Embed Size (px)
DESCRIPTION
biochimie
Citation preview

Hormoni 2 4.03.2014
Mecanism de acţiune H hidrosolubili– proteine G– sisteme generatoare de mesageri secunzi– Hormonii tiroidieni– Homoni pancreatici

• R membranari pot fi împărţiţi în patru clase:
-R cuplaţi cu tirozin kinaze citosolice;
-R cu activitate enzimatică intrinsecă;
-R cu rol de canale ionice;
-R care se leagă de proteine G.

R care formează canale pentru diferiţi ioni
• Trecerea ionului prin canal modifică potenţialul electric de-a lungul membranei.
• Ex, la joncţiunea nerv-muşchi, acetil colina se leagă la un R specific ce permite Na+
să migreze în interior şi K+ să migreze în exteriorul celulei ţintă.
Ligand
Membranã
Citosol
IonLoc de legare pentruligand

Receptori cuplaţi cu tirozin kinaze citosolice
• Pentru: citokine, interferoni, hormonul de creştere, prolactina
• Receptori nu au activitate catalitică intrinsecă

Ligand
Protein-tirozin kinaze (inactive)
P P
ATPADPP P
Proteinã substratATP
ADP
Proteinã substratfosforilatã
P
OH
Receptori cuplaţi cu tirozin kinaze citosolice.

R cu activitate enzimatică intrinsecă localizată în domeniul citosolic
Ex.
• R specific factorului atrial natriuretic catalizează
transformarea GTP la GMPc.
• Alţi R acţionează ca protein – fosfataze activând proteine
prin defosforilarea resturilor de fosfotirozină din structura
acestora.

Ligand
Citosol
Exterior
GTP
3`,5`-GMPc
PPa
R cu activitate guanilat ciclazică generează GMPc la legarea factorului atrial
natriuretic.

R cu activitate tirozin kinazică intrinsecă
• fosforilează resturi de Tyr proprii sau resturi de Tyr ale unor
proteine ţintă. Fosforilarea Tyr este corelată cu proliferarea
celulară.
• Normal, în celulele mamiferelor, din 3000 de resturi
aminoacidice fosforilate, doar unul este de Tyr, restul fiind de
Ser şi Thr.

• 4 clase de receptori cu activitate tirozin kinazică intrinsecă
• Agoniştii în acest caz sunt: insulina şi factorii de creştere.

Factorii de creştere
• sunt polipeptide sintetizate în diferite ţesuturi, care după secreţie pot acţiona:
- autocrin
-paracrin
-endocrin.
• acţionează asupra celulelor:
nervoase
epiteliale
mezenchimale
sanguine

• Clasificare factori de creştere:
-factori de creştere care acţionează asupra unei varietăţi mari de tipuri celulare:
IGF-I (factori de creştere asemănători cu insulina)
EGF (factorul de creştere al epidermei)
PDGF (factorul de creştere eliberat de plachete)
FGF (factorul de creştere derivat din fibroblaşti).
-factori de creştere cu specificitate înaltă de ţesut: NGF (factorul de creştere al nervilor)
eritropoietina (produs de rinichi şi ficat)
limfokine (factori de creştere ai limfocitelor).

Membranã
Domeniu
Domeniuextracelular
N N
C C C
N
intracelular
N N
C C
Domeniutirozinkinazic
1) 2) 3) 4)
Receptori tirozin kinazici.Tipul 1 (pentru EGF) şi tipul 2 (pentru insulină), conţin domenii extracelulare bogate în
cisteină. Tipurile 3 (pentru PDGF) şi 4 (pentru FGF) conţin domenii extracelulare asemănătoare
anticorpilor, iar domeniile kinazice nu sunt continue. In receptorul de tip 2 (pentru insulină), fiecare lanţ este legat prin punţi disulfurice la lanţul şi la celălalt lanţ . Capetele N-terminale ale receptorilor, sunt extracelulare.

• La legarea H la R se activează domeniul catalitic citosolic
al R
Proteinele ţintă, care induc creşterea şi diferenţierea
celulară, sunt activate prin fosforilarea resturilor de Tyr din
structura lor.
• Mutaţiile, care determină activitatea tirozin kinazică
persistentă în aceşti receptori, duc la cancer.

Receptorul insulinic
• sintetizat sub forma unui singur polipeptid
• este glicozilat şi
• clivat în
– două subunităţi şi
– două subunităţi
• care se asociază prin punţi disulfurice într-un tetramer

• La legarea insulinei R se autofosforilează, în prezenţă de ATP, la Tyr
din domeniul catalitic citosolic (protein kinazic-tyrozin specific) al
subunităţilor
• Receptorul fosforilat activează proteine substrat pentru receptorul
insulinic din citosol.
– Sunt cel puţin 4 molecule numite IRS 1-4 (insulin receptor substrates)
• cu structură similară
• dar distribuţie tisulară diferită,
– care ca răspuns la acţiunea hormonului, reglează anumite evenimente intracelulare

