Embed Size (px)
Citation preview

Fausto Caruana
Dip. di Neuroscienze,
Sezione di Fisiologia
Università di Parma
Azioni, intenzioni ed emozioni: Le basi neurali della comprensione
del comportamento degli altri
Pomaia, 13.06.2009

• “In Principio era l’Azione”: cosa sappiamo sul cervello motorio
• Comprendere le Azioni:- i neuroni mirror e le scoperte sulla scimmia- il sistema mirror nell’uomo
• Alla ricerca di altri specchi: le emozioni e le sensazioni

“… in principio era l’azione”
Il Sistema Motorio e lo Scopo delle Azioni
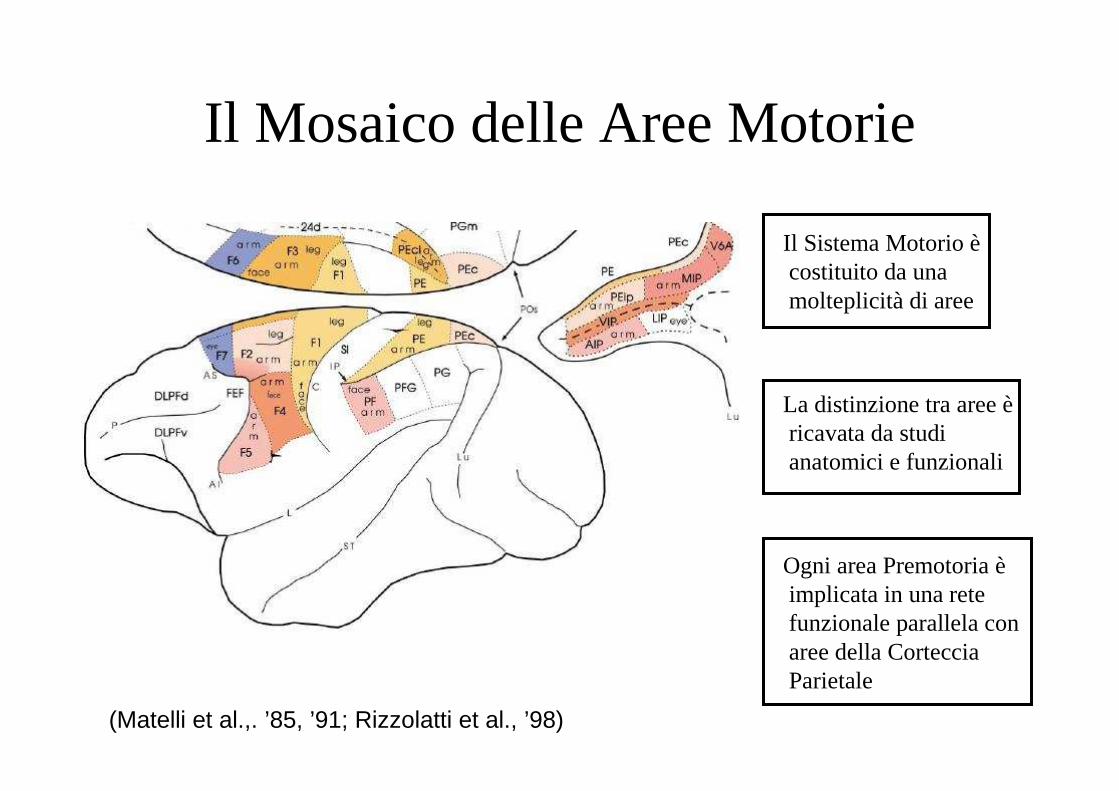
Il Mosaico delle Aree Motorie
Il Sistema Motorio ècostituito da una molteplicità di aree
La distinzione tra aree èricavata da studi anatomici e funzionali
Ogni area Premotoria èimplicata in una rete funzionale parallela con aree della Corteccia Parietale
(Matelli et al.,. ’85, ’91; Rizzolatti et al., ’98)

L’idea classica è che il Sistema Motorio ha un controllo esecutivo;
Secondo tale idea, in particolare, il Sistema Motorio codifica e controlla movimenti

Suddivisione funzionale classica
PERCEZIONE
COGNIZIONE
AZIONE
AREE SENSORIALI
AREE ASSOCIATIVE
AREE MOTORIE
La suddivisione funzionale classica del cervello e’stata fortemente influenzata dall’idea ingenua di ciò che pensiamo faccia “la mente”
[ 1 ] attività percettiva e immagazzinamento di informazioni --- Aree sensoriali
[ 2 ] elaborazione cognitiva --- Aree associative
[ 3 ] uscita motoria --- Aree motorie
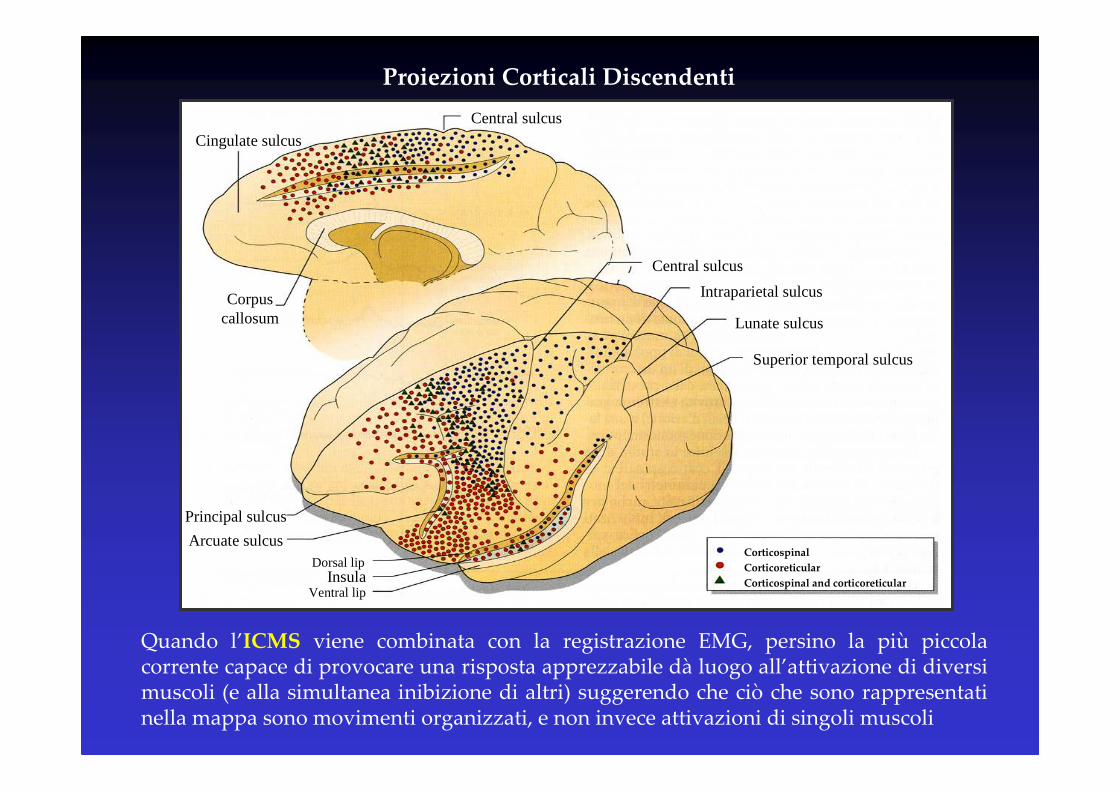
Cingulate sulcus
Central sulcus
Corpus callosum
Central sulcus
Intraparietal sulcus
Lunate sulcus
Superior temporal sulcus
Principal sulcus
Arcuate sulcusDorsal lip
Ventral lipInsula
Corticospinal
Corticoreticular
Corticospinal and corticoreticular
Proiezioni Corticali Discendenti
Quando l’ICMS viene combinata con la registrazione EMG, persino la più piccola corrente capace di provocare una risposta apprezzabile dà luogo all’attivazione di diversi muscoli (e alla simultanea inibizione di altri) suggerendo che ciò che sono rappresentati nella mappa sono movimenti organizzati, e non invece attivazioni di singoli muscoli
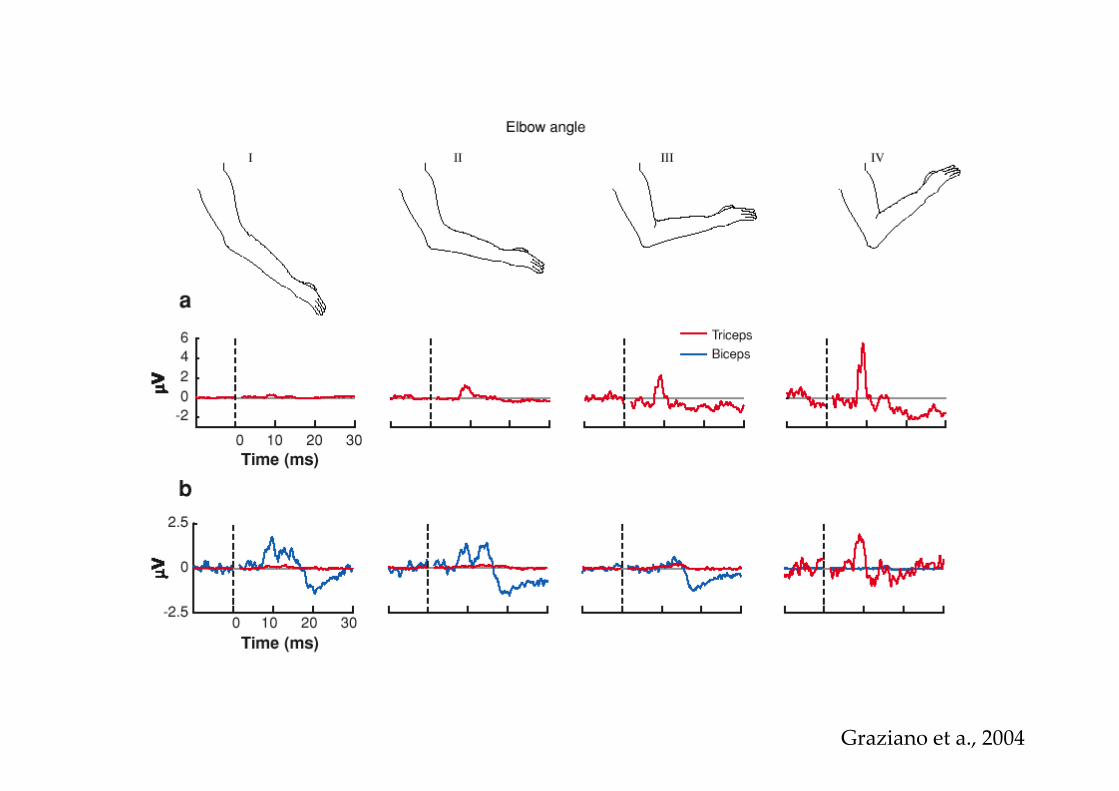
Graziano et a., 2004

QUALE RUOLO PER LA CORTECCIA MOTORIA?

A Afferramento con la bocca
B Afferramento con la mano destra
C Afferramento con la mano destra
Rizzolatti et al., 1988: Nella corteccia premotoria ci sono neuroni che siattivano ogni volta che la scimmia afferra un oggetto, indipendentemente dal tipo di effettore usato: la mano destra, la manosinistra, la bocca.
Hp: In F5 esistono neuroni che codificano lo scopo dell’azione –afferrare, tirare, strappare, etc. – e non il movimento necessario per raggiungerlo.
(Rizzolatti et al. 1988)

Cosa succede se l’azione viene dissociatadal movimento richiesto per compierla?
Studio delle proprieta’ dei neuroni motori durante atti motori dotati dello stesso scopo finale (afferrare il cibo) ma ottenuti mediante movimenti opposti della mano.
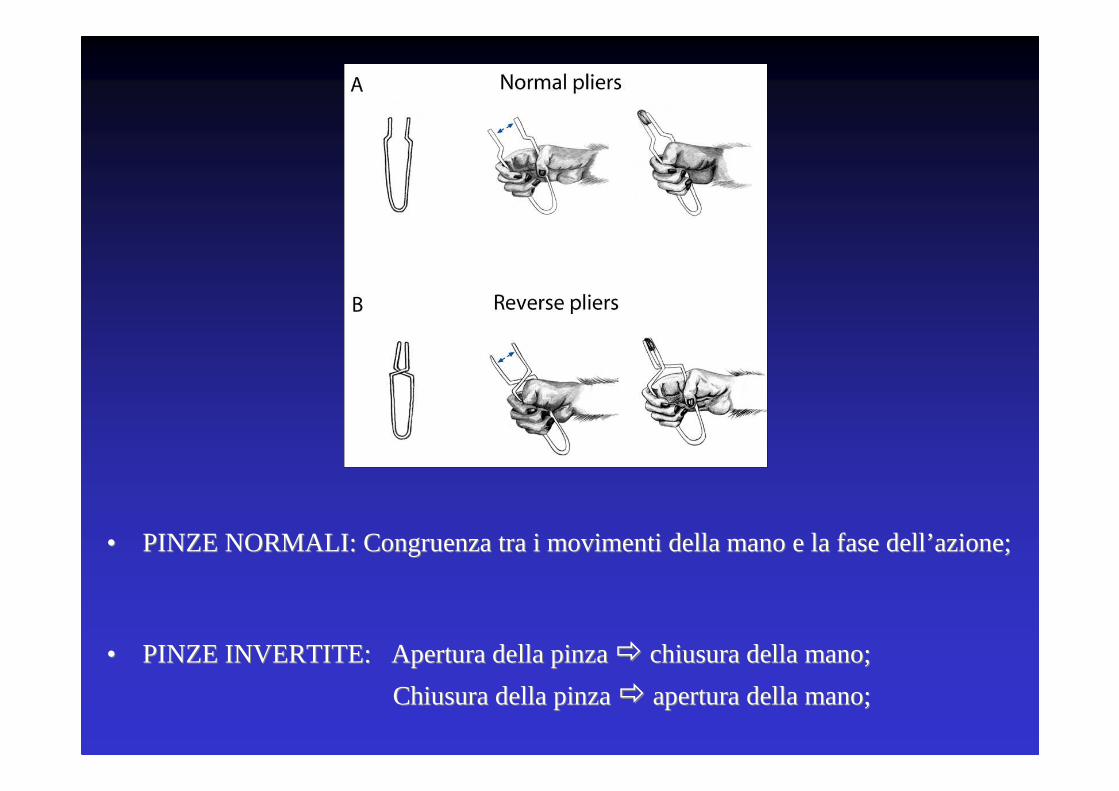
•• PINZE PINZE NORMALNORMALII: : Congruenza tra i movimenti della mano e la fase dellCongruenza tra i movimenti della mano e la fase dell’’ azioneazione;;
•• PINZE INVERTITEPINZE INVERTITE: : Apertura della pinza Apertura della pinza �� chiusura della mano;chiusura della mano;
Chiusura della pinza Chiusura della pinza �� apertura della mano;apertura della mano;
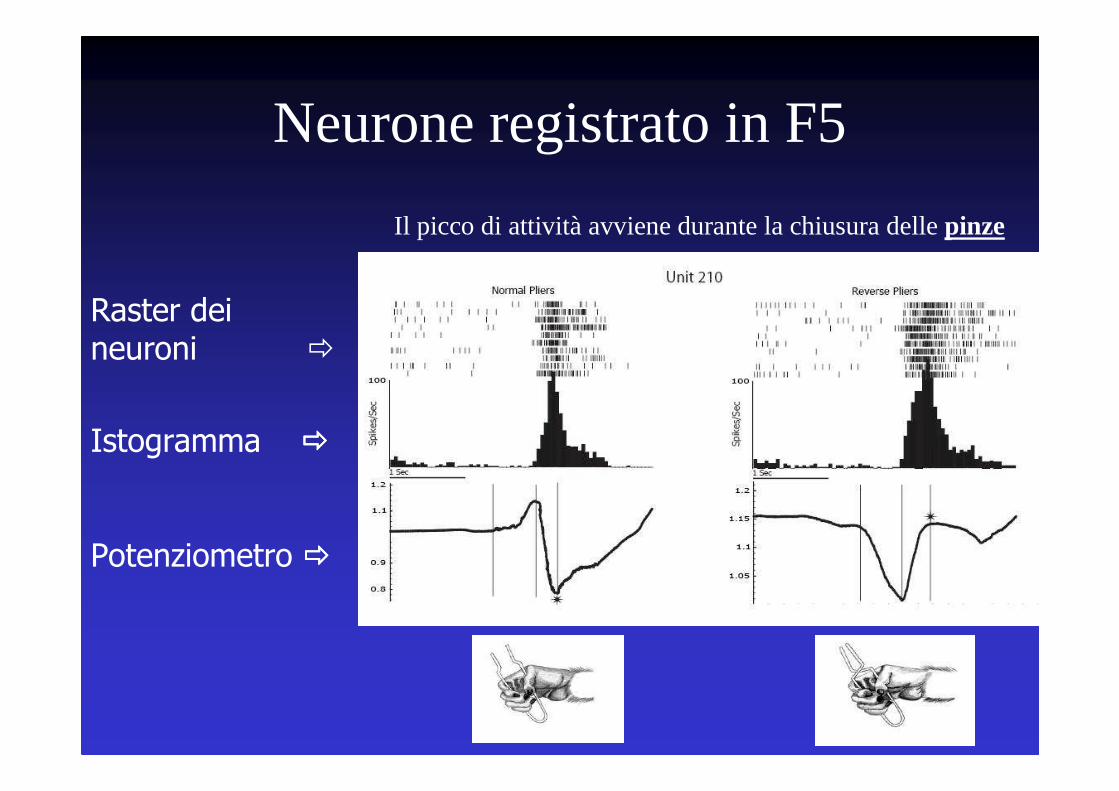
Il picco di attività avviene durante la chiusura delle pinze
Neurone registrato in F5
Raster dei neuroni �
Istogramma ����
Potenziometro ����
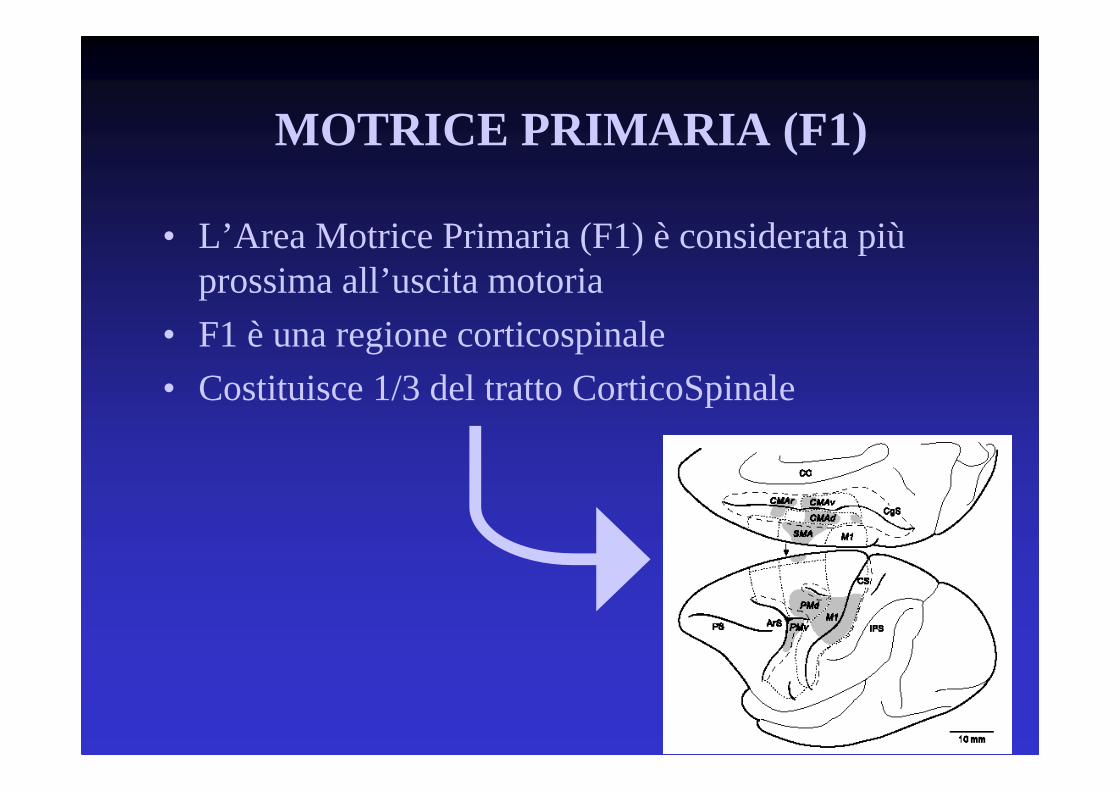
• L’Area Motrice Primaria (F1) è considerata piùprossima all’uscita motoria
• F1 è una regione corticospinale
• Costituisce 1/3 del tratto CorticoSpinale
MOTRICE PRIMARIA (F1)

