Embed Size (px)
Citation preview

Univerzitet u Nišu Prirodno-matematički fakultet
Departman za biologiju i ekologiju
Ivan D. Todosijević
Indikatorska sposobnost dve grupe
makrozoobentosa za procenu kvaliteta vode sliva reke Nišave
Master rad
Niš, 2013. godina

2
Univerzitet u Nišu Prirodno-matematički fakultet
Departman za biologiju i ekologiju
MASTER RAD
Indikatorska sposobnost dve grupe makrozoobentosa za procenu kvaliteta
vode sliva reke Nišave
Mentor: Student: Prof. dr Slaviša Stamenković Ivan Todosijević
Niš, Februar 2013.

3
University of Niš Faculty of sciences and mathematics Department of Biology and Ecology
Ivan D. Todosijević
Two groups of macrozoobenthos as a water quality indicators in the Nišava river
basin
Master thesis
Niš, February 2013.

4
Ovom prilikom se zahvaljujem prof. dr Slaviši Stamenkoviću na korektnom i profesionalnom odnosu, stručnoj pomoći i korisnim sugestijama pri izradi master rada. Zahvaljujem se asistentima Djuradju Miloševiću i Milici Stojković na strpljenju, posvećenom vremenu, ukazanom poverenju i ustupljenoj literaturi. Posebno se zahvaljujem svojoj porodici i prijateljima koji su me podržavali u svojoj nameri da istrajem do kraja. Podaci za ovaj master rad korišćeni su iz doktorske disertacije Djuradja Miloševića i doktorske disertacije Milice Stojković.

5
INDIKATORSKA SPOSOBNOST DVE GRUPE MAKROZOOBENTOSA ZA PROCENU KVALITETA VODE SLIVA REKE NIŠAVE
Rezime
Cilj ovog istraživanja je bio da uporedimo sposobnost familije hironomida i grupe EPT u razdvajanju lokaliteta različitog nivoa kvaliteta vode.
Rezultati nMDS analize pokazali su koje su grupe lokaliteta
kvalitativno i kvantitativno slični. Ova studija je pokazala da se na nivou zajednice hironomida i
EPT grupe, sa taksonomskog rezolucijom do nivoa roda, mogu formirati grupe sa lokalitetima različitog kvaliteta i stepena antropogenog uticaja.
BioEnv analiza je izdvojila nadmorsku visinu kao jedan od
najbitnijih faktora sredine koji značajno utiče na struktuiranost kako zajednice hironomida tako i na grupe EPT.

6
TWO GROUPS OF MACROZOOBENTHOS AS A WATER QUALITY INDICATORS IN THE NIŠAVA RIVER BASIN Summary The main objective of this study was to compare discimination strenghts in assessment of water quality using the Chironomidae family and EPT group. Results of nMDS analyses revealed which group of sites are qualitative or quantitative similar. This study pointed out that information about community pattern of Chironomid and EPT group, identified up to genus level can be sufficient for reliable classification of sites with different level of anthropogenic impact. According to BioEnv analyses, the elevation is on of the most important environmental factor which has a great influence on community structure of Chironomids and EPT group.

7
Sadržaj
1. Uvod ...................................................................................... 8
1.1. Biomonitoring programi zasnovani na makrofauni dna ............... 9
1.2. Ephemeroptera, Plecoptera i Trichoptera kao bioindikatori ........ 12
1.3. Chironomidae kao bioindikatori .............................................. 12
1.4. Odnos EPT i Chironomidae .................................................... 14
1.5. Uticaj sredinskih faktora na zajednice makroinvertebrata .......... 14
1.6. Cilj ..................................................................................... 20
2. Materijal i metode ................................................................... 20
2.1. Oblast istraživanja ............................................................... 20
2.1.1. Nišava .......................................................................... 22
2.1.2. Temska sa Visočicom ...................................................... 25
2.1.3. Jerma ........................................................................... 27
2.2. Hidrološke karakteristike pritoka Nišave ................................. 28
2.3. Metode uzorkovanja ............................................................. 29
2.4. Analiza podataka ................................................................. 31
3. Rezultati ................................................................................ 31
3.1 Taksonomski sastav zajednice EPT i Chironomidae .................... 31
3.2 Prostorna distribucija zajednica EPT i Chironomidae .................. 37
4. Diskusija ............................................................................... 39
5. Zaključak .............................................................................. 42
6. Literatura .............................................................................. 43

8
1. Uvod Izračunato je da na površini Zemlje ima 1.6 milijardi km2 vode, što predstavlja 71% njene površine. Voda je na Zemlji prisutna u različitim rezervoarima kao što su okeani i mora (97% od ukupne količine vode), polarne ledene kape (2%) i u rekama, jezerima i podzemnim tokovima (svega 1%). Hidrobiologija je nauka o vodenim ekosistemima, pri čemu istražuje njihovu strukturu i funkciju, proučava fizičke, hemijske i biološke karakteristike akvatičnih sistema, uticaj koji vodena sredina ima na živi svet, kao i međusobne uticaje živih bića. Hidrobiologija se deli na okeanologiju koja proučava Svetske okeane i mora, i limnologiju koja proučava kopnene slatke vode. U užem smislu limnologija je nauka koja proučava jezerske ekosisteme, dok potamologija proučava rečne ekosisteme. Svaku reku karakteriše longitudinalni tok od izvora ka ušću. Na tom putu se menjaju životni uslovi i životna sredina, pa se na različitim delovima reke može naći drugačiji sastav životne zajednice. Rečni tok možemo podeliti na gornji tok (ritron) i donji tok (potamon). Razlikuju se po tome što gornji tok karakteriše brza, hladna i dobro aerisana voda, dno je peskovito i šljunkovito, vegetacija je slabo razvijena i organizmi su specifično prilagodjeni na veliku brzinu toka (dorzoventralno spljošteno delo, organi za pričvršćivanje i povećanje telesne mase), dok u donjem toku brzina i prozirnost se smanjuju, temperatura vode raste, ima manje kiseonika. Podloga je peskovita, muljevita i glinovita, pa zbog toga većina organizama koji žive na dnu zarivaju se u podlogu. Zagađena voda je voda koja kvantitativno i kvalitativno odstupa od prirodnih fizičkih, hemijskih i bioloških osobina, što ograničava mogućnost njene upotrebe i izaziva nepoželjne posledice u vodenim ekosistemima nanoseći štete ljudskom zdravlju i ekonomiji. Pod monitoringom stanja prirodne sredine, tačnije monitoringom antropogenih promena životne sredine, podrazumeva se organizovanje posebnog informacionog sistema za praćenje i analizu životne sredine, pre svega u pogledu zagađivanja i efekata zagađivanja u biosferi.

9
1.1. Biomonitoring programi zasnovani na makrofauni dna
Upotreba akvatičnih organizama za procenu kvaliteta vode datira još od početka prošlog veka (Kolkwitz i Marson), ali je njihova primena ostala nezapažena sve do sedamdesetih godina zato što su se dotadašnji monitoring programi oslanjali isključivo na hemijski i fizički monitoring. Jedan od problema oslanjanja samo na hemijske i fizičke metode je taj što one obezbedjuju podatke o stanju ekosistema samo u trenutku kada je uzrorak uzet. Suprotno tome, biološki monitoring daje podatke o prošlom i sadašnjem stanju ekosistema. Ovu razliku mozemo ilustrovati kao razliku izmedju slike i filma, gde bi fizičke i hemijske metode bile slika, a biološke metode film. Od svih slatkovodnih organizama koji mogu da se koriste u biomonitoringu makrozoobentos se pokazao kao najpouzdaniji (Hellawell 1986, Bonada et al. 2006, Carter et al. 2006). Makroinvertebrate se koriste za procenu efekta antropogenog stresa vodenih ekosistema na svim nivoima biološke organizacije (Rosenberg i Resh 1993). Na nivou organizma, promene u rastu i reprodukciji i stopi morfoloskih deformiteta su ocenjeni kao odgovor na povećano zagadjenje (Martinez et al. 2002). Najčešće, dinamika i struktura zajednica i populacija se ocenjuju kada se ispituju efekti zagadjenja u prirodi. Na primer, abundantnost, bogatstvo vrsta i ujednačenost vrsta zajednice makroinvertebrata se rutinski koriste u studijama o kvalitetu vode već decenijama. Naslovi radova koji su obeležili prethodne decenije mogu se dobiti na sledećoj referenci: Djuradj Milosevic: Barbour MT, Gerritsen J, Snyder BD, Stribling JB (1999) Rapid bioassessment protocols for use in streams and wadeable rivers: periphyton, bentic macroinvertebrates and fish, 2nd edn. EPA 841-b-99-002 U.S. Environmental Protection Agency, Office of Water, Washington DC
Bentosne makroinvertebrate predstavljaju integralni deo lotičkih sistema preradom organske materije i snabdevanjem energijom viših trofičkih nivoa; dakle, razumevanje efekta antrpogenih, takodje i prirodnih stresora, na njihovu distribuciju i abundancu je kritično za sveobuhvatnu procenu uticaja na potoke i reke.

10
Osnovni princip procene zagadjenja vrednovanjem strukture i funkcije makroinvertebrata je poredjenje potencijalno zagadjenih lokaliteta sa nezagadjenim. Nezagadjeni lokaliteti označavaju se kao kontrolni ili referentni lokaliteti, a lokaliteti za koje pretpostavlja da su zagadjeni kao test lokaliteti. Fraza kontrolni lokalitet retko se koristi u današnjici i zamenjen je konceptom referentni lokalitet ili referentno stanje. Dakle, studije su najčešće ograničene na područja koja imaju sličnu bruto fiziografiju. Ekoregioni, subekoregioni, tip zemljšnog pokrivača, veličina potoka i nadmorska visina su samo neki od kriterijuma za razmatranje kada ograničeni opsezi fizičkih varijabli mogu dovesti u zabunu poredjenjem zajednice makroinvertebrata izmedju lokaliteta u proceni uticaja.
Analitički pristup Analiziranje zajednice makroinvertebrata za biološku procenu je
često podeljena na dva pristupa: multimetrijski i multivarijantni. Glavna razlika izmedju ova dva pristupa leži u tome kako su varijable definisane.
Multimetrijski pristup Multimetrijski pristup je baziran na premisi da neke mere
bentosne zajednice mogu biti upotrebljene da indikuju svoje ekološko stanje i, u nastavku, stanje lotičkih ekosistema. Najsavremeniji istraživački postupci se oslanjaju na više mera strukture i funkcije zajednice. Ove mere mogu biti grupisane u nekoliko kategorija, kao sto su (1) taksonomsko bogatstvo (npr, nivo familije, roda i vrste) cele bentosne zajednice ili specifične kategorije koje su naizgled tolerantne (npr. Chironomidae) ili netolerantne (Ephemeroptera, Plecoptera i Trichoptera – nazvani EPT takson) na zagadjenje; (2) nabrajanje (npr. broj svih prikupljenih makroinvertebrata) ili proporcija izabranih redova kao sto su EPT; (3) indeksi diverziteta zajednice, koji generalno oslikavaju dominantnost (npr. Šenonov indeks); (4) funkcionalni odnos trofičkih gildi (procenat “shredder” funkcionalnih grupa); i (5) biotički indeksi. Biotički indeksi: u Velikoj Britaniji, za procenu kvaliteta vode najčešće se koristi Trent biotički indeks.

