Embed Size (px)
Citation preview

STRUTTURE DEGLI ACIDI STRUTTURE DEGLI ACIDI STRUTTURE DEGLI ACIDI
NUCLEICI,NUCLEICI,NUCLEICI,
CODICE GENETICO CODICE GENETICO CODICE GENETICO
EEE
REPLICAZIONE DEL DNAREPLICAZIONE DEL DNAREPLICAZIONE DEL DNA
26 Novembre 2011Auditorium Presidio Ospedaliero S.Chiara,
Trento
IL LABORATORIO DI BIOLOGIA MOLECOLARE:
Introduzione alle tecniche e alle loro applicazioni
Relatore: Lorenzo Zeni
Il DN
Anell’a
rte: S
usan R
ankaitis
2002, “D
NA
2”

Il corso ha l’obbiettivo di dare le
conoscenze di base
sulla struttura e
sulle funzioni
degli acidi nucleici
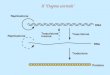
Introduzione
La struttura primaria:La struttura primaria:
i componenti degli acidi nucleicii componenti degli acidi nucleici
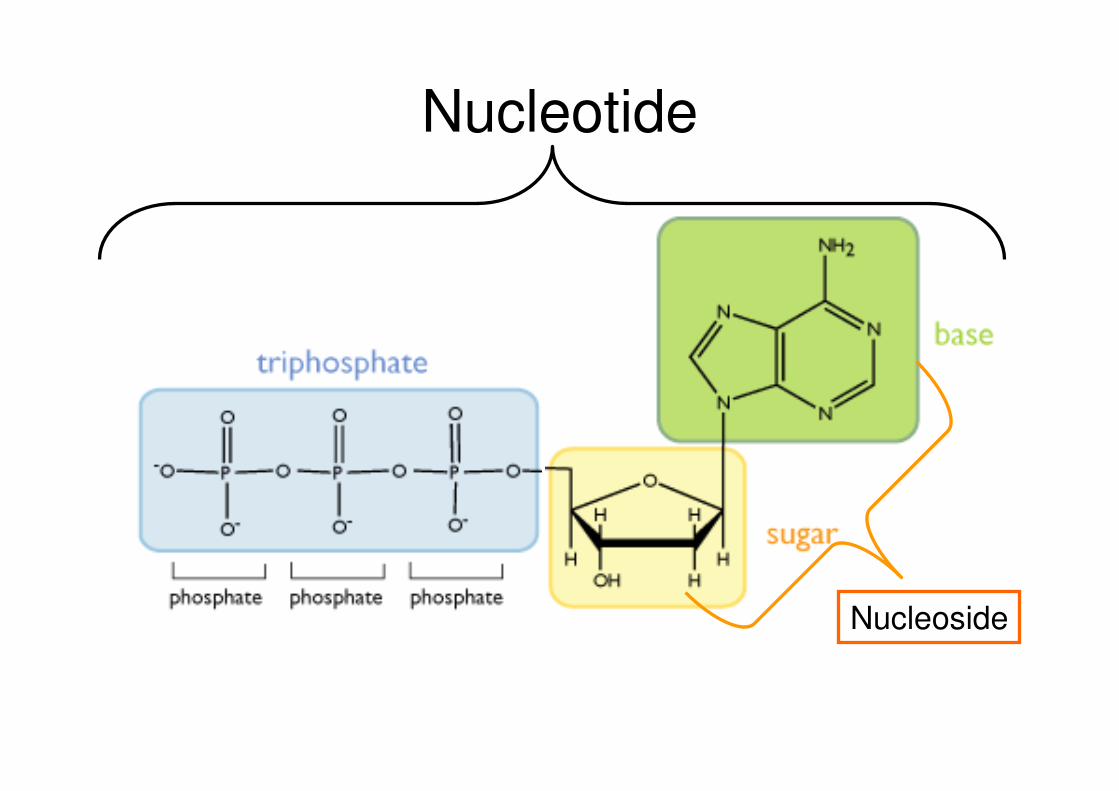
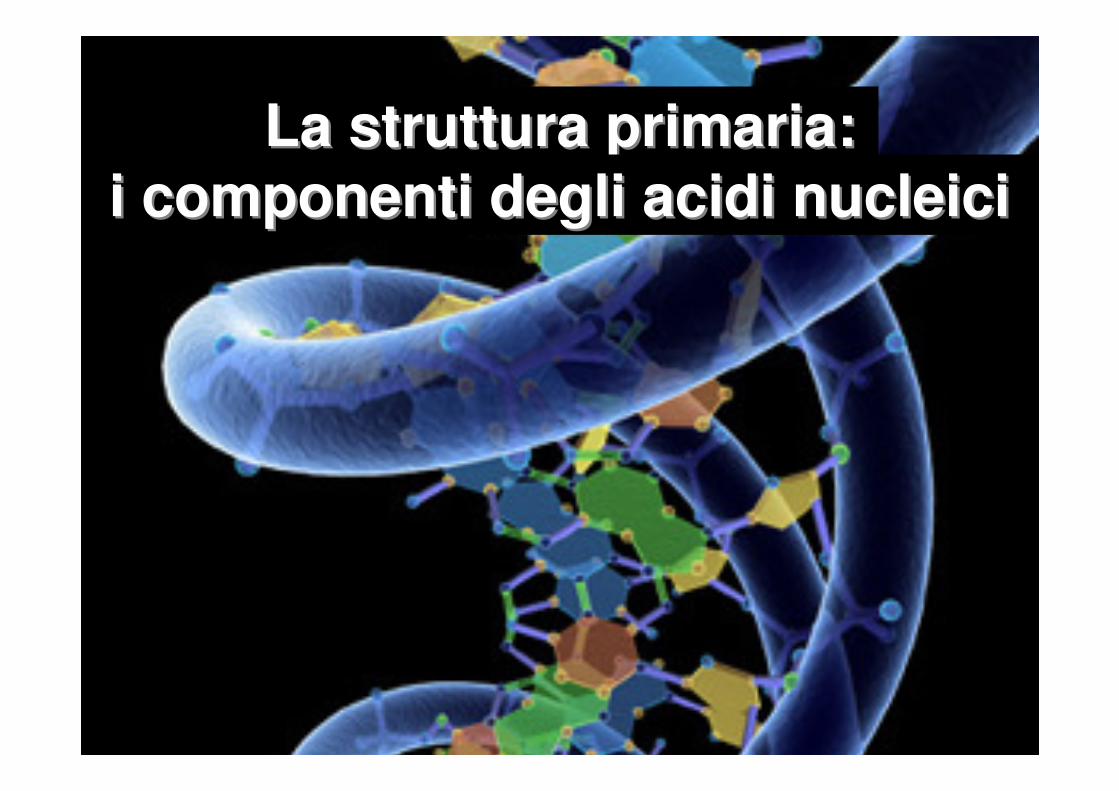
Nucleotide
Nucleoside
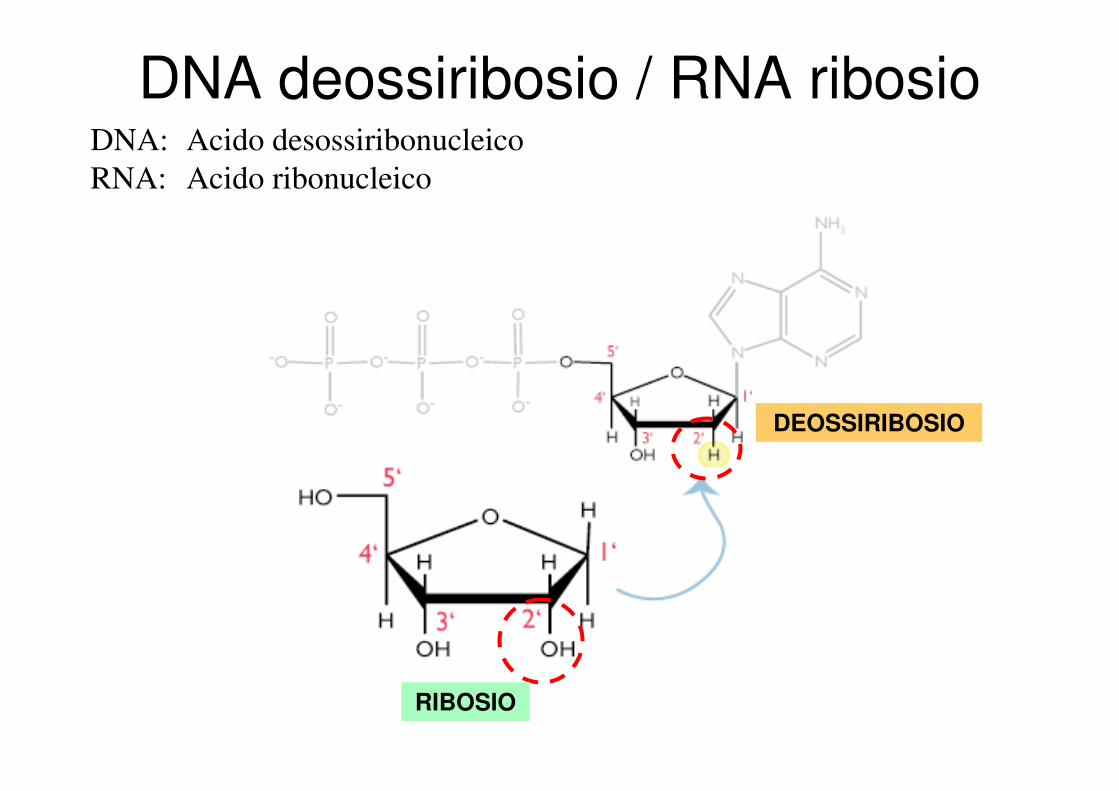
DNA deossiribosio / RNA ribosioDNA: Acido desossiribonucleico
RNA: Acido ribonucleico
RIBOSIO
DEOSSIRIBOSIO
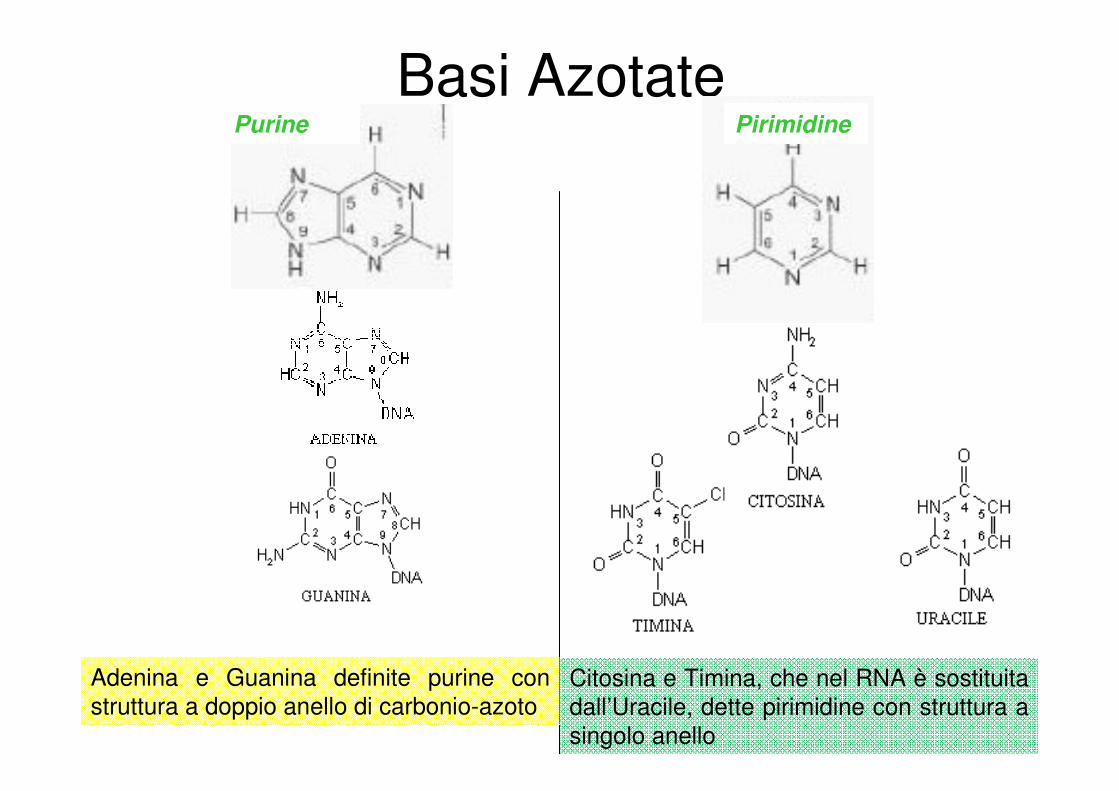
Basi AzotatePurine Pirimidine
Adenina e Guanina definite purine con
struttura a doppio anello di carbonio-azoto Citosina e Timina, che nel RNA è sostituita
dall’Uracile, dette pirimidine con struttura a
singolo anello
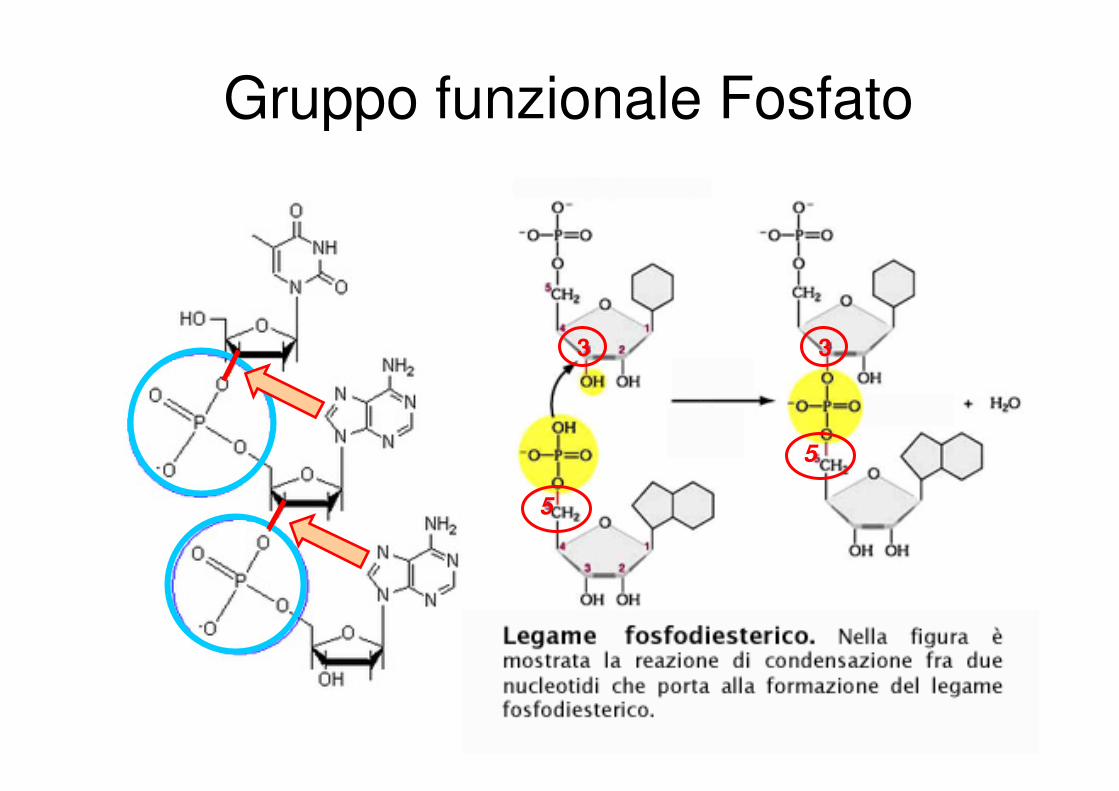
Gruppo funzionale Fosfato
3 3
5
5
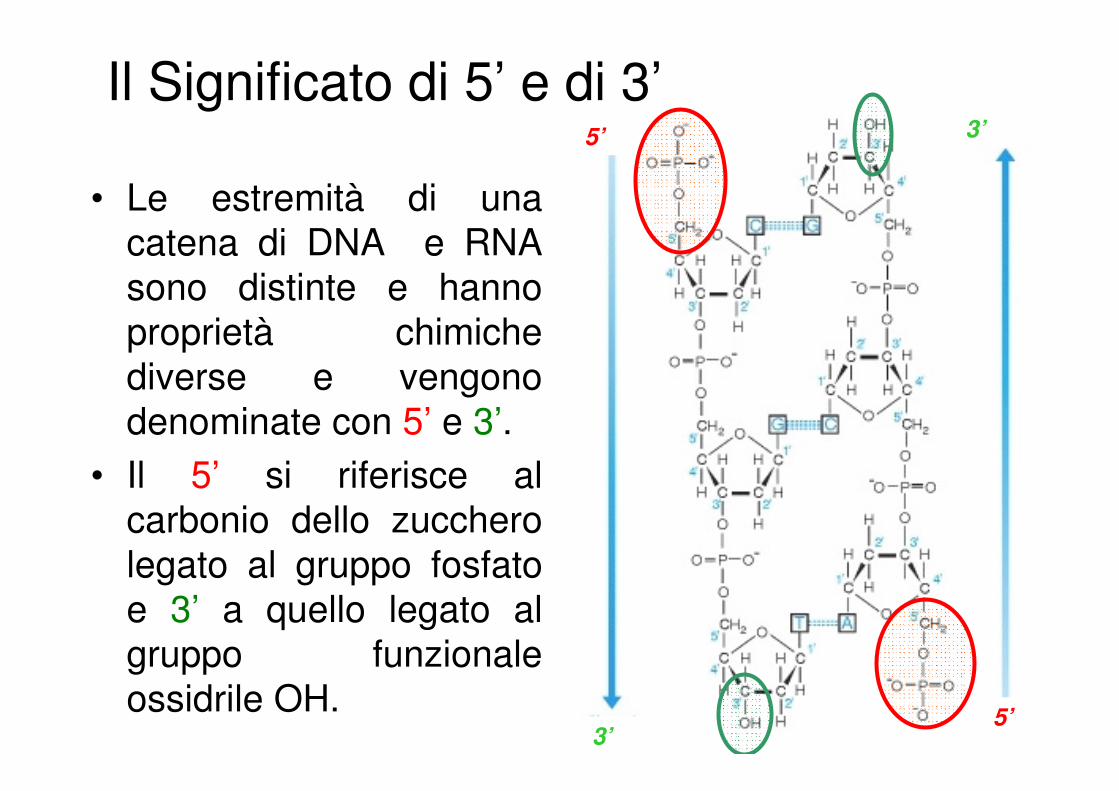
Il Significato di 5’ e di 3’
• Le estremità di una
catena di DNA e RNA
sono distinte e hanno
proprietà chimiche
diverse e vengono
denominate con 5’ e 3’.