Due distinte categorie di neuroni sono state trovate nell’area motrice primaria F1:
1° - Neuroni che scaricano in relazione allo scopo dell’azione dell’atto motorio (F5-like)
2° - Neuroni che scaricano in relazione ai movimenti della mano (F1m)
Registrazione nell’Area Motrice Primaria (F1)
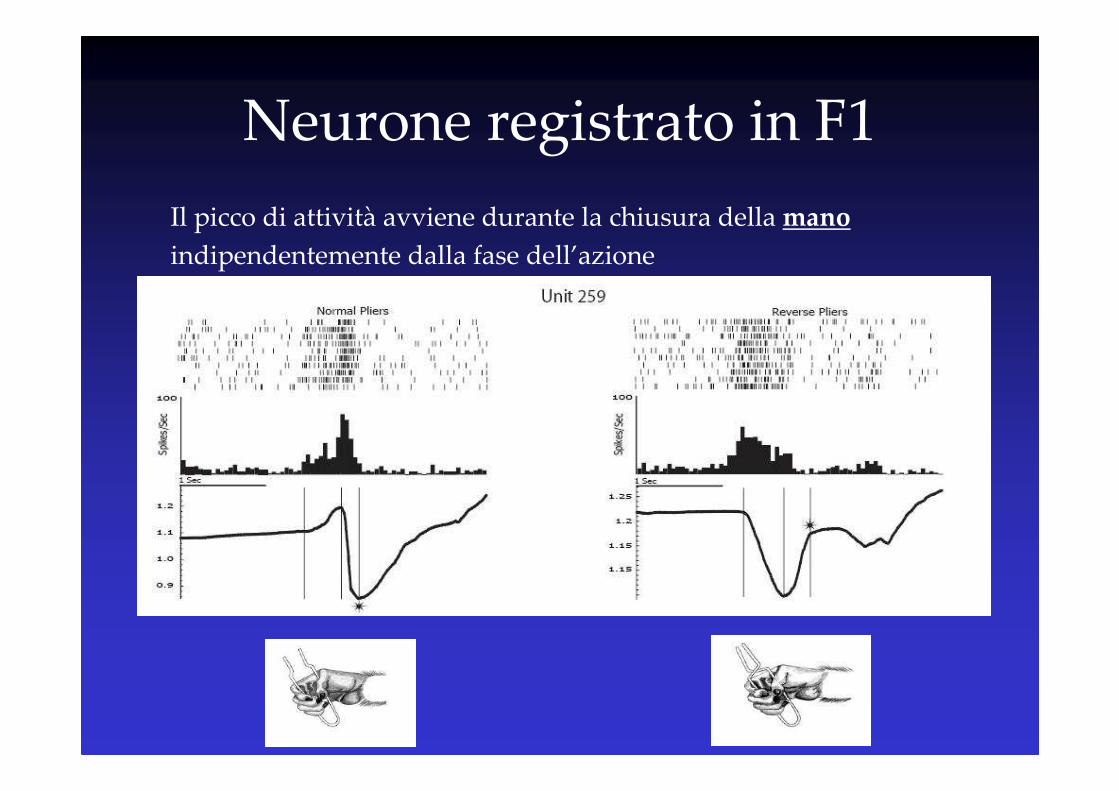
Neurone registrato in F1
Il picco di attività avviene durante la chiusura della mano
indipendentemente dalla fase dell’azione

• Se dissociamo lo scopo dell’azione dal movimento richiestoper compierla, F5 e parte della Motoria Primaria codificano l’azione dell’effettore finale, piuttosto che il movimento della mano biologica.
• Il sistema motorio e’ organizzato intorno allo scopo, al proposito dei nostri movimenti, e non intorno ai movimentistessi
… LE PINZE DIVENTANO DITA …

Un altro argomento a favore
dell’organizzazione ecologica, o
intenzionale, del Sistema Motorio
proviene da particolari studi di
microstimolazione intracorticale
Organizzazione “intenzionale”del repertorio motorio
Stimolazione IntracorticaleStimolazione Intracorticale
Tempi di Tempi di Scala Temporale
Stimolazione Stimolazione Comportamentale
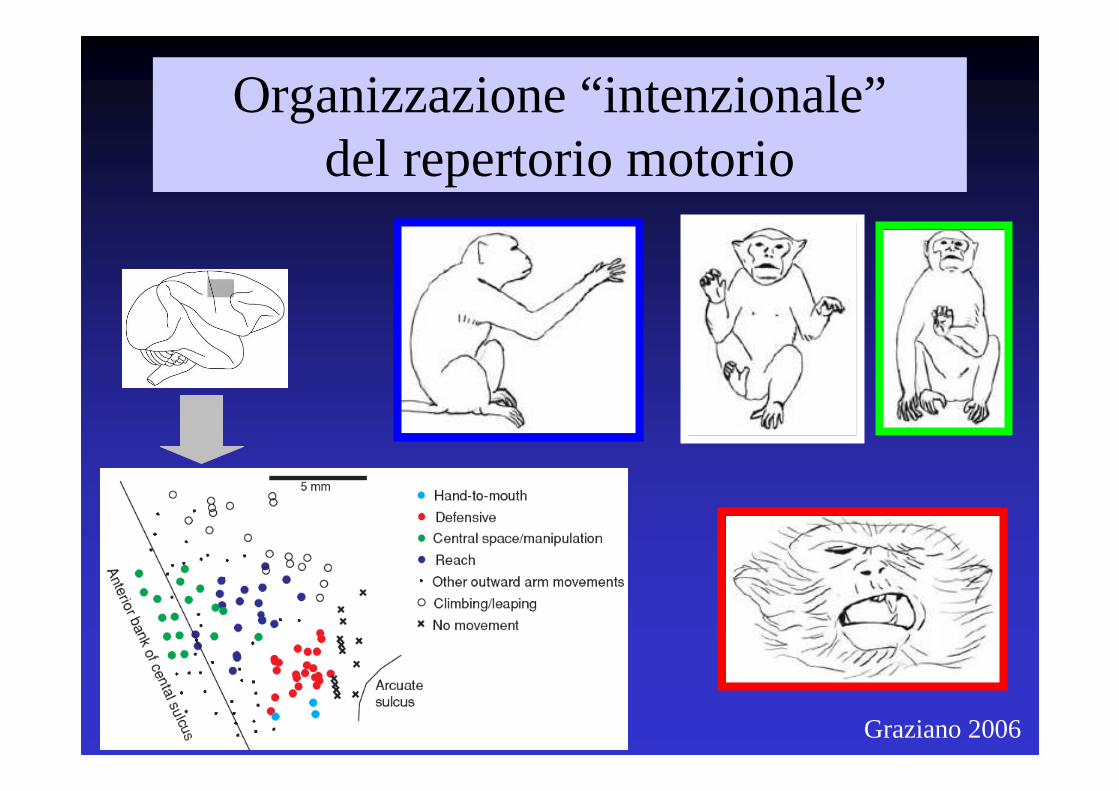
Graziano 2006
Organizzazione “intenzionale”del repertorio motorio

Il Sistema Motorio ha un’organizzazione centrata sullo scopo dell’atto motorio, e non invece sulla mappa muscolare
Una variabile importante per il sistema motorio è il “che cosa fare” (“what”) più che il “come fare”

IL “MODELLO DELLE CATENE”E IL CASO DELLE AZIONI COMPLESSE
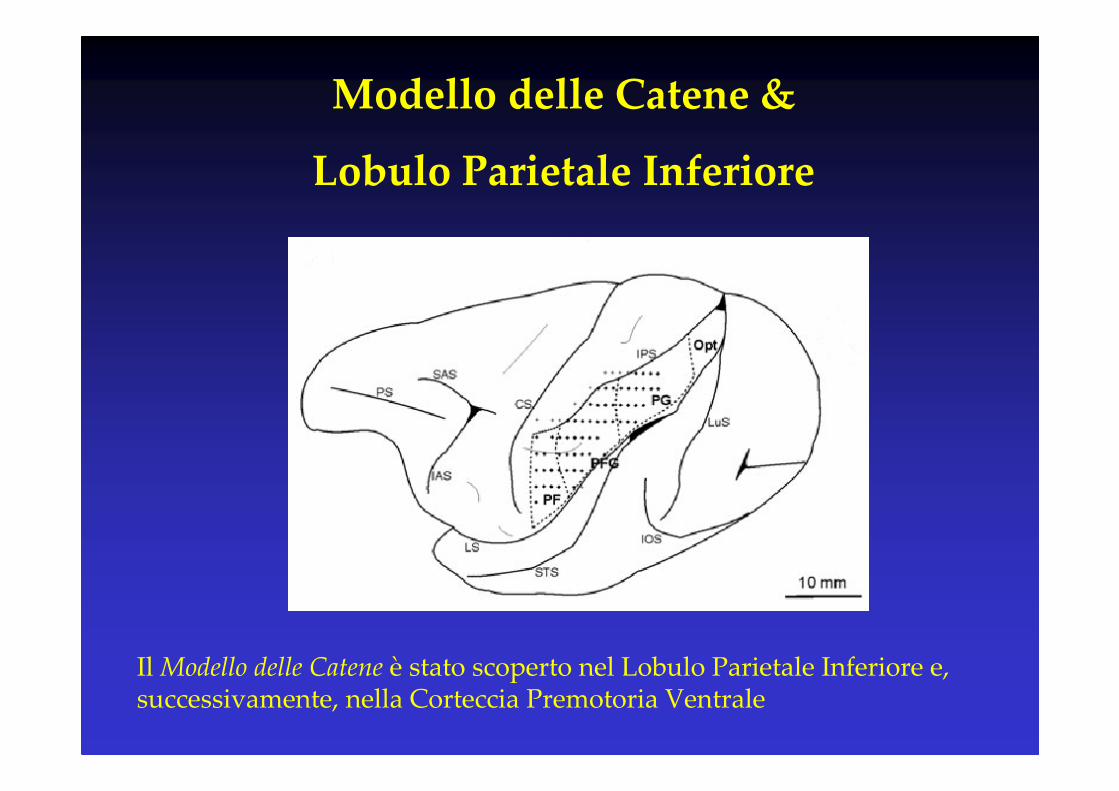
Il Modello delle Catene è stato scoperto nel Lobulo Parietale Inferiore e, successivamente, nella Corteccia Premotoria Ventrale
Modello delle Catene &
Lobulo Parietale Inferiore

La corteccia parietale posteriore (PPC)
• Classicamente la corteccia parietale posteriore (PPC) èstata considerata una larga area associativa, in cui afferenze da differenti modalità sensoriali vengono integrate per fornire la base per processi percettivi (percezione dello spazio, percezione del proprio corpo)
• Questi processi percettivi erano considerati indipendenti dalle funzioni motorie
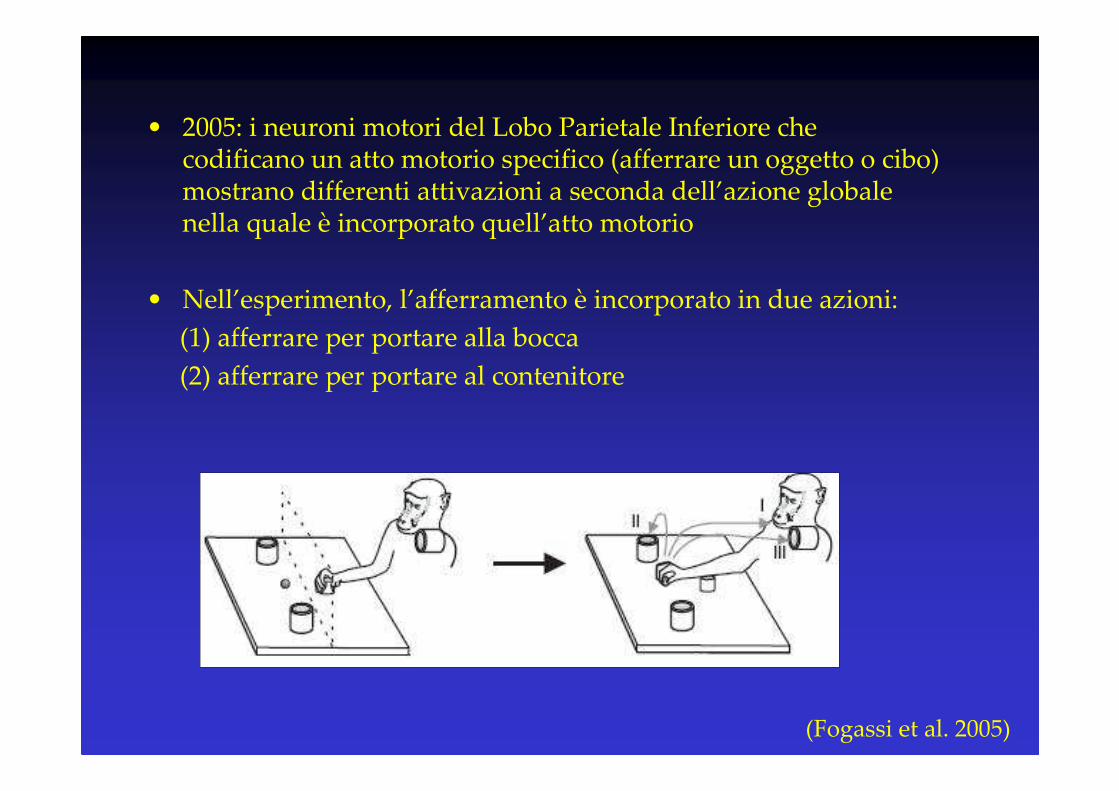
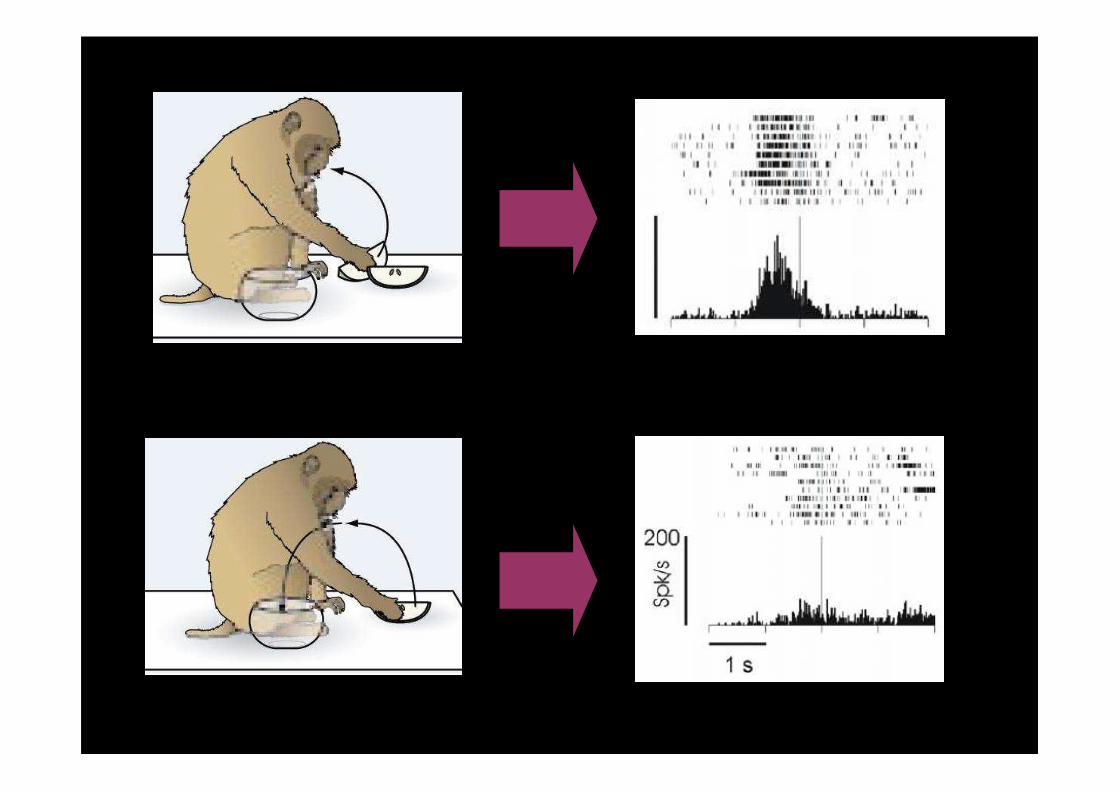
• 2005: i neuroni motori del Lobo Parietale Inferiore che codificano un atto motorio specifico (afferrare un oggetto o cibo) mostrano differenti attivazioni a seconda dell’azione globale nella quale è incorporato quell’atto motorio
• Nell’esperimento, l’afferramento è incorporato in due azioni:
(1) afferrare per portare alla bocca
(2) afferrare per portare al contenitore
(Fogassi et al. 2005)
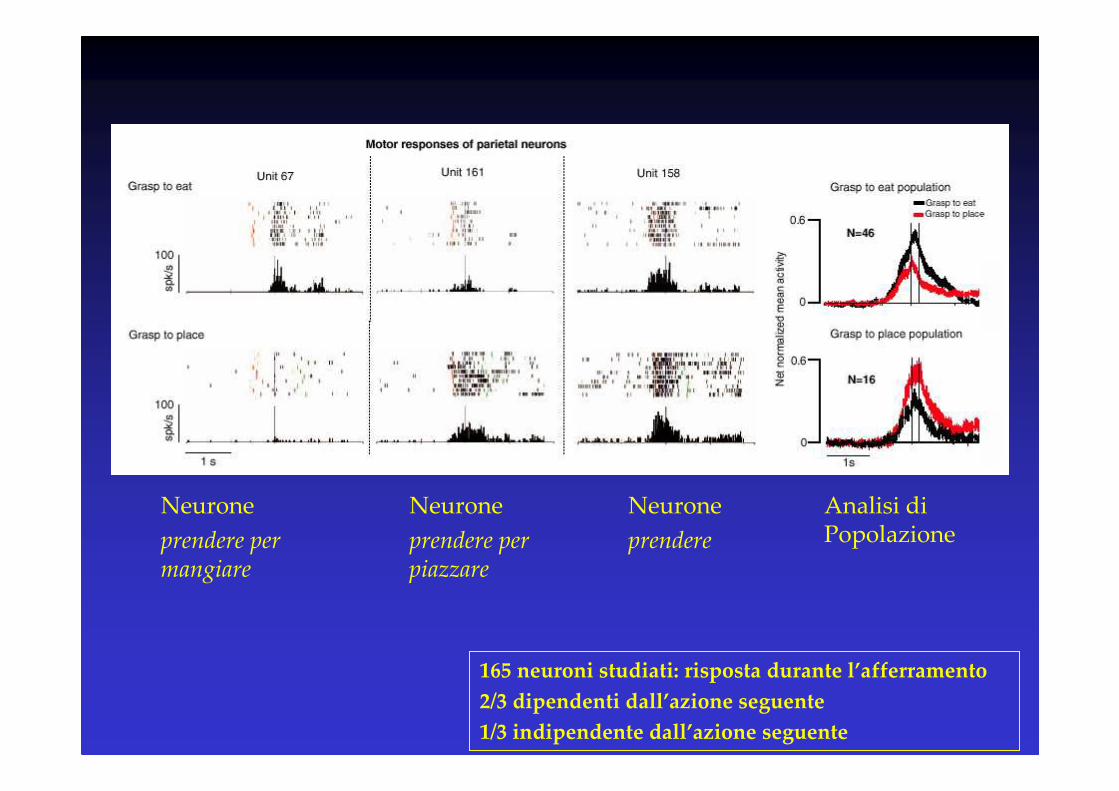
Neurone
prendere per mangiare
Neurone
prendere per piazzare
Neurone
prendere
Analisi di Popolazione
165 neuroni studiati: risposta durante l’afferramento
2/3 dipendenti dall’azione seguente
1/3 indipendente dall’azione seguente

Dati recenti hanno dimostrato con lo stesso compito sperimentale che la stessa organizzazione presente nel lobulo parietale inferiore esiste nella corteccia premotoria ventrale

Il Modello delle Catene
Gli atti motori non sono correlati l’uno con l’altro indipendentemente dallo scopo finale dell’azione, ma sembrano appartenere ad una catena intenzionale precostituita, nella quale ogni atto motorio è facilitato dal precedente

