Embed Size (px)
Citation preview

Atherosclerosis 150 (2000) 343–355
In vivo and in vitro evidence for the glycoxidation of low densitylipoprotein in human atherosclerotic plaques
Yoshinobu Imanaga a, Noriyuki Sakata a,*, Shigeo Takebayashi a, Akira Matsunaga b,Jun Sasaki b, Kikuo Arakawa b, Ryoji Nagai c, Seikoh Horiuchi c, Hiroyuki Itabe d,
Tatsuya Takano d
a Second Department of Pathology, School of Medicine, Fukuoka Uni6ersity, 45-1, 7-chome Nanakuma, Jonan-ku, Fukuoka 814-0180, Japanb Second Department of Medicine, School of Medicine, Fukuoka Uni6ersity, 45-1, 7-chome Nanakuma, Fukuoka 814-0180, Japan
c Department of Biochemistry, Kumamoto Uni6ersity School of Medicine, 2-2-1, Honjo, Kumamoto 860-0811, Japand Department of Microbiology and Molecular Pathology, Faculty of Pharmaceutical Sciences, Teikyo Uni6ersity, 1091-1 Suarashi,
Sagamiko-machi, Kanagawa 199-0195, Japan
Received 25 January 1999; received in revised form 6 September 1999; accepted 15 September 1999
Abstract
Although there have been suggestions that the glycation and oxidation of low density lipoprotein (LDL) might increase itsatherogenic potential, little is known about the presence of glycoxidative LDL in human atherosclerotic lesions. We developedspecific antibodies against different immunological epitopes of AGE structures, including N o-(carboxymethyl)lysine-protein adduct(CML), a glycoxidation product, and structure(s) other than CML (nonCML), and a monoclonal antibody against oxidizedphosphatidylcholine (oxPC), as an epitope of oxidized LDL. Immunohistochemical analysis demonstrated that the CML- andoxPC-epitopes were accumulated mainly in macrophage-derived foam cells in atherosclerotic lesions, including fatty streaks andatherosclerotic plaques. On the other hand, the nonCML-epitope and apolipoprotein B were localized mainly in extracellularmatrices of atherosclerotic lesions. The CML- and oxPC-epitopes were characterized by a model antigen-generating system usingthe copper ion-induced peroxidation and/or glucose-induced glycation of LDL. The glycoxidation of LDL caused the formationof CML-epitope with increasing concentrations of copper ion and glucose. It was also formed to some extent in LDL incubatedwith high concentrations (500 mM) of glucose. However, no CML-epitope was observed in oxidized LDL induced by copper ionalone. On the other hand, the formation of oxPC-epitope in LDL was dependent on copper ion-induced peroxidation, butindependent of glucose-induced glycation. The addition of chelators, ethylenediaminetetraacetic acid and diethylenetriaminepen-taacetic acid, reduced the increase in electrophoretic mobility and TBARS caused by the peroxidation and glycoxidation of LDL,but had no effects on the formation of fructosamine caused by the glycation and glycoxidation of LDL. Chelators as well asaminoguanidine protected the formation of CML-epitope in glycated or glycoxidative LDL. Although the formation ofoxPC-epitope was completely inhibited by the addition of chelators, it was partially protected by aminoguanidine. These in vitroresults suggest that the glycoxidative modification of LDL may occur in the arterial intima, and may contribute to thedevelopment of human atherosclerotic lesions. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Atherogenesis; Glycoxidation of LDL; Oxidized phosphatidylcholine; No-(carboxymethyl)lysine (CML)-protein adduct; ELISA;Immunohistochemistry
www.elsevier.com/locate/atherosclerosis
1. Introduction
Although oxidized low density lipoprotein (LDL)reportedly plays an important role in atherogenesis, itcannot sufficiently account for the increased susceptibil-
ity of patients with diabetes mellitus to atherosclerosis.Advanced glycation end products (AGEs) are formedfrom a series of non-enzymatic chemical reactions be-tween reducing sugars, proteins, lipids and nucleic acid[1]. The final step of these reactions gives rise to AGEswhich consist of heterogeneous chemical compounds.We [2] previously reported that AGEs accumulate inhuman atherosclerotic lesions and their location de-
* Corresponding author. Tel.: +81-92-8011011, ext 3285; fax:+81-92-8638383.
0021-9150/00/$ - see front matter © 2000 Elsevier Science Ireland Ltd. All rights reserved.PII: S 0 0 2 1 -9150 (99 )00396 -2

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355344
pends on the different epitopes of AGEs. Palinski et al.[3] demonstrated the colocalization of oxidation-specificadducts and AGE-specific epitopes within intimal le-sions of euglycemic WHHL rabbits. Thus, it is possiblethat in vivo lipid peroxidation and glycation reactionsmay be closely linked and enhance atheroscleroticlesions.
N o-(carboxymethyl)lysine-protein adduct (CML) andpentosidine, major products of glycoxidation, havebeen suggested to be general markers of the oxidativemodification of glycated proteins in atherosclerosis anddiabetes [4–6]. A close link between glycoxidation andlipid peroxidation has been shown to exist and to playa role in tissue damage in atherogenesis, diabetes andaging [6,7]. Glycated LDL has been shown to be moresusceptible to oxidative modification of its lipids andprotein than native LDL, suggesting that glycoxidationcontributes to the formation of oxidized LDL [8,9].Superoxides are likely to play an important role in themechanism by which glycoxidation enhances the oxi-dized modification of LDL, because Amadori com-pounds and intermediates of the browning reaction areknown to be a source of superoxides [10]. CML isformed by the oxidative cleavage of Amadori com-pounds [11]. The formation of CML has been recentlyshown to be mediated by hydroxy radical which isgenerated by a Fenton reaction between Fe2+ andAmadori compounds [12]. An immunohistochemicalanalysis with monoclonal anti-CML antibody (6D12)showed that CML accumulated in macrophage-derivedfoam cells and extracellular matrix in atheroscleroticlesions [13,14]. On the other hand, oxidized phos-phatidylcholine (oxPC), which is recognized by themonoclonal antibody FOH1a/DLH3, has been shownto be formed by the copper ion-induced oxidation ofLDL and to exist in human atherosclerotic plaques andplasma [15,16]. Thus, it is possible that glycoxidativemodification may be involved in the formation of bothCML- and oxPC-epitopes in LDL and may play a rolein atherogenesis.
