Embed Size (px)
Citation preview

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•185
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Camélidos sudamericanos
Grupo 5

186 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
IntRODUCCIónLos camélidos sudamericanos son especies muy importantes en la economía andina; por constituir fuente de carne, fibra y trabajo para los criadores que habitan las zonas altoandinas por encima de los 4,000 msnm. Estos animales utilizan extensas áreas de praderas naturales, que debido a factores asociados a la altitud no podrían ser aprovechadas de manera eficiente por otros animales domésticos (Novoa y Flores, 1991).
Actualmente esta crianza se desarrolla en condiciones de comunidades campesinas el 95%, por lo tanto, se requiere seguir trabajando en el campo de la investigación y validación para contribuir a mejorar los niveles de producción y productividad, si se tiene en cuenta que el 70% de los productores desarrollan una crianza tradicional. La producción de camélidos como en otras especies, está sustentada en cuatro factores importantes, tres de ellos relacionados al medio ambiente: La sanidad, la alimentación y el manejo; y uno relacionado a la biología propia del animal como es la genética. En la zona altoandina a nivel de las comunidades campesinas, donde se encuentra la mayor concentración de llamas y alpacas, la producción y productividad es muy baja, en lo referente a peso vellón, peso vivo, características supeditadas al medio ambiente, Sin embargo por influencia de la demanda del mercado se observa el blanqueo de los rebaños, la desaparición de los animales de color y una mayor saca de las llamas para incrementar las alpacas, en estas condiciones existe el peligro de la desaparición de la variabilidad genética de los camélidos.
El Programa Nacional de Investigación en Camélidos (P.N.I.C.) consciente de su rol protagónico en la generación de tecnología, presenta los avances que se vienen logrando en el fortalecimiento del Banco de Germoplasma del INIA en la EE. Illpa – CIP Quimsachata.
AntECEDEntESEn 1985 el Instituto de Investigación y Promo-ción Agraria - INIPA, creó el Programa Nacional de
experienCia del inia en el FortaleCimiento del BanCo de germoplasma de Camélidos doméstiCos
huanca, t1; Apaza, n1; Gonzáles, M1.
1 Instituto nacional de Investigación Agraria. Prog nac. Invest. en Camélidos
Camélidos Domésticos Andinos para generar tecnolo-gía y apoyar la promoción de su crianza, para ello reci-bió como transferencia el predio Quimsachata donde estableció un núcleo inicial de alpacas y llamas.
Posteriormente, en 1987, con el apoyo técnico, financiero del Proyecto A]pacas (PAL), Convenio de Cooperación Técnica del Gobierno Suizo COTESU INIA, se estableció en la Estación Experimental IIlpa Puno, Anexo Quimsachata, un Banco de Germoplasma de Alpacas y Llamas orientado inicialmente a la recu-peración de alpacas de color de raza Suri y Huacayo y llamas en sus dos ecotipos, a partir de esta fecha se viene estabilizando el capital pecuario; asimismo generando reproductores para orientarlos al mercado de comunidades vecinas. A partir de 1993, el Programa Nacional de Investigación de Camélidos Domésticos Andinos, pasa a constituirse como órgano de acción dentro de la Estructura del INIA, como Programa Nacional de Investigación en Camélidos.
IMPORtAnCIA DEL BAnCO DE GERMOPLASMA DE ALPACAS DE COLOR y LLAMASAnte el blanqueo inminente de los rebaños de alpacas que se encuentra en poder de las comunidades campesinas, pequeños, medianos y grandes criadores se hace necesario contar con Bancos de Germoplasma de alpacas de color en sus dos razas y las llamas en sus dos ecotipos por las siguientes razones:
- Permite recuperar la variabilidad de las alpacas de color, que por influencia del mercado externo, van camino a la desaparición.
- Disponer de una variabilidad de colores que presentan las alpacas de acuerdo a las tonalidades identificadas por la industria.
- Diseñar, un manejo técnico de los animales en una determinada extensión de terreno.
- Implementar un sistema de empadre que permita manejar los animales de color.
- Recuperar, animales de colores raros o muy escasos en nuestro país mediante la biotecnología reproductiva: Inseminación artificial y transferencia de embriones.
- Realizar cruzamientos planificados entre colores para determinar la herencia de colores.
- Estudios específicos sobre las características