• Il 5’ si riferisce al
carbonio dello zucchero
legato al gruppo fosfato
e 3’ a quello legato al
gruppo funzionale
ossidrile OH.
5’
5’3’
3’
La struttura secondaria del DNALa struttura secondaria del DNA
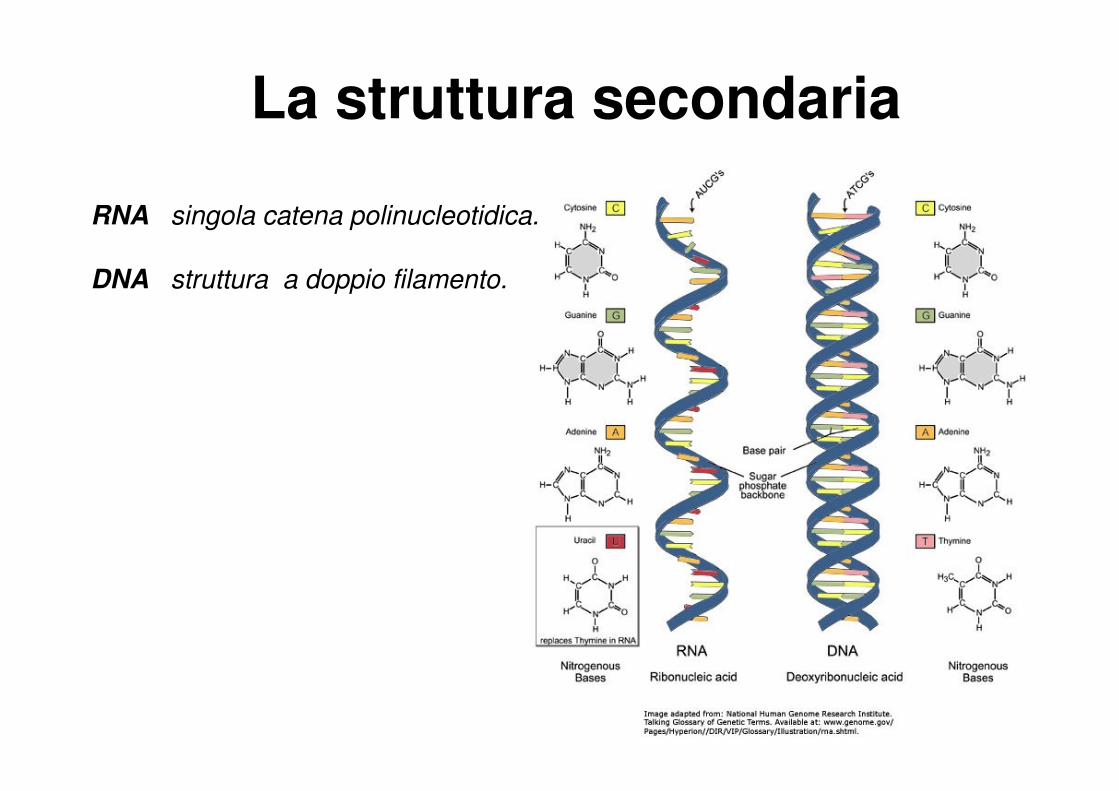
RNA singola catena polinucleotidica.
DNA struttura a doppio filamento.
La struttura secondaria
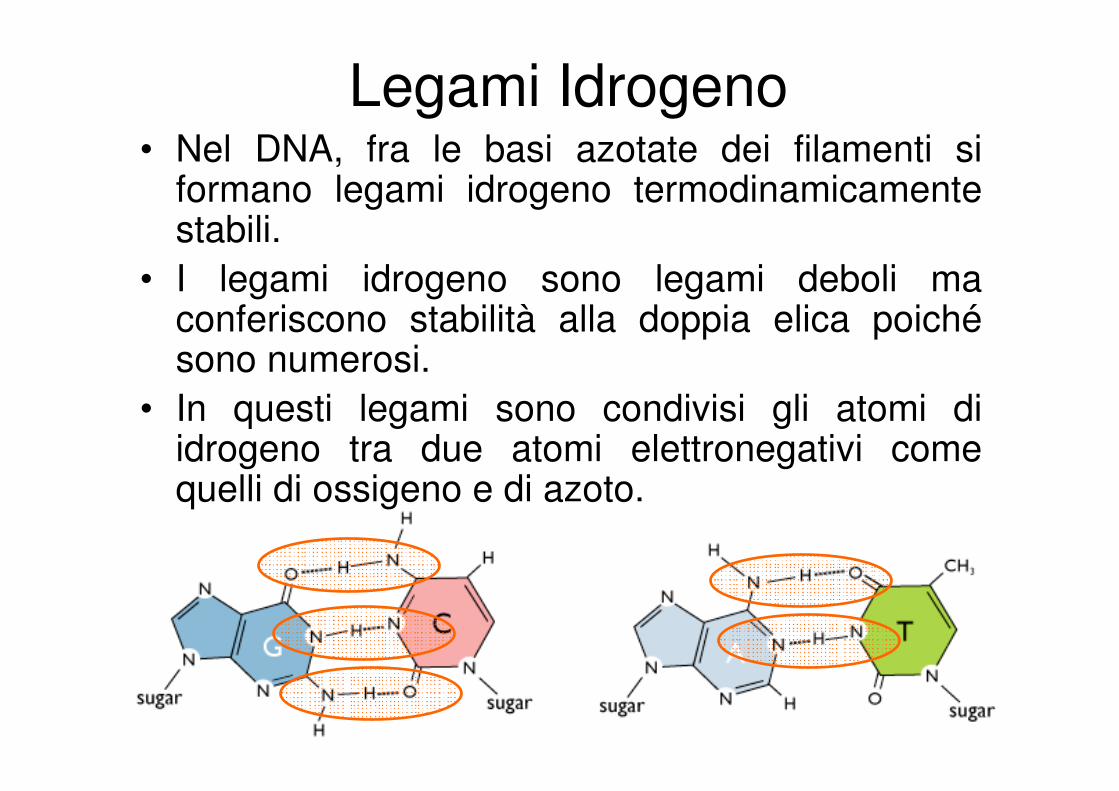
Legami Idrogeno• Nel DNA, fra le basi azotate dei filamenti si
formano legami idrogeno termodinamicamente stabili.
• I legami idrogeno sono legami deboli ma conferiscono stabilità alla doppia elica poichésono numerosi.
• In questi legami sono condivisi gli atomi di idrogeno tra due atomi elettronegativi come quelli di ossigeno e di azoto.
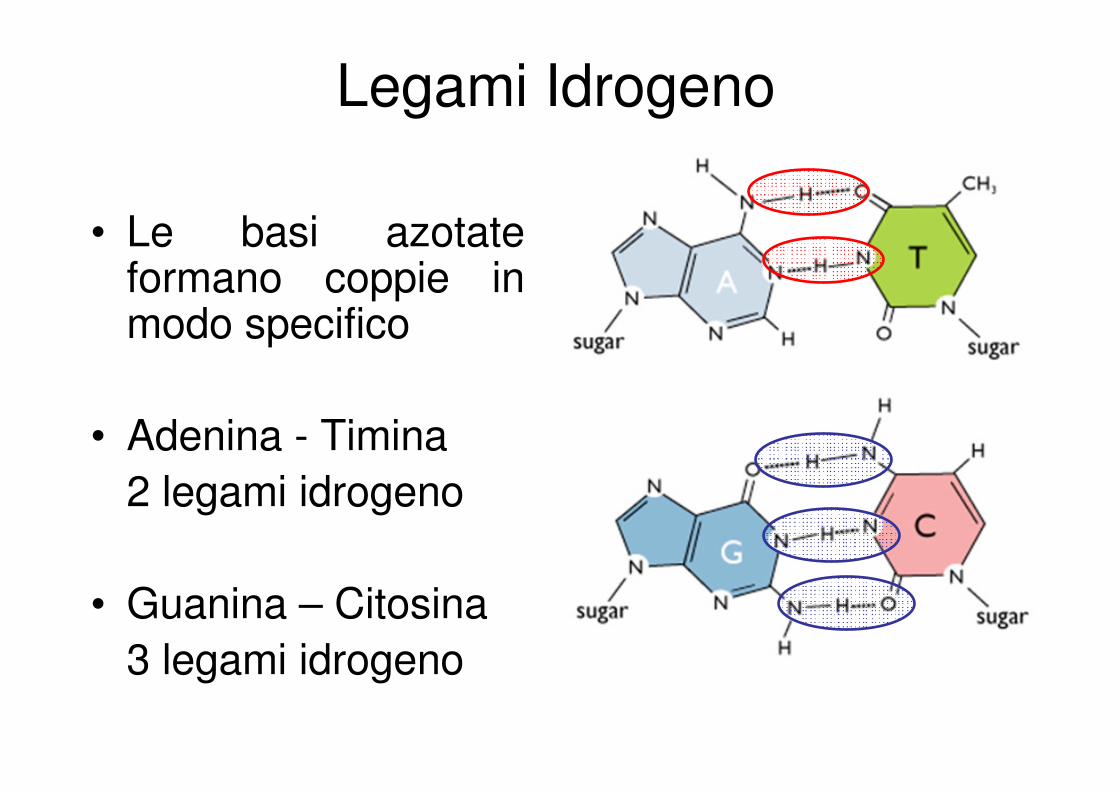
Legami Idrogeno
• Le basi azotate formano coppie in modo specifico
• Adenina - Timina
2 legami idrogeno
• Guanina – Citosina
3 legami idrogeno
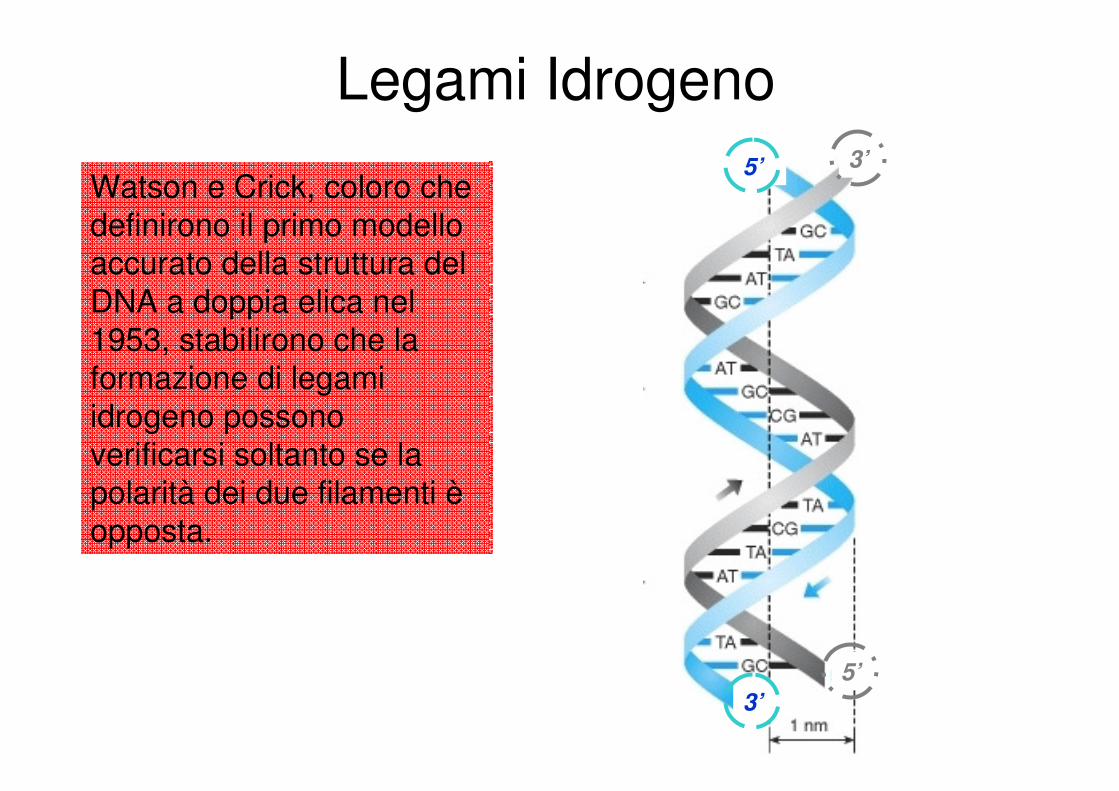
Legami Idrogeno
1 nm
5’
3’
5’
3’
Watson e Crick, coloro che
definirono il primo modello accurato della struttura del
DNA a doppia elica nel
1953, stabilirono che la formazione di legami
idrogeno possono verificarsi soltanto se la
polarità dei due filamenti èopposta.
