Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Aug. 1989, p. 2036-2040 Vol. 55, No. 80099-2240/89/082036-05$02.00/0Copyright C) 1989, American Society for Microbiology
Mechanism of Resistance to Benzalkonium Chloride byPseudomonas aeruginosa
YOSHIKAZU SAKAGAMI,l* HIROSHI YOKOYAMA,1 HIROSHI NISHIMURA,'YOUKI OSE, AND TOSHIO TASHIMA3
Osaka Prefectural Institute of Public Health, 1-3-69 Nakamichi, Higashinari-ku, Osaka 537,1 Gifu PharmaceuticalUniversity, 5-6-1 Mitahora-higashi, Gifai 502,2 and Central Laboratory of Maruishi Pharmaceutical Co., Ltd.,
2-2-18 Imazia-naka, Tsurumi-ku, Osaka 535,3 JapanReceived 21 November 1988/Accepted 23 March 1989
The mechanisms of resistance of Pseudomonas aeruginosa to benzalkonium chloride (BC) were studied. Theeffluence of cell components was observed in susceptible P. aeruginosa by electron microscopy, but resistant P.aeruginosa seemed to be undamaged. No marked changes in cell surface potential between Escherichia coliNIHJC-2 and a spheroplast strain were found. The contents of phospholipids (PL) and fatty and neutral lipids(FNL) in the cell walls of resistant P. aeruginosa were higher than those in the cell walls of susceptible P.aeruginosa. The amounts of BC adsorbed to PL and FNL of cell walls of BC-resistant P. aeruginosa were lowerthan those for BC-susceptible P. aeruginosa. Fifteen species of cellular fatty acids were identified by capillarygas chromatography and gas chromatography-mass spectrometry. The ability of BC to permeate the cell wallwas reduced because of the increase in cellular fatty acids. These results suggested that the resistance of P.aeruginosa to BC is mainly a result of increases in the contents of PL and FNL. In resistant P. aeruginosa, thedecrease in the amount of BC adsorbed is likely to be the result of increases in the contents of PL and FNL.
There are many reports on the mechanisms of bacterialresistance to antibiotics (5, 6, 12, 13, 21, 24, 30, 32, 34,36-38), but only a few reports on the mechanisms of bacte-rial resistance to disinfectants have been presented. Furtherdetailed studies seem to be necessary for clarifying the lattermechanisms of resistance.
Richards and Cavill (26) investigated changes of cell shapecaused by 800 pg of benzalkonium chloride (BC) per ml inPseudomonas aeruginosa by electron microscopy. Theyreported that BC stripped the outer cell membrane, that cellsretained their shape after the outer membrane was removed,and that the peptidoglycan layer remained intact.
In 1986 Russell (27) reviewed the possible two majormechanisms of bacterial resistance to antibacterial agentssuch as antibiotics. The first is acquired resistance, and thesecond is intrinsic resistance. As acquired resistance, Ten-nent et al. (35) reported plasmid-encoded resistance toquarternary ammonium salts in gram-positive bacteria suchas Staphylococcus aureus. However, in gram-negative rod-shaped bacteria no such resistance was found. Resistance tohydrophilic and hydrophobic chemicals is an example ofintrinsic resistance (17-19, 23). Hydrophilic chemicals of lowmolecular weight (below about 650) are able to enter bacte-rial cells via porins. In P. aeruginosa the porins appear to belarger, despite the fact that this organism is frequently highlyresistant to disinfectants. P. aeruginosa may possess fewerporins, and many may exist in a nonfunctional state. Thesusceptibility of P. aeruginosa to hydrophilic chemicals suchas phenols and dyes is reduced, and the cause has beenlinked to outer membrane lipopolysaccharide. Susceptibilityand resistance are related to the appearance of phospholipids(PL) in the outer layer of the outer membrane.