This study was undertaken to investigate the possibil-ity that the glycoxidative modification of LDL occursin human aorta, and contributes to the progression ofatherosclerosis. We examined the localization of CML-and oxPC-epitopes in atherosclerotic lesions of humanaorta and the mechanism of CML and oxPC formationin LDL in vitro.
2. Materials and methods
2.1. Chemicals
O-Phenylenediamine dihydrochloride and naphthol-AS-BI-phosphoric acid were purchased from Sigma (St.Louis, MO). Crystallized bovine serum albumin (BSA)
was purchased from the Seikagaku Kogyo Co. (Tokyo,Japan). Horseradish peroxidase (HRP)-conjugated rab-bit anti-mouse IgG antibody and goat anti-rabbit IgGantibody were purchased from CAPPEL (Durham,NC). Monoclonal anti-smooth muscle actin antibody,monoclonal anti-human macrophage (HAM56) anti-body, biotinylated rabbit anti-mouse IgG antibody,biotinylated swine anti-rabbit antibody and alkalinephosphatase-labeled streptavidin were obtained fromDAKO (Carpinteria, CA). Biotinylated goat anti-mouse IgM antibody was obtained from VECTOR(Burlingame, CA). Monoclonal anti-human apolipo-protein B (Apo-B) antibody was obtained fromMONOSAN (Netherlands). The monoclonal antibodyagainst oxidized LDL, FOH1a/DLH3, was prepared byItabe et al. and has been shown to recognize oxidizedphosphatidylcholine (oxPC) as a marker of oxidizedLDL [15]. The monoclonal anti-CML (6D12) and poly-clonal anti-nonCML antibodies were prepared as previ-ously described [2,17,18]. Non-immune, chromato-graphically purified rabbit and mouse IgG were ob-tained from ZYMED (San Francisco, CA). A 96-wellmicrotitration plate was purchased from the GreinerCo. (Frickenhausen, Germany). All other chemicalswere of the best grade available from commercialsources.
2.2. Tissue samples
For the immunohistochemical analysis, tissue sam-ples of the aorta were obtained at autopsy from 20nondiabetic patients (ten males and ten females; meanage, 53.8 years; range, 1–75 years). The autopsy wascarried out within 2–6 h postmortem. The causes ofdeath are listed in Table 1. All of the subjects were freeboth clinically and pathologically of diabetes mellitus.The samples consisted of grossly normal regions (dif-fuse intimal thickening), fatty streaks, atheroscleroticplaques and complicated lesions. The tissue specimenswere washed with PBS, and embedded in OCT com-pound. The embedded samples were rapidly frozen indry ice-acetone. Serial frozen sections were made by acryostat and stored at−80°C until use. The sectionswere stained with hematoxylin and eosin or oil red O toevaluate lipid deposition in the lesions.
2.3. Immunohistochemical methods
Immunohistochemical localization of various anti-gens, including CML, nonCML, oxPC and Apo-B, wasexamined using an avidin/biotin/alkaline phosphatasesystem. Briefly, serial tissue sections on microscopeslides were fixed in cold ethanol for 10 min and thenincubated for 5 min with 6% BSA to block nonspecificbinding. The slides were then reacted with the primaryantibody at room temperature for 60 min. The primary

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355 345
antibodies used in this study included polyclonal anti-nonCML, monoclonal anti-CML (6D12), anti-oxPC(FOH1a/DLH3) and anti-Apo-B antibodies, and mon-oclonal anti-a smooth muscle actin and anti-humanmacrophage (HAM 56) antibodies. After additionalwashes (in PBS, three times), biotinylated second anti-body (1:200) was added and the samples were allowedto sit for 30 min at room temperature. The slides werethen rinsed with PBS and incubated for 30 min withalkaline phosphatase-labeled streptavidin (1:100) atroom temperature. The alkaline phosphatase coloringreaction was carried out by incubating the slides withnaphthol-AS-BI-phosphoric acid as a substrate andhexazotized new fuchsin as a coupler in 0.2 M Tris–HCl buffer (pH 8.2). The reaction mixture containedlevamisole (24 mg/ml) to block endogenous alkalinephosphatase activity. After washing with PBS, the sec-tions were counterstained with Meyer hematoxylin.Control tests for the specificity of immunostaining in-cluded the substitution of non-immune sera or PBS forthe primary antibodies. To further verify the specificityof the staining of monoclonal anti-CML and polyclonalanti-nonCML antibodies, controls were performed bythe preadsorption of anti-AGEs and anti-CML fromthese primary antisera with purified AGE-BSA andCML-BSA, respectively. Three atherosclerotic lesions,i.e. diffuse intimal thickening (DIT), fatty streak (FS)and atherosclerotic plaque (AP), were examined in eachcase under a light microscope. Complicated lesions with
calcification and ulceration were excluded from thisstudy. The degree of extracellular staining of antigenswas evaluated by calculating the number of cases withpositively stained extracellular matrix as a percentageof the total number of cases examined. Regarding thedegree of staining of antigens in macrophage/foamcells, we counted the number of positively stained cellsas a percentage of the total number of cells within thedefined field, and then classified the values into threedegrees, including no stained cells (− ), and less (+ ) ormore (+ + ) than 50% under low-power lightmicroscopy.
2.4. Preparation of low density lipoprotein
LDL was isolated from freshly drawn plasmasamples (250 ml) from different human donors.Native LDL (nLDL) was prepared in the presence of1 mM ethylenediaminetetraacetic acid (EDTA) bya potassium bromide stepwise density gradient ultra-centrifugation [19]. After centrifugation, the fractionswith a density of 1.019–1.063 were obtained as LDLand then dialyzed at 4°C against excessive 0.15 M NaCland 1 mM EDTA (pH7.4). nLDL was sterilized byMillipore filtration (0.45 mm) and stored in the dark at4°C.
