Embed Size (px)
Citation preview

INFORME FINAL DEL PROYECTO DE
INVESTIGACION CGPI-20040267
PURIFICACION Y CARACTERIZACION DE
UNA AMINOPEPTIDASA DE
Rhizobium tropici
RESPONSABLE
Dra. Rosa Ma. Ahidé López Merino
Laboratorio de Microbiología General Escuela Nacional de Ciencias Biológicas
1

RESUMEN Se purificó una aminopeptidasa de Rhizobium tropici cepa 299. El proceso de
purificación consistió en un fraccionamiento con sulfato de amonio y tres pasos de
cromatografía. Este procedimiento dió por resultado un grado de recuperación de
4.13% y un incremento de 700 veces en la actividad específica. La aminopeptidasa
parece ser una enzima monomérica con masa molecular de 98 kDa y un punto
isoeléctrico de 3.9. La actividad fue óptima a pH de 7 y a 37° C.
La actividad enzimática fue inhibida con EDTA, 1,10 – fenantrolina y cationes
divalentes (Zn+2, Ca+2 y Hg+2), lo cual sugirió que se trataba de una metalo
aminopeptidasa. La enzima actuó preferentemente sobre la alanina del derivado
aminoacil del extremo N terminal . Los valores de Km para Ala-pNA y Lys-pNA fueron
0.51 mM y 0.23 mM respectivamente. Se determinó la secuencia del extremo amino
terminal de la aminopeptidasa y presentó homología con alanil aminopetidasas (PepN)
putativas de Agrobacterium tumefaciens, Sinorhizobium meliloti and Mesorhizobium loti.
2

PURIFICACION Y CARACTERIZACION DE UNA AMINOPEPTIDASA DE Rhizobium tropici
De Ley fue el primer investigador que asoció a las bacterias de rápido
crecimiento como Rhizobium, Agrobacterium y Phillobacterium con bacterias de mas
lento crecimiento del género Brucella. Con ello se estableció por vez primera una
filiación taxonómica con base a similitudes en el cistrón rRNA. Análisis de las
secuencias del gen ribosomal 16S posteriores, permitieron ubicar a estas bacterias en
el grupo de las alfa Proteobacterias. En la división 2 alfa se encuentra Brucella
relacionada específicamente con Rickettsiae, Agrobacteria y Rhizobiae, que son
bacterias que viven en asociación estrecha con células eucarióticas. La familia
Rhizobiaceae comprende géneros como Rhizobium, Sinorhizobium y Mesorhizobium
que forman nódulos fijadores de nitrógeno en plantas, a Agrobacterium que produce
tumores en las plantas y entre otros géneros se tiene a Brucella que está conformado
por patógenos intracelulares facultativos de animales. Investigaciones recientes han
puesto de manifiesto la presencia de algunos genes involucrados en la simbiosis de
Sinorhizobium meliloti que aparentemente son homólogos a los implicados en la
patogénesis de Brucella. Por otro lado, los genes bac de las rhizobiae que participan en
la diferenciación hacia bacteroide, se ha visto que son homólogos a los genes de
Brucella que participan en su sobrevivencia en el interior del macrófago, los que
también participan en la patogénesis en los ratones. De igual forma, se ha reportado
que el sistema regulador de dos componentes BvrR y BvrS, que está relacionado con
la virulencia de Brucella, se encuentra representado por genes Exo en S. meliloti. La
proteasa periplásmica de Sinorhizobium meliloti codificada por el gen degP es mas
parecida al gene correspondiente de B. abortus que al de E. coli.
Estudios recientes realizados por nuestro grupo de investigación, empleando el
método de electroforesis de enzimas metabólicas multilocus (MLEE), demostraron que
Brucella presenta una relación estrecha con Rhizobium tropici.
Finalmente, nuestro grupo de investigación publicó la purificación y
caracterización de una aminopeptidasa N inmunogénica (APE97) que fue obtenida de
una cepa de Brucella melitensis. Al comparar la secuencia de amino ácidos obtenida
usando el programa BLASTpsi se constató que la estructura de la secuencia primaria
3

de la APE97 de B. melitensis era similar (99%) a una aminopeptidasa N putativa de
Brucella suis 1330. Asimismo, se encontraron identidades menores con una
aminopeptidase N putativa de Mesorhizobium loti (63%), con una aminopeptidasa N
putativa de Sinorhizobium meliloti (59%) y con una aminoeptidasa N putativa de
Agrobacterium tumefaciens (58%).
La degradación de proteínas intracelular es un proceso esencial en las bacterias
para llevar a cabo: la eliminación de proteínas dañadas, la modulación de los niveles de
proteínas, y el mantenimiento del nivel de los diferentes aminoácidos. Tradicionalmente
se aceptaba, que la función primaria de las enzimas proteolíticas tenía por fin la
adquisición de nutrientes para el crecimiento. Sin embargo, estudios recientes han
colocado a las enzimas proteolíticas, derivadas de patógenos, en otro contexto ya que
se ha observado que pueden tener una participación en la sobrevivencia de la bacteria
en medios ambientes hostiles. Las aminopeptidasas (APEs) de muchas bacterias se
han estudiado desde el punto de vista estructural y enzimático. En forma específica, las
de Porphyromonas gingivalis se han relacionado con procesos como periodontitis
severas y enfermedades cardiovasculares. En el caso Aeromonas spp, se ha sugerido
que las enzimas extracelulares juegan un papel importante en la invasión al tejido y en
el establecimiento de la infección.
Rhizobium tropici es una bacteria que fija nitrógeno en forma simbiótica con
algunas leguminosas tropicales, formando nódulos en sus raíces. Es muy resistente al
choque ácido y a concentraciones altas de aluminio, que son condiciones prevalentes
en los ecosistemas donde se desarrollan las plantas. R. tropici se ha seleccionado
como modelo para el estudio de las interacciones planta – bacteria, esta bacteria porta
un plásmido simbiótico que ha sido expresado con éxito en diferentes bacterias.
