Embed Size (px)
Citation preview

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƯỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN
---------------------
Trần Thúy Hằng
PHÂN LẬP, NGHIÊN CỨU ĐẶC ĐIỂM SINH HỌC CỦA
MỘT SỐ CHỦNG VI SINH VẬT CÓ KHẢ NĂNG TẠO
MÀNG SINH VẬT (BIOFILM) Ở VIỆT NAM
LUẬN VĂN THẠC SĨ KHOA HỌC
Hà Nội – 2011

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƯỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN
---------------------
Trần Thúy Hằng
PHÂN LẬP, NGHIÊN CỨU ĐẶC ĐIỂM SINH HỌC CỦA
MỘT SỐ CHỦNG VI SINH VẬT CÓ KHẢ NĂNG TẠO
MÀNG SINH VẬT (BIOFILM) PHÂN LẬP Ở VIỆT NAM
Chuyên ngành: Sinh học thực nghiệm
Mã số: 60 42 30
LUẬN VĂN THẠC SĨ KHOA HỌC
NGƯỜI HƯỚNG DẪN KHOA HỌC:
TS. NGUYỄN QUANG HUY
Hà Nội - 2011

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN i
LỜI CẢM ƠN
Lời đầu tiên, tôi xin gửi lời cảm ơn chân thành và sâu sắc nhất tới TS.
Nguyễn Quang Huy, người thầy mẫu mực đã tận tụy hướng dẫn và truyền đạt kiến
thức cho tôi trong suốt quá trình thực hiện đề tài cũng như luôn quan tâm, động viên
và giúp đỡ chúng tôi trong cuộc sống.
Tôi xin gửi lời cảm ơn chân thành tới các thầy cô trong Khoa Sinh học,
Trường Đại học Khoa học Tự nhiên, Đại học Quốc gia Hà Nội đã dành mọi tâm
huyết giảng dạy, trang bị kiến thức cho chúng tôi trong suốt khóa học này.
Tôi cũng xin gửi lời cảm ơn đến tập thể các anh, chị, em trong Phòng
Enzyme học và Phân tích hoạt tính sinh học, Phòng thí nghiệm trọng điểm Công
nghệ Protein và Enzyme; Bộ môn Sinh lý thực vật và Hóa sinh, Bộ môn Vi sinh vật
học đã nhiệt tình cộng tác và giúp đỡ tôi trong suốt quá trình thực hiện đề tài này.
Cuối cùng, tôi xin được gửi lời cảm ơn đến gia đình, bạn bè đặc biệt là chồng
tôi đã luôn ở bên động viên, chia sẻ khó khăn, giúp đỡ tôi trong quá trình học tập và
thực hiện luận văn này.
Hà Nội, ngày 20 tháng 11 năm 2011
Học viên cao học
Trần Thúy Hằng

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN ii
MỤC LỤC
Trang
MỞ ĐẦU ................................................................................................................1
Chương 1. TỔNG QUAN TÀI LIỆU.......................................................................2
1.1 Khái niệm về màng sinh vật (biofilm)................................................................2
1.2 Các dạng màng sinh vật trong tự nhiên và vai trò đối với vi sinh vật..................5
1.2.1 Các chủng vi sinh vật có khả năng tạo màng sinh vật trong tự nhiên..............5
1.2.2 Các dạng tồn tại của màng sinh vật .................................................................7
1.2.3 Ảnh hưởng của màng sinh vật đối với vi sinh vật............................................8
1.3 Thành phần, cấu trúc và đặc điểm của màng sinh vật .......................................11
1.3.1 Mạng lưới ngoại bào.....................................................................................11
1.3.2 Các thành phần khác .....................................................................................13
1.4 Các yếu tố ảnh hưởng đến sự hình thành và phát triển của màng sinh vật ........15
1.4.1 Các giai đoạn tạo thành màng sinh vật ..........................................................15
1.4.2 Các yếu tố ảnh hưởng đến sự hình thành màng sinh vật ................................20
1.4.3 Điều hòa quá trình hình thành màng sinh vật ................................................22
1.5 Nghiên cứu ứng dụng màng sinh vật................................................................24
1.5.1 Ứng dụng màng sinh vật trong việc xử lý nước thải ......................................24
1.5.2 Ứng dụng màng sinh vật trong việc ức chế các vi sinh vật gây hại ................25
1.5.3 Một số nghiên cứu ứng dụng khác ................................................................26
Chương 2. NGUYÊN LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU.......................29
2.1 Nguyên liệu .....................................................................................................29
2.1.1 Chủng vi sinh vật nghiên cứu........................................................................29
2.1.2 Vi sinh vật kiểm định....................................................................................29
2.2 Hóa chất, thiết bị và dụng cụ thí nghiệm ..........................................................30
2.2.1 Môi trường nuôi cấy .....................................................................................30
2.2.2 Máy móc, thiết bị..........................................................................................31
2.3 Phương pháp nghiên cứu .................................................................................31
2.3.1 Phương pháp phân lập vi khuẩn ....................................................................31

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN iii
2.3.2 Phương pháp bảo quản giống vi sinh vật.......................................................32
2.3.3 Phương pháp nghiên cứu đánh giá khả năng tạo màng sinh vật của các chủng
vi sinh vật..............................................................................................................32
2.3.4 Tối ưu hóa các điều kiện tạo màng sinh vật...................................................33
2.3.5 Phương pháp đánh giá khả năng tạo chất hoạt động bề mặt...........................35
2.3.6 Phương pháp đánh giá khả năng kháng khuẩn...............................................35
2.3.7 Phương pháp nhuộm Gram ...........................................................................36
2.3.8 Quan sát cấu trúc màng sinh vật bằng ảnh chụp trên kính hiển vi điện tử quét
..............................................................................................................................37
2.3.9 Phương pháp phân loại phân tử dựa trên gen 16S rDNA...............................37
Chương 3. KẾT QUẢ VÀ THẢO LUẬN..............................................................39
3.1 Phân lập, tuyển chọn các chủng vi sinh vật có khả năng tạo màng sinh vật ......39
3.1.1 Phân lập vi sinh vật.......................................................................................39
3.1.2 Khả năng phát triển và tạo màng sinh vật của các chủng phân lập.................40
3.2 Các yếu tố ảnh hưởng đến sự tạo màng sinh vật của các chủng phân lập..........42
3.2.1 Ảnh hưởng của nhiệt độ................................................................................42
3.2.2 Ảnh hưởng của pH môi trường .....................................................................43
3.2.3 Ảnh hưởng của nguồn cacbon .......................................................................44
3.2.4 Ảnh hưởng của nguồn nitơ............................................................................46
3.2.5 Ảnh hưởng của giá thể ..................................................................................47
3.3 Một số đặc tính sinh học và phân loại các chủng vi sinh vật phân lập ..............50
3.3.1 Khả năng tạo chất hoạt động bề mặt.............................................................50
3.3.3 Đặc điểm hình thái........................................................................................53
3.3.4 Phân loại các chủng vi sinh vật phân lập dựa trên gen 16S rDNA .................55
KẾT LUẬN VÀ KIẾN NGHỊ ...............................................................................59
TÀI LIỆU THAM KHẢO .....................................................................................60
PHỤ LỤC..............................................................................................................69

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN iv
DANH MỤC BẢNG
Bảng 1: Ảnh hưởng của pH môi trường đến sự tạo màng sinh vật của các chủng vi
sinh vật phân lập....................................................................................................44
Bảng 2: Ảnh hưởng của nguồn cacbon đến sự tạo màng sinh vật của các chủng vi
sinh vật phân lập....................................................................................................45
Bảng 3: Ảnh hưởng của các nguồn nitơ đến sự tạo màng sinh vật của các chủng vi
sinh vật phân lập....................................................................................................47
Bảng 4: Hoạt tính kháng khuẩn của các chủng vi khuẩn nghiên cứu ......................52

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN v
DANH MỤC HÌNH
Hình 1. Các vi sinh vật gắn kết với bề mặt nhựa nhân tạo và liên kết với nhau thông qua mạng lưới polysaccarit dưới kính hiển vi điện tử ..............................................3 Hình 2. Ảnh SEM một màng sinh vật do Staphylococcus aureus tạo nên.................4 Hình 3. Ảnh SEM màng sinh vật nổi được hình thành bởi chủng Bacillus subtilis B-1 ..........................................................................................................................5 Hình 4. Một số ví dụ về màng sinh vật ....................................................................8 Hình 5. Mô hình phát triển của màng sinh vật .......................................................16 Hình 6. Khuẩn lạc một số chủng vi sinh vật phân lập trên môi trường thạch ..........39 Hình 7. Khả năng tạo màng sinh vật của một số chủng vi sinh vật phân lập từ mẫu nước thải làng miến Lại Trạch ...............................................................................40 Hình 8. Khả năng tạo màng sinh vật của một số chủng vi sinh vật phân lập từ mẫu nước thải nhà máy sản xuất bia..............................................................................41 Hình 9. Khả năng tạo màng sinh vật của một số chủng vi sinh vật phân lập từ mẫu nước thải làng nghề bún Phú Đô............................................................................42 Hình 10. Ảnh hưởng của nhiệt độ đến khả năng tạo màng sinh vật từ các chủng phân lập.................................................................................................................43 Hình 11. Màng nổi của 4 chủng vi sinh vật sau 5 ngày nuôi cấy ............................48 Hình 12. Màng sinh vật trên bề mặt nhựa ..............................................................48 Hình 13. Cấu trúc màng sinh vật của các chủng phân lập ......................................49 Hình 14. Khả năng nhũ tương hóa dầu ăn của các chủng vi sinh vật ......................51 Hình 15. Hình thái khuẩn lạc và hình dạng vi khuẩn dưới kính hiển vi quang học .54 Hình 16. Hình thái tế bào các chủng vi khuẩn trong màng sinh vật ........................55 Hình 17. Vị trí phân loại của các chủng M3.8, M4.9 với các loài có quan hệ họ hàng dựa vào trình tự gen 16S rDNA .............................................................................56 Hình 18. Vị trí phân loại của chủng U1.3 và U3.7 với các loài có quan hệ họ hàng dựa vào trình tự gen 16S rDNA .............................................................................57

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN vi
BẢNG CHỮ VIẾT TẮT
Acyl - HSL Acyl - homoserine lactone
COD Chemical Oxygen Demand (nhu cầu oxy hóa học)
dd NTP Dideoxynucleotide
đvC đơn vị Cacbon
E24 Emulsion index (chỉ số nhũ tương hóa)
OD Optical Density (Mật độ quang học)
SDS Sodium dodecyl sulfate
SEM Scanning electron microscopy
TCA Tricarboxylic acid
TOC Total Organic Carbon
w/v Khối lượng (g)/thể tích (ml)

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 1
MỞ ĐẦU
Nghiên cứu vi sinh vật học là mô hình nghiên cứu ưu việt để tìm hiểu bản
chất của các quá trình sống, đồng thời nó cũng đóng vai trò quan trọng trong các
lĩnh vực y học, nông nghiệp, công nghiệp và môi trường.
Tuy nhiên, trong tự nhiên vi sinh vật ít khi tồn tại dưới dạng các tế bào đơn
lẻ mà chúng thường được tìm thấy dưới dạng tập hợp các tế bào liên kết chặt chẽ
với nhau và với các bề mặt thông qua mạng lưới chất ngoại bào gọi là màng sinh vật
(biofilm).
Nghiên cứu về màng sinh vật giúp chúng ta có một cái nhìn tổng quát hơn về
sự tăng trưởng, phát triển và thích nghi của vi sinh vật trong mối quan hệ với nhau
cũng như với các điều kiện môi trường. Đồng thời, việc tìm hiểu về màng sinh vật
cũng giúp chúng ta có những hiểu biết sâu hơn về mối liên hệ bên trong của các tế
bào trong một màng sinh vật cũng như các cơ chế điều hòa quá trình tạo màng sinh
vật.
Nghiên cứu về màng sinh vật góp phần tạo ra những sản phẩm ứng dụng cao
trong cuộc sống như tạo các công nghệ sinh học xử lý ô nhiễm môi trường, xử lý
các sự cố tràn dầu, ứng dụng trong nghiên cứu phòng bệnh cho cây trồng cũng như
các nghiên cứu trong công nghiệp thực phẩm, hóa mỹ phẩm v.v… mà hiện nay tại
Việt Nam các nghiên cứu về vi sinh vật tạo màng sinh vật và ứng dụng của chúng
còn rất mới mẻ.
Chính từ những ý nghĩa thực tiễn trên, chúng tôi quyết định thực hiện đề tài:
“Phân lập, nghiên cứu đặc điểm sinh học của một số chủng vi sinh vật có khả
năng tạo màng sinh vật phân lập ở Việt Nam”.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 2
Chương 1. TỔNG QUAN TÀI LIỆU
1.1 Khái niệm về màng sinh vật (biofilm)
Vi sinh vật là tên gọi chung để chỉ tất cả các sinh vật có hình thể bé nhỏ,
muốn thấy rõ được người ta phải sử dụng tới kính hiển vi. Vi sinh vật không phải là
một nhóm riêng biệt trong sinh giới. Chúng thậm chí thuộc về nhiều giới sinh vật
khác nhau và giữa các nhóm có thể không có quan hệ mật thiết với nhau [1].
Lịch sử nghiên cứu và phát triển của vi sinh vật học đã ghi nhận người có
công phát hiện ra thế giới vi sinh vật và cũng là người đầu tiên mô tả hình thái nhiều
loại vi sinh vật - Antonie van Leeuwenhoek (1632 - 1723). Với việc tự chế tạo ra
trên 400 chiếc kính hiển vi, ông đã lần lượt quan sát mọi thứ có xung quanh mình,
trong đó có cả các vi khuẩn và động vật nguyên sinh mà ông đã gọi là những “động
vật vô cùng nhỏ bé”. Ông là người đầu tiên phát hiện ra hiện tượng bám dính và
phát triển phổ biến của vi khuẩn trên bề mặt răng tạo thành các mảng bám răng, một
dạng của màng sinh vật sau này [1].
Cùng với sự ra đời của kính hiển vi quang học hoàn chỉnh vào đầu thế kỷ 19,
đặc biệt là việc chế tạo thành công chiếc kính hiển vi điện tử đầu tiên (1934) đã góp
phần tạo nên những cống hiến lớn lao của các nhà khoa học trong lĩnh vực vi sinh
vật học [1]. Năm 1936, Zobell và cộng sự [87] đã nghiên cứu cho thấy số lượng vi
khuẩn bám dính tại vị trí tiếp xúc giữa nước biển và bề mặt vật rắn lớn hơn nhiều so
với các vị trí xung quanh. Lợi ích của bề mặt chất rắn mang lại được đánh giá như
một nơi cư trú của các vi khuẩn giúp tập trung hấp thụ chất dinh dưỡng, tăng cường
hoạt động của các enzyme và hấp thụ các chất chuyển hóa. Trong những nghiên cứu
về hiệu quả của bề mặt rắn đối với hoạt tính của vi khuẩn, Zobell [86] đã chỉ ra rằng
bên cạnh việc cung cấp nơi khu trú và tập trung chất dinh dưỡng, bề mặt chất rắn
còn làm chậm sự khuếch tán của các enzyme ngoại bào, thúc đẩy sự đồng hóa các
chất dinh dưỡng thông qua quá trình thủy phân trước khi chúng được hấp thụ.
Một loạt những nghiên cứu tiếp theo đó cũng đề cập đến khả năng hoạt động
và tăng trưởng đáng kể của vi sinh vật bằng cách bám dính vào một bề mặt xác

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 3
định. Nghiên cứu của Heukelekian và cộng sự [39] cho thấy giới hạn nồng độ chất
dinh dưỡng không cố định mà phụ thuộc vào tổng diện tích bề mặt tiếp xúc với môi
trường.
Jones và cộng sự [43] đã sử dụng kính hiển vi điện tử quét (SEM) để chỉ ra
sự xuất hiện của màng sinh vật trong bộ lọc nhỏ giọt của một nhà máy xử lý nước
thải và cho thấy rằng chúng bao gồm nhiều loại, nhóm các vi sinh vật khác nhau
(dựa trên đặc điểm hình thái tế bào). Bằng cách sử dụng chất nhuộm màu
polysaccarit đặc biệt là đỏ Ruthenimum và cố định bởi Osmium tetroxide (OsO4),
các nhà nghiên cứu đã chứng minh rằng vật liệu chất nền bao xung quanh và kết
dính các tế bào trong cấu trúc màng sinh vật chính là polysaccarit.
Hình 1. Các vi sinh vật gắn kết với bề mặt nhựa nhân tạo và liên kết với nhau thông qua
mạng lưới polysaccarit dưới kính hiển vi điện tử ( 57700) [43].
Dựa trên những quan sát ở mảng bám răng và một số thí nghiệm khác, năm
1978, Costerton và cộng sự [14] đã đưa ra giả thuyết về màng sinh học: “Trong tự
nhiên, các tế bào vi khuẩn gắn kết với nhau và bám dính trên một bề mặt nhất định
nhờ hệ thống sợi glycocalyx”. Giả thuyết này đã góp phần giải thích cơ chế bám
dính của vi sinh vật trên các vật liệu vô sinh và hữu sinh và những lợi ích thu được
nhờ phương thức sinh thái thích hợp này.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 4
Cùng với những tiến bộ trong quá trình nghiên cứu, khái niệm về màng sinh
vật được đưa ra ngày càng hoàn thiện và đầy đủ hơn. Hiện nay, khái niệm màng
sinh vật được hiểu là tập hợp các quần xã vi sinh vật bám dính và phát triển trên bề
mặt các môi trường khác nhau thông qua mạng lưới chất ngoại bào do chính chúng
tạo ra [26], [56]. Màng sinh vật có thể hình thành trên bề mặt môi trường vô sinh
hay hữu sinh và là một hiện tượng phổ biến xuất hiện trong tự nhiên, trong đời sống
hay trong nhiều ngành công nghiệp khác nhau.
Hình 2. Ảnh SEM một màng sinh vật do Staphylococcus aureus tạo nên [26]
Các tế bào vi sinh vật trong một màng sinh vật khác biệt với các tế bào sống
trôi nổi tự do bởi việc tổng hợp các chất ngoại bào giúp các tế bào bám dính với
nhau trên bề mặt, sự giảm tỷ lệ tăng trưởng, và sự điều hòa tăng hoặc giảm của các
gen đặc biệt nào đó. Khả năng bám dính của vi sinh vật là một quá trình phức tạp
được điều hòa bởi các đặc điểm khác nhau về môi trường nuôi cấy, chất nền ngoại
bào và bề mặt tế bào [57].
Vi khuẩn bắt đầu quá trình tạo màng sinh vật để đáp ứng với những tác động
cụ thể từ môi trường sống như nguồn chất dinh dưỡng và oxy. Quá trình tạo thành
màng sinh vật cũng trải qua các biến đổi động học trong việc chuyển từ đời sống tự

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 5
Hình 3. Ảnh SEM màng sinh vật nổi được hình thành bởi chủng Bacillus subtilis B-1 [57]
do sang dạng sống bám dính trong cấu trúc màng sinh vật, bao gồm cả việc sản xuất
các chất chuyển hóa thứ cấp và gia tăng các chất chống lại các tác nhân vật lý, hóa
học và sinh học gây hại [56].
1.2 Các dạng màng sinh vật trong tự nhiên và vai trò đối với vi sinh vật
1.2.1 Các chủng vi sinh vật có khả năng tạo màng sinh vật trong tự nhiên
Các vi sinh vật trong tự nhiên ít khi tồn tại riêng rẽ mà thường hình thành tập
hợp quần xã vi sinh vật với một loạt các hoạt động chức năng sinh lý và sinh hóa.
Sự tạo thành màng sinh vật diễn ra tại bề mặt rắn tiếp xúc với môi trường chất lỏng.
Tại đây, các mảnh vụn hữu cơ và chất khoáng tập trung lại tạo điều kiện cho phép
các vi sinh vật có thể sinh trưởng và phát triển thành các vi khuẩn lạc và dần hình
thành nên màng sinh vật trưởng thành [13].
Phân tích thành phần vi sinh vật của màng cho thấy sự hiện diện của vi tảo
và vi khuẩn Gallionella spp. Đặc biệt khả năng oxi hóa sắt của chủng Gallionella
spp gây kết tủa sắt trong đường ống tạo ra những thay đổi không mong muốn về độ
đục, màu sắc và mùi của nước [58]. Các chủng vi khuẩn hiếu khí như Pseudomonas
putrefaciens, Escherichia coli, Bacillus sp, Serratia sp cũng được phân lập. Trong
10 m 1 m

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 6
đó, nghiên cứu cho thấy chủng P. putrefaciens có khả năng tạo chất ngoại bào giúp
gắn kết các nhóm vi sinh vật với nhau để hình thành màng sinh vật [66].
Trong công nghiệp thực phẩm, khả năng bám dính của các vi sinh vật trên bề
mặt các thiết bị chế biến có thể ảnh hưởng nghiêm trọng đến giá trị cảm quan của
sản phẩm thực phẩm. Nghiên cứu của Sule và cộng sự [78] cho thấy chủng E. coli
O157:H7 là tác nhân gây bệnh có thể phân lập từ các mẫu thịt tươi. Bằng kỹ thuật
Real-time PCR, 13 trong số 15 gen ở chủng E. coli này được nghiên cứu cho thấy
có liên quan đến các chức năng sống như trao đổi chất, phân chia tế bào, hình thành
màng sinh học và khả năng gây bệnh.
Nghiên cứu của Kubota và cộng sự [47] về khả năng hình thành màng sinh
vật trên 3 chủng vi khuẩn Lactobacillus plantarum, Lactobacillus brevis và
Lactobacillus fructivorans đại diện cho nhóm vi khuẩn lactic gây hư hỏng thực
phẩm. Đặc điểm của sự hình thành màng sinh vật ở các chủng vi khuẩn này cung
cấp cho chúng ta một hướng nghiên cứu trong việc kiểm soát các tác nhân gây bệnh
trong các sản phẩm thực phẩm.
Một trong những hướng nghiên cứu ứng dụng của màng sinh vật là kiểm soát
sinh học các tác nhân gây bệnh hại ở cây trồng. Các chủng vi khuẩn được phân lập
từ rễ cây hoặc đất trồng ở các khu vực canh tác nông nghiệp cũng cho thấy khả
năng hình thành màng sinh vật tương đối cao như: Bacillus subtilis, Agrobacterium
tumefaciens, Xylella fastidiosa, Rhizobium leguminosarum… Nghiên cứu của Fall
và cộng sự [30] cho thấy Bacillus subtilis bám dính ở rễ cây có khả năng hình thành
màng sinh vật tốt ở nồng độ K+ cao trong môi trường nuôi cấy. Bacillus subtilis
cũng có khả năng ức chế sự lây nhiễm của tác nhân gây bệnh Pseudomonas
syringae pv tomato DC3000 ở rễ cây Arabidopsis nhờ khả năng tạo thành màng
sinh vật và sản xuất surfactin [5]. Khả năng cộng sinh với mô thực vật cũng như sự
tạo thành màng sinh vật trên bề mặt chóp rễ của các chủng Agrobacterium
tumefaciens, Xylella fastidiosa [69] cho thấy sự tương tác cùng có lợi giữa thực vật
và vi khuẩn trong việc cung cấp chất dinh dưỡng và hạn chế các tác nhân gây bệnh.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 7
Ngoài ra, một số chủng vi khuẩn được thu thập từ khu vực nước thải của các
nhà máy, làng nghề cũng được phân lập nhằm tìm ra những chủng có khả năng hình
thành màng sinh vật tốt, ứng dụng trong việc xử lý nước thải và vệ sinh môi trường.
1.2.2 Các dạng tồn tại của màng sinh vật
Trong tự nhiên, có thể bắt gặp và quan sát thấy màng sinh vật trong rất nhiều
môi trường khác nhau: từ môi trường tự nhiên như trên bề mặt của các viên đá nằm
dưới đáy sông suối, trên bề mặt nước của các hồ, ao tù đến các hệ thống nhân tạo
như vòi hoa sen, ống dẫn nước.
1.2.2.1 Trong môi trường tự nhiên
Môi trường nước trong các hồ, ao, sông, suối là điều kiện thuận lợi nhất cho
việc hình thành và phát triển một mạng lưới màng sinh vật. Màng sinh vật có thể
được tạo thành ngay trên bề mặt nước (khoảng tiếp xúc với không khí) để hình
thành nên dạng cấu trúc màng nổi (floating biofilm), có thể quan sát dưới dạng
những cặn hay váng của vi sinh vật trên mặt ao, hồ hay bể lọc nước.
Một dạng khác của màng sinh vật trong tự nhiên được tìm thấy là khi các vi
sinh vật bám dính trên bề mặt vật liệu rắn như các viên sỏi, đá trong nước tạo thành
dạng màng sinh vật bề mặt (hình 4a).
1.2.2.2 Trong các hệ thống thiết bị nhân tạo
Màng sinh vật cũng tồn tại trên bề mặt các thiết bị nhân tạo được cấu tạo chủ
yếu từ vật liệu vô sinh (nhựa, thủy tinh, thép …) như trên vỏ của tàu thuyền, trong
lòng các ống dẫn nước, ống dẫn dầu hay dẫn khí đốt, trên sàn các quầy hàng thực
phẩm.
Trong các thiết bị, đồ dùng gia đình cũng có sự xuất hiện của màng sinh vật
khi các vi sinh vật bám dính trong hệ thống vòi hoa sen, bồn rửa mặt.
1.2.2.3 Trong cơ thể sinh vật sống
Ngay trong cơ thể sống con người cũng xuất hiện màng sinh vật chủ yếu là
của những loài vi sinh vật gây bệnh. Màng sinh vật có thể hình thành trên bề mặt

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 8
lớp tế bào biểu mô như biểu mô ống dẫn niệu, xoang mũi, xoang miệng hay trên
răng tạo thành cấu trúc màng sinh vật gọi là mảng bám răng (hình 4c). Thậm chí bề
mặt của những dụng cụ y tế đặt trong cơ thể như van tim, niệu quản nhân tạo cũng
có thể là vị trí tồn tại của màng sinh vật.
(a) (b) (c)
Hình 4. Một số ví dụ về màng sinh vật
(a): Màng sinh vật trên tảng đá; (b): Màng sinh vật trên bề mặt bàn chải đánh răng;
(c): Màng sinh vật của vi khuẩn sâu răng hình thành nên mảng bám răng
1.2.3 Ảnh hưởng của màng sinh vật đối với vi sinh vật
Sự tạo thành màng sinh vật là một hiện tượng phổ biến trong đời sống của
các vi sinh vật và trở thành phương thức giúp chúng tồn tại và phát triển trong tự
nhiên.
1.2.3.1 Bảo vệ tế bào trước những bất lợi của môi trường
Mạng lưới ngoại bào của màng sinh vật cung cấp nơi khu trú và một hằng số
nội môi thích hợp cho các vi khuẩn tồn tại. Nó đóng vai trò quan trọng trong cấu
trúc và chức năng của các màng sinh vật. Chất nền ngoại bào này cũng có khả năng
ngăn chặn sự xâm nhập của các tác nhân kháng khuẩn vào trong màng nhờ hoạt tính
trao đổi anion. Điều này cũng có nghĩa là nó sẽ làm hạn chế sự khuếch tán của một
số hợp chất từ môi trường xung quanh vào trong màng sinh vật [45].
Màng sinh vật của các chủng vi sinh vật tạo màng giúp chúng kháng lại các
tác nhân là các vi khuẩn, các chất kháng sinh hay chất sát trùng. Cơ chế bảo vệ của
màng sinh vật có thể là: ngăn chặn sự xâm nhập của các chất này vào trong màng

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 9
sinh vật; thay đổi tốc độ tăng trưởng của các vi sinh vật trong màng hay các biến đổi
chức năng sinh lý khác thông qua các phương thức tăng trưởng. Đặc tính này phần
lớn phụ thuộc vào tính chất của cả mạng lưới biofim và tác nhân kháng khuẩn và
thường thể hiện rõ với các chất kháng sinh thuộc nhóm ưa nước và tích điện dương
như các aminoglycoside [36].
Mạng lưới chất ngoại bào cũng được ghi nhận là có khả năng giúp tế bào
chống lại tác động của một số kim loại nặng, các cation và chất độc; đồng thời bảo
vệ tế bào tránh khỏi nhiều yếu tố stress từ môi trường như sự thay đổi độ pH, bức xạ
tia cực tím, áp suất thẩm thấu và sự khô hạn [45]. Thành phần chính của màng sinh
vật chiếm tới 97% là nước. Khả năng giữ nước cao của mạng lưới ngoại bào thông
qua các liên kết hydro trong cấu trúc màng giúp bảo vệ màng chống lại sự khô hạn
trong môi trường tự nhiên. Quá trình hấp thụ các nguyên tố kim loại nặng được biết
đến do tác dụng của nhóm mang điện tích âm trong mạng lưới chất ngoại bào như
nhóm phosphate, lưu huỳnh, hay nhóm chức axit [56].
1.2.3.2 Thu nhận nguồn chất dinh dưỡng từ môi trường
Môi trường nội bào trong cấu trúc màng sinh vật cung cấp phương tiện trao
đổi dinh dưỡng và chuyển hóa chất hiệu quả thông qua các pha dung dịch lớn, tăng
cường khả năng hấp thụ dinh dưỡng cũng như loại bỏ những sản phẩm trao đổi chất
có nguy cơ độc hại [23]. Các vi khuẩn trong mạng lưới màng sinh vật thường bao
gồm nhiều quần thể vi khuẩn khác nhau. Chúng là kết quả của mối liên hệ giữa các
loài sinh vật đồng trao đổi chất. Mối liên kết chặt chẽ này tạo điều kiện thuận lợi
cho sự trao đổi, loại bỏ và phân phối các sản phẩm trao đổi chất trung gian giữa các
loài.
Màng sinh vật cung cấp một môi trường lý tưởng cho sự thiết lập mối quan
hệ hợp dưỡng giữa các loài vi sinh vật. Hợp dưỡng là một trường hợp đặc biệt của
mối quan hệ cộng sinh, trong đó hai loài (hoặc hai chủng) vi sinh vật khác nhau phụ
thuộc lẫn nhau về mặt trao đổi chất để sử dụng một số cơ chất nhất định, đặc biệt là
cho các yêu cầu về năng lượng [31].