In vitro oxidation, glycation and glycoxidation oflow density lipoprotein. All solutions were sterilized byfiltration through 0.45-mm pore membranes into sterileconical tubes, and the modifications were carried out inthe absence of EDTA at 37°C for 2 weeks under sterileconditions. The oxidative modification of LDL wasperformed by incubating freshly prepared LDL with 5or 10 mM CuSO4. Glycated LDL was prepared byincubating freshly prepared LDL with 25 or 500 mMglucose under a stream of N2 gas. The glycoxidativemodification of LDL was performed by incubatingfreshly prepared LDL with both 5–10 mM CuSO4 and25–500 mM glucose. The modification was stopped byextensive dialysis of the reaction mixtures against 0.15M NaCl and 1 mM EDTA at 4°C. Furthermore, LDLwas incubated in PBS without glucose and copper ionat 37°C for 2 weeks, which was referred to as nongly-cated/nonoxidized LDL. Native or nonglycated/nonox-idized LDL was used as control.
2.5. Analysis of TBARS, electrophoretic mobility andearly glycation products
The extent of oxidation in modified LDL was esti-mated by the formation of thiobarbituric acid reactivesubstrate (TBARS) and by changes in the elec-trophoretic mobility on agarose gel [20]. The formationof early glycation products in modified LDL was as-
Table 1Age, sex and diagnosis of subjects examined
Sex Diagnosis at autopsyAgeNo
Malignant lymphomaF1 1212 F Malignant lymphoma243 M Acute myelocyticleukemia
4 28 M Acute myelocyticleukemiaM315 Dilatated cardiomyopathy
476 M Gastric adenocarcinoma.7 50 F Cervical carcinoma of uterus
Carcinoma of gall bladder8 F60609 F Pulmonary tuberculosis61 F10 Mucinous adenocarcinoma of the ovary
Cerebellar thromboembolism, myocardialF11 63infarctionAcute gastric ulcer with arterial rupture6612 M
F13 Mixed lineageleukemia686814 M Gastric adenocarcinoma.
15 M Prostaticadenocarcinoma, gastric69adenocarcinoma
M69 Mitral valve regurgitation16F17 Angiotropic lymphoma, breast cancer71
7218 M Lung cancer, renal cell carcinomaLung cancerM7319
7520 F Rupture of the duodenal varices

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355346
sessed by determining fructosamine using the Fruc-tosamine Test (Hoffman-La Roche, Switzerland), aspreviously described [21].
2.6. E6aluation of the formation of CML- and oxPC-epitopes by noncompetiti6e ELISA
The formation of CML and oxPC was estimated bynoncompetitive ELISA with anti-CML and anti-oxPCantibodies, respectively. All of the procedures werecarried out at room temperature. The wells of microt-iter plates were coated with various concentrations ofsamples in 0.1 ml of carbonate buffer for 60 min. Thewells were washed three times with washing buffer (PBScontaining 0.05% Tween 20), and then incubated with0.5% gelatin for 60 min to block nonspecific binding.After the wells were washed with washing buffer, 100 mlof anti-CML or anti-oxPC antibody in PBS containing0.1% BSA and 0.05% Tween 20 was added to each welland incubated for 60 min. After further washings, thewells were incubated with 100 ml of peroxidase-conju-gated second antibody for 60 min. The reactivity ofperoxidase was determined by incubation with o-phenylenediamine dihydrochloride for appropriate in-tervals, and the absorbance at 492 nm was measured onan ELISA plate reader.
2.7. Effects of chelators and aminoguanidine onoxidation, glycation and the formation of CML- andoxPC-epitopes
Incubation of LDL with copper ion and/or glucosewas carried out at 37°C for 2 weeks in the presence ofmetal chelators, 1 mM diethylenetriaminepentaaceticacid (DTPA) and 1 mM EDTA, or 100 mM amino-guanidine. After extensive dialysis against PBS, elec-trophoretic mobility, TBARS, fructosamine contentsand the formation of CML- and oxPC-epitopes weredetermined as described above.
Fig. 1. Immunohistochemical localization of nonCML-epitope andapolipoprotein B in diffuse intimal thickening (DIT) of aorta. The redalkaline phosphatase reaction product is localized at antigenic sites.Although nonCML was not formed in DIT of aorta from a 21-year-old subject (A); apolipoprotein B was observed in the intima (B). Anextra-cellular deposit of nonCML was seen in collagen fibers in DITfrom a 68-year-old subject (C, arrows). As shown in D, apolipo-protein B was observed not only in nonCML-positively stainedcollagen (arrows), but also in other extracellular spaces. Originalmagnification is ×350 for all micrographs.Fig. 2. Comparative immunolocalization of CML (A)-, oxPC (B)-and nonCML (C)-epitopes and apolipoprotein B (D) in the fattystreaks of aorta from a 63-year-old subject. In a fatty streak, theCML- and oxPC-epitopes were accumulated exclusively in foam cellswhich were positive for HAM-56 (E). In contrast, the nonCML-epi-tope and apolipoprotein B were found mainly in the extracellularspaces in the intima. Original magnification is ×210 for all micro-graphs.

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355 347
Fig. 3. Comparative immunostaining of the CML (A)-, oxPC (B)- and nonCML (C)-epitopes and apolipoprotein B (D) in the atheroscleroticplaques from a 68-year-old subject. Immunoreactivity for the CML-epitope is found both in foam cells and in the extracellular fibrous area(indicated by an arrow). The oxPC-epitope was also found in most of the foam cells. Extracellular matrices were faintly positive for oxPC-epitope(indicated by arrows). On the other hand, immunoreactivity for nonCML and apolipoprotein B was seen exclusively in the extracellular fibrousarea, but not in foam cells. Original magnification is ×170 for all micrographs.
Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355348
2.8. Statistical analysis
Numerical data are expressed as the mean9SE.Statistical analyses were performed using Mann–Whit-ney U-test, Chi-square test and unpaired Student’s t-test.P-values of less than 0.05 were considered significant.