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•187
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
productivas, reproductivas y/o otras que sean potenciales de los animales de color frente a las blancas.
- Constituir un centro de producción de reproductores de alpacas de color para implementar nuevos centros, sean estas para productores, CC y/o instituciones estatales u ONGs.
- Constituir centros de transferencia tecnológica de una crianza tecnificada.
OBJEtIVOS GEnERALESContribuir al incremento de los niveles de producción y productividad de la crianza de camélidos, generando alternativas tecnológicas para impulsar la crianza sostenible y conservación de la biodiversidad genética.
MEtODOLOGíA DE tRABAJOEl Programa de Conservación y Mejoramiento Genético de las alpacas considera 3 fases:
Fase I: 1983 -1997 -Acopio de vientres y reproductores de color
- Capitalización del rebaño
- Definición y registro de base de datos
- Contar con el 95% de las tonalidades de color identificado por la industria
Fase II: 1998 - 2002 - Consolidación del capital pecuario
- Implementación del empadre controlado- Registro de información - producción reproducción- Iniciar trabajos de investigación- Selección por color definido- Lograr crias de color definido similar y/o mejores
que sus progenitores
Fase III: 2003 - 2010 - Consolidación del Banco de Germoplasma de las alpacas de color
- Producir reproductores de alpacas de color de probada calidad para su difusión
- Consolidar el funcionamiento de la red de Banco de Germoplasma
- Promover la implementación de Bancos de Germoplasma
- Promover el funcionamiento de una red de Banco de Germoplasma
- Desarrollar trabajos de investigación- Realizar selección por color, finura de fibra y fenotipo definido
CARACtERíStICAS DEL AnExO qUIMSAChAtALa E.E. IlIpa - Anexo Quimsachata, lugar donde el P.N.I.C. realiza acciones de investigación se encuentra ubicada entre los distritos de Santa Lucia y Cabanillas de las provincias de Lampa y San Román respectivamente en el departamento de Puno, a 15°04’00” latitud sur y 70°78’00” longitud oeste, a una altitud promedio de 4 300 msnm y a 118 km de la ciudad de Puno. Esta constituida por sectores: Central Quimsachata, Compuerta Huata, Tincopalca y Huasicara, que en conjunto abarca una extensión total de 6 281 has. La temperatura fluctúa entre 3°C de Mayo a Julio y 15°C entre setiembre y diciembre; siendo el promedio durante el año aproximadamente 7°C.
POBLACIónLos Cuadros 1 y 2, muestran la población inicial y actual del capital pecuario, entre alpacas y llamas del Banco de Germoplasma del CEQ, en ella se observa de que existe una diferencia significativa entre la población inicial y la población actual, ello se debe a que se viene buscando la estabilización de los rebaños en función de la pradera nativa y la soportabilidad que esta puede ofrecer.
Cuadro 1. Población inicial y actual del banco de germoplasma de alpacas - Quimsachata.
Sexo Campaña Clase Inicial 1996Actual 2007
MachosPadresTuísCrías
247138
84
13734
267
HembrasmadresTuísCrías
734124
79
994132229
TOTAL 1 406 1 793
Por las características de la pradera nativa que ofrece el CIP Quimsachata, presenta condiciones apropiadas para la crianza de llamas en sus dos ecotipos Káras y Chákus, en razon de este diagnostico agrostoedafologico es que a traves de los años se ha venido incrementado el capital pecuario con vientres y reproductores con fenotipo definido, por ello es que se cuenta con el 50% de cada variedad Cuadro 2
188 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
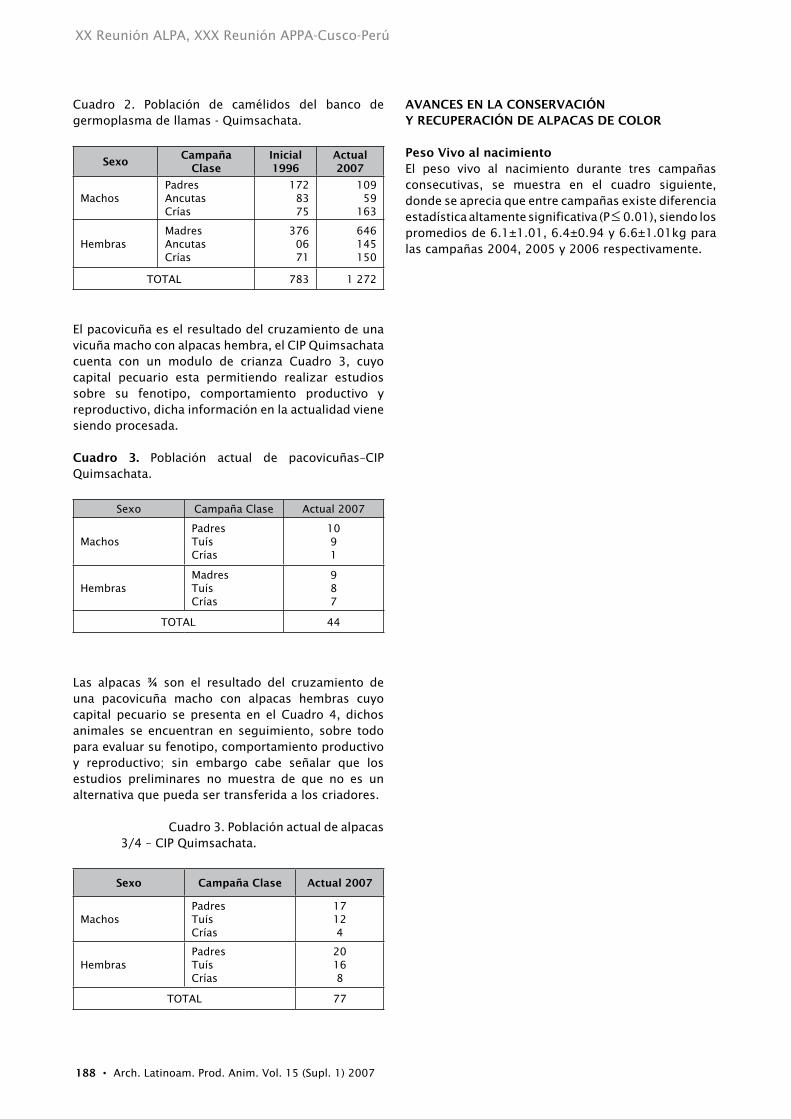
Cuadro 2. Población de camélidos del banco de germoplasma de llamas - Quimsachata.
SexoCampaña
ClaseInicial 1996
Actual 2007
MachosPadresAncutasCrías
1728375
10959
163
HembrasMadresAncutasCrías
3760671
646145150
TOTAL 783 1 272
El pacovicuña es el resultado del cruzamiento de una vicuña macho con alpacas hembra, el CIP Quimsachata cuenta con un modulo de crianza Cuadro 3, cuyo capital pecuario esta permitiendo realizar estudios sobre su fenotipo, comportamiento productivo y reproductivo, dicha información en la actualidad viene siendo procesada.
Cuadro 3. Población actual de pacovicuñas–CIP Quimsachata.
Sexo Campaña Clase Actual 2007
MachosPadresTuísCrías
1091
HembrasMadresTuís Crías
987
TOTAL 44
Las alpacas ¾ son el resultado del cruzamiento de una pacovicuña macho con alpacas hembras cuyo capital pecuario se presenta en el Cuadro 4, dichos animales se encuentran en seguimiento, sobre todo para evaluar su fenotipo, comportamiento productivo y reproductivo; sin embargo cabe señalar que los estudios preliminares no muestra de que no es un alternativa que pueda ser transferida a los criadores.
Cuadro 3. Población actual de alpacas 3/4 – CIP Quimsachata.
Sexo Campaña Clase Actual 2007
MachosPadresTuís Crías
17124
HembrasPadresTuísCrías
20168
TOTAL 77
AVAnCES En LA COnSERVACIón y RECUPERACIón DE ALPACAS DE COLOR
Peso Vivo al nacimientoEl peso vivo al nacimiento durante tres campañas consecutivas, se muestra en el cuadro siguiente, donde se aprecia que entre campañas existe diferencia estadística altamente significativa (P≤ 0.01), siendo los promedios de 6.1±1.01, 6.4±0.94 y 6.6±1.01kg para las campañas 2004, 2005 y 2006 respectivamente.
Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•189
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
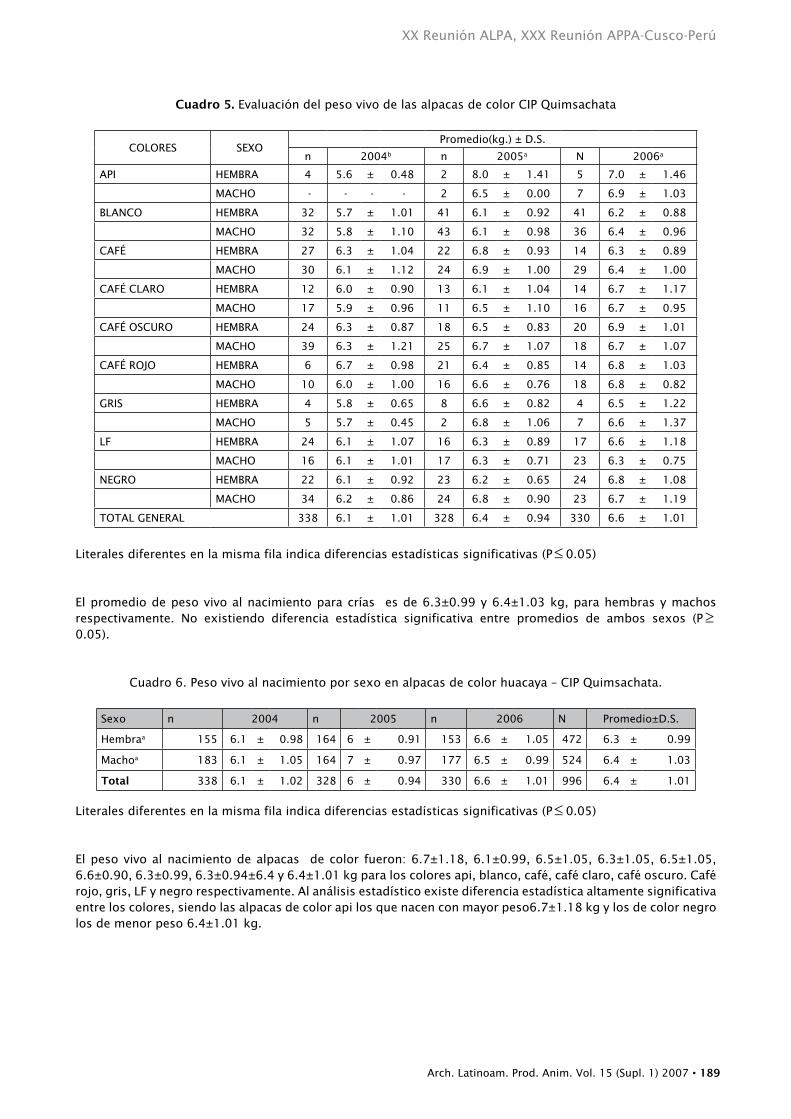
Cuadro 5. Evaluación del peso vivo de las alpacas de color CIP Quimsachata
COLORES SEXOPromedio(kg.) ± D.S.
n 2004b n 2005a N 2006a
API HEMBRA 4 5.6 ± 0.48 2 8.0 ± 1.41 5 7.0 ± 1.46
MACHO - - - - 2 6.5 ± 0.00 7 6.9 ± 1.03
BLANCO HEMBRA 32 5.7 ± 1.01 41 6.1 ± 0.92 41 6.2 ± 0.88
MACHO 32 5.8 ± 1.10 43 6.1 ± 0.98 36 6.4 ± 0.96
CAFÉ HEMBRA 27 6.3 ± 1.04 22 6.8 ± 0.93 14 6.3 ± 0.89
MACHO 30 6.1 ± 1.12 24 6.9 ± 1.00 29 6.4 ± 1.00
CAFÉ CLARO HEMBRA 12 6.0 ± 0.90 13 6.1 ± 1.04 14 6.7 ± 1.17
MACHO 17 5.9 ± 0.96 11 6.5 ± 1.10 16 6.7 ± 0.95
CAFÉ OSCURO HEMBRA 24 6.3 ± 0.87 18 6.5 ± 0.83 20 6.9 ± 1.01
MACHO 39 6.3 ± 1.21 25 6.7 ± 1.07 18 6.7 ± 1.07
CAFÉ ROJO HEMBRA 6 6.7 ± 0.98 21 6.4 ± 0.85 14 6.8 ± 1.03
MACHO 10 6.0 ± 1.00 16 6.6 ± 0.76 18 6.8 ± 0.82
GRIS HEMBRA 4 5.8 ± 0.65 8 6.6 ± 0.82 4 6.5 ± 1.22
MACHO 5 5.7 ± 0.45 2 6.8 ± 1.06 7 6.6 ± 1.37
LF HEMBRA 24 6.1 ± 1.07 16 6.3 ± 0.89 17 6.6 ± 1.18
MACHO 16 6.1 ± 1.01 17 6.3 ± 0.71 23 6.3 ± 0.75
NEGRO HEMBRA 22 6.1 ± 0.92 23 6.2 ± 0.65 24 6.8 ± 1.08
MACHO 34 6.2 ± 0.86 24 6.8 ± 0.90 23 6.7 ± 1.19
TOTAL GENERAL 338 6.1 ± 1.01 328 6.4 ± 0.94 330 6.6 ± 1.01
Literales diferentes en la misma fila indica diferencias estadísticas significativas (P≤ 0.05)
El promedio de peso vivo al nacimiento para crías es de 6.3±0.99 y 6.4±1.03 kg, para hembras y machos respectivamente. No existiendo diferencia estadística significativa entre promedios de ambos sexos (P≥0.05).
Cuadro 6. Peso vivo al nacimiento por sexo en alpacas de color huacaya – CIP Quimsachata.
Sexo n 2004 n 2005 n 2006 N Promedio±D.S.
Hembraa 155 6.1 ± 0.98 164 6 ± 0.91 153 6.6 ± 1.05 472 6.3 ± 0.99
Machoa 183 6.1 ± 1.05 164 7 ± 0.97 177 6.5 ± 0.99 524 6.4 ± 1.03
total 338 6.1 ± 1.02 328 6 ± 0.94 330 6.6 ± 1.01 996 6.4 ± 1.01
Literales diferentes en la misma fila indica diferencias estadísticas significativas (P≤ 0.05)
El peso vivo al nacimiento de alpacas de color fueron: 6.7±1.18, 6.1±0.99, 6.5±1.05, 6.3±1.05, 6.5±1.05, 6.6±0.90, 6.3±0.99, 6.3±0.94±6.4 y 6.4±1.01 kg para los colores api, blanco, café, café claro, café oscuro. Café rojo, gris, LF y negro respectivamente. Al análisis estadístico existe diferencia estadística altamente significativa entre los colores, siendo las alpacas de color api los que nacen con mayor peso6.7±1.18 kg y los de color negro los de menor peso 6.4±1.01 kg.
190 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
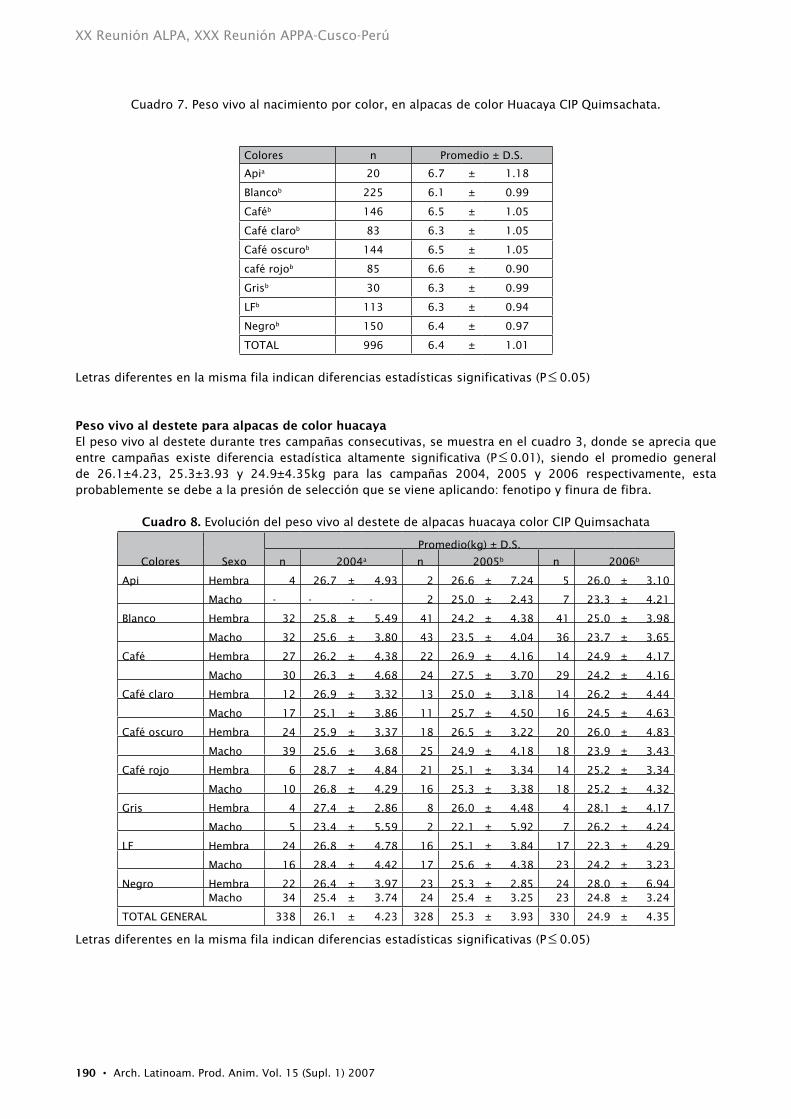
Cuadro 7. Peso vivo al nacimiento por color, en alpacas de color Huacaya CIP Quimsachata.
Colores n Promedio ± D.S.
Apia 20 6.7 ± 1.18
Blancob 225 6.1 ± 0.99
Caféb 146 6.5 ± 1.05
Café clarob 83 6.3 ± 1.05
Café oscurob 144 6.5 ± 1.05
café rojob 85 6.6 ± 0.90
Grisb 30 6.3 ± 0.99
LFb 113 6.3 ± 0.94
Negrob 150 6.4 ± 0.97
TOTAL 996 6.4 ± 1.01
Letras diferentes en la misma fila indican diferencias estadísticas significativas (P≤ 0.05)
Peso vivo al destete para alpacas de color huacaya El peso vivo al destete durante tres campañas consecutivas, se muestra en el cuadro 3, donde se aprecia que entre campañas existe diferencia estadística altamente significativa (P≤ 0.01), siendo el promedio general de 26.1±4.23, 25.3±3.93 y 24.9±4.35kg para las campañas 2004, 2005 y 2006 respectivamente, esta probablemente se debe a la presión de selección que se viene aplicando: fenotipo y finura de fibra.
Cuadro 8. Evolución del peso vivo al destete de alpacas huacaya color CIP Quimsachata
Colores Sexo
Promedio(kg) ± D.S.
n 2004a n 2005b n 2006b
Api Hembra 4 26.7 ± 4.93 2 26.6 ± 7.24 5 26.0 ± 3.10
Macho - - - - 2 25.0 ± 2.43 7 23.3 ± 4.21
Blanco Hembra 32 25.8 ± 5.49 41 24.2 ± 4.38 41 25.0 ± 3.98
Macho 32 25.6 ± 3.80 43 23.5 ± 4.04 36 23.7 ± 3.65
Café Hembra 27 26.2 ± 4.38 22 26.9 ± 4.16 14 24.9 ± 4.17
Macho 30 26.3 ± 4.68 24 27.5 ± 3.70 29 24.2 ± 4.16
Café claro Hembra 12 26.9 ± 3.32 13 25.0 ± 3.18 14 26.2 ± 4.44
Macho 17 25.1 ± 3.86 11 25.7 ± 4.50 16 24.5 ± 4.63
Café oscuro Hembra 24 25.9 ± 3.37 18 26.5 ± 3.22 20 26.0 ± 4.83
Macho 39 25.6 ± 3.68 25 24.9 ± 4.18 18 23.9 ± 3.43
Café rojo Hembra 6 28.7 ± 4.84 21 25.1 ± 3.34 14 25.2 ± 3.34
Macho 10 26.8 ± 4.29 16 25.3 ± 3.38 18 25.2 ± 4.32
Gris Hembra 4 27.4 ± 2.86 8 26.0 ± 4.48 4 28.1 ± 4.17
Macho 5 23.4 ± 5.59 2 22.1 ± 5.92 7 26.2 ± 4.24
LF Hembra 24 26.8 ± 4.78 16 25.1 ± 3.84 17 22.3 ± 4.29
Macho 16 28.4 ± 4.42 17 25.6 ± 4.38 23 24.2 ± 3.23
Negro Hembra 22 26.4 ± 3.97 23 25.3 ± 2.85 24 28.0 ± 6.94 Macho 34 25.4 ± 3.74 24 25.4 ± 3.25 23 24.8 ± 3.24
TOTAL GENERAL 338 26.1 ± 4.23 328 25.3 ± 3.93 330 24.9 ± 4.35
Letras diferentes en la misma fila indican diferencias estadísticas significativas (P≤ 0.05)
Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•191
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
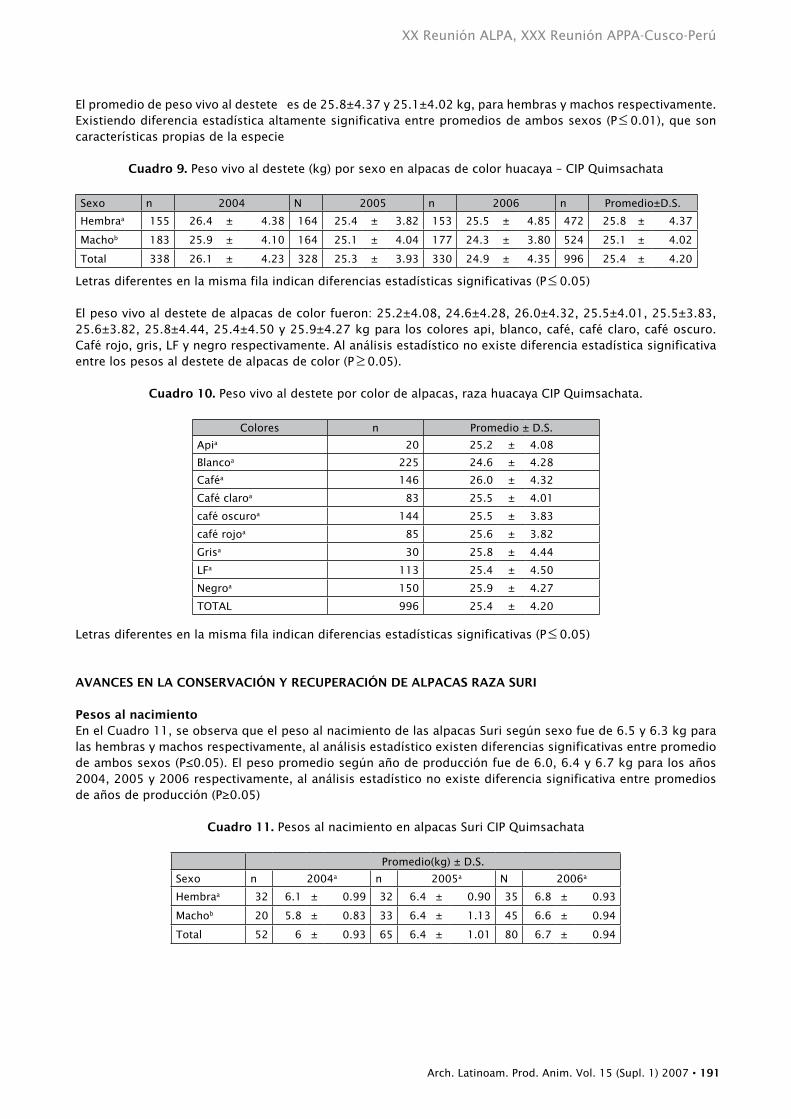
El promedio de peso vivo al destete es de 25.8±4.37 y 25.1±4.02 kg, para hembras y machos respectivamente. Existiendo diferencia estadística altamente significativa entre promedios de ambos sexos (P≤ 0.01), que son características propias de la especie
Cuadro 9. Peso vivo al destete (kg) por sexo en alpacas de color huacaya – CIP Quimsachata
Sexo n 2004 N 2005 n 2006 n Promedio±D.S.
Hembraa 155 26.4 ± 4.38 164 25.4 ± 3.82 153 25.5 ± 4.85 472 25.8 ± 4.37
Machob 183 25.9 ± 4.10 164 25.1 ± 4.04 177 24.3 ± 3.80 524 25.1 ± 4.02
Total 338 26.1 ± 4.23 328 25.3 ± 3.93 330 24.9 ± 4.35 996 25.4 ± 4.20
Letras diferentes en la misma fila indican diferencias estadísticas significativas (P≤ 0.05)
El peso vivo al destete de alpacas de color fueron: 25.2±4.08, 24.6±4.28, 26.0±4.32, 25.5±4.01, 25.5±3.83, 25.6±3.82, 25.8±4.44, 25.4±4.50 y 25.9±4.27 kg para los colores api, blanco, café, café claro, café oscuro. Café rojo, gris, LF y negro respectivamente. Al análisis estadístico no existe diferencia estadística significativa entre los pesos al destete de alpacas de color (P≥ 0.05).
Cuadro 10. Peso vivo al destete por color de alpacas, raza huacaya CIP Quimsachata.
Colores n Promedio ± D.S.
Apia 20 25.2 ± 4.08
Blancoa 225 24.6 ± 4.28
Caféa 146 26.0 ± 4.32
Café claroa 83 25.5 ± 4.01
café oscuroa 144 25.5 ± 3.83
café rojoa 85 25.6 ± 3.82
Grisa 30 25.8 ± 4.44
LFa 113 25.4 ± 4.50
Negroa 150 25.9 ± 4.27
TOTAL 996 25.4 ± 4.20
Letras diferentes en la misma fila indican diferencias estadísticas significativas (P≤ 0.05)
AVAnCES En LA COnSERVACIón y RECUPERACIón DE ALPACAS RAzA SURI
Pesos al nacimientoEn el Cuadro 11, se observa que el peso al nacimiento de las alpacas Suri según sexo fue de 6.5 y 6.3 kg para las hembras y machos respectivamente, al análisis estadístico existen diferencias significativas entre promedio de ambos sexos (P≤0.05). El peso promedio según año de producción fue de 6.0, 6.4 y 6.7 kg para los años 2004, 2005 y 2006 respectivamente, al análisis estadístico no existe diferencia significativa entre promedios de años de producción (P≥0.05)
Cuadro 11. Pesos al nacimiento en alpacas Suri CIP Quimsachata
Promedio(kg) ± D.S.
Sexo n 2004a n 2005a N 2006a
Hembraa 32 6.1 ± 0.99 32 6.4 ± 0.90 35 6.8 ± 0.93
Machob 20 5.8 ± 0.83 33 6.4 ± 1.13 45 6.6 ± 0.94
Total 52 6 ± 0.93 65 6.4 ± 1.01 80 6.7 ± 0.94
192 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
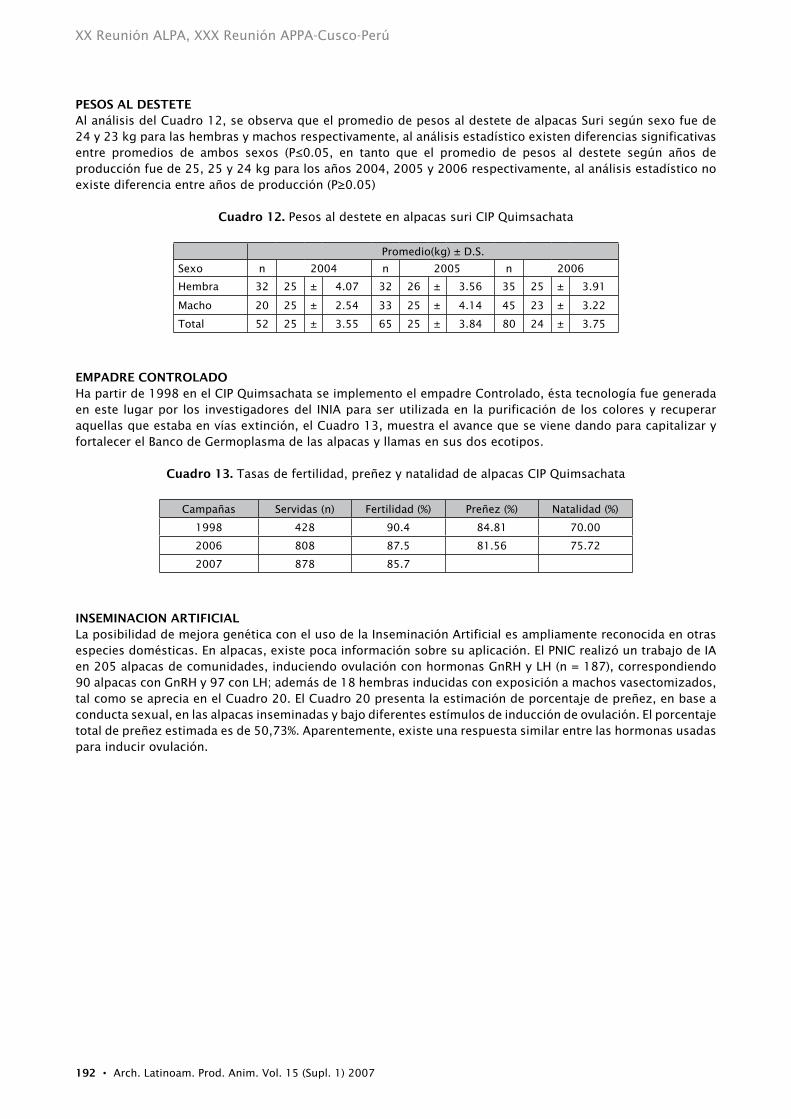
PESOS AL DEStEtEAl análisis del Cuadro 12, se observa que el promedio de pesos al destete de alpacas Suri según sexo fue de 24 y 23 kg para las hembras y machos respectivamente, al análisis estadístico existen diferencias significativas entre promedios de ambos sexos (P≤0.05, en tanto que el promedio de pesos al destete según años de producción fue de 25, 25 y 24 kg para los años 2004, 2005 y 2006 respectivamente, al análisis estadístico no existe diferencia entre años de producción (P≥0.05)
Cuadro 12. Pesos al destete en alpacas suri CIP Quimsachata
Promedio(kg) ± D.S.
Sexo n 2004 n 2005 n 2006
Hembra 32 25 ± 4.07 32 26 ± 3.56 35 25 ± 3.91
Macho 20 25 ± 2.54 33 25 ± 4.14 45 23 ± 3.22
Total 52 25 ± 3.55 65 25 ± 3.84 80 24 ± 3.75
EMPADRE COntROLADOHa partir de 1998 en el CIP Quimsachata se implemento el empadre Controlado, ésta tecnología fue generada en este lugar por los investigadores del INIA para ser utilizada en la purificación de los colores y recuperar aquellas que estaba en vías extinción, el Cuadro 13, muestra el avance que se viene dando para capitalizar y fortalecer el Banco de Germoplasma de las alpacas y llamas en sus dos ecotipos.
Cuadro 13. Tasas de fertilidad, preñez y natalidad de alpacas CIP Quimsachata
Campañas Servidas (n) Fertilidad (%) Preñez (%) Natalidad (%)
1998 428 90.4 84.81 70.00
2006 808 87.5 81.56 75.72
2007 878 85.7
InSEMInACIOn ARtIFICIALLa posibilidad de mejora genética con el uso de la Inseminación Artificial es ampliamente reconocida en otras especies domésticas. En alpacas, existe poca información sobre su aplicación. El PNIC realizó un trabajo de IA en 205 alpacas de comunidades, induciendo ovulación con hormonas GnRH y LH (n = 187), correspondiendo 90 alpacas con GnRH y 97 con LH; además de 18 hembras inducidas con exposición a machos vasectomizados, tal como se aprecia en el Cuadro 20. El Cuadro 20 presenta la estimación de porcentaje de preñez, en base a conducta sexual, en las alpacas inseminadas y bajo diferentes estímulos de inducción de ovulación. El porcentaje total de preñez estimada es de 50,73%. Aparentemente, existe una respuesta similar entre las hormonas usadas para inducir ovulación.