YY
Y
Y
YY
Y
YPP
PP
Insulinã
IRS - YIRS - Y -
IRS fosforilat fosforileazã alte kinaze si fosfataze ducând la realizarea rolurilor biologice ale insulinei
Translocarea GLUT-4 Cresterea celularã
Sinteza ADNîn adipocite si muschi scheletici din citosol în membranã
Transcrierea genelor pentru cca. 100 proteine: Glucokinaza, PEPCK, FFK-1, Piruvat kinaza
Activarea enzimelorFosfodiesterazaProtein fosfataze
P

NH3+
-OOC
Extracelular
Intracelular
-helix transmembranar

• Opsina este o proteina transmembranara cu 7 segmente
All-trans-retinal 11-cis-retinal
Opsina
Rodopsinah
impuls nervos

1.H hidrosolubili nu pătrund în celulă
Receptorii pentru H hidrosolubili sunt glicoproteine:
oligomere
multidomeniale
membranare
cu specificitate şi afinitate mare pentru un anumit ligand:

☺☺H hidrosolubili
modifică activitatea:
unor enzime sau
factori de transcriere
existenţi în celule

• Intensitatea răspunsului celular la acţiunea unui H depinde de:
-numărul de receptori
-de gradul de ocupare al acestora cu ligand.

Numărul de receptori poate fi reglat de concentraţia H în sânge:
up regulation (prolactină, angiotensină II)
down regulation (insulină în diabetul zaharat)

• Durata de acţiune
• a H hidrosolubili este mai mică, iar răspunsul celular
este mai rapid
• în comparaţie cu H liposolubili.


Mecanism de acţiune H hidrosolubili– proteine G– sisteme generatoare de mesageri secunzi– Hormonii tiroidieni– Homoni pancreatici

• Hormoni mesageri primi
transmit semnale prin intermediul receptorilor

NH3+
-OOC
Extracelular
Intracelular
-helix transmembranar

• Complexul H–R
– transmite semnale în membrană
• unui sistem efector
– prin intermediul unui sistem de cuplare, reprezentat de proteinele G.
Mesager prim
Receptor Proteinã G Sistem efector
Mesager secund(AMPc, GMPc, IP3, DAG, CA2+)
Rãspuns biologic

Sisteme de cuplare
• Proteinele G
-se află pe partea citosolică a membranei, asociate cu complexul H–R
-leagă nucleotide cu guanină: GDP sau GTP.
-sunt proteine oligomere, formate din trei subunităţi proteice cu
structuri diferite.
leagă nucleotidele GTP sau GDP şi conferă identitatea proteinei G.
, inhibă activitatea subunităţii .

Tipuri de proteine G:
• Gs stimulează adenilat ciclaza;
• Gi inhibă adenilat ciclaza;
• Gq activează fosfolipaza C
• Gt sau transducina activează GMPc–fosfodiesteraza după
stimularea rodopsinei, ca urmare a absorbţiei unui foton de
către retinal

H R
HR
GTP(mediu)
GDP(mediu)
GDP. GTP(forma inactivã) (forma activã)
toxina holericã GDP(inactiv)
H2O
H3PO4
sistemul efector

Mecanismul de actiune al proteinelor G
• înlocuirea GDP cu GTP din mediu
• formarea subunităţii - GTP activă
• inactivarea subunităţii -GTP, ca urmare a activităţii
enzimatice GTP-hidrolazice proprii
• toxina holerică (enzimă produsă de bacteria Vibrio
cholerae) inactivează GTP-aza intrinsecă subunităţii s–
GTP, proteina G rămînînd în forma activă

Sisteme efectoare generatoare de mesageri secunzi
• sistemul adenilat ciclazei;
• sistemul fosfolipazei C;
• sistemul guanilat ciclazei;
• sistemul tirozin kinazei.

Sistemul adenilat ciclazei
• conţine:
-receptori hormonali
-proteine Gs, Gi
-adenilat ciclaza
-fosfodiesteraza


• H care activează AC sunt, prin proteine Gs:
– Glucagonul
– Adrenalina prin receptori -adrenergici 1 şi 2
– ACTH (adrenocorticotrop)
– Parathormonul, Calcitonina
– MSH, LH, TSH, somatostatina
• H care inhibă activitatea AC, prin proteine Gi:
– adrenalina prin receptori 2

Adenilat ciclaza • enzimă membranară, activată de Gs şi inhibată de Gi
• catalizează formarea AMPc din ATP
NO
H2CO
HOHO
P
O
O-
P
O
O
O-
O-
O-
O
O
P N
NN
NH2
ATP
OH2CO
OHO
N N
NN
NH2
P
O-
O
Adenilatciclaza
HR
AMPc

Fosfodiesteraza • Se află în citosol şi catalizează transformarea AMPc în AMP.
• Este activată de: Ca2+, insulină, prostaglandine, şi este inhibată de
metilxantine (cofeină, teofilină), tiroidieni, steroizi.
OH2CO
OHO
N N
NN
NH2
P
O-
O
AMPc
Fosfodiesteraza
Metilxantine
InsulinãCa2+-Calmodulinã 5`-AMP
NO
H2CO
HOHO
P
O
O-
O- N
NN
NH2

Mesagerii secunzi
• poartă informaţia transmisă de primul mesager
(hormonul):
-modifică activitatea unor proteine intracelulare
-intervin în contracţie sau în asigurarea permeabilităţii membranare.