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 10
1.2.3.3 Thu nhận những đặc tính di truyền mới
Quá trình trao đổi gen (horizontal gene transfer) đóng vai trò quan trọng
trong sự tiến hóa và đa dạng di truyền của cộng đồng các vi sinh vật trong tự nhiên.
Trao đổi gen được biết đến là sự di chuyển vật liệu di truyền giữa các loài vi sinh
vật khác nhau. Hiện tượng này xảy ra trong vi khuẩn là cách kết hợp trực tiếp DNA
tự do của các tế bào vi khuẩn. Nhờ việc thu nhận những đặc tính di truyền mới giúp
cộng đồng vi sinh vật trong màng tiếp nhận được những gen cần thiết tham gia tích
cực trong hoạt động sống của màng sinh vật [45].
Tầm quan trọng của quá trình trao đổi gen đã được ứng dụng trong những
nghiên cứu về khả năng kháng thuốc của vi sinh vật, kỹ thuật di truyền tạo nên
chủng vi sinh vật mới trong các ngành công nghiệp. Trong đó cơ chế trao đổi gen
phổ biến ở vi sinh vật là truyền gen thông qua plasmid và cầu tiếp hợp. Tuy nhiên
khi các vi khuẩn trong tự nhiên tồn tại dưới dạng màng sinh vật, liên kết với nhau
thông qua mạng lưới chất ngoại bào thì việc tiếp hợp giống như là cơ chế mà nhờ
đó vi khuẩn trong màng sinh vật có thể truyền gen từ tế bào này sang tế bào khác
[26].
1.2.3.4 Mối quan hệ hợp tác giữa các loài
Màng sinh vật được hình thành nhờ sự hợp tác cùng chung sống của nhiều
loài vi sinh vật tạo nên một quần xã vi sinh vật có cấu trúc không gian phức tạp. Do
đó, các loài sinh vật cùng tồn tại trong màng sinh vật thích nghi với những điều kiện
dinh dưỡng, nồng độ khác nhau tạo nên những “vi ổ sinh thái” trong màng. Ngoài
ra, khả năng thích nghi với nhiều điều kiện dinh dưỡng khác nhau giúp các vi sinh
vật tận dụng được tối đa nguồn dinh dưỡng từ môi trường đồng thời hỗ trợ lẫn nhau
theo hướng cùng có lợi trong quá trình chuyển hóa vật chất [4].
Mối quan hệ hợp tác giữa các loài trong màng sinh vật cũng có tác động lớn
đến chu trình tuần hoàn của các nguyên tố trong tự nhiên. Sự phối hợp của nhiều
nhóm vi khuẩn có cơ chế trao đổi chất khác nhau để cùng phân giải một hợp chất
hữu cơ và việc cùng cư trú trong màng sinh vật của các nhóm vi sinh vật sẽ góp
phần thúc đẩy các quá trình này diễn ra nhanh hơn [45].

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 11
1.3 Thành phần, cấu trúc và đặc điểm của màng sinh vật
Quá trình tạo màng sinh vật cần có sự tham gia của nhiều yếu tố để hình
thành nên cấu trúc đặc trưng của màng. Tỷ lệ và mức độ bám dính của tế bào vi
khuẩn vào một bề mặt phụ thuộc vào đặc tính kỵ nước của bề mặt tế bào, sự hiện
diện của lông roi, tiêm mao và chất kết dính, ngoài ra còn có protein màng tế bào và
sự sản suất mạng lưới chất ngoại bào [26]. Thêm vào đó, các bằng chứng thực
nghiệm cho thấy rằng sự phát triển chủ yếu của một màng sinh vật có thể được điều
hòa bởi mật độ tế bào - phụ thuộc vào mức độ biểu hiện của gen được điều khiển
bởi các phân tử tín hiệu ngoại bào.
1.3.1 Mạng lưới ngoại bào
Mạng lưới chất ngoại bào chứa hàm lượng cacbon chiếm 50 - 90% tổng
lượng cacbon hữu cơ trong màng sinh vật và được xem như là vật liệu chất nền
chính của màng sinh vật.
Mạng lưới ngoại bào có thể khác nhau về một số tính chất vật lý và hóa học
nhưng nó bao gồm chủ yếu là các polysaccarit. Một số polysaccarit là trung tính hay
mang điện tích âm, như trường hợp chất ngoại bào ở vi khuẩn gram âm. Sự hiện
diện của axit uronic (ví dụ như D - glucuronic, D - galacturonic, và axit
mannuronic) hoặc liên kết xeton trong phân tử axit pyruvic đưa đến tính chất của
ion âm [79]. Tính chất này rất quan trọng bởi nó cho phép các phân tử polysaccarit
ngoại bào liên kết được với các ion dương hóa trị II như Ca2+ và Mg2+. Từ đó hình
thành liên kết chéo giữa các sợi polymer và tạo ra lực liên kết lớn hơn trong cấu trúc
màng sinh vật. Trong trường hợp của một số vi khuẩn gram dương như
Staphylococcus, thành phần hóa học của mạng lưới ngoại bào có thể hoàn toàn khác
nhau và gồm chủ yếu là các ion dương. Hussain và cộng sự [41] đã phát hiện ra
rằng màng nhầy của tụ cầu khuẩn tạo coagulase - một loại enzyme cho phép chuyển
đổi fibrinogen thành fibrin gây đông máu - âm tính có chứa hỗn hợp axit teichoic và
một lượng nhỏ protein.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 12
Mạng lưới ngoại bào có thể là ưa nước do kết hợp với một lượng lớn các
phân tử nước thông qua liên kết hydro hoặc cũng có thể là kỵ nước. Tuy nhiên,
phần lớn các dạng chất ngoại bào bao gồm cả ưa nước và kỵ nước. Đồng thời mạng
lưới ngoại bào cũng có thể thay đổi độ hòa tan của nó [26].
Sutherland [79] đã ghi nhận hai thuộc tính quan trọng của mạng lưới ngoại
bào. Thứ nhất, thành phần và cấu trúc của các polysaccarit xác định cấu tạo chính
của mạng lưới ngoại bào. Ví dụ, cấu trúc bộ khung của mạng lưới ngoại bào ở nhiều
vi khuẩn chứa nhiều liên kết 1,3 hoặc 1,4--D-fructan và có xu hướng làm cho nó
cứng chắc hơn, ít bị biến dạng và trong một số trường hợp trở nên kém hòa tan hoặc
không hòa tan. Các liên kết khác trong phân tử polysaccarit như liên kết 1,2 hoặc
1,6--D-glucan giúp cấu trúc mạng lưới ngoại bào trở nên linh hoạt hơn. Thứ hai,
mạng lưới ngoại bào của màng sinh vật thường không đồng nhất, có thể thay đổi
theo không gian và thời gian.
Leriche và cộng sự [53] đã sử dụng liên kết đặc hiệu của lectin với một loại
đường đơn để đánh giá sự phát triển của màng sinh vật ở các vi sinh vật khác nhau.
Kết quả nghiên cứu cho thấy các sinh vật khác nhau sản xuất một lượng chất ngoại
bào khác nhau và lượng chất ngoại bào này tăng lên theo thời gian. Mạng lưới chất
ngoại bào có thể liên kết với các ion kim loại, các cation hóa trị hai hay các đại phân
tử khác như protein, DNA, lipit… Điều kiện dinh dưỡng của môi trường nuôi cấy
cũng được chứng minh là có ảnh hưởng đến sự sản xuất chất ngoại bào. Nghiên cứu
cho thấy sự dư thừa nguồn carbon sẵn có và sự hạn chế của nitơ, kali, phospho
trong môi trường nuôi cấy đã thúc đẩy quá trình tổng hợp mạng lưới ngoại bào [79].
Sự tăng trưởng chậm của vi khuẩn cũng sẽ làm tăng cường sản xuất chất
ngoại bào. Sự tích nước trong mạng lưới chất ngoại bào là rất lớn, do đó nó ngăn
chặn sự khô hạn của màng sinh vật trong tự nhiên. Mạng lưới chất ngoại bào cũng
góp phần vào đặc tính kháng kháng sinh của màng sinh vật bằng cách cản trở sự
vận chuyển lượng chất kháng sinh qua màng, có lẽ là thông qua việc liên kết trực
tiếp với các chất này [25].

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 13
1.3.2 Các thành phần khác
1.3.2.1 Vai trò của lông roi, tiêm mao và các phân tử bám dính
Những nghiên cứu trước đây đã đưa ra mô hình hóa học đơn giản dựa trên
nồng độ chất điện phân và lực liên kết van der Waals để giải thích sự bám dính của
vi khuẩn lên bề mặt [55]. Nhưng trong những nghiên cứu gần đây, chủ yếu dựa trên
sự phát sinh đột biến của gen nhảy, lại cho thấy chính các thành phần cấu trúc của tế
bào vi khuẩn như lông roi, tiêm mao và các phân tử bám dính đóng vai trò quan
trọng trong sự xâm nhiễm của vi khuẩn lên một bề mặt nào đó.
Chức năng chính của lông roi trong sự hình thành màng sinh vật là giúp cho
vi sinh vật di chuyển trong môi trường nước tốt hơn, tạo nên những tương tác ban
đầu giữa bề mặt và tế bào. Thí nghiệm của De Flaun, De Weger cùng các cộng sự
[22], [24] đã chứng minh sự thiếu vắng của lông roi làm suy giảm khả năng xâm
nhiễm của Pseudomonas fluorescens lên rễ cây khoai tây, lúa mì và làm giảm độ
bám dính tế bào của P. aeruginosa và P. fluorescens lên bề mặt polystyrene [63].
Tương tự, nghiên cứu trên các chủng Vibrio cholerae và Escherichia coli đột biến
thiếu lông roi đã cho thấy không có sự hình thành màng sinh vật như ở các dạng
hoang dại của chúng vẫn thực hiện trên bề mặt nhựa polyvinylchloride (PVC) [84].
Tiêm mao và các phân tử bám dính liên kết với tiêm mao cũng có vai trò
quan trọng trong sự bám dính và xâm nhiễm bề mặt. Nghiên cứu của Schmoll và
cộng sự [73] trên chủng vi khuẩn gây bệnh E. coli cho thấy biểu hiện của gen sfaA,
một gen mã hóa cho các phân tử bám dính tiêm mao, được điều hòa tăng cường khi
có sự tiếp xúc của vi khuẩn với bề mặt. Ở E. coli, khả năng bám dính bề mặt giảm
đi khi có sự đột biến ở gen sinh tổng hợp curlin csgA và gen sinh tổng hợp tiêm mao
type I fim H, một loại tiêm mao chứa các phân tử đặc hiệu với mannose [68]. Tương
tự, đột biến gen tổng hợp tiêm mao hemagglutinin có độ nhạy cao với mannose ở
V. cholerae cũng làm giảm độ bám dính bề mặt [84].
Như vậy, có thể nói rằng các loài vi sinh vật có lông roi và tiêm mao cũng sẽ
có ưu thế hơn trong việc di chuyển đến một bề mặt giá thể xác định - nơi có điều

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 14
kiện thuận lợi cho việc hình thành màng sinh vật, đồng thời giúp cho việc bám dính
ban đầu của tế bào với bề mặt tốt hơn [26].
1.3.2.2 Vai trò của protein màng
Protein màng tế bào được ghi nhận là có ảnh hưởng đáng kể trong sự bám
dính cũng như trong giai đoạn phát triển sớm của màng sinh vật. Nghiên cứu cho
thấy sự gắn kết của E. coli lên bề mặt vô sinh dẫn đến những thay đổi trong thành
phần protein ngoại vi của màng tế bào hay chính là những tương tác vật lý của tế
bào với bề mặt đã làm biến đổi đặc tính bề mặt của lớp màng ngoại bào [65].
Đột biến protein màng tế bào, bao gồm protein liên kết với Ca2+, hemolysin,
protein vận chuyển và protein bơm K+, đã gây nên khiếm khuyết trong quá trình
bám dính của P. putida KT 2440 lên hạt ngô giống [29].
Nghiên cứu của Whiteley và cộng sự [85] ở Pseudomonas aeruginosa cho
thấy biểu hiện của 2 gen tatA và tatB cùng mã hóa cho protein vận chuyển tolA,
đóng vai trò quan trọng trong cấu trúc lớp lipopolysaccarit của màng. Một nghiên
cứu khác của Sauer và cộng sự [72] cũng chứng minh rằng protein porin E1 và
protein vận chuyển ABC cũng được biểu hiện tăng cường ở các tế bào màng sinh
vật P. aeruginosa.
1.3.2.3 Vai trò của sự cảm ứng mật độ tế bào
Tín hiệu giữa các tế bào với nhau gần đây đã được chứng minh là đóng vai
trò quan trọng trong sự bám dính cũng như tách rời của tế bào ra khỏi màng sinh
vật.
Xie và cộng sự [85] đã chỉ ra rằng mảng bám răng là một màng sinh học
phức tạp bao gồm hơn 30 chi đại diện cho hơn 500 loài vi sinh vật khác nhau. Mặc
dù phức tạp nhưng màng sinh vật này có tổ chức rất cao. Những khuẩn lạc đầu tiên
hình thành là các vi khuẩn Gram dương, chủ yếu là liên cầu khuẩn, kế đến là một
loạt các loài vi sinh vật khác và sự xuất hiện lên đến đỉnh điểm trong sự hình thành
màng là nhóm vi khuẩn Gram âm như Porphyromonas gingivalis chiếm ưu thế.
Nghiên cứu cho thấy những khuẩn lạc ban đầu của Streptococcus gordonii cung cấp

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 15
chất nền bám dính cho Porphyromonas gingivalis thông qua sự tương tác của các
cặp thụ thể bám dính, được biết đến là các phân tử protein FimA [11]. Ngược lại,
tương tác phân tử giữa các tế bào S. cristatus và P. gingivalis đã kìm hãm sự biểu
hiện của gen fimA và kết quả là P. gingivalis không thể tiếp tục bám dính và sẽ
được tách rời khỏi màng sinh vật [48].
Davies và cộng sự [20] đã đưa ra hai hệ thống tín hiệu ngoại bào khác nhau ở
P. aeruginosa có liên quan đến sự hình thành màng sinh vật, đó là lasR - las I và
rhlR - rhlI. Ở mật độ quần thể tế bào đủ lớn, các tín hiệu này đạt đến nồng độ cần
thiết để kích hoạt các gen liên quan đến sự khác biệt màng sinh vật. Các dạng đột
biến không thể tạo ra cả hai tín hiệu sẽ hình thành một màng khác nhiều so với
màng sinh vật của dạng hoang dại như màng mỏng hơn, các tế bào được đóng gói
dày đặc hơn và cấu trúc của màng cũng chưa hoàn thiện.
Vai trò của tín hiệu nội bào trong màng sinh vật của nhiều loài khác biệt
đáng kể so với màng sinh vật của một loài. Những tín hiệu này được phân loại như
là một sản phẩm truyền tin tích cực hay thụ động làm thay đổi trạng thái của các tế
bào vi khuẩn lân cận. Chúng bao gồm các sản phẩm trao đổi chất của vi khuẩn, các
phân tử acyl - homoserine lactone, vật chất di truyền như DNA hay RNA, v.v…Các
tín hiệu này có thể làm thay đổi sự phân bố của các loài cụ thể trong màng sinh vật,
thay đổi sự biểu hiện protein trong các tế bào lân cận, đưa đến những đặc điểm di
truyền mới ở các tế bào lân cận và kết hợp các vi khuẩn với nhau trong màng sinh
vật [83].
1.4 Các yếu tố ảnh hưởng đến sự hình thành và phát triển của màng sinh vật
1.4.1 Các giai đoạn tạo thành màng sinh vật
Dựa trên các phương pháp phân tích di truyền học, proteomics và sinh học
phân tử, cùng với những phân tích về mặt cấu trúc, hóa học màng sinh vật, các nhà
khoa học đã đưa ra một mô hình cấu trúc màng sinh học cơ bản [16]. Trong mô
hình này, vi khuẩn hình thành nên các vi khuẩn lạc và được bao quanh bởi một
mạng lưới chất ngoại bào giúp các thành phần tế bào liên kết với nhau một cách có

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 16
trật tự đảm bảo sự trao đổi thông tin liên tục diễn ra giữa các tế bào đồng thời tạo
nên những kênh dẫn truyền dịch ngoại bào bên trong màng sinh vật. Nhờ đó, dịch tế
bào có thể đi qua màng sinh vật tạo điều kiện cho việc khuếch tán, phân phối chất
dinh dưỡng đến khắp các tế bào trong màng cũng như loại bỏ các chất thải [76].
Sự tạo thành màng sinh vật cũng giống như một quá trình phát triển của vi
sinh vật và cần phải trải qua một số bước. Cơ chế phân tử giữa các vi sinh vật có thể
khác nhau nhưng các giai đoạn trong sự phát triển của màng sinh vật thì luôn được
bảo tồn. Quá trình này lần lượt bao gồm: sự gắn kết của các tế bào vi sinh vật trôi
nổi tự do lên một bề mặt, sự tăng trưởng và liên kết các tế bào thành vi khuẩn lạc,
sự tạo thành màng sinh vật trưởng thành (hoàn chỉnh), và cuối cùng là sự tách rời
của các tế bào vi sinh vật thành dạng dịch lỏng tế bào (hình 5).
Hình 5. Mô hình phát triển của màng sinh vật [76]
1.4.1.1 Giai đoạn 1: Giai đoạn gắn kết thuận nghịch
Trước khi vi sinh vật bám dính lên bề mặt, các phân tử hữu cơ, protein,
glycoprotein đã tiếp xúc và hình thành nên một màng điều kiện, đó là một khu vực
giàu dinh dưỡng giúp thuận lợi cho sự trao đổi chất của các tế bào vi sinh vật [8].
Gắn kết Gắn kết Hình thành Hình thành Tách rời thuận nghịch không thuận các vi khuẩn màng sinh vật nghịch lạc trưởng thành

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 17
Khi bề mặt điều kiện được hình thành, các thuộc tính của nó thay đổi để thu hút các
vi sinh vật. Sự gắn kết của các vi sinh vật lên bề mặt nhờ lực liên kết van der Waals,
lực hút tĩnh điện, tương tác ưa nước và các tương tác đặc biệt khác, hoặc bởi một sự
kết hợp các liên kết này, tùy theo mức độ gần gũi của các vi sinh vật với bề mặt
bám dính. Nhờ khả năng di chuyển độc lập bằng các cử động co rút tế bào hay sử
dụng các tiêm mao, và khả năng tiết các chất ngoại bào giúp các tế bào riêng rẽ
được bao bọc trong một mạng lưới và bắt đầu sự hình thành màng sinh vật [62].
Tuy nhiên, các tế bào này chưa hẳn đã đi vào quá trình hình thành màng sinh vật và
có thể rời bề mặt để tiếp tục đời sống phù du.
1.4.1.2 Giai đoạn 2: Gắn kết không thuận nghịch
Sau khi gắn kết thuận nghịch ban đầu lên một bề mặt, vi sinh vật không
những phải giữ liên kết với bề mặt giá thể mà còn phải tăng trưởng để hình thành
một màng sinh vật hoàn chỉnh. Vì vậy, giai đoạn tiếp theo là sự sản xuất các chất
ngoại bào nhằm làm tăng tính bám dính ổn định thông qua các cầu nối hữu cơ giữa
tế bào và giá thể [60].
Các nghiên cứu trên gen chỉ thị cho thấy biểu hiện của gen sinh tổng hợp
alginate ở Pseudomonas aeruginosa algC và algD được điều hòa tăng lên trong 15
phút ngay sau khi có sự bám dính đầu tiên của vi khuẩn lên một bề mặt. Mặc dù sự
sản xuất alginate cũng được xem như là phương thức để hình thành nên bộ khung
cấu trúc và hóa học của màng sinh vật [77], nhưng những nghiên cứu gần đây lại
chứng minh rằng chính sự hiện diện của mạng lưới chất ngoại bào chứ không phải
alginate là thực sự cần thiết cho sự hình thành màng sinh vật của P.aeruginosa [54].
Việc chuyển từ giai đoạn bám dính thuận nghịch sang giai đoạn bám dính
không thuận nghịch được thực hiện nhờ lông roi, tiêm mao vào các sợi bám dính.
Trong khi sự vận động thông qua trung gian lông roi được đánh giá là quan trọng
trong bước đầu thiết lập sự bám dính của vi sinh vật lên bề mặt thì vận động co rút
được chỉ ra là cần thiết cho sự trưởng thành của màng sinh vật trong điều kiện tĩnh.
Cụ thể, nhu động co rút giúp cho sự hình thành nên các vi khuẩn lạc trong màng

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 18
sinh vật bằng cách tạo điều kiện thuận lợi cho tương tác giữa các vi khuẩn với bề
mặt để hình thành nên các nhóm tế bào, qua đó giúp tăng cường mức độ bám dính
với bề mặt [62].
1.4.1.3 Giai đoạn 3: Hình thành mạng lưới vi khuẩn lạc
Các hợp chất polymer ngoại bào tiếp tục được tạo ra bởi các tế bào để liên
kết các tế bào với nhau một cách có tổ chức đồng thời tạo thành cầu nối giữa các vi
khuẩn lạc. Chúng cũng có vai trò trong việc thu hút các tế bào sống trôi nổi (có thể
là từ nhiều loài khác nhau) trong môi trường. Kết quả là mật độ tế bào trong một
màng sinh vật cũng như lượng các polymer ngoại bào tạo ra tăng lên. Một mạng
lưới màng sinh vật dần được hình thành [76].
1.4.1.4 Giai đoạn 4: Hình thành một màng sinh vật hoàn chỉnh
Khi tế bào vi sinh vật bám dính không thuận nghịch lên bề mặt thì quá trình
trưởng thành của màng sinh vật bắt đầu. Trong suốt quá trình này, sự phân chia trực
phân của các tế bào vi sinh vật bám dính không thuận nghịch là nguyên nhân giúp
các tế bào phân chia để lan rộng và phát triển đầy lên từ các điểm gắn kết để hình
thành các vi khuẩn lạc hay các cụm tế bào [40].
Bản chất của bề mặt là nơi bám dính của các vi khuẩn lạc và chính các điều
kiện vật lý và hóa học của môi trường sẽ quyết định cơ chế hình thành màng sinh
vật nào là chiếm ưu thế. Sự trưởng thành của màng sinh vật dẫn đến sự kế tiếp của
các cấu trúc dạng nấm hay dạng cột xen kẽ với các kênh chứa đầy dịch, và một khi
phát triển đến đầy đủ thì màng sinh vật sẽ cho thấy những mô hình biến đổi trong sự
tăng trưởng của vi khuẩn cũng như sự tương tác sinh lý và hiệu quả trao đổi chất
[82].
1.4.1.5 Giai đoạn 5: Tách rời
Khả năng phát triển của màng sinh vật giới hạn trong điều kiện dinh dưỡng
của môi trường nuôi cấy và biểu hiện của các phân tử cảm ứng mật độ tế bào. Các
phân tử này được giải phóng ra nhằm đáp ứng với những hạn chế về dinh dưỡng,
sự tích tụ các sản phẩm độc hại và một số nhân tố khác, bao gồm các yếu tố pH,