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•193
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Cuadro 14. Diagnóstico de preñez de las alpacas inseminadas por comunidades.
Nombre de lacomunidad
TRATAMIENTOSN° GnRH LH Machos Vasectomizados
n Vacía Preñada n Vacía Preñada N Vacía Preñada01 Chocorasi 17 07 1002 San José 19 06 13 12 10 0203 Chunguilluni 06 03 0304 Phorke 01 01 11 04 0705 Lacotuyo 08 04 04 01 0106 Sullcanaca 06 02 04 03 01 02 03 01 0207 Ayupalca 09 05 0408 Providencia 05 02 0309 Mazocruz 04 03 0110 Apopata 02 01 01 06 04 0211 Orcoyo 03 01 02 05 03 02 08 02 0612 Casana 07 05 02 01 0113 Chapilaca 04 02 02 01 0114 Chichillapi 09 08 0115 Tupala 33 16 17 07 03 04
16 Capaso 14 06 08
TOTAL 90 43 47 97 52 45 18 06 12
El protocolo sobre inseminación artificial con semen fresco desarrollado por el INIA viene siendo aplicado por la institución DESCO en condiciones de comunidades, los resultados son muy alentadores por la participación de los productores en asumir la propuesta Cuadro 15, con ello se viene demostrando de que esta alternativa puede ser utilizada por las organizaciones de los criadores e instituciones que viene promoviendo el mejoramiento genético de los camélidos domésticos
Cuadro 15. Resultados de inseminación artificial – DESCO 2007
Distrito nAlpacas inducidas
Alpacas inseminadas
Diagnostico de preñez
% fertilidad
Bilabial 3 3 3 3 100
Lampa 52 38 32 16 50
Palca 85 70 62 30 48.38
Palca 101 75 73 46 63.01
Palca 104 78 75 48 64
TOTAL 345 264 245 143 58.36
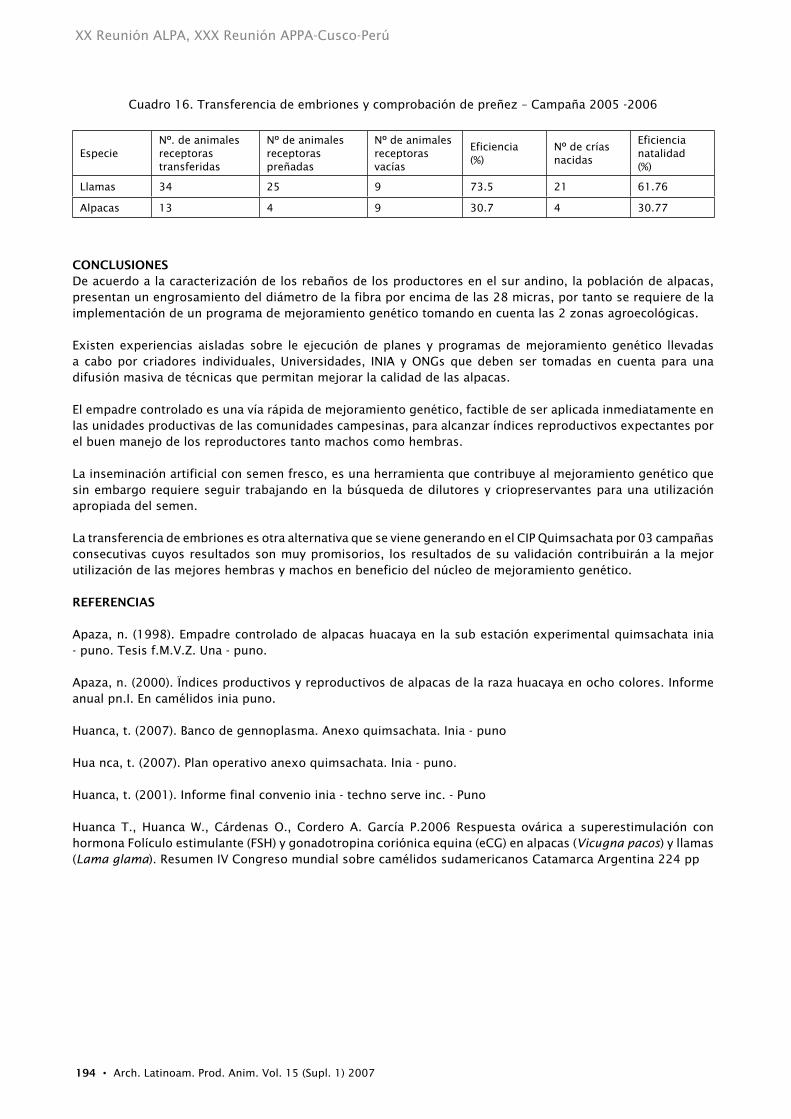
tRAnSFEREnCIA DE EMBRIOnESLos resultados obtenidos Cuadro 16, son alentadores, es así que, en el CIP Quimsachata, durante la campaña 2005, donde se realizó por primera vez una transferencia de embriones en un número representativo a nivel mundial, en 34 llamas y 13 alpacas, al diagnostico de gestación mediante ecografía de 34 llamas, resultaron preñadas 25 animales, representando el 73.5 %, de las cuales, en el año del 2006 nacieron 25 crías llamas obteniéndose una natalidad del 61.76%, en alpacas nacieron 4 crías que representa el 30.77%; las causas de estas diferencias por especie requieren mayor evaluación, estas podrían ser atribuidas entre otros factores a que el desarrollo embrionario en alpacas puede ser diferente a las llamas y ello implicaría que al existir un desarrollo embrionario más temprano hacia el estadio de blastocisto expandido no pueda ser recuperado en los lavados realizados a los 7 días post monta.
194 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Cuadro 16. Transferencia de embriones y comprobación de preñez – Campaña 2005 -2006
EspecieNº. de animales receptoras transferidas
Nº de animales receptoras preñadas
Nº de animales receptoras vacías
Eficiencia(%)
Nº de crías nacidas
Eficiencianatalidad(%)
Llamas 34 25 9 73.5 21 61.76
Alpacas 13 4 9 30.7 4 30.77
COnCLUSIOnESDe acuerdo a la caracterización de los rebaños de los productores en el sur andino, la población de alpacas, presentan un engrosamiento del diámetro de la fibra por encima de las 28 micras, por tanto se requiere de la implementación de un programa de mejoramiento genético tomando en cuenta las 2 zonas agroecológicas.
Existen experiencias aisladas sobre le ejecución de planes y programas de mejoramiento genético llevadas a cabo por criadores individuales, Universidades, INIA y ONGs que deben ser tomadas en cuenta para una difusión masiva de técnicas que permitan mejorar la calidad de las alpacas.
El empadre controlado es una vía rápida de mejoramiento genético, factible de ser aplicada inmediatamente en las unidades productivas de las comunidades campesinas, para alcanzar índices reproductivos expectantes por el buen manejo de los reproductores tanto machos como hembras.
La inseminación artificial con semen fresco, es una herramienta que contribuye al mejoramiento genético que sin embargo requiere seguir trabajando en la búsqueda de dilutores y criopreservantes para una utilización apropiada del semen.
La transferencia de embriones es otra alternativa que se viene generando en el CIP Quimsachata por 03 campañas consecutivas cuyos resultados son muy promisorios, los resultados de su validación contribuirán a la mejor utilización de las mejores hembras y machos en beneficio del núcleo de mejoramiento genético. REFEREnCIAS
Apaza, n. (1998). Empadre controlado de alpacas huacaya en la sub estación experimental quimsachata inia - puno. Tesis f.M.V.Z. Una - puno.
Apaza, n. (2000). Ïndices productivos y reproductivos de alpacas de la raza huacaya en ocho colores. Informe anual pn.I. En camélidos inia puno.
Huanca, t. (2007). Banco de gennoplasma. Anexo quimsachata. Inia - puno
Hua nca, t. (2007). Plan operativo anexo quimsachata. Inia - puno.
Huanca, t. (2001). Informe final convenio inia - techno serve inc. - Puno
Huanca T., Huanca W., Cárdenas O., Cordero A. García P.2006 Respuesta ovárica a superestimulación con hormona Folículo estimulante (FSH) y gonadotropina coriónica equina (eCG) en alpacas (vicugna pacos) y llamas (Lama glama). Resumen IV Congreso mundial sobre camélidos sudamericanos Catamarca Argentina 224 pp

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•195
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
IntRODUCCIónLa crianza de los camélidos domésticos, alpacas y llamas, es una de las actividades de mayor importancia e impacto en el desarrollo socio económico de la población alto andina de nuestro país, no solo por su capacidad de adaptación a las difíciles condiciones medioambientales, alturas sobre los 4,000 metros snm, sino por su utilización como una fuente alimenticia de proteína de origen animal y medio de transporte y en el caso de la alpaca, como un recurso para la producción de fibra de buena calidad.
El Perú tiene más de 3 millones de alpacas (87 % de la población mundial) y la segunda población mundial en llamas con más de 1 millón de animales; sin embargo, Las deficiencias en los esquemas de crianza tradicional, como la crianza conjunta de alpacas y llamas, con los consiguientes cruzamientos no programados, han contribuido a disminuir la calidad genética de los animales, originando una pérdida en la cantidad y calidad de fibra, reportándose que el 45 % de la producción de fibra tiene una finura de 26.0 micras y el 46 % una de 33.0 micras y solo un 8 % presenta una fibra del 22.0 micras (Freyre G. 2006).
La aplicación de biotecnologías reproductivas como la Inseminación Artificial (IA) ha contribuido al progreso genético obtenido en especies domesticas como los bovinos de leche, contribuyendo a obtener los actuales niveles de producción láctea. En camélidos, la posibilidad de mejora genética de los rebaños de productores mediante la prueba de progenie, con la formación de núcleos de reproductores, requiere años de trabajo y esta limitada, entre otros factores, por el
BioteCnologias reproduCtivas en Camelidos sudameriCanos domestiCos: avanCes Y perspeCtivas
Wilfredo huanca1, Aída Cordero2, teodosio huanca3 y Gregg P. Adams4
1 Laboratorio de Reproducción Animal – Facultad de Medicina Veterinaria Universidad nacional Mayor de San Marcos2 Departamento de nutrición – Facultad de zootecnia – Universidad nacional Agraria La Molina3 Programa nacional de Investigación en Camélidos – EE ILLPA – InIA – Puno4 Veterinary Biomedical Sciences, Western Collage of Veterinary Medicine – University of Saskatchewan, Saskatoon CanadaE-mail: [email protected]
largo intervalo generacional y la capacidad fisiológica de una hembra que solo puede tener hasta 4 crías, durante toda su vida reproductiva (Novoa C. 1999).
Los esfuerzos para el desarrollo y aplicación de biotecnologías reproductivas no siempre han sido desarrollados en forma programada y continua, sino como esfuerzos individuales y aislados e incluso algunos investigadores consideran que no es posible desarrollar y aplicar estas tecnologías por los pobres resultados obtenidos. Sin embargo, el desarrollo de estas biotecnologías ha requerido la mejora del conocimiento sobre la fisiología reproductiva de esta especie., contando con la ayuda de técnicas como la ultrasonografía.
El objetivo del presente trabajo es realizar una breve revisión del estado actual del conocimiento, sustentado en estudios realizados en nuestro país, especialmente en Inseminación Artificial, Transferencia de Embriones y Fertilización In Vitro; con el propósito de disponer de alternativas tecnológicas que puedan servir como herramientas para la mejora genética de los camélidos domésticos, alpacas y llamas; así como la posibilidad de su potencial uso en los camélidos no domésticos.
InSEMInACIón ARtIFICIALLa IA con semen congelado es una de las más importantes tecnologías reproductivas en la producción de animales domésticos y combinada con una prueba de progenie, ha contribuido sustancialmente en el mejoramiento genético de bovinos lecheros, especialmente cuando fue posible disponer de semen congelado de toros de alta calidad genética En camélidos, si bien existen reportes sobre el desarrollo de la IA y aun cuando puede ser considerada una alternativa tecnológica, aun no se han superado las limitantes existentes, como el desarrollo de protocolos de criopreservación, limitando por ahora al desarrollo de la Inseminación Artificial con el uso de semen fresco (Huanca y Adams 2007), con las consiguientes dificultades para permitir una amplia difusión de reproductores genéticamente superiores.
Existen diferencias fisiológicas entre los camélidos y otras especies domésticas, siendo una primera diferencia el hecho que las hembras camélidas