3. Results
3.1. Immunohistochemical localization of CML- andoxPC-epitopes in atherosclerotic lesions
AGEs, including CML- and nonCML-epitopes,showed an age-dependent staining pattern in diffuseintimal thickening (DIT). As shown in Fig. 1A and B,neither CML- nor nonCML-epitope was formed in DIT
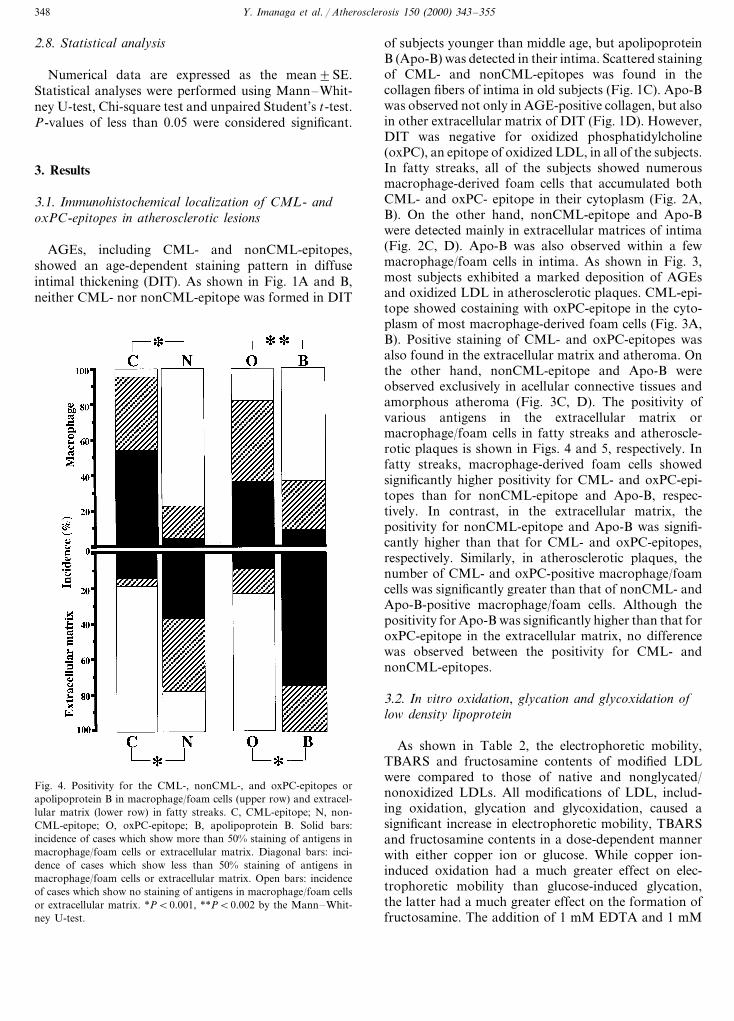
of subjects younger than middle age, but apolipoproteinB (Apo-B) was detected in their intima. Scattered stainingof CML- and nonCML-epitopes was found in thecollagen fibers of intima in old subjects (Fig. 1C). Apo-Bwas observed not only in AGE-positive collagen, but alsoin other extracellular matrix of DIT (Fig. 1D). However,DIT was negative for oxidized phosphatidylcholine(oxPC), an epitope of oxidized LDL, in all of the subjects.In fatty streaks, all of the subjects showed numerousmacrophage-derived foam cells that accumulated bothCML- and oxPC- epitope in their cytoplasm (Fig. 2A,B). On the other hand, nonCML-epitope and Apo-Bwere detected mainly in extracellular matrices of intima(Fig. 2C, D). Apo-B was also observed within a fewmacrophage/foam cells in intima. As shown in Fig. 3,most subjects exhibited a marked deposition of AGEsand oxidized LDL in atherosclerotic plaques. CML-epi-tope showed costaining with oxPC-epitope in the cyto-plasm of most macrophage-derived foam cells (Fig. 3A,B). Positive staining of CML- and oxPC-epitopes wasalso found in the extracellular matrix and atheroma. Onthe other hand, nonCML-epitope and Apo-B wereobserved exclusively in acellular connective tissues andamorphous atheroma (Fig. 3C, D). The positivity ofvarious antigens in the extracellular matrix ormacrophage/foam cells in fatty streaks and atheroscle-rotic plaques is shown in Figs. 4 and 5, respectively. Infatty streaks, macrophage-derived foam cells showedsignificantly higher positivity for CML- and oxPC-epi-topes than for nonCML-epitope and Apo-B, respec-tively. In contrast, in the extracellular matrix, thepositivity for nonCML-epitope and Apo-B was signifi-cantly higher than that for CML- and oxPC-epitopes,respectively. Similarly, in atherosclerotic plaques, thenumber of CML- and oxPC-positive macrophage/foamcells was significantly greater than that of nonCML- andApo-B-positive macrophage/foam cells. Although thepositivity for Apo-B was significantly higher than that foroxPC-epitope in the extracellular matrix, no differencewas observed between the positivity for CML- andnonCML-epitopes.
3.2. In 6itro oxidation, glycation and glycoxidation oflow density lipoprotein
As shown in Table 2, the electrophoretic mobility,TBARS and fructosamine contents of modified LDLwere compared to those of native and nonglycated/nonoxidized LDLs. All modifications of LDL, includ-ing oxidation, glycation and glycoxidation, caused asignificant increase in electrophoretic mobility, TBARSand fructosamine contents in a dose-dependent mannerwith either copper ion or glucose. While copper ion-induced oxidation had a much greater effect on elec-trophoretic mobility than glucose-induced glycation,the latter had a much greater effect on the formation offructosamine. The addition of 1 mM EDTA and 1 mM
Fig. 4. Positivity for the CML-, nonCML-, and oxPC-epitopes orapolipoprotein B in macrophage/foam cells (upper row) and extracel-lular matrix (lower row) in fatty streaks. C, CML-epitope; N, non-CML-epitope; O, oxPC-epitope; B, apolipoprotein B. Solid bars:incidence of cases which show more than 50% staining of antigens inmacrophage/foam cells or extracellular matrix. Diagonal bars: inci-dence of cases which show less than 50% staining of antigens inmacrophage/foam cells or extracellular matrix. Open bars: incidenceof cases which show no staining of antigens in macrophage/foam cellsor extracellular matrix. *PB0.001, **PB0.002 by the Mann–Whit-ney U-test.