I NEURONI ‘MIRROR’
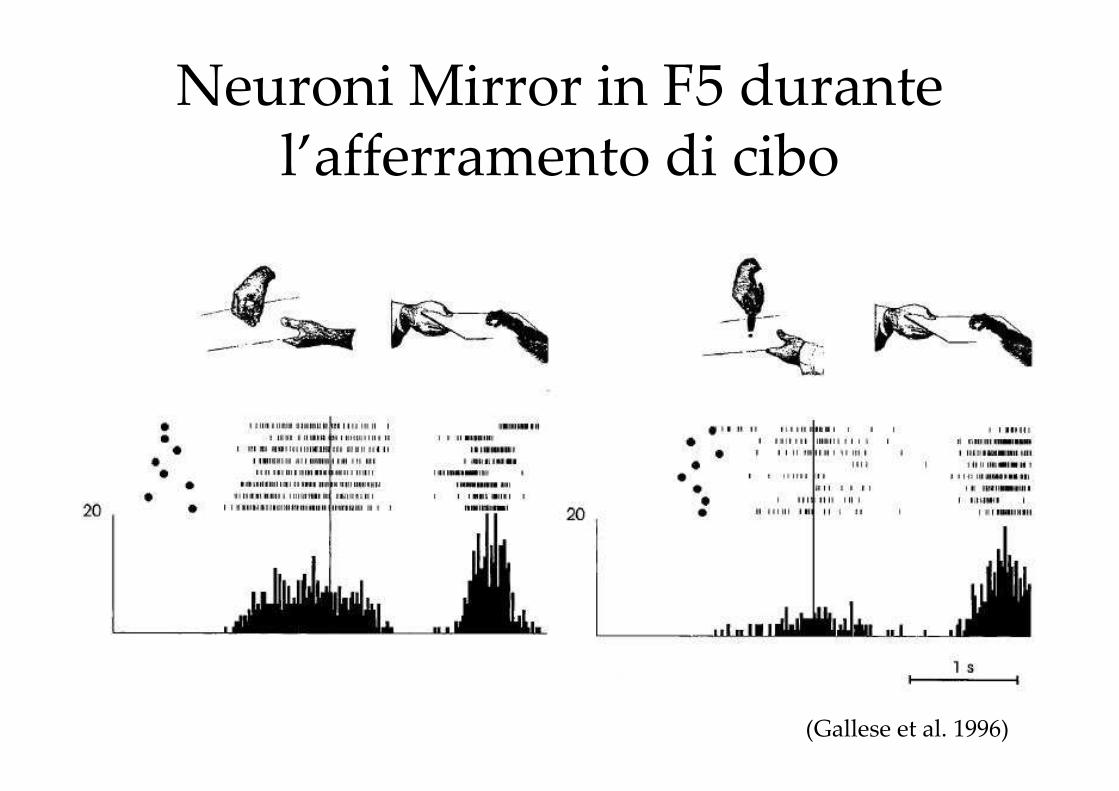
Neuroni Mirror in F5 durante l’afferramento di cibo
(Gallese et al. 1996)
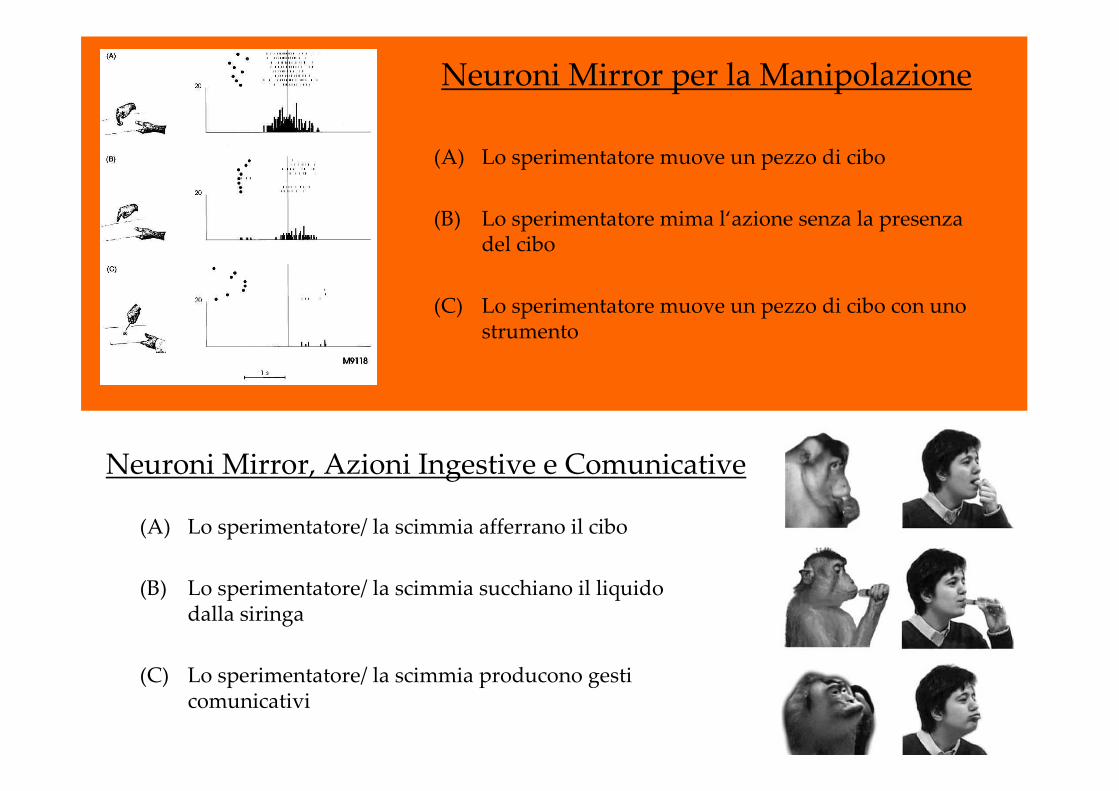
Neuroni Mirror per la Manipolazione
(A) Lo sperimentatore muove un pezzo di cibo
(B) Lo sperimentatore mima l‘azione senza la presenza del cibo
(C) Lo sperimentatore muove un pezzo di cibo con uno strumento
Neuroni Mirror, Azioni Ingestive e Comunicative
(A) Lo sperimentatore/ la scimmia afferrano il cibo
(B) Lo sperimentatore/ la scimmia succhiano il liquido dalla siringa
(C) Lo sperimentatore/ la scimmia producono gesti comunicativi

L’importanza dei ‘neuroni mirror’ di per le azioni di bocca
La presenza di neuroni mirror di bocca può essere un punto di partenza particolare per spiegare il fenomeno di imitazione neonatale
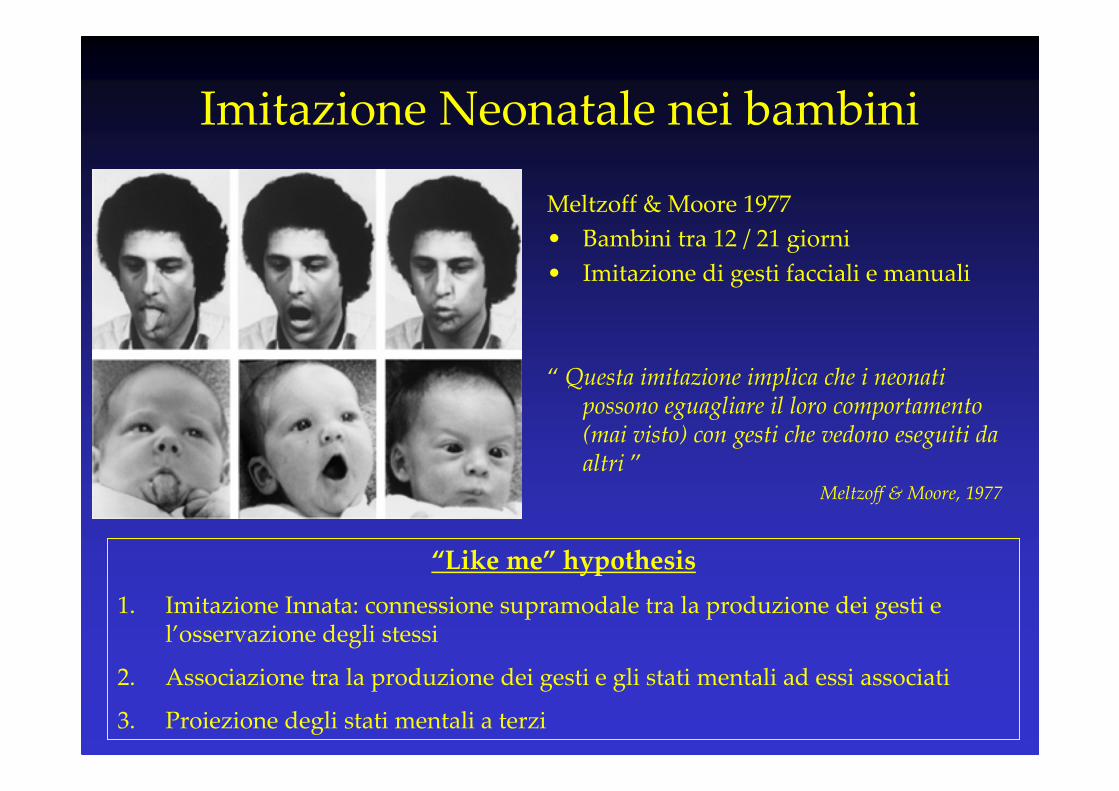
Imitazione Neonatale nei bambini
Meltzoff & Moore 1977
• Bambini tra 12 / 21 giorni
• Imitazione di gesti facciali e manuali
“ Questa imitazione implica che i neonati possono eguagliare il loro comportamento (mai visto) con gesti che vedono eseguiti da altri ”
Meltzoff & Moore, 1977
“Like me” hypothesis
1. Imitazione Innata: connessione supramodale tra la produzione dei gesti e l’osservazione degli stessi
2. Associazione tra la produzione dei gesti e gli stati mentali ad essi associati
3. Proiezione degli stati mentali a terzi

Imitazione Neonatale nei Macachi
Apertura della bocca
Protrusione della lingua
Ferrari et al., 2006

Un’ipotesi è che:
• i neonati possiedano già alla nascita un sistema di neuroni specchio che rende possibile l’imitazione
• il sistema inibitore dell’imitazione (lobo prefrontale) è ancora debole e di modesta funzionalità (scarsa mielinizzazione)
• Il sistema dei neuroni specchio potrebbe dunque essere:(1) innato(2) basato su uno schema corporeo multimodale (poiché i neonati non hanno mai visto la propria bocca)

Che cosa viene mappato sul Sistema Mirror?
Movimenti? Azioni?

• I Neuroni Mirror sono modulati dall’osservazione della fase di chiusura dell’effettore e dal raggiungimento dello scopo finale…e non dall’osservazione del movimento biologico che compone l’azione
• Il Sistema Mirror (nella scimmia) non mappa i movimenti corporei

Ruolo dell’Esperienza Motoria

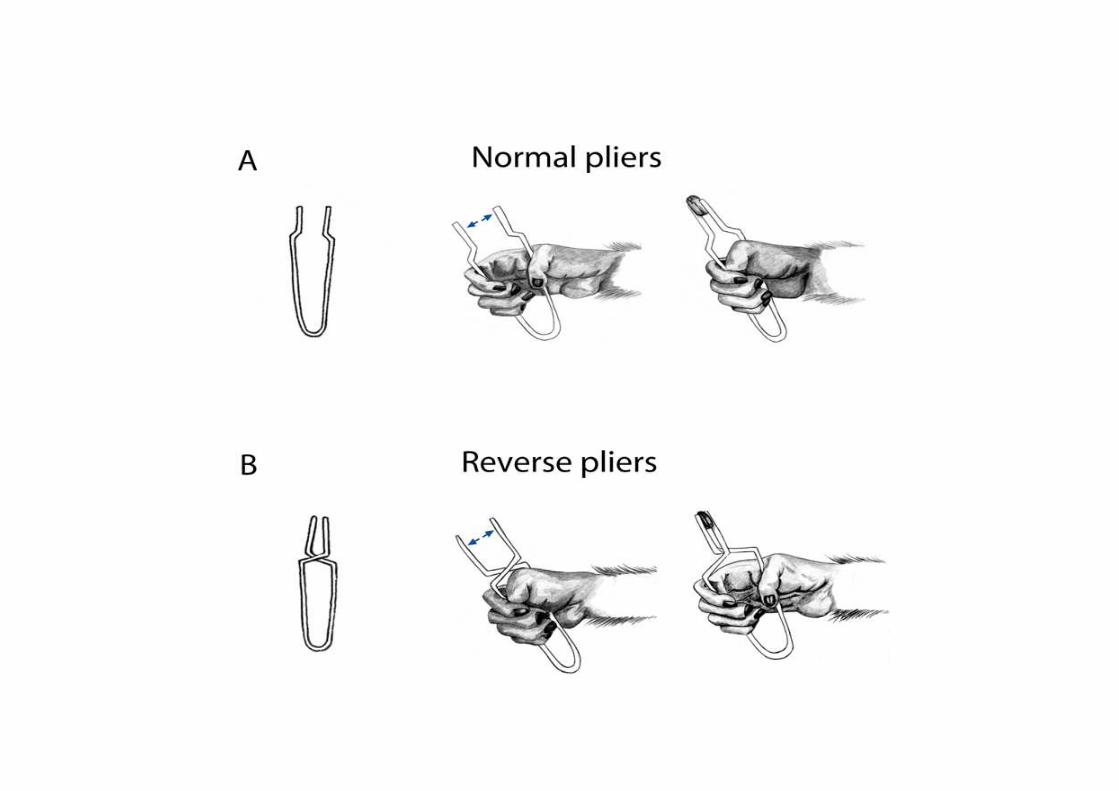
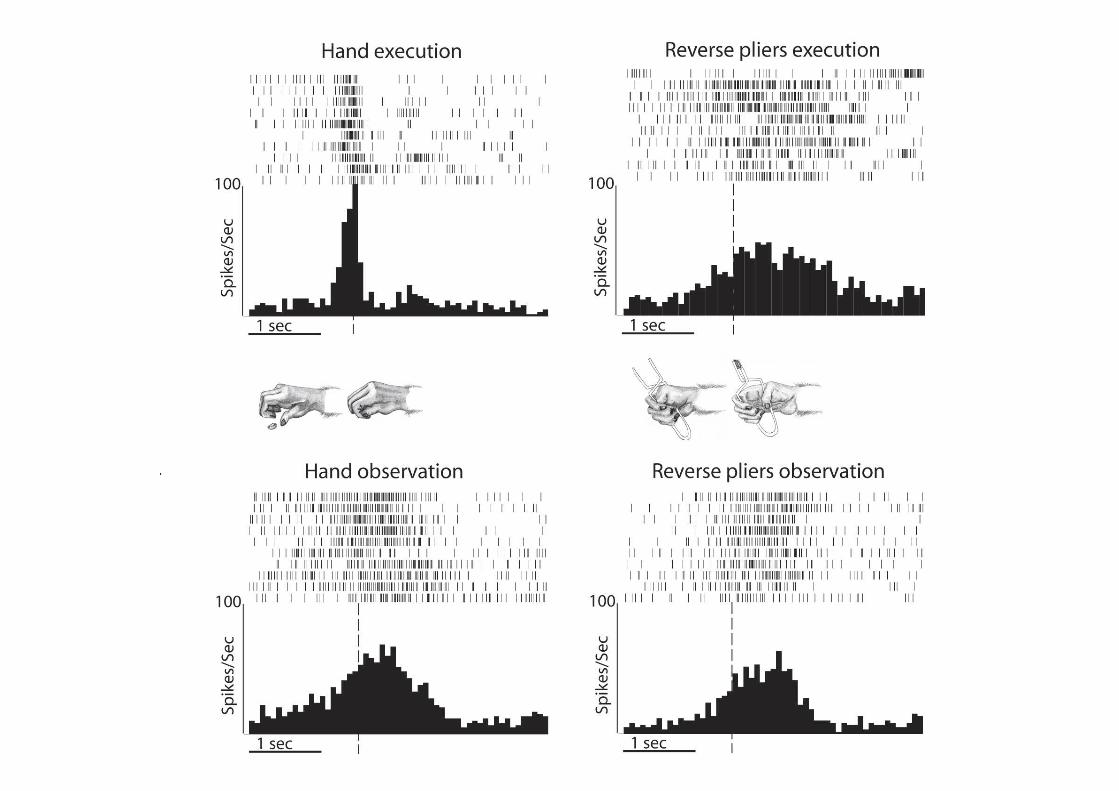
I neuroni mirror della premotoria ventrale sono stati nuovamentetestati durante l’osservazione di:
• Afferramento manuale;
• Afferramento con uno strumento che la scimmia sa utilizzare (pinze invertite);
• Afferramento con uno strumento che la scimmia non sa utilizzare (bastone con punta);
‘Tool-responding mirror neurons’
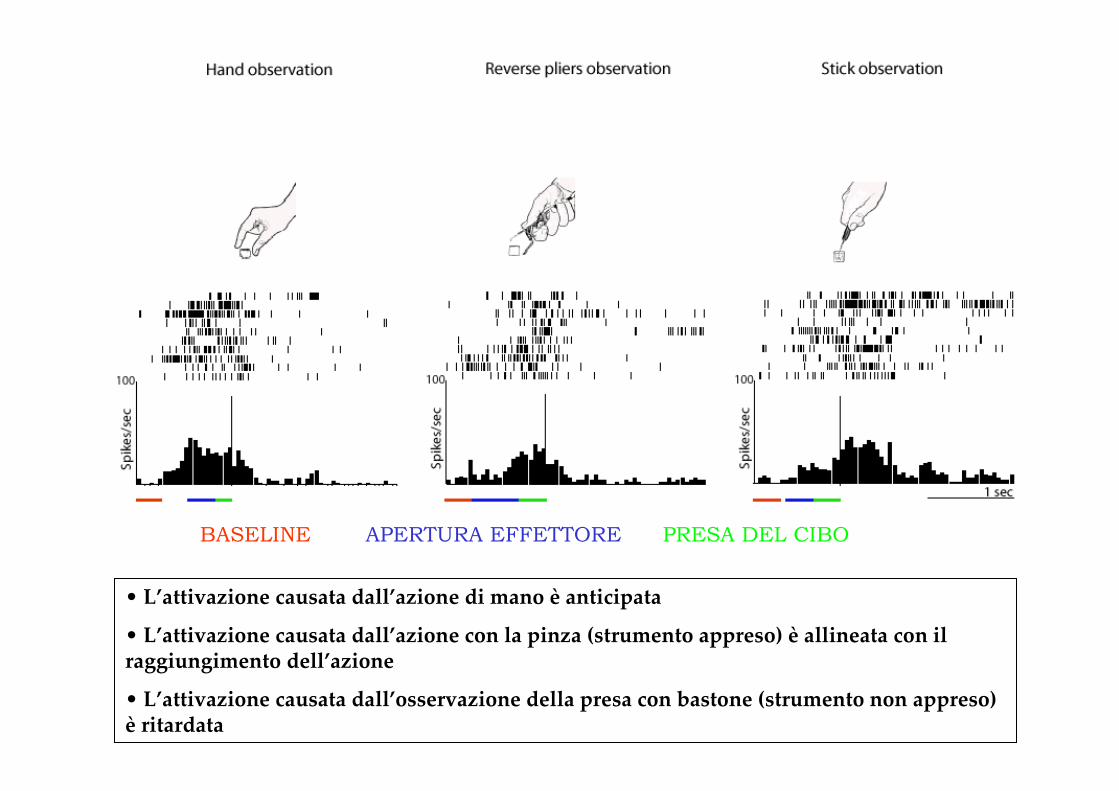
• L’attivazione causata dall’azione di mano è anticipata
• L’attivazione causata dall’azione con la pinza (strumento appreso) è allineata con il raggiungimento dell’azione
• L’attivazione causata dall’osservazione della presa con bastone (strumento non appreso) è ritardata
BASELINE APERTURA EFFETTORE PRESA DEL CIBO

La capacità di attivarsi e quindi di anticipare lo scopo dell’azione osservata è dipendente dal grado di familiarità del sistema motorio con l’azione stessa
Comprendiamo prima (e anticipiamo prima) le azioni che sappiamo eseguire

• Sappiamo che alcuni neuroni dell’area F5 partecipano all’azione con una codifica astratta dello scopo
• L’osservazione dell’azione eseguita da un conspecificoattiva nel cervello la stessa rete di neuroni motori usualmente attivata per eseguire quella particolare azione
• La percezionedell’azione NON E’ puramente pittorica, ma implica invece un’attivazione inconscia e automatica del Sistema Motorio

La percezione dell’azione
come attività costruttiva

(Umiltà et al. 2001)
Oggetto visibile Oggetto non visibile
Azione mimata visibile Azione mimata non visibile
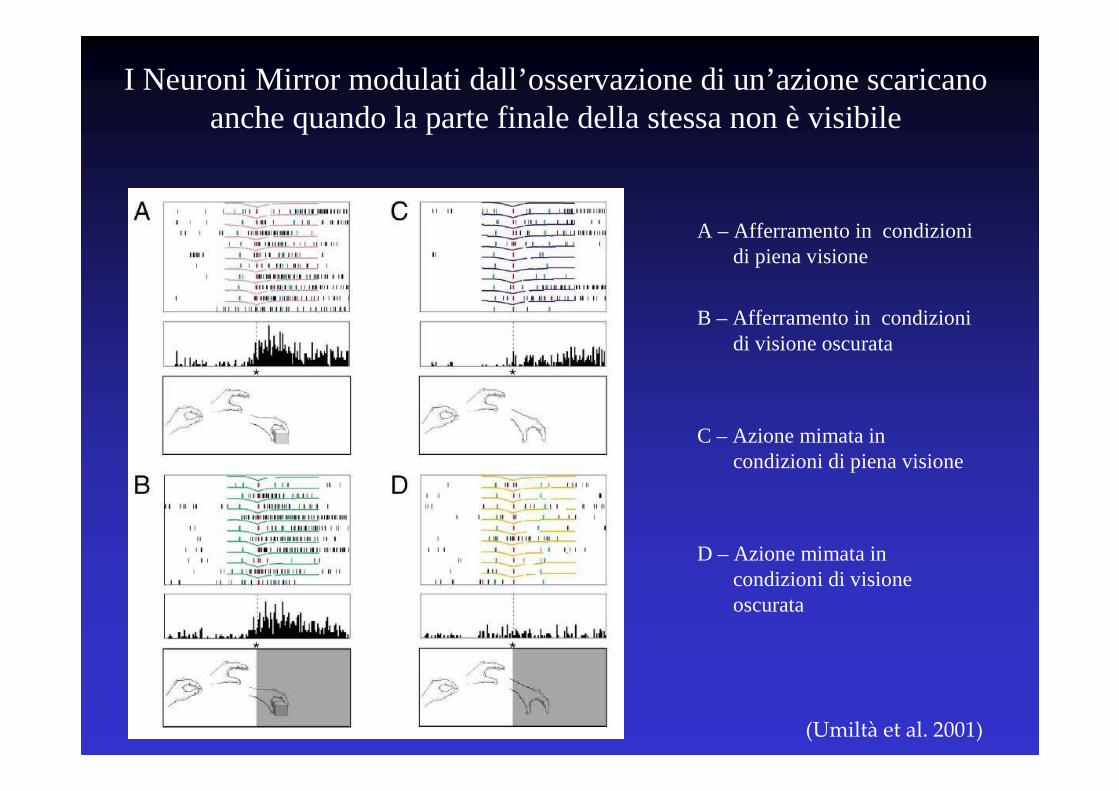
A – Afferramento in condizioni di piena visione
B – Afferramento in condizioni di visione oscurata
C – Azione mimata in condizioni di piena visione
D – Azione mimata in condizioni di visione oscurata
(Umiltà et al. 2001)
I Neuroni Mirror modulati dall’osservazione di un’azione scaricano anche quando la parte finale della stessa non è visibile

Perché ‘percezione’?
• Nelle Scienze Cognitive la percezione non è implicata con l’idea empirista di una “impressione” passiva sulla mente, ma èconsiderata come un’attività costruttiva
• Secondo la Psicologia della Gestalt “l’elemento minimo della percezione è la forma”
• Nella condizione oscurata cogliamo l’intenzione motoria dell’agente reclutando quella parte del proprio sistema motorio usualmente coinvolta nella stessa azione
• Il salto che si pone tra “ciò che vede” e “ciò che percepisce” viene riempito dal reclutamento del sistema motorio
• La percezione dell’azione è dunque un processo costruttivo, in cui il sistema motorio risulta essere direttamente coinvolto

“L’appareil sensoriel, tel que la physiologie moderne
se le représente, n’est plus propre au rôle de
trasmetteur que la science classique lui faisait jouer.
[…] Le fonctionnement normal doit être compris
comme un processus d’intégration où le texte du
monde extérieur est non pas recopié, mais constitué”
M. Merleau-Ponty, Phenomenologie de la Perception

“A mere visual perception, without involvement of
the motor system would only provide a description
of the visible aspects of the movements of the
agent, but it would not give precise information
about the intrisic components of the observed
action which are critical for understanding what
the action is about, what is its goal, and how to
reproduce it”
M. Jeannerod , Action from within
I neuroni Mirror audio-motori
…se la percezione dell’azione è un processo costruttivo, allora le cose dovrebbero valere anche per la percezione uditiva di un’azione!