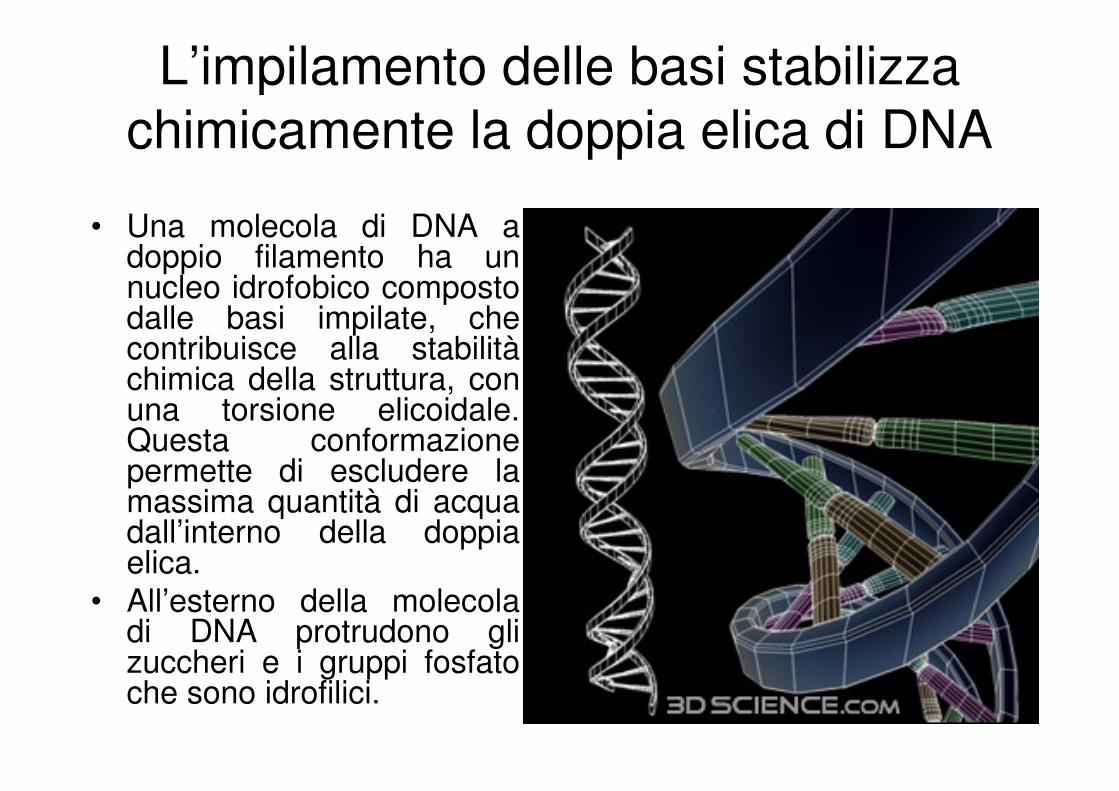
L’impilamento delle basi stabilizza
chimicamente la doppia elica di DNA
• Una molecola di DNA a doppio filamento ha un nucleo idrofobico composto dalle basi impilate, che contribuisce alla stabilitàchimica della struttura, con una torsione elicoidale. Questa conformazione permette di escludere la massima quantità di acqua dall’interno della doppia elica.
• All’esterno della molecola di DNA protrudono gli zuccheri e i gruppi fosfato che sono idrofilici.
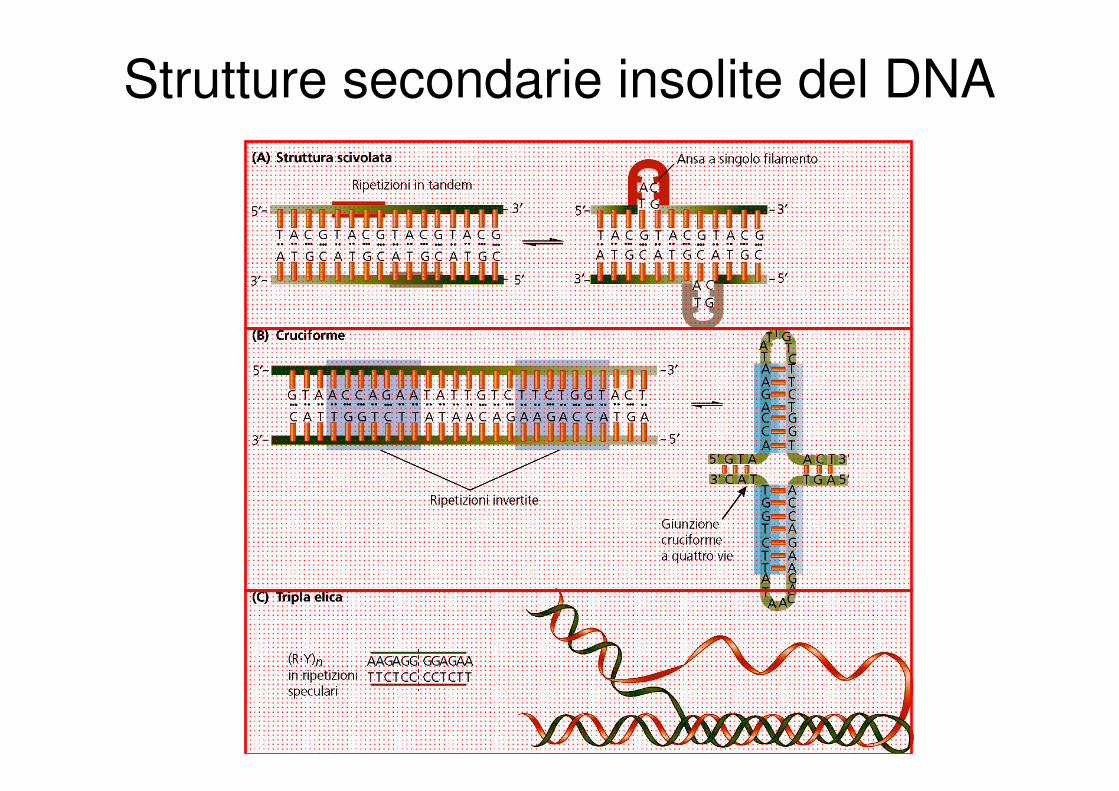
Strutture secondarie insolite del DNA
La struttura terziaria del DNALa struttura terziaria del DNA
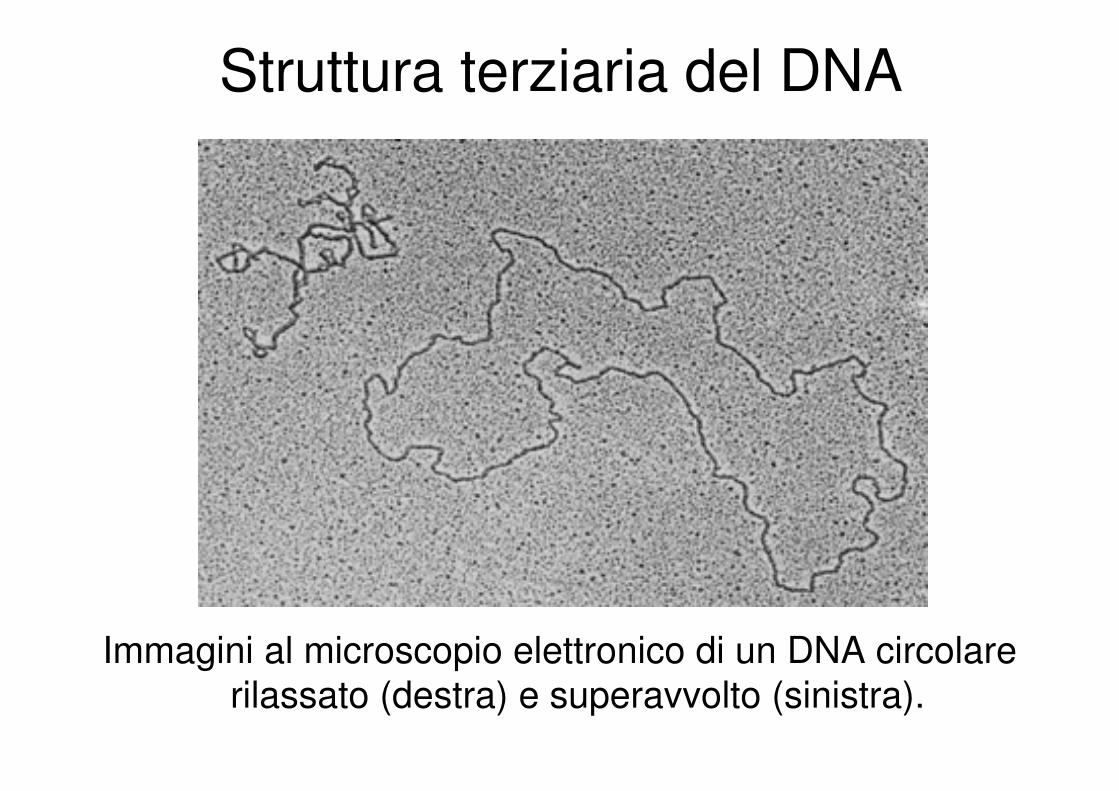
Struttura terziaria del DNA
Immagini al microscopio elettronico di un DNA circolare
rilassato (destra) e superavvolto (sinistra).
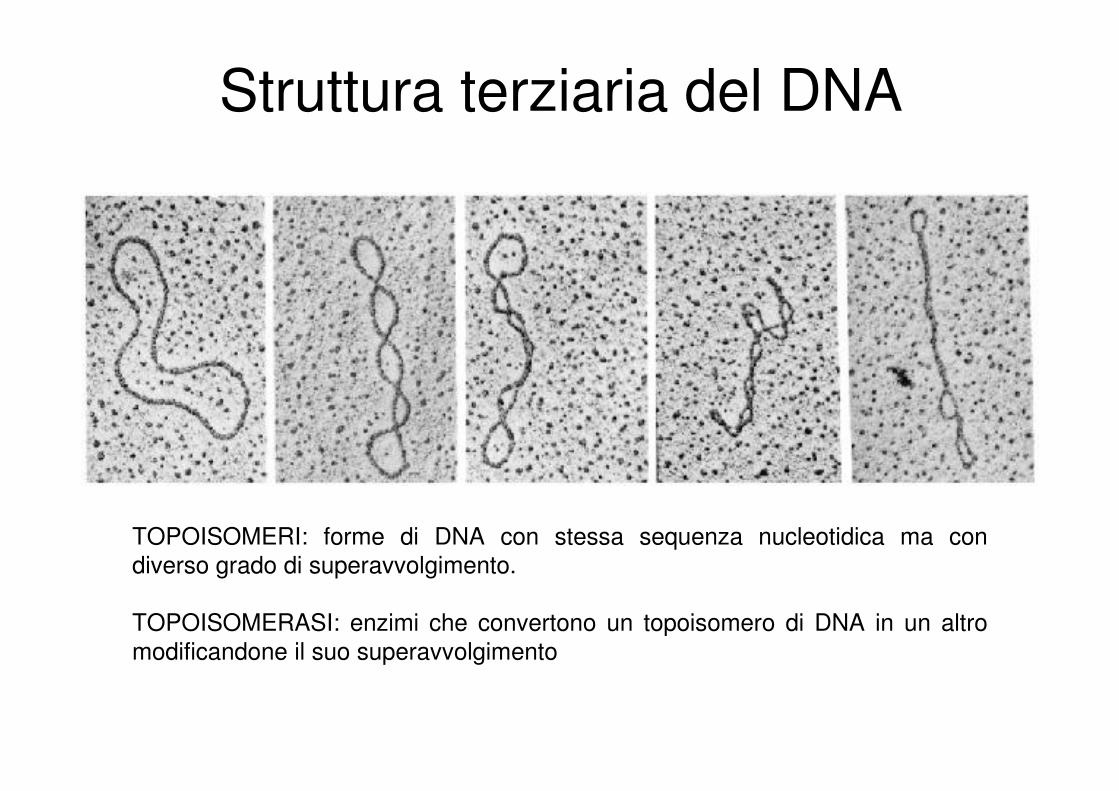
Struttura terziaria del DNA
TOPOISOMERI: forme di DNA con stessa sequenza nucleotidica ma con
diverso grado di superavvolgimento.