196 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
son especies de ovulación inducida, es decir que se requiere el estímulo de la copula para inducir ovulación (San Martín et al 1968), pero esta diferencia más que una dificultad puede ser una ventaja para la aplicación de la IA. Sin embargo, otras diferencias, como la dificultad para la colección de semen debido a las características de cópula de la hembra (posición y duración); así como el manejo del semen debido a su naturaleza viscosa; han sido los principales obstáculos para el desarrollo masivo de la IA, a los que hay que agregar la baja concentración de espermatozoides y el alto porcentaje de anormales (Fernández-Baca 1993).
Los reportes sobre métodos de colección de semen son diversos, desde el uso de sacos vaginales (Mogrovejo et al 1952), esponjas vaginales (San Martín 1961) y electroeyaculación (Fernández-Baca y Calderón 1965; Calderón W. 1968); con las consiguientes dificultades para los machos durante la copula y la calidad del semen. Posteriormente se reporta el uso de una vagina artificial adaptada de ovinos (Sumar J y Leyva V.1981), que si bien mejora la técnica de colección, aún presenta dificultades para mantener una temperatura adecuada durante el largo de la cópula. El uso de una frazadilla eléctrica cubriendo la vagina Artificial, permite algunas mejoras en la técnica de colección, facilitando el mantenimiento de la temperatura (Gauly and Leindiger , 1996). Igualmente, el uso de un maniquí (Sumar y Leyva 1981) o la colección con hembra receptiva (Gauly and Leindiger 1996; Huanca y Gauly 2001) se presentan como alternativas para la colección de una muestra de semen fisiológicamente normal; sin embargo, se requiere un entrenamiento de los animales y no siempre todos los machos llegan a aceptar el maniquí; mientras que el uso de hembra receptiva genera incomodidades en el operador. Otras alternativas de colección incluyen la colección de semen mediante una fístula uretral (Pérez G. 2006) y una más reciente ha sido reportada por Giulano et al (2007) mediante la colección de semen con uso de electroeyaculación pero con una previa anestesia del animal, con resultados interesantes y sin contaminación del semen con orina.
Los estudios sobre conservación de semen no son totalmente satisfactorios, particularidades como la alta viscosidad del semen de alpacas y llamas (Lichtenwalner et al 1996; Bravo et al 1997) se constituyen en factores que dificultan el desarrollo de las técnicas de conservación., dificultando la determinación de la concentración, morfología y motilidad espermática, además del alto porcentaje de espermatozoides anormales (Fernández- Baca 1993) y una motilidad oscilatoria (Sumar y García 1986, Garnica et al 1993). Intentos para reducir la alta viscosidad ha sido reportada, mediante con el uso de enzimas como la colagenasa, hyaluronidasa y tripsina (Bravo W. et 2000) o mediante una acción mecánica (Valdivia M.
1999). Los resultados observados son variables y si bien con el uso de la enzima se observa motilidad espermática, esta característica no siempre se ha reflejado en mejoras en la tasa de preñez. Igualmente existe poca información sobre el uso de dilutores, reportándose el uso de Yema – citrato (Pacheco et al 1996), solución de Suero de Albúmina Bovina (BSA) y Glucosa (Huanca y Gauly 2001), Tris – Glucosa - Yema Huevo (Raymundo et al 2000).
El primer reporte sobre IA en alpacas fue realizado por Fernández-Baca y Novoa (1968), utilizando semen sin diluir de 2 vicuñas y 4 paco-vicuñas, obteniéndose una sola cría de 42 alpacas inseminadas. Posteriormente se ha reportado la inseminación de 83 alpacas y 11 llamas con semen fresco obtenido por electroeyaculación de una vicuña y 4 paco-vicuñas, con una tasa de 48 % en hembras inducidas a ovulación con Gonadotropica Corionica Humana (hCG) y 11 % en hembras inducidas a ovulación con machos vasectomizados (Leyva et al 1977). Igualmente se reporta una tasa de preñez del 73 % a la IA con semen fresco y depositado en los cuernos uterinos, así como un 67 % de preñez a la IA por laparoscopia (Bravo et al 1997), similar al reporte de Pacheco (1996). Sin embargo, todas esas experiencias fueron realizadas en centros experimentales, a diferencia de una experiencia a nivel de criadores particulares, donde se reporta una tasa de preñez del 51 % de 207 alpacas inseminadas con semen fresco diluido con una solución de BSA + Glucosa e induciendo la ovulación con un análogo de GnRH o LH, entre 24 a 26 horas antes de la Inseminación (Apaza et al 2001).
Los reportes sobre semen congelado son muy escasos; así tenemos que un estudio realizado con semen obtenido por electroeyaculación y diluido con Tris – Yema huevo – Glicerol, solo se observo un 10 % de motilidad pos descongelamiento (McEvoy et al 1992); así mismo, otro estudio con semen tratado con colagenasa y posteriormente diluido con citrato de sodio – Yema huevo – Glicerol (7 %) permite obtener una motilidad entre el 30 – 40 % (Bravo 1996). El uso de etilinglicol como agente crioprotector ha sido reportado, permitiendo una tasa de motilidad pos descongelamiento del 20 % (Santiani et al 2005). A pesar de estos resultados a la fecha no se ha reportado estudios que nos permitan señalar la factibilidad de congelar semen de camélidos. Posiblemente, como sucede con otras especies, no va a ser fácil congelar semen, más aún si a las dificultades de contar con un dilutor apropiado, hay que considerar los altos porcentajes de anormalidades presentes en los eyaculados.
Los resultados obtenidos sugieren que es posible la colección de semen con vagina artificial colocada en un maniquí y envuelta con una frazadilla eléctrica,

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•197
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
así como el desarrollo de la IA con semen fresco; sin embargo se requieren nuevos estudios que permitan desarrollo protocolos de criopreservación de semen, por lo que parece que el uso de la IA en camélidos va a estar, por ahora, restringido al uso de semen fresco diluido y utilizado inmediatamente, con resultados cercanos al 50 %.
tRAnSFEREnCIA DE EMBRIOnESEn camélidos, el primer reporte sobre transferencia de embriones fue realizado por Sumar et al. (1974), posteriormente otros reportes confirman la factibilidad de la aplicación de la técnica pero con una variabilidad en la respuesta ovárica a los protocolos de superestimulacion, así como la respuesta en número y calidad de embriones recuperados (Del Campo et al 1995).
La ultrasonografía ha contribuido a mejorar el conocimiento sobre la fisiología ovárica en especies domésticas ((Pierson and Ginther 1984) y no domésticas; contribuyendo al conocimiento de la dinámica folicular ovárica en llamas (Adams et al 1990) y alpacas (J. Vaughan 2001). La diferencia sustancial, como ocurre en las especies de ovulación inducida, que si no ocurre copula, el folículo dominante ingresa a una fase de regresión y se produce una nueva onda de crecimiento folicular (Sumar J. 2000).
La ovulación en alpacas y llamas puede ser inducida por la administración de hormonas exógenos como la Gonadotropina Corionica Humana (hCG), Hormona Liberadora de las Gonadotropina (GnRH) (Bourke et al 1995) o Hormona Luetinizante (LH) (Huanca et al 2001). La ovulación ocurre alrededor de las 30 horas después de la aplicación de GnRH, LH o por estimulo de monta (Ratto et al 2006, Huanca et al 2001).
Una primera etapa en el desarrollo de protocolos de Ovulación Múltiple y Transferencia de Embriones (MOET) involucra la superestimulación ovárica, con el propósito de inducir el crecimiento, maduración y ovulación de un gran número de folículos. Los protocolos de estimulación ovárica aplicados en camélidos incluyen el uso de Hormona Folículo Estimulante (FSH) o Gonadotropina Corionica Equina (eCG), durante una fase luteal inducida con la aplicación de Hormonas hipotalámicas (GnRH); fase luteal artificial con la aplicación de progestágenos exógenos y durante una fase de receptividad sexual (Bourke et. al 1992; Correa et al 1994). Las respuestas obtenidas son variables, alta incidencia de folículos luteinizados, baja tasa de recuperación embrionaria (0 – 2.3 embriones/ donadora) y de calidad variable (Del Campo M. et al 1995).
Los estudios orientados a la inhibición del desarrollo folicular con tratamientos con progesterona (Leyva
y Garcia 1999), señalan que ante la inserción de un progestageno intravaginal en llamas a cualquier momento del estadio folicular inhibe el crecimiento folicular y si al final del tratamiento se aplica 500 UI de eCG, se obtiene 5.2 ± 2.5 folículos y 2.1 ± 2.9 embriones recuperados (Chaves et al 2002). Similares resultados han sido reportados por otros autores. En alpacas, se realizo un estudio con la aplicación de un progestágeno intravaginal y la administración de 1000 UI de eCG, obteniéndose una alta respuesta ovárica con el numero de Cuerpos lúteos (Velásquez y Novoa 1999). Estudios realizados por nuestro grupo de investigación sugieren que es factible obtener una adecuada respuesta ovárica en alpacas y llamas, cuando los animales son sometidos previamente a un protocolo de sincronización de onda folicular y el estimulo hormonal se inicial durante la emergencia de las ondas de crecimiento folicular, sustentado en el reporte de Ratto et al (2003).
Referente a la hormona a utilizar, se ha reportado un estudio para evaluar la respuesta de ambas hormonas, FSH y eCG, en llamas, habiéndose determinado que el número de folículos 6 mm sometidas a un protocolo de estimulación ovárica fue de 17.9 ± 2.2 al tratamiento con FSH y 17.7 ± 2.2 folículos con eCG (Ratto et al 2005). Estos resultados permitieron obtener evidencia que existe una buena respuesta ovárica a la acción de ambas hormonas, sin diferencias entre ambas. Alpacas y llamas sometidas a un protocolo hormonal de estimulación ovárica con eCG, iniciando el tratamiento durante la emergencia de las ondas de crecimiento folicular, permite obtener el desarrollo de 12.8 ± 1.4 folículos y 8.1 ± 1.0 cuerpos Lúteo en llamas y 7.5 ± 1.2 folículos y 5.9 ± 1.3 cuerpos lúteo en alpacas (Huanca et al 2006)
La recuperación embrionaria puede ser realizada vía quirúrgica o no quirúrgica. El primer reporte sobre colección de embriones fue realizado de oviductos de alpaca, previa laparotomía (Novoa y Sumar, 1968) y la primera cría nacida por transferencia fue reportada por Sumar J. (1974). Posteriormente se reporta la colección de embriones mediante una técnica no quirúrgica a los 7 días en llamas y la transferencia en fresco con el nacimiento de una cría (Wiepz and Chapman 1985) Otros estudios confirman que la recuperación de embriones en estadio de blastocisto puede ser realizada a los 7 días pos servicio mediante técnicas no quirúrgicas en llamas y alpacas. La técnica de recuperación no quirúrgica utilizada en camélidos es similar a la reportada en vacunos, mediante la inclusión de una solución de Dulbecco PBS y los embriones recuperados observados y evaluados en un estereoscopio (Huanca et al 2004).
Resultados sobre recuperación y transferencia de embriones en llamas han sido descritos por Huanca

198 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
et al (2006) señalando la recuperación de 4.8 ± 0.9 embriones, con una tasa de recuperación 66,1 % del número posible de embriones, determinado en base al número de cuerpos lúteos. La tasa de preñez fue del 68.9 %. En alpacas, se recuperaron 1.6 ± 0.3, respuesta diferente de la observada en llamas, con una tasa de recuperación del 23.6 % de embriones posibles, con una tasa de preñez del 30.0 %. Igualmente, se ha reportado la recuperación de 37 embriones de 47 hembras no estimuladas (79 %), de las cuales resultaron en un 41 % de preñez al ser transferidas a receptoras (Taylor et al 2001).
La recuperación de las hembras donadoras fue evaluada, habiéndose observado una recuperación del tracto reproductivo, determinado por ecografía, a las 3 semanas posteriores al lavado uterino y una tasa de preñez luego de ser servidas, del 40 %, con un solo servicio (Huanca et al 2006). Esta observación permite plantear la posibilidad de realizar tratamientos de superestimualción y recuperación embrionaria al inicio de la época de empadre y luego de un descanso, realizar el servicio de las donadoras y obtener tasas de preñez similares a las observadas bajo condiciones de empadre continuo.
Estos resultados sugieren la posibilidad de obtener resultados satisfactorios con los protocolos de superestimulación ovárica en alpacas y llamas, permitiendo obtener embriones de buena calidad y viabilidad y que al ser transferidos permiten obtener tasas de preñez sobre el 50 %. Igualmente, la rápida recuperación de las hembras donadoras, permite señalar que una vez realizado el protocolo de estimulación hormonal, las hembras requieren un descanso aproximado de un mes, con tasas de preñez muy similares a las obtenidas bajo condiciones de crianza de los productores.
FERtILIzACIOn In VItROLa técnica de Fertilización In Vitro (FIV) se presenta como una de las tecnologías que mayor desarrollo esta experimentando en los últimos tiempos. La colección de ovocitos del folículo ovárico para la posterior maduración nuclear y citoplasmática, es la primera fase en el desarrollo de esta técnica (Miragaya et al 2006). En camélidos existe escasa información referida a la colección de ovocitos de ovarios de mataderos y madurados In Vitro. Un estudio realizado con ovocitos obtenidos de ovarios de matadero y madurados a 38 °C y 5 %de CO2 , en los que se utilizaron medios de maduración de uso en bovinos, reporta una alta tasa de ovocitos en MII (62 %) a las 32 horas de incubación (Del Campo et al 1992).
La obtención de ovocitos puede ser realizada de ovarios procedentes de mataderos o por aspiración transvaginal. En el primer caso, los mataderos no
siempre se encuentran cerca a los laboratorio, por lo que en una primera fase se evaluaron las diferencias en la calidad de ovocitos obtenidos de ovarios de alpacas y transportados desde el matadero, bajo dos diferentes temperaturas (4° y 35 ° C), los resultados sugieren una mejor calidad de los ovocitos procedentes de ovarios transportados a 35 °C , respecto a los 4°C, (Huanca et al 2007).En el segundo caso, colección de ovocitos vía transvaginal, Brogliatti et al (2000) utilizando un transductor transvaginal de 7.5 MHZ y aplicado vía transvaginal reporta la colección de 76 COCs de 134 folículos (57 %), mientras que la recuperación de ovocitos via transvaginal en llamas, sometidas a estimulo hormonal permitió la recuperación de 10.7±2.1 y 11.2 ± 2.3 COCs (71 y 74 % ) (Ratto et al 2005). En alpacas, un ensayo preliminar señala la factibilidad de recuperar ovocitos vía transvaginal (Huanca et al 2006) y aún cuando se requieren mas investigación, estos resultados sugieren que es posible recuperar ovocitos vía transvaginal sin originar daños mayores en llamas y alpacas.
Miragaya et al (2002) estudio la maduración In Vitro de ovocitos, obtenidos por aspiración quirúrgica a las 22 horas después de la administración de un análogo de GnRH, en llamas estimuladas con un progestageno y estimuladas con 500 UI de eCG,obteniendo una maduración de 62 %, similar al reporte de Del Campo et al (1992). Estudios sobre Fertilización In Vitro señalan una tasa de desarrollo al estadio de pro núcleo del 29.2 y 57.1 % con dos diferentes dosis de heparina (2 y 5 ug/ml), pero solo una tasa de desarrollo del 5.6 %, 6.0 % y 4.7 % para los estadios de morula, blastocisto temprano y blastocisto expandido, luego de 9 días de cultivo (Del Campo et al 1994).
Los resultados obtenidos señalan que el desarrollo de protocolos de FIV deberá ser una de las prioridades de investigación, en forma sistemática y conjuntamente con la Transferencia de embriones permitirán contribuir a diseñar alternativas tecnológicas para los programas de mejoramiento genético en los camélidos domésticos y en base a los resultados obtenidos, evaluar la posibilidad de su aplicación en camélidos no domésticos.
COnCLUSIónLos resultados obtenidos a la fecha señalan que el desarrollo de la IA esta limitada al uso de semen fresco y se requiere estudios que permitan desarrollar protocolos orientados a obtener semen congelado, con resultados expresados en tasa de preñez similares a los obtenidos con semen fresco. En Transferencia de Embriones se han establecido protocolos de superovulación que permiten obtener un número alrededor de 5 embriones viables/ donadora y con una tasa de preñez mayor al 40 %. De otro lado, los estudios señalan que la recuperación de las hembras

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•199
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
donadoras se produce en un tiempo no mayor a los 40 días, permitiendo una tasa de preñez, al ser expuestas al macho, similar a las reportadas en las condiciones de campo. Los estudios iníciales sobre Fertilización In Vitro nos permiten sugerir lo promisorio de esta técnica y que contribuirá, conjuntamente con la transferencia de embriones, al progreso genético de los camélidos domésticos y como una primera fase para la aplicación de tecnologías reproductivas en los camélidos no domésticos.
AGRADECIMIEntOSLos autores expresan su agradecimiento por el financiamiento recibido del Concejo Nacional de Ciencia y Tecnología (CONCYTEC), (Proyecto PROCOM 166 – 2006) y al Proyecto INCAGRO - Subproyecto FDSE Biotecnologías reproductivas 05-0015 - INCAGRO FDSE – UNMSM.
REFEREnCIAS
Adams GP, Sumar J., Ginther OJ. 1990. Effect of lactational and reproductive status on ovarian follicular waves in llamas. J. Reprod. Fertil. 90: 535-545.
Adams GP and Ratto MH 2001. Reproductive Biotechnology in south American camelids. Rev Inv. Vet Perú. Suppl. 1: 134 -141.
Apaza N, Sapana R, Huanca T y Huanca W. 2001. Inseminación Artificial en alpacas con semen frescoen comunidades campesinas. Rev. Invest. Vet. Perú. Suppl. 1: 435 - 438.
Bourke SA, Kyle CE, McEvoy TG, Young O, Adam CL. 1995. Superovulatory responses to eCG in llamas (Lama glama). Theriogenology 44, 255-268.
Bourke D., Adam C. and Kyle C., Young P., and Mc Evoy TG. 1992. Ovulation, superovulation and embryo recovery in llamas. Procc. 12th Congress Animal Reprod. 1:193-195.
Bravo W.; Sidkmore JA; Zhao X. 2000. Reproductive aspects and storage of semen camelids. Anim.Reprod. Sci. 62: 173-193.
Bravo W, Flores U, Garnica J, Ordoñez C 1997. Collection of semen and artificial insemination of alpacas. Theriogenology 47: 619-626.
Calderón W., Sumar J., Franco E. 1968. Avances en la Inseminación Artificial de la Alpaca (Lama paco). Vol 2. Lima – Perú, Facultad de Medicina Veterinaria – Universidad Nacional Mayor de San Marcos.pp 19 -25.
Correa J.,Ratto MH, and Gatica R. 1994. Actividad estral y respuesta ovárica en alpacas y llamas tratadas
con progesterona y gonadotropinas. Arch.Med.Vet.26:59–64.
Chaves MG, Aba MA, Agüero A, Egey J, Berestin V, Rutter B. 2002. Ovarian follicular wave pattern and the effect of exogenous progesterone on follicular activity in non-mated llamas. Anim. Reprod. Sci. 69, 37-46.
Del Campo MR, Donoso MX, Del Campo CH, Rojo R, Barros C, Parrish JJ, Mapletoft RJ 1992. In Vitro maduration of llama (Lama glama) oocytes IN: Procc. 12th International Congress on Animal Reproduction. Vol 1 pp 324-326.
Del Campo MR,Del Campo CH, Donoso MX, Berland M, Mapletoft RJ. 1994. In vitro fertilization and development of llama (Lama glama ) oocytes using epididymal spermatozoa and oviductal cell co-culture. Theriogenology. 41(6):1219-29.
Del Campo MR., Del Campo CH., Adams GP et al. 1995.The application of new reproductive technologies to south American camelids. Theriogenology, 43: 21 – 30.
Fernández-Baca S.1993. Manipulation of reproductive functions in male and female New World camelids. Anim Reprod. Sci. 33:307-323.
Fernández-Baca S. Y Calderón W. 1965. Métodos de colección de semen de la alpaca. Rev. IVITA – Facultad de Medicina Veterinaria, Universidad Nacional Mayor de San Marcos.
Freyre G 2006. Experiencias de Transformación y Comercialización de la fibra de alpacas. Conferencia Internacional de Camélidos Sudamericanos. 30-31 de Marzo, Arequipa – Perú.
Gomez C, Ratto MH, Berland M, Wolter M, Adams GP 2002. Superstimulatory response and oocyte collection in alpacas. Theriogenology 57, 584. Abst.
Gauly M and Leindinger H. 1995. Semen quality characteristics , volume distribution and hypo-osmoitic sensitivity of spermatozoa of Lama glama and Lama guanicoe. In Gerken M, Renieri C. Eds: Procc. 2nd European Symposium on South American Camelids. Universita Degli Studi Di Camerino. Pp 235-244.
Garnica J, Flores E, Bravo W. 1995. Citric Acid and Fructuose concentration in seminal plasma of alpacas. Small ruminants Res 18: 95-98.
Giuliano S., Director A., Gambarotta M, Trasorras V., Miragaya M. 2007. Collection method, season and individual variation on seminal characteristics in the llama (lama glama). Anim. Reprod. Sci. Feb. 27.