Con base en los antecedentes expuestos en este proyecto se proyectó llevar a
cabo la purificación y caracterización de una aminopeptidasa de Rhizobium tropici y
una vez realizada esta parte, se compararía con la APE97 de Brucella melitensis, que
como se señaló en los párrafos anteriores, es una bacteria filogenéticamente cercana.
La importancia fisiológica que tendrá esta enzima durante el proceso simbiótico de
fijación del nitrógeno, será motivo de una investigación posterior a la que se propuso
desarrollar en este proyecto.
4

OBJETIVO GENERAL
Purificar y caracterizar una aminopeptidasa de Rhizobium tropici y compararla con la
aminopeptidasa de Brucella melitensis reportada
OBJETIVOS PARTICULARES
1. Obtener un esquema para la purificación de una aminopeptidasa de Rhizobium
tropici mediante diferentes métodos.
2. Caracterizar la aminopeptidasa de Rhizobium tropici mediante diferentes métodos
3. Comparar la aminopeptidasa purificada con la de Brucella melitensis descrita
previamente
MATERIAL Y METODOS 1.- Cepas bacterianas. a) Cepa de Rhizobium tropici CFN 299 perteneciente a la colección del Centro de
Investigación sobre Fijación de Nitrógeno de la UNAM
2.- Cultivo de las cepas y preparación de los extractos crudos. Se partió de una placa de la cepa que fue proporcionada por la Dra. Esperanza
Martínez del Centro de Investigación sobre Fijación de Nitrógeno de la UNAM. Se
sembró en placas con agar peptona extracto de levaduras con CaCl2 (PY). Una vez
verificada la pureza de la cepa, se sembraron los matraces semilla que contenían el
medio PY. A partir de ellos, se sembraron matraces de 2 lt con 1400 ml de medio PYE.
Se realizarán tinciones de Gram, para determinar la pureza y la fase de la cepa en cada
una de las etapas. La cepa se resembró en cajas con PY y se almacenó a 4° C.
5

Se cosecharon las células por centrifugación a 9,000 x g durante 20 minutos
para recuperar la masa bacteriana y se lavaron las células tres veces con regulador de
imidazol 10 mM estéril. Se pesaron para obtener el rendimiento en peso húmedo y se
realizó una suspensión al 10 % (peso/vol) en Imidazol 10 mM. Se rompieron las células
con varios ciclos de ultrasonido. Se mantuvieron en baño de hielo para evitar que se
dañaran las enzimas del extracto crudo.
DIAGRAMA DE TRABAJO
Crecimiento celular de 48 h. en caldo PY-caldo PY- CaCl2 a 28° C)
Rompimiento con ultrasonido en baño de hielo a 4°C
Centrifugación 14,000 rpm, 30 min. a 4°C
Ultracentrifugación (40,000 rpm, 2h, 4°C)
Precipitación con sulfato de amonio (40-70%)
Precipitación con sulfato de estreptomocina
Posteriormente se centrifugaron las células rotas a 9,000 x g durante 30 minutos,
se separó el sobrenadante (extracto crudo de la cepa, ExC-Rt). El extracto crudo se le
determinó la concentración de proteínas por el método de Pierce. El extracto crudo se
sometió a una ultra centrifugación a 40,000 rpm durante 1.5 hs, a 4° C.
Al sobrenadante resultante se precipitó con sulfato de amonio entre 20-70%. Se
dializó exhaustivamente contra el mismo amortiguador hasta la desaparición de la sal.
Posteriormente se le realizó una precipitación con sulfato de estreptomicina para
eliminar los ácidos nucleicos. La muestra se filtró a través de una membrana Millipore
de 0.22 micras de poro y se almacenó en congelación a –20° C.
3.- Purificación de las proteínas de interés. Se purificó la proteína enzimática usando pasos de cromatografía secuenciales
mediante el sistema FPLC (Fast Protein Liquid Chromatography, Amersham
Pharmacia), se incluyeron las columnas siguientes:
6

Para la cromatografía de intercambio iónico se empleó una columna Q Sepharose High
Performance (60ml) equilibrada con imidazol 10 mM (pH 7.0). Se le aplicó un gradiente
de 0 a 1 M de NaCl y se colectaron las fracciones a las que se les determinó la
actividad empleando Lys-pNA. Las fracciones con actividad enzimática se juntaron en
un tubo. Esta muestra se depositó en una columna Phenyl superose HR 5/5 equilibrada
con fosfato de sodio 0.1 M y sulfato de amonio 1.5 M (pH 7.0). La fracción activa se
depositó en una columna Superose 12HR .
4.- Determinación de la actividad aminopeptidasa.
La mezcla de reacción se llevó a cabo con 100 µl de muestra, 120 µl de agua,
250 µl de Tris-HCl 0.1 M pH 7 y 30 µl L-lisina-p-nitroanilida 10 mM (Lys-pNA). La
mezcla de reacción se incubó a 37° C hasta la aparición de un color amarillo. La
reacción se detuvo adicionando 500 µl de sulfato de zinc al 5% y 100 µl de hidróxido de
bario al 7.5 %. La mezcla se centrifugó a 14,000 rpm durante 10 minutos y el
sobrenadante se leyó en un espectrofotómetro a 405 nm para valorar la cantidad de
nitroanilida liberada. Una unidad de actividad de esta proteasa se definió como la
cantidad de enzima que hidroliza 1 µM de sustrato por minuto (Hirsh y col., 1988).