TOPOISOMERASI: enzimi che convertono un topoisomero di DNA in un altro
modificandone il suo superavvolgimento
LL’’organizzazione del DNA organizzazione del DNA
negli eucariotinegli eucarioti
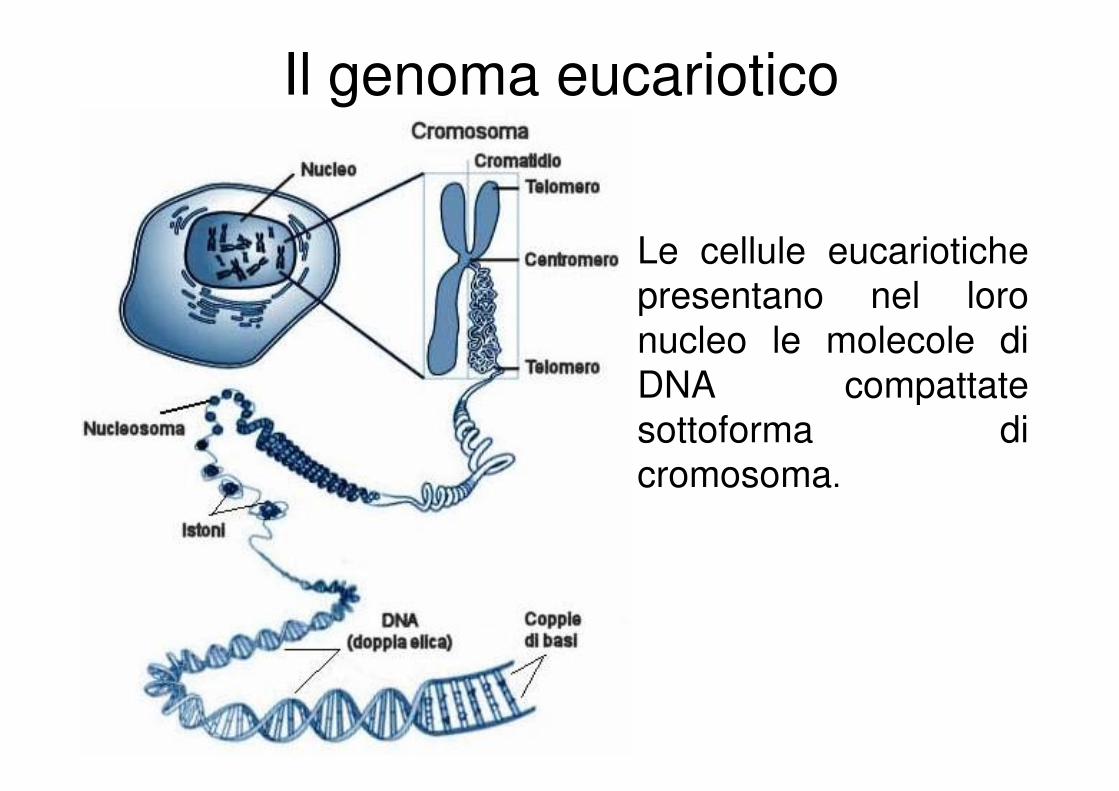
Il genoma eucariotico
Le cellule eucariotiche
presentano nel loro
nucleo le molecole di
DNA compattate
sottoforma di
cromosoma.
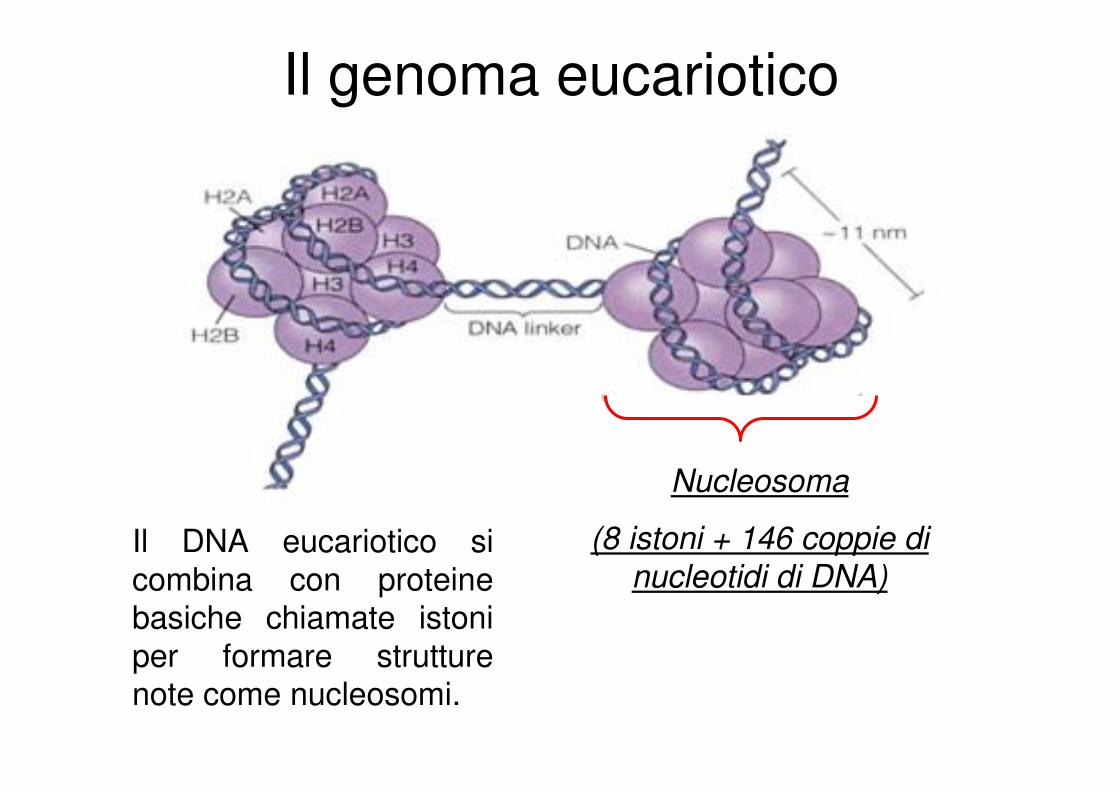
Il genoma eucariotico
Nucleosoma
(8 istoni + 146 coppie di
nucleotidi di DNA)Il DNA eucariotico si
combina con proteine basiche chiamate istoni
per formare strutture note come nucleosomi.
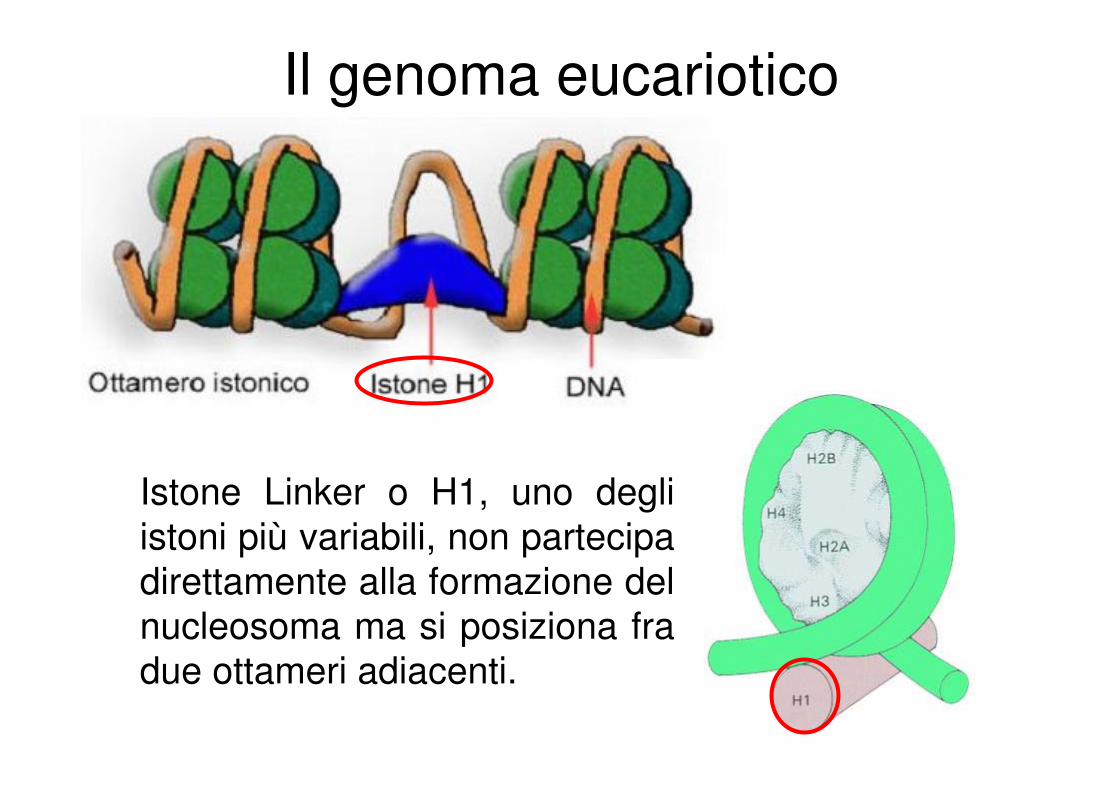
Il genoma eucariotico
Istone Linker o H1, uno degli
istoni più variabili, non partecipa
direttamente alla formazione del
nucleosoma ma si posiziona fra
due ottameri adiacenti.
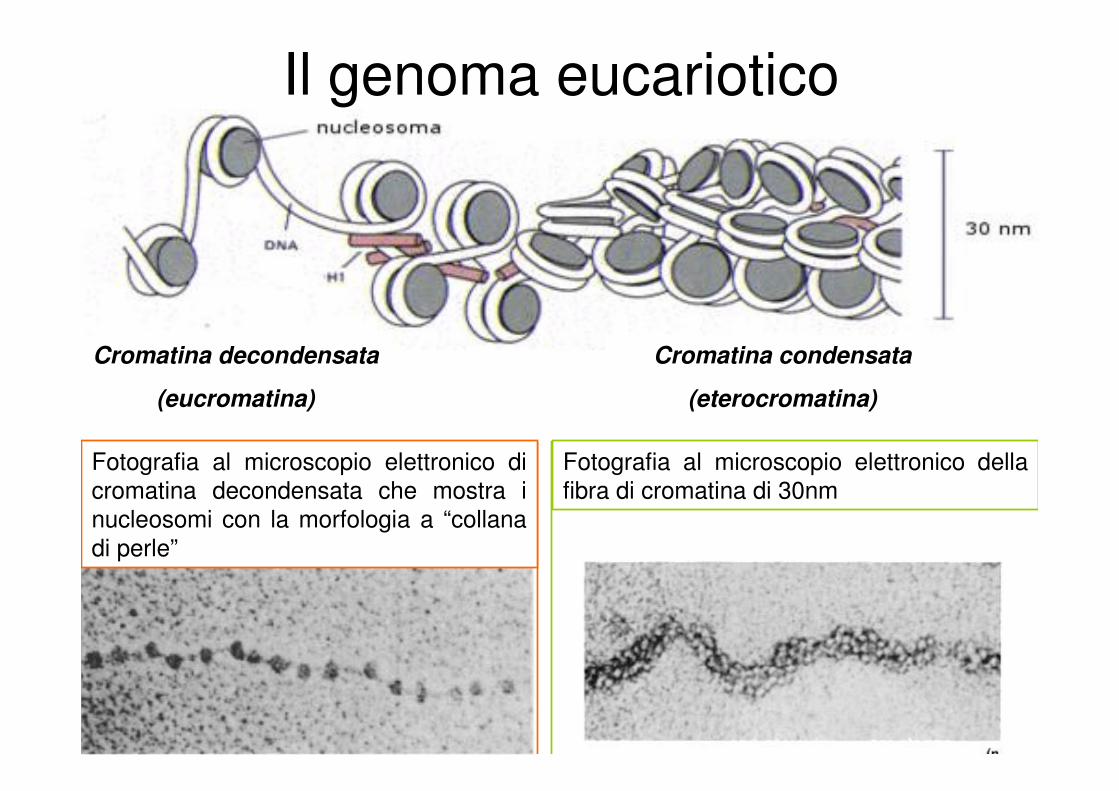
Il genoma eucariotico
Cromatina decondensata
(eucromatina)
Cromatina condensata
(eterocromatina)
Fotografia al microscopio elettronico di
cromatina decondensata che mostra i
nucleosomi con la morfologia a “collana
di perle”
Fotografia al microscopio elettronico della
fibra di cromatina di 30nm
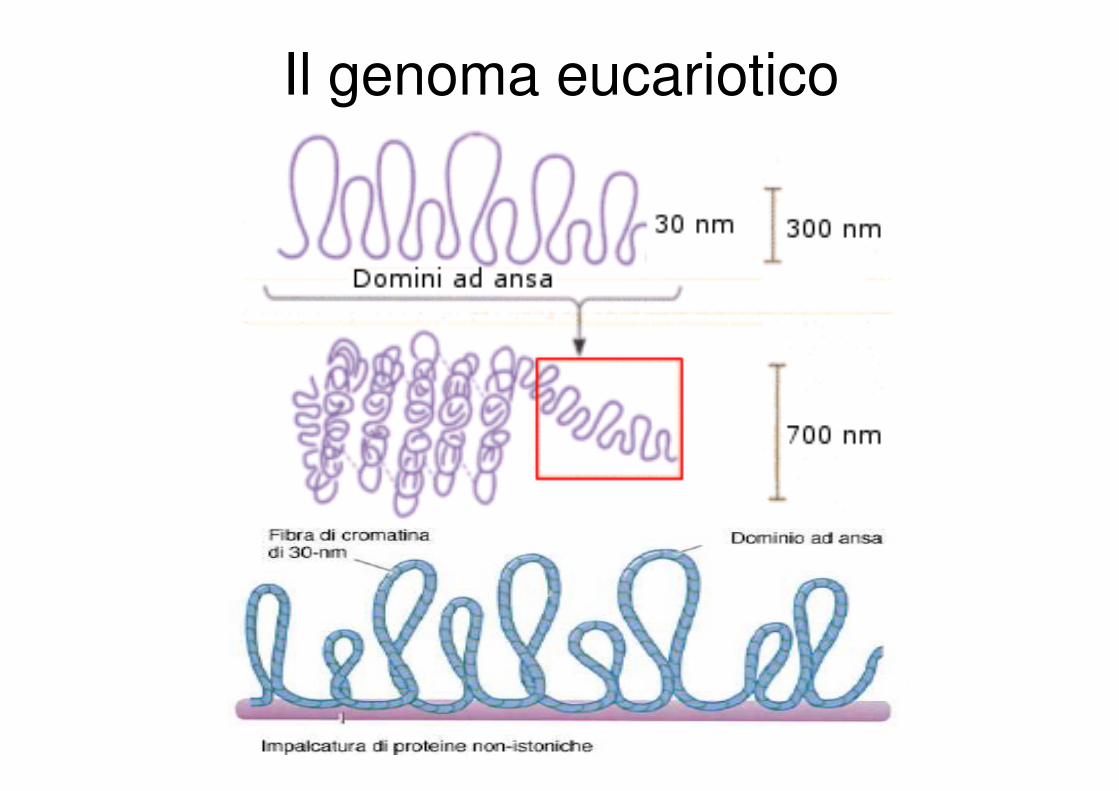
Il genoma eucariotico
Il codice genetico Il codice genetico
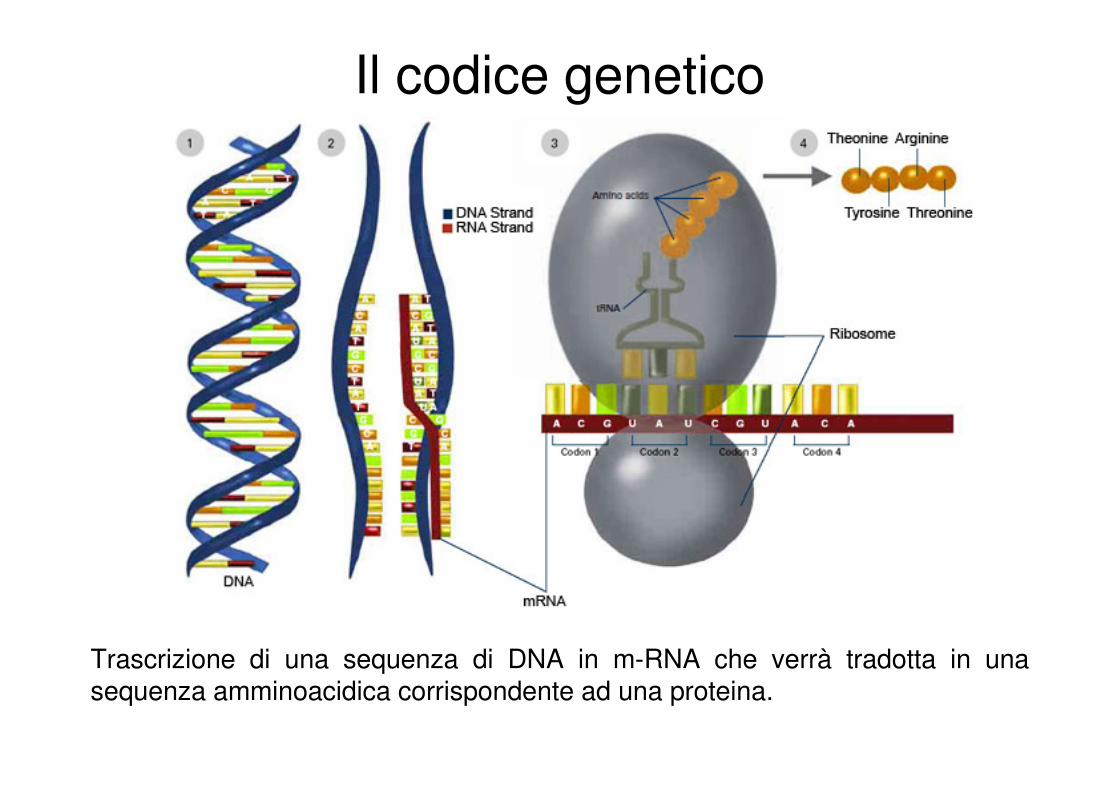
Il codice genetico
Trascrizione di una sequenza di DNA in m-RNA che verrà tradotta in una sequenza amminoacidica corrispondente ad una proteina.