11
Ovaj indeks ne uzima u obzir abundantnost: jedna jedinka nekog taksona doprinosi isto kao i hiljadu jedinki nekog drugog taksona.
Kod ovog indeksa postoji 11 mogućih skorova (0-10), što za posledicu ima neosetljivost indeksa za većinu razlika koje se javljaju u karakteristikama zajednica. Sve nedostatke koje je imao Trent biotički sistem prevazidjeni su razvijanjem osetljivijeg Biotičkog skor sistema (Chandler and Biol, 1970). Ovaj indeks ima veći opseg mogućih razultata, što ga čini znatno osetljivijim nego Trent biotički sistem i samim tim boljim indeksom. U skorije vreme u oblastima gde su se biotički indeksi široko upotrebljavali pojavio se BMWP score (Armitage et al. 1983; ISO/BMWP, 1979) koji je zamenio sve prethodne indekse (inicijali označavaju Biological Monitoring Working Party). Za razliku od ranijih indeksa, BMWP score nije vezan za bilo koji region i može se podjednako koristiti u kontinentalnim razmerama.
Multivarijantni pristup Multivarijantni pristup razmatra svaki takson kao varijablu i
prisustvo ili abundantnost svakog taksona kao atribut lokacije ili kao vremensku tačku (Norris i Georges 1993). U suprotnosti sa multimetrijskim pristupom, vrednost povezana sa datim lokalitetom je funkcija tog taksonomskog sastava lokaliteta u odnosu na sastav svih drugih lokaliteta u analizi. Multivarijantni pristup je češće upotrebljavan za procenu stepena zagađenosti širokih razmera nego na studije tačkastih izvora.

12
1.2. Ephemeroptera, Plecoptera i Trichoptera kao bioindikatori Primena makrozoobetosa u svrhu biomonitoringa je proces koji zahteva vreme i ekspertizu. U poslednje vreme iz grupe makroinvertebrata se izdvajaju tri reda insekata (Ephemeroptera, Plecoptera i Trichoptera) i koriste u metodama biološke procene kvaliteta vode kao metrička osobina EPT indeks, koja na pouzdan način razdvaja zagađene od čistih lokaliteta (Wallace, Grubaugh i Whiles 1996). Naime, većina vrsta iz ove tri grupe je osetljiva na organsko zagađenje što ih čini dobrim i pouzdanim indikatorima kvaliteta vode. Zbog toga ukupan broj vrsta zabeleženih u uzorku opada kako se kvalitet životne sredine pogoršava. Takođe jednostavna identifikacija ovih grupa olakšava primenu EPT indeksa. Ephemeroptera, Plecoptera i Trichoptera (EPT) indeksi prikazuju bogatstvo unutar insekatskih grupa, dakle njihova brojnost treba da raste sa rastom kvaliteta vode. Prvobitno razvijen za indetifikaciju na nivou vrste, ovaj indeks važi za upotrebu na nivou familije (Plafkin et all. 1989). EPT indeks je jednak ukupnom broju familija reprezentovan unutar ova tri reda u uzorku.
1.3. Chironomidae kao bioindikatori Familija Chironomidae predstavlja grupu organizama iz podreda Nematocera, red Diptera. Svi predstavnici Chrironomidae predstavljaju holometabolne organizme, prolazeći kroz stadijume jajeta, larve, lutke i adulta. Ženke obično polažu jaja u želatinastoj masi na stabilnom supstratu poput kamenja, grana ili jakih biljaka. Dužina razvića jaja zavisi od spoljašnje temperature i obično traje nekoliko dana. Jaja se takodje mogu upotrebiti za determinaciju vrste (Vallenduuk i Moller Pillot 2007). Upotreba hironomida kao bioindikatora ima dugu istoriju u biomonitoringu kvaliteta voda (Richardson 1928, Graufin 1958, Saether 1979). Paine i Graufin su 1956. godine izveli jednogodišnja istraživanja kako bi odredili ekologiju i distribuciju ove grupe

13
organizama. Oni su zabeležili 94 vrsta iz 11 različitih vrsta koje su podelili u tri kategorije: 1. vrste zagadjenih voda koje opstaju samo pri niskoj koncentraciji kiseonika i visokoj koncentraciji organskih materija; 2. fakultativne ili tolerantne vrtse koje imaju široku ekološku valencu, te mogu izdržati velike promene u ekosistemima; 3. vrste čistih voda koje zahtevaju vodu visokog kvaliteta i sa dosta rastvorenog kiseonika. Zbog određene adaptiranosti i selektivnosti za različita staništa hironomide su se pokazali kao dobri bioindikatori stanja ekosistema. Postoji širok spektar heterogenosti u distribuciji larva hironomida, zato istraživanje treba sprovesti samo na jednom mikrostaništu ili uključiti sva postojeća mikrostaništa koja možemo naći na određenom lokalitetu. Drugi parametar koji može značajno varirati je vremensko variranje komponenti. Veliki taksonomski diverzitet familije Hironomida ima za posledicu veliko variranje životnih ciklusa kod različitih vrsta. Kod mnogih vrsta dolazi do preklapanja kohorti, što stvara poteškoće u odredjivanju kome stadijumu jedinka pripada. Treba uzeti u obzir da tokom leta većina larvi Hironomida se nalazi u blizini obale, dok se zimi povlače ka matici reke. Zbog poteškoća prilikom upotrebe hironomida kao bioindikatora Rabeni i Wang (2001) su testirali hipotezu da li isključivanje familije Chironomidae ima uticaj na osetljivost bioprocene kvaliteta vode. Na osnovu dobijenih rezultata naučnici su došli do zaključka da eliminacija hironomida iz istraživanja neće značajno izmeniti rezultate. Medjutim, neke metričke osobine kao što su biotički indeks, indeks diverziteta i ukupan broj taksona ne mogu biti izmereni sve dok se hironomide ne determinišu barem do nivoa roda, što je vremenski veoma zahtevno. U našoj zemlji su 70-tih i 80-tih godina prošlog veka radjena opsežna istraživanja faune hironomida, uglavnom od strane jednog autora (Janković 1979, Janković 1980, Janković 1983, Janković 1985). U svom radu “Larve hironomida kao bioindikatori zagadjenja“ Janković (1982) je pokušao da proceni kvalitet vode Južne Morave na osnovu zajednice hironomida. Uočeno je da prisustvo larvi hironimida najviše zavisi od brzine potoka i kvaliteta podloge, dok nešto manje zavisi od prisustva organske materije u sedimentu i količine rastvorenog kiseonika.

14
1.4. Odnos EPT i Chironomidae Abundantnost EPT i Chironomidae je metrička osobina koja ukazuje na balans u zajednici, pošto se smatra da su EPT osetljivije, a hironomide manje osetljive na sredinski stres (Plafkin et al 1989). U zajednici koja se smatra da je u dobrom biotičkom stanju pokazaće ravnomernu raspodelu izmedju ove četiri grupe, dok zajednice sa velikim brojem hironomida mogu ukazati na delovanje nekog sredinskog stresa (Plafkin et al. 1989). Medju svim analiziranim indeksima, utvrdjeno je da je EPT visoko korelisan sa nekoliko ostalih indeksa, uključujući ETO (Ephemeroptera, Trichoptera, Odonata). Uklanjanje ETO indeksa prikladno iz nekoliko razloga. Prvo, ETO indeks nema referencu i jedino se upotrebljava za uporedjivanje tokom vremena za studiju istog lokaliteta. EPT indeks takodje može biti analiziran na ovaj način. Drugo, kao i EPT, ovaj indeks obezbedjuje taksonomsko bogatstvo za nekoliko grupa makroinvertebrata za koje smatramo da su osetljive na zagadjenja i, dakle, ponovljive. Štaviše, i ETO i EPT uključuju taksonomsko bogatstvo Ephemeroptera i Trichoptera i razlikuju se samo u jednoj grupi (Odonata ili Plecoptera). Najveća korelacija dolazi izmedju % EPT i % ne-dipterskih insekata.
1.5. Uticaj sredinskih faktora na zajednice makroinvertebrata
Fizičke karakteristike kopnenih voda: kompleks abiotičkih faktora deluje na životnu zajednicu i odredjuje njenu strukturu i dinamiku. To je složen sistem velikog broja uticaja jer nijedan ekološki faktor ne deluje pojedinačno na životnu zajednicu. Ovi faktori su odredjeni kako horizontalnim položajem (polarne, umerene, suptropske i tropske oblasti) tako i vertikalnim položajem akvatičnih ekosistema (nadmorska visina).

15
U važne fizičke abiotičke faktore spadaju: 1. Temperatura 2. Svetlosni režim 3. Vodena kretanja 4. Struktura i sastav sedimenata Temperatura vode je jedan od najbitnijih ekoloških faktora koji primarno odredjuje sastav životne zajednice. Temperatura tekućih voda zavisi od geografske širine, godišnjeg doba, količine vode koju reka nosi i dubine na kojoj se voda nalazi. Termičke stratifikacije u rekama obično nema, sem u donji tokovima velikih reka gde voda teče veoma sporo. Svetlost i boja vode su vrlo bitni za metabolizam i primarnu produkciju vodenog ekosistema. Proces fotosinteze direktno je zavistan od količine i kvaliteta svetlosti. Boja rečne vode prvenstveno zavisi od stepena apsorpcije. U gornjem toku zbog dvostruke refleksije boja vode prvenstveno zavisi od boje podloge ili razvijene vegetacije. Veoma bistre planinske reke obično imaju plavu ili svetlozelenu boju. U donjim tokovima reka boja sve manje zavisi od boje podloge, već zavisi od količine suspendovanih čestica organske i neorganske prirode. Kretanje vode takođe je bitan ekološki faktor za tekuće vode. Hidrobionti se mogu zadržati u vodi čija brzina varira izmedju 1 do 3,5 m/s. Ako je brzina veća, nijedan hidrobiont ne može to podneti jer ga voda nosi, a ako je brzina manja taj tekući sistem mogu naseliti organizmi koji žive u stajaćim vodama. Konfiguracija korita, naročito u gornjim tokovima reka, dovodi do naimeničnog smenjivanja područja u kome je nagib terena veliki i gde voda teče brzo (brzaci) i taj deo reke se naziva lotičko područje, i područja sa malim nagibom gde je voda dublja i teče sporije – lentičko područje. Ova dva područja se ekološki jako razlikuju pa su i zajednice koje u njima žive različite. Od brzine protoka vode zavisi i tip dna reke (Tabela 3).

16
Tabela 3: Zavisnost tipa dna tekućica od brzine vode (Simić i Simić 2009)
Brzina vode Tip dna 3-20 cm/s Organski detritus, mulj 20-40 cm/s Sitan pesak 40-60 cm/s Grubi pesak, sitan šljunak 50-120 cm/s Šljunak do veličine šake
120-200 cm/s Veliko kamenje i kameni
blokovi Tipovi rečnih dna se mogu razlikovati i prema veličini čestica i to na sledeći način: 1. malozrnast tip: gline (peliti), mulj (seliti, aleuriti), pesak sa veličinom zrnaca od 0,1 do 1 mm. 2. tvrda dna: krupniji pesak, šljunak (oblutak), kamen oblutak, kameni blokovi 3. finozrnasti tip podloge određuje se u zavisnosti od sadržaja finih frakcija, a to su čestice manje od 0,01 mm: pesak, muljeviti pesak, peščani mulj, mulj, glinoviti mulj, detritus Hemijske karakteristike kopnenih voda U prirodi nema hemijski čiste vode, u njoj se uvek nalaze neke primese u suspendovanom ili rastvorenom stanju i samo u takvim vodama je moguć opstanak živih organizama. Hemijski sastav voda nije svuda isti, najbitnije razlike su u koncentraciji rastvorenih: 1. gasova 2. mineralnih materija 3. organiskih materija u obliku koloida, suspenzija i emulzija. Hemijski sastav vode zavisi od medjusobne interakcije vode i okolnih stena, zemljišta, atmosfere i od aktivnosti živih organizama. Rastvoreni gasovi u vodi Najvažniji gasovi rastvoreni u kopnenim vodama su kiseonik (O2) i ugljen diokid (CO2).

