Embed Size (px)
Citation preview

MASARYKOVA UNIVERZITA
PŘÍRODOVĚDECKÁ FAKULTA
ÚSTAV BOTANIKY A ZOOLOGIE
VVaarr iiaabbii ll ii ttaa ssppoolleeččeennsstteevv mměěkkkkýýššůů bbaazziicckkýýcchh ssllaatt iinniiššťť
PPoollsskkaa aa jj iižžnníí SSkkaannddiinnáávviiee
Diplomová práce
Veronika Schenková
Vedoucí práce: doc. RNDr. Michal Horsák, Ph.D. Brno 2012

Bibliografický záznam
Autor: Bc. Veronika Schenková Přírodovědecká fakulta, Masarykova univerzita Ústav botaniky a zoologie
Název práce: Variabilita společenstev měkkýšů bazických slatinišť Polska a jižní Skandinávie
Studijní program: Biologie
Studijní obor: Systematická biologie a ekologie
Vedoucí práce: doc. RNDr. Michal Horsák, Ph.D.
Akademický rok: 2011/2012
Počet stran: 88
Klíčová slova: Měkkýši; Bazická slatiniště; Polsko; Skandinávie; Latitudinální gradient; Produktivita

Bibliographic entry
Author: Bc. Veronika Schenková Faculty of Science, Masaryk University Department of Botany and Zoology
Title of Thesis: Variation in mollusc assemblages of alkaline fens in Poland and southern Scandinavia
Degree programme: Biology
Field of Study: Systematic Biology and Ecology
Supervisor: doc. RNDr. Michal Horsák, Ph.D.
Academic Year: 2011/2012
Number of Pages: 88
Keywords: Molluscs; Alkaline fens; Poland; Scandinavia; Latitudinal gradient; Productivity

Abstrakt
Bazická slatiniště se vyznačují mimořádnou diverzitou a unikátní skladbou
společenstev měkkýšů. Přesto však nebyla ve většině evropských zemí věnována
malakofauně těchto biotopů přílišná pozornost. Hlavními cíli diplomové práce bylo
prozkoumat malakologicky dosud opomíjená bazická slatiniště Polska, a na základě
srovnání s bazickými slatiništi jižní Skandinávie, studovanými v rámci mé bakalářské
práce, vyhodnotit změny společenstev měkkýšů podél zeměpisné šířky.
V letech 2002–2011 bylo vzorkováno celkem 62 bazických slatinišť, 39 na území
východního Polska a 23 na území jižního Švédska a Norska. V terénu bylo měřeno pH a
konduktivita vody, nadmořská výška a zeměpisné souřadnice. Na základě souřadnic byly
k jednotlivým lokalitám doplněny klimatické proměnné. Data byla zpracována
v programech Statistica, CANOCO a R.
Na studovaných lokalitách bylo nalezeno celkem 72 druhů měkkýšů, ze kterých se 64
druhů vyskytovalo na území Polska a 40 na území jižní Skandinávie. Počty druhů i
abundance měkkýšů signifikantně klesaly směrem k vyšším zeměpisným šířkám. Velikost
mediánového jedince se v rámci společenstev suchozemských plžů podél zeměpisné šířky
neměnila, neboť na všech studovaných slatiništích zcela dominovali jedinci velmi drobných
druhů dosahujících nejvýše 4 mm. Směrem k vyšším zeměpisným šířkám signifikantně
rostlo zastoupení jedinců čel. Vertiginidae ve společenstvech, ačkoli druhová bohatost
tohoto taxonu zůstávala konstantní. Za nejvýznamnější rozdíly ve skladbě suchozemských
plžů odpovídal komplexní geografický gradient, související především s průměrnými
červencovými teplotami. Podél tohoto gradientu došlo k jasnému vymezení lokalit boreální
zóny Skandinávie, typických chladnomilnými druhy Vertigo genesii, V. modesta, V.
lilljeborgi a Columella columella, od lokalit mírného pásu Polska a jižního Švédska,
s výskytem klimaticky méně tolerantních druhů jako Vertigo angustior, V. antivertigo a
Vallonia enniensis. Na území Polska byly učiněny nové významné nálezy druhů Vertigo
geyeri, V. moulinsiana, Pupilla alpicola, P. pratensis a Sphaerium nucleus. Pro obě oblasti,
Polsko i jižní Skandinávii, byla prokázána logaritmická závislost počtu druhů na
produktivitě společenstev plžů, naznačující absenci kompetičních interakcí v rámci
slatiništních malakocenóz.

Abstract
Alkaline fens are characterized by a high diversity and unique species composition of
mollusc assemblages. However, in most of the European countries only few researches
have been done on fen mollusc fauna. The aims of this study were to survey mollusc
assemblages in previously unexplored alkaline fens of Poland, to compare them with fen
malacocenoses of southern Scandinavia, which were studied in my bachelor thesis, and to
assess changes in mollusc assemblages along the gradient of latitude.
Field work was conducted from 2002 to 2011. Altogether 62 alkaline fen sites were
sampled, 39 in eastern Poland and 23 in southern Sweden and Norway. Water
conductivity and pH, geographic coordinates and altitude were measured in the field, and
climatic parameters were compiled for each site. The data were processed using
Statistica, CANOCO and R programmes.
A total of 72 species were found in the studied fens, from which 64 species occurred
in Poland and 40 in Scandinavia. Both numbers of species and individuals decreased with
increasing latitude. In land snail assemblages, body size of a median specimen did not
change along the latitude, due to the predominance of minute snail species (up to 4 mm)
in all studied fens. Representation of individuals of the family Vertiginidae in land snail
assemblages increased significantly with increasing latitude, although species richness of
this taxon did not significantly change. The main direction of land snail species turnover
was associated with the complex geographical gradient, strongly correlated with mean July
temperatures. Along this gradient, sites in the boreal zone of Scandinavia, with the
occurrence of cold-climate species Vertigo genesii, V. modesta, V. lilljeborgi and
Columella columella, were clearly separated from the temperate sites of Poland and
southernmost Sweden, with the occurrence of less climatically tolerant species, e.g.
Vertigo angustior, V. antivertigo and Vallonia enniensis. Several new important records
were conducted for the territory of Poland, namely Vertigo geyeri, V. moulisiana, Pupilla
alpicola, P. pratensis and Sphaerium nucleus. In both regions, a logarithmic dependence
between the number of species and productivity of land snail assemblages was found,
suggesting the absence of interspecific competition among fen mollusc assemblages.



Poděkování
Ze všeho nejvíce bych ráda poděkovala svému školiteli Michalu Horsákovi za cenné
rady a podnětné připomínky v průběhu celého mého studia, a především za nikdy
neutuchající a značně nakažlivé nadšení pro malakologii a výzkum slatinišť. Speciálně
děkuji také za pomoc se zpracováním dat v programu R. Za skvělé terény v Polsku děkuji
botanikům Michalu Hájkovi, Danu Dítěti a nejvíc Zuzce Pleskové, která mi pomáhala při
prvním vzorkování polských slatinišť. Mockrát děkuji Pawłu Pawlikowskému za ochotnou
spolupráci a velkou pomoc v terénu – bez něj bychom některé z úchvatných polských
lokalit nikdy neobjevili. Poděkování si zaslouží také organizace IMCG, díky které jsem
poprvé nahlédla na polská slatiniště. Ondrovi Hájkovi děkuji za výrobu map a klimatická
data. Upřímně děkuji celé své rodině za to, že je tak fajn a že mě vždy podporuje v tom,
čemu se věnuji a co mě baví. Srdečně děkuji také všem svým milým přátelům za spoustu
pohody, smíchu a tolik potřebného odreagování ve volném čase. Na závěr děkuji všem
okolnostem, které mě dovedly ke studiu biologie a k velmi zajímavému tématu práce, díky
kterému pro mě psaní diplomky nebylo pouze povinností, ale především příjemnou a tvůrčí
náplní času.

Prohlášení Prohlašuji, že jsem svoji diplomovou práci vypracovala samostatně s využitím
informačních zdrojů, které jsou v práci citovány.
Souhlasím s uložením této diplomové práce v knihovně Ústavu botaniky a zoologie PřF
MU v Brně, případně v jiné knihovně MU, s jejím veřejným půjčováním a využitím pro
vědecké, vzdělávací nebo jiné veřejně prospěšné účely, a to za předpokladu, že převzaté
informace budou řádně citovány a nebudou využívány komerčně.
Brno 3. května 2012 …………………………………..
Veronika Schenková

OBSAH
1. ÚVOD ............................................................................................................................ 1
1.1. Cíle diplomové práce............................................................................................... 3
2. SLATINIŠTĚ A JEJICH MALAKOFAUNA.............................. ...................................... 4
2.1. Obecná charakteristika slatinišť .............................................................................. 4
2.2. Vliv ekologických gradientů na biotu slatinišť .......................................................... 5
2.3. Malakofauna slatinišť .............................................................................................. 7
2.3.1. Měkkýši jako modelová skupina ....................................................................... 7
2.3.2. Faktory ovlivňující diverzitu měkkýšů slatinišť .................................................. 7
2.3.3. Výzkumy slatiništních malakocenóz v Evropě .................................................. 9
2.4. Počet druhů, produktivita a slatiniště..................................................................... 10
3. ZMĚNY DIVERZITY PODÉL LATITUDINÁLNÍHO GRADIENTU....... ......................... 12
3.1. Obecný úvod k latitudinálnímu gradientu .............................................................. 12
3.2. Měkkýši a latitudinální gradient ............................................................................. 13
4. CHARAKTERISTIKA STUDOVANÝCH OBLASTÍ ............. ........................................ 16
4.1. Polsko ................................................................................................................... 16
4.1.1. Variabilita polských slatinišť............................................................................ 17
4.2. Jižní Skandinávie .................................................................................................. 18
4.2.1. Variabilita skandinávských slatinišť ................................................................ 19
5. METODIKA PRÁCE .................................. .................................................................. 20
5.1. Studované lokality ................................................................................................. 20
5.2. Terénní práce a zpracování vzorků ....................................................................... 20
5.3. Vysvětlující proměnné........................................................................................... 23
5.4. Statistické vyhodnocení dat................................................................................... 23
6. VÝSLEDKY ........................................ ......................................................................... 26
6.1. Společenstva měkkýšů Polska a jižní Skandinávie ............................................... 26
6.1.1. Změny druhové bohatosti a velikosti mediánového jedince............................ 26
6.1.2. Rozdíly v druhové skladbě společenstev měkkýšů......................................... 28
6.1.3. Specifika druhové skladby měkkýšů ve studovaných oblastech..................... 34
6.2. Malakocenózy polských slatinišť ........................................................................... 36
6.3. Produktivita společenstev suchozemských plžů.................................................... 39
7. DISKUZE..................................................................................................................... 42
7.1. Malakocenózy bazických slatinišť studovaných oblastí......................................... 42
7.1.1. Změny druhové bohatosti ............................................................................... 42

7.1.2. Změny velikosti mediánového jedince ............................................................ 43
7.1.3. Faktory ovlivňující skladbu společenstev suchozemských plžů ...................... 44
7.1.4. Druhové složení suchozemských plžů ve studovaných oblastech.................. 46
7.1.5. Změny diverzity čeledi Vertiginidae ................................................................ 47
7.2. Malakofauna bazických slatinišť východního Polska............................................. 48
7.2.1. Slatiniště polské strany Západních Karpat...................................................... 48
7.2.2. Slatiniště střední a severní části Polska ......................................................... 49
7.2.3. Významné nálezy pro Polsko.......................................................................... 50
7.3. Vztah druhové bohatosti a produktivity malakocenóz............................................ 52
8. ZÁVĚR......................................................................................................................... 54
9. CITOVANÁ LITERATURA ............................. ............................................................. 56
10. PŘÍLOHY................................................................................................................... 68

1
1. ÚVOD
Bazická slatiniště patří bezesporu mezi ideální modelové biotopy pro studium
společenstev měkkýšů. Z malakologického pohledu byla tato stanoviště překvapivě dlouho
přehlížena, a to i přes příznivé podmínky v podobě trvalé vlhkosti a vysokého obsahu
dostupného vápníku v podzemní vodě, odpovídající za mimořádnou diverzitu měkkýšů.
Právě vápník je pro měkkýše zcela stěžejním prvkem, nezbytným pro stavbu schránek a
potřebným také pro správné fungování rozmnožování (WÄREBORN 1970, 1979). Rozdíly ve
vápnitosti podzemní vody proto představují v lokálním měřítku nejvýznamnější ekologický
gradient, udávající změny druhové bohatosti a složení slatiništních malakocenóz (např.
POKRYSZKO 1993, HORSÁK & HÁJEK 2003). Počet druhů roste od chudých přechodových
rašelinišť po bazická slatiniště se srážením uhličitanu vápenatého ve formě pěnovce
(HORSÁK & HÁJEK 2003, HORSÁK & CERNOHORSKY 2008, HORSÁK et al. 2011a). Namísto
kompletní obměny druhové skladby však dochází spíše k víceméně plynulému přibývání
druhů náročnějších na množství dostupného vápníku (HORSÁK 2005a, HORSÁK &
CERNOHORSKY 2008).
V rámci mé bakalářské práce byly vyhodnoceny změny měkkýších společenstev podél
kompletního gradientu vápnitosti na slatiništích jižní Skandinávie. V navazující diplomové
práci bude prostudována také oblast východního Polska, a pozornost v ní bude věnována
pouze slatiništím bazické části gradientu minerální bohatosti. Na lokalitách, kde je vápník
již v nadbytku, nabývají na významu jiné ekologické faktory, jako např. zachovalost
lokality, nadmořská výška či klimatické poměry oblasti. Podstatnou roli může hrát stáří a
historický vývoj lokality, neboť výskyt některých vzácných druhů stanovištních specialistů a
glaciálních reliktů je často omezen na velmi starobylá slatiniště s přímou návazností na
konec posledního glaciálu (HORSÁK et al. 2007a, HÁJEK et al. 2011).
Ačkoli většina prací týkajících se ekologie společenstev slatiništních měkkýšů se
omezuje na výzkum území jednoho státu či geografického regionu (např. POKRYSZKO
1993, HORSÁK & HÁJEK 2003, HORSÁK et al. 2011a), existence některých zajímavých trendů
a rozdílů ve skladbě a bohatosti malakocenóz je patrná až při studiu větších prostorových
škál. Mezi základní, velmi obecné a univerzální jevy, které se na větších prostorových
škálách promítají do podoby společenstev organizmů, patří pokles druhové bohatosti
směrem od nižších k vyšším zeměpisným šířkám (např. ROSENZWEIG 1995, BROWN &
LOMOLINO 1998, WILLIG et al. 2003, HILLEBRAND 2004). Změna společenstev podél
severojižního gradientu se označuje jako latitudinální gradient diverzity. Jde o fenomén

2
známý již od 2. poloviny 19. století, potvrzený makroekologickými studiemi mnoha různých
skupin organizmů, jehož příčiny jsou však stále živě diskutovány a dosud nebyly
uspokojivě vysvětleny (např. PIANKA 1966, ROHDE 1992, GASTON 2000).
Bylo prokázáno, že latitudinální gradient ovlivňuje také diverzitu terestrických měkkýšů
(např. NEKOLA 2005, POKRYSZKO & CAMERON 2005, HAUSDORF 2006). Změny společenstev
podél tohoto gradientu se netýkají jen samotné druhové bohatosti malakocenóz, ale mění
se také skladba druhů, zastoupení některých taxonů či velikostní struktura společenstev
(HAUSDORF 2003, NEKOLA et al. v tisku). Žádná ze studií vlivu latitudinálního gradientu na
společenstva měkkýšů se však doposud nevěnovala slatiništím, která jsou oproti jiným
typům biotopů značně konzervativní, a díky přítomnosti v různých geografických regionech
je lze velmi dobře využít pro srovnání diverzity měkkýšů na velkých prostorových škálách
(HORSÁK et al. 2011a). Zjistit, jak rozdíly v zeměpisné šířce ovlivňují společenstva
měkkýšů na bazických slatiništích, bude proto jedním z cílů této diplomové práce.
Studované lokality se nacházejí na území Polska, Norska a Švédska, a pokrývají tak
poměrně širokou geografickou škálu, od mírného pásu střední Evropy až po
skandinávskou chladnou boreální zónu.
Důležitým cílem této práce bude také získání prvních detailních znalostí o
společenstvech měkkýšů slatinišť východní části Polska. Polská bazická slatiniště, ač
mnohdy rozsáhlá, zachovalá a známá výskytem populací vzácných a reliktních druhů
rostlin, byla z malakologického hlediska doposud opomíjena. Diplomová práce by tak měla
přinést zcela nové informace o skladbě a ekologii malakocenóz polských slatinišť a
případně přispět k ochraně těchto výjimečných biotopů před degradací a negativními
dopady lidské činnosti.
Jak již bylo výše uvedeno, pro studium rozdílů ve společenstvech měkkýšů podél
gradientu zeměpisné šířky budou použita také data z bazických slatinišť jižního Norska a
Švédska, zpracovaná v rámci bakalářské práce. Ve Skandinávii se díky vhodným
klimatickým podmínkám a postglaciálnímu vývoji krajiny po ústupu Skandinávského
ledovce nacházejí obrovské plochy různých typů rašeliništních biotopů, ovšem výskyt
bazických slatinišť je spíše vzácný a ojedinělý. Na většině území dominují chudá slatiniště
s rašeliníky a kyselá ombrotrofní vrchoviště, neboť podzemní voda je vlivem převažujícího
minerálně chudého krystalinického podloží nevápnitá (TAHVANAINEN 2004). Pro účely této
práce budou proto z datového souboru vybrány pouze lokality nacházející se v oblastech
s minerálně bohatším podložím, umožňujícím vznik bazických slatinišť s rozmanitými
společenstvy měkkýšů.

3
Úvodní kapitoly diplomové práce budou pojednávat o slatiništích a obecných
aspektech ekologie slatiništních malakocenóz, a charakterizovány zde budou rovněž
přírodní poměry zkoumaných oblastí. Pozornost bude věnována také latitudinálnímu
gradientu, především pak jeho vlivu na diverzitu společenstev měkkýšů. Stručně bude dále
zmíněna problematika vztahu druhové bohatosti a produktivity, která bude v rámci
diplomové práce nově studována na příkladu suchozemských slatiništních měkkýšů.
Zbývající část textu se bude týkat vyhodnocení vlastního výzkumu společenstev měkkýšů
na polských a skandinávských slatiništích, tedy metodice, zpracování a interpretaci
získaných výsledků. Hlavním posláním této diplomové práce je charakterizovat druhovou
skladbu, ekologii a specifické rysy malakocenóz dosud neprozkoumaných polských
slatinišť, a také vyhodnotit změny druhové skladby a bohatosti společenstev měkkýšů
podél gradientu zeměpisné šířky.
1.1. Cíle diplomové práce
1) shromáždit informace z literatury týkající se slatinišť, ekologie společenstev měkkýšů,
charakteru studovaných oblastí a problematiky latitudinálního gradientu diverzity
2) provést malakologický průzkum bazických slatinišť v oblasti východního Polska
3) zpracovat a vyhodnotit odebrané vzorky, prostudovat druhové složení, bohatost a
specifika společenstev měkkýšů na polských slatiništích
4) provést společnou analýzu dat z bazických slatinišť Polska, zpracovaných v rámci
diplomové práce, a bazických slatinišť v oblasti jižní Skandinávie, studovaných v rámci
bakalářské práce, a na základě výsledků:
a) vyhodnotit změny druhové skladby a bohatosti měkkýších společenstev podél
gradientu zeměpisné šířky
b) analyzovat změny velikostní struktury společenstev měkkýšů podél gradientu
zeměpisné šířky
5) pro každý vzorek zvážit jedince suchozemských ulitnatých plžů (v době vzorkování
živých) a podle stanovené hmotnosti biomasy popsat vztah produktivity a druhové
bohatosti společenstev; porovnat zjištěné údaje s analogickými výsledky pro
skandinávská slatiniště, získanými v rámci bakalářské práce

4
2. SLATINIŠTĚ A JEJICH MALAKOFAUNA
2.1. Obecná charakteristika slatiniš ť Slatiniště jsou vzácné, biologicky vyhraněné biotopy, vyznačující se mimořádnou
diverzitou rostlinných a živočišných společenstev a velmi často také výskytem ohrožených
a reliktních druhů (např. WASSEN et al. 2005, HORSÁK & CERNOHORSKY 2008, HÁJEK et al.
2011). Specifickou vlastností slatinišť je jejich trvalé podmáčení způsobené vyvěráním či
průsakem podzemní vody na povrch. Díky vzestupným hydrologickým gradientům
nedochází na slatiništích k výraznějšímu kolísání hladiny podzemní vody, a kořenová zóna
rostlin je tak saturována i v obdobích déletrvajícího sucha. Důsledkem nadměrného
zamokření je anoxie, pomalý a nedokonalý rozklad organické hmoty, limitované množství
živin a výskyt vegetace s převahou mechů a nízkých ostřic (HÁJEK et al. 2006). Dochází
k akumulaci organického materiálu ve formě rašeliny a často také ke vzniku sedimentů
uhličitanu vápenatého ve formě jílu či pěnovce (AMON et al. 2002).
Stálý kontakt povrchu slatiniště s podzemní vodou, a tedy i minerálně bohatým
podložím, odlišuje slatiniště od kyselých ombrotrofních vrchovišť, sycených výhradně
vodou srážkovou (ØKLAND et al. 2001, SJÖRS & GUNNARSSON 2002). Pro souhrnné
označení slatinišť a vrchovišť se tradičně používá termín rašeliniště (např. HÁJEK et al.
2006). Pro vymezení slatinišť oproti jiným typům mokřadních ekosystémů (slatinné louky,
aluviální sezónně přeplavovaná rašeliniště, močály…), je nutné kromě chemismu vody
zohlednit kombinaci působení dalších ekologických faktorů. Pro slatiniště se jedná
především o trvalé nasycení povrchu podzemní vodou, absenci dlouhodobého zaplavení,
akumulaci organických či uhličitanových sedimentů a typickou strukturu vegetace (AMON et
al. 2002). Někdy je kladen důraz také na přítomnost indikátorových druhů rostlin,
regionálně charakteristických pro konkrétní zkoumaná území (ØKLAND et al. 2001, AMON et
al. 2002, BRAGAZZA et al. 2005).
Největší koncentrace slatinišť a rašeliništních ekosystémů obecně se nachází
v boreální a subarktické zóně, v Evropě tedy především ve skandinávských zemích (RYDIN
& JEGLUM 2006). Obdobím vrcholného rozmachu slatinišť ve střední Evropě byl konec
posledního glaciálu až střední holocén (LOŽEK 1990), v současnosti již tyto biotopy
představují ve většině oblastí spíše roztroušené a izolované fragmenty. Hlavní příčinou
mizení slatinišť z krajiny jsou negativní dopady lidské činnosti, jako odvodňování, změny
hydrologie a převedení na zemědělské plochy, kterými byla drasticky zasažena především
slatiniště střední Evropy (JOOSTEN & CLARKE 2002, GROOTJANS et al. 2006, VAN DIGGELEN et

5
al. 2006). Vzhledem k unikátní skladbě organizmů, přítomnosti vzácných a reliktních
druhů, mimořádné diverzitě a především extrémní citlivosti na vnější zásahy, si tyto
biotopy zasluhují v současnosti zvláštní pozornost a ochranu (JOOSTEN & CLARKE 2002,
VAN DIGGELEN et al. 2006, HORSÁK & CERNOHORSKY 2008).
2.2. Vliv ekologických gradient ů na biotu slatiniš ť
O diverzitě slatiništní bioty spolurozhoduje komplex lokálních podmínek prostředí a
ekologických gradientů. Mezi základní determinanty druhové rozmanitosti patří minerální
bohatost, tedy především obsah vápníku v podzemní vodě napájející slatiniště. Rozdíly ve
vápnitosti vytvářejí nejvýznamnější a nejdelší ekologický gradient v rámci slatinišť, tzv.
gradient minerální bohatosti (např. MALMER 1986, ØKLAND et al. 2001, HÁJEK et al. 2002).
Vápnitost slatinišť je dána chemickou povahou geologického podloží a poměrně spolehlivě
ji zastupují parametry pH a konduktivita vody; pH především na minerálně chudších
lokalitách (TAHVANAINEN 2004), konduktivita na bazičtějších, vápníkem bohatších typech
slatinišť (HORSÁK 2006).
Minerální bohatost slatiniště lze poměrně spolehlivě odhadnout dle charakteru
rostlinných společenstev (HÁJEK et al. 2002, HÁJKOVÁ et al. 2004). Na velmi kyselých
slatiništích zcela dominují rašeliníky; s rostoucí koncentrací minerálů se skladba
mechového patra obměňuje, narůstá druhová bohatost, objevují se kalcikolní cévnaté
rostliny a přežívají pouze kalcitolerantní druhy rašeliníků. Při vysokých koncentracích
vápníku a hydrogenuhličitanů je zcela znemožněn růst rašeliníků a utváří se rostlinný
pokryv s převahou hnědých mechů a rostlin z čeledi Cyperaceae (podle HÁJEK et al.
2006).
Významnou charakteristikou bazických slatinišť je rovněž extrémní nedostatek
dostupného fosforu. Nízké množství tohoto limitujícího prvku zásadním způsobem
ovlivňuje charakter slatiništní vegetace, neboť prakticky vylučuje výskyt náročnějších
širolistých bylin a snižuje úživnost a produktivitu těchto stanovišť (BOYER & WHEELER 1989,
BOEYE et al. 1997, BEDFORD et al. 1999). Změny hydrologického režimu však mohou vést
k fluktuacím či trvalému snížení hladiny vody, a tím k vyššímu prokysličení, zrychlení
dekompozice a následnému uvolnění živin (ALMENDINGER & LEETE 1998). Obohacení
nízkoproduktivních slatiništních ekosystémů o živiny způsobí sukcesní posun ke
společenstvům lučních, křovinných a lesních druhů a souběžné mizení konkurenčně
slabších stanovištních specialistů a druhově rozmanitých slatiništních společenstev (např.
WASSEN et al. 2005, OLDE VENTERINK et al. 2001, AMON et al. 2002). Tento posun směrem

6
k produktivnějším stanovištím se označuje jako gradient fertility či trofický gradient
(WHEELER & PROCTOR 2000, HÁJEK et al. 2006). Zvýšené množství živin může být
podmíněno mnoha antropogenně vyvolanými procesy, např. eutrofizací, odvodněním
slatinišť a jejich okolí, přerušením trvalého odběru biomasy v podobě pastvy nebo kosení,
erozí či atmosférickou depozicí (viz např. JOOSTEN & CLARKE 2002, SJÖRS & GUNNARSSON
2002, VAN DIGGELEN et al. 2006, SKEFFINGTON et al. 2006).
Rozdíly v minerální bohatosti a fertilitě stanoviště lze poměrně snadno rozpoznat na
základě vyhodnocení lokálních ekologických parametrů. Může však dojít také k situaci, kdy
se biotopy zcela totožné, co se chemismu vody, množství dostupných živin a klimatických
podmínek týče, nápadně odlišují svým druhovým složením. Možným vysvětlením popsané
skutečnosti je existence tzv. geografického gradientu, odrážejícího rozdílný historický
vývoj a stáří těchto biotopů (HORSÁK et al. 2007a). Slatiniště představují refugia pro
specifická společenstva rostlin a živočichů a klíčem k pochopení rozdílů v druhové skladbě
je poznání, zda se jedná o paleorefugia, nebo neorefugia (NEKOLA 1999). Mnohá evropská
slatiniště mají svůj původ v pozdním glaciálu či starém holocénu, a v kontextu mladší
okolní krajiny proto představují typická paleorefugia. Naproti tomu lokality relativně mladé,
vzniklé následkem antropogenního odlesňování krajiny před několika staletími, lze označit
jako neorefugia (HORSÁK et al. 2007a, HÁJEK et al. 2009). Paleorefugia, reliktní fragmenty
dříve široce rozšířeného typu ekosystému, si uchovávají stabilní kombinaci specifických
podmínek prostředí. Díky tomu na nich do současnosti přežilo množství vzácných a úzce
specializovaných reliktních druhů, které nejsou vlivem omezené schopnosti migrace a
pasivního šíření schopny efektivně kolonizovat nově vzniklá stanoviště (neorefugia)
podobného typu (HÁJEK et al. 2011).
Existence gradientu historického vývoje pomáhá pochopit fakt, že významnost faktorů
působících na skladbu společenstev má vždy prostorově omezenou platnost a závisí na
škále, v rámci níž společenstva studujeme. Je zřejmé, že bez znalosti historického a
krajinného kontextu, pouze na základě stanovení lokálních podmínek prostředí, by nebylo
možné vysvětlit variabilitu společenstev větších a geograficky heterogenních území. O
regionálních rozdílech ve společenstvech na velkých prostorových škálách bude dále
pojednáno v kapitole 3 (str. 12) o latitudinálním gradientu.