AMPc • este un nucleotid ciclic, mesager secund pentru unii hormoni.
• concentraţia intracelulară a AMPc depinde de: AC şi
fosfodiesterază
• Activitatea reglatoare a AMPc:
-la procariote, controlează exprimarea unor gene prin legarea
cu o proteină reglatoare;
-la eucariote, activează protein-kinaze specifice, ex. protein
kinaza A.

Protein kinaze AMPc dependente • Protein kinaza-AMPc dependentă este un tetramer :
-2 subunităţi reglatoare (R);
-2 subunităţi catalitice (C).
• Tetramerul R2C2 este inactiv.
– subunităţile R din tetramerul inactiv, leagă cîte două molecule de
AMPc
• Protein kinaza activă, activează prin fosforilare o proteină
celulară specifică.

»Cine confera specificitate raspunsului hormonal ?

• Particularitatea răspunsului celular depinde de:
– natura proteinelor substrat pentru protein kinaze şi
– de modul în care fosforilarea afectează funcţia proteinei.

• Cum se stinge mesajul hormonal ?

Fosfoprotein–fosfatazele• aduc proteinele fosforilate la starea anterioară
• prezintă multiple izoforme
• activitatea lor este reglată hormonal
• intervin în:
– procesele de diviziune celulară
– transcrierea şi traducerea genelor
– permeabilitatea membranară
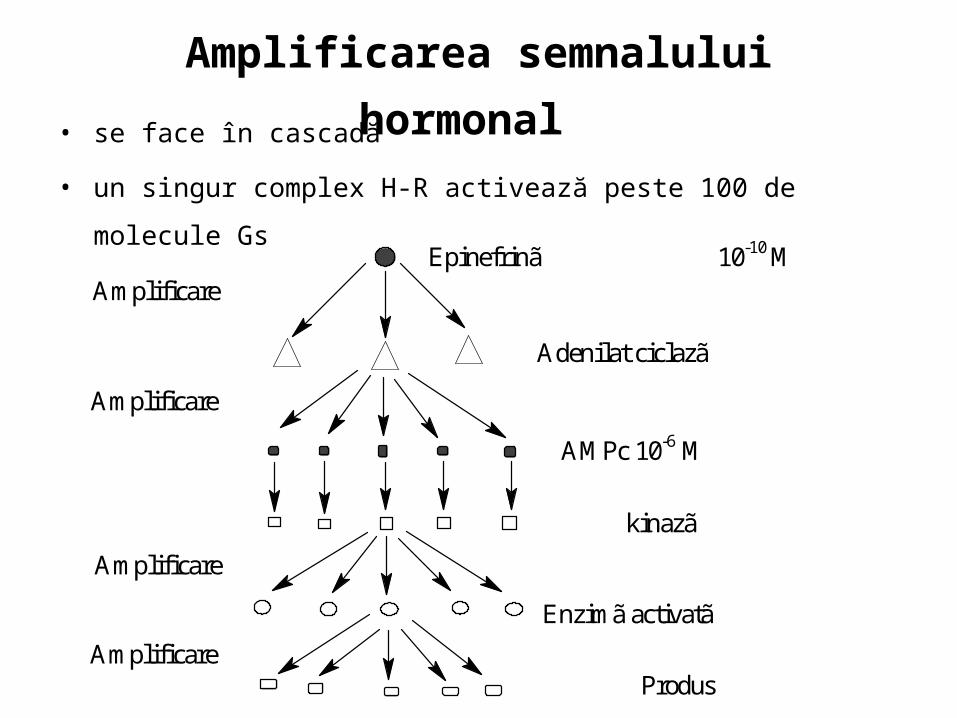
Amplificarea semnalului hormonal • se face în cascadă
• un singur complex H-R activează peste 100 de molecule Gs
Epinefrinã 10-10 MAmplificare
Adenilat ciclazã
AMPc 10-6 M
kinazã
Enzimã activatã
Produs
Amplificare
Amplificare
Amplificare





STINGEREA MESAJULUIHORMONAL ÎN SISTEMULADENILAT CICLAZEI
au loc astfel:
1) PKA fosforilează R şiîi inactivează2) Fosfodiesteraza scade conc. intracelulară de AMPc3) Fosfoprotein fosfatazele defosforilează Ser şi Thr din proteine


Sistemul fosfolipazei C
• receptori hormonali,
• proteine Gq,
• fosfolipaza C,
• protein kinaza C


Fosfolipaza C
• enzimă membranară
• catalizează hidroliza fosfatidil inozitol 4,5-bisfosfatului (PIP2)
• Produşii reacţiei de hidroliză sunt:
– inozitol 1,4,5-trisfosfatul (IP3), şi
– diacilglicerolul (DAG).