… vediamo e ascoltiamouna azione
… vediamouna azione
… ascoltiamouna azione
Cosa fanno i Neuroni Mirror mentre …
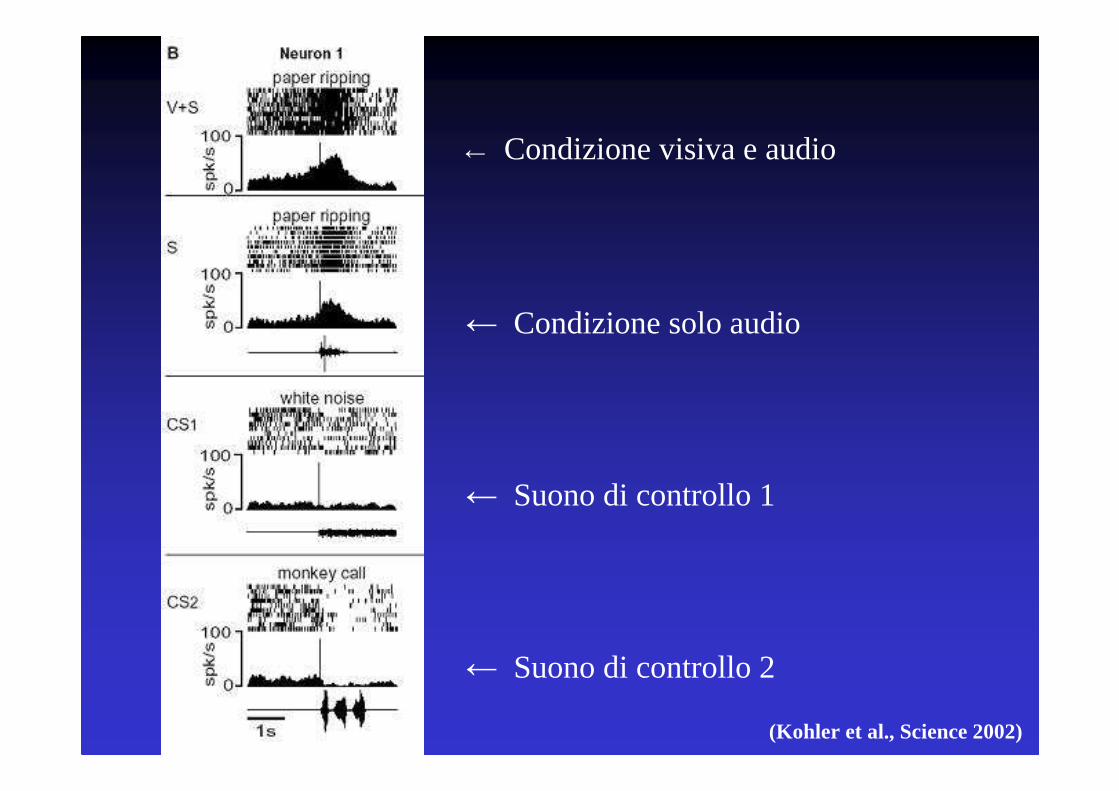
(Kohler et al., Science 2002)
← Condizione visiva e audio
← Condizione solo audio
← Suono di controllo 1
← Suono di controllo 2

Il Sistema Mirror ci aiuta a predire le azioni?
Il Sistema Mirror e la comprensione delle intenzioni
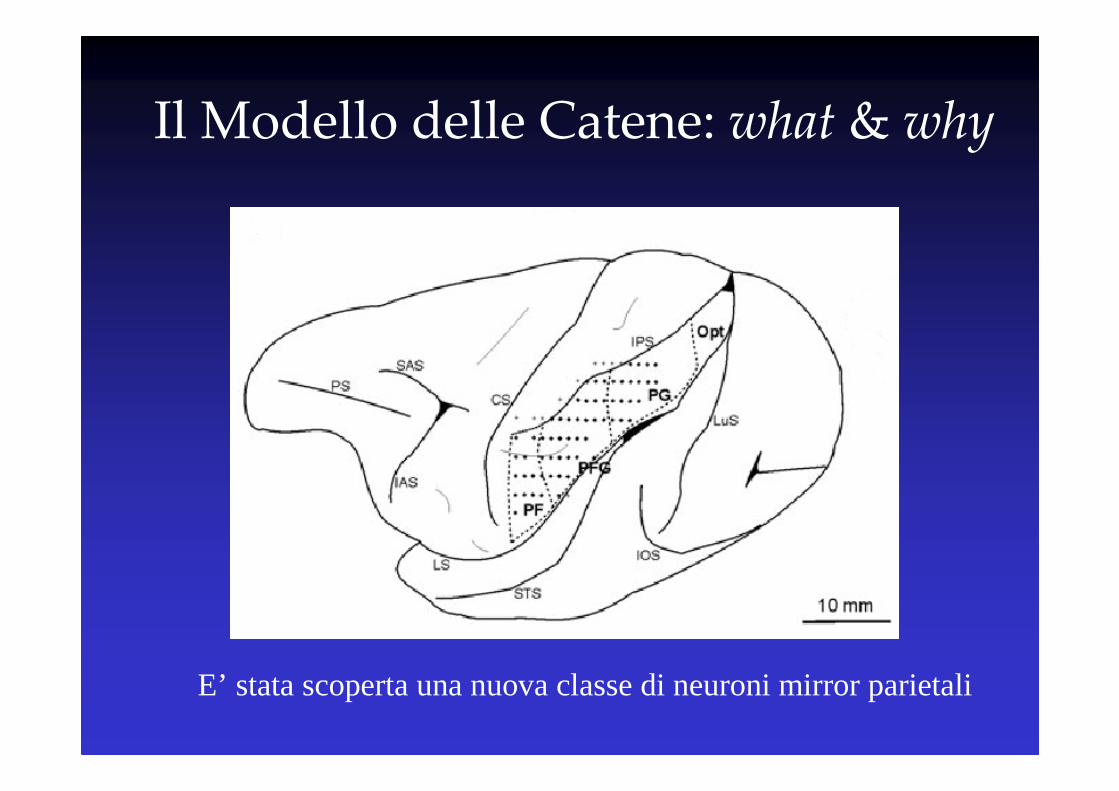
Il Modello delle Catene: what & why
E’ stata scoperta una nuova classe di neuroni mirror parietali
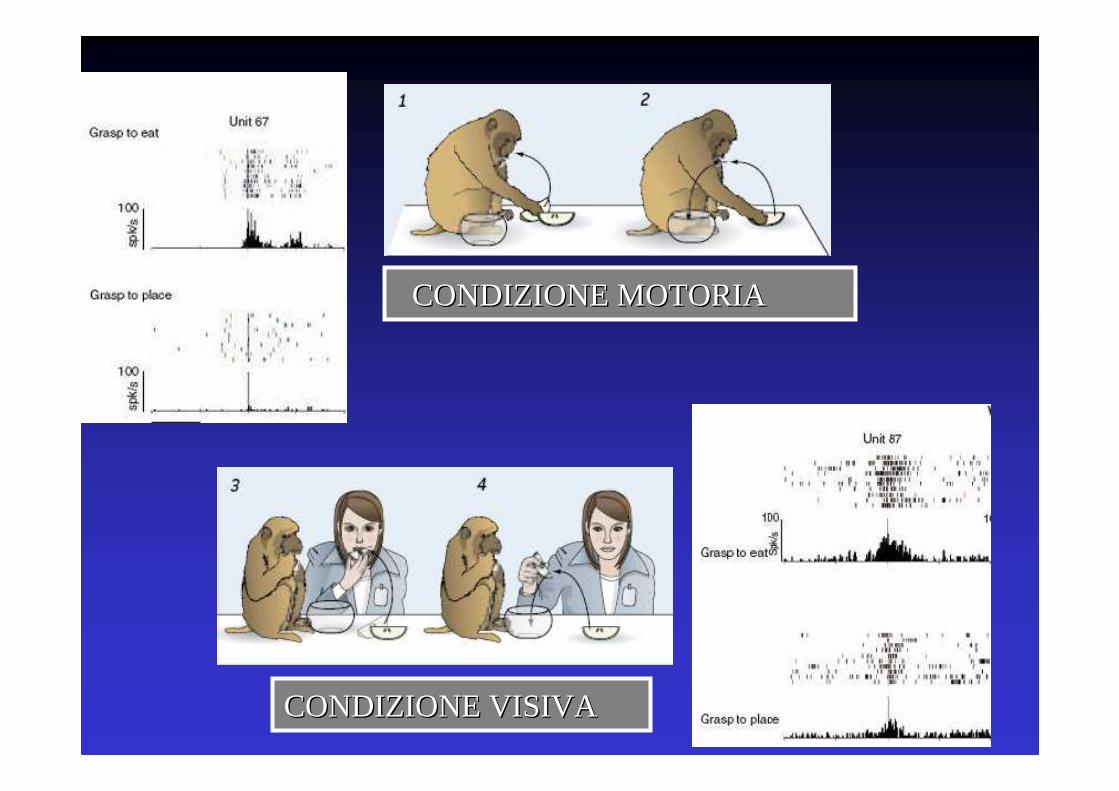
CONDIZIONE MOTORIACONDIZIONE MOTORIA
CONDIZIONE VISIVA CONDIZIONE VISIVA

Il Modello delle Catene
VISIVO
Durante l’osservazione, l’atto motorio osservato recluta un’intera e specifica catena intenzionale.
A seconda del tipo di catena motoria attivata, l’osservatore avrà una spontanea previsione del perché(“ why”) l’agente osservato fa quello che vediamo (per mangiare / per posare).
MOTORIO
Gli atti motori non sono correlati l’uno con l’altro indipendentementedallo scopo finale dell’azione, ma sembrano appartenere ad una catena intenzionale precostituita, nella quale ogni atto motorio èfacilitato dal precedente

What & Why
What:i Neuroni Mirror indicano il “cosa” fa l’altro (prende, morde, strappa, etc)
Why: le Catene Motorie mettono a disposizione del Sistema Mirror il “perché”l’altro fa qualcosa (prende per mangiare, prende per posare)
scarica corollaria?

IL SISTEMA MIRROR NELL’UOMO
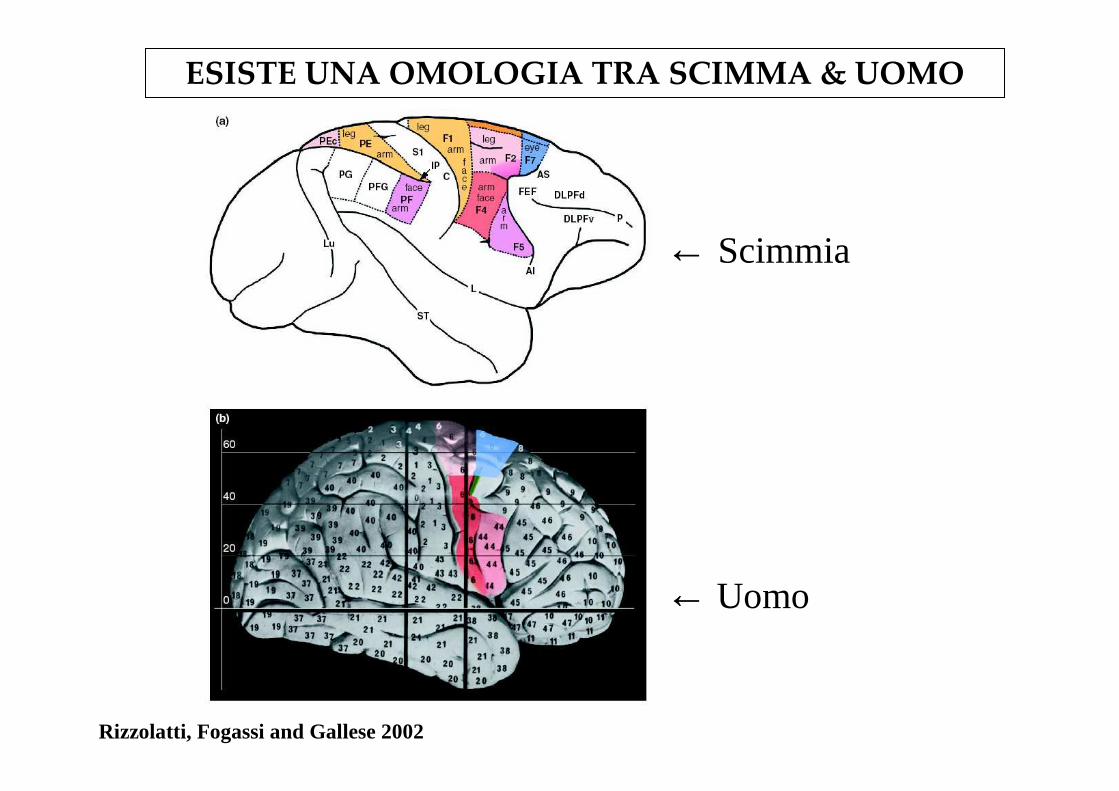
Rizzolatti, Fogassi and Gallese 2002
← Scimmia
← Uomo
ESISTE UNA OMOLOGIA TRA SCIMMA & UOMO
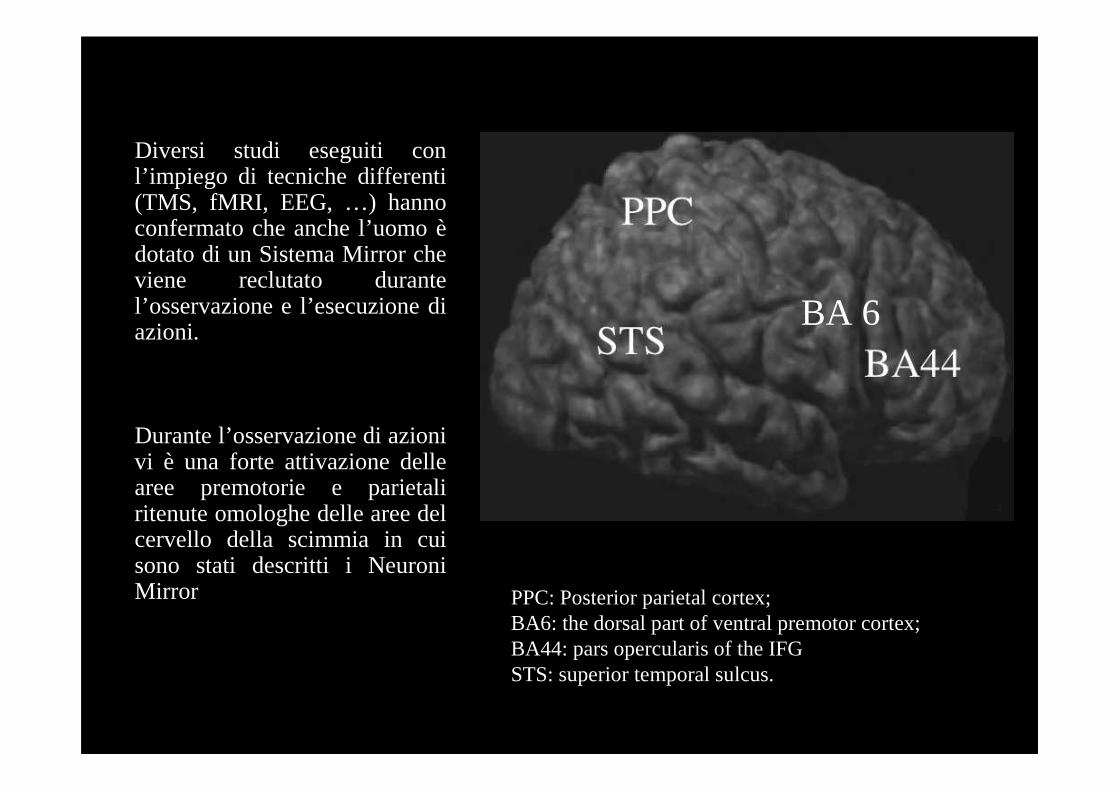
Diversi studi eseguiti con l’impiego di tecniche differenti(TMS, fMRI, EEG, …) hannoconfermato che anche l’uomo èdotato di un Sistema Mirror cheviene reclutato durantel’osservazione e l’esecuzione diazioni.
Durante l’osservazione di azionivi è una forte attivazione dellearee premotorie e parietaliritenute omologhe delle aree del cervello della scimmia in cui sono stati descritti i NeuroniMirror PPC: Posterior parietal cortex;
BA6: the dorsal part of ventral premotor cortex;BA44: pars opercularis of the IFGSTS: superior temporal sulcus.
BA 6

Somatotopia dell’Osservazione di Azioni
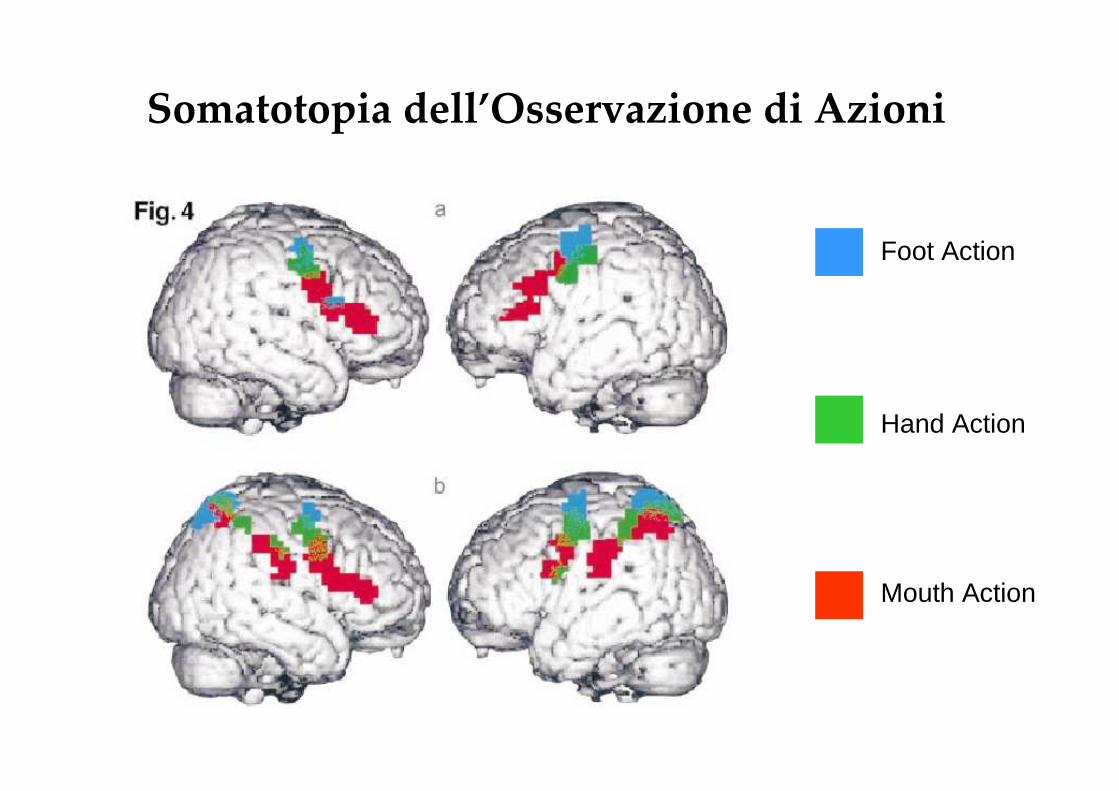
Somatotopia dell’Osservazione di Azioni
Foot Action
Hand Action
Mouth Action