200 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Huanca W, Cardenas O, Olazabal C, Ratto M, Adams GP 2001. Efecto hormonal y empadre sobre el intervalo a la ovulación en llamas. Revista de Investigaciones Veterinarias del Peru. 1: 462-463.
Huanca W., M.Ratto, A. Santiani, A, Cordero and T. Huanca. 2004. Embryo Transfer in camelids: Study of a reliable superovulatory treatment in llamas. 4th European Symposium on South American Camelids and DECAMA European Seminar, Gottingen, 7 – 9 October, 2004, Germany. Abstracts. Ed. M. Gerken, C. Renieri, M. Gauly and A. Riek.
Huanca W and Adams GP. 2007. Semen Collection and Artificial Insemination in Llamas and Alpacas. En: Current Therapy in Large Animal Theriogenology, Youngquist R and Threlfall W. 2° Edition Saunders. Elsevier Inc. pp 869-873.
Huanca W. y Gauly M. 2001. Conservación de semen refrigerado de llamas. Rev. Inv. Vet. Perú Suppl 1: 460-461.
Huanca W, Ratto M, Vásquez M, Cervantes M, Cordero A, Huanca T and Adams GP. 2006. Fertilización In Vitro en camelids: Recuperación de ovocitos via transvaginal. Resumen XXIX Reunión Asociación Peruana de Producción Animal – Huancayo, Perú.
Huanca W, Gonzalez M, Cordero A, Huanca T. 2006. Comportamiento reproductivo de donadoras de embriones, después de un protocolo de superovulación en llamas. Resumen V Congreso Mundial de Camélidos, Catamarca – Argentina.
Huanca W, Ratto M, Cordero A, Santiani A, Huanca T, Cárdenas O, Adams GP. 2006. Respuesta ovárica y transferencia de embriones en alpacas y llamas en la zona altoandina del Perú. Resumen del V Congreso Mundial de Camélidos, Catamarca – Argentina.
Huanca W, Palomino J, Cervantes M, Cordero A, Huanca T. 2007. Efecto de temperatura de transporte (35° y 4°C) sobre la calidad morfológica de ovocitos colectados desde ovarios de camal. Procc. XX Reunión ALPA y XXX Reunión APPA- Cusco, Perú.
Leyva V. Y García W. 1999. Efecto de la progesterona exogena sobre la función del cuerpo lúteo de alpacas. Resumen del II Congreso Mundial de Camelidos–pp 87, Cuzco.
Lichtenwalner AB, Woods GL, Weber JA, 1996. Eyaculatory patterns of llama during copulation . Theriogenology 46: 285-291.
McEvoy TG, Kyle CE, Slater D. 1992. Collection, evaluation and cryopreservation of llama semen. J. Reprod. Fert. 9: 48 (Abstract)
Miragaya M, Chaves MG, Capdevielle EF, Ferrer MS, Pinto MR, Rutter B.,Neild DM. 2002. In Vitro maduration of llama (Lama glama) oocytes obtained surgically using follicle aspiration. Theriogenology 57 (1), 731.
Miragaya M, Chavez MG, Agüero A. 2006. Reproductive biotechnology in South American camelids. Small Ruminant Research 61: 299-310.
Mogrovejo D. Estudios del semen de la alpaca. 1952. BS Thesis, Facultad de Medicina Veterinaria, Universidad Nacional Mayor de San Marcos, Lima Perú, 21 pp.
Novoa C. Sumar J. 1968. Colección de huevos in vivo y ensayo de transferencia en alpacas. Boletín extraordinario IVITA – UNMSM, Lima, Perú, 3: 31 – 34.
Novoa C., E. Franco; W. García; D.Pezo. 1999. Dosis de Gonadotropinas (eCG y hCG), superovulación y obtención de embriones en alpacas. RIVEP.Perú 10 (1):48-53.
Pierson RA, Ginther OJ. 1984. Ultrasonography of the bovine ovary. Theriogenology 21, 495-504.
Pacheco C. 1996. Efecto de la tripsina y colagenasa sobre el acrosoma del espermatozoide y su relación con la fertilidad del semen de alpaca. 1996. Tesis MVZ Programa de Medicina Veterinaria y Zootecnia, Universidad Catolica de Santa María, Arequipa, Perú. 52 pp.
Ratto M.H., Singh J., Huanca W., Adams G.P. 2003. Ovarian follicular wave synchronization and pregnancy rate after fixed-time natural mating in llamas.Theriogenology 60, 1645-1656.
Ratto MH, Berland M, Huanca W, Singh j, Adams GP. 2005. In Vitro and In Vivo maturation of llama oocytes. Theriogenology 63, 2445 –2457.
Ratto M, Huanca W, Singh J, Adams GP. 2006. Comparison of the effect of natural mating, LH and GnRH on interval to ovulation and luteal function in llamas. Anim. Reprod. Sci. 91 (3-4): 299-306.
Raymundo F, Huanca W, Huertas S, Gauly M. 2000. Influence of diferents extenders on the motility in alpaca (Lama paco) semen. Procc. 2nd International Camelids Conference on Agroeconomics of Camelids Farming. Almaty, Kazakstan, p 79 (Abstract)
San Martín M, Copaira M, Zuñoga J., Rodríguez R., Bustinza G. Acosta L. 1968. Aspects of reproduction in the alpaca. J. Reprod. Fertility 16, 395-399.
San Martín M. 1961. Fisiología de la reproducción de la alpaca, Anim. Symp. Sobre problemas ganaderos. Lima Peru, pp 121- 131.
Sumar J y Leyva V. 1981. Colección de semen mediante

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•201
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
la vagina artificial en alpaca. Proceeding of the IVth International Conference on South America Camelids. Punta Arenas, Chile, 12 pp.
Sumar J. 2000. Llamas an Alpacas.IN: Reproduction in farm Animals. Seventh Edition. E.S.E. Hafez, B. Hafez
Sumar J. Y Franco E. 1974. Ensayos de Transferencia de Embriones en Camélidos Sudamericanos. IN: Informe Final (IVITA) UNMSM Lima, Perú.
Santiani A, Huanca W, Sapana R, Huanca T, Sepulveda N, Sánchez R. 2005. Effects of the quality of frozen-thawed alpaca (Lama paco) semen using two different cryoprotectants and extenders. Asian J. Androl. 7(3) : 303-309.
Taylor S., Taylor PJ, James AN., Godke R. 2000. Successful commercial embryo transfer in the Llama (Lama glama). Theriogenology. 53, 1, 344.
Vaughan J.L. 2004. Artificial Breeding in alpacas. 4th European Symposium on South American Camelids and DECAMA European Seminar, Gottingen, 7 – 9 October, 2004, Germany. Abstracts. Ed. M. Gerken, C. Renieri, M. Gauly and A. Riek.
Vaughan JL. 2001. Control of Follicular waves in alpacas. Rev. Inv. Vet. Peru: Suplem. 1:112 – 114. Lima – Perú.
Wilson Wiepz D;Chapman RJ. 1985. Non surgical embryo transfer and live birth in a lama. Theriogenology. 24 (2) : 251 – 257.
Valdivia M,Ruiz M, Bermúdez L, Quinteros S, Gonzales A, Manosalva I, Ponce C, Olazábal J, Dávalos R. 1999. Criopreservacion de semen de alpacas. Resumen. II congreso Mundial sobre camelidos, Cusco, Peru 81abstract.

202 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
CaraCteriZaCiÓn histolÓgiCa de las memBranas útero-plaCentarias
Luis Olivera Marocho*
* Universidad nacional del Altiplano - [email protected]
La biología de la placenta de alpaca muestra la interacción de estructuras materno-embrionario-fetales, que son responsables para mantener la estabilidad y viabilidad del nuevo ser, desde su concepción hasta el nacimiento. Es un sistema biológico que obedece a cambios hormonales y moleculares cuya expresión es visualizada en la morfodinámica de los elementos que lo componen. La placenta de alpaca debe desarrollar una alta capacidad para transportar nutrientes desde el lado materno al fetal y metabolitos desde el fetal al materno.
1.- CARACtERIzACIOn DEL UtERO DE ALPACAS nO PREÑADASEl útero de alpaca es bicorne, en las no preñadas la superficie endometrial muestra leves sinuosidades, es revestido por epitelio simple cilíndrico, con núcleos situados en la región basal; las células epiteliales están adheridas lateralmente por uniones complejas, en su superficie apical son observados numerosos y cortos microvillos. En el citoplasma de todas las células, mitocondrias esféricas y alargadas y polirribosomas están distribuidos por todo el citoplasma, estando más concentrado en las regiones basal y apical de la célula. En la fase luteal la región apical es más acidófila. Las glándulas uterinas son tubulares simples, ligeramente sinuosas y están presentes en toda la espesura del endometrio; son conformadas por epitelio simple cilíndrico, siendo mas altas próximas al epitelio uterino que en el resto del endometrio. La superficie de las células glandulares muestra microvillos más largos que las células epiteliales y algunas presentan cilios. Las mitocondrias son mas frecuentes en la región apical de la célula. La presencia de estas glándulas en fase luteal es mas frecuente y de mayor tortuosidad y ocasionalmente es observado entre los haces de músculo liso del miométrio. El estroma endometrial próximo al epitelio es más celularizado, semejante a la capa compacta presente en el endometrio de otros mamíferos, conformado en su mayoría por fibroblastos, frecuentes macrófagos, plasmocitos y eventuales leucocitos. Esta densidad
Histological Characterization utero- placentaries membranes of the alpaca
celular se incrementa en la fase luteal y la distribución de los macrófagos ocupan una región distante del epitelio y conteniendo en su citoplasma material fagocitado PAS+. Próximo al miométrio, el estroma es menos celularizado; el citoplasma de los fibroblastos exhibe retículo endoplasmático granular y reducido número de mitocondrias. La matriz extracelular es amorfa con haces de fibrillas de colágeno (Olivera, 2003a). Una evidente red capilar está presente en el estrato compacto y particularmente próximo al epitelio. Las arterias que originan esta red capilar presentan un trayecto tortuoso (Valencia, 2007).
2.-IMPLAntACIón EMBRIOnARIAA pesar de la universalidad del proceso, la implantación embrionaria no envuelve los mismos mecanismos, tampoco es un evento que se establezca en el mismo momento para las diferentes especies de mamíferos. Por lo que es difícil comprender en su generalidad, los avances en el esclarecimiento de este proceso. Solo es posible cuando la secuencia de eventos es explorada separadamente en cada una de las especies.
El proceso de implantación es un estado de transición en el progreso de la gestación, durante el cual, el blastocisto asume una posición fija e inicia una interrelación fisiológica con el útero. Para garantizar este proceso, en camélidos es necesaria la presencia del cuerpo lúteo (Sumar, 1988; Fernández-Baca et al, 1979). La localización del embrión es en el cuerno uterino izquierdo, siendo similar en el camello y dromedario (Skidmore et al, 1996; Elwishy, 1987). Independiente del fijador utilizado, la interface de muestras útero-embrionarias hasta los 30 días de la preñez (ddp) no resisten al proceso histológico, fácilmente se separaban. El trofoblasto esta formado por una única capa de células, apreciándose frecuentes figuras de mitosis, células binucleadas y células poliplóides. Estas células son acompañadas por células mesenquimales que forman una capa única y continua. El trofoblasto y las mesenquimales dan origen al corion.
Hasta el primer mes de preñez, las características estructurales del trofoblasto muestran núcleo eucromático, en el citoplasma existe gran cantidad

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•203
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
de mitocondrias con crestas paralelas, cisternas del retículo endoplasmático granular y del aparato de Golgi además polirribosomas y ribosomas libre, filamentos intermediarios y acúmulos ocasionales de filamentos finos están distribuidas por toda la célula. Llama la atención en el citoplasma, la presencia de grandes cuerpos vesiculosos revestidos por membrana, de formas variadas y diferentes grados de electrodensidades. El epitelio uterino se presenta con diferentes aspectos, en lugares donde no existe contacto con el trofoblasto son cilíndricas altas, mientras en áreas de contacto, las células epiteliales son tanto altas cuanto bajas. Su ultra-estructura muestra organelas compatibles con células de intensa actividad, además se observa discretos depósitos de glucógeno; en la región apical esta presente numerosos y pequeños gránulos revestidos por membrana, conteniendo material de acentuada electrodensidad; la superficie apical de estas células es irregular presentando cortas proyecciones. Las glándulas muestran un pleomorfismo nuclear, depósito de glucógeno y vesículas electrodensas de reacción positiva para PAS+ y fosfatasa ácida. El estrato compacto del estroma endometrial aumenta en densidad celular, favorecido por la mayor red capilar y presencia de vénulas dilatadas. La interface trofoblasto-epitelio uterino muestra una completa yuxtaposición de superficies de ambos organismos, en la cual las proyecciones de la superficie materna se encajan en depresiones de la superficie embrionaria. Entre ambas superficies exhibe presencia de material PAS+ y diferentes respuestas a lectinas (Jones et al, 2002), de homogénea electrodensidad; en la superficie de contacto, se observa una gradual interdigitación de membranas, donde cortos microvillos del epitelio uterino se complementan con las del trofoblasto, además es posible observar la formación de vesículas en el citoplasma apical de las células trofoblásticas y sugiere un pasaje de material del medio extracelular para el interior de estas células. Estas graduales modificaciones en la interface materno-embrionaria son características de las fases de aproximación, contacto y adhesión del proceso de implantación embrionaria (Olivera et al, 2003a).
Entre los 30 a 45 ddp, numerosos gránulos secretorios son observados en el epitelio uterino, mientras que células gigantes multinucleadas aparecen entre las células trofoblásticas mostrando intensa actividad a la fosfatasa alcalina y gránulos PAS+. Por el día 45 entre el trofoblasto y el conectivo extraembrionario es evidente una vascularización que es indicativo para la formación de la placenta. Los capilares fetales y materno indentan el epitelio y el trofoblasto, reduciendo en espesura a ambos tipos celulares y formando áreas especializadas de intercambio biomolecular a lo largo de la superficie de contacto.
3.-PLACEntACIOnDesde los 60 a los 347 ddp la superficie de la placenta en contacto con el feto se muestra lisa, siendo lo contrario en la superficie del trofoblasto relacionado al organismo materno, observándose proyecciones esféricas o convexas, que según el avance de la preñez se ramifican y se proyectan al lado materno, alcanzando regiones profundas del endometrio. En la región central de estas proyecciones se aprecia tejido mesenquimal e indentando a las células trofoblásticas existe una red de capilar. Las células gigantes trofoblásticas ocupan mayormente la porción apical de las proyecciones que aumentan en número hasta el final de la preñez. Estas células muestran un elevado contenido de DNA evidenciado por la reacción de Feulgen y el número de regiones organizadores nucleolares (Ag NORs) varian de 15 a 100, mientras que en las células trofoblásticas mononucleares presentan uno o dos AgNORs; no existe citocinesis por lo que estas células gigantes evidencian una endoreduplicación o mitosis incompleta lo que caracteriza a una poliploidización mitótica (Klisch et al 2006).
La superficie uterina muestra depresiones esféricas en diferentes grados de desarrollo donde las proyecciones del trofoblasto hacen correspondencia. El epitelio uterino se presenta sobre la forma de un epitelio simple plano mostrando actividad de síntesis celular y próximo a la interface materno-fetal exhibe una conjunción de material electrodenso conformado por haces de microfilamentos; en la región apical de estas células hubo una disminución de vesículas, lo cual es indicativo de una rápida transferencia de biomoléculas al lado fetal. La delgada espesura del citoplasma uterino y del trofoblasto y las finas paredes de la célula endotelial forman la barrera placentaria. Se observa tanto en el trofoblasto y epitelio uterino procesos de lisis celular y mitosis.
La interface materno-fetal de reacción positiva a la fosfatasa alcalina es evidenciada por la interdigitación de microvellosidades de ambos tejidos, materno y fetal, siendo de mayores dimensiones las del trofoblasto por lo que la interdigitación es incompleta. Con el progreso de la preñez, la superficie de contacto aumenta por la presencia de sinuosidades y ramificaciones de las proyecciones del trofoblasto y endometrio, ello probablemente facilite el pasaje de nutrientes.
El pasaje de substancias es evidenciada en las paredes de los capilares materno y fetal, este intercambio molecular parece ser favorecido por la reducida distancia entre ambos capilares debido a la indentación de los capilares fetales en la capa trofoblástica y la disminución en la espesura del epitelio uterino y evidente aumento de la red vascular capilar subepitelial.