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 19
nguồn cung cấp cacbon, oxy [61]. Trong một số trường hợp, khi màng sinh vật đạt
đến khối lượng và một mức cân bằng động tối đa thì các tế bào trong đó sẽ tự tách
rời và cùng với các tế bào của một màng khác hình thành nên các vi khuẩn lạc [46].
Những nghiên cứu gần đây đã cho thấy rằng quá trình tách rời là hết sức
phức tạp. Sự khác biệt về mặt tế bào và hoạt tính của nó tại các trung tâm của một
cấu trúc màng sinh vật trưởng thành dẫn đến sự phân tách tế bào từ bên trong cấu
trúc màng, chuyển sang lấp đầy các khoảng trống của một màng sinh vật khác chưa
hoàn thiện. Cơ chế tách rời của màng sinh vật liên quan đến sự phân hủy các tế bào
bên trong cấu trúc của nó. Enzyme lyase phân hủy polysaccarit đóng vai trò quan
trọng trong sự phân rã màng sinh vật ở nhiều loài vi sinh vật bằng cách phân hủy
mạng lưới chất nền ngoại bào được tạo ra từ các tế bào [44].
Boyd và Chakarabarty [9] đã chứng minh sự cảm ứng biểu hiện của enzyme
lyase phân giải alginate ở P. aeruginosa làm giảm đáng kể tổng lượng sản xuất
alginate, tương ứng với sự tăng số lượng tế bào bị phân tách. Điều này giúp các nhà
nghiên cứu khẳng định vai trò của enzyme phân giải alginate ở dạng P. aeruginosa
hoang dại có thể là nguyên nhân giải phóng các tế bào khỏi các bề mặt rắn hay các
màng sinh vật của chúng, làm phát tán những vi sinh vật vào trong môi trường nuôi
cấy.
Quá trình tách rời cũng có nguyên nhân bởi các lực tương tác vật lý. Brading
và cộng sự [10] đã nhấn mạnh tầm quan trọng của các tương tác vật lý trong sự tách
rời, bao gồm 3 quá trình chính đó là: sự xói mòn hay phân cắt (liên tục loại bỏ
những phần nhỏ của màng sinh vật), sự bóc tách (loại bỏ nhanh chóng và với lượng
lớn) và sự bào mòn (sự tách rời do va chạm của lượng lớn các hạt phân tử chất lỏng
với màng sinh vật). Characklis [12] nhận thấy rằng tỷ lệ xói mòn tăng lên cùng với
độ dày màng sinh vật và dòng chất lỏng tại bề mặt giao diện với màng sinh vật mà ở
đó màng dày lên. Với sự gia tăng của tốc độ dòng chảy, thủy động lực học của lớp
ranh giới giảm, dẫn đến sự pha trộn và nhiễu loạn ở gần bề mặt màng sinh vật. Sự
bóc tách xảy ra ngẫu nhiên hơn sự xói mòn và được cho là kết quả của sự suy giảm
chất dinh dưỡng và oxy trong cấu trúc màng sinh vật.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 20
1.4.2 Các yếu tố ảnh hưởng đến sự hình thành màng sinh vật
Trong tự nhiên, màng sinh vật có thể được hình thành trên các bề mặt khác
nhau bao gồm cả các mô sống, các thiết bị, dụng cụ y tế, các hệ thống ống nước cấu
tạo từ các vật liệu khác nhau. Do vậy, để chuyển từ dạng sống tự do sang dạng cấu
trúc màng sinh vật đòi hỏi một loạt những điều kiện nhất định. Trong đó, ba yếu tố
chính quy định nên sự hình thành màng sinh vật bao gồm:
Tính chất bề mặt giá thể
Điều kiện môi trường
Đặc tính tế bào của các chủng vi sinh vật [45]
1.4.2.1 Tính chất bề mặt giá thể
Đây là yếu tố quyết định đến việc hấp thụ chất hữu cơ và bám dính của tế
bào bởi vậy mỗi loài vi khuẩn chỉ hình thành màng sinh vật trên một số loại bề mặt
với tính chất nhất định.
Sự phát triển của các tế bào bên trong màng sinh vật đã được chứng minh là
có tăng lên khi tăng mức độ thô ráp của bề mặt. Characklis và cộng sự [12] đã ghi
nhận mức độ bám dính của khuẩn lạc vi sinh vật gia tăng khi các bề mặt càng ghồ
ghề. Điều này được giải thích là do ở bề mặt thô nhám, lực tương tác giữa các tế
bào với bề mặt giảm đi và diện tích tiếp xúc được tăng lên đáng kể so với các bề
mặt trơn nhẵn.
Tính chất hóa lý của bề mặt vật liệu cũng ảnh hưởng mạnh mẽ đến tốc độ và
mức độ bám dính của tế bào lên bề mặt. Hầu hết các nghiên cứu cho thấy rằng các
vi sinh vật gắn kết với một bề mặt kị nước, không phân cực như Teflon và nhựa
nhanh hơn và tốt hơn so với bề mặt một vật liệu ưa nước như thủy tinh hay kim loại
[7].
Bên cạnh đó, diện tích bề mặt là một trong những yếu tố quan trọng ảnh
hưởng đến sự phát triển của màng sinh vật. Theo nguyên tắc, diện tích bề mặt càng
lớn càng làm tăng khả năng tiếp xúc với tế bào, qua đó tạo điều kiện cho việc bám
dính lên bề mặt giá thể. Các hệ thống ống dẫn khác với hầu hết các môi trường tự

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 21
nhiên (ao hồ, sông …) là thường có một diện tích bề mặt khá lớn tạo điều kiện cho
việc tiếp xúc giữa tế bào vi khuẩn và bề mặt.
1.4.2.2 Điều kiện môi trường
Các đặc trưng hóa lý của môi trường như nhiệt độ, pH, mức độ dinh dưỡng,
nồng độ các ion, đóng một vai trò quan trọng ảnh hưởng đến tốc độ gắn kết của vi
sinh vật lên bề mặt.
Các nghiên cứu đã chỉ ra rằng khả năng bám dính của vi sinh vật và sự hình
thành màng sinh vật cũng chịu ảnh hưởng của nhịp điệu mùa ở các lưu vực nước
khác nhau [27]. Hiệu ứng này có được là do ảnh hưởng của nhiệt độ nước theo mùa
lên các thông số tăng trưởng của vi sinh vật. Fletcher và cộng sự [32] cũng đã
chứng minh sự gia tăng nồng độ của một số cation (Na+, Ca2+, Fe3+, La+) ảnh hưởng
đến khả năng bám dính của chủng Pseudomonas fluorescens lên bề mặt thủy tinh,
bằng cách làm giảm lực tương tác giữa các tế bào vi khuẩn với bề mặt kính. Nghiên
cứu của Cowan và cộng sự [17] cho thấy sự gia tăng nồng độ chất dinh dưỡng tỷ lệ
thuận với số lượng gắn kết của các tế bào vi khuẩn lên bề mặt.
Việc tạo màng sinh vật có thể coi như là một cách thức tồn tại, phát triển của
vi sinh vật trong những điều kiện dinh dưỡng thấp của môi trường. Cơ chế của quá
trình này có thể được hiểu như sau: khi lượng chất dinh dưỡng bao gồm nguồn
cacbon, nitơ bị giảm sút thì các vi sinh vật sống trôi nổi trong môi trường nước sẽ
có xu hướng tập trung đến nơi có nguồn dinh dưỡng tích tụ. Khi các hợp chất hữu
cơ tích tụ lại trên bề mặt, chúng sẽ thu hút các vi khuẩn, tảo và động vật nguyên
sinh đến, theo thời gian sẽ phát triển thành một màng sinh vật.
1.4.2.3 Đặc tính của tế bào
Mặc dù màng sinh vật là hình thức tồn tại phổ biến của vi sinh vật trong môi
trường tự nhiên nhưng không phải vi sinh vật nào cũng có khả năng hình thành
màng sinh vật. Các đặc tính của tế bào bao gồm các cấu trúc phụ trợ như lông roi,
tiêm mao, khả năng di động, bám dính, khả năng tạo các chất ngoại bào (protein,
polysaccarit), cảm biến mật độ (quorum sensing) ảnh hưởng lớn đến việc hình thành

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 22
màng sinh vật. Thí nghiệm so sánh giữa hai chủng Pseudomonas fluorescens của
Davey và cộng sự [19] đã cho thấy chủng di động có khả năng hình thành màng
sinh vật nhanh hơn so với chủng không di động.
Tuy nhiên, một số loài không có phần phụ trợ tế bào nhưng vẫn có khả năng
hình thành màng sinh vật mạnh dựa vào khả năng tự tổng hợp các chất ngoại bào
như lipopolysaccarit, glycoprotein tạo thành cấu trúc màng giáp (capsule), màng
nhày (slime) bao quanh tế bào. Đặc tính này tạo tính tự kết dính cho tế bào và chủ
yếu có trong những loài vi sinh vật gây bệnh như Streptococcus mutants,
Streptococcus salivarius, Xanthomonas, Bacillus anthracis [15].
1.4.3 Điều hòa quá trình hình thành màng sinh vật
Quá trình chuyển từ dạng sống phù du sang kiểu tăng trưởng bám dính trên
bề mặt thực hiện theo một con đường phức tạp và cần một cơ chế điều hòa kiểm
soát sự hoạt động của các gen biểu hiện sự hình thành màng sinh vật. Bằng kỹ thuật
di truyền và sinh học phân tử kết hợp với phương pháp kính hiển vi tiên tiến đã góp
phần làm sáng tỏ cơ chế điều hòa và các yếu tố cơ bản của quá trình hình thành
màng sinh vật.
1.4.3.1 Con đường dẫn truyền tín hiệu hai thành phần
Sự bám dính của tế bào với bề mặt phụ thuộc vào cả yếu tố cơ hội (tức là khả
năng có hay không sự tiếp xúc trực tiếp của vi khuẩn với bề mặt) và cả sự tương tác
thuận lợi giữa tế bào và bề mặt tiếp xúc để có thể vượt qua lực đẩy giữa hai bề mặt
[35].
Mặc dù sự tiếp xúc ban đầu giữa tế bào vi sinh vật và bề mặt không nhất thiết
đòi hỏi sự điều hòa, nhưng các bằng chứng trong nghiên cứu đã cho thấy rằng sự
hình thành những tương tác ổn định giữa tế bào và bề mặt được điều hòa bởi hệ
thống tín hiệu hai thành phần. Hệ thống này bao gồm hai protein: một protein cảm
biến kinase đóng vai trò nhận biết tín hiệu từ môi trường, tự động phosphoryl hóa,
sau đó kích hoạt thành phần protein còn lại, đó là một phân tử điều hòa phản ứng.
Phân tử điều hòa phản ứng này hoạt động như là một nhân tố hoạt hóa gen, kích

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 23
thích sự phiên mã cho phép vi khuẩn nhanh chóng thích nghi với các điều kiện của
môi trường [52].
1.4.3.2 Các yếu tố điều hòa sự trao đổi Cacbon
Trong hệ gen của một số vi khuẩn có chứa gen mã hóa cho một nhân tố điều
hòa trao đổi nguồn cacbon cần thiết cho sự hình thành màng sinh vật. Ở
P.aeruginosa là gen crc còn ở E.coli là csrA. Nghiên cứu của O’Toole và cộng sự
[61] chứng minh rằng gen crc được kích hoạt bởi các sản phẩm trung gian của chu
trình TCA, là nguồn cacbon ưa thích của P. aeruginosa. Bên cạnh đó, nhân tố Crc
còn hoạt hóa sự phiên mã của gen pilA, mã hóa cho các tiểu đơn vị cấu trúc cần
thiết cho quá trình sinh tổng hợp tiêm mao type IV. Đây là một bước giúp các tế bào
nối kết với nhau để hình thành các vi khuẩn lạc, từ đó tạo thành màng sinh vật.
Nghiên cứu của Jackson và cộng sự [42] về sự ảnh hưởng của nhân tố CsrA
lên sự hình thành màng sinh vật cho thấy: Khác với chủng P. aeruginosa, sự gián
đoạn trong biểu hiện của gen csrA làm tăng khả năng hình thành màng sinh vật hơn
so với chủng hoang dại. Tác dụng chính của nhân tố CsrA được biết đến chỉ như là
một chất điều hòa sự chuyển hóa glycogen - nguồn cacbon chính trong giai đoạn
tổng hợp các yếu tố cần thiết như tiêm mao và các chất bám dính trong quá trình
hình thành màng sinh vật. Cho nên một sự tăng cường quá mức CsrA có thể dẫn
đến ức chế sự hình thành màng sinh vật.
1.4.3.3 Sự điều hòa phụ thuộc các giai đoạn
Trong nhiều nghiên cứu, người ta đã chứng minh được rằng nhân tố RpoS
đóng vai trò quan trọng trong sự hình thành và phát triển của màng sinh vật. Tuy
nhiên yếu tố này lại xuất hiện không giống nhau trong các giai đoạn phát triển của
từng loài vi sinh vật. Nếu như ở E. coli, người ta thấy được sự xuất hiện của RpoS
với nồng độ thấp ở pha tăng trưởng lũy thừa và có sự tích tụ lại thì ở P. aeruginosa,
sự hiện diện của RpoS được ghi nhận ở đầu pha cân bằng [33].
Sự sản xuất RpoS được điều hòa ở nhiều mức độ khác nhau giúp tế bào đáp
ứng với các điều kiện stress của môi trường, bao gồm cả sự giới hạn về mặt dinh

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 24
dưỡng. RpoS điều chỉnh độ dày của cấu trúc màng sinh vật cho phép tế bào thu
nhận tối đa nguồn chất dinh dưỡng [75].
Khi màng sinh vật đạt kích thước đủ lớn, các tế bào trung tâm giảm hấp thụ
các chất dinh dưỡng, dẫn đến kích hoạt RpoS, là tín hiệu giúp tế bào nhận biết sự
giới hạn của nguồn chất dinh dưỡng trong màng sinh vật. Do đó, các tế bào liên kết
trong màng sinh vật được giải phóng ở dạng tế bào trôi nổi tự do di chuyển đến môi
trường mới thuận lợi hơn [76].
1.4.3.4 Cơ chế cảm biến tới hạn (Quorum sensing)
Thông tin liên lạc nội bào giữa các vi sinh vật được thực hiện chủ yếu thông
qua các sản phẩm của vi sinh vật khuếch tán từ tế bào này sang tế bào khác. Một
sản phẩm tiêu biểu cho các phân tử cảm biến tín hiệu được biết đến là acyl - HSL
[45].
Nghiên cứu trên chủng P. aeruginosa cho thấy các phân tử acyl - HSL chịu
trách nhiệm xác định sự tách biệt các khối tế bào vi sinh vật trong cấu trúc không
gian ba chiều của màng sinh vật, góp phần giữ cho cấu trúc màng sinh vật ổn định.
Các chủng P. aeruginosa đột biến không sản xuất được acyl - HSL dẫn đến các tế
bào trong màng sinh vật được đóng gói chặt chẽ với nhau và dễ dàng bị phá vỡ bởi
SDS. Acyl - HSL cũng là các phân tử trung gian trong bề mặt bám dính ở chủng
Pseudomonas fluorescens [83].
Mặc dù còn khá ít những hiểu biết về vai trò của các tín hiệu nội bào trong
màng sinh vật đa loài, song các nhà nghiên cứu tin rằng có một sự khác biệt đáng kể
giữa chúng với các màng sinh vật đơn loài [45]. Những nghiên cứu về các tín hiệu
nội bào ảnh hưởng đến sự hình thành màng sinh vật vẫn đang là một trong những
vấn đề vô cùng thú vị cần được nghiên cứu.
1.5 Nghiên cứu ứng dụng màng sinh vật
1.5.1 Ứng dụng màng sinh vật trong việc xử lý nước thải
Xử lý nước thải môi trường, nhất là trong tình hình thực tiễn hiện nay là một
vấn đề mang tính thời sự cấp thiết. Bởi lẽ xử lý nước thải không chỉ nhằm mục đích

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 25
cải thiện điều kiện vệ sinh môi trường sống của con người mà xa hơn còn nhằm duy
trì cân bằng sinh thái, tạo điều kiện phát triển bền vững lâu dài cho loài người. Hiện
nay, việc xử lý nước thải nói chung theo hướng áp dụng các kỹ thuật sinh học rất
được chú trọng do chúng có tính bền vững, thích nghi với nhiều điều kiện trong tự
nhiên.
Trong cấu trúc màng sinh vật, các vi sinh vật liên kết với nhau chặt chẽ, tạo
ra một cấu trúc bền vững, hoạt động có hiệu quả hơn trong việc xử lý nước thải.
Đồng thời, các vi sinh vật trong mạng lưới màng sinh vật sẽ cùng hợp tác trao đổi
chất giúp cho quá trình loại bỏ các chất gây ô nhiễm trong nước diễn ra dễ dàng và
hiệu quả [76].
So sánh với việc sử dụng các chủng vi sinh vật trôi nổi để xử lý nước thải thì
công nghệ xử lý sinh học nước thải bằng màng sinh vật mang lại nhiều lợi ích đáng
kể. Một là, mật độ các chủng vi sinh vật trong màng sinh vật cao hơn nhiều, tạo
điều kiện xử lý tối đa nguồn nước thải. Hai là, ngoài việc loại bỏ các chất không
mong muốn trong nước thải thì quá trình tiếp theo là loại bỏ các vi sinh vật này khỏi
môi trường. Đối với các tế bào trôi nổi, việc khử trùng nguồn nước sẽ tốn một chi
phí lớn, do vậy việc áp dụng màng sinh vật trên các giá thể cố định thể hiện ưu thế
lớn. Với những ưu điểm này, các nhà khoa học đã đề xuất công nghệ xử lý nước
thải ứng dụng màng sinh vật được xem là giải pháp thân thiện môi trường [50].
1.5.2 Ứng dụng màng sinh vật trong việc ức chế các vi sinh vật gây hại
Đấu tranh phòng trừ bệnh gây hại cho cây trồng đã và đang là hướng nghiên
cứu rất được quan tâm ứng dụng trong ngành nông nghiệp. Và một trong những
biện pháp được ưa chuộng đó là đấu tranh sinh học. Trong đó, nhiều nghiên cứu cho
thấy rằng các vi sinh vật có thể hoạt động như một tác nhân đối kháng với nhiều
mầm bệnh khác nhau ở thực vật bao gồm các loại sâu bọ và các mầm bệnh vi sinh
vật như nấm, xạ khuẩn, vi khuẩn.
Các vi sinh vật có thể tồn tại tự do trong đất hoặc bám dính với bề mặt mô
thực vật thành từng cụm tế bào. Mối quan hệ này phần lớn mang lại lợi ích cho cả
hai bên: thực vật là nguồn cung cấp chất dinh dưỡng và nơi khu trú lâu dài cho các

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 26
quần thể vi sinh vật. Mặt khác, các vi sinh vật trong mối tương tác này có thể làm
thay đổi môi trường sống xung quanh theo hướng có lợi cho sự phát triển của thực
vật [69].
Một trong những cơ chế giúp các vi sinh vật ức chế mầm bệnh gây hại ở cây
trồng đó là thông qua chất kháng sinh. Ví dụ, chủng Bacillus cereus UW85 có khả
năng tổng hợp cả zwittermycin và kanosamine [69]. Khả năng tổng hợp chất kháng
sinh giúp làm tăng tính cạnh tranh giữa các nhóm vi sinh vật; đồng thời cũng là cơ
sở để tạo ra các chế phẩm vi sinh vật có khả năng phòng trừ bệnh gây hại ở thực vật
chủ yếu dưới dạng phân bón vi sinh bổ sung vào nguồn đất trồng.
Bais và cộng sự [5] đã chứng minh rằng nhờ quá trình hình thành màng sinh
vật trên rễ cây Arabidopsis, chủng vi khuẩn Bacillus subtilis 6051 đã tạo ra surfactin
ức chế sự xâm nhiễm của chủng P. syringae gây hại cây trồng.
Khả năng đối kháng của chủng Bacillus thuringiensis chống lại tác nhân gây
bệnh cây trồng Erwinia carotovora đã được chứng minh qua nghiên cứu của
Morikiwa [56]. E. carotovora sản xuất các phân tử tín hiệu cảm ứng mật độ tế bào
acyl - HSL và biểu hiện gen gây độc, trong khi đó các chủng B. thuringiensis có
enzyme acyl - homoserine lactonase, làm giảm mạnh acyl - HSL. Do vậy, B.
thuringiensis làm giảm đáng kể khả năng nhiễm và phát triển của E. carotovora -
tác nhân gây bệnh thối củ ở khoai tây.
1.5.3 Một số nghiên cứu ứng dụng khác
Ứng dụng của màng sinh vật làm giảm sự ăn mòn kim loại
Vi khuẩn gây ra sự ăn mòn kim loại được biết đến là các chủng vi khuẩn khử
sulfate Desulfosporosinus orientis và vi khuẩn oxy hóa sắt Leptothrix discophora
SP-6. D. orientis gây ra sự ăn mòn ở gang, thép cacbon, thép không gỉ và một số
hợp kim khác. Hằng năm, thiệt hại do sự ăn mòn kim loại gây ra ở Mỹ là 4 - 6 tỷ
USD [34]. Vi khuẩn này có chứa enzyme hydrogenase sử dụng hydrogen như một
chất cho điện tử để thu nhận năng lượng gây ra hiện tượng ăn mòn điện hóa ở các
bề mặt kim loại. Trong khi đó, vi khuẩn L. discophora SP-6 oxy hóa sắt, tự nó
không gây ra sự ăn mòn đáng kể thép nhẹ (thép ít cacbon), nhưng khi có sự kết hợp

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 27
với D. orientis và Paenibacillus polymyxa 10401 thì tốc độ ăn mòn thép nhẹ sẽ tăng
lên rất lớn [28]. Nghiên cứu của Morikawa [56] cho thấy sự hình thành màng sinh
vật ở chủng Bacillus brevis 18-3 đã tạo ra gramicidin làm giảm đáng kể tốc độ ăn
mòn kim loại bằng cách ức chế cả hai chủng vi khuẩn D. orientis và L. discophora
SP-6.
Trong công nghiệp lên men
Trong các bồn lên men ở quy mô công nghiệp, màng sinh vật là hình thức
hiệu quả để giữ lại sinh khối vi sinh vật [47].
Sau mỗi mẻ lên men, các tế bào ở dạng tự do khó có khả năng giữ lại trong
các bồn lên men. Do đó, mỗi khi tiếp tục một quy trình mới lại phải bổ sung thêm
một lượng sinh khối nhất định và đợi thời gian để vi sinh vật có thể sinh trưởng,
phát triển tới một nồng độ nhất định mới. Quy trình này gây tốn kém ở khâu nguyên
liệu đầu vào cũng như mất thời gian vận hành.
Ngược lại, khi đã được bám giữ trên bề mặt giá thể bằng mạng lưới màng
sinh vật sinh khối vi sinh vật có thể được giữ lại một cách có hiệu quả sau mỗi mẻ
xử lý cũng như tái sử dụng được ở những lần xử lý tiếp theo mà không cần phải bổ
sung thêm vi sinh vật cũng như đợi thời gian phát triển.
Ứng dụng trong công nghiệp dầu khí
Nghiên cứu khả năng tổng hợp chất hoạt động bề mặt sinh học
(biosurfactant) hoạt tính cao từ chủng vi khuẩn Pseudomonas aeruginosa, được
phân lập từ nguồn nước thải nhiễm dầu [67] mở ra triển vọng ứng dụng trong công
nghiệp dầu khí với các mục đích như nâng cao hệ số thu hồi dầu, xử lý môi trường,
làm chất phân tán và nhũ hóa cặn dầu.
Nghiên cứu về màng sinh vật đã và đang là lĩnh vực thu hút sự quan tâm của
rất nhiều nhà khoa học, giúp chúng ta hiểu rõ hơn về tác hại cũng như lợi ích ứng
dụng của màng sinh vật trong đời sống. Tuy nhiên, các nghiên cứu chủ yếu thu nhận
được từ các công trình khoa học nước ngoài. Tài liệu về màng sinh vật ở Việt Nam
còn tương đối ít và nghiên cứu về màng sinh vật còn chưa nhiều, chưa sâu. Đề tài
nghiên cứu của chúng tôi với mục đích tạo thêm cơ sở dữ liệu khoa học về các

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 28
chủng vi sinh vật có khả năng tạo thành màng sinh vật từ môi trường nước thải giàu
nguồn cacbon ở các khu vực làng nghề và nhà máy sản xuất và đưa ra một số ứng
dụng theo hướng xử lý nước thải môi trường.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 29
Chương 2. NGUYÊN LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1 Nguyên liệu
2.1.1 Chủng vi sinh vật nghiên cứu
Với mục tiêu phân lập, nghiên cứu đặc điểm sinh học của các chủng vi sinh
vật có khả năng tạo thành màng sinh vật và ứng dụng trong xử lý nước thải cũng
như ức chế các vi sinh vật gây hại, chúng tôi đã tiến hành lựa chọn và lấy mẫu nước
thải tại một số khu vực ô nhiễm làng nghề ở Việt Nam. Mẫu được lấy vào thời điểm
các buổi sáng trong ngày, chứa trong các ống falcon đã khử trùng rồi đem về phòng
thí nghiệm để phân tích. Cụ thể, tại các địa điểm sau:
Thứ nhất, nguồn nước thải tại làng miến Lại Trạch - Yên Phú - Yên Mỹ -
Hưng Yên: Hệ thống thoát nước và hồ lắng của làng miến chứa lượng nước thải lớn
lẫn cặn bột và bã bột dong. Mẫu nước thải được lấy tại các hồ lắng này.
Thứ hai, nguồn nước thải tại làng bún Phú Đô - Mễ Trì - Từ Liêm - Hà Nội:
Toàn bộ nước thải của quá trình làm bún được thải trực tiếp ra các cống, rãnh của cả
làng. Do vậy, khu vực nước thải làng nghề nơi chúng tôi lấy mẫu có một màu trắng
đục, chua và hôi.
Thứ ba, nguồn nước thải tại nhà máy sản xuất bia - Viện Công nghiệp thực
phẩm - Thanh Xuân - Hà Nội: Nguồn nước thải được sử dụng là nước thải của
xưởng bia - Viện Công nghiệp Thực phẩm đã được tách cặn thông qua bể lắng sơ
bộ. Nguồn nước thải có thành phần COD 2500 - 3000 (mg/l), BOD5 1500 - 2000
(mg/l), pH 4,5 - 5,5. Chỉ số COD cao chủ yếu là do trong thành phần nước thải có
chứa nhiều tinh bột và các chất hữu cơ.
2.1.2 Vi sinh vật kiểm định
Các vi sinh vật kiểm định được sử dụng trong đề tài bao gồm:
Staphylococcus aureus; Ralstonia solanacaerum; Samonella typhi; E. coli; Vibrio
parahaemolyticus do bộ môn Vi sinh thuộc trường Đại học Khoa học Tự nhiên -
Đại học Quốc gia Hà Nội phối hợp cung cấp.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 30
2.2 Hóa chất, thiết bị và dụng cụ thí nghiệm
2.2.1 Môi trường nuôi cấy
Môi trường LB (Luria - Bertani broth hay Luria broth):
Tryptone 10g
Cao nấm men 5g
NaCl 10g
Nước cất 1 lít
Môi trường LB là môi trường thường được sử dụng trong việc nuôi cấy các
chủng vi khuẩn hiếu khí. Môi trường có đầy đủ các chất dinh dưỡng nguồn cacbon,
nitơ cho vi sinh vật sinh trưởng và phát triển.
Môi trường khoáng cơ bản:
(NH4)2SO4 2g
MgSO4.7H2O 0,2g
NaH2PO4.H2O 0,5g
CaCl2.2H2O 0,1g
K2HPO4 0,5g
Nước cất 1 lít
Môi trường khoáng cơ sở:
KH2PO4 1,36g
CaCl2 0,03g
Na2HPO4 2,13g
MgSO4.7H2O 0,2g
FeSO4.7H2O 0,01g
Glucose 10g
Nước cất 1 lít