AZIONI DI BOCCA COMUNICATIVE

UOMO CHE PARLA
LIP SMACKING DELLA SCIMMIA
ABBAIARE DEL CANE
Buccino et al., 2004.
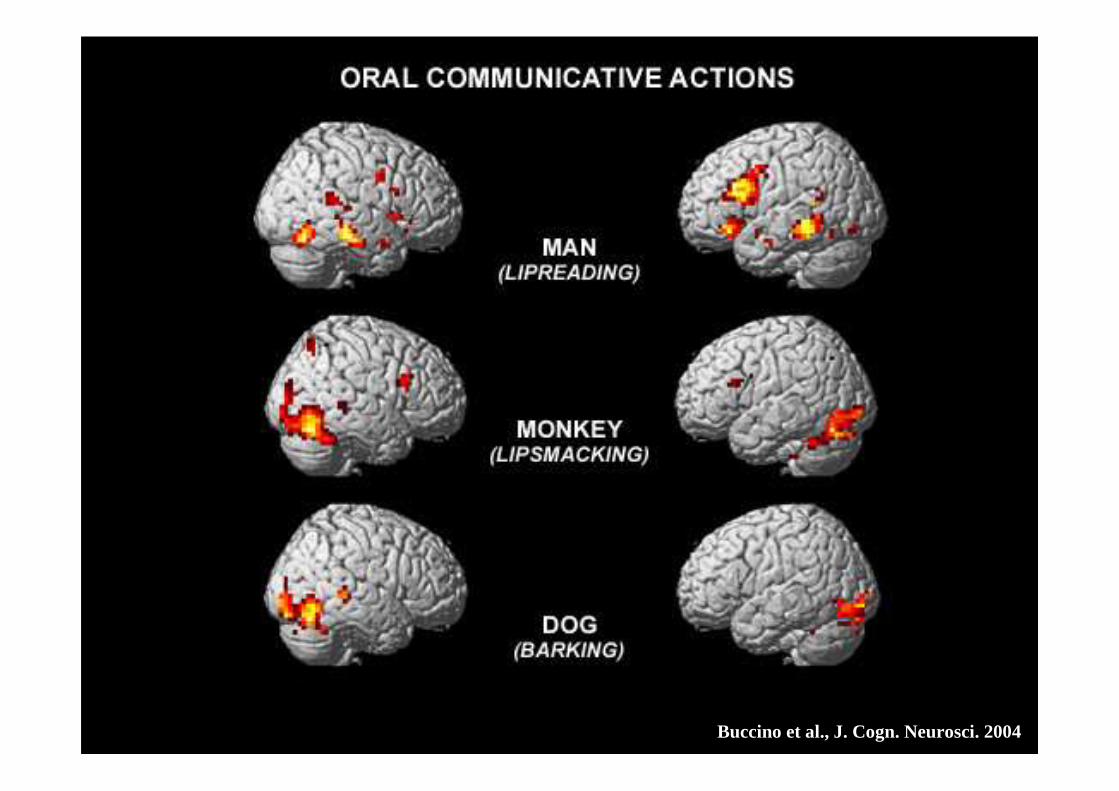
Buccino et al., J. Cogn. Neurosci. 2004

UOMO: pars opercularis del giro frontale inferiore, un settore dell’area di Broca
SCIMMIA: porzione minore della stessa regione
CANE: esclusivamente le aree visive exstrastriate
• Esistono dunque differenti modi di comprendere un’azione comunicativa, a seconda del soggetto che stiamo osservando
• In particolare, se l’azione appartiene al repertorio motorio dell’osservatore- o è simile ad un’azione che appartiene al repertorio motoriodell’osservatore - allora questa viene mappata anche nel sistema motorio
• Altrimenti, come nel caso dell’abbaiare, la comprensione dell’azione vieneaffidata principalmente ai sistemi implicati nell’analisi visiva

Fino a che punto è rilevante che un’azione appartenga al repertorio motorio dell’osservatore?
Fino a che punto questa congruenza modula l’attivazione del Sistema Mirror?

Abbiamo visto dal modello animale che i neuroni mirror rispondono all’osservazione di azioni con tempistiche diverse a seconda del grado di familiaritàmotoria
Le abilità motorie umane apprese offrono una via preferenziale per testare la funzione del Sistema Mirror, poiché le persone differiscono reciprocamente nel repertorio di azioni conosciute

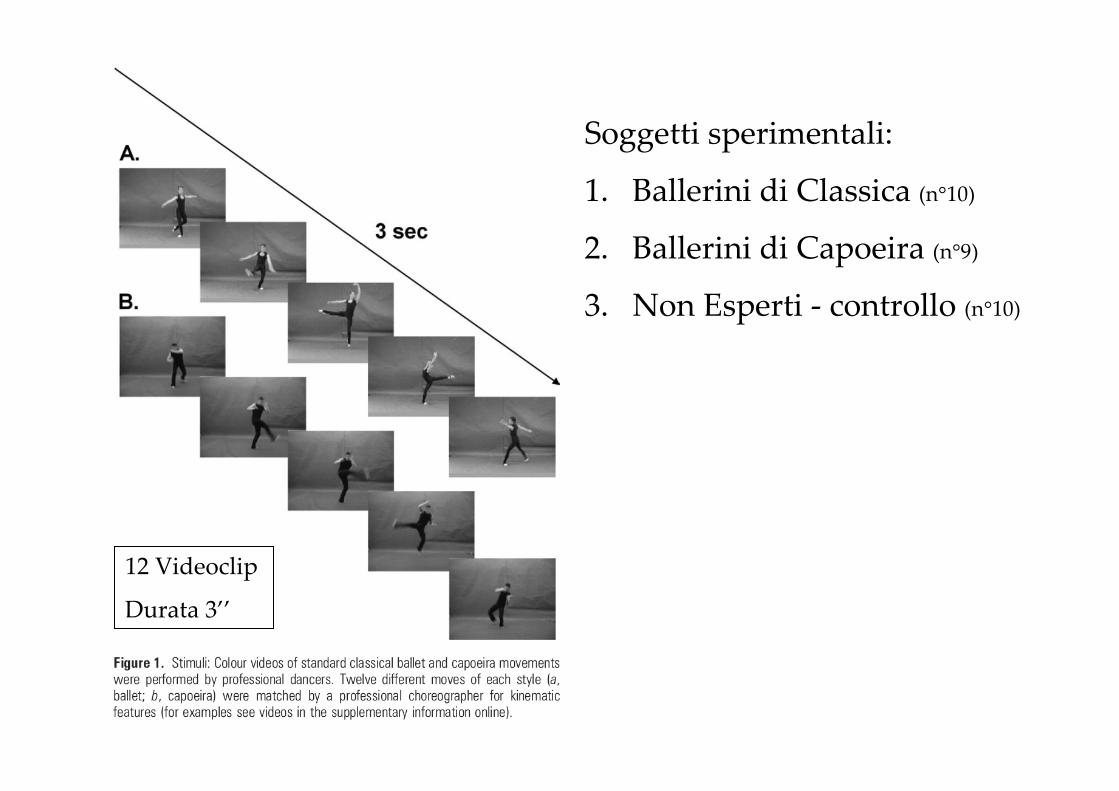
Soggetti sperimentali:
1. Ballerini di Classica (n°10)
2. Ballerini di Capoeira (n°9)
3. Non Esperti - controllo (n°10)
12 Videoclip
Durata 3’’
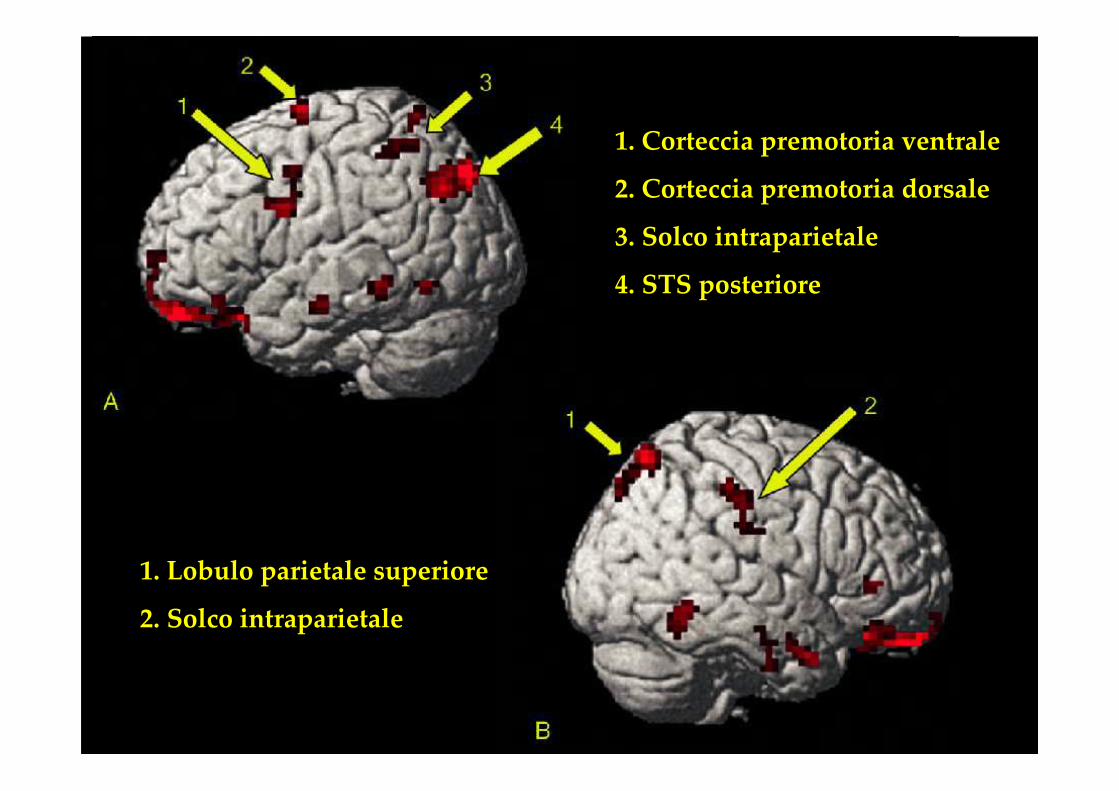
1. Corteccia premotoria ventrale
2. Corteccia premotoria dorsale
3. Solco intraparietale
4. STS posteriore
1. Lobulo parietale superiore
2. Solco intraparietale

1. Attivazione del Sistema Mirror nei 3 gruppi
2. Maggiore attivazione durante la condizione congruente
3. Insensibilità al tipo di stimolo nei soggetti Non Esperti

• Il confronto dimostra la rilevanza dell’esperienza motoria nell’osservazione dell’azione
• “Our results show that this ‘mirror system’ integrates observed actions of others with an individual’s personal motor repertoire, and suggest that the human brain understands actions by motor simulation”
• Gli autori evidenziano che i movimenti, e quindi i gruppi muscolariimplicati nelle due discipline, sono sostanzialmente gli stessi.
• Conseguentemente, la differenza tra i due gruppi dimostra che ilsistema mirror non mappa i movimenti, ma le abilità motorie

IL SISTEMA MIRROR
&
LA MOTOR IMAGERY
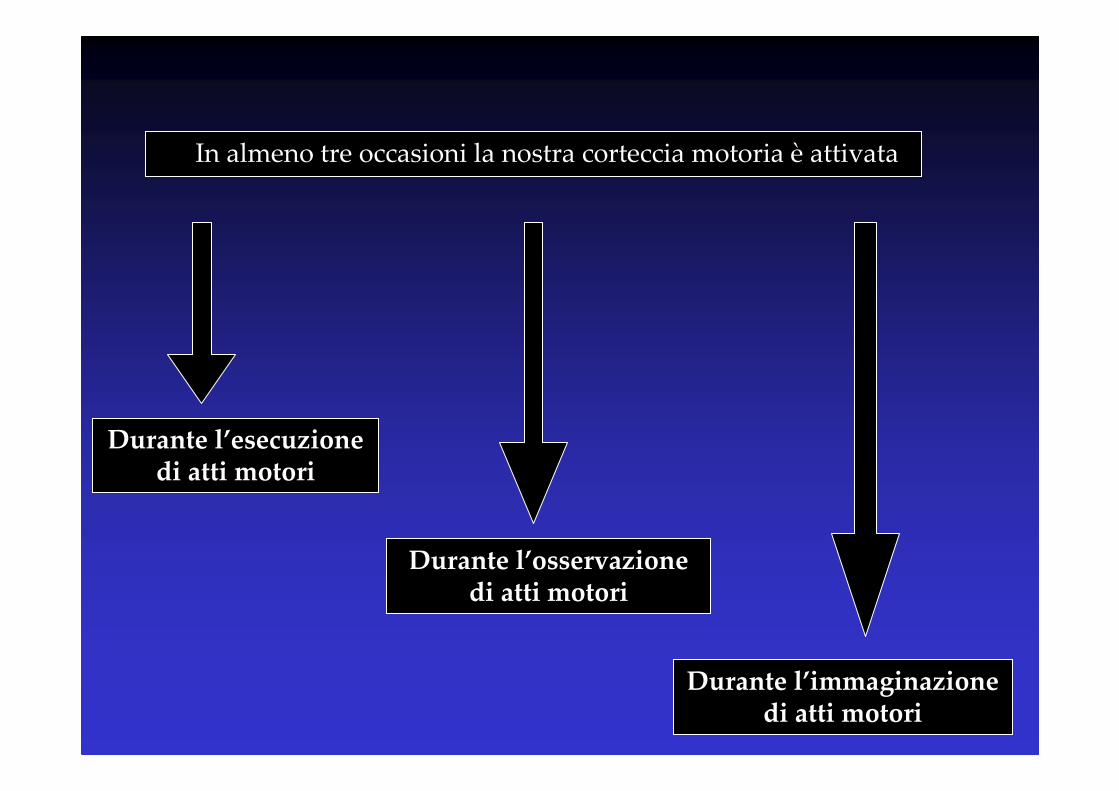
In almeno tre occasioni la nostra corteccia motoria è attivata
Durante l’osservazione di atti motori
Durante l’immaginazione di atti motori
Durante l’esecuzione di atti motori

Motor imagery
Studi apparsi negli ultimi 20 anni hanno dimostrato che l’immaginazione di una azione richiede le stesse caratteristiche temporali corrispondenti alla reale esecuzione.
Camminare verso punti diversi di una stanza, richiede gli stessi tempi, sia che il compito sia eseguito fisicamente o mentalmente.

Anche nelle operazioni ‘non coscienti’ si verifica questa analogia: immaginare di afferrare un oggetto piazzato in differenti orientamenti, richiede tempi diversi a seconda dell’orientamento.
Questo Suggerisce che il soggetto debba mentalmente ruotare il braccio nella posizione appropriata, con tempi corrispondenti alla reale esecuzione
Motor imagery

Decety et al., 1991:
Il grado di aumento di frequenza cardiaca e ventilazione polmonare di un soggetto che corre mentalmente a 12km/h èparagonabile a quello di un soggetto che corre realmente a 5km/h
Non può essere spiegato come un aumento della domanda metabolica periferica
Negli esperimenti di Motor Imagery sono stati rilevate variazioni dei parametri del SNA

Il Sistema Mirror
è un ‘sottoprodotto’ della
Motor Imagery?
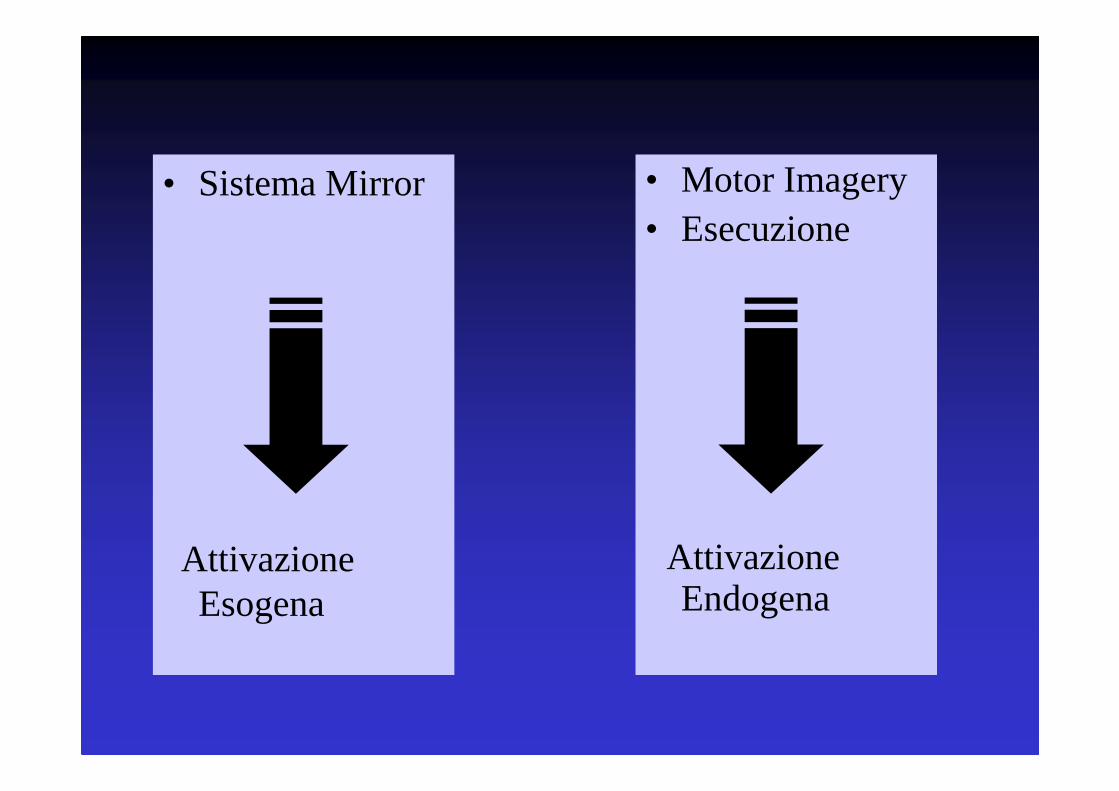
• Sistema Mirror
Attivazione Esogena
• Motor Imagery• Esecuzione
Attivazione Endogena

Che cosa viene mappato dai due sistemi?
Movimenti? Azioni?

• Abbiamo detto che nella Scimmia, sia durante l’esecuzione motoria che durante l’osservazione, la dissociazione tra movimento e scopo dell’azione dimostra che i Neuroni Mirror della corteccia premotoria F5 sono modulati dallo scopo, indipendentemente dalla strategia muscolare
1. Sistema Mirror
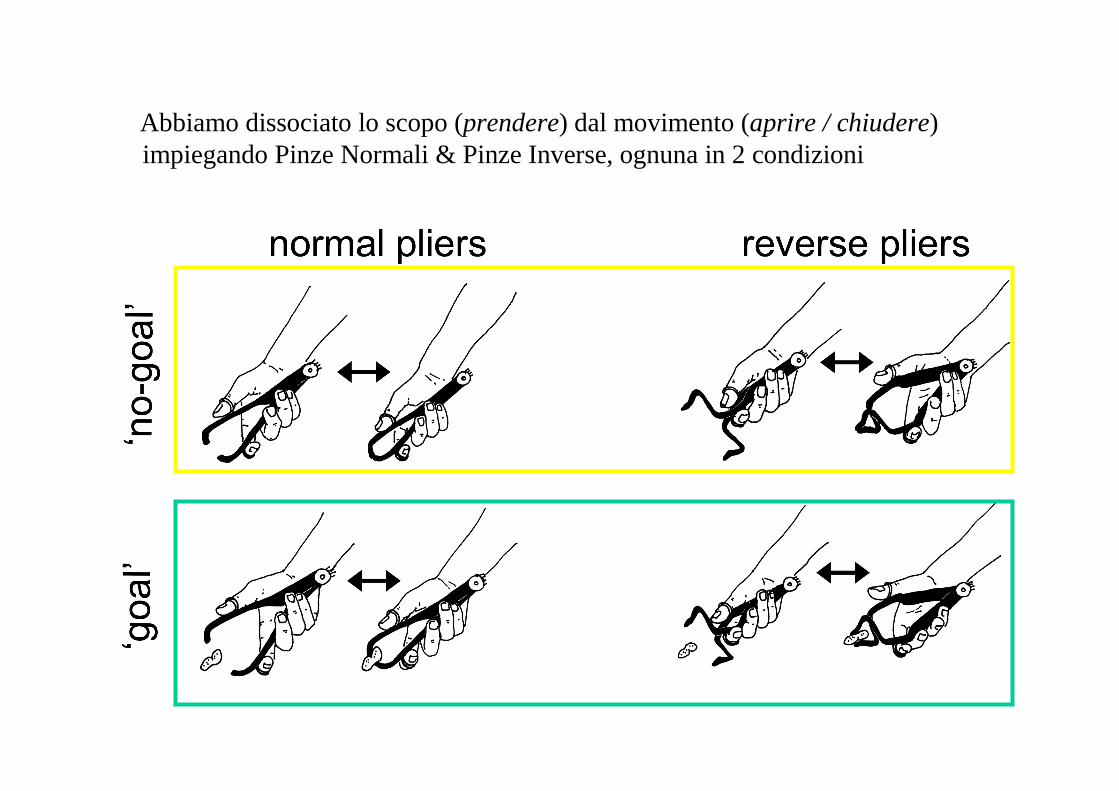
Abbiamo dissociato lo scopo (prendere) dal movimento (aprire / chiudere) impiegando Pinze Normali & Pinze Inverse, ognuna in 2 condizioni

Pinze normaliNo scopo
Pinze invertiteNo scopo
Osservare movimenti senza scopo
Facilitazione durante l’osservazione della chiusura
della mano
Facilitazione durante l’osservazione della chiusura
della mano
(mentre la pinza si apre)

Pinze normaliScopo
Pinze invertiteScopo
Osservare movimenti con uno scopo
Facilitazione durante l’osservazione della chiusura
della mano (e della pinza)
Facilitazione durante l’osservazione della chiusura
della pinza!!!
(e non della mano!!!)