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355 349
Fig. 5. Positivity for the CML-, nonCML-, and oxPC-epitopes orapolipoprotein B in macrophage/foam cells (upper row) and extracel-lular matrix (lower row) in atherosclerotic plaques. C, CML-epitope;N, nonCML-epitope; O, oxPC-epitope; B, apolipoprotein B. Solidbars: incidence of cases which show more than 50% staining ofantigens in macrophage/foam cells or extracellular matrix. Diagonalbars: incidence of cases which show less than 50% staining of antigensin macrophage/foam cells or extracellular matrix. Open bars: inci-dence of cases which show no staining of antigens in macrophage/foam cells or extracellular matrix. *PB0.0001, **PB0.001,***PB0.005 by the Mann–Whitney U-test.
concentration (25 mM) of glucose, but was generated tosome extent by incubation with a high concentration(500 mM) of glucose (Fig. 6A). As shown in Fig. 7A,incubation of LDL with glucose alone did not generatethe oxPC-epitope. On the other hand, the oxidative andglycoxidative modification of LDL caused the forma-tion of oxPC-epitope, which was dependent on copperion, but independent of glucose (Fig. 7B, C).
3.4. Effects of chelators and aminoguanidine on theformation of CML- and oxPC-epitopes in low densitylipoprotein
As shown in Fig. 8A, the addition of chelatorscompletely blocked the formation of CML in LDLincubated with copper ion and/or a high concentration(500 mM) of glucose. Aminoguanidine decreased theformation of CML-epitope in glycated and glycoxida-tive LDL by approximately 70–98% (Fig. 8B). Fig. 9shows the effects of chelators and aminoguanidine onthe formation of oxPC-epitope. The chelators com-pletely blocked the formation of oxPC in glycoxidativeand oxidized LDL (Fig. 9A). On the other hand, theaddition of aminoguanidine reduced the formation ofoxPC-epitope by approximately 50% in oxidized andglycoxidative LDL incubated with 10 mM copper ion orboth 500 mM glucose and 10 mM copper ion (Fig. 9B).
4. Discussion
The present study demonstrates for the first timeindirect evidence for the presence of glycoxidative LDL,which may contribute to the progression of humanatherogenesis.
Advanced glycation end products (AGEs) consist ofheterogeneous chemical compounds, including N o-(car-boxymethyl)lysine (CML) [11], pentosidine [22] andpyrraline [23]. Although the in vivo localization ofAGEs has been extensively studied in various tissues,such as artery, skin and kidney, some controversy stillexists about its location in atherosclerotic lesions. We[2] previously demonstrated that CML-epitope was lo-cated within macrophage-derived foam cells while non-CML-epitope was located mainly in the extracellularspace in atherosclerotic lesions. The formation of CMLand pentosidine has been shown to depend on theoxidation of glycated protein [24]. On the other hand,although the chemical structure of nonCML-epitopehas not yet been identified, we showed that nonCML-epitope was formed by oxidation-independent glycation[25]. Thus, glycoxidative products may be accumulatedin macrophage-derived foam cells, and oxidation-inde-pendent glycated proteins may be formed in the extra-cellular matrices in atherosclerotic lesions.
DTPA inhibited the increased electrophoretic mobilityand TBARS in oxidative, glycated and glycoxidativeLDL, but did not have any effect on the enhancedformation of fructosamine in glycated and glycoxida-tive LDL.
3.3. In 6itro formation of CML- and oxPC-epitopes inlow density lipoprotein by oxidation, glycation andglycoxidation
Fig. 6 shows the formation of CML-epitope of LDLusing noncompetitive ELISA. The incubation of LDLwith both copper ion and glucose (glycoxidative modifi-cation of LDL) increased the immunoreactivity to anti-CML antibody in a dose-dependent manner (Fig. 6C).On the other hand, copper ion-induced oxidation ofLDL failed to form the CML-epitope (Fig. 6B). CML-epitope was not generated by incubation with a low

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355350
Table 2Electrophoretic mobility, TBARS and fructosamine of modified low density lipoprotein
ElectrophoreticAdministration TBARS (nmolLDL Fructosamine(m mol/l)MDA/mg)mobility
Cu++ (mM) EDTA (mM) DTPA (mM)Glucose (mM)
– 1 1.5990.10– 6.7990.67––Native LDL–Nonglycated/ –– – 1.34 1.5990.20 13.390.63
nonoxidizedLDL
– 2.1925 6.1990.50*– 22.590.84*Glycated LDL –500 – – 2.72 4.1390.45 47.492.10*–
– – – 2.75 5.3890.37* 18.690.53*Oxidised LDL 0.5– 3.50 7.2590.26*– 18.490.88*10–
Glycoxidative 0.5 – – 2.63 6.2390.98 21.091.00*25LDL
– 3.19 5.0690.13*10 24.190.80*–25500 – – 2.94 4.6490.22* 52.691.40*0.5
–500 – 3.13 3.2390.24 56.290.89*101.0 1.0 1.25 1.1590.61– 43.091.31*Effect of 500
chelators1.0 1.10– 0.6990.08 8.6791.2010 1.01.0 1.21 1.1490.291.0 48.291.36*0.5500
1.0500 1.0 1.21 0.9090.09 57.891.60*10
* PB0.0001, compared with native LDL, n=4.
Fig. 6. CML-epitope formed by incubating LDL with glucose (A), copper ion (B) or glucose and copper ion (C). The formation of CML-epitopewas estimated by non-competitive ELISA with anti-CML antibody (6D12). LDL incubated with both glucose and copper ion showed an increasein immunoreactivity to anti-CML antibody with increasing concentrations of either glucose or copper ion. In contrast, incubation with copper ionor 25 mM glucose alone failed to increase the immunoreactivity of LDL to anti-CML antibody. LDL incubated with 500 mM glucose showeda modest increase in immunoreactivity to anti-CML antibody. The experiments were repeated three times. The data represent the mean values fortriplicate determinations.