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 31
Các môi trường đều được khử trùng ở nhiệt độ 121oC trong thời gian 120
phút. Các hóa chất khác sử dụng trong nghiên cứu, đánh giá khả năng tạo màng sinh
vật như các loại đường Fructose (Sigma - Mỹ); Arabinose (Merk - Đức), Mannose
(Merk - Đức), NaCl (Công ty cổ phần Hóa chất Đức Giang)… đều đạt độ tinh sạch
cho mục đích nghiên cứu.
2.2.2 Máy móc, thiết bị
Nồi khử trùng (ALP - Nhật Bản)
Máy lắc ổn nhiệt (Satorius - Đức)
Tủ cấy vi sinh vật (Aura vertical - Ý)
Cân điện tử 2 số lẻ (Kern - Đức)
Cân phân tích (Presica - Thụy Sỹ)
Máy đo pH (Horiba - Nhật Bản)
Máy đo mật độ quang học (Bionate - Anh)
Tủ ấm (Memmert - Đức)
Tủ sấy (Memmert - Đức)
Máy khuấy từ gia nhiệt (IKA RET - Đức)
Kính hiển vi điện tử quét JSM - 5421LV (Nhật)
2.3 Phương pháp nghiên cứu
2.3.1 Phương pháp phân lập vi khuẩn
Vi khuẩn được phân lập từ các mẫu nước thải, sử dụng phương pháp pha
loãng mẫu trong nước cất vô trùng.
Mẫu sau khi pha loãng được cấy gạt dịch ở các nồng độ từ 10-1, 10-2 đến 10-5
trên các đĩa petri chứa môi trường LB - thạch. Các đĩa sau khi cấy trải được nuôi
cấy trong tủ ấm với nhiệt độ 37oC trong thời gian 24 giờ. Các khuẩn lạc xuất hiện
và được quan sát sau thời gian ít nhất là 24 giờ. Các khuẩn lạc sau khi phát triển sẽ
được tách riêng rẽ và được nuôi cấy trên môi trường phù hợp hoặc LB có bổ sung
thạch.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 32
2.3.2 Phương pháp bảo quản giống vi sinh vật
Phương pháp giữ giống trên môi trường thạch nghiêng
Sau khi phân lập riêng rẽ trên các đĩa petri chứa môi trường LB - thạch, các
vi sinh vật được cấy truyền trên môi trường thạch nghiêng thích hợp đã được khử
trùng.
Các ống thạch nghiêng sau khi cấy được nuôi trong tủ ấm với nhiệt độ 37oC
trong thời gian 24 giờ. Các ống giống sau khi phát triển sẽ được lấy ra và cho vào tủ
lạnh giữ ở 4oC. Hàng tháng, các ống giống được lấy ra và cấy truyền lại vào môi
trường mới.
Phương pháp bảo quản giống lâu dài
Các chủng vi sinh vật sau khi phân lập riêng rẽ được bảo quản lâu dài trong
môi trường glycerol theo tỷ lệ 1:1 ở nhiệt độ lạnh sâu -80oC.
2.3.3 Phương pháp nghiên cứu đánh giá khả năng tạo màng sinh vật của các
chủng vi sinh vật
Trong môi trường dinh dưỡng thích hợp và điều kiện nuôi cấy tĩnh, một số
chủng vi sinh vật có khả năng tạo thành màng sinh vật trên bề mặt giá thể. Việc phát
hiện, quan sát sự tạo thành màng sinh vật được thực hiện bằng phương pháp nhuộm
màu với dung dịch tím kết tinh 1% (w/v). Tím kết tinh (crystal violet hay Gentian
violet) là một thuốc nhuộm hóa học triarylmethane có công thức phân tử là
C25H30N3Cl, công thức cấu tạo là [C(C6H4N(CH3)2)3]Cl - Tris (4-(dimethylamino)
phenyl) methylium chloride. Dung dịch tím kết tinh 1% có khả năng bắt màu với
các tế bào sống, sự thay đổi về cường độ màu thể hiện mức độ và số lượng của các
tế bào vi sinh vật.
Thí nghiệm được tiến hành theo phương pháp của O’Toole và cộng sự [62]
Khuẩn lạc các chủng vi sinh vật sau khi tách riêng rẽ sẽ được nuôi cấy kích
hoạt trong bình tam giác chứa 10ml môi trường LB lỏng trong 24 giờ ở 37oC sao
cho mật độ tế bào OD620 ở vào khoảng 0,3 - 0,4. Hút 100l dịch nuôi cấy vi khuẩn

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 33
bổ sung vào 700l LB lỏng trong các ống eppendorf đã khử trùng và ủ trong điều
kiện tĩnh ở 37oC.
Sau 24 giờ, các dịch nuôi cấy được loại bỏ khỏi các ống eppendorf. Đánh giá
mật độ tế bào sống trôi nổi trong môi trường bằng phương pháp đo mật độ quang
học ở bước sóng 620 nm (OD620) dịch nuôi cấy vi khuẩn.
Phương pháp quan sát khả năng tạo thành màng sinh vật:
Mỗi ống eppendorf được rửa sạch 2 lần bằng nước cất khử trùng. Sau đó mỗi
ống eppendorf được bổ sung 1ml dung dịch tím kết tinh 1% và giữ trong 20 phút ở
nhiệt độ phòng. Dung dịch nhuộm tím kết tinh sau đó được loại bỏ, rửa sạch 2 lần
bằng nước cất và quan sát sự bắt màu của các tế bào bám trên thành ống với tím kết
tinh.
Mật độ tế bào trong màng sinh vật tạo ra được đánh giá bằng độ hấp thụ ánh
sáng ở bước sóng 570 nm theo phương pháp của Morikawa và cộng sự [57]: Sau
khi rửa sạch 2 lần bằng nước cất, các tinh thể tím bám trên thành eppendorf được
hòa tan trong 1ml etanol 70o. Mật độ tế bào trong màng sinh vật được xác định bằng
cách đo độ hấp thụ OD 570 nm.
2.3.4 Tối ưu hóa các điều kiện tạo màng sinh vật
Mỗi chủng vi sinh vật tùy theo đặc tính sinh học của chúng chỉ có thể tạo
thành màng sinh vật tốt nhất ở những điều kiện thích hợp về nhiệt độ, pH, các
nguồn carbon, nitơ.
2.3.4.1 Ảnh hưởng của nhiệt độ môi trường
Các chủng vi sinh vật sau khi được lựa chọn, có hoạt tính tạo màng sinh vật
sẽ được nghiên cứu về khả năng phát triển trong các điều kiện nhiệt độ môi trường
khác nhau. Các nhiệt độ được lựa chọn nghiên cứu là: 20oC, 30oC, 37oC, 40oC, 50oC
và 60oC. Sau 24 giờ nuôi cấy, tiến hành quan sát, đánh giá khả năng tạo thành màng
sinh vật của các chủng vi sinh vật nghiên cứu bằng cách đo độ hấp thụ OD 570 nm
theo phương pháp của Morikawa và cộng sự [57].

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 34
2.3.4.2 Ảnh hưởng của pH môi trường nuôi cấy
Các chủng vi sinh vật sau khi được lựa chọn, có hoạt tính tạo màng sinh vật
sẽ được nghiên cứu về khả năng phát triển trong các điều kiện pH môi trường khác
nhau. Các giá trị pH môi trường khác nhau được lựa chọn nghiên cứu là: pH 4; 4,5;
5; 5,5; 6; 6,5; 7; 7,5; 8; 8,5. Sau 24 giờ nuôi cấy, mật độ tế bào trong màng sinh vật
tạo ra được đánh giá bằng cách đo độ hấp thụ ánh sáng OD 570 nm theo phương
pháp của Morikawa và cộng sự [57].
2.3.4.3 Ảnh hưởng của nồng độ cacon trong môi trường
Bổ sung các nguồn cacbon vào môi trường khoáng cơ bản với nồng độ 1%.
Các nguồn carbon được sử dụng trong thí nghiệm bao gồm:
Đường 5 cacbon: Arabinose, Fucose, Rhamnose
Đường 6 cacbon: Glucose, Mannose, Fructose, Galactose
Đường đôi: Saccharose, Lactose
Polysaccarit : Tinh bột
Khử trùng môi trường ở nhiệt độ 121oC trong 20 phút
Các chủng vi sinh vật sau khi được lựa chọn, có hoạt tính tạo màng sinh vật
sẽ được nuôi cấy trong môi trường khoáng cơ bản có bổ sung các nguồn cacbon
khác nhau, điều kiện tĩnh, ở 37oC. Sau 24 giờ nuôi cấy, mật độ tế bào trong màng
sinh vật tạo ra được đánh giá bằng cách đo độ hấp thụ ánh sáng OD 570 nm theo
phương pháp của Morikawa và cộng sự [57].
2.3.4.4 Ảnh hưởng của nguồn nitơ trong môi trường
Bổ sung các nguồn nitơ khác nhau vào môi trường khoáng cơ sở với nồng độ
0,1%. Chỉnh pH tới 7,0 - 7,2.
Các nguồn nitơ được sử dụng trong thí nghiệm bao gồm:
(NH4)2SO4, NaNO3, KNO3, (NH4)2C6H5O7, Pepton, Cao nấm men.
Khử trùng môi trường ở nhiệt độ 121oC trong 20 phút

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 35
Các chủng vi sinh vật sau khi được lựa chọn, có hoạt tính tạo màng sinh vật
sẽ được nuôi cấy trong môi trường khoáng cơ sở có bổ sung các nguồn nitơ khác
nhau với điều kiện tĩnh, ở 37oC. Sau 24 giờ nuôi cấy, mật độ tế bào trong màng sinh
vật tạo ra được đánh giá bằng cách đo độ hấp thụ ánh sáng OD 570 nm theo phương
pháp của Morikawa và cộng sự [57].
2.3.5 Phương pháp đánh giá khả năng tạo chất hoạt động bề mặt
Khả năng tạo thành chất hoạt động bề mặt của các chủng vi sinh vật được
đánh giá bằng mức độ nhũ tương hóa dung dịch dầu ăn của dịch nuôi cấy tế bào
thông qua chỉ số E24 (emulsion index) theo phương pháp của Suwansukho và cộng
sự [80].
Các chủng vi sinh vật sau khi được lựa chọn, có hoạt tính tạo màng sinh vật
sẽ được nuôi cấy trong môi trường LB lỏng, lắc với tốc độ 160 vòng/phút trong 24
giờ ở nhiệt độ 37oC. Sau đó, dịch nuôi cấy lắc được ly tâm với tốc độ 10.000
vòng/phút trong 10 phút để loại bỏ cặn tế bào, thu lấy phần dịch nổi.
Bổ sung 2ml dầu ăn vào 2ml dịch vi khuẩn thu được sau khi ly tâm trong các
ống nghiệm nhỏ, đường kính 1cm.
Vortex với tốc độ cao trong 1 phút
Sau 24 giờ, lấy ra đo mức độ nhũ tương hóa.
Chỉ số nhũ tương hóa E24 được tính theo công thức sau:
E24 = [(chiều cao cột nhũ tương hóa)/(tổng chiều cao cột)] × 100% [80].
2.3.6 Phương pháp đánh giá khả năng kháng khuẩn
Khả năng kháng khuẩn của các chủng vi sinh vật thử nghiệm đối với các
chủng vi khuẩn kiểm định được xác định theo phương pháp của De Angelis và cộng
sự [21].
Các chủng vi sinh vật (bao gồm các chủng vi sinh vật đã phân lập được thử
nghiệm khả năng kháng khuẩn và các chủng vi sinh vật kiểm định) được nuôi cấy
qua đêm trong môi trường LB lỏng trên máy lắc với tốc độ 160 vòng/phút, ở 37oC.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 36
Sau đó, dịch nuôi cấy vi sinh vật thử nghiệm được ly tâm ở 10.000 vòng/phút
trong 10 phút để loại bỏ hoàn toàn tế bào vi khuẩn. Dịch lắc của vi khuẩn kiểm định
được cấy trải trên các đĩa môi trường LB - thạch với thể tích 100l. Các lỗ được đục
với đường kính 6mm trên các đĩa thạch.
Dịch lọc của từng chủng vi sinh vật thử nghiệm được cho vào các lỗ thạch
với thể tích 30l. Các đĩa thạch được ủ qua đêm ở 37oC. Quan sát và chụp ảnh trên
các đĩa thạch.
Khả năng kháng khuẩn của các chủng vi sinh vật thử nghiệm được xác định
dựa vào đường kính vòng kháng khuẩn (D) xuất hiện xung quanh lỗ thạch.
D = D - d
D: đường kính vòng vô khuẩn (mm)
d: đường kính lỗ thạch (mm).
2.3.7 Phương pháp nhuộm Gram
Nguyên tắc:
Dựa vào sự khác biệt giữa thành tế bào vi khuẩn Gram (+) và Gram (-). Vi
khuẩn Gram (+) có peptidoglican hoạt động như một hàng rào thẩm thấu ngăn cản
sự thất thoát của tím kết tinh. Ban đầu, vi khuẩn được nhuộm bằng tím kết tinh sau
đó được xử lý bằng iot để tăng độ giữ màu. Sau đó được tẩy màu bằng cồn làm co
các lỗ của lớp peptidoglican dày lại. Do vậy phức chất tím kết tinh và iot được giữ
lại, vi khuẩn có màu tím. Peptidoglican ở vi khuẩn Gram (-) rất mỏng, ít liên kết
chéo và có lỗ lớn. Do vậy, ở bước rửa bằng cồn đã loại bỏ phức chất màu tím của
tím kết tinh - iot. Khi nhuộm lại bằng safranin thì vi khuẩn có màu hồng [3].
Tiến hành:
Chuẩn bị vết bôi: dùng que cấy vô trùng lấy một ít vi khuẩn từ đĩa thạch (sau
khi cấy 24 giờ) hòa vào 1 giọt nước cất ở giữa phiến kính, làm khô trong không khí.
Cố định tế bào: hơ nhanh vết bôi trên ngọn lửa đèn cồn 2 - 3 lần.
Nhuộm bằng dung dịch tím kết tinh trong 1 phút, rửa nước, thấm khô.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 37
Nhuộm lại bằng dung dịch Lugol (1% iot, 2% KI) trong 1 phút, rửa nước,
thấm khô.
Nhỏ dịch tẩy màu (ethanol 95%), giữ khoảng 30 giây, rửa nước, thấm khô.
Nhuộm bổ sung bằng dung dịch Safranin trong 2 - 3 phút, rửa nước, để khô
trong không khí.
Quan sát dưới kính hiển vi: dùng vật kính dầu với độ phóng đại 100 lần [3].
2.3.8 Quan sát cấu trúc màng sinh vật bằng ảnh chụp trên kính hiển vi điện tử
quét
Chuẩn bị mẫu màng sinh vật nổi: dịch nuôi cấy lắc vi khuẩn phân lập có khả
năng tạo màng sinh vật tốt được bổ sung vào bình tam giác chứa 20ml môi trường
LB lỏng. Nuôi cấy tĩnh trong 24 giờ, ở 37oC.
Mẫu màng sinh vật được gắn lên lamelle, hơ nhẹ trên ngọn lửa đèn cồn để cố
định mẫu.
Rửa nhẹ mẫu gắn màng sinh vật bằng nước cất khử trùng, để khô tự nhiên.
Gắn mẫu lên đế
Mạ phủ mẫu bằng vàng trên máy JFC - 1200 trong 5 phút ở 30mA
Quan sát và chụp ảnh trên kính hiển vi điện tử quét JSM - 5421LV (Nhật) tại
phòng chụp hiển vi điện tử quét thuộc Trung tâm Khoa học Vật liệu - Khoa Vật Lý
- Trường Đại học Khoa học Tự nhiên - Đại học Quốc gia Hà Nội.
2.3.9 Phương pháp phân loại phân tử dựa trên gen 16S rDNA
Phương pháp này dựa trên phương pháp Sanger có cải tiến: Dựa vào sự tổng
hợp mạch bổ sung cho trình tự cần xác định nhờ DNA - polymerase. Với việc sử
dụng thêm các ddNTP cùng các dNTP thông thường, kết quả là sự hình thành một
tập hợp nhiều đoạn DNA có kích thước khác nhau.
Vectơ tái tổ hợp có chứa đoạn gen 16S rRNA mang hai trình tự chuyên biệt,
mỗi trình tự nằm trên một mạch. Khi cần xác định trình tự của mạch nào, trước hết

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 38
phải biến tính tách rời 2 mạch rồi sử dụng mồi bắt cặp với trình tự chuyên biệt nằm
trên mạch đó.
Mỗi dideoxynucleotit được đánh dấu bằng một hóa chất huỳnh quang có màu
khác nhau. Mỗi thuốc nhuộm huỳnh quang phát ra một phổ ánh sáng hẹp với một
đỉnh khác biệt. Các số liệu phát ra được lưu ghi lại trong máy tính. Sau khi quá trình
chạy điện di kết thúc, chuỗi các tín hiệu huỳnh quang được chuyển ngược thành
thông tin trình tự nucleotit [71].
Kết quả được xử lý trên phần mềm PC/Gene, Gendoc, BioEdit và được so
sánh với trình tự đã được công bố trên ngân hàng dữ liệu gen quốc tế bằng chương
trình BLAST (Basic Local Aignment Search Tool) qua dữ liệu của DDGJ EMBL để
tìm xem mức độ tương đồng của các chủng nghiên cứu với loài nào là lớn nhất [2].
Kết quả đọc trình tự 16S rDNA của các chủng vi sinh vật có khả năng tạo
màng sinh vật mạnh được thực hiện với sự giúp đỡ của Viện Vi sinh vật và Công
nghệ Sinh học - Đại học Quốc gia Hà Nội.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 39
Chương 3. KẾT QUẢ VÀ THẢO LUẬN
3.1 Phân lập, tuyển chọn các chủng vi sinh vật có khả năng tạo màng sinh vật
3.1.1 Phân lập vi sinh vật
Từ mẫu nước thải khu vực làng nghề miến Lại Trạch, chúng tôi phân lập
được 23 chủng vi sinh vật, số lượng các chủng lần lượt thu được từ phần nước thải
tầng mặt là 6 chủng, phần giữa cách tầng mặt 30cm là 11 chủng và phần mặt đáy là
6 chủng, số chủng phân lập từ phần giữa chiếm 47,83%.
Phân lập mẫu nước thải nhà máy sản xuất bia chúng tôi thu được 6 chủng vi
sinh vật ở tầng mặt, 8 chủng từ khu vực phần giữa cách tầng mặt 30cm và phần mặt
đáy là 11 chủng, số chủng phân lập được từ phần mặt đáy chiếm 44%.
Từ mẫu nước thải làng bún Phú Đô chúng tôi phân lập được 27 chủng vi sinh
vật với số lượng chủng hiện diện ở tầng mặt là 5 chủng, khu vực nước phần giữa
cách mặt nước 15cm là 12 chủng và phần mặt đáy là 10 chủng, số chủng phân lập
được từ phần giữa chiếm 44,44%.
Hình 6. Khuẩn lạc một số chủng vi sinh vật phân lập trên môi trường thạch
Kết quả thu được cho phép chúng tôi nhận định: phần lớn các chủng vi sinh
vật phân lập được từ các mẫu nước thải tập trung ở khu vực nước phần giữa và phần
mặt đáy. Các địa điểm lấy mẫu lại chủ yếu là những khu vực nước thải chứa hàm
lượng cacbon cao. Điều này phù hợp với giải thích về khả năng bám dính của các
chủng vi sinh vật ở các phần nước này. Đây là phần nước thải ít có sự biến động và
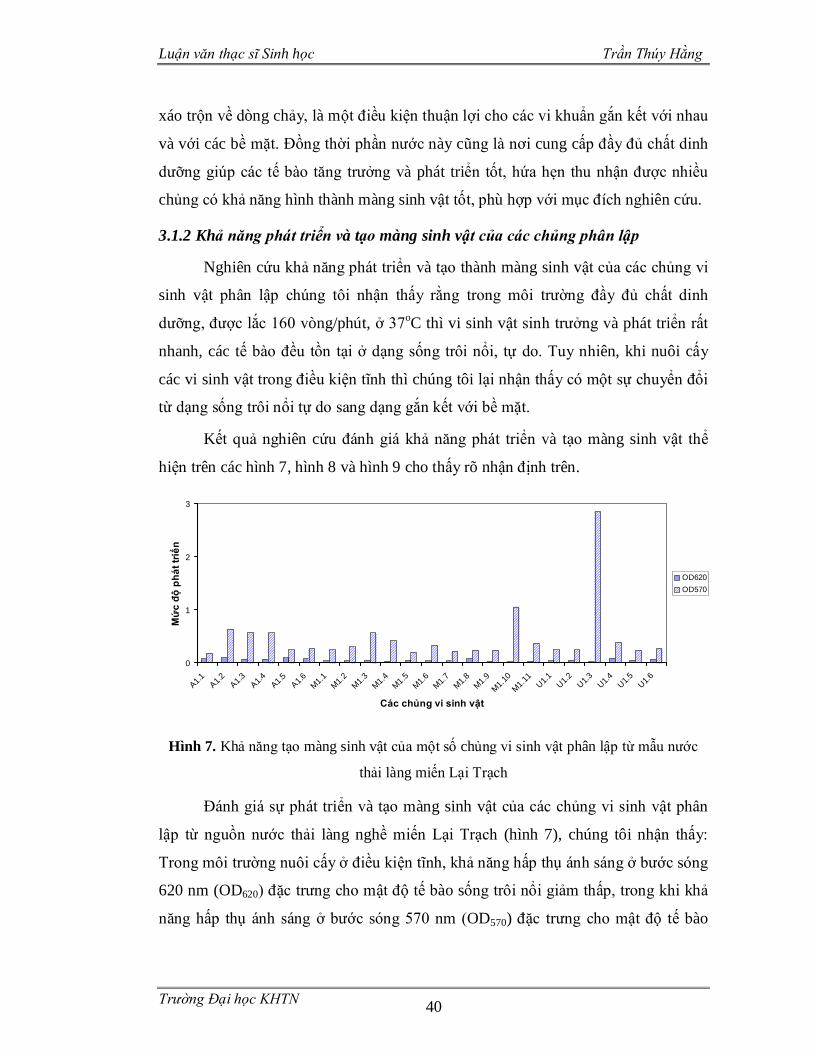
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 40
xáo trộn về dòng chảy, là một điều kiện thuận lợi cho các vi khuẩn gắn kết với nhau
và với các bề mặt. Đồng thời phần nước này cũng là nơi cung cấp đầy đủ chất dinh
dưỡng giúp các tế bào tăng trưởng và phát triển tốt, hứa hẹn thu nhận được nhiều
chủng có khả năng hình thành màng sinh vật tốt, phù hợp với mục đích nghiên cứu.
3.1.2 Khả năng phát triển và tạo màng sinh vật của các chủng phân lập
Nghiên cứu khả năng phát triển và tạo thành màng sinh vật của các chủng vi
sinh vật phân lập chúng tôi nhận thấy rằng trong môi trường đầy đủ chất dinh
dưỡng, được lắc 160 vòng/phút, ở 37oC thì vi sinh vật sinh trưởng và phát triển rất
nhanh, các tế bào đều tồn tại ở dạng sống trôi nổi, tự do. Tuy nhiên, khi nuôi cấy
các vi sinh vật trong điều kiện tĩnh thì chúng tôi lại nhận thấy có một sự chuyển đổi
từ dạng sống trôi nổi tự do sang dạng gắn kết với bề mặt.
Kết quả nghiên cứu đánh giá khả năng phát triển và tạo màng sinh vật thể
hiện trên các hình 7, hình 8 và hình 9 cho thấy rõ nhận định trên.
0
1
2
3
A1.1A1.2A1.3 A1.4 A1.5A1.6 M1.1 M1.2 M1.3 M1.4 M1.5M1.6M1.7 M1.8 M1.9
M1.10
M1.11
U1.1 U1.2 U1.3 U1.4 U1.5 U1.6
Các chủng vi sinh vật
Mứ
c độ
phá
t triể
n
OD620OD570
Hình 7. Khả năng tạo màng sinh vật của một số chủng vi sinh vật phân lập từ mẫu nước
thải làng miến Lại Trạch
Đánh giá sự phát triển và tạo màng sinh vật của các chủng vi sinh vật phân
lập từ nguồn nước thải làng nghề miến Lại Trạch (hình 7), chúng tôi nhận thấy:
Trong môi trường nuôi cấy ở điều kiện tĩnh, khả năng hấp thụ ánh sáng ở bước sóng
620 nm (OD620) đặc trưng cho mật độ tế bào sống trôi nổi giảm thấp, trong khi khả
năng hấp thụ ánh sáng ở bước sóng 570 nm (OD570) đặc trưng cho mật độ tế bào
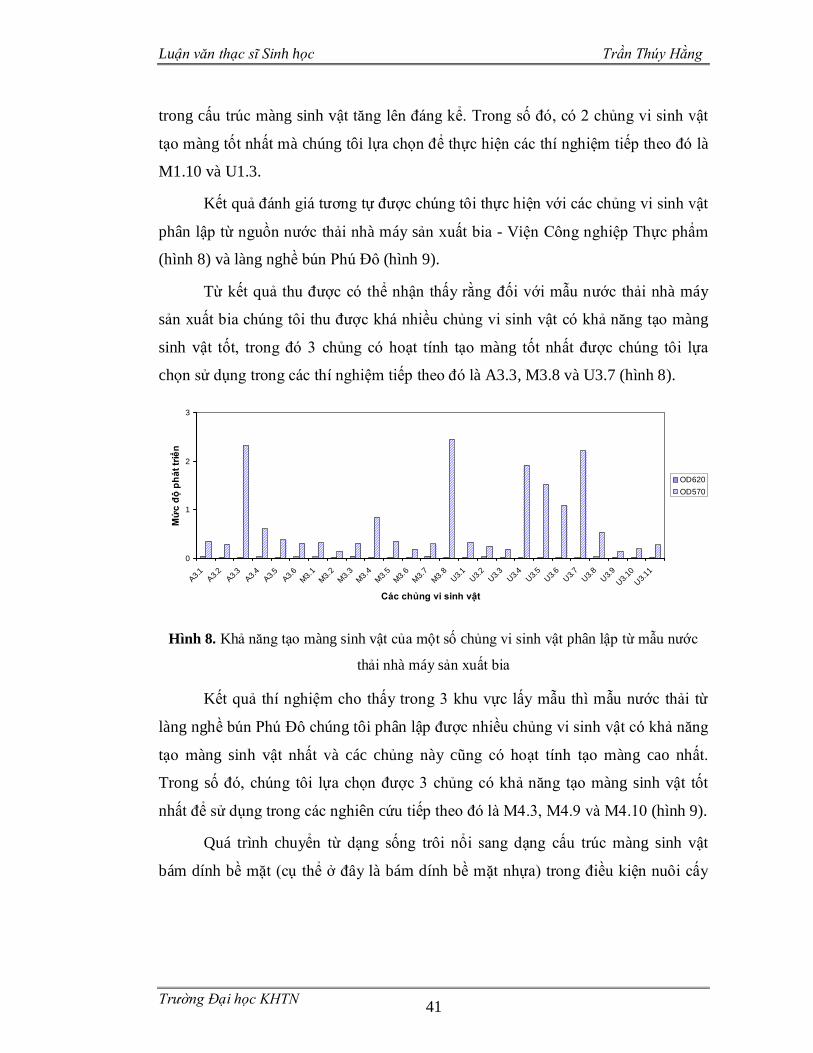
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 41
trong cấu trúc màng sinh vật tăng lên đáng kể. Trong số đó, có 2 chủng vi sinh vật
tạo màng tốt nhất mà chúng tôi lựa chọn để thực hiện các thí nghiệm tiếp theo đó là
M1.10 và U1.3.
Kết quả đánh giá tương tự được chúng tôi thực hiện với các chủng vi sinh vật
phân lập từ nguồn nước thải nhà máy sản xuất bia - Viện Công nghiệp Thực phẩm
(hình 8) và làng nghề bún Phú Đô (hình 9).
Từ kết quả thu được có thể nhận thấy rằng đối với mẫu nước thải nhà máy
sản xuất bia chúng tôi thu được khá nhiều chủng vi sinh vật có khả năng tạo màng
sinh vật tốt, trong đó 3 chủng có hoạt tính tạo màng tốt nhất được chúng tôi lựa
chọn sử dụng trong các thí nghiệm tiếp theo đó là A3.3, M3.8 và U3.7 (hình 8).
0
1
2
3
A3.1 A3.2 A3.3 A3.4 A3.5 A3.6 M3.1 M3.2 M3.3 M3.4 M3.5 M3.6 M3.7 M3.8 U3.1 U3.2 U3.3 U3.4 U3.5 U3.6 U3.7 U3.8 U3.9U3.1
0U3.1
1
Các chủng vi sinh vật
Mứ
c độ
phá
t triể
n
OD620OD570
Hình 8. Khả năng tạo màng sinh vật của một số chủng vi sinh vật phân lập từ mẫu nước
thải nhà máy sản xuất bia
Kết quả thí nghiệm cho thấy trong 3 khu vực lấy mẫu thì mẫu nước thải từ
làng nghề bún Phú Đô chúng tôi phân lập được nhiều chủng vi sinh vật có khả năng
tạo màng sinh vật nhất và các chủng này cũng có hoạt tính tạo màng cao nhất.
Trong số đó, chúng tôi lựa chọn được 3 chủng có khả năng tạo màng sinh vật tốt
nhất để sử dụng trong các nghiên cứu tiếp theo đó là M4.3, M4.9 và M4.10 (hình 9).
Quá trình chuyển từ dạng sống trôi nổi sang dạng cấu trúc màng sinh vật
bám dính bề mặt (cụ thể ở đây là bám dính bề mặt nhựa) trong điều kiện nuôi cấy