5.- Electroforesis en geles de poliacrilamida (PAGE – SDS). Las electroforesis en gel de poliacrilamida al 10 % (acrilamida – bisacrilamida
30:0.8), se realizaron según el método de Laemmli, y se emplearon para monitorear la
purificación de las aminopeptidasas contenidas en el extracto ExC-Rt, así como para
estimar la masa molecular de la enzima. Los geles se prepararon en condiciones
desnaturalizantes, se corrieron en una cámara de electroforesis vertical (Mini-
PROTEAN II, BIO-RAD).
Una vez realizada la separación electroforética se procedió a la tinción de los
geles con azul brillante de Coomasie R-250 (BIO-RAD Cat. N° ) al 0.125 % en metanol,
agua bidestilada y ácido acético (relación 5:5:1) durante 2 horas. Posteriormente los
geles de poliacrilamida se destiñeron con una mezcla de metanol, agua y ácido acético
en proporción 5:5:1.
7

6.- Caracterización de la proteína purificada. Se estimó el Peso molecular de la aminopetidasa mediante PAGE-SDS y por
FPLC filtración en gel. La filtración en gel se llevó a cabo en una columna de Superosa
12 HR 10/30 calibrada con proteínas de peso molecular conocido (estándares).
Se medió el efecto del pH sobre la actividad de la aminopeptidasa en un intervalo de
pH de 3 a 11. El efecto de la temperatura sobre la actividad de la aminopeptidasa se
medió en un intervalo de 4 a 70° C. El punto isoeléctrico se determinó en una columna
Mono P por cromatoenfoque eluida con polibuffer 74 pH 4 / imidazole 25mM pH 7.4.
7. - Cinética enzimática.
La enzima se incubó con varias concentraciones de L-lisina-p-nitroanilida, a una
concentración final de 0.025-1mM. La determinación de la Vmax y Km se realizó por triplicado.
8.- Secuenciación del extremo amino terminal La proteína purificada con actividad de aminopeptidasa, se transfirió a una
membrana PVDF y se envió para que le secuenciaran el extremo amino terminal
(University of Virginia Health System, Charlottesville, VA). Se realizó un análisis por
BLAST con la secuencia obtenida.
RESULTADOS Purificación de la actividad aminopeptidasa. Se purificó una actividad Ala-pNA a
partir de un extracto de células viables de Rhizobium tropici A cepa 299 sonicadas. La
precipitación con sulfato de amonio (40-70%) y los subsecuentes pasos de
cromatografía (Figuras 1, 2 y 3) dieron por resultado final el aislamiento de una enzima
homogénea. La aminopeptidasa fue purificada 700-veces con un porcentaje de
recuperación de 4.13 % de la actividad de la enzima (Tabla 1).
8
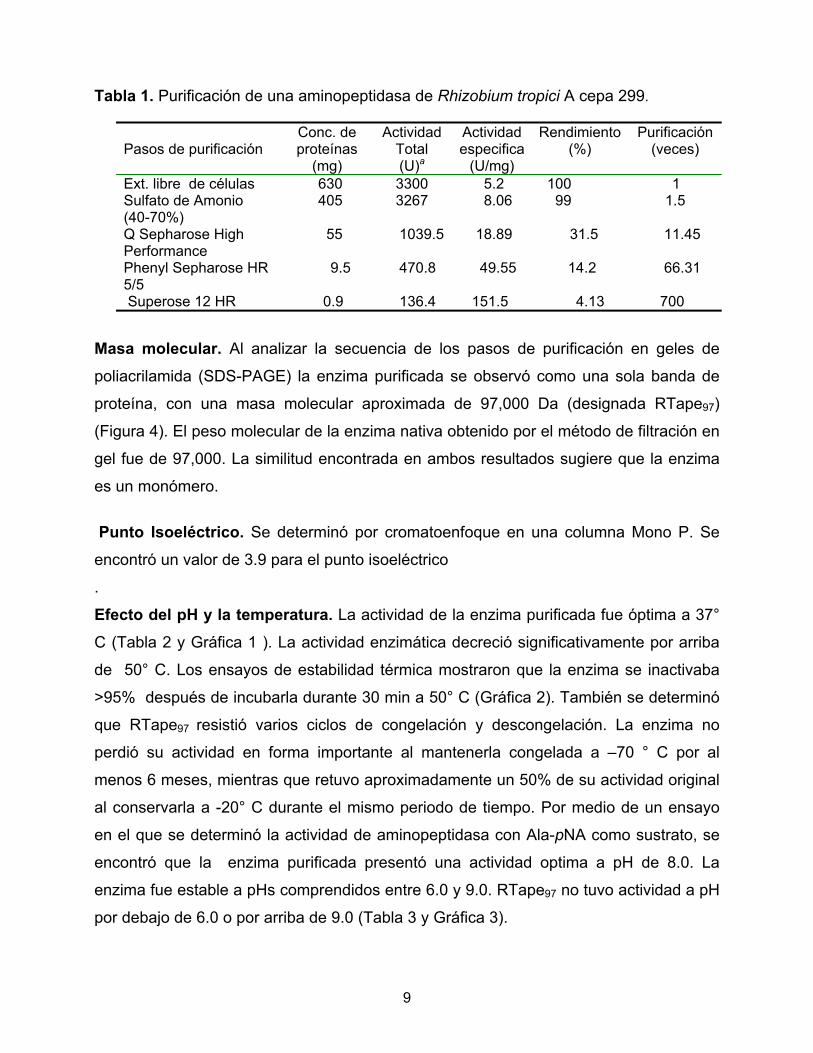
Tabla 1. Purificación de una aminopeptidasa de Rhizobium tropici A cepa 299.
