3.5, yet normal gliding ratios recorded were only 1-3. One reason for this fact is that P. leucogenys home ranges are limited to an area of just a few hectares : (0.46-5.16 ha, Baba et al., 1982 ; and 0.4-4.7 ha, Kawamichi, 1984) and long distance glides would be unnecessary in such small home ranges with continuous vegetation. Further- more, visiting more trees during a journey may also be advantageous explora- tory behavior, because the total distance traveled per night by P. leucogenys is only 111-620 m, and thus the number of visiting places is limited (Ando & Imaizumi, 1982).
A further reason for not reaching maximum gliding ratios may involve the psychological and technical difficulties of long distance gliding. In the case of the longest glide recorded, of 160.2 m, only one individual was able to glide the route initially, however several other individuals in the shrine grove subse- quently learned to glide the same route (Imaizumi, 1988).
Thirdly, wind condition is a significant environmental factor affecting P. leucogenys, since they cannot glide against the wind. During the field study, wind velocities of 3.4-5.5 m/sec (3 on the Beaufort scale) were common. Judging from their gliding speeds (7-13.3 m/sec), the glide ratio may drop to less than one half of that in windless conditions if it dares to glide against the

30 And0 & Shiraishi
wind. Because the major gliding routes of an individual in its own home range tend to be fixed (Ando & Imaizumi, 1982), such routes have to be chosen conservatively to allow gliding even in windy conditions. Reduced gliding activity under windy conditions also indicates the adverse effects of wind. The similar glide speeds (8-15 m/sec) observed in Petaurus breviceps (Nach- tigall et al., 1974) suggests that wind may be an important environmental factor
. .
for other gliding mammals as well. During a constant glide, velocity and gliding angle are consequences of
wing loading, aspect ratio and the coefficient of lift. The gliding ratios of marsupial and rodent gliders are generally less than three (Table 6). Glide ratios less than three are considerably smaller than those obtained for bats and birds in wind tunnel tests; e.g., 6.8 in Rousettus aegyptiacus (Pennycuick, 1971) and 11.0 in Parabuteo unicinctus (Tucker & Heine, 1990). This is probably the result of the differences in wing loadings and aspect ratios. Wing loadings in the Petauristinae (30- 110 N/m2) are greater than those (10-20 N/m2) in the Chiroptera (Thorington & Heaney, 1981). The aspect ratios of Petaurista spp. range from 1.0 to 2.2 (Thorington & Heaney, 1981) and that for P. leucogenys is 1.76 (And6 & Shiraishi, 1984), whereas the figures in all bats are more than 5, and for Aquila chrysaetos it is 6.17 (Azuma, 1979).
High wing loadings reduce the maneuverability of gliding mammals in the air. For example, a smaller glider with a lower wing loading (Idiurus zenkeri) has been observed to glide from a 10 m tall tree and land lower down the same tree trunk after a spiral glide (Kingdon, 1974). In contrast, P. leucogenys with a much higher wing loading of 81 N/m2 (And6 & Shiraishi, 1984) can glide from a 25 m high tree top and barely reach the base of the same tree after a large U- turn (Imaizumi, 1988).
Birds and bats are able to vary their gliding speeds by controlling their wing loadings and aspect ratios through wing position during flight (Tucker & Parrott, 1970). In a wind tunnel test, Columba livia, for example, was found to be able to glide at speeds ranging from 8.6 to 22.1 m/sec (Pennycuick, 1968), while Rousettus aegyptiacus was able to glide a t speeds from 5.3 to 11.0 m/sec (Pennycuick, 1971). In contrast, the wing area and aspect ratios of P. leucogenys are kept almost constant during flight because it glides by fully extending its patagial membranes. Although P. leucogenys was capable of controlling its camber and angle of attack, the range of control had to be within an aerodynamically permissive extent to prevent stalling. The fact that the gliding speed of P. leucogenys a t a certain glide ratio was limited to a relatively narrow range, i.e., 8.5-11.0 m/sec a t a glide ratio of 2 and 9.0-13.3 m/sec a t a glide ratio 3, is indicative that it is unable to vary its gliding posture.
Gliding is more energetically efficient than quadrupedal locomotion (Tucker, 1970), though this does not apply to steeply dropping glides. Over short distances, P. leucogenys was found to prefer non-gliding locomotion to gliding. This was considered to be because over short distances time is too short for P. leucogenys to attain the optimal glide ratio of a constant glide. Moreover, gliding with a very low glide ratio, as happens in a parachute