Chaplin (7) reported that the lipoprotein content on thecell surface was increased significantly in BC-resistant Ser-ratia marcescens, and Nishikawa and Yamamoto (20) re-ported an increase in lipid content in BC-resistant Entero-
* Corresponding author.
bacter cloacae. In some studies the relationship betweenresistance to disinfectants and the fatty acids in the cellcomponents was addressed. Shigeta et al. (31) reported that3-hydroxytetradecanoic acid levels were increased in resis-tant Achromobac ter xylosoxidans, P. cepacia, and A. faeca-lis, but Ohta and Zenda (S. Ohta and H. Zenda, 102nd Annu.Meet. Jpn. Pharm. Sci., p. 751, 1982) reported that 3-hydroxytetradecanoic acid levels were not increased signif-icantly in resistant A. xylosoxidans. Uyeda et al. (39) iden-tified six kinds of fatty acids in S. marcescens, but the fattyacid contents in the resistant cells were nearly the same asthose in the susceptible cells. Recently, Russell et al. (27, 28)suggested that in gram-negative rod-shaped bacteria the cellmembrane acts as a barrier to disinfectants and preserva-tives.
In this study, we examined the mechanisms of resistanceto BC of strains of susceptible and resistant P. aeruginosaand of Escherichia coli and a spheroplast strain in variousexperiments.
MATERIALS AND METHODS
Disinfectants. Twenty samples of commercial 10% BC(Nihon Pharmaceutical Co., Ltd. and other sources) wereobtained in pharmacies.
Fatty acids. Lauric acid, 2-hydroxydodecanoic acid,myristic acid, palmitoleic acid, palmitic acid, oleic acid, andstearic acid were used in this study. They were purchasedfrom Funakoshi Co., Ltd.
Bacterial strains. (i) P. aeruginosa. Three groups withdifferent susceptibilities to disinfectants were used. Resis-tant group A included three strains of P. aeruginosa isolatedfrom activated sludge and acclimatized to a mixture ofglutaraldehyde, BC, and chlorhexidine digluconate. Resis-tant group B included three strains of BC-resistant P.aeruiginosa acclimatized to BC at 800 p.g/ml. Susceptiblegroup S included two strains of P. aeruginosa as well asstandard P. aeruiginosa IFO 13275; the two strains were
2036
on January 15, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

P. AERUGINOSA RESISTANCE TO BENZALKONIUM CHtLORIDE
isolated from activated sludge, and the standard strain waspurchased from the Institute for Fermentation of Osaka.
(ii) E. coli. E. coli NIHJC-2 and E. coli ES-114 (sphero-plast strain) were kindly donated by Fujisawa Pharmaceuti-cal Co., Ltd.MIC. The MIC of BC was measured by a standard method
(9).Measurement of cell surface potential. BC at 1, 10, 25, or 50
[Lg/ml was added to a test tube containing 10 ml of anovernight culture and incubated at 25°C for 5 min. Sterilizedphysiological saline (40 ml) was added and mixed. The cellsurface potential of the tested bacteria in the mixture wasmeasured by using a model 501 Lazer Zee Meter (Pen KemCo., Ltd.).
Morphological investigation by electron microscopy. P.(leruginosa strains (resistant and susceptible) were treatedwith 200, 400, and 800 pg/ml of BC. After fixation with 2%OS04 solution, shadowing with palladium platinum wasperformed.PL and FNL. The method of Broxton et al. (4) was used to
determine lipid content. Freeze-dried cells (500 mg) wereplaced into a preweighed flask and extracted twice withchloroform-methanol (2:1). The extract was dried underdiminished pressure in a desiccator containing phosphoruspentoxide overnight. After the residue was extracted with 30ml of diethyl ether, 30 ml of acetone was added and shakenat -10°C. The acetone layer was evaporated under dimin-ished pressure, and the contents were measured as PL (4).After extraction with diethyl ether the contents of theresidue in the flask were measured as fatty and neutral lipids(FNL) (4).
Adsorption of BC to cell wall lipids. BC (5 ml) at 100 VLg/mlwas added to the flasks containing PL or FNL and shakenfor 60 min. BC adsorbed to PL or FNL was calculated asfollows: adsorption contents of BC (pLg/mg) = 5[100 ,ug - BCconcentration (>1g/ml) in the supernatant]/PL or FNL (mg) inthe flask.