Pasos de purificación
Conc. de proteínas
(mg)
Actividad Total (U)a
Actividad especifica
(U/mg)
Rendimiento (%)
Purificación (veces)
Ext. libre de células 630 3300 5.2 100 1 Sulfato de Amonio (40-70%)
405 3267 8.06 99 1.5
Q Sepharose High Performance
55 1039.5 18.89 31.5 11.45
Phenyl Sepharose HR 5/5
9.5 470.8 49.55 14.2 66.31
Superose 12 HR 0.9 136.4 151.5 4.13 700 Masa molecular. Al analizar la secuencia de los pasos de purificación en geles de
poliacrilamida (SDS-PAGE) la enzima purificada se observó como una sola banda de
proteína, con una masa molecular aproximada de 97,000 Da (designada RTape97)
(Figura 4). El peso molecular de la enzima nativa obtenido por el método de filtración en
gel fue de 97,000. La similitud encontrada en ambos resultados sugiere que la enzima
es un monómero.
Punto Isoeléctrico. Se determinó por cromatoenfoque en una columna Mono P. Se
encontró un valor de 3.9 para el punto isoeléctrico
.
Efecto del pH y la temperatura. La actividad de la enzima purificada fue óptima a 37°
C (Tabla 2 y Gráfica 1 ). La actividad enzimática decreció significativamente por arriba
de 50° C. Los ensayos de estabilidad térmica mostraron que la enzima se inactivaba
>95% después de incubarla durante 30 min a 50° C (Gráfica 2). También se determinó
que RTape97 resistió varios ciclos de congelación y descongelación. La enzima no
perdió su actividad en forma importante al mantenerla congelada a –70 ° C por al
menos 6 meses, mientras que retuvo aproximadamente un 50% de su actividad original
al conservarla a -20° C durante el mismo periodo de tiempo. Por medio de un ensayo
en el que se determinó la actividad de aminopeptidasa con Ala-pNA como sustrato, se
encontró que la enzima purificada presentó una actividad optima a pH de 8.0. La
enzima fue estable a pHs comprendidos entre 6.0 y 9.0. RTape97 no tuvo actividad a pH
por debajo de 6.0 o por arriba de 9.0 (Tabla 3 y Gráfica 3).
9
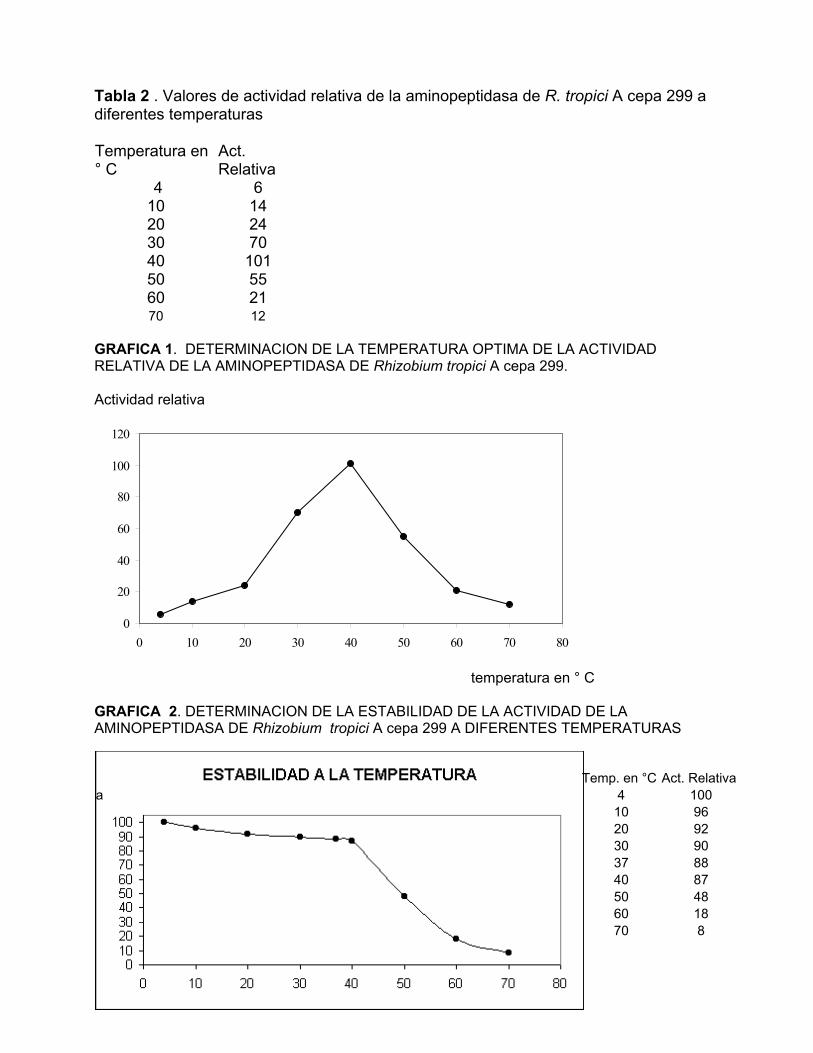
Tabla 2 . Valores de actividad relativa de la aminopeptidasa de R. tropici A cepa 299 a diferentes temperaturas Temperatura en ° C
Act. Relativa
4 6 10 14 20 24 30 70 40 101 50 55 60 21 70 12
GRAFICA 1. DETERMINACION DE LA TEMPERATURA OPTIMA DE LA ACTIVIDAD RELATIVA DE LA AMINOPEPTIDASA DE Rhizobium tropici A cepa 299. Actividad relativa
0
20
40
60
80
100
120
0 10 20 30 40 50 60 70 80
temperatura en ° C GRAFICA 2. DETERMINACION DE LA ESTABILIDAD DE LA ACTIVIDAD DE LA AMINOPEPTIDASA DE Rhizobium tropici A cepa 299 A DIFERENTES TEMPERATURAS
10