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355 351
Fig. 7. oxPC-epitope formed by incubating LDL with glucose (A), copper ion (B) or glucose and copper ion (C). The formation of oxPC-epitopewas estimated by non- competitive ELISA with anti-oxPC antibody (FOH1a/DLH3). LDL incubated with copper ion alone or both glucose andcopper ion showed increased immunoreactivity to anti-oxPC antibody, which depended on the concentration of copper ion, but not on that ofglucose. Incubation with glucose alone did not increase the immunoreactivity of LDL to anti-oxPC antibody. The experiments were repeated threetimes. The data represent the mean values for triplicate determinations.
Several studies have shown that the oxidation of lowdensity lipoprotein (LDL) is implicated in atherogene-sis. Foam cells, which were found in atheroscleroticlesions, were formed from macrophages by taking upoxidized LDL through several receptors, and con-tributed to the progression of the lesions [26,27]. Im-munochemical and immunohistochemical analyses havedemonstrated the presence of oxidized LDL in humanplasma and atherosclerotic lesions [15,28]. A ‘model’compound of AGEs, 2-furoyl-4[5]-(2-furanyl)-1H-imi-dazole, has been shown to colocalize with epitopesgenerated during the oxidation of LDL, e.g. MDA-lysine and 4-hydroxynoneal-lysine, in the atheroscle-rotic lesions of euglycemic rabbits [3]. We [14]previously showed the colocalization of epitopes ofAGEs, CML and nonCML, and apolipoprotein B inintimal lesions of human aorta. Therefore, it is possiblethat the nonenzymatic glycation of proteins, includingLDL and extracellular matrix components, may beclosely linked to lipid peroxidation in the arterial in-tima. The present study demonstrated that CML-epi-tope, a major product of the oxidative modification ofglycated proteins, and oxPC, an epitope of oxidizedLDL, existed mainly in the cytoplasm of macrophage-derived foam cells in atherosclerotic lesions. On theother hand, nonCML, an oxidation-independent epi-tope, and apolipoprotein B were located mainly in the
extracellular matrix of intimal lesions. These resultssuggest that glycoxidation may be involved in the oxi-dative modification of LDL in atherosclerotic lesion.
AGEs have been shown to increase with age incollagen of various tissues, including arteries and skin[7,29]. Our previous studies [2,14] showed that AGE-modified collagen significantly increased with age inDIT of human aorta. In this study, DIT of the subjectsyounger than middle age was positive for apolipo-protein B, but negative for AGEs, including CML andnonCML. In DIT of elderly persons, apolipoprotein Bwas observed not only in nonCML- positive collagen,but also in other extracellular matrices. However, DITwas negative for oxPC, an epitope of oxidized LDL, inall of the subjects. LDL has been shown to be boundspecifically by AGE of collagen [30]. Thus, AGEs ob-served in collagen may contribute to increased LDLtrapping in DIT of aorta.
In vitro experiments have shown that CML is formedby the metal ion-catalyzed oxidative cleavage of fruc-tose-lysine or glycated human serum albumin [11,12].Glycated LDL has been shown to be more susceptibleto metal ion-catalyzed oxidative modification than na-tive LDL [8,9]. Therefore, we examined the possibilityof CML formation by incubating LDL with both glu-cose and copper ion as an in vitro model of glycoxida-tive modification. Upon incubation with both glucose

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355352
and copper ion, LDL dramatically increased its im-munoreactivity to anti-CML antibody, and this de-pended on the concentration of either glucose or copperion. Moreover, the formation of CML-epitope in gly-coxidative LDL was inhibited by chelators andaminoguanidine. On the other hand, CML was notformed by incubating LDL with copper ion alone orwith 25 mM of glucose. These results suggest that theCML-epitope may be formed by the glycoxidative mod-ification of LDL. Although CML was not detected inLDL incubated with 25 mM of glucose alone, theincubation with a high concentration (500 mM) ofglucose led to the formation of CML in LDL to someextent. AGEs generate in a manner dose-dependent onglucose, and its formation requires a period of severalweeks. The time-course study showed that a period ofover 3 weeks was required to detect CML in BSAincubated with glucose alone [18]. Therefore the incu-bation time may be too short to achieve adequate CMLlevels in LDL incubated with 25 mM of glucose. On theother hand, the formation of CML in LDL incubatedwith a high concentration of glucose might be causedby the autoxidation of glucose [31].
Recently, CML has been shown to be formed by thecopper-ion induced oxidation of LDL, which may bederived from products of lipid peroxidation rather thanfrom carbohydrates [32]. However, in this study, the
CML-epitope was not detected in LDL incubated withcopper ion alone. This discrepancy may be due to thedifferent assay systems for CML. On the other hand,the incubation of LDL with copper ion in the presenceor absence of glucose generated the oxPC-epitope. Theformation of oxPC-epitope in LDL was dependent oncopper ion-catalyzed oxidation, but independent of in-cubation with glucose. These results suggest that theoxPC-epitope may be formed only by the copper ion-induced peroxidation of LDL.
Although the mechanism of CML formation is notfully understood, CML has been proposed to beformed in vitro through three pathways, i.e. via theoxidative fragmentation of Amadori products [11],from a Schiff base [33], and through modification withglyoxal generated directly by the autoxidation of glu-cose [31]. In vivo, CML is thought to be primarilyformed by the oxidative cleavage of Amadori products[34]. In this study, the glycoxidative modification ofLDL generated CML as well as fructosamine, an earlyglycation product. The formation of CML was inhib-ited not only by chelators, but also by aminoguanidine.Thus, CML might be generated mainly by the copperion-catalyzed oxidation of Amadori products in this invitro model of glycoxidation of LDL.
Where does LDL undergo glycoxidative modificationin arterial tissues? In this study, an immunohistochemi-
Fig. 8. Effects of chelators (A) and aminoguanidine (B) on the formation of CML-epitope in LDL incubated with 500 mM glucose and/or copperion. The addition of chelators, including 1 mM EDTA and 1 mM DTPA, and aminoguanidine significantly inhibited the formation ofCML-epitope in LDL incubated with 500 mM glucose and/or copper ion. The experiments were repeated three times. The data represent the meanvalues for triplicate determinations.