Fatty acids in the tested strains. Fatty acids were deter-mined as described by Moss and Lewis (15). Freeze-driedcells of bacteria (100 mg) were saponified with 10 ml of 15%KOH-50% methanol, extracted with 30 ml of ether-hexane(1:1) under acidic conditions, and evaporated. The residuewas methylated with saturated diazomethane solution (0.5ml) and injected into a gas chromatograph and a gas chro-matograph-mass spectrometer equipped with a flame ioniza-tion detector. The peaks were identified. Identification levelA represents identifications confirmed by comparison withstandard fatty acid methyl esters. Identification level B isbased on mass data books. The fatty acid contents weremeasured from the calibration curve. For the determinationof contents of nine kinds of fatty acids for which standardscould not be purchased, calibration curves for the mostsimilar fatty acids were used. The analytical conditions forgas chromatography were as follows: apparatus, Hewlett-Packard 5890A gas chromatograph; column, 5% phenylme-thyl silicone; column temperature, 150 to 250°C; detectoroven temperature, 270°C; injection port temperature, 250°C;rate, 5.0°C/min; and line speed, 27 cm/s. The analyticalconditions for gas chromatography-mass spectrometry wereas follows: apparatus, JEOL DX-303 mass spectrometerequipped with a DA 5000 data system analyzer; chambertemperature, 300°C; ionic current, 300 MA; ionic voltage, 70eV; pressure voltage, 3 kV; mass scan range, 40 MZ; andcyclic scan range, 0.5 s.Measurement of partition coefficients with an n-octanol-
water system. The partition coefficients (log P) of fatty acids
and BC in n-octanol-water were measured by followingslightly modified OECD guidelines (G,iideliniesfji Testilig ofClemicals, p. 107, OECD, 1982). ni-Octanol-water (20 ml)was added to a centrifugation tube containing a test sample(5 or 10 mg), and the mixture was shaken for 1 h at a slowspeed at 20°C. After centrifugation (2,000 rpm, 15 min), themixture was separated into water and n-octanol layers.The concentrations of fatty acids were measured by cap-
illary gas chromatography as described above. The log Pvalues of fatty acids for which standards could not bepurchased were calculated by the Tr method (10). BC wasmeasured by a colorimetric method with bromphenol blue(1).
RESULTS
MIC. The MICs for group A and group B were two andeight times high, respectively, than those for group S (400,1,600, and 200 [Lg/ml, respectively).
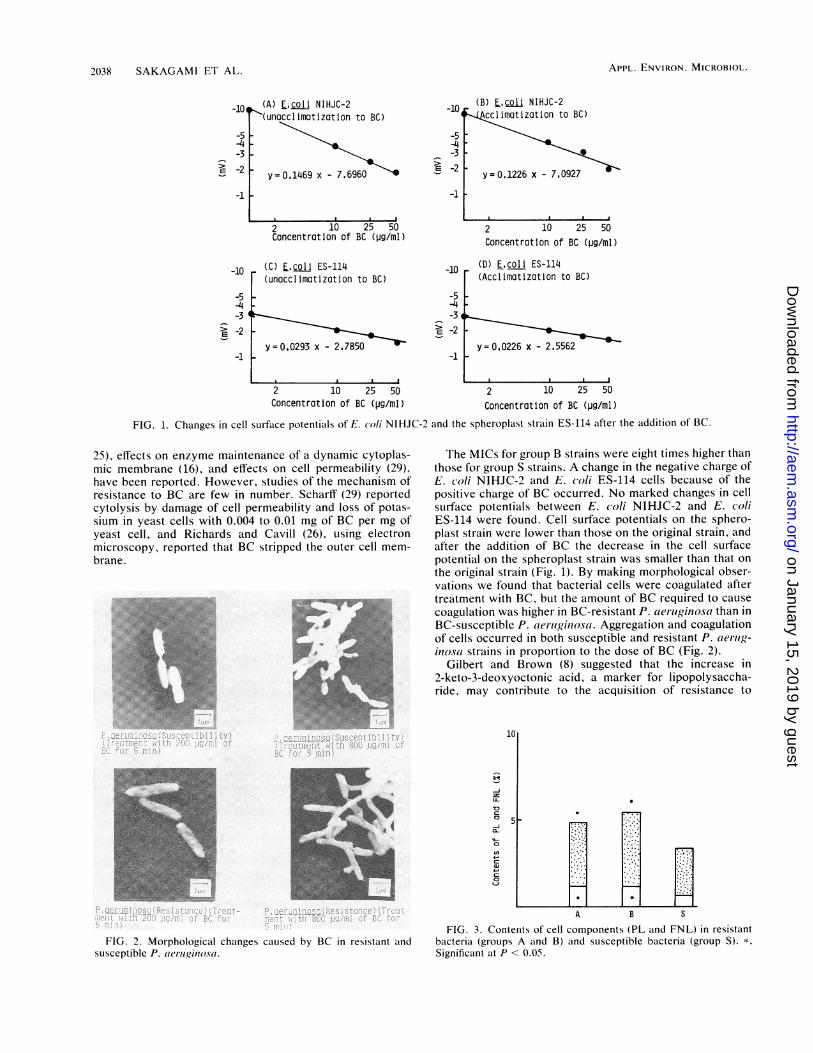
Cell surface potentials. A difference in cell surface poten-tials tested with BC was not recognized between nonaccli-matized and acclimatized strains. However, the difference inpotentials between the original E. coli strain and the sphero-plast strain ES-114 was significant (Fig. 1).