Temp. en °C Act. Relativa a 4 100 10 96 20 92 30 90 37 88 40 87 50 48 60 18 70 8
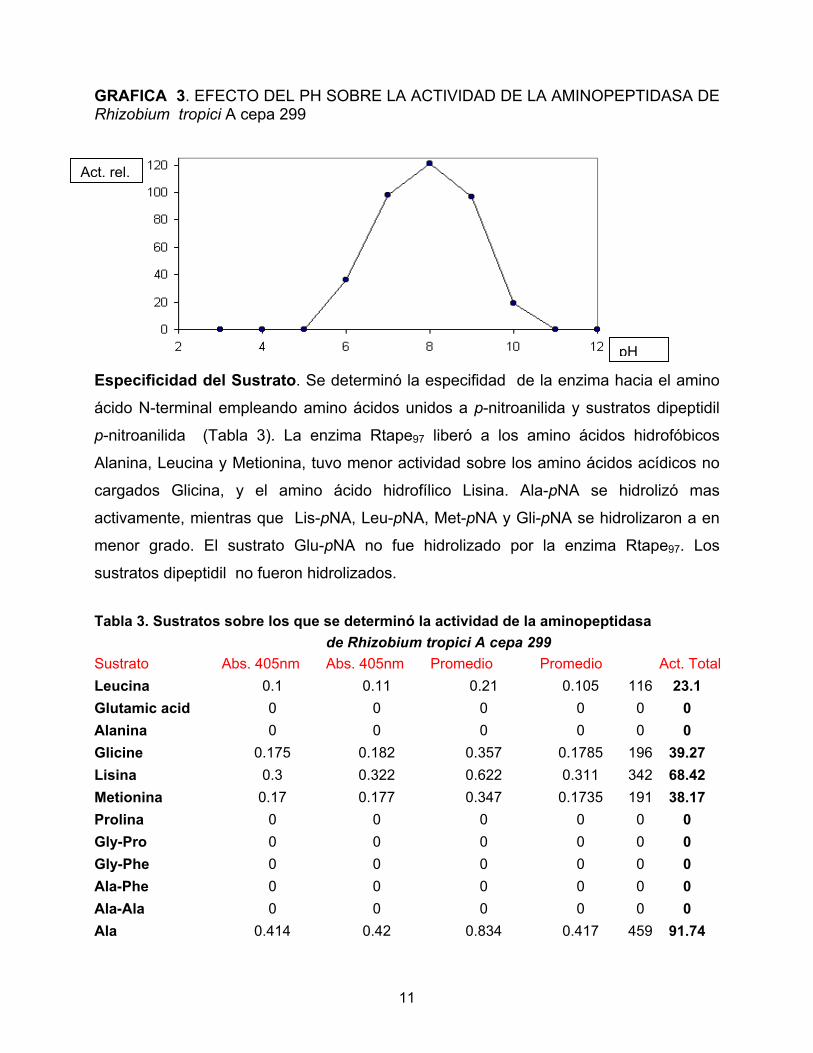
GRAFICA 3. EFECTO DEL PH SOBRE LA ACTIVIDAD DE LA AMINOPEPTIDASA DE Rhizobium tropici A cepa 299
Especificidad del Sustrato. Se determinó la especifidad de la enzima
ácido N-terminal empleando amino ácidos unidos a p-nitroanilida y sus
p-nitroanilida (Tabla 3). La enzima Rtape97 liberó a los amino ácido
Alanina, Leucina y Metionina, tuvo menor actividad sobre los amino ácid
cargados Glicina, y el amino ácido hidrofílico Lisina. Ala-pNA se
activamente, mientras que Lis-pNA, Leu-pNA, Met-pNA y Gli-pNA se hi
menor grado. El sustrato Glu-pNA no fue hidrolizado por la enzima
sustratos dipeptidil no fueron hidrolizados. Tabla 3. Sustratos sobre los que se determinó la actividad de la aminopept de Rhizobium tropici A cepa 299 Sustrato Abs. 405nm Abs. 405nm Promedio Promedio Leucina 0.1 0.11 0.21 0.105 Glutamic acid 0 0 0 0 Alanina 0 0 0 0 Glicine 0.175 0.182 0.357 0.1785 Lisina 0.3 0.322 0.622 0.311 Metionina 0.17 0.177 0.347 0.1735 Prolina 0 0 0 0 Gly-Pro 0 0 0 0 Gly-Phe 0 0 0 0 Ala-Phe 0 0 0 0 Ala-Ala 0 0 0 0 Ala 0.414 0.42 0.834 0.417
11
pH
Act. rel.
hacia el amino
tratos dipeptidil
s hidrofóbicos
os acídicos no
hidrolizó mas
drolizaron a en
Rtape97. Los
idasa
Act. Total 116 23.1
0 0 0 0
196 39.27 342 68.42 191 38.17
0 0 0 0 0 0 0 0 0 0
459 91.74

Sustrato Act. Total por 100 Act. Relativa Act. Relativa Leucina 23.1 2310 25.17986 25 Glutamic acid 0 0 0 0 Alanina-Pro 0 0 0 0 Glicine 39.27 3927 42.80576 43 Lisina 68.42 6842 74.58034 75 Metionina 38.17 3817 41.60671 42 Prolina 0 0 0 0 Gly-Pro 0 0 0 0 Gly-Phe 0 0 0 0 Ala-Phe 0 0 0 0 Ala-Ala 0 0 0 0 Ala 91.74 9174 100 100 Perfil de Inhibición. La influencia de algunos agentes sobre la actividad de Rtape97 se
encuentran sumariados en la Tabla 4. La actividad de Rtape97 se inhibió casi
completamente en presencia de agentes quelantes como la 1,10 -phenanthrolina (10
µM) y EDTA (200 mM). Bestatina, que es un inhibidor típico de exopeptidasas, inhibió
la actividad en mas del 50%. Rtape97 no fue sensible a los inhibidores de la proteasa de
serina como leupeptina, pefabloc, y PMSF; E64, que es un inhibidor de proteinasas de
cisteina y la pepstatina, inhibidor específico del aspártico, no tuvieron efecto sobre la
actividad de Rtape97. La presencia de Zn+2, Hg+2, Ca+2 y Co+2 causó una inhibición
completa a una concentración de 1.0 mM, mientras que se observó un efecto inhibitorio
de mas del 50% en presencia de Mg+2 (tabla 4 ).
