Embed Size (px)
Citation preview

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM BOTÂNICA
DIVERSIDADE E COMPOSIÇÃO ECOLÓGICA DO CONTEÚDO PALINOLÓGICO
DE DEPÓSITOS NEOGÊNICOS (FORMAÇÃO SOLIMÕES) E HOLOCÊNICOS DA
BACIA DO SOLIMÕES.
BIANCA TACORONTE GOMES
Manaus, Amazonas
Março, 2019


i
BIANCA TACORONTE GOMES
DIVERSIDADE E COMPOSIÇÃO ECOLÓGICA DO CONTEÚDO PALINOLÓGICO
DE DEPÓSITOS NEOGÊNICOS (FORMAÇÃO SOLIMÕES) E HOLOCÊNICOS DA
BACIA DO SOLIMÕES.
ORIENTADORA: Dra. Maria Lúcia Absy
COORIENTADOR: Dr. Carlos D’Apolito Junior
Dissertação apresentada ao Instituto Nacional
de Pesquisas da Amazônia como parte dos
requisitos para obtenção do título de Mestre em
Botânica.
Manaus, Amazonas
Março, 2019

ii
BIANCA TACORONTE GOMES
DIVERSIDADE E COMPOSIÇÃO ECOLÓGICA DO CONTEÚDO PALINOLÓGICO
DE DEPÓSITOS NEOGÊNICOS (FORMAÇÃO SOLIMÕES) E HOLOCÊNICOS DA
BACIA DO SOLIMÕES.
Dissertação apresentada ao Instituto Nacional de Pesquisas da Amazônia como parte dos
requisitos para obtenção do título de Mestre em Botânica.
Aprovado em : 27/03/2019
Orientadora: Dra. Maria Lúcia Absy
Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus-AM.
Co-orientador: Dr. Carlos D’Apolito Junior
Faculdade de Geociências (FAGEO) - Universidade Federal do Mato Grosso (UFMT),
Cuiabá-MT.
Membro Titular Dr. Emílio Alberto Soares – Aprovado
Universidade Federal do Amazonas (UFAM), Manaus-AM.
Membro Titular Dra. Rosemery Rocha da Silveira- Aprovado
Universidade Federal Amazonas (UFAM), Manaus-AM.
Membro Titular Dr. Antônio Carlos Marques Souza –Aprovado
Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus-AM.

iii
G633d Gomes, Bianca Tacoronte
DIVERSIDADE E COMPOSIÇÃO ECOLÓGICA DO CONTEÚDO
PALINOLÓGICO DE DEPÓSITOS NEOGÊNICOS (FORMAÇÃO
SOLIMÕES) E HOLOCÊNICOS DA BACIA DO SOLIMÕES. /
Bianca Tacoronte Gomes; orientadora Maria Lúcia
Absy; coorientador Carlos D’Apolito
Junior . -- Manaus:[s.l], 2019.
146 f.
Dissertação (Mestrado - Programa de Pós Graduação
em Botânica) -- Coordenação do Programa de Pós-
Graduação, INPA, 2019.
1. Palinologia. 2. Amazônia ocidental. 3. .
Ecologia. 4. Formação Solimões. I. Absy, Maria
Lúcia Absy, orient. II. Junior, Carlos D”Apolito
Junior, coorient. III. Título.
CDD: 580
Sinopse:
Estudou-se a composição palinológica de sedimentos da Formação Solimões (Neógeno) e de
sedimentos recentes da Amazônia ocidental a fim de fazer uma comparação da diversidade
florística, ambientes ecológicos e deposicionais entre o período Neógeno e o atual da
Amazônia ocidental. Além disso, os palinomorfos encontrados também foram utilizados para
datar os sedimentos da Formação Solimões com base em palinoestratigrafia.
Palavras-chave: Palinologia, Amazônia ocidental, Ecologia, Formação Solimões.

iv

v
AGRADECIMENTOS
A minha família pela educação, dedicação e incentivo aos estudos. Em especial a minha
avó Valentina Tacoronte, avô Jose Martiniano Bento, mãe Fátima Tacoronte e Irmão José
Otávio Tacoronte dos Santos;
Ao Conselho Nacional de desenvolvimento Cientifico e Tecnológico (CNPq) pela bolsa
de mestrado concedida (Proc. 134058/2017-8);
Ao Laboratório de Palinologia do Instituto Nacional de Pesquisas da Amazônia (INPA)
pela infraestrutura oferecida para que pudesse realizar minha pesquisa;
Ao Programa de Pós-graduação em Botânica do INPA pelos cursos e ensinamentos
oferecidos;
A minha orientadora Maria Lúcia Absy por ter aceitado me orientar e por toda a ajuda,
ao meu coorientador Carlos D’Apolito por tudo, sem você esse trabalho não teria sido possível;
Ao Dr. Carlos Jaramillo por ceder as amostras de pólen fóssil e por me aceitar para fazer
um estágio no Smithsonian Tropical Research Institute, pela orientação e ensinamentos;
A Enrique Moreno por todos os ensinamentos sobre palinologia;
A equipe do Laboratório de Palinologia do Inpa, Marcos Gonçalves Ferreira, Alinne
Costa Cavalcanete Rezende, Mayra Nina Araújo e Alyne Daniele Pimentel pelas contribuições
no trabalho, risadas e lanchinhos;
A turma de mestrado da botânica do INPA de 2017, em especial Oscarina Prado, José
de Anchieta e Guilherme pelas conversas risadas e churrascos;
A minha melhor amiga Carolina Toledo Cavalcante que me conhece há tantos anos e
que esteve comigo durante todas as etapas dessa jornada.
Muito obrigada!


vii
“Muitos pensam que a pesquisa científica é uma
atividade puramente racional, na qual o objetivismo
lógico é o único mecanismo capaz de gerar
conhecimento. Como resultado, os cientistas são
vistos como insensíveis e limitados, um grupo de
pessoas que corrompe a beleza da Natureza ao
analisá-la matematicamente. Essa generalização,
como a maioria das generalizações, me parece
profundamente injusta, já que ela não incorpora a
motivação mais importante do cientista, o seu
fascínio pela Natureza e seus mistérios. Que outro
motivo justificaria a dedicação de toda uma vida ao
estudo dos fenômenos naturais, senão uma profunda
veneração pela sua beleza? A ciência vai muito além
da sua mera prática. Por trás das fórmulas
complicadas, das tabelas de dados experimentais e
da linguagem técnica, encontra-se uma pessoa
tentando transcender as barreiras imediatas da vida
diária, guiada por um insaciável desejo de adquirir
um nível mais profundo de conhecimento e de
realização própria. Sob esse prisma, o processo
criativo científico não é assim tão diferente do
processo criativo nas artes, isto é, um veículo de
autodescoberta que se manifesta ao tentarmos
capturar a nossa essência e lugar no Universo”.
(Marcelo Gleiser, A dança do universo).


ix
RESUMO
O presente trabalho teve por objetivos principais responder questões bioestratigráficas e
paleoecológicas sobre a paleoflora da Amazônia ocidental do período Neógeno (23 milhões de
anos Antes do Presente) através do estudo do conteúdo palinológico presente em um furo de
sondagem (1AS-15-AM) coletado da Formação Solimões (AM), Brasil (Idade Neógeno) e
também o estudo de conteúdo palinológico de sedimentos recentes (Holoceno) coletados em
diferentes subambientes deposicionais da Bacia do Solimões. Foi contado um mínimo de 300
esporos e grãos de pólen por amostra quando possível e paralelamente outros palinomorfos
presentes nas lâminas (cistos de dinoflagelados, células de algas, acritarcos, partes de
foraminíferos). Foram analisadas 51 amostras no total (30 do perfil 1AS-15-AM e 21 do
material holocênico). Após a análise taxônomica e contagem do material foram tabulados dados
de FAD (First Appearance Datum) e LAD (Last Appearance Datum) para os palinomorfos do
perfil 1-AS-15-AM a fim de datar os sedimentos pelo método da correlação gráfica. Os taxa
foram classificados de acordo com sua ecologia e também foi feito o uso de índices estatísticos
para caracterizar a diversidade dos palinomorfos. Foram encontrados 284 tipos polínicos nas
30 amostras analisadas da palinoflora fóssil do poço 1AS-15-AM, (234 de angiospermas e 50
de pteridófitas), nas 21 amostras analisadas do material recente foram encontrados 231 tipos
polínicos (202 de angiospermas e 29 de pteridófitas). Um total de 46 novas espécies são
descritas para o material fóssil, 10 novas espécies de esporos e 36 novas espécies de pólen e
através da correlação gráfica é atribuída idade de final do Mioceno inicial a início do Mioceno
tardio (16.1 a 7.1 Ma) aos sedimentos do perfil 1-AS-15-AM. Os tipos predominantes na
palinoflora fóssil foram M. franciscoi e Poaceae e tipos associados a várzea e ambientes de
planícies amazônicas indiferenciadasterras para a palinoflora holocênica. Os índices estatísticos
não evidenciaram diferenças das palinofloras fósseis e holocênicas com relação à diversidade,
mas, por outro lado, mostraram grande diferença com relação a composição. Os resultados deste
trabalho permitiram concluir que a vegetação do Noroeste da Bacia Amazônica passou por
grandes mudanças desde o período Néogeno (23 Ma) até o presente, refletindo a presença do
sistema de grandes lagos (Mioceno inferior-médio), sua extinção (Mioceno superior) e
estabelecimento de um sistema fluvial erosivo (Plioceno até período atual). Por outro lado, sua
vegetação sempre foi a de uma floresta tipicamente tropical como conhecemos atualmente.


xi
ABSTRACT
The present study have by its principal objectives to answer bioestratigraphic and paleoecologic
questions about the western Amazon paleoflora of the Neogene period (23 million years Before
Present ) through the study of palynological content present in a core (1A-15-AM) collected
from Solimões Formation (AM), Brasil (Neogene) and also the study of the palynological
content from recent sediments (Holocene) collected in different deposition subambients of
Solimões Basin. It was counted a number of 300 spores and grains of pollen per sample and
parallelly other palynomorphs present in the slides (dinoflagellate cists, algae cellule,
acritarchs, foraminifers). 51 samples were analyzed (30 from the 1AS-15-AM profile and 21 of
the holocenic material). After the taxonomic analyses and the count of the material were
tabulated datas of FAD (First Appearance Datum) and LAD (Last Appearance Datum) for the
palynomorphs present in the profile 1AS-15-AM aiming to date the sediments by the graphic
correlation method. The taxa were classified according to its ecology and also was made use of
statistical indices to characterize the palynomorph diversity. A total of 284 pollen types were
found in the 30 analyzed samples of the fossil palinoflora from the 1-AS-15-AM core (234 of
angiosperms and 50 of pteridophytes). In the 21 analyzed samples of the recent material, 231
pollen types were found (202 of angiosperms and 29 of pteridophytes). A total of 46 new
species are described for fossil material, 10 new species of spores and 36 new species of pollen
and through graphical correlation is attributed the final age of late early Miocene to early late
Miocene (16.1 to 7.1 Ma) to sediments of core 1-AS-15-AM. The predominant types in the
fossil palinoflora were M. franciscoi and Poaceae and types associated with lowland floodplains
and undifferentiated lowland amazonian environments for Holocene palinoflora. The statistical
indices evidenced a lack of difference of the fossil and Holocene palinofloras in relation to
diversity, but a great difference in relation to the composition. It was possible to conclude with
the work that the vegetation of the Northwest of the Amazon Basin underwent great changes
from the Neogene period (23 Ma) to the present, reflecting the presence of the system of large
lakes (Early-middle Miocene) its extinction (Late Miocene) and establishment of an erosive
fluvial system (Pliocene until current period). On the other hand, its vegetation has always been
that of a typically tropical forest as we know it today.


xiii
SUMÁRIO
1 INTRODUÇÃO ________________________________________________________ 17
1.1 Bioestratigrafia _______________________________________________________ 20
2 OBJETIVOS ___________________________________________________________ 23
2.1 Objetivo geral ________________________________________________________ 23
2.2 Objetivos específicos ___________________________________________________ 23
3 MATERIAIS E MÉTODOS ______________________________________________ 25
3.1 Área de estudo ________________________________________________________ 25
3.1.1 Formação Solimões ____________________________________________________ 25
3.1.2 Material fóssil ________________________________________________________ 26
3.1.3 Material recente _______________________________________________________ 28
3.2 Análise palinológica ____________________________________________________ 29
3.3 Análise bioestratigráfica ________________________________________________ 29
3.4 Análise numérica ______________________________________________________ 30
4 RESULTADOS ________________________________________________________ 33
4.1 Análise palinológica ____________________________________________________ 33
4.1.1 Palinologia sistemática _________________________________________________ 33
4.1.2 Esporos _____________________________________________________________ 34
4.1.2.1 Monolete ___________________________________________________________ 34
4.1.2.2 Trilete _____________________________________________________________ 34
4.1.3 Polen _______________________________________________________________ 40
4.1.3.1 Inaperturado ________________________________________________________ 40
4.1.3.2 Monosulcado ________________________________________________________ 40
4.1.3.3 Tricolpado __________________________________________________________ 42
4.1.3.4 Tricolporado ________________________________________________________ 44
4.1.3.5 Triporado __________________________________________________________ 56
4.1.3.6 Stephanocolpado _____________________________________________________ 57
4.1.3.7 Periporado _________________________________________________________ 58
4.1.3.8 Stephanoporado _____________________________________________________ 59
4.2 Bioestratigrafia ______________________________________________________ 111
4.3 Análise numérica _____________________________________________________ 112
4.3.1 Análise de similaridade ________________________________________________ 112
4.3.2 Análise de riqueza e diversidade _________________________________________ 113

xiv
4.3.3 Ordenação das comunidades ___________________________________________ 114
4.3.4 Análise de composição das amostras recentes ______________________________ 115
4.3.5 Análise ecológica ____________________________________________________ 116
5 DISCUSSÃO _________________________________________________________ 135
6 CONCLUSÃO ________________________________________________________ 137
REFERÊNCIAS _________________________________________________________ 139

xv
LISTA DE ILUSTRAÇÕES
Quadro 1 - Quadro comparativo do zoneamento palinoestratigráfico do Neógeno estabelecido
para o norte da América do Sul ................................................................................................ 20
Figura 1 - Mapa do Brasil (1) e extensão da Formação Solimões (2) com as linhas
de Isópacas da Formação .......................................................................................................... 25
Figura 2 - Perfil 1-AS-15-AM de 0 a 262.8 metros de profundidade destacando as amostras que
foram analisadas no presente trabalho ...................................................................................... 27
Figura 3 - Mapa destacando a área de coleta (a). Poço 1AS-15-AM e perfis de sedimento
superficial (b) ............................................................................................................................ 28
Figura 4 - Prancha 1 ................................................................................................................. 62
Figura 5 - Prancha 2 ................................................................................................................. 64
Figura 6 - Prancha 3 ................................................................................................................. 66
Figura 7 - Prancha 4 ................................................................................................................. 68
Figura 8 - Prancha 5 ................................................................................................................. 70
Figura 9 - Prancha 6 ................................................................................................................. 72
Figura 10 - Prancha 7 ............................................................................................................... 74
Figura 11 - Prancha 8 ............................................................................................................... 76
Figura 12 - Prancha 9 ............................................................................................................... 78
Figura 13 - Prancha 10 ............................................................................................................. 80
Figura 14 - Prancha 11 ............................................................................................................. 82
Figura 15 - Prancha 12 ............................................................................................................. 84
Figura 16 - Prancha 13 ............................................................................................................. 86
Figura 17 - Prancha 14 ............................................................................................................. 88
Figura 18 - Prancha 15 ............................................................................................................. 90
Figura 19 - Prancha 16 ............................................................................................................. 92
Figura 20 - Prancha 17 ............................................................................................................. 94
Figura 21 - Prancha 18 ............................................................................................................. 96
Figura 22 - Prancha 19 ............................................................................................................. 98
Figura 23 - Prancha 20 ........................................................................................................... 100
Figura 24 - Prancha 21 ........................................................................................................... 102
Figura 25 - Prancha 22 ........................................................................................................... 104

xvi
Figura 26 - Prancha 23 ........................................................................................................... 106
Figura 27 - Prancha 24 ........................................................................................................... 108
Figura 28 - Prancha 25 ........................................................................................................... 110
Figura 29 - Distribuição dos principais marcadores palinoestratigráficos do perfil 1AS-15-AM
................................................................................................................................................ 111
Figura 30 - Correlação gráfica do perfil 1-AS-15-AM com o perfil 1-AS-105-AM ............. 112
Tabela 1 - Índices de similaridade comparando as palinofloras fóssil e holocênica ............. 113
Figura 31 - Riqueza rarefeita (a) e diversidade Shannon (b) das palinofloras fóssil e recente
................................................................................................................................................ 113
Figura 32 - Curva de acumulação de espécies paras as amostras fósseis (linhas cinzas) e
recentes (linhas pretas). Intervalo de confiança mostrado de 95% ........................................ 114
Figura 33 - Análise nMDS para as palinofloras fóssil e recente ............................................ 115
Figura 34 - Subambientes de coleta comparados com a diversidade (a) e scores nMDS do eixo
1 (b) ........................................................................................................................................ 116
Tabela 2 - Lista de afinidades botânicas e ecológicas para pólen e esporos deste estudo.
Gimnospermas (G), Angiospérmicas (A) e Pteridófitas (P); Afinidades ecológicas: florestas
secas (fs), exógeno (ex), hidrosseral (hi), terras baixas (tb), lianas de terras baixas (li tb),
manguezal (ma), montano (mo), ripário (ri), terrestre (te) , várzea (va), campinarana (ca) .. 117
Figura 35 - Composição ecológica das palinofloras fóssil e recente ..................................... 133

17
1 INTRODUÇÃO
A Bacia hidrográfica amazônica constitui-se de uma vasta planície central ladeada por
planaltos, sendo limitada a oeste pela Cordilheira dos Andes, ao norte pelo Escudo das Guianas
e ao sul pelo Escudo Brasileiro, desaguando no ocêano Atlântico (Victoria et al., 2010). O
sistema fluvial Solimões-Amazonas caracteriza-se por leve predominância de processos
deposicionais com relação a processos erosivos com elevado padrão de estabilidade dos canais
e um estilo fluvial anabranching (Passos e Soares, 2017).
A floresta que recobre a bacia é reconhecida como sendo a mais extensa floresta tropical
do mundo (Thomas, 1999). Possui 14.003 espécies de plantas com sementes em 1.788 gêneros,
sendo 45% desses com mais ou equivalente a 10cm a altura do peito (DAP) e os outros 55%
constituídos por arbustos, árvores pequenas, lianas e ervas. As famílias mais ricas em espécies,
em ordem decrescente, são: Leguminosae, Rubiaceae, Orchidaceae, Melastomataceae,
Araceae, Poaceae e Euphorbiaceae (Cardoso et al., 2017).
Sua paisagem é caracterizada por grande heterogeneidade e os principais tipos de
vegetação que ocorrem nela, são: floresta de terra firme (presente em áreas não alagadas);
floresta de várzea (planícies de inundação de rios de água branca); floresta de igapó (áreas muito
encharcadas, com alagação permanente ou não, águas paradas ou quase paradas); manguezais
(região costeira); campos de várzea (áreas sob influência de alagações procedentes de rios de
águas brancas); campos de terra firme (áreas de vegetação herbácea entre áreas de vegetação
de terra firme); campinas (solo arenoso); vegetação serrana (Escudo das Guianas); e vegetação
de restinga (praias costeiras e dunas do litoral do Pará, Amapá e Maranhão) (Braga, 1979).
Devido à grande importância da Amazônia para o clima mundial, com sua contribuição
ao suprimento de oxigênio (cerca de 20% da produção global), nutrientes para o oceano e
produção de biomassa (cerca de 10% da produção global), o conhecimento dos processos
históricos que levaram ao desenvolvimento de sua biodiversidade, que ainda é pequeno
comparado com suas enormes proporções, é muito importante para sua conservação (Hoorn et
al., 2010).
Uma das explicações postuladas para o surgimento de sua biodiversidade é a Teoria dos
Refúgios, formulada por Haffer (1969), em que fases de contração e fragmentação da floresta
tropical amazônica, durante os períodos glaciais do Quaternário (últimos 2.59 milhões de anos
(Ma)), teriam resultado na formação de refúgios florestais onde populações de plantas e animais
isoladas poderiam se diferenciar em distintas espécies.

18
Porém, evidências posteriores postularam outras origens históricas de diversidade para
a Amazônia, como o estudo de Hoorn et al. (2010) que cita que os mecanismos de diversificação
mais importantes ocorreram antes do Quaternário. Ricklefs e Renner (2012) afirmam que a
floresta tropical dos Neotropicos permaneceu parecida com a atual por todo o Cenozóico (65.5
a 5.33 Ma). Wing et al. (2009) indicam que já no Paleoceno (65.5 a 55.8 Ma) a composição
florística no norte da América do Sul era basicamente a mesma que a atual (embora menos
diversa). Os estudos de Jaramillo et al. (2006; 2010a), evidenciam eventos de diversificação
durante o Máximo Climático do Paleoceno tardio-Eoceno (56.3 Ma), com estimativas de
diversidade de pólen maiores que as atuais. Com relação ao Neógeno (23.03 a 2.59 Ma) não
existe uma boa estimativa de comparação de diversidade e composição ecológica (Jaramillo et
al., 2010a).
Especial destaque se dá ao período Neógeno para a Amazônia por ter sido um período
decisivo no estabelecimento da geografia e composição biótica como é conhecida hoje (Hoorn
et al., 2010). No Mioceno inicial (23.03 a 15.97 Ma), o sistema de rios existente na Amazônia
corria em direção oeste, ao contrário da direção atual (Caputo e Soares, 2016). Com a
intensificação da orogenia andina, um sistema de grandes lagos e pântanos se desenvolveram
na Amazônia ocidental (Wesselingh et al., 2002). Esse ambiente aquático denominado
“Sistema Pebas” foi colonizado por moluscos e ostracodes, muitos dos quais eram endêmicos
(Wesselingh, 2006). Incursões marinhas durante esse período são comprovadas pela existência
de microfósseis de origem marinha, como foraminíferos, pólen de plantas de mangue (indicador
de ambiente com influência marinha) e dinoflagelados (Hoorn et al., 2010; Jaramillo et al.,
2017). No Mioceno tardio (11.63 a 5.33 Ma) e Plioceno (5.33 a 2.59 Ma), o sistema fluvial da
Amazônia foi estabelecido como é conhecido atualmente, com o Rio Amazonas nascendo na
bacia sedimentar dos Andes e correndo em direção ao Oceano Atlântico (Caputo e Soares,
2016).
Para conhecer a biodiversidade florística e ambientes deposicionais do Neógeno da
Amazônia ocidental, estudos palinológicos são muito interessantes, dado que palinomorfos são
encontrados em rochas sedimentares de todas as idades de cerca de dois bilhões de anos até o
presente, em todos os tipos de ambientes, de água doce a sedimentos marinhos. Palinomorfos
podem também ser sensíveis indicadores dos processos de sedimentação e da proveniência
sedimentar. Enquanto a origem de alguns palinomorfos (dinoflagelados) é primariamente
marinha, outros palinomorfos, como esporos e pólen, indicam a presença de um ambiente
continental e revelam muito sobre o clima e vegetação dos paleoambientes (Traverse, 2007).

19
Para o Neógeno do Norte da América do Sul, os trabalhos pioneiros de Van der Hammen
(1954) (Colômbia), Germeraad et al. (1968) (Colômbia e Venezuela), Regali et al. (1974)
(Brasil), Lorente (1986) (Venezuela), Muller et al. (1987) (Colômbia, Venezuela, Brasil) e
Hoorn (1993; 1994a) (Brasil e Colômbia) contribuíram para o conhecimento da palinoflora da
época e o estabelecimento de marcadores bioestratigráficos. Dentre os estudos mais recentes
para a região, destacam-se: Silva-Caminha et al. (2010), Kachinasz e Silva-Caminha (2016),
Silveira e Souza (2015; 2016; 2017); D’Apolito (2016), Leite et al. (2017), Jaramillo et al.
(2017) e Linhares et al. (2017), que estudaram sedimentos provenientes da Formação Solimões
(Neógeno).
Os estudos apresentam resultados conflitantes sobre os ambientes deposicionais do
período Neógeno. No trabalho de Silva-Caminha et al. (2010), não foi encontrada nenhuma
evidência de incursão marinha ou sistema de grandes lagos. Já no trabalho de Leite et al. (2017)
foram encontradas evidências de duas fases deposicionais, uma correspondente ao sistema
Pebas de grandes lagos e outra correspondente a um sistema fluvial. No trabalho de Jaramillo
et al. (2017) foram observados dois eventos de incursões marinhas, um no Mioceno tardio e
outro no Mioceno médio. Linhares et al. (2017) também apontaram a existência de incursões
marinhas, porém, além das inferidas por Jaramillo et al. (2017), apontam a presença de uma
incursão no Mioceno inicial. Silveira e Souza (2015; 2016; 2017) indicam ambiente
deposicional de lagoas de água doce, lagos e/ou rios lentos, possivelmente em condições
eutróficas e a possibilidade de influência marinha no Plioceno.
Com relação à composição florística, o trabalho de Silva-Caminha et al. (2010) relata a
presença de táxons indicadores de florestas de várzea (Malvaceae-Bombacoideae, Mauritia,
Melastomataceae, Malpighiaceae, Humiriaceae, Araceae, Euphorbiaceae, entre outros) e de
ambientes de lagoas e rios (Pediastrum, Botryococcus, Salviniaceae, Ceratopteris). No trabalho
de Leite et al. (2017), houve a dominância de esporos.Os tipos polínicos encontrados indicaram
pântano (Mauritia), florestas de terra firme (Anacardiaceae, Crudia/Macrolobium, Pachira
aquatica) e florestas de terra firme associadas com rios (Amanoa), além de Pediastrum sp.
(indicador de lago). O trabalho de D’Apolito (2016) indicou a presença de dinocistos,
foraminíferos e pólen de mangue, indicadores de ambientes marinhos (Rhizophora +
Pelliciera), além das algas Pediastrum sp. e Botryococcus sp., indicadoras de ambientes
lacustres. Foi constatada a dominância de palmeiras, herbáceas, plantas aquáticas e uma rica
assembleia de árvores, incluindo táxons de florestas sazonalmente inundáveis (Malvaceae-

20
Bombacoideae), bem como grande quantidade de esporos indicando florestas de várzea,
pântanos e florestas de terra firme.
1.1 Bioestratigrafia
Os trabalhos de Germeraad et al. (1968), Lorente (1986), Muller et al. (1987) e Hoorn
(1993; 1994a) estabeleceram zonas palinoestratigráficas que foram usadas para datar os
sedimentos da Formação Solimões nas últimas décadas (Quadro 1).
Quadro 1 - Quadro comparativo do zoneamento palinoestratigráfico do Neógeno estabelecido
para o norte da América do Sul
ÉPOCA
AUTOR
Germeraad et al.
(1968) Muller et al. 1987 Lorente (1986) Hoorn (1993)
Jaramillo et al.
2011
PLIOCENO
(5.33 -
2.59 Ma)
Echitricolporites
mcneillyi
Echitricolporites
mcneillyi Fenestrites
longispinosus
Cyatheacidites
annulatus
(T-17)
MIOCENO
(23.03 -
5.33 Ma)
TARDIO
(11.63 -
5.33 Ma)
Pachidermites
diederixii
Echitricolporites
spinosus
Asteraceae
Fenestrites
spinosus
(T-16)
Grimsdalea
Magnaclavata
Grimsdalea
Magnaclavata
MÉDIO
(15.97 -
11.63 Ma)
Crassoretitriletes
vanraadshooveni
Crassoretitriletes
vanraadshooveni
Crassoretitriletes
vanraadshooveni
Crassoretitrile-
tes
vanraadshooveni
(T-15)
Grimsdalea
Magnaclavata
(T-14)
NICIAL
(23.03 -
15.97 Ma)
G. Magnaclavata
E. maristellae/
Psiladiporites
Psiladiporites Psiladiporites/
Crototricolporites
M. vanderhamenii Verrutricolporites Retitricolporites
Psiladiporites
Verrutricolporites/
Cicatricosisporites Verrutricolporites
V. rotundiporus
E. barbeitoensis
Echitricolpori-
tes
Maristellae
(T-13)
J. saemrogiformis
Horniella
lunerensis
(T-12)
Fonte: Da autora (2019), adaptado de Germeraad et al. (1968), Muller et al. (1987), Lorente (1986), Hoorn (1993)
e Jaramillo et al. (2011).
Silva-Caminha et al. (2010) atribuíram aos sedimentos da Formação Solimões idades
de Mioceno tardio (11.63 a 5.33 Ma) a Plioceno (5.33 a 2.59 Ma), com base na presença dos

21
marcadores Cichoreacidites longispinosus e Ladakhipollenites caribbiensis, que localizam os
sedimentos na zona F. longispinosus (Lorente, 1986). O trabalho de Leite et al. (2017), através
da presença dos marcadores Crassoretitriletes vanraadshoovenii, Fenestrites spinosus,
Cichoreacidites longispinosus, Ladakhipollenites? caribbiensis e Echitricolporites mcneillyi,
reconheceu 4 biozonas de acordo com Lorente (1986): Crassoretitriletes, Asteraceae,
Psilatricolporites caribbiensis e Echitricolporites–Alnipollenites, estabelecendo idade de
Mioceno médio (15.97 a 11.63 Ma) a Plioceno para a formação. Kachniasz e Silva-Caminha
(2016) através da presença de espécies índice nos sedimentos, como Grimsdalea magnaclavata,
Ladakhipollenites? caribbiensis e Echitricolporites mcneillyi, incluíram suas seções dentro das
subzonas 8 Psilatricolporites caribbiensis e Echitricolporites-Alnipollenites (Lorente, 1986),
indicando também idade Mioceno tardio-Plioceno.
Estes zoneamentos sofrem com a falta de maior detalhamento e calibração das zonas.
Jaramillo et al. (2011) propôs um zoneamento para os sedimentos cenozóicos das Bacias e
sopés dos Lanos colômbianos, utilizando dados de 70 seções estratigráficas e 18 pontos de
calibração, incluindo isótopos de carbono, foraminíferos e magnetoestratigrafia. Foi produzida
uma seção composta pelo método da correlação gráfica (Shaw 1964; Edwards 1989) e
estabelecidas 18 zonas palinoestratigráficas relacionadas a escala geológica, sendo 6 zonas (T-
12, T-13, T-14, T-15, T-16 e T-17) compreendidas no período Neógeno. O poço 1-AS-105-AM
da Formação Solimões (Jaramillo et al., 2017) foi datado por correlação gráfica com a seção
composta produzida por Jaramillo et al. (2011), obtendo idade de final do Mioceno inicial a
inicio do Mioceno tardio (18.7 a 10.7 Ma) e permitindo uma melhor estimativa de idade para a
Formação.
Apesar de os trabalhos citados apresentarem importantes informações sobre os
processos geológicos que ocorreram no Neógeno da Bacia Amazônica, ainda falta ampliar o
conhecimento de como esses processos influenciaram na diversidade e composição ecológica
florística da época, além de se obter mais dados bioestratigráficos para o período. Objetivos aos
quais este trabalho se propõe.