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 42
0
1
2
3
4
A4.1 A4.2 A4.3 A4.4 A4.5 M4.1 M4.2 M4.3 M4.4 M4.5 M4.6 M4.7 M4.8 M4.9M4.1
0M4.1
1M4.12 U4.1 U4.2 U4.3 U4.4 U4.5 U4.6 U4.7 U4.8 U4.9U4.1
0
Các chủng vi sinh vật
Mứ
c độ
phá
t triể
n
OD620OD570
Hình 9. Khả năng tạo màng sinh vật của một số chủng vi sinh vật phân lập từ mẫu nước
thải làng nghề bún Phú Đô
tĩnh đã chứng tỏ đây là một kiểu phản ứng thích nghi với sự thay đổi về điều kiện
sống của vi sinh vật. Khi sống trong một điều kiện không thuận lợi cho sự tăng
trưởng tự do, các tế bào sống có xu hướng gắn kết nhau trên một bề mặt hữu sinh
hoặc vô sinh thông qua mạng lưới chất ngoại bào để có thể trao đổi vật chất và
thông tin với nhau, kết thành một tổ chức vững chắc nhằm đáp ứng với những hạn
chế của môi trường, thông qua đó giúp chúng tồn tại và phát triển.
Như vậy, trong số các chủng vi sinh vật phân lập được từ các khu vực nước
thải giàu nguồn cacbon chúng tôi đã lựa chọn được 8 chủng có hoạt tính tạo màng
sinh vật mạnh nhất cho các nghiên cứu tiếp theo. Đó là các chủng được ký hiệu lần
lượt là M1.10, U1.3, A3.3, M3.8, U3.7, M4.3, M4.9, M4.10. Các chủng này hầu hết
đều được phân lập từ mẫu nước thải khu vực phần giữa và mặt đáy. Điều này phù
hợp với nhận định ban đầu về khả năng bám dính của vi sinh vật với hoạt tính tạo
màng sinh vật của chúng.
3.2 Các yếu tố ảnh hưởng đến sự tạo màng sinh vật của các chủng phân lập
3.2.1 Ảnh hưởng của nhiệt độ
Khảo sát sự tạo thành màng sinh vật của các chủng vi sinh vật nghiên cứu
với các điều kiện nhiệt độ khác nhau để tìm ra khoảng nhiệt độ thích hợp nhất,
chúng tôi thu được kết quả như sau:
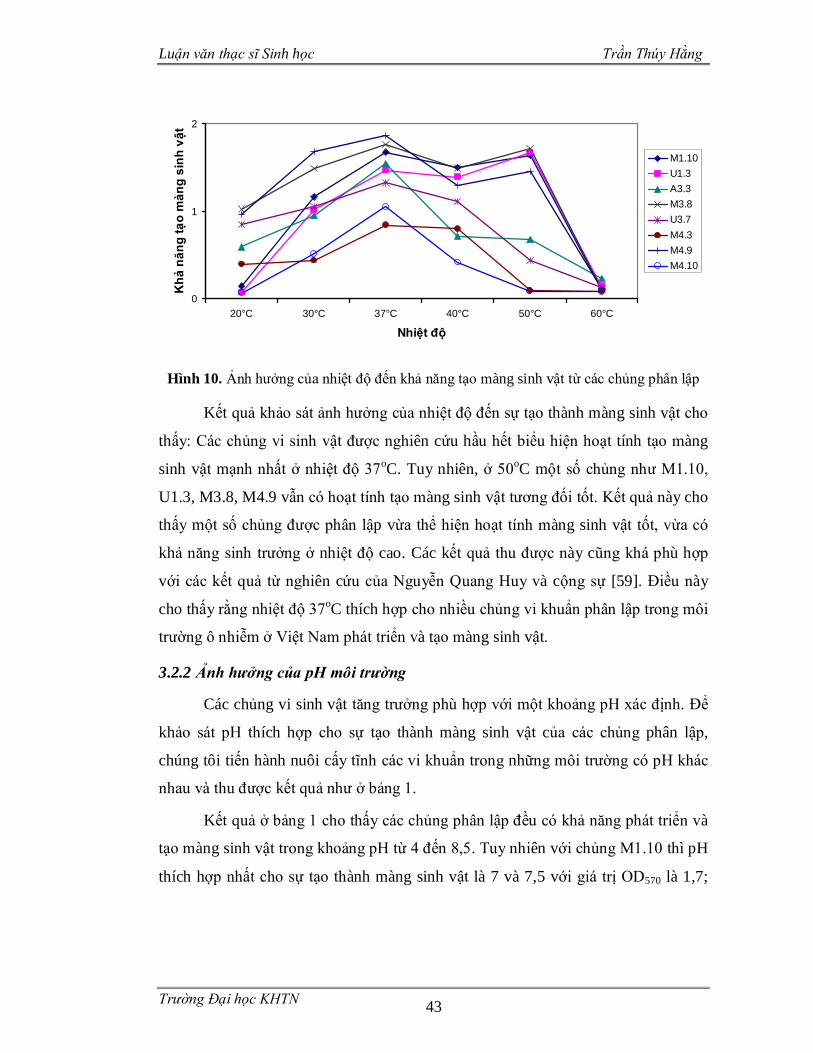
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 43
0
1
2
20°C 30°C 37°C 40°C 50°C 60°C
Nhiệt độ
Khả
năn
g tạ
o m
àng
sinh
vật
M1.10U1.3A3.3M3.8U3.7M4.3M4.9M4.10
Hình 10. Ảnh hưởng của nhiệt độ đến khả năng tạo màng sinh vật từ các chủng phân lập
Kết quả khảo sát ảnh hưởng của nhiệt độ đến sự tạo thành màng sinh vật cho
thấy: Các chủng vi sinh vật được nghiên cứu hầu hết biểu hiện hoạt tính tạo màng
sinh vật mạnh nhất ở nhiệt độ 37oC. Tuy nhiên, ở 50oC một số chủng như M1.10,
U1.3, M3.8, M4.9 vẫn có hoạt tính tạo màng sinh vật tương đối tốt. Kết quả này cho
thấy một số chủng được phân lập vừa thể hiện hoạt tính màng sinh vật tốt, vừa có
khả năng sinh trưởng ở nhiệt độ cao. Các kết quả thu được này cũng khá phù hợp
với các kết quả từ nghiên cứu của Nguyễn Quang Huy và cộng sự [59]. Điều này
cho thấy rằng nhiệt độ 37oC thích hợp cho nhiều chủng vi khuẩn phân lập trong môi
trường ô nhiễm ở Việt Nam phát triển và tạo màng sinh vật.
3.2.2 Ảnh hưởng của pH môi trường
Các chủng vi sinh vật tăng trưởng phù hợp với một khoảng pH xác định. Để
khảo sát pH thích hợp cho sự tạo thành màng sinh vật của các chủng phân lập,
chúng tôi tiến hành nuôi cấy tĩnh các vi khuẩn trong những môi trường có pH khác
nhau và thu được kết quả như ở bảng 1.
Kết quả ở bảng 1 cho thấy các chủng phân lập đều có khả năng phát triển và
tạo màng sinh vật trong khoảng pH từ 4 đến 8,5. Tuy nhiên với chủng M1.10 thì pH
thích hợp nhất cho sự tạo thành màng sinh vật là 7 và 7,5 với giá trị OD570 là 1,7;
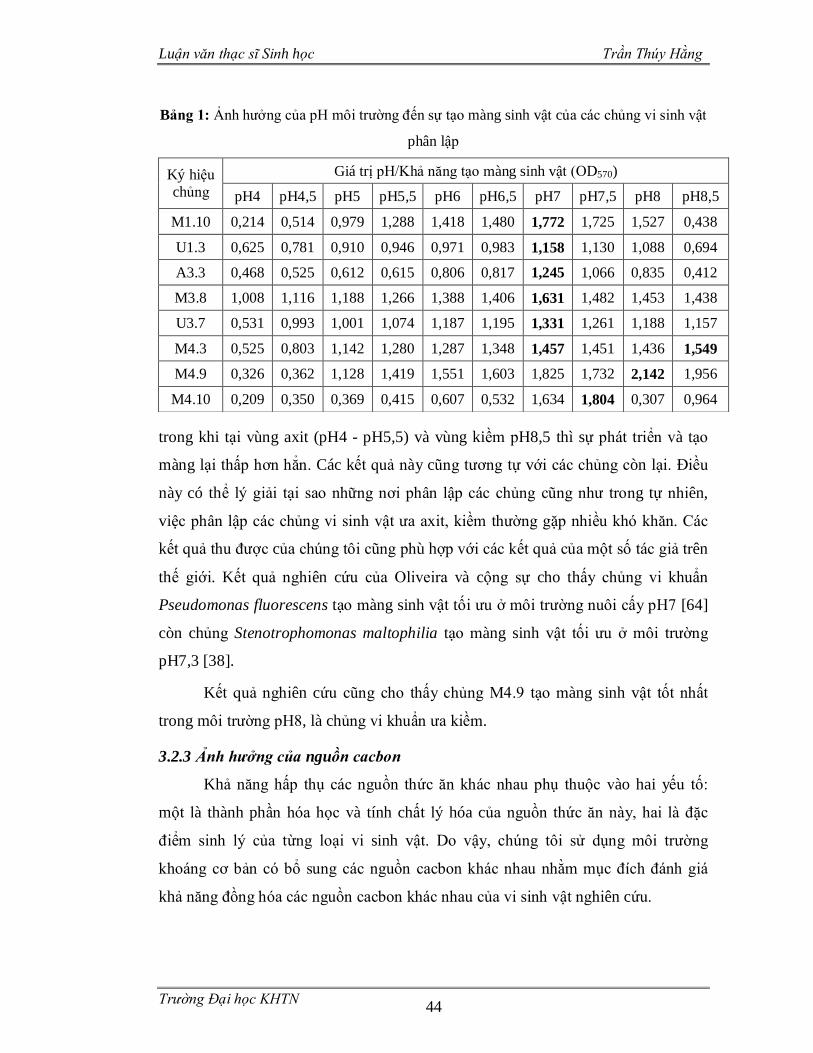
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 44
Bảng 1: Ảnh hưởng của pH môi trường đến sự tạo màng sinh vật của các chủng vi sinh vật
phân lập
Giá trị pH/Khả năng tạo màng sinh vật (OD570) Ký hiệu chủng pH4 pH4,5 pH5 pH5,5 pH6 pH6,5 pH7 pH7,5 pH8 pH8,5
M1.10 0,214 0,514 0,979 1,288 1,418 1,480 1,772 1,725 1,527 0,438
U1.3 0,625 0,781 0,910 0,946 0,971 0,983 1,158 1,130 1,088 0,694
A3.3 0,468 0,525 0,612 0,615 0,806 0,817 1,245 1,066 0,835 0,412
M3.8 1,008 1,116 1,188 1,266 1,388 1,406 1,631 1,482 1,453 1,438
U3.7 0,531 0,993 1,001 1,074 1,187 1,195 1,331 1,261 1,188 1,157
M4.3 0,525 0,803 1,142 1,280 1,287 1,348 1,457 1,451 1,436 1,549
M4.9 0,326 0,362 1,128 1,419 1,551 1,603 1,825 1,732 2,142 1,956
M4.10 0,209 0,350 0,369 0,415 0,607 0,532 1,634 1,804 0,307 0,964
trong khi tại vùng axit (pH4 - pH5,5) và vùng kiềm pH8,5 thì sự phát triển và tạo
màng lại thấp hơn hẳn. Các kết quả này cũng tương tự với các chủng còn lại. Điều
này có thể lý giải tại sao những nơi phân lập các chủng cũng như trong tự nhiên,
việc phân lập các chủng vi sinh vật ưa axit, kiềm thường gặp nhiều khó khăn. Các
kết quả thu được của chúng tôi cũng phù hợp với các kết quả của một số tác giả trên
thế giới. Kết quả nghiên cứu của Oliveira và cộng sự cho thấy chủng vi khuẩn
Pseudomonas fluorescens tạo màng sinh vật tối ưu ở môi trường nuôi cấy pH7 [64]
còn chủng Stenotrophomonas maltophilia tạo màng sinh vật tối ưu ở môi trường
pH7,3 [38].
Kết quả nghiên cứu cũng cho thấy chủng M4.9 tạo màng sinh vật tốt nhất
trong môi trường pH8, là chủng vi khuẩn ưa kiềm.
3.2.3 Ảnh hưởng của nguồn cacbon
Khả năng hấp thụ các nguồn thức ăn khác nhau phụ thuộc vào hai yếu tố:
một là thành phần hóa học và tính chất lý hóa của nguồn thức ăn này, hai là đặc
điểm sinh lý của từng loại vi sinh vật. Do vậy, chúng tôi sử dụng môi trường
khoáng cơ bản có bổ sung các nguồn cacbon khác nhau nhằm mục đích đánh giá
khả năng đồng hóa các nguồn cacbon khác nhau của vi sinh vật nghiên cứu.
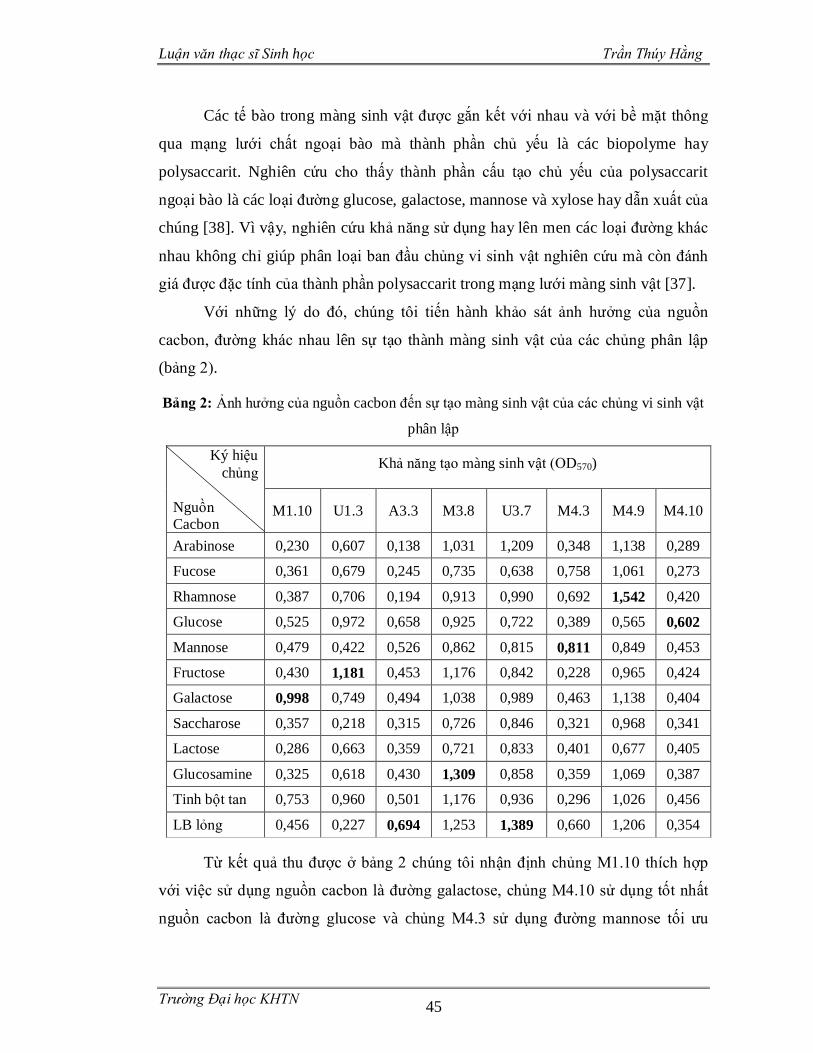
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 45
Các tế bào trong màng sinh vật được gắn kết với nhau và với bề mặt thông
qua mạng lưới chất ngoại bào mà thành phần chủ yếu là các biopolyme hay
polysaccarit. Nghiên cứu cho thấy thành phần cấu tạo chủ yếu của polysaccarit
ngoại bào là các loại đường glucose, galactose, mannose và xylose hay dẫn xuất của
chúng [38]. Vì vậy, nghiên cứu khả năng sử dụng hay lên men các loại đường khác
nhau không chỉ giúp phân loại ban đầu chủng vi sinh vật nghiên cứu mà còn đánh
giá được đặc tính của thành phần polysaccarit trong mạng lưới màng sinh vật [37].
Với những lý do đó, chúng tôi tiến hành khảo sát ảnh hưởng của nguồn
cacbon, đường khác nhau lên sự tạo thành màng sinh vật của các chủng phân lập
(bảng 2).
Bảng 2: Ảnh hưởng của nguồn cacbon đến sự tạo màng sinh vật của các chủng vi sinh vật
phân lập
Khả năng tạo màng sinh vật (OD570) Ký hiệu
chủng Nguồn Cacbon
M1.10 U1.3 A3.3 M3.8 U3.7 M4.3 M4.9 M4.10
Arabinose 0,230 0,607 0,138 1,031 1,209 0,348 1,138 0,289
Fucose 0,361 0,679 0,245 0,735 0,638 0,758 1,061 0,273
Rhamnose 0,387 0,706 0,194 0,913 0,990 0,692 1,542 0,420
Glucose 0,525 0,972 0,658 0,925 0,722 0,389 0,565 0,602
Mannose 0,479 0,422 0,526 0,862 0,815 0,811 0,849 0,453
Fructose 0,430 1,181 0,453 1,176 0,842 0,228 0,965 0,424
Galactose 0,998 0,749 0,494 1,038 0,989 0,463 1,138 0,404
Saccharose 0,357 0,218 0,315 0,726 0,846 0,321 0,968 0,341
Lactose 0,286 0,663 0,359 0,721 0,833 0,401 0,677 0,405
Glucosamine 0,325 0,618 0,430 1,309 0,858 0,359 1,069 0,387
Tinh bột tan 0,753 0,960 0,501 1,176 0,936 0,296 1,026 0,456
LB lỏng 0,456 0,227 0,694 1,253 1,389 0,660 1,206 0,354
Từ kết quả thu được ở bảng 2 chúng tôi nhận định chủng M1.10 thích hợp
với việc sử dụng nguồn cacbon là đường galactose, chủng M4.10 sử dụng tốt nhất
nguồn cacbon là đường glucose và chủng M4.3 sử dụng đường mannose tối ưu