Parametros Cinéticos. Los valores encontrados para la constante de Michaelis (Km) y
para la velocidad máxima de reacción (Vmax) fueron 0.51 mM y 63 mmol min-1 mg de
proteína para L-Ala-pNA respectivamente; y 0.23 mM, y 15 mmol min –1 mg de proteína
para L-Lys-pNA respectivamente.
Determinación de la secuencia Amino terminal. Se determinó la secuencia N-
terminal que fue MRTDTGQIINLADYXPTDFVL. La secuencia fue usada como plantilla
para buscar en el sitio web NCBI secuencias de proteínas similares. La búsqueda
12

reveló similitudes con aminopeptidasas N putativas de Agrobacterium tumefaciens
(NP_354008), Sinorhizobium meliloti (NP_385132) y Mesorhizobium loti (NP_107963 Tabla 4. EFECTO DE ALGUNOS INHIBIDORES SOBRE LA ACTIVIDAD DE LA AMINOPEPTIDASA DE Rhizobium tropici A cepa 299 Inhibidor Concentración
En mM % de actividad residual a
Ninguno 100 Zn2+ 1 0 Co2+ 1 94 Ca2+ 1 0 Hg2+ 1 0 Mg2+ 1 48 EDTA 2.5 80 200 0 Bestatina 0.25 52 Pepstatina 0.025 95 0.05 98 PMSF 0.005 74 E-64 0.05 89 1,10-phenanthroline 0.01 0 a Expresado como el porcentaje de la actividad obtenida en ausencia de ningún agente químico, al cual se le asignó un valor de 100 %
13
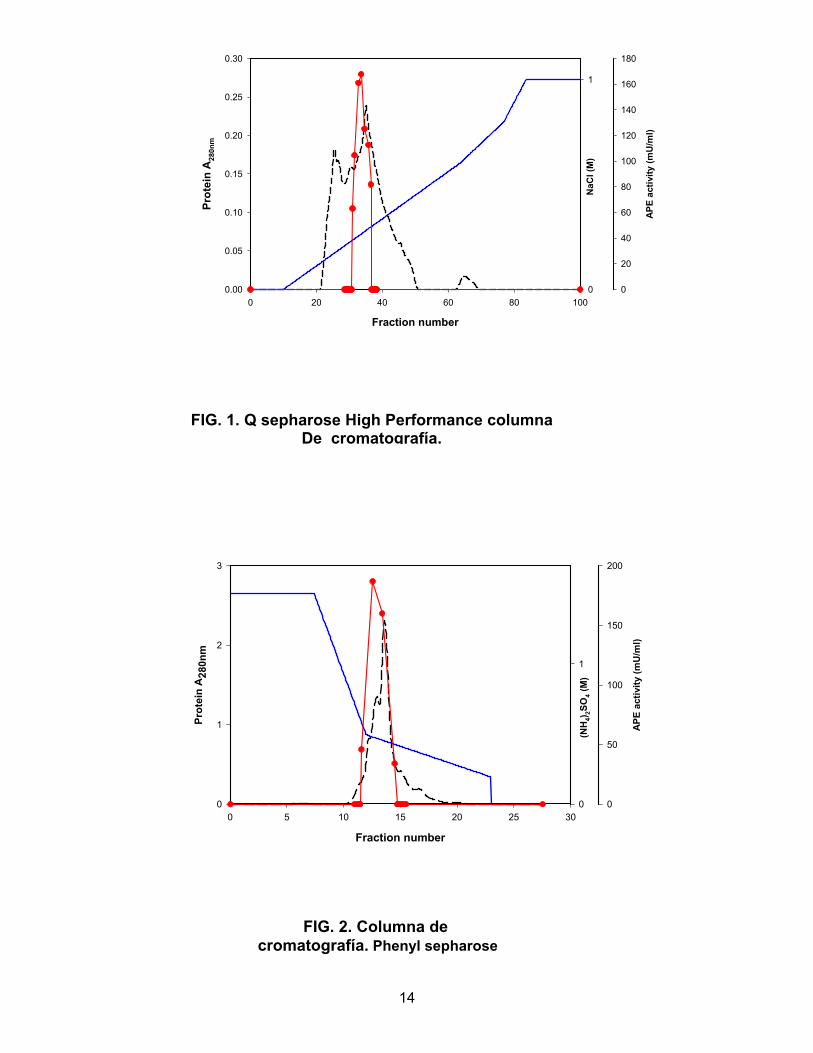
Fraction number
0 20 40 60 80 100
Prot
ein
A28
0nm
0.00
0.05
0.10
0.15
0.20
0.25
0.30
NaC
l (M
)
0
1
APE
activ
ity (m
U/m
l)
0
20
40
60
80
100
120
140
160
180
FIG. 1. Q sepharose High Performance columna De cromatografía.