23
2 OBJETIVOS
2.1 Objetivo geral
Responder questões bioestratigráficas e paleoecológicas sobre a flora da Amazônia
ocidental do período Neógeno através do estudo do conteúdo palinológico presente em um furo
de sondagem (1AS-15-AM) coletado da Formação Solimões (AM), Brasil (Idade Neógeno) e
também o estudo de conteúdo palinológico de sedimentos recentes (Holoceno) coletados em
diferentes subambientes deposicionais da Bacia do Solimões.
2.2 Objetivos específicos
a) Datar o furo de sondagem 1AS-15-AM, usando marcadores palinoestratigráficos do
Neógeno do norte da América do Sul;
b) Estabelecer a diversidade e composição ecológica da palinoflora do furo de
sondagem 1AS-15-AM;
c) Estabelecer diversidade e composição ecológica da palinoflora do material
holocênico;
d) Comparar as diversidades e a composição ecológica do furo de sondagem 1-AS-15-
AM com a do material recente.


25
3 MATERIAIS E MÉTODOS
3.1 Área de estudo
3.1.1 Formação Solimões
Na Amazônia sul ocidental brasileira se encontra a Bacia do Solimões, sendo essa uma
bacia sedimentar intracratônica, com cerca de 500.000 km2 de área, quase toda contida no
estado do Amazonas (Figura 1). A oeste é limitada pelo Arco de Iquitos e a leste pelo Arco de
Purus, sendo que, ao norte e ao sul, é limitada pelas rochas proterozóicas dos escudos das
Guianas e do Brasil central, respectivamente. A bacia pode ser dividida em duas sequências de
primeira ordem, uma paleozóica e outra cretácea e mesocenozóica, que se sobrepõe
discordantemente à primeira sequência e que predomina em sua superfície (Maia e Marmos,
2010).
Figura 1 - Mapa do Brasil (1) e extensão da Formação destacada em bege (2) com as linhas
de Isópacas da Formação
Fonte: Adaptado de D’Apolito (2016) e localização dos furos da CPRM estudados por Latrubesse et al. (2010) (a),
Leite et al. (2016) (b), Kachniasz e Silva-Caminha et al. (2016) (c) (k), Hoorn (1993) (d), Linhares et al. (2017)
(e), (f), Jaramillo et al. (2017) (g), este estudo (h), Silva-Caminha et al. (2010) (i) (j).
Depositada sobre a Bacia do Solimões e do Acre, encontra-se a Formação Solimões,
composta por arenitos, siltitos e argilitos com camadas de conchas e linhito, muito rica em
fósseis vegetais e animais (Maia et al., 1977; Eiras et al., 1994, B. Geoci. Petrobras, 2007).

26
Associações esporopolínicas sugerem idades entre Mioceno e Plioceno (Cruz, 1984; Hoorn,
1994a; Silva-Caminha et al., 2010; Leite et al., 2017; Jaramillo et al., 2017). É sugerido um
ambiente de deposição fluviolacustre e essa unidade forma uma cunha sedimentar desde o Arco
de Purus até as bacias subandinas, atingindo mais de 2000 metros de espessura (Maia et al.,
1977; Maia e Marmos, 2010).
3.1.2 Material fóssil
O furo de sondagem 1AS-15-AM (04°01’S/69°29’O) (Figura 1) se localiza próximo à
cidade de Tabatinga (Amazonas, Brasil), na bacia do alto Solimões, e foi perfurado pela
Companhia de Pesquisa de Recursos Minerais do Brasil (CPRM). O furo está totalmente
localizado na Formação Solimões e é composto, em sua maioria, por argilitos, arenitos e siltitos
com presenças pontuais de calcário e linhito segundo os critérios de Tucker 2003. O perfil foi
construído utilizando o pacote SDAR (Ortiz et al., 2015) do software R. (Figura 2). Foram
processadas e analisadas 30 amostras quanto ao conteúdo palinológico (pólen e esporos). As
amostras palinológicas foram selecionadas com um intervalo aproximado de 10 metros entre
cada uma, sendo duas delas em amostras com litologia de arenito, onze em siltito e dezesseis
em argilito. O processamento seguiu o procedimento padrão para palinologia: desagregação
das amostras com martelo e separação de 10cm3 de sedimento, eliminação dos carbonatos com
ácido clorídrico (HCL) concentrado por um período mínimo de 12 horas; em seguida,
eliminação dos silicatos com ácido fluorídrico (HF) concentrado, por um período mínimo de
24 horas (Wood et al., 1996). Após estas etapas, o material foi lavado com água destilada para
neutralizar a ação dos ácidos, sendo, após, peneirado em malhas de 250µm e 10µm para
eliminação dos resíduos orgânicos grosseiros e muito finos, respectivamente. Depois, a fração
<10μm foi desagregada em banho de ultrassom e a porção de matéria orgânica menos densa
recuperada. Esse resíduo foi limpo em banho de ultrassom por alguns segundos e concentrado
em uma centrífuga. As lâminas foram montadas com a resina Epoxy (Sivaguru et al., 2016).
Todo processamento foi realizado pela Paleoflora Ltda., Colômbia.

27
Figura 2 - Perfil 1-AS-15-AM de 0 a 262.8 metros de profundidade destacando as amostras que foram
analisadas no presente trabalho
Fonte: Da autora (2019), produzido utilizando o software R.

28
3.1.3 Material recente
Foram coletados sete perfis de sedimentos superficiais de cerca de 60cm cada em
ambientes de sedimentação recente, sendo os pontos L1, L2, L3 e L6 coletados em subambiente
deposicional de várzea, os pontos L5 e L8 em margem de lago e o ponto L7 em pântano de
Mauritia flexuosa L. (Figura 3). Os locais de coleta foram próximos às cidades de Atalaia do
Norte, Benjamim Constant e Tabatinga (Figura 3). A área de coleta é caracterizada por
ambientes de deposição fluvial (várzea), lacustre e de terra firme. Ocorre na região, uma floresta
tropical densa (IBGE, 1977). Para cada perfil, foram selecionadas 3 amostras obtidas a cada
10 cm, totalizando 21 amostras. O processamento do material foi feito baseado nas seguintes
fases: tratamento com potassa cáustica (KOH) diluída à 10% (Faegri e Iversen, 1966) para
retirada de ácidos húmicos, filtragem em malha de 250 µm para retirar os resíduos mais
grosseiros, acetólise (Erdtman, 1952) para retirada do conteúdo protoplasmático dos
palinomorfos e separação gravitativa com líquido pesado, usando-se uma mistura de
bromofórmio e álcool etílico em uma proporção de 2:1 (Kummel e Raup, 1965). Após a
preparação, a montagem das lâminas foi feita em gelatina glicerinada e selagem com parafina.
Esse processamento foi realizado no Laboratório de Palinologia do Instituto Nacional de
Pesquisas da Amazônia (INPA).
Figura 3 - Mapa destacando a área de coleta (a). Poço 1AS-15-AM e perfis de sedimento
superficial (b)
Fonte: Da autora (2019), produzido utilizando o software R.

29
3.2 Análise palinológica
As lâminas de pólen foram analisadas no Laboratório de Palinologia do INPA utilizando
microscópio óptico Primo Star (Zeiss) nos aumentos de 40X e 100X e os tipos polínicos
fotografados com uma câmera Axiocam IC em aumento de 100X. Para cada amostra, foi feita
a contagem de um mínimo de 300 esporos e grãos de pólen. A identificação taxonômica dos
tipos polínicos encontrados no material do perfil 1AS-15-AM foi feita com a consulta de
trabalhos sobre palinologia do Neógeno para o norte da América do Sul (Germeraad et al.,
1968; Lorente, 1986; Muller et al., 1987; Hoorn, 1994a; Silva-Caminha et al., 2010; D’Apolito,
2016) e da base de dados eletrônica de Jaramillo e Rueda (2018), que possui informações
atualizadas sobre pólens e esporos do período Cretáceo e Cenozoico para a região norte da
América do Sul.
A identificação taxonômica dos tipos polínicos encontrados em sedimentos recentes foi
feita por consulta à Palinoteca do Laboratório de Palinologia do INPA e aos trabalhos de Absy
(1979), Roubik e Moreno (1991), Colinvaux et al. (1999) e Lorente et al. (2017). Os
palinomorfos recentes também receberam nomes de acordo com a taxonomia de polinomorfos
fósseis, igualmente ao material do perfil 1AS-15-AM, a fim de possibilitar a comparação direta
entre Mioceno e recente.
3.3 Análise bioestratigráfica
Para inferir a idade relativa dos sedimentos presentes no perfil 1AS-15 AM, foram
tabulados dados de FAD (First Appearance Datum - dado de primeira aparição), LAD (Last
Appearance Datum - dado de última aparição), FCO (First consistent occurrence-primeira
aparição consistente) e ACME (peak of abundance zone- zona aonde ocorre a maior abundância
de um taxon) para os palinomorfos encontrados. Estes eventos foram comparados com os
mesmos eventos do furo de sondagem 105AM (Jaramillo et al., 2017) utilizando o método da
Correlação Gráfica (Shaw, 1964; Edwards, 1989). Nesse método, os eventos da seção de
referência (neste cado o furo 105AM) são correlacionados com os eventos do furo 15AM e uma
linha de correlação (LOC) é criada manualmente. Dado que os eventos do furo 105-AM já tem
calibração cronológica referente ao biozoneamento de Jaramillo et al. (2011) para os Llanos
colombianos, idades relativas puderam ser sugeridas para o poço aqui estudado.

30
3.4 Análise numérica
3.4.1 Análise de similaridade
Para comparar a similaridade de composição de espécies das palinofloras, foi utilizado
o índice de Jaccard. Esse índice varia de 0 a 1 e quanto mais próximo de 1 mais as amostras são
diferentes entre si. Para comparar a similaridade de abundância de espécies das palinofloras,
foram utilizados os índices de distância euclidiana (que mostra a distância em números
absolutos) e o de Bray-Curtis (que varia de 0 a 1). As análises foram realizadas no programa R
(R Development Core Team, 2018) com o pacote Vegan (Oksanen et al., 2017).
3.4.2 Análise de diversidade
Foram calculadas as riquezas rarefeitas das palinofloras utilizando o método da sample
coverage ou cobertura amostral (Chao e Jost, 2012) e depois comparadas entre si (test-t), essa
análise foi feita no programa R, utilizando o pacote iNEXT (Chao et al., 2018). Também foi
computada a diversidade das palinofloras (Índice de Shannon) e esta foi comparada entre as
palinofloras no geral (teste-t) e separando a palinoflora fóssil de acordo com sua litologia
(arenito, argilito, siltito e linhito) (ANOVA). Por fim, foi feita a curva de acumulação de
espécies para ambas as palinofloras. Todas essas análises foram feitas no programa R utilizando
o pacote Vegan (Oksanen et al., 2017).
3.4.3 Ordenação das comunidades
Foi feito um escalonamento multidimensional não métrico (nMDS) a fim de analisar
como as comunidades das palinofloras estão ordenadas com relação a sua composição e uma
análise de espécie indicadora (Indval) para identificar as espécies mais importantes que
explicam essa ordenação. As análises foram feitas utilizando o programa R e os pacotes Vegan
e Labdsv (Roberts, 2016). Os scores do eixo 1 da análise nMDS foram comparados entre as
amostras holocênicas e as amostras fósseis separadas de acordo com sua litologia (arenito,
argilito, siltito e linhito) (ANOVA).

31
3.4.4 Análise de composição das amostras holocênicas
Foi testado se existe diferença significativa nas diversidades e scores nMDS das
amostras holocênicas com relação aos três subambientes deposicionais de onde estas provêm
(várzea, pântano e lago). Para isso, foi utilizado o teste ANOVA usando o programa R.
3.4.5 Análise de afinidades ecológicas
Para as palinofloras fóssil e holocênica, foi feita uma lista de afinidades botânicas para
os tipos polínicos encontrados, com base nos trabalhos de D’Apolito (2016) e Jaramillo et al.
(2010a). Os tipos foram classificados quanto à ecologia (ervas, ervas aquáticas, várzea/ripária,
árvores de terras baixas amazônicas indiferenciadas, indeterminados e esporos) e a proporção
de cada grupo ecológico foi comparada entre as palinofloras. Também foram comparadas
separadamente as proporções dos tipos polínicos de Mauritiidites franciscoi (=Mauritia spp.) e
Poaceae.


33
4 RESULTADOS
4.1 Análise palinológica
Foram encontrados 284 tipos polínicos nas 30 amostras analisadas da palinoflora fóssil
do poço 1AS-15-AM, com 234 de angiospermas e 50 de pteridófitas, com 39 famílias (33 de
angiospermas e 6 de pteridófitas) e 43 gêneros identificados (35 de angiospermas e 8 de
pteridófitas). Nas 21 amostras analisadas do material recente foram encontrados 231 tipos
polínicos (202 de angiospermas e 29 de pteridófitas) com 51 famílias (46 de angiospermas e 5
de pteridófitas) e 68 gêneros (63 de angiospermas e 5 de pteridófitas) identificados. No total,
somando as espécies encontradas em ambas as palinofloras, foram encontrados 463 tipos
polínicos, 62 famílias e 89 gêneros, sendo que, em comum entre a palinoflora fóssil e a recente,
foram encontrados 52 tipos (11,23%), 28 famílias (45,16%) e 22 gêneros (24,71%).
4.1.1 Palinologia sistemática
A palinologia do perfil 1AS-15-AM é descrita nesta seção e ilustrada. E dada a
descrição morfológica para 47 novas formas (11 esporos e 36 polens). Espécies formalmente
descritas na literatura foram ilustradas, mas não descritas. As descrições das novas espécies
foram organizadas em grupos morfológicos (por exemplo, Monoletes, Triletes, Monocolpado,
Tricolpado, etc.) e depois alfabeticamente dentro de cada seção. As espécies novas foram
comparadas a espécies já existentes utilizando a literatura e a base morfológica de Jaramillo e
Rueda (2018). A terminologia usada nas descrições está baseada em Punt et al. (2007), com
algumas modificações de Jaramillo e Dilcher (2001). Todos os grãos foram localizados
utilizando o sistema de coordenadas England Finder (EF).
Para os esporos a seguinte terminologia foi utilizada:
a) TLI se refere ao índice trilete, que é ‘comprimento do radius/(diâmetro do
esporo/2)’;
b) MLI se refere ao índice monolete, que é ‘comprimento da lesura/comprimento do
esporo’.
Para os grãos de pólen os termos usados são:

34
a) Cpi que é calculado na vista equatorial, sendo ‘comprimento do colpo/diâmetro
polar’;
b) Cei que é calculado na vista polar, sendo ‘comprimento do colpo/ diâmetro
equatorial’.
4.1.2 Esporos
4.1.2.1 Monolete
Gênero: Reticulosporis Krtuzsch (1959).
Espécie tipo: Reticulosporis miocenicus (Selling) Krutzsch (1959).
Reticulosporis irregularis sp. nov.
Prancha 1, figuras 8-9.
Holótipo: Prancha 1, figura 8, amostra 22690, EF V-5
Parátipo: Prancha 1, figura 9, amostra 22713, EF J44
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, reniforme; monolete, curvatura ausente, lesura
distinta, reta, extremidades pontudas, lesura com 24μm de comprimento, MLI 0.62, margem
indistinta, comissura indistinta, intexina com 1μm de espessura, perina com 2.5μm de espessura
na face proximal e 1μm de espessura na face distal. Reticulado, heterobrocado, lumina com 4-
7μm de comprimento, muri com 1.5μm de comprimento e 1μm de espessura, reticulo formado
pela perina.
Dimensões: Diâmetro equatorial 24-(24.5)-25μm; diâmetro polar 35-(38.5)-42μm;
polar/equatorial 1.68; nm=2, no=2.
Observações: Reticulosporis foveolatus (Pierce, 1961) Skarby (1964) e Reticulosporis
cretaceous var. africanus (Kedves, 1995) possuem luminas menores e Reticulosporis diversus
(D’Apolito et al., in prep) tem os luminas em menor número.
Derivação do nome: Por conta de seu lumina irregular.
4.1.2.2 Trilete
Gênero: Baculatisporites Thomson et al. (1953).
Espécie tipo: Baculastisporites primarius (Wolfe, 1934) Thomson et al. (1953)

35
Baculatisporites indecisus sp. nov.
Prancha 2, figuras 2-3.
Holótipo: Prancha 2, figuras 2-3, amostra 22749, EF K-38-2
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, triangular-obtuso-convexo; trilete, curvatura
ausente, lesura distinta, levemente ondulada, extremidades pontudas, raio longo, com 12μm,
TLI 1, margem distinta, muito fina, 0.5μm de espessura, intexina com 0.5μm de espessura.
Baculado/equinado na face proximal, bacula com 1μm de comprimento; 0.5μm de largura,
equina com 1μm de comprimento, 1μm de largura na base e 0.5μm de largura no topo, baculas
e equinas distribuídas aleatoriamente, separadas 2μm. Laevigado na face distal.
Dimensões: Comprimento do diâmetro equatorial 24μm; Largura do diâmetro equatorial 20μm,
comprimento / largura 1.2; nm=1, no=1.
Observações: Neoraistrickia dubiosa D’Apolito et al., in prep é apenas baculado, com baculas
distribuídas por todo o esporo.
Derivação do nome: Pela presença de equinas e baculas ao mesmo tempo.
Gênero: Camarozonosporites Pant ex Potonié (1956), emend. Klaus (1960).
Espécie tipo: Camarozonosporites cretaceous (Weyland and Krieger, 1953; Potonié, 1956).
Camarozonosporites ondulatus sp. nov.
Prancha 2, figuras 6-7.
Holótipo: Prancha 2, figuras 6-7, amostra 23528, EF P-41
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, triangular-obtuso-concavo; trilete, curvatura
ausente, lesura distinta, reta, extremidades pontudas, raio curto, com 18μm, TLI 0.72, margem
indistinta, comissura distinta, intexina com 1μm de espessura. Laevigado na face proximal,
verrugado na face distal, verruga com 1,5μm de largura, separada 1μm. Ao redor do equador,
verrugas se fundem e formam uma crassitude interradial.
Dimensões: Comprimento do diâmetro equatorial 25μm; Largura do diâmetro equatorial 25μm,
comprimento / largura=1; nm=1, no=1.
Observações: Camarozonosporites trilobatus (D’Apolito et al., in prep) possui verrugas em
toda sua superfície.
Derivação do nome: Por conta da crassitude inter-radial ser ondulada.

36
Camarozonosporites segmentatus sp. nov.
Prancha 2, figuras 8-9.
Holótipo: Prancha 2, figuras 8-9, amostra 22666, EF T-17-4
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, triangular-obtuso-reto; curvatura ausente, lesura
distinta, ondulada, segmentada, extremidades pontudas, raio longo, com 12μm, TLI 0.8,
margem distinta; com 1.5μm de espessura, intexina com 1,5μm de espessura; crassitude
interradial, intexina com 4μm na região interradial e 2μm nos vértices. Laevigado na face
proximal. Rugulado na face distal, rugula com 1μm de largura e ± 5μm de comprimento,
separada 2μm.
Dimensões: Comprimento do diâmetro equatorial 30μm; Largura do diâmetro equatoria 36μm,
comprimento / largura 0.83; nm=1, no=1.
Observações: Camarozonosporites crassus Silva-Caminha et al. (2010) tem ornamentação
verrugada. Camarozonosporites australiensis Norvick & Burger (1975) possui as rugulas com
lumina e muri maior.
Derivação do nome: Por conta da segmentação de sua lesura.
Gênero: Distaverrusporites Muller (1968).
Espécie tipo: Distaverrusporites simplex Muller (1968).
Distaverrusporites muratus sp. nov.
Prancha 3, figuras 3-4.
Holótipo: Prancha 3, figuras 3-4, amostra 22530, EF J-21
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, triangular-obtuso-côncavo; curvatura ausente,
lesura distinta, ondulando levemente, extremidades pontudas, raio médio, 10μm, TLI 0.8,
margem indistinta, intexina com 1μm de espessura. Laevigada na face proximal, baculada na
face distal, bacula com 2μm de comprimento, 1-3 de espessura, separadas 0.5μm.
Dimensões: Comprimento do diâmetro equatorial 25μm; Largura do diâmetro equatorial 27μm,
comprimento / largura 0.90; nm=1, no=1.
Observações: Distaverrusporites simplex Muller, 1968 possui o tamanho das baculas maiores
e seu formato equatorial diferente.
Derivação do nome: Pela ornamentação que lembra um muro feito de tijolos de barro.
Gênero: Echinatisporis Krutzsch (1959).

37
Espécie tipo: Echinatisporis longechinus Krutzsch (1959).
Echinatisporis trianguloconvexus sp. nov.
Prancha 4, figuras3-4.
Holótipo: Prancha 4, figuras 3-4, amostra 23582, EF B-44
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, triangular-obtuso-convexo; trilete, curvatura
ausente, lesura distinta, ondulando, extremidades pontudas, raio médio 10μm, TLI 1 margem
distinta, com 1μm de espessura, intexina com 2μm de espessura. Equinado na superfície inteira
do esporo, equina com 0.5μm de comprimento e 1μm de largura, separadas 4μm.
Dimensões: Comprimento do diâmetro equatorial 20μm; Largura do diâmetro equatoria 20μm,
comprimento/largura 1; nm=1, no=1.
Observações: Echinatisporis infantus (D’Apolito et al., in prep) possui maior tamanho,
intexina mais fina e equina mais comprida.
Derivação do nome: Por conta do seu formato equatorial.
Gênero: Foveotriletes Van der Hammen ex Potonié (1956).
Espécie tipo: Foveotriletes scrobiculatus Potonié (1956).
Foveotriletes noctis sp. nov.
Prancha 4, figuras 5-6
Holótipo: Prancha 4, figuras 5-6, amostra 22749, EF K-21
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, triangular-obtuso-convexo; trilete, curvatura
ausente, lesura distinta, reta, extremidades pontudas, raio médio, 10μm, TLI 0.76, margem
indistinta, intexina com 1μm de espessura. Fossulado em toda a superfície do esporo, fossula
anamastosada, muri com 1μm de largura, lumina com 1μm de largura.
Dimensões: Comprimento do diâmetro equatorial 26μm; Largura do diâmetro equatoria 28μm,
comprimento / largura 0.92; nm=1, no=1.
Observações: Não existe gênero específico para esporos com ornamentação fossulada, então o
esporo ficara provisoriamente neste gênero. Ischyosporites problematicus Jaramillo and
Dilcher (2001) possui tamanho maior e fossulas maiores. Concavissimisporites fossulatus
Dueñas (1980) tem formato equatorial concavo.

38
Derivação do nome: Em homenagem ao período noturno do dia.
Foveotriletes pseudornatus sp. nov.
Prancha 4, figuras 7-8.
Holótipo: Prancha 4, figuras 7-8, amostra 22629, EF T-18/1
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, triangular-obtuso-convexo; trilete, sem curvatura,
lesura distinta, reta, extremidades pontudas, radii longo, 14μm, TLI 0.96, margem indistinta,
intexina com 1μm de espessura. Foveolado na vista distal, lumina com 1μm de largura, muri
com 2μm de largura. Laevigado na vista proximal.
Dimensões: Comprimento do diâmetro equatorial 29μm; Largura do diâmetro equatoria 25μm,
comprimento / largura 1.16; nm=1, no=1.
Observações: Foveotriletes ornatus Regali et al., 1974 tem ornamentação foveolada em toda
sua superfície.
Derivação do nome: Pela semelhança com o esporo Foveotriletes ornatus.
Gênero: Rugulatisporites Thomson et al. (1953).
Espécie tipo: Rugulatisporites quintus Thomson et al. (1953).
Rugulatisporites reticulatus sp. nov.
Prancha 6, figuras 3-4.
Holótipo: Prancha 6, figuras 3-4, amostra 22690, EF Y-35
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, triangular-obtuso-concavo; trilete, sem curvatura,
radii longo, 11μm, TLI 0.91, margem indistinta; intexina com 1μm de espessura. Rugulado na
vista proximal, lumina com 0.5μm de espessura, muri com 2μm de espessura e 0.5μm de
comprimento. Reticulado na vista distal, lumina com 0.5μm de largura e muri com 0.5μm de
largura.
Dimensões: Comprimento do diâmetro equatorial 24μm; Largura do diâmetro equatoria 29μm,
comprimento / largura 0.82; nm=1, no=1.
Observações: Rugulatisporites polysculptilis Herngreen (1975) e Rugulatisporites maculosus
Jaramilo et al. (2007) possuem rugulas maiores.
Derivação do nome: Por conta da ornamentação reticulada da vista distal.

39
Rugulatisporites rugulatus sp. nov.
Prancha 6, figuras 5-6.
Holótipo: Prancha 6, figuras 5-6, amostra 22530, EF L-32-1
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, triangular-obtuso-convexo; trilete, sem curvatura,
lesura distinta, reta, extremidades pontudas, radii longo, 10μm, TLI 0.86, margem distinta,
intexina com 2μm de espessura; Microverrugado na face proximal, verrugas com 1μm de
comprimento, 1μm de largura, separadas 1μm. Rugulado na face distal, rugula com 5μm de
comprimento, 2μm de largura, separadas 1μm.
Dimensões: Comprimento do diâmetro equatorial 23μm; Largura do diâmetro equatoria 30μm,
comprimento / largura 0.76; nm=1, no=1.
Observações: Distaverrusporites simplex Muller, 1968 e Distaverrusporites muratus sp. nov.
possuem ornamentação baculada na face distal,
Derivação do nome: Por conta da ornamentação rugulada.
Gênero. Verrucatotriletes van Hoeken-Klinkenberg, 1964
Espécie tipo: Verrucatotriletes bullatus Van Hoeken-Klinkenberg (1964).
Verrucatotriletes circularis sp. nov.
Prancha 6, figuras 9-10.
Holótipo: Prancha 6, figuras 9-10, amostra 22749, EF R-28/1
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, simetricamente radial, circular; trilete, sem curvatura, lesura distinta, radii
longo, 11μm, TLI 0.76, ondulado, margem distinta, 0.5μm de largura; intexina com 1μm de
largura. Verrugado em todo o grão, verrugas aumentam de tamanho em direção ao equador,
verrugas com 1μm de largura na região da marca trilete e com 2μm de largura no equador,
separadas 0.5μm, 0.5μm de comprimento.
Dimensões: Comprimento do diâmetro equatorial 21μm; Largura do diâmetro equatorial 24μm,
comprimento / largura 0.87; nm=1, no=1.
Observações: Verrucatotriletes etayoi Dueñas 1980 possui verrugas muito maiores, maior
tamanho e não possui margem.
Derivação do nome: Por conta de seu formato equatorial circular.