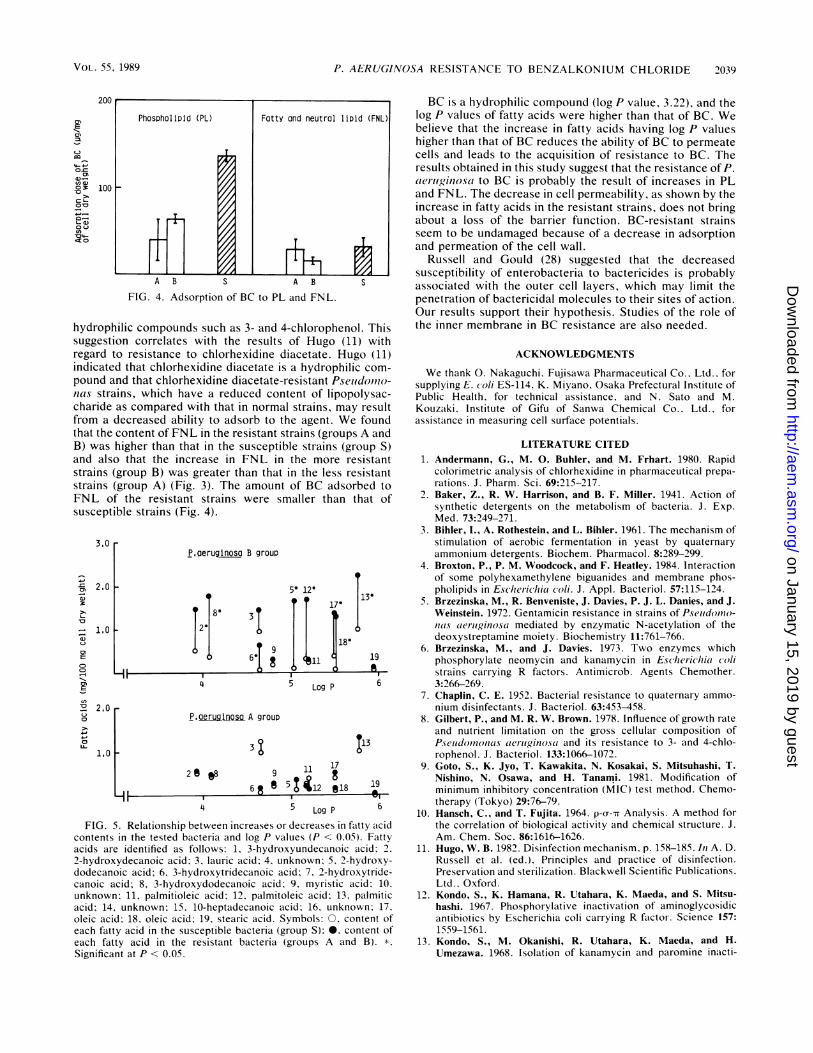
Morphological changes. After treatment with 800 pLg of BCper ml, aggregation and coagulation of cells and removal ofthe cell membrane were recognized in both resistant andsusceptible P. ae-iruginosa strains (Fig. 2).