HC
H2C
O
O
CO
CO R1
R2
H2C O P
O
O
OH
OPO3H2
OHOH
OH
OPO3H2
Fosfatidilinozitol 4,5-bisfosfat (PIP2)
Fosfolipaza CHC
H2C
O
O
CO
CO R1
R2
H2C OH
PO3H2O
OPO3H2
OHOH
OH
OPO3H2
+
(IP3)(DAG)Diacilglicerol Inozitol 1,4, 5-trisfosfat

IP3
• determină eliberarea Ca2+ din reticulul endoplasmic în citosol
• Ca2+ este o entitate chimică permanentă
• concentraţia Ca2+ poate fi modulată numai prin deplasări între diverse
compartimente
• Ca2+ citosolic pentru o celulă de mamifer este de 10-7-10-6 M
• Ca2+ extracelular este 10-3 M

• Concentraţia mică a Ca2+ în citosol este menţinută prin:
– pompe de Ca2+
– permeabilitatea naturală foarte mică a membranelor pentru
cationi
– mecanisme care depozitează Ca2+ în mitocondrii şi reticul
endoplasmic (reticul sarcoplasmic în muşchi)

• Ca2+ acţionează ca mesager secund împreună cu:
– AMPc
– GMPc
– DAG sau
– IP3
• sau
independent de aceştia
• Rolurile reglatoare ale Ca2+ sunt mediate de proteine specifice de
legare a cationului

Calmodulina
• proteină formată din 148 aa
• prezentă în toate celulele eucariote în
concentraţie mare
• nu este limitantă pentru un proces dat
• poate fixa 4 ioni de Ca2+/ moleculă


• complexul Ca2+- calmodulină:
-reglează contracţia muşchilor netezi
-activează pompa de Ca2+ din membrana plasmică
-reglează activitatea unor protein kinaze modulatori ai altor enzime
-interacţionează cu celelalte sisteme mesageriale prin reglarea
activităţii AC, GC, fosfodiesterazelor.

DAG
• moleculă liposolubilă, activează protein kinaza C.
• DAG este inactivat prin hidroliză.

• Hormonii care au ca mesager secund IP3, Ca2+, sau DAG sunt:
-epinefrina prin receptori 1-adrenergici
-gastrina
-oxitocina
-colecistokinina
-hormonul eliberator al gonadotropinelor.

Protein kinaza C
• enzimă membranară cu multe izoforme
– unele sunt activate de DAG
– altele necesită pe lîngă DAG şi Ca2+
• are specificitate pentru resturile de serină şi treonină.


Sistemul guanilat ciclazei
• guanilat ciclaza
• GMPc mesager secund
• protein kinaze GMPc dependente.


GMPc
• GMPc antagonizează efectele AMPc.
• GMPc este mediatorul semnalelor luminoase în celulele retiniene cu
conuri şi bastonaşe.
• hiperpolarizarea membranei cu perceperea senzaţiei de lumină.
Rodopsina+h
R
(opsina - proteinã cu 7 segmente transmembranare)
T-GTPTransducinã
PDEDeschide canale pentru Na+
Inchide canale pentru Na+
GMPc
GMP(opsinã + 11-cis retinal)

All-trans-retinal 11-cis-retinal
Opsina
Rodopsinah
impuls nervos

Guanilat ciclaza
• prezintă cel puţin 2 izoforme:
membranară şi una
citoplasmatică.

• Guanilat ciclaza membranară :
-funcţia de receptor al semnalului extern
-cît şi funcţia de sistem efector care
generează (GMPc).

Factorul atrial natriuretic (natriopeptină) - 28 aa la om -secretat de celulele cardiace atriale -acţionează:
renal zona glomerulară a suprarenalelor
prin:creşte volumul urinar, creşte eliminarea de , scade secreţa de renină şi aldosteron.
Na

Guanilat ciclaza citoplasmatică
• sistem efector pentru oxidul de azot
(endothelium-derived relaxing factor, EDRF)
NOS endotelială - EDRFNOS inductibilă – macrofage sau hepatocite
-stimulare imunologică sau reacţii inflamatoriiNOS neuronală
.NO
L-Argininã L-Citrulinã
NO.
NO-sintazã

Nitric oxid sintaza (NOS) are structură asemănătoare cu citocromul P-450.
Pentru elucidarea rolului acestei enzime în stările normale şi patologice, s-a primit premiul Nobel în 1998.
Au fost identificate 3 izoenzime ale NOS: NOS-I (de origine neuronală), NOS-II (din macrofage sau inductibilă), NOS-III (din endoteliul vascular).
FAD FMN
NADPH
e- e-
e-
HEM
BH4
L-Argininã
Calmodulinã
ReductazãOxigenazã
L-Citrulinã + NO.