7
2.3. Malakofauna slatiniš ť
2.3.1. Měkkýši jako modelová skupina
Měkkýši, především suchozemští plži, představují z mnoha důvodů ideální skupinu
modelových organizmů. Determinace měkkýšů do druhové úrovně je většinou možná i bez
studia anatomických znaků, pouze na základě schránky nebo dokonce jejích úlomků. Také
jejich biologie, stanovištní nároky a rozšíření jsou v Evropě poměrně dobře prostudovány.
Většina druhů je stenoekních a kvůli úzké vazbě na specifická stanoviště, omezené
pohyblivosti a nízké schopnosti aktivního šíření jen velmi málo odolných vůči disturbancím
a změnám v podmínkách prostředí (PAWŁOWSKA & POKRYSZKO 1998). Z uvedených
důvodů lze na měkkýše pohlížet jako na spolehlivé bioindikátory podmínek prostředí (např.
BARKER 2001, FOECKLER et al. 2006). Díky zachování schránek ve vápnitých sedimentech
se jejich fosilie také hojně využívají pro paleorekonstrukci klimatu a vývoje krajiny
v průběhu kvartéru (např. LOŽEK 1964, ROUSSEAU et al. 1994, HORSÁK 2011). Podrobné
znalosti o ekologii měkkýšů mohou významně posloužit jak vědeckým, tak ochranářským
účelům (LOŽEK 1981).
2.3.2. Faktory ovliv ňující diverzitu m ěkkýš ů slatiniš ť
Mezi biotopy s vůbec nejvyšší rozmanitostí malakocenóz patří bazická slatiniště.
Setkáváme se na nich s pestrou mozaikou měkkýšů otevřených stanovišť, obohacenou o
druhy silně vlhkomilné, vodní, kalcifilní a taktéž o slatiništní specialisty a glaciální relikty
(HORSÁK & CERNOHORSKY 2008). Slatiniště jsou navíc charakteristická mimořádně
vysokými abundancemi měkkýšů na velmi malých plochách, neboť zde zcela převažují
drobné druhy se schránkami menšími než 5 mm (HORSÁK 2005a). Příčinou dominance
drobných velikostních forem je pravděpodobně nízká produktivita a homogenita slatiništní
vegetace, spjatá s absencí potenciálních úkrytů pro přežití mrazů a výrazně limitující
dlouhodobější přežívání velkých druhů plžů (SCHAMP et al. 2010).
Měkkýši se podobně jako rostliny vyznačují jen omezenou pohyblivostí a úzkou
vazbou na konkrétní stanoviště, a jejich výskyt na slatiništích je proto předurčen
analogickým komplexem podmínek prostředí a ekologických gradientů (HORSÁK & HÁJEK
2003). Skladbu vegetace je dokonce možné využít jako nejkvalitnější prediktor druhového
složení malakocenóz (HORSÁK & HÁJEK 2003, HORSÁK et al. 2011a). Složení rostlinných

8
společenstev je totiž daleko méně variabilní v čase než jednorázově měřené proměnné
prostředí, a odráží kromě aktuálních podmínek také jejich dlouhodobý vývoj působící na
finální podobu slatiništní bioty (HÁJEK et al. 2006).
Diverzita malakofauny na slatiništích plynule narůstá podél gradientu minerální
bohatosti a největších hodnot dosahuje na bazických slatiništích se srážením uhličitanu
vápenatého ve formě pěnovce (HORSÁK & HÁJEK 2003). Zvláště suchozemští plži jsou
charakterističtí svou preferencí vápnitých substrátů a silná korelace počtů druhů i
abundancí s množstvím dostupného vápníku byla dokumentována ve studiích
z nejrůznějších typů biotopů (např. WÄREBORN 1970, WALDÉN 1981, POKRYSZKO 1993,
HYLANDER et al. 2005, SULIKOWSKA-DROZD & HORSÁK 2007). Vápník je prvek nezbytný
především pro stavbu měkkýších schránek a pozitivně působí také na rozmnožování a
další fyziologické procesy (např. WÄREBORN 1970, 1979, FOURNIÉ & CHÉTAIL 1984, LUCHTEL
& DEYRUP-OLSEN 2001). Podobně významnou roli hraje také vlhkost stanoviště, která se
s ohledem na náchylnost měkkýších těl k vyschnutí může za určitých podmínek stát
limitujícím faktorem (WÄREBORN 1969, MARTIN & SOMMER 2004). To se však netýká
slatinišť, která jsou trvale vlhká, a o jejich druhové bohatosti proto rozhoduje především
množství dostupného vápníku (např. POKRYSZKO 1993, HORSÁK & HÁJEK 2003, HORSÁK
2006).
Místo kompletní výměny druhové skladby, charakteristické pro slatiništní vegetaci
(např. HÁJEK et al. 2002, TAHVANAINEN 2004), však podél gradientu minerální bohatosti
pozorujeme spíše plynulé přibývání druhů s užší ekologickou valencí a silnější vazbou na
vysoké množství dostupného vápníku (HORSÁK 2005a, HORSÁK & CERNOHORSKY 2008).
Nutno zdůraznit, že i tento trend má svá omezení a na nejextrémnějších stanovištích
v podobě vápnitých slatinišť s vysokou koncentrací solí a tvorbou travertinu dochází
k prudkému poklesu druhové bohatosti měkkýšů (HORSÁK 2005a, HORSÁK 2006, HORSÁK &
CERNOHORSKY 2008). Jen velmi málo specializovaných kalcifilních druhů je schopno přežít
v těchto extrémních podmínkách, daných vysokou koncentrací toxických iontů a absencí
mikrostanovišť vlivem kompletního pokrytí povrchu krustou z vysráženého travertinu
(HORSÁK 2006).
To, co bylo popsáno pro rostlinná společenstva a jejich vztah k měnící se fertilitě
slatiniště (trofickému gradientu), platí obdobně i pro společenstva měkkýšů. Při zvýšeném
přísunu živin a nárůstu produktivity vegetace mizí vlivem zastínění a narušení
hydrologického režimu náročné slatinné druhy a nahrazují je nespecializovaní ubikvisté,
luční, křovinné a lesní druhy (HÁJEK et al. 2006). Pozoruhodná shoda v ekologii rostlin a
měkkýšů je patrná také při bližším studiu rozšíření druhů ve vztahu ke stáří lokalit a

9
geografickému gradientu. Recentněji vzniklé lokality (neorefugia) představují refugia na
úrovni společenstev s unikátní druhovou kombinací nemající obdobu v jiných biotopech
(HORSÁK 2005a). Pouze historicky starší slatiniště (paleorefugia) jsou však typická také
výskytem glaciálních reliktů, které na těchto izolovaných biotopech přežily z pozdního
glaciálu až do současnosti. Vztah mezi druhovým složením a stářím stanovišť byl
prostudován na slatiništích Západních Karpat, kde se reliktním rozšířením vyznačovaly
druhy Vertigo geyeri a Pupilla alpicola (HORSÁK et al. 2007a, HÁJEK et al. 2011).
Vzhledem k popsaným aspektům vegetace a malakocenóz je často klíčem
k pochopení podmínek a procesů formujících společenstva slatinišť souběžné studium
těchto taxonomicky velmi vzdálených, a přesto ekologicky velmi blízkých skupin
organizmů. Kombinace malakologického a botanického přístupu se logicky vhodně
doplňuje a umožňuje získat komplexní představu o fungování slatiništních ekosystémů.
2.3.3. Výzkumy slatiništních malakocenóz v Evrop ě
S ohledem na fragmentární a roztroušený charakter byla slatiniště střední Evropy
z malakologického hlediska v minulosti spíše přehlížena. Pozornost byla věnována studiu
jiných, v krajině běžnějších a snadněji dosažitelných biotopů, a podrobných studií úzce
zaměřených na ekologii recentních slatiništních malakocenóz je spíše poskrovnu.
Poznatky o fauně měkkýšů na slatiništích jsou součástí několika prací z oblasti Velké
Británie (např. CAMERON 1978, DAVIES et al. 1996), zahrnujících i studium dalších
terestrických biotopů. Poněkud podrobněji se na malakofaunu slatinišť Britských ostrovů
zaměřili BISHOP (1981) a AUSDEN et al. (2005). Z dalších oblastí Evropy pochází spíše
omezené množství studií zabývajících se především ekologií jednotlivých druhů (viz
SPEIGHT et al. 2003). Výjimku tvoří region Západních Karpat, kde již více než 15 let
probíhají intenzivní a podrobné výzkumy ekologie společenstev měkkýšů slatinišť se
zaměřením na vliv rozmanitých ekologických gradientů, vztahy mezi malakocenózami a
vegetací a reliktní povahu slatinišť (např. HORSÁK & HÁJEK 2003, HÁJEK et al. 2006,
HORSÁK et al. 2007a, b, HORSÁK & CERNOHORSKY 2008, HÁJEK et al. 2011). Teprve velmi
nedávno byla prozkoumána také slatiniště Bulharska (HORSÁK et al. 2011a).
V severní Evropě jsou znalosti o slatiništní malakofauně o něco úplnější, neboť
slatiniště zde pokrývají obrovské plochy a představují tak jeden z velmi charakteristických
biotopů této krajiny. Údaje o měkkýších slatinišť jsou součástí výsledků mnoha
faunistických výzkumů, které probíhaly na území Norska a Švédska ve 20. století (např.
KERNEY & CAMERON 1979, ANDERSEN & HALVORSEN 1984, WALDÉN 1995). Celá řada autorů

10
se také ve svých studiích věnovala detailně právě biotopu slatinišť (např. NILSSON 1987,
POKRYSZKO 1993, VON PROSCHWITZ 2003). I z oblasti Skandinávie však stále chybí
podrobné studie zabývající se ekologickými determinanty druhové skladby měkkýších
společenstev.
2.4. Počet druh ů, produktivita a slatiništ ě
Ačkoli i tato podkapitola souvisí s jedním z cílů této práce, totiž s porozuměním vztahu
mezi druhovou bohatostí a produktivitou (hmotností biomasy) společenstev měkkýšů na
slatiništích, bude nutné ji pro pochopení potřebných souvislostí pojmout více ze široka, a
poukázat tak na obecně platné závěry použitelné pro následné srovnání se získanými
výsledky.
Vztah druhové diverzity a produktivity (hmotnosti biomasy) společenstev je poměrně
kontroverzní a již několik desetiletí živě diskutovanou ekologickou otázkou (např. PIANKA
1966, HUSTON 1979, TILMAN et al. 1996), které se podrobně věnují desítky prací
zaměřených především na rostlinná společenstva (WAIDE et al. 1999, MITTELBACH et al.
2001). Situace je komplikovaná a vztah počtu druhů a rostlinné biomasy může být jak
pozitivně, tak i negativně rostoucí, v závislosti na mnoha faktorech a okolnostech
provedení výzkumu (viz např. GRACE 1999). Za obecně platný a nejčastěji pozorovaný
model je však považována unimodální závislost s maximem druhové bohatosti při
středních hodnotách produktivity (např. GRIME 1973, AL-MUFTI et al. 1977, GRACE 1999).
Nízká druhová bohatost je přičítána silně stresovým podmínkám a disturbancím při
nízkých hodnotách produktivity, a opětovný pokles druhové bohatosti směrem k vyšší
produktivitě je vysvětlován mezidruhovou kompeticí – ve vysokoproduktivním
společenstvu převládne jen několik málo silně dominantních druhů, které vyloučí druhy
kompetičně slabší (GRIME 1979). Bylo ovšem zjištěno, že samotné kompetiční vyloučení
nemusí být příčinou pozorovaného poklesu druhové bohatosti směrem k vyšší
produktivitě. Velmi významná může být role tzv. „species-pool“, tedy množství druhů
v okolní krajině potenciálně schopných kolonizovat daný typ biotopu (TAYLOR et al. 1990,
ZOBEL 1997). Nízká druhová bohatost tak může být pouze důsledkem omezeného počtu
druhů adaptovaných na přežití v podmínkách vysoké produktivity, především v regionech,
kde jsou tyto vysokoproduktivní typy stanovišť vzácné (PÄRTEL et al. 2007, AXMANOVÁ et al.
odesláno). Oba zmíněné mechanizmy, kompetice i vliv „species pool“ se mohou doplňovat
a působit současně (SAFFORD et al. 2001).

11
Unimodální závislost je převažujícím výsledkem i v pracích zabývajících se vegetací
slatinišť (např. WHEELER & GILLER 1982, OLDE VENTERINK et al. 2001, 2003). Vzhledem
k mnoha ekologickým analogiím propojujícím společenstva rostlin a měkkýšů tak vyvstává
otázka, zda se budou zmíněné taxony shodovat i svou odpovědí na rostoucí množství
biomasy. Na rozdíl od rostlin pravděpodobně nebude mít na tvar křivky vliv kompetiční
vyloučení, neboť se předpokládá, že kompetice hraje ve společenstvech měkkýšů spíše
méně významnou roli (BOYCOTT 1934, WALDÉN 1981). V současnosti však neexistuje
studie týkající se terestrických společenstev měkkýšů, produktivity a druhové bohatosti.
Mé výsledky budou proto zcela nové a s ohledem na velikost datového souboru spíše
orientační. Vzhledem k úzkému zaměření a omezenému rozsahu studie není cílem získat
zobecnitelné a široce aplikovatelné závěry, ale především poukázat na možné trendy a
vlastnosti společenstev suchozemských měkkýšů z hlediska doposud neprobádaného
vztahu počtu druhů a produktivity.

12
3. ZMĚNY DIVERZITY PODÉL LATITUDINÁLNÍHO GRADIENTU
3.1. Obecný úvod k latitudinálnímu gradientu
Jedním ze základních, univerzálních a nejdéle známých makroekologických fenoménů
je pokles druhové bohatosti směrem k vyšším zeměpisným šířkám, označovaný jako
latitudinální gradient diverzity (např. ROSENZWEIG 1995, BROWN & LOMOLINO 1998, WILLIG et
al. 2003, HILLEBRAND 2004). Přestože existence latitudinálního gradientu zásadním
způsobem ovlivňuje rostlinná i živočišná společenstva nejrůznějších ekosystémů, jeho
příčiny nebyly doposud uspokojivě objasněny a nalezení obecně akceptovatelného
vysvětlení představuje dlouhodobou výzvu pro ekology a biogeografy (např. PIANKA 1966,
ROHDE 1992, GASTON 2000, CARDILLO et al. 2005).
Ve snaze odhalit obecný mechanismus odpovídající za latitudinální gradient diverzity
bylo navrženo a testováno velkého množství hypotéz. Recentní studie a meta-analýzy
rozsáhlých datových souborů však naznačují, že spíše než o jediném univerzálním
mechanismu je třeba uvažovat o spolupůsobení mnoha vzájemně se doplňujících faktorů,
ovlivňujících výsledný pozorovaný trend (WHITTAKER et al. 2001, WILLIG et al. 2003).
Velká skupina hypotéz poukazuje na souvislosti mezi druhovou bohatostí a
klimatickými faktory . Vyšší přísun solární energie, dostupnost vody, teploty blízké
fyziologickým optimům mnoha druhů a relativní stabilita klimatu vedou k vyšší druhové
bohatosti v oblastech blíže rovníku. Příznivé a stálé klima bez výraznějších sezónních
výkyvů podporuje vznik početnějších populací organizmů, snižuje riziko extinkce,
umožňuje vyšší míru specializace, diverzifikaci ekologických nik a vznik nových druhů
(HAWKINS et al. 2003).
Další skupina hypotéz klade důraz především na geografické a prostorové faktory .
Geografické teorie vychází z faktu, že oblasti kolem rovníku jsou plošně nejrozsáhlejší, a
tedy druhově nejbohatší, díky obecně platnému vztahu mezi počtem druhů a plochou,
nižšímu riziku extinkce a vyšší míře speciace (TERBORGH 1973, ROSENZWEIG 2003). Jiné
modelové vysvětlení představuje tzv. efekt středu domény, předpokládající čistě náhodný
překryv výskytů jednotlivých druhů ve středu geograficky ohraničeného území (domény),
na Zemi tedy právě v oblastech kolem rovníku (COLWELL & HURTT 1994, COLWELL & LEES
2000, CONNOLLY 2005).
Silnou podporu si díky množství věrohodných důkazů získávají také historické a
evolu ční teorie . Podle evolučních teorií se vlivem vyšších teplot v nižších zeměpisných

13
šířkách zrychlují fyziologické procesy, zkracuje generační čas a zvyšuje mutační rychlost a
selekční tlak. Uvedené faktory podporují mikroevoluční procesy, následnou speciaci a
vznik nových druhů (ROHDE 1992). Historická vysvětlení zohledňují především čas, po
který jednotlivé ekosystémy setrvávají ve stabilním stavu, bez výrazných disturbancí a
výkyvů v podmínkách prostředí. Oblasti vyšších zeměpisných šířek byly v minulosti více
postiženy změnami klimatu a rozsáhlým zaledněním, čímž se zpomalila kolonizace a
následná speciace druhů (CLARKE & CRAME 2003).
Navrženo bylo velké množství dalších hypotéz, stejně tak jako empiricky dobře
podložených argumentů popírajících jejich platnost a poukazujících na řadu zjevných
výjimek. Vyčerpávající shrnutí veškerých dostupných poznatků by vzhledem k rozsahu
problematiky dalece přesahovalo rámec této diplomové práce. Uvedený výčet má sloužit
pouze jako stručný přehled nejvýznamnějších recentně uznávaných teorií vysvětlujících
příčiny latitudinálního gradientu diverzity, který bude dále rozebírán v souvislosti se
společenstvy měkkýšů. Podrobné informace o aktuálních přístupech k tématice
latitudinálního gradientu shrnují např. práce WILLIG et al. 2003, HILLEBRAND 2004,
JABLONSKI et al. 2006, MITTELBACH et al. 2007.
3.2. Měkkýši a latitudinální gradient
Existence latitudinálního gradientu byla opakovaně dokumentována pro nejrůznější
skupiny organismů, ačkoli práce zabývající se bezobratlými živočichy jsou spíše
v menšině. Měkkýši však tvoří výjimečně vhodnou modelovou skupinu pro tento typ studií,
neboť navzdory vysoké taxonomické rozmanitosti jsou jednotlivé druhy dobře popsány a
metodika výzkumu spjatá s determinací do druhové úrovně je daleko méně problematická
než u jiných taxonů bezobratlých (NEKOLA 2005).
Negativní vztah mezi druhovou bohatostí a zeměpisnou šířkou byl studován
především u měkkýšů oceánských a mořských ekosystémů (např. TAYLOR & TAYLOR 1977,
ROY et al. 1996, JABLONSKI et al. 2006). Zkoumána však byla také fauna měkkýšů
terestrických biotopů, přičemž studie z Evropy i Severní Ameriky potvrdily pokles počtu
druhů měkkýšů směrem k vyšším zeměpisným šířkám (WALDÉN 1995, HAUSDORF 2006,
NEKOLA 2005). Zjištěn byl i pokles β-diverzity společenstev, což zjednodušeně znamená,
že společenstva ve vyšších zeměpisných šířkách si jsou vzájemně podobnější (NEKOLA
2005). HAUSDORF (2006) usuzuje, že současné rozšíření evropských měkkýšů je silně
ovlivněno pleistocénním zaledněním, které většina druhů přežila v glaciálních refugiích na
jihu Evropy (HAUSDORF & HENNIG 2003). V holocénu tyto druhy následně expandovaly do

14
vyšších zeměpisných šířek, a severní hranice jejich areálů jsou dány schopnostmi šíření a
rozdílnou ekologickou tolerancí co se klimatických podmínek týče.
Při studiu obecně platných skutečností je vždy nutné uvažovat o působení rozmanité
škály ekologických faktorů a existenci „výjimek potvrzujících pravidlo“. ANDERSEN &
HALVORSEN (1984) např. dokumentovali prudký pokles počtu druhů podél
východozápadního gradientu, na úrovni stejné zeměpisné šířky a na zcela shodném typu
biotopu, způsobený rozdíly v mírném oceánickém a drsném kontinentálním klimatu.
VALDOVINOS et al. (2003) prokázali, že diverzita měkkýšů podél Pacifického šelfu v jižní
Americe směrem k severním šířkám roste, což je v přímém protikladu s latitudinálním
gradientem diverzity. Uvedené příklady poukazují na fakt, že zcela univerzální trend
v ekologii společenstev najdeme jen stěží, a při interpretaci výsledků je nezbytné důsledně
zohlednit okolnosti provedení výzkumu a případná specifika studované taxonomické
skupiny.
Změny společenstev podél latitudinálního gradientu se netýkají pouze druhové
bohatosti – u terestrických měkkýšů byly také zaznamenány rozdíly ve velikostní struktuře.
Ukázalo se, že směrem k vyšším zeměpisným šířkám roste zastoupení drobných druhů ve
společenstvech suchozemských ulitnatých plžů (HAUSDORF 2003, NEKOLA 2005, NEKOLA et
al. v tisku). Toto zjištění je v rozporu s Bergmannovým pravidlem, podle kterého velikost
těla v rámci skupiny blízce příbuzných organizmů pozitivně koreluje se zeměpisnou šířkou
(BERGMANN 1847, BLACKBURN et al. 1999). U suchozemských plžů tento jev
pravděpodobně souvisí s extrémně nízkou schopností aktivní migrace a naopak
převažujícím pasivním způsobem šíření. Pasivní disperze je snadnější právě pro drobné
taxony (HAUSDORF & HENNIG 2003), které jsou efektivnějšími kolonizátory i díky častějšímu
výskytu samooplození (POKRYSZKO & CAMERON 2005). Výhodou drobnějších taxonů je také
vyšší pravděpodobnost přežití mrazů díky nižšímu objemu vody v těle (ANSART & VERNON
2003, NEKOLA et al. v tisku). Podél severojižního gradientu se mění také procento
zastoupení některých taxonů ve společenstvech. Např. čeleď Vertiginidae dosahuje
nečekaně nejvyšší diverzity v severských boreálních regionech (VON PROSCHWITZ 2003,
POKRYSZKO & CAMERON 2005). Tyto rozdíly mohou také přispívat k vysvětlení
pozorovaných latitudinálních trendů ve velikostní struktuře (HAUSDORF 2003).
Neexistuje doposud žádná studie zaměřená na vliv latitudinálního gradientu na
malakofaunu slatinišť, ačkoli z hlediska diverzity a druhové skladby měkkýšů se jedná o
zcela vyhraněné a unikátní biotopy. Zjistit, zda malakocenózy slatinišť odpovídají na
latitudinální gradient obdobně jako společenstva jiných biotopů, či zda mají slatiniště svá
zjevná specifika (daná např. izolovaností, historickou kontinuitou, převahou velmi malých

15
druhů apod.), bude jedním z významných cílů této diplomové práce. Zkoumané lokality
svým geografickým rozsahem (od 49˚ do 63° s.š.) zah rnují přechod mírného pásu do
boreální zóny, a pokrývají tak poměrně širokou geografickou škálu, umožňující sledovat
potenciální změny ve společenstvech měkkýšů podél latitudinálního gradientu.

16
4. CHARAKTERISTIKA STUDOVANÝCH OBLASTÍ
4.1. Polsko
Obsah diplomové páce bude zaměřen na východní část Polska, která se vyznačuje
mimořádně vysokou koncentrací zachovalých slatiništních ekosystémů. Většina rozlohy
zkoumaného území náleží do rozsáhlého pásu středoevropských nížin, pouze na
jihovýchod zasahuje okrajově karpatský oblouk. Reliéf nížinné polské krajiny nese zjevné
stopy činnosti Skandinávského ledovce z období opakovaných pleistocénních zalednění
(MARKS 2004). Důsledky glaciálních geomorfologických procesů jsou nejvíce patrné
v severní části země, kam zasahoval ledovec i při posledním, Viselském zalednění
(MARKS 2010). Mezi charakteristické rysy mladé postglaciální krajiny Polska patří
morénové valy, mírně zvlněný povrch s velkým množstvím ledovcových jezer, hustá říční
síť a rozsáhlé mokřadní komplexy (HILLBRICHT-ILKOWSKA et al. 2000).
Klimaticky spadá Polsko do zóny mírného pásu, ačkoli severovýchod území má
z hlediska podnebí a skladby vegetace již spíše hemiboreální ráz (SJÖRS 1965a). Pro
polské klima je typická vysoká variabilita a přechodný charakter, neboť na území dochází
ke střetu chladného arktického a vlhkého atlantického proudění vzduchu. Obecně je
podnebí severní a západní části Polska spíše vlhké, oceánické a teplotně vyrovnanější,
zatímco východ a jih Polska je sušší a více kontinentální, vyznačující se většími teplotními
extrémy a sezónními rozdíly.
Opakovaná kvartérní zalednění měla vliv nejen na modelaci reliéfu polské krajiny, ale
také na horninovou skladbu geologického podloží. Na severovýchod zasahoval i v období
posledního glaciálu jižní okraj pevninského Skandinávského ledovce, a povrch tohoto
území proto v současnosti pokrývají štěrkové, písčité a jílovité nánosy, vzniklé procesy
glaciofluviální, glaciolakustrinní a glaciální sedimentace (BER 2006). Centrální část
východního Polska představuje krajinu geologicky výrazně starší, pokrytou vápnitými
sedimenty ze svrchní křídy, sestávajícími především z jílů, křídy a vápence. V některých
oblastech dominují vápníkem bohaté miocénní mořské sedimenty a výjimkou nejsou ani
mocné vrstvy spraší. Na jihovýchod Polska zasahuje svým severním okrajem oblouk
Západních a Východních Karpat. Karpatské pohoří vznikalo alpínským vrásněním
v průběhu svrchní křídy a třetihor a jeho geologická stavba je velmi rozmanitá a složitá.
Převážná část karpatského oblouku spadá v Polsku do vnějšího flyšového pásma
tvořeného souvrstvími střídajících se pískovců a jílovců. Částečně na území Polska vybíhá

17
i jádrové pohoří složené z prvohorních krystalických břidlic, žul a granitoidů, překrytých
vrstvami vyvrásněných druhohorních sedimentů. Kromě orogenních procesů ovlivnila
geologickou skladbu nejjižnější hornaté části Polska také přítomnost horských ledovců
v průběhu kvartérních zalednění.
4.1.1. Variabilita polských slatiniš ť
Slatiniště jsou na území Polska zcela převažujícím typem rašeliništních ekosystémů
(SIENKIEWICZ 2008). Především ve východní a severovýchodní části země se dosud
vyskytují velmi rozsáhlé a na středoevropské poměry výjimečně zachovalé slatiništní
komplexy, typické přítomností mnoha vzácných a ohrožených rostlinných druhů (např.
PAWLIKOWSKI et al. 2009, JABŁOŃSKA et al. 2011). Současná rozloha však představuje
pouze zlomek z původní plochy pokryté slatiništi, neboť naprostá většina rašeliništních
ekosystémů byla v průběhu posledních desetiletí zničena vlivem intenzivního
zemědělského a průmyslového využití krajiny. Odhadované ztráty původní rozlohy
rašelinišť činí v Polsku přibližně 84 % (WOLEJKO et al. 2005), přičemž rašeliniště a
mokřady patří stále mezi extrémně ohrožené a nadále mizející biotopy (např. GROOTJANS
et al. 2006, OLDE VENTERINK et al. 2009). K ochraně rašelinišť přispívá zejména existence
soustavy přírodních rezervací, chráněných území, národních parků a území zahrnutých do
soustavy Natura 2000 (GROOTJANS & WOLEJKO 2007).
Nejrozsáhlejší, lidskou činností dosud nenarušené komplexy slatinišť se nacházejí v
mladé postglaciální krajině severního Polska, v okolí četných jezer a v rozlehlých aluviích
velkých říčních toků. Velká část z nich náleží díky vápnitému podloží a minerálně bohaté
podzemní vodě mezi bazická slatiniště (WOLEJKO et al. 2005). Další oblastí s vysokou
koncentrací bazických slatinišť je okolí města Chełm na jihovýchodě Polska, pokryté
vápencovými sedimenty ze svrchní křídy. Ve zbytku nížinné části východního Polska se
minerálně bohatá slatiniště vyskytují spíše roztroušeně a ojediněle na vápnitém
horninovém podkladu, obvykle na průsacích podzemní vody v terénních sníženinách a
říčních údolích (WOLEJKO et al. 2005). Poslední oblastí nápadně bohatou na slatiniště je
hornatý jihovýchod Polska náležící ke karpatskému oblouku a jeho podhůří. Setkáváme se
zde především s menšími bazickými svahovými prameništi, neboť geomorfologie a reliéf
terénu neumožňují vznik plošně rozsáhlejších slatinišť. Výjimku tvoří území Oravsko-
Nowotarské kotliny na úpatí Tater, kde však byla značná část cenných rašeliništních
biotopů a jejich biodiverzity nenávratně zničena vlivem destruktivních dopadů těžby
rašeliny (KOCZUR 2006).

18
4.2. Jižní Skandinávie
Pro diplomovou práci budou použity také výsledky práce bakalářské, v níž byla
studována slatiniště Norska a Švédska. Obě země se rozkládají na Skandinávském
poloostrově a jejich jižní části spadají převážně do boreální a hemiboreální zóny. Výjimku
představuje nejjižnější cíp Švédska, který se klimatem, geologií i skladbou vegetace
výrazně podobá střední Evropě, a je proto součástí zóny mírného pásu (SJÖRS 1965a,
DIERSSEN 1996). Současný charakter skandinávské bioty je velmi mladý, neboť byl
formován až v průběhu holocénu po ústupu Skandinávského ledovce. Opakovaná
pleistocénní zalednění se zásadním způsobem promítla také do geologie a geomorfologie
Skandinávie. Pozůstatky ledovcové činnosti v podobě členitého pobřeží, fjordů, morén, či
glaciálních jezer patří mezi nezaměnitelné rysy severoevropské krajiny.
Klimatické poměry Skandinávie jsou vlivem Golfského proudu a teplého jihozápadního
proudění větru daleko příznivější, než je pro tyto zeměpisné šířky typické. Díky tomu se
pobřeží Norska vyznačuje mírným oceánickým podnebím a jen relativně malými teplotními
rozdíly podél severojižního gradientu, zatímco východ Norska a Švédsko jsou více
kontinentální. Především severovýchodní část Skandinávie je v zimě ovlivňována
přísunem studeného vzduchu z polárních oblastí, zatímco na jih Švédska působí teplé
atlantické proudění. Severojižní gradient teplotních rozdílů je proto ve Švédsku daleko
výraznější než v Norsku.
Po celé délce západní části skandinávských zemí se táhne hřbet skandinávského
pohoří, díky kterému se norský terén vyznačuje mimořádnou hornatostí. Pohoří zasahuje
značnou částí také na území Švédska, kde však dosahuje pouze nižších nadmořských
výšek. Skandinávské pohoří vznikalo v průběhu prvohorního Kaledonského vrásnění a
jeho geologické podloží tvoří mozaika kambro-silurských hornin, jako jsou žuly, ruly, fylity,
svory, místy i horniny s vysokým obsahem vápníku, např. slepence, vápence a dolomity.
V nižších polohách směrem na východ a na jih od skandinávského pohoří došlo vlivem
rozsáhlých zalednění a glaciální eroze k obroušení a odplavení málo odolných vápnitých
sedimentů a obnažení hornin prekambrijského stáří. Toto tzv. krystalinické podloží je
extrémně minerálně chudé, tvořené převážně kyselými žulami a rulami. Lokálně se však i
zde vyskytují fragmenty minerálně bohatého podloží tvořené horninami s vysokým
obsahem vápníku. Nejrozsáhlejší plochy kambro-silurských vápnitých sedimentů se
zachovaly v provincii Skåne na jižním pobřeží Švédska a ve vnitrozemské provincii
Jämtland (SJÖRS 1965b).

19
4.2.1. Variabilita skandinávských slatiniš ť
Chladné a vlhké boreální klima, vhodná topografie terénu a postglaciální charakter
krajiny s mnoha stopami pleistocénní ledovcové činnosti, přispěly ve Skandinávii ke vzniku
velmi početných a rozsáhlých komplexů rašelinišť. Ačkoli především v nížinných částech
skandinávských zemí mnohá rašeliniště zanikla v důsledku odvodňování, pastvy a
zemědělství, destrukce mokřadních biotopů zde neměla zdaleka takový rozsah jako ve
střední Evropě (GROOTJANS et al. 2006). V Norsku i ve Švédsku se i přes značné ztráty
zachovala více než polovina z původní rozlohy rašelinišť (JOOSTEN & CLARKE 2002).
Kromě klimatu a reliéfu terénu předurčuje charakter rašeliništních ekosystémů
především horninová skladba geologického podloží, udávající chemické parametry
podzemní vody (TAHVANAINEN 2004). Ve Skandinávii, kde převažuje nevápnité
krystalinické podloží, se rašeliniště obvykle omezují na kyselá ombrotrofní vrchoviště a
minerálně chudá slatiniště s dominancí rašeliníků. Na většině území proto reprezentují
bazický konec gradientu minerální bohatosti slatiniště s kalcitolerantními rašeliníky (HÁJEK
et al. 2006).
Bazická slatiniště s vegetací tvořenou převážně hnědými mechy jsou na území Norska
a Švédska velmi vzácná, situovaná výhradně do oblastí s minerálně bohatším horninovým
podkladem. Jak již bylo výše zmíněno, jedním z takových míst je provincie Skåne na jihu
Švédska. Přítomnost písčitých, jílovitých a křídových sedimentů zde umožnila vznik
vysoce bazických slatinišť, často dokonce se srážením uhličitanu vápenatého (MALMER
1965). Vápnitá pěnovcová slatiniště představují v rámci Skandinávie vskutku ojedinělý
fenomén, typický dále jen pro provincii Jämtland ve středním Švédsku. Bazická slatiniště
se zde utvořila na mocných nánosech vápníkem bohatých jílovitých sedimentů,
zatlačených na toto území masami ledovce v průběhu pleistocénních zalednění (SJÖRS
1965b). Bazická slatiniště (bez srážení pěnovce) se lokálně vyskytují i v hornaté krajině
Norska, např. na minerálně bohatých horninách v Národním parku Dovrefjell (POKRYSZKO
1993).