Cell wall components. The statistical differences in thecontents of PL and FNL among the groups were evaluatedby the t test (P < 0.01 and P < 0.05). The content of FNL ingroup S strains was lower than that in group B strains (P <0.01), and the increase in FNL content in group B strainswas greater than that in group A strains (P < 0.01) (Fig. 3).
Adsorption of BC to cell wall components. The amounts ofBC adsorbed to PL and FNL of resistant group A and Bstrains were lower than those of susceptible group S strains(Fig. 4).
Fatty acids in the tested bacteria. Nineteen kinds of fattyacids were detected in the tested bacteria. They includedfour kinds of linear saturated fatty acids (lauric acid, myristicacid, palmitic acid, and stearic acid), four kinds of linearunsaturated fatty acids (two each of isomers of palmitoleicacid and oleic acid), seven kinds of saturated hydroxy fattyacids (3-hydroxyundecanoic acid, 2-hydroxydecanoic acid,2-hydroxydodecanoic acid, and 10-heptadecanoic acid), andfour kinds of unknown fatty acids.
Partition coefficients in n-octanol-water. The log P value ofBC was 3.22. BC was a hydrophilic compound.
Correlation between the fatty acids in the cell wall compo-nents and their partition coefficients. The significance of theincreases or decreases in fatty acid contents in the cell wallcomponents was evaluated by the t test. No significantincreases in fatty acid contents in the group A strains werefound. In group B strains the contents of 2-hydroxydecanoicacid (peak 2), 2-hydroxydodecanoic acid (peak 5), 3-hydroxy-tridecanoic acid (peak 6), 3-hydroxydodecanoic acid (peak8), palmitoleic acid (peak 12). palmitic acid (peak 13). andoleic acids (peaks 17 and 18) were increased significantly(Fig. 5).
DISCUSSION
Many types of disinfection mechanisms, such as effects onmetabolic reactions with glucose in a variety of bacteria (2,3, 22, 33), protein denaturation and enzyme inactivation (14,
VOL. 55, 1989 2037
on January 15, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
2038 SAKAGAMI ET AL.
-104
E
-5-4-3-2
-1
(A) E.coli NIHJC-2(unacclimatization to BC)
y=0.1469 x - 7.6960
APPL. ENVIRON. MICROBIOL.
-10o
E
-5-4-3-2
-1
2 10 25 50Concentration of BC (pg/ml)
(B) E.coli NIHJC-2ccl imtization to BC)
Y= 0.1226 x - 7.0927
a I
2 10 25 50Concentration of BC (pg/ml)
(C) E.coli ES-114r (unacclimatization to BC)
y=0.023x-275
-10 (D) E.coli ES-114r (Acclimatization to BC)
E
-5-4-3-2
-1
2 10 25 50Concentration of BC (pg/ml)
y=0.0226 x 256
2 10 25 50
Concentration of BC (pg/ml)
FIG. 1. Changes in cell surface potentials of E. coli NIHJC-2 and the spheroplast strain ES-114 after the addition of BC.
25), effects on enzyme maintenance of a dynamic cytoplas-mic membrane (16), and effects on cell permeability (29),have been reported. However, studies of the mechanism ofresistance to BC are few in number. Scharff (29) reportedcytolysis by damage of cell permeability and loss of potas-sium in yeast cells with 0.004 to 0.01 mg of BC per mg ofyeast cell, and Richards and Cavill (26), using electronmicroscopy, reported that BC stripped the outer cell mem-
brane.