17
Kiseonik (O2) u vodene ekosistema najvećim delom dospeva difuzijom iz vazduha, dok manji deo nastaje fotosintezom vodenih biljaka. Koncentracija kiseonika u vazduhu je konstantna, dok njegova koncentracija u vodama zavisi od temperature vode, kretanja vode, prisustva organskih i neorganskih materija i životnih aktivnosti hidrobionata. Kiseonika ima više u brzim i hladnim voda, a i njegova koncentracija manje varira nego u sporotekućim vodama. Kiseonik se u vodama neprekidno troši na disanje organizama koji žive u vodi, ali i na razlaganje autohtone i alohtone organske materije. Proces razlaganja je brži na višim temperaturama, kiseonik se brže troši, tako da vode sa prosečno višom temperaturom imaju manje rastvorenog kiseonika. Dodatno, u toploj vodi se slabije rastvara kiseonik što dodatno utiče na smanjenje koncentracije kiseonika pri povećavanju temperature vode. Do manjka kiseonika može doći i tokom zime kada je površina vode zaledjena, tada led predstavlja barijeru za difuziju kiseonika iz atmosfere, a kiseonik u vodi se konstantno troši od strane hidrobionata. Zbog toga se prave otvori u ledu da bi se pospešila aeracija. Ugljen-dioksid (CO2) u vodu najčešće dospeva difuzijom iz vazduha, ali može nastati i procesima u vodi kao što su disanje organizama, radom bakterija i u procesu truljenja. Ugljen-dioksid u reakciji sa vodom stvara slabo postojanu ugljenu kiselinu. Ugljena kiselina reaguje sa krečnjakom i gradi rastvorljiv bikarbonat:
H2CO3 + CaCO3 ↔ Ca(HCO3)2
Ova reakcija je povratna, što znači da ako iz nekog razloga opadne koncentracija ugljen-dioksida u vodi, bikarbonat Ca(HCO3)2 se razlaže na slabo rastvorljiv krečnjak (CaCO3) i ugljenu kiselinu (H2CO3) koja se kasnije razlaže na vodu (H2O) i ugljen dioksid (CO2), tako da izmedju rastvorenog ugljen-dioksida (CO2) i bikarbonata Ca(HCO3)2
postoji odredjena ravnoteža u vodi. Ova reakcija reguliše jonsku reakciju prirodnih voda, odnosno
njenu pH vrednost.

18
Nakon ove reakcije uvek ostaje višak neutrošenih OH― jona, zbog čega su kopnene vode slabo bazne. Koncentracija ugljen-dioksida i kiseonika duž toka reke zavisi od ekoloških uslova datih lokaliteta. Reke koje protiču kroz krečnjačku podlogu imaju veliku koncentraciju ugljen-dioksida, a malu koncentraciju kiseonika u izvorišnom delu, pa kako teče reka koncentracija ugljen diokida se smanjuje, a koncentracija kiseonika se povećava zbog asimilacionih procesa, brzine i rasprskivanja vode. Mineralne materije u vodi U vodi se mineralne materija sreću kao makroelementi (Na, K, Ca, Mg) i mikroelementi (Fe, Cu, Co, Mn, Ni, Zn, J, F, Se). U kopnenim vodama registrovano je više od 70 elemenata. Joni Ca2+ i Mg2+ su stalno prisutni u vodi i od njihovog sadržaja zavisi ukupna tvrdoća vode. Ovi joni dospevaju u vodu iz stena bogatih krečnjacima. Kao najznačajnije mineralne materije za živi svet voda možemo izdvojiti Azot (N) i fosfor (P) koji su biogene soli i ulaze u sastav živih bića i njihov metabolizam. Mineralna jedinjanja azota (N) u vodi najčešće se javljaju u obliku amonijaka (NH3), nitrita (NO2-), nitrata (NO3- jon) i organskih materija azota nastala razlaganjem proteina i aminokiselina. Ako je fitoplankton veoma razvijen soli azota mogu potpuno da nestanu iz površinskog sloja vode. Na većim dubinama koncentracija azota je konstantna i znatno veća nego u površinskim slojevima. Mineralna jedinjenja fosfora (P) u vodi se najčešće javljaju u obliku fosfata. Fosfati se u vodama najčešće nalaze u malim količinama i obično se vezuju za druge molekule i grade nerastvorna jedinjenja. Soli fosfata su veoma važne za rast autotrofnih organizama. Fosfor se u prirodnim vodama najčešće nalazi u obliku soli fosfata: ortofosfata i polifosfata. Ortofosfat i polifosfat se formiraju kao produkt razlaganja organske materije. Čovek je u prirodu uveo različite vrste fosfatnih jedinjenja (djubriva, deterdženti, industrijske otpadne vode). Neke fosforne soli su nerastvorljive u vodi pa se kao čestice vezuju za podlogu (sediment, mulj), pa zato u vodi uvek ima manje fosfornih soli nego azotnih soli koje su uvek rastvorljive.

19
Količina biogenih soli u rekama je u stalnom porastu zbog upotrebe deterdženata sa fosfatima i intenzivnim razvojem poljoprivrede. Sve to dovodi do procesa antropogene eutrofizacije koje se manifestuje kao bujanje biomase algi i viših biljaka. Organske materije u vodi mogu biti različitog porekla, pa prema tome ih delimo na autohtone i alohtone. Autohtona organska materija je ona koja se stvara u samom vodenom ekosistemu od strane autotrofnih organizama ili raspadanjem heterotrofnih organizama. U nezagadjenim vodama osnovnu organsku materiju čini humusna jedinjenja nastala razlaganjem biljnih ostataka. Alohtona organska materija je ona koja se iz okolnih terestričnih ekosistema unosi u vodene ekosisteme. Unos alohtone materije može biti prirodnim putem pri obilnim kišnim padavinama ili pri otapanju snežnog pokrivača i veštačkim putem preko otpadnih voda iz domaćinstva i industrije. Pri razlaganju ovih materija troši se velika količina kiseonika, menja pH i povećava koncentracija soli. Ovakvi uslovi ne pogoduju većini organizama koje žive u vodi pa oni nestaju, a ostaju samo vrste koje su otporne na zagadjenje. U ovakvim uslovima javlja se mali broj vrsta sa velikim brojem individua, jer vrste koje su opstale ne podležu interspecijskoj kompeticiji za hranu i prostor. Biološka potrošnja kiseonika (BPK) je količina kiseonika koja je potrebna da se izvrši biološka oksidacija prisutnih, biološki razgradljivih, sastojaka vode. Standardna metoda definiše vreme inkubacije od 5 dana, čime se određuje tzv. BPK5. Stepen zagađenosti vode organskim jedinjenjima definisan je pored HPK i ovim parametrom (BPK5). Temperatura i vreme razgradnje utiču na veličinu BPK5, tj. sa povećanjem temperature raste i brzina potrošnje kiseonika (biohemijska oksidacija). Smatra se da je potrebno 5 dana kako bi se razgradio veći deo (70%) prisutnih organskih materija, i to na stalnoj temperaturi vode od 20°C, što se i uzima za jedinicu. Prvo se odredi količina rastvorenog kiseonika na početku inkubacionog perioda, odnosno prvog dana, a zatim se izmeri količina rastvorenog kiseonika u uzorku nakon 5 dana inkubacije na 20 ºC. Razlika između ove dve vrednosti količine rastvorenog kiseonika u uzorku predstavlja vrednost BPK5.

20
1.6. Cilj
Cilj ovog istraživanja je bio da uporedimo sposobnost familije hironomida i grupe EPT u razdvajanju lokaliteta različitog nivoa kvaliteta vode. Takođe testirano je koji faktori staništa u najznačajnijem stepenu utiču na kvalitativni i kvantitativni sastav zajednice hironomida, odnosno EPT.
2. Materijal i metode
2.1. Oblast istraživanja Geografska oblast istraživanja u ovom radu je sliv reke Nišave. Uzorkovano je sa 10 tačaka, uključujući reku Nišavu i njene
pritoke Visočicu, Temsku i Jermu, koje takođe pripadaju slivu reke Nišave. Lokaliteti su birani tako da budu razmatrani gornji tokovi i donji tokovi. Na samoj Nišavi određena su četiri lokalitet zbog veličine i dužine reke, a na ostalim pomenutim pritokama po dva.
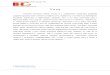
Na slici 1 prikazan je sliv Nišave sa podslivovima prekograničnih vodotoka Temske (Visočice), Gaberske i Jerme, dok su na tabeli 4 date geografske koordinate ispitivanih lokaliteta.
21
Slika 1: Sliv reke sa obeleženim lokalitetima Tabela 4: Geografske koordinate lokaliteta
Reka Lokalitet Geografske koordinate N E Nišava 1 43° 00’ 50’’ 22° 48’ 02’’ Jerma 2 43° 03’ 40’’ 22° 41’ 24’’ Jerma 3 42° 56’ 41’’ 22° 37’ 07’’ Visočica 4 43° 07’ 12’’ 22° 48’ 57’’ Temska 5 43° 19’ 21’’ 22° 39’ 36’’ Visočica 6 43° 17’ 48’’ 22° 36’ 42’’ Temska 7 43° 15’ 39’’ 22° 33’ 06’’ Nišava 8 43° 13’ 25’’ 22° 25’ 47’’ Nišava 9 43° 19’ 51’’ 22° 04’ 24’’ Nišava 10 43° 49’ 20’’ 21° 52’ 21’’

22
2.1.1. Nišava
Nišava protiče kroz Bugarsku i Srbiju, ima dužinu od 218 km i
najveća je pritoka Južne Morave. Nišava postaje od Ginske reke i Vrbnice koje se sastaju na 640m nadmorske visine u blizini sela Toden u Bugarskoj. Ginska reka je vodom bogatija i duža od Vrbnice, pa se vodi kao glavni izvorišni krak Nišave. Ginska reka izvire u Bugarskoj ispod vrha Kom na Staroj planini. Nišava ulazi u Srbiju 6 km uzvodno od Dimitrovgrada. Tu je široka samo 8-10 m i duboka 30-50 cm. Od Dimitrovgrada uglavnom teče prema zapadu, protiče još kroz Pirot, Belu Palanku, Nišku Banju i Niš, da bi se na kraju oko 10 km zapadno od Niša (kod sela Trupala) ulivala u Južnu Moravu. Nišava pripada Crnomorskom slivu i njen sliv prekriva teritoriju od 3950 km2, od čega 1237 km2 (ili 31%) pripada Bugarskoj, a 2713 km2 (ili 69%) pripada Srbiji. Prosečan protok vode je 36 m3/s.
Najveće pritoke Nišave su Temska sa Visočicom sa desne i Kutinska, Crvena reka, Koritnička, Rogoz, Jerma i Gaberska sa leve strane. Sve ove reke se ulivaju u Nišavu i zajedno čine najveću pritoku Južne Morave. Najveći vodostaj reke obično je u martu i aprilu nakon otapanja snežnog pokrivača, a najmanji vodostaj je u septembru i oktombru.
Geološka podloga kojom teče Nišava je dvojaka. Od ušća u Južnu Moravu pa uzvodno sve do Sićevačke klisure Nišava je široka nizijska reka na silikatnom supstratu sa granularnim sedimentima. Od Sićevačke klisure uzvodno Nišava je široka brdska reka na krečnjačkom supstratu sa granularnim sedimentima.
Na putu kroz našu zemlju Nišava teče kroz nekoliko kotlina i
klisura. One se smenjuju sledećim redosledom: Pirotska kotlina, Sopotski tesnac, Đurđevpoljska kotlina, tesnac kod Sv. Oca, Belopalanačka kotlina, Sićevačka klisura i Niška kotlina.
Pirotska kotlina je dugačka 14, a široka 3 – 5 km. Prolaskom Nišave kroz Pirot ona se dosta menja usled antropološkog dejstva, i to na dva načina. Prvi način je izgled same reke, jer je korito Nišave kroz Pirot kanalisano i obloženo kamenom u dužini od 2 – 2,5 km (slika 2). Oko korita su nasipi koji štite grad u slučaju visokog vodostaja. Drugo,