NO acţionează paracrin asupra multor celule:
-reglează presiunea sanguină,
-mediator în citotoxicitate şi în neurotransmisie.
-în celulele ţintă
-Hemoglobina are un rol important în transportul oxidului de azot.
NO. Guanilat
ciclaza
GTP
GMPc Mediazã relaxarea muschilor netezi
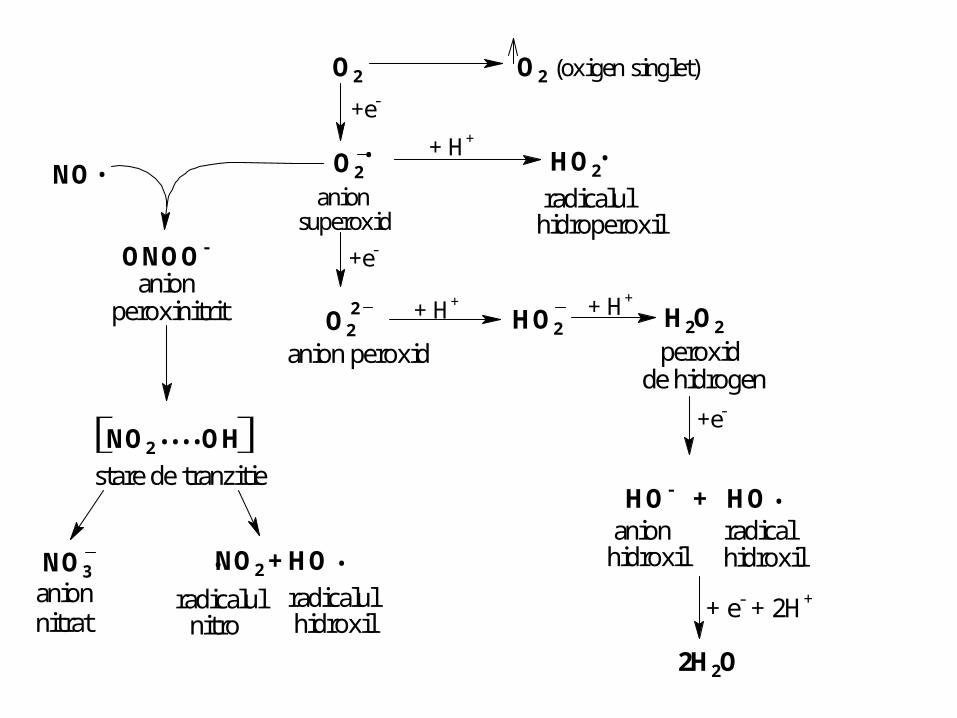
.O2
.
O2 O2 (oxigen singlet)
+e-
anionsuperoxid
+ H+
HO2
radicalulhidroperoxil
+e-
O22
anion peroxid
+ H+HO2
+ H+
H2O2
peroxidde hidrogen
NO.
ONOO-
anionperoxinitrit
NO2 OH....stare de tranzitie
NO3anion nitrat
NO2 + HO..radicalul
nitroradicalulhidroxil
+e-
HO- + HOanion radical
.
hidroxil hidroxil
+ e- + 2H+
2H2O


Hormonii tiroidieni
• Hormonii tiroidieni sunt derivaţi ioduraţi ai compusului
ipotetic, tironină, derivat de la tirozină.
OOH CH2 CH
NH2
COOH1
23
4
5 6
1`
2`3`
4`
5` 6`
Tironina
OOH CH2 CH
NH2
COOH
I I
I
OOH CH2 CH
NH2
COOH
I
I
I
I
3, 5, 3`, 5` - Tetraiodotironinã (Tiroxinã) T4
3, 5, 3` - Triiodotironinã (T3)
OOH CH2 CH
NH2
COOH
I
I
I
3, 3`, 5` - Triiodotironina (revers T3) (rT3)

Biosinteza
Unde are loc sinteza ?
• în celulele tiroidiene şi
• la interfaţa dintre celulele tiroidiene şi coloid
• Ce este coloidul ?– o soluţie omogenă de tireoglobulină

• Ce ete tireoglobulina ?
– Glicoproteină (Proteina + resturi glucidice):
• 8-10 % glucide şi peste
• 100 resturi tirozil pe moleculă.

• Etapele biosintezei:
Captarea iodului în tiroidă
-prin difuzie simplă
-prin transport activ, dependent de ATP (pompa de iod),
Iodul din plasmă, (I-) poate fi:
-exogen, din apa potabilă, alimente, sare iodată;
-endogen, rezultat din catabolismul hormonilor tiroidieni

• Oxidarea I- I+ (agent electrofil) – de către H2O2 formată intracelular sub acţiunea NADPH
oxidazei
(localizată pe suprafaţa apicală a celulelor tiroidiene)
NADPH + H+ + O2 NADP+ + H2O2 tireoperoxidazã
2 I- + H2O2 + 2 H+ 2 I+ + 2 H2O

• Iodurarea resturilor Tyr din tireoglobulină – glicoproteină cu:
• 8-10 % glucide şi peste
• 100 resturi tirozil pe moleculă.
Tireoglobulina este iodurată (de tireoperoxidază)
monoiodotirozină (MIT) şi
diiodotirozină (DIT).

CO
NH
CH CH2 OH
CO
NH
CH CH2 OH
Tireoglobulinã
+ n I+
CO
NH
CH CH2 OH
I
CO
NH
CH CH2 OH
I
I
Tireoglobulinã ioduratã
(MIT)
(DIT)

• Cuplarea
– MIT cu DIT ;
–DIT cu DIT
cu formarea de T3 şi T4 ca părţi componente ale
tireoglobulinei

CO
NH
CH CH2 OH
I
CO
NH
CH CH2 OH
I
I
(MIT)
(DIT)
CO
NH
CH CH2 OH
I
I
CO
NH
CH CH2 OH
I
I (DIT)
CO
NH
CH CH2 O
I
OH
I
I
CO
NH
CH CH3
CO
NH
CH CH2 O
I
I
OH
I
I
T3
CO
NH
CH CH3
T4
(DIT)

• Endocitoza tireoglobulinei din coloid în celule,
eliberarea de T3 şi T4 prin hidroliza
tireoglobulinei şi secreţia acestora în sânge.
• Eliberare: T3 ,T4 , MIT, DIT şi diverşi aminoacizi
care intră în fondul metabolic celular.