40
4.1.3 Polen
4.1.3.1 Inaperturado
Gênero: Inaperturopollenites (Pflug) Potonié (1958).
Espécie tipo: Inaperturopollenites dubius (Potonié & Venitz) Thomson et al. (1953).
Inapertuporpollenites baculatus sp. nov.
Prancha 7, figuras 5-6.
Holótipo: Prancha 7, figuras 5-6, amostra 22749, EF B-44-4
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, âmbito circular, inaperturado; intectado, exina com
2μm de espessura. Baculado, com baculas maiores alternando com baculas menores. Baculas
maiores com 2μm de comprimento, 1.5μm de largura, menores com 1μm de comprimento,
separadas 0.5μm.
Dimensões: Comprimento do diâmetro equatorial 25μm, Largura do diâmetro equatorial 21μm,
nm=1, no=1.
Observações: Inaperturopollenites microclavatus Regali et al. (1974) e Clavainaperturites
cordatus Gonzalez-Guzman (1967) tem ornamentação clavada.
Derivação do nome: Por conta da ornamentação baculada.
4.1.3.2 Monosulcado
Gênero: Foveomonocolpites Wijmstra (1971)
Espécie tipo: Foveomonocolpites regalis Wijmstra (1971)
Foveomonocolpites amadus sp. nov.
Prancha 7, figuras 15-16.
Holótipo: Prancha 7, figuras 15-16, amostra 22727, EF H-15-1
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, bilateralmente simétrico, heteropolar, perprolado; monocolpado, colpo
longo, extremidades pontudas, borda reta, colpi 30μm de comprimento, CPi 0.66, colpo
costado, costa com 1μm de largura; tectado, exina com 3μm de espessura, nexina com 1μm de

41
espessura, columelas distintas, com 1μm de comprimento, 1μm de largura, separadas 1μm, teto
com 1μm de espessura. Foveolado, lumina com 1.5μm de largura e muri com 1μm de largura.
Dimensões: Diâmetro polar 45μm, Diâmetro equatorial 20μm, polar/equatorial 2.25; nm=1,
no=1.
Observações: Foveomonocolpites bauchensis Adegoke et al. (1978) possui o colpo simples e
a exina mais fina.
Derivação do nome: Palavra “amado” traduzida do português para o latim.
Gênero: Monocolpopollenites Potonié e Venitz (1934) emend. Nichols et al. (1973).
Espécie tipo: Monocolpopollenites tranquillus Potonié e Venitz (1934) ex Thomson et al.
(1953).
Monocolpopollenites digitus sp. nov.
Prancha 8, figuras 5-6
Holótipo: Prancha 8, figuras 5-6, amostra 23656, EF D-18-1
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, bilateralmente simétrico, heteropolar, prolato-esferoidal; monocolpado,
colpo longo, extremidades arredondadas, borda levemente arredondada, colpo com 22μm de
comprimento, CPi 0.81, colpo simples; atectado, exina com 1μm de espessura. Microreticulado,
lumina com largura menor que 0.5μm e muri com largura menor que 0.5μm.
Dimensões: Diâmetro polar 27μm, diâmetro equatorial 25μm, polar/equatorial 1.08; nm=1,
no=1.
Observações: Psilamonocolpites operculatus Pardo et al. (2003) possui opérculo e exina mais
grossa, Monocolpopollenites ovatus Jaramillo e Dilcher (2001) possui reticulo maior e formato
ovoide.
Derivação do nome: Por conta de seu formato equatorial que lembra um dedo.
Gênero: Psilamonocolpites Van der Hammen e Garcia (1965).
Espécie tipo: Psilamonocolpites grandis (Van der Hammen, 1954) Van der Hammen e Garcia
(1965).
Psilamonocolpites delicatus sp. nov.
Prancha 8, figuras 9-10

42
Holótipo: Prancha 9, figuras 9-10, amostra 23539, EF H-42-2
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, bilateralmente simétrico, heteropolar, prolato; monocolpado, colpo médio,
extremidades pontudas, borda reta, colpo com 12μm de comprimento, CPi 0.6; atectado, exina
com 0.5μm de espessura. Psilado.
Dimensões: Diâmetro polar 20μm, diâmetro equatorial 15μm, polar/equatorial 1.33; nm=1,
no=1.
Observações: Psilamonocolpites minor Jan du Chene et al. (1978) tem as extremidades do
colpo arredondadas, Psilamonocolpites nanus Hoorn, 1993 tem a exina tectada.
Derivação do nome: Por conta de sua exina muito fina.
4.1.3.3 Tricolpado
Gênero: Loranthacites Mchedlishvili, 1961
Espécie tipo: Loranthacites macrosolenoides Mchedlishvili (1961).
Loranthacites apocirculoreticulatus sp. nov.
Prancha 10, figuras 13-17
Holótipo: Prancha 10, figuras 13-15, amostra 23596, EF D-46-1
Parátipo: Prancha 10, figuras 16-17, amostra 23570, C-36
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, âmbito triangular-obtuso-reto; tricolpado,
sincolpado, levemente marginado, margem com 1μm de largura produzida pela diminuição de
tamanho das columelas perto dos colpos, CEi 0.5, colpos parecem estar abertos, com dois
grande poros em cada polo, aberturas com 13μm de comprimento e 11μm de largura; tectado,
exina com 2.5μm de espessura, nexina com 0.5μm de espessura, columela com 2μm de
comprimento, distintas, com 1μm de largura, separadas 1μm, teto com 1μm de espessura.
Microreticulado, homobrochado, simplicolumelado, lumina com 1μm de largura e muri com
0.5μm de comprimento.
Dimensões: Comprimento do diâmetro equatorial 22-(26)-30μm, Largura do diâmetro
equatorial 25-(28)-31μm , comprimento/largura: 0.96; nm=2, no=2.
Observações: Loranthacites digitatus Silva-Caminha et al 2010 possui columelas digitadas e
não possui orifícios na região polar.

43
Derivação do nome: Por conta da presença de orifícios nos polos e da ornamentação reticulada.
Gênero: Rhoipites Wodehouse (1933).
Espécie tipo: Rhoipites bradleyi Wodehouse (1933).
Rhoipites? hoornii sp. nov.
Prancha 11, figuras 7-8.
Holótipo: Prancha 11, figuras 7-8, amostra 23503, EF C-51
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, perprolado; tricolpado, colpo com 25μm
de comprimento, médio, CPi 0.69, borda reta, extremidades pontudas; semitectado, exina com
1μm de espessura, nexina com 0.3μm de espessura, columela com 0.4μm de espessura,
indistinta, teto com 0.3μm de espessura. Microreticulado, homobrochado, lumina de circular a
levemente alongada, com 0.5μm de largura e muri com 0.5μm de comprimento
Dimensões: Diâmetro polar 36μm, diâmetro equatorial 15μm, polar/equatorial: 2.4, nm=1,
no=1.
Observações: Retitricolpites minutus Gonzalez-Guzman (1967) possui formato prolato, colpo
marginado e menor. Retitricolpites obtusus Van Hoeken-Klinkenberg (1966) possui um
espessamento da exina nos polos. Retritricolpites simplex Gonzalez-Guzman (1967) possui
tamanho maior e colpo costado. Retitricolpites wijningae Hoorn (1994a) possui luminas
maiores, Rhoipites? basicus D’Apolito et al. (2018) possui exina mais grossa.
Derivação do nome: Em homenagem a palinológa holandesa Carina Hoorn.
Gênero: Rousea Srivastava (1969).
Espécie tipo: Rousea subtilis Srivastava (1969).
Rousea? simplex sp. nov.
Prancha 11, figuras 9-10.
Holótipo: Prancha 11, figuras 9-10, amostra 23547, EF B7-2
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, subprolato; tricolpado, colpo com 30μm
de comprimento, CPi 0.90, borda reta, extremidades pontudas, marginado, margem com 2μm
de largura, costado, costa com 1μm de largura; semitectado, exina com 2μm de espessura,

44
nexina com 0.5μm de espessura, columela com 1μm de espessura, distinta, com 0.5μm de
largura e separadas 1μm, teto com 0.5μm de espessura. Reticulado, lumina diminui em tamanho
em direção aos colpos, lumina no mesocolpia com 1μm de largura e perto dos colpos com
0.5μm de largura.
Dimensões: Diâmetro polar 33μm, Diâmetro equatorial 25μm, polar/equatorial: 1.32; nm=1,
no=1.
Observações: Rousea florentina (Gonzalez-Guzman, 1967; Jaramillo and Dilcher, 2001
e Rousea georgensis (Brenner) Dettmann (1973) tem a diminuição do seu lumina em direção
aos polos.
Derivação do nome: Por conta da morfologia simples.
4.1.3.4 Tricolporado
Gênero: Echitricolporites Van der Hammen (1956b) ex Germeraad et al. (1968).
Espécie tipo: Echitricolporites spinosus Van der Hammen 1956 ex Germeraad et al. (1968).
Echitricolporites distintus sp. nov.
Prancha 12, figuras 11-12.
Holótipo: Prancha 12, figuras 11-12, amostra 23656, EF B 41/1
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, suboblato; tricolporado, ectocolpo de
tamanho médio, com 10μm de comprimento, borda reta, extremidades pontudas, CPi 0.58,
colpo costado, costa com 1μm de largura, endoporo indistinto, tectado, exina com 1.5μm de
espessura, nexina com < 0.5 de espessura, columela com 1μm de espessura, indistinta, teto com
< 0.5μm de espessura. Equinado, equinas com 1μm de comprimento, largura de 0.5μm na base,
bem espaçadas e homogeneamente distribuídas, separadas 2μm. Superfície entre as equinas
psilada.
Dimensões: Diâmetro polar17μm, diâmetro equatorial20μm. comprimento/largura: 0.85;
nm=1, no=1
Observações: Echitricolporites mcneillyi Germeraad et al. (1968), Echitricolporites minutus
Regali et al. (1974) e Echitricolporites spinosus Van der Hammen (1956b) possuem equinas
mais grossas e abundantes.
Derivação do nome: Por conta de sua morfologia distinta.

45
Echitricolporites insanus sp. nov.
Prancha 12, figuras 13-14.
Holótipo: Prancha 12, figuras 13-14, amostra 23582, EF U33
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, prolato; tricolporado, ectocolpo longo,
com 20μm de comprimento, borda reta, extremidades pontudas, CPi 0.74, colpo costado, costa
com 2μm de largura, endoporo lolongado, com 1μm de largura e 5μm de comprimento, simples;
intectado, exina com 1μm de espessura. Equinado, equinas com 0.5μm de comprimento, com
0.5μm de largura na base, separadas 2μm. Superfície entre as equinas psilada.
Dimensões: Diâmetro polar 27μm, diâmetro equatorial 15μm, polar/equatorial: 1.8; nm=1,
no=1
Observações: Echitricolporites mcneillyi Germeraad et al.,1968, Echitricolporites minutus
Regali et al.,1974 e Echitricolporites spinosus Van der Hammen (1956b) possuem equinas mais
grossas e abundantes e formato esferoidal.
Derivação do nome: Palavra “louco” traduzido do português para o latim.
Gênero: Foveotricolporites Pierce (1961).
Espécie tipo: Foveotricolporites rhombohedralis Pierce (1961).
Foveotricolporites miritiensis sp. nov.
Prancha 12, figuras 17-20.
Holótipo: Prancha 12, figuras 17-18, amostra 23656, EF E48-2
Parátipo: Prancha 12, figuras 19-20, amostra 23656, EF T11-4
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, esferoidal; tricolporado, ectocolpo curto,
com 7μm de comprimento, extremidades pontudas, CPi 0.33, costado, costa com 0.5μm de
largura, endoporos circulares, com 4μm de comprimento e 4μm de largura, simples; tectado,
exina com 2μm de espessura, nexina com 0.5μm espessura, columela com 1μm de espessura,
indistinta, teto com 0.5μm de espessura. Foveolado, lumina irregular, variando de tamanho de
0.5 a 2μm de largura, muri com 1μm de largura.
Dimensões: Diâmetro polar 21-(23)-25, diâmetro equatorial 21-(22)-23μm, polar/equatorial 1;
nm=2, no=2.

46
Observações. Foveotricolporites pseudodubiosus Silva-Caminha et al. (2010) possui poros
lalongados e lumina multicolumelado, Foveotricolporites brevicolpatus Jaramillo et al. (2007)
possui a lumina menor.
Derivação do nome: Por conta da cidade amazônica de Miriti.
Foveotricolporites parintensis sp. nov.
Prancha 13, figuras 1-2.
Holótipo: Prancha 13, figuras 1-2, amostra 23582, EF V-32-1
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, esferoidal; tricolporado, ectocolpo longo,
com 29μm de comprimento, borda reta, extremidades pontudas, CPi 1, costado, costa com 1μm
de largura, endoporo lolongado, com 5μm de comprimento e 2 de largura, costado, costa com
1μm de largura; tectado, exina com 1.5μm de espessura, nexina com 1μm de espessura,
columela com 0.4μm de espessura, indistinta, teto com 0.3μm de espessura. Foveolado, lumina
com 0.5μm de largura e muri com 1 μm de largura.
Dimensões: Diâmetro polar 29, diâmetro equatorial 29μm, polar/equatorial 1; nm=1, no=1.
Observações: Foveotricolporites marginatus Gonzalez-Guzman (1967) possui colpo
marginado.
Derivação do nome: Pela cidade amazônica de Parintins.
Gênero: Ladakhipollenites Mathur et al. (1980).
Espécie tipo: Ladakhipollenites levis (Sah e Dutta, 1966; Mathur et al., 1980).
Ladakhipollenites? compressus nov.
Prancha 13, figuras 9-10.
Holótipo: Prancha 13, figuras 9-10, amostra 23656, EF B-53
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, perprolato; tricolporado, ectocolpo com
25μm de comprimento, médio, CPi 0.73, borda reta, extremidade pontuda, costado, costa com
1μm de espessura, 0.5μm de largura, endoporos lalongados, com 2μm de comprimento e 3μm
de largura, simples; tectado, exina com 2μm de espessura, nexina com 0.5μm de espessura,
columela com 1μm de comprimento, empacotadas, teto com 0.5μm de espessura. Psilado a
microreticulado.

47
Dimensões: Diâmetro polar 34μm, diâmetro equatorial 16μm polar/equatorial 2.12; nm=1,
no=1.
Observações: Psilatricolporites magniporatus Hoorn (1993) possui a costa do colpo mais larga
e poros circulares, Psilatricolporites atalayensis Hoorn (1993) possui formato prolato, costa do
colpo mais larga e poro constrito. Ladakhipollenites? caribbiensis (Muller et al., 1987; Silva-
Caminha et al., 2010) possui tamanho muito maior.
Derivação do nome: Por conta das columelas empacotadas.
Ladakhipollenites? irandubaensis sp. nov.
Prancha 13, figuras 15-16.
Holótipo: Prancha 13, figuras 15-16, amostra 23596, EF O15-4
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, subprolato; tricolporado, ectocolpo com
21μm de comprimento, CPi 1.31, borda reta, extremidades pontudas; colpo costado, com 1μm
de largura, endoporo lolongado, 4μm de comprimento e 2 μm de largura; tectado, exina com
2μm de espessura, nexina com 0.5μm de espessura, columela com 1μm de comprimento,
columelas empacotadas, teto com 0.5μm de espessura. Psilado.
Dimensões: Diâmetro polar 25μm, diâmetro equatorial 19μm polar/equatorial 1.31; nm=1,
no=1.
Observações: Psilatricolporites cryptoporus Boltenhagen (1976) possui as extremidades dos
colpos arredondadas e poros indistintos, Psilatricolporites crassoexinatus Hoorn (1993) possui
a exina mais grossa, poros lalongados e constritos. Ladakhipollenites? lolongatus D’Apolito et
al. (2018) possui exina mais fina e formato diferente.
Derivação do nome: Pela cidade amazônica de Iranduba.
Ladakhipollenites? posterus sp. nov.
Prancha 13, figuras 23-24.
Holótipo: Prancha 13, figuras 23-24, amostra 22727, EF E5-2
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, subprolato; tricolporado, ectocolpo com
7μm de comprimento, curto, CPi 0.36, borda reta, extremidades pontudas; costado, costa com
1μm de largura, endoporo lalongado, com 2μm de comprimento e 5μm de largura, costado,
costa com 1μm de largura; tectado, exina com 1μm de espessura, nexina com 0.3μm de

48
espessura, columelas com 0.4μm de comprimento, indistintas, teto com 0.3μm de espessura.
Psilado.
Dimensões: Diâmetro polar 19μm, diâmetro equatorial 15μm polar/equatorial 1.26; nm=1,
no=1.
Observações: Psilatricolporites pachydermatus Lorente (1986) possui exina mais grossa.
Derivação do nome: Palavra latina para “posterior”.
Gênero: Retibrevitricolporites Legoux (1978).
Espécie tipo: Retibrevitricolporites obodoensis Legoux (1978).
Retibrevitricolporites superbus sp. nov.
Prancha 15, figuras 7-8.
Holótipo: Prancha 15, figuras 7-8, amostra 22629, EF O 14-4
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, âmbito circular; tricolporado, colpo com
8μm de comprimento, CPi 0.29, borda reta, extremidades pontudas, simples, poro lolongado,
com 4μm de comprimento e 3μm de largura, costado, costa com 2μm de largura; semitectado,
exina com 1μm de espessura, nexina com 0.3μm de espessura, columela com 0.4μm de
comprimento, distinta, com 1μm de largura e separada 1μm, teto com 0.3μm de espessura.
Reticulado, simplicolumelado, heterobrochado, lumina diminuindo de tamanho dos polos em
direção ao mesocolpia, lumina com 0.5μm de largura nos polos e 1μm de largura no mesolcopia,
muri com 0.5μm de largura, lumina de circular a levemente alongado.
Dimensões: Comprimento do diâmetro equatorial 27μm, Largura do diâmetro equatorial 25μm,
comprimento / largura 1.08; nm=1, no=1.
Observações: Retibrevitricolporites yavarensis (Hoorn, 1993; Silva-Caminha et al., 2010)
possui retículo homobrochado e colpo costado. Retibrevitricolpites retibolus Leidelmeyer
(1966) possui retículo homobrochado.
Derivação do nome: Por conta de sua bonita morfologia.
Retibrevitricolporites tabatingaensis sp. nov.
Prancha 15, figuras 9-10.
Holótipo: Prancha 15, figuras 9-10, amostra 22684, EF L35-3
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.

49
Descrição: Mônade, radialmente simétrico, isopolar, âmbito circular; tricolporado, ectocolpo
com 14μm de comprimento, protudindo, curto, CEi 0.51, borda reta, extremidades
arredondadas; marginado, margem produzida pela diminuição de tamanho da sexina perto do
colpo, margo com 0.5μm de espessura, costado, costa com 2μm de comprimento e 3μm de
largura; semitectado, exina com 1μm de espessura, nexina com 0.3μm de espessura, columelas
com 0.4μm de espessura, distintas, com 0.5μm de largura e separadas 0.5μm, teto com 0.3μm
de espessura, endoporo circular, com 3 x 3μm de comprimento, simples. Reticulado, lumina
circular a levemente alongado, com 0.5 de largura, muri com 0.5μm de espessura,
homobrochado.
Dimensões: Comprimento do diâmetro equatorial 31μm, largura do diâmetro equatorial 33μm,
comprimento/largura: 0.93; nm=1, no=1
Observações: Retibrevitricolporites grandis Jaramillo e Dilcher (2001) possui poro costado e
Retibrevitricolporites yavarensis (Hoorn, 1993; Silva-Caminha et al., 2010) possui colpos
menores.
Derivação do nome: Pela cidade amazônica de Tabatinga.
Gênero: Rhoipites Wodehouse (1933).
Espécie tipo: Rhoipites bradleyi Wodehouse (1933).
Rhoipites crassiexinatus sp. nov.
Prancha 16, figuras 3-4.
Holótipo: Prancha 16, figuras 3-4, amostra 22749, P-11-3.
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, subprolato; tricolporado, colpo com 11μm
de comprimento, curto, CPi 0.55, costado, costa com 1μm de largura, endoporo lolongado com
4μm de comprimento e 3μm de largura, costado, costa com 1μm de largura; tectado, exina com
1.5μm de espessura, exina aumenta de espessura em direção aos polos onde possui 3μm de
espessura, nexina com 0.5μm de espessura, columela com 0.5μm de comprimento, com 0.5μm
de largura e separadas 0.5μm, teto com 0.5μm de espessura. Reticulado, simplicolumelado,
heterobrochado, aumentando de tamanho em direção aos polos, lumina com 1μm de largura na
região equatorial e com 2μm de largura na região polar, lumina circular a alongado, muri com
0.5μm de largura,

50
Dimensões: Diâmetro polar 20μm, diâmetro equatorial 17μm, polar/equatorial 1.17; nm=1,
no=1.
Observações: Retitricolporites belmontensis Regali et al. (1974) possui maior tamanho e exina
mais grossa. Rhoipites caricatus D’Apolito et al., in prep possui reticulo homobrochado,
Retitrescolpites benjaminense D’Apolito et al., in prep é homobrochado e possui poro simples.
Derivação do nome: Pelo fato de a exina ser mais grossa nos polos.
Rhoipites dignus sp. nov.
Prancha 16, figuras 5-6.
Holótipo: Prancha 16, figuras 5-6, amostra 23596, B-41-2
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, prolato; tricolporado, colpo com 25μm de
comprimento, médio, CPi 0.73, difícil de distinguir suas características, poro indistinto;
semitectado, exina com 2μm de espessura, nexina com 0.5μm de espessura, columelas com
1μm de comprimento, distintas, 0.5μm de largura e separadas 0.5μm, teto com 0.5μm de
espessura. Reticulado, simplicolumelado, lumina poligonal, com 3μm de largura e muri com
1μm de largura.
Dimensões: Diâmetro polar 34μm, Diâmetro equatorial 20μm, comprimento/largura: 1.7;
nm=1, no=1.
Observações: Retitrescolpites magnus (Gonzalez-Guzman, 1967; Jaramillo e Dilcher, 2001) é
tricolpado.
Derivação do nome: Por conta do seu muro grosso.
Rhoipites figueiredoensis sp. nov
Prancha 16, figuras 7-8.
Holótipo: Prancha 16, figuras 7-8, amostra 22684, E-11-2
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, prolato; tricolporado, ectocolpo com
21μm de comprimento, longo, CPi 0.87, marginado, margo com 1μm delargura, costado, costa
com 1μm de espessura, 1μm de largura; endoporo lalongado com 3μm de largura e 2μm de
comprimento, simples; tectado, exina com 1μm de espessura, nexina com 0.3μm de espessura,
columela com 0.4μm de comprimento, indistinta, teto com 0.3μm de espessura.
Microcroreticulado, simplicolumelado, homobrochado, lumina com 0.5μm de largura, muri
com 0.5μm de largura.

51
Dimensões: Diâmetro polar 24μm, diâmetro equatorial 17μm, polar/equatorial 1.41; nm=1,
no=1.
Observações: Retitricolporites ellipticus Van Hoeken-Klinkenberg (1966) possui tamanho
menor e colpo apenas colpado.
Derivação do nome: Pela cidade amazônica de Presidente Figueiredo.
Rhoipites normalis sp. nov.
Prancha 16, figuras 15-16
Holótipo: Prancha 16, figuras 15-16, amostra 22727, EF= K-25-3
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, prolato; tricolporado, colpo longo, com
20μm de comprimento, longo, CPi 0.90, costado, costa com 2μm de largura, poro lolongado
com 3μm de comprimento e 2μm de largura; semitectado, exina com 1μm de espessura, nexina
com 0.3μm de espessura, columela com 0.4μm de comprimento, distinta, com menos de 0.5μm
de largura, separadas menos de 0.5μm, teto com 0.3μm de espessura. Reticulado,
homobrochado, lumina de circular a levemente alongada, lumina com 1μm de largura, muri
com 0.5μm de largura.
Dimensões: Diâmetro polar 22μm, diâmetro equatorial 14μm, polar/equatorial 1.5; nm=1,
no=1.
Observações: Retitricolporites milnei Hoorn (1993) possui colpo menor e poro lalongado,
Retitricolporites ellipticus Van Hoeken-Klinkenberg (1966) possui poro lalongado.
Derivação do nome: Por conta de sua morfologia simples
Rhoipites superlolongatus sp. nov.
Prancha 16, figuras 21-22.
Holótipo: Prancha 16, figuras 21-22, amostra 23547, EF= O-18
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, prolato; tricolporado, ectocolpo com
15μm de comprimento, longo, CPi 0.78; costado, costa com
0.5μm de largura; endoporo lolongado com 4μm de comprimento e 3μm de largura; costado,
costa com 0.5μm de largura; semitectado, exina com 1μm de espessura, nexina com 0.3μm de
espessura, columela com 0.4μm de comprimento, indistinta, teto com 0.3μm de espessura.

52
Reticulado, simplicolumelado, homobrochado, lumina com 1μm de largura, muri com 0.5μm
de largura, lumina de circular a levemente alongado.
Dimensões: Diâmetro polar19μm, diâmetro equatorial 14μm, polar/equatorial 1.35; nm=1,
no=1.
Observações: Ladakhipollenoites? lolongatus D’Apolito et al. (2018) é foveolado, Rhoipites
pseudopilatus sp.nov D’Apolito et al., in prep possui poros maiores.
Derivação do nome: Pela presença de poros lolongados.
Gênero: Rugutricolporites Gonzalez-Guzman (1967).
Espécie tipo: Rugutricolporites felix Gonzalez-Guzman (1967).
Rugutricolporites perfuratus sp. nov
Prancha 17, figuras 3-4.
Holótipo: Prancha 17, figuras 3-4, amostra 23698, EF =Q-34
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, subprolato; tricolporado, colpo com 15μm
de comprimento, curto, CPi 0.65, costado, costa com 1μm de largura, poro simples, circular,
com 2μm de comprimento e 2μm de largura; atectado, exina com 2μm de espessura. Verrucado
a rugulado, verruca-rugula com 2μm de largura, separadas 0.5μm, nexina perfurada,
perfurações com 0,5μm de largura separadas 1μm.
Dimensões: Diâmetro polar23μm, diâmetro equatorial 20μm, polar/equatorial 1.15; nm=1,
no=1.
Observações: Rugutricolporites felix Gonzalez-Guzman (1967) possui ornamentação apenas
rugulada, Verrutricolporites haplites Gonzalez-Guzman (1967) possui verrugas em muito mais
quantidade e poro costado.
Derivação do nome: Pela presença de nexina perforada.
Gênero: Siltaria Traverse (1955)
Espécie tipo: Siltaria scabriextima Traverse (1955).
Siltaria pseudodilcheri sp. nov.
Prancha 17, figuras 7-8
Holótipo: Prancha 17, figuras 7-8, amostra 23650, EF= F-38
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.