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355 353
Fig. 9. Effects of chelators (A) and aminoguanidine (B) on the formation of oxPC-epitope in LDL incubated with 500 mM glucose and/or copperion. The addition of chelators, including 1 mM EDTA and 1 mM DTPA, completely blocked the formation of oxPC-epitope in the copperion-induced oxidation or glycoxidation of LDL. Aminoguanidine inhibited the formation of oxPC-epitope in the glycoxidative modification ofLDL to some extent, but had no effect on that in oxidized LDL. The experiments were repeated three times. The data represent the mean valuesfor triplicate determinations.
cal analysis showed that macrophage-derived foam cellswere strongly stained by both anti-CML and anti-oxPCantibodies. However, CML- and oxPC-epitopes werealso observed in extracellular matrices including colla-gen fibers and atheroma. The nonCML epitope andApo-B were localized mainly in extracellular spaces inall intimal lesions. Recently, malondialdehyde, a glyca-tion-independent lipid peroxidation product, was de-tected in diabetic nodular lesions with the accumulationof CML and pentosidine [35]. Macrophages are knownto mediate the endocytic uptake of oxidized LDL andAGEs through scavenger receptors as well as AGEreceptor [26,27,36,37]. Thus, at least in advanced lesions,LDL can undergo glycoxidative modification in extra-cellular spaces and be taken up by macrophages, whichmay lead to the intracellular accumulation of glycoxida-tive LDL recognized by both anti-CML and anti-oxPCantibodies. However, this study could not rule out thepossibility that modified LDL may undergo endocytosisby macrophages, followed by glycoxidative modifica-tion.
Diabetes is one of the main risk factors of atheroscle-rosis. The metabolic events responsible for its develop-
ment are not fully understood. We [2] previously showedthat different AGE- epitopes, including CML and non-CML, were found in human atherosclerotic lesions.CML and pentosidine have been shown to increase inplasma and tissues of patients with diabetes mellitus[5,35,38]. In this study, CML- and oxPC-epiotpes werecolocalized in macrophage-derived foam cells inatherosclerotic lesions. In vitro experiments showed theformation of CML- and oxPC-epitopes in glycoxidativeLDL. Therefore, hyperglycemia may enhance the gly-coxidative modification in LDL, which may thus con-tribute to the development of diabetic macroangiopathy.
In summary, an immunohistochemical analysis forCML- and oxPC-epitopes in atherosclerotic lesionsshowed their colocalization in the cytoplasm ofmacrophage-derived foam cells. In contrast, nonCML-epitope and apolipoprotein B were found mainly in theextracellular matrix of these lesions. In vitro experimentsshowed that incubation of LDL with both copper ionand glucose caused the formation CML-epitope, whichwas inhibited by the addition of either chelators oraminoguanidine. On the other hand, oxPC-epitope wasformed by incubating LDL with copper ion regardless of

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355354
the presence or absence of glucose. These results sug-gest that LDL undergoes glycoxidative modification,which may contribute to the development of humanathero-sclerotic lesions.
Acknowledgements
The authors are grateful to Yuko Aoyagi for herexcellent technical assistance. This work was funded bya Grant-in-Aid from the Ministry of Education, Scienceand Culture of Japan (No. 10670183).
References
[1] Vlassara H, Bucala R, Striker L. Pathogenic effects of advancedglycosylation: biochemical, biologic, and clinical implications fordiabetes and aging. Lab Invest 1994;70:138–51.
[2] Sakata N, Imanaga Y, Meng J, Tachikawa Y, Takebayashi S,Nagai R, et al. Immunohistochemical localization of differentepitopes of advanced glycation end products in humanatherosclerotic lesions. Atherosclerosis 1998;141:61–75.
[3] Palinski W, Koschinsky T, Butler SW, Miller E, Vlassara H,Cerami A, et al. Immunological evidence for the presence ofadvanced glycosylation end products in atherosclerotic lesions ofeuglycemic rabbits. Arterioscler Thromb Vasc Biol 1995;15:571–82.
[4] Baynes JW. Role of oxidative stress in development of complica-tions in diabetes. Diabetes 1991;40:405–12.
[5] Schleicher ED, Wagner E, Nerlich AG. Increased accumulationof the glycoxidation product No-(carboxymethyl)lysine in humantissues in diabetes and aging. J Clin Invest 1997;99:457–68.
[6] Meng J, Sakata N, Takebayashi S, Asano T, Futata T, Nagai R,et al. Glycoxidation in aortic collagen from STZ-induced dia-betic rats and its relevance to vascular damage. Atherosclerosis1998;136:355–65.
[7] Odetti P, Pronzato MA, noberasco G, Cosso L, Traverso N,Cottalasso D, et al. Relationships between glycation and oxida-tion-related fluorescences in rat collagen during aging. Lab In-vest 1994;70:61–7.
[8] Sakurai T, Kimura S, Nakano M, Kimura H. Oxidative modifi-cation of glycated low density lipoprotein in the presence of iron.Biochem Biophys Res Comm 1991;177:433–9.
[9] Kawamura M, Heinecke JW, Chait A. Pathophysiological con-centrations of glucose promote oxidative modification of lowdensity lipoprotein by a superoxide-dependent pathway. J ClinInvest 1995;94:771–8.
[10] Sakurai T, Tsuchiya S. Superoxide production from nonenzy-matically glycated protein. FEBS Lett 1988;236:406–10.
[11] Ahmed MU, Thorpe SR, Baynes JW. Identification of No-(car-boxymethyl)lysine as a degradation product of fructoselysine inglycated protein. J Biol Chem 1986;261:4889–94.
[12] Nagai R, Ikeda K, Higashi T, Sano H, Jinnouchi Y, Araki T, etal. Hydroxyl radical mediates No-(carboxymethyl)lysine forma-tion from Amadori product. Biochem Biophys Res Comm1997;234:167–72.
[13] Kume S, Takeya M, Mori T, Araki N, Suzuki H, Horiuchi S, etal. Immunohistochemical and ultrastructural detection of ad-vanced glycation end products in atherosclerotic lesions of hu-man aorta with a novel specific monoclonal antibody. Am JPathol 1995;147:654–67.