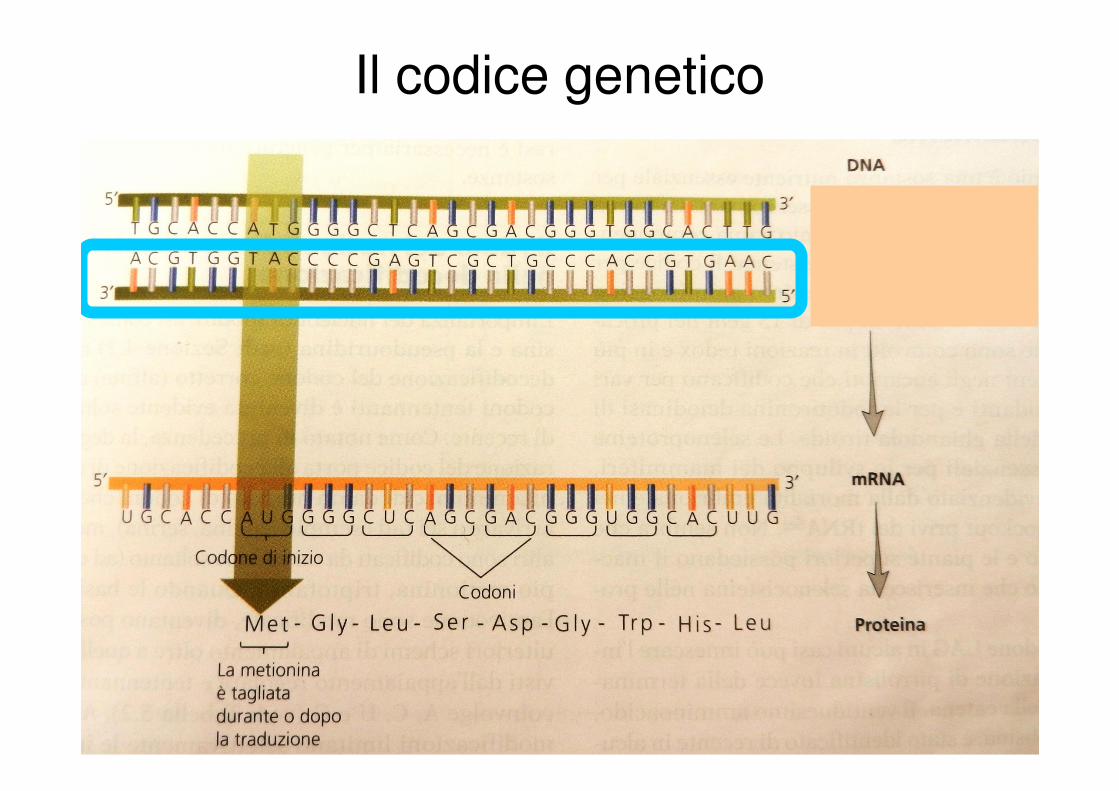
Il codice genetico
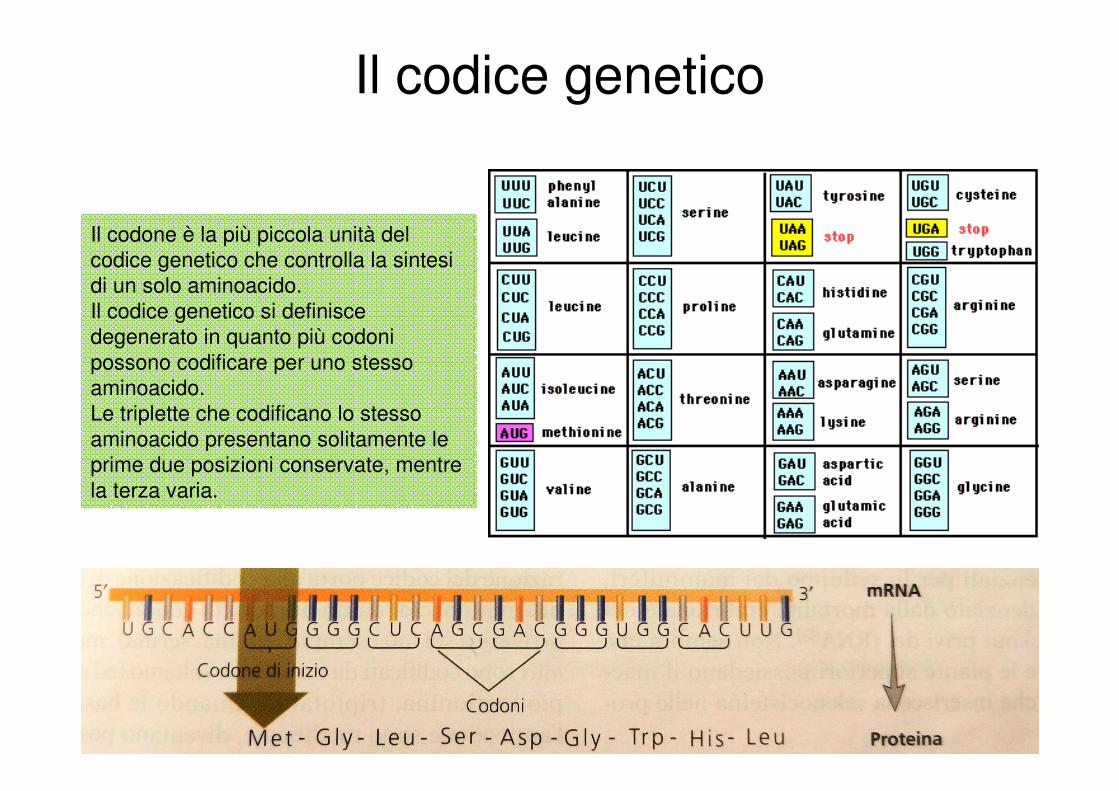
Il codice genetico
Il codone è la più piccola unità del
codice genetico che controlla la sintesi
di un solo aminoacido.
Il codice genetico si definisce
degenerato in quanto più codoni
possono codificare per uno stesso
aminoacido.
Le triplette che codificano lo stesso
aminoacido presentano solitamente le
prime due posizioni conservate, mentre
la terza varia.
La replicazione del DNA La replicazione del DNA
negli eucariotinegli eucarioti
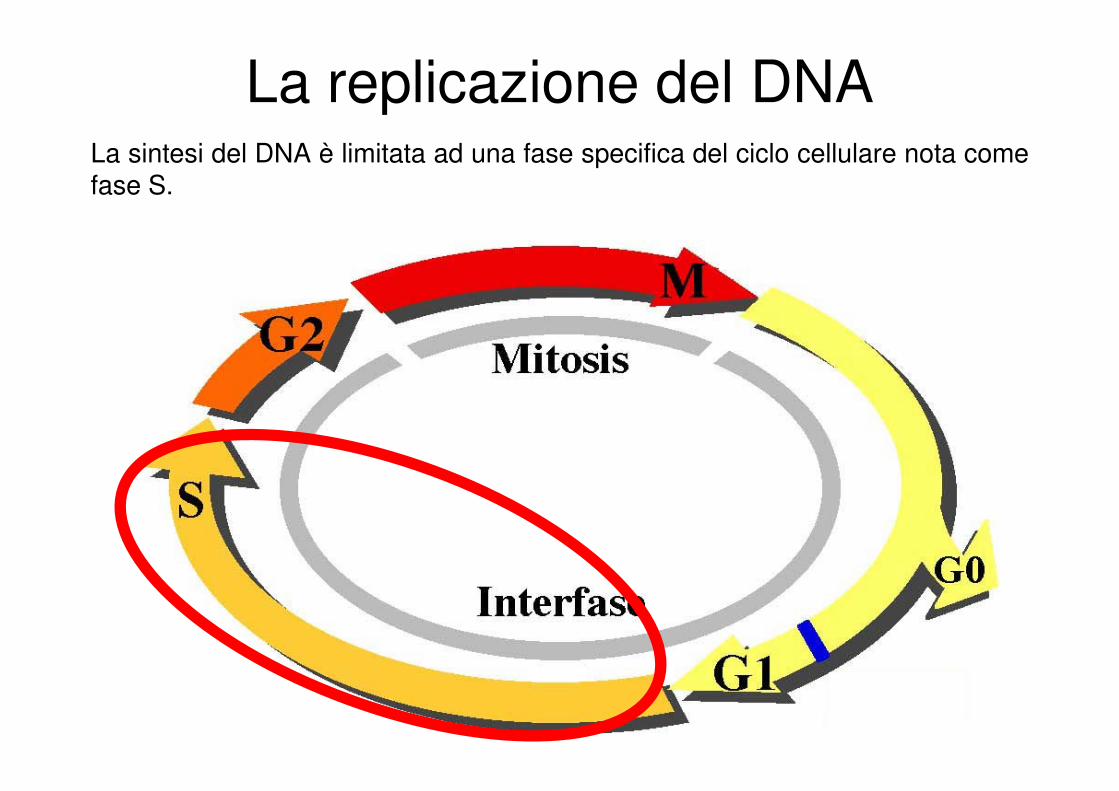
La sintesi del DNA è limitata ad una fase specifica del ciclo cellulare nota come fase S.