The MICs for group B strains were eight times higher thanthose for group S strains. A change in the negative charge ofE. coli NIHJC-2 and E. coli ES-114 cells because of thepositive charge of BC occurred. No marked changes in cellsurface potentials between E. coli NIHJC-2 and E. coliES-114 were found. Cell surface potentials on the sphero-plast strain were lower than those on the original strain, andafter the addition of BC the decrease in the cell surfacepotential on the spheroplast strain was smaller than that on
the original strain (Fig. 1). By making morphological obser-vations we found that bacterial cells were coagulated aftertreatment with BC, but the amount of BC required to cause
coagulation was higher in BC-resistant P. aerulginosa than inBC-susceptible P. aer-i/giniosa. Aggregation and coagulationof cells occurred in both susceptible and resistant P. aeruig-
inos(l strains in proportion to the dose of BC (Fig. 2).Gilbert and Brown (8) suggested that the increase in
2-keto-3-deoxyoctonic acid, a marker for lipopolysaccha-ride, may contribute to the acquisition of resistance to~~~~~~~~~ Jte I
P.QerLo inosa(Susceptibility)(Treotment with 200 pg/ml ofBC for 5 min)
P,aeruginoso(SuscePtiblity)(Treatment with 800 pg/ml ofBC for 5 min)
P cEq inosao(Resistance)(Treat- P. gerusin (Resistance)(Treot-,,,ent wl~ti 0 ug/ml of BC for ment with O0 p,g/mi of BC for5 min) 5 mlTli)FIG. 2. Morphological changes caused by BC in resistant and
susceptible P. aeriuginosa.
A B S
FIG. 3. Contents of cell components (PL and FNL) in resistantbacteria (groups A aind B) and susceptible bacteria (group S). *,
Significant at P < 0.05.
-10
-5-4-3-2E
-1
.A-
-i2=LL.
'OC=0
-iCl-
4-0
(n41C=(L)4 -
c0L.)
on January 15, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
P. AERUGINOSA RESISTANCE TO BENZALKONIUM CHLORIDE 2039
200
Phospholipid (PL) Fatty and neutral liPid (FNL)
Mr 100'o
A B S A B S
FIG. 4. Adsorption of BC to PL and FNL.
hydrophilic compounds such as 3- and 4-chlorophenol. Thissuggestion correlates with the results of Hugo (11) withregard to resistance to chlorhexidine diacetate. Hugo (11)indicated that chlorhexidine diacetate is a hydrophilic com-pound and that chlorhexidine diacetate-resistant Pseiudouzzo-nas strains, which have a reduced content of lipopolysac-charide as compared with that in normal strains, may resultfrom a decreased ability to adsorb to the agent. We foundthat the content of FNL in the resistant strains (groups A andB) was higher than that in the susceptible strains (group S)and also that the increase in FNL in the more resistantstrains (group B) was greater than that in the less resistantstrains (group A) (Fig. 3). The amount of BC adsorbed toFNL of the resistant strains were smaller than that ofsusceptible strains (Fig. 4).
3.0
=, 2 .0
3c
_ 1.0z
-1ID9
P.aerugLnosa B group
8 J31,2 t6v 9
4 5 Log P
P.aerugLnosa A group
3
28 88 11 187S82 81
5 Log P
FIG. 5. Relationship between increases or decreases in fatty acidcontents in the tested bacteria and log P values (P < 0.05). Fattyacids are identified as follows: 1, 3-hydroxyundecanoic acid: 2,
2-hydroxydecanoic acid; 3, lauric acid: 4. unknown; 5, 2-hydroxy-dodecanoic acid; 6, 3-hydroxytridecanoic acid; 7, 2-hydroxytride-canoic acid: 8, 3-hydroxydodecanoic acid: 9, myristic acid: 10.unknown; 11, palmitioleic acid; 12, palmitoleic acid; 13, palmiticacid; 14, unknown: 15, 10-heptadecanoic acid; 16, unknown; 17,oleic acid: 18, oleic acid; 19, stearic acid. Symbols: 0, content ofeach fatty acid in the susceptible bacteria (group S) *0. content ofeach fatty acid in the resistant bacteria (groups A and B).Significant at P < 0.05.
BC is a hydrophilic compound (log P value, 3.22), and thelog P values of fatty acids were higher than that of BC. Webelieve that the increase in fatty acids having log P valueshigher than that of BC reduces the ability of BC to permeatecells and leads to the acquisition of resistance to BC. Theresults obtained in this study suggest that the resistance of P.wierluginiostI to BC is probably the result of increases in PLand FNL. The decrease in cell permeability, as shown by theincrease in fatty acids in the resistant strains, does not bringabout a loss of the barrier function. BC-resistant strainsseem to be undamaged because of a decrease in adsorptionand permeation of the cell wall.