20
5. METODIKA PRÁCE
5.1. Studované lokality
V rámci diplomové práce bylo studováno celkem 39 bazických slatinišť na území
Polska (Obr. 1) a 23 bazických slatinišť na území jižní Skandinávie (Obr. 2). Bazická
slatiniště byla definována jako lokality: (1) s dominancí hnědých mechů a úplnou absencí
rašeliníků, tedy vápnitá pěnovcová a extrémně bohatá slatiniště bez srážení pěnovce; (2)
s výskytem hnědých mechů i kalcitolerantních druhů rašeliníků, pokud jejich konduktivita
přesahovala hodnotu 100 µS*cm-1. Zaměření výzkumu na bazická slatiniště si kladlo za cíl
vytvořit srovnatelné datové soubory zahrnující lokality s nejvyšší diverzitou měkkýšů
v obou oblastech, a částečně také odfiltrovat vliv gradientu minerální bohatosti, který byl
detailně prozkoumán v celé řadě předcházejících studií včetně bakalářské práce. Omezení
výzkumu na bazická slatiniště umožňuje zdůraznit působení dalších ekologických faktorů,
projevujících se především na větších prostorových škálách.
Pro výzkum byla vybrána zachovalá otevřená slatiniště sycená minerálně bohatou
podzemní vodou. Na území Polska se jednalo o první komplexní průzkum společenstev
měkkýšů slatinišť – až doposud zde byly malakologické studie zaměřeny na jiné typy
biotopů, a slatiniště byla z hlediska druhové skladby malakocenóz zcela přehlížena.
Vzorkování bylo provedeno ve východní části země, která je charakteristická relativně
vysokou koncentrací rozsáhlých a zachovalých slatiništních komplexů (WOLEJKO et al.
2005). Přesto však na mnoha místech, především ve střední nížinné části východního
Polska, představují zachovalá bazická slatiniště jen velmi roztroušené izolované ostrůvky,
obklopené převážně zemědělsky využívanou a průmyslovou krajinou. Možnost pokrýt
oblast vzorkovanými lokalitami rovnoměrně podél severojižního gradientu tak byla
omezena absencí vhodných biotopů v některých částech studovaného území (Obr. 1).
5.2. Terénní práce a zpracování vzork ů
Seznam všech vzorkovaných lokalit je uveden v Příloze 1 (str. 68). Do datového
souboru polských slatinišť byly zahrnuty celkem 4 lokality (P1, P2, P6, P10) vzorkované M.
Horsákem v letech 2002–2008 v rámci rozsáhlých průzkumů slatinišť Západních Karpat. V
červenci 2010 jsem v průběhu terénního symposia pořádaného Mezinárodní organizací
pro ochranu rašelinišť (IMCG) navštívila dalších 6 polských lokalit (P12–15, P19, P38),

21
rozmístěných od jihu až po severní hranici s Ruskem. V květnu a červnu 2011 bylo na tyto
pilotní průzkumy navázáno systematickým vzorkováním polských bazických slatinišť pro
účely mé diplomové práce, s cílem detailně poznat specifika druhové skladby
společenstev měkkýšů a nasbírat reprezentativní soubor dat pro srovnání druhové skladby
malakocenóz podél gradientu zeměpisné šířky, od jižního Polska až po boreální zónu
Skandinávie. Studované polské lokality se nacházely v geografickém rozmezí
49˚17’01,7”–54˚21’03,4” s.š. a 19˚39’17,1”–23˚36’0,1” v.d. (Obr. 1). Vzorky měkkýšů na
území Polska jsem odebírala společně s M. Horsákem. Pro diplomovou práci bylo dále
vybráno 23 bazických slatinišť na území jižní Skandinávie, zpracovaných v rámci
bakalářské práce (11 lokalit v oblasti Švédska, 12 lokalit v oblasti Norska). Vzorkování
skandinávských lokalit proběhlo v srpnu 2006, lokality se nacházely v geografickém
rozmezí 55˚31’35,2”–63˚43’21,0” s.š. a 09˚35’07,8”–15˚23’33,2” v.d. (Obr. 2). Vzorky
měkkýšů ve Skandinávii odebírali M. Horsák a N. Černohorská.
Obrázek 1. Mapa území Polska s vyznačenými studovanými lokalitami (charakteristika lokalit viz Příloha 1,
str. 68).

22
Obrázek 2. Mapa území jižní Skandinávie s vyznačenými studovanými lokalitami (charakteristika lokalit viz
Příloha 1, str. 68).
V centrální části každé lokality byla vymezena vegetačně homogenní plocha o
rozměrech 4 x 4 m2, ze které byl odebrán vzorek o objemu 12 l, zahrnující svrchní vrstvu
půdy, rostlinný opad, mechy i cévnaté rostliny. Měkkýši byli ze vzorku získáni metodou
mokrého prosevu (HORSÁK 2003), při níž jsou vzorky postupně promývány v mírně tekoucí
vodě pomocí polokulovitého síta s velikostí ok 0,5 mm. Tím dochází k odplavení jemných
anorganických částic a usazení měkkýších ulit (jak živých jedinců, tak i prázdných, vodou
naplněných schránek) na dno síta. Rostlinný materiál plovoucí na hladině je průběžně
ručně odstraňován, což vede k výrazné redukci konečného objemu vzorku a usnadnění
jeho laboratorního zpracování.
Po usušení byly ze vzorků pod binokulární lupou ručně vybrány schránky měkkýšů a
určeny do druhů s pomocí příslušné literatury (LOŽEK 1956, KERNEY et al. 1983). Pro každý

23
druh byl zaznamenán počet jedinců v době vzorkování živých a počet prázdných
schránek. Ze vzorků byli vytříděni živí jedinci suchozemských ulitnatých plžů, usušeni po
dobu 24 hodin při 60 °C a po 30 minutách chladnutí v exsikátoru zvá ženi na laboratorních
vahách s přesností 0,00001 g. Tak byla stanovena celková hmotnost suché biomasy pro
každý vzorek. Pro vzorky z polských slatinišť byly jednotlivé druhy rovněž zváženy zvlášť,
za účelem vyhodnocení korelací mezi biomasou jednotlivých druhů a jejich abundancí na
lokalitách. Pro živé suchozemské ulitnaté plže byly dále zkoumány rozdíly v mediánové
velikosti jedinců na jednotlivých lokalitách. Velikost těla plže byla definována jako objem
schránky, spočítaný podle vzorce uvedeného v práci MCCLAIN & NEKOLA (2008) jako objem
kužele spočívajícího na vrcholu válce:
+−=
3))1((
22 thr
htrVππ
kde V je objem schránky, r je polovina šířky schránky, h je výška schránky a t je podíl
z celkové výšky tvořený kónickou částí schránky, určený na základě vizuálního
vyhodnocení fotografií měkkýších ulit. Rozměry schránek byly vypočítány jako průměry
z hodnot uvedených v publikaci KERNEY et al. (1983).
5.3. Vysvětlující prom ěnné
V terénu byly s využitím přenosných přístrojů PH 119 a CM 101 (Snail Instruments,
Beroun, Česká republika) změřeny chemické parametry indikující minerální bohatost
slatiniště, tedy pH a konduktivita vody. Pomocí GPS přijímače byly zaznamenány
geografické souřadnice a nadmořská výška lokalit. K jednotlivým lokalitám byly zjištěny
klimatické parametry na základě souřadnic a informací z databáze WorldClim 1.4 (HIJMANS
et al. 2005), zpracovaných v programu ArcGIS 8.3 (ESRI 2003), konkrétně průměrné
červencové, lednové a roční teploty a celkové letní, zimní a roční srážkové úhrny. Hodnoty
všech proměnných prostředí charakterizujících vzorkované lokality jsou uvedeny v Příloze
1 (str. 68).
5.4. Statistické vyhodnocení dat
Pro výpočty korelačních koeficientů a grafické znázornění korelací byl využit program
Statistica 10 (HILL & LEWICKI 2007). Korelace byly spočítány pomocí neparametrického
Spearmanova korelačního koeficientu a hladina významnosti byla ve všech případech

24
mnohonásobeného testování upravena Bonferroniho korekcí (HOLM 1979). Studovány byly
korelace druhových bohatostí a abundancí měkkýšů se zeměpisnou šířkou a rovněž
procentuální zastoupení druhů a jedinců čeledi Vertiginidae ve společenstvech
suchozemských ulitnatých plžů podél gradientu zeměpisné šířky. Druhové zastoupení bylo
počítáno jako procento z celkového počtu všech druhů suchozemských plžů včetně těch, u
kterých byly nalezeny jen prázdné schránky, zastoupení jedinců bylo počítáno jako
procento z počtu živých jedinců suchozemských plžů. Prázdné schránky pomáhají zachytit
kompletní druhové spektrum lokality, neboť se mezi nimi mohou nacházet i některé velmi
vzácné druhy (CAMERON & POKRYSZKO 2005), zatímco počty živých jedinců lépe odrážejí
reálné abundance měkkýšů na slatiništích. Rychlost rozkladu vápnitých měkkýších ulit se
totiž výrazně liší v závislosti na minerální bohatosti lokalit, přičemž především na velmi
vápnitých slatiništích dochází k dlouhodobé akumulaci prázdných schránek
(CERNOHORSKY et al. 2010).
Pro ordinační analýzy byl použit programový balík CANOCO 4.5 (TER BRAAK &
ŠMILAUER 2002). Do analýz byli zahrnuti pouze suchozemští ulitnatí plži, neboť vodní
měkkýši mají obecně rozdílné ekologické nároky a jejich výskyt může být na slatiništích
ovlivněn jinými faktory. Z ordinačních analýz byli vyloučeni nazí plži (Arion fuscus a
Deroceras laeve) a dále druhy, pro které nebyli nalezeni živí jedinci na žádné ze
studovaných lokalit (Discus ruderatus, Isognomostoma isognomostomos, Monachoides
vicinus, Semilimax semilimax, Urticicola umbrosus a Vitrea transsylvanica), neboť se
patrně jedná o náhodné nálezy druhů obývajících jiné typy biotopů v okolí slatiniště. Pro
účely ordinačních analýz byly sloučeny druhy Pupilla alpicola a Pupilla pratensis, které se
vyznačují podobnými ekologickými požadavky, a podle předběžných výsledků
molekulárních analýz se nejspíše nejedná o dobře odlišitelné druhy, ale pouze o
geografické poddruhy (viz HORSÁK et al. 2012).
Data byla před vstupem do ordinačních analýz upravena logaritmickou transformací
(log (x+1)) a bylo zvoleno podvážení vzácných druhů. Pro zjištění délky hlavního gradientu
ve studovaných datech byla využita nepřímá ordinace DCA (detrendovaná
korespondenční analýza). Délka hlavního gradientu přesahovala 3 SDU (jednotky
směrodatné odchylky), a data byla proto dále zpracovávána pomocí ordinačních technik
předpokládajících unimodální odpověď dat na gradienty prostředí. Prostřednictvím analýzy
DCA byly určeny hlavní směry variability v datech o druhové skladbě měkkýšů. Pomocí
Spearmanova korelačního koeficientu byly následně spočítány korelace proměnných
prostředí, počtů druhů a abundancí se skóre vzorků na 1.–4. ose DCA.

25
Datový soubor studovaných lokalit byl na základě znalostí geomorfologie, klimatu a
charakteru bioty jednotlivých částí zkoumaného území rozdělen do 3 hlavních
biogeografických oblastí: (1) slatiniště Západních Karpat a jejich podhůří na jihu Polska,
(2) slatiniště v nížinné oblasti východního a severovýchodního Polska a nejjižnějšího
Švédska, náležející do zóny mírného pásu, a (3) slatiniště jižního Švédska a Norska,
náležející do boreální zóny (zařazení lokalit k uvedeným oblastem viz Příloha 1, str. 68).
Pro ověření rozdílů mezi biogeografickými oblastmi, vyjádřenými pomocí binárních
proměnných, byla využita přímá ordinace CCA (kanonická korespondenční analýza).
Statistická významnost proměnných byla otestována Monte Carlo permutačním testem
(999 permutací, p < 0,05). Pro skupinu temperátních lokalit, náležejících do oblastí (1) a
(2), a skupinu boreálních lokalit, náležejících do oblasti (3), byly vybrány nejhojnější druhy
měkkýšů, tj. druhy vyskytující se více než na 1/3 z celkového počtu lokalit v daných
skupinách. Statistická významnost rozdílů ve frekvenci výskytu těchto druhů
v temperátním a boreálním regionu byla spočítána pomocí Fisherova exaktního testu.
Závislost počtu druhů na biomase společenstev živých suchozemských ulitnatých plžů
byla modelována v softwaru R 2.14.2 (R DEVELOPMENT CORE TEAM 2012). Studovaná data
splňovala předpoklady lineárního regresního modelu (viz např. PEKÁR & BRABEC 2009), a
k jejich analýze byla proto využita lineární regrese. Vzhledem k vizuálně dobře patrnému
zakřivení průběhu hodnot počtu druhů byly zvoleny modely lineární regrese logaritmicky
transformované biomasy a lineární regrese s kvadratickým členem. Statistická významnost
parametrů modelu byla testována pomocí dílčích t-testů a vhodnost použití jednotlivých
regresních modelů byla ověřena analýzou rozptylu (ANOVA, F-test). Kvalita regresních
modelů byla vyhodnocena pomocí adjustovaného koeficientu determinance (Adj. R2), který
zohledňuje náhodný nárůst procenta vysvětlené variability při obohacení modelu o
vysvětlující proměnné. Pro srovnání logaritmického a kvadratického modelu pro data
z Polska, jižní Skandinávie a z obou oblastí dohromady bylo využito Akaikeho Informační
Kritérium (AIC). Jako statisticky významně vhodnější byly vybrány modely s hodnotou AIC
nižší nejméně o 10 jednotek (BURNHAM & ANDERSON 2004). Skript popisující průběh analýz
je uveden v Příloze 2 (str. 70).

26
6. VÝSLEDKY
Celkem bylo studováno 62 bazických slatinišť, 39 v oblasti Polska a 23 v oblasti jižní
Skandinávie. Na 39 polských slatiništích bylo nalezeno 64 druhů měkkýšů, z nichž 44
druhů náleželo mezi suchozemské plže, 15 mezi vodní plže a 5 mezi vodní mlže (Příloha
3, str. 73). Z celkového počtu 13 928 nalezených jedinců bylo 6 828 jedinců 53 druhů v
době vzorkování živých, zbytek tvořily prázdné schránky. Na 23 slatiništích jižní
Skandinávie bylo nalezeno 40 druhů měkkýšů, z nichž 29 druhů náleželo mezi
suchozemské plže, 8 mezi vodní plže a 3 mezi vodní mlže (Příloha 3, str. 73). Z celkového
počtu 3 816 jedinců bylo 1 861 jedinců 34 druhů v době vzorkování živých. Druhovou
bohatost Skandinávie značně navyšovala 3 slatiniště na jižním cípu Švédska (S1–3, Obr.
2) s diverzitou a druhovou skladbou takřka středoevropského charakteru. V rámci vlastní
boreální zóny Skandinávie (lokality S4–23, Obr. 2) bylo nalezeno pouze 30 druhů měkkýšů
(22 suchozemských plžů, 6 vodních plžů, 2 vodní mlži). Počet všech druhů měkkýšů na
lokalitu se v Polsku pohyboval od 5 do 27 s mediánovou hodnotou 15 druhů, ve
Skandinávii (včetně 3 lokalit na jižním cípu Švédska) od 3 do 17 druhů s mediánovou
hodnotou 8 druhů na lokalitu (Příloha 4, str. 75). Celý datový soubor zahrnoval 72 druhů
měkkýšů (50 suchozemských plžů, 17 vodních plžů a 5 vodních mlžů) zastoupených
17 744 jedinci (Příloha 3, str. 73). Druhově nejbohatší lokalitou bylo velmi malé silně
pěnovcové prameniště v blízkosti obce Rowele na severovýchodě Polska u hranic s Litvou
(P37, Obr. 1), na kterém bylo nalezeno 27 druhů měkkýšů zastoupených 1 116 jedinci.
6.1. Společenstva m ěkkýš ů Polska a jižní Skandinávie
6.1.1. Změny druhové bohatosti a velikosti mediánového jedinc e
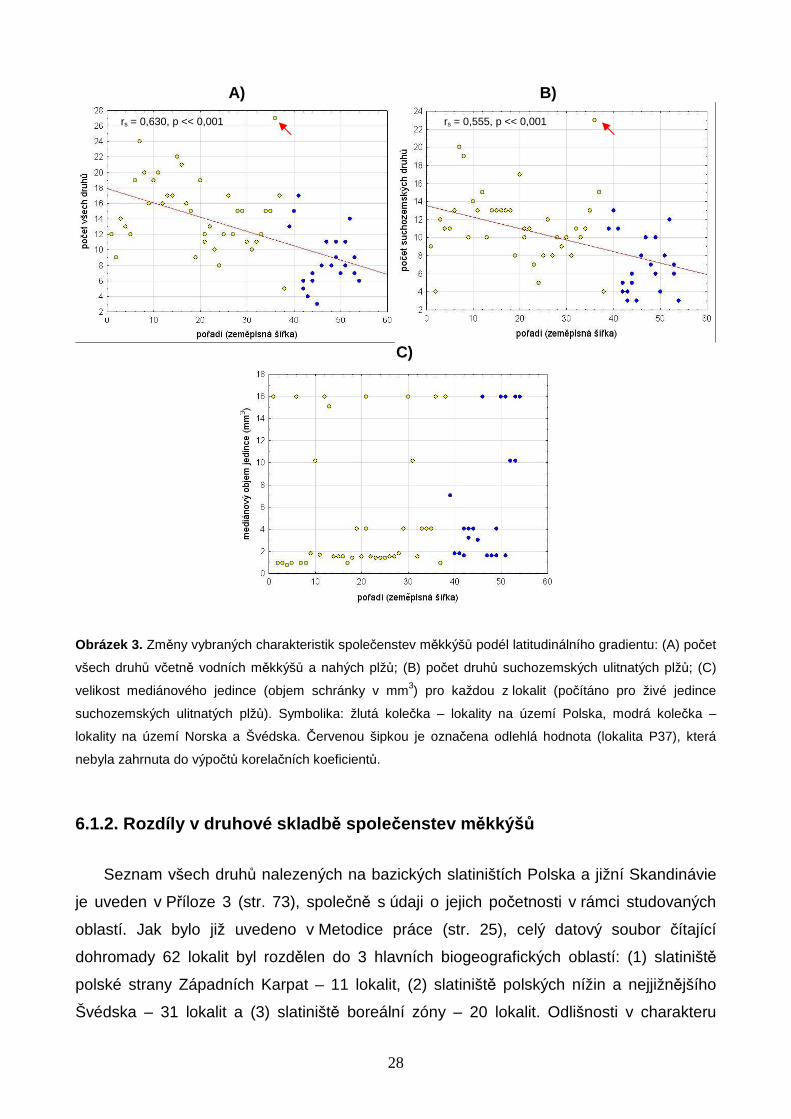
Tabulka 1 obsahuje výsledky korelací druhové bohatosti a abundancí měkkýšů se
zeměpisnou šířkou; vybrané korelace jsou znázorněny graficky (Obr. 3A, B). Nejbohatší
polská lokalita (P37) se vyznačovala extrémně vysokým počtem druhů, a v rámci souboru
lokalit byla proto považována za odlehlou hodnotu, která nebyla zahrnuta do výpočtů
korelačních koeficientů. Výsledky prokázaly signifikantně negativní vztah počtu druhů
měkkýšů a zeměpisné šířky (Tabulka 1, Obr. 3A, B). Hodnoty korelačních koeficientů byly
vyšší pro počty všech druhů včetně vodních měkkýšů než pouze pro počty druhů
suchozemských ulitnatých plžů (Tabulka 1, Obr. 3A, B). Pozorovaný rozdíl je ovlivněn

27
výskytem velkého počtu vodních druhů měkkýšů na polských slatiništích – skandinávská
slatiniště byla z hlediska vodní malakofauny daleko méně pestrá (Příloha 3, str. 73).
Obdobně také abundance měkkýšů klesaly směrem k vyšším zeměpisným šířkám, a tento
trend byl výraznější pro všechny druhy měkkýšů (Tabulka 1).
Tabulka 1. Spearmanovy korelace počtů druhů a jedinců se zeměpisnou šířkou. Hladina významnosti: * <
0,05; ** (tučně) < 0,0055 (upraveno Bonferroniho korekcí).
Spearmanovy korelace
počet druh ů zem. šířka po čet jedinc ů zem. šířka
všichni měkkýši -0,630** všichni měkkýši -0,540** živí jedinci i prázdné schránky suchozemští ulitnatí plži -0,555** suchozemští ulitnatí plži -0,458**
všichni měkkýši -0,607** všichni měkkýši -0,528** pouze živí jedinci
suchozemští ulitnatí plži -0,452** suchozemští ulitnatí plži -0,276*
Z grafů na Obr. 3A, B je patrné, že tři slatiniště v nejjižnější části Švédska se počtem
druhů blížila spíše polským lokalitám, a výrazný skokový pokles druhové bohatosti se
odehrál až mezi nimi a lokalitami situovanými dále na sever v boreální zóně Norska a
Švédska. Tři lokality na jihu Švédska, spadající ještě do klimaticky příznivé zóny mírného
pásu, se vyznačovaly velmi vysokými hodnotami konduktivity (Příloha 1, str. 69) a náležely
mezi silně bazická slatiniště se srážením pěnovce. Ve srovnání s ostatními lokalitami na
území Norska a Švédska se jednalo o druhově výjimečně bohatá slatiniště. Přesto však
ani tyto, na skandinávské poměry v mnoha ohledech unikátní lokality, nedosahovaly
druhové rozmanitosti nejbohatších polských slatinišť (Obr. 3A, B).
Studovány byly také rozdíly v mediánové velikosti živých jedinců suchozemských
ulitnatých plžů podél latitudinálního gradientu, přičemž "velikost" těla byla definována jako
objem schránky (výpočet viz Metodika práce, str. 23). Z Obr. 3C je patrné, že objem
mediánového jedince se na většině lokalit pohyboval mezi 0,7 až 4,1 mm3, což odpovídá
např. objemům schránek plžů r. Vertigo, Carychium, Columella a druhu Puncum
pygmaeum, a výjimečně dosahoval ke zhruba 16 mm3, což je objem schránky druhu
Perpolita hammonis (Příloha 5, str. 77). Z uvedených výsledků vyplývá, že na všech
studovaných lokalitách dominovaly druhy nepřesahující v žádném z rozměrů schránky
velikost 4 mm, přičemž přibližně na polovině z těchto lokalit se jednalo o druhy velké
kolem pouhých 2 mm (viz Obr. 3C a Příloha 5, str. 77). Výsledky však neprokázaly žádný
trend v mediánovém objemu jedinců podél zeměpisné šířky (Obr. 3C).
28
Obrázek 3. Změny vybraných charakteristik společenstev měkkýšů podél latitudinálního gradientu: (A) počet
všech druhů včetně vodních měkkýšů a nahých plžů; (B) počet druhů suchozemských ulitnatých plžů; (C)
velikost mediánového jedince (objem schránky v mm3) pro každou z lokalit (počítáno pro živé jedince
suchozemských ulitnatých plžů). Symbolika: žlutá kolečka – lokality na území Polska, modrá kolečka –
lokality na území Norska a Švédska. Červenou šipkou je označena odlehlá hodnota (lokalita P37), která
nebyla zahrnuta do výpočtů korelačních koeficientů.
6.1.2. Rozdíly v druhové skladb ě spole čenstev m ěkkýš ů
Seznam všech druhů nalezených na bazických slatiništích Polska a jižní Skandinávie
je uveden v Příloze 3 (str. 73), společně s údaji o jejich početnosti v rámci studovaných
oblastí. Jak bylo již uvedeno v Metodice práce (str. 25), celý datový soubor čítající
dohromady 62 lokalit byl rozdělen do 3 hlavních biogeografických oblastí: (1) slatiniště
polské strany Západních Karpat – 11 lokalit, (2) slatiniště polských nížin a nejjižnějšího
Švédska – 31 lokalit a (3) slatiniště boreální zóny – 20 lokalit. Odlišnosti v charakteru
A) B)
C)
rs = 0,630, p << 0,001 rs = 0,555, p << 0,001

29
společenstev měkkýšů mezi jednotlivými lokalitami navzájem a mezi vymezenými
biogeografickými oblastmi byly následně testovány pomocí ordinačních analýz.
Pro zjištění hlavních směrů variability druhového složení malakocenóz byla využita
nepřímá ordinace DCA (analyzováni byli pouze suchozemští ulitnatí plži). Obrázek 4
zobrazuje pozice lokalit v diagramu podél 1. a 2. osy DCA, barevnými symboly je odlišena
příslušnost lokalit ke třem výše definovaným biogeografickým oblastem. Nejjižněji
položené švédské lokality (S1–3) se dle očekávání začlenily mezi polská slatiniště, a došlo
tak k jednoznačnému oddělení slatinišť boreální zóny Norska a Švédska od slatinišť
mírného pásu v oblastech Polska a nejjižnějšího Švédska (Obr. 4). Vymezení lokalit v
oblasti Západních Karpat oproti ostatním lokalitám mírného pásu bylo již méně
jednoznačné. Malakofauna většiny karpatských slatinišť byla sice patrně druhově
specifická, o čemž vypovídá vytvoření shluku karpatských lokalit v levé horní části
diagramu, ovšem některá karpatská slatiniště (P2, P9–11) se zcela začlenila mezi lokality
temperátní nížinné oblasti (Obr. 4). Z výsledků analýzy tak vyplývá, že malakocenózy
boreálních skandinávských slatinišť, tvořící samostatný shluk na pravé straně diagramu
DCA, se výrazně odlišovaly od slatinišť mírného pásu v oblastech Karpat, polských nížin a
nejjižnějšího Švédska, která si naopak byla vzájemně podobnější a nedošlo v rámci nich k
vytvoření jasně vymezitelných shluků.
Pro stanovení faktorů odpovídajících za hlavní rozdíly v druhovém složení
společenstev měkkýšů byly provedeny korelace proměnných prostředí se skóre vzorků na
1.–4. ose DCA. Jak ukázaly hodnoty Spearmanových korelačních koeficientů, za hlavní
gradient v datech o skladbě malakocenóz odpovídaly především rozdíly v klimatických
parametrech zkoumaných oblastí (Tabulka 2). Nejsilnější byly negativní korelace 1. osy
DCA s průměrnými teplotami a zeměpisnou šířkou, ovšem signifikantní byly rovněž
korelace s dalšími faktory: nadmořskou výškou, zimními srážkami a konduktivitou (Tabulka
2). První osa tak představovala komplexní ekologický gradient, podél něhož se měnilo
několik, často vzájemně provázaných parametrů prostředí. Vysoké hodnoty korelačních
koeficientů pro průměrné teploty a zeměpisnou šířku však naznačují, že složení
společenstev měkkýšů bylo ovlivněno především klimatickými poměry, danými
geografickou polohou studovaných lokalit. Kromě druhové skladby se podél 1. osy
signifikantně měnila i diverzita společenstev měkkýšů – jak počty druhů, tak abundance
klesaly směrem k chladnějším oblastem ve vyšších zeměpisných šířkách (Tabulka 2).
Stanovení ekologických parametrů odpovídajících za změny společenstev podél 2., 3.
a 4. osy DCA již bylo obtížnější. Ekologický gradient vyjádřený 2. osou DCA měl vliv
především na druhovou skladbu lokalit na území Polska (Obr. 4) a signifikantně koreloval
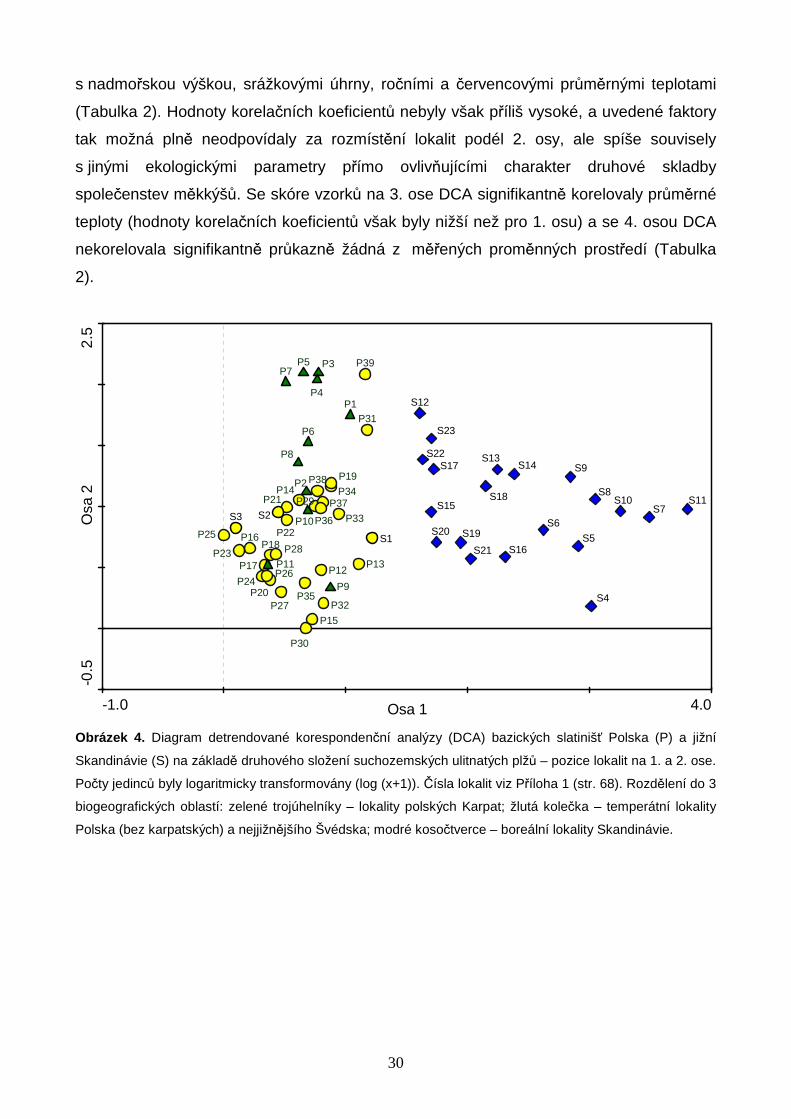
30
s nadmořskou výškou, srážkovými úhrny, ročními a červencovými průměrnými teplotami
(Tabulka 2). Hodnoty korelačních koeficientů nebyly však příliš vysoké, a uvedené faktory
tak možná plně neodpovídaly za rozmístění lokalit podél 2. osy, ale spíše souvisely
s jinými ekologickými parametry přímo ovlivňujícími charakter druhové skladby
společenstev měkkýšů. Se skóre vzorků na 3. ose DCA signifikantně korelovaly průměrné
teploty (hodnoty korelačních koeficientů však byly nižší než pro 1. osu) a se 4. osou DCA
nekorelovala signifikantně průkazně žádná z měřených proměnných prostředí (Tabulka
2).
-1.0 4.0
-0.5
2.5
P1
P38
P16P18
P17
P2
P20P24
P25
P23
P21
P22
P28
P27
P6
P30
P29
P26
P33
P34
P35
P10
P31
P32
P37
P39
P36
P9
P11
P3
P4
P5
P8
P7
P19
P12P13
P14
P15
S2
S18
S19
S15
S16
S23
S17 S14S13
S12
S1
S8
S9
S10
S6
S4
S5
S11
S3
S21
S22
S20
S7
Osa 1
Osa
2
Obrázek 4. Diagram detrendované korespondenční analýzy (DCA) bazických slatinišť Polska (P) a jižní
Skandinávie (S) na základě druhového složení suchozemských ulitnatých plžů – pozice lokalit na 1. a 2. ose.
Počty jedinců byly logaritmicky transformovány (log (x+1)). Čísla lokalit viz Příloha 1 (str. 68). Rozdělení do 3
biogeografických oblastí: zelené trojúhelníky – lokality polských Karpat; žlutá kolečka – temperátní lokality
Polska (bez karpatských) a nejjižnějšího Švédska; modré kosočtverce – boreální lokality Skandinávie.
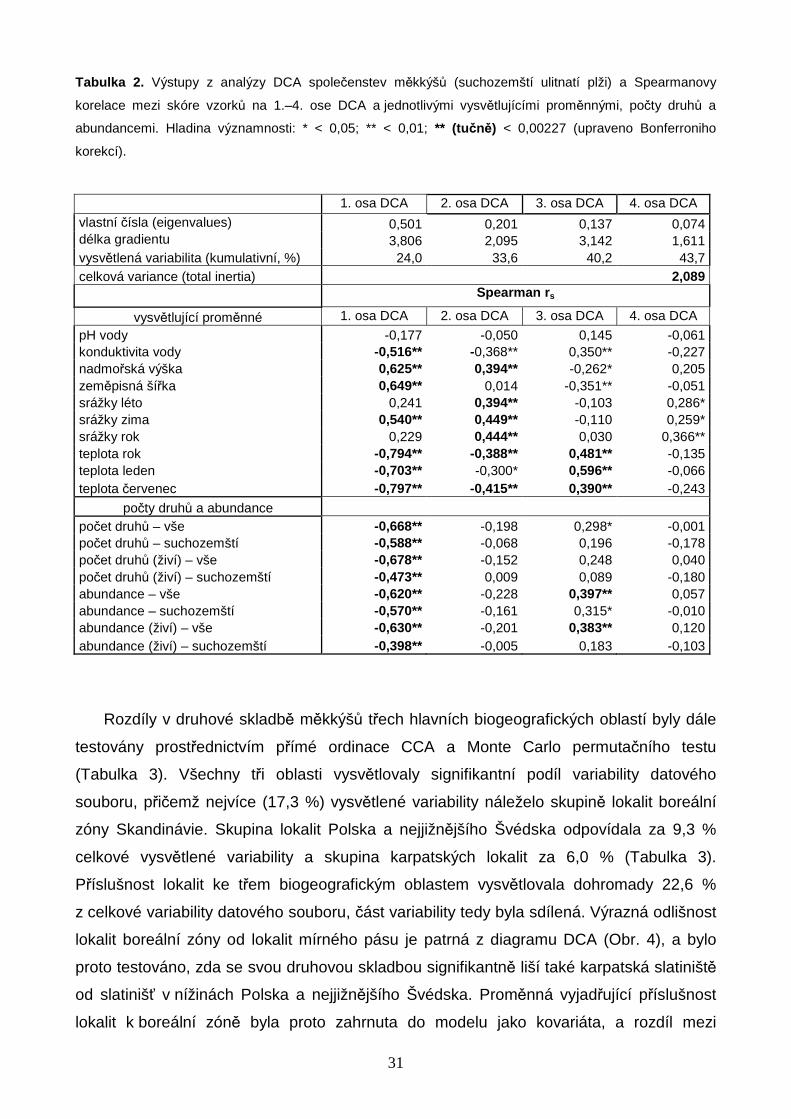
31
Tabulka 2. Výstupy z analýzy DCA společenstev měkkýšů (suchozemští ulitnatí plži) a Spearmanovy
korelace mezi skóre vzorků na 1.–4. ose DCA a jednotlivými vysvětlujícími proměnnými, počty druhů a
abundancemi. Hladina významnosti: * < 0,05; ** < 0,01; ** (tučně) < 0,00227 (upraveno Bonferroniho
korekcí).
1. osa DCA 2. osa DCA 3. osa DCA 4. osa DCA
vlastní čísla (eigenvalues) 0,501 0,201 0,137 0,074 délka gradientu 3,806 2,095 3,142 1,611 vysvětlená variabilita (kumulativní, %) 24,0 33,6 40,2 43,7 celková variance (total inertia) 2,089
Spearman r s
vysvětlující proměnné 1. osa DCA 2. osa DCA 3. osa DCA 4. osa DCA
pH vody -0,177 -0,050 0,145 -0,061 konduktivita vody -0,516** -0,368** 0,350** -0,227 nadmořská výška 0,625** 0,394** -0,262* 0,205 zeměpisná šířka 0,649** 0,014 -0,351** -0,051 srážky léto 0,241 0,394** -0,103 0,286* srážky zima 0,540** 0,449** -0,110 0,259* srážky rok 0,229 0,444** 0,030 0,366** teplota rok -0,794** -0,388** 0,481** -0,135 teplota leden -0,703** -0,300* 0,596** -0,066 teplota červenec -0,797** -0,415** 0,390** -0,243
počty druhů a abundance
počet druhů – vše -0,668** -0,198 0,298* -0,001 počet druhů – suchozemští -0,588** -0,068 0,196 -0,178 počet druhů (živí) – vše -0,678** -0,152 0,248 0,040 počet druhů (živí) – suchozemští -0,473** 0,009 0,089 -0,180 abundance – vše -0,620** -0,228 0,397** 0,057 abundance – suchozemští -0,570** -0,161 0,315* -0,010 abundance (živí) – vše -0,630** -0,201 0,383** 0,120 abundance (živí) – suchozemští -0,398** -0,005 0,183 -0,103
Rozdíly v druhové skladbě měkkýšů třech hlavních biogeografických oblastí byly dále
testovány prostřednictvím přímé ordinace CCA a Monte Carlo permutačního testu
(Tabulka 3). Všechny tři oblasti vysvětlovaly signifikantní podíl variability datového
souboru, přičemž nejvíce (17,3 %) vysvětlené variability náleželo skupině lokalit boreální
zóny Skandinávie. Skupina lokalit Polska a nejjižnějšího Švédska odpovídala za 9,3 %
celkové vysvětlené variability a skupina karpatských lokalit za 6,0 % (Tabulka 3).
Příslušnost lokalit ke třem biogeografickým oblastem vysvětlovala dohromady 22,6 %
z celkové variability datového souboru, část variability tedy byla sdílená. Výrazná odlišnost
lokalit boreální zóny od lokalit mírného pásu je patrná z diagramu DCA (Obr. 4), a bylo
proto testováno, zda se svou druhovou skladbou signifikantně liší také karpatská slatiniště
od slatinišť v nížinách Polska a nejjižnějšího Švédska. Proměnná vyjadřující příslušnost
lokalit k boreální zóně byla proto zahrnuta do modelu jako kovariáta, a rozdíl mezi