La replicazione del DNA
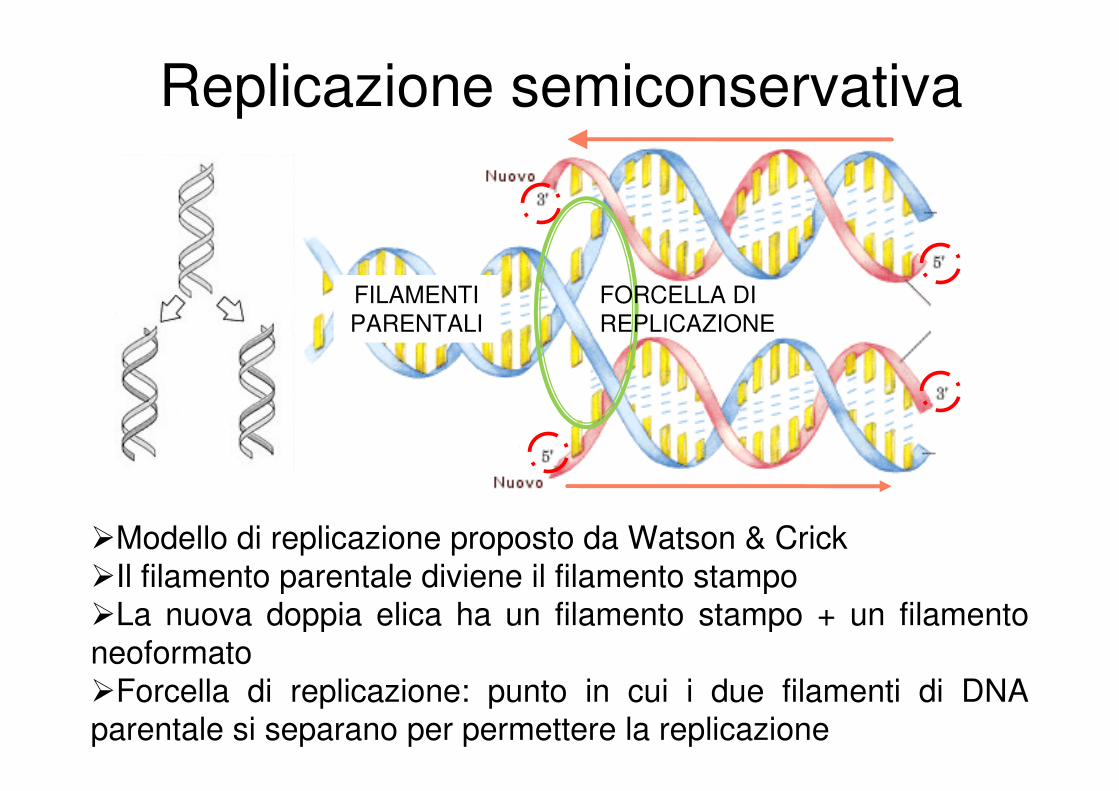
Replicazione semiconservativa
FILAMENTI
PARENTALI
�Modello di replicazione proposto da Watson & Crick
�Il filamento parentale diviene il filamento stampo�La nuova doppia elica ha un filamento stampo + un filamento
neoformato
�Forcella di replicazione: punto in cui i due filamenti di DNA parentale si separano per permettere la replicazione
FORCELLA DI
REPLICAZIONE
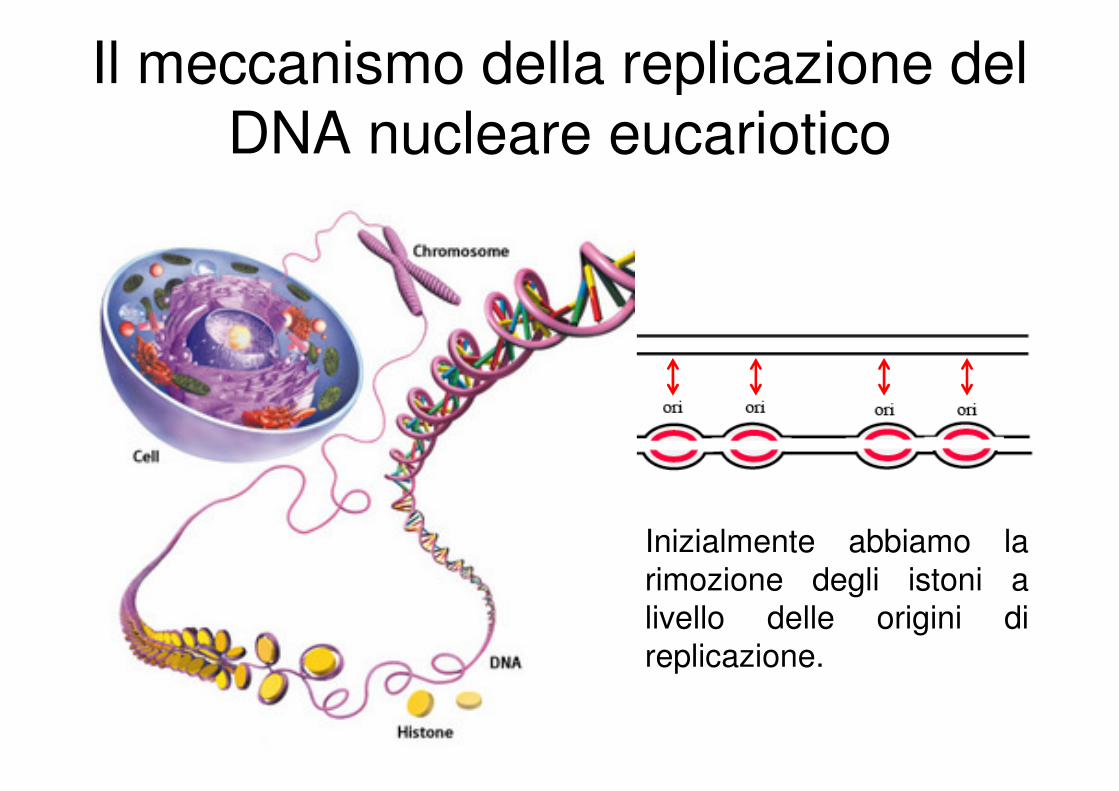
Il meccanismo della replicazione del
DNA nucleare eucariotico
Inizialmente abbiamo la
rimozione degli istoni a livello delle origini di
replicazione.
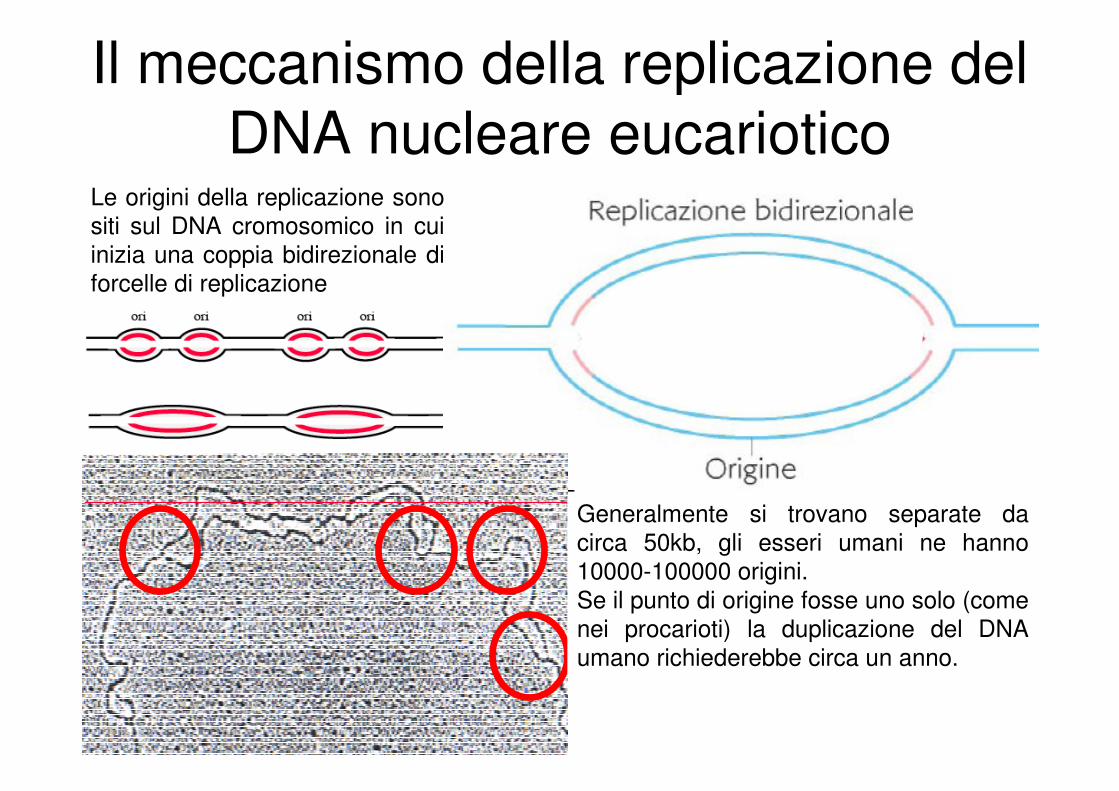
Il meccanismo della replicazione del
DNA nucleare eucarioticoLe origini della replicazione sono
siti sul DNA cromosomico in cui
inizia una coppia bidirezionale di
forcelle di replicazione
Generalmente si trovano separate da
circa 50kb, gli esseri umani ne hanno
10000-100000 origini.
Se il punto di origine fosse uno solo (come
nei procarioti) la duplicazione del DNA
umano richiederebbe circa un anno.
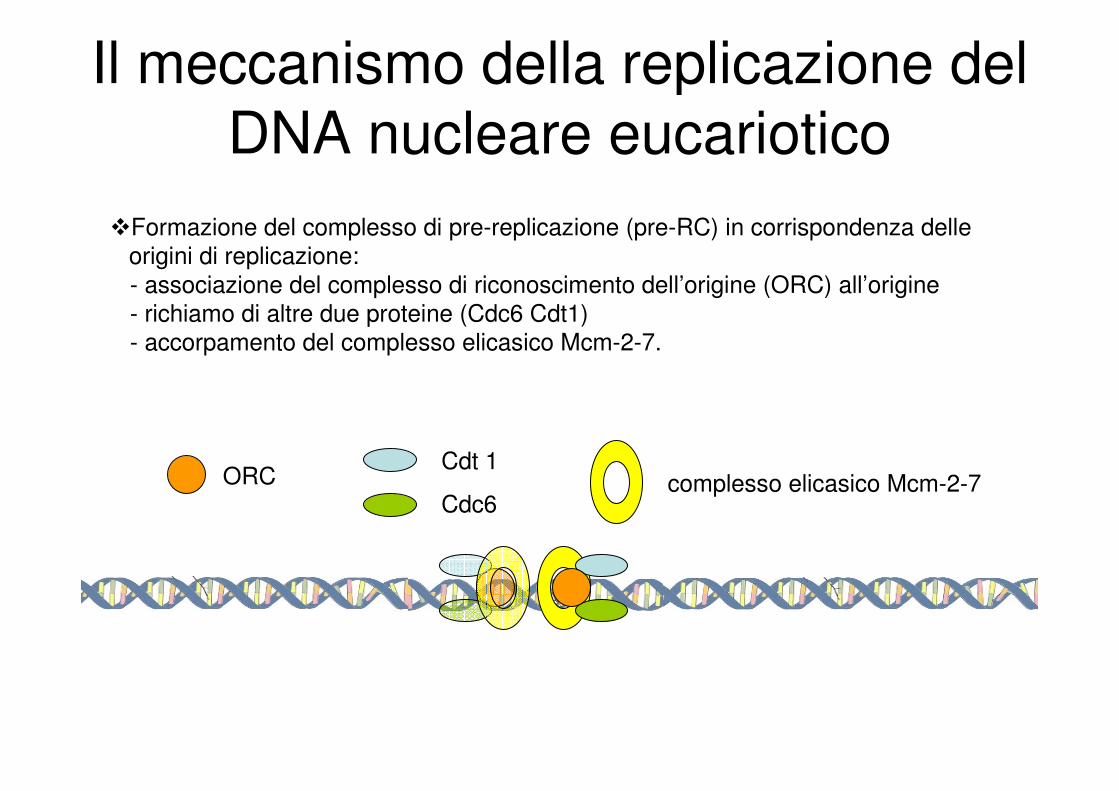
Il meccanismo della replicazione del
DNA nucleare eucariotico
�Formazione del complesso di pre-replicazione (pre-RC) in corrispondenza delle
origini di replicazione:
- associazione del complesso di riconoscimento dell’origine (ORC) all’origine
- richiamo di altre due proteine (Cdc6 Cdt1)
- accorpamento del complesso elicasico Mcm-2-7.