Se è presente uno scopo, il sistema mirror mappa lo scopo e quindi l’effettore finale
Per il nostro Sistema Mirror, le uniche dita da con siderare sono le “dita” della pinza!
Se NON è presente uno scopo, il sistema mirror mappa il movimento biologico della mano osservata

Cosa succede se invece di osservare queste azioni, le immaginiamo ad occhi chiusi?
2. Motor imagery

• Abbiamo eseguito lo stesso esperimento, ma anziché osservarele 4 condizioni, i soggetti dovevano immaginarle
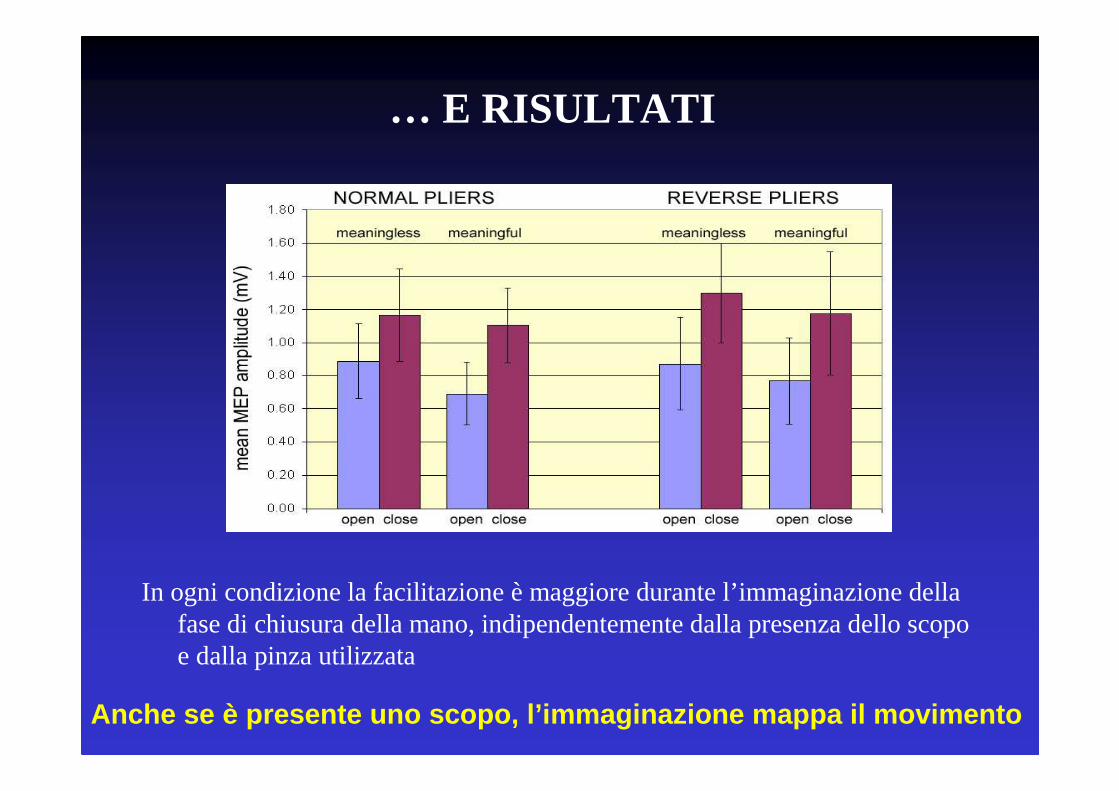
… E RISULTATI
In ogni condizione la facilitazione è maggiore durante l’immaginazione della fase di chiusura della mano, indipendentemente dalla presenza dello scopo e dalla pinza utilizzata
Anche se è presente uno scopo, l’immaginazione mappa il movimento

Le due attivazioni ‘silenti’ della corteccia motoria mostrano un comportamento diverso se testate in un compito simile.
• Quando immaginiamodi eseguire un’azione, il nostro sistema motorio simula l’esecuzione dell’atto (parametri di basso livello)
• Quando osserviamoun’azione, il nostro sistema motorio ha un comportamento diverso a seconda che l’azione sia finalizzata o meno:
(a) se non-finalizzata, il sistema motorio replica i singoli movimenti che costituiscono l’azione osservata;
(b) se finalizzata, il sistema motorio è modulato dallo scopo finale dell’azione;

ALTRI SPECCHI?

Il contagio del disgusto
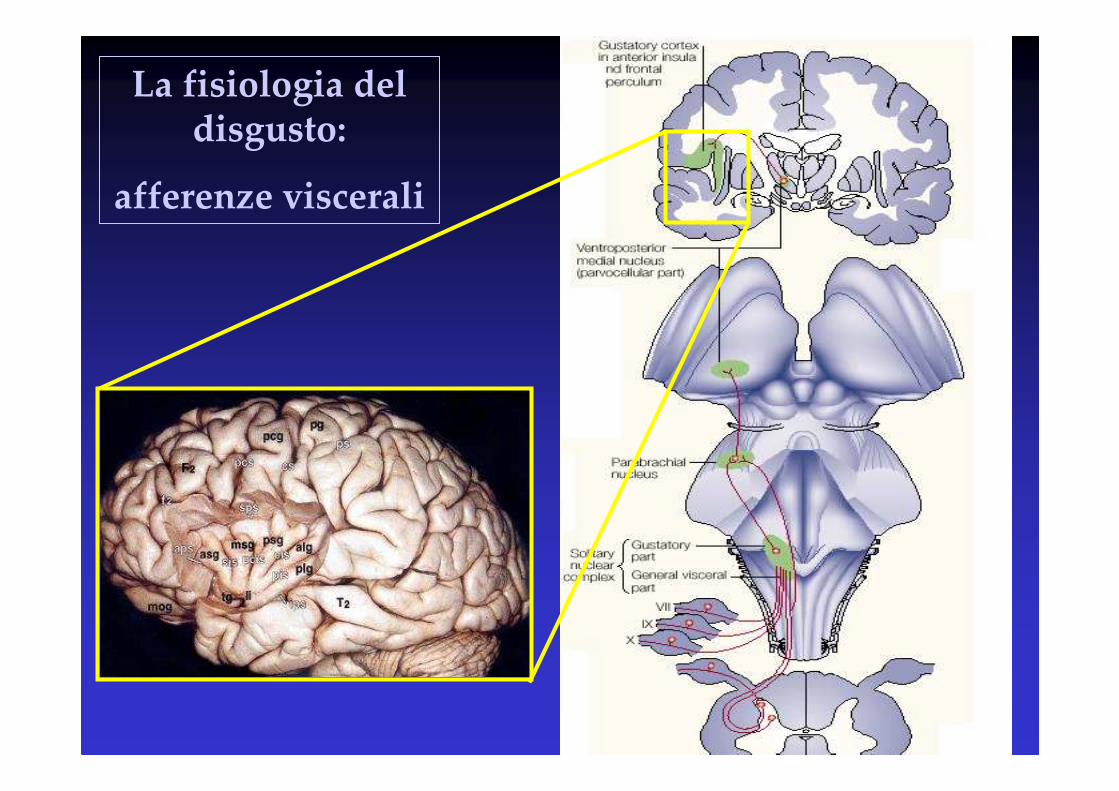
La fisiologia del disgusto:
afferenze viscerali
Buried in the depth of the lateral fissure lies the enigmatic island of Reil, whose functions and affinities are totally unknown.
Von Bonin G. & Bailey P., 1947
La notion d’épilepsie insulaire, restée dans un certain anonymat après les travaux de Penfield, a bénéficié d’un regain d’intérêt au cours des dix dernières années (…). Cependant, aucune étude systématique de l’épilepsie insulaire n’aété conduite et la sémiologie clinique des crises insulaires demeure inconnue.
Isnard J. & Mauguière F., 2005
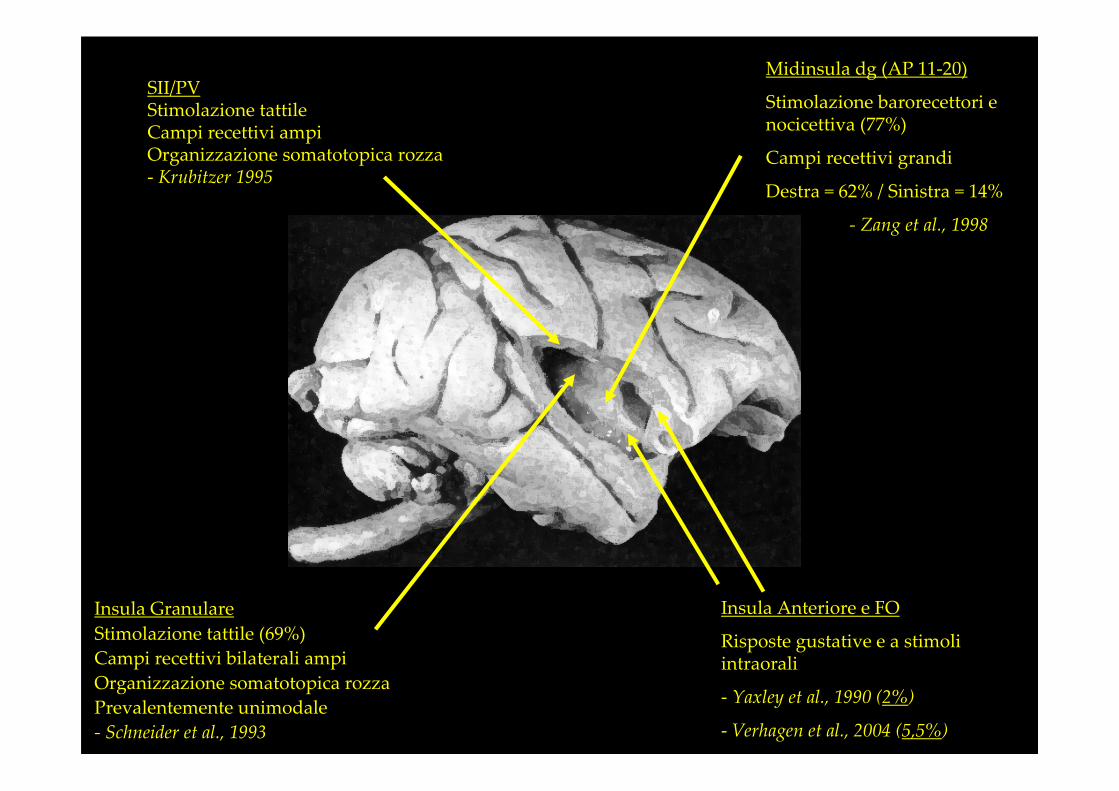
Insula Anteriore e FO
Risposte gustative e a stimoli intraorali
- Yaxley et al., 1990 (2%)
- Verhagen et al., 2004 (5,5%)
Insula Granulare
Stimolazione tattile (69%)
Campi recettivi bilaterali ampi
Organizzazione somatotopica rozza
Prevalentemente unimodale
- Schneider et al., 1993
Midinsula dg (AP 11-20)
Stimolazione barorecettori e nocicettiva (77%)
Campi recettivi grandi
Destra = 62% / Sinistra = 14%
- Zang et al., 1998
SII/PVStimolazione tattileCampi recettivi ampiOrganizzazione somatotopica rozza- Krubitzer 1995
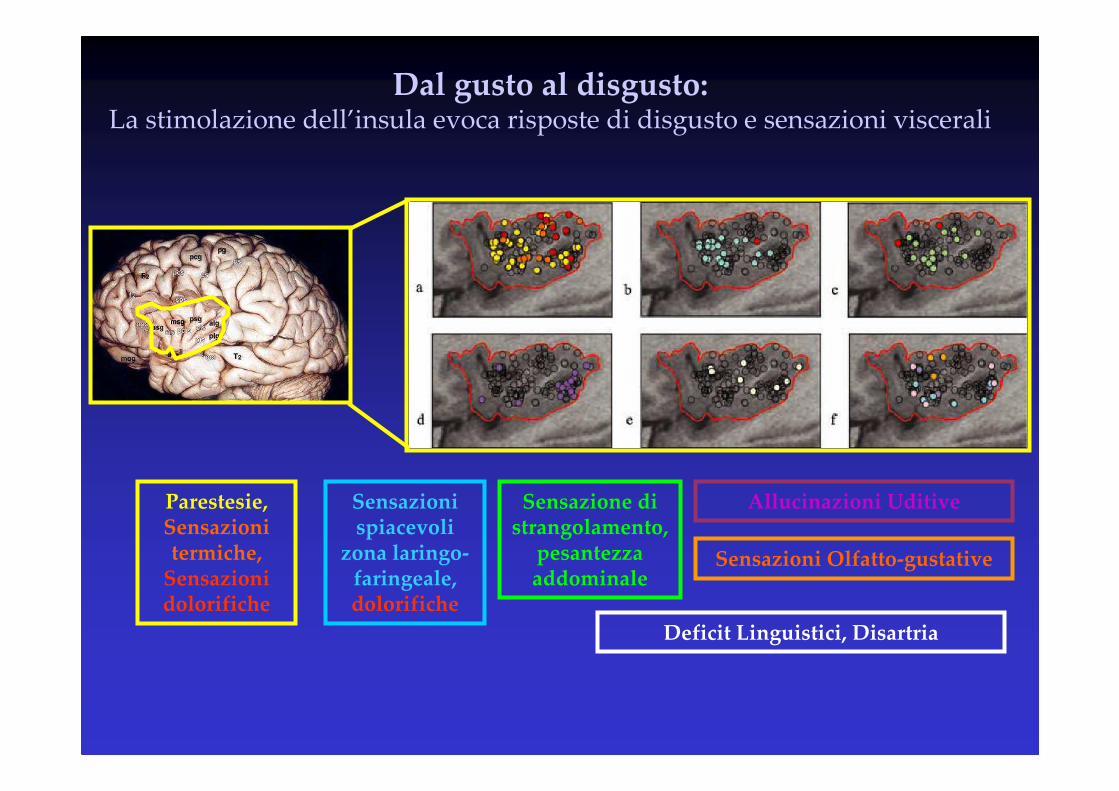
Dal gusto al disgusto:La stimolazione dell’insula evoca risposte di disgusto e sensazioni viscerali
Parestesie, Sensazioni termiche,
Sensazioni dolorifiche
Sensazioni spiacevoli
zona laringo-faringeale, dolorifiche
Sensazione di strangolamento,
pesantezza addominale
Allucinazioni Uditive
Deficit Linguistici, Disartria
Sensazioni Olfatto-gustative
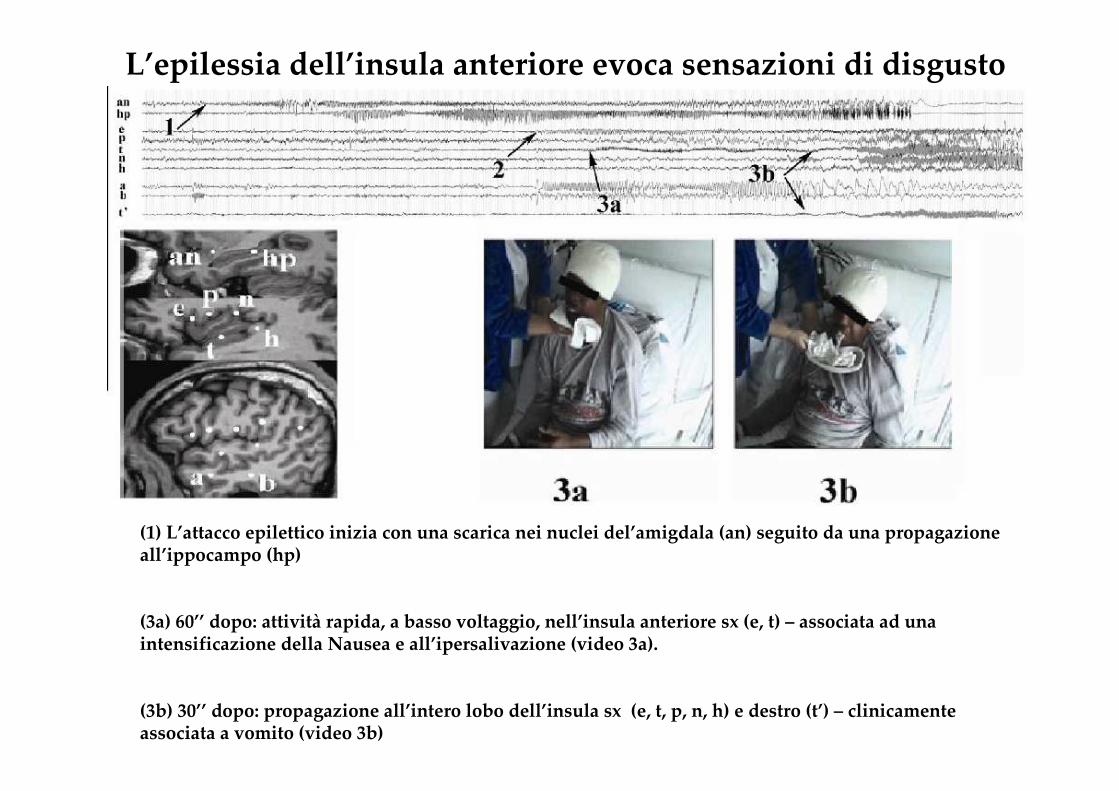
L’epilessia dell’insula anteriore evoca sensazioni di disgusto
(1) L’attacco epilettico inizia con una scarica nei nuclei del’amigdala (an) seguito da una propagazione all’ippocampo (hp)
(3a) 60’’ dopo: attività rapida, a basso voltaggio, nell’insula anteriore sx (e, t) – associata ad una intensificazione della Nausea e all’ipersalivazione (video 3a).
(3b) 30’’ dopo: propagazione all’intero lobo dell’insula sx (e, t, p, n, h) e destro (t’) – clinicamente associata a vomito (video 3b)
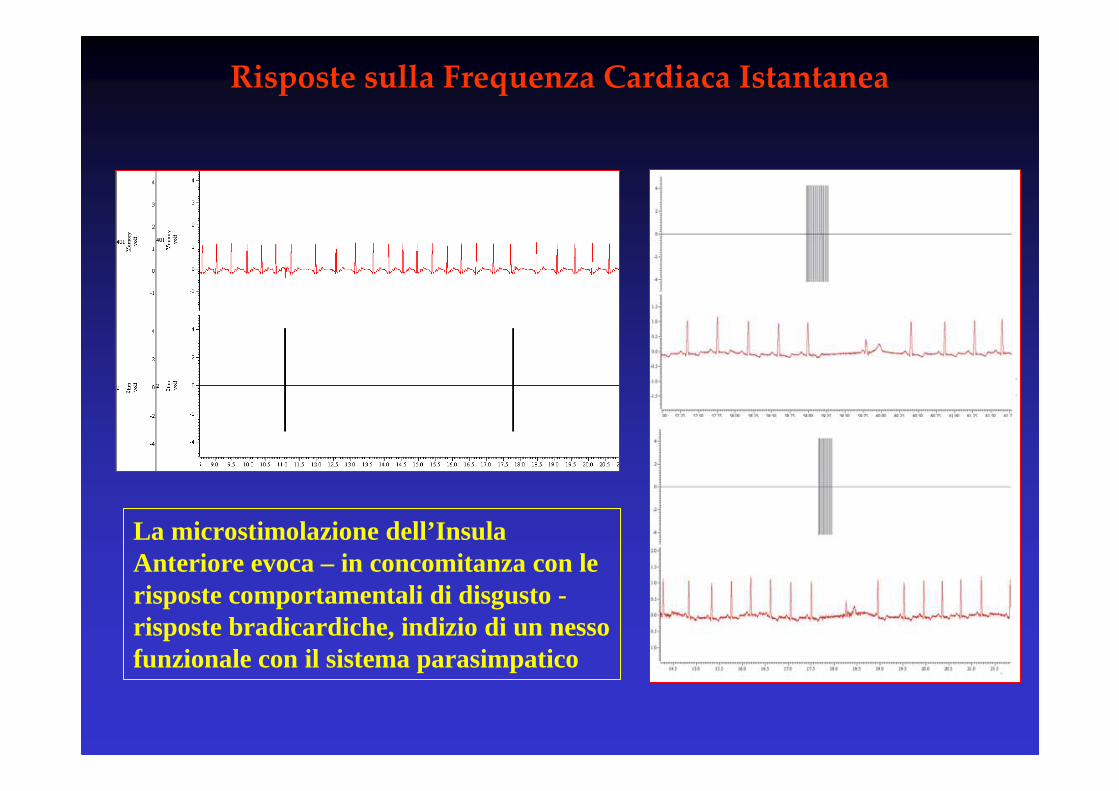
Risposte sulla Frequenza Cardiaca Istantanea
La microstimolazione dell’Insula Anteriore evoca – in concomitanza con le risposte comportamentali di disgusto -risposte bradicardiche, indizio di un nesso funzionale con il sistema parasimpatico