Fraction number
0 5 10 15 20 25 30
Prot
ein
A28
0nm
0
1
2
3
(NH 4) 2S
O4 (
M)
0
1
APE
activ
ity (m
U/m
l)
0
50
100
150
200
FIG. 2. Columna de cromatografía. Phenyl sepharose
14
Fraction number
0 10 20 30
Prot
ein
A28
0nm
0.000
0.002
0.004
0.006
0.008
0.010
0.012
0.014
0.016
0.018
APE
activ
ity (m
U/m
l)
0
20
40
60
80
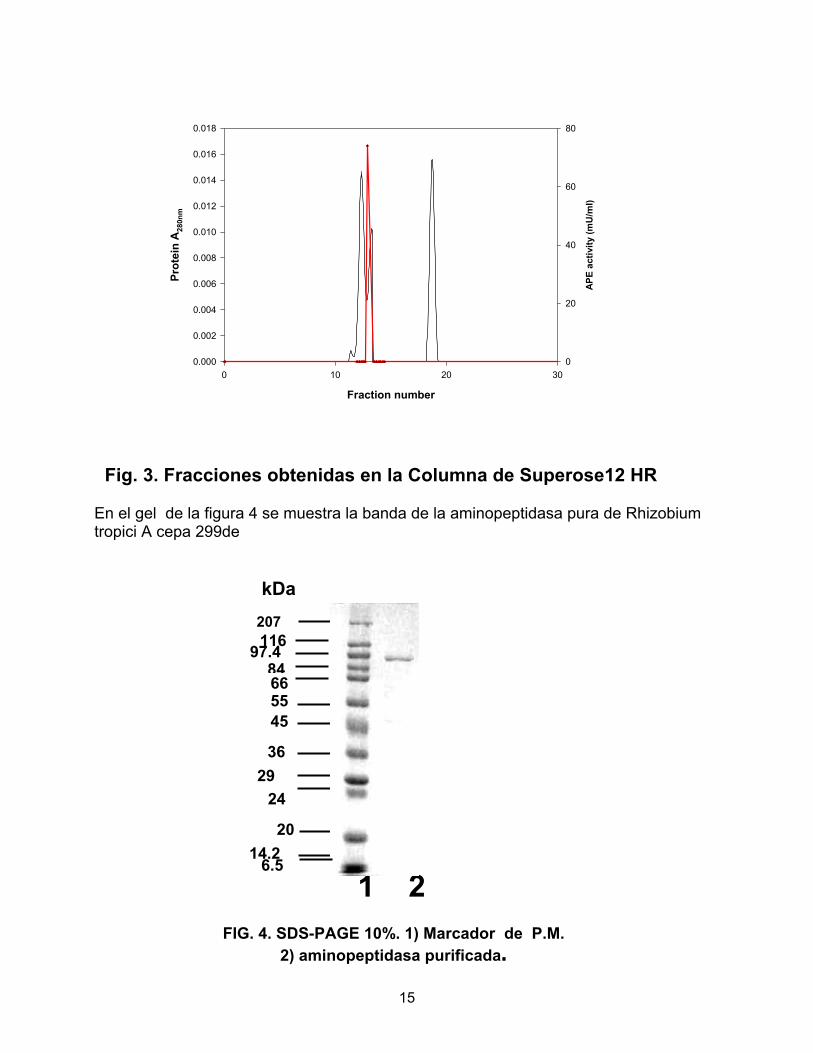
Fig. 3. Fracciones obtenidas en la Columna de Superose12 HR En el gel de la figura 4 se muestra la banda de la aminopeptidasa pura de Rhizobium tropici A cepa 299de kDa
207
1 2
20
24 29
36
116 97.4 8466 55 45
14.2
6.5 FIG. 4. SDS-PAGE 10%. 1) Marcador de P.M.
2) aminopeptidasa purificada.
15

IMPACTO. En el presente estudio se describen los pasos que se siguieron para lograr la
purificación de una aminopeptidasa de Rhizobium tropici cepa 299, asimismo se
reporta la caracterización bioquímica y con base en los análisis realizados en las bases
de datos, se postula que es muy similar a la aminopeptidasa
Este tipo de estudios experimentales básicos que analizan procesos fisiológicos de la
bacteria, no tienen un impacto inmediato ya que se debe de continuar la investigación
hasta determinar el papel que juega esta enzima en la fisiología de la bacteria y en el
establecimiento de la simbiosis con la planta.
La investigación sin embargo tiene una gran importancia ya que contribuye en la
dilucidación de procesos proteolíticos que se dan en estas bacterias Esta enzima no ha
sido estudiada con anterioridad ni reportada y de acuerdo con la búsqueda bibliográfica
parecería que somos los primeros en abordar este aspecto en la fisiología de la
bacteria.
REFERENCIAS
1. Alen-Boerrigter, I. J. van, R. Bankreis, and W. M de Vos. 1991. Characterization and overexpression of the Lactococcus lactis pepN gene and localization of its product, aminopeptidase N. Appl. Environ. Microbiol. 57:2555-2561.
2. Altschul, S. F., T. L. Madden, A. A. Schäffer, J. Z., Z. Zhang, W. Miller, and D. J. Lipman.
1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res.25:3389-3402.
3. Banbula, A., P. Mak, M. Bugno, J. Silberring, A. Dubin, D. Nelson, J. Travis, and J. Potempa. 1999. Prolyl trypeptidyl peptidase from Porphyromonas gingivalis. J. Biol. Chem. 274:9246-9252.
4. Barret, A. J., N. D. Rawlings, and J. F. Woessner. 1998. Handbook of proteolytic
enzymes. Academic Press, San Diego, Calif.
5. Buchmeier, A. N, & Heffron F. 1989. Induction of Salmonella stress proteins upon infection of macrophages. Science. 248:730-732.
6. Céspedes, S., E. Andrews, H. Folch & A. Oñate. 2000. Identification and partial
characterization of a new protective antigen of Brucella abortus. J. Med. Microbiol. 49:165-170.