53
Descrição: Mônade, radialmente simétrico, circular; tricolporado, colpo com 20μm de
comprimento, CPi 0.76, marginado, margem com 0.5μm de largura, produzida pela diminuição
da exina perto dos colpos, poro lalongado com 1μm de comprimento e 4μm de largura; tectado,
exina com 1μm de espessura, nexina com menos de 0.5μm de espessura, columelas com 0.5μm
de comprimento, indistinta, teto com menos de 0.5μm de espessura. Microreticulado, lumina
com menos de 0.5μm de largura, muri com menos de 1μm de largura.
Dimensões: Comprimento do diâmetro equatorial 26μm, Largura do diâmetro equatorial 22μm,
comprimento/largura 1.18; nm=1, no=1.
Observações: Siltaria dilcheri Silva-Caminha et al. (2010) possui exina mais grossa e poros
costados.
Derivação do nome: Pela semelhança com S. dilcheri.
Gênero: Striatricolporites (Van der Hammen, 1956a; Leidelmeyer, 1966).
Espécie tipo: Striatricolporites pimulis (Van der Hammen, 1956a; Leidelmeyer, 1966).
Striatricolporites caminhae sp. nov.
Prancha 17, figuras 17-18
Holótipo: Prancha 17, figuras 17-18, amostra 22629, EF= C-33
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, prolato; tricolporado, colpo com 15μm de
comprimento, médio, Cpi 0.71, costado, costa com 1μm de espessura 1μm e 1μm de largura,
poro lalongado com 2μm de comprimento e 3μm de largura, simples, tectado, exina com 1μm
de espessura, nexina com 0.3μm de espessura, columela com 0.5μm de comprimento, indistinta,
teto com 0.4μm de espessura. Estriado, estrias anamastosando, não paralelas aos colpos, muito
finas, com 0.5μm de largura e muri com 0.5μm de largura.
Dimensões: Diâmetro polar 21, diâmetro equatorial 12μm, polar/equatorial 1.75; nm=2, no=4.
Observações: Striatricolporites agustinus Gonzalez-Guzman (1967) possui as estrias paralelas
aos colpos. Striatricolporites poloreticulatus Silva-Caminha et al. (2010) também é reticulado.
Derivação do nome: Em homenagem à palinóloga brasileira Silane Silva-Caminha
Striatricolporites magnificus sp. nov.
Prancha 17, figuras 19-20.
Holótipo: Prancha 17, figuras 19-20, amostra 23570, N-35-3

54
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, prolato; tricolporado, colpo com 20μm de
comprimento, médio, CPi 0.76, costado, costa com 1μm de espessura e 1μm de largura, poro
circular, com 4μm de comprimento e 4μm de largura, simples; tectado, exina com 1μm de
espessura, nexina com 0.3μm de espessura, columela com 0.4μm de comprimento, distinta, com
0.5μm de largura e separada 0.5μm, teto com 0.3μm de espessura. Estriado, estrias paralelas
entre si mas não paralelas aos colpos, com 0.5μm de largura e muri com 0.5μm.
Dimensões: Diâmetro polar 20μm, diâmetro equatorial 19μm, polar/equatorial 1.36, nm=2,
no=6.
Observações: Striatricolporites archangelskyi Herngreen (1975) possui poros lalongados e
maior tamanho. Striatricolporites tenuissimus Dueñas (1980) possui as estrias mais finas
Derivação do nome: Por conta de seu bonito formato.
Gênero: Syncolporites Van der Hammen (1954).
Espécie tipo: Syncolporites lisamae Van der Hammen (1954).
Syncolporites microreticulatus sp. nov.
Prancha 18, figuras 3-5
Holótipo: Prancha 18, figuras 3-4 amostra 23503, E-11-1
Parátipo: Prancha 18, figura 5, amostra 23582, N-38-2
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, tricolporado, sincolpado, ectocolpo com 28μm de
comprimento, marginado, margem produzida pelo afinamento da exina perto dos colpos, com
1μm de largura e 0.5μm de espessura, costado, costa com 1μm de largura, endoporo lalongado
com 4μm de comprimento e 6μm de largura, costado, costa com 0.5μm de largura; exina com
1μm de espessura, nexina com 0.3μm de espessura, columela com 0.4μm de comprimento,
indistinta, teto com 0.3μm de espessura. Microreticulado, lumina com 0.5μm de largura, muri
com 0.5μm de largura.
Dimensões: Diâmetro polar22-(22.5)-23μm, diâmetro equatorial 20-(21)-22μm,
polar/equatorial 1.1; nm=2, no=2.
Observações: Syncolporites marginatus Van Hoeken-Klinkenberg (1964) possui a costa do
poro mais grossa e o reticulo com o lumina maior.
Derivação do nome: Por ser microreticulado.

55
Syncolporites tenuicolpatus
Syncolporites sp.1 Silva-caminha et al. (2010).
Espécimes: Prancha 18, figuras 6-7, amostra 22690, T-24-1
Prancha 18, figura 8, amostra 23653, D16
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, âmbito triangular-agudo-reto; tricolporado,
sincolpado, ectocolpo simples, borda reta, área do apocolpio presente, com 5μm de largura,
endoporo circular com 1μm de comprimento e 1μm de largura, fastigiado e costado, costa com
2 μm de largura, fastígio com 3μm de espessura e 1μm de largura; tectado, exina com 1.3μm
de espessura, nexina com 0.5μm de espessura, columela com 0.5μm de comprimento, indistinta,
teto com 0.3μm de espessura. Psilado.
Dimensões: Comprimento do diâmetro equatorial 25-(25.5)-26μm, Largura do diâmetro
equatorial 25μm, comprimento/largura 1.04; nm=2,no=2.
Observações: Syncolporites poricostatus van Hoeken-Klinkenberg (1966) possui o poro
vestibulado. Syncolporites reymenti Jan du Chene (1977) possui ornamentação escabrada e
vestibulo.
Derivação do nome: Pelos colpos finos.
Gênero: Verrutricolporites Van der Hammen e Wijmstra (1964).
Espécie tipo: Verrutricolporites rotundiporus Van der Hammen e Wijmstra (1964).
Verrutricolporites intectatussp.nov.
Prancha 18, figuras 9-10
Holótipo: Prancha 18, figuras 9-10, amostra 23679, EF M-20-3
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, prolato; tricolporado, colpo com 15μm de
comprimento, médio, CPI 0.75, costado, costa com 1μm de largura, poro lalongado, com 2μm
de comprimento e 3μm de largura, costado, costa com 0.5μm de largura; intectado, exina com
0.5μm de espessura. Verrugado, verrugas com 0.5μm de largura, 0.3μm de comprimento,
separadas 1μm.
Dimensões: Diâmetro polar20μm, diâmetro equatorial 15μm, polar/equatorial 1.33; nm=1,
no=1.

56
Observações: Verrutricolporites haplites Gonzalez-Guzman (1967) possui as verrugas
maiores. Verrutricolporites pequenus D’Apolito et al., in prep é tectado, Verrutricolporites
simplex D’Apolito et al., in prep possui verrugas com tamanho muito maior.
Derivação do nome: Por conta de sua exina intectada.
Verrutricolporites poricostatussp.nov.
Prancha 18, figuras 11-12
Holótipo: Prancha 18, figuras 11-12, amostra 23698 EF J-16-3
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, esferoidal; tricolporado, colpo com 10μm
de comprimento, curto, CPi 0.55, costado, costa com 0.5μm de largura, poro indistinto, costado,
costa com 1μm de largura; tectado, exina com 2μm de espessura, nexina com 0.5μm de
espessura, columela com 1μm de comprimento, indistinta, teto com 0.5μm de espessura.
Verrugado, verrugas com 0.5μm de largura, 0.5μm de comprimento e separadas 0.5μm.
Dimensões: Diâmetro polar18μm, diâmetro equatorial 18μm, polar/equatorial 1; nm=1, no=1.
Observações: Verrutricolporites intectatus sp. nov. é intectado, Verrutricolporites
rotundiporus Van der Hammen e Wijmstra (1964) possui o colpo indistinto e poro circular.
Verrutricolporites pequenus D’Apolito et al., in prep tem os poros maiores e Verrutricolporites
simplex D’Apolito et al., in prep temporos maiores e lologandos e verrugas maiores.
Derivação do nome: Por conta de seus poros costados.
4.1.3.5 Triporado
Gênero: Echitriporites Van der Hammen 1956 (Van Hoeken-Klinkenberg, 1964)
Espécie tipo: Echitriporites trianguliformis Van der Hammen 1956 (Van Hoeken-Klinkenberg,
1964)
Echitriporites nanuspinosus sp. nov.
Prancha 18, figuras 21-22.
Holótipo: Prancha 18, figuras 21-22, amostra 23639, EF F12-4
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.

57
Descrição: Mônade, radialmente simétrico, isopolar, âmbito circular; triporado, poros com 1μm
de comprimento, costado, costa com 1μm de espessura e 0.5μm de largura, marginado, margem
com 0.5μm de espessura, margem criada pelo engrossamento da exina perto dos poros; tectado,
exina com 2.5μm de espessura, nexina com 1μm de espessura, columela com 1μm de espessura,
indistinta, teto com 0.5μm de espessura. Equinado, equinas com 1μm de comprimento, 0.5μm
de largura, separadas 2μm, exina entre as equinas psilada.
Dimensões: Comprimento do diâmetro equatorial 17μm, Largura do diâmetro equatorial17μm,
comprimento/largura 1; nm=1, no=1.
Observações: Echitriporites kwaense Salard-Cheboldaeff (1974) possui poros simples e é
intectado, Echitriporites trianguliformis var. orbicularis Jaramillo e Dilcher (2001) é intectado.
Derivação do nome: Por conta das equinas pequenas.
Gênero: Scabratriporites T. Van der Hammen ex P.M.J. Vvan Hoeken-Klinkenberg (1964).
Espécie tipo: Scabratriporites annellus P.M.J. van Hoeken-Klinkenberg (1964).
Scabratriporites annulatus sp. nov.
Prancha 19, figuras 9-10.
Holótipo: Prancha 9, figuras 9-10, amostra 23519, EF Z-46-2
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, âmbito circular; triporado, poros com 4μm
de comprimento e 4μm de largura, marginado, margo com 2μm de largura e 1μm de espessura,
margo formado pelo espessamento da exina perto dos poros, costado, costa muito fina, com
0.5μm de largura e 0.5μm espessura; tectado, exina com 1μm de espessura, nexina com 0.3μm
de espessura, columela com 0.4μm de espessura, indistinta, teto com 0.3μm de espesssura.
Scabrado, escabras com 0.5μm de largura separadas 0.5μm.
Dimensões: Comprimento do diâmetro equatorial 21μm, Largura do diâmetro equatorial 20μm,
comprimento / largura 1.05; nm=1, no=1.
Observações: Scabratriporites moderatus Gonzalez-Guzman (1967) possui poros simples,
Scabratriporites simpliformis Van Hoeken-Klinkenberg (1966) possui âmbito triangular e
poros apenas costados.
4.1.3.6 Stephanocolpado

58
Gênero: Verrustephanocolpites T. van der Hammen & Garcia, 1966
Espécie tipo: Verrustephanocolpites verrucatus T. van der Hammen & Garcia, 1966
Verrustephanocolpites guimaraesensis sp. nov.
Prancha 20, figuras 5-6
Holótipo: Prancha 20, figuras 5-6, amostra 23528, EF Q-33-3
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, âmbito circular, isopolar, stephanocolpado, com 4
colpos curtos, com 14μm de comprimento, CEi 0.46, borda reta, com extremidades pontudas,
levemente costado, costa com 0.5μm de largura; tectado, exina com 2μm de espessura, nexina
com 0.5μm de espessura, columela com 1μm de comprimento, distinta, com 0.5μm de largura,
separada 0.5μm, teto com 0.5μm de espessura.Verrucado a rugulado, verrugas com de 2 a 3μm
de largura, separadas 0.5μm.
Dimensões: Comprimento do diâmetro equatorial 30μm, Largura do diâmetro equatorial 30μm,
comprimento/largura 1; nm=1, no=1.
Observações: Verrustephanocolpites rugulatus Jaramillo e Dilcher (2001) tem os colpos
cercados por elevações da exina. Verrustephanocolpites verrucatus Van der Hammen e Garcia
(1966) possui 5 colpos.
Derivação do nome: Em homenagem a Chapada dos Guimarães.
4.1.3.7 Periporado
Gênero: Cyperaceapollis W. Krutzsch (1970).
Espécie tipo: Cyperaceapollis cf. C. neogenicus Krutzsch (1970).
Cyperaceapollis rotundus sp. nov.
Prancha 20, figuras 23-26
Holótipo: Prancha 20, figuras 23-24, amostra 22530, EF R-37-2
Parátipo: Prancha 20, figuras 25-26, amostra 23519, EF G-34
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, anisopolar, bilateralmente simétrico, prolatocom 4 colpóides, colpóides
com 10μm de comprimento, simples; tectado, exina com 1.5μm de espessura, nexina com
menos de 0.5μm, columela com 1μm de comprimento, distinta, com 0.5μm de largura e

59
separadas 0.5μm, teto com 0.5μm de espessura. Psilado, possível ver o topo das columelas em
vista frontal. Psilado.
Dimensões: Diâmetro polar27-(28.5)-30μm, Diâmetro equatorial 19-(19.5)-20μm,
comprimento /equatorial 1.57; nm=2, no=31
Observações: Cyperaceapollis cf. C. neogenicus Krutzsch (1970) aparentemente possui a exina
mais fina e as semi-aberturas circulares.
Derivação do nome: Por conta dos cantos arredondados.
Psilaperiporites annulatus sp. nov.
Prancha 21, figuras 15-16.
Holótipo: Prancha 21, figuras 15-16, amostra 22713, EF H-8-3
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, âmbito circular; pantoporado, com 5-6
poros, marginado, margem com 1μm de largura e 1μm de espessura, poro circular, com 2μm
de comprimento e 2μm de largura, simples, tectado, nexina com 0.5μm de espessura, columela
com 1μm de espessura, teto com 0.5μm de espessura. Psilado.
Dimensões: Comprimento do diâmetro equatorial 26μm; Largura do diâmetro equatorial 22μm;
comprimento/largura 1.18; nm=1, no=1.
Observações: Psilaperiporites multiporatus Hoorn (1994b) possui maior quantidade de poros
e maior tamanho.
Derivação do nome: Pela presença de poros anulados.
4.1.3.8 Stephanoporado
Gênero: Pachydermites Germeraad et al. (1968).
Espécie tipo: Pachydermites diederixi Germeraad et al. (1968).
Pachydermites pseudoalnus sp. nov.
Prancha 22, figuras 3-4.
Holótipo: Prancha 22, figuras 3-4, amostra 23698, EF Y31
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, âmbito cirular a pentagonal;
stephanoporado, poro circular com 2μm de comprimento e 2μm de largura, marginado, margem

60
com 1μm de espessura, formada pelo espessamento da sexina perto dos poros, costado, costa
com 2μm de largura; tectado, nexina com 0.3μm de espessura, columela com 0.4μm de
espessura, teto com 0.3μm de espessura. Psilado.
Dimensões: Comprimento do diâmetro equatorial 21μm; Largura do diâmetro equatorial 20μm;
comprimento/largura 1.18; nm=1, no=1.
Observações: Pachydermites diederixi Germeraad et al. (1968) possui a exina mais grossa,
ornamentação verrugada e endoporo vestibulado.
Derivação do nome: Pela semelhança com o tipo polínico de Alnus.
Gênero: Verrustephanoporites Leidelmeyer (1966).
Espécie tipo: Verrustephanoporites simplex Leidelmeyer (1966).
Verrustephanoporites? uatumaensis sp. nov.
Prancha 22, figuras 17-18
Holótipo: Prancha 22, figuras 17-18, amostra 23639, EF Q 46
Localidade do tipo: Poço 1-AS-15-AM, Formação Solimões, Amazonas, Brasil.
Descrição: Mônade, radialmente simétrico, isopolar, circular a retangular; stephanoporado, 4
poros circulares com 2μm de comprimento e 2μm de largura, marginado, margem com 2μm de
espessura e 2μm de largura produzida pelo espessamento da sexina perto dos poros; costado,
costa com 1.5μm de largura; tectado, nexina com 0.3μm de espessura, columelas com 0.4μm
de comprimento, indistintas, teto com 0.3μm de espessura. Escabrado, escabras com 0.5μm de
largura separadas 0.5μm.
Dimensões: Comprimento do diâmetro equatorial 25μm; Largura do diâmetro equatorial 23μm;
comprimento/largura1.08; nm=1, no=1.
Observações: Não existe gênero para pólen escabrado e stephanoporado, então o tipo será
colocado provisoriamente neste gênero que descreve grãos stephanoporados e verrugados.
Derivação do nome: Em homenagem ao rio Uatumã da Amazônia.

61
Prancha 1
1. Echinosporis densiechinatus D’Apolito et al., in prep
2. Laevigatosporites caoticus D'Apolito et al., in prep
3. Laevigatosporites fossulatus D'Apolito et al., in prep
4. Laevigatosporites tibuensis (Van der Hammen 1956a) Jaramillo e Dilcher (2001)
5. Polypodiisporites discretus D’Apolito et al., in prep
6. Polypodiisporites aff. speciosus Sah (1967)
7. Polypodiisporites usmensis (Van der Hammen, 1956a) Khan e Martin (1972)
8. Reticulosporis irregularis sp. nov. (Holotipo)
9. Reticulosporis irregularis sp. nov. (Parátipo)
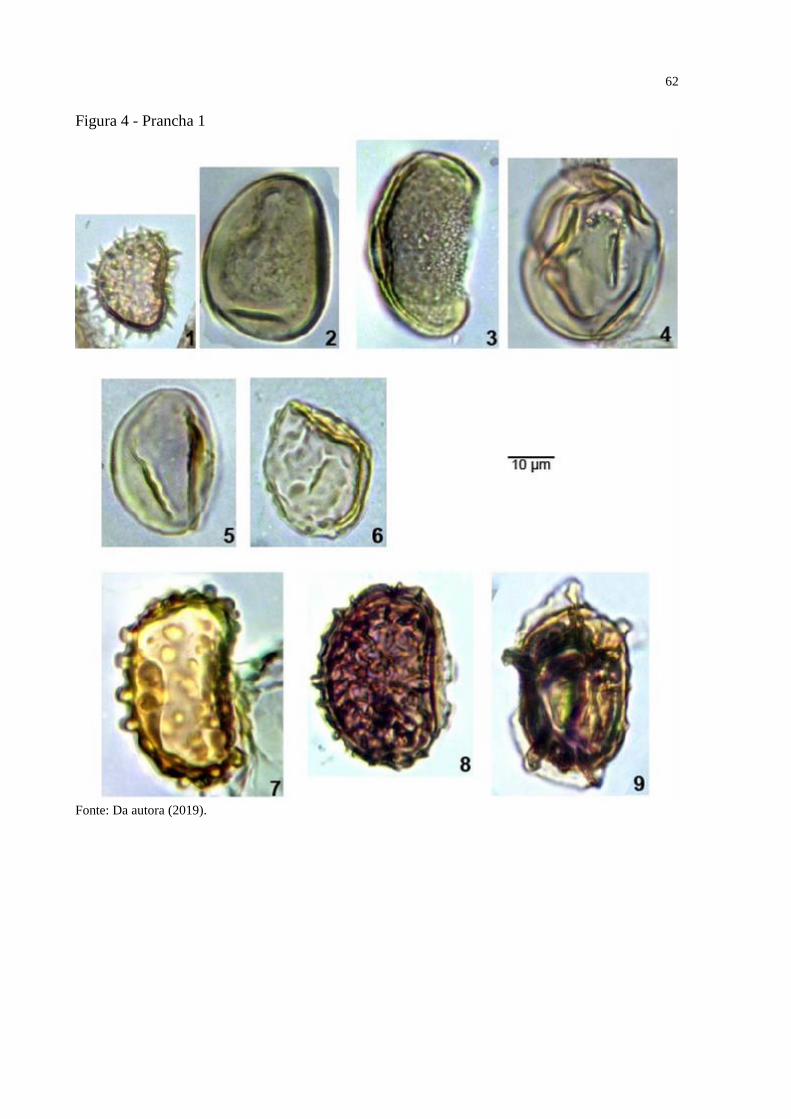
62
Figura 4 - Prancha 1
Fonte: Da autora (2019).

63
Prancha 2
1*. Azolla sp.
2-3. Baculatisporites indecisus sp. nov (Holotipo)
4-5. Camarozonosporites crassus Silva-Caminha et al. (2010)
6-7. Camarozonosporites ondulatus sp. nov.
8-9. Camarozonosporites segmentatus sp. nov.
10-11. Cicatricosisporites pseudograndiosus D'Apolito et al., in prep
12. Crassoretitriletes vanraadshoovenii Germeraad et al. (1968)

64
Figura 5 - Prancha 2
Fonte: Da autora (2019).

65
Prancha 3
1-2. Deltaidospora adriennis (Potonié e Gelletich, 1933) Frederiksen (1983)
3-4. Distaverrusporites muratus sp. nov.
5-6. Echinatisporis brevispinosus Jaramillo e Dilcher (2001)
7-8. Echinatisporis circularis Silva-Caminha et al. (2010)
9-10. Echinatisporis muelleri (Regali et al., 1974) Silva-Caminha et al. (2010)
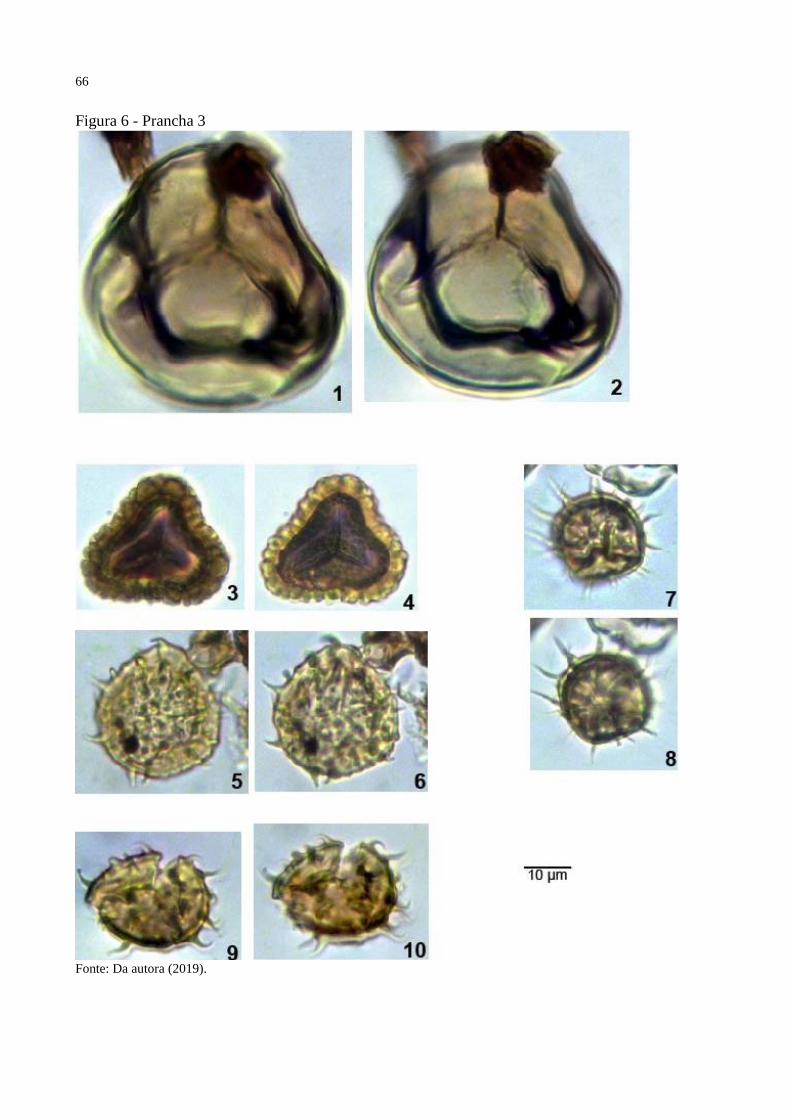
66
Figura 6 - Prancha 3
Fonte: Da autora (2019).

67
Prancha 4
1-2. Echinatisporis trianguloconvexus sp. nov.
3-4. Foveotriletes noctis sp. nov
5-6. Foveotriletes pseudornatus sp. nov.
7-8. Hamulatisporis bare D'Apolito et al., in prep
9-10. Hydrosporis minor Silva-Caminha et al. (2010)
11-12. Kuylisporites waterbolkii Potonié 195613-14. Magnastriatites grandiosus
(Kedves and Sole de Porta, 1963) Dueñas (1980)
15-16. Neoraistrickia dubiosa D'Apolito et al., in prep
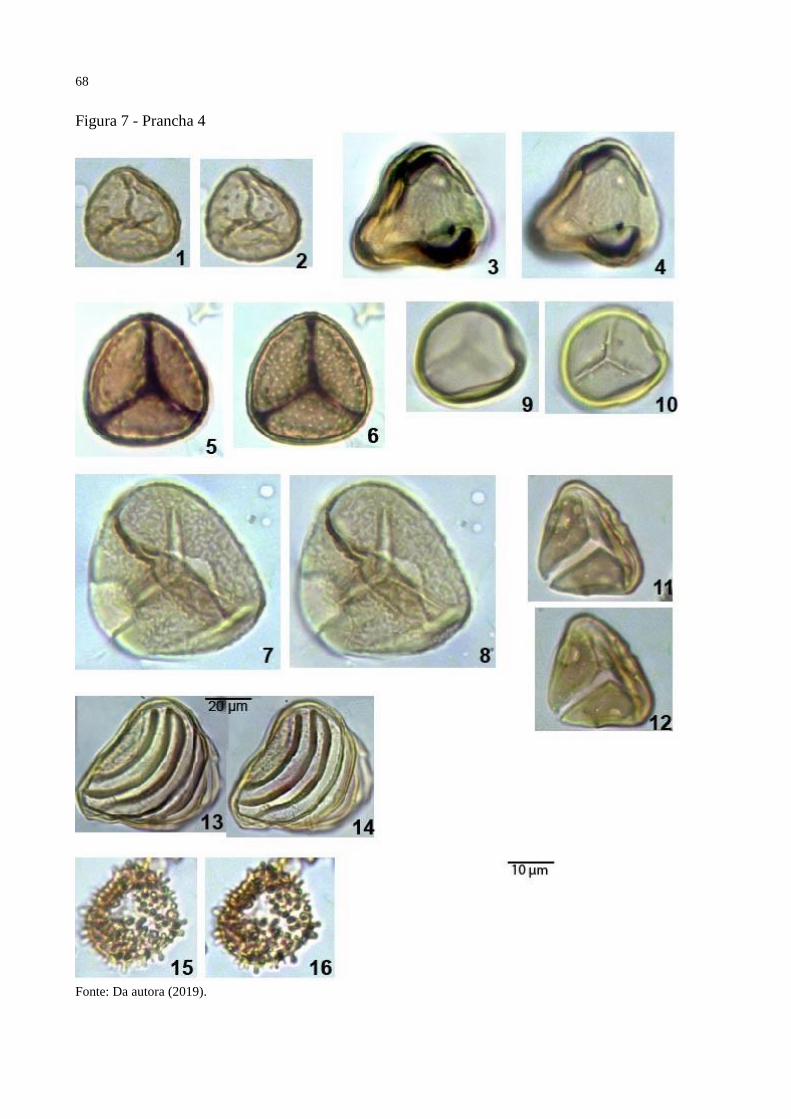
68
Figura 7 - Prancha 4
Fonte: Da autora (2019).

69
Prancha 5
1-2. Nijssenosporites fossulatus Lorente (1986)
3-4. Polypodiaceoisporites? amazonensis Silva-Caminha et al. (2010)
5-6. Polypodiaceoisporites pseudopsilatus Lorente (1986)
7-8. Psilatriletes lobatus Hoorn (1994) 9-10.Psilatriletes marginatus D'Apolito et al.,
in prep
11-12. Pteridaceoisporis gemmatus Silva-Caminha et al. (2010)

70
Figura 8 - Prancha 5
Fonte: Da autora (2019).

71
Prancha 6
1-2. Retitriletes altimuratus Silva-Caminha et al. (2010)
3-4. Rugulatisporites reticulatuss sp. nov.
5-6. Rugulatisporites rugulatus sp. nov.
7-8. Striatriletes saccolomoides Jaramillo et al. (2010)
9-10. Verrucatotriletes circularis sp. nov.
11-12. Verrucatotriletes etayoi Dueñas (1980)

72
Figura 9 - Prancha 6
Fonte: Da autora (2019).

73
Prancha 7
1-2. Clavainaperturites microclavatus Hoorn (1994a)
3-4. Crotonoidaepollenites reticulatus Silva-Caminha et al. (2010)
5-6. Inapertuporpollenites baculatus sp. nov.
7-8. Polyadopollenites macroreticulatus Salard-Cheboldaeff (1974)
9-10. Arecipites? polaris Silva-Caminha et al. (2010)
11-12. Arecipites regio (Van der Hammen e Garcia, 1966) Jaramillo e Dilcher (2001)
13-14. Clavamonocolpites lorentei Muller et al. (1987)
15-16. Foveomonocolpites amadus sp. nov.
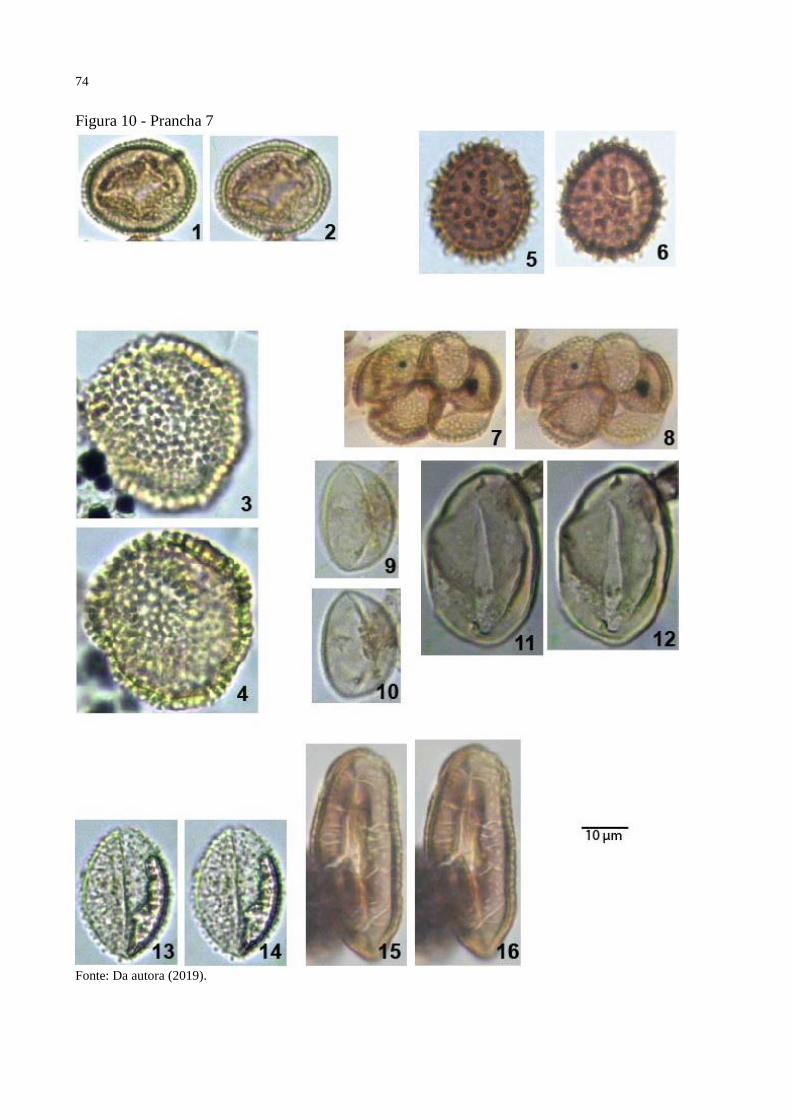
74
Figura 10 - Prancha 7
Fonte: Da autora (2019).