204 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
En todos los periodos analizados, fueron encontrados en la interface materno-fetal y a intervalos regulares, pequeñas áreas de acumulo de material amorfo, de superficie convexa que por analogía a las estructuras semejantes encontradas en la placenta de suínos (Leiser and Dantzer, 1994; Dantzer, 1984) fueron denominados de aréolas. En placentas de 347 las aréolas mostraron mayor desarrollo. Estas regiones corresponden a la abertura de las glándulas uterinas, donde el corion no esta en contacto con la superficie endometrial, permitiendo un acumulo de material eletrodenso, homogéneo, PAS+, fosfatasa ácida + proveniente de las glándulas uterinas. El corion en estos locales mostraron cortas proyecciones digitiformes en contacto directo con la secreción glandular; los capilares situados en el tejido mesenquimal son Fe+. Las células trofoblásticas muestran hipertrofia, cuyo citoplasma apical esta repleto de gránulos de diversos tamaños y PAS+. La misma reacción fue observada en la secreción presente en la luz de la areola. La ultra-estructura de estas células mostró presencia de numerosos y largo microvillos, figuras de endocitosis y /o exocitosis, acumulación de gránulos de electro densidad variada y presencia de material de características lipídicas ocupan grandes proporciones en el citoplasma. La inmunolocalización y western blotting para la hormona lactogeno placentario es observado en todas las células trofoblásticas, identificando pesos moleculares entre 35 y 76 kDa para el anticuerpo policlonal IgG de conejo anti-hormona lactógeno placentario bovino. En base a la similitud del peso molecular estimado, probablemente la banda 35 kDa corresponda a la isoforma de la alpaca de la hormona lactógena placentaria y la segunda puede estar relacionada a las proteínas específicas de la preñez encontradas en varias especies (Olivera et al, 2003b).
Para todas las edades, las estructuras que garantizan la adhesión de ambos organismos corresponden a la bien elaborada interdigitación de microvellosidades y presencia de material electrodenso situado entre ambas superficies, que actúa como medio adhesivo y próximo al parto estas estructuras pierden contacto físico y entre ellas muestran presencia de material filamentoso electrodenso.
La presencia de células comprometidas con procesos de muerte celular por apoptosis fue evaluada por la técnica del TUNEL. La reacción se delimito en el núcleo de las células trofoblásticas de muestras provenientes de placentas post-parto y en algunas de placenta a término. Estas apreciaciones son coherentes con la fisiología de la placenta, un órgano extremadamente importante, pero transitorio.
Los datos mostrados en la caracterización de la placenta de alpaca sugieren que a pesar de ser
considerado una placenta difusa, epiteliocorial, esta contiene regiones especializadas funcionando como sitios histotróficos, incrementando el intercambio molecular.
REFEREnCIAS
Dantzer V (1984). Scanning electron microscopy of exposed surfaces of the porcine placenta. Acta Anat. 118: 96-106.
Elwishy A. (1987). Reproduction in the female dromedary (Camelus dromedaries): A Review. Anim Reprod Sci 15: 273-97
Fernandez-Baca S, Hansel W, Saatman R, Sumar J, Novoa C. (1979). Differential luteolytic effects of right and left uterine horns in the alpaca. Biol Reprod. 20: 586-95
Jones CJ, Abd-Elnaeim M, Bevilacqua E, Oliveira LV, Leiser R. (2002). Comparison of uteroplacental glycosylation in the camel (Camelus dromedarius) and alpaca (Lama pacos). Reproduction. 123:115-26.
Klisch K, Bevilacqua E, Olivera LV. (2005). Mitotic polyploidization in trophoblast giant cells of the alpaca. Cells Tissues Organs. 181:103-8
Leiser R, Dantzer V. (1994). Initial vascularisation in the pig placenta: II. Demonstration of gland and areola-gland subunits by histology and corrosion casts. Anat Rec.238: 326-34.
Olivera L, Zago D, Leiser R, Jones C, Bevilacqua E. (2003). Placentation in the alpaca, Lama pacos. Anat Embryol.. 207: 45-62.
Olivera LV, Zago DA, Jones CJ, Bevilacqua E ( 2003). Developmental changes at the materno-embryonic interface in early pregnancy of the alpaca, Lamos pacos. Anat Embryol. 207: 317-31.
Skidmore JA, Wooding FB, Allen WR. (1996). Implantation and early placentation in the one-humped camel (Camelus dromedarius). Placenta. 17: 253-62.
Sumar J. (1988) Removal of the ovaries or ablation of the corpus luteum and its effect on the maintenance of gestation in the alpaca and llama. Acta Vet Scand Suppl. 83: 133-41.
Valencia M. (2007) Microvascularización del útero de la alpaca. Tesis, Facultad de Medicina Veterinaria y Zootecnia. Puno-Perú.

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•205
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
programa de meJoramiento genetiCo en Camelidos domestiCos
Renieri, C1; Pacheco C. 2, Valbonesi, A1; Frank, E3; Antonini; M4
1Departamiento de Ciencias Ambientales, Universidad de Camerino, Italia. Email: [email protected]; 2DESCO CEDAt, Arequipa, Peru, email : 3Facultad de Ciencias Veterinarias, Universidad Catolica de Cordoba, Argentina. Email ...; 3ILO, Universidad de Camerino, Italia. Email : [email protected]
Selection program in south american domestic camelids
1. IntRODUCCIón El documento presenta la experiencia de selección genética que se esta conduciendo en la población de alpacas de la provincia de Caylloma en Perú (PROMEGE, programa de mejoramiento genético) a través de un esfuerzo de cooperación entre el Centro de Estudios y Promoción del Desarrollo (desco) – Perú, el departamento de ciencia ambiental de la universidad de camerino – Italia, el Centro para la nueva tecnología, la energía y el ambiente ENEA - Italia y el programa SUPPRAD de la universidad Católica de Córdoba – Argentina.
El programa utiliza un esquema de selección de núcleo abierto, caracterizado por un núcleo manejado por desco – CEDAT, 20 planteleros (850 animales aproximadamente) y más de 200 criadores (familias campesinas) (15000 animales aproximadamente) .La población total de la provincia de Caylloma se estima en 200 000 animales aproximadamente, 90 % Huacaya y 10 % Suri, con un 60% de animales blancos y un 40 % de animales de color (Gonzales y Renieri 1998). El programa de mejoramiento genético se inicio en 1985 con la creación por Desco del programa de desarrollo rural valle del Colca, y continuó en 1996 con la creación del Centro de Desarrollo Alpaquero de Toccra (CEDAT) En 1998 se ha formulado el plan de mejora genética (Gonzales y Renieri 1998) que entró en funcionamiento en el 2005, después de los resultados de dos programas de investigación para el desarrollo financiados por la Unión Europea (U.E. INCO) SUPREME (1997 – 2000) DECAMA (2002 – 2006) que han permitido adquirir toda la información genética necesaria. En este momento el programa
esta en pleno funcionamiento. PROMEGE es el primer programa de selección genética hasta ahora hecho para operar en las poblaciones de alpacas y llamas criadas en el altiplano andino. El único otro ejemplo en ese sentido es en efecto el programa AGE, que funciona en Australia y Nueva Zelanda e ITALPACA, que funciona desde el 2004 en Italia.
2. PROMEGE
2.1 LOS OBJEtIVOS DE SELECCIón Los caracteres que se toman en cuenta como objetivos de selección son los siguientes:
a.- Caracteres cualitativos Tipo de vellón: Suri o Huacaya El color del vellón, en este caso la selección es: A favor del blanco uniforme no albinoA favor del vellón de color uniforme (Negro, marrón, LF y salvaje) Contra el vellón de dos colores, contra las manchas irregulares
b.- Características cuantitativas Cantidad de fibra producidaFinura de la fibra (diámetro)Coeficiente de variabilidad del diámetro de fibra
2.2.1 CRItERIOS DE SELECCIón Representan criterios de selección:La observación directa de los animales al nacimiento, por el tipo de vellón y el color El peso de la fibra producida en la primera esquila que debe realizarse al año de edad, con un intervalo de confianza de mas menos 2 meses (Antonini et al 2004).El diámetro medio de la fibra y el coeficiente de variabilidad obtenidos de la muestra estándar del lado derecho del animal tomado en la primera esquila .
206 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
2.3 LA BASE GEnétICA DE LOS CARACtERES
2.3.1. SURI VS hUACAyA
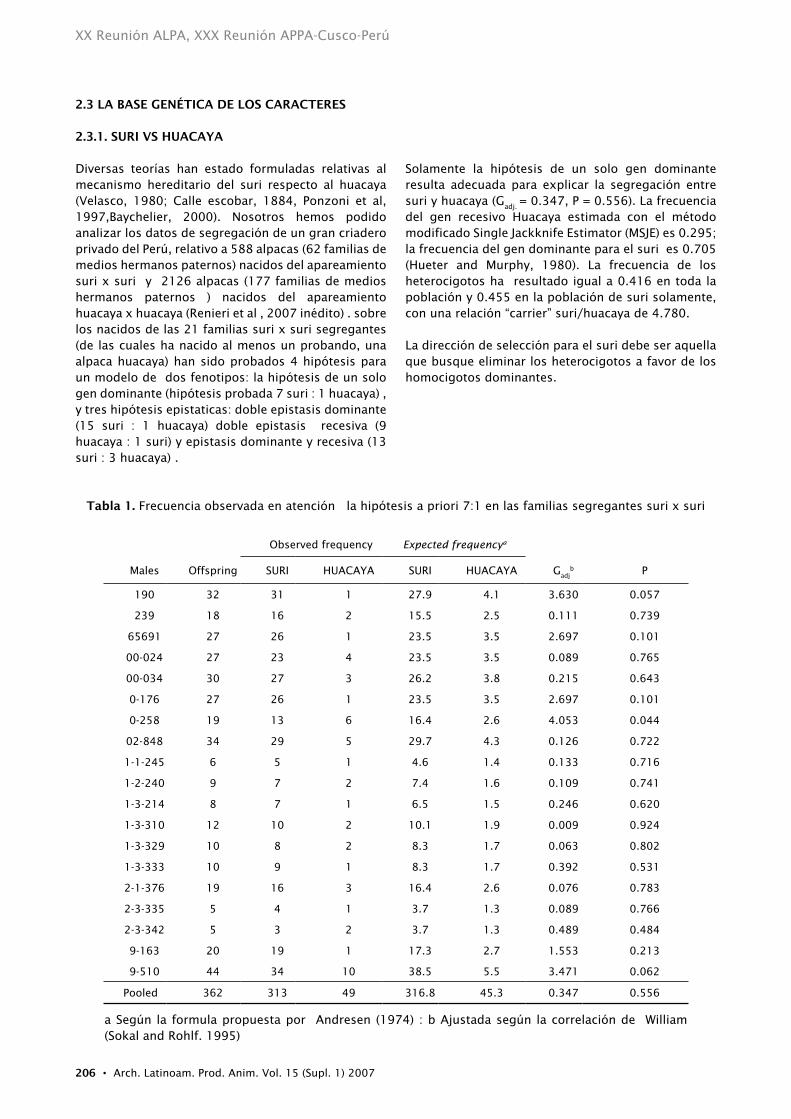
Diversas teorías han estado formuladas relativas al mecanismo hereditario del suri respecto al huacaya (Velasco, 1980; Calle escobar, 1884, Ponzoni et al, 1997,Baychelier, 2000). Nosotros hemos podido analizar los datos de segregación de un gran criadero privado del Perú, relativo a 588 alpacas (62 familias de medios hermanos paternos) nacidos del apareamiento suri x suri y 2126 alpacas (177 familias de medios hermanos paternos ) nacidos del apareamiento huacaya x huacaya (Renieri et al , 2007 inédito) . sobre los nacidos de las 21 familias suri x suri segregantes (de las cuales ha nacido al menos un probando, una alpaca huacaya) han sido probados 4 hipótesis para un modelo de dos fenotipos: la hipótesis de un solo gen dominante (hipótesis probada 7 suri : 1 huacaya) , y tres hipótesis epistaticas: doble epistasis dominante (15 suri : 1 huacaya) doble epistasis recesiva (9 huacaya : 1 suri) y epistasis dominante y recesiva (13 suri : 3 huacaya) .
Solamente la hipótesis de un solo gen dominante resulta adecuada para explicar la segregación entre suri y huacaya (G
adj. = 0.347, P = 0.556). La frecuencia
del gen recesivo Huacaya estimada con el método modificado Single Jackknife Estimator (MSJE) es 0.295; la frecuencia del gen dominante para el suri es 0.705 (Hueter and Murphy, 1980). La frecuencia de los heterocigotos ha resultado igual a 0.416 en toda la población y 0.455 en la población de suri solamente, con una relación “carrier” suri/huacaya de 4.780.
La dirección de selección para el suri debe ser aquella que busque eliminar los heterocigotos a favor de los homocigotos dominantes.
tabla 1. Frecuencia observada en atención la hipótesis a priori 7:1 en las familias segregantes suri x suri
Observed frequency expected frequencya
Males Offspring SURI HUACAYA SURI HUACAYA Gadj
b P
190 32 31 1 27.9 4.1 3.630 0.057
239 18 16 2 15.5 2.5 0.111 0.739
65691 27 26 1 23.5 3.5 2.697 0.101
00-024 27 23 4 23.5 3.5 0.089 0.765
00-034 30 27 3 26.2 3.8 0.215 0.643
0-176 27 26 1 23.5 3.5 2.697 0.101
0-258 19 13 6 16.4 2.6 4.053 0.044
02-848 34 29 5 29.7 4.3 0.126 0.722
1-1-245 6 5 1 4.6 1.4 0.133 0.716
1-2-240 9 7 2 7.4 1.6 0.109 0.741
1-3-214 8 7 1 6.5 1.5 0.246 0.620
1-3-310 12 10 2 10.1 1.9 0.009 0.924
1-3-329 10 8 2 8.3 1.7 0.063 0.802
1-3-333 10 9 1 8.3 1.7 0.392 0.531
2-1-376 19 16 3 16.4 2.6 0.076 0.783
2-3-335 5 4 1 3.7 1.3 0.089 0.766
2-3-342 5 3 2 3.7 1.3 0.489 0.484
9-163 20 19 1 17.3 2.7 1.553 0.213
9-510 44 34 10 38.5 5.5 3.471 0.062
Pooled 362 313 49 316.8 45.3 0.347 0.556
a Según la formula propuesta por Andresen (1974) : b Ajustada según la correlación de William (Sokal and Rohlf. 1995)