23
Pirot ima razvijenu industriju (Tigar), tako da se ovde prvi put javlja značajno antropološko zagadjenje.
Slika 2: Nišava u Pirotu Belopalanačko polje je duže (16 km), ali dva puta uže od
Pirotskog. Ovde je reka široka do 60 m i duboka do 2 m. U vreme visokih voda deo Belopalanačke kotline je često plavljen, iako su uz rečno korito podignuti nasipi.
Sićevačka klisura deo je rečne doline Nišave nastala intenzivnim usecanjem njenog vodenog toka u stensku masu. Dugačka je 17 km, a mestimično dublja od 350 m. Nalazi se 14 km istočno od Niša, izmedju sela Prosek i Dolac. Spaja Belopalanačku kotlinu i Nišku kotlinu.
Sićevačku klisuru možemo podeliti na četiri dela. Prvi deo se naziva Kusača, dug je 5,4 km i pruža se od naselja Prosek do betonske brane električne centrale na Nišavi kod sela Sićeva. Drugi deo Sićevačke klisure počinje od brane Hidrocentrale na Nišavi, u rejonu “Selišta” i prostire se uzvodno do “Banjice” u ataru naselja Ostrovica. Treći deo, u dužini od 2 km, poznat i kao Ostrovička kotlina, počinje od “Banjice” i proteže se do železničkog mosta na Nišavi, kod Ostravičkog kamenoloma u ataru naselja Ostrovica. Četvrti deo klisure, u dužini 5,5 km, pripada kanjonskom delu klisure, poznatom pod imenom Gradištanski kanjon. Prvi i drugi deo pripadaju klisurastom delu, treći kotlinskom i četvrti kanjonskom delu.

24
Zbog svojih geomorfoloških odlika i bogatstva flore i faune, Sićevačka klisura je proglašena za park prirode. U njoj možemo naći endemične i reliktne vrste.
Zbog velikog hidroenergetskog potencijala koji stvara Nišava prolaskom kroz usku Sićevačku klisuru izgrađene su dve male hidroelektrane: Sićevo i Ostrovica.
Slika 3: Kanjonski deo Sićevačke klisure Niška kotlina predstavlja završni deo toka Nišave, ona je
veoma prostrana (40 km dugačka i 23 km široka i ima površinu od 630 km2). Na jugu počinje od Kurvingradskog suženja, završava se Mezgrajskim suženjem na severu. Ovde reka ima karakteristike ravničarskog toka. Rečno korito u Nišu je podzidano kamenom, nizvodno od grada su izgradjeni nasipi koji prestavljaju zaštitu od poplava u mesecima sa visokim vodostajem. Treba istaći da ovde Nišava trpi značajno antropološko zagadjenje, jer je Niš najveći grad kroz koji protiče Nišava. To se veoma lako može dokazati biološkim (izmenjen sastav zajednica u odnosu na gornji tok) i fizičko-hemijskim analizama (merenje kvaliteta vode).

25
Slika 4: Niška kotlina
2.1.2. Temska sa Visočicom
Temska sa Visočicom je najveća desna pritoka Nišave. Ukupna dužina je 86 km, od čega 69 km protiče kroz Srbiju. Površina njenog sliva obuhvata 820 km2 (u Srbiji 715 km2). Visočica nastaje od Brlske i Srednje reke koje izviru na planini Kom u Bugarskoj. Od ušća Toplodolske reke u Visočicu kod sela Temska, Visočica dobija naziv Temska (ili Temštica) sve do ušća reke u Nišavu.
Visočica u Srbiju ulazi nedaleko od sela Donji Krivodol, a u Nišavu se uliva kod sela Staničenje, 16 km nizvodno od Pirota, dajući joj prosečno 10,5 m3/s vode. U toku jedne godine na sliv reke Visočice se izluči 688 miliona m3 vodenog taloga.

26
Slika 5: Visočica Kod sela Zavoj, u februaru 1963. godine skliznulo je oko 2
miliona kubnih metara zemljišta i kamena u dolini Visočice i obrazovana je prirodna brana visoka 36 m. Ta brana je predstavljala prepreku za vodeni tok i stvorena je akumulacija vode od koje je kasnije nastalo jezero Zavoj. Jezera nastala na ovakav način nazivaju se urniska. Voda iz ovog jezera se sprovode podzemnim tunelom dugačkim 9 km do hidroelektrane Pirot koja ima kapacitet od 80 MW.
Slika 6: Zavojsko jezero
Sliv Temske je asimetričan. S desne strane toka Visočica prima
više pritoka. Desne pritoke su Vodenička reka, Rosomačka reka, Dojkinačka reka, Gostuška reka, Toplodolska reka, Rudinska i Klajća.

27
Veća leva pritoka je Pokrovštica. Naime, padine Stare planine na kojima je razvijen osnovni (desni) deo sliva sastavljene su od permskih peščara i trijaskih krečnjaka, a levi deo sliva na kom su zastupljeni kredni i jurski krečnjaci je više karstifikovan. Zbog ogolelosti sliva i geološkog sastava terena, sliv Visočice je podložan svim vidovima erozije.
Geološka podloga kojim teče reka Temska sa Visočicom je heterogena. Temska je mala brdska reka na silikatnoj podlozi. Visočica je od 0 do 17,4 km mala, srednjeplaninska reka na silikatnoj podlozi, a od 17,4 do 43,4 km mala, srednjeplaninska reka na karbonatnoj podlozi.
2.1.3. Jerma Reka Jerma je najveća leva pritoka Nišave. Duga je 74 km, a površina sliva je 799,3 km2. Nastaje od planinskih potoka sa severoistočne padine planine Strešer na krajnjem Jugoistoku Srbije. Jerma nastaje od Vučje i Grubinske reke istočno od Vlasinskog jezera. Nedugo nakon svog formiranja kao jak planinski potok prelazi na teritoriji Republike Bugarske kod sela Strezimirovci i svoj put nastavlja preko planinske visoravni poznate kao Trnsko polje.
Jerma na teritoriju Srbije prelazi kod sela Petačinci i tu već predstavlja jaču planinsku reku. Jerma teče pretežno kroz kanjonske doline i njen tok je pristupačan. Na svom putu Jerma prolazi kroz Odorovačko ždrelo koja je jedna od najživopisnijih klisura u Srbiji. Korito je mestimično kamenito, amestimično usečeno u rečne sediment. Prosečna dubina je oko 1m, a njen pad je tokom celog toka veoma ujednačen pa nema nekih velikih konfiguracijskih lomova. Jerma se uliva u Nišavu 1 km nizvodno od sela Gradišta unoseći 5 m3/s vode u Nišavu.
Zbog svega navedenog možemo zaključiti da je ihtiofauna salmonidnog karaktera.
Od važnijih pritoka treba istaći Jablanicu, Liškovicu, Poganovsku reku. Nijedna od njih nema površinu sliva veću od 100 km2.
Geološka podloga Jerme: Jerma je od 0 do 9,5 km mala brdska reka na karbonatnoj podlozi, od 9,5 do 28,5 km je mala srednjeplaninska reka na silikatnoj podlozi, a od 53,6 do 59,6 km je

28
mala visokoplaninska reka na silikatnoj podlozi. Treba napomenuti da je ovde uzet u obzir samo tok Jerme kroz Srbiju.
Slika 7: Jerma
2.2. Hidrološke karakteristike pritoka Nišave
Tabela 5: Morfološke karateristike Visočice, Temske i Jerme
Vodotok Profil F
(km2) L
(km)
Zmin
(mnm)
Zmax
(mnm)
Srednji pad toka (%)
Temska sa Visočicom
Ušće u Nišavu
827,5 59,8 338 835 0,83
Jerma Ušće u Nišavu
799,3 59,6 412 870 0,77

29
Prosečni proticaji Tabela 6: Prosečni srednje mesečni i godišnji proticaji na Visočici, Temskoj i Jermi u period 1946-1991. godine (Aneks 7)
Profil
Qsr.mes (m3/s) Qsr.go
d
(m3/s)
I II III IV V VI VII
VIII
IX X XI XII
Staničenje 8,24
12,03
17,56
22,55
18,63
11,33
6,20
3,02
3,18
4,03
5,59
8,08
10,04
Izatovac 0,96 1,49 2,11 2,37 2,03 1,26 0,6
2 0,24
0,25
0,36
0,58
0,95 1,10
Visočka Ržana 4,75 6,80 10,2
5 13,6
0 11,1
8 6,89 3,45
1,65
1,65
2,20
2,96
4,53 5,83
Trnski i Odorovci 2,93 4,67 6,96 6,81 5,49 4,22 2,4
6 1,42
1,32
1,58
1,96
2,60 3,53
Sukovo 4,69 7,00 10,4
4 10,3
5 8,39 6,58 3,88
2,20
2,09
2,42
3,03
4,18 5,44
Lokacija Staničenje se nalazi na Temskoj, Izatovac i Visočka
Ržana se nalaze na Visočici, a Trnski i odorovci i Sukovo se nalaze na Jermi
Iz tabele možemo zaključiti da su najveći proticaji beleže u aprilu, tj. u period kada se otapaju velike količine snežnog pokrivača sa obližnje Stare planine. Najmanji proticaji se beleže u avgustu i septembru, tj. periodu posle letnjih suša.
2.3. Metode uzorkovanja
Istraživanje makrozoobentosa Nišavskog sliva sprovedeno je 2011. godine tokom svih sezona . Prvi put material je sakupljan u januaru (zima), drugi put u martu (proleće), treći put u avgustu (leto) i četvrti put u novembru (jesen).
Uzorkovano je na deset reprezentativno odabranih lokaliteta na reci Nišavi i njenim pritokama sa ciljem pokrivanja što većeg diverziteta kako u biotičkom tako i u abiotičkom smislu (Slika 1). Birani su lokaliteti sa različitom geološkom podlogom, nadmorskom visinom, nagibom terena, brzinom protoka vode, tipom dna, mikroklimatskim uslovima itd.