75
Prancha 8
1-2. Mauritiidites franciscoi var. franciscoi (Van der Hammen, 1956) Van Hoeken
Klinkenberg (1964)
3-4. Mauritiidites franciscoi var. minutus Van der Hammen e Garcia (1966)
5-6. Monocolpopollenites digitus sp. nov.
7-8. Psilamonocolpites amazonicus Hoorn (1993)
9-10. Psilamonocolpites delicatus sp. nov.
11-12. Psilamonocolpites medius (Van der Hammen, 1956) Van der Hammen e Garcia
(1966)
13-14. Retimonocolpites absyae Hoorn (1993)
15-16. Proxapertites psilatus Sarmiento (1992)

76
Figura 11 - Prancha 8
Fonte: Da autora (2019).

77
Prancha 9
1-2. Proxapertites tertiaria Van der Hammen e Garcia (1966)
3-4. Trichotomosulcites amazonicusD'Apolito et al., in prep
5-6. Grimsdalea magnaclavata Germeraad et al. (1968)
7-8. Monoporopollenites annulatus (Van der Hammen, 1954) Jaramillo e Dilcher
(2001)
9-10. Monoporopollenites annuloides D'Apolito et al., in prep
11-12. Dicolpopollis? costatus D'Apolito et al., in prep
13-14. Bombacacidites lorenteae (Hoorn, 1993) D'Apolito et al., in prep

78
Figura 12 - Prancha 9
Fonte: Da autora (2019).

79
Prancha 10
1-2. Crassiectoapertites columbianus (Dueñas, 1980) Lorente (1986)
3-4. Crototricolpites finitus Silva-Caminha et al. (2010)
5-6. Foveotricolpites colpiconstrictus (Hoorn, 1994) D'Apolito et al., in prep
7-8. Foveotricolpites simplex Gonzalez-Guzman (1967) (D'Apolito et al., in prep)
9-10. Ladakhipollenites baitacolumellatus D'Apolito et al., in prep
11-12. Ladakhipollenites floratusSilva-Caminha et al. (2010)
13-15. Loranthacites apocirculoreticulatussp. nov. (Holotipo)
16-17. Loranthacites apocirculoreticulatussp. nov. (Paratipo)
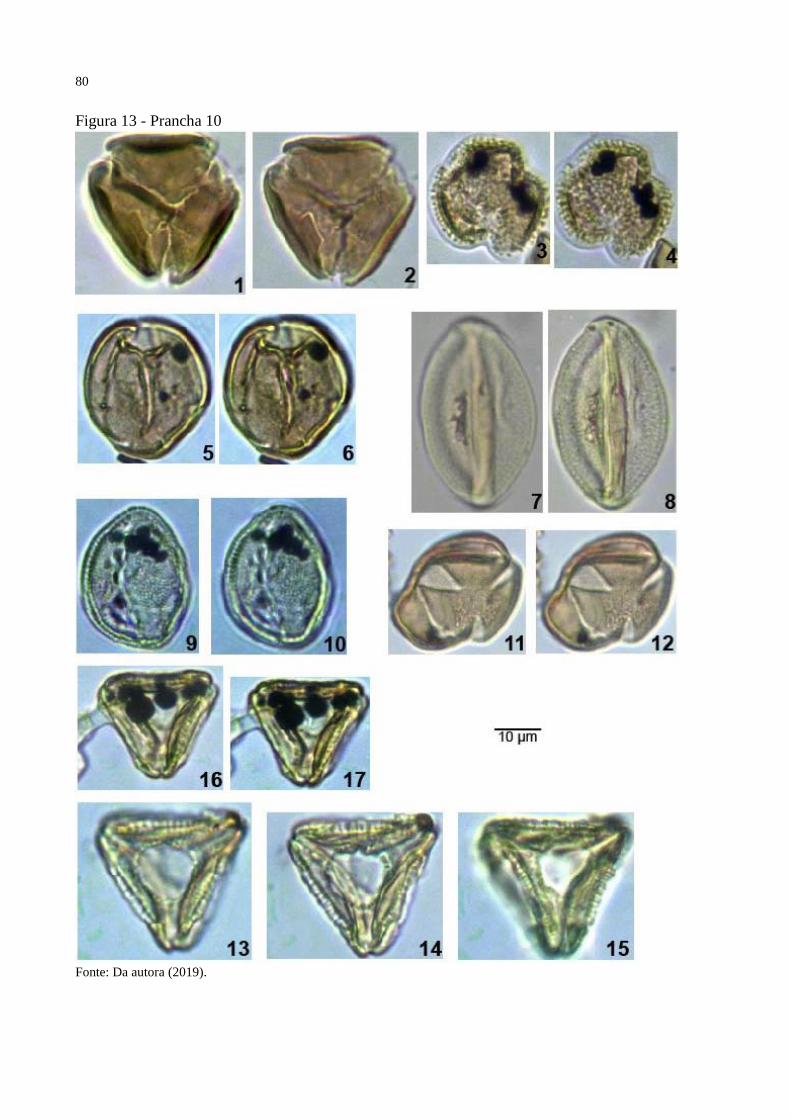
80
Figura 13 - Prancha 10
Fonte: Da autora (2019).

81
Prancha 11
1-2. Perfotricolpites digitatus Gonzalez-Guzman (1967)
3-4. Psilatricolpites colpiconstrictus Van Hoeken-Klinkenberg (1966)
5-6. Retibrevitricolpites pseudoretibolus D'Apolito et al., in prep
7-8. Rhoipites? hoornii sp. nov.
9-10. Rousea? simplex sp. nov.
11-12. Tricolpites? pseudoclarensis Silva-Caminha et al. (2010)
13-14. Bombacacidites araracuarensis Hoorn (1994a)
15-16. Bombacacidites baculatus Muller et al. (1987)

82
Figura 14 - Prancha 11
Fonte: Da autora (2019).

83
Prancha 12
1-2. Bombacacidites brevis (Dueñas, 1980) Muller et al. (1987)
3-4. Bombacacidites nacimientoensis (Anderson, 1960) Elsik (1968)
5-6. Cichoreacidites? flamulatus Leite et al., in prep
7-8. Cichoreacidites longispinosus (Lorente, 1986) Silva-Caminha et al. (2010)
9-10. Echitricolporites cruzi Leite et al., in prep
11-12. Echitricolporites distintussp. nov.
13-14. Echitricolporites insanussp. nov.
15-16. Echitricolporites spinosus Van der Hammen (1956)
17-18. Foveotricolporites miritiense sp. nov. (Holotipo)
19-20. Foveotricolporites miritiense sp. nov. (Paratipo)

84
Figura 15 - Prancha 12
Fonte: Da autora (2019).

85
Prancha 13
1-2. Foveotricolporites parintinsense sp. nov.
3-4. Ilexipollenites tropicallis Silva-Caminha et al. (2010)
5-6. Ladakhipollenites? caribbiensis (Muller et al 1987) Silva-Caminha et al. (2010)
7-8. Ladakhipollenites? cassioides D'Apolito et al., in prep
9-10. Ladakhipollenites? compressus sp. nov.
11-12. Ladakhipollenites? corvattatus D'Apolito et al., in prep
13-14. Ladakhipollenites? garzonni Hoorn (1993) (D'Apolito et al., in prep)
15-16. Ladakhipollenites? irandubaensis sp. nov.
17-18. Ladakhipollenites? labiatus Hoorn (1993) ( D'Apolito et al., in prep)
19-20. Ladakhipollenites? lolongatus D'Apolito et al. (2018)
21-22. Ladakhipollenites? magniporatus Hoorn (1993) (D'Apolito et al., in prep)
23-24. Ladakhipollenites? posterus sp. nov.
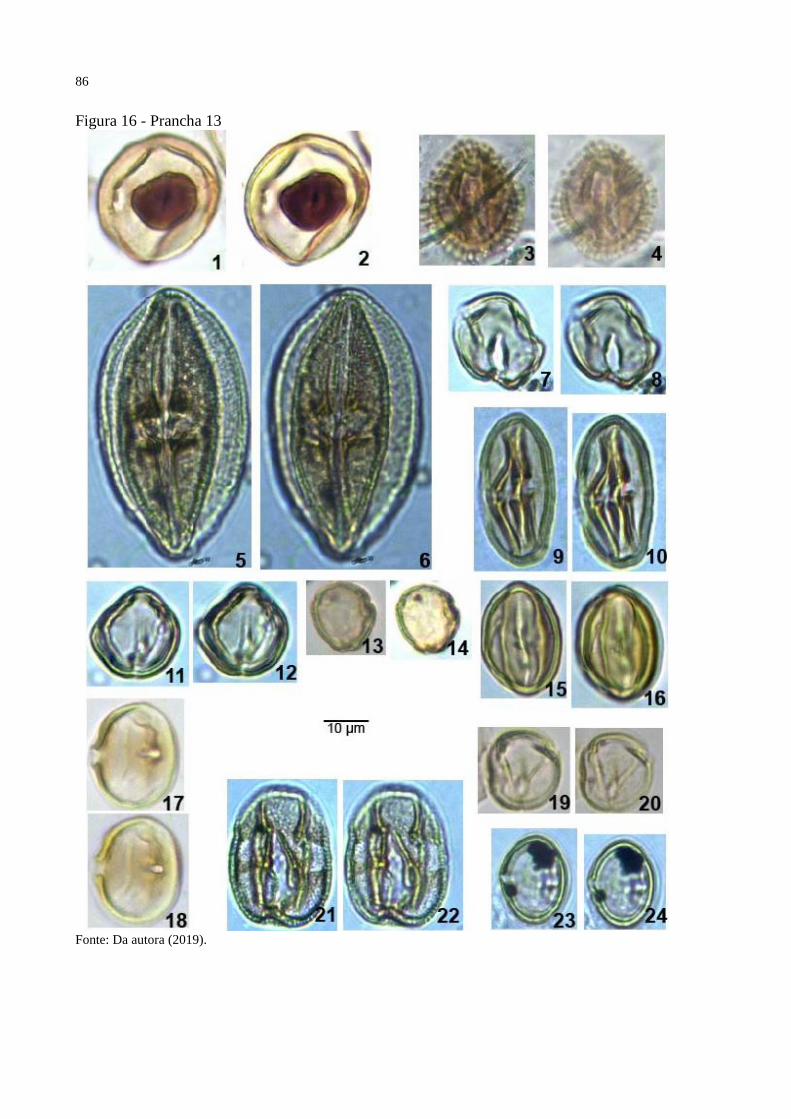
86
Figura 16 - Prancha 13
Fonte: Da autora (2019).

87
Prancha 14
1-2. Ladakhipollenites? silvaticus Hoorn (1993) (D'Apolito et al., in prep)
3-4. Ladakhipollenites? sphericus D'Apolito et al., in prep
5-6. Ladakhipollenites? xatanawensis D'Apolito et al., in prep
7-8. Lakhiapollis costatus Silva-Caminha et al. (2010)
9-10. Lanagiopollis crassa (Van der Hammen e Wymstra, 1964) Frederiksen (1988)
11-12. Malvacipolloides maristellae (Muller et al., 1987) Silva-Caminha et al. (2010)
13-14. Margocolporites carinae Leite et al., in prep
15-16. Retibrevitricolporites retibolus Leidelmeyer (1966)
17-18. Paleosantalaceaepites cingulatusJaramillo et al. (2010)
19-20. Psilabrevitricolporites triangularis (Van der Hammen e Wymstra (1964)
Jaramillo e Dilcher (2001)
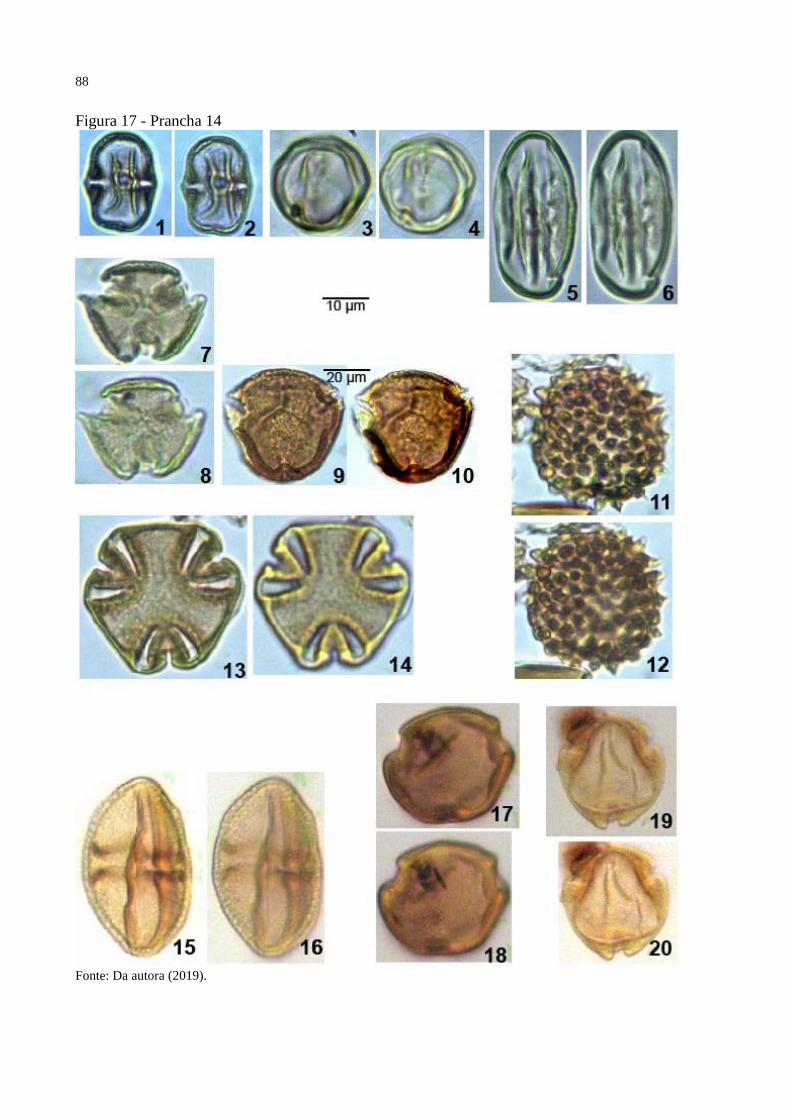
88
Figura 17 - Prancha 14
Fonte: Da autora (2019).

89
Prancha 15
1-2. Raunculacidites operculatus (Van der Hammen e Wymstra, 1964) Jaramillo e
Dilcher (2001)
3-4. Retibrevitricolporites retibolus Leidelmeyer (1966)
5-6. Retibrevitricolporites solimoensis Hoorn (1993) (D'Apolito et al., in prep)
7-8. Retibrevitricolporites superbus sp. nov.
9-10. Retibrevitricolporites tabatinguenses sp. nov.
11-12. Retitrescolpites benjaminense D'Apolito et al., in prep
13-14. Retitrescolpites grossus D'Apolito et al., in prep
15-16. Retitrescolpites? irregularis (Van der Hammen e Wymstra, 1964) Jaramillo e
Dilcher (2001)
17-18. Retitrescolpites? traversei Silva-Caminha et al. (2010)
19-20. Retitricolporites ticoneorum Hoorn (1993)
21-22. Rhoipites caputoi Hoorn (1993) (D'Apolito et al., in prep)
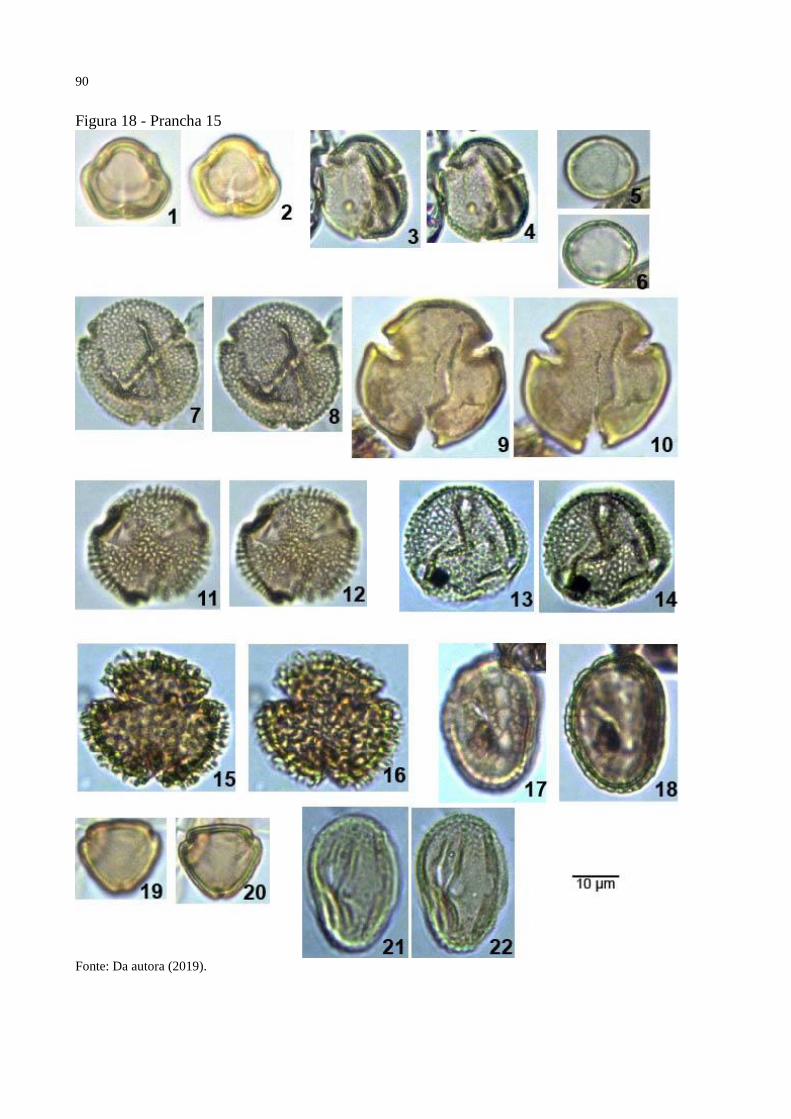
90
Figura 18 - Prancha 15
Fonte: Da autora (2019).

91
Prancha 16
1-2. Rhoipites crassicostatus Van der Hammen e Wymstra (1964) (D'Apolito et al., in
prep)
3-4. Rhoipites crassiexinatus sp. nov.
5-6. Rhoipites dignus sp. nov.
7-8. Rhoipites figueiredoensis sp. nov. tirar foto em que se evidencia retículo
9-10. Rhoipites grossomurus D'Apolito et al., in prep.
11-12. Rhoipites guianensis (Van der Hammen e Wymstra, 1964) Jaramillo e Dilcher
(2001)
13-14. Rhoipites manausensis D'Apolito et al. (2018)
15-16. Rhoipites normalis sp. nov.
17-18. Rhoipites oblatus (Hoorn, 1994) D'Apolito et al., in prep
19-20. Rhoipites poropartitus D'Apolito et al., in prep
21-22. Rhoipites superlolongatus sp. nov.
23-24. Rugutricolporites arcus Hoorn (1993)

92
Figura 19 - Prancha 16
Fonte: Da autora (2019).

93
Prancha 17
1-2. Rugutricolporites felix sp. nov.
3-4. Rugutricolporites perfuratus sp. nov.
5-6. Siltaria dilcheri Silva-Caminha et al. (2010)
7-8. Siltaria pseudodilcheri sp. nov.
9-10. Siltaria santaisabelensis (Hoorn, 1994) Silva-Caminha et al. (2010)
11-12. Striasyncolpites anastomosatus Silva-Caminha et al. (2010)
13-14. Striatopollis catatumbus (Gonzalez-Gusman, 1967) Takahashi e Jux (1989)
15-16. Striatopollis macrolobium D'Apolito et al., in prep
17-18. Striatricolporites caminhae sp. nov.
19-20. Striatricolporites magnificus sp. nov.
21-22. Striatricolporites poloreticulatusSilva-Caminha et al. (2010)
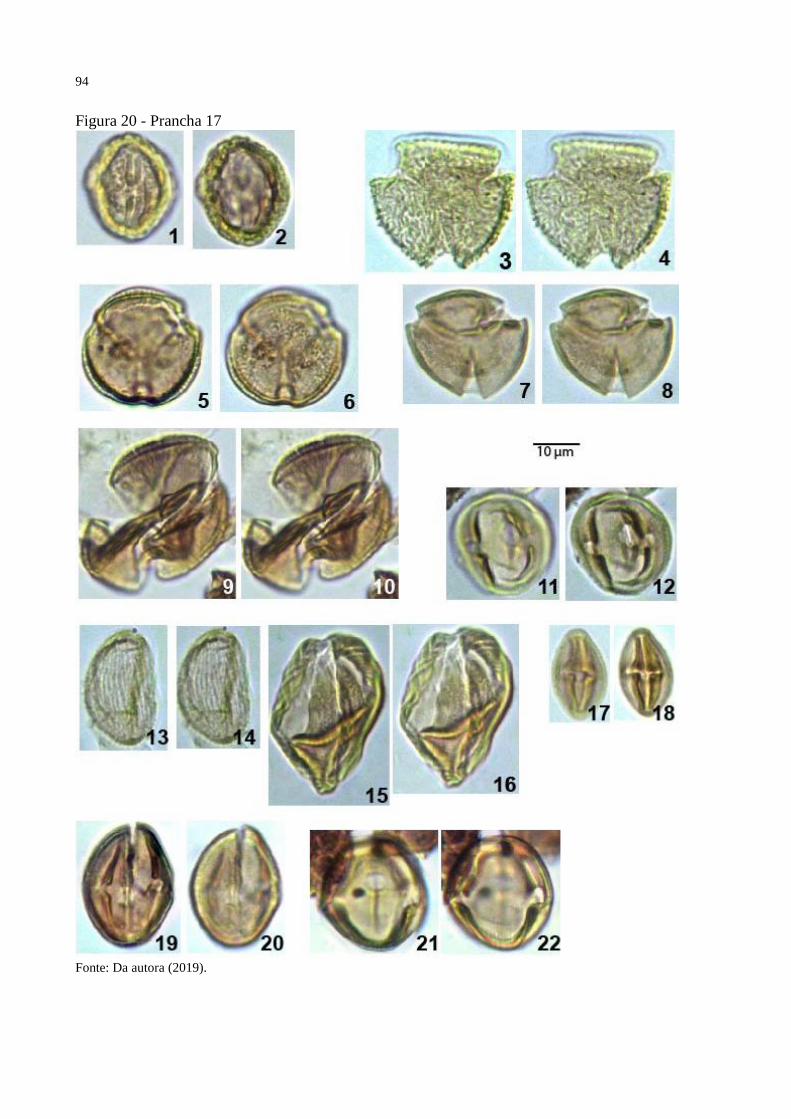
94
Figura 20 - Prancha 17
Fonte: Da autora (2019).

95
Prancha 18
1-2. Striatricolporites tenuissimus Dueñas (1980)
3-4. Syncolporites microreticulatus sp. nov. (Holotipo)
5. Syncolporites microreticulatus sp. nov. (Parátipo)
6-7. Syncolporites tenuicolpatus sp.nov. (Holotipo)
8. Syncolporites tenuicolpatus sp. nov. (Parátipo)
9-10. Verrutricolporites intectatus sp. nov.
11-12. Verrutricolporites poricostatus sp. nov.
13-14. Verrutricolporites simplex D'Apolito et al., in prep
15-16. Zonocostites ramonae Dueñas (1980)
17-18. Byttneripollis ruedae Silva-Caminha et al. (2010)
19-20. Corsinipollenites oculusnoctis (Thiergart, 1940) Nakoman (1965)
21-22. Echitriporites nanuspinosus sp. nov.
23-24. Florschuetzia salafraria D'Apolito et al., in prep

96
Figura 21 - Prancha 18
Fonte: Da autora (2019).

97
Prancha 19
1-2. Proteacidites triangulatus Lorente (1986)
3-4. Psilatriporites sarmientoi Hoorn (1993)
5-6. Retitriporites crotononicollumelatus (Jaramillo et al. 2010) D'Apolito et al., in prep
7-8. Retitriporites typicus Gonzalez-Guzman (1967)
9-10. Scabratriporites annulatus sp. nov.
11-12. Clavastephanocolpites crotonoides Van der Hammen e Wymstra (1964)
13-14. Ctenolophonidites suigeneris Silva-Caminha et al. (2010)
15-16. Lymingtonia splendens D'Apolito et al., in prep
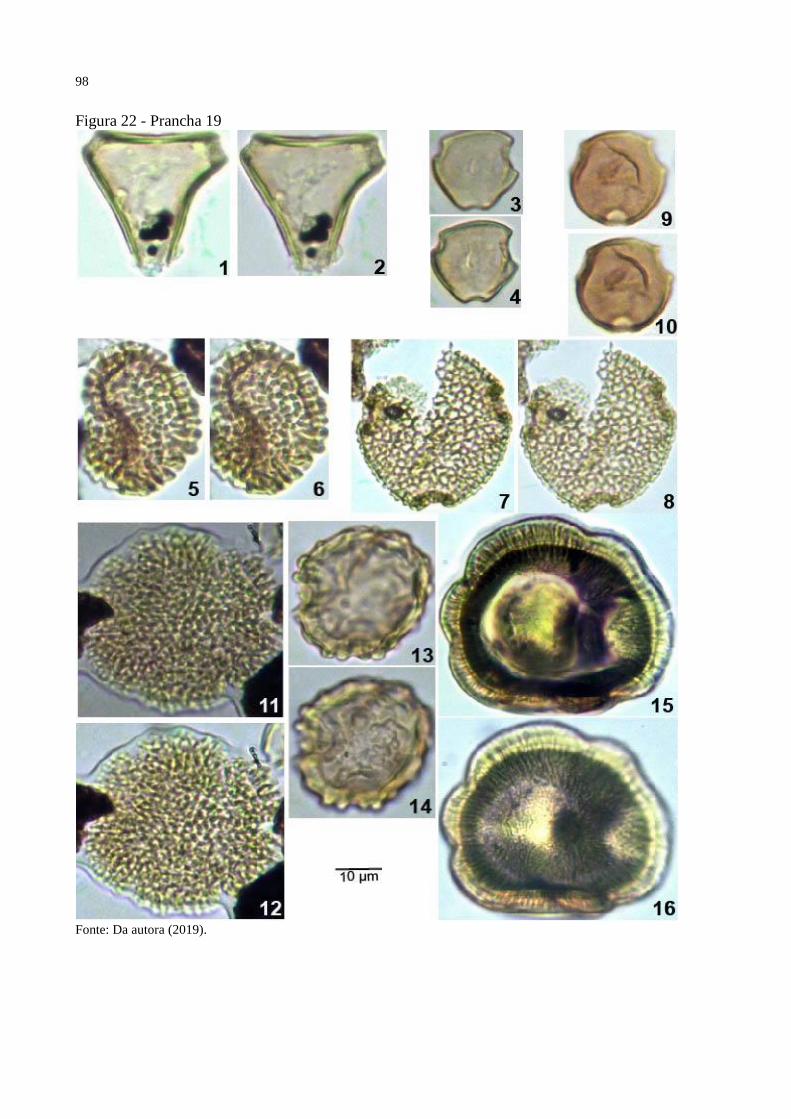
98
Figura 22 - Prancha 19
Fonte: Da autora (2019).