32
zbývajícími dvěma oblastmi následně činil 5,3 % (p = 0,001) z celkové variability datového
souboru. Výsledky analýzy CCA, v souladu s výsledky nepřímé ordinace DCA (Obr. 4),
dokumentovaly specifický ráz druhové skladby lokalit v oblasti boreální zóny Skandinávie,
a potvrdily také méně zřetelnou, avšak signifikantní odlišnost druhové skladby obou oblastí
mírného pásu.
Tabulka 3. Procenta vysvětlené variability na základě analýzy CCA s použitím Monte Carlo permutačního
testu. Oblast 1 – lokality polských Karpat, Oblast 2 – lokality polských nížin a nejjižnějšího Švédska, Oblast 3
– boreální lokality Skandinávie. Signifikantní faktory (p < 0,05) jsou označeny tučně a kurzívou.
CCA
Monte Carlo permuta ční test
vysvětlená variabilita
signifikance (p)
Oblast 3 17,3 % 0,001 Oblast 2 9,3 % 0,001 Oblast 1 6,0 % 0,001 variabilita vysvětlená Oblastmi 1–3 22,6 %
Které konkrétní druhy tedy odpovídaly za pozorované rozdíly ve skladbě společenstev
měkkýšů oblastí Polska a Skandinávie? Obrázek 5A zobrazuje rozmístění druhů
suchozemských ulitnatých plžů v diagramu DCA podél 1. a 2. osy. V levé horní části
diagramu se vyčlenily druhy Macrogastra tumida, Aegopinella epipedostoma, Platyla polita
a Trochulus hispidus (zelené trojúhelníky), nalezené pouze na slatiništích Karpat. Jejich
výskyt však byl i zde spíše vzácný a ojedinělý (Příloha 3, str. 73), což indikuje, že se
nejednalo o typické slatiništní specialisty, ale spíše o druhy pronikající na drobná svahová
karpatská slatiniště z okolních lesních a křovinných porostů. V pravé části diagramu DCA
došlo k oddělení chladnomilných severských druhů Vertigo genesii, V. lilljeborgi, V.
modesta a Columella columella (modré kosočtverce), jejichž výskyt se omezoval výhradně
na slatiniště boreální zóny Skandinávie (Příloha 3, str. 73). Ostatní druhy měkkýšů byly
pomocí symbolů rozlišeny na druhy vyskytující se od Polska po nejjižnější Švédsko, avšak
nezasahující na slatiniště boreální zóny Skandinávie (žlutá kolečka) a druhy vyskytující se
na slatiništích všech částí studovaného území (červené čtverce). Tyto dvě skupiny
netvořily výrazně oddělené shluky, avšak umístění temperátních druhů blíže k levé
polovině diagramu a druhů zasahujících až do boreální zóny blíže k pravé polovině
diagramu je dobře patrné (Obr. 5A). Mezi teplomilnější prvky středoevropské fauny,
nepřesahující hranici boreální zóny, patřily např. Vertigo angustior, V. moulinsiana a
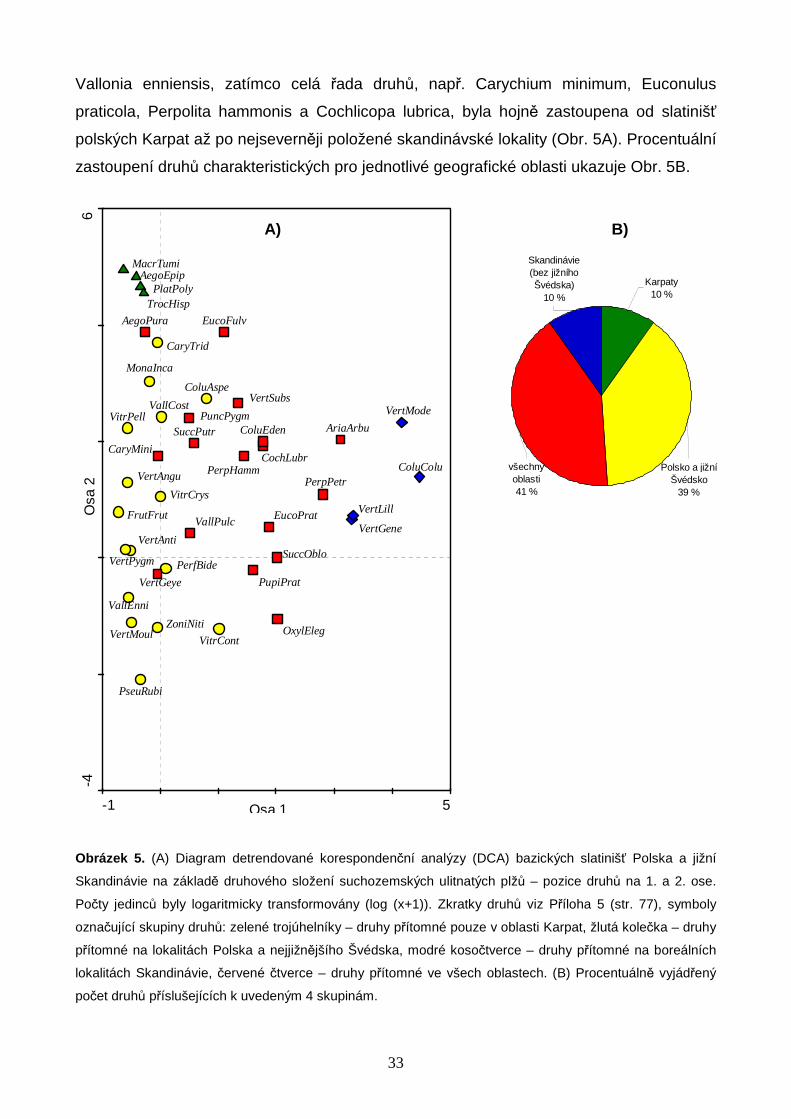
33
Vallonia enniensis, zatímco celá řada druhů, např. Carychium minimum, Euconulus
praticola, Perpolita hammonis a Cochlicopa lubrica, byla hojně zastoupena od slatinišť
polských Karpat až po nejseverněji položené skandinávské lokality (Obr. 5A). Procentuální
zastoupení druhů charakteristických pro jednotlivé geografické oblasti ukazuje Obr. 5B.
-1 5
-46
AegoEpip
AegoPura
AriaArbu
CaryMini
CaryTrid
CochLubr
ColuAspe
ColuColu
ColuEden
EucoFulv
EucoPratFrutFrut
MacrTumi
MonaInca
OxylEleg
PerfBide
PerpHammPerpPetr
PlatPoly
PseuRubi
PuncPygm
PupiPrat
SuccPutr
SuccOblo
TrocHisp
VallCost
VallEnni
VallPulc
VertAngu
VertAnti
VertMode
VertGene
VertGeye
VertLill
VertMoul
VertPygm
VertSubs
VitrCont
VitrCrys
VitrPell
ZoniNiti
Osa 1
Osa
2
Obrázek 5. (A) Diagram detrendované korespondenční analýzy (DCA) bazických slatinišť Polska a jižní
Skandinávie na základě druhového složení suchozemských ulitnatých plžů – pozice druhů na 1. a 2. ose.
Počty jedinců byly logaritmicky transformovány (log (x+1)). Zkratky druhů viz Příloha 5 (str. 77), symboly
označující skupiny druhů: zelené trojúhelníky – druhy přítomné pouze v oblasti Karpat, žlutá kolečka – druhy
přítomné na lokalitách Polska a nejjižnějšího Švédska, modré kosočtverce – druhy přítomné na boreálních
lokalitách Skandinávie, červené čtverce – druhy přítomné ve všech oblastech. (B) Procentuálně vyjádřený
počet druhů příslušejících k uvedeným 4 skupinám.
A) B)
Karpaty10 %
Polsko a jižní Švédsko
39 %
všechny oblasti41 %
Skandinávie (bez jižního Švédska)
10 %

34
6.1.3. Specifika druhové skladby m ěkkýš ů ve studovaných oblastech
Výsledky prezentované v předchozí kapitole ukazují, že z hlediska druhové skladby
společenstev měkkýšů se výrazně liší slatiniště boreální zóny Skandinávie od slatinišť
mírného pásu v oblastech polských Karpat, nížinné části Polska a nejjižnější části
Švédska (Obr. 4, Tabulka 3). Byla proto studována otázka, zda se temperátní a boreální
region kromě přítomnosti specifických druhů měkkýšů liší také frekvencí výskytu
jednotlivých druhů. V rámci studovaných oblastí byly vybrány nejhojnější druhy měkkýšů
(viz Metodika práce, str. 25) a následně byla srovnána frekvence výskytu těchto druhů
v obou oblastech (Tabulka 4).
Z celkem 22 nejhojnějších druhů měkkýšů patřilo 15 mezi druhy vyskytující se v obou
studovaných oblastech. Ze zbývajících 7 druhů bylo 5 nalezeno pouze na temperátních
slatiništích a 2 pouze na boreálních slatiništích (Tabulka 4). Obdobně hojnými v obou
oblastech byly druhy Euconulus praticola, Cochlicopa lubrica, Oxyloma elegans, Vertigo
substriata a Succinella oblonga (frekvence jejich výskytu v obou oblastech nebyla na
základě výsledků Fisherova testu signifikantně odlišná). V rámci datového souboru se tedy
jednalo o druhy nejrovnoměrněji pokrývající celé studované území. V ostatních případech
se frekvence výskytu druhů signifikantně odlišovala. Naprostá většina druhů byla relativně
hojnější v oblasti polských a jihošvédských slatinišť (Tabulka 4), pouze druhy Perpolita
petronella, Galba truncatula, Columella columella a Vertigo genesii byly četnější na
slatiništích Norska a Švédska, přičemž poslední dva jmenované se vůbec nevyskytovaly
mimo boreální zónu (Tabulka 4). Naopak druhy Vertigo pygmaea, V. antivertigo, V.
angustior, Pisidium personatum a Vallonia enniensis obývaly velmi hojně slatiniště
temperátní oblasti a dále na severu v boreální zóně nebyly vůbec nalezeny (Tabulka 4).
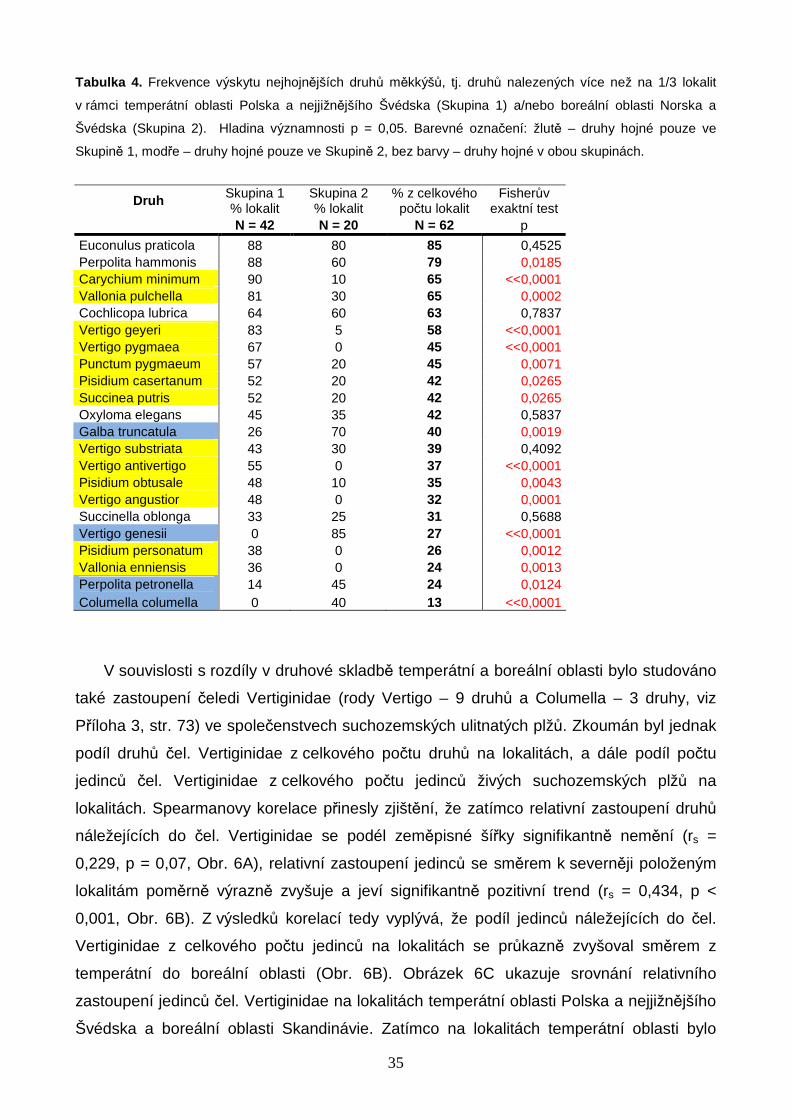
35
Tabulka 4. Frekvence výskytu nejhojnějších druhů měkkýšů, tj. druhů nalezených více než na 1/3 lokalit
v rámci temperátní oblasti Polska a nejjižnějšího Švédska (Skupina 1) a/nebo boreální oblasti Norska a
Švédska (Skupina 2). Hladina významnosti p = 0,05. Barevné označení: žlutě – druhy hojné pouze ve
Skupině 1, modře – druhy hojné pouze ve Skupině 2, bez barvy – druhy hojné v obou skupinách.
Druh Skupina 1 % lokalit
Skupina 2 % lokalit
% z celkového počtu lokalit
Fisherův exaktní test
N = 42 N = 20 N = 62 p
Euconulus praticola 88 80 85 0,4525 Perpolita hammonis 88 60 79 0,0185 Carychium minimum 90 10 65 <<0,0001 Vallonia pulchella 81 30 65 0,0002 Cochlicopa lubrica 64 60 63 0,7837 Vertigo geyeri 83 5 58 <<0,0001 Vertigo pygmaea 67 0 45 <<0,0001 Punctum pygmaeum 57 20 45 0,0071 Pisidium casertanum 52 20 42 0,0265 Succinea putris 52 20 42 0,0265 Oxyloma elegans 45 35 42 0,5837 Galba truncatula 26 70 40 0,0019 Vertigo substriata 43 30 39 0,4092 Vertigo antivertigo 55 0 37 <<0,0001 Pisidium obtusale 48 10 35 0,0043 Vertigo angustior 48 0 32 0,0001 Succinella oblonga 33 25 31 0,5688 Vertigo genesii 0 85 27 <<0,0001 Pisidium personatum 38 0 26 0,0012 Vallonia enniensis 36 0 24 0,0013 Perpolita petronella 14 45 24 0,0124 Columella columella 0 40 13 <<0,0001
V souvislosti s rozdíly v druhové skladbě temperátní a boreální oblasti bylo studováno
také zastoupení čeledi Vertiginidae (rody Vertigo – 9 druhů a Columella – 3 druhy, viz
Příloha 3, str. 73) ve společenstvech suchozemských ulitnatých plžů. Zkoumán byl jednak
podíl druhů čel. Vertiginidae z celkového počtu druhů na lokalitách, a dále podíl počtu
jedinců čel. Vertiginidae z celkového počtu jedinců živých suchozemských plžů na
lokalitách. Spearmanovy korelace přinesly zjištění, že zatímco relativní zastoupení druhů
náležejících do čel. Vertiginidae se podél zeměpisné šířky signifikantně nemění (rs =
0,229, p = 0,07, Obr. 6A), relativní zastoupení jedinců se směrem k severněji položeným
lokalitám poměrně výrazně zvyšuje a jeví signifikantně pozitivní trend (rs = 0,434, p <
0,001, Obr. 6B). Z výsledků korelací tedy vyplývá, že podíl jedinců náležejících do čel.
Vertiginidae z celkového počtu jedinců na lokalitách se průkazně zvyšoval směrem z
temperátní do boreální oblasti (Obr. 6B). Obrázek 6C ukazuje srovnání relativního
zastoupení jedinců čel. Vertiginidae na lokalitách temperátní oblasti Polska a nejjižnějšího
Švédska a boreální oblasti Skandinávie. Zatímco na lokalitách temperátní oblasti bylo
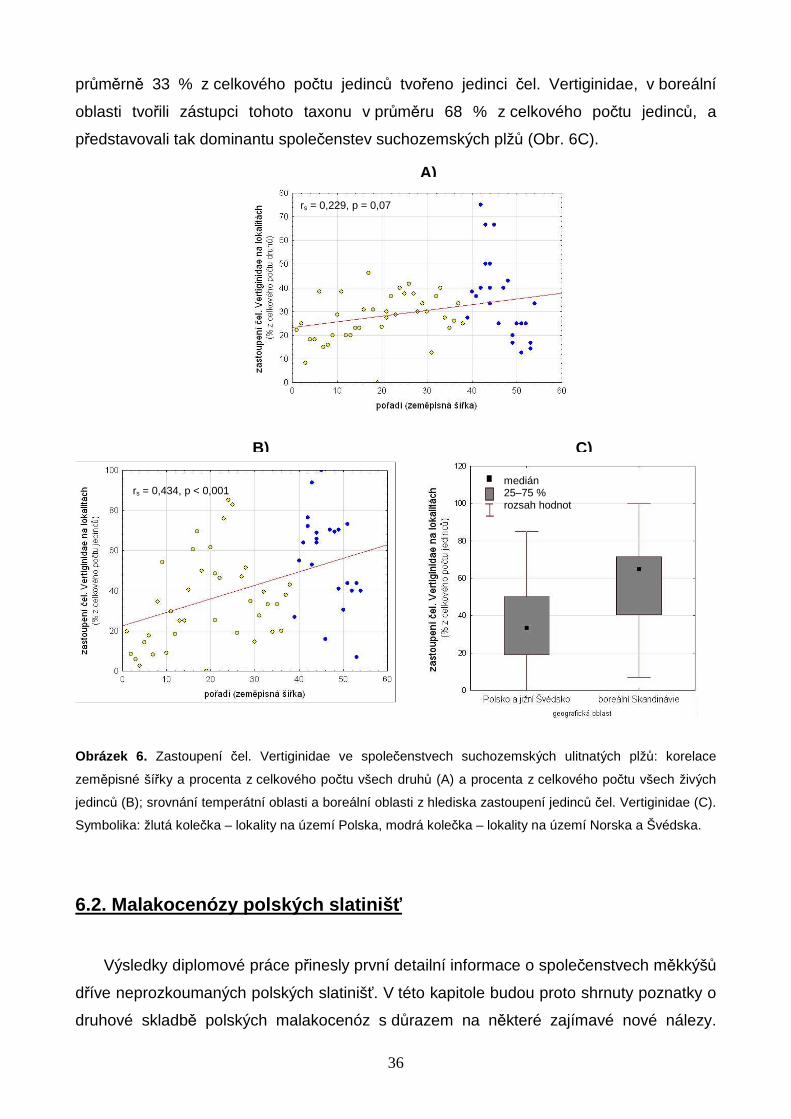
36
průměrně 33 % z celkového počtu jedinců tvořeno jedinci čel. Vertiginidae, v boreální
oblasti tvořili zástupci tohoto taxonu v průměru 68 % z celkového počtu jedinců, a
představovali tak dominantu společenstev suchozemských plžů (Obr. 6C).
Obrázek 6. Zastoupení čel. Vertiginidae ve společenstvech suchozemských ulitnatých plžů: korelace
zeměpisné šířky a procenta z celkového počtu všech druhů (A) a procenta z celkového počtu všech živých
jedinců (B); srovnání temperátní oblasti a boreální oblasti z hlediska zastoupení jedinců čel. Vertiginidae (C).
Symbolika: žlutá kolečka – lokality na území Polska, modrá kolečka – lokality na území Norska a Švédska.
6.2. Malakocenózy polských slatiniš ť
Výsledky diplomové práce přinesly první detailní informace o společenstvech měkkýšů
dříve neprozkoumaných polských slatinišť. V této kapitole budou proto shrnuty poznatky o
druhové skladbě polských malakocenóz s důrazem na některé zajímavé nové nálezy.
B) C)
medián 25–75 % rozsah hodnot
rs = 0,434, p < 0,001
A)
rs = 0,229, p = 0,07

37
Polská bazická slatiniště se vyznačovala jak vysokou diverzitou společenstev měkkýšů,
tak i značnou heterogenitou druhového složení mezi lokalitami jednotlivých částí
studovaného území. Jak ukazuje Příloha 3 (str. 73), rozdíly ve skladbě společenstev
souvisely především s odlišností malakofauny slatinišť regionu Západních Karpat oproti
malakofauně slatinišť rozsáhlých polských nížin dále na východ a severovýchod
zkoumaného území. Mezi druhy nalezené pouze na karpatských slatiništích patřily např.
Platyla polita, Macrogastra tumida, Aegopinella epipedostoma a Bythinella austriaca.
Specifikem slatinišť severovýchodní nížinné části země byla pozoruhodně bohatá vodní
malakofauna. Okolí četných jezer a zachovalá aluvia přirozeně meandrujících říčních toků
s množstvím periodicky zaplavovaných mokřadů, tůní a rašelinišť představovala unikátní
životní prostředí pro druhy jako Valvata cristata, Valvata macrostoma, Gyraulus riparius,
Bithynia leachii nebo Anisus septemgyratus. Nejhojnějšími druhy rozšířenými na
slatiništích všech částí Polska byly Carychium minimum, Euconulus praticola, Perpolita
hammonis, Vallonia pulchella a Vertigo geyeri.
Vzorkování polských slatinišť přineslo první spolehlivou recentní dokumentaci druhů
Vertigo geyeri, Pupilla pratensis a Sphaerium nucleus pro území Polska, a nově byly
objeveny také lokality druhů Pupilla alpicola a Vertigo moulinsiana. Status ohrožení nově
nalezených druhů měkkýšů je uveden v Tabulce 5 a o jejich ekologických nárocích,
rozšíření a ohroženosti bude detailněji pojednáno v Diskuzi (str. 50). V této kapitole budou
stručně charakterizovány lokality, na nichž byly výše uvedené druhy nalezeny (poloha
lokalit viz mapa území Polska, Obr. 1).
Drobný reliktní plž vrkoč Geyerův (Vertigo geyeri) byl do r. 2010 znám ze tří lokalit
v oblasti Oravsko-Nowotarské kotliny u hranic se Slovenskem (P2, P6, P10). Teprve při
systematických průzkumech východního Polska v letech 2010 až 2011 se ukázalo, že
výskyt vrkoče Geyerova se neomezuje pouze na slatiniště v oblasti Západních Karpat,
nýbrž že se jedná o druh poměrně hojně rozšířený po celém polském území v rámci
vhodných typů stanovišť (blíže viz Diskuze, str. 51). Druh byl nově dokumentován na
dalších 29 lokalitách od jihu přes východ až po severovýchod zkoumaného území (P5, P9,
P11–18, P20–38) a vyznačoval se relativně širokou ekologickou amplitudou ve vztahu
k minerální bohatosti – jeho lokality náležely jak mezi bazická až silně bazická slatiniště se
srážením pěnovce, tak mezi slatiniště s kalcitolernantními rašeliníky.
Dalším v Polsku nově zjištěným druhem byla zrnovka slatinná (Pupilla pratensis).
Objevena byla na slatiništi u obce Rowele (P37) při vzorkování v r. 2011. Jednalo se o
drobné, značně vodnaté prameniště se silným srážením pěnovce na mírném svahu,
obklopené křovinami a lesem. Nalezeni byli pouze 2 živí jedinci (a 34 prázdných

38
schránek), takže zrnovka slatinná zde pravděpodobně netvoří příliš početnou populaci.
Tentýž druh byl nejspíš nalezen také na svahovém pěnovcovém prameništi u obce
Bieniowce (P20) na severovýchodě Polska, kde však kvůli přítomnosti pouze prázdných a
nedostatečně vyvinutých schránek nebyla možná spolehlivá determinace do druhové
úrovně.
Druhem, který nebyl z území Polska recentně spolehlivě dokumentován, je také
drobný mlž okružanka kulovitá (Sphaerium nucleus), nalezený na lokalitě Brzeźno v údolí
řeky Nida ve střední části východního Polska (P15) v r. 2010 a na severovýchodě Polska
v údolí řeky Rospuda (P28) v r. 2011. V prvním případě se jednalo o vápnité slatiniště se
srážením pěnovce, tvořící ostrůvek ve zvodnělém mokřadu porostlém rákosinami. Ve
druhém případě šlo o vodnatou bazickou plochu na rozlehlém aluviálním slatiništi.
Nález reliktní zrnovky alpské (Pupilla alpicola) na rašeliništi Baligówka v blízkosti obce
Czarny Dunajec (P11) v r. 2011 představoval druhou známou populaci druhu na území
Polska. Rašeliniště Baligówka zahrnuje rozsáhlý komplex vrchovišť a bazických slatinišť s
lokálním srážením uhličitanu vápenatého, jehož značná část však byla v minulosti
znehodnocena těžbou rašeliny. Populace zrnovky alpské je zde pravděpodobně jen velmi
slabá, neboť v odebraném 12-ti litrovém vzorku se podařilo nalézt pouze 1 živého jedince
a 1 prázdnou schránku.
Nově byly v r. 2011 objeveny také dvě lokality vrkoče bažinného (Vertigo moulinsiana).
Jednou z nich byla Přírodní rezervace Torfy Orońskie, bazické slatiniště bez srážení
pěnovce situované asi 75 km jižně od Varšavy (P17), druhá lokalita se nacházela v údolí
řeky Rospuda (P27), kde se vrkoč bažinný vyskytoval na dosti heterogenní ploše tvořené
zvodnělými šlenky s převahou hnědých mechů a odrostlými bulty s výskytem
kalcitolerantních rašeliníků. V obou případech byl nalezen 1 živý jedinec a 2 prázdné
schránky.
Tabulka 5. Přehled významných nálezů měkkýšů pro území Polska a status jejich ohrožení.
Druh Status ohrožení
Pupilla alpicola (Charpentier, 1837) KO (Červená kniha PL)* Pupilla pratensis (Clessin, 1871) druh nový pro Polsko – status ohrožení není definován Vertigo geyeri Lindholm, 1925 Annex II **; Natura 2000 ***; druh nový pro Polsko Vertigo moulinsiana (Dupuy, 1849) KO (Červená kniha PL); Annex II ; Natura 2000 Sphaerium nucleus (Studer, 1820) druh nový pro Polsko – status ohrožení není definován
* druh kriticky ohrožený podle polské Červené knihy (Polish Red Data Book of Animals) ** druh zahrnut v Příloze II Směrnice o stanovištích - Annex II of the EU Habitats Directive (92/43/EEC) *** druh chráněný v rámci soustavy Natura 2000