16

7. Contreras-Rodríguez A. , B. Ramírez Z., A. Contreras, G. Schurig, N Sriranganathan & A. López Merino. 2004. Purification and characterization of an immunogenic aminopeptidase of Brucella melitensis. Infect. Immun. 71: 5238-5244
8. Corbel M.J. Brucellosis: an overview. 1997. Emerg. Infect. Dis. 3:213-221.
9. DelVecchio, V., V. Kapatral, R. J. Redkar, G. Patra, C. Mujer, T. Los, N. Ivanova, I.
Anderson, A. Bhattacharyya, A. Lykidis, G. Reznik, L. Jablonski, N. Larsen, M. D´Souza, A. Bernal, M. Mazur, E. Goltsman, E. Selkiv, P. H. Elzer, S. Hagius, D. Oócallaghan, J. J. Letesson, R. Haselkon, N. Kyrpides & R. Overbeek. 2002. The genome sequence of the facultative intracellular pathogen Brucella melitensis. Proc. Natl. Acad. Sci. 99:443-448.
10. Ekaza E., L. Guilloteau, J. Teyssier, J.P. Lituard S. Kohleer. 2000. Functional analysis of
the ClpATPase ClpA of Brucella suis, and persistence of a knockout mutant in BALB/c mice. Microbiology. 146:1605-1616.
11. Exterkate, F.A., De Jong M., G. J. C. M. De Veer, and Baankreis, R. 1992. Localization
and characterization of aminopeptidase N in Lactococcus lactis subsp. cremoris HP. Appl. Microbiol. Biotechnol. 37:46-54.
12. Gándara- Jasso, B., A. López-Merino, M. A. Rogel, and E. Mártinez-Romero. 2001.
Limited genetic diversity of Brucella spp. J. Clin. Microbiol. 39: 235-240
13. Hemmen, F., V. Weynants, T. Scarcez, J.J. Letesson and E. Saman. 1995. Cloning and sequence analysis of a newly identified Brucella abortus gene and serological evaluation of the 17-kilodalton antigen that it encodes. Clin. Diag. Lab. Immun. 2:263-267.
14. Hirsh, H.H., R. Suárez, T. Achstetter & D.H. Wolf. 1988. Aminopeptidase yscll of yeast.
Isolation of mutants and their biochemical and gnetic analysis. Eur. J. Biochem. 173: 589-598
15. Laemmli, K. 1970. Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature. 227:680-685.
16. LeVier, K., R.W. Phillips, V. K. Grippe, R. M. Roop II, & G.C. Walker. 2000. Similar requirements of a plant symbiont and a mammalian pathogen for prolonged intracellular survival. Science. 287:2492-2493.
17. Leung, K. Y., and R. M. Stevenson. 1988. Characteristics and distribution of extracellular
proteases from Aeromonas hydrophila. J. Gen. Microbiol. 134:151-160.
18. Lin J., L.G.Adams and T.A. Ficht. 1992. Characterization of the heat shock response in Brucella abortus and isolation of the genes encoding the GroE heat shock proteins.Infect. Immun.60:2425-2431
19. Maeda H., and T. Yamamoto. 1996. Pathogenic mechanism induced by microbial
proteases in microbial infections. Biol. Chem. 377:217-226.
20. McCaman M. T., and M. R. Villarejo. 1982. Structural and catalytic properties of peptidase N from Escherichia coli K-12. Arch. Biochem. Biophys. 213:384-394.
17

21. Maurizi MR. 1992. Proteases and protein degradation in Escherichia coli. Experientia. 48:178-201
22. Moreno, E., E. Stackebrandt, M. Dorsch, J. Wolters, M. Busch, and H. Mayer. 1990.
Brucella abortus 16S rRNA and lipid A reveal a phylogenetic relationship with members of the alpha-2 subdivision of the class Proteobacteria. J. Bacteriol. 172:3569-3576.
23. Sanz, Y., and F. Toldrá. 2002. Purification and characterization of an arginine
aminopeptidase from Lactobacillus sakei. Appl. Environ. Microbiol. 64:1980-1987.
24. Stabel TJ, Sha Z, Mayfield JE. 1994. Periplasmic location of Brucella abortus Cu/Zn superoxide dismutase. Vet. Microbiol.38(4):307-14
25. Texeira-Gomes A. P., A. Clockaert, G. Bézard, G. Dubray and M. S. Zingmunt. 1997.
Mapping and identification of Brucella melitensis proteins by two-dimensional electrophoresis and microsequencing. Electrophoresis. 18:156-162.
26. Texeira-Gomes A. P., A. Clockaert, and M. S. Zingmunt. 2000. Characterization of heat,
oxidative, and acid stress responses in Brucella melitensis. Infect. Immun. 68:2954-2961.
27. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretic transfer of proteins from polyacrilamide gels to nitrocellulose sheets. Procedure and some applications. Proc. Natl. Acad. Sci. 76:4350-4354
28. Woods, M. L., II, R. Bonfiglioli, Z. A. McGee, & C. Georgopoulos.1990.Synteshis of a
selected group of proteins by Neiseria gonorrheae in response to thermal stress. Infect. Inmmun. 58:719-725.
1. Se purificó una aminopeptidasa de Rhizobium tropici reveló similitudes con aminopeptidasas N putativas de Agrobacterium tumefaciens
(NP_354008), Sinorhizobium meliloti (NP_385132) y Mesorhizobium loti (NP_107963
18






![coloquio [Modo de Compatibilidade] - UFJF · Pesar cerca de 0,075 g de o-fenantrolina. Solubilizar e transferir quantitativamente para um balão volumétrico de 25,00 mL](https://img.pdfslide.tips/doc/110x75/5c4b2db493f3c34c5065d14c/coloquio-modo-de-compatibilidade-pesar-cerca-de-0075-g-de-o-fenantrolina.jpg)






