[14] Sakata N, Imanaga Y, Meng J, Tachikawa Y, Takebayashi S,Nagai R, et al. Increased advanced glycation end products in
atherosclerotic lesions of patients with end-stage renal disease.Atherosclerosis 1999;142:67–77.
[15] Itabe H, Takeshima E, Iwasaki H, Kimura J, Yoshida Y,Imanaka T, et al. A monoclonal antibody against oxidizedlipoprotein recognizes foam cells in atherosclerotic lesions. J BiolChem 1994;269:15274–9.
[16] Itabe H, Yamamoto H, Imanaka T, Shimamura K, UchiyamaH, Kimura J, et al. Sensitive detection of oxidatively modifiedlow density lipoprotein using a monoclonal antibody. J LipidRes 1996;37:45–53.
[17] Horiuchi S, Araki N, Morino Y. Immunochemical approach tocharacterize advanced glycation end products of the Maillardreaction: evidence for the presence of a common structure. J BiolChem 1991;266:7329–32.
[18] Ikeda K, Higashi T, Sano H, Jinnouchi Y, Yoshida M, Araki T,et al. No-(carboxymethyl)lysine protein adduct is a major im-munological epitope in proteins modified with advanced glyca-tion end products of the Maillard reaction. Biochemistry1996;35:8075–83.
[19] Radgrave TG, Rogert DCK, West CE. Separation of plasmalipoproteins by density gradient ultracentrifugation. AnalBiochem 1975;65:42–9.
[20] Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides inanimal tissues by thiobarbituric acid reaction. Anal Biochem1979;95:351–8.
[21] Johnson RN, Metcalf PA, Baker JR. Fructosamine: a newapproach to the estimation of serum glycosylation. An index ofdiabetic control. Clin Chim Acta 1982;127:87–95.
[22] Sell DR, Monnier VM. Structure elucidation of a senescencecross-link from human extracellular matrix: implication of pen-toses in aging process. J Biol Chem 1989;264:21597–602.
[23] Hayase F, Nagaraj RH, Miyata S, Njoroge FG, Monnier VM.Aging of proteins: immunological detection of a glucose-derivedpyrrole formed during Maillard reaction in vivo. J Biol Chem1989;264:3758–64.
[24] Dyer DG, Blackledge JA, Thorpe SR, Baynes JW. Formation ofpentosidine during nonenzymatic browning of protein by glu-cose: identification of glucose and other carbohydrates as possi-ble precursors of pentosidine in vivo. J Biol Chem1991;266:11654–60.
[25] Ikeda K, Nagai R, Sakamoto T, Sano H, Araki T, Sakata N, etal. Immunochemical approaches to AGE-structures: characteri-zation of anti-AGE antibodies. J Immunol Methods1998;215:95–104.
[26] Endemann G, Stanton LW, Maddem KS, Bryant CM, WhiteRT, Protter AA. CD36 is a receptor for oxidized low densitylipoprotein. J Biol Chem 1993;269:11811–6.
[27] Freeman MY, Ekkel Y, Rohrer L, Penmen M, Freeman NJ,Chisolm GM, et al. Expression of type I and type II bovinescavenger receptors in Chinese hamster ovary cells: lipid propletaccumulation and nonreciprocal cross competition by acetylatedand oxidized low density lipoprotein. Proc Natl Acad Sci USA1991;88:4931–5.
[28] Palinski W, Rosenfeld ME, Yla-Herittuala S, Gurtner GC,Socher SS, Butler SW, et al. Low density lipoprotein undergoesoxidative modification in vivo. Proc Natl Acad Sci USA1989;86:1372–6.
[29] Reiser KM. Nonenzymatic glycation of collagen in aging anddiabetes: minireview. Proc Soc Exp Biol Med 1991;196:17–29.
[30] Brownlee M, Vlassara H, Cerami A. Nonenzymatic glycosyla-tion products on collagen covalently trap low-density lipo-protein. Diabetes 1985;34:938–41.
[31] Wells-Knecht KJ, Zyzak DV, Litchfield JE, Thorpes SR, BaynesJW. Mechanism of autoxidative glycosylation: identification ofglyoxal and arabinose as intermediates in the autoxidative mod-ification of proteins by glucose. Biochemistry 1995;34:3702–9.

Y. Imanaga et al. / Atherosclerosis 150 (2000) 343–355 355
[32] Requena JR, Fu MX, Ahmed MU, Jenkins AJ, Lyons TJ,Thorpe SR. Lipoxidation products as biomarkers of oxidativedamage to proteins during lipid peroxidation reactions. NephrolDial Transplant 1996;11(Suppl 5):48–53.
[33] Glomb MA, Monnier VM. Mechanism of protein modificationby glyoxal and glycoaldehyde, reactive intermediates of theMaillard reaction. J Biol Chem 1995;270:10017–26.
[34] Wells-Knecht MC, Thorpe SR, Baynes JW. Pathways of forma-tion of glycoxidation products during glycation of collagen.Biochemistry 1995;34:15134–41.
[35] Horie K, Miyata T, Maeda K, Miyata S, Sugiyama S, SakaiH, et al. Immunohistochemical colocalization of glycoxida-tion products and lipid peroxidation products in dia-
betic renal glomerular lesions. J Clin Invest 1997;100:2995–3004.
[36] Yui S, Sasaki T, Araki N, Horiuchi S, Yamasaki M. Inductionof macrophage growth by advanced glycation end products ofthe Maillard reaction. J Immunol 1994;152:1943–9.
[37] Goldstein JL, Ho YK, Basu SK, Brown MS. Binding site onmacrophages that mediates uptake and degradation of acetylatedlow density lipoprotein, producing massive cholesterol deposi-tion. Proc Natl Acad Sci USA 1979;76:333–7.
[38] Sell DR, Lapolla A, Odetti P, Fogarty J, Monnier VM. Pento-sidine formation in skin correlates with severity of complicationin individuals with long-standing IDDM. Diabetes 1992;41:1286–92.
..





