ORCCdt 1
Cdc6complesso elicasico Mcm-2-7
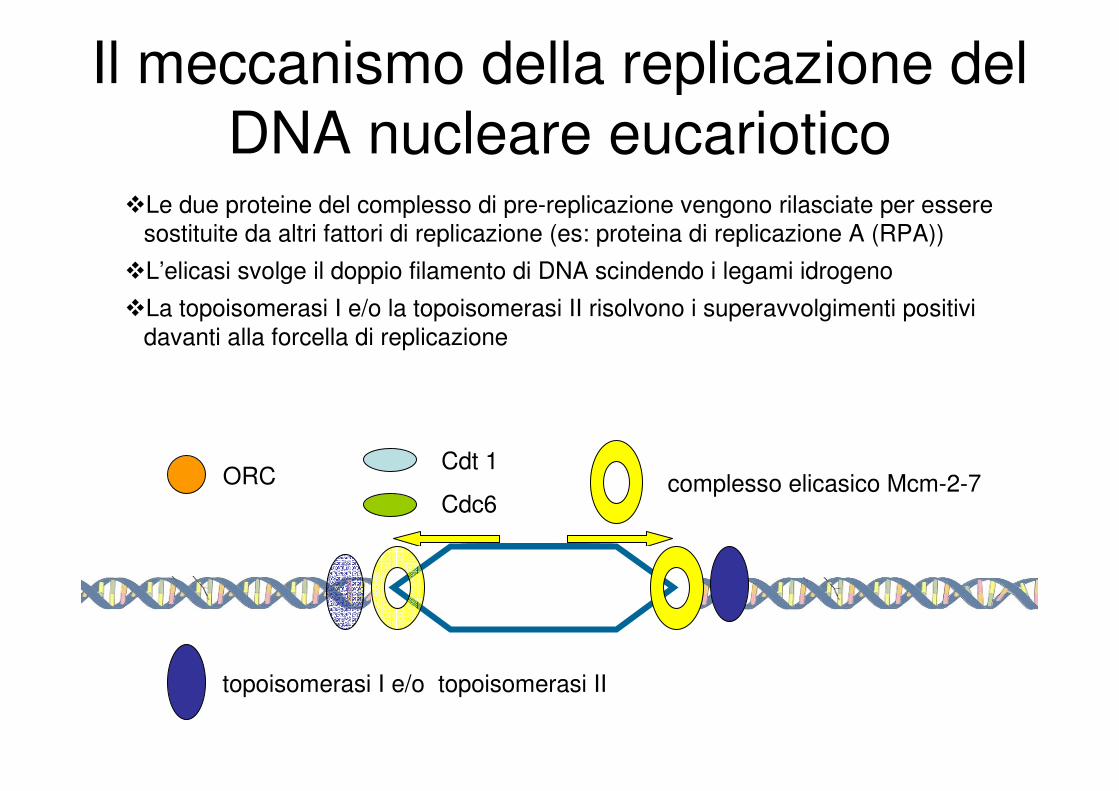
Il meccanismo della replicazione del
DNA nucleare eucariotico
ORCCdt 1
Cdc6complesso elicasico Mcm-2-7
�Le due proteine del complesso di pre-replicazione vengono rilasciate per essere
sostituite da altri fattori di replicazione (es: proteina di replicazione A (RPA))
�L’elicasi svolge il doppio filamento di DNA scindendo i legami idrogeno
�La topoisomerasi I e/o la topoisomerasi II risolvono i superavvolgimenti positivi
davanti alla forcella di replicazione
topoisomerasi I e/o topoisomerasi II
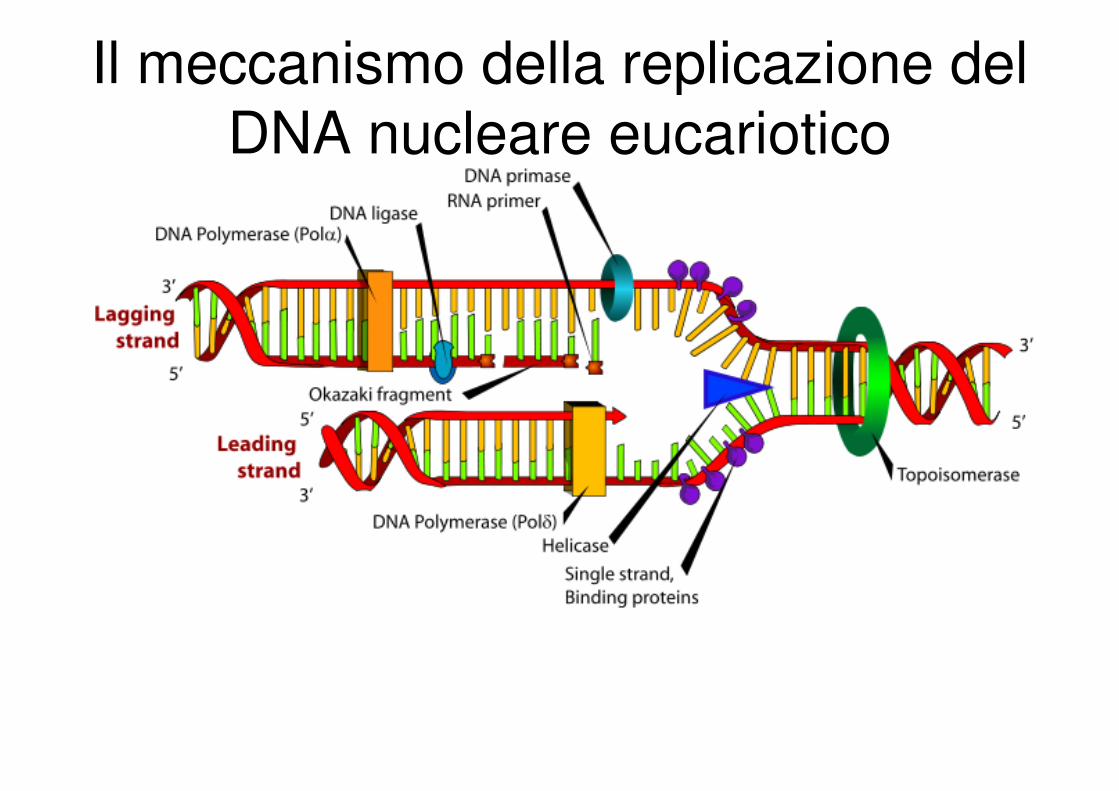
Il meccanismo della replicazione del
DNA nucleare eucariotico
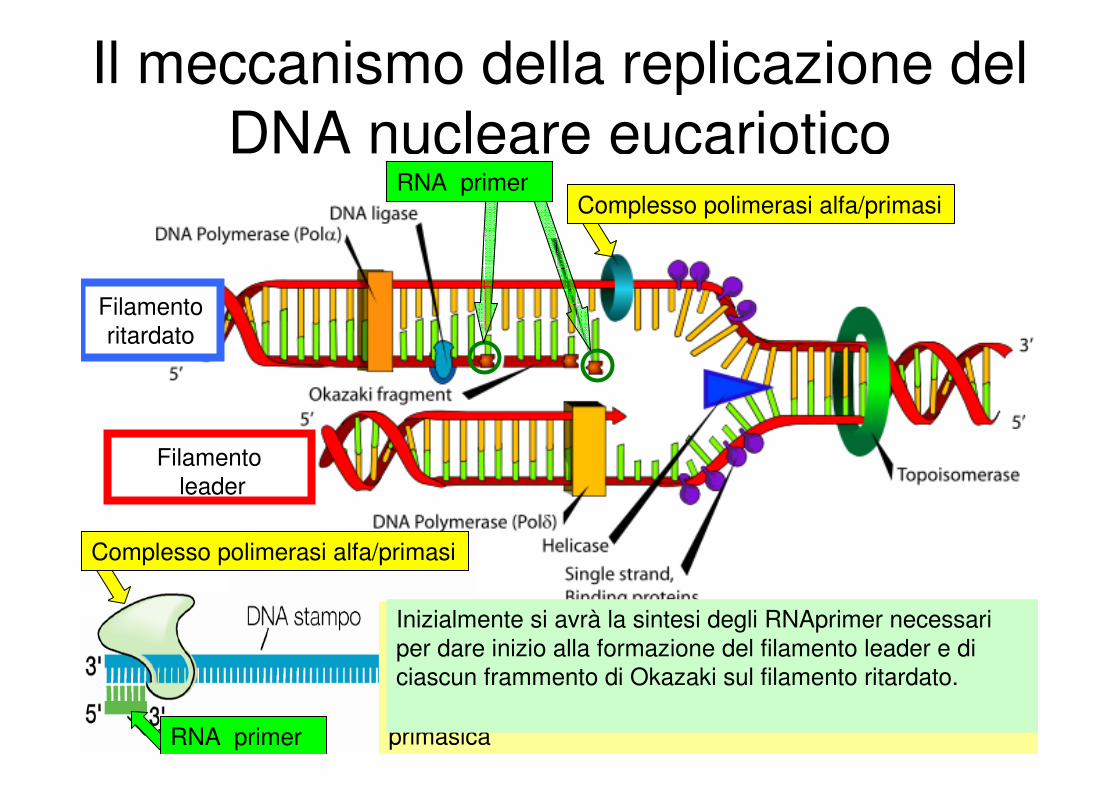
Il meccanismo della replicazione del
DNA nucleare eucariotico
Complesso polimerasi alfa/primasi
Complesso polimerasi alfa/primasi
RNA primer
RNA primer
Complesso polimerasi alfa/primasi :
�subunità con attività DNA polimerasica
�subunità necessaria per l’assemblaggio del primer
�due piccole proteine che assieme forniscono l’attività
primasica
Inizialmente si avrà la sintesi degli RNAprimer necessari
per dare inizio alla formazione del filamento leader e di
ciascun frammento di Okazaki sul filamento ritardato.
Filamento
leader
Filamento
ritardato
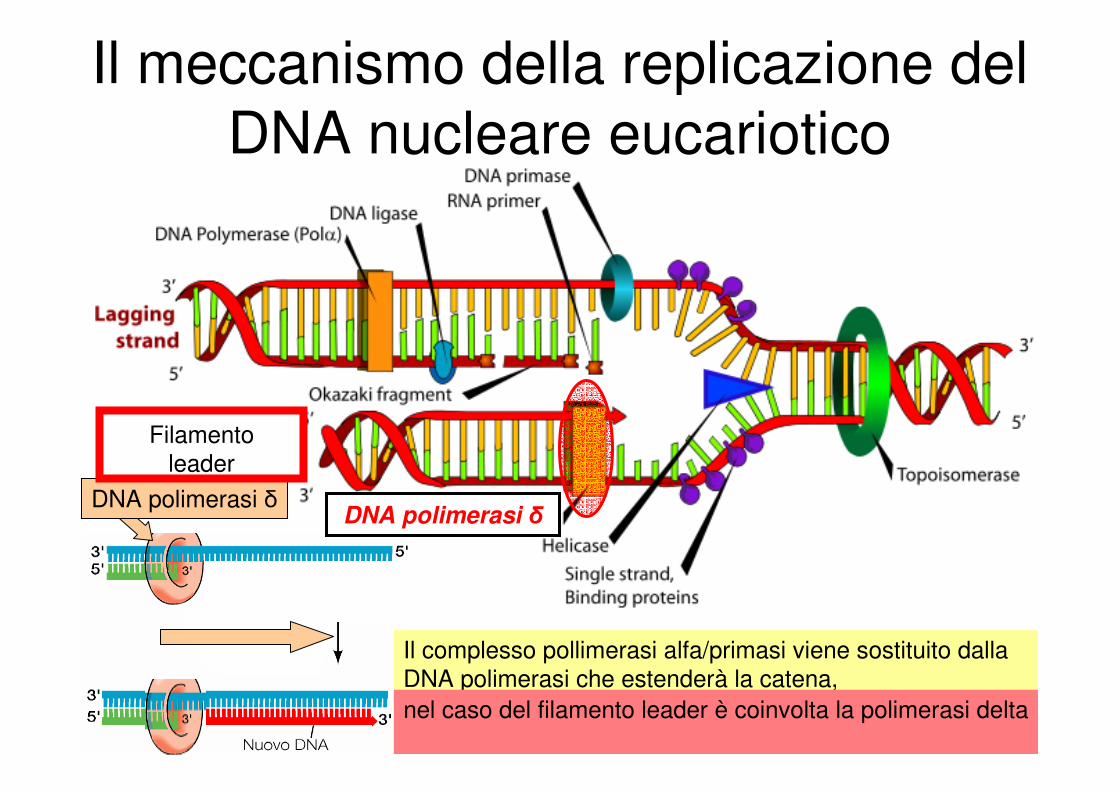
Il meccanismo della replicazione del
DNA nucleare eucariotico
DNA polimerasi δ
Complesso pol alfa/primasi
DNA polimerasi δ
Filamento
leader
Il complesso pollimerasi alfa/primasi viene sostituito dalla
DNA polimerasi che estenderà la catena,
nel caso del filamento leader è coinvolta la polimerasi delta
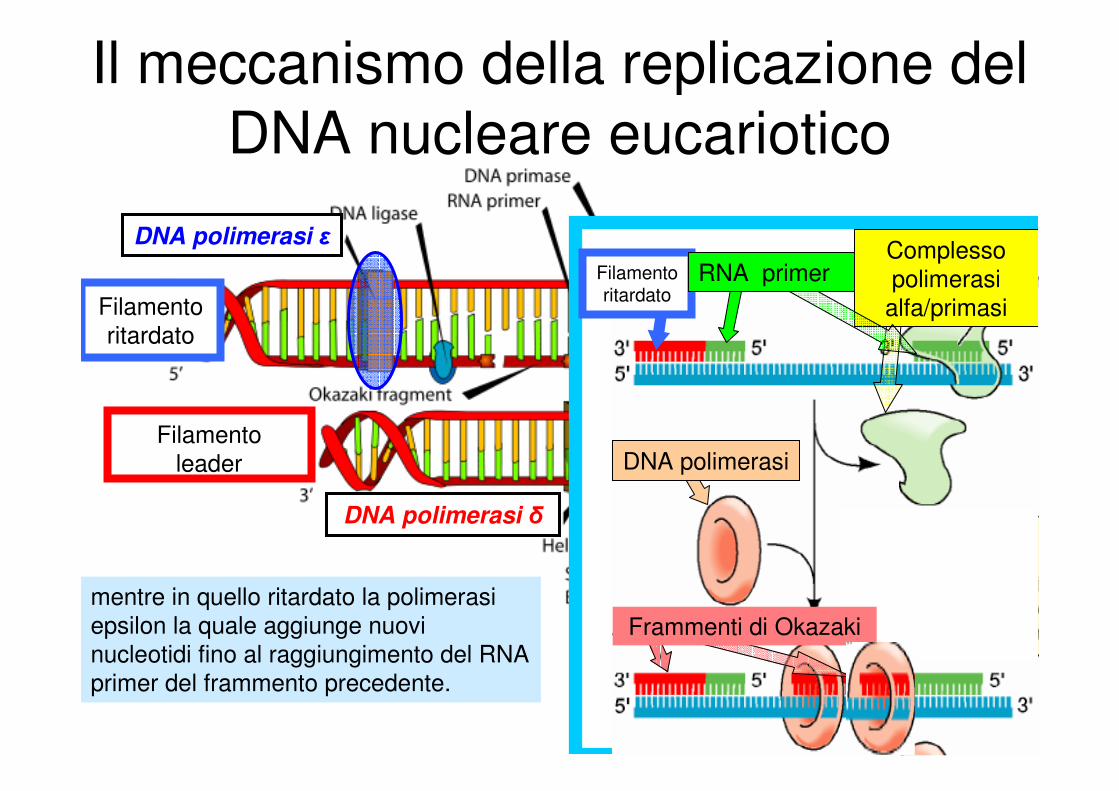
Il meccanismo della replicazione del
DNA nucleare eucariotico
DNA polimerasi ε
Filamento
leader
Filamento
ritardato
mentre in quello ritardato la polimerasi
epsilon la quale aggiunge nuovi
nucleotidi fino al raggiungimento del RNA
primer del frammento precedente.
DNA polimerasi δ
DNA polimerasi
Filamento ritardato
Complesso
polimerasi
alfa/primasi
Frammenti di Okazaki
RNA primer
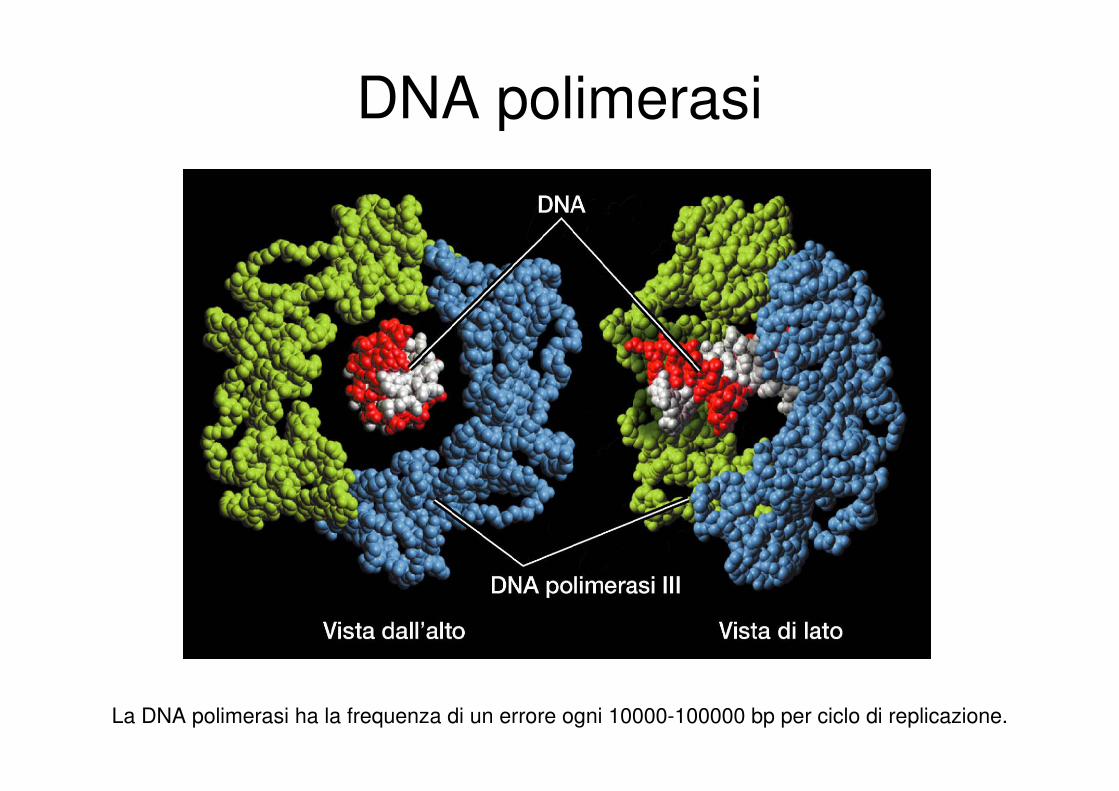
DNA polimerasi
La DNA polimerasi ha la frequenza di un errore ogni 10000-100000 bp per ciclo di replicazione.