• Tiroida secretă în principal T4 şi
mai puţin T3.
• T3 din sânge provine şi din deiodurarea
periferică a T4.
• T3 este de câteva ori mai activ biologic decât
T4.

• Transportul HT:
– TBG (thyroxine binding globulin), cu afinitate mare pentru T4.
Sinteza acestei proteine este stimulată de estrogeni şi micşorată de androgeni şi glucocorticoizi.
– TBPA (thyroxine-binding prealbumin), leagă numai T4
– Serumalbumina

Metabolismul hormonilor tiroidieni
• T1/2:
– 6-7 zile pentru T4
– 1,5 zile pentru T3
• Deiodurarea în ţesuturile periferice: ficat, rinichi, muşchi.
Prin monodeiodurare:
- 5`-deioduraza transformă 30-40% din T4 T3 biologic activ
- 5 -deioduraza transformă 40% din T4 rT3 biologic inactiv
-T3 şi rT3 sunt deioduraţi la diiodotironine şi monoiodotironine
inactive.

OOH CH2 CH
NH2
COOH
I I
I
OOH CH2 CH
NH2
COOH
I
I
I
I
3, 5, 3`, 5` - Tetraiodotironinã (Tiroxinã) T4
3, 5, 3` - Triiodotironinã (T3)
OOH CH2 CH
NH2
COOH
I
I
I
3, 3`, 5` - Triiodotironina (revers T3) (rT3)

• Conjugarea cu acidul glucuronic sau sulfuric şi
eliminarea prin bilă a produşilor de conjugare
• Transaminare şi decarboxilare oxidativă a lui T4
şi T3 cu formarea cataboliţilor solubili, excretabili
prin urină.

Roluri fiziologice
• La concentraţii normale sau mici manifestă efecte anabolizante.
• stimulează sinteza hormonului de creştere.• controlează metabolismul oxidativ. • stimulează oxigenarea ţesuturilor.• creşte sinteza Na+/K+-ATP azei (pompă
consumatoare de ATP), prin acţiune la nivel nuclear.
• Au roluri esenţiale în dezvoltarea fetală şi postnatală.

• La concentraţii mari manifestă efecte catabolizante.
-creşte viteza metabolismului bazal;
-micşorează rezervele energetice glucidice şi lipidice;
-catabolismul proteic se intensifică.
-nivelul colesterolului scade în hipertiroidism.

Reglarea secreţiei
• Inhibiţia prin feed-back pe axul hipotalamo
hipofizar a eliberării HT
– participă numai T3, prezent ca atare în sânge sau
format prin deiodurarea T4.
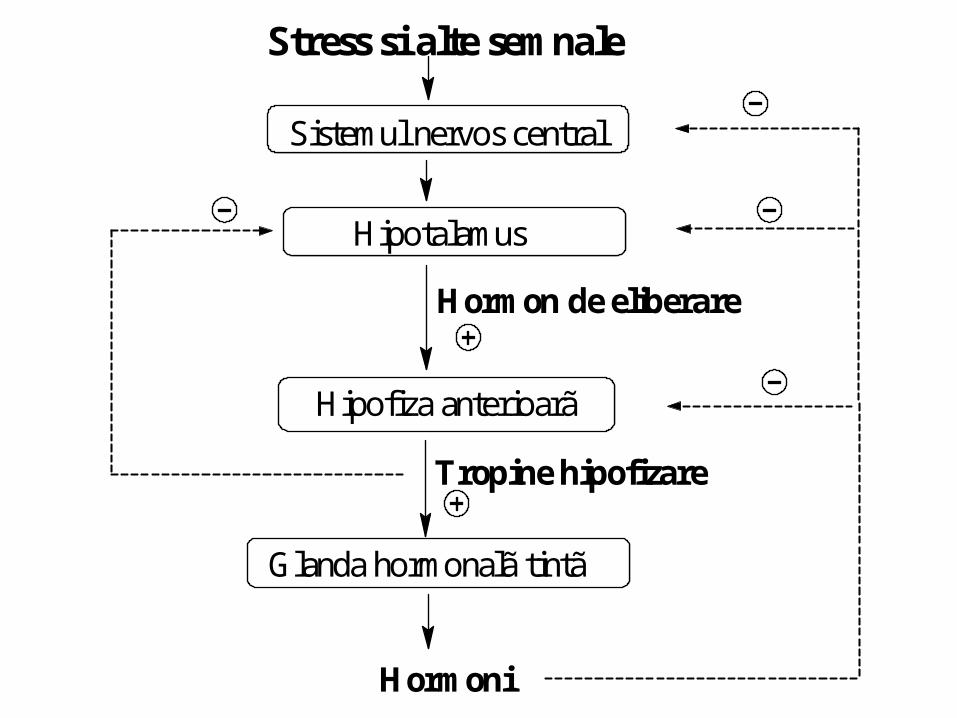
Stress si alte semnale
Sistemul nervos central
Hipotalamus
Hipofiza anterioarã
Glanda hormonalã tintã
Hormoni
Hormon de eliberare
Tropine hipofizare

• Somatostatina (“antihormonul“ de creştere) inhibă
secreţia de TSH.
– secreţia de somatostatină este stimulată de concentraţiile mari de
T3.
• Reglarea prin transformarea T4 în T3 activ biologic şi
rT3 inactiv biologic.
– conversia T4 —> rT3 este favorizată în inaniţie pentru a diminua
arderile şi consumul de materiale energogene.