99
Prancha 20
1-2. Retistephanocolpites angeli Leidelmeyer (1966)
3-4. Retistephanocolpites pardoi Leite et al., in prep
5-6. Verrustephanocolpites guimaraesensis sp. nov.
7-8. Heterocolpites brevicolpatus Silva-Caminha et al. (2010)
9-10. Heterocolpites incomptus Hoorn (1993)
11-12. Heterocolpites rotundus Hoorn (1993)
13-14. Heterocolpites verrucosus Hoorn (1993)
15-16. Jandufouria minor Germeraad et al. (1968)
17-18. Psilastephanocolporites fissilis Lorente (1986)
19-20. Psilastephanocolporites marinamensis Hoorn (1994)
21-22. Psilastephanocolporites meliosus D'Apolito et al., in prep
23-24. Cyperaceapolis rotundus sp. nov. (Holotipo)
25-26. Cyperaceapolis rotundus sp. nov. (Parátipo)
27-28. Echiperiporites akanthos Van der Hammen e Wymstra (1964)
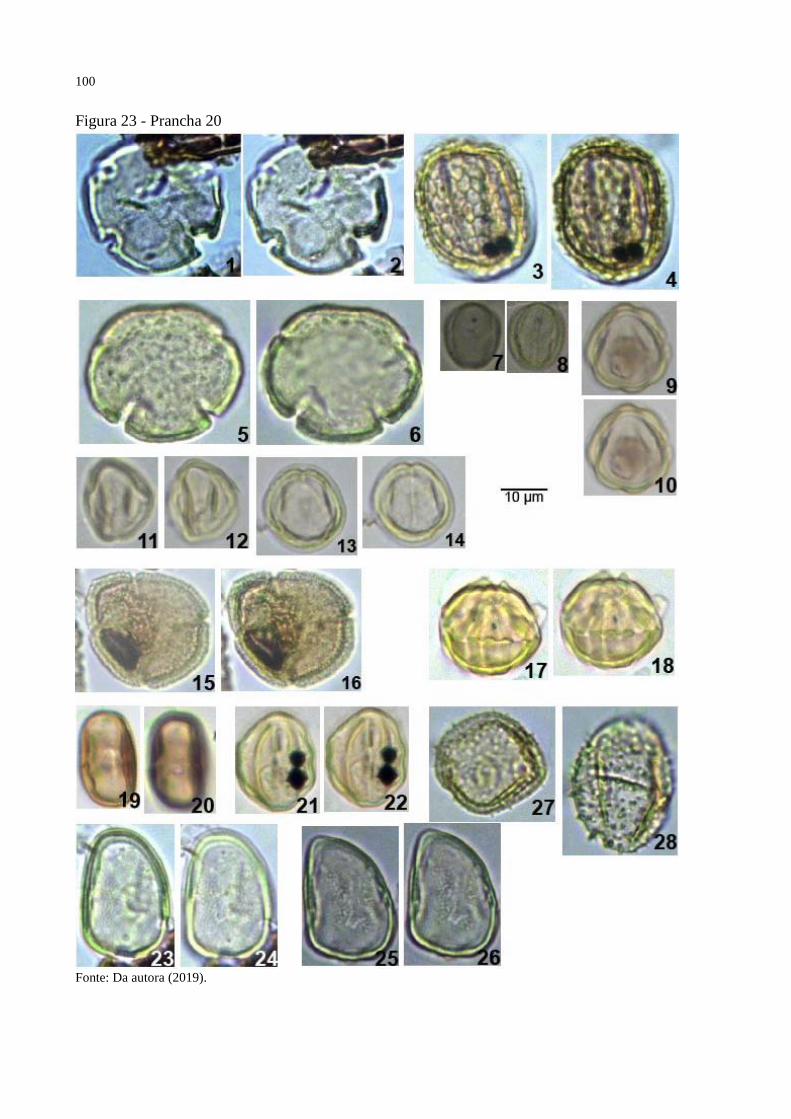
100
Figura 23 - Prancha 20
Fonte: Da autora (2019).

101
Prancha 21
1-2. Echiperiporites estelae Germeraad et al. (1968)
3-4. Echiperiporites germeraadii Leite et al., in prep
5-6. Echiperiporites jutaiensis Silva-Caminha et al. (2010)
7-8. Echiperiporites lophatus Silva-Caminha et al. (2010)
9-10. Multiporopollenites intermedius D'Apolito et al., in prep
11-12. Parsonsidites? brenacii Silva-Caminha et al. (2010)
13-14. Perisyncolporites pokornyi Germeraad et al. (1968)
15-16. Psilaperiporites annulatus sp. nov.
17-18. Psilaperiporites multiporatus Hoorn (1994)

102
Figura 24 - Prancha 21
Fonte: Da autora (2019).

103
Prancha 22
1-2. Malvacipollis minutispinulosa D'Apolito et al., in prep
3-4. Pachidermites pseudoalnus sp. nov.
5-6. Psilastephanoporites herngreenii Hoorn (1993)
7-8. Psilastephanoporites tesseroporus Regali et al. (1974)
9-10. Retistephanoporites crassiannulatus Lorente (1986)
11-12. Retistephanoporites mimosus sp. nov.
13-14. Thymelipollis amazonicus D'Apolito et al., in prep
15-16. Verrustephanoporites? uatumaensis sp. nov.

104
Figura 25 - Prancha 22
Fonte: Da autora (2019).

105
Prancha 23
1. Pediastrum spp.
2. Cleistosphaeridium spp.
3. Leiosphaeridia spp.
4. Lejeunecysta? spp.
5. Lingoludinium machaerophorum (Deflandre e Cookson, 1955) Wall (1967)
6. Operculidinium israelianum (Rossignol) Wall (1967)
7. Quadrinia? condida De Verteuil e Norris (1996)

106
Figura 26 - Prancha 23
Fonte: Da autora (2019).

107
Prancha 24
1. Quadrinia “variabilis”
2. Rounded Brown Cysts (RBCs)
3. Spiniferites spp.
4. Spiniferites spp.
5. Spiniferites spp.
6. Trinovantedinium ferugnomatum De Verteuil e Norris (1996)
7. Trinovantedinium? spp.

108
Figura 27 - Prancha 24
Fonte: Da autora (2019).

109
Prancha 25
1. Ammonidium “belissimum”RW
2. Ammonidium spp. RW
3. Ammonidium spp. RW
4. Ammonidium spp. RW
5. Oligosphaeridium spp. RW
6. Verihachium spp. RW
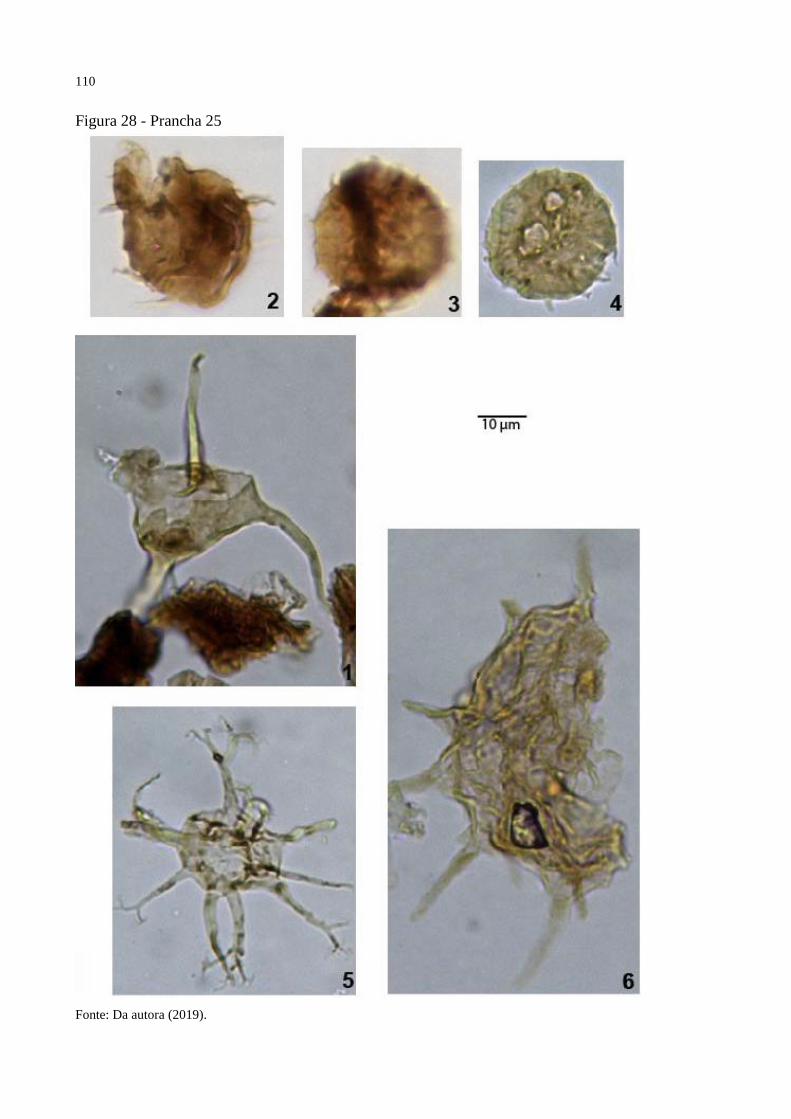
110
Figura 28 - Prancha 25
Fonte: Da autora (2019).

111
4.2 Bioestratigrafia
A LOC feita pela correlação do perfil 1-AS-15 AM com o perfil 105-AM foi construída
utilizando os pontos: FAD do Retimonocolpites absyae em -255.5m, FAD do Malvacipolloides
maristellae em -229.4m, ACME do Mauritiidites franciscoi em -203.2m, FCO da Grimsdalea
Magnaclavata em -156.4m, FAD do Echiperiporites germeraadi em -94.5m e a FAD do
Retitrescolpites? traversei em -66.1m. Os sedimentos foram estabelecidos como pertencentes
as palinozonas T16,T15,T14 e T13 de Jaramillo et al. (2011), da seguinte maneira: palinozona
T16 (12.7 a 7.1 Ma) do topo (-37.5m) até -94.5m, palinozona T15 (14.2 a 12.7 Ma) de -94.5m
a -109.25, palinozona T14 (16.1 a 14.2 Ma) de -109.25Ma -146.9m e palinozona T13 (17.7 a
16.1 Ma) de -146.9m até a base (-255.5 m) (Figuras 29, 30).
Figura 29 - Distribuição dos principais marcadores palinoestratigráficos do perfil 1AS-15-AM
Fonte: Da autora (2019), produzido utilizando o software R.

112
Figura 30 - Correlação gráfica do perfil 1-AS-15-AM com o perfil 1-AS-105-AM
Fonte: Da autora (2019), produzido utilizando o software R.
4.3 Análise numérica
4.3.1 Análise de similaridade
O índice de Jaccard obtido (Tabela 1), por estar próximo de 1, indica grande
dessemelhança das amostras com relação a sua composição, enquanto o índice de Distância
Euclidiana, por seu alto valor, indica a grande diferença das palinofloras com relação a sua
abundância. O índice de Bray-curtis também evidencia essa diferença com relação à
abundância, com um valor próximo de 1 (Tabela 1).
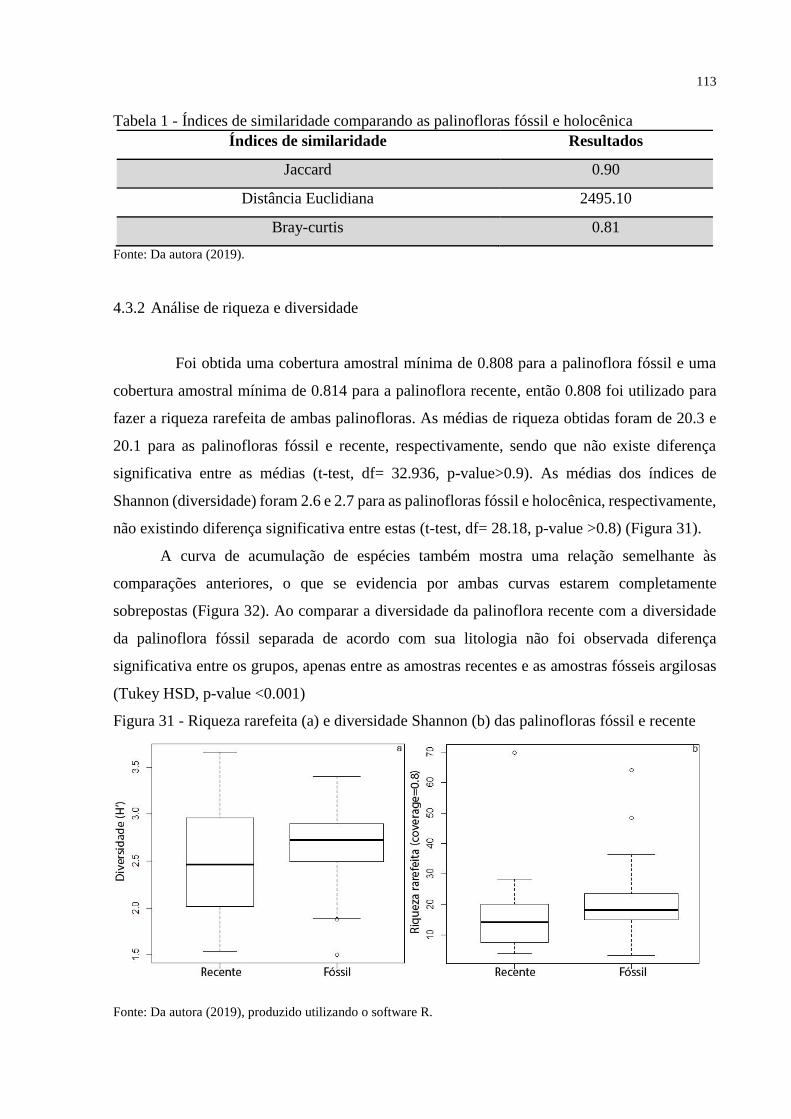
113
Tabela 1 - Índices de similaridade comparando as palinofloras fóssil e holocênica
Índices de similaridade Resultados
Jaccard 0.90
Distância Euclidiana 2495.10
Bray-curtis 0.81
Fonte: Da autora (2019).
4.3.2 Análise de riqueza e diversidade
Foi obtida uma cobertura amostral mínima de 0.808 para a palinoflora fóssil e uma
cobertura amostral mínima de 0.814 para a palinoflora recente, então 0.808 foi utilizado para
fazer a riqueza rarefeita de ambas palinofloras. As médias de riqueza obtidas foram de 20.3 e
20.1 para as palinofloras fóssil e recente, respectivamente, sendo que não existe diferença
significativa entre as médias (t-test, df= 32.936, p-value>0.9). As médias dos índices de
Shannon (diversidade) foram 2.6 e 2.7 para as palinofloras fóssil e holocênica, respectivamente,
não existindo diferença significativa entre estas (t-test, df= 28.18, p-value >0.8) (Figura 31).
A curva de acumulação de espécies também mostra uma relação semelhante às
comparações anteriores, o que se evidencia por ambas curvas estarem completamente
sobrepostas (Figura 32). Ao comparar a diversidade da palinoflora recente com a diversidade
da palinoflora fóssil separada de acordo com sua litologia não foi observada diferença
significativa entre os grupos, apenas entre as amostras recentes e as amostras fósseis argilosas
(Tukey HSD, p-value <0.001)
Figura 31 - Riqueza rarefeita (a) e diversidade Shannon (b) das palinofloras fóssil e recente
Fonte: Da autora (2019), produzido utilizando o software R.

114
Figura 32 - Curva de acumulação de espécies paras as amostras fósseis (linhas cinzas) e
recentes (linhas pretas). Intervalo de confiança mostrado de 95%.
Fonte: Da autora (2019), produzido utilizando o software R.
4.3.3 Ordenação das comunidades
O estresse obtido para a análise nMDS foi ≈ 0.118, o que indica que a análise representa
de maneira boa, porém não excelente, os dados. Ao comparar os scores do eixo 1 da análise foi
possível observar a clara segregação da palinoflora fóssil da palinoflora recente (t.test, df=
21.518, p-value<0.001). Entre as amostras recentes, foi evidenciada separação do subambiente
de várzea dos subambientes de margem de lago e pântano (t.test, df= 11.926, p-value<0.001),
segregando as amostras recentes em dois grupos. A palinoflora fóssil também mostrou grande
diferença com relação às amostras recente de várzea (t.test, df=16.878, p-value<0.001) e de
margem de lago e pântano (t.test, df= 7.612, p-value<0.001). A análise de espécie indicadora
(Indval) apontou 109 tipos polínicos indicadores, 33 para o grupo fóssil, 49 para o grupo várzea
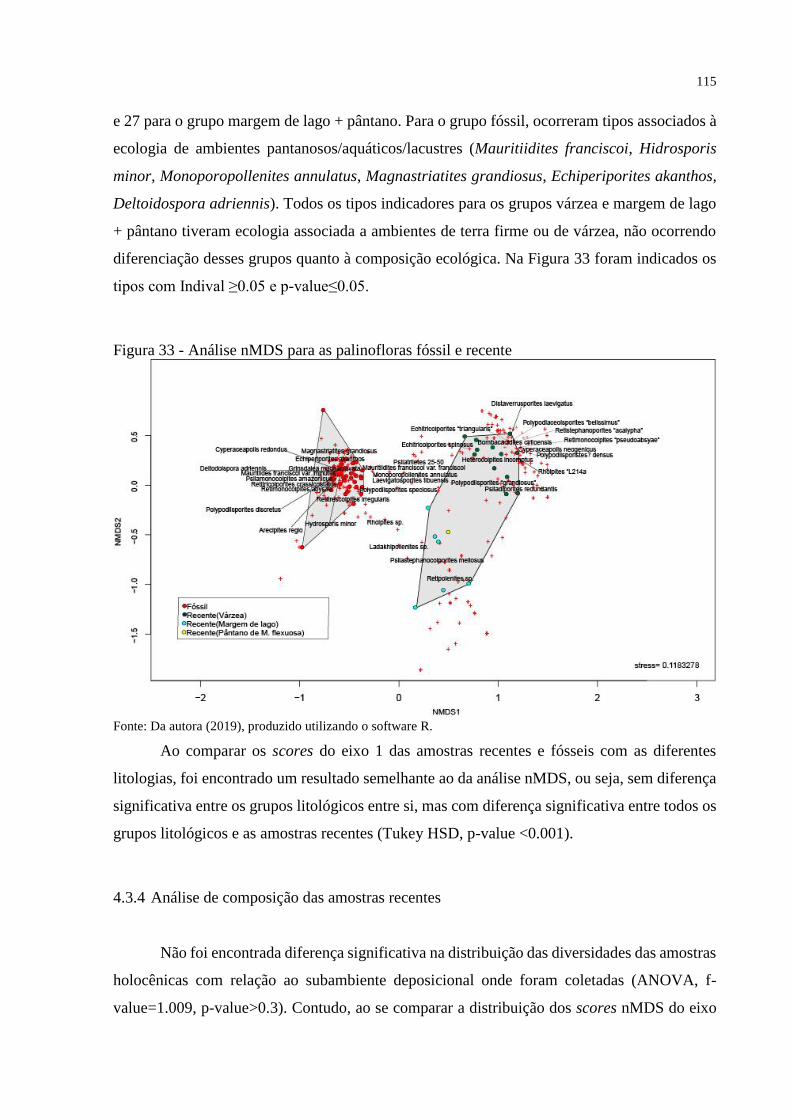
115
e 27 para o grupo margem de lago + pântano. Para o grupo fóssil, ocorreram tipos associados à
ecologia de ambientes pantanosos/aquáticos/lacustres (Mauritiidites franciscoi, Hidrosporis
minor, Monoporopollenites annulatus, Magnastriatites grandiosus, Echiperiporites akanthos,
Deltoidospora adriennis). Todos os tipos indicadores para os grupos várzea e margem de lago
+ pântano tiveram ecologia associada a ambientes de terra firme ou de várzea, não ocorrendo
diferenciação desses grupos quanto à composição ecológica. Na Figura 33 foram indicados os
tipos com Indival ≥0.05 e p-value≤0.05.
Figura 33 - Análise nMDS para as palinofloras fóssil e recente
Fonte: Da autora (2019), produzido utilizando o software R.
Ao comparar os scores do eixo 1 das amostras recentes e fósseis com as diferentes
litologias, foi encontrado um resultado semelhante ao da análise nMDS, ou seja, sem diferença
significativa entre os grupos litológicos entre si, mas com diferença significativa entre todos os
grupos litológicos e as amostras recentes (Tukey HSD, p-value <0.001).
4.3.4 Análise de composição das amostras recentes
Não foi encontrada diferença significativa na distribuição das diversidades das amostras
holocênicas com relação ao subambiente deposicional onde foram coletadas (ANOVA, f-
value=1.009, p-value>0.3). Contudo, ao se comparar a distribuição dos scores nMDS do eixo

116
1 com o subambiente de coleta, foi observada uma diferença significativa (ANOVA, f-
value=21.18, p-value <0.001) com as amostras de várzea e margem de lago, com diferença
significativa entre si (Tukey HSD, p-value <0.001) (Figura 34).
Figura 34 - Subambientes de coleta comparados com a diversidade (a) e scores nMDS do eixo
1 (b)
Fonte: Da autora (2019), produzido utilizando o software R.
4.3.5 Análise ecológica
Do total de 463 tipos polínicos encontrados no presente trabalho foi possível encontrar
afinidades botânicas para 150 destes, sendo atribuída família e quando possível genero e
espécie. Os ambientes ecológicos aos quais esses tipos estão associados foram definidos como
floresta seca, exógeno(não amazônico), hidrosseral (ambiente aquático de água doce), terras
baixas (terras baixas amazônicas não diferenciadas), lianas de terras baixas, manguezal,
montano, ripário (margem de rios), terra firme,várzea e campinarana (Tabela 2).

117
Tabela 2 - Lista de afinidades botânicas e ecológicas para pólen e esporos deste estudo. Gimnospermas (G), Angiospérmicas (A) e Pteridófitas
(P); Afinidades ecológicas: florestas secas (fs), exógeno (ex), hidrosseral (hi), terras baixas (tb), lianas de terras baixas (li tb), manguezal
(ma), montano (mo), ripário (ri), terrestre (te) , várzea (va), campinarana (ca)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
G Podocarpidites sp.
(informal) -- Podocarpaceae Podocarpus árvore
ex, tb,
mo 29
A Alnipollenites verus 27 Betulaceae Alnus árvore ex, mo 2
A Arecipites “euterpe” 7 Arecaceae Euterpe árvore tb 7
A Arecipites? polaris 35 Arecaceae -- árvore tb 35
A Arecipites regio 45 Arecaceae -- árvore tb 44
A Bombacacidites
araracuarensis 12 Malvaceae (Bombac.) Ceiba árvore tb, va 12
A Bombacacidites
baculatus 25 Malvaceae (Bombac.) Pachira aquatica árvore tb, va 25,33
A Bombacacidites brevis 6 Malvaceae (Bombac.) -- árvore tb 6
A Bombacacidites
“caoticus” 7 Malvaceae (Bombac.) -- árvore tb 7
A Bombacacidites
ciriloensis 25 Malvaceae (Bombac.) -- árvore tb 25
(Continua)

118
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Bombacacidites
nacimientoensis 1 Malvaceae (Bombac.) Bombax árvore tb, va 1
A Bombacacidites
"L1.0.a" 7 Malvaceae (Bombac.) -- árvore tb 7
A Byttneripollis ruedae 35 Malvaceae Byttneria árvore tb 35
A Cichoreacidites?
flamulatus 22 Asteraceae -- erva te, hi 22
A Cichoreacidites
longispinosus 24 Asteraceae -- erva te, hi 23
A Clavainaperturites
microclavatus 12 Chloranthaceae Hedyosmum árvore mo, ri 12, 2
A Clavamonocolpites
lorentei 25 Arecaceae Iriartea? árvore tb 9
A Corsinipollenites
oculusnoctis 37 Onagraceae Ludwigia erva hi 37
A Crassiectoapertites
columbianus 23 Fabaceae (Papilion.) Dioclea/Canavalia liana tb, li, ri
5, 33, 15,
18

119
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Crotonoidaepollenites
“L2.68.b” 7 Euphorbiaceae Croton árvore tb 7
A Crototricolpites finitus 35 Convolvulaceae
Dicranostyles liana te, tb 7
A Ctenolophonidites
suigeneris 35 Apocynaceae Geissospermum árvore tb 35
A Cyperaceaepollis cf
neogenicus 23 Cyperaceae -- erva te, hi 23
A Cyperaceapolis
“rotundus” 7 Cyperaceae -- erva te, hi 23, 7
A Cyperaceopolis l5.18a 7 Cyperaceae -- erva te, hi 23, 7
A Dicolpopollis?
“costatus” 4 Apocynaceae Macoubea árvore tb 4
A Echiperiporites
akanthos 47 Alismataceae Sagittaria/Echinodorus erva hi 4, 13
A Echiperiporites estelae 8 Malvaceae Hampea/Hibiscus árvore tb 8, 18
A Echiperiporites
lophatus 35 Convolvulaceae? -- erva tb, li 4

120
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Echitricolporites
mcneillyi 8 Asteraceae Ambrosia erva te, hi 8, 18
A Echitricolporites
spinosus 40 Asteraceae -- erva te, hi 8
A Echitricolporites
“triangularis” 7 Asteraceae -- erva te, hi 7
A Foveotricolpites
simplex 9 Euphorbiaceae Sapium árvore tb 9
A Foveotricolporites
“gordus” 7 Bignoniaceae Fredericia liana, arbusto te, lo 7
A Grimsdalea
magnaclavata 8 Arecaceae extinct genus árvore tb, hi 8, 26
A Heterocolpites
brevicolpatus 35 Melastomataceae -- árvore tb 35
A Heterocolpites
incomptus 11 Melastomataceae Miconia? árvore tb 11
A Hetercolpites
“reticulates” 7 Melastomataceae -- árvore tb 7

121
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Heterocolpites
rotundus 11 Melastomataceae Combretum/Terminalia árvore tb, va 11,33
A Heterocolpites
verrucosus 11 Melastomataceae -- árvore tb 11
A Heterocolpites
“L2.14.d” 7 Melastomataceae -- árvore tb 7
A Ilexpollenites tropicalis 35 Aquifoliaceae Ilex árvore tb, ca 35, 36
A Inaperturopollenites
“gordus” 7 Annonaceae Anona árvore tb 7
A Jandufouria minor 14 Malvaceae Catostemma? árvore tb, va 8
A Ladakhipollenites
“baitacolumellatus” 4 Lamiaceae Vitex árvore tb, va 4
A Ladakhipollenites?
caribbiensis 25 Euphorbiaceae Sapium árvore tb, va 25
A Ladakhipollenites?
“cassioides” 4 Fabaceae (Caesalpin.) Cassia? árvore -- 4
A Ladakhipollenites?
“duplus” 7 Meliaceae Trichilia árvore tb, va 7

122
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Ladakhipollenites?
“escheweilera” 7 Lecythidaceae Eschweilera árvore te, tb 7
A Ladakhipolenites?
garzonii 11 Sapotaceae -- árvore tb 11
A Ladakhipolenites?
labiatus 11 Sapotaceae -- árvore tb 11
A Ladakhipolenites?
obesus 11 Sapotaceae -- árvore tb 11
A Ladakhipollenites?
lolongatus 3 Polygonaceae Symmeria árvore va 3
A Ladakhipollenites?
“solanum” 7 Solanaceae Solanum erva te, tb 7
A Ladakhipollenites?
“L3.2.a” 7 Fabaceae Machaerium árvore tb 7
A Lanagiopollis crassa 46 Tetrameristaceae Pelliciera árvore ma 15, 13
A Lymingtonia
“splendens” 4 Convolvulaceae Evolvulus/Jacquemontia erva tb, li 4

123
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Malvacipolloides
maristellae 25 Malvaceae aff. Abutilon erva tb, fs 25, 33, 4
A Margocolporites
carinae 22 Apocynaceae Rauvolfia árvore tb 4
A Mauritiidites franciscoi
var. franciscoi 43 Arecaceae Mauritia árvore hi 43
A Mauritiidites franciscoi
var. minutus 44 Arecaceae Mauritia árvore hi 43
A Monoporopollenites
annulatus 38 Poaceae -- erva te, hi 38
A Monoporopollenites
annuloides 22 Poaceae -- erva te, hi 4
A Multiareolites formosus 41 Acanthaceae Justicia erva tb, va 8
A Myrtaceidites “ L1.0.a” 7 Myrtaceae Eugenia árvore tb,va 7
A Myrtaceidites
“L2.68.b” 7 Myrtaceae Myrcia árvore te, tb 7