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 46
nhất. Các kết quả này phù hợp với đánh giá của các tác giả trong nước như Nguyễn
Quang Huy và cộng sự [59] hay tác giả nước ngoài như Haggag [38]. Tuy nhiên khả
năng tạo màng sinh vật của các chủng này trong kết quả nghiên cứu là không cao
đối với từng loại đường riêng biệt.
Qua bảng kết quả nghiên cứu, chúng tôi nhận thấy ngoài khả năng sử dụng
các nguồn đường đã được một số tác giả công bố, các chủng vi sinh vật phân lập
của chúng tôi còn có khả năng sử dụng nhiều nguồn khác tối ưu cho việc tạo màng
sinh vật như fructose, rhamnose hay glucosamine. Ngoài ra, trong số 8 chủng vi
sinh vật được lựa chọn nghiên cứu, chúng tôi nhận thấy 4 chủng có hoạt tính tạo
màng sinh vật cao nhất là M3.8, M4.9, U1.3, U3.7; và cả 4 chủng này đều có khả
năng sử dụng tất cả các loại đường cho sự tạo thành màng sinh vật với các mức độ
khác nhau.
3.2.4 Ảnh hưởng của nguồn nitơ
Thành phần chủ yếu của mạng lưới ngoại bào trong cấu trúc màng sinh vật là
polysaccarit, bên cạnh đó còn có các đại phân tử khác như protein, axit nucleic,
glycoprotein, photpholipit. Sự tổng hợp mạng lưới ngoại bào của các tế bào vi
khuẩn phụ thuộc vào nguồn cacbon và nitơ sẵn có trong môi trường nuôi cấy. Các
nguồn nitơ vi sinh vật có thể đồng hóa được là NH3, NH4+, NO3
-, N2 [18]. Tuy
nhiên, trong điều kiện phòng thí nghiệm chúng tôi sử dụng hai nguồn cung cấp nitơ
là muối nitrat và muối amon để khảo sát khả năng đồng hóa các nguồn nitơ khác
nhau của vi sinh vật nghiên cứu.
Tác giả Sleytr [74] đã chỉ ra rằng nồng độ nitơ thấp trong môi trường nuôi
cấy làm tăng khả năng hấp thụ đường và ảnh hưởng đáng kể đến sự tạo thành mạng
lưới ngoại bào giúp cho sự tạo thành màng sinh vật được thuận lợi.
Kết quả bảng nghiên cứu số 3 (bảng 3) cho thấy một số chủng như U1.3,
M3.8, M4.9, M4.10 tạo màng sinh vật tương đối tốt với nguồn nitơ từ muối
(NH4)2SO4. Kết quả này tương đối phù hợp với nghiên cứu của Lee và cộng sự [51]
cho thấy chủng Aureobasidium pullulans tạo màng sinh vật tốt với nguồn nitơ từ
muối (NH4)2SO4. Bên cạnh đó, kết quả nghiên cứu cũng cho thấy cao nấm men là
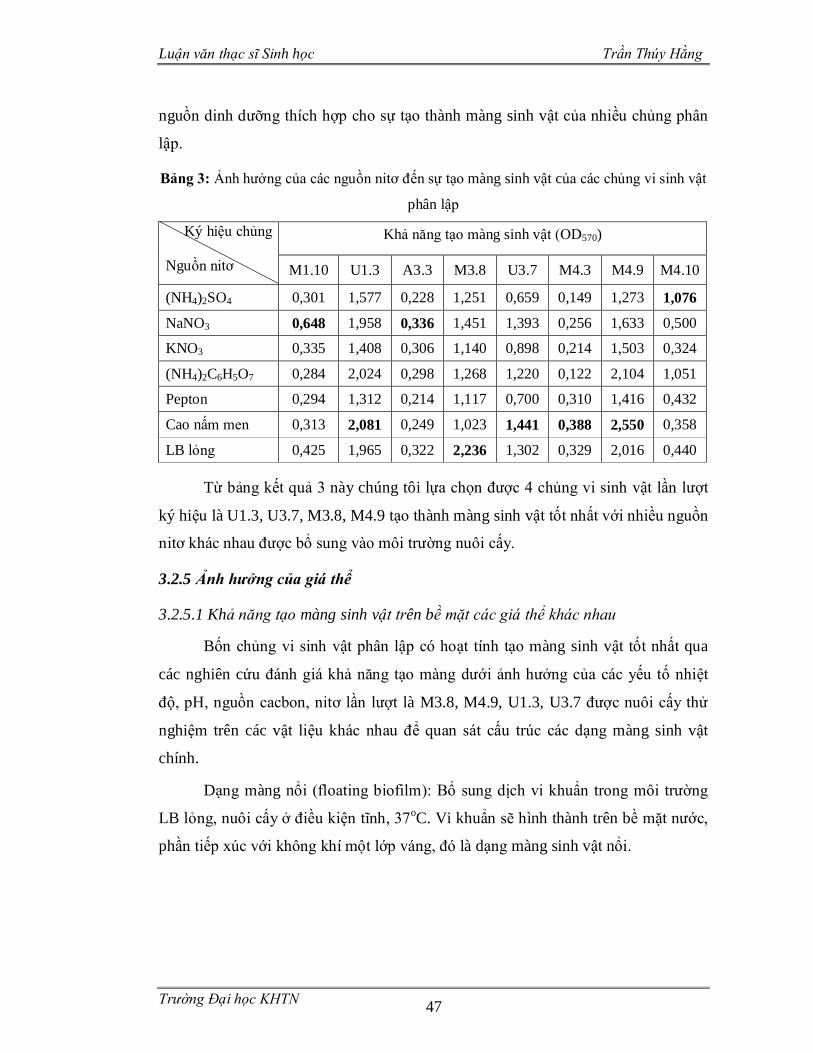
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 47
nguồn dinh dưỡng thích hợp cho sự tạo thành màng sinh vật của nhiều chủng phân
lập.
Bảng 3: Ảnh hưởng của các nguồn nitơ đến sự tạo màng sinh vật của các chủng vi sinh vật
phân lập
Khả năng tạo màng sinh vật (OD570) Ký hiệu chủng Nguồn nitơ M1.10 U1.3 A3.3 M3.8 U3.7 M4.3 M4.9 M4.10
(NH4)2SO4 0,301 1,577 0,228 1,251 0,659 0,149 1,273 1,076
NaNO3 0,648 1,958 0,336 1,451 1,393 0,256 1,633 0,500
KNO3 0,335 1,408 0,306 1,140 0,898 0,214 1,503 0,324
(NH4)2C6H5O7 0,284 2,024 0,298 1,268 1,220 0,122 2,104 1,051
Pepton 0,294 1,312 0,214 1,117 0,700 0,310 1,416 0,432
Cao nấm men 0,313 2,081 0,249 1,023 1,441 0,388 2,550 0,358
LB lỏng 0,425 1,965 0,322 2,236 1,302 0,329 2,016 0,440
Từ bảng kết quả 3 này chúng tôi lựa chọn được 4 chủng vi sinh vật lần lượt
ký hiệu là U1.3, U3.7, M3.8, M4.9 tạo thành màng sinh vật tốt nhất với nhiều nguồn
nitơ khác nhau được bổ sung vào môi trường nuôi cấy.
3.2.5 Ảnh hưởng của giá thể
3.2.5.1 Khả năng tạo màng sinh vật trên bề mặt các giá thể khác nhau
Bốn chủng vi sinh vật phân lập có hoạt tính tạo màng sinh vật tốt nhất qua
các nghiên cứu đánh giá khả năng tạo màng dưới ảnh hưởng của các yếu tố nhiệt
độ, pH, nguồn cacbon, nitơ lần lượt là M3.8, M4.9, U1.3, U3.7 được nuôi cấy thử
nghiệm trên các vật liệu khác nhau để quan sát cấu trúc các dạng màng sinh vật
chính.
Dạng màng nổi (floating biofilm): Bổ sung dịch vi khuẩn trong môi trường
LB lỏng, nuôi cấy ở điều kiện tĩnh, 37oC. Vi khuẩn sẽ hình thành trên bề mặt nước,
phần tiếp xúc với không khí một lớp váng, đó là dạng màng sinh vật nổi.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 48
Hình 11 cho thấy màng nổi của các chủng vi khuẩn. Sau 1 ngày nuôi cấy,
xuất hiện lớp màng mỏng, màu trắng, dạng váng nổi trên môi trường LB lỏng. Lớp
màng nổi dày thêm sau 2, 3, 4 ngày nuôi cấy và đạt tối đa sau 5 ngày nuôi cấy.
Hình 11. Màng nổi của 4 chủng vi sinh vật sau 5 ngày nuôi cấy
M3.8 M4.9 U1.3 U3.7 ĐC
Hình 12. Màng sinh vật trên bề mặt nhựa
Dạng màng sinh vật bám dính bề mặt: là dạng màng sinh vật được hình
thành trên bề mặt vật rắn. Bằng cách thực hiện các thí nghiệm nuôi cấy vi khuẩn
trên các bề mặt vật liệu khác nhau như nhựa, thủy tinh. chúng tôi nhận thấy rằng
khả năng bám dính và hình thành màng sinh vật trên bề mặt nhựa là tốt nhất (hình
12).
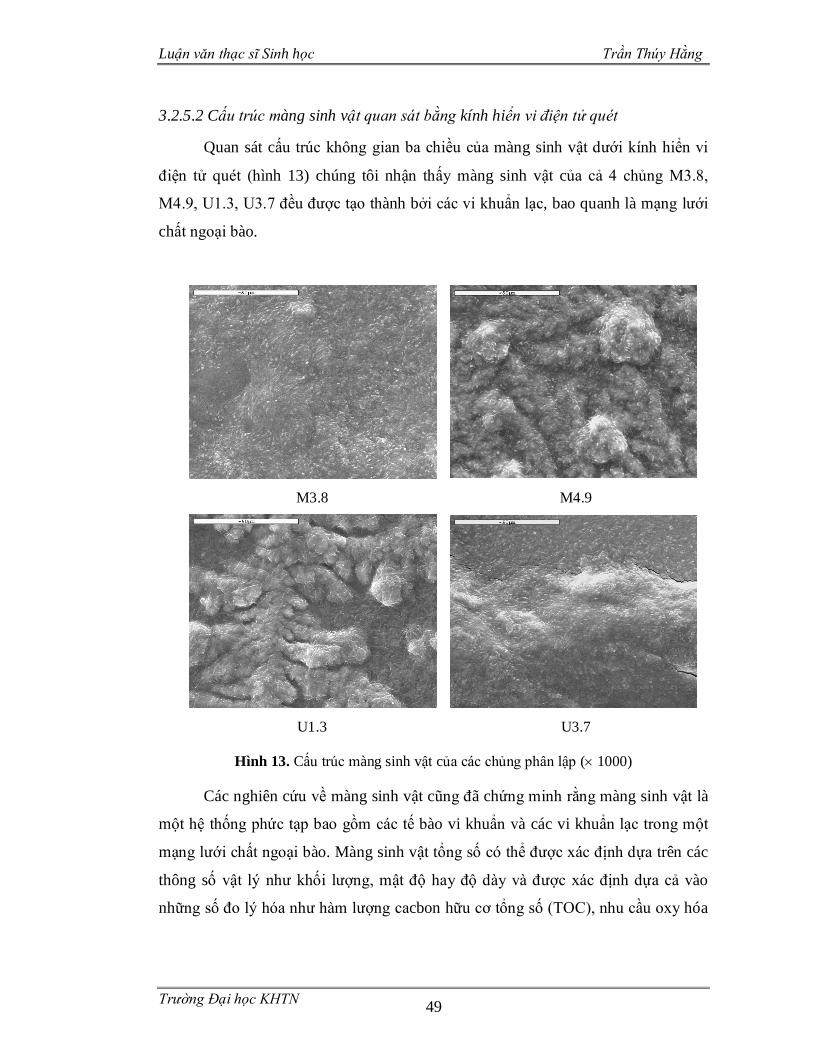
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 49
3.2.5.2 Cấu trúc màng sinh vật quan sát bằng kính hiển vi điện tử quét
Quan sát cấu trúc không gian ba chiều của màng sinh vật dưới kính hiển vi
điện tử quét (hình 13) chúng tôi nhận thấy màng sinh vật của cả 4 chủng M3.8,
M4.9, U1.3, U3.7 đều được tạo thành bởi các vi khuẩn lạc, bao quanh là mạng lưới
chất ngoại bào.
M3.8 M4.9
U1.3 U3.7
Hình 13. Cấu trúc màng sinh vật của các chủng phân lập ( 1000)
Các nghiên cứu về màng sinh vật cũng đã chứng minh rằng màng sinh vật là
một hệ thống phức tạp bao gồm các tế bào vi khuẩn và các vi khuẩn lạc trong một
mạng lưới chất ngoại bào. Màng sinh vật tổng số có thể được xác định dựa trên các
thông số vật lý như khối lượng, mật độ hay độ dày và được xác định dựa cả vào
những số đo lý hóa như hàm lượng cacbon hữu cơ tổng số (TOC), nhu cầu oxy hóa

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 50
học (COD). Cấu tạo của màng sinh vật được mô tả chi tiết thông qua phương pháp
đo các thành phần cụ thể như hàm lượng polysaccarit ngoại bào, protein, tổng số tế
bào hay các thành phần tế bào khác như peptidoglycan, lipopolysaccarit, lipit [5].
Polysaccarit cũng được chứng minh chiếm trên 65% thành phần của chất nền
ngoại bào, ngoài ra còn có một số các hợp chất khác như protein, axit nucleic, lipit,
glycoprotein, glycoside hay glucophotphat. Vì vậy, có khi mạng lưới chất ngoại bào
còn có cái tên khác là “biopolymer” hay “polysaccarit” [5].
3.3 Một số đặc tính sinh học và phân loại các chủng vi sinh vật phân lập
3.3.1 Khả năng tạo chất hoạt động bề mặt
Chất hoạt động bề mặt sinh học (biosurfactant) có vai trò quan trọng trong
công nghiệp thực phẩm như khả năng nhũ tương hóa và tạo bọt trong chế biến thực
phẩm, tạo nhũ hóa cho các sản phẩm mỹ phẩm [80]. Ngoài ra, chất hoạt động bề
mặt còn được ứng dụng trong công nghệ môi trường như xử lý các sự cố tràn dầu
[6].
Chất hoạt động bề mặt thuộc 4 nhóm glycolipit, photpholipit, lipoprotein và
lipopeptit. Trong đó, chất hoạt động bề mặt do chủng vi khuẩn Bacillus subtilis tạo
ra là surfactin thuộc nhóm lipopeptit có tính chất hoạt động bề mặt và khả năng
kháng khuẩn cao [80].
Nhằm mục đích đánh giá khả năng tạo chất hoạt động bề mặt của các chủng
vi sinh vật phân lập, chúng tôi tiến hành nghiên cứu khả năng nhũ tương hóa dầu ăn
của chúng và thu được kết quả như ở hình 14.
Kết quả nhũ tương hóa dầu ăn cho thấy một số chủng vi sinh vật có hoạt tính
hình thành màng sinh vật mạnh đồng thời cũng có khả năng tạo thành chất hoạt
động bề mặt. Chất hoạt động bề mặt sinh học do vi sinh vật tạo ra là chất có thành
phần cấu trúc đa dạng với bề mặt rất hoạt động. Bởi vậy, có một mối liên hệ giữa sự
hình thành màng sinh vật và khả năng tạo chất hoạt động bề mặt. Nhiều nghiên cứu
đã chỉ ra rằng các phân tử chất hoạt động bề mặt có thể tạo điều kiện thuận lợi cho
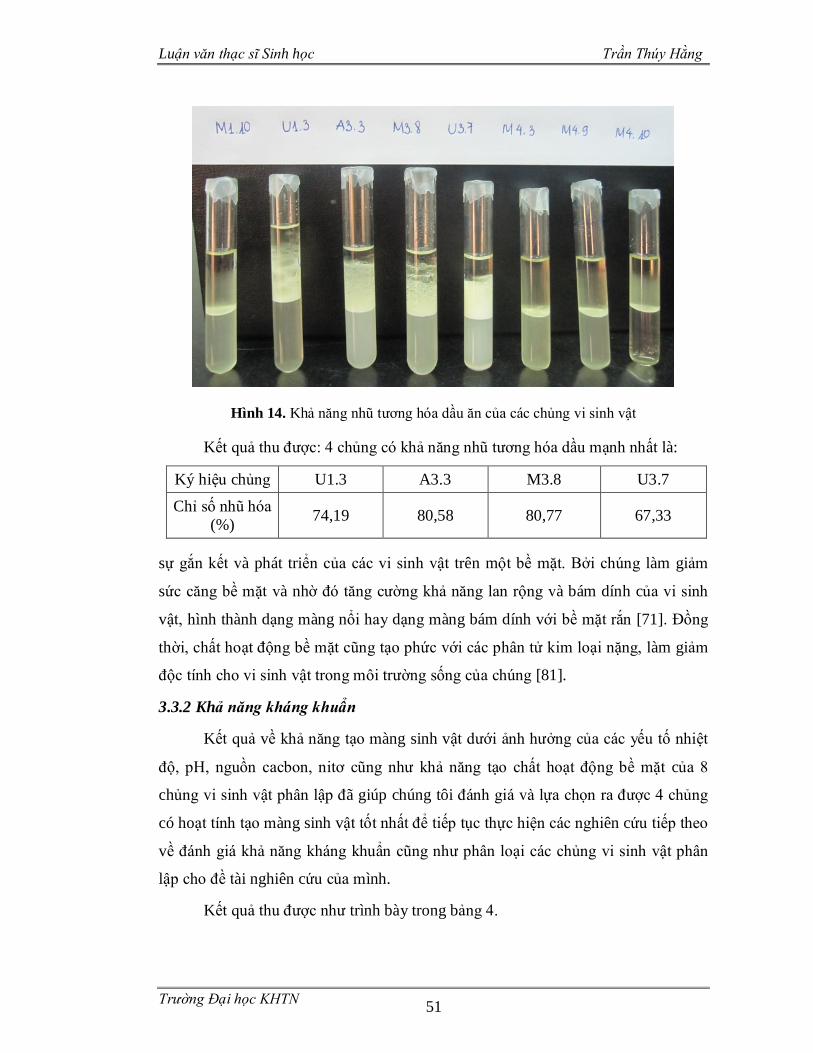
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 51
Hình 14. Khả năng nhũ tương hóa dầu ăn của các chủng vi sinh vật
Kết quả thu được: 4 chủng có khả năng nhũ tương hóa dầu mạnh nhất là:
Ký hiệu chủng U1.3 A3.3 M3.8 U3.7
Chỉ số nhũ hóa (%) 74,19 80,58 80,77 67,33
sự gắn kết và phát triển của các vi sinh vật trên một bề mặt. Bởi chúng làm giảm
sức căng bề mặt và nhờ đó tăng cường khả năng lan rộng và bám dính của vi sinh
vật, hình thành dạng màng nổi hay dạng màng bám dính với bề mặt rắn [71]. Đồng
thời, chất hoạt động bề mặt cũng tạo phức với các phân tử kim loại nặng, làm giảm
độc tính cho vi sinh vật trong môi trường sống của chúng [81].
3.3.2 Khả năng kháng khuẩn
Kết quả về khả năng tạo màng sinh vật dưới ảnh hưởng của các yếu tố nhiệt
độ, pH, nguồn cacbon, nitơ cũng như khả năng tạo chất hoạt động bề mặt của 8
chủng vi sinh vật phân lập đã giúp chúng tôi đánh giá và lựa chọn ra được 4 chủng
có hoạt tính tạo màng sinh vật tốt nhất để tiếp tục thực hiện các nghiên cứu tiếp theo
về đánh giá khả năng kháng khuẩn cũng như phân loại các chủng vi sinh vật phân
lập cho đề tài nghiên cứu của mình.
Kết quả thu được như trình bày trong bảng 4.
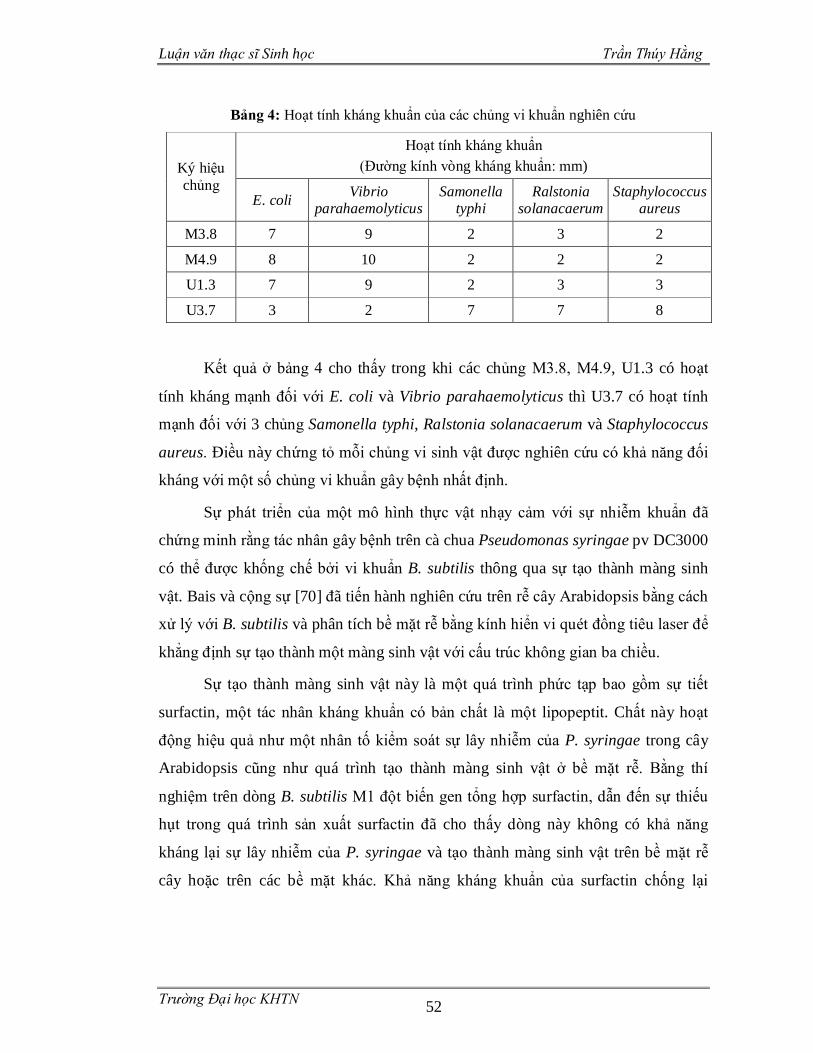
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 52
Bảng 4: Hoạt tính kháng khuẩn của các chủng vi khuẩn nghiên cứu
Hoạt tính kháng khuẩn (Đường kính vòng kháng khuẩn: mm) Ký hiệu
chủng E. coli Vibrio
parahaemolyticus Samonella
typhi Ralstonia
solanacaerum Staphylococcus
aureus
M3.8 7 9 2 3 2
M4.9 8 10 2 2 2
U1.3 7 9 2 3 3
U3.7 3 2 7 7 8
Kết quả ở bảng 4 cho thấy trong khi các chủng M3.8, M4.9, U1.3 có hoạt
tính kháng mạnh đối với E. coli và Vibrio parahaemolyticus thì U3.7 có hoạt tính
mạnh đối với 3 chủng Samonella typhi, Ralstonia solanacaerum và Staphylococcus
aureus. Điều này chứng tỏ mỗi chủng vi sinh vật được nghiên cứu có khả năng đối
kháng với một số chủng vi khuẩn gây bệnh nhất định.
Sự phát triển của một mô hình thực vật nhạy cảm với sự nhiễm khuẩn đã
chứng minh rằng tác nhân gây bệnh trên cà chua Pseudomonas syringae pv DC3000
có thể được khống chế bởi vi khuẩn B. subtilis thông qua sự tạo thành màng sinh
vật. Bais và cộng sự [70] đã tiến hành nghiên cứu trên rễ cây Arabidopsis bằng cách
xử lý với B. subtilis và phân tích bề mặt rễ bằng kính hiển vi quét đồng tiêu laser để
khẳng định sự tạo thành một màng sinh vật với cấu trúc không gian ba chiều.
Sự tạo thành màng sinh vật này là một quá trình phức tạp bao gồm sự tiết
surfactin, một tác nhân kháng khuẩn có bản chất là một lipopeptit. Chất này hoạt
động hiệu quả như một nhân tố kiểm soát sự lây nhiễm của P. syringae trong cây
Arabidopsis cũng như quá trình tạo thành màng sinh vật ở bề mặt rễ. Bằng thí
nghiệm trên dòng B. subtilis M1 đột biến gen tổng hợp surfactin, dẫn đến sự thiếu
hụt trong quá trình sản xuất surfactin đã cho thấy dòng này không có khả năng
kháng lại sự lây nhiễm của P. syringae và tạo thành màng sinh vật trên bề mặt rễ
cây hoặc trên các bề mặt khác. Khả năng kháng khuẩn của surfactin chống lại

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 53
P. syringae cũng được xác định trong cả môi trường nuôi cấy dạng dịch và trên bề
mặt thạch.
Nghiên cứu về vai trò của màng sinh vật trong kiểm soát sinh vật dịch bệnh ở
cây trồng [5] cũng cho thấy sự tạo thành màng sinh vật trên vùng rễ cây vừa có tác
dụng như một bể chứa chất dinh dưỡng cho vùng rễ vừa giúp giảm khả năng lây
nhiễm của các mầm bệnh gây hại cho cây trồng.
3.3.3 Đặc điểm hình thái
Để phân loại các chủng phân lập có khả năng tạo thành màng sinh vật mạnh
M3.8, M4.9, U1.3, U3.7, chúng tôi quan sát hình dạng khuẩn lạc và nhuộm Gram
các chủng, kết quả cho thấy (hình 15):
- Chủng M3.8: Khuẩn lạc tròn, lồi, nhẵn, bóng, màu trắng đục, có diềm xung
quanh; đường kính khuẩn lạc 2 - 3 mm, thuộc nhóm vi khuẩn Gram dương.
- Chủng M4.9: Khuẩn lạc tròn, lồi, nhẵn, bóng, màu trắng đục, mọng nước,
thuộc nhóm vi khuẩn Gram dương.
- Chủng U1.3: Khuẩn lạc tròn, không lồi, màu trắng, hơi xốp, thuộc nhóm vi
khuẩn Gram dương.
- Chủng U3.7: Khuẩn lạc tròn, to, màu trắng, bề mặt khuẩn lạc hơi nhăn
nheo, thuộc nhóm vi khuẩn Gram dương.
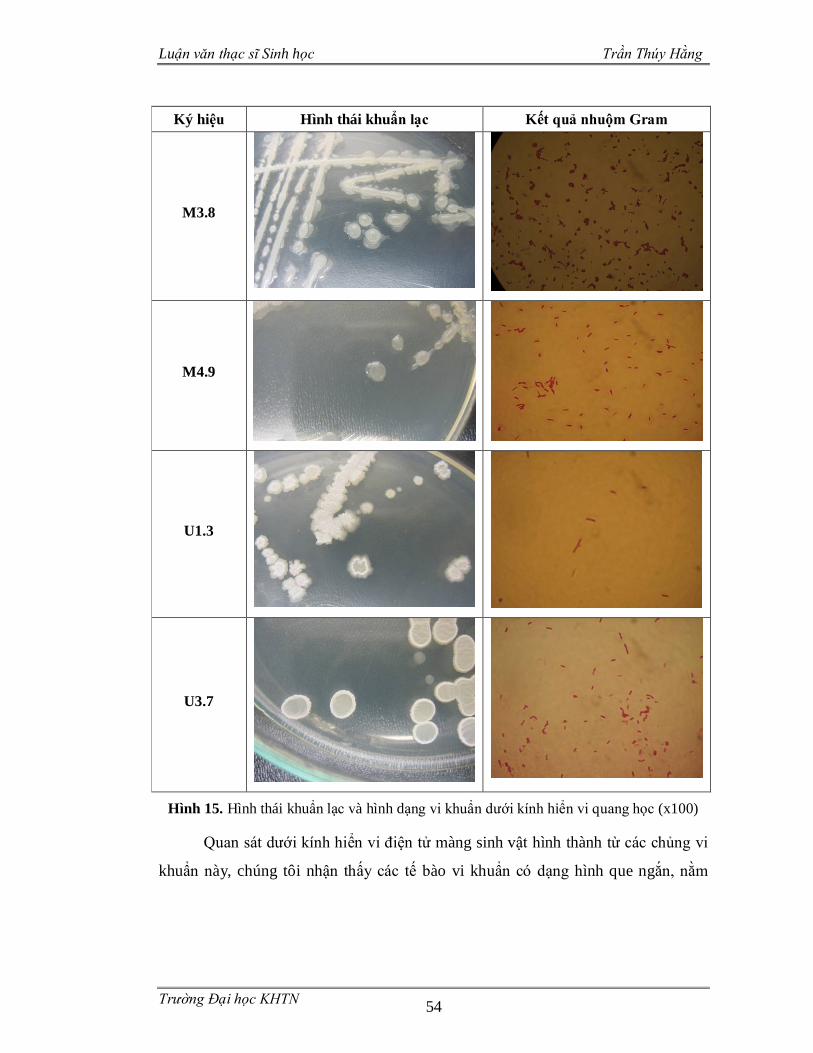
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 54
Ký hiệu Hình thái khuẩn lạc Kết quả nhuộm Gram
M3.8
M4.9
U1.3
U3.7
Hình 15. Hình thái khuẩn lạc và hình dạng vi khuẩn dưới kính hiển vi quang học (x100)
Quan sát dưới kính hiển vi điện tử màng sinh vật hình thành từ các chủng vi
khuẩn này, chúng tôi nhận thấy các tế bào vi khuẩn có dạng hình que ngắn, nằm
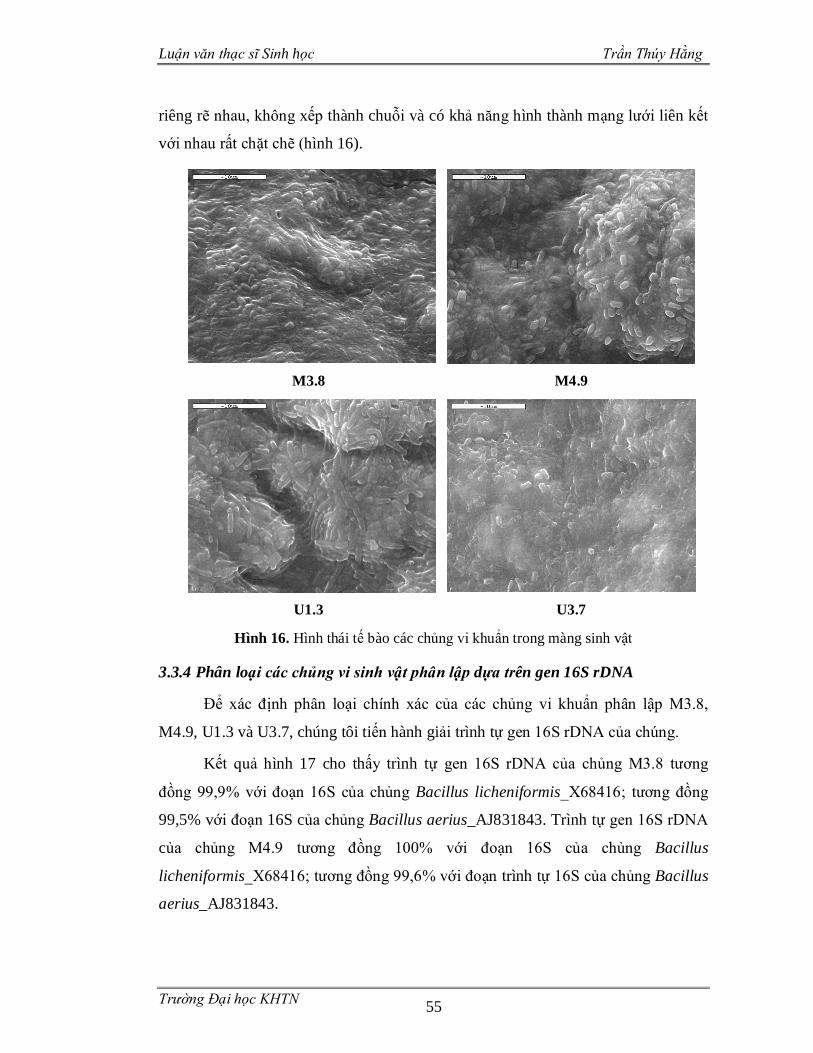
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 55
riêng rẽ nhau, không xếp thành chuỗi và có khả năng hình thành mạng lưới liên kết
với nhau rất chặt chẽ (hình 16).
M3.8 M4.9
U1.3 U3.7
Hình 16. Hình thái tế bào các chủng vi khuẩn trong màng sinh vật
3.3.4 Phân loại các chủng vi sinh vật phân lập dựa trên gen 16S rDNA
Để xác định phân loại chính xác của các chủng vi khuẩn phân lập M3.8,
M4.9, U1.3 và U3.7, chúng tôi tiến hành giải trình tự gen 16S rDNA của chúng.
Kết quả hình 17 cho thấy trình tự gen 16S rDNA của chủng M3.8 tương
đồng 99,9% với đoạn 16S của chủng Bacillus licheniformis_X68416; tương đồng
99,5% với đoạn 16S của chủng Bacillus aerius_AJ831843. Trình tự gen 16S rDNA
của chủng M4.9 tương đồng 100% với đoạn 16S của chủng Bacillus
licheniformis_X68416; tương đồng 99,6% với đoạn trình tự 16S của chủng Bacillus
aerius_AJ831843.
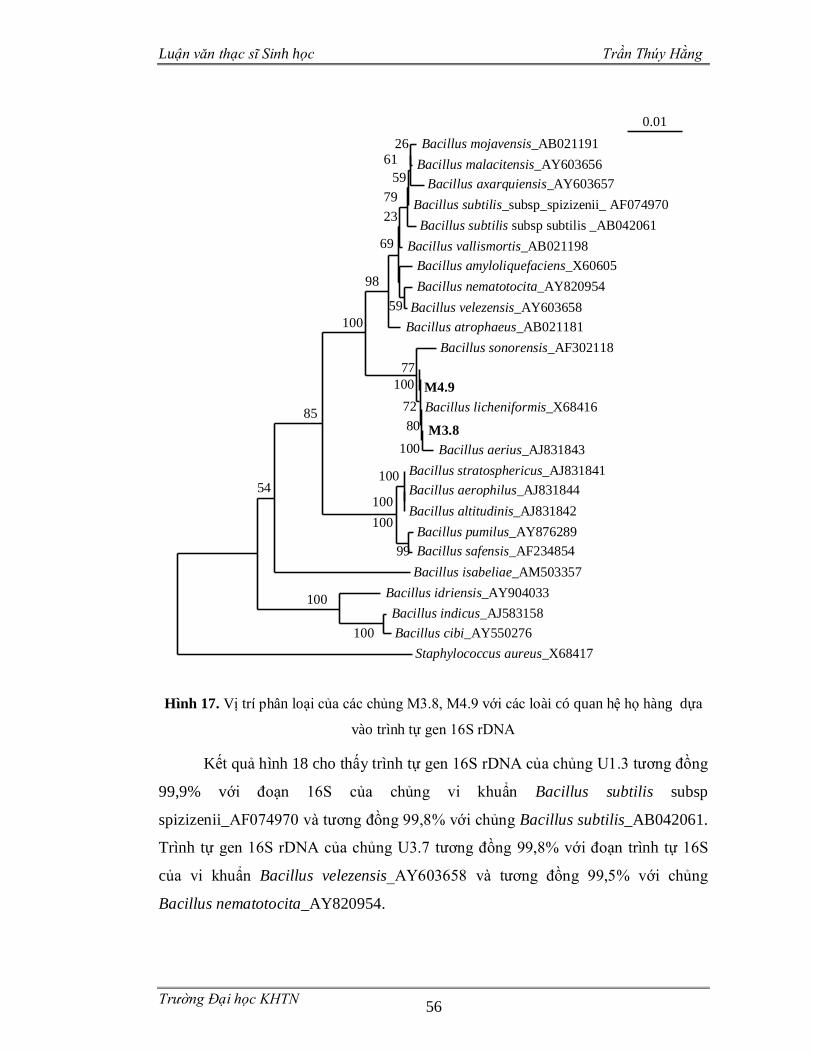
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 56
Hình 17. Vị trí phân loại của các chủng M3.8, M4.9 với các loài có quan hệ họ hàng dựa
vào trình tự gen 16S rDNA
Kết quả hình 18 cho thấy trình tự gen 16S rDNA của chủng U1.3 tương đồng
99,9% với đoạn 16S của chủng vi khuẩn Bacillus subtilis subsp
spizizenii_AF074970 và tương đồng 99,8% với chủng Bacillus subtilis_AB042061.
Trình tự gen 16S rDNA của chủng U3.7 tương đồng 99,8% với đoạn trình tự 16S
của vi khuẩn Bacillus velezensis_AY603658 và tương đồng 99,5% với chủng
Bacillus nematotocita_AY820954.
Staphylococcus aureus_X68417 Bacillus cibi_AY550276 Bacillus indicus_AJ583158
Bacillus idriensis_AY904033
100
Bacillus isabeliae_AM503357 Bacillus safensis_AF234854 Bacillus pumilus_AY876289
Bacillus altitudinis_AJ831842 Bacillus aerophilus_AJ831844 Bacillus stratosphericus_AJ831841
100
99
100
Bacillus aerius_AJ831843
Bacillus licheniformis_X68416
M3.8 100
M4.9
80
77 Bacillus sonorensis_AF302118
72
Bacillus atrophaeus_AB021181 Bacillus velezensis_AY603658 Bacillus nematotocita_AY820954 Bacillus amyloliquefaciens_X60605
59
Bacillus vallismortis_AB021198 Bacillus subtilis subsp subtilis _AB042061
Bacillus subtilis_subsp_spizizenii_ AF074970 Bacillus axarquiensis_AY603657
Bacillus malacitensis_AY603656 Bacillus mojavensis_AB021191 26
61 59
79 23
69
100
98
100
100
85
100
54
0.01