Cu privire la sinteza hormonilor tiroidieni, sunt corecte următoarele afirmaţii:A.Iodurarea se realizează le nivelul tirozinei libereB.Triiodotironina este precursor al tiroxineiC.Se află sub controlul unui sistem de feed-back pozitiv la care participă TSHD.Revers T3 se obtine din T4 sub acţiunea unei 5’ -deiodurazeE.Apa oxigenată necesară oxidării I- la I+ este furnizată de NADPH oxidază


Hormonii pancreatici
• Pancreasul format din ţesuturi:
-endocrine
-exocrine.

☺Funcţia endocrină
insulele Langerhans (1-2 % din masa totală) conţin:
-celule A (28% din numărul total de celule), care produc şi secretă glucagon;
-celule B (70%) -insulină;
-celule D (5%) - somatostatină;
-celule F, polipeptidul pancreatic care reglează diverse funcţii ale tractului intestinal.

Insulina
• domină faza anabolică a metabolismului
• exercită acţiuni mitogene specifice factorilor
de creştere

Structura şi biosinteza
• 2 lanţuri peptidice, unite prin punţi de sulf:
–lanţul A - 21 aa şi
–lanţul B - 30 aa.
pre - pro - insulinãreticul
endoplasmicpro - insulinã
aparatGolgi
insulinã


Reglarea secreţieiFactorii care activează secreţia de insulină:
• -creşterea glicemiei glicemia normală este: 80-100 mg/dl ser
• aa (Leu, Lyz, Arg), acizii graşi, corpii cetonici;
• “Hormoni candidaţi”: enteroglucagon, GIP (gastric inhibitory peptid), VIP (vasoactive intestinal peptide)

Factori care inhibă secreţia de insulină
• adrenalina şi noradrenalina
prin receptori de tip 2
• somatostatina produsă de celulele D din pancreas,
prin acţiune paracrină.

Metabolizare
• T1/2 este de 3-5 minute.
• Insulina este inactivată în ficat prin:
-desfacerea legăturilor disulfurice dintre lanţuri, cu
participarea G-SH;
-cu ajutorul proteazelor specifice.

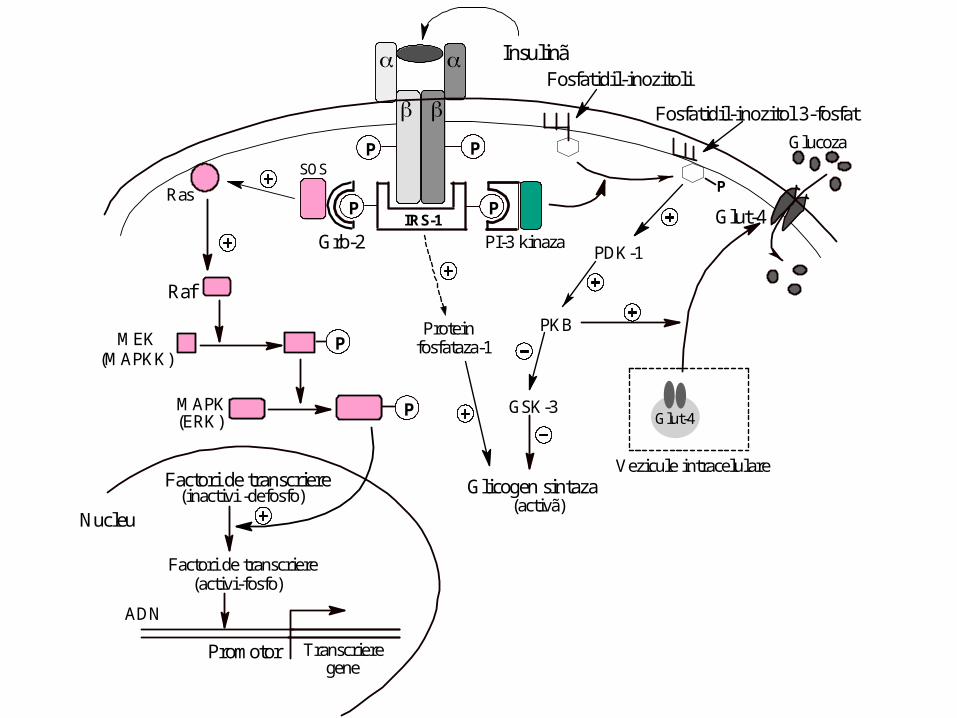
Mecanisme de acţiune
• Nr. Receptori
• La nivel membranar
-stimulează transportul glucozei în ţesuturile insulino dependente
(muscular şi adipos) prin translocarea transportorului pentru glucoză
GLUT 4 din veziculele intracelulare la nivelul membranei (secunde)
52 1010
Insulinã
P P
IRS-1P P
Grb-2
SOS
PI-3 kinaza
Ras
Raf
(MAPKK)MEK P
MAPK(ERK)
P
Nucleu
ADN
Promotor
Factori de transcriere (inactivi -defosfo)
Factori de transcriere(activi-fosfo)
Transcrieregene
Fosfatidil-inozitoli
Fosfatidil-inozitol 3-fosfat
P
Glucoza
Vezicule intracelulare
Glut-4
PDK-1
PKB
GSK-3
Glicogen sintaza (activã)
Proteinfosfataza-1
Glut-4

• modifică activitatea enzimelor din multe tipuri de celule
(minute pînă la ore) prin modificarea stării de fosforilare a
enzimelor existente:
-proteinfosfataze
-fosfodiesteraze
-lipoprotein lipaza
-glicogen sintaz kinaza 3 (GSK 3), etc).