124
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Myrtaceidites
“L8.20. e” 7 Myrtaceae -- árvore tb 7
A Myrtaceidites spp.
(informal) -- Myrtaceae -- árvore tb 4
A Pachydermites
diederixi 8 Clusiaceae Symphonia globulifera árvore te, tb 8
A Paleosantalaceaepites
cingulatus 16 Euphorbiaceae Sapium paucinervium árvore te, va, tb 16
A Parsonsidites? “L1.16” 7 Caryophillaceae -- erva te, tb 7
A Parsonsidites “L2.14” 7 Chenopodiaceae -- erva te, tb 7
A Perfotricolpites
digitatus 9 Convolvulaceae Merremia erva tb, li 9, 13
A Perisyncolporites
pokornyi 8 Malpighiaceae -- -- tb 8
A Polyadopollenites
macroreticulatus 34 Hippocrateaceae Hippocratea (volubilis?) árvore tb 34
A Polyadopollenites
mariae 5 Fabaceae (Mimos.) Acacia árvore tb, va, fs 5
125
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Polyadopollenites
pseudomariae 7 Fabaceae (Mimos.) -- árvore tb 7
A Proteacidites dehaani 8 Proteaceae -- árvore tb 8
A Proteacidites
triangulatus 23 Sapindaceae/Proteaceae Allophylus? árvore tb 23, 18, 33
A Psiladiporites
“cecropia” 7 Cecropiaceae Cecropia árvore va, df, tb 7
A Proxapertites
operculatus 40 Araceae -- erva tb 40
A Proxapertites psilatus 35 Nymphaeaceae Nymphaea erva hi 4
A Proxapertites tertiaria 44 Annonaceae Crematosperma árvore tb 44, 13
A Psilabrevitricolporites
devriesii 24 Humiriaceae Humiria árvore tb, ri, ca 23
A Psilabrevitricolporites
triangularis 48 Sapindaceae? -- -- -- 23
A Psiladiporites minimus 46 Moraceae Ficus/Artocarpus/Sorocea árvore tb 46
A Psiladiporites
redundantis 9 Moraceae -- árvore tb 9

126
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Psilamonocolpites
amazonicus 11 Arecaceae Euterpe?/Geonoma?/Chelyocarpus? árvore tb, va 11, 18, 13
A Psilamonocolpites
“delicates” 7 Arecaceae -- árvore tb 7
A Psilamonocolpites
“pinguim “ 7 Arecaceae -- árvore tb 7
A Psilamonocolpites
“simplissimum” 7 Arecaceae -- árvore tb 7
A Psilastephanocolporites
fissilis 21 Polygalaceae Polygala/Monnina? erva hi 23, 33
A Psilastephanocolporites
marinamensis 12 Sapotaceae -- árvore tb 12
A Psilastephanocolporites
meliosus 4 Meliaceae? -- árvore tb 4
A Psilastephanocolporites
“ovnis” 7 Lentibulariaceae Utricularia erva tb, va 7
A Psilastephanoporites
herngreeni 11 Apocynaceae -- -- -- 18

127
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Psilastephanoporites
“redondus” 7 Apocynaceae Mesechites trifida liana tb, te, ca 7
A Psilatricolporites
silvaticus 11 Burseraceae/Sapotaceae -- árvore tb 11
A Psilatriporites
“minimum” 4 Moraceae -- árvore tb 4
A Ranunculacidites
operculatus 48 Euphorbiaceae Alchornea árvore tb, va 48
A Retimonocolpites
absyae 11 Myristicaceae Virola árvore tb, va 11
A Retimonocolpites
maximus 11 Arecaceae -- árvore tb 11
A Retimonocolpites
“pseudoabsyae” 7 Arecaceae -- árvore tb 7
A Retiperiporites
“annulatus” 7 Apocynaceae Forsteronia myriantha liana tb, li 7
A Retistephanoporites
“acalypha” 7 Euphorbiaceae Acalypha árvore tb 7
128
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Retistephanoporites
crassiannulatus 23 Malvaceae Quararibea árvore tb, va 23, 31, 33
A Retitrescolpites?
irregularis 48 Phyllanthaceae Amanoa árvore tb, va, ri 48, 33
A Retitrescolpites?
traversei 35 Acanthaceae Teliostachya árvore tb 35
A Retitricolpites lorenteae 11 Malvaceae (Bombac.) Bombax árvore tb, va 11
A Rhoipites “byrsonima” 7 Malpighiaceae Byrsonima árvore te, tb 7
A Rhoipites “circulus” 7 Dilleniaceae Doliocarpus liana, arbusto te, tb 7
A Rhoipites “diversum” 7 Acanthaceae Lepidagathis erva te, tb 7
A Rhoipites “formosus” 7 Euphorbiaceae Mabea árvore te, tb 7
A Rhoipites
“guarantenses” 7 Bignoniaceae Fredericia liana, arbusto te, tb 7
A Rhoipites guianensis 48 Malvaceae aff. Vasivaea árvore tb 33
A Rhoipites
“heteroluminados” 7 Passifloraceae Turnera erva te, tb 7
A Rhoipites? “longus” 7 Acanthaceae Aphelandra erva te, tb 7

129
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Rhoipites manausensis 3 Araliaceae Schefflera árvore tb 4
A Rhoipites “obesus” 7 Euphorbiaceae Mabea árvore te, tb 7
A Rhoipites
“perprolatus” 7 Euphorbiaceae Hyeronima árvore te, tb 7
A Rhoipites
“pseudocassioides” 7 Fabaceae Cassia árvore te, va, tb 7
A Rugutricolporites arcus 11 Chrysobalanaceae Licania/Chrysobalanus árvore tb 11, 13
A Rugutricolporites
“floratus” 7 Chrysobalanaceae Couepia árvore tb 7
A Scabratricolporites
“dalbergia” 7 Fabaceae Dalbergia liana, arbusto te, tb 7
A Spirosyncolpites
spiralis 9 Bignoniaceae? -- -- -- 31
A Stephanocolpites
evansii 25 Spermacoce -- árvore tb 25
A Striatopollis
catatumbus 10 Fabaceae (Caesalpin.) Crudia árvore tb, va 18

130
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
A Striatopollis
“crassitectatus” 4 Fabaceae (Caesalpin.) Macrolobium/Crudia árvore tb 4
A Striatricolporites
“oblates” 7 Anacardiaceae Spondias árvore te, va, tb 7
A Syncolporites
“cupania” 7 Sapindaceae Cupania árvore ca, te 7
A Syncolporites “L6.2.a” 7 Loranthaceae Struthanthus liana ca, te, va 7
A Trichotomosulcites
“amazonicus” 4 Arecaceae -- árvore tb 4
A Verrustephanoporites
“aramaca” 7 Araceae Anthurium erva te, va, tb 7
A Verrutricolporites
“L1.0.a” 7 Rubiaceae Coussarea árvore te, tb 7
A Zonocostites ramonae 8 Rhizophoraceae Rhizophora árvore ma 5,8
P Azolla sp. (informal) -- Salviniaceae Azolla esporo hi 4
P Crassoretitriletes
vanraadshooveni 8 Schizaeaceae Lygodium microphyllum esporo hi 8
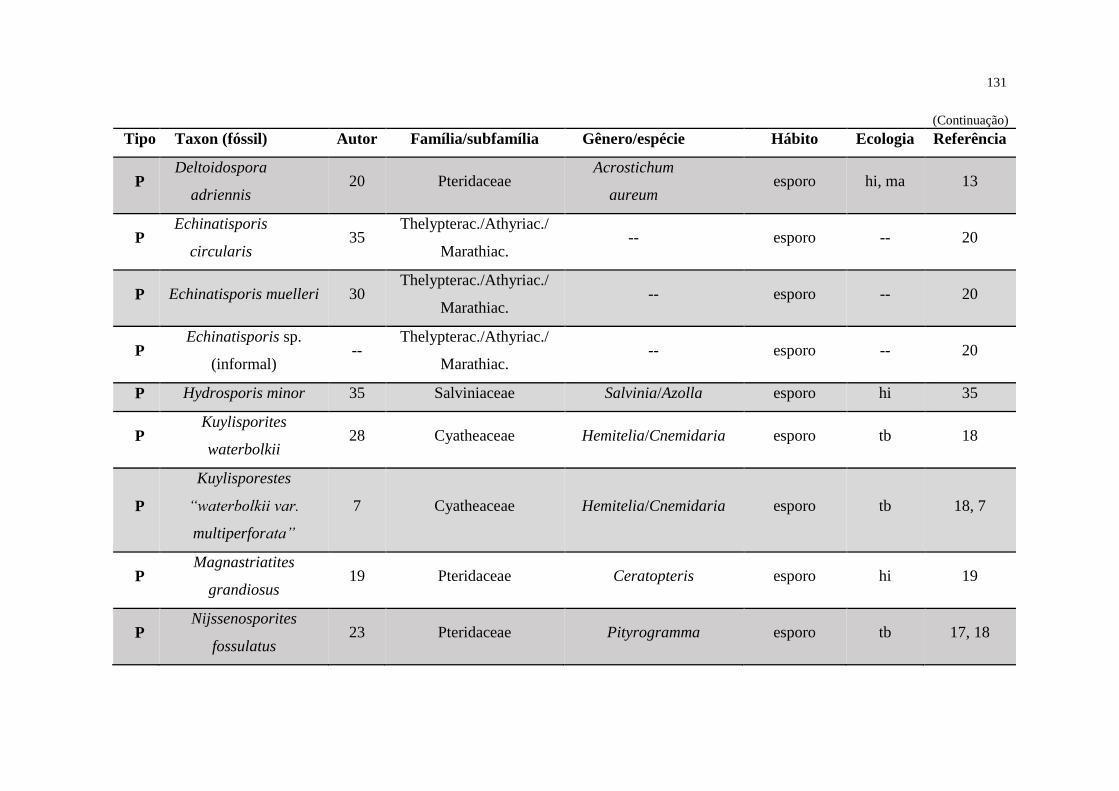
131
(Continuação)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
P Deltoidospora
adriennis 20 Pteridaceae
Acrostichum
aureum esporo hi, ma 13
P Echinatisporis
circularis 35
Thelypterac./Athyriac./
Marathiac. -- esporo -- 20
P Echinatisporis muelleri 30 Thelypterac./Athyriac./
Marathiac. -- esporo -- 20
P Echinatisporis sp.
(informal) --
Thelypterac./Athyriac./
Marathiac. -- esporo -- 20
P Hydrosporis minor 35 Salviniaceae Salvinia/Azolla esporo hi 35
P Kuylisporites
waterbolkii 28 Cyatheaceae Hemitelia/Cnemidaria esporo tb 18
P
Kuylisporestes
“waterbolkii var.
multiperforata”
7 Cyatheaceae Hemitelia/Cnemidaria esporo tb 18, 7
P Magnastriatites
grandiosus 19 Pteridaceae Ceratopteris esporo hi 19
P Nijssenosporites
fossulatus 23 Pteridaceae Pityrogramma esporo tb 17, 18
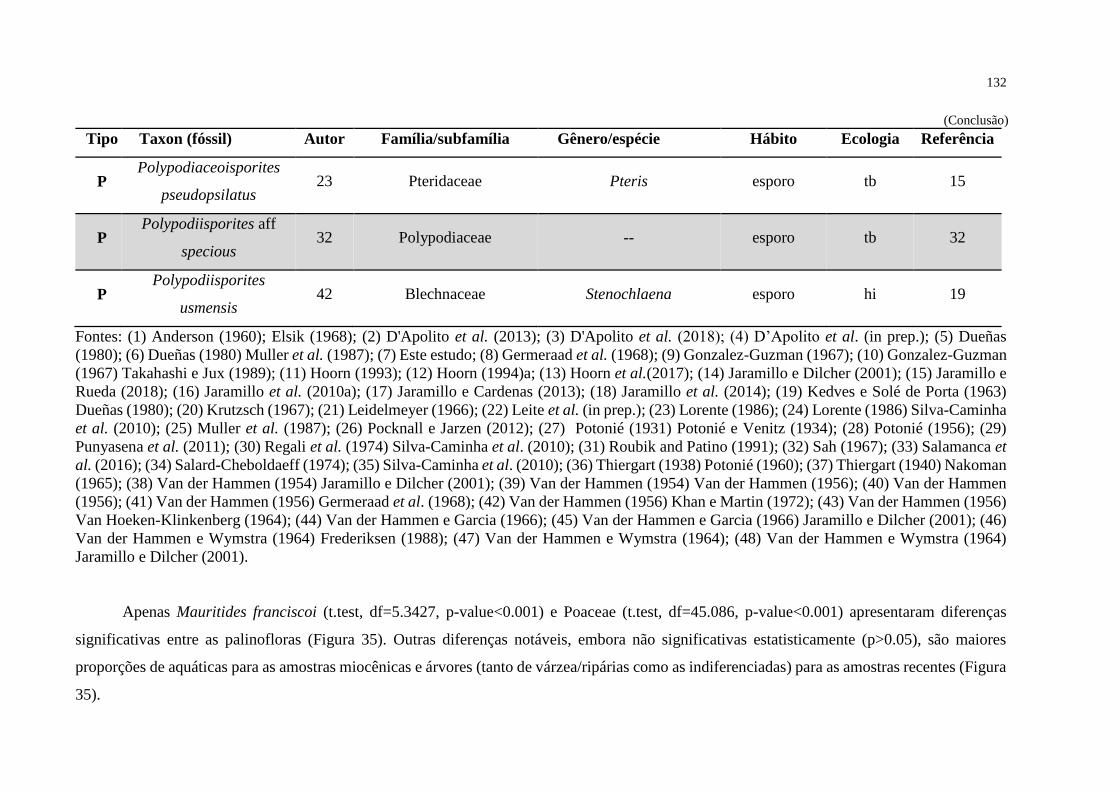
132
(Conclusão)
Tipo Taxon (fóssil) Autor Família/subfamília Gênero/espécie Hábito Ecologia Referência
P Polypodiaceoisporites
pseudopsilatus 23 Pteridaceae Pteris esporo tb 15
P Polypodiisporites aff
specious 32 Polypodiaceae -- esporo tb 32
P Polypodiisporites
usmensis 42 Blechnaceae Stenochlaena esporo hi 19
Fontes: (1) Anderson (1960); Elsik (1968); (2) D'Apolito et al. (2013); (3) D'Apolito et al. (2018); (4) D’Apolito et al. (in prep.); (5) Dueñas
(1980); (6) Dueñas (1980) Muller et al. (1987); (7) Este estudo; (8) Germeraad et al. (1968); (9) Gonzalez-Guzman (1967); (10) Gonzalez-Guzman
(1967) Takahashi e Jux (1989); (11) Hoorn (1993); (12) Hoorn (1994)a; (13) Hoorn et al.(2017); (14) Jaramillo e Dilcher (2001); (15) Jaramillo e
Rueda (2018); (16) Jaramillo et al. (2010a); (17) Jaramillo e Cardenas (2013); (18) Jaramillo et al. (2014); (19) Kedves e Solé de Porta (1963)
Dueñas (1980); (20) Krutzsch (1967); (21) Leidelmeyer (1966); (22) Leite et al. (in prep.); (23) Lorente (1986); (24) Lorente (1986) Silva-Caminha
et al. (2010); (25) Muller et al. (1987); (26) Pocknall e Jarzen (2012); (27) Potonié (1931) Potonié e Venitz (1934); (28) Potonié (1956); (29)
Punyasena et al. (2011); (30) Regali et al. (1974) Silva-Caminha et al. (2010); (31) Roubik and Patino (1991); (32) Sah (1967); (33) Salamanca et
al. (2016); (34) Salard-Cheboldaeff (1974); (35) Silva-Caminha et al. (2010); (36) Thiergart (1938) Potonié (1960); (37) Thiergart (1940) Nakoman
(1965); (38) Van der Hammen (1954) Jaramillo e Dilcher (2001); (39) Van der Hammen (1954) Van der Hammen (1956); (40) Van der Hammen
(1956); (41) Van der Hammen (1956) Germeraad et al. (1968); (42) Van der Hammen (1956) Khan e Martin (1972); (43) Van der Hammen (1956)
Van Hoeken-Klinkenberg (1964); (44) Van der Hammen e Garcia (1966); (45) Van der Hammen e Garcia (1966) Jaramillo e Dilcher (2001); (46)
Van der Hammen e Wymstra (1964) Frederiksen (1988); (47) Van der Hammen e Wymstra (1964); (48) Van der Hammen e Wymstra (1964)
Jaramillo e Dilcher (2001).
Apenas Mauritides franciscoi (t.test, df=5.3427, p-value<0.001) e Poaceae (t.test, df=45.086, p-value<0.001) apresentaram diferenças
significativas entre as palinofloras (Figura 35). Outras diferenças notáveis, embora não significativas estatisticamente (p>0.05), são maiores
proporções de aquáticas para as amostras miocênicas e árvores (tanto de várzea/ripárias como as indiferenciadas) para as amostras recentes (Figura
35).
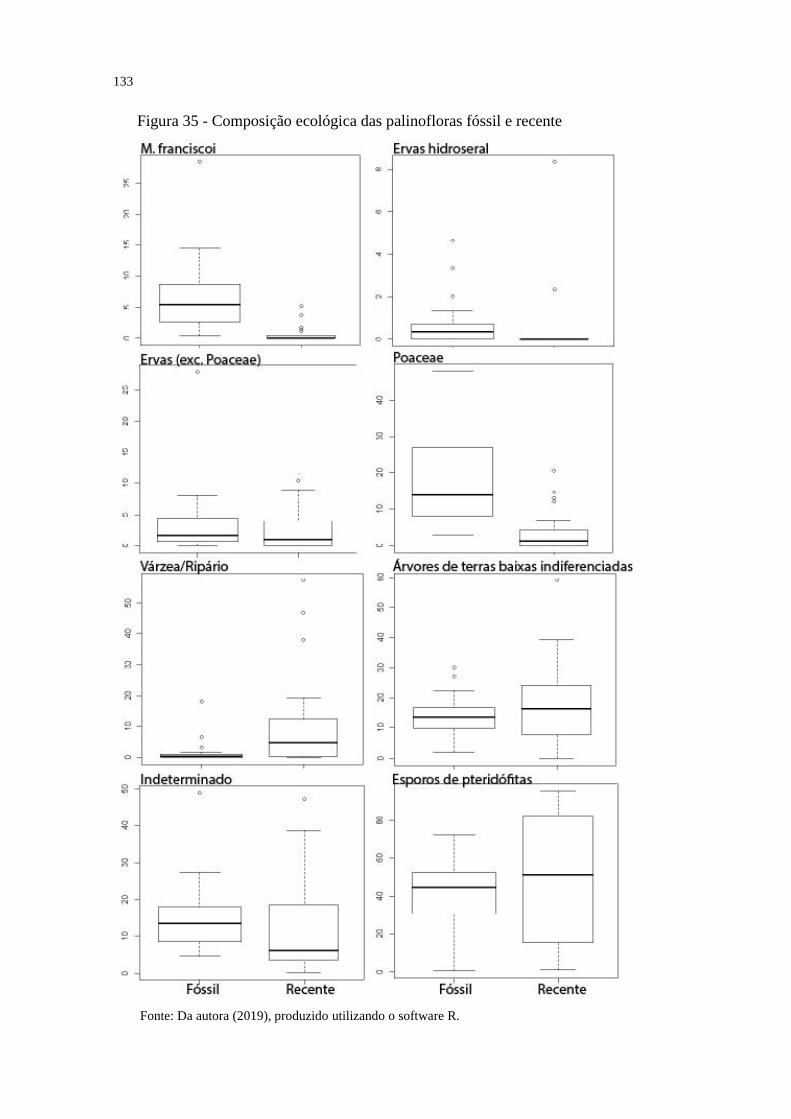
133
Figura 35 - Composição ecológica das palinofloras fóssil e recente
Fonte: Da autora (2019), produzido utilizando o software R.

134

135
5 DISCUSSÃO
A idade atribuída ao perfil 1AS-15-AM com base na correlação gráfica é mais
semelhante à dos perfis1-AS-4A-AM (Mioceno inferior a superior (Hoorn, 1993)) e 1AS-105-
AM (final do Mioceno inferior a início do Mioceno superior (Jaramillo et al., 2017)) do que a
de outros furos estudados da Formação Solimões. O perfil 1AS-15-AM é quase perfeitamente
correlato com o perfil 1-AS-105-AM com seus 255.5 m de extensão correspondendo aos
primeiros 200 m do perfil 1-AS-105-AM.
A composição ecológica da palinoflora miocênica, com o predomínio dos tipos
associados à Mauritia flexuosa L. e à Poaceae e maior presença de herbáceas e plantas aquáticas
(Figura 35), poderia estar relacionada à presença do sistema Pebas. Os grandes lagos do sistema
Pebas durante o Mioceno inferior e médio (Hoorn et al., 2010), ofereceram um ambiente
favorável para existência de floras de lagos, pântanos e ambientes associados, o que foi
confirmado pelos dados palinológicos apresentados. Já a menor presença destes táxons de
ambientes alagados e maior representatividade de táxons associados a ambientes de terras
baixas amazônicas e várzea para o grupo de amostras recentes se relaciona à extinção do sistema
Peba no Mioceno superior (Hoorn et al., 2010).
A presença do sistema de grandes lagos (Mioceno inferior-médio), sua extinção
(Mioceno superior) e estabelecimento de um sistema fluvial erosivo (Plioceno até período atual)
produziram depósitos sedimentares característicos que delimitam a distribuição florística da
Amazônia atualmente (Higgins et al., 2011). A existência do sistema Pebas atuava como uma
barreira de dispersão florística entre os Andes e as terras baixas amazônicas, impedindo a ampla
dispersão das espécies e concentrando sua distribuição em um ou outro ambiente (Antonelli et
al., 2009). O final do sistema e o reestabelecimento de conexões terrestres entre os Andes e as
terras baixas no Mioceno superior possibilitaram a dispersão das espécies na Amazônia
ocidental/central e também vários eventos de colonização (Antonelli et al., 2009), o que
explicaria a grande diferença das palinofloras com relação a sua composição.
No período Neógeno, a diversidade florística da floresta Amazônica já estava instalada
como a que se conhece hoje, como evidenciado pelos trabalhos de Hoorn et al. (1993; 1994a;
1994b), Silva-Caminha et al. (2010) e Jaramillo et al. (2010a). Esses trabalhos mostram
afinidades botânicas dos tipos polínicos fósseis associadas à 9 das 10 principais famílias de
angiospermas existentes hoje para a Amazônia (Fabaceae, Orchidaceae, Rubiaceae,

136
Melastomataceae, Poaceae, Apocynaceae, Annonaceae, Cyperaceae, Araceae, Euphorbiaceae)
(Cardoso et al., 2017; Forzza et al., 2010). No presente estudo, as afinidades encontradas
também são condizentes com a flora Amazônica atual (Tabela 2) e inclui ambientes que
ocorrem na paisagem deste bioma.O clima do Mioceno foi diferente do atual, com temperaturas
predominantemente maiores (Zachos et al. 2001; 2008) e com a presença de um ótimo climático
(Máximo térmico do Mioceno médio; MTMM, cerca de 17 a 15 Ma) (Zachos et al. 2001), em
que as temperaturas foram aproximadamente de 3 a 4 graus mais altas que as de hoje. Mesmo
assim, esta diferença climática não parece ter sido suficiente para afetar significativamente a
diversidade florística para o período quando comparada ao presente. Dick et al. (2013) mostram
a origem de espécies de árvores amplamente distribuídas na Amazônia para o período
Miocênico, demonstrando que a floresta Amazônica tolera temperaturas maiores como as que
ocorreram neste período.
Apesar das diferenças em ambientes prevalentes entre o registro fóssil e o atual, as
estimativas de diversidade similares mostram que, potencialmente, a flora miocênica não se
distinguia da atual quanto ao número de espécies. Ao comparar o número de tipos polínicos
encontrados nas palinofloras, é observada semelhança com os resultados obtidos nos trabalhos
de Hooghiemstra e Van der Hammen (1998) e Hooghiemstra e Van der Hammen (2000), com
um maior número de tipos polínicos para a palinoflora fóssil do que para a palinoflora
holocênica. Já para as medidas de diversidade, o presente trabalho confirma o observado em
Jaramillo et al. (2010a), em que é evidenciada uma falta de diferença significativa na
diversidade entre Néogeno e recente.
Um maior número de amostras para ambas as palinofloras provavelmente beneficiaria
muito as análises. Sendo importante destacar as amostras recentes, dado que estas foram
coletadas basicamente em ambiente ecológico de várzea e, assim, os ambientes amazônicos
atuais estão subrepresentados. A fim de poder representar mais fielmente a vegetação
amazônica através de chuva polínica, seria necessária uma amostragem palinológica da
interface sedimento/água (melhores amostras superficiais para a calibragem de uma fração
regional de chuva polínica) (Colinvaux et al., 1999) direcionada por vários transectos da bacia
amazônica, nos seus vários ambientes ecológicos (terra firme, várzea, igapó, campinas, etc.)
correlacionando-a com vegetação local e parâmetros climáticos.

137
6 CONCLUSÃO
Um total de 47 novas espécies foram descritas para o material fóssil, 11 novas espécies
de esporos e 36 novas espécies de pólen e é atribuída idade de final do Mioceno Inicial a início
do Mioceno Tardio (16.1 a 7.1 Ma) aos sedimentos do perfil 1-AS-15-AM. Os tipos
predominantes na palinoflora fóssil foram M. Franciscoi e Poaceae e tipos associados a várzea
e ambientes de terras baixas amazônicas indiferenciadas para a palinoflora holocênica.
A vegetação do Noroeste da Bacia Amazônica passou por grandes mudanças desde o
período Néogeno (23 Ma) até o presente, refletindo as drásticas alterações
geológicas/geográficas pelas quais foi submetida em todo esse período de tempo. Por outro
lado, sua vegetação, desde essa época, foi a de uma floresta tipicamente tropical como se
conhece atualmente.


139
REFERÊNCIAS
Absy, M.L. 1979. A palynological study of the Holocene sediments in the Amazon basin. Tese
de Doutorado, Universidade de Amsterdam, Amsterdam. 102pp.
Adegoke, O.S.; Jan Du Chéne, R.E.; Agumanu, A.E.; Ajayi, P.O. 1978. Palynology and age
of the Kerri-Kerri Formation, Nigeria. Revista Espanola de Micropaleontologa, 10(2): 267-
283.
Anderson, R.Y. 1960. Cretaceous-Tertiary palynology, eastern side of the San Juan Basin,
New Mexico. State Bureau of Mines and Mineral Resources/New Mexico Institute of Mining
and Technology, New Mexico, USA. 58pp.
Antonelli, A.; Nylander, J.A.A.; Persson, C.; Sanmartín, I. 2009. Tracing the impact of the
Andean uplift on Neotropical plant evolution. Proceedings of the National Academy of
Sciences, 106(24): 9749-9754.
B. Geoci. Petrobras, Rio de Janeiro, v. 15, n. 2, p. 217-225, maio/nov. 2007
Boltenhagen, E. 1976. Senonian microflora of Gabon. Revue de Micropaleontologie, 18(4):
191-199.
Braga, P.I.S. 1979. Subdivisão fitogeográfica, tipos de vegetação, conservação e inventário
florístico da floresta amazônica. Acta Amazonica, 9(4): 53-80.
Caputo, M.V.; Soares, E.A.A. 2016. Eustatic and tectonic change effects in the reversion of
the transcontinental Amazon River drainage system. Brazilian Journal of Geology, 46(2):
301-328.
Cardoso, D.; Särkinen, T.; Alexander, S.; Amorim, A.M.; Bittrich, V.; Celis, M. et al. 2017.
Amazon plant diversity revealed by a taxonomically verified species list. Proceedings of the
National Academy of Sciences, 114(40): 10695-10700.
Chao, A.; Hsieh, E.L.; Ma, K.H. 2018. iNEXT: iNterpolation and EXTrapolation for species
diversity. R package version 2.0.15. (http://chao.stat.nthu.edu.tw/blog/software-download/).
Acesso: 26/08/18.
Chao, A.; Jost. L. 2012. Coverage-based rarefaction and extrapolation: standardizing samples
by completeness rather than size. Ecology, 93(12): 2533-2547.
Colinvaux, P.; Oliveira, P.E.; Patiño, J.E.M. 1999. Manual e Atlas palinológico da
Amazônia. Harwood Academic Publishers, Amsterdam, NL. 322pp.
Cruz, N.M.C. 1984. Palinologia do Linhito do Solimões no Estado do Amazonas. Anais do
II Simpósio Amazônico, Manaus, BR. p. 473-480.