30
Uzorkovanje je sprovedeno korišćenjem Surberove mreže (Slika 8) sa dimenzijama rama 25x25 cm, što znači da je zahvatna površina mrežice 0,625 m2. Sa svakog lokaliteta su uzeta po tri uzoraka. Na terenu na licu mesta su uzeti podaci za sledeće abiotičke faktore: pH, elektroprovodljivost, temperatura, saturacija kiseonikom, koncentracija kiseonika i mutnoća.
Slika 8: Izgled Surberove mreže
Uzorkovani material je iz Surberove mreže prebačen direktno u plastične kese zapremine 3l u kojima se ubacuju papirići sa nazivom lokaliteta i datumom koji se ispisuju isključivo grafitnom olovkom. Odmah nakon uzorkovanja materijal je obrađen na Prirodno – matematičkom fakultetu u Nišu. Uzorak iz svake kese je ispran pod mlazom vode kroz sistem sita. Uzorkovani material prvo prolazi kroz sito čija su okca promera 2,25mm, a zatim odmah i kroz drugo sito koje je sitnije od prvog, sa promerom okca od 0,5mm. Nakon toga izolovani material se prenosi u providne, plitke posude odakle se pincetama i iglama odvaja od ostalih nečistoća, a same jedinke se prenose u male staklene boce ispunjene 70%-tnim alkoholom gde se čuvaju do determinacije. Na svakoj flašici stoji etiketa sa lokalitetom i datumom uzorkovanja.
Sakupljeni materijal determinisan je pomoću binokularne lupe i mikroskopa takođe na Prirodno matematičkom fakultetu u Nišu. Za potrebe ovog rada organizmi su determinisani do nivoa roda.

31
2.4. Analiza podataka Nemetričko multidimenzionalno skaliranje (Nonmetric
multidimensional scaling, nMDS) je korišćeno kako bi se grupisali lokaliteti sa sličnim kvalitativnim i kvantitativnim sastavom ispitivanih grupa. U ekološkim studijama, koje analiziraju prostornu i vremensku dinamiku zajednice makroinvertebrata, koriste se ordinacione metode kao sto su PCA, CCA i Cluster analiza. U novije vreme za vizualizaciju prostornog i vremenskog obrasca u primeni je nMDS zbog svojih prednosti u odnosu na pomenute ordinacione metode. Rezultat nMDS-a prikazuje set tačaka (lokaliteta) u n-dimenzionom prostoru raspoređenih tako da distanca među njima je direktno proporcionalna stepenu sličnosti. Parametar „nonmetric stress“ predstavlja stepen neuspešnosti u takoj vizualizaciji. nMDS moze predstaviti rezultate u opsegu od 2 do 5 dimenzija gde sa povećavanje broja dimenzija raste i vrednost stresa. Vrednost stresa ne bi trebalo da pređe vrednost 0.25, da bi dobijeni rezultati bili pouzdani. Sa ciljem utvrđivanja kombinacije abiotičkih faktora koji u najvećem stepenu utiču na kvalitativno-kvantitativni sastav zajednice, korišćena je multipna regresiona analiza BEST. Ova analiza formira najbolji model koji uključuje kombinaciju nezavisnih promenljivih (abiotički faktori) koja u najvećem procentu objašnjava varijabilnost zavisne promenljive (zajednice makroinvertebrata). Sve analize u urađene u statističkom paketu PRIMER v6.
3. Rezultati
3.1 Taksonomski sastav zajednice EPT i Chironomidae
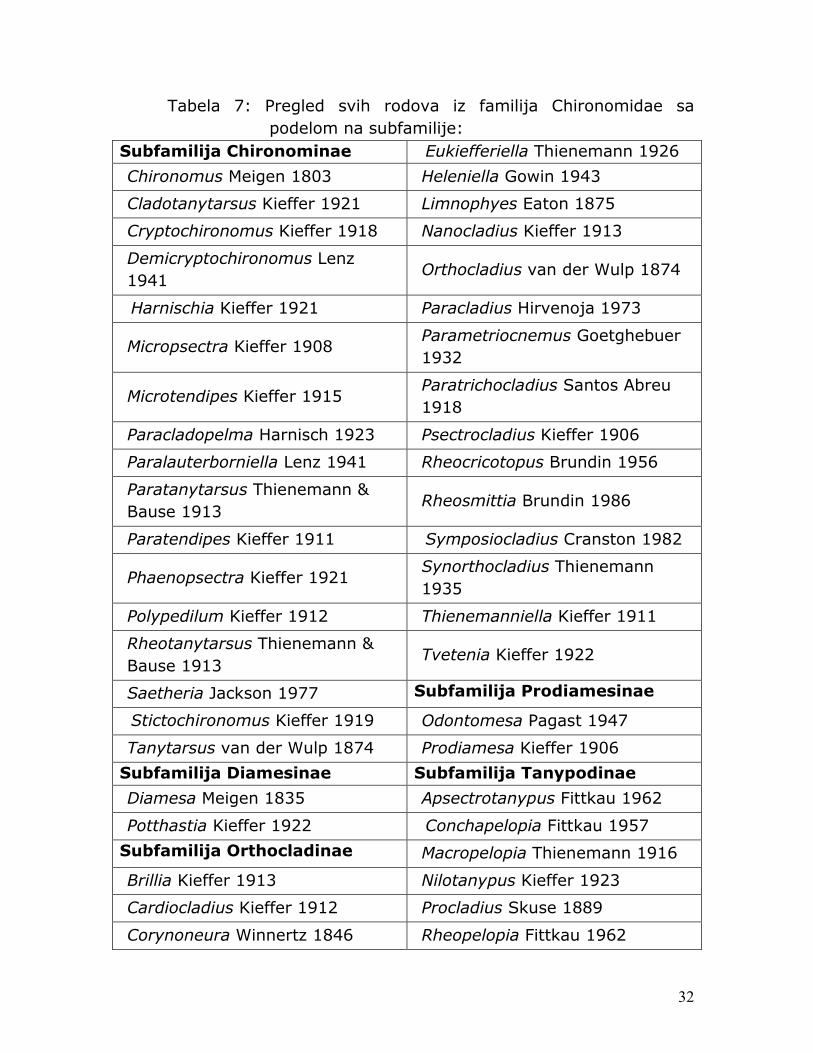
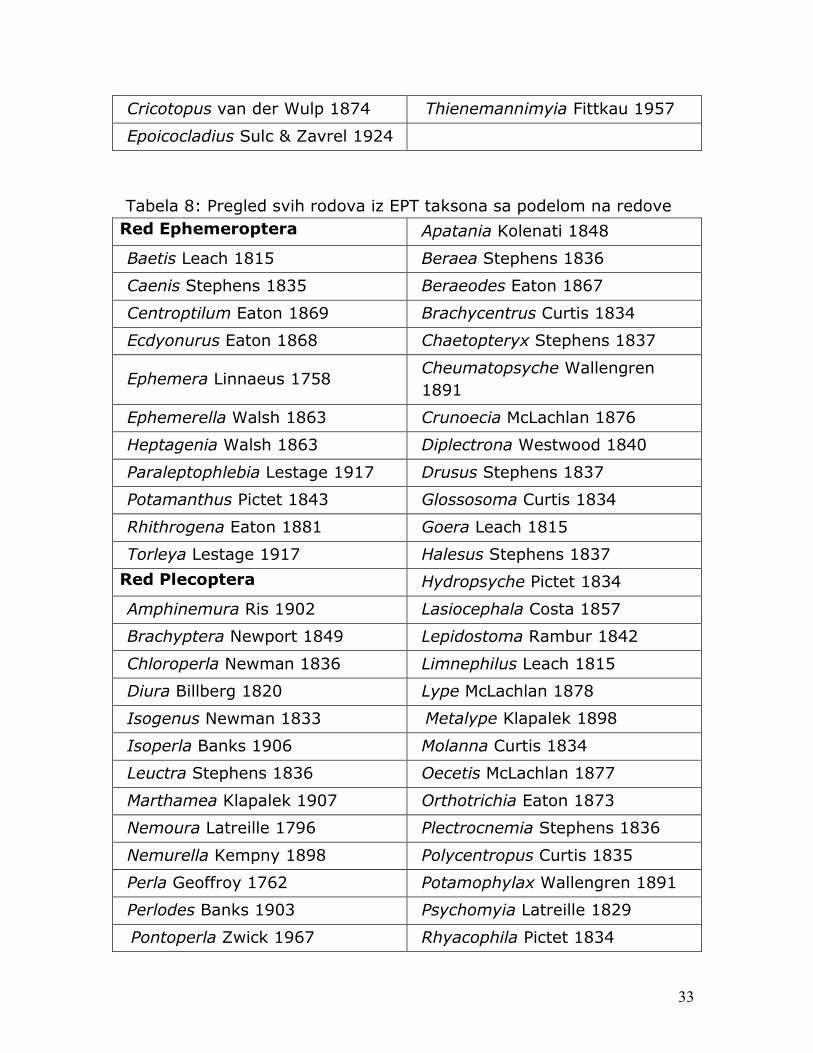
Tokom istraživanja od ukupno 1901 jedinke hironomida i 2007 jedinki grupe EPT, determinisano je 48 taksona iz familije Chironomidae (videti tabelu 6) i 59 taksona iz grupe EPT (videti tabelu 7).
32
Tabela 7: Pregled svih rodova iz familija Chironomidae sa podelom na subfamilije:
Subfamilija Chironominae Eukiefferiella Thienemann 1926
Chironomus Meigen 1803 Heleniella Gowin 1943
Cladotanytarsus Kieffer 1921 Limnophyes Eaton 1875
Cryptochironomus Kieffer 1918 Nanocladius Kieffer 1913
Demicryptochironomus Lenz 1941
Orthocladius van der Wulp 1874
Harnischia Kieffer 1921 Paracladius Hirvenoja 1973
Micropsectra Kieffer 1908 Parametriocnemus Goetghebuer 1932
Microtendipes Kieffer 1915 Paratrichocladius Santos Abreu 1918
Paracladopelma Harnisch 1923 Psectrocladius Kieffer 1906
Paralauterborniella Lenz 1941 Rheocricotopus Brundin 1956
Paratanytarsus Thienemann & Bause 1913
Rheosmittia Brundin 1986
Paratendipes Kieffer 1911 Symposiocladius Cranston 1982
Phaenopsectra Kieffer 1921 Synorthocladius Thienemann 1935
Polypedilum Kieffer 1912 Thienemanniella Kieffer 1911
Rheotanytarsus Thienemann & Bause 1913
Tvetenia Kieffer 1922
Saetheria Jackson 1977 Subfamilija Prodiamesinae
Stictochironomus Kieffer 1919 Odontomesa Pagast 1947
Tanytarsus van der Wulp 1874 Prodiamesa Kieffer 1906
Subfamilija Diamesinae Subfamilija Tanypodinae
Diamesa Meigen 1835 Apsectrotanypus Fittkau 1962
Potthastia Kieffer 1922 Conchapelopia Fittkau 1957
Subfamilija Orthocladinae Macropelopia Thienemann 1916
Brillia Kieffer 1913 Nilotanypus Kieffer 1923
Cardiocladius Kieffer 1912 Procladius Skuse 1889
Corynoneura Winnertz 1846 Rheopelopia Fittkau 1962
33
Cricotopus van der Wulp 1874 Thienemannimyia Fittkau 1957
Epoicocladius Sulc & Zavrel 1924
Tabela 8: Pregled svih rodova iz EPT taksona sa podelom na redove Red Ephemeroptera Apatania Kolenati 1848
Baetis Leach 1815 Beraea Stephens 1836
Caenis Stephens 1835 Beraeodes Eaton 1867
Centroptilum Eaton 1869 Brachycentrus Curtis 1834
Ecdyonurus Eaton 1868 Chaetopteryx Stephens 1837
Ephemera Linnaeus 1758 Cheumatopsyche Wallengren 1891
Ephemerella Walsh 1863 Crunoecia McLachlan 1876
Heptagenia Walsh 1863 Diplectrona Westwood 1840
Paraleptophlebia Lestage 1917 Drusus Stephens 1837
Potamanthus Pictet 1843 Glossosoma Curtis 1834
Rhithrogena Eaton 1881 Goera Leach 1815
Torleya Lestage 1917 Halesus Stephens 1837
Red Plecoptera Hydropsyche Pictet 1834
Amphinemura Ris 1902 Lasiocephala Costa 1857
Brachyptera Newport 1849 Lepidostoma Rambur 1842
Chloroperla Newman 1836 Limnephilus Leach 1815
Diura Billberg 1820 Lype McLachlan 1878
Isogenus Newman 1833 Metalype Klapalek 1898
Isoperla Banks 1906 Molanna Curtis 1834
Leuctra Stephens 1836 Oecetis McLachlan 1877
Marthamea Klapalek 1907 Orthotrichia Eaton 1873
Nemoura Latreille 1796 Plectrocnemia Stephens 1836
Nemurella Kempny 1898 Polycentropus Curtis 1835
Perla Geoffroy 1762 Potamophylax Wallengren 1891
Perlodes Banks 1903 Psychomyia Latreille 1829
Pontoperla Zwick 1967 Rhyacophila Pictet 1834