• iniţiază procesul de creştere a concentraţiei multor enzime (ore
până la zile):
-glucokinaza
-fosfofructokinaza
-piruvat kinaza
prin intensificarea transcrierii genelor, sinteza ARNm şi în final
sinteza enzimelor.


Implicatiile metabolice ale insulinei
• Hormon
-anabolizant
-hipoglicemiant

☺In absenţa insulinei ce ţesuturi utilizează
glucoza pentru a produce energie ?
☺ numai creierul şi eritrocitul

☺ Dar în prezenţa insulinei ?
☺ toate celulele din organism comută pe
utilizarea glucozei

Insulina efecte metabolice:
-intensifică transportul glucozei în celule
-intensifică glicoliza în scop anabolic
-stimulează sinteza de glicogen în ficat şi muşchi
-inhibă gluconeogeneza
-stimulează lipogeneza în adipos.

În ficat
• stimulează glicoliza în scop anabolic. – favorizează indirect influxul de glucoză în
hepatocite– activează glucokinaza
• favorizează glicogeneza=glicogenogeneza
• stimulează biosinteza proteică prin accelerarea transportului aa prin membrane


În muşchii scheletici
• stimulează sinteza de glicogen
• stimulează sinteza proteică
• încetineşte degradarea proteinelor.

În ţesutul adipos
• facilitează transformarea glucozei în glicerolfosfat.
• activează lipoprotein-lipaza
• stimulează lipogeneza şi inhibă lipaza hormon sensibilă
• scade producţia de corpi cetonici (acid aceto acetic şi acid -hidroxibutiric).

• inhibă lipoliza în adipocite la concentraţii mici ale
insulinei (5mU/ml);
• activează lipogeneza (sinteza de lipide), în aceleaşi
celule, la concentraţii ale insulinei de 10 ori mai mari.

Insulina acţionează ca factor de creştere
• stimulează proliferarea unor tipuri de celule.
• Insulina şi receptorul insulinic, prezintă analogie structurală cu IGF-I şi IGF-II (insuline-like growth factors).

Diabetul
• Diabetul insipid
-sete şi urinare frecventă (urina este insipidă - nu conţine glucoză)
-lipsa H antidiuretic = vasopresină sau
-defect al receptorilor renali ai acestui hormon.

Diabetul insulino dependent IDDM • distrugerea celulelor secretorii din insulele
Langerhans, auto imun.
• În IDDM netratat domină starea catabolică
-degradarea proteinelor tisulare şi
-mobilizarea rezervelor energetice.

• Lipsa de insulină
• secreţia de glucagon este crescută
• Stare generală de stress
– Se activează medulosuprarenalele

• Acizi graşi neesterificaţi pot ajunge la 3-4 mmol/l (normal 0,2-1,0 mmol/l)
• corpi cetonici 0,2 mmol/l cresc la 10-20 mmoli/l
• pH-ul sanguin poate să scadă de la valoarea normală 7,4 la aprox. 7,0, situaţie cunoscută sub numele de ceto-acidoză diabetică.

Diabetul insulino independent
• se caracterizează prin rezistenţă la insulină
caracteristică a obezităţii.
• În NIDDM nu se dezvoltă cetoacidoza.
• NIDDM apare la persoane adulte.

Glucagonul
• secretat de celulele A din insulele Langerhans
• hormon hiperglicemiant.

Structură, biosinteză
• Metabolizare în ficat
-T1/2 -5 minute.
pre - pro - glucagonreticul
endoplasmicpro - glucagon
aparatGolgi
glucagon

• Reglarea secreţiei.
Factorul reglator este glucoza
• Factori care stimulează secreţia:
-aminoacizi;
-agonişti -adrenergici
• Factori care inhibă secreţia:
-glucoza;
-somatostatina, produsă de celulele D din pancreas.

Mecanism de acţiune
• receptor membranar
– proteine Gs
• mesageri secunzi (AMPc)

NH3+
-OOC
Extracelular
Intracelular
-helix transmembranar


Rol fiziologic• Acţiune metabolică opusă insulinei.• În ficat:
-stimulează gluconeogeneza
-inhibă glicoliza
-activează glicogenoliza
-stimulează -oxidarea acizilor graşi şi cetogeneza.
-conversia glucozei în acizi graşi şi sinteza de trigliceride este diminuată.

• În muşchi, metabolismul glucidic nu este influenţat de glucagon.
• În ţesutul adipos
-activează lipoliza, AG eliberaţi în sânge fiind preluaţi de ficat şi supuşi -oxidării mitocondriale.
-inhibă lipogeneza





