Gliding Flight of Petaurista leucogenys 31
descent, lacks stability in the air and does not fully permit maneuverability in the air (Azuma, 1979).. This lack of stability and control would be disadvanta- geous for P, leucogenys as it must retain maneuverability during flight to evade small branches. The fact that P. leucogenys also favors landing on large tree trunk may also indicate the importance of safer landing.
The stout hind-foot of P. leucogenys is particularly suited to clinging firmly to tree trunks, but a t the cost of lacking agility on narrow twigs (And6 & Shiraishi, 1991). As is indicated by such morphological specialization, 'failure free' landing is of vital importance for this gliding species. When landing on a vertical tree trunk after a glide with a low glide ratio, the remaining vertical speed acts as a vector forcing the body downward, and it exerts considerable load on the claws of the fore- and hind-feet. This may be another factor discouraging short distance glides.
In order to ensure a safe landing, it is essential to reduce landing speed, because the impact increases in proportion to a square of the gliding speed at landing. For example, if the animal lands on a target at a speed of around 10 m/sec, without deceleration, the impact would be more than ten times as much as that of a short leap (3 -4.5 m/sec). The deep stooping posture of the animal a t landing seems an effective way of buffering the impact.
In conclusion, P. leucogenys is capable of utilizing gliding locomotion in forests with a canopy height of more than 10 m. However, typical gliding distances and ratios are modest in comparison to its maximum gliding capacity.
Acknowledgments
We wish to express our sincere thanks to Drs. Andrew S. Fraser, John S. Scheibe and Mr. Joe Banerjee for improving the manuscript.
References
Ando, M. & Y. Imaizumi. 1982. Habitat utilization of the white-cheeked giant flying squirrel Petaurista leucogenys in a small shrine grove. J. Mamm. Soc. Japan, 9 : 70-81 (in Japanese with English abstract).
AndG, M. & S. Shiraishi. 1983. The nest and nest-building behavior of the Japanese giant flying squirrel, Petaurista leucogenys. Sci. Bull. Fac. Agr., Kyushu Univ., 38 : 59-69 (in Japanese with English summary).
AndB, M. & S. Shiraishi. 1984. Relative growth and gliding adaptations in the Japanese giant flying squirrel, Petaurista leucogenys. Sci. Bull. Fac. Agr., Kyushu Univ., 39 : 49-57 (in Japanese with English summary).
AndG, M. & S. Shiraishi. 1991. Arboreal quadrupedalism and gliding adaptations in the Japanese giant flying squirrel, Petaurista leucogenys. Honyurui Kagaku [Mammalian Science], 30 : 167- 181 (in Japanese with English abstract).
And6, M., S. Shiraishi & T. A. Uchida. 1985. Food habits of the Japanese giant flying squirrel, Petaurista leucogenys. J. Fac. Agr., Kyushu Univ., 29 : 189- 202.
Azuma, A. 1979. Flight of Organisms. Kodansha, Tokyo, 255 pp. (in Japanese). Baba, M., T. Doi & Y. Ono. 1982. Home range utilization and nocturnal activity of the giant flying
squirrel, Petaurista leucogenys. Jpn. J. Ecol., 32 : 189 - 198.

32 AndD & Shiraishi
Davis, D. D. 1962. Mammals of the lowland rainforest of north Borneo. Bull. Natl. Mus., Singapore, 31 : 1- 129.
Grzimelt, B. 1972. Grzimek's Animal Life Encyclopedia. Vol.10. Van Nostrand Reinhold Company, New York, 627 pp.
Imaizumi, Y. 1983. Behavioral modifications of some flying squirrels. Anima, l l (3) : 56-63 (in Japanese).
Imaizumi, Y. 1988. Animal champions: the longest glide of the Japanese giant flying squirrel. Anima, 16(1): 16 - 19 (in Japanese).
Imaizumi, Y. & H. Obara. 1966. A Natural History of Mammals of the World: Monotremata & Marsupialia. Shinshichyosha, Tokyo, 559 pp. (in Japanese).
Johnson-Murray, J. L. 1977. Myology of the gliding membranes of some petauristine rodents (Genera : Glazicomys, Pteromys, Petinojnys and Petazirista). J. Mamm., 58 : 374-384.
Johnson-Murray, J. L. 1987. The comparative myology of the gliding membranes of Acrobates, Petazrroides and Petaztnis contrasted with the cutaneous myology of Hemibelidezis and Pseudo- clzeinn (Marsupialia : Phalangeridae) and with selected gliding Rodentia (Sciuridae and Anomaluridae). Aust. J, Zool., 35 : 101 - 113.
Kawamichi, T. 1984. The world of giant flying squirrels gliding in the dark (2). Shizen [Nature], 39(2) : 64-72 (in Japanese).
Kingdon, J. 1974. East African Mammals, Vo1.2, Part B. Academic Press, New York, 762 pp. Nachtigall, W. 1979. Gleitflug des Flugbeutlers Petaums breuiceps papuanzis : 11. Filmanalysen zur
Einstellung von Gleitbahn und Rumpf sowie zur Steuerung des Gleitflugs. J. Comp. Physiol., 133 : 89-95.
Nachtigall, W., R. Grosch & T. Schultze-Westrum. 1974. Gleitflug des Flugbeutlers Pe taum breuiceps papuanus (Thomas) : Flugverhalten und Flugsteuerung. J. Comp. Physiol., 92 : 105- 115.
Nowak, R. M. 1991. Walker's Mammals of the World, Fifth Edition. The Johns Hopkins Univ. Press, Baltimore, 1,629 pp.
Pennycuick, C. J. 1968. A wind-tunnel study of gliding flight in the pigeon Columba livia. J. Exp. Biol., 49 : 509-526.
Pennycuick, C. J. 1971. Gliding flight of the dog-faced bat Rousettus aegyPtiacus observed in a wind tunnel. J. Exp. Biol., 55 : 833-845.
Scheibe, J. S., D. Figgs & J. Heiland. 1990. Morphological attributes of gliding rodents : A prelimi- nary analysis. Trans. Missouri Acad. Sci., 24 : 49-56.
Thorington, R. W. Jr. & L. R. Heaney. 1981. Body proportions and gliding adaptations of flying squirrels (Petauristinae). J. Mamm., 62 : 101 - 114.
Tucker, V. A. 1970. Energetic cost of locomotion in animals. Comp. Biochem. Physiol, 34 : 841-846. Tucker, V. A. & C. Heine. 1990. Aerodynamics of gliding flight in a Harris' hawk, Parabuteo
unicinctzis. J. Exp. Biol., 149 : 469-489. Tucker, V. A. & G. C. Parrott. 1970. Aerodynamics of gliding flight in a falcon and other birds. J.
Exp. Biol., 52 : 345-367. Wells-Gosling, N. 1985. Flying Squirrels: Gliders in the Dark. Smithsonian Institution Press,
Washington, D. C., 128 pp.





