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•207
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Entre los 2126 (1009 hembras y 1117 machos) nacidos del apareamiento huacaya x huacaya en 177 familias de medios hermanos paternos, 2123 resultaron huacaya y 3 suris nacidos en tres familias diferentes. puesto que los padres pertenecen a líneas huacaya de varias generaciones, no se puede pensar en un error de clasificación por las madres. Se trata entonces del resultado de una nueva mutación con sobre expresión genética (mutación inversa) a nivel de líneas gameticas de los progenitores. El gen responsable para la síntesis de huacaya y suri estaría por lo tanto el resultado de una mutación recurrente, que aparece con una tasa de mutación constante en cada generación. La tasa de mutación seria igual a 3/(2126) = 0.001411101. El nacimiento de suri productos de apareamiento huacaya x huacaya explica porque a veces a sido considerado recesivo. algunos autores describen la existencia de un tipo intermedio denominado chili (anónimo, 1994: C Tuckwel, comunicación personal y Ponzoni et al, 1997) descrito también en la llama (Frank et al .; 2001). El estudio de la penetración del carácter suri en la heterocigosis será seguido en el centro experimental INIA – Quimsachata gracias al cruce entre suri y huacaya según el siguiente esquema : 9 machos suris x 135 hembras huacaya y 8 machos huacaya x 95 hembras suri.
2.3.2 COLOR DEL VELLónSon favoritos para la selección el blanco uniforme no albino y el modelo de pigmentación uniforme negro y marrón (Renieri et al., 1994 a, b; Renieri, 1995) blanco significa ausencia de pigmentación. Porque los melanocitos, las células que producen pigmentos no se encuentran solo sobre la piel de los animales provistos de cobertura pilífera (melanocitos foliculares) pero también en el ojo, en el oído interno, en las meninges y en otros órganos internos, una primera distinción importante que hacer es entre una ausencia generalizada de pigmentación o una ausencia de pigmentación limitada al folículo piloso. El primer tipo de blanco es decir ALBINISMO OCULOCUTANEO, da origen a animales con problemas también auditivos y visuales , puede derivarse de dos mecanismos genéticos:
• Albinismo “tirosinasas negativo” (OCA 1): consigue una mutación con perdida de funciones de los alelos salvajes al locus estructural para la tirosinasa (albino), el cual es la vía para la síntesis de todas los pigmentos (pasaje de tiroxina a DOPA quinona)
• Albinismo “Tirosinasa positivo” consigue una mutación de loci asociados como pink eyed (OCA 2), TYRP1 (OCA3), MATP (OCA4)
Varias formas hipomelanosis (Hermansky-Pudlack syndrome, ChediaK-Higashi syndrome, Griscelli sindrome, Angelman syndrome, Prader-Willi syndrome,
Ataxia Telengiectasia, Hallerman-Streiff syndrome, Histidinemia, Homocystinuria, Oculocerebral syndrome with Hypopigmentation, Tietz syndrome, Kappa-Chain deficiency, Menkes’Kinky Hair syndrome, Phenilketonuria), nelle quali sono coinvolti molti geni (King y Oetting, 2006).
Ninguna de estas situaciones patologicas pueden ser tomadas en consideración para obtener un blanco uniforme ( Renieri, 2003). El Blanco uniforme no albino característico de varias especies de mamíferos es obtenido como mutación con perdida de funciones de genes involucrados en la migración embrionaria de melanoblastos, de la cresta neural al folículo piloso del los animales en su desarrollo local (Bennet y Lamoreaux 2003). Están actualmente identificados y clonados en mamíferos 22 genes que intervienen en esta función ( Baxter et al 2004). La mayor parte de las mutaciones asociadas al blanco en tales loci se comportan como letales. En dos casos a la inversa para el loci MITF ( microftalmia ) y C – Kit ( Dominant White Spootting) es posible obtener individuos vitales completamente blancos. Estos podrían ser los genes implicados en el blanco.
En colaboración con INIA, en la estación experimental de Quimsachata se esta llevando acabo una experimentación orientada a comprender el mecanismo biológico del blanco uniforme y su base genética.
El vellón negro uniforme deriva de la prevalecía de eumelanina en el vellón del animal (Renieri et al 1991, Renieri et al 1995; Cecchi et al 2006). Estas Eumelaninas son negras por que en el heteropolimero que le caracteriza se tiene una prevalencia neta del monómero DHI (Dihidroxindol) .
En el caso de una prevalencia de la forma acidificada al contrario (DHICA ) las eumelaninas son marrones. Genéticamente el negro uniforme puede tener dos orígenes. Puede derivarse de una mutación con perdida de función al locus Agouti, en este caso se comporta como recesivo con respecto a los otros modelos de pigmentación. o también puede derivarse de mutaciones con sobre expresión génica al locus MC1R el cual en este caso es dominante ( Lauvergne et al 1996) de los datos de segregación en nuestro proceso existe seguramente un negro recesivo en la alpaca. No es posible excluir también la existencia de un negro dominante.
El marrón representa un modelo Eumelanico con prevalencia de eumelaninas marrones, por la presencia de eumelanosomas marrones (Cozzali et al 2001) genéticamente el marrón deriva de una mutación con perdidas de funciones del locus TYRP1 (Brown)
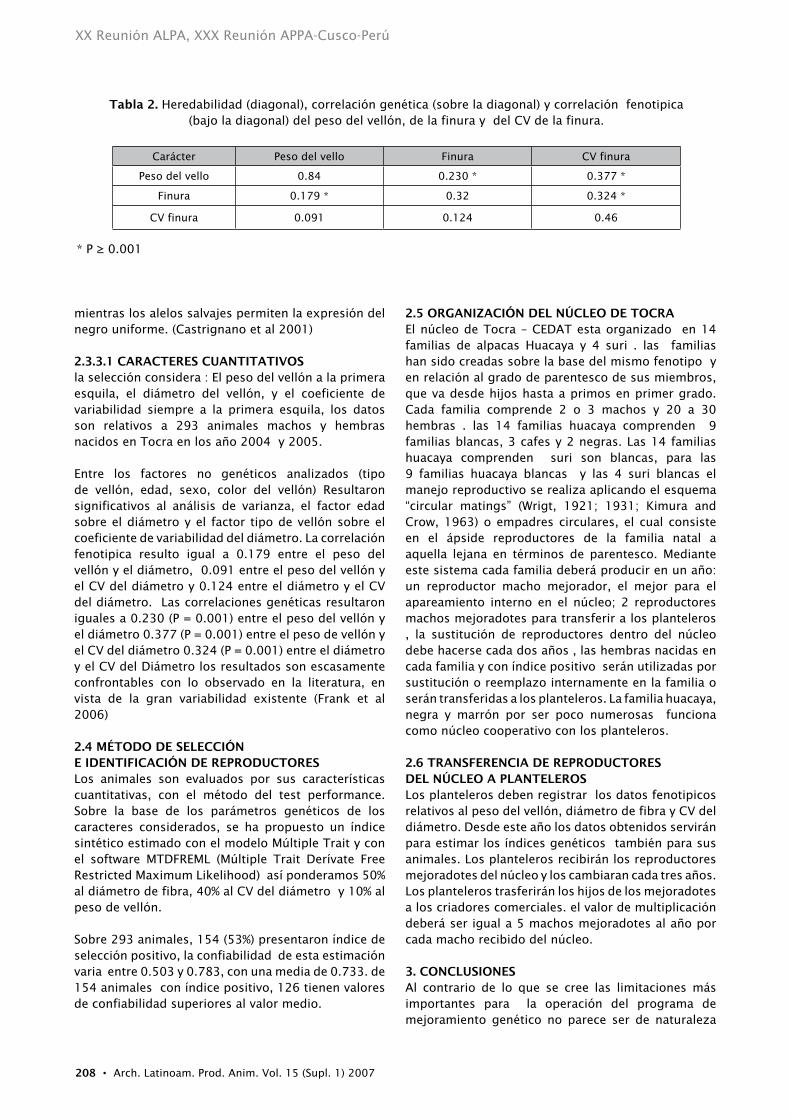
208 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
mientras los alelos salvajes permiten la expresión del negro uniforme. (Castrignano et al 2001)
2.3.3.1 CARACtERES CUAntItAtIVOS la selección considera : El peso del vellón a la primera esquila, el diámetro del vellón, y el coeficiente de variabilidad siempre a la primera esquila, los datos son relativos a 293 animales machos y hembras nacidos en Tocra en los año 2004 y 2005.
Entre los factores no genéticos analizados (tipo de vellón, edad, sexo, color del vellón) Resultaron significativos al análisis de varianza, el factor edad sobre el diámetro y el factor tipo de vellón sobre el coeficiente de variabilidad del diámetro. La correlación fenotipica resulto igual a 0.179 entre el peso del vellón y el diámetro, 0.091 entre el peso del vellón y el CV del diámetro y 0.124 entre el diámetro y el CV del diámetro. Las correlaciones genéticas resultaron iguales a 0.230 (P = 0.001) entre el peso del vellón y el diámetro 0.377 (P = 0.001) entre el peso de vellón y el CV del diámetro 0.324 (P = 0.001) entre el diámetro y el CV del Diámetro los resultados son escasamente confrontables con lo observado en la literatura, en vista de la gran variabilidad existente (Frank et al 2006)
2.4 MétODO DE SELECCIón E IDEntIFICACIón DE REPRODUCtORES Los animales son evaluados por sus características cuantitativas, con el método del test performance. Sobre la base de los parámetros genéticos de los caracteres considerados, se ha propuesto un índice sintético estimado con el modelo Múltiple Trait y con el software MTDFREML (Múltiple Trait Derívate Free Restricted Maximum Likelihood) así ponderamos 50% al diámetro de fibra, 40% al CV del diámetro y 10% al peso de vellón.
Sobre 293 animales, 154 (53%) presentaron índice de selección positivo, la confiabilidad de esta estimación varia entre 0.503 y 0.783, con una media de 0.733. de 154 animales con índice positivo, 126 tienen valores de confiabilidad superiores al valor medio.
2.5 ORGAnIzACIón DEL núCLEO DE tOCRA El núcleo de Tocra – CEDAT esta organizado en 14 familias de alpacas Huacaya y 4 suri . las familias han sido creadas sobre la base del mismo fenotipo y en relación al grado de parentesco de sus miembros, que va desde hijos hasta a primos en primer grado. Cada familia comprende 2 o 3 machos y 20 a 30 hembras . las 14 familias huacaya comprenden 9 familias blancas, 3 cafes y 2 negras. Las 14 familias huacaya comprenden suri son blancas, para las 9 familias huacaya blancas y las 4 suri blancas el manejo reproductivo se realiza aplicando el esquema “circular matings” (Wrigt, 1921; 1931; Kimura and Crow, 1963) o empadres circulares, el cual consiste en el ápside reproductores de la familia natal a aquella lejana en términos de parentesco. Mediante este sistema cada familia deberá producir en un año: un reproductor macho mejorador, el mejor para el apareamiento interno en el núcleo; 2 reproductores machos mejoradotes para transferir a los planteleros , la sustitución de reproductores dentro del núcleo debe hacerse cada dos años , las hembras nacidas en cada familia y con índice positivo serán utilizadas por sustitución o reemplazo internamente en la familia o serán transferidas a los planteleros. La familia huacaya, negra y marrón por ser poco numerosas funciona como núcleo cooperativo con los planteleros.
2.6 tRAnSFEREnCIA DE REPRODUCtORES DEL núCLEO A PLAntELEROS Los planteleros deben registrar los datos fenotipicos relativos al peso del vellón, diámetro de fibra y CV del diámetro. Desde este año los datos obtenidos servirán para estimar los índices genéticos también para sus animales. Los planteleros recibirán los reproductores mejoradotes del núcleo y los cambiaran cada tres años. Los planteleros trasferirán los hijos de los mejoradotes a los criadores comerciales. el valor de multiplicación deberá ser igual a 5 machos mejoradotes al año por cada macho recibido del núcleo.
3. COnCLUSIOnES Al contrario de lo que se cree las limitaciones más importantes para la operación del programa de mejoramiento genético no parece ser de naturaleza
tabla 2. Heredabilidad (diagonal), correlación genética (sobre la diagonal) y correlación fenotipica (bajo la diagonal) del peso del vellón, de la finura y del CV de la finura.
Carácter Peso del vello Finura CV finura
Peso del vello 0.84 0.230 * 0.377 *
Finura 0.179 * 0.32 0.324 *
CV finura 0.091 0.124 0.46
* P ≥ 0.001

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•209
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
ambiental (complejidad del ambiente andino) ni de tipo económico, pero esencialmente de tipo cultural.Un programa de mejoramiento genético requiere que se defina bien la población que donde se aplicará tal programa.
Las razas existentes en la alpaca son esencialmente del tipo primario, resultado de la primera repartición del pool genético sucesivo a la domesticación y en los camélidos domésticos algunos efectos de deriva genética provocados por el shock de la conquista (Renieri et al 2007) las razas primaras se caracterizan clásicamente por una elevada variabilidad fenotipica sea de los caracteres cualitativos (color, tipo de vellón) como de los caracteres cuantitativos (parámetros biométricos, producción)
Para lo cuál el acercamiento a la selección exige particularmente opciones impactantes en la misma población. Un acercamiento bastante erróneo ha llevado a prepreferir los efectos de la combinación genética (incrocio) a los de la selección. En particular tal elección ha estado fuertemente valorada en Perú. De la creación de un L.G. nacional que permitía la inscripción de de animales pertenecientes a cualquier población, en la idea herrada que un mejorador pueda comportarse en tal modo en cualquier población donde se reproduzca. REFEREnCIAS
Andresen, E., 1974. The effect of ascertainment by truncate selection on segregation ratios. 1st World Congress Genetic Applied Livestock Production, Madrid, 111-114.
Anonymous, 1994. Alpacas Australia, Issue Number I, p. 47.
Antonini M., Gonzales M. Y Valbonesi A., 2004. Relaccion entre la edad y el desarrollo postnatal de los fullicolos pilosos en tres tipos de camelidos sudamericanos domesticos. Livestock Production Science, 90, 241-246.
Baychelier P., 2000. Suri and Huacaya : Two alleles or Two Genes ?. Proc. 2000 Australian Alpaca Association National Conf., Camberra, pp 79-85.
Baxter L.L., Ling Hou, S.K. Loftus and W.J. Pavan, 2004. Spotlight on Spotted Mice: A Review of White Spotting Mouse Mutants and Associated Human Pigmentation Disorders. Pigment Cell Research, 17, 215-224.
Bennett D.C. and Lamoreax M.L., 2003. The Color Loci of Mice – A Genetic Century. Pigment Cell Research, 16, 333-344.
Calle Escobar R., 1984. Animal Breeding and Animal Production of American Camelids. Ron Henning – Patience, Lima, Peru.
Castrignanò F., Antonini M., Misiti S., Cristofanelli S., Renieri C., 2001. Sequence of tyrosinase related protein-1 (TRP-1) in alpaca. In : Gerken M. e Renieri C Eds “Progress in South American camelids research”, EAAP publ n° 105, Wageningen Pers, Wageningen, 2001, 199-206.
Cozzali C., Dell’Aglio C., Gargiulo A.M., Frank E., Hick M., Ceccarelli P., 2001. SUPREME-Project : Pigmentation on South American camelids: II. Morphological features of follicular melanocytes. In : Gerken M. e Renieri C Eds “Progress in South American camelids research”, EAAP publ n° 105, Wageningen Pers, Wageningen, 2001, 237-238.
Frank E. N., Hick M.H.V., Pesarini M., Hick, P.M.L. Hick, Capelli C. I., Ahumada M.R., 2001. SUPREME-Project: Classification of fibres of different types of fleeces in Argentine llamas. In : Gerken M. e Renieri C Eds “Progress in South American camelids research”, EAAP publ n° 105, Wageningen Pers, Wageningen, 2001, 251.
Frank E.N., Hick M.V.H., Gauna C.D., Lamas H.E., Renieri C., Antonini M., 2006. Phenotypic and genetic description of fibre traits in South American Domestic Camelids (Llamas y Alpacas). Small Ruminant Research, 61, 113-129.
Gonzales M. y Renieri C., 1998. Propuesta de un plan de seleccion de la poblacion de alpacas en la provincia de Caylloma, Arequipa. En Frank E., Renieri C. Y Lauvergne J.J., Actas del Tercer Seminario de Camelidos Sudamericanos Domesticos y primer Seminario Proyecto SUPREME, Universidad Catolica de Cordoba, pp. 27-38.
Huether C.A. and Murphy E.A., 1980. Reduction of bias estimating the frequency of recessive genes. Am. J. Hum. Genet., 32: 212-222.
Kimura M., Crow CJ.F., 1963. On the maximum avoidance of inbreeding. Genet. Res., 4, 399-415.
King R.A. and Oetting W.S., 2006. Oculocutaneous Albinism. In “Genetic Hypomelanoses : Generalized Hypopigmentation”. In Nordlung J.J. et al. (eds) The Pigmentary System. Blackwell Publishing, Oxford University Press, pp599-621.
Lauvergne J.J., Renieri C., Frank E., 1996. Identification of some allelic series for coat colour in domestic camelids of Argentina. Proc. 2° European Symposium on South American Camelids, Università di Camerino, Italy, 39-50.

210 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Ponzoni R.W., Hubbard D.J., Kenyoni R.V., Tuckwell C.D., McGregor B.A., Howse A., Carmichael I., Judson G.J., 1997. Phenotypes resulting from huacaya by huacaya, suri by huacaya and suri by suri alpaca crossing. Proc. Intern. Alpaca Industry Sem., pp 11-13.
Renieri C., 1994 a. Pigmentation in domestic mammals with reference to fine fibre producing species.Eur. Fine Fibre Network, Occ. Publ. No. 1, 113-136.
Renieri C., 1994 b. The genetic basis of pigmentation in South American Camelids. Proc. Eur. Symp. South American Camelids, M. Gerken & C. Renieri Ed., Univ. di Camerino,1994, 31-41.
Renieri C., 1995. Biologia y genetica de las capas de los mamiferos y su extension a los camelidos. Actas Primer Seminario Internacional de Camelidos Sudamericanos Domesticos, Frank E.N. e C. Renieri Eds, Univ. di Camerino, 69-87.
Renieri C., 2003. Selection for coat colour in alpaca (Lama pacos) and llama (Lama glama). Proc. III Congreso Mundial sobre Camelids y I Taller Internacional de DECAMA, Potosi, Bolivia, 11-19.
Renieri C., Trabalza Marinucci M., Martino G., Giordano G., 1991. Preliminary report on qualità of hair and coat colour in pigmented alpaca. Proc. IX Italian ASPA, 905-914.
Renieri C., Lauvergne J.J., Antonimi M., Lundie R., 1995. Quantitative determination of melanins in black, brown, grey and red phenotypes of sheep, goat and alpaca. Proc. 9th Int. Wool Textile Res. Conf., II, 580-586
Renieri1 C. ,Frank2 E.N., Rosati3 A.Y Macias Serrano4 J. A. , 2007. El concepto de raza en zootecnia y su applicaciòn a la llama y a la alpaca. In Frank E., Antonini M., Toro O.(eds) South American Camelids Research. Volume II, Wageningen Academic Publisher, in press.
Renieri C., Valbonesi A., La Manna V., Antonimi M., Asparrin M., 2007. Inheritance of Suri and Huacaya type of fleece in alpaca. Small Ruminant Research, submitted.
Sokal R.R., Rohlf F.J., 2003. Biometry. 3rd ed. Freeman WH and Company, New York, 887 p.
Velasco J., 1980. Mejoramiento Genetico de Alpacas. Anales III Reunion Cientifica Annual Soc. Peruana de Prod. Anim., Lima.
Wright S., 1921. Systems of mating. Genetics, 6, 111-178.
Wright S., 1931. Evolution in Mendelian population. Genetics, 16, 97-159.