34
Protonemura Kempny 1898 Sericostoma Latreille 1825
Taeniopteryx Pictet 1841 Silo Curtis 1833
Red Trichoptera Stenophylax Kolenati 1848
Agapetus Curtis 1834 Tinodes Leach 1815
Allogamus Schmid 1955
U okviru grupe Chironomidae najveću brojnost imaju jedinke iz rodova Orthocladius sa 17,1%, zatim slede Parametriocnemus sa 9,2% i Paratendipes sa 8,9% učešća od ukupnog broja jedinki. Najfrekventniji rodovi su Chonchapelopia, Parametriocnemus i Polypedilum koji se pojavljuju na 90% lokaliteta. Najmanja brojnost je zabeležena kod rodova Psectrocladius i Phaenospectra sa 0,01% i Cardiocladus i Saetheria sa 0,04%. Taksoni sa najmanjom frekventnošću su Apsectrotanytarsus, Cardiocladus, Paratanytarsus, Phaenospectra, Psectrocladius, Rheosmittia i Saetheria koji se pojavljuju samo na jednom lokalitetu.
Ako se pogleda procentualno učešće subfamilija hironomida dobijamo sledeće rezultate: Orthocladiinae 43,8%, Chironominae 33,3%, Tanypodinae 14,6%, Diamesinae 4,1% i Prodiamesinae 4,1%. Lokalitet sa najvećim bogatstvom taksona je lokalitet 1 sa 29 različitih taksona, slede ga lokaliteti 5 i 9 sa 28 različitih taksona. Lokaliteti sa najmanjim brojem taksona su lokalitet 10 sa 12 različitih taksona i lokalitet 8 sa 14 različitih takosna. Lokalitet sa najvećim brojem jedinki je lokalitet 6 sa 3574 jedinki/m2, slede ga lokalitet 5 sa 1142 jedinki/m2 i lokalitet 2 sa 1120 jedinki/m2. Najmanji broj jedinki je zabeležen na lokalitetima 10 sa 289 jedinki/m2 i 7 sa 305 jedinki/m2. Na slici 9 je predstavljeno udeo svakog lokaliteta u ukupnom broju hironomida.
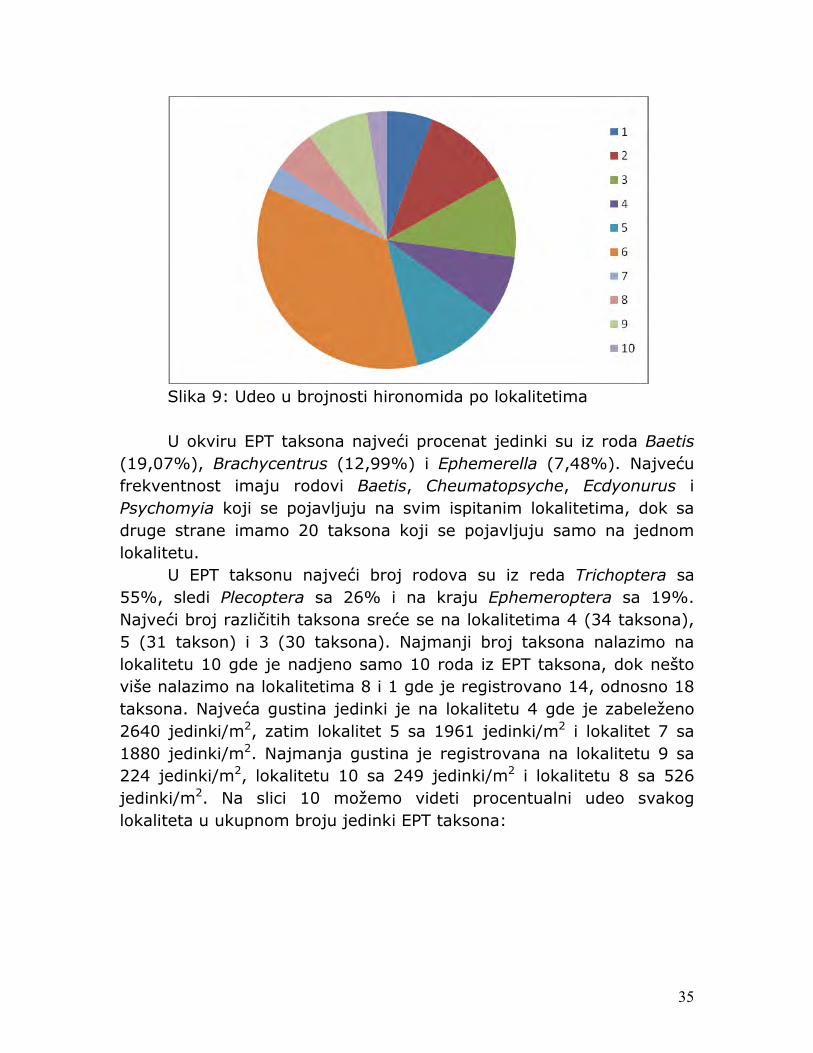
35
Slika 9: Udeo u brojnosti hironomida po lokalitetima U okviru EPT taksona najveći procenat jedinki su iz roda Baetis
(19,07%), Brachycentrus (12,99%) i Ephemerella (7,48%). Najveću frekventnost imaju rodovi Baetis, Cheumatopsyche, Ecdyonurus i Psychomyia koji se pojavljuju na svim ispitanim lokalitetima, dok sa druge strane imamo 20 taksona koji se pojavljuju samo na jednom lokalitetu. U EPT taksonu najveći broj rodova su iz reda Trichoptera sa 55%, sledi Plecoptera sa 26% i na kraju Ephemeroptera sa 19%. Najveći broj različitih taksona sreće se na lokalitetima 4 (34 taksona), 5 (31 takson) i 3 (30 taksona). Najmanji broj taksona nalazimo na lokalitetu 10 gde je nadjeno samo 10 roda iz EPT taksona, dok nešto više nalazimo na lokalitetima 8 i 1 gde je registrovano 14, odnosno 18 taksona. Najveća gustina jedinki je na lokalitetu 4 gde je zabeleženo 2640 jedinki/m2, zatim lokalitet 5 sa 1961 jedinki/m2 i lokalitet 7 sa 1880 jedinki/m2. Najmanja gustina je registrovana na lokalitetu 9 sa 224 jedinki/m2, lokalitetu 10 sa 249 jedinki/m2 i lokalitetu 8 sa 526 jedinki/m2. Na slici 10 možemo videti procentualni udeo svakog lokaliteta u ukupnom broju jedinki EPT taksona:
36
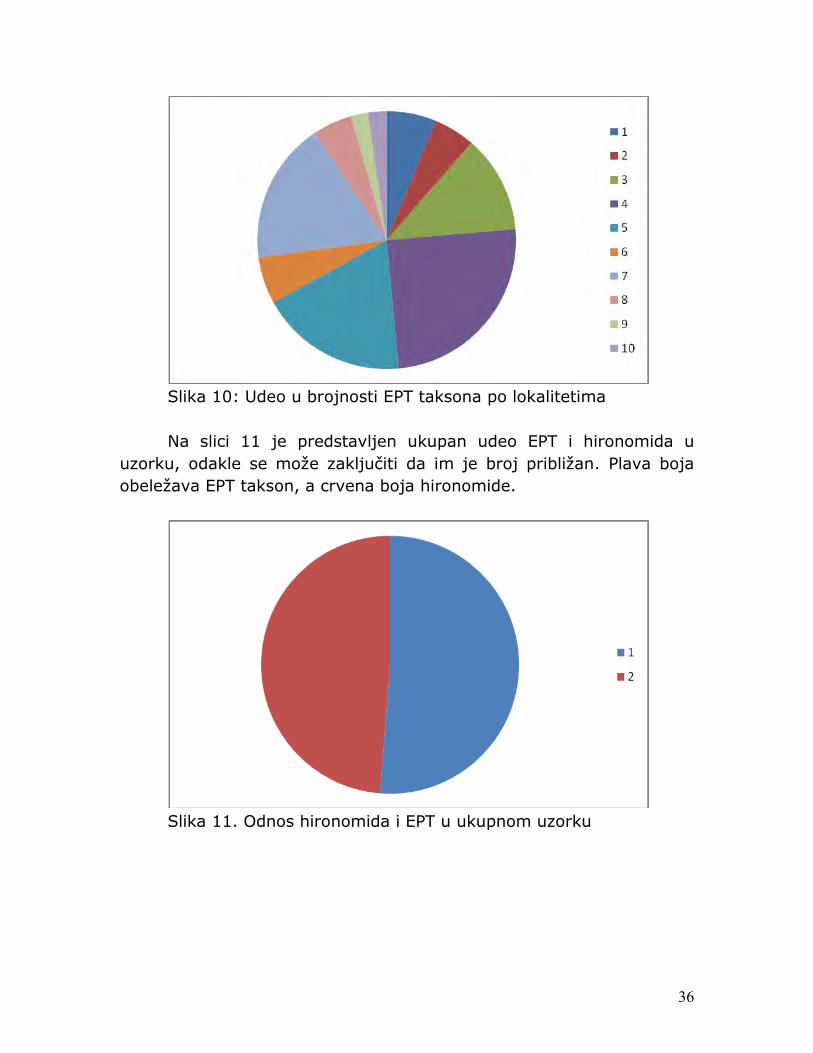
Slika 10: Udeo u brojnosti EPT taksona po lokalitetima Na slici 11 je predstavljen ukupan udeo EPT i hironomida u uzorku, odakle se može zaključiti da im je broj približan. Plava boja obeležava EPT takson, a crvena boja hironomide.
Slika 11. Odnos hironomida i EPT u ukupnom uzorku

37
3.2 Prostorna distribucija zajednica EPT i Chironomidae
Rezultati nMDS analize pokazali su da se na osnovu kvalitativnog
i kvantitativnog sastava EPT taksona mogu izdvojiti 2 grupe lokaliteta sa sličnošću od 40% (Slika 10). U prvoj grupi su se izdvojili lokaliteti koji pripadaju srednjem i donjem toku reke Nišave (8, 9, 10). Nasuprot tome, drugoj grupi pripadaju lokaliteti (1, 2, 3, 4, 5, 6, 7), uglavnom smešteni na pritokama Nišave. U pogledu zajednice hironomida, klasifikaciona analiza je takodje izdvojila dve grupe sličnog sastava lokaliteta (slika 9). U okviru prve grupe izdvojili su se lokaliteti na Nišavi (8 i 10), dok drugoj grupi najvećim delom pripadaju lokaliteti na pritokama. nMDS analiza je korišćenjem sredinskih varijabli formirala tri grupe lokaliteta (slika 11). Prvoj grupi pripadaju lokaliteti sa srednjeg i donjeg toka Nišave (8, 9 i 10). Ova staništa su okarakterisana malim nadmorskim visinama, višim vrednostima temperature, širine i dubinom korita. Kod ovih lokaliteta je takodje zabeleženo prisustvo veće koncentracije nitrata, fosfata, amonijaka i visoke vrednosti BPK5. Druga grupa je predstavljena lokalitetima na pritokama. Staništa koja pripadaju ovoj grupi se nalaze na višim nadmorskim visinama što je praćeno i niskim vrednostima temperature, dubine i širine korita kao i niskim mineralnim sastavom (nitrati, fosfati i amonijak). Trećoj grupi pripada gornji tok Nišave (1) i donji tok Jerme (2).
Sa ciljem pronalaženja grupe abiotičkih faktora koji na najbolji način objašnjavaju multidimenzionalnu strukturu zajednice, BEST analiza je pokazala da na obrazac EPT grupe u najvećem stepenu utiče nadmorska visina i koncetracija nitrata sa statistički značajnim koeficijentom Spearman-ove korelacije (BioEnv, r = 0.764, p = 0.01). Istom analizom je utvrđeno da je obrazac zajednice hironomida pod uticajem tri najznačajnija faktora (pH, BPK5 i nadmorska visina) (BioEnv, r = 0.699, p = 0.01). BioEnv je potvrdio koncept rečnog kontinuuma i izdvojio nadmorsku visinu kao jedan od najbitnijih parametara koji značajno utiče na strukturu zajednice hironomida.