39
6.3. Produktivita spole čenstev suchozemských plž ů
Závislost počtu druhů na produktivitě společenstev živých suchozemských ulitnatých
plžů byla modelována pomocí lineární regrese. Regresní modely byly vytvořeny jak pro
data z obou studovaných oblastí dohromady, tak odděleně pro Polsko a jižní Skandinávii.
Srovnání lineární regrese logaritmicky transformované biomasy a lineární regrese
s kvadratickým členem ukázalo, že větší procento z celkové variability dat vysvětluje
logaritmický model, ovšem pouze pro oblast Polska a pro data z obou oblastí dohromady
(Tabulka 6). Data ze Skandinávie se vyznačovala mírným unimodálním trendem, tedy
poklesem druhové bohatosti při nejvyšších hodnotách produktivity, a lépe je proto
popisoval model s kvadratickým členem (Tabulka 6, Obr. 7B). Srovnání obou testovaných
modelů pomocí Akaikeho informačního kritéria (AIC) však neprokázalo signifikantní rozdíl
mezi logaritmickým a kvadratickým modelem pro oblast Skandinávie, a potvrdilo vhodnost
logaritmického modelu pro Polsko a pro obě oblasti analyzované dohromady (Tabulka 6).
Pro data z obou studovaných oblastí byl proto zvolen logaritmický model, tedy křivka
druhové bohatosti dosahující asymptoty při maximálních hodnotách produktivity (Obr. 7A).
Výsledky dílčích t-testů a analýzy rozptylu ověřily, že vybraný logaritmický model je v obou
oblastech signifikantně významný a vysvětluje vysoký podíl variability dat. Průběh
závislosti počtu druhů na produktivitě se mezi oblastmi Polska a jižní Skandinávie
signifikantně nelišil; signifikantně odlišný byl pouze intercept lineární regrese, neboť
studované oblasti se vyznačovaly rozdílnou druhovou bohatostí (Obr. 7A).
Tabulka 6. Srovnání lineární regrese přirozeným logaritmem transformované biomasy (mg) a lineární
regrese s kvadratickým členem pro oba studované regiony dohromady a odděleně. Červeně jsou označeny
modely vysvětlující větší procento z celkové variability dat (Adj. R2).
model AIC F - statistika reziduální
st. volnosti p Adj. R 2
Polsko a Skandinávie 244,53 60,35 2, 59 <<0,0001 0,661 Polsko 154,83 63,43 1, 37 <<0,0001 0,622 Skandinávie
lineární regrese log. transformované
biomasy 91,98 18,27 1, 21 0,0003 0,440 Polsko a Skandinávie 255,89 31,30 3, 58 <<0,0001 0,598 Polsko 165.72 20,90 2, 36 <<0,0001 0,512 Skandinávie
lineární regrese s kvadratickým
členem 90,21 12,03 2, 20 0,0004 0,501
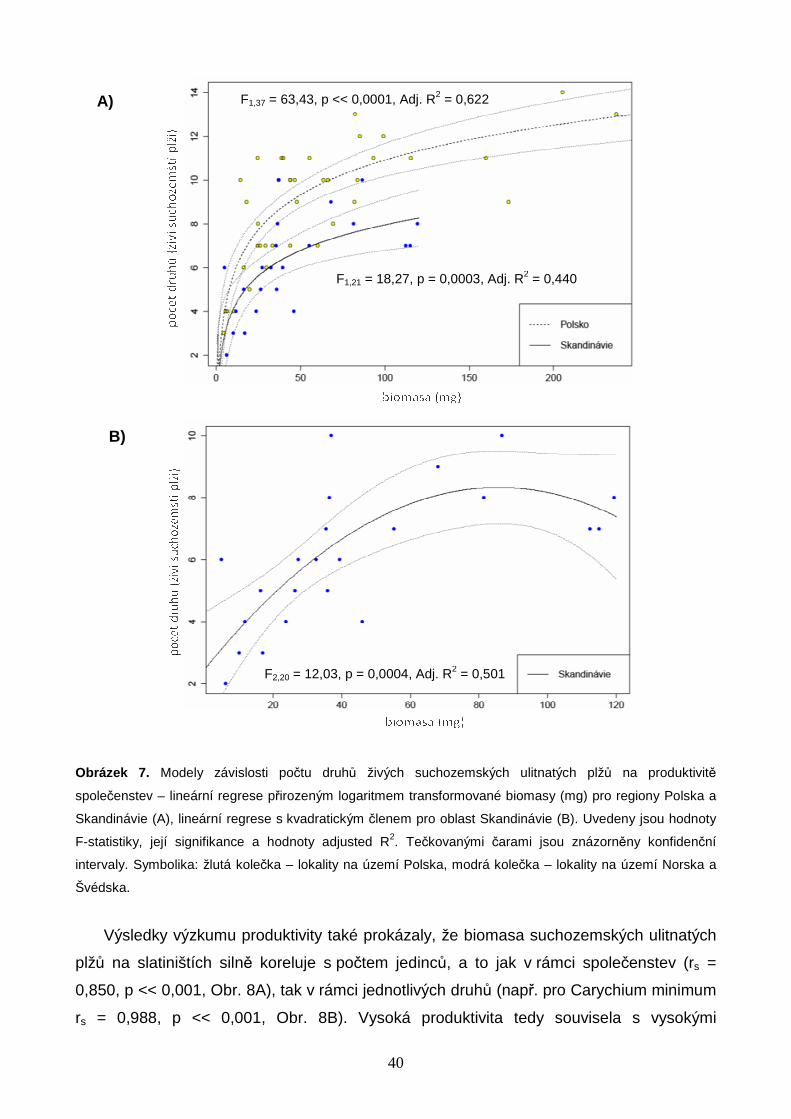
40
Obrázek 7. Modely závislosti počtu druhů živých suchozemských ulitnatých plžů na produktivitě
společenstev – lineární regrese přirozeným logaritmem transformované biomasy (mg) pro regiony Polska a
Skandinávie (A), lineární regrese s kvadratickým členem pro oblast Skandinávie (B). Uvedeny jsou hodnoty
F-statistiky, její signifikance a hodnoty adjusted R2. Tečkovanými čarami jsou znázorněny konfidenční
intervaly. Symbolika: žlutá kolečka – lokality na území Polska, modrá kolečka – lokality na území Norska a
Švédska.
Výsledky výzkumu produktivity také prokázaly, že biomasa suchozemských ulitnatých
plžů na slatiništích silně koreluje s počtem jedinců, a to jak v rámci společenstev (rs =
0,850, p << 0,001, Obr. 8A), tak v rámci jednotlivých druhů (např. pro Carychium minimum
rs = 0,988, p << 0,001, Obr. 8B). Vysoká produktivita tedy souvisela s vysokými
F1,37 = 63,43, p << 0,0001, Adj. R2 = 0,622
F2,20 = 12,03, p = 0,0004, Adj. R2 = 0,501
A)
B)
F1,21 = 18,27, p = 0,0003, Adj. R2 = 0,440
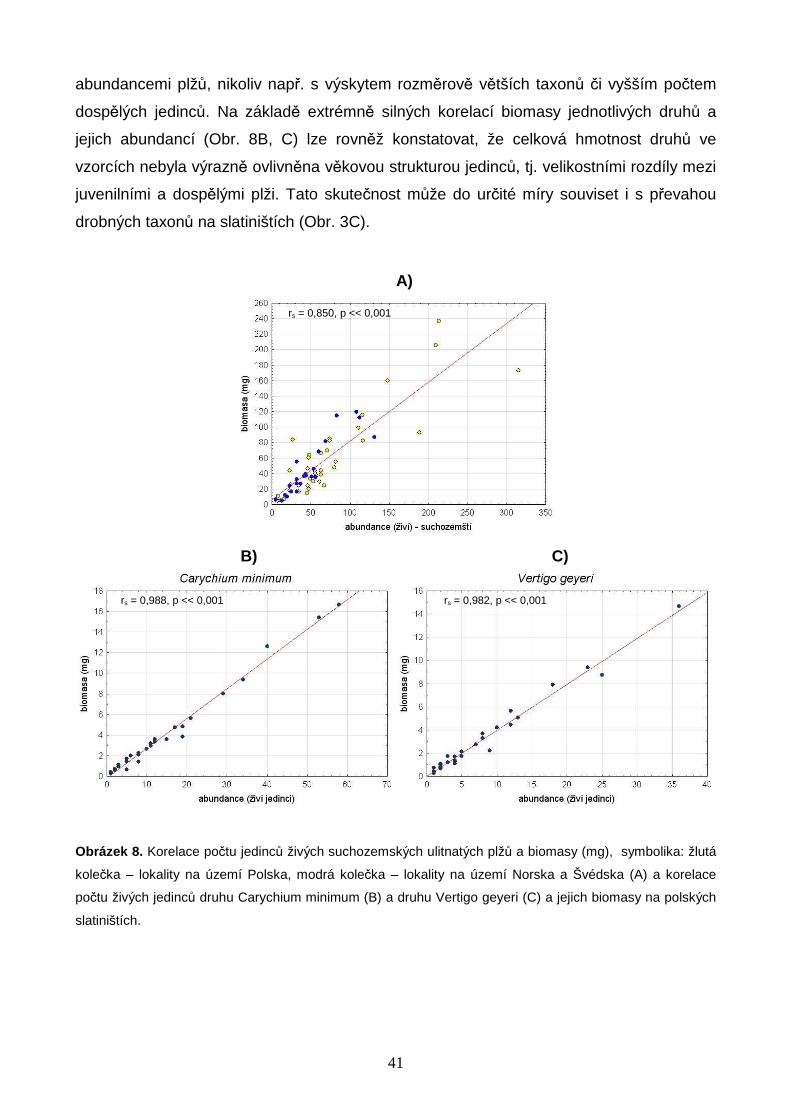
41
abundancemi plžů, nikoliv např. s výskytem rozměrově větších taxonů či vyšším počtem
dospělých jedinců. Na základě extrémně silných korelací biomasy jednotlivých druhů a
jejich abundancí (Obr. 8B, C) lze rovněž konstatovat, že celková hmotnost druhů ve
vzorcích nebyla výrazně ovlivněna věkovou strukturou jedinců, tj. velikostními rozdíly mezi
juvenilními a dospělými plži. Tato skutečnost může do určité míry souviset i s převahou
drobných taxonů na slatiništích (Obr. 3C).
Obrázek 8. Korelace počtu jedinců živých suchozemských ulitnatých plžů a biomasy (mg), symbolika: žlutá
kolečka – lokality na území Polska, modrá kolečka – lokality na území Norska a Švédska (A) a korelace
počtu živých jedinců druhu Carychium minimum (B) a druhu Vertigo geyeri (C) a jejich biomasy na polských
slatiništích.
A)
B) C)
rs = 0,850, p << 0,001
rs = 0,988, p << 0,001 rs = 0,982, p << 0,001

42
7. DISKUZE
7.1. Malakocenózy bazických slatiniš ť studovaných oblastí
7.1.1. Změny druhové bohatosti Výsledky diplomové práce potvrdily, že pokles druhové bohatosti měkkýšů směrem
k vyšším zeměpisným šířkám se týká i geograficky značně konzervativních biotopů, tedy
bazických slatinišť (Obr. 3A, B). Ve studované oblasti, zahrnující území od Západních
Karpat na jihu Polska po boreální zónu skandinávských zemí, souvisí pravděpodobně
pozorovaný trend s kvartérními změnami v rozšíření druhů, především s postglaciální
kolonizací severní Evropy z jižně položených refugií po ústupu Skandinávského ledovce.
Druhy klimaticky tolerantnější a schopné se efektivně šířit expandovaly po skončení
posledního glaciálu dále na sever a postupně osídlily i chladné, boreální a arktické oblasti
(HAUSDORF 2006). K významným evropským refugiím patřilo pravděpodobně karpatské
pohoří, představující v současnosti geografické centrum areálů rozšíření mnoha druhů
suchozemských plžů (HAUSDORF & HENNIG 2003). Význam karpatského regionu jakožto
refugia v dobách pleistocénních zalednění dokládá např. mimořádná rozmanitost lesních
společenstev měkkýšů oproti jiným regionům severní a západní Evropy (POKRYSZKO &
CAMERON 2005). Rovněž v rámci studovaného datového souboru patří karpatská slatiniště
k druhově nejpestřejším lokalitám (Příloha 3, str. 73).
Z grafů na Obr. 3A, B je patrné, že pokles druhové bohatosti měkkýšů podél
latitudinálního gradientu není zcela plynulý – výraznější odchylky od očekávaného trendu
jsou dány rozdíly v chemismu vody, rozloze, druhové skladbě okolních biotopů, stáří či
sukcesním stádiu vegetace jednotlivých lokalit. Nejnápadněji působí takřka skokový pokles
počtu druhů mezi třemi lokalitami v temperátní oblasti jižního cípu Švédska a zbývajícími
lokalitami boreální zóny Skandinávie, tedy zhruba mezi 55° a 62° severní ší řky. Právě
tímto územím, konkrétně zónou kolem 60. rovnoběžky v jižním Švédsku, prochází
významné biologické hraniční pásmo, v literatuře označované jako „Limes Norrlandicus“
(FRANSSON 1965), oddělující smíšené lesy mírného pásu od jehličnaté boreální tajgy. K
této přechodové zóně zasahují severní hranice areálů rozšíření mnoha evropských druhů
měkkýšů, které se již v chladnějších oblastech dále na sever nevyskytují (viz např. KERNEY
et al. 1983, VON PROSCHWITZ 2003). Z toho důvodu jsou lokality na nejjižnějším cípu

43
Švédska svou druhovou bohatostí bližší polským nežli ostatním skandinávským lokalitám
(Obr. 3A, B).
Poněkud překvapivě se jeví skutečnost, že negativní korelace se zeměpisnou šířkou
jsou silnější při zahrnutí vodních měkkýšů do celkových součtů druhů a jedinců (Tabulka
1). S různými ekologickými gradienty jsou totiž na slatiništích obvykle těsněji svázáni
suchozemští ulitnatí plži (HORSÁK et al. 2011a). Výrazný úbytek vodních druhů měkkýšů
směrem k lokalitám v oblasti Skandinávie souvisí však nejspíše se značným obohacením
nížinných polských slatinišť o druhy zachovalých mokřadních ekosystémů v blízkém okolí
(viz kapitola 7.2.2), nikoliv přímo s postupnými změnami diverzity vodní malakofauny
směrem na sever studovaného území.
7.1.2. Změny velikosti mediánového jedince Studie zabývající se velikostní strukturou malakocenóz obvykle dokumentují pokles
průměrné velikosti těla ve společenstvech suchozemských ulitnatých plžů směrem
k vyšším zeměpisným šířkám (HAUSDORF 2003, POKRYSZKO & CAMERON 2005, NEKOLA et
al. v tisku). Drobné druhy totiž lépe přežívají mrazy v zimním období, díky minimálnímu
obsahu vody v těle a nižšímu riziku poškození tkání vlivem vymrznutí (např. ANSART &
VERNON 2003). Výsledky diplomové práce však žádný velikostní trend nepotvrdily – na
všech slatiništích, nezávisle na geografické poloze, dominovaly ve společenstvech jedinci
náležející k velmi drobným druhům plžů, dosahujícím maximálních rozměrů 1,4 až 4 mm
(Obr. 3C a Příloha 5, str. 77). Slatiniště se totiž ve srovnání s mnoha jinými typy
terestrických biotopů vyznačují nižší produktivitou a homogenitou vegetačního pokryvu,
spojenou s nedostatkem vhodných úkrytů pro přežití zimních měsíců (SCHAMP et al. 2010).
Zdá se, že tyto extrémní podmínky společně s častou izolovaností slatinišť představují
jakýsi přírodní filtr, zvýhodňující právě drobné druhy. Ty se mohou na slatiniště snáze
rozšířit prostřednictvím pasivní disperze, a dlouhodobě zde přežívat díky schopnosti snížit
obsah vody v těle před obdobím mrazů na nutné minimum (NEKOLA et al. v tisku). Většina
druhů obývajících slatiniště tak reprezentuje konec možného velikostního spektra v rámci
suchozemských plžů, a průměrná velikost jedinců již téměř nemůže směrem k chladným
severně položeným lokalitám dále klesat.
Drobné rozdíly ve velikosti mediánového se nicméně mezi studovanými slatiništi přece
jen vyskytují. Poměrně nápadné je rozdělení lokalit do dvou velikostních skupin, kdy
v první z nich převažují plži se schránkou o objemu maximálně 4 mm3, a ve druhé plži
dosahující objemu 16 mm3 (s několika málo výjimkami na pomezí těchto skupin, viz Obr.

44
3C). Uvedené skupiny lokalit se od sebe výrazně neliší žádnou ze studovaných
vysvětlujících proměnných včetně pH a konduktivity vody, ačkoliv podle práce SCHAMP et
al. (2010) je zastoupení drobných druhů ve společenstvech slatiništních plžů nejvyšší na
extrémně vápnitých lokalitách. Určitou roli mohou hrát mírné rozdíly v postavení lokalit
podél trofického gradientu – zatímco první skupina lokalit se vyznačuje převahou typicky
slatiništních druhů měkkýšů, druhá skupina je charakteristická vyšším podílem méně
náročných ubikvistů jako Perpolita hammonis a Cochlicopa lubrica, preferujících živinami
bohatší, a tím produktivnější stanoviště lučního charakteru (HORSÁK et al. 2007a). Tuto
domněnku však nelze podložit konkrétními daty, neboť pro studované lokality nejsou k
dispozici přesné údaje o množství živin a produktivitě vegetace.
7.1.3. Faktory ovliv ňující skladbu spole čenstev suchozemských plž ů
Hlavní směr variability v datech o druhové skladbě společenstev suchozemských
ulitnatých plžů udávají především geograficky podmíněné rozdíly v klimatu studovaných
lokalit, jak ukazuje rozmístění vzorků v diagramu DCA a silné negativní korelace 1. osy s
průměrnými teplotami a zeměpisnou šířkou (Obr. 4, Tabulka 2). Podél gradientu daného 1.
osou došlo k vytvoření dvou jednoznačně vymezených shluků, představujících lokality na
území Polska, respektive jižní Skandinávie (Obr. 4), přičemž mezi polské lokality se dle
očekávání začlenila také 3 slatiniště na nejjižnějším cípu Švédska, náležející klimaticky i
vegetačně do zóny mírnému pásu (SJÖRS 1965a, DIERSSEN 1996). Podobně jako druhová
bohatost, také skladba společenstev měkkýšů se tedy nejvíce liší mezi lokalitami v oblasti
temperátních smíšených lesů a lokalitami v oblasti boreální jehličnaté tajgy, oddělenými
přechodovou zónou kolem 60. rovnoběžky v jižním Švédsku.
Příčinou nižší korelace 1. osy DCA s průměrnými lednovými teplotami oproti korelaci s
průměrnými červencovými teplotami (Tabulka 2) je patrně velmi mírné klima lokalit na
jižním cípu Švédska. Toto území se díky oceánickému charakteru vyznačuje daleko
příznivějšími podmínkami v zimním období než je pro regiony v obdobných zeměpisných
šířkách typické − jedná se dokonce o oblast s nejvyššími průměrnými lednovými teplotami
v rámci celého datového souboru včetně Polska (Příloha 1, str. 68).
Poměrně silná negativní korelace 1. osy DCA s konduktivitou svědčí o vzájemné
provázanosti klimatického gradientu a gradientu minerální bohatosti (Tabulka 2). Míra
vápnitosti jednotlivých lokalit totiž souvisí s regionálními rozdíly v charakteru geologického
podloží, které odpovídají např. za úplnou absenci vápnitých slatinišť v oblasti centrální
Skandinávie (HÁJEK et al. 2006). Mohlo by se proto zdát, že vliv gradientu minerální

45
bohatosti nelze plně oddělit od vlivu geografické polohy lokalit. Na základě srovnání
hodnot korelačních koeficientů (Tabulka 2) a s využitím znalosti druhové skladby
jednotlivých lokalit lze však spolehlivě zhodnotit, že minerální bohatost nebyla hlavním
faktorem odpovídajícím za rozdíly ve druhové skladbě malakocenóz – naprostá většina
skandinávských lokalit byla z hlediska konduktivity srovnatelná s lokalitami na území
Polska, ačkoliv jejich druhové složení se zásadním způsobem lišilo (viz Obr. 4 a Příloha 1,
str. 68).
Stejně tak i silná korelace 1. osy DCA s nadmořskou výškou (Tabulka 2) vypovídá
spíše o vlastnostech datového souboru než o přímé souvislosti se skladbou společenstev
plžů. Většina lokalit situovaných nejdále na severu je totiž součástí skandinávského
pohoří, a dosahuje proto nejvyšších nadmořských výšek (Příloha 1, str. 69). Nadmořská
výška má obvykle negativní vliv na diverzitu měkkýšů, zprostředkovaný zejména
klesajícími teplotami směrem k vyšším polohám (HORSÁK & CERNOHORSKY 2008, HORSÁK
et al. 2011a). V boreální zóně Skandinávie je však celá řada druhů dobře adaptována na
chladné klima (SOLHØY 1976, POKRYSZKO 1993), přičemž Golfský proud a teplé
jihozápadní proudění navíc výrazně zeslabují zimní teplotní extrémy, s ohledem na
zeměpisné šířky a hornatý charakter západní části skandinávských zemí. Opět lze proto
konstatovat, že nadmořská výška není v rámci studovaných dat rozhodujícím faktorem pro
skladbu společenstev plžů.
Změny malakocenóz podél 2. osy DCA zjevně souvisí s rozdíly ve srážkách, teplotách
a nadmořské výšce lokalit, ovšem korelace jsou s výjimkou ročních a letních srážkových
úhrnů slabší než korelace těchto proměnných s 1. osou (Tabulka 2). Faktory signifikantně
korelované s 2. osou tak mohou být svázány s jinými ekologickými parametry studovaných
lokalit, které jsou v terénu obtížněji měřitelné a které přímo odpovídají za rozdíly v druhové
skladbě společenstev. Klíčem k pochopení gradientu 2. osy je nápadné rozmístění
polských lokalit, od malých svahových pramenišť flyšových Karpat, vzniklých v důsledku
intenzivního odlesňování krajiny teprve před několika staletími (HORSÁK & HÁJKOVÁ 2005),
v horní části diagramu, po rozsáhlé a starobylé lokality s glaciální návazností včetně
reliktních slatinišť v oblasti Oravsko-Nowotarské kotliny (RYBNÍČEK & RYBNÍČKOVÁ 2003)
v dolní části diagramu (Obr. 4). Důležité parametry, které se podél tohoto gradientu mění,
jsou kromě nadmořské výšky a klimatických podmínek např. rozloha, vzdálenost k
druhově bohatému lesnatému okolí či míra zastínění lokalit. V souvislosti se znalostmi
vlivu historického vývoje na malakofaunu slatinišť v regionu Západních Karpat (např.
HORSÁK et al. 2007a, HÁJEK et al. 2011) lze proto 2. osu interpretovat jako komplexní
ekologický gradient podmíněný stářím a rozdílnou historií studovaných lokalit. Rozsáhlá

46
skandinávská slatiniště, pravděpodobně značně podobného stáří, se podél 2. osy výrazně
nerozmístila (Obr. 4), což by odpovídalo navržené interpretaci.
7.1.4. Druhové složení suchozemských plž ů ve studovaných oblastech
Jak bylo výše popsáno, podél latitudinálního gradientu dochází u společenstev
měkkýšů jak k očekávanému poklesu diverzity (Tabulka 1), tak k částečné obměně
druhového složení. Podstatně chudší skandinávská slatiniště se vyznačují přítomností
druhů specializovaných na chladné klimatické podmínky severských regionů (Obr. 5,
Tabulka 4). Tím se latitudinální gradient zcela zásadně odlišuje od gradientu minerální
bohatosti, podél kterého počet druhů směrem k méně bazickým slatiništím klesá, ovšem
namísto obměny druhového složení dochází pouze k úbytku náročných druhů s užší
vazbou na vápnitá stanoviště (HORSÁK & CERNOHORSKY 2008). Jak ukazují výsledky
diplomové práce, směrem ke klimaticky méně příznivému konci latitudinálního gradientu
dochází sice k vymizení teplomilnějších druhů, ale zároveň se objevují druhy vázané
výhradně na boreální zónu, s ekologickými optimy v chladných severských regionech.
Konkrétně se jedná o druhy Vertigo genesii, V. modesta, V. lilljeborgi a Columella
columella (Obr. 5), tedy glaciální relikty, které byly ve střední Evropě poměrně hojně
rozšířené v dobách pleistocénních zalednění, a teprve v průběhu holocénu došlo v rámci
Evropy k omezení jejich areálů na boreální zónu Skandinávie, případně vysokohorské
oblasti alpínského pásma ve střední Evropě (HAUSDORF & HENNIG 2003).
Skandinávská slatiniště jsou naopak ochuzena o celou řadu druhů typických pro
střední Evropu, zasahujících pouze okrajově do jižního Švédska, jako např. Vertigo
pygmaea, V. antivertigo, Vallonia enniensis a Pseudotrichia rubuginosa (Obr. 5).
Vzhledem k charakteru okolních biotopů, tvořených často tundrovou vegetací či
rozvolněnými jehličnatými porosty, zde pochopitelně chybí také lesní druhy plžů, běžně
pronikající na slatiniště mírného pásu (např. Monachoides incarnatus, Perforatella
bidentata a Vitrea crystallina). Některé druhy, které nebyly na lokalitách boreální zóny
Skandinávie nalezeny, se však v této oblasti vyskytovat můžou (viz KERNEY et al. 1983),
pouze nebyly vlivem nízkého počtu lokalit v průběhu vzorkování zachyceny, např. Vallonia
costata, Carychium tridentatum a Columella aspera (Obr. 5).
Zatímco druhová skladba lokalit v boreální zóně je velmi specifická a vyhraněná oproti
lokalitám mírného pásu, očekávaný rozdíl druhové skladby slatinišť Západních Karpat
oproti ostatním slatiništím mírného pásu je méně dobře patrný. Část karpatských slatinišť
se svou druhovou skladbou podobá nížinným polským lokalitám (P2, P9–11, Obr. 4), a

47
vyčlenění několika karpatských slatinišť v levé horní části diagramu DCA je způsobeno
především jejich obohacením o lesní druhy měkkýšů (viz kapitola 7.2.1.). Kromě
přítomnosti druhů pronikajících na karpatská slatiniště z okolních biotopů není fauna
suchozemských měkkýšů těchto lokalit nijak výjimečná (Příloha 3, str. 73). Všechny
typicky slatiništní a vlhkomilné druhy otevřených stanovišť byly nalezeny jak na lokalitách
v regionu Západních Karpat, tak i na ostatních lokalitách mírného pásu nížinné části
Polska a nejjižnějšího Švédska (s výjimkou druhu Pupilla alpicola, který byl v severněji
položených oblastech nahrazen druhem Pupilla pratensis, viz kapitola 7.2.3.).
7.1.5. Změny diverzity čeledi Vertiginidae
Fauna drobných suchozemských plžů čeledi Vertiginidae (zastoupená na slatiništích
rody Vertigo a Columella) je pozoruhodná vysokou druhovou diverzitou ve vyšších
zeměpisných šířkách, v rámci Evropy tedy na území Skandinávie (VON PROSCHWITZ 1993).
Skandinávské země se vyznačují takřka kompletním druhovým spektrem všech
evropských zástupců čel. Vertiginidae, přičemž mnohé z nich jsou zde plošně rozšířené, a
v ostatních částech Evropy se vyskytují jen roztroušeně (viz KERNEY et al. 1983). Patří k
nim i všechny čtyři druhy vázané na boreální zónu v rámci studovaných bazických slatinišť
(Obr. 5). Výsledky diplomové práce potvrdily signifikantní nárůst procentuálního
zastoupení jedinců čel. Vertiginidae směrem k vyšším zeměpisným šířkám (Obr. 6B, C),
známý také např. z rozsáhlého výzkumu společenstev suchozemských plžů v Severní
Americe (J.C. NEKOLA, nepublikovaná data). Mnohé druhy rodů Vertigo a Columella jsou
plně adaptovány na klimatické podmínky chladných severských oblastí, takže jako jedny z
mála zde dosahují vysokých populačních hustot a představují časté dominanty
společenstev (POKRYSZKO 1993). V rámci skandinávských slatinišť platí uvedené tvrzení
zvláště pro druh Vertigo genesii, který byl nalezen na 17 z 20 studovaných lokalit (Tabulka
4) a představuje nejhojnější druh boreálních slatinišť jak z hlediska frekvence výskytu, tak
z hlediska abundancí (z celkového počtu 1464 jedinců suchozemských plžů náleželo 607
jedinců druhu Vertigo genesii, Příloha 3, str. 73). Zajímavé je, že s výjimkou studovaných
regionů s vápnitým podložím ve skandinávském pohoří a švédské provincii Jämtland je
tento druh po celém území Skandinávie jen velmi vzácný, lokální a značně roztroušený
(VON PROSCHWITZ 2003).
Změny druhové bohatosti čel. Vertiginidae podél latitudinálního gradientu však
v diplomové práci prokázány nebyly: na slatiništích Polska a jižního cípu Švédska se
vyskytuje dohromady 8 druhů, na slatiništích boreální zóny Skandinávie 7 druhů čel.

48
Vertiginidae (Příloha 3, str. 73) a procento druhů příslušejících tomuto taxonu z celkového
počtu druhů na lokalitách signifikantně nekoreluje se zeměpisnou šířkou (ačkoli rozložení
hodnot v grafu na Obr. 6A naznačuje pozitivní trend směrem k severně položeným
oblastem). Podíl čel. Vertiginidae na celkové druhové bohatosti lokalit je po celé délce
latitudinálního gradientu relativně vysoký a konstantní, ačkoli oblast boreální zóny se
v souvislosti s velmi nízkými počty druhů vyznačuje vyššími výkyvy v diverzitě tohoto
taxonu. Velmi zřetelně se ovšem obměňuje druhová skladba čel. Vertiginidae, kdy pro
boreální a temperátní region jsou společné pouze druhy Vertigo geyeri, V. substriata a
Columella edentula (Příloha 3, str. 73).
7.2. Malakofauna bazických slatiniš ť východního Polska
Společenstva měkkýšů 39 polských slatinišť sestávala z 64 druhů, z nichž celkem 53
druhů lze považovat za typicky slatiništní nebo alespoň schopné zde po určitou dobu
přežívat – zbývajících 11 druhů měkkýšů nebylo na žádné ze studovaných lokalit nalezeno
živých, a nejspíše se tedy jednalo o náhodný výskyt prázdných schránek z okolních
biotopů. Druhová bohatost polských slatinišť je tak v rámci střední Evropy srovnatelná
např. s bohatostí vnějších Západních Karpat na moravsko-slovenském pomezí, kde bylo
nalezeno 57 měkkýších druhů na 48 lokalitách (HORSÁK & HÁJEK 2003), a značně
převyšuje druhovou rozmanitost bulharských slatinišť, na kterých bylo při obdobném počtu
lokalit dokumentováno 44 druhů (HORSÁK et al. 2011a). Nové nálezy vzácných druhů,
bohatost a unikátní skladba společenstev měkkýšů svědčí o velmi zachovalém charakteru
polských slatinišť, která by si v mnoha případech zasluhovala vhodný management a
přísnější ochranu. Stejně jako celá střední Evropa je i Polsko značně ohrožené
pokračujícím úbytkem slatinišť, doprovázeným nevratnými ztrátami biodiverzity vázané na
tyto výjimečné ekosystémy (GROOTJANS et al. 2006, VAN DIGGELEN et al. 2006).
7.2.1. Slatiništ ě polské strany Západních Karpat
Charakter slatinišť na polské straně Karpat je dosti odlišný od slatinišť v nížinných
oblastech východního a severovýchodního Polska. Studované karpatské lokality náleží do
flyšového pásma vnějších Západních Karpat a většinu z nich reprezentují menší svahová
prameniště, uzavřená v křovinatých a lesnatých porostech. Je známo, že otevřený
charakter těchto slatinišť je relativně mladý a souvisí s intenzivním odlesňováním krajiny

49
započatém před asi 600–700 lety (HORSÁK & HÁJKOVÁ 2005). Výjimku představují reliktní
slatiniště v oblasti Oravsko-Nowotarské kotliny, jejichž původ spadá až do období starého
holocénu (RYBNÍČEK & RYBNÍČKOVÁ 2003).
Druhová skladba malakocenóz na svahových prameništích flyšových Karpat je často
obohacována působením tzv. okrajového efektu, kterým se rozumí pronikání lesních druhů
plžů na otevřenou plochu slatiniště (LACINA 2010). Díky malé rozloze a blízkosti druhově
bohatého lesnatého okolí se proto na studovaných karpatských slatiništích vyskytují jak
živí jedinci přísně lesních druhů plžů (Aegopinella epipedostoma, A. pura, Macrogastra
tumida, Monachoides incarnatus a Platyla polita), tak i četné prázdné schránky mnoha
dalších lesních druhů (např. Isognomostoma isognomostomos, Monachoides vicinus a
Urticicola umborsus). Naproti tomu na plošně rozsáhlých a pravděpodobně velmi
starobylých lokalitách v oblasti Oravsko-Nowotarské kotliny (P9–11, Obr. 1) se druhy
náležející k lesní malakofauně vůbec nevyskytují. Díky fenoménu okrajového efektu
převyšuje 11 karpatských slatinišť s počtem 34 druhů suchozemských plžů zbývajících 28
lokalit v nížinné části Polska s pouhými 30 druhy (Příloha 3, str. 73).
7.2.2. Slatiništ ě střední a severní části Polska
Jelikož slatiniště Západních Karpat byla a stále jsou předmětem četných
malakologických výzkumů (např. HORSÁK & HÁJEK 2003, HORSÁK et al. 2007a, b, HÁJEK et
al. 2011), větší neznámou představovala doposud zcela neprobádaná slatiniště střední a
severní části východního Polska. Výsledky průzkumu nížinných polských slatinišť
vypovídají o návaznosti těchto lokalit na konec posledního glaciálu a počátek holocénu,
jak naznačuje druhová skladba blízká starobylým lokalitám v centrální části Západních
Karpat (viz HORSÁK et al. 2007a) a přítomnost glaciálního reliktu Vertigo geyeri (HÁJEK et
al. 2011), který byl nalezen téměř na všech vzorkovaných nížinných slatiništích (viz
kapitola 7.2.3.).
Díky zachovalému charakteru a obvykle značné rozloze sestává fauna suchozemských
plžů nížinných lokalit téměř výhradně z druhů otevřených stanovišť a slatiništních
specialistů, jakými jsou např. Euconulus praticola, Vallonia enniensis, Vertigo geyeri a V.
angustior, a druhy okolních lesních biotopů (Fruticicola fruticum, Monachoides incarnatus,
Perforatella bidentata a Vitrea crystallina) se zde vyskytují jen zcela ojediněle (Příloha 3,
str. 73). Při bližším pohledu je patrné, že malakofauna suchozemských plžů nížinných
lokalit je takřka podmnožinou druhového spektra karpatských lokalit, ochuzenou díky
absenci okrajového efektu o většinu lesních druhů. Druhy, jejichž výskyt byl zaznamenán