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•211
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
realidades Y mitos soBre los Camelidos sudameriCanos
Julio Sumar*
* MV,MSc,FRVCS.Profesor Emerito de la UnMSM.
El trasiego que se ha hecho hacia los EEUU y Europa de los Camélidos Sudamericanos Domésticos en los últimos 27 años, ha cambiado totalmente la realidad del monopolio que se tenía sobre estas especies, así como ha variado el número poblacional en los Andes Centrales. Se considera que el Perú encabeza las encuestas en número y calidad, seguidos por Bolivia, Chile y Argentina. Hoy los EEUU ha pasado a tercer lugar en población, y pronto Australia estará disputando con ese país el tercer lugar en el mundo.
Sin embargo, no es el número que ha salido lo que nos preocupa, sino, la calidad genética que hemos perdido. Los criadores motivados por su pobreza y por los altos precios que alcanzaron sus mejores animales, han terminado vendiendo la “gallina de los huevos de oro”. Después de algunos años de contínua exportación, los EEUU, Canadá y otros países han cerrado sus Registros genealógicos, porque consideran que ya tienen la cantidad y calidad de alpacas necesarias, pudiendo competir en calidad con el Perú. El que os habla es Juez Internacional y conoce de primera mano, lo que está sucediendo en cuanto al mejoramiento y calidad de fibra que se ha logrado en otros países.
Es pues importante comenzar a entender que si no hacemos un Programa Nacional que realmente impulse el Mejoramiento Genético de alpacas y llamas, ya lo he dicho en varias ocasiones, la fibra fina tendrá que ser importada, lo mismo que los reproductores. Lo que sucede es que existen algunos Realidades positivas y también negativas, así como varios mitos, sobre los que me referiré más adelante, manejados por los criadores, y también, por algunas autoridades del sector Agricultura alpaquero, que no son técnicos especializados, sino, políticos ambiciosos, metidos a prosperar económicamente, sin importarles absolutamente el mejoramiento de los criadores y de su nivel de vida..
Sin embargo y a pesar de la “erosión genética” que hemos sufrido y que seguimos sufriendo, es una
Realidad que el Perú sigue conservando una gran variabilidad genética, la mayor población de alpacas y llamas, y numerosos rebaños de alpacas y llamas, de gran calidad genética, que con un Programa bien estructurado, hecho entre criadores exitosos y profesionales de experiencia e inteligencia, se podría rápidamente diseminar el material genético, evitar la consanguinidad y los cruces ínterespecíficos e interraciales, poniendo al país en los niveles Pre-Reforma Agraria (1968), remontando lo que pudieran estar haciendo otros países en el rubro de mejoramiento genético.
En los últimos años hemos sido testigos de un desarrollo más acelerado de la industria textil artesanal de la fibra de alpaca, y eso es sumamente alentador, generada por algunas ONGs y por el sector turismo, dando mano de obra a miles de mujeres y varones, que con sus métodos de hilado y tejido ecológico, están promoviendo el desarrollo y expansión de esta industria. Sin embargo existe una industria textil intermedia, es decir que no es del tamaño industrial, ni es artesanal, y produce notables cantidades de tejidos planos y otros, que se comercializan con membretes de calidad baby alpaca, siendo en realidad de baja calidad; o como sucede en otros casos, donde se usan ciertas mezclas, ya sea con lana de ovino u otras fibras sintéticas, mezclas que no se pueden analizar y certificar, por carecer a la fecha de un laboratorio de fibras que pueda analizar las muestras y cuantificar las mezclas. Urge la intervención del Estado en este rubro.
Otra Realidad es que el país cuenta con algunas Estaciones Experimentales trabajando “in situ” en investigación de alpacas y llamas; y claro, no es de extrañar que lo hagan muy bien por amor al Perú y al sector alpaquero; sin embargo sus recursos económicos son escasos y sobreviven a base de esfuerzos sobrehumanos. Así por ejemplo, el INIA, el IVITA de San Marcos, Machuhuasi-La Raya de la UNA-Puno y una Asociación Civil Peruana, están trabajando arduamente en lo que es la Transferencia de Embriones (TE) en alpacas y llamas, como un medio de mejoramiento acelerado de la calidad genética de nuestros camélidos, con notables resultados Me he enterado recientemente, que el mayor ejecutivo del Programa Sierra Exportadora, ha manifestado su interés de importar embriones congelados de alpacas

212 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
desde Australia, para mejorar posiblemente la calidad genética , y esa noticia me ha dejado aterrado por las siguientes consideraciones: a).- El Perú ya ha producido crías en alpacas y llamas por TE, y esta en camino de congelar embriones; porque entonces no se ayuda a esas instituciones a realizarlo, faltándoles solamente los equipos apropiados, en lugar de la peregrina idea de importar embriones de Australia, que además huele mal, ya que parecen que hay intereses económicos listos a ofrecernos como salvación dichos embriones.b).-Resulta que la TE puede servir para diseminar enfermedades virales, que si están presentes en Australia, tales como el BVD, Lengua Azul, etc. De manera que esos y otros países, no son la mejor fuente de material genético de alpacas. Resulta que en Australia están en el mismo desarrollo de TE en alpacas, y seria contraproducente ayudar a Australia, en lugar de hacerlo con los nuestros.
Según tengo entendido las vicuñas pertenecen al Estado peruano, que las da a las Comunidades Campesinas, para que adecuadamente y bajo supervisión, obtengan beneficios económicos por la esquila de ellos o su venta controlada hacia los criadores peruanos. Sin embargo, no hay orden ni concierto en el manejo de la vicuña, que se ha convertido en un campo de aficionados, de politiqueros y de corruptos. No me voy a referir a los últimos acontecimientos referidos a la transferencia de 2000 vicuñas de Puno a la Sierra de Trujillo; de eso se encargaran los jueces. Me voy a referir a la exportación de Paco-vicuñas hacia los EEUU, que se ha producido por cruces de alpaca y vicuña realizados en el Norte de Chile, y enviados a los EEUU, en alpacas gestantes. Hemos vendido pues, genes de vicuña en los cruces. Y para atreverme a decir lo que digo, tengo las pruebas documentales y los registros aduaneros de embarques de alpacas de Chile a USA, figurando un contingente de alpacas que se exportó cuando el Registro de alpacas de los EEUU estaba cerrado, y el ingreso de esos animales descartaba su registro y su valor económico. No eran las alpacas lo que se quería, sino, lo que traían en sus vientres. Y no seria raro que algunas entidades inescrupulosas o criadores, estén pensando en exportar la vicuña en vientre de alpaca. Por eso, resulta irritable, que algunos criadores estén manejando las vicuñas en cruces y en otros experimentos, sin control o permiso del ente regulador de esas especies. Urge que la institución estatal a quien corresponde velar por esas especies prohibidas de exportar, se pongan a hacer lo que les corresponde por ley.
Otra Realidad nos dice de la falta de un mejoramiento genético en la mayoría de nuestros rebaños, que se expresa en el porcentaje bajo de fibra fina que se produce en el país. ¿Quien es el responsable de esta situación? Me pongo en el pellejo de un criador de Macusani, y dando respuesta a esa pregunta, diría
que, por los precios que me dan por mi fibra, creo que no me interesa el mejoramiento genético.
Podría aventurarme a decir que el Perú es un país muy rico en Mitos, tan rico como la antigua Grecia de Socrates; vayamos nomás a una librería o preguntemos a algunos de los antropólogos. Huelga enumerar a tantos autores. Sin embargo, creo que podría escribir un libro sobre los Mitos que sobre la crianza de alpacas andan de boca en boca. He aquí algunos:
Mito de la altura.- Cuando las alpacas salían ilegalmente del Perú (1984 hasta la fecha), hacia los EEUU y otros países, los criadores decían que “las alpacas se van a morir en esos climas, que no son de altura”. El tiempo se encargaría de mostrarnos a todos, que no solo sobrevivían, sino, producían tanto o mejor que en su país de origen.
Mito de la finura.- La gran finura que muestran algunos ejemplares se debe a “la ingestión de algunas plantitas que les dan gran finura y que crecen en los bofedales de altura”. La finura que muestran algunas alpacas en la altura, se debe a la desnutrición, es decir a lo que se llama “finura por hambre”. La finura de la fibra es producto de los genes y el ambiente.
Mito del engrosamiento de la fibra.- Se inicio con la exportación de alpacas. Entre los exportadores americanos, aparecieron algunos periodistas investigadores espontáneos, que decidieron averiguar qué pasaba con la finura de la fibra de las alpacas que compraban en la altura y a las que se tomaba una muestra de fibra, y luego eran internadas en las cuarentenas del país y del extranjero, que duraban en promedio 8 meses, al termino del cual tomaban otra muestra y “Dios mió el micronaje se había incrementado en mas de 6µ lo que quiere decir que la buena alimentación no sirve para estos animales”. Claro que si un animal en la altura vive desnutrido y luego lo superalimentamos, se incrementará el grosor de la fibra; lo que no nos dijeron era cuántos de los animales no cambiaban o cambiaban muy poco su finura. No he leído ningún trabajo que dé esa información. Lo que sabemos a ciencia cierta es que existen numerosos animales que cambian muy poco de una esquila a otra, y esos deben ser los que nos interesen genéticamente.
Mito del Paco-Vicuña.- Tiene sus orígenes en un premio que le concedió el Congreso de la Republica, al cura de Macusani, Padre Cabrera, allá por 1850, por haber cruzado la vicuña con la alpaca y “haber creado una nueva raza de abundosa y fina fibra: el Paco-vicuña”. Del Padre Cabrera nunca mas se oyó hablar y tampoco de su nueva raza. Volvemos a escuchar sobre ellos, cuando algunos hacendados-exportadores que tenían licencia para exportar fibra de

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•213
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
vicuña, introducían a sus rebaños de vicuñas, algunas alpacas para producir el cruce y obtener especímenes de mayor producción de fibra. Cuando trabajaba en la Raya en la década del 60, encontramos un rebaño de Paco-vicuñas de todos los colores y de diversas producciones, herencia de la ex-Granja Modelo de Auquénidos de la Raya del Ministerio de Agricultura Sin embargo no había información valedera de los cruces de manera que no sabíamos con que cruce o generación estábamos liando. Decidimos entonces realizar algunos cruces experimentales, llegando hasta el F2, es decir cruces de dos paco-vicuñas. Los resultados fueron que, primero, el F1 solo producía un 14% mas de fibra fina; segundo, que el temperamento del F1 era similar al de la vicuña, haciéndose más intratable e inmanejable, dada su mayor fortaleza, heredada de la alpaca, y tercero, la frecuencia de corte, era de 3 a 4 años, lo que lo hacia un animal poco económico; cuarto, los cruces de dos F2, disgregaban las características, hacia especimenes que no eran ni alpacas ni vicuñas, por lo que fijar las características era difícil, sino, imposible. Si bien el cruce es viable, siendo dominante el color y el temperamento de la vicuña, su producción no es rentable.
Mito del Pacuña.- Este Mito es auténticamente Norteamericano. Algún tiempo después de la introducción ilegal del Paco-vicuña a ese país se creó, el Registro de la Raza Pacuña. El cruce de los F1 con alpacas, formó el tipo 1/4vicuña- 3/4 alpaca, y luego todas las combinaciones, acercándose en todo caso a la alpaca. Y de los cruces de dos F1, el F2, les producía de todo, menos vicuñas. Con ese contingente y algunos ejemplares adquiridos de algún zoológico europeo, crearon la Raza Pacuña. Personalmente he visto los ejemplares llamados Pacuñas y …… prefiero no hacer comentarios, ya que conozco a más de una persona en el país, que son asesores de dicho Registro.
Mito de la Raza Surillama.- Se inicio cuando un criador de llamas de los EEUU, el Dr. Barnet, se cruzo en su camino a Nuñoa, Puno, con unos ejemplares cruzados entre llama y alpaca Suri, de propiedad del Sr. Díaz. Y bueno, le gustaron esos animales y los exporto a EEUU con el nombre de la Raza Surillama. Con el tiempo crearon un Registro Nacional de estos ejemplares, que a mi buen entender tienen una buena talla y presencia, pero su fibra sobrepasa las 30 micras. Claro que algunos ejemplares F1 cruzados con Suri, tienden a afinarse, pero dudo mucho que puedan fijar las características de manera que la talla sea de la llama y la finura del Suri. En fin cada loco con su tema.
Mito de las miles de hectáreas necesarias para criar alpacas.- Todos los estudiantes Universitarios de las especialidades agropecuarias, recibimos información como parte de nuestra formación profesional, que
la crianza de alpacas, se realiza extensivamente en praderas naturales de las grandes altiplanicies del sur del Perú, y en otras alturas de la cordillera. Así por ejemplo las praderas se clasifican en número de alpacas por hectárea que va de 2 Has. para una alpaca, hasta 2-3 Has. por alpaca. A nadie se le ocurrió que se las podría criar semi-extensivamente y aun intensivamente [claro, que con los precios de la fibra en el Perú, seria una imprudencia criar estos animales). Y así la crianza de alpacas, sin cambio alguno, pasó sin pena ni gloria. Esta vez fueron los norteamericanos, los que nos sacaron de nuestro letargo, criando a nuestras alpacas en espacios reducidos, con alimento variado, concentrados, sales minerales y vitaminas, produciendo vellones más pesados en promedio y tan finos como los nuestros. Ni que decir de las tasas de natalidad y sobrevivencia del neonato.
Mito de los Bancos de Germoplasma.- Los Bancos de Germoplasma, se crearon en la década de los años 50 del siglo pasado, con la finalidad de asegurar la permanencia de muchas plantas económicamente necesarias para el hombre, y que estaban despareciendo, ya sea por el abandono de ciertos cultivos, o la introducción de nuevas variedades industriales de alta producción. Estos Bancos guardan en cuartos refrigerados especiales, semillas de miles de plantas útiles en la alimentación, plantas maderables, ornamentales, medicinales, etc. Contienen las variedades domesticadas, y sus variedades silvestres, que pueden ser fuente de genes especiales, necesarios en el futuro para la bioingeniería.
Los Bancos de Germoplasma para animales domésticos y silvestres, se crearon muy posteriormente, cuando se desarrollaron la Inseminación Artificial con el congelamiento de semen, y también la congelación de embriones, oocitos, etc, (Conservación Ex-Situ). Considero que seria importante que el país desarrolle un Banco de Germoplasma para animales domésticos, y también para la variada y riquísima fauna silvestre. Pero se debe hacer con semen y embriones congelados, y no como algunos Proyectos que con el nombre de Bancos de Germoplasma, funcionan como un repositorio de animales de diversas razas, colores, ecotipos, cruces, etc.,( Conservación In-Situ) que no tienen ningún valor, ni de preservar las especies, ni de asegurar la supervivencia de las mismas. Si el objetivo es conservar ciertos colores de alpacas o llamas, habría que llamarlos, algo así como Centros de preservación y mejoramiento de los colores nativos de alpacas. La manera práctica de lograr la conservación In-Situ de los recursos genéticos de los camélidos, es a través de la utilización sustentable de las diversas razas o tipos de animales. Las estrategias de crianza de las diversas poblaciones o grupos de alpacas han fallado debido a un número de razones, entre las cuales están las propuestas técnicas inadecuadas, equivocados o muy

214 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
pobres objetivos de selección y logros, además de falta de infraestructura y presupuestos, con incentivos económicos realmente bajos.
Bueno, creo que ya es hora de soltar amarras con el pasado, y criar a los camélidos con técnicas modernas y rentables. Terminaría esta charla diciendo, que no necesitamos miles de hectáreas de pastos nativos para criar alpacas y llamas; bastan un número de Has. para los corrales y sistemas de manejo, y la producción de forrajes diversos, como la reina de ellos, la avena; puede ser también la cebada, la alfalfa, y la harina de soya como fuentes de proteína. Algo de sales minerales, comenzando con la sal común, abundante en el país, así como la inoculación de ciertas vitaminas y minerales estratégicos, faltantes en algunas zonas, como el cobre, azufre, selenio.
El mejoramiento genético debe estar dirigido por genetistas de campo y no por aquellos salidos de los escritorios burocráticos de Lima, concertando con los criadores y el Estado, usando todas las técnicas conocidas, especialmente la inseminación artificial y el congelamiento de embriones. Pero donde yo realizaría la batalla principal, es en realizar concertados esfuerzos, para que el criador reciba un precio justo por su producto, y eleve su nivel de vida y el de su familia. Que se invierta la figura y que el criador de alpacas y llamas, “se lleve la parte del león”.





