38
Slika 12: Vizualizacija grupisanja lokaliteta prema sličnosti u kvalitativnom i kvantitativnom sastavu zajednice hironomida
Slika 13: Vizualizacija grupisanja lokaliteta prema sličnosti u kvalitativnom i kvantitativnom sastavu EPT grupe

39
Slika 14: Vizualizacija grupisanja lokaliteta prema sličnostima u kvalitetu sredinskih parametara
4. Diskusija
Korišćenjem grupe EPT, kao input matrice za nMDS analizu, formirale su se dve grupe lokaliteta u 2-d prostoru. Prva grupa obuhvata skoro sve lokalitete na glavnom toku reke Nišave osim lokaliteta 1 koji pripada gornjem delu reke, smeštenom iznad Dimitrovgrada što ga izdvaja van uticaja antropogenog stresa. Sličan rezultat je prezentovao Savić (2010), gde se lokalitet smešten iznad Dimitrovgrada izdvojio od ostalih lokaliteta na Nišavi. Ovo izdvajanje je bilo bazirano na grupi Ephemeroptera. Nasuprot tome, lokaliteti prve grupe su većinom pozicionirani neposredno ispod velikih urbanih naselja, što doprinosi njihovoj izloženosti tačkastim izvorima zagadjenja (otpadne vode industrijskog i komunalnog porekla). S obzirom da je udaljenost u nMDS 2-d prostoru proporcionalna realnoj razlici prikazanih lokaliteta u pogledu sastava ispitivanih grupa, na slici 13 se jasno može uočiti horizontalni gradijent kvaliteta vode koji sa leve ka desnoj strani postepeno raste. Ako se sagledaju fizičko-hemijski parametri (slika 14) može se uočiti sličan obrazac gde su lokaliteti takodje rasporedjeni duž horizontalnog gradijenta kvaliteta

40
vode. U tom smislu, na slici 13 lokalitet 10 zauzima dijametralno suprotan položaj od lokaliteta 4 i 5. Ovo je očekivano, s obzirom na činjenicu da se lokalitet 10 nalazi ispod komunalnog izliva kanalizacije Grada Niša, dok su lokaliteti 4 i 5 daleko od ikakvog antropogenog uticaja pa ih možemo smatrati referentnim lokalitetima. Na taj način je ponovo potvrdjeno da EPT grupa poseduje sposobnost da jasno izdvoji delove reka različitog stepena zagađenosti. Sve pogodosnoti prilikom primene ove grupe u monitoring sistemima, navedene u poglavlju Uvod, upućuju na korišćenje EPT grupe u brzim metodama procene kvaliteta vode (AQEM, RBP). Ovim pristupom se na brz, jeftin i pouzdan načun može doći do podataka o stepenu zagadjenja vodenog staništa.
Sa druge strane, na osnovu kvalitativno-kvantitativnog sastava zajednice hironomida, nMDS analiza je jasno razdvojila lokalitete u dve grupe. Sa jedne strane 2-d prostora su se izdvojili lokaliteti 8 i 10, slično kao u prethodnoj analizi sa EPT grupom, koji su izloženi snažnim antropogenim uticajem jer se nalaze neposredno ispod velikih gradova (Pirot i Niš). Na drugom kraju 2-d prosora su grupisani ostali lokaliteti iz kojih se izdvaja lokalitet 6. Ostali lokaliteti druge grupe su veoma slični na osnovu zajednice hironomida. Izdvajanje lokaliteta 6 od ostatka druge grupe se moze objasniti činjenicom da se radi o lokalitetu koji je neposredno ispod brane akumulacije Zavoj. Zbog toga su uslovi sredine drastično izmenjeni, pa je i samim tim struktura zajednice hiromonida drugačija (Milošević, 2012). Rezultati klasterovanja se ne mogu pripisati samo antropogenom uticaju već i velikoj prirodnoj varijabilnosti koja je karakteristična za zajednicu familije Chironomidae. Mnoge studije su potvrdile značajnu varijabilnost hironomida koja nekada i premašuje varijabilnost izazvanu antropogenim stresom. Punti (2009) je prikazao klasifikaciju lotičkih sistema u Mediteranskoj oblasti na osnovu zajednice hironomida gde ova grupa razdvaja lokalitete u tri, jasno definisane grupe (gornji, srednji i donji tok). Milošević (2013) je potrvdio ovakav rezultat prikazavši sličan distribucioni obrazac u rekama umerenog pojasa (ekoregion 5). S obzirom da je prvi korak u primeni neke grupe kao indikatora u biomonitoring programima otkrivanje i definisanje prostorno-vremenskog obrasca, studije kao što je ova su jako korisne. Ukoliko je prirodna varijabilnost nepoznata, registrovane promene u sastavu zajednice mogu dati lažni signal o kvalitetu staništa. Prema

41
tome jako je bitno a priori odrediti prirodan obrazac grupe, pravljenjem modela koji će pokriti svu prirodnu varijabilnost, pa tek onda definisati metričke osobine i implementirati ih u određeni bioassesment program.
Uticaj sredinskih varijabli na makroinvertebrate je odavno potrvdjen i poznat. Istraživanjem svih grupa makroinvertebrata došlo se do zaključka da je nadmorska visina najvažniji parametar koji uslovljava pravilnu promenu ostalih sredinskih parametara (na primer, brzina, dubina širina korita temperatura itd). Vannote et al. (1980) je svojim konceptom rečnog kontinuuma predstavio kako se duž longitudinalnog gradijenta menjaju sa odredjenom pravilnošću svi sredinski parametri, izdvojivši nadmorsku visinu kao glavnog pokretača ovog gradijenta. BioEnv analizom je potvrđeno da je nadmorska visina jedan od najbitnijih parametara i u ovoj studiji, utičući bitno na sastav i strukturu zajednice hironomida i grupe EPT. Na primer, Carter et al. (1996) i Marchant et al. (1995) su potvrdili da je nadmorska visina najvažnji faktor u struktuiranju zajednice Ephemeroptera, Trichoptera i Plecoptera. Na isti način Lencioi i Rossaro (2005) svojom studijom pokazuju da je zajednica familije Chironomidae snažno izložena gradijentu nadmorske visine koga prate pravilne promene i ostalih parametara (brzina vode, širina i dubina korita, temperatura). Ovakave činjenice jednim delom objašnjavaju podelu lokaliteta na dve grupe prema zajednicama hironomida i EPT gde su se sa jedne strane izdvojili donji tok a sa druge gornji tokovi reka.
Pored ovog parametara i BPK5 značajno utiče na promenu zajednice hironomida. S obzirom da je BPK5 indikator organskog zagadjenja, ovakav rezultat je očekivan (Wilson, 1992, Milošević, 2012). Chironomidae su poznati indikatori organskog zagadjenja i njihova primena je danas učestala u nekim indeksima bioprocene koje su u primeni u AQEM sistemu (AQEM 2002). Jedina mana ove grupe je komplikovan proces determinacije koji je vremenski jako zahtevan. To je osnovni razlog zašto se često hironomide isključuju iz rutinskih skrining sistema procene kvaliteta vode (Raunio, 2011). Medjutim, i ovaj problem se prevazilazi uvodjenjem novog pristupa u konstruisanju ključeva za determinaciju (Vallenduuk i Moller Pillot 2007, Orendt i Spies 2012) koji osim spiska vrsta sadrže i autekološke

42
podatke koji značajno olakšavaju proces identifikacije (Milosević, 2013).
Za distribuciju predstavnika EPT grupe koncentracija nitrata je predstavljala jedan do značajnih parametara koji utiče na strukturu zajednice ove grupe. Ovaj sredinski parametar takođe ukazuje na količinu organskog zagadjenja vode. Značaj ovog parametra u distribuciji vrsta EPT grupe se objašnjava njihovom osetljivošću na organsko opterećenje (Wenn C.L. 2008). Upravo zahvaljujući ovoj osobini, da se EPT taksoni smatraju najmanje tolerantnim na organsko zagadjenje, EPT grupa se koristi u većini multimetrijskih sistema za procenu kvaliteta vode.
5. Zaključak Ova studija je pokazala da se na osnovu zajednice hironomida i
EPT grupe, sa taksonomskom rezolucijom do nivoa roda, mogu formirati grupe sa lokalitetima različitog kvaliteta i stepena antropogenog uticaja. Razlika u sposobnosti razdvajanja lokaliteta na osnovu zajednica se ogleda u većoj osetljivosti EPT grupe u odnosu na familiju hironomida prikazujući veću nijansiranost u razdvajanju lokaliteta. BioEnv analiza je izdvojila nadmorsku visinu kao jedan od najbitnijih faktora sredine koji značajno utiče na struktuiranost kako zajednice hironomida tako i grupe EPT. Takodje su izdvojeni i indikatori organskog zagadjenja (BPK5 i koncentracija nitrata) kao faktori koji značajno utiču na distribuciju taksona obe istraživane grupe.

43
6. Literatura
Armitage, P. D.; Moss, D. Wright, J. F. & Furse M. T. 1983. The performance of a new biological water quality score system based on macroinvertebrates over a wide range of unpolluted running-water sites. Water Research, 17: 333-347.
AQEM, 2002. Manual for the application of the AQEM system. A comprehensive method to assess European streams using benthic macroinvertebrates, developed for the purpose of the Water Framework Directive. Contract No: EVK1-CT1999-00027)
Bailey, R. C.; Norris, R. H. & Reynoldson, T. B. 2001. Taxonomic resolution of benthic macroinvertebrate communities in bioassessments. J. N. Am. Benthol. Soc. 20:280–86
Berg, M. B. & Hellenthal, R. A. 1990. Data variability and the use of Chironomids in environmental studies: the standard error of the midge. In Davis, W.S. (ed.), Proceedings of the 1990 Midwest Pollution Control Biologists Meeting. U.S. Environmental Protection Agency Region V, Environmental Sciences Division, Chicago, IL. EPA-905–9-9-/005: 1–8.
Bonada, N.; Rieradevall, M. Prat, N. & Resh, V. H. 2006. Benthic
macroinvertebrate assemblages and macrohabitat connectivity in mediterranean-climate streams of northern California. Journal of the North American Benthological Society 25:32–43.
Calle-Martínez, D. & Casas, J.J. 2006. Chironomid species, stream classification, and water-quality assessment: the case of 2 Iberian Mediterranean mountain regions. Journal of the North American Benthological Society 25:465-476. doi: 10.1899/0887-3593(2006)25[465:CSSCAW]2.0.CO;2
Carter, J. L.; Fend, S. V. & Kenelly, S. S. 1996. The relationships
among three habitat scales and stream benthic invertebrate community structure. Freshwater Biology., 35: 109-124.