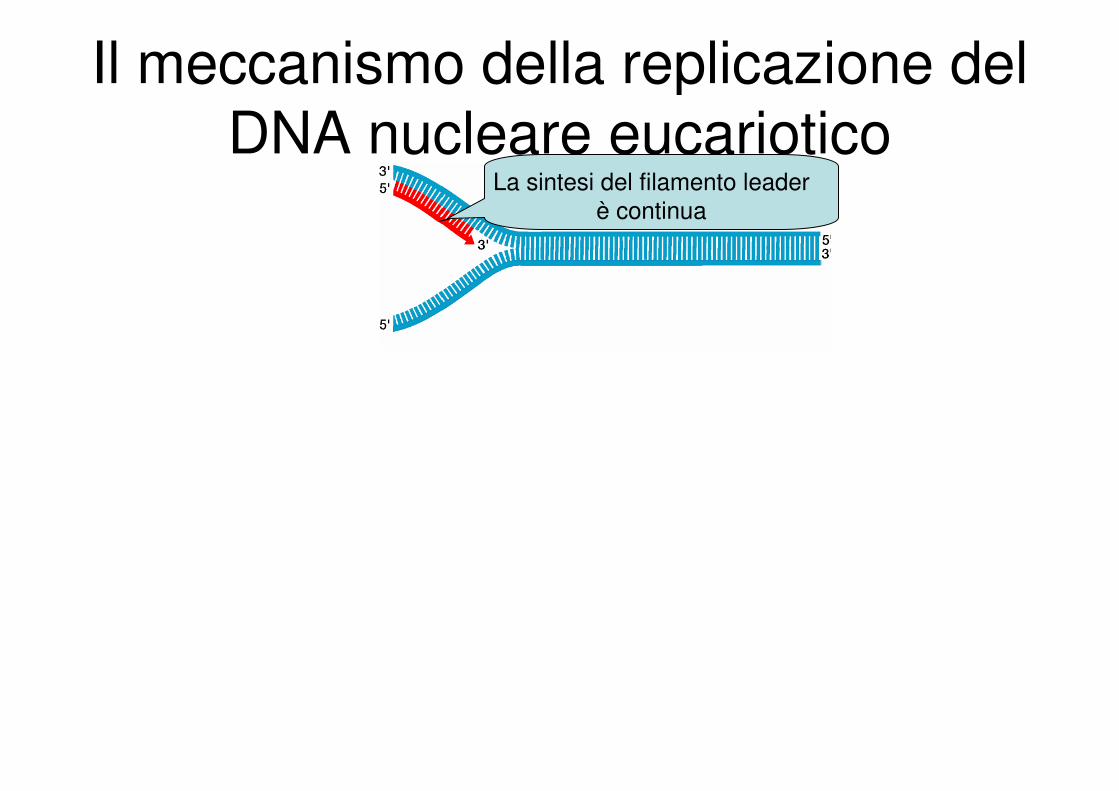
Il meccanismo della replicazione del
DNA nucleare eucarioticoLa sintesi del filamento leader
è continua
Il filamento ritardato viene
sintetizzato sottoforma di
frammenti di Okazaki
Frammenti di Okazaki
Nel frattempo la forcella di
replicazione cresce
… e cresce

Matu
razio
ne d
el D
NA
di n
uova s
inte
si:
Ligasi I
unione dei frammenti di
Okazaki sul filamento
ritardato.
riempimento delle
interruzioni
DNA Polimerasi ε
rimozione dell’RNA
primer
Il meccanismo della replicazione del
DNA nucleare eucariotico
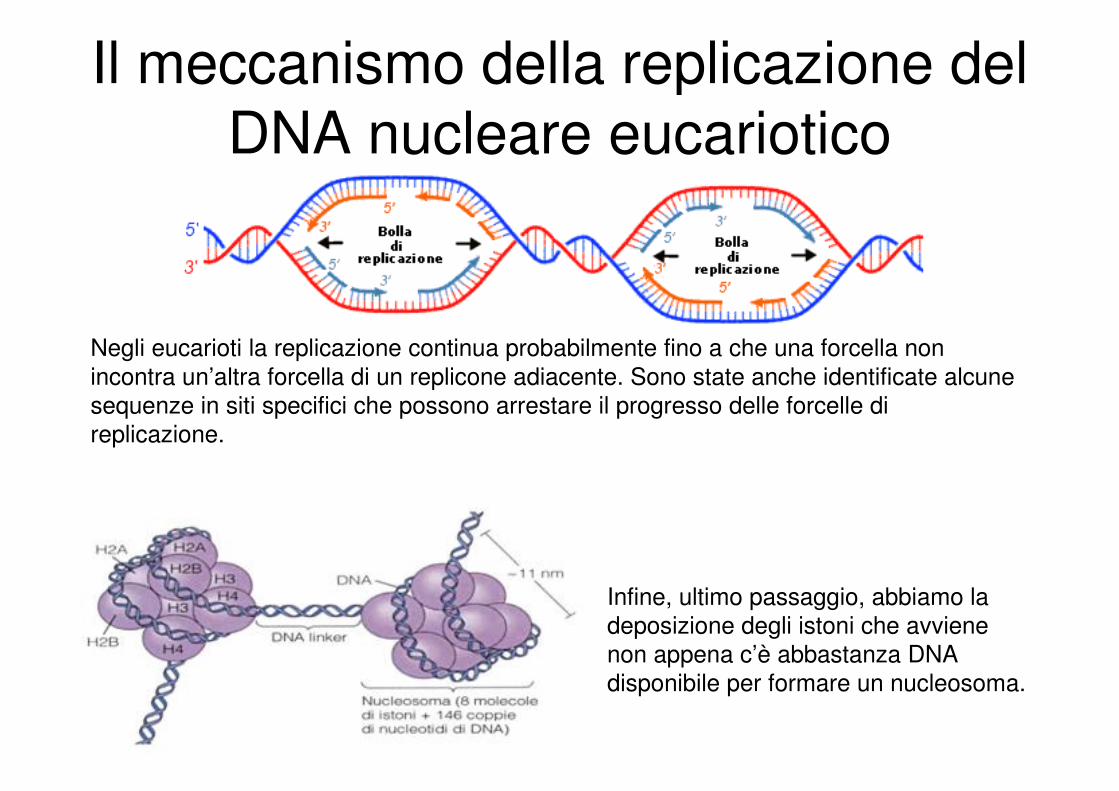
Il meccanismo della replicazione del
DNA nucleare eucariotico
Negli eucarioti la replicazione continua probabilmente fino a che una forcella non
incontra un’altra forcella di un replicone adiacente. Sono state anche identificate alcune
sequenze in siti specifici che possono arrestare il progresso delle forcelle di
replicazione.
Infine, ultimo passaggio, abbiamo la
deposizione degli istoni che avviene
non appena c’è abbastanza DNA
disponibile per formare un nucleosoma.
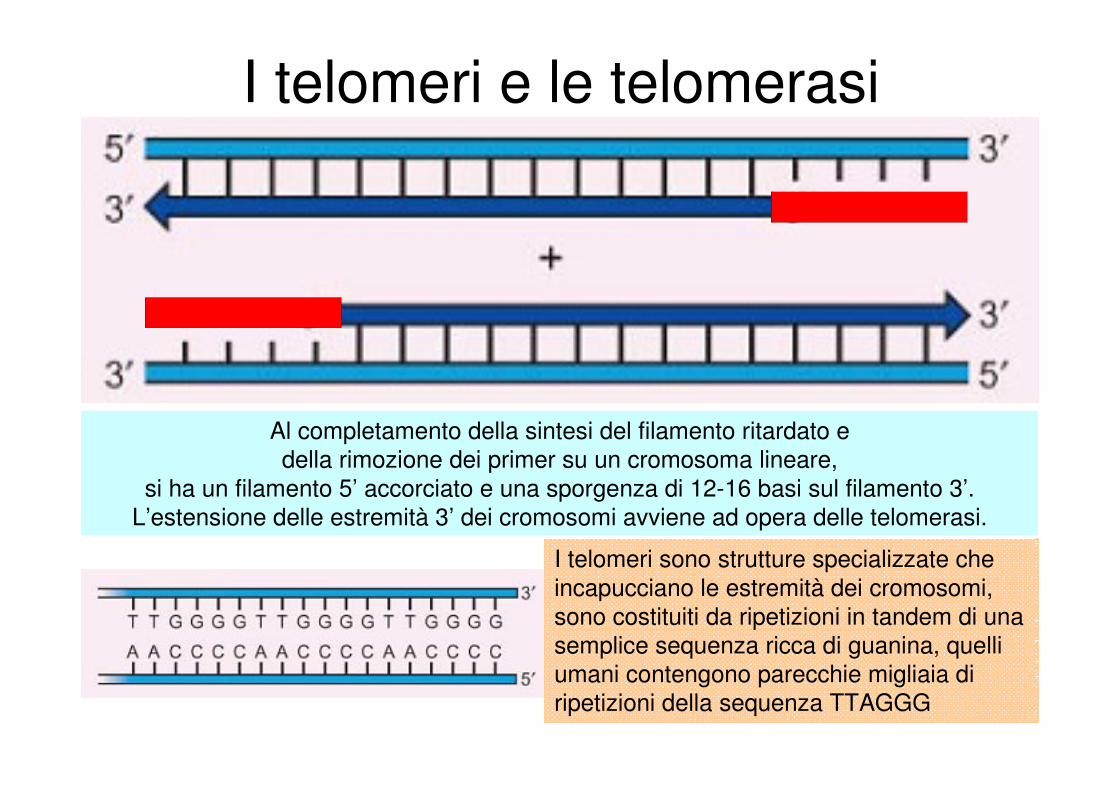
I telomeri e le telomerasi
Al completamento della sintesi del filamento ritardato e
della rimozione dei primer su un cromosoma lineare,
si ha un filamento 5’ accorciato e una sporgenza di 12-16 basi sul filamento 3’.
L’estensione delle estremità 3’ dei cromosomi avviene ad opera delle telomerasi.
I telomeri sono strutture specializzate che
incapucciano le estremità dei cromosomi,
sono costituiti da ripetizioni in tandem di una
semplice sequenza ricca di guanina, quelli
umani contengono parecchie migliaia di
ripetizioni della sequenza TTAGGG
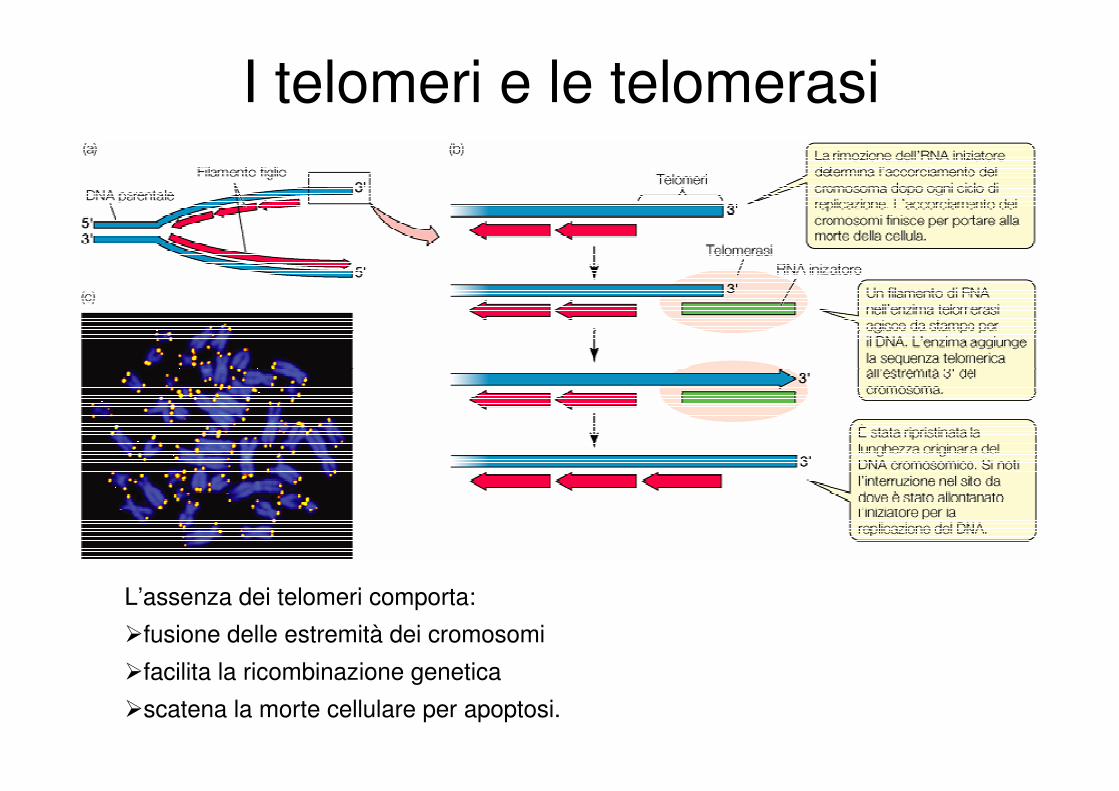
I telomeri e le telomerasi
L’assenza dei telomeri comporta:
�fusione delle estremità dei cromosomi
�facilita la ricombinazione genetica
�scatena la morte cellulare per apoptosi.

Grazie per l’attenzione






![5 Clonaggio [modalità compatibilità] - Giancarlo Accattatis · Successivamente si avrà la replicazione di questo DNA modificato (RICOMBINANTE) migliaia o milioni di volte fornendo](https://img.pdfslide.tips/doc/110x75/5c68496a09d3f226188d1aee/5-clonaggio-modalita-compatibilita-giancarlo-accattatis-successivamente.jpg)






![index [] · replicazione virale nell‟influenza A e B 4-amino-4-deoxy-Neu5Ac2en ... fasi di trascrizione e traslazione Early proteins: sintetizzano nuovo DNA virale usando il DNA](https://img.pdfslide.tips/doc/110x75/5c6589f309d3f2876e8cca7c/index-replicazione-virale-nellinfluenza-a-e-b-4-amino-4-deoxy-neu5ac2en.jpg)