L’effetto sulla frequenza istantanea è visibile nella prima coppia di qRsdopo l’inizio della stimolazione
L’insula è dunque implicata in risposte non solo sensoriali (gustative) ma anche viscero-motorie
L’attivazione dell’insula viene percepita dal soggetto in termini di sensazione di disgusto, ma al contempo un pattern viscero-motorioviene attivato modulando i parametri vegetativi

L’insula sembra un candidato adatto per un ‘meccanismo mirror’ per il disgusto
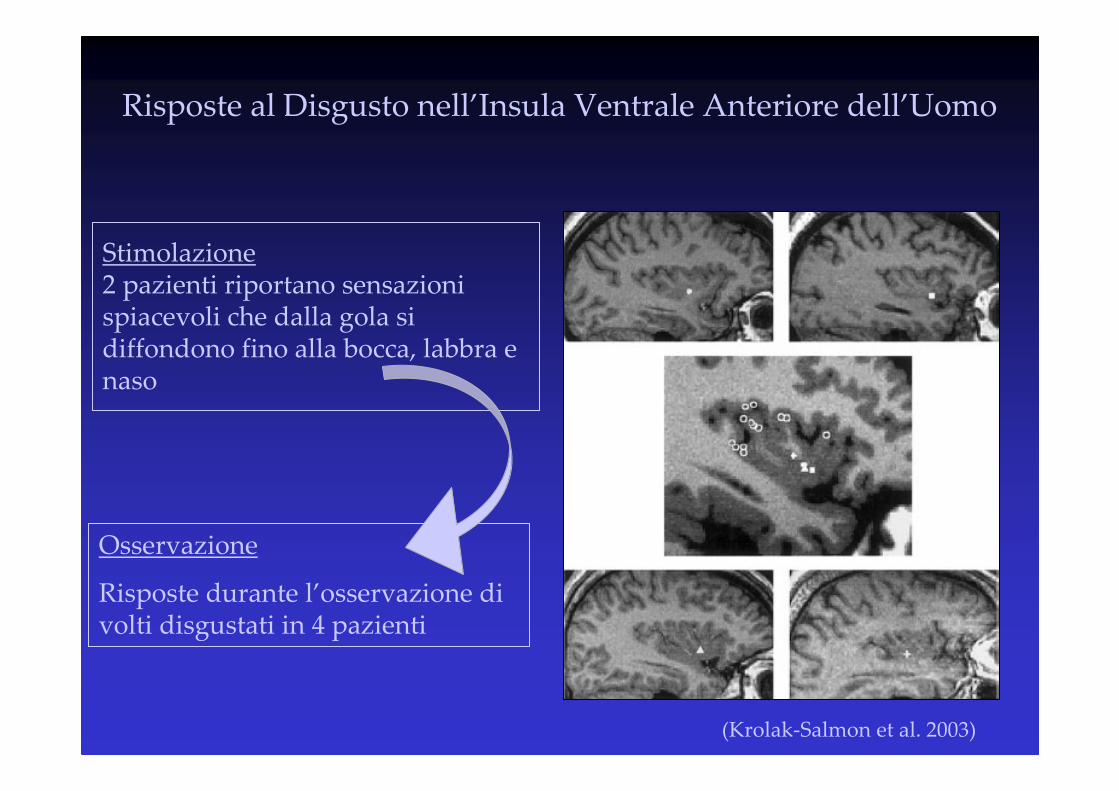
(Krolak-Salmon et al. 2003)
Risposte al Disgusto nell’Insula Ventrale Anteriore dell’Uomo
Stimolazione2 pazienti riportano sensazioni spiacevoli che dalla gola si diffondono fino alla bocca, labbra e naso
Osservazione
Risposte durante l’osservazione di volti disgustati in 4 pazienti
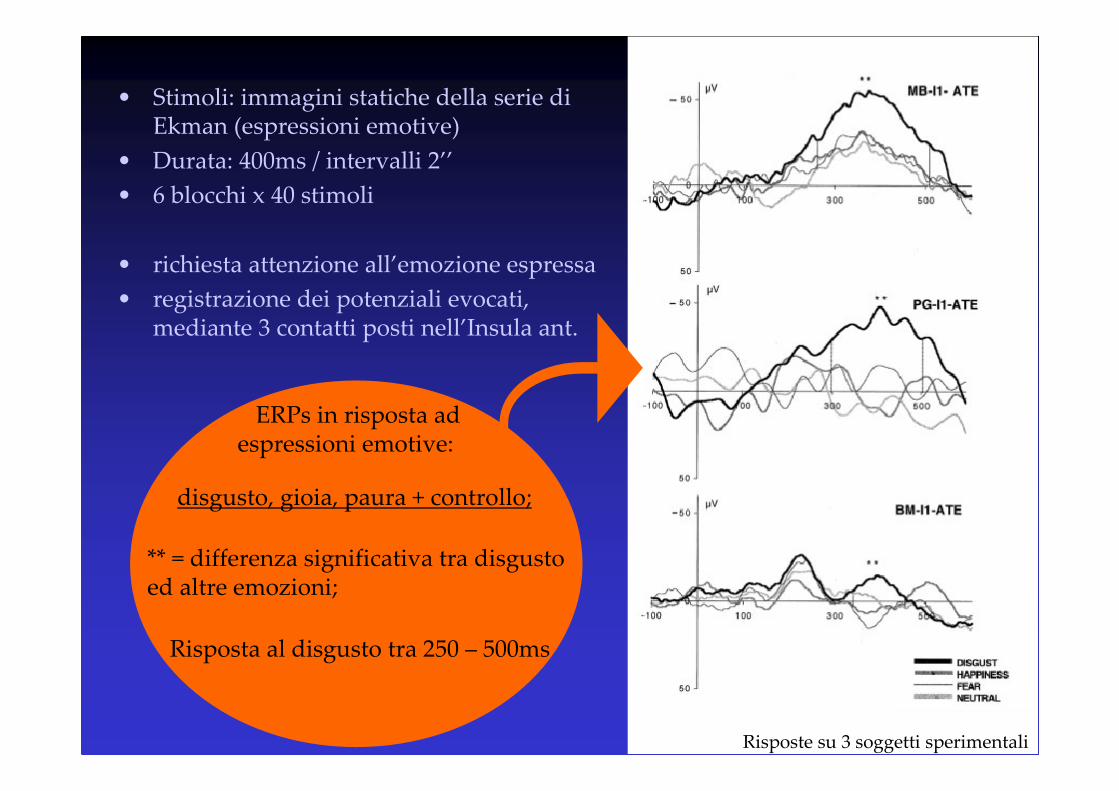
• Stimoli: immagini statiche della serie di Ekman (espressioni emotive)
• Durata: 400ms / intervalli 2’’
• 6 blocchi x 40 stimoli
• richiesta attenzione all’emozione espressa
• registrazione dei potenziali evocati, mediante 3 contatti posti nell’Insula ant.
ERPs in risposta ad espressioni emotive:
** = differenza significativa tra disgusto ed altre emozioni;
Risposta al disgusto tra 250 – 500ms
disgusto, gioia, paura + controllo;
Risposte su 3 soggetti sperimentali

DISGUSTO OLFATTIVO:“BOTH OF US DISGUSTED IN MY INSULA”
1. Un odore disgustoso o piacevole viene soffiato all’interno della maschera del soggetto nella fMRI
2. Scopo: mappare le aree corticali reclutate durante l’odore disgustoso
3. Visione di video in cui gli attori mostrano un’espressione di disgusto, di piacere, o neutra
(Wicker et al. 2003)
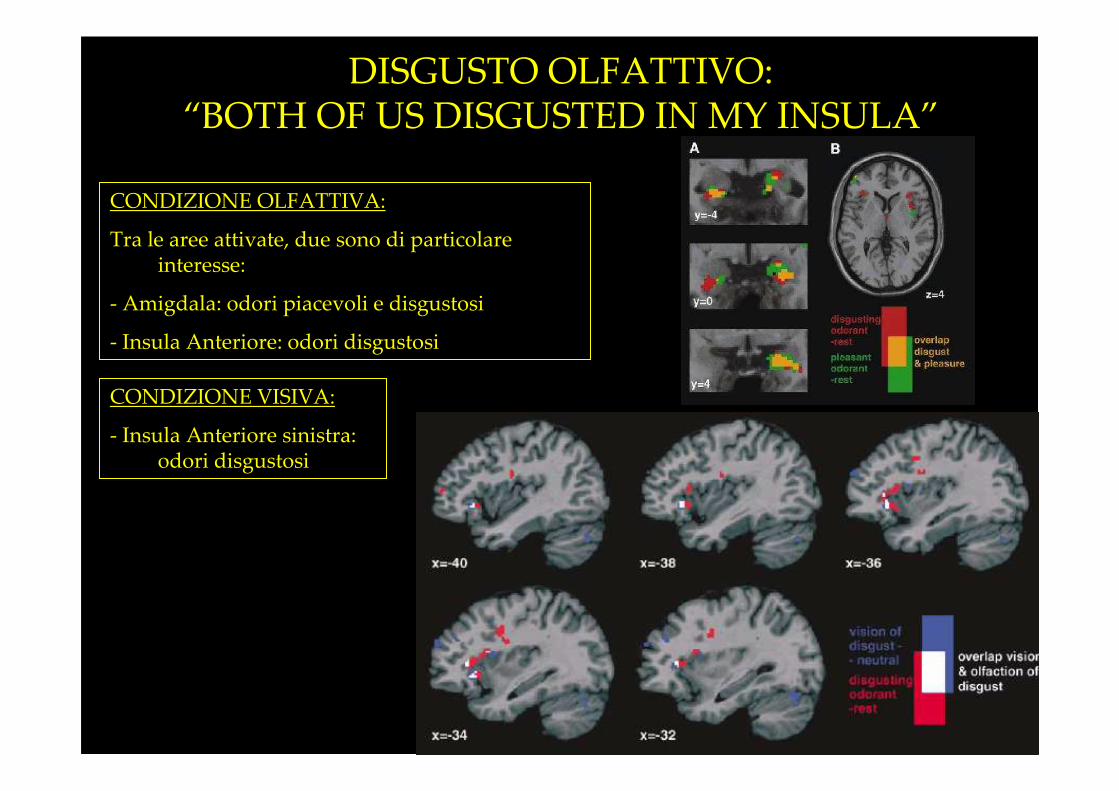
DISGUSTO OLFATTIVO:“BOTH OF US DISGUSTED IN MY INSULA”
CONDIZIONE OLFATTIVA:
Tra le aree attivate, due sono di particolare interesse:
- Amigdala: odori piacevoli e disgustosi
- Insula Anteriore: odori disgustosi
CONDIZIONE VISIVA:
- Insula Anteriore sinistra: odori disgustosi
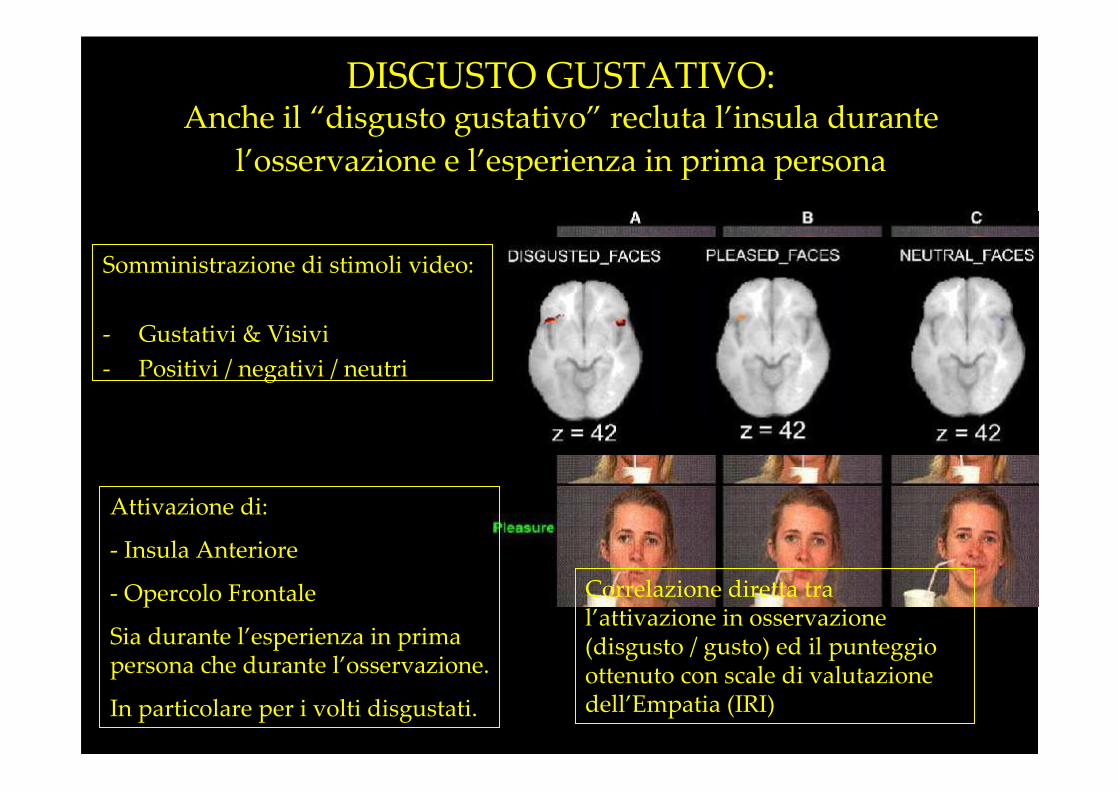
Somministrazione di stimoli video:
- Gustativi & Visivi
- Positivi / negativi / neutri
DISGUSTO GUSTATIVO:Anche il “disgusto gustativo” recluta l’insula durante
l’osservazione e l’esperienza in prima persona
Attivazione di:
- Insula Anteriore
- Opercolo Frontale
Sia durante l’esperienza in prima persona che durante l’osservazione.
In particolare per i volti disgustati.
Correlazione diretta tra l’attivazione in osservazione (disgusto / gusto) ed il punteggio ottenuto con scale di valutazione dell’Empatia (IRI)

Il Sistema Mirror per le azioni ha mostrato una parziale sovrapposizione con il substrato corticale del Motor Imagery.
Così come per il caso delle azioni, anche esperire / osservare / immaginare il disgusto mostra una parziale sovrapposizione corticale
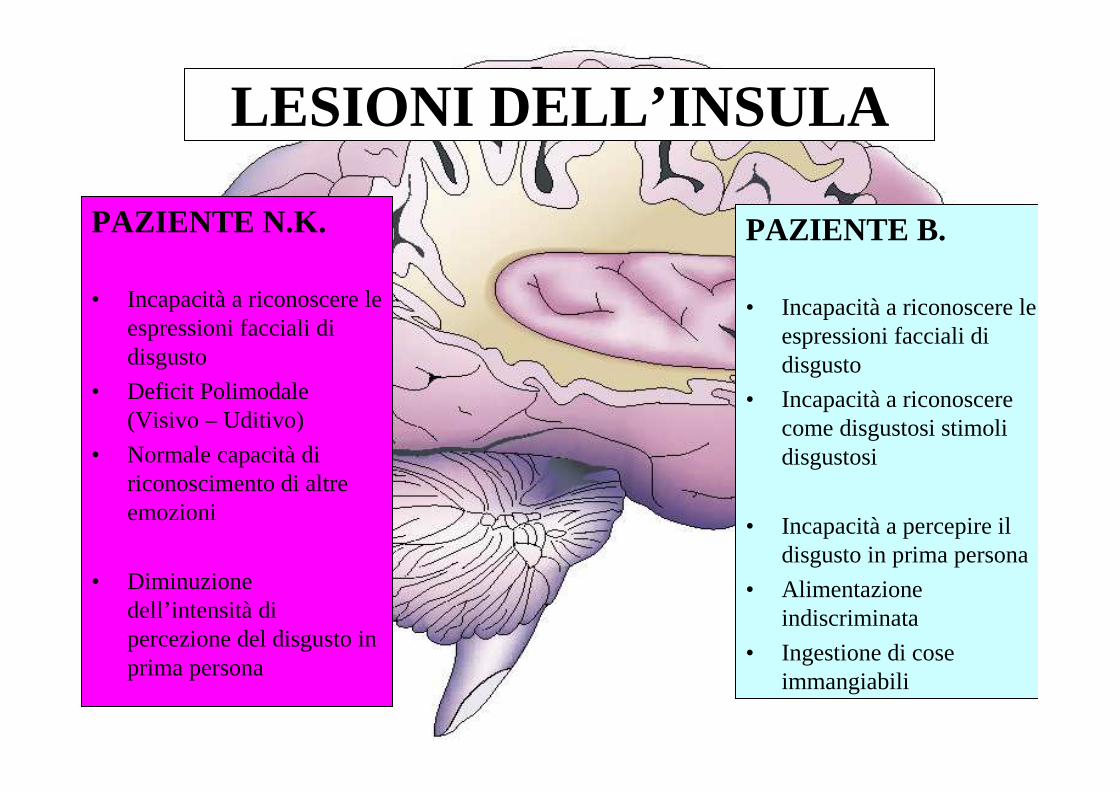
LESIONI DELL’INSULA
PAZIENTE N.K.
• Incapacità a riconoscere le espressioni facciali di disgusto
• Deficit Polimodale(Visivo – Uditivo)
• Normale capacità di riconoscimento di altre emozioni
• Diminuzione dell’intensità di percezione del disgusto in prima persona
PAZIENTE B.
• Incapacitàa riconoscere le espressioni facciali di disgusto
• Incapacità a riconoscere come disgustosi stimoli disgustosi
• Incapacità a percepire il disgusto in prima persona
• Alimentazione indiscriminata
• Ingestione di cose immangiabili

Le informazioni provenienti dalle aree visive che descrivono i volti arrivano all’insula
L’insula è la regione in cui sono rappresentati gli stati interni del corpo. Un centro di integrazione viscero-motoria la cui attivazione provoca la trasformazione degli input sensoriali in reazioni viscerali
“Questo meccanismo implica una simulazione interna che consiste nella rapida modificazione delle mappe dello stato corrente del corpo”
(Antonio Damasio)

IL DOLORE DEGLI ALTRI

La capacità di esperire il dolore degli altri è forse il tratto piùcaratteristico dell’Empatia
Il dolore degli altri può essere letto:
- dall’espressione facciale prodotta;
- dall’immagine di una parte del corpo colpita;
Entrambi i casi sono compatibili in linea di principio con l’eventualità di un meccanismo mirror per il riconoscimento del dolore altrui

Empathy for Pain Involves the Affective but not Sensory Components of Pain
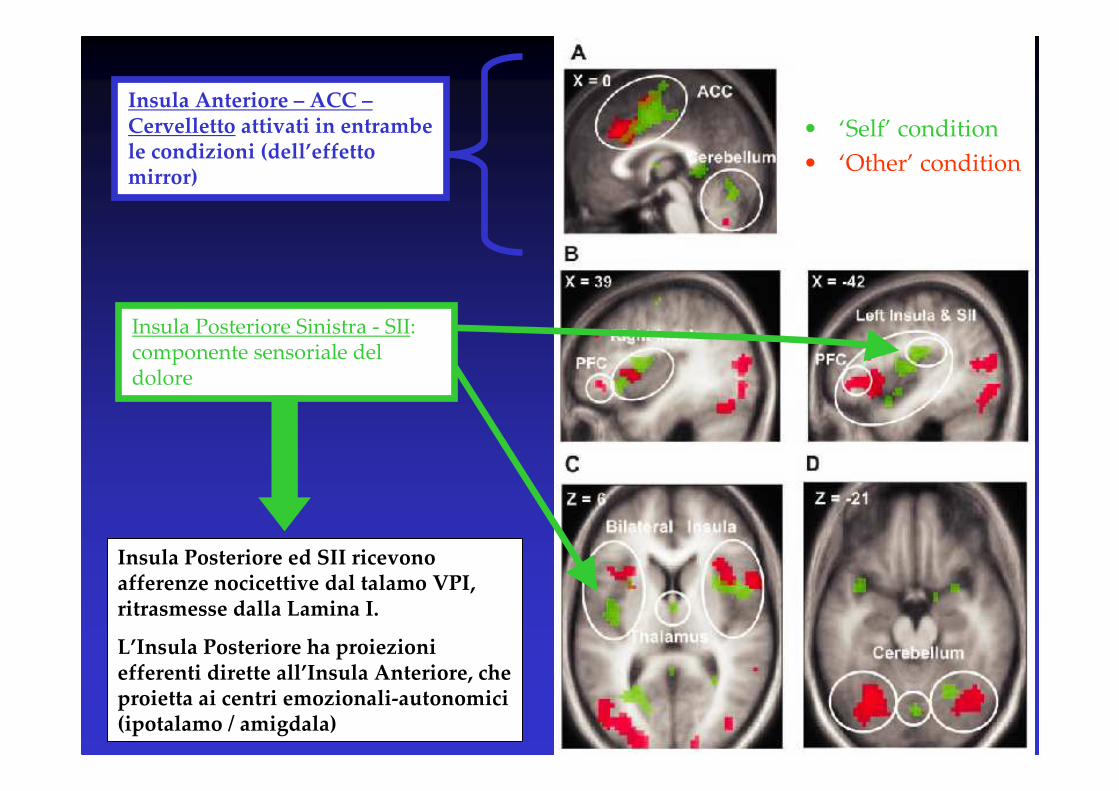
• ‘Self’ condition
• ‘Other’ condition
Insula Posteriore Sinistra - SII: componente sensoriale del dolore
Insula Anteriore – ACC –Cervelletto attivati in entrambe le condizioni (dell’effetto mirror)
Insula Posteriore ed SII ricevono afferenze nocicettive dal talamo VPI, ritrasmesse dalla Lamina I.
L’Insula Posteriore ha proiezioni efferenti dirette all’Insula Anteriore, che proietta ai centri emozionali-autonomici(ipotalamo / amigdala)

Risultati principali
• L’osservazione del dolore altrui recluta circuiti normalmente reclutati durante l’esperienza del dolore (‘meccanismo mirror’)
• Non tutta la matrice neurale implicata nel senso del dolore viene attivata (non viene reclutata la parte “sensoriale”)
• Le differenze nell’attivazione neurale co-variano con le differenze di punteggio ottenuto ai test di valutazione dell’empatia (in ACC e AI sinistra)

Cosa succede quando soffre il cattivo?