50
pouze na nížinných slatiništích (např. Columella aspera, Pseudotrichia rubiginosa,
Perpolita petronella a Vitrea crystallina), se mohou vyskytovat také na slatiništích
Západních Karpat (HORSÁK 2005b, CERNOHORSKY et al. 2010), a při vzorování pouze
nebyly zachyceny vlivem nízkého počtu lokalit. Výjimku tvoří pouze druhy Pupilla alpicola
a P. pratensis, jejichž areály rozšíření se v rámci Evropy nepřekrývají (viz kapitola 7.2.3.).
Zajímavým atributem polských nížinných lokalit je velmi bohatá malakofauna vodních
druhů měkkýšů. Především východ a severovýchod studovaného území (např. NP
Poleski, NP Biebrza, NP Wigry či údolí řeky Rospuda) se vyznačuje unikátními
mokřadními komplexy v rozsáhlých aluviích říčních toků a jezer, s množstvím periodických
tůní a sezónně přeplavovaných rašelinišť. Tato zachovalá mokřadní stanoviště představují
optimální prostředí pro společenstva citlivých vodních plžů jako Gyraulus riparius, Valvata
macrostoma, Anisus septemgyratus nebo Bithynia leachii, které se běžně vyskytují také na
bazických slatiništích v jejich blízkém okolí. Mnohé z nich patří mezi druhy značně
ohrožené úbytkem a degradací biotopů v důsledku regulací řek a následného zarůstání a
zazemňování mokřadů (viz např. BERAN 1998). Slatiniště na východě a severovýchodě
Polska jsou díky obohacení o druhy zachovalých nížinných mokřadů výrazně bohatší na
vodní plže nežli karpatská slatiniště (12 oproti 6 druhům; Příloha 3, str. 74).
Jen velmi zřídka se v severní části Polska setkáváme s drobnými svahovými
prameništi karpatského typu – k takovým patří v rámci navštívených lokalit pouze dvě
slatiniště se srážením pěnovce u obcí Rowele (P37) a Bieniowce (P20) na severovýchodě
Polska, obě uzavřené v lesnatém či křovinatém okolí. Není s podivem, že i přes malou
rozlohu jsou tyto lokality v rámci nížinných polských slatinišť výrazně nejbohatší co se týče
suchozemských plžů, a naopak zcela postrádají výše popisovanou faunu náročných
mokřadních druhů.
7.2.3. Významné nálezy pro Polsko
Velmi potěšující byly četné nové nálezy drobného plže vrkoče Geyerova (Vertigo
geyeri), představujícího ve střední Evropě vzácný pozdně glaciální relikt (LOŽEK 1992),
chráněný mezinárodní směrnicí EU o stanovištích (Annex II of the Habitats and Species
Directive). Vrkoč Geyerův se vyznačuje boreo-alpínským rozšířením, a jeho výskyt je proto
vázán především na území Skandinávie (POKRYSZKO 1993, VON PROSCHWITZ 2003) a
roztroušené reliktní lokality ve střední Evropě (HÁJEK et al. 2011). V Polsku byl po dlouhou
dobu známý pouze jediný nález druhu z oblasti Svatokřížských hor, u kterého byla navíc
později odhalena nesprávná determinace a záměna s jiným druhem r. Vertigo (viz

51
POKRYSZKO 1990). Jedinou spolehlivě dokumentovanou populaci poté představoval nález
na lokalitě Chochołów (P6), publikovaný v práci HORSÁK & HÁJEK (2005). Protože vrkoč
Geyerův poměrně hojně obývá starobylá slatiniště vnitřních Západních Karpat (VAVROVÁ et
al. 2009), jeho výskyt na polské straně pohoří byl relativně předvídatelný. Poněkud
překvapivější byly nálezy velkého množství lokalit ve východní a severovýchodní části
země. Celkem bylo objeveno 31 nových lokalit, takže se zdá, že nedostatečná znalost
rozšíření vrkoče Geyerova v Polsku pramení především z naprosté absence výzkumů
biotopů, které druh obývá. Těmi jsou výhradně zachovalá otevřená slatiniště reliktního
charakteru s velmi stabilním hydrologickým režimem, od slatinišť s výskytem
kalcitolerantních rašeliníků až po vápnitá pěnovcová prameniště (např. HORSÁK & HÁJEK
2005, KUCZYŃSKA & MOORKENS 2010). Znalosti o rozšíření vrkoče Geyerova jsou v celé
střední Evropě nejspíš dosti neúplné, především kvůli jeho nenápadnosti, vyhraněným
ekologickým nárokům a úzké vazbě na specifické a vzácné biotopy (SCHENKOVÁ et al.
2012).
K významným nálezům patří také dva druhy zrnovek, zrnovka alpská (Pupilla alpicola)
a z. slatinná (P. pratensis). Zrnovka alpská představuje v Evropě vzácný a ohrožený relikt
z pozdního glaciálu s roztroušeným výskytem v Alpách a Západních Karpatech (HORSÁK et
al. 2011b). Jde o druh vázaný na bezlesá, silně vápnitá pěnovcová slatiniště, obývající
podobně jako vrkoč Geyerův starobylé reliktní lokality (např. HORSÁK et al. 2011b, HÁJEK et
al. 2011). V polské Červené knize je druh označen jako kriticky ohrožený a jeho jediná
populace byla dříve známa z lokality u obce Niedzica v Pieninách (ALEXANDROWICZ 1994).
Při vzorkování pro diplomovou práci byla zrnovka alpská objevena asi o 30 km dále na
západ na slatiništi Baligówka (P11).
Nález zrnovky slatinné na slatiništi Rowele (P37) představuje první recentně a
spolehlivě doloženou populaci druhu na území Polska. Až do současnosti byla známa
pouze jedna její lokalita, a to na základě 120 let starého dokladového materiálu (viz VON
PROSCHWITZ et al. 2009), bez aktuálního ověření výskytu druhu. Ekologické nároky
zrnovky slatinné se velmi podobají nárokům zrnovky alpské, ovšem areály jejich rozšíření
se v rámci Evropy zřejmě nepřekrývají – zrnovka slatinná se vyskytuje především v
zemích severní a západní Evropy (VON PROSCHWITZ et al. 2009). Situaci však komplikují
předběžné výsledky molekulárních analýz, podle kterých zrnovka alpská a slatinná
reprezentují možná pouze geografické poddruhy jediného, hojněji rozšířeného druhu
(HORSÁK et al. 2012). To však nic nemění na faktu, že oba druhy jsou na polských
slatiništích extrémně vzácné – ačkoli bylo vzorkováno velké množství potenciálně

52
vhodných stanovišť, populace zrnovek (navíc jen velmi málo početné) byly nalezeny
pouze na dvou z nich.
Příjemné byly rovněž dva nové nálezy v Polsku kriticky ohroženého vrkoče bažinného
(Vertigo moulinsiana), mezinárodně chráněného směrnicí EU o stanovištích. Vrkoč
bažinný je v Evropě považován za relikt z raných fází holocénu (LOŽEK 1964) a v
současnosti je souvisleji rozšířen na jihu Evropy (SPEIGHT et al. 2003). Ve střední Evropě
tvoří jeho areál izolované ostrůvky v podobě trvale zvodnělých vápnitých mokřadů a
rákosových či ostřicových porostů v nížinných oblastech (např. POKRYSZKO 1990,
CAMERON et al. 2003, JUEG 2004). Vrkoč bažinný tedy není přísným slatiništním
specialistou, ačkoliv může obývat i otevřená vápnitá slatiniště (VAVROVÁ et al. 2009). Za
lokální a ojedinělý výskyt vrkoče bažinného v rámci střední Evropy odpovídá především
drastický úbytek zachovalých mokřadních stanovišť. V Polsku byly donedávna známy
pouze 2 populace druhu (viz KSIĄŻKIEWICZ 2009), v současnosti však spolehlivě
doložených nálezů přibývá, takže se zdá, že vrkoč bažinný je podobně jako vrkoč Geyerův
v Polsku rozšířen daleko více, než se dříve předpokládalo (KSIĄŻKIEWICZ 2009, BARGA-
WIĘCŁAWSKA 2012). Díky stále výjimečně zachovalému charakteru mokřadních
ekosystémů má Polsko značný potenciál pro nové nálezy a trvalé udržení populací tohoto
vzácného reliktního druhu.
Co se týče polské vodní malakofauny je důležité zmínit dva nálezy drobného
mokřadního mlže, okružanky kulovité (Sphaerium nucleus), který byl teprve nedávno
vyčleněn z druhového komplexu Sphaerium corneum agg. a povýšen na samostatný druh
(KORNIUSHIN 2001). Kvůli dřívějšímu slučování uvedených druhů vyžaduje poznání
současného rozšíření revizi staršího dokladového materiálu, neboť celá řada nálezů
označovaných jako Sphaerium corneum přísluší ve skutečnosti druhu S. nucleus
(KOŘÍNKOVÁ et al. 2008). Objevené lokality tak nejspíše představují první recentní a
ověřený výskyt populací spolehlivě determinovaných jako Sphaerium nucleus na území
Polska.
7.3. Vztah druhové bohatosti a produktivity malakoc enóz
Jedním z cílů diplomové práce bylo také prostudovat vztah druhové bohatosti a
produktivity společenstev suchozemských ulitnatých plžů. Pro rostlinná společenstva
různých biotopů včetně slatinišť bývá vztah mezi uvedenými proměnnými často popisován
jako unimodální, kvůli stresovým podmínkám při nízkých hodnotách produktivity a zvýšené
mezidruhové kompetici při vysokých hodnotách produktivity (např. GRIME 1973, GRACE

53
1999, OLDE VENTERINK et al. 2001). Výsledky diplomové práce však ukazují, že ve
společenstvech suchozemských ulitnatých plžů slatinišť se v rámci studovaného rozsahu
proměnných mezidruhové interakce nejspíše neprojevují (Obr. 7A). Náznak poklesu
druhové bohatosti při nejvyšších hodnotách produktivity v datech ze Skandinávie je patrně
pouze náhodný a souvisí s nízkým počtem vzorků. Logaritmický model navíc lépe
odpovídá realitě při nízkých hodnotách produktivity, neboť kvadratický model předpokládá
určitý počet druhů i při nulové produktivitě (Obr. 7B).
Na lokalitách s nejvyšší produktivitou dochází tedy pravděpodobně k nasycení
společenstev vlivem vyčerpání regionální zásoby druhů schopných obývat tento typ
biotopu, tzv. „species pool“ (PÄRTEL et al. 1996), a vliv kompetice je pouze zanedbatelný.
Ani na nejproduktivnějších lokalitách, tj. lokalitách s největší koncentrací jedinců (Obr. 8A),
nedochází totiž k výraznějšímu poklesu druhové bohatosti (Obr. 7A). Ačkoli existují i
důkazy kompetičního vyloučení mezi suchozemskými plži (např. MORDAN 1977), obecně je
kompetici ve společenstvech měkkýšů přikládána spíše menší váha (např. BOYCOTT 1934,
WALDÉN 1981). Důsledky mezidruhových interakcí mezi plži nebyly dosud prokázány ani
na slatiništích (HORSÁK 2006, HORSÁK & CERNOHORSKY 2008), obývaných převážně
herbivorními a detritovorními druhy (SCHAMP et al. 2010). Limitující faktor pro nárůst
druhové bohatosti plžů podél gradientu produktivity tedy představuje regionální zásoba
druhů, která je v rámci studovaných území signifikantně vyšší pro oblast Polska nežli pro
oblast jižní Skandinávie (Obr. 7A).
O možném vlivu kompetice jsem na základě unimodálního vztahu mezi produktivitou a
počtem druhů skandinávských slatinišť (Obr. 7B) spekulovala ve své bakalářské práci.
S ohledem na nízký počet vzorků, signifikantně nevýznamný rozdíl mezi kvadratickým a
logaritmickým modelem pro oblast jižní Skandinávie (Tabulka 6) a vyšší druhovou
bohatost na slatiništích Polska při obdobných i vyšších počtech jedinců (Obr. 7A) lze tuto
úvahu pravděpodobně zamítnout.
Pro účely budoucích studií může být užitečná informace, že biomasa společenstev
ulitnatých plžů silně koreluje s počtem jedinců (Obr. 8A). Tato korelace nejspíše souvisí s
převahou drobných taxonů na slatiništích (Obr. 3C), a taktéž s rovnoměrným zastoupením
jedinců různých věkových (hmotnostních) kategorií v jednotlivých vzorcích (viz Obr. 8B,
C). V obdobných výzkumech by proto bylo možné aproximovat hmotnost biomasy
celkovým počtem živých jedinců (Obr. 8A).

54
8. ZÁVĚR
V rámci této diplomové práce byla poprvé podrobně studována druhová skladba a
bohatost malakocenóz bazických slatinišť východního Polska. Důležitou náplní práce bylo
rovněž vyhodnocení změn společenstev měkkýšů podél gradientu zeměpisné šířky, od
slatinišť na území Polska po slatiniště v jižní části Švédska a Norska, studovaných
v bakalářské práci.
V teoretické části diplomové práce byly shrnuty poznatky o ekologii slatinišť a
specifických rysech malakofauny těchto biotopů, a obecně charakterizovány přírodní
poměry studovaných území. Pozornost byla věnována také ekologickým fenoménům
latitudinálního gradientu diverzity a vztahu mezi produktivitou a druhovou bohatostí, které
byly v diplomové práci nově studovány na příkladu suchozemských slatiništních měkkýšů.
(Cíl 1)
Vlastní terénní průzkum byl proveden na 39 bazických slatiništích východního Polska,
od jižně položených lokalit v oblasti Západních Karpat až po lokality v severovýchodní
části území při hranicích s Ruskem a Litvou. Z datového souboru zpracovaného v rámci
bakalářské práce bylo vybráno 23 bazických slatinišť na území jižní Skandinávie, a na
základě vyhodnocení obou studovaných oblastí bylo zjištěno, že:
• druhová bohatost společenstev měkkýšů klesá směrem k vyšším zeměpisným šířkám,
přičemž nejnápadnější je pokles počtu druhů mezi slatiništi jižního cípu Švédska,
náležejícími do mírného pásu, a slatiništi boreální zóny Skandinávie;
• velikost těla suchozemských plžů se podél zeměpisné šířky nemění, neboť na všech
studovaných slatiništích zcela dominují drobné druhy, které jsou díky nízkému obsahu
vody v těle dobře adaptovány na přežití mrazů v podmínkách nízké produktivity
slatiništní vegetace;
• směrem k severně položeným lokalitám výrazně roste zastoupení jedinců čel.
Vertiginidae ve společenstvech suchozemských plžů, takže v boreálním regionu tvoří
zástupci tohoto taxonu (především boreo-alpínský druh Vertigo genesii) naprostou
dominantu společenstev;
• nejvýznamnější ekologický gradient odpovídající za rozdíly ve druhové skladbě
malakocenóz souvisí především s průměrnými teplotami a geografickou polohou
lokalit; podél tohoto gradientu se studované lokality dělí do dvou dobře vymezených
skupin: (1) slatiniště boreální zóny Norska a Švédska, charakteristické chladnomilnými
severskými druhy Vertigo genesii, V. modesta, V. lilljeborgi a Columella columella, a (2)

55
slatiniště mírného pásu v Polsku a nejjižnějším Švédsku, s výskytem klimaticky méně
tolerantních druhů, které do boreální zóny nezasahují, např. Vertigo angustior, V.
antivertigo a Vallonia enniensis. (Cíl 4)
Na studovaných polských slatiništích bylo nalezeno celkem 64 druhů měkkýšů s
maximem 27 druhů na lokalitu. Učiněny byly významné nové nálezy pro území Polska,
představované druhy Vertigo geyeri, V. moulinsiana, Pupilla alpicola, P. pratensis a
Sphaerium nucleus. Patrné byly rozdíly mezi slatiništi flyšového pásma polských
Západních Karpat, obohacenými vlivem okrajového efektu o druhy z lesnatého okolí, a
rozsáhlými slatiništi nížinné části země, s četným výskytem vodních druhů měkkýšů
z okolních zachovalých mokřadních biotopů. (Cíle 2 a 3)
Mezi produktivitou a druhovou bohatostí byla v obou studovaných oblastech nalezena
logaritmická závislost – počet druhů roste lineárně s přirozeným logaritmem biomasy
společenstev, a maximální dosažená druhová bohatost je omezena regionální zásobou
druhů potenciálně schopných obývat daný typ biotopu. Vliv kompetice, který by se projevil
poklesem počtu druhů na nejproduktivnějších lokalitách, je ve společenstvech slatiništních
plžů nejspíše pouze zanedbatelný. (Cíl 5)
Prezentované výsledky diplomové práce přinesly první ucelené údaje o unikátních,
zachovalých a druhově velmi pestrých bazických slatiništích východního Polska, a přispěly
taktéž k poznání obecných ekologických aspektů slatiništních malakocenóz na velkých
prostorových škálách.

56
9. CITOVANÁ LITERATURA
ALEXANDROWICZ S.W. (1994) Pupilla alpicola (Charpentier, 1837) from Niedzica. Geologia,
20, 325-331.
ALMENDINGER J.E., LEETE J.H. (1998) Peat characteristics and groundwater geochemistry
of calcareous fens in the Minnesota River Basin, U.S.A. Biogeochemistry, 43, 17-41.
AL-MUFTI M.M., SYDES C.L., FURNESS S.B., GRIME J.P., BAND S.R. (1977) A quantitative
analysis of shoot menology and dominance in herbaceous vegetation. Journal of
Ecology, 65, 759-791.
AMON J.P., THOMPSON C.A., CARPENTER Q.J., MINER J. (2002) Temperate zone fens of the
glaciated Midwestern USA. Wetlands, 22, 301-317.
ANDERSEN J., HALVORSEN O. (1984) Species composition, abundance, habitat requirements
and regional distribution of terrestrial Gastropods in Arctic Norway. Polar Biology, 3,
45-53.
ANSART A., VERNON P. (2003) Cold hardiness in molluscs. Acta Oecologica, 24, 95-102.
AUSDEN M., HALL M., PEARSON P., STRUDWICK T. (2005) The effect of cattle grazing on tall-
herb fen vegetation and molluscs. Biological Conservation, 122, 317-326.
AXMANOVÁ I., CHYTRÝ M., DANIHELKA J., LUSTYK P., KOČÍ M., KUBEŠOVÁ S., HORSÁK M.,
CHEROSOV M.M., GOGOLEVA A. Plant species richness-productivity relationships in a
low-productive region: exploring the effects of competition versus species-pool
limitation. Odesláno.
BARGA-WIĘCŁAWSKA J.A. (2012) New Localities of Vertigo moulinsiana (Dupuy, 1849) in
south-eastern, central and northern Poland. Folia Malacologica, 20, 43-71.
BARKER G.M. (2001) The biology of terrestrial molluscs. CABI publishing, New Zealand.
BEDFORD B.L., WALBRIDGE M.R., ALDOUS A. (1999) Patterns in nutrient availability and plant
diversity of temperate North American wetlands. Ecology, 80, 2151-2169.
BER A. (2006) Pleistocene interglacials and glaciations of northeastern Poland compared
to neighbouring areas. Quaternary International, 149, 12-23.
BERAN L. (1998) Vodní měkkýši ČR. Metodika Českého svazu ochránců přírody 17, ZO
ČSOP Vlašim.
BERGMANN C. (1847) Über die Verhältnisse der Wärmeökonomie der Thiere zu ihrer
Grösse. Gottinger Studien, 3, 595-708.

57
BISHOP M.J. (1981) Quantitative studies on some living British wetland mollusc faunas.
Biological Journal of the Linnean Society, 15, 299-326.
BLACKBURN T.M., GASTON K.J., LODER N. (1999) Geographic gradients in body size: a
clarification of Bergmann's rule. Diversity and Distributions, 5, 165-174.
BOEYE D., VERHAGEN B., VAN HAESEBROECK V., VERHEYEN R.F. (1997) Nutrient limitation in
species-rich lowland fens. Journal of Vegetation Science, 8, 415-424.
BOYCOTT A.E. (1934) The habitats of land mollusca in Britain. Journal of Ecology, 22, 1-38.
BOYER M.L.H., WHEELER B.D. (1989) Vegetation patterns in spring-fed calcareous fens:
calcite precipitation and constraints on fertility. Journal of Ecology, 77, 597-609.
BRAGAZZA L., RYDIN H., GERDOL R. (2005) Multiple gradients in mire vegetation: a
comparison of a Swedish and an Italian bog. Plant Ecology, 177, 223-236.
BROWN J.H., LOMOLINO M.V. (1998) Biogeography. Sinauer, Sunderland.
BURNHAM K.P., ANDERSON D.R. (2004) Multimodel inference: understanding AIC and BIC in
Model Selection. Sociological Methods and Research, 33, 261-304.
CAMERON R.A.D. (1978) Terrestrial snail faunas of the Malham area. Field Studies, 4, 715-
728.
CAMERON R.A.D., COLVILLE B., FALKNER G., HOLYOAK G.A., HORNUNG E., KILLEEN I.J.,
MOORKENS E.A., POKRYSZKO B.M., VON PROSCHWITZ T., TATTERSFIELD P., VALOVIRTA I.
(2003) Species accounts for snails of the genus Vertigo listed in Annex II of the
Habitats Directive: V. angustior, V. genesii, V. geyeri and V. moulinsiana (Gastropoda,
Pulmonata: Vertiginidae). In: M.C.D. Speight, E. Moorkens, G. Falkner (eds)
Proceedings of the workshop on conservation biology of European Vertigo species
(Dublin, April 2002). Heldia, 5, pp. 151-170.
CAMERON R.A.D., POKRYSZKO B.M. (2005) Estimating the species richness and composition
of land mollusc communities: problems, consequences and practical advice. Journal of
Conchology, 38, 529-547.
CARDILLO M., ORME C.D.L., OWENS I.P.F. (2005) Testing for latitudinal bias in diversification
rates: an example using New World birds. Ecology, 86, 2278-2287.
CERNOHORSKY N.H., HORSÁK M., CAMERON R.A.D. (2010) Land snail species richness and
abundance at small scales: the effects of distinguishing between live individuals and
empty shells. Journal of Conchology, 40, 233-241.
CLARKE A., CRAME J.A. (2003) The importance of historical processes in global patterns of
diversity. In: T.M. Blackburn, and K.J. Gaston (eds) Macroecology Concepts and
Consequences. Blackwell Scientific, Oxford, pp. 130-151.

58
COLWELL R.K., HURTT G.C. (1994) Nonbiological gradients in species richness and a
spurious Rapoport effect. American Naturalist, 144, 570-595.
COLWELL R.K., LEES D.C (2000) The mid-domain effect: geometric constraints on the
geography of species richness. Trends in Ecology and Evolution, 15, 70-76.
CONNOLLY S.R. (2005) Process-based models of species distributions and the mid-domain
effect. American Naturalist, 166, 1-11.
DAVIES P., GALE C.H., LEES M. (1996) Quantitative studies of modern wet-ground
molluscan faunas from Bossington, Hampshire. Journal of Biogeography, 23, 371-377.
DIERSSEN K. (1996) Vegetation nordeuropas. Verlag Eugen Ulmer, Stuttgart.
ESRI (2003) ArcGIS 8.3. Environmental Systems Research Institute, Redlands, CA, USA.
Available at: http://www.esri.com.
FOECKLER F., DEICHNER O., SCHMIDT H., CASTELLA E. (2006) Suitability of molluscs as
bioindicators for meadow- and flood-channels of the Elbe-floodplains. International
Review of Hydrobiology, 91, 314-325.
FOURNIÉ J., CHÉTAIL M. (1984) Calcium dynamics in land gastropods. American Zoologist,
24, 857-870.
FRANSSON S. (1965) The borderland. In: The plant cover of Sweden. Acta
Phytogeographica Suecica, 50, 167-175.
GASTON K.J. (2000) Global patterns in biodiversity. Nature, 405, 220-227.
GRACE J.B. (1999) The factors controlling species density in herbaceous plant
communities: an assessment. Perspectives in Plant Ecology, Evolution and
Systematics, 2, 1-28.
GRIME J.P. (1973) Competitive exclusion in herbaceous vegetation. Nature, 242, 344–347.
GRIME J.P. (1979) Plant strategies and vegetation processes. John Wiley and Sons,
London.
GROOTJANS A.P., ADEMA E.B., BLEUTEN W., JOOSTEN H., MADARAS M., JANÁKOVÁ M. (2006)
Hydrological landscape settings of base-rich fen mires and fen meadows: an overview.
Applied Vegetation Science, 9, 175-184.
GROOTJANS A., WOŁEJKO L. (2007) Conservation of wetlands in Polish agricultural
landscapes / Ochrona mokradeł w rolniczych krajobrazach Polski. Wydawnictvo Klubu
Przyrodników.
HÁJEK M., HEKERA P., HÁJKOVÁ M. (2002) Spring fen vegetation and water chemistry in the
Western Carpathian flysch zone. Folia Geobotanica, 37, 205-224.
HÁJEK M., HORSÁK M., HÁJKOVÁ P., DÍTĚ D. (2006) Habitat diversity of central European
fens in relation to environmental gradients and an effort to standardise fen terminology

59
in ecological studies. Perspectives in Plant Ecology, Evolution and Systematics, 8, 97-
114.
HÁJEK M., HÁJKOVÁ P., APOSTOLOVA I., HORSÁK M., PLÁŠEK V., SHAW B., LAZAROVA M. (2009)
Disjunct occurrences of plant species in the refugial mires of Bulgaria. Folia
Geobotanica, 44, 365-386.
HÁJEK M., HORSÁK M., TICHÝ L., HÁJKOVÁ P., DÍTĚ D., JAMRICHOVÁ E. (2011) Testing a relict
distributional pattern of fen plant and terrestrial snail species at the Holocene scale: a
null model approach. Journal of Biogeography, 38, 742-755.
HÁJKOVÁ P., WOLF P., HÁJEK M. (2004) Environmental factors and Carpathian spring fen
vegetation: the importance of scale and temporal variation. Annales Botanici Fennici,
41, 249-262.
HAUSDORF B. (2003) Latitudinal and altitudinal body size variation among north-west
European land snail species. Global Ecology and Biogeography, 12, 389-394.
HAUSDORF B. (2006) Latitudinal and altitudinal diversity patterns and Rapoport effects in
north-west European land snails and their causes. Biological Journal of the Linnean
Society, 87, 309-323.
HAUSDORF B., HENNIG C. (2003) Nestedness of north-west European land snail ranges as a
consequence of differential immigration from Pleistocene glacial refuges. Oecologia,
135, 102-109.
HAWKINS B.A., FIELD R., CORNELL H.V., CURRIE D.J., GUÉGAN J., KAUFMAN D.M., KERR J.T.,
MITTELBACH G.G., OBERDORFF T., O'BRIEN E.M., PORTER E.E., TURNER J.R.G. (2003)
Energy, water and broad-scale geographic patterns of species richness. Ecology, 84,
3105-3117.
HIJMANS R.J., CAMERON S.E., PARRA J.L., JONES P.G., JARVIS A. (2005) Very high resolution
interpolated climate surfaces for global land areas. International Journal of Climatology,
25, 1965-1978.
HILL T., LEWICKI P. (2007) STATISTICS Methods and Applications. StatSoft, Tulsa, OK.
(http://www.statsoft.com/textbook/stathome.html).
HILLBRICHT-ILKOWSKA A., RYBAK J., RZEPECKI M. (2000) Ecohydrological research of lake–
watershed relations in diversified landscape (Masurian Lakland, Poland). Ecological
Engineering, 16, 91-98.
HILLEBRAND H. (2004) On the generality of the latitudinal diversity gradient. The American
Naturalist, 163, 192-211.
HOLM S. (1979) A simple sequentially rejective multiple test procedure. Scandinavian
Journal of Statistics, 6, 65-70.

60
HORSÁK M. (2003) How to sample mollusc communities in mires easily. Malacologica
Bohemoslovaca, 2, 11-14.
HORSÁK M. (a) (2005) The uniqueness of spring fens and malacological consequences.
Malacologica Bohemoslovaca, 3, 89-99.
HORSÁK M. (b) (2005) Molluscs. In: A. Poulíčková, M. Hájek, K. Rybníček (eds) Ecology
and palaecology of spring fens in the western part of the Carpathians, Palacký
University, Olomouc, pp. 197-208.
HORSÁK M. (2006) Mollusc community patterns and species response curves along a
mineral richness gradient: a case study in fens. Journal of Biogeography, 33, 98-107.
HORSÁK M. (2011) Mollusc assemblages in palaeoecological reconstructions: an
investigation of their predictive power using transfer function models. Boreas, 40, 459-
467.
HORSÁK M., HÁJEK M. (2003) Composition and species richness of mollusc communities in
relation to vegetation and water chemistry in the Western Carpathian spring fens: the
poor-rich gradient. Journal of Molluscan Studies, 69, 349-357.
HORSÁK M., HÁJEK M. (2005) Habitat requirements and distribution of Vertigo geyeri
(Gastropoda: Pulmonata) in Western Carpathian rich fens. Journal of Conchology, 38,
683-700.
HORSÁK M., HÁJKOVÁ P. (2005) The historical development of the White Carpathian spring
fens based on palaeomalacological data. In: A. Poulíčková, M. Hájek, K. Rybníček
(eds) Ecology and palaecology of spring fens in the western part of the Carpathians,
Palacký University, Olomouc, pp. 63-68.
HORSÁK M., HÁJEK M., DÍTĚ D., TICHÝ L. (a) (2007) Modern distribution patterns of snails
and plants in the Western Carpathian spring fens: is it a result of historical
development? Journal of Molluscan Studies, 73, 53-60.
HORSÁK M., HÁJEK M., TICHÝ L., JUŘÍČKOVÁ L. (b) (2007) Plant indicator values as a tool for
land mollusc autecology assessment. Acta Oecologica, 32, 161-171.
HORSÁK M., CERNOHORSKY N. (2008) Mollusc diversity patterns in Central European fens:
hotspots and conservation priorities. Journal of Biogeography, 35, 1215-1225.
HORSÁK M., HÁJEK M., HÁJKOVÁ P., CERNOHORSKY N., APOŠTOLOVÁ I. (a) (2011) Mollusc
communities in Bulgarian fens: predictive power of the environment, vegetation and
spatial structure. Naturwissenschaften, 98, 671-681.
HORSÁK M., ŠKODOVÁ J., CERNOHORSKY N.H. (b) (2011) Ecological and historical
determinants of Western Carpathian populations of Pupilla alpicola (Charpentier, 1837)

61
in relation to its present range and conservation. Journal of Molluscan Studies, 77, 248-
254.
HORSÁK M., SCHENKOVÁ V. MYŠÁK J. (2012) The second site of Pupilla alpicola
(Charpentier, 1837) and the first recent record of Pupilla pratensis (Clessin, 1871) in
Poland. Folia Malacologica, 20, 21-26.
HUSTON M. (1979) A general hypothesis of species diversity. The American Naturalist, 113,
81-101.
HYLANDER K., NILSSON C., JONSSON B.G., GÖTHNER T. (2005) Differences in habitat quality
explain nestedness in a land snail meta-community. Oikos, 108, 351-361.
JABŁOŃSKA E., PAWLIKOWSKI P., JARZOMBKOWSKI F., CHORMAŃSKI J., OKRUSZKO T.,
KŁOSOWSKI S. (2011) Importance of water level dynamics for vegetation patterns in a
natural percolation mire (Rospuda fen, NE Poland). Hydrobiologia, 674, 105-117.
JABLONSKI D., ROY K., VALENTINE J.W. (2006) Out of the tropics: evolutionary dynamics of
the latitudinal diversity gradient. Science, 314, 102-106.
JOOSTEN H., CLARKE D. (2002) Wise use of mires and peatlands - Background and
principles including a framework for decision making. International Mire Conservation
Group and International Peat Society, NHBS, Totnes, Devon.
JUEG U. (2004) Die Verbreitung und Ökologie von Vertigo moulinsiana (Dupuy, 1849) in
Mecklenburg – Vorpommern (Gastropoda: Stylommatophora: Vertiginidae).
Malakologische Abhandlungen, 22, 87-124.
KERNEY M.P., CAMERON R.A.D. (1979) Land snails of Britain and North-West Europe.
Collins, London.
KERNEY M.P., CAMERON R.A.D., JUNGBLUTH J.H. (1983) Die Landschnecken Nord- und
Mitteleuropas. Paul Parey, Hamburg and Berlin, Germany.
KOCZUR A. (2006) Importance of vegetation in the Orawsko-Nowotarskie peat bogs to
biological diversity in the Polish Carpathians. Acta Agrophysica, 7, 383-393.
KORNIUSHIN A.V. (2001) Taxonomic revision of the genus Sphaerium s.l. in the palaearctic
region, with some notes on the North American species. Archiv für Molluskenkunde,
129, 77-122.
KOŘÍNKOVÁ T., BERNA L., HORSÁK M. (2008) Recent distribution of Sphaerium nucleus
(Studer, 1820) (Bivalvia: Sphaeriidae) in the Czech Republic. Malacologica
Bohemoslovaca, 7, 26-32.
KSIĄŻKIEWICZ Z. (2009) New localities of Vertigo moulinsiana (Dupuy, 1849) (Gastropoda:
Pulmonata: Vertiginidae) in Northwestern Poland. Folia Malacologica, 17, 219-222.