44
Carter, J. L.; Resh, V. H.; Rosenberg, D. M. & Reynolds, T. B.
2006. Biomonitoring in North American rivers: a comparison of methods used for benthic macroinvertebrates in Canada and the United States. Pages 203–228 in G. Ziglio, M. Siligardi, and G. Flaim (editors). Biological monitoring of rivers. John Wiley and Sons, West Sussex, UK
Chandler, J. R. 1970 A biological approach to water quality management. Water Pollution Control 69, 415±422.
Davies, P.E. 2000. Development of a national river bioassessment system (AUSRIVAS) in Australia. In: Wright, J.F., D.W. Sutcliffe & M.T. Furse (Eds), Assessing the Biological Quality of Fresh Waters: RIVPACS and other Techniques. Freshwater Biological Association, Ambleside (Cumbria, UK): 113-124.
Hellawell, J. M. 1986. Biological indicators of freshwater
pollution and environmental management. Elsevier Applied Science Publishers, New York, 546 pp.
Hilsenhoff, W.L. 1987. An improved biotic index of organic steam pollution. Great lakes Entomologist 20:31-39.
Johnson, R. K.; Wiederholm, T. & Rosenberg, D. M. 1993.
Freshwater biomonitoring using individual organisms, populations and species assemblages of benthic macroinvertebrates. In Rosenberg, D. M. & V. H. Resh (eds), Freshwater Biomonitoring and Benthic Macroinvertebrates. Chapman and Hall, New York, 40–158.
Lenat, D .R. & Resh, V. H. 2001. Taxonomy and stream ecology—the benefits of genus- and species-level identifications. Journal of the North American Benthological Society 20:287–98
Lencioni, V. & Rossaro, B. 2005. Microdistribution of chironomids (Diptera: Chironomidae) in Alpine streams: an autoecological perspective. Hydrobiologia 533:61-76. doi: 10.1007/s10750-004-2393-x

45
Marchant, R.; Barmuta, L. A. & Chessman, B. C. 1995. Influence
of sample quantification and taxonomic resolution on the ordination of macroinvertebrate comunities from running waters in Victoria, Australia. Marine & Freshwater Research., 46: 501-506.
Metcalfe, J. L. 1989. Biological water quality assessment of running waters based on macroinvertebrate communities: history and present status in Europe. Environmental Pollution Jurnal 60:101-139. doi: 10.1016/0269-7491(89)90223-6.
Milošević, D.; Simić, V.; Stojković, M.; Čerba, D.; Mančev, D.; Petrović, A. & Paunović, M. 2013. Spatio-temporal pattern of the Chironomidae community: towards the use of non-biting midges in bioassessment programs. Aquaric Ecology. In press.
Milošević, D.; Simić, V.; Stojković, M. & Živić, I. 2012. Chironomid faunal composition represented by taxonomic distinctness index reveals environmental change in a lotic system over three decades. Hydrobiologia 683:62-82. doi: 10.1007/s10750-011-0941-8
Norris, R. H. & Georges, A. 1993. Analysis and interpretation of benthic macroinvertebrate surveys. Pages 234–286 in D. M. Rosenberg and V.H. Resh (editors). Freshwater biomonitoring and benthic macroinvertebrates. Chapman and Hall, New York
Orendt, C. & Spies, M. 2012. Chironomini (Diptera: Chironomidae: Chironominae). Keys to Central European larvae using mainly macroscopic characters. Second revised edition, Leipzig
Plafkin, J. L.; Barbour, M. T.; Porter, K. D.; Gross, S. K. & Hughes, R. M. 1989. Rapid bioassessment protocols for use in streams and rivers. EPA/444/4-89-001. Environmental Protection Agency, Washington, D.C.,USA.
Puntí, T.; Rieradevall, M. & Prat, N. 2009. Environmental factors,
spatial variation, and specific requirements of Chironomidae in

46
Mediterranean reference streams. Journal of the North American Benthological Society 28:247-265. doi: 10.1899/07-172.1
Rabeni, C. F. & Wang, N. 2001. Bioassessment of streams using macroinvertebrates: Are the chironomidae necessary? Environmental Monitoring and Assessment 71:177-185. doi: 10.1023/A:1017523115381
Raunio, J.; Heino, J. & Paasivirta, L. 2011. Non-biting midges in biodiversity conservation and environmental assessment: Findings from boreal freshwater ecosystems. Ecol Indic 11:1057–1064. doi: 10.1016/j.ecolind.2010.12.002
Resh, V. H. & Jackson J. K. 1993. Rapid assessment approaches to biomonitorg using benthic macroinvertebrates. Freshwater Biomonitoring and Benthic Macroinvertebrates (Eds D.M. Rosenberg and V.H. Resh), pp. 195-223. Chapman & Hall, New York.
Resh V.H. & Unzicker J.D. (1975) Water quality monitoring and aquatic organisms: the importance of species identification. Journat of the Water Pol/ution Control Federation, 47, 9--19.
Reynoldson, T. B.; Bailey, R. C.; Day, K. E. & Norris, R. H. 1995. Biological guidelines for freshwater sediment based on BEnthic Assessment of SedimenT (the BEAST) using a multivariate approach for predicting biological state. Australian Journal of Ecology 20:198–219 Rosenberg, D. M. 1993. Freshwater biomonitoring and Chironomidae. Aquatic Ecology 26:101-122. doi: 10.1007/BF02255231 Saether, O.A. 1979. Chironomid communities as water quality indicator. Holarctic ecology 2: 65-74.
Savić, A.; Ranđelović, V. & Krpo-Ćetković, J. 2010. Seasonal
variability in community structure and habitat selction of mayflies (Ephemeroptera) in the Nišava River (Serbia), Biotechnology and

47
Biotechnological Equipment., special edition, Plovdiv. Vol. 24(2): 639–645.
Vallenduuk, H.J. & Moller Pillot, H.K.M. 2007. Chironomidae Larvae Vol. I: General Ecology and Tanypodinae. KNNV Publishing, Zeist.
Vannote, R. L.; Minshall, G. W.; Cummins, K. W.; Sedell, J.R. & Cushing, C. E. 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences Sci. 37:130-37
Wallace, J. B.; Grubaugh, J. W. & Whiles, M. R. 1996. Biotic indices and stream ecosystem processes: results from an experimental study. Ecological Applications 6:140-151.
Wenn, C.L. 2008. Do freshwater macroinvertebrates reflect
water quality improvements following the removal of point source pollution from Spen Beck, West Yorkshire? Earth & E-nvironment 3: 369-406
Wilson, R. 1992. Monitoring organic enrichment of rivers using chironomid pupal exuvial assemblages. Aquatic Ecology 26:521-525. doi: 10.1007/BF02255285 Wright, J.F.; Moss, D.; Furse, M.T. & Armitage, P.D. 1984. A preliminary classification of running-water sites in Great Britain based on macroinvertebrate species and the prediction of community type using environmental data. Freshwater Biology, 14, 211–256. Wright, J. F.; Sutcliffe, D. W. & Furse, M. T. (eds), 2000. Assessing the Biological Quality of Freshwaters: RIVPACS and Similar Techniques. Freshwater Biological Association, Ambleside.
Прилог 5/1

48
ПРИРОДНO - MАТЕМАТИЧКИ ФАКУЛТЕТ НИШ
КЉУЧНА ДОКУМЕНТАЦИЈСКА ИНФОРМАЦИЈА
Редни број, РБР: Идентификациони број, ИБР: Тип документације, ТД: монографска
Тип записа, ТЗ: текстуални / графички
Врста рада, ВР: мастер рад
Аутор, АУ: Иван Тодосијевић
Ментор, МН: Славиша Стаменковић
Наслов рада, НР: Индикаторска способност две групе макрозообентоса за процену квалитета воде слива реке Нишаве
Језик публикације, ЈП: српски
Језик извода, ЈИ: енглески
Земља публиковања, ЗП: Р. Србија
Уже географско подручје, УГП: Р. Србија
Година, ГО: 2013.
Издавач, ИЗ: ауторски репринт
Место и адреса, МА: Ниш, Вишеградска 33.
Физички опис рада, ФО: (поглавља/страна/ цитата/табела/слика/графика/прилога)
43 стр. ; граф. прикази
Научна област, НО: биологија
Научна дисциплина, НД: екологија
Предметна одредница/Кључне речи, ПО: Фамилија хирономида, ЕПТ група, мултидимензионално скалирање, средински фактори
УДК 574.587:556.53(497.11 Ниш)
Чува се, ЧУ: библиотека
Важна напомена, ВН:
Извод, ИЗ: Циљ овог истраживања је био да упоредимо способност фамилије хирономида и групе ЕПТ у радвајању локалитета различитог нивоа квалитета воде. Резултати nMDS анализе показали су које су групе локалитета квалитативно и квантитативно слични. Ова студија је показала да се на основу заједнице хирономида и ЕПТ групе, са таксономском резолуцијом до нивоа рода, могу формирати групе са локалитетима различитог квалитета и степена антропогеног утицаја.
Датум прихватања теме, ДП: 27.12.2012
Датум одбране, ДО:
Чланови комисије, КО: Председник: Члан: Члан, ментор:
Образац Q4.09.13 - Издање 1

49
Прилог 5/2
ПРИРОДНО - МАТЕМАТИЧКИ ФАКУЛТЕТ НИШ
KEY WORDS DOCUMENTATION
Accession number, ANO:
Identification number, INO:
Document type, DT: monograph
Type of record, TR: textual / graphic
Contents code, CC: master thesis
Author, AU: Ivan Todosijević
Mentor, MN: Slaviša Stamenković
Title, TI: Two groups of macrozoobenthos as a water quality indicators in the Nisava river basin
Language of text, LT: Serbian
Language of abstract, LA: English
Country of publication, CP: Republic of Serbia
Locality of publication, LP: Serbia
Publication year, PY: 2013
Publisher, PB: author’s reprint
Publication place, PP: Niš, Višegradska 33.
Physical description, PD: (chapters/pages/ref./tables/pictures/graphs/appendixes)
43 p. ; graphic representations
Scientific field, SF: biology
Scientific discipline, SD: ecology
Subject/Key words, S/KW: Chironomidae family, EPT group, multidimensional scaling, environmental factors
UC 574.587:556.53(497.11 Niš)
Holding data, HD: library
Note, N:
Abstract, AB: The main objective of this study was to compare discimination strenghts in assessment of water quality using the Chironomidae family and EPT group. Results of nMDS analyses revealed which group of sites are qualitative or quantitative similar. This study pointed out that information about community pattern of Chironomid and EPT group, identified up to genus level can be sufficient for reliable classification of sites with different level of anthropogenic impact.
Accepted by the Scientific Board on, ASB: 27.12.2012
Defended on, DE:
Defended Board, DB: President:
Member:
Member, Mentor:
Образац Q4.09.13 - Издање 1