• I soggetti giocano ad un gioco economico in cui gli altri giocatori si comportano in maniera
(a) leale
(b) sleale
• In una seconda fase, il soggetto nello scanner assiste al rilascio di stimolazioni dolorose a:
(a) Se stessi
(b) Giocatore leale
(c) Giocatore sleale

Risultati principali
• Esistono differenze significative tra uomo / donna (attivazione di ACC significativa nelle donne, e non significativa negli uomini) – con il giocatore sleale
• L’osservazione del dolore del giocatore leale recluta circuiti normalmente reclutati durante l’esperienza del dolore (‘meccanismo mirror’), sia negli uomini che nelle donne.
• L’osservazione del dolore del giocatore sleale, nell’uomo, recluta in maniera significativamente minore l’Insula Anteriore
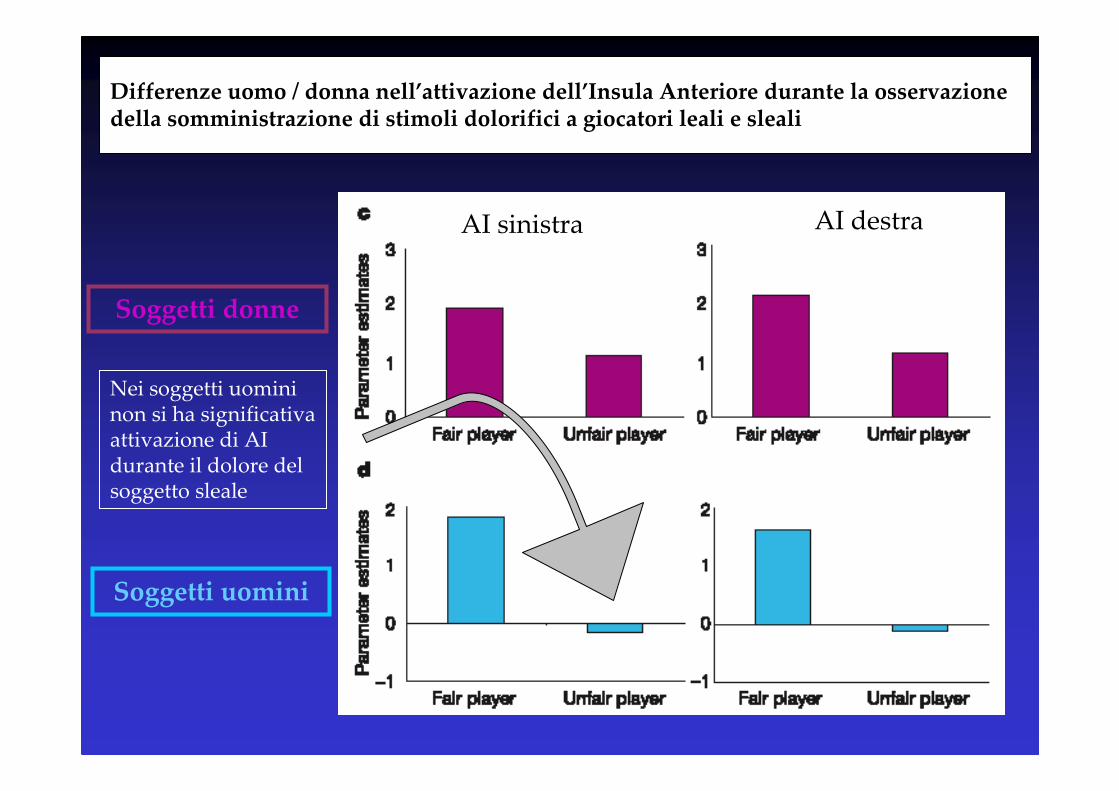
Differenze uomo / donna nell’attivazione dell’Insula Anteriore durante la osservazione della somministrazione di stimoli dolorifici a giocatori leali e sleali
AI sinistra AI destra
Soggetti donne
Soggetti uomini
Nei soggetti uomini non si ha significativa attivazione di AI durante il dolore del soggetto sleale
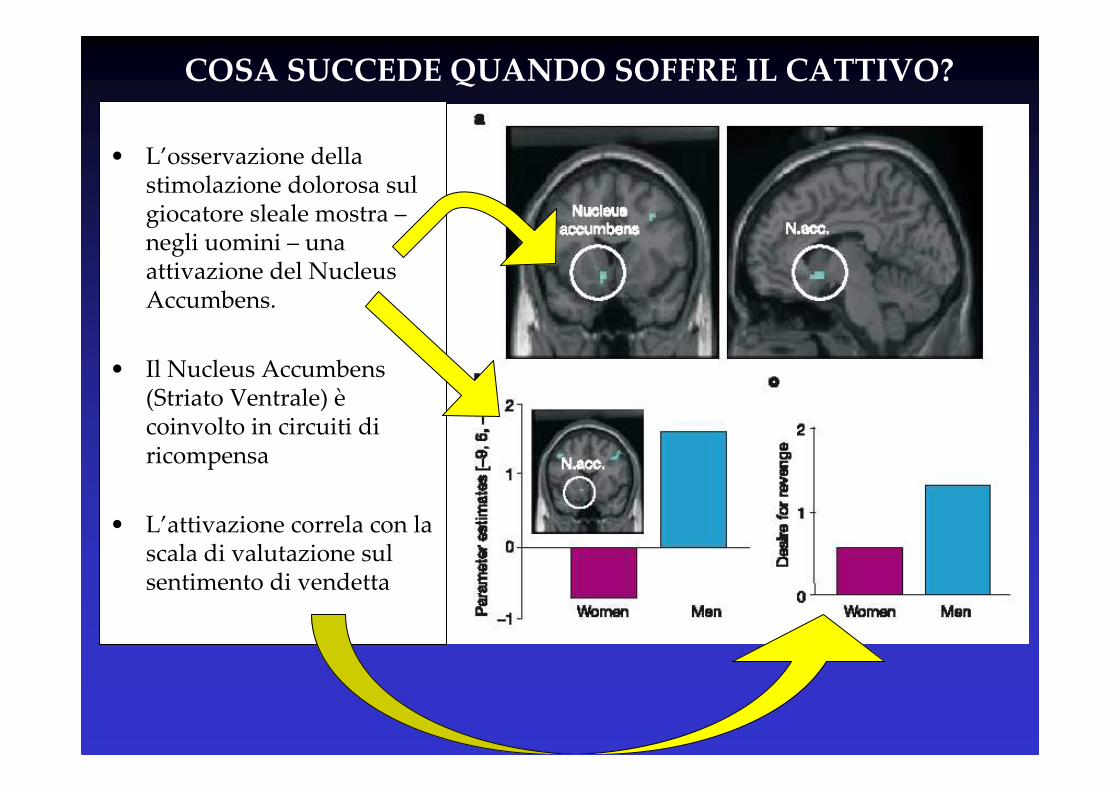
• L’osservazione della stimolazione dolorosa sul giocatore sleale mostra –negli uomini – una attivazione del NucleusAccumbens.
• Il Nucleus Accumbens(Striato Ventrale) ècoinvolto in circuiti di ricompensa
• L’attivazione correla con la scala di valutazione sul sentimento di vendetta
COSA SUCCEDE QUANDO SOFFRE IL CATTIVO?
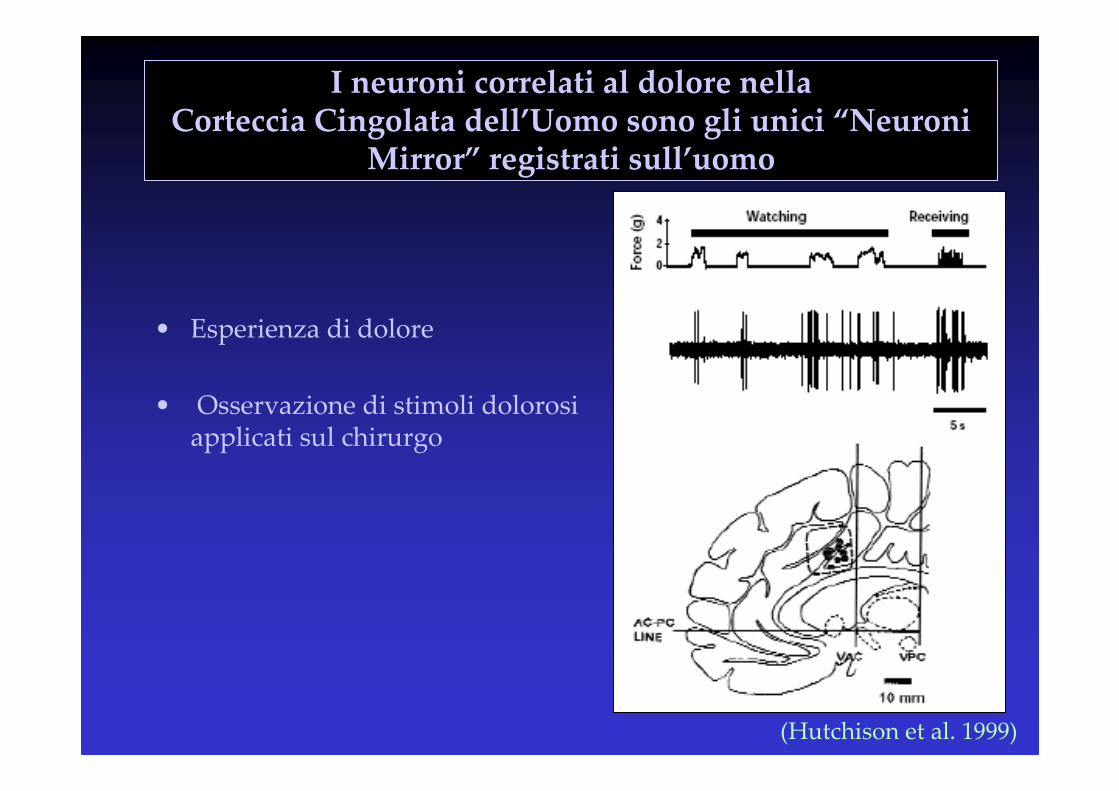
I neuroni correlati al dolore nella Corteccia Cingolata dell’Uomo sono gli unici “Neuroni
Mirror” registrati sull’uomo
• Esperienza di dolore
• Osservazione di stimoli dolorosi applicati sul chirurgo
(Hutchison et al. 1999)

LE SENSAZIONI TATTILI E LE
AREE SOMATICHE PARIETALI

ANCHE LE SENSAZIONI PROVATE DAGLI ALTRI VENGONO COMPRESE MEDIANTE UN ‘MECCANISMO MIRROR’?
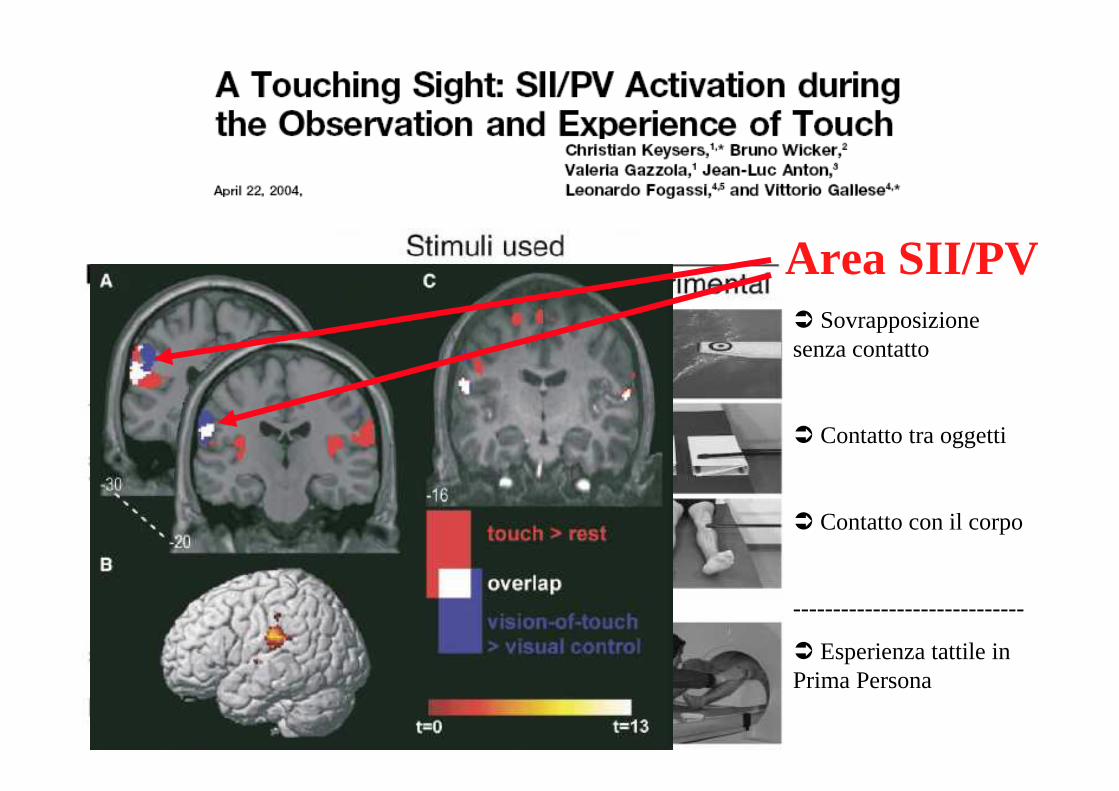
� Sovrapposizione senza contatto
� Contatto tra oggetti
� Contatto con il corpo
-----------------------------
� Esperienza tattile in Prima Persona
Area SII/PV
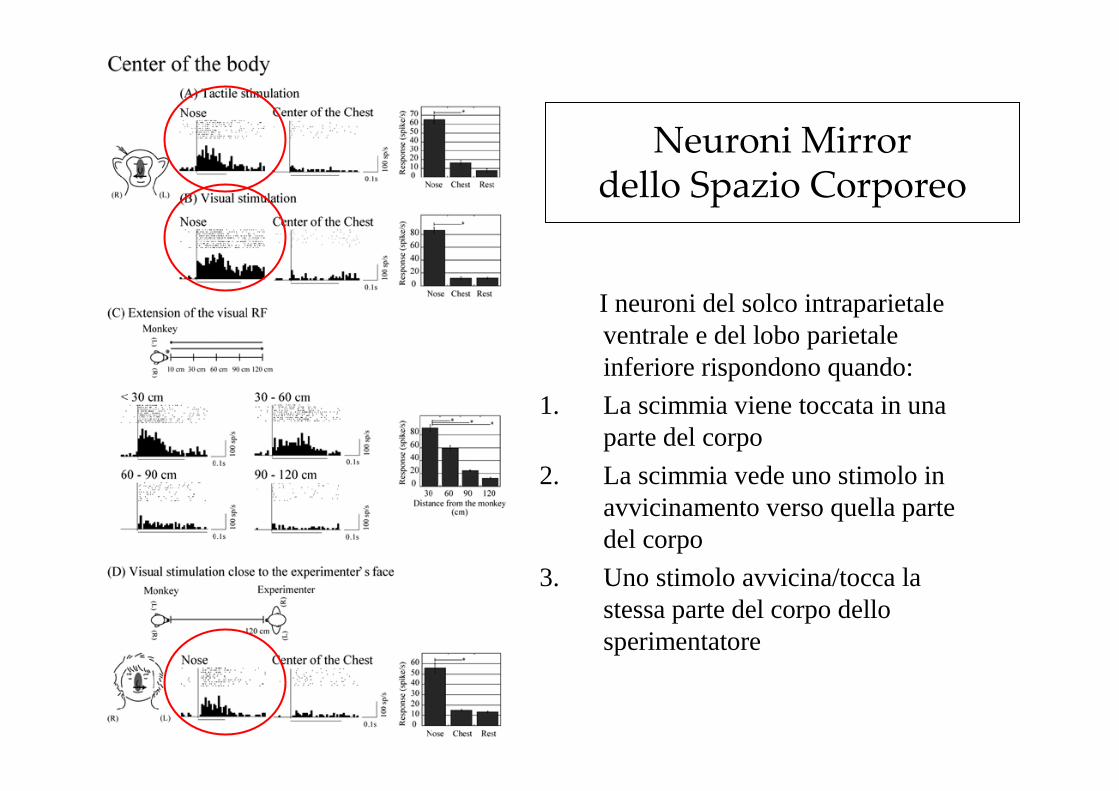
Neuroni Mirror dello Spazio Corporeo
I neuroni del solco intraparietale ventrale e del lobo parietale inferiore rispondono quando:
1. La scimmia viene toccata in una parte del corpo
2. La scimmia vede uno stimolo in avvicinamento verso quella parte del corpo
3. Uno stimolo avvicina/tocca la stessa parte del corpo dello sperimentatore
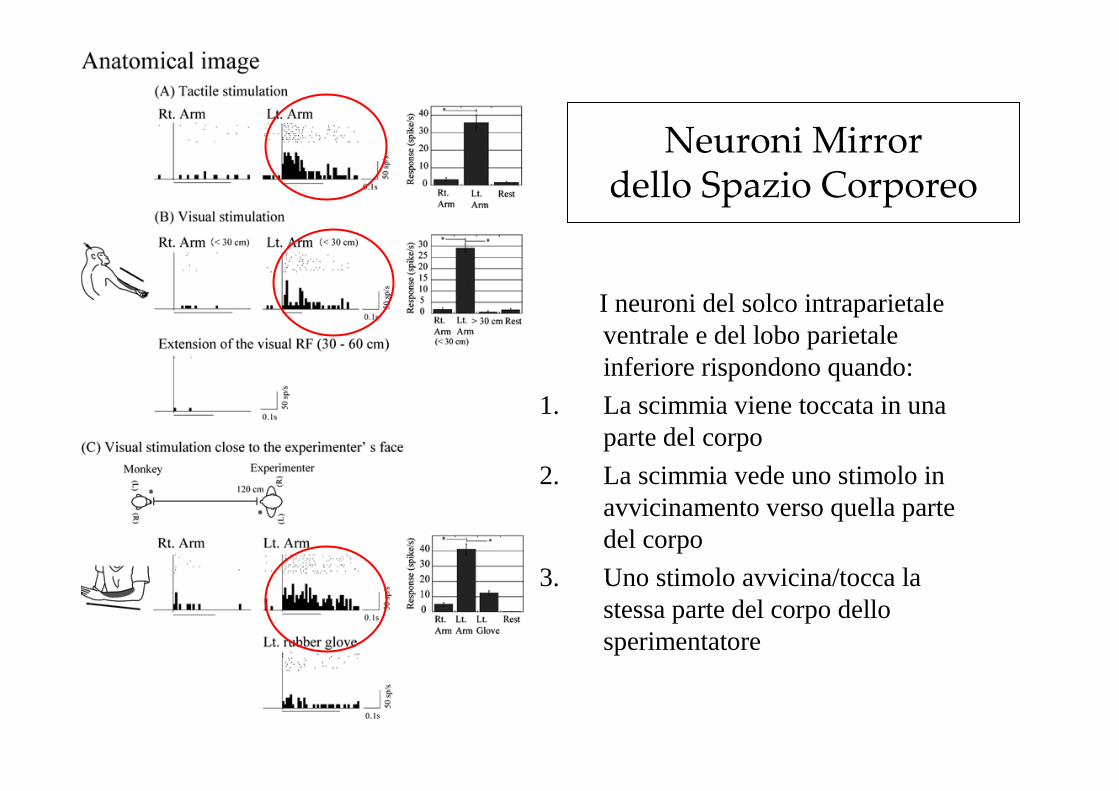
Neuroni Mirror dello Spazio Corporeo
I neuroni del solco intraparietale ventrale e del lobo parietale inferiore rispondono quando:
1. La scimmia viene toccata in una parte del corpo
2. La scimmia vede uno stimolo in avvicinamento verso quella parte del corpo
3. Uno stimolo avvicina/tocca la stessa parte del corpo dello sperimentatore

QUANTI SISTEMI MIRROR?
• Azioni • Intenzioni• Linguaggio• Disgusto• Dolore • Sensazione• […]

FINE





