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 57
Hình 18. Vị trí phân loại của chủng U1.3 và U3.7 với các loài có quan hệ họ hàng dựa vào
trình tự gen 16S rDNA
Kết quả nghiên cứu cho thấy rằng 4 chủng vi khuẩn có khả năng hình thành
màng sinh vật mạnh nhất phân lập được từ các khu vực nước thải của các làng nghề
đều thuộc chi Bacillus. Chi này bao gồm các vi khuẩn hình que, Gram dương, có
khả năng sinh trưởng ở nhiệt độ cao, hình thành bào tử và đa số là không gây bệnh.
Nhờ khả năng tạo được các sản phẩm thương mại có giá trị, như hầu hết các
protease và amylase, cũng như thao tác di truyền dễ dàng mà các vi khuẩn chi
Bacillus được xem là mô hình nghiên cứu hiệu quả và có tính ứng dụng cao[49].
Sự tạo thành màng sinh vật ở chủng Bacillus subtilis đóng vai trò quan trọng
ứng dụng trong nhiều lĩnh vực: công nghiệp, nông nghiệp, thực phẩm, hóa mỹ phẩm
Staphylococcus aureus_X68417 Bacillus cibi_AY550276 Bacillus indicus_AJ583158
Bacillus idriensis_AY904033 100
Bacillus isabeliae_AM503357 Bacillus safensis_AF234854 Bacillus pumilus_AY876289
Bacillus altitudinis_AJ831842 Bacillus aerophilus_AJ831844 Bacillus stratosphericus_AJ831841
99 100
99 Bacillus aerius_AJ831843
Bacillus licheniformis_X68416 Bacillus sonorensis_AF302118
100
Bacillus atrophaeus_AB021181 Bacillus velezensis_AY603658
U3.7 Bacillus nematotocita_AY820954
82
Bacillus amyloliquefaciens_X60605
79
Bacillus vallismortis_AB021198 Bacillus axarquiensis_AY603657
Bacillus malacitensis_AY603656 Bacillus mojavensis_AB021191
78
Bacillus subtilis subsp spizizenii_ AF074970
59
U1.3 Bacillus subtilis_AB042061
67
54 79
70
72
100
99
100
98
77
100
66
0.01

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 58
[56]. Bằng cách tạo ra hợp chất surfactin trong quá trình tạo màng sinh vật,
B. subtilis giúp các cây trồng có khả năng kháng lại một số vi khuẩn gây bệnh như
Erwina, Pseudomonas, Xanthomonas. Khả năng tạo gramicidin của chủng Bacillus
brevis 18-3 có tác dụng làm giảm đáng kể tốc độ ăn mòn kim loại bằng cách ức chế
cả hai chủng vi khuẩn D. orientis và L. discophora SP-6 [5]. Mặt khác, khả năng tạo
chất hoạt động bề mặt của các chủng vi sinh vật trong quá trình tạo màng cũng làm
tăng mức độ nhũ tương hóa và tạo bọt trong chế biến thực phẩm và tạo nhũ hóa cho
các sản phẩm mỹ phẩm [80]. Đồng thời, đặc tính này cũng đã mở ra triển vọng mới
trong việc chế tạo chất hoạt động bề mặt sinh học hoạt tính cao từ nguồn vi sinh vật
ứng dụng cho công nghiệp dầu khí để xử lý các sự cố tràn dầu [6].

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 59
KẾT LUẬN VÀ KIẾN NGHỊ
Kết luận
1. Từ mẫu 75 chủng vi sinh vật phân lập được từ nước thải các làng nghề ở
Việt Nam, chúng tôi đã lựa chọn được 8 chủng M1.10, U1.3, A3.3, M3.8, U3.7,
M4.3, M4.9 và M4.10 có hoạt tính tạo màng sinh vật cao hơn các chủng còn lại.
2. Tám chủng vi khuẩn phân lập có khả năng sinh trưởng và tạo màng sinh
vật tốt nhất ở 37oC và pH từ 7 - 7,5. Trong số các chủng này, bốn chủng M3.8,
M4.9, U1.3, U3.7 có khả năng sinh trưởng và tạo màng sinh vật tốt nhất trong môi
trường khoáng cơ bản có bổ sung các nguồn cacbon (fructose, rhamnose,
glucosamine) và trong môi trường khoáng cơ sở có bổ sung các nguồn nitơ
((NH4)2SO4, cao nấm men) khác nhau.
3. Các chủng M3.8, M4.9 và U1.3 có hoạt tính kháng mạnh đối với E. coli và
Vibrio parahaemolyticus, với đường kính vòng kháng khuẩn lần lượt là 7 mm và 9
mm; 8 mm và 10 mm; 7 mm và 9 mm; trong khi đó chủng U3.7 có hoạt tính kháng
mạnh đối với 3 chủng Staphylococcus aureus, Samonella typhi và Ralstonia
solanacaerum với đường kính vòng kháng khuẩn lần lượt là 7 mm, 7 mm và 8 mm.
4. Những phân tích về đặc điểm hình thái và phân tích gen 16S rDNA của 4
chủng cho phép chúng tôi nhận định 4 chủng M3.8, M4.9, U1.3, U3.7 là vi khuẩn
Gram dương thuộc chi Bacillus. Trong đó, chủng M3.8, M4.9 gần với loài Bacillus
licheniformis; chủng U1.3 gần với loài Bacillus subtilis còn chủng U3.7 gần với loài
Bacillus velezensis.
Kiến nghị
Tiếp tục nghiên cứu sâu hơn về khả năng tạo thành chất hoạt động bề mặt và
đặc tính kháng khuẩn của bốn chủng vi khuẩn phân lập M3.8, M4.9, U1.3, U3.7 để
ứng dụng trong công nghệ xử lý nước thải và trong phòng chống dịch hại gây bệnh
trên cây trồng.
Tìm hiểu thành phần protein và vai trò của sự điều hòa biểu hiện gen trong
quá trình tạo thành màng sinh vật của bốn chủng vi khuẩn phân lập.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 60
TÀI LIỆU THAM KHẢO
Tài liệu tiếng Việt:
1. Nguyễn Lân Dũng, Nguyễn Đình Quyến, Phạm Văn Ty (2002), Vi sinh vật học,
Nxb Giáo dục, Hà Nội.
2. Hồ Huỳnh Thùy Dương (2002), Sinh học phân tử, Nxb Giáo dục, Hà Nội.
3. Vũ Thị Minh Đức (2001), Thực tập vi sinh vật học, Nxb Đại học Quốc Gia,
Hà Nội.
Tài liệu tiếng Anh:
4. Allison D. G., Gilbert P., Wilson M. (2000), Community ctructure and
co-operation in biofilms, Cambridge University Press, Cambridge, England.
5. Bais H. P., Fall R., Vivanco J. M. (2004), “Biocontrol of Bacillus subtilis against
infection of arabidopsis roots by Pseudomonas syringae is facilitated by
biofilm formation and surfactin production”, Plant Physiology, 134(1),
pp. 307-319.
6. Banat I. M. (1995), “Characterization of biosurfactants and their use in pollution
removal-state of the art”, Acta biotechnologica, 15, pp. 251-267.
7. Bendinger B., Rijnaarts H. H. M., Altendorf K., Zehnder A. J. B. (1993),
“Physicochemical cell surface and adhesive properties of coryneform
bacteria related to the presence and chain length of mycolic acids”, Applied
and Environmental Microbiology, 59, pp. 3973-3977.
8. Beveridge T. J., Makin S. A., Kadurugamuwa J. L., Li Z. (1997), “Interactions
between biofilms and the environment”, FEMS Microbiology Reviews, 20,
pp. 291-303.
9. Boyd A., Chakrabarty A. M. (1994), “Role of alginate lyase in cell detachment of
Pseudomonas aeruginosa”, Applied and Environmental Microbiology, 60,
pp. 2355-2359.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 61
10. Brading M. G., Jass J., Lappin-Scott H. M. (1995), “Dynamics of bacterial
biofilm formation”, Microbial biofilms, Cambridge University Press,
Cambridge, pp. 46-63.
11. Brooks W., Demuth D. R., Gil S., Lamont R. J. (1997), “Identification of a
Streptococcus gordonii SspB domain that mediates adhesion to
Porphyromonas gingivalis”, Infection and Immunity, 65, pp. 3753-3758.
12. Characklis W. G. (1990), “Biofilm processes”, Biofilms, Wiley-Interscience,
New York, pp. 195-231.
13. Characklis W. G., Marshall K. C. (1990), “Biofilms: a basis for an
interdisciplinary approach”, Biofilms, John Wiley & Sons, New York,
pp. 3-15.
14. Costerton J. W., Geesey G. G., Cheng K. J. (1978), “How bacteria stick”,
Scientific American, 238(1), pp. 86-95.
15. Costerton J. W., Ingram J. M., Cheng K. J. (1974), “Structure and function of
the cell envelope of gram-negative bacteria”, Bacteriological Reviews, 38(1),
pp. 87-110.
16. Costerton J. W., Lewandowski Z., Caldwell D. E., Korber D. R., Lappin-Scott
H. M. (1995), “Microbial biofilms”, Annual Review of Microbiology, 49,
pp. 711-745.
17. Cowan M. M., Warren T. M., Fletcher M. (1991), “Mixed species colonization
of solid surfaces in laboratory biofilms”, Biofouling, 3, pp. 23-34.
18. Czaczyk K., Myszka K. (2007), “Biosynthesis of extracellular polymeric
substances (EPS) and its role in microbial biofilm formation”, Polish Journal
of Environmental Studies, 16(6), pp. 799-806.
19. Davey M. E., O'toole G. A. (2000), “Microbial biofilms: from ecology to
molecular genetics”, Microbiology and Molecular Biology Reviews, 64(B),
pp. 847-867.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 62
20. Davies D. G., Parsek M. R., Pearson J. P., Iglewski B. H., Costerton J. W.,
Greenberg E. P. (1998), “The involvement of cell-to-cell signals in the
development of a bacterial biofilm”, Science, 280, pp. 295-298.
21. De Angelis M., Siragusa S., Berloco M., Caputo L., Settanni L., Alfonsi G.,
Amerio M., Grandi A., Gobbetti M. (2006), “Selection of potential probiotic
lactobacilli from pig feces to be used as additives in pelleted feeding”,
Research in Microbiology, 157, pp. 792-801.
22. De Weger L. A., Van Der Vlught C. I. M., Wijfjes A. H. M., Bakker P. A. H.
M., Schippers B., Lugtenberg B. (1987), “Flagella of a plant growth-
stimulating Pseudomonas fluorescens strain are required for colonization of
potato roots”, Journal of Bacteriology, 169, pp. 2769-2773.
23. Decho A. W. (1990), “Microbial exopolymer secretion in ocean environment:
Their role(s) in food web and marine process”, Oceanography and Marine
Biology: Annual Review, 28, pp. 73-153.
24. Deflaun M. F., Marshall B. M., Kulle E. P., Levy S. B. (1994), “Tn5 insertion
mutants of Pseudomonas fluorescens defective in adhesion to soil and
seeds”, Applied and Environmental Microbiology, 60, pp. 2637-2642.
25. Donlan R. M. (2000), “Role of biofilms in antimicrobial resistance”, American
Society for Artificial Internal Organs Journal, 46(6), pp. S47-S52.
26. Donlan R. M. (2002), “Biofilms: Microbial life on surfaces”, Emerging
Infectious Diseases, 8(9), pp. 881-890.
27. Donlan R. M., Pipes W. O., Yohe T. L. (1994), “Biofilm formation on cast iron
substrata in water distribution systems”, Water Research, 28, pp. 1497-1503.
28. Emerson D., Ghiorse W. C. (1992), “Isolation, cultural maintenance, and
taxonomy of a sheath-forming strain of Leptothrix discophora and
characterization of manganese-oxidizing activity associated with the sheath”,
Applied and Environmental Microbiology, 58, pp. 4001-4010.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 63
29. Espinosa-Urgel M., Salido A., Ramos J. L. (2000), “Genetic analysis of
functions involved in adhesion of Pseudomonas putida to seeds”, Journal of
Bacteriology, 182, pp. 2363-2369.
30. Fall R., Kinsinger R. F., Wheeler K. A. (2004), “A simple method to isolate
biofilm-forming Bacillus subtilis and related species from plant roots”,
Systematic and Applied Microbiology, 27, pp. 372-379.
31. Flemming H. C. (1993), “Biofilm and environmental protection”, Water Science
and Technology, 27, pp. 1-10.
32. Fletcher M. (1988), “Attachment of Pseudomonas fluorescens to glass and
influence of electrolytes on bacterium-substratum separation distance”,
Journal of Bacteriology, 170, pp. 2027-2030.
33. Fujita M., Tanaka K., Takahashi H., Amemura A. (1994), “Transcription of the
principal sigma-factor genes, rpoD and rpoS, in Pseudomonas aeruginosa is
controlled according to the growth phase”, Molecular Microbiology, 13,
pp. 1071-1077.
34. Geesey G. G. (1990), Biofouling and biocorrosion in industrial water systems:
Proceedings of the international workshop on industrial biofouling and
biocorrosion, stuttg, Springer Verlag, Berlin, Germany.
35. Geesey G. G. (2001), “Bacterial behaviour at surfaces”, Current Opinion in
Biotechnology, 4, pp. 296-300.
36. Gilbert P., Das J., Foley I. (1997), “Biofilm susceptibility to antimicrobials”,
Advances in Dental Research, 11(1), pp. 160-167.
37. Haggag W. (2010), “The role of biofilm exopolysaccharides on biocontrol of
plant diseases”, Biopolymers, InTech, pp. 217-184.
38. Haggag W. M. (2007), “Colonization of exopolysaccharide-producing
Paenibacillus polymyxa on peanut roots for enhancing resistance against
crown rot disease”, African Journal of Biotechnology, 6(3), pp. 1568-1577.
39. Heukelekian H., Heller A. (1940), “Relation between food concentration and
surface for bacterial growth”, Journal of Bacteriology, 40(4), pp. 547-558.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 64
40. Heydorn A., Nielsen A. T., Hentzer M., Sternberg C., Givskov M., Ersboll B.
K., Molin S. (2000), “Quantification of biofilm structures by the novel
computer program COMSTAT”, Microbiology, 146, pp. 2395-2407.
41. Hussain M., Wilcox M. H., White P. J. (1993), “The slime of coagulase -
negative staphylococci: biochemistry and relation to adherence”, FEMS
Microbiology Reviews, 104, pp. 191-208.
42. Jackson D. W., Simecka J. W., Romeo T. (2002), “Catabolite repression of
Escherichia coli biofilm formation”, Journal of Bacteriology, 184,
pp. 3406-3410.
43. Jones H. C., Roth I. L., Sanders W. M. (1969), “Electron microscopic study of a
slime layer”, Journal of Bacteriology, 99(1), pp. 316-325.
44. Kaplan J. B., Ragunath C., Ramasubbu N., Fine D. H. (2003), “Detachment of
Actinobacillus actinomycetemcomitans biofilm cells by an endogenous beta-
hexosaminidase activity”, Journal of Bacteriology, 185, pp. 4693-4698.
45. Kokare C. R., Chakraborty S., Khopade A. N., Mahadik K. R. (2009), “Biofilm:
Importance and applications”, Indian Journal of Biotechnology, 8,
pp. 159-168.
46. Korber D. R., Lawrence J. R., Sutton B., Caldwell D. E. (1989), “Effects of
laminar flow velocity on the kinetics of surface colonization by mot+ and
mot- Pseudomonas fluorescens”, Microbial Ecology, 18, pp. 1-19.
47. Kubota H., Senda S., Nomura M., Tokuda H., Uchiyama H. (2008), “Biofilm
formation by lactic acid bacteria and resistance to environmental stress”,
Journal of Bioscience and Bioengineering, 106(4), pp. 381-386.
48. Lamont R. J., Jenkinson H. F. (1998), “Life below the gum line: pathogenic
mechanisms of Porphyromonas gingivalis”, Microbiology and Molecular
Biology Reviews, 62, pp. 1244-1263.
49. Lazarova V., Manem J. (1995), “Biofilm characterization and activity analysis
in water and wastewater treatment”, Water Research, 29(10), pp. 2227-2245.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 65
50. Lazarova V., Manem J. (2000), “Innovative biofilm treatment technologies for
water and wastewater treatment”, Biofilms II: process analysis and
applications, Wiley-Liss, New York, United States, pp. 159-206.
51. Lee J. W., Yeomans W. G., Allen A. L., Deng F., Gross R. A., Kaplan D. L.
(1999), “Biosynthesis of novel exopolymers by Aureobasidium pullulans”,
Applied and Environmental Microbiology, 65(12), pp. 5265-5271.
52. Lengeler J. W., Drews G., Schlegel H. G. (1999), Biology of the Prokaryotes,
Wiley-Blackwell, New Jersey, United States.
53. Leriche V., Sibille P., Carpentier B. (2000), “Use of an enzyme-linked
lectinsorbent assay to monitor the shift in polysaccharide composition in
bacterial biofilms”, Applied and Environmental Microbiology, 66,
pp. 1851-1856.
54. Matsukawa M., Greenberg E. P. (2004), “Putative exopolysaccharide synthesis
genes influence Pseudomonas aeruginosa biofilm development”, Journal of
Bacteriology, 186, pp. 4449-4456.
55. Mceldowney S., Fletcher M. (1986), “Variability of the influence of
physicochemical factors affecting bacterial adhesion to polystyrene
substrata”, Applied and Environmental Microbiology, 52(3), pp. 460–465.
56. Morikawa M. (2006), “Beneficial biofilm formation by industrial bacteria
Bacillus subtilis and related species”, Journal of Bioscience and
Bioengineering, 101(1), pp. 1-8.
57. Morikawa M., Kagihiro S., Haruki M., Takano K., Branda S., Kolter R., Kanaya
S. (2006), “Biofilm formation by a Bacillus subtilis strain that produces
γ - polyglutamate”, Microbiology, 152, pp. 2801-2807.
58. National Association of Corrosion Engineers (1976), “The role of bacteria in the
corrosion of oil field equipment”, Technical Practices Committee No 3,
National Association of Corrosion Engineers, Houston, Texas, pp. 23.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 66
59. Nguyen Q. H., Nguyen T. P. L., Tran T. H. (2011), “Characterization of biofilm-forming bacteria isolated from soil in Vietnam”, VNU Journal of Science, Natural Sciences and Technology, 27(2S), pp. 187-193.
60. Notermans S., Doormans J. A., Mead G. C. (1991), “Contribution of surface attachment to the establishment of microorganisms in food processing plants: A review”, Biofouling, 5, pp. 21-36.
61. O'toole G. A., Gibbs K. A., Hager P. W., Phibbs P. V. J., Kolter R. (2000), “The global carbon metabolism regulator crc is a component of a signal transduction pathway required for biofilm development by Pseudomonas aeruginosa”, Journal of Bacteriology, 182(2), pp. 425-431.
62. O’toole G. A., Kolter R. (1998), “Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development”, Molecular Microbiology, 30, pp. 295-304.
63. O’toole G. A., Kolter R. (1998), “Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis”, Molecular Microbiology, 28, pp. 449-461.
64. Oliveira R., Melo L., Oliveira A., Salgueiro R. (1994), “Polysaccharide production and biofilm formation by Pseudomonas fluorescen: effects of pH and surface material”, Colloids and Surfaces B: Biointerfaces, 2(1-3), pp. 41-46.
65. Otto K., Norbeck J., Larsson T., Karlsson K. A., Hermansson M. (2001), “Adhesion of type 1-fimbriated Escherichia coli to abiotic surfaces leads to altered composition of outer membrane proteins”, Journal of Bacteriology, 183, pp. 2445-2453.
66. Page S., Gaylarde C. (1990), “Biocide activity against Legionella and Pseudomonas”, International Biodeterioration, 26(2-4), pp. 139-148.
67. Pornsunthorntawee O., Maksung S., Huayyai O. (2009), “Biosurfactant production by Pseudonmonas aeruginosa SP4 using sequencing batch reactors: Effect of oil loading rate and cycle time”, Bioresource Technology 100, pp. 812-818.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 67
68. Pratt L. A., Kolter R. (1998), “Genetic analysis of Escherichia coli biofilm formation: roles of flagella, motility, chemotaxis and type I pili”, Molecular Microbiology, 30(2), pp. 285-289.
69. Ramey B. E., Koutsoudis M., Von Bodman S. B., Fuqua C. (2004), “Biofilm formation in plant-microbe associations”, Current Opinion in Microbiology, 7(6), pp. 602-609.
70. Ron E. Z., Rosenberg E. (2001), “Natural roles for biosurfactants”, Environmental Microbiology, 3, pp. 229-236.
71. Sanger F., Nicklen E. F., Conlson A. R. (1997), “DNA Sequencing with chain terminating inhibitor”, Proceedings of the National Academy of Sciences of the United States of America, 74(12), pp. 5403-5460.
72. Sauer K., Camper A. K., Ehrlich G. D., Costerton J. W., Davies D. G. (2002), “Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm”, Journal of Bacteriology, 184, pp. 1140-1154.
73. Schmoll T., Ott M., Oudega B., Hacker J. (1990), “Use of a wild-type gene fusion to determine the influence of environmental conditions on expression of the S fimbrial adhesin in Escherichia coli pathogen”, Journal of Bacteriology, 172(5), pp. 5103-5111.
74. Sleytr U. B. (1997), “Basic and applied S-layer research: an overview”, FEMS Microbiology Reviews, 20(1-2), pp. 5–12.
75. Stanley N. R., Lazazzera B. A. (2004), “Environmental signals and regulatory pathways that influence biofilm formation”, Molecular Microbiology, 52, pp. 917-924.
76. Steyn B. (2005), Proteomic analysis of the biofilm and biofilm – associated phenotypes of Pseudomonas aeruginosa cultured in batch, PhD dissertation, The Faculty of Natural and Agricultural Sciences, University of Pretoria, Pretoria.
77. Stoodley P., Sauer K., Davies D. G., Costerton J. W., Microbiol. A. R. (2002), “Biofilms as complex differentiated communities”, Annual review of microbiology, 56, pp. 187-209.

Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 68
78. Sule P., Horne S. M., Logue C. M., Prüß B. M. (2011), “Regulation of cell division, biofilm formation, and virulence by FlhC in Escherichia coli O157:H7 grown on meat”, Applied and Environmental Microbiology, 77(11), pp. 3653–3662.
79. Sutherland I. W. (2001), “Biofilm exopolysaccharides: a strong and sticky framework”, Microbiology, 147, pp. 3-9.
80. Suwansukho P., Rukachisirikul V., Kawai F., H-Kittikun A. (2008), “Production and applications of biosurfactant from Bacillus subtilis MUV4”, Songklanakarin Journal of Science and Technology, 30, pp. 87-93.
81. Thys S. (2009), Effect of biosurfactants isolated from endophytes on biofilm formation by Candida albicans, Master dissertation, Universiteit gent, Novara, Italy.
82. Tolker-Nielsen T., Brinch U. C., Ragas P. C., Andersen J. B., Jacobsen C. S., Molin S. (2000), “Development and dynamics of Pseudomonas sp. biofilms”, Journal of Bacteriology, 182, pp. 6482-6489.
83. Watnick P., Kolter P. (2000), “Biofilm, city of microbes”, Journal of Bacteriology, 182(10), pp. 2675-2679.
84. Watnick P. I., Fulner K. J., Kolter R. (1999), “A role for the mannose-sensitive hemagglutinin in biofilm formation by Vibrio cholera El Tor”, Journal of Bacteriology, 181, pp. 3606-3609.
85. Xie H., Cook G. S., Costerton J. W., Bruce G., Rose T. M., Lamont R. J. (2000), “Intergeneric communication in dental plaque biofilms”, Journal of Bacteriology, 182(24), pp. 7067-7069.
86. Zobell C. E. (1943), “The effect of solid surfaces upon bacteria activity”, Journal of Bacteriology, 46(1), pp. 39-56.
87. Zobell C. E., Anderson D. Q. (1936), “Observations on the multiplication of bacteria in different volumes of stored sea water and the influence of oxygen tension and solid surfaces”, The Biological bulletin, 71, pp. 324-342.
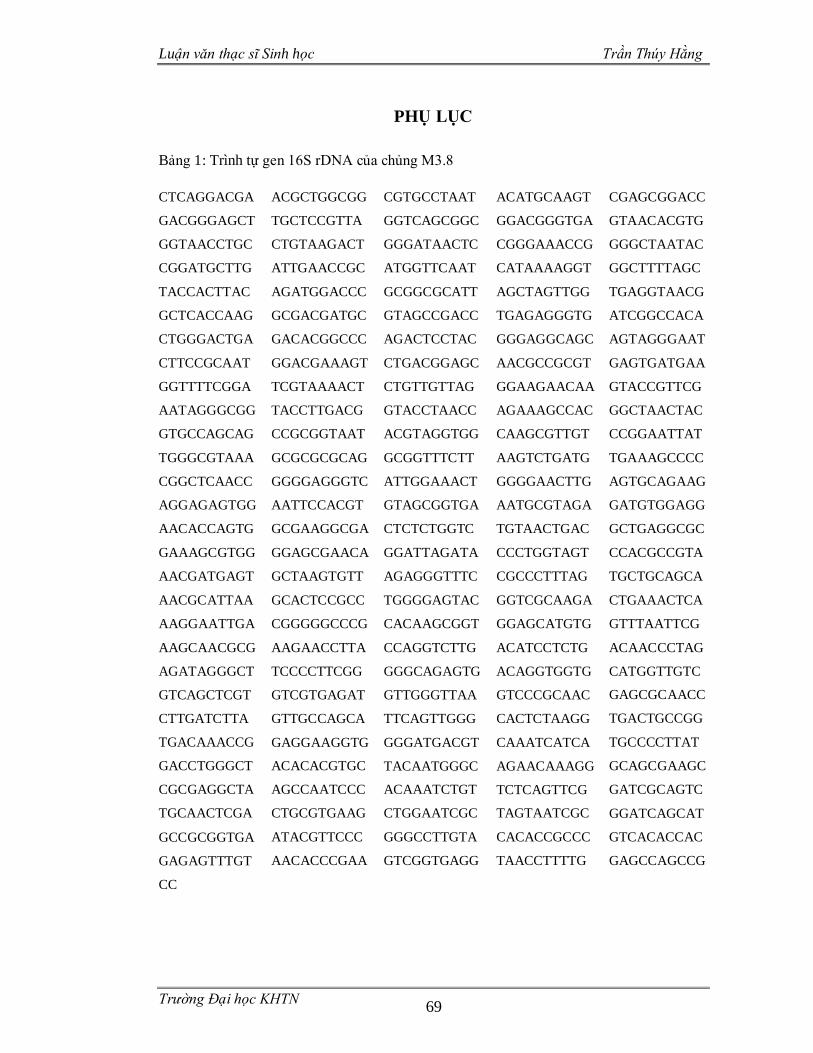
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 69
PHỤ LỤC
Bảng 1: Trình tự gen 16S rDNA của chủng M3.8 CTCAGGACGA ACGCTGGCGG CGTGCCTAAT ACATGCAAGT CGAGCGGACC
GACGGGAGCT TGCTCCGTTA GGTCAGCGGC GGACGGGTGA GTAACACGTG GGTAACCTGC CTGTAAGACT GGGATAACTC CGGGAAACCG GGGCTAATAC CGGATGCTTG ATTGAACCGC ATGGTTCAAT CATAAAAGGT GGCTTTTAGC
TACCACTTAC AGATGGACCC GCGGCGCATT AGCTAGTTGG TGAGGTAACG
GCTCACCAAG GCGACGATGC GTAGCCGACC TGAGAGGGTG ATCGGCCACA
CTGGGACTGA GACACGGCCC AGACTCCTAC GGGAGGCAGC AGTAGGGAAT
CTTCCGCAAT GGACGAAAGT CTGACGGAGC AACGCCGCGT GAGTGATGAA
GGTTTTCGGA TCGTAAAACT CTGTTGTTAG GGAAGAACAA GTACCGTTCG
AATAGGGCGG TACCTTGACG GTACCTAACC AGAAAGCCAC GGCTAACTAC
GTGCCAGCAG CCGCGGTAAT ACGTAGGTGG CAAGCGTTGT CCGGAATTAT
TGGGCGTAAA GCGCGCGCAG GCGGTTTCTT AAGTCTGATG TGAAAGCCCC
CGGCTCAACC GGGGAGGGTC ATTGGAAACT GGGGAACTTG AGTGCAGAAG
AGGAGAGTGG AATTCCACGT GTAGCGGTGA AATGCGTAGA GATGTGGAGG
AACACCAGTG GCGAAGGCGA CTCTCTGGTC TGTAACTGAC GCTGAGGCGC
GAAAGCGTGG GGAGCGAACA GGATTAGATA CCCTGGTAGT CCACGCCGTA
AACGATGAGT GCTAAGTGTT AGAGGGTTTC CGCCCTTTAG TGCTGCAGCA
AACGCATTAA GCACTCCGCC TGGGGAGTAC GGTCGCAAGA CTGAAACTCA
AAGGAATTGA CGGGGGCCCG CACAAGCGGT GGAGCATGTG GTTTAATTCG
AAGCAACGCG AAGAACCTTA CCAGGTCTTG ACATCCTCTG ACAACCCTAG
AGATAGGGCT TCCCCTTCGG GGGCAGAGTG ACAGGTGGTG CATGGTTGTC
GTCAGCTCGT GTCGTGAGAT GTTGGGTTAA GTCCCGCAAC GAGCGCAACC
CTTGATCTTA GTTGCCAGCA TTCAGTTGGG CACTCTAAGG TGACTGCCGG
TGACAAACCG GAGGAAGGTG GGGATGACGT CAAATCATCA TGCCCCTTAT
GACCTGGGCT ACACACGTGC TACAATGGGC AGAACAAAGG GCAGCGAAGC
CGCGAGGCTA AGCCAATCCC ACAAATCTGT TCTCAGTTCG GATCGCAGTC
TGCAACTCGA CTGCGTGAAG CTGGAATCGC TAGTAATCGC GGATCAGCAT
GCCGCGGTGA ATACGTTCCC GGGCCTTGTA CACACCGCCC GTCACACCAC
GAGAGTTTGT AACACCCGAA GTCGGTGAGG TAACCTTTTG GAGCCAGCCG
CC
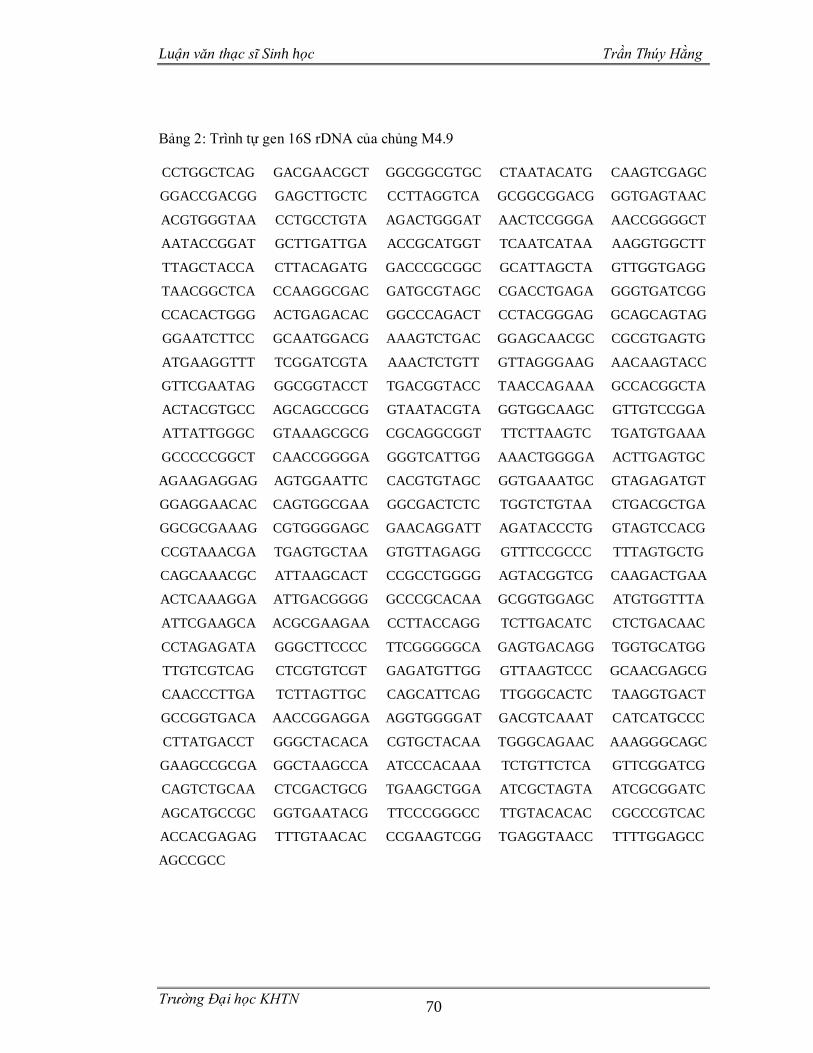
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 70
Bảng 2: Trình tự gen 16S rDNA của chủng M4.9 CCTGGCTCAG GACGAACGCT GGCGGCGTGC CTAATACATG CAAGTCGAGC
GGACCGACGG GAGCTTGCTC CCTTAGGTCA GCGGCGGACG GGTGAGTAAC
ACGTGGGTAA CCTGCCTGTA AGACTGGGAT AACTCCGGGA AACCGGGGCT
AATACCGGAT GCTTGATTGA ACCGCATGGT TCAATCATAA AAGGTGGCTT
TTAGCTACCA CTTACAGATG GACCCGCGGC GCATTAGCTA GTTGGTGAGG
TAACGGCTCA CCAAGGCGAC GATGCGTAGC CGACCTGAGA GGGTGATCGG
CCACACTGGG ACTGAGACAC GGCCCAGACT CCTACGGGAG GCAGCAGTAG
GGAATCTTCC GCAATGGACG AAAGTCTGAC GGAGCAACGC CGCGTGAGTG
ATGAAGGTTT TCGGATCGTA AAACTCTGTT GTTAGGGAAG AACAAGTACC
GTTCGAATAG GGCGGTACCT TGACGGTACC TAACCAGAAA GCCACGGCTA
ACTACGTGCC AGCAGCCGCG GTAATACGTA GGTGGCAAGC GTTGTCCGGA
ATTATTGGGC GTAAAGCGCG CGCAGGCGGT TTCTTAAGTC TGATGTGAAA
GCCCCCGGCT CAACCGGGGA GGGTCATTGG AAACTGGGGA ACTTGAGTGC
AGAAGAGGAG AGTGGAATTC CACGTGTAGC GGTGAAATGC GTAGAGATGT
GGAGGAACAC CAGTGGCGAA GGCGACTCTC TGGTCTGTAA CTGACGCTGA
GGCGCGAAAG CGTGGGGAGC GAACAGGATT AGATACCCTG GTAGTCCACG
CCGTAAACGA TGAGTGCTAA GTGTTAGAGG GTTTCCGCCC TTTAGTGCTG
CAGCAAACGC ATTAAGCACT CCGCCTGGGG AGTACGGTCG CAAGACTGAA
ACTCAAAGGA ATTGACGGGG GCCCGCACAA GCGGTGGAGC ATGTGGTTTA
ATTCGAAGCA ACGCGAAGAA CCTTACCAGG TCTTGACATC CTCTGACAAC
CCTAGAGATA GGGCTTCCCC TTCGGGGGCA GAGTGACAGG TGGTGCATGG
TTGTCGTCAG CTCGTGTCGT GAGATGTTGG GTTAAGTCCC GCAACGAGCG
CAACCCTTGA TCTTAGTTGC CAGCATTCAG TTGGGCACTC TAAGGTGACT
GCCGGTGACA AACCGGAGGA AGGTGGGGAT GACGTCAAAT CATCATGCCC
CTTATGACCT GGGCTACACA CGTGCTACAA TGGGCAGAAC AAAGGGCAGC
GAAGCCGCGA GGCTAAGCCA ATCCCACAAA TCTGTTCTCA GTTCGGATCG
CAGTCTGCAA CTCGACTGCG TGAAGCTGGA ATCGCTAGTA ATCGCGGATC
AGCATGCCGC GGTGAATACG TTCCCGGGCC TTGTACACAC CGCCCGTCAC
ACCACGAGAG TTTGTAACAC CCGAAGTCGG TGAGGTAACC TTTTGGAGCC
AGCCGCC
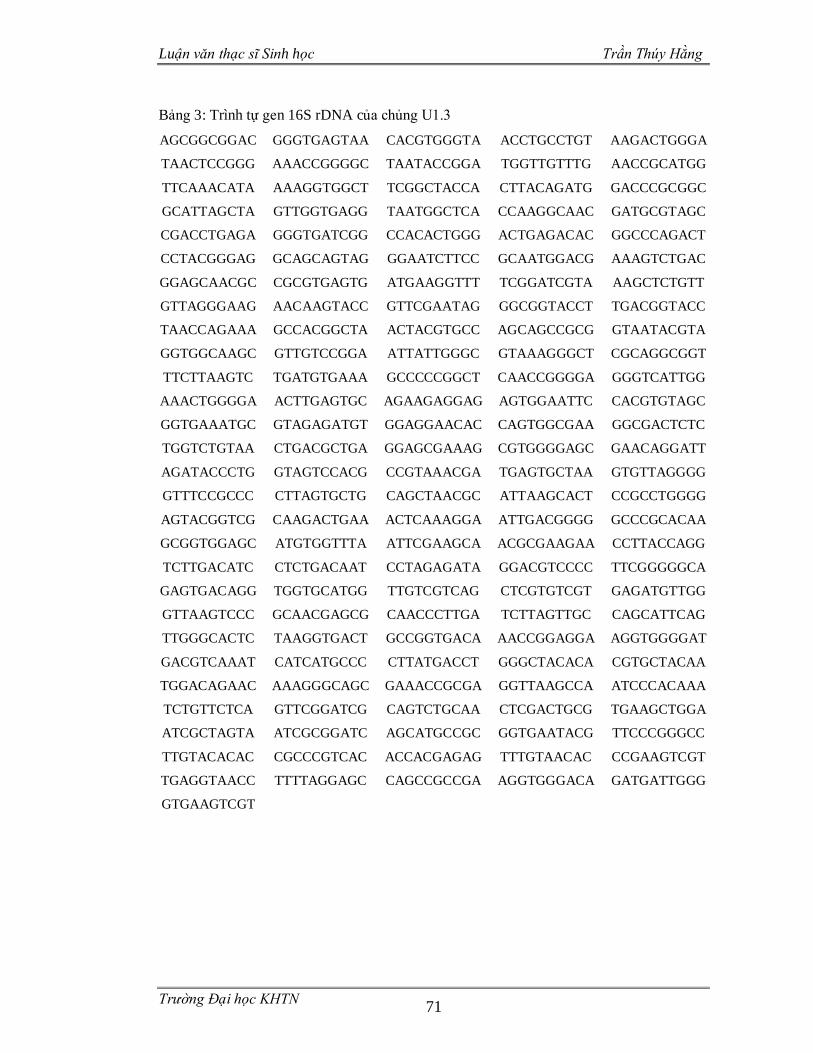
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 71
Bảng 3: Trình tự gen 16S rDNA của chủng U1.3
AGCGGCGGAC GGGTGAGTAA CACGTGGGTA ACCTGCCTGT AAGACTGGGA
TAACTCCGGG AAACCGGGGC TAATACCGGA TGGTTGTTTG AACCGCATGG
TTCAAACATA AAAGGTGGCT TCGGCTACCA CTTACAGATG GACCCGCGGC
GCATTAGCTA GTTGGTGAGG TAATGGCTCA CCAAGGCAAC GATGCGTAGC
CGACCTGAGA GGGTGATCGG CCACACTGGG ACTGAGACAC GGCCCAGACT
CCTACGGGAG GCAGCAGTAG GGAATCTTCC GCAATGGACG AAAGTCTGAC
GGAGCAACGC CGCGTGAGTG ATGAAGGTTT TCGGATCGTA AAGCTCTGTT
GTTAGGGAAG AACAAGTACC GTTCGAATAG GGCGGTACCT TGACGGTACC
TAACCAGAAA GCCACGGCTA ACTACGTGCC AGCAGCCGCG GTAATACGTA
GGTGGCAAGC GTTGTCCGGA ATTATTGGGC GTAAAGGGCT CGCAGGCGGT
TTCTTAAGTC TGATGTGAAA GCCCCCGGCT CAACCGGGGA GGGTCATTGG
AAACTGGGGA ACTTGAGTGC AGAAGAGGAG AGTGGAATTC CACGTGTAGC
GGTGAAATGC GTAGAGATGT GGAGGAACAC CAGTGGCGAA GGCGACTCTC
TGGTCTGTAA CTGACGCTGA GGAGCGAAAG CGTGGGGAGC GAACAGGATT
AGATACCCTG GTAGTCCACG CCGTAAACGA TGAGTGCTAA GTGTTAGGGG
GTTTCCGCCC CTTAGTGCTG CAGCTAACGC ATTAAGCACT CCGCCTGGGG
AGTACGGTCG CAAGACTGAA ACTCAAAGGA ATTGACGGGG GCCCGCACAA
GCGGTGGAGC ATGTGGTTTA ATTCGAAGCA ACGCGAAGAA CCTTACCAGG
TCTTGACATC CTCTGACAAT CCTAGAGATA GGACGTCCCC TTCGGGGGCA
GAGTGACAGG TGGTGCATGG TTGTCGTCAG CTCGTGTCGT GAGATGTTGG
GTTAAGTCCC GCAACGAGCG CAACCCTTGA TCTTAGTTGC CAGCATTCAG
TTGGGCACTC TAAGGTGACT GCCGGTGACA AACCGGAGGA AGGTGGGGAT
GACGTCAAAT CATCATGCCC CTTATGACCT GGGCTACACA CGTGCTACAA
TGGACAGAAC AAAGGGCAGC GAAACCGCGA GGTTAAGCCA ATCCCACAAA
TCTGTTCTCA GTTCGGATCG CAGTCTGCAA CTCGACTGCG TGAAGCTGGA
ATCGCTAGTA ATCGCGGATC AGCATGCCGC GGTGAATACG TTCCCGGGCC
TTGTACACAC CGCCCGTCAC ACCACGAGAG TTTGTAACAC CCGAAGTCGT
TGAGGTAACC TTTTAGGAGC CAGCCGCCGA AGGTGGGACA GATGATTGGG
GTGAAGTCGT
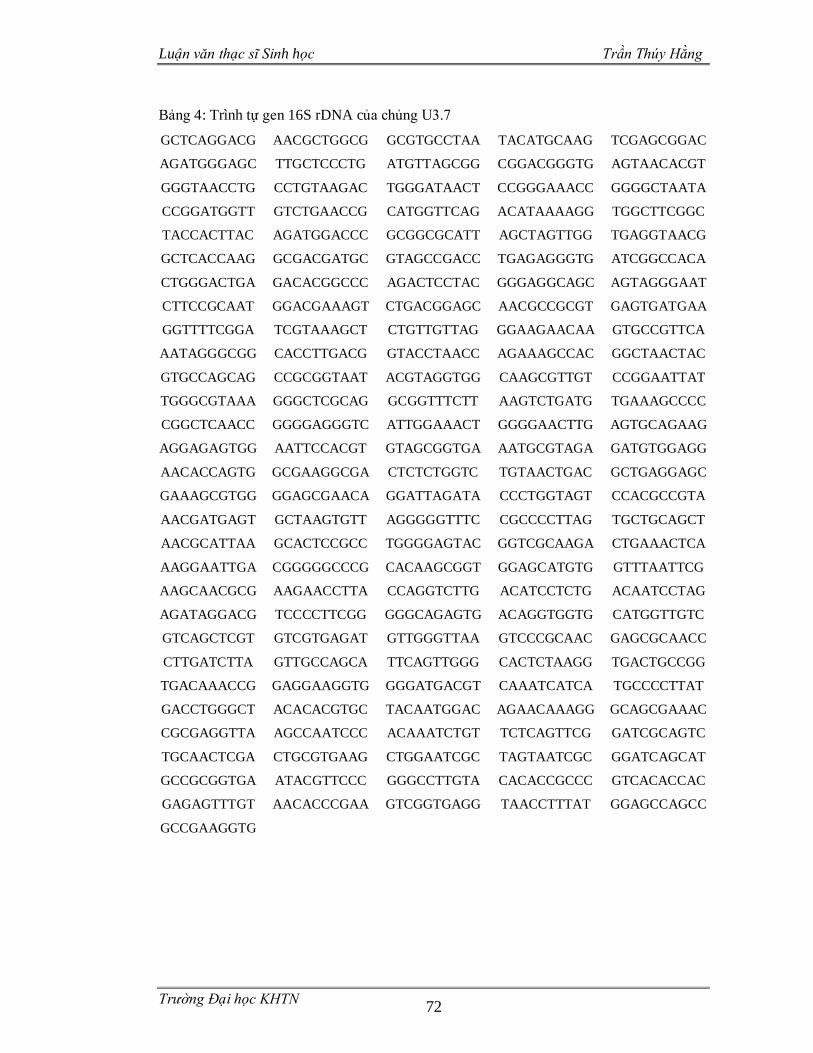
Luận văn thạc sĩ Sinh học Trần Thúy Hằng
Trường Đại học KHTN 72
Bảng 4: Trình tự gen 16S rDNA của chủng U3.7
GCTCAGGACG AACGCTGGCG GCGTGCCTAA TACATGCAAG TCGAGCGGAC
AGATGGGAGC TTGCTCCCTG ATGTTAGCGG CGGACGGGTG AGTAACACGT
GGGTAACCTG CCTGTAAGAC TGGGATAACT CCGGGAAACC GGGGCTAATA
CCGGATGGTT GTCTGAACCG CATGGTTCAG ACATAAAAGG TGGCTTCGGC
TACCACTTAC AGATGGACCC GCGGCGCATT AGCTAGTTGG TGAGGTAACG
GCTCACCAAG GCGACGATGC GTAGCCGACC TGAGAGGGTG ATCGGCCACA
CTGGGACTGA GACACGGCCC AGACTCCTAC GGGAGGCAGC AGTAGGGAAT
CTTCCGCAAT GGACGAAAGT CTGACGGAGC AACGCCGCGT GAGTGATGAA
GGTTTTCGGA TCGTAAAGCT CTGTTGTTAG GGAAGAACAA GTGCCGTTCA
AATAGGGCGG CACCTTGACG GTACCTAACC AGAAAGCCAC GGCTAACTAC
GTGCCAGCAG CCGCGGTAAT ACGTAGGTGG CAAGCGTTGT CCGGAATTAT
TGGGCGTAAA GGGCTCGCAG GCGGTTTCTT AAGTCTGATG TGAAAGCCCC
CGGCTCAACC GGGGAGGGTC ATTGGAAACT GGGGAACTTG AGTGCAGAAG
AGGAGAGTGG AATTCCACGT GTAGCGGTGA AATGCGTAGA GATGTGGAGG
AACACCAGTG GCGAAGGCGA CTCTCTGGTC TGTAACTGAC GCTGAGGAGC
GAAAGCGTGG GGAGCGAACA GGATTAGATA CCCTGGTAGT CCACGCCGTA
AACGATGAGT GCTAAGTGTT AGGGGGTTTC CGCCCCTTAG TGCTGCAGCT
AACGCATTAA GCACTCCGCC TGGGGAGTAC GGTCGCAAGA CTGAAACTCA
AAGGAATTGA CGGGGGCCCG CACAAGCGGT GGAGCATGTG GTTTAATTCG
AAGCAACGCG AAGAACCTTA CCAGGTCTTG ACATCCTCTG ACAATCCTAG
AGATAGGACG TCCCCTTCGG GGGCAGAGTG ACAGGTGGTG CATGGTTGTC
GTCAGCTCGT GTCGTGAGAT GTTGGGTTAA GTCCCGCAAC GAGCGCAACC
CTTGATCTTA GTTGCCAGCA TTCAGTTGGG CACTCTAAGG TGACTGCCGG
TGACAAACCG GAGGAAGGTG GGGATGACGT CAAATCATCA TGCCCCTTAT
GACCTGGGCT ACACACGTGC TACAATGGAC AGAACAAAGG GCAGCGAAAC
CGCGAGGTTA AGCCAATCCC ACAAATCTGT TCTCAGTTCG GATCGCAGTC
TGCAACTCGA CTGCGTGAAG CTGGAATCGC TAGTAATCGC GGATCAGCAT
GCCGCGGTGA ATACGTTCCC GGGCCTTGTA CACACCGCCC GTCACACCAC
GAGAGTTTGT AACACCCGAA GTCGGTGAGG TAACCTTTAT GGAGCCAGCC
GCCGAAGGTG





