62
KUCZYŃSKA A., MOORKENS E. (2010) Micro-hydrological and micro-meteorological controls
on survival and population growth of the whorl snail Vertigo geyeri Lindholm, 1925 in
groundwater fed wetlands. Biological Conservation, 143, 1868-1875.
LACINA A. (2010) Okrajový efekt jako fenomén určující skladbu malakofauny otevřených
vápnitých slatinišť na kontaktu s lesními stanovišti. Diplomová práce, Masarykova
univerzita, Brno.
LOŽEK V. (1956) Klíč Československych měkkyšů. Vydavatelstvo Slovenskej akademie
vied, Bratislava.
LOŽEK V. (1964) Quartärmollusken der Tschechoslowakei. Rozpravy Ústředního ústavu
geologického, Praha, 31, 1-374.
LOŽEK V. (1981) Měkkýši jako modelová skupina v ochranářském výzkumu. Památky a
příroda, 6, 171-178.
LOŽEK V. (1990) Mokřady v historickém pohledu. Památky a příroda, 15, 611-618.
LOŽEK V. (1992) Měkkýši (Mollusca). In: L. Škapec (ed.) Červená kniha ohrožených a
vzácných rostlin a živočichů ČSFR. 3. Bezobratlí. Príroda, Bratislava, pp. 22-33.
LUCHTEL D.L., DEYRUP-OLSEN I. (2001) Body wall: form and function. In: G.M. Barker (ed)
The biology of terrestrial molluscs. CABI Publishing, London, pp. 147-178.
Malmer N. (1965) The southern mires. Acta phytogeographica Suecica, 50, 149-158.
Malmer N. (1986) Vegetational gradients in relation to environmental conditions in
northwestern European mires. Canadian Journal of Botany, 64, 375-383.
MARKS L. (2004) Pleistocene glacial limits in Poland. In: J. Ehlers and P.L. Gibbard (eds)
Quaternary Glaciations – Extents and Chronology. Elsevier BV, pp. 295-300.
MARKS L. (2010) Timing of the Late Vistulian (Weichselian) glacial phases in Poland.
Quaternary Science Reviews, doi:10.1016/j.quascirev.2010.08.008.
MARTIN K., SOMMER M. (2004) Relationships between land snail assemblage patterns and
soil properties in temperate-humid forest ecosystems. Journal of Biogeography, 31,
531-545.
MCCLAIN C.R., NEKOLA J.C. (2008) The role of local-scale processes on terrestrial and
deep-sea gastropod body size distributions across multiple scales. Evolutionary
Ecology Research, 10, 129-146.
MITTELBACH G.G., STEINER C.F., SCHEINER S.M., GROSS K.L., REYNOLDS H.L., WAIDE R.B.,
WILLIG M.R., DODSON S.I., GOUGH L. (2001) What is the observed relationship between
species richness and productivity? Ecology, 82 , 2381-2396.

63
MITTELBACH G.G., SCHEMSKE D.W., CORNELL H.V., ALLEN A.P., BROWN J.M. et al. (2007)
Evolution and the latitudinal diversity gradient: speciation, extinction and biogeography.
Ecology Letters, 10, 315-31.
MORDAN P.B. (1977) Factors affecting the distribution and abundance of Aegopinella and
Nesovitrea (Pulmonata:Zonitidae) at the Monks Wood National Nature Reserve,
Huntingdonshire. Biological Journal of the Linnean Society, 9, 59-72.
NEKOLA J.C. (1999) Paleorefugia and neorefugia: the influence of colonization history on
community pattern and process. Ecology, 80, 2459-2473.
NEKOLA J.C. (2005) Geographic variation in richness and shell size of eastern North
American land snail communities. Records of the Western Australian Museum,
Supplement No. 68, 39-51.
NEKOLA J.C., BARKER G.M., CAMERON R.A.D., POKRYSZKO B.M. Latitudinal and longitudinal
variation of body size in land snail populations and communities. In: F. Smith, K. Lyons
(eds) Global Patterns of Body Size. University of Chicago Press. V tisku.
NILSSON A. (1987) Terrestrial molluscs from the western part of the Torneträsk area. Fauna
Norrlandica, 5, 1-16.
OLDE VENTERINK H., WASSEN M.J., BELGERS D.M., VERHOEVEN J.T. (2001) Control of
environmental variables on species density in fens and meadows: importance of direct
effects and effects through community biomass. Journal of Ecology, 89, 1033-1040.
OLDE VENTERINK H., WASSEN M.J., VERKROOST A.W.M., DE RUITER P.C. (2003) Species
richness-productivity patterns differ between N-, P-, and K-limited wetlands. Ecology,
84, 2191-2199.
OLDE VENTERINK H., KARDEL I., KOTOWSKI W., PEETERS W., WASSEN M.J. (2009) Long-term
effects of drainage and hay-removal on nutrient dynamics and limitation in the Biebrza
mires, Poland. Biogeochemistry, 93, 235-252.
ØKLAND R.H., ØKLAND T., RYDGREN K. (2001) A Scandinavian perspective on ecological
gradients in north-west European mires: reply to Wheeler and Proctor. Journal of
Ecology, 89, 481-486.
PÄRTEL M., ZOBEL M., ZOBEL K., VAN DER MAAREL E. (1996) The species pool and its relation
to species richness: evidence from estonian plant communities. Oikos, 75, 111-117.
PÄRTEL M., LAANISTO L., ZOBEL M. (2007) Contrasting plant productivity-diversity
relationships across latitude: the role of evolutionary history. Ecology, 88, 1091-1097.
PAWLIKOWSKI P., JARZOMBKOWSKI F., WOŁKOWYCKI D, KOZUB Ł., ZANIEWSKI P., BAKANOWSKA
O., BANASIAK Ł, BARAŃSKA K., BIELSKA A., BIEREŻNOJ U., GALUS M., GRZYBOWSKA M.,
KAPLER A., KARPOWICZ J., SADOWSKA I, ZARZECKI R. (2009) Rare and threatened plants

64
of the mires in the intensively managed landscape of the Góry Sudawskie region
(north-eastern Poland). Botanika – Steciana, 13, 29-36.
PAWŁOWSKA E., POKRYSZKO B.M. (1998) Why are terrestrial gastropods of Poland
threatened? Folia Malacologica, 6, 63-71.
PEKÁR S., BRABEC M. (2009) Moderní analýza biologických dat. Zobecněné lineární modely
v prostředí R. Scientia, Praha.
PIANKA E.R. (1966) Latitudinal gradients in species diversity: a review of concepts. The
American Naturalist, 100, 33-46.
POKRYSZKO B.M. (1990) The Vertiginidae of Poland (Gastropoda: Pulmonata: Pupilloidea)
– a systematic monograph. Annales Zoologici, 43, 133-257.
POKRYSZKO B.M. (1993) Fen malacocenoses in Dovrefjell (S.Norway). Fauna norvegica
Series A, 14, 27-38.
POKRYSZKO B.M., CAMERON R.A.D. (2005) Geographical variation in the composition and
richness of forest snail faunas in northern Europe. Records of the Western Australian
Museum, 68, 115-132.
R DEVELOPMENT CORE TEAM (2010) R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-
07-0, URL http://www.R-project.org.
ROHDE K. (1992) Latitudinal gradients in species diversity: the search for the primary
cause. Oikos, 65, 514-527.
ROSENZWEIG M.L. (1995) Species diversity in space and time. Cambridge University Press,
Cambridge.
ROSENZWEIG M.L. (2003) How to reject the area hypothesis of latitudinal gradients. In: T.M.
Blackburn, K.J. Gaston (eds) Macroecology: Concepts and Consequences. Blackwell
Publishing, Oxford, pp. 87-106.
ROUSSEAU D.D., LIMONDIN N., MAGNIN F., PUISSEGUR J.J. (1994) Temperature oscillations
over the last 10,000 years in western Europe estimated from terrestrial mollusc
assemblages. Boreas, 23, 66-73.
ROY K., JABLONSKI D., VALENTINE J.W. (1996) Higher taxa in biodiversity studies: patterns
from eastern Pacific marine molluscs. Philosophical Transactions of the Royal Society
of London B, 351, 1605-1613.
RYBNÍČEK K., RYBNÍČKOVÁ E. (2003) Vegetation history of the Upper Orava Region in the
last 11000 years. Acta Palaeobotanica, 42, 153-170.
RYDIN H., JEGLUM J.K. (2006) The biology of peatlands. Oxford University Press, Oxford.

65
SAFFORD H.D., REJMÁNEK M., HADAČ E. (2001) Species pools and the ‘‘hump-back’’ model
of plant species diversity: an empirical analysis at a relevant spatial scale. Oikos, 95,
282-290.
SCHAMP B., HORSÁK M., HÁJEK M. (2010) Deterministic assembly of land snail communities
according to species size and diet. Journal of Animal Ecology, 79, 803-810.
SCHENKOVÁ V., HORSÁK M., PLESKOVÁ Z., PAWLIKOWSKI P. (2012) Habitat preferences and
conservation of Vertigo geyeri (Gastropoda: Pulmonata) in Slovakia and Poland.
Journal of Molluscan Studies, 78, 105-111.
SIENKIEWICZ J. (2008) Ramsar sites in Poland. Institute of Environmental Protection,
Warsaw.
SJÖRS H. (a) (1965) Features of land and climate. Acta phytogeographica Suecica, 50, 1-
12.
SJÖRS H. (b) (1965) Northern mires – Regional ecology of mire sites and vegetation. Acta
phytogeographica Suecica, 50, 180-188.
SJÖRS H., GUNNARSSON U. (2002) Calcium and pH in north and central Swedish mire
waters. Journal of Ecology, 90, 650-657.
SKEFFINGTON M.S., MORAN J., CONNOR Á.O., REGAN E., COXON C.E., SCOTT N.E., GORMALLY
M. (2006) Turloughs - Ireland's unique wetland habitat. Biological Conservation, 133,
265-290.
SOLHØY T. (1976) Terrestrial gastropods (Mollusca, Gastropoda: Basommatophora and
Stylommatophora). Fauna of the Hardangervidda, 10, 23-45.
SPEIGHT M.C.D., MOORKENS E., FALKNER G. (eds) (2003) Proceedings of the workshop on
conservation biology of European Vertigo species (Dublin, April 2002). Heldia, 5, 1-
183.
SULIKOWSKA-DROZD A., HORSÁK M. (2007) Woodland mollusc communities along
environmental gradients in the East Carpathians. Biologia, 62, 201-209.
TAHVANAINEN T. (2004) Water chemistry of mires in relation to the poor-rich vegetation
gradient and contrasting geochemical zones of the north-eastern fennoscandian
Shield. Folia Geobotanica, 39, 353-369.
TAYLOR J.D., TAYLOR C.N. (1977) Latitudinal distribution of predatory gastropods on the
eastern Atlantic shelf. Journal of Biogeography, 4, 73-81.
TAYLOR D.R., AARSSEN L.W., LOEHLE C. (1990) On the relationship between r/K selection
and environmental carrying capacity: a new habitat templet for plant life history
strategies. Oikos, 58, 239-250.

66
TERBORGH J. (1973) On the notion of favorableness in plant ecology. American Naturalist,
107, 481-501
TER BRAAK C.J.F., ŠMILAUER P. (2002) CANOCO Reference manual and Canodraw for
Windows user’s guide. Software for Canonical Community Ordination (version 4.5).
Biometris, Wageningen & České Budějovice.
TILMAN D., WEDIN D., KNOPS J. (1996) Productivity and sustainability influenced by
biodiversity in grassland ecosystems. Nature, 379, 718-720.
VALDOVINOS C., NAVARRETE S.A., MARQUET P.A. (2003) Mollusk species diversity in the
Southeastern Pacific: why are there more species towards the pole? Ecography, 26,
139-144.
VAVROVÁ Ľ., HORSÁK M., ŠTEFEK J. & ČEJKA T. (2009) Ecology, distribution and conservation
of Vertigo species of European importance in Slovakia. Journal of Conchology, 40, 61-
71.
VAN DIGGELEN R., MIDDLETON B., BAKKER J., GROOTJANS A., WASSEN M. (2006) Fens and
floodplains of the temperate zone: Present status, threats, conservation and
restoration. Applied Vegetation Science, 9, 157-162.
VON PROSCHWITZ T. (1993) Habitat selection and distribution of ten vertiginid species in the
province of Dalsland (SW. Sweden) (Gastropoda. Pulmonata: Vertiginidae).
Malakologische Abhandlungen , 16, 177-217.
VON PROSCHWITZ T. (2003) A review of the distribution, habitat selection and conservation
status of the species of the genus Vertigo in Scandinavia (Denmark, Norway and
Sweden) (Gastropoda, Pulmonata: Vertiginidae). In: M.C.D. Speight, E. Moorkens, G.
Falkner (eds) Proceedings of the workshop on conservation biology of European
Vertigo species (Dublin, April 2002). Heldia, 5, pp. 27-50.
VON PROSCHWITZ T., SCHANDER C., JUEG U., THORKILDSEN S. (2009) Morphology, ecology
and DNA-barcoding distinguish Pupilla pratensis (Clessin, 1871) from Pupilla
muscorum (Linnaeus, 1758) (Pulmonata: Pupillidae). Journal of Molluscan Studies, 75,
315-322.
WAIDE R.B., WILLIG M.R., STEINER C.F., MITTELBACH G., GOUGH L., DODSON S.I., JUDAY G.P.,
PARMENTER R. (1999) The relationship between productivity and species richness.
Annual Review of Ecology and Systematics, 30, 257-300.
WALDÉN H.W. (1981) Communities and diversity of land molluscs in Scandinavian
woodlands. I. High diversity communities in taluses and boulder slope in SW Sweden.
Journal of Conchology, 30, 351-372.

67
WALDÉN H.W. (1995) Norway as an environment for terrestrial molluscs, with viewpoints on
threats against species and diversity. In: A.C. van Bruggen, S.M. Wells & T.h.C.M.
Kemperman (eds) Proceedings of the Alan Solen Memorial Symposium on the
Biodiversity and Conservation of the Mollusca, 11th International Malacological
Congress (Siena, Italy, 1992), pp. 111-132.
WÄREBORN I. (1969) Land molluscs and their environments in an oligotrophic area in
southern Sweden. Oikos, 20, 461-479.
WÄREBORN I. (1970) Environmental factors influencing the distribution of land molluscs of
an oligotrophic area in southern Sweden. Oikos, 21, 285-291.
WÄREBORN I. (1979) Reproduction of two species of land snails in relation to calcium salts
in the Foerna layer. Malacologia, 18, 177-180.
WASSEN M.J., VENTERINK H.O., LAPSHINA E.D., TANNEBERGER F. (2005) Endangered plants
persist under phosphorus limitation. Nature, 437, 547-550.
WHEELER B.D., GILLER K.E. (1982) Species richness of herbaceous fen vegetation in
broadland, Norfolk in relation to the quantity of above-ground plant material. Journal of
Ecology, 70, 179-200.
WHEELER B.D., PROCTOR M.C.F. (2000) Ecological gradients, subdivisions and terminology
of north-west European mires. Journal of Ecology, 88, 187-203.
WHITTAKER R.H., WILLIS K.J., FIELD R. (2001) Scale and species richness: towards a
general, hierarchical theory of species diversity. Journal of Biogeography, 28, 453-70.
WILLIG M.R., KAUFMAN D.M., STEVENS R.D. (2003) Latitudinal gradients of biodiversity:
pattern, process, scale and synthesis. Annual Review of Ecology, Evolution and
Systematics, 34, 273-309.
WOLEJKO L., HERBICHOWA M., POTOCKA J. (2005) Typological differentiation and status of
Natura 2000 mire habitats in Poland. Stapfia 85, zugleich Kataloge der OÖ.
Landesmuseen. Neue Serie 35, 175-219.
ZOBEL M. (1997) The relative role of species pools in determining plant species richness:
an alternative explanation of species coexistence. Trends in Ecology and Evolution, 12,
266-269.
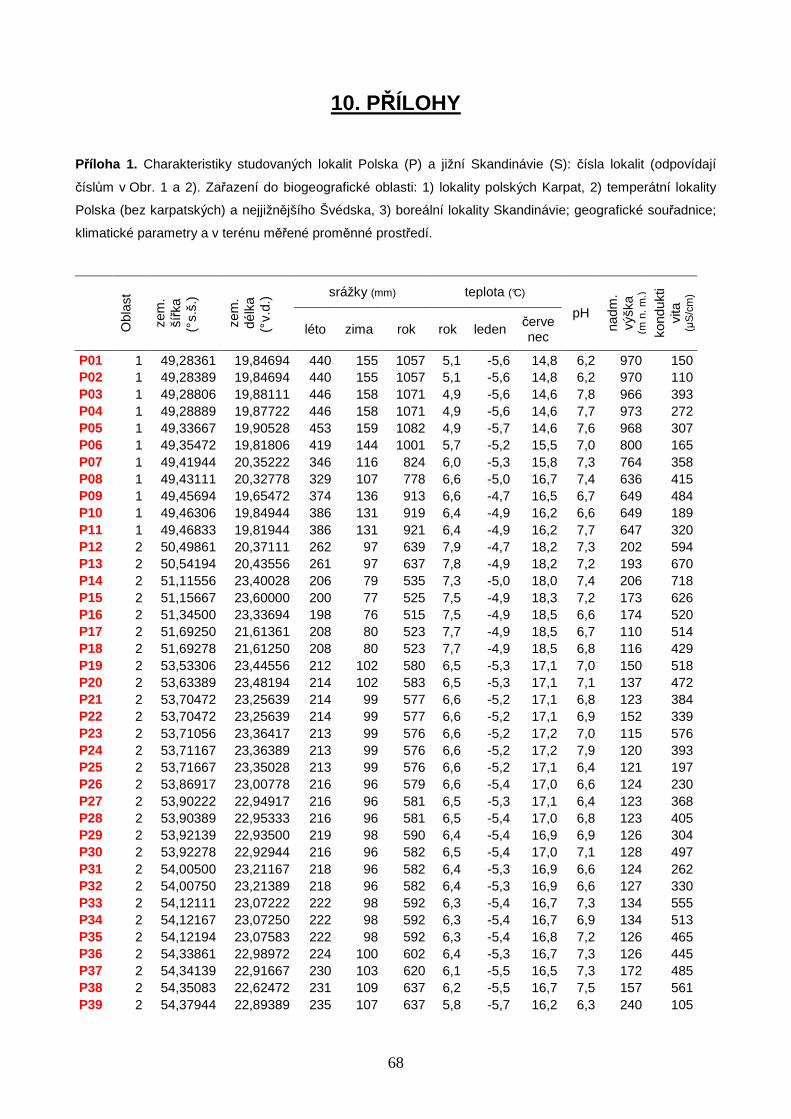
68
10. PŘÍLOHY
Příloha 1. Charakteristiky studovaných lokalit Polska (P) a jižní Skandinávie (S): čísla lokalit (odpovídají
číslům v Obr. 1 a 2). Zařazení do biogeografické oblasti: 1) lokality polských Karpat, 2) temperátní lokality
Polska (bez karpatských) a nejjižnějšího Švédska, 3) boreální lokality Skandinávie; geografické souřadnice;
klimatické parametry a v terénu měřené proměnné prostředí.
srážky (mm) teplota (°C)
Obl
ast
zem
. ší
řka
(°
s.š.
)
zem
. dé
lka
(°
v.d.
)
léto zima rok rok leden červenec
pH
nadm
. vý
ška
(m
n. m
.)
kond
ukti
vita
(µ
S/c
m)
P01 1 49,28361 19,84694 440 155 1057 5,1 -5,6 14,8 6,2 970 150 P02 1 49,28389 19,84694 440 155 1057 5,1 -5,6 14,8 6,2 970 110 P03 1 49,28806 19,88111 446 158 1071 4,9 -5,6 14,6 7,8 966 393 P04 1 49,28889 19,87722 446 158 1071 4,9 -5,6 14,6 7,7 973 272 P05 1 49,33667 19,90528 453 159 1082 4,9 -5,7 14,6 7,6 968 307 P06 1 49,35472 19,81806 419 144 1001 5,7 -5,2 15,5 7,0 800 165 P07 1 49,41944 20,35222 346 116 824 6,0 -5,3 15,8 7,3 764 358 P08 1 49,43111 20,32778 329 107 778 6,6 -5,0 16,7 7,4 636 415 P09 1 49,45694 19,65472 374 136 913 6,6 -4,7 16,5 6,7 649 484 P10 1 49,46306 19,84944 386 131 919 6,4 -4,9 16,2 6,6 649 189 P11 1 49,46833 19,81944 386 131 921 6,4 -4,9 16,2 7,7 647 320 P12 2 50,49861 20,37111 262 97 639 7,9 -4,7 18,2 7,3 202 594 P13 2 50,54194 20,43556 261 97 637 7,8 -4,9 18,2 7,2 193 670 P14 2 51,11556 23,40028 206 79 535 7,3 -5,0 18,0 7,4 206 718 P15 2 51,15667 23,60000 200 77 525 7,5 -4,9 18,3 7,2 173 626 P16 2 51,34500 23,33694 198 76 515 7,5 -4,9 18,5 6,6 174 520 P17 2 51,69250 21,61361 208 80 523 7,7 -4,9 18,5 6,7 110 514 P18 2 51,69278 21,61250 208 80 523 7,7 -4,9 18,5 6,8 116 429 P19 2 53,53306 23,44556 212 102 580 6,5 -5,3 17,1 7,0 150 518 P20 2 53,63389 23,48194 214 102 583 6,5 -5,3 17,1 7,1 137 472 P21 2 53,70472 23,25639 214 99 577 6,6 -5,2 17,1 6,8 123 384 P22 2 53,70472 23,25639 214 99 577 6,6 -5,2 17,1 6,9 152 339 P23 2 53,71056 23,36417 213 99 576 6,6 -5,2 17,2 7,0 115 576 P24 2 53,71167 23,36389 213 99 576 6,6 -5,2 17,2 7,9 120 393 P25 2 53,71667 23,35028 213 99 576 6,6 -5,2 17,1 6,4 121 197 P26 2 53,86917 23,00778 216 96 579 6,6 -5,4 17,0 6,6 124 230 P27 2 53,90222 22,94917 216 96 581 6,5 -5,3 17,1 6,4 123 368 P28 2 53,90389 22,95333 216 96 581 6,5 -5,4 17,0 6,8 123 405 P29 2 53,92139 22,93500 219 98 590 6,4 -5,4 16,9 6,9 126 304 P30 2 53,92278 22,92944 216 96 582 6,5 -5,4 17,0 7,1 128 497 P31 2 54,00500 23,21167 218 96 582 6,4 -5,3 16,9 6,6 124 262 P32 2 54,00750 23,21389 218 96 582 6,4 -5,3 16,9 6,6 127 330 P33 2 54,12111 23,07222 222 98 592 6,3 -5,4 16,7 7,3 134 555 P34 2 54,12167 23,07250 222 98 592 6,3 -5,4 16,7 6,9 134 513 P35 2 54,12194 23,07583 222 98 592 6,3 -5,4 16,8 7,2 126 465 P36 2 54,33861 22,98972 224 100 602 6,4 -5,3 16,7 7,3 126 445 P37 2 54,34139 22,91667 230 103 620 6,1 -5,5 16,5 7,3 172 485 P38 2 54,35083 22,62472 231 109 637 6,2 -5,5 16,7 7,5 157 561 P39 2 54,37944 22,89389 235 107 637 5,8 -5,7 16,2 6,3 240 105
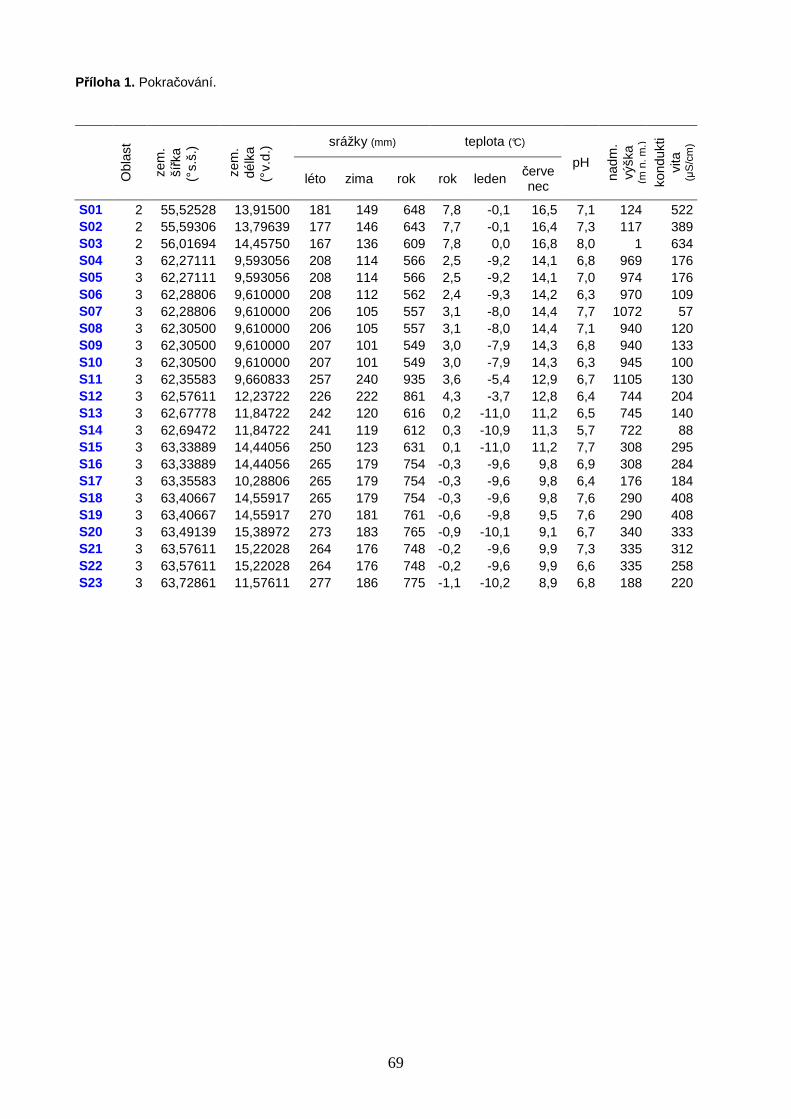
69
Příloha 1. Pokračování.
srážky (mm) teplota (°C)
Obl
ast
zem
. ší
řka
(°
s.š.
)
zem
. dé
lka
(°
v.d.
)
léto zima rok rok leden červenec
pH
nadm
. vý
ška
(m
n. m
.)
kond
ukti
vita
(µ
S/c
m)
S01 2 55,52528 13,91500 181 149 648 7,8 -0,1 16,5 7,1 124 522 S02 2 55,59306 13,79639 177 146 643 7,7 -0,1 16,4 7,3 117 389 S03 2 56,01694 14,45750 167 136 609 7,8 0,0 16,8 8,0 1 634 S04 3 62,27111 9,593056 208 114 566 2,5 -9,2 14,1 6,8 969 176 S05 3 62,27111 9,593056 208 114 566 2,5 -9,2 14,1 7,0 974 176 S06 3 62,28806 9,610000 208 112 562 2,4 -9,3 14,2 6,3 970 109 S07 3 62,28806 9,610000 206 105 557 3,1 -8,0 14,4 7,7 1072 57 S08 3 62,30500 9,610000 206 105 557 3,1 -8,0 14,4 7,1 940 120 S09 3 62,30500 9,610000 207 101 549 3,0 -7,9 14,3 6,8 940 133 S10 3 62,30500 9,610000 207 101 549 3,0 -7,9 14,3 6,3 945 100 S11 3 62,35583 9,660833 257 240 935 3,6 -5,4 12,9 6,7 1105 130 S12 3 62,57611 12,23722 226 222 861 4,3 -3,7 12,8 6,4 744 204 S13 3 62,67778 11,84722 242 120 616 0,2 -11,0 11,2 6,5 745 140 S14 3 62,69472 11,84722 241 119 612 0,3 -10,9 11,3 5,7 722 88 S15 3 63,33889 14,44056 250 123 631 0,1 -11,0 11,2 7,7 308 295 S16 3 63,33889 14,44056 265 179 754 -0,3 -9,6 9,8 6,9 308 284 S17 3 63,35583 10,28806 265 179 754 -0,3 -9,6 9,8 6,4 176 184 S18 3 63,40667 14,55917 265 179 754 -0,3 -9,6 9,8 7,6 290 408 S19 3 63,40667 14,55917 270 181 761 -0,6 -9,8 9,5 7,6 290 408 S20 3 63,49139 15,38972 273 183 765 -0,9 -10,1 9,1 6,7 340 333 S21 3 63,57611 15,22028 264 176 748 -0,2 -9,6 9,9 7,3 335 312 S22 3 63,57611 15,22028 264 176 748 -0,2 -9,6 9,9 6,6 335 258 S23 3 63,72861 11,57611 277 186 775 -1,1 -10,2 8,9 6,8 188 220

70
Příloha 2. Skript popisující modely závislosti počtu druhů živých suchozemských ulitnatých plžů na biomase
společenstev (mg) v programu R (černě) a souhrnné výstupy z analýz (modře): A) lineární regrese
přirozeným logaritmem transformované biomasy pro data z Polska a jižní Skandinávie, B) lineární regrese
s kvadratickým členem pro data z Polska a jižní Skandinávie, C) srovnání logaritmického a kvadratického
modelu pro data ze Skandinávie, D) srovnání logaritmického a kvadratického modelu pro data z Polska.
A)
m1<-lm(spTerLiv~log(prod)*reg); anova(m1)
m2<-lm(spTerLiv~log(prod)+reg); anova(m2)
anova(m1, m2)
plot(m2, which=1:4); pr<-resid(m2, type="pearson"); plot(prod, pr)
summary(m2)
Call:
lm(formula = spTerLiv ~ log(prod) + reg)
Residuals:
Min 1Q Median 3Q Max
-2.8873 -1.2301 -0.1242 1.2055 3.8541
Coefficients:
Estimate Std. Error t value Pr(>|t|)
(Intercept) 1.1237 0.9106 1.234 0.222
log(prod) 2.0877 0.2337 8.932 1.49e-12 ***
regScan -2.3211 0.4427 -5.243 2.23e-06 ***
---
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ’ 1
Residual standard error: 1.671 on 59 degrees of freedom
Multiple R-squared: 0.6717, Adjusted R-squared: 0.6606
F-statistic: 60.35 on 2 and 59 DF, p-value: 5.376e-15
B)
m1<-lm(spTerLiv~prod*reg+I(prod^2)*reg); anova(m1)
m2<-lm(spTerLiv~prod*reg+I(prod^2)); anova(m2)
anova(m1, m2)
m3<-lm(spTerLiv~prod+I(prod^2)+reg); anova(m3)
anova(m2, m3)
plot(m3, which=1:4); pr<-resid(m3, type="pearson"); plot(prod, pr)
summary(m3)
Call:
lm(formula = spTerLiv ~ prod + I(prod^2) + reg)
Residuals:
Min 1Q Median 3Q Max
-3.3366 -1.4639 -0.0536 1.3613 4.1451
Coefficients:

71
Estimate Std. Error t value Pr(>|t|)
(Intercept) 5.695e+00 5.417e-01 10.514 4.68e-15 ***
prod 7.700e-02 1.395e-02 5.521 8.28e-07 ***
I(prod^2) -2.232e-04 6.764e-05 -3.300 0.00166 **
regScan -2.380e+00 4.841e-01 -4.916 7.61e-06 ***
---
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ’ 1
Residual standard error: 1.817 on 58 degrees of freedom
Multiple R-squared: 0.6182, Adjusted R-squared: 0.5984
F-statistic: 31.3 on 3 and 58 DF, p-value: 3.662e-12
C)
summary(m3) # logaritmický model pro Skandinávii
Call:
lm(formula = spTerLiv ~ log(prod))
Residuals:
Min 1Q Median 3Q Max
-2.6584 -1.1787 -0.2606 0.5815 3.7115
Coefficients:
Estimate Std. Error t value Pr(>|t|)
(Intercept) 0.2220 1.4141 0.157 0.876775
log(prod) 1.6809 0.3932 4.274 0.000337 ***
---
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ’ 1
Residual standard error: 1.642 on 21 degrees of freedom
Multiple R-squared: 0.4653, Adjusted R-squared: 0.4398
F-statistic: 18.27 on 1 and 21 DF, p-value: 0.0003371
summary(m1) # kvadratický model pro Skandinávii
Call:
lm(formula = spTerLiv ~ prod + I(prod^2))
Residuals:
Min 1Q Median 3Q Max
-3.0763 -0.7682 -0.3222 0.6258 3.5672
Coefficients:
Estimate Std. Error t value Pr(>|t|)
(Intercept) 2.4584750 0.8820891 2.787 0.01138 *
prod 0.1371638 0.0380124 3.608 0.00175 **
I(prod^2) -0.0008001 0.0002988 -2.677 0.01447 *
---
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ’ 1
Residual standard error: 1.55 on 20 degrees of freedom
Multiple R-squared: 0.546, Adjusted R-squared: 0.5006

72
F-statistic: 12.03 on 2 and 20 DF, p-value: 0.0003717
AIC(m1, m3)
df AIC
m1 4 90.21213
m3 3 91.97892
D)
summary(m3) # logaritmický model pro Polsko
Call:
lm(formula = spTerLiv ~ log(prod))
Residuals:
Min 1Q Median 3Q Max
-3.1888 -1.0497 -0.2306 1.2253 3.5350
Coefficients:
Estimate Std. Error t value Pr(>|t|)
(Intercept) 0.3397 1.1075 0.307 0.761
log(prod) 2.2983 0.2886 7.964 1.53e-09 ***
---
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ’ 1
Residual standard error: 1.674 on 37 degrees of freedom
Multiple R-squared: 0.6316, Adjusted R-squared: 0.6216
F-statistic: 63.43 on 1 and 37 DF, p-value: 1.534e-09
summary(m2) # kvadratický model pro Polsko
Call:
lm(formula = spTerLiv ~ prod + I(prod^2))
Residuals:
Min 1Q Median 3Q Max
-3.6422 -1.6379 -0.1126 1.3756 3.6705
Coefficients:
Estimate Std. Error t value Pr(>|t|)
(Intercept) 5.410e+00 6.883e-01 7.859 2.53e-09 ***
prod 8.387e-02 1.831e-02 4.582 5.36e-05 ***
I(prod^2) -2.431e-04 8.274e-05 -2.939 0.00572 **
---
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ’ 1
Residual standard error: 1.902 on 36 degrees of freedom
Multiple R-squared: 0.5373, Adjusted R-squared: 0.5116
F-statistic: 20.9 on 2 and 36 DF, p-value: 9.438e-07
AIC(m2, m3)
df AIC
m2 4 165.717
m3 3 154.834
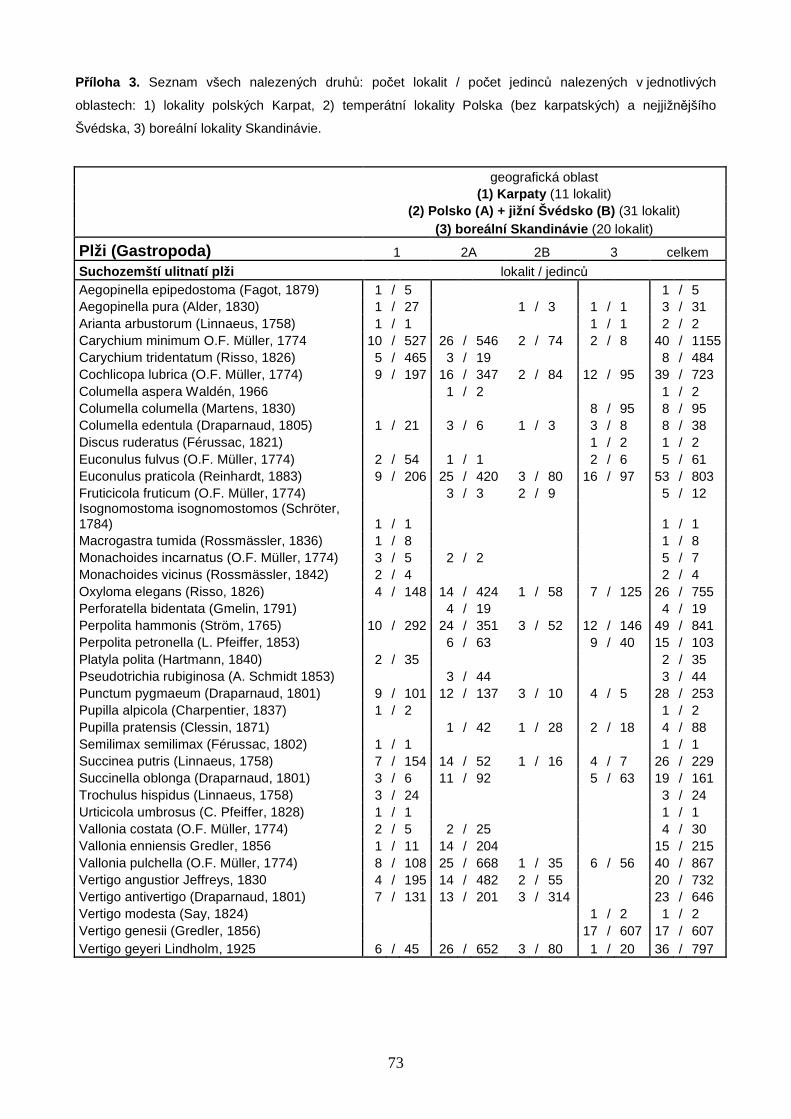
73
Příloha 3. Seznam všech nalezených druhů: počet lokalit / počet jedinců nalezených v jednotlivých
oblastech: 1) lokality polských Karpat, 2) temperátní lokality Polska (bez karpatských) a nejjižnějšího
Švédska, 3) boreální lokality Skandinávie.
geografická oblast (1) Karpaty (11 lokalit) (2) Polsko (A) + jižní Švédsko (B) (31 lokalit) (3) boreální Skandinávie (20 lokalit)
Plži (Gastropoda) 1 2A 2B 3 celkem
Suchozemští ulitnatí plži lokalit / jedinců Aegopinella epipedostoma (Fagot, 1879) 1 / 5 1 / 5 Aegopinella pura (Alder, 1830) 1 / 27 1 / 3 1 / 1 3 / 31 Arianta arbustorum (Linnaeus, 1758) 1 / 1 1 / 1 2 / 2 Carychium minimum O.F. Müller, 1774 10 / 527 26 / 546 2 / 74 2 / 8 40 / 1155 Carychium tridentatum (Risso, 1826) 5 / 465 3 / 19 8 / 484 Cochlicopa lubrica (O.F. Müller, 1774) 9 / 197 16 / 347 2 / 84 12 / 95 39 / 723 Columella aspera Waldén, 1966 1 / 2 1 / 2 Columella columella (Martens, 1830) 8 / 95 8 / 95 Columella edentula (Draparnaud, 1805) 1 / 21 3 / 6 1 / 3 3 / 8 8 / 38 Discus ruderatus (Férussac, 1821) 1 / 2 1 / 2 Euconulus fulvus (O.F. Müller, 1774) 2 / 54 1 / 1 2 / 6 5 / 61 Euconulus praticola (Reinhardt, 1883) 9 / 206 25 / 420 3 / 80 16 / 97 53 / 803 Fruticicola fruticum (O.F. Müller, 1774) 3 / 3 2 / 9 5 / 12 Isognomostoma isognomostomos (Schröter, 1784) 1 / 1 1 / 1 Macrogastra tumida (Rossmässler, 1836) 1 / 8 1 / 8 Monachoides incarnatus (O.F. Müller, 1774) 3 / 5 2 / 2 5 / 7 Monachoides vicinus (Rossmässler, 1842) 2 / 4 2 / 4 Oxyloma elegans (Risso, 1826) 4 / 148 14 / 424 1 / 58 7 / 125 26 / 755 Perforatella bidentata (Gmelin, 1791) 4 / 19 4 / 19 Perpolita hammonis (Ström, 1765) 10 / 292 24 / 351 3 / 52 12 / 146 49 / 841 Perpolita petronella (L. Pfeiffer, 1853) 6 / 63 9 / 40 15 / 103 Platyla polita (Hartmann, 1840) 2 / 35 2 / 35 Pseudotrichia rubiginosa (A. Schmidt 1853) 3 / 44 3 / 44 Punctum pygmaeum (Draparnaud, 1801) 9 / 101 12 / 137 3 / 10 4 / 5 28 / 253 Pupilla alpicola (Charpentier, 1837) 1 / 2 1 / 2 Pupilla pratensis (Clessin, 1871) 1 / 42 1 / 28 2 / 18 4 / 88 Semilimax semilimax (Férussac, 1802) 1 / 1 1 / 1 Succinea putris (Linnaeus, 1758) 7 / 154 14 / 52 1 / 16 4 / 7 26 / 229 Succinella oblonga (Draparnaud, 1801) 3 / 6 11 / 92 5 / 63 19 / 161 Trochulus hispidus (Linnaeus, 1758) 3 / 24 3 / 24 Urticicola umbrosus (C. Pfeiffer, 1828) 1 / 1 1 / 1 Vallonia costata (O.F. Müller, 1774) 2 / 5 2 / 25 4 / 30 Vallonia enniensis Gredler, 1856 1 / 11 14 / 204 15 / 215 Vallonia pulchella (O.F. Müller, 1774) 8 / 108 25 / 668 1 / 35 6 / 56 40 / 867 Vertigo angustior Jeffreys, 1830 4 / 195 14 / 482 2 / 55 20 / 732 Vertigo antivertigo (Draparnaud, 1801) 7 / 131 13 / 201 3 / 314 23 / 646 Vertigo modesta (Say, 1824) 1 / 2 1 / 2 Vertigo genesii (Gredler, 1856) 17 / 607 17 / 607 Vertigo geyeri Lindholm, 1925 6 / 45 26 / 652 3 / 80 1 / 20 36 / 797
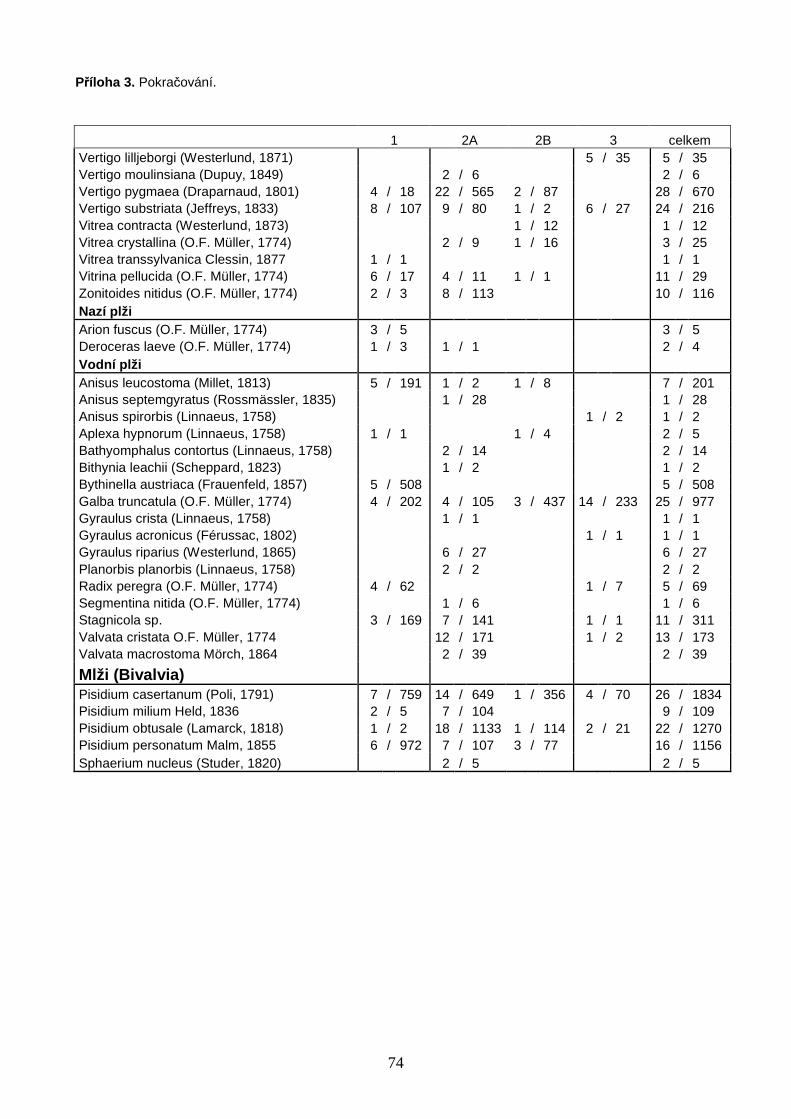
74
Příloha 3. Pokračování.
1 2A 2B 3 celkem Vertigo lilljeborgi (Westerlund, 1871) 5 / 35 5 / 35 Vertigo moulinsiana (Dupuy, 1849) 2 / 6 2 / 6 Vertigo pygmaea (Draparnaud, 1801) 4 / 18 22 / 565 2 / 87 28 / 670 Vertigo substriata (Jeffreys, 1833) 8 / 107 9 / 80 1 / 2 6 / 27 24 / 216 Vitrea contracta (Westerlund, 1873) 1 / 12 1 / 12 Vitrea crystallina (O.F. Müller, 1774) 2 / 9 1 / 16 3 / 25 Vitrea transsylvanica Clessin, 1877 1 / 1 1 / 1 Vitrina pellucida (O.F. Müller, 1774) 6 / 17 4 / 11 1 / 1 11 / 29 Zonitoides nitidus (O.F. Müller, 1774) 2 / 3 8 / 113 10 / 116 Nazí plži Arion fuscus (O.F. Müller, 1774) 3 / 5 3 / 5 Deroceras laeve (O.F. Müller, 1774) 1 / 3 1 / 1 2 / 4 Vodní plži Anisus leucostoma (Millet, 1813) 5 / 191 1 / 2 1 / 8 7 / 201 Anisus septemgyratus (Rossmässler, 1835) 1 / 28 1 / 28 Anisus spirorbis (Linnaeus, 1758) 1 / 2 1 / 2 Aplexa hypnorum (Linnaeus, 1758) 1 / 1 1 / 4 2 / 5 Bathyomphalus contortus (Linnaeus, 1758) 2 / 14 2 / 14 Bithynia leachii (Scheppard, 1823) 1 / 2 1 / 2 Bythinella austriaca (Frauenfeld, 1857) 5 / 508 5 / 508 Galba truncatula (O.F. Müller, 1774) 4 / 202 4 / 105 3 / 437 14 / 233 25 / 977 Gyraulus crista (Linnaeus, 1758) 1 / 1 1 / 1 Gyraulus acronicus (Férussac, 1802) 1 / 1 1 / 1 Gyraulus riparius (Westerlund, 1865) 6 / 27 6 / 27 Planorbis planorbis (Linnaeus, 1758) 2 / 2 2 / 2 Radix peregra (O.F. Müller, 1774) 4 / 62 1 / 7 5 / 69 Segmentina nitida (O.F. Müller, 1774) 1 / 6 1 / 6 Stagnicola sp. 3 / 169 7 / 141 1 / 1 11 / 311 Valvata cristata O.F. Müller, 1774 12 / 171 1 / 2 13 / 173 Valvata macrostoma Mörch, 1864 2 / 39 2 / 39
Mlži (Bivalvia) Pisidium casertanum (Poli, 1791) 7 / 759 14 / 649 1 / 356 4 / 70 26 / 1834 Pisidium milium Held, 1836 2 / 5 7 / 104 9 / 109 Pisidium obtusale (Lamarck, 1818) 1 / 2 18 / 1133 1 / 114 2 / 21 22 / 1270 Pisidium personatum Malm, 1855 6 / 972 7 / 107 3 / 77 16 / 1156 Sphaerium nucleus (Studer, 1820) 2 / 5 2 / 5
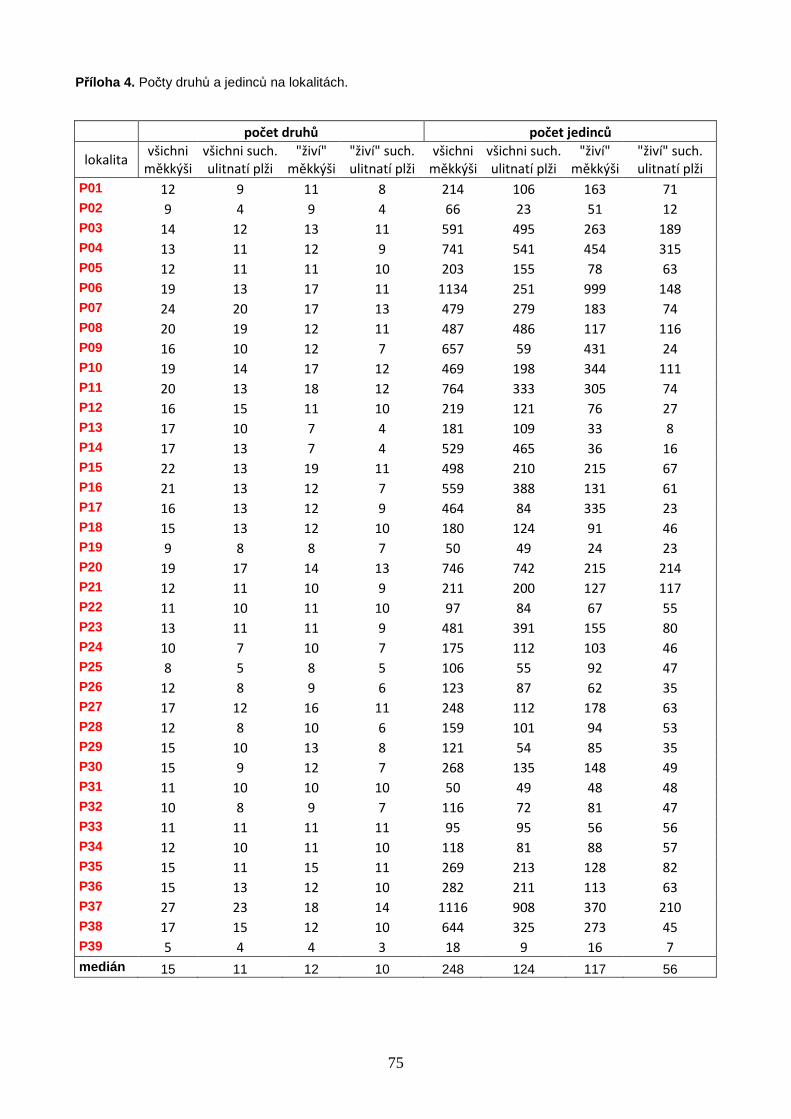
75
Příloha 4. Počty druhů a jedinců na lokalitách.
počet druhů počet jedinců
lokalita všichni
měkkýši
všichni such.
ulitnatí plži
"živí"
měkkýši
"živí" such.
ulitnatí plži
všichni
měkkýši
všichni such.
ulitnatí plži
"živí"
měkkýši
"živí" such.
ulitnatí plži
P01 12 9 11 8 214 106 163 71
P02 9 4 9 4 66 23 51 12
P03 14 12 13 11 591 495 263 189
P04 13 11 12 9 741 541 454 315
P05 12 11 11 10 203 155 78 63
P06 19 13 17 11 1134 251 999 148
P07 24 20 17 13 479 279 183 74
P08 20 19 12 11 487 486 117 116
P09 16 10 12 7 657 59 431 24
P10 19 14 17 12 469 198 344 111
P11 20 13 18 12 764 333 305 74
P12 16 15 11 10 219 121 76 27
P13 17 10 7 4 181 109 33 8
P14 17 13 7 4 529 465 36 16
P15 22 13 19 11 498 210 215 67
P16 21 13 12 7 559 388 131 61
P17 16 13 12 9 464 84 335 23
P18 15 13 12 10 180 124 91 46
P19 9 8 8 7 50 49 24 23
P20 19 17 14 13 746 742 215 214
P21 12 11 10 9 211 200 127 117
P22 11 10 11 10 97 84 67 55
P23 13 11 11 9 481 391 155 80
P24 10 7 10 7 175 112 103 46
P25 8 5 8 5 106 55 92 47
P26 12 8 9 6 123 87 62 35
P27 17 12 16 11 248 112 178 63
P28 12 8 10 6 159 101 94 53
P29 15 10 13 8 121 54 85 35
P30 15 9 12 7 268 135 148 49
P31 11 10 10 10 50 49 48 48
P32 10 8 9 7 116 72 81 47
P33 11 11 11 11 95 95 56 56
P34 12 10 11 10 118 81 88 57
P35 15 11 15 11 269 213 128 82
P36 15 13 12 10 282 211 113 63
P37 27 23 18 14 1116 908 370 210
P38 17 15 12 10 644 325 273 45
P39 5 4 4 3 18 9 16 7
medián 15 11 12 10 248 124 117 56
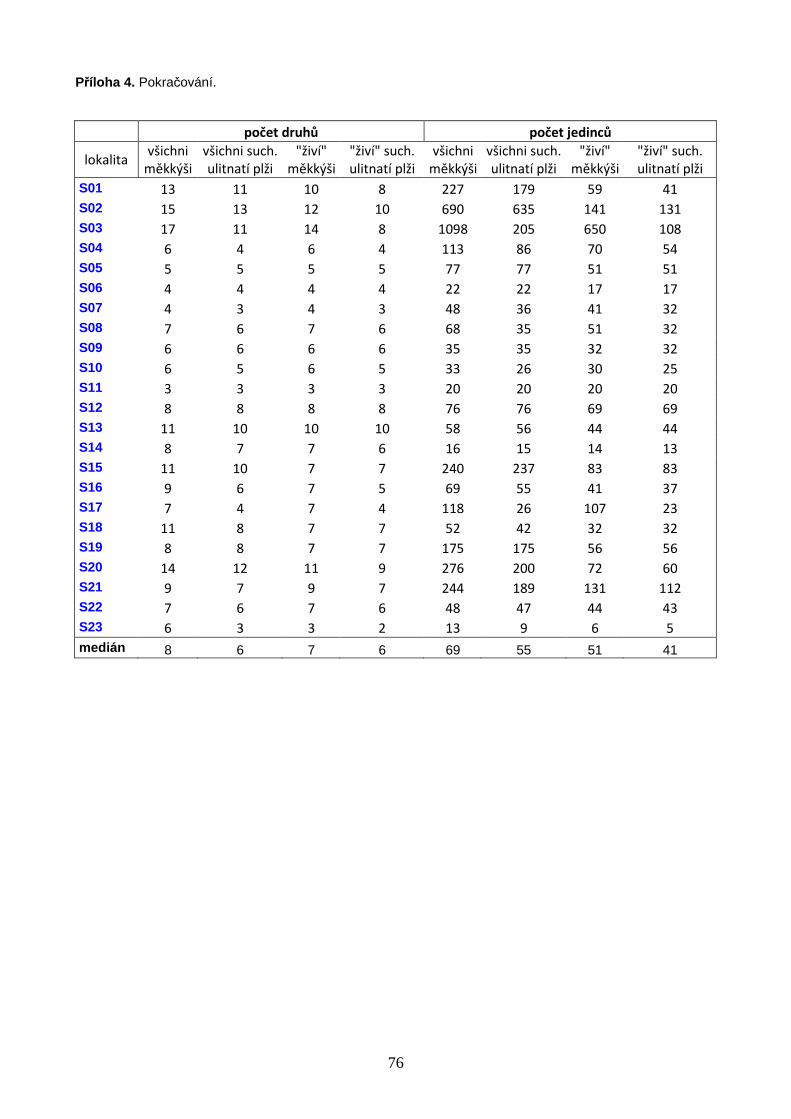
76
Příloha 4. Pokračování.
počet druhů počet jedinců
lokalita všichni
měkkýši
všichni such.
ulitnatí plži
"živí"
měkkýši
"živí" such.
ulitnatí plži
všichni
měkkýši
všichni such.
ulitnatí plži
"živí"
měkkýši
"živí" such.
ulitnatí plži
S01 13 11 10 8 227 179 59 41
S02 15 13 12 10 690 635 141 131
S03 17 11 14 8 1098 205 650 108
S04 6 4 6 4 113 86 70 54
S05 5 5 5 5 77 77 51 51
S06 4 4 4 4 22 22 17 17
S07 4 3 4 3 48 36 41 32
S08 7 6 7 6 68 35 51 32
S09 6 6 6 6 35 35 32 32
S10 6 5 6 5 33 26 30 25
S11 3 3 3 3 20 20 20 20
S12 8 8 8 8 76 76 69 69
S13 11 10 10 10 58 56 44 44
S14 8 7 7 6 16 15 14 13
S15 11 10 7 7 240 237 83 83
S16 9 6 7 5 69 55 41 37
S17 7 4 7 4 118 26 107 23
S18 11 8 7 7 52 42 32 32
S19 8 8 7 7 175 175 56 56
S20 14 12 11 9 276 200 72 60
S21 9 7 9 7 244 189 131 112
S22 7 6 7 6 48 47 44 43
S23 6 3 3 2 13 9 6 5
medián 8 6 7 6 69 55 51 41
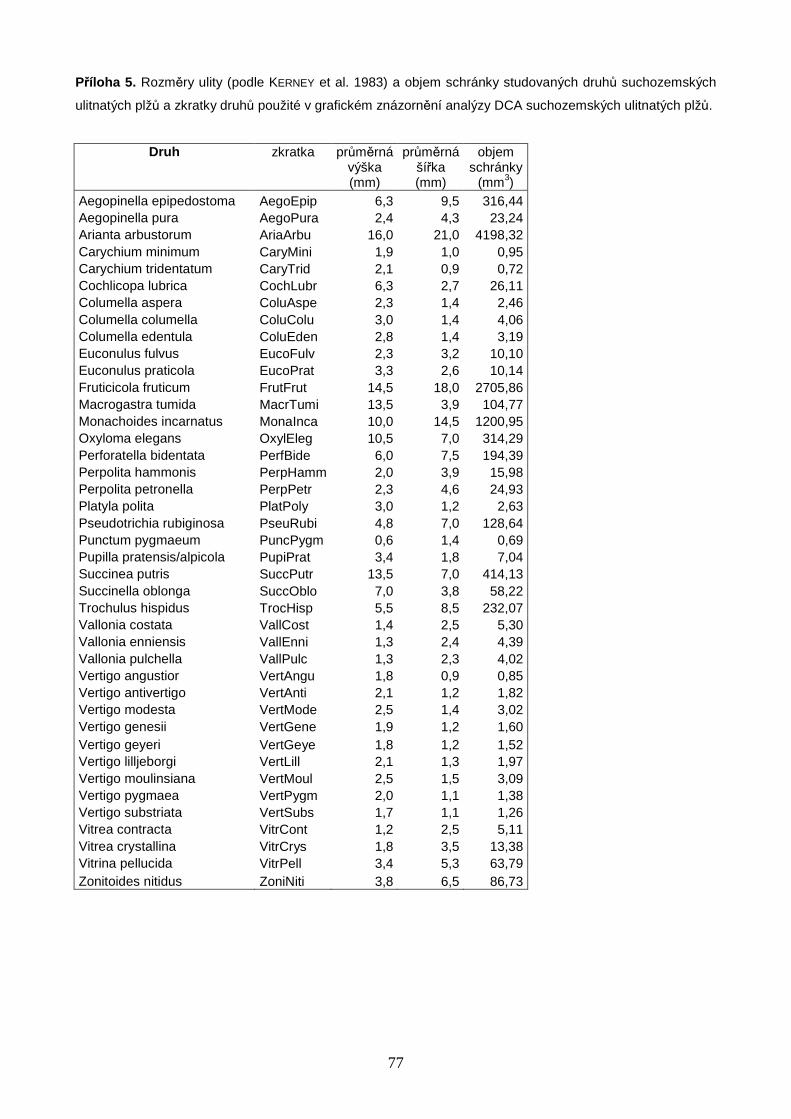
77
Příloha 5. Rozměry ulity (podle KERNEY et al. 1983) a objem schránky studovaných druhů suchozemských
ulitnatých plžů a zkratky druhů použité v grafickém znázornění analýzy DCA suchozemských ulitnatých plžů.
Druh zkratka průměrná výška (mm)
průměrná šířka (mm)
objem schránky
(mm3) Aegopinella epipedostoma AegoEpip 6,3 9,5 316,44 Aegopinella pura AegoPura 2,4 4,3 23,24 Arianta arbustorum AriaArbu 16,0 21,0 4198,32 Carychium minimum CaryMini 1,9 1,0 0,95 Carychium tridentatum CaryTrid 2,1 0,9 0,72 Cochlicopa lubrica CochLubr 6,3 2,7 26,11 Columella aspera ColuAspe 2,3 1,4 2,46 Columella columella ColuColu 3,0 1,4 4,06 Columella edentula ColuEden 2,8 1,4 3,19 Euconulus fulvus EucoFulv 2,3 3,2 10,10 Euconulus praticola EucoPrat 3,3 2,6 10,14 Fruticicola fruticum FrutFrut 14,5 18,0 2705,86 Macrogastra tumida MacrTumi 13,5 3,9 104,77 Monachoides incarnatus MonaInca 10,0 14,5 1200,95 Oxyloma elegans OxylEleg 10,5 7,0 314,29 Perforatella bidentata PerfBide 6,0 7,5 194,39 Perpolita hammonis PerpHamm 2,0 3,9 15,98 Perpolita petronella PerpPetr 2,3 4,6 24,93 Platyla polita PlatPoly 3,0 1,2 2,63 Pseudotrichia rubiginosa PseuRubi 4,8 7,0 128,64 Punctum pygmaeum PuncPygm 0,6 1,4 0,69 Pupilla pratensis/alpicola PupiPrat 3,4 1,8 7,04 Succinea putris SuccPutr 13,5 7,0 414,13 Succinella oblonga SuccOblo 7,0 3,8 58,22 Trochulus hispidus TrocHisp 5,5 8,5 232,07 Vallonia costata VallCost 1,4 2,5 5,30 Vallonia enniensis VallEnni 1,3 2,4 4,39 Vallonia pulchella VallPulc 1,3 2,3 4,02 Vertigo angustior VertAngu 1,8 0,9 0,85 Vertigo antivertigo VertAnti 2,1 1,2 1,82 Vertigo modesta VertMode 2,5 1,4 3,02 Vertigo genesii VertGene 1,9 1,2 1,60 Vertigo geyeri VertGeye 1,8 1,2 1,52 Vertigo lilljeborgi VertLill 2,1 1,3 1,97 Vertigo moulinsiana VertMoul 2,5 1,5 3,09 Vertigo pygmaea VertPygm 2,0 1,1 1,38 Vertigo substriata VertSubs 1,7 1,1 1,26 Vitrea contracta VitrCont 1,2 2,5 5,11 Vitrea crystallina VitrCrys 1,8 3,5 13,38 Vitrina pellucida VitrPell 3,4 5,3 63,79 Zonitoides nitidus ZoniNiti 3,8 6,5 86,73





