140
D’Apolito, C. 2016. Landscape evolution in Western Amazonia: palynostratigraphy,
palaeoenvironments and diversity of the miocence Solimões formation, Brazil. Tese de
Doutorado, School of Geography, Earth and Environmental Sciences College of Life and
Environmental Sciences/University of Birmingham, Birmingham. 350pp.
D’Apolito, C.; Absy, M.L.; Latrubesse, E.M. 2013. The Hill of Six Lakes revisited: new data
and re-evaluation of a key Pleistocene Amazon site. Quaternary Science Reviews, 76: 140-
155.
D’Apolito, C.; Jaramillo, C.; Harrington, G. in prep. Miocene palynology of the Solimões
Formation, western Amazonia.
D'Apolito, C.; Silva-Caminha, S. A. F.; Jaramillo, C.; Dino, R.; Soares, E.A.A. 2018. The
Pliocene-Pleistocene palynology of the Negro River, Brazil. Palynology, 1-21.
De Verteuil, L.; Norris, G. 1996. Miocene dinoflagellate stratigraphy and systematics of
Maryland and Virginia. Micropaleontology, 42, 1-172.
Deflandre, G.; Cookson, I.C. 1955. Fossil microplankton from Australian Late Mesozoic and
Tertiary sediments. Australian Journal of Marine and Freshwater Research, 6(2): 242-313.
Dettmann, M.E., 1973. Angiospermous pollen from Albian to Turonian sediments of
Australia. Geological Society of Australia Special Publication, 4: 3-34.
Dick, C.W.; Lewis, S.L.; Maslin, M.; Bermingham, E. 2013. Neogene origins and implied
warmth tolerance of Amazon árvore species. Ecology and Evolution, 3(1): 162-169.
Dueñas, H. 1980. Palynology of Oligocene-Miocene strata of borehole Q-E-22, Planeta Rica,
Northern Colombia. Review of Palaeobotay and Palynology, 30: 313-328.
Edwards, L.E. 1989. Supplemented graphic correlation: a powerful tool for palaeontologists
and nonpaleontologists. Palaios, 4(2): 127-143.
Eiras, J.F.; Becker, C.R.; Souza, E.M.; Gonzaga, F.G.; Silva, J.G.F.; Daniel, L.M.F. et al.
1994. Bacia do Solimões. Boletim de Geociências da Petrobrás, 8: 17-45.
Elsik, W.C. 1968. Palynology of a Paleocene Rockdale Lignite, Milam County, Texas. II.
Morphology and taxonomy. Pollen and Spores, 10(2): 263-314.
Erdtman, G. 1952. Pollen morphology and plant taxonomy - angiosperms: an introduction to
palynology. v. 1. Almqvist & Wiksell, Stockholm, SWE. 539pp.
Faegri, K.; Iversen, J. 1966. Textbook of modern pollen analysis. 2. ed. Ejner Munksgaard,
Copenhagen, DK. 108pp.
Frederiksen, N.O. 1983. Middle Eocene palynomorphs from San Diego, California. Part II.
Angiosperm pollen and miscellanea. American Association of Stratigraphic Palynologists, 12:
32-155.

141
Frederiksen, N.O. 1988. Sporomorph biostratigraphy, floral changes, and paleoclimatology,
Eocene and earliest Oligocene of the Eastern Gulf Coast. U.S. Geological Survey
professional paper 1448, Denver, USA. 68pp.
Forzza, R.C. et al. (Org.). 2010. Catálogo de plantas e fungos do Brasil. Andrea Jakobsson
Estúdio; Instituto de Pesquisa Jardim Botânico do Rio de Janeiro, Rio de Janeiro, BR. 875pp.
Germeraad, J.H.; Hopping, C.A.; Muller, J. 1968. Palynology of tertiary sediments from
tropical areas. Review of Palaeobotany and Palynology, 6(3-4): 189-348.
Gonzalez-Guzman, A.E. 1967. A palynological study on the Upper Los Cuervos and Mirador
Formations (Lower and Middle Eocene; Tibu area, Colombia). E.J. Brill, Leiden, NL. 68pp.
Haffer, J. 1969. Speciation in Amazonian forest birds. Science, 165(3889): 131-137.
Herngreen, G.F.W. 1975. An Upper Senonian pollen assemblage of Borehole 3-PIA-10-AL,
State of Alagoas, Brazil. Pollen et Spores, 17(1): 93-140.
Higgins, M.A.; Ruokolainen, K.; Tuomisto, H.; Llerena, N.; Cardenas, G.; Phillips, O.L. et al.
2011. Geological control of floristic composition in Amazonian forests. Journal of
Biogeography, 38(11): 2136-2149.
Hooghiemstra, H.; Van Der Hammen, T. 1998. Neogene and Quaternary development of the
neotropical rain forest: the forest refugia hypothesis, and a literature overview. Earth Sciences
Reviews, 44(3-4): 147-183.
Hooghiemstra, H.; Van Der Hammen, T. 2000. Neogene and Quaternary history of
vegetation, climate, and plant diversity in Amazonia. Quaternary Science Review, 19(8): 725-
742.
Hoorn, C. 1993. Marine incursions and the influence of Andean tectonics on the Miocene
depositional history of northwestern Amazonia: Results of a palynostratigraphic study.
Palaeogeography, Palaeoclimatology, Palaeoecology, 105(3-4): 267-309.
Hoorn, C. 1994a. An environmental reconstruction of the palaeo-Amazon River system
(Middle to Late Miocene, NW Amazonia). Palaeogeography, Palaeoclimatology,
Palaeoecology, 112(3-4): 187-238.
Hoorn, C. 1994b. Fluvial palaeoenvironments in the intracratonic Amazonas Basin (Early
Miocene-early Middle Miocene, Colombia). Palaeogeography, Palaeoclimatology,
Palaeoecology, 109(1): 1-54.
Hoorn, C.; Bogotá-A, R.G.; Romero-Baez, M.; Lammertsma, E.I.; Flantua, S.; Dantas, E.L. et
al. 2017. The Amazon at sea: onset and stages of the Amazon River from a marine record in
the Foz do Amazonas Basin (Brazilian Equatorial Margin), with special reference to
vegetation turnover in the Plio-Pleistocene. Global and Planetary Change, 153: 15-65.
Hoorn, C.; Wesselingh, F.P.; Ter Steege, H.; Bermudez, M.A.; Mora, A.; Sanmartín, J.S.I.; et
al. 2010. Amazonia through time: Andean uplift, climate change, landscape evolution and
biodiversity. Science, 330(6006): 927-931.

142
Ibge. 1977. Folha SB-19 Juruá. Instituto Brasileiro de Geografia e Estatística, Rio de Janeiro,
BR. 421pp.
Ibrahim, A.C. 1933. Sporenformen des Aegir-horizonts des Ruhr-Reviers. Dissertação de
Mestrado, University of Berlin, Konrad Triltsch, Wurzburg, DE. 47pp.
Jan du Chene, R.E. 1977. Some new pollen species of the Upper Maastrichtian, Tar Sand,
Abeokuta Formation, Southern Nigeria. Revista Espanola de Micropaleontologia, 9(2): 191-
201.
Jan du Chene, R.E., Klasz, I., Archibong, E.E. 1978. Biostratigraphic study of the Borehole
Ojo-1, SW Nigeria, with special emphasis on the Cretaceous microflora. Revue de
Micropaleontology, 21(3): 123-139.
Jaramillo, C.; Bayona, G.; Pardo-Trujillo, A.; Rueda, M.; Torres, V.; Harrington, G. et al.
2007. The palynology of the Cerrejón Formation (upper Paleocene) of northern Colombia.
Palynology, 31: 153-189.
Jaramillo, C.A.; Cardenas, A. 2013. Global warming and neotropical rainforests: a historical
perspective. Annual Review of Earth and Planetary Sciences, 41: 741-766.
Jaramillo, C.A.; Dilcher, D.L. 2001. Middle Paleogene palynology of central Colombia,
South America: a study of pollen and spores from tropical latitudes. Palaeontographica
Abteilung B, 258: 87-213.
Jaramillo, C.A.; Hoorn, C.; Silva, S.A.F.; Leite, F.; Herrera, F.; Quiroz, L. et al. 2010a. The
origin of the modern Amazon rainforest: implications of the palynolgical and palaeobotanical
record. In: Hoorn, C.; Wesselingh, F.P. (Eds.). Amazonia: landscape and species evolution –
a look into the past. Blackwell Publishing, Oxford. p. 317-334.
Jaramillo, C.A.; Moreno, E.; Ramírez, V.; Silva, S.A.F.; Barrera, A.; Barrera, A.; et al. 2014.
Palynological Record of the last 20 million years in Panama. In: Stevens, W.D.; Montiel,
O.M.; Raven, P. Paleobotany and biogeography: a festschrift for Alan Graham in his 80th
year. Missouri Botanical Garden Press, St. Louis. p. 134-253.
Jaramillo, C.A.; Romero, I.; D’Apolito, C.; Bayona, G.; Duarte, E.; Louwye, S.; et al.
2017. Miocene flooding events of western Amazonia. Science Advances, 3(5): e1601693.
Jaramillo, C.A.; Rueda, J.M. 2018. A Morphological Electronic Database of Cretaceous-
Tertiary and Extant pollen and spores from Northern South America. v.
2018. (http://biogeodb.stri.si.edu/jaramillo/palynomorph/). Acesso: 12/09/18.
Jaramillo, C.A.; Rueda, J.M.; Mora, G. 2006. Cenozoic plant diversity in the neotropics.
Science, 311(5769): 1893-1896.
Jaramillo, C.A.; Rueda, M.; Vladimir, T. 2011. A palynological zonation for the Cenozoic of
the Llanos and Llanos Foothills of Colombia. Palynology, 35(1): 46-84.

143
Linhares, A.P.; Gaia, V.S.; Ramos, M.I.F. 2017. The significance of marine microfossils for
paleoenvironmental reconstruction of the Solimões Formation (Miocene), western Amazonia,
Brazil. Journal of South American Earth Sciences, 79: 57-66.
Kachinasz, K.R.; Silva-Caminha, S.A.F. 2016. Palinoestratigrafia da Formação Solimões:
comparação entre bioestratigrafia tradicional e o método de associações unitárias. Revista
Brasileira de Paleontologia, 19(3): 481-490.
Kedves, M. 1995. Upper Cretaceous spores from Egypt. Szeged, HUN. 87pp.
Kedves, M.; Solé de Porta, N. 1963. Comparacion de las esporas del género
Cicatricosisporites R. Pot y Gell, 1933 de Hungria y Colombia. Algunos problemas referentes
a su significado estratigráfico. Boletin Geologico, 12: 51-76.
Khan, A.M.; Martin, A.R.H. 1972. A note on genus Polypodiisporites R. Potonié. Pollen et
Spores, 13: 475-480.
Klaus, W. 1960. Sporen der Karnischen Stufe der ostalpinen Trias. Jahrbuch der
Geologischen Bundesanstalt, 5: 107-184.
Krutzsch, W. 1959. Mikropaläontologische (sporenpaläontologische) Untersuchungen in der
Braunkohle des Geiseltales-[Part] I, Die Sporen und die Sporenartigen sowie ehemals im
Geiseltal zu Sporites gestellten Formeinheiten der Sporae dispersae der mitteleozänen
Braunkohle des mittleren Geisel tales (Tagebau Neumark-West i.w.S.) unter
Berücksichtigung und Revision weiterer Sporenformen aus der bisherigen Literatur. Geologie,
8: 21-22.
Krutzsch, W. 1967. Atlas der mittel- und jungtertiaren dispersen Sporen- und Pollen-sowie
der Mikroplanktonformen des nordlichen Mitteleuropas. Lieferung 4 und 5: Weitere
azonotrilete (apiculate, murornate), monolete und alete Sporenformen. VEB Gustav Fischer
Verlag Jena, Berlin, DE. 232pp.
Krutzsch, W. 1970. Atlas of the Middle and Upper Tertiary dispersed spores and pollen as
well as the mikroplankton forms of northern central Europe. VII: Monoporate, monocolpate,
longicolpate dicolpate an dephedroide (polyplicate) pollen forms. Atlas der mittel- und
jungterti¤ren dispersen Sporen- und pollen - Sowie der Mikroplanktonformen des nжrdlichen
Mitteleuropas. Lieferung VII: monoporate, monocolpate, longicolpate, dicolpate und
ephedrioide (polyplicate) Pollenformen. VEB Gustav Fischer Verlag Jena, Berlin, DE. 175pp.
Kummel, B.; Raup, D. 1965. Handbook of paleontological techniques. Freeman, San
Francisco, USA. 852pp.
Latrubesse, E.M.; Cozzuol, M.; Silva-Caminha, S.A.F.; Rigsby, C.A.; Absy, M.L.; Jaramillo,
C. 2010. The late Miocene paleogeography of the Amazon basin and the evolution of the
Amazon River system. Earth-Science Reviews, 99(3-4): 99-124.
Legoux, O. 1978. Some characteristic species of pollen from the Neogene of Nigeria.
Quelques especes de pollen Caracteristiques du Neogene du Nigeria. Bulletin des Centres de
Recherches Exploration-Production Elf-Aquitaine, 2(2): 265-317.

144
Leidelmeyer, P. 1966. The Paleogene and Lower Eocene pollen flora of Guyana. Leidse
geologische mededelingen, 38(1): 49-57.
Leite, F.P.R. et al. in prep. New taxa for the palinostratigrafic framework of the Amazon
eogene, Brazil.
Leite, F.P.R.; Paz, J.; Demerval, A.D.; Silva-Caminha, S. A. 2017. The effects of the
inception of Amazonian transcontinental drainage during the Neogene on the landscape and
vegetation of the Solimões Basin, Brazil. Palynology, 41(3): 412-422.
Lorente, F.L.; Junior, A.A.B.; Oliveira, P.E.; Pessenda, L.C.R. 2017. Atlas palinologico:
laboratório14C-Cena/USP. FEALQ, Piracicaba, BR. 333pp.
Lorente, M.A. 1986. Palynology and palynofacies of the upper tertiary in Venezuela.
Dissertationes Botanicae, Stuttgart, DE. 222pp.
Maia, M.A.M.; Marmos, J. L. (Org.). 2010. Geodiversidade do estado do Amazonas. CPRM,
Manaus, BR. 275pp.
Maia, R.G.; Godoy, H.K.; Yamaguti, H.S.; Moura, P.A.; Costa, F.S.; Holanda, M.A. et al.
1977. Projeto de carvão no Alto Solimões. DNPM/CPRM, Manaus, BR. 137pp.
Mathur, Y.K. et al. 1980. Palynology and age of the Dras Volcanics near Shergol, Ladakh,
Jammu and Kashmir, India. Geoscience Journal, 1(1): 55-74.
Mchedlishvili, N.D. 1961. Liliaceae. In: Samoilovich, S.R.; Mtchedlishvili, N.D. (Eds). Pyltsa
i spory zapadnoi Sibiri, yura paleotsen. Trudy Vsesoyuznogo Neftyanogo Nauchno-
Issledovatel'skogo Geologorazvedochnogo Instituta, Leningrad, RUS.
Muller, J. 1968. Palynology of the Pedawan and Plateau Sandstone formations (Cretaceous-
Eocene) in Sarawak, Malaysia. Micropaleontology, 14(1): 1-37.
Muller, J.; Giacomo, E.; Van Erve, A.W. 1987. A palynological zonation for the Cretaceous,
Tertiary, and Quaternary of northern South America. American Association of Stratigraphic
Palynologists, Nottingham, UK. 76pp.
Nakoman, E. 1965. Description d'un nouveau genre de forme: Corsinipollenites. Annales de
la Societé Géologique du Nord, 85: 155-158.
Nichols, D.J.; Ames, H.T.; Traverse, A. 1973. On Arecipites Wodehouse,
Monocolpopollenite Thomson & Pflug, and the species “Monocolpopollenites tranquillus”.
Taxon, 22: 241-56.
Norvick, M.S.; Burger, D. 1975. Palynology of the Cenomanian of Bathurst Island, Northern
Territory, Australia. Bureau of Mineral Resources Australia, 151: 1-169.
Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D. et al. 2017.
Vegan: Community Ecology Package. (https://CRAN.R-
project.org/package=vegan). Acesso: 21/12/17.

145
Ortiz, J., Moreno, C., Cardenas, A., and Jaramillo, C. 2015. SDAR 1.0 a New Quantitative
Toolkit for Analyze Stratigraphic Data. Geophysical Research Abstracts 17:2015-2790.
Passos,M.S.;Soares,E.A.A.2017. Análise multitemporal do sistema fluvial Solimões-
Amazonas entre os tributários Purus e Negro, Amazônia Ocidental, Brasil. Geol. USP, Sér.
cient, 17(1):61:74.
Pierce, R.L. 1961. Lower Upper Cretaceous plant microfossils from Minnesota. Minnesota
Geological Survey Bulletin, 42: 1-86.
Pocknall, D.T.; Jarzen, D.M. 2012. Grimsdalea magnaclavata Germeraad, Hopping & Muller:
an enigmatic pollen type from the Neogene of northern South
America. Palynology, 36(1): 134-143.
Potonié, R. 1931. Zur Mikroskopie der Braunkohlen. (4) Tertiare Sporen- und
Blutenstaubformen (Vierte Mitteilung). Braunkohle, 30: 554-556.
Potonié, R. 1956. Synopsis der Gattungen der Sporae dispersae. I. Teil: Sporites. Beihefte zum
Geologischen Jahrbuch, 23: 103.
Potonié, R. 1958. Synopsis der Gattungen der Sporae dispersae. II. Teil: Sporites (Nachtage),
Saccites, Aletes, Praecolpates, Polyplicates, Monocolpates. Beihefte zum Geologischen
Jahrbuch, 31: 114.
Potonié, R. 1960. Synopsis der Gattungen der Sporae dispersae. III. Teil: Nachtrage Sporites,
Fortsetzung Pollenites. Mit Generalregister zu Teil I-III. Beihefte zum Geologischen
Jahrbuch, 39: 189.
Potonié, R.; Gelletich, J. 1933. Uber Pteridophyten-sporen einer eozanen Braunkohle aus
Dorog in Ungarn. Sitzungsberichte der Gesellschaft naturforschender Freundes zu Berlin, 33:
517-526.
Potonié, R.; Venitz, A. 1934. Zur Mikrobotanik des miozanen Humodils der
niederrheinischen Bucht. Arbeiten aus dem Institut fur Palaobotanik und Petrographie der
Brennsteine, 5: 5-54.
Punt, W., Hoen, P. P., Blackmore, S., Nilsson, S., Le-Thomas, A. 2007. Glossary of pollen
and spore terminology. Review of Paleobotany and Palynology, 143: 1-81.
Punyasena, S.W.; Dalling, J.W.; Jaramillo, C.; Turner, B.L. 2011. Comment on “The response
of vegetation on the Andean flank in Western Amazonia to Pleistocene climate change”.
Science, 333(6051): 1825a-b.
R Development Core Team. 2018. R: A language and environment for statistical computing.
R Foundation for Statical Computing, Vienna, Austria. (https://www.r-project.org/). Acesso:
20/06/17.
Raine, J.I. 1981. Palynological correlation of the Dunollie/Rewanui Member boundary in
drillholes 621 and 622, Greymouth Coalfield. New Zealand Geological Survey Report, 47: 11.

146
Regali, M.S.; Uesegui, N.; Santos, A. 1974. Palinologia dos sedimentos Meso–Cenozoicos do
Brasil (I). Boletim Técnico da Petrobrás, 17: 263-362.
Ricklefs, R.E.; Renner, S.S. 2012. Global Correlations in Tropical Arbore Species Richness
and Abundance Reject Neutrality. Science, 335(6067): 464-467.
Roberts, D.W. 2016. Ordination and Multivariate Analysis for Ecology. R package version
1.8-0. (https://cran.r-project.org/web/packages/labdsv/index.html). Acesso: 26/08/18.
Roubik, D.W.; Patino, J.E.M. 1991. Pollen and Spores of Barro Colorado Island. Missouri
Botanical Garden, St. Louis, USA. 268pp.
Sah, S.C.D. 1967. Palynology of an upper Neogene profile from Rusizi Valley (Burundi).
Annales du Musée Royal de l'Afrique Centrale, 57: 173.
Sah, S.C.D.; Dutta, S.K. 1966. Palynostratigraphy of the sedimentary formations of Assam-I.
Stratigraphical position of the Cherra Formation. Palaeobotanist, 15(1-2): 72-86.
Salamanca, S.; Van Soelen, E. E.; Van Manen, M. T; Flantua, S.; Santos, R.; Roddaz, M. et
al. 2016. Amazon forest dynamics under changing abiotic conditions in the early Miocene.
(Colombian Amazonia). Journal of Biogeography, 43(12): 2424-2437.
Salard-Cheboldaeff, M. 1974. Pollens tertiaries du Cameroun rapportés a la famille des
Hippocratéacées. Pollen et Spores, 16: 499-506.
Sarmiento, G. 1992. Palinología de la Formación Guaduas - estratigrafía y sistemática.
Boletín Geológico, 32: 45-126.
Shaw, A.B. 1964. Time in stratigraphy. McGraw- Hill, New York, USA. 365pp.
Silva-Caminha, S.A.F.; Jaramillo, C.; Absy, M.L. 2010. Neogene palynology of the Solimões
Basin, Brazilian Amazonia. Palaeontographica Abteilung B, 283(1): 1-67.
Silveira, R.R.; Souza, P.A. 2015. Palinologia (grãos de pólen de angiospermas) das formações
Solimões e Içá (bacia do Solimões), nas regiões de Coari e Alto Solimões, Amazonas. Revista
Brasileira de Paleontologia, 18(3): 455-474.
Silveira, R.R.; Souza, P.A. 2016. Palinologia (esporos de fungos e pteridófitas, grãos de
pólens de gimnospermas, cistos de algas e escolecodonte) das formações Solimões e Içá
(Neogeno e Pleistoceno, Bacia do Solimões), Amazonas, Brasil. Pesquisas em Geociências,
43(1): 17-39.
Silveira, R.R.; Souza, P.A. 2017. Palinoestratigrafia da Formação Solimões na Região do Alto
Solimões (Atalaia do Norte e Tabatinga), Amazonas, Brasil. Geociências, 36(1): 100-117.
Sivaguru, M.; Urban, M.A.; Fried, G.; Wesseln, C.J.; Mander, L.; Punyasena, S.W. 2016.
Comparative performance of airyscan and structured illumination superresolution microscopy
in the study of the surface texture and 3D shape of pollen. Microscopy Research and
Technique, 81(2): 1-14.

147
Skarby, A. 1964. Revision of Gleicheniidites senonicus Ross. Acta Universitatis
Stockholmiensis, 11(3): 59-77.
Srivastava, S.K. 1969. Assorted angiosperm pollen from the Edmonton Formation
(Maestrichtian), Alberta, Canada. Canadian Journal of Botany, 47(6): 975-989.
Takahashi, K.; Jux, U. 1989. Palynology of Middle Tertiary lacustrine deposits from the Jos
Plateau, Nigeria. Bulletin of the Faculty of Liberal Arts, Nagasaki University
(NaturalScience), 29(2): 181-367.
Thiergart, F. 1938. Die Pollenflora der Niederlausitzer Braunkohle, besonders im Profil der
Grube Marga bei Senftenberg. Jahrbuch der Preussische geologischen Landesanstalt und
Bergakademie zu Berlin, 58: 282-356.
Thiergart, F. 1940. Die Mikropaläontologie als Pollenanalyse in Dienst der
Braunkohlenforschung. Brennstoff-Geologie, 13: 1-82.
Thomas, W.W. 1999. Conservation and monographic research on the flora of tropical
America. Biodiversity and Conservation, 8(8): 1007-1015.
Thomson, P.W.; Pflug, H.; Hans, D. 1953. Pollen und Sporen des mitteleuropäischen tertiärs.
Palaeontographica Abteilung B, 94: 1-138.
Traverse, A. 1955. Pollen analysis of the Brandon Lignite of Vermont. U.S. Dept. of the
Interior, Bureau of Mines, Washington, USA. 107pp.
Traverse, A. 2007. Paleopalynology. 2nd ed. Dordrecht: Springer, pp. 813.
Tucker, Maurice. 2003. Sedimentary Rocks in the Field. 3nd ed. John Wiley & Sons Ltd,
234pp.
Van der Hammen, T. 1954. The development of Colombian flora throughout geologic
periods: I, Maestrichtian to Lower Tertiary. Boletín Geológico, 2: 49-106.
Van der Hammen, T. 1956a. A palynological systematic nomenclature. Boletín Geológico, 4:
63-101.
Van der Hammen, T., 1956b. Description of some genera and species of fossil pollen and
spores. Boletín Geológico IV(2-3): 111-117.
Van der Hammen, T.; Garcia, C. 1965. The Paleocene pollen flora of Colombia. Leidse
Geologische Mededelingen, 35(1): 105-114.
Van der Hammen, T.; Wymstra, T. A. 1964. A palynological study on the Tertiary and Upper
Cretaceous of British Guayana. Leidse Geologische Mededelingen, 30(1): 183-241.
Van Hoeken-Klinkenberg, P.M.J. 1964. A palynological investigation of some Upper
Cretaceous sediments in Nigeria. Pollen et Spores, 6(1): 209-231.

148
Van Hoeken-Klinkenberg, P.M.J. 1966. Maastrichtian Paleocene and Eocene pollen spores
from Nigeria. Leidse Geologische Mededelingen, 38(1): 37-44.
Victoria, R.L; Martinelli, L.A.; Cunha, H.B.; Richey, J.E. et al. 2010. The Amazon basin and
its natural cycles. In: Salati, E.; Absy, M.L.; Victoria, R.L. (Eds). Amazonia, um ecossistema
em transformação. INPA, Manaus. p. 163-214.
Wall, D. 1967. Fossil microplankton in deep-sea cores from the Caribbean Sea.
Palaeontology, 10(1): 95-123.
Wesselingh, F.P. 2006. Molluscs from the Miocene Pebas Formation of Peruvian and
Colombian Amazonia. Scripta Geologica, 133: 19-290.
Wesselingh, F.P.; Räsänen, M.E.; Irion, G.; Vonhof, H.B.; Kaandorp, R.; Renema, W. et al.
2002. Lake Pebas: a palaeoecological reconstruction of a Miocene, long-lived lake complex in
western Amazonia. Cainozoic Research, 1: 35-81.
Weyland, H.; Krieger, W. 1953. Die Sporen und Pollen der Aachener Kreide und ihre
Bedeutung fur die Charakterisierung des Mittleren Senons. Palaeontographica Abteilung B,
95: 2-29.
Wijmstra, T.A. 1971. The palynology of the Guiana Coastal Basin. PhD Thesis, University of
Amsterdam, Amsterdam. 62pp.
Wing, S.L.; Herrera, F.; Jaramillo, C.; Gómez-Navarro, C.; Wilf, P.; Labandeira, C.C. 2009.
Late Paleocene fossils from the Cerrejón Formation, Colombia, are the earliest record of
Neotropicalrainforest. PNAS, 106(44): 18627-18632.
Wodehouse, R.P. 1933. Tertiary pollen II. The oil shales of the Eocene Green River
Formation. Bulletin of the Torrey Botanical Club, 60: 479-524.
Wood, G.D.; Gabriel, A.M.; Lawson, J.C. 1996. Palynological techniques e processing and
microscopy. In: Jansonius, J.; McGregor, D.C. (Ed.). Palynology, principles and applications.
v. 1. American of Stratigraphic Palynologists Foundation, Dallas. p. 29-50.
Wolfe, H. 1934. Mikrofossilien des pliozanen Humodils der Grube Freigericht bei Dettingen
a. M. und Vergleich mit alteren Schichten des Tertiars sowie post-Tertiaren Ablagerungen.
Arbeiten aus dem Institut fur Palaobotanik und Petrographie der Brennsteine, 5: 55-86.
Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. 2001. Trends, rhythms, and
aberrations in global climate 65 Ma to present. Science, 292(5517): 686-693.
Zachos, J.C.; Dickens, G.R.; Zeebe, R.E. 2008. An early Cenozoic perspective on greenhouse
warming and carbon-cycle dynamics. Nature, 451(7176): 279-283.

149
APÊNDICES

150

151























![Apostila Dança Moderna [Captulo IV - A Dança Moderna] (1)](https://img.pdfslide.tips/doc/110x75/55cf9191550346f57b8e82fb/apostila-danca-moderna-captulo-iv-a-danca-moderna-1.jpg)