Russell and Gould (28) suggested that the decreasedsusceptibility of enterobacteria to bactericides is probablyassociated with the outer cell layers, which may limit thepenetration of bactericidal molecules to their sites of action.Our results support their hypothesis. Studies of the role ofthe inner membrane in BC resistance are also needed.
ACKNOWLEDGMENTS
We thank 0. Nakaguchi. Fujisawa Pharmaceutical Co.. Ltd.. forsupplying E. coli ES-114. K. Miyano, Osaka Prefectural Institute ofPublic Health, for technical assistance, and N. Sato and M.Kouzaki. Institute of Gifu of Sanwa Chemical Co.. Ltd., forassistance in measuring cell surface potentials.
LITERATURE CITED1. Andermann, G., M. 0. Buhler, and M. Frhart. 1980. Rapid
colorimetric analysis of chlorhexidine in pharmaceutical prepa-rations. J. Pharm. Sci. 69:215-217.
2. Baker, Z., R. W. Harrison, and B. F. Miller. 1941. Action ofsynthetic detergents on the metabolism of bacteria. J. Exp.Med. 73:249-271.
3. Bihler, I., A. Rothestein, and L. Bihler. 1961. The mechanism ofstimulation of aerobic fermentation in yeast by quaternaryammonium detergents. Biochem. Pharmacol. 8:289-299.
4. Broxton, P., P. M. Woodcock, and F. Heatley. 1984. Interactionof some polyhexamethylene biguanides and membrane phos-pholipids in Escherichia co/i. J. AppI. Bacteriol. 57:115-124.
5. Brzezinska, M., R. Benveniste, J. Davies, P. J. L. Danies, and J.Weinstein. 1972. Gentamicin resistance in strains of Pseiudomtto-nIius aei(iginosa mediated by enzymatic N-acetylation of thedeoxystreptamine moiety. Biochemistry 11:761-766.
6. Brzezinska, M., and J. Davies. 1973. Two enzymes whichphosphorylate neomycin and kanamycin in Eschlerichia (/listrains carrying R factors. Antimicrob. Agents Chemother.3:266-269.
7. Chaplin, C. E. 1952. Bacterial resistance to quaternary ammo-nium disinfectants. J. Bacteriol. 63:453-458.
8. Gilbert, P., and M. R. W. Brown. 1978. Influence of growth rateand nutrient limitation on the gross cellular composition ofPseuldoitnona(is aierulginosai and its resistance to 3- and 4-chlo-rophenol. J. Bacteriol. 133:1066-1072.
9. Goto, S., K. Jyo, T. Kawakita, N. Kosakai, S. Mitsuhashi, T.Nishino, N. Osawa, and H. Tanami. 1981. Modification ofminimum inhibitory concentration (MIC) test method. Chemo-therapy (Tokyo) 29:76-79.
10. Hansch, C., and T. Fujita. 1964. p-a-ir Analysis. A method forthe correlation of biological activity and chemical structure. J.Am. Chem. Soc. 86:1616-1626.
11. Hugo, W. B. 1982. Disinfection mechanism. p. 158-185. In A. D.Russell et al. (ed.), Principles and practice of disinfection.Preservation and sterilization. Blackwell Scientific Publications,Ltd., Oxford.
12. Kondo, S., K. Hamana, R. Utahara, K. Maeda, and S. Mitsu-hashi. 1967. Phosphorylative inactivation of aminoglycosidicantibiotics by Escherichia coli carrying R factor. Science 157:1559-1561.
13. Kondo, S., M. Okanishi, R. Utahara, K. Maeda, and H.Umezawa. 1968. Isolation of kanamycin and paromine inacti-
cE 2,0 -
1.0 _
Hk
VOL. 55, 1989
on January 15, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

2040 SAKAGAMI ET AL.
vated by E. coli. J. Antibiot. 21:22-29.14. Kuhn, R., and 0. Dann. Uber Invertseifen. 11. Butyl-octyl-
lauryl- und cetyl-dimethyl sulfonium iodine. Ber. Dtsch. Chem.Ges. 73:1092-1102.
15. Moss, C. W., and V. J. Lewis. 1967. Characterization ofclostridia by gas chromatography. AppI. Microbiol. 15:390-397.
16. Newton, B. A. 1958. The strategy of chemotherapy. Surfaceactive bactericides. Symp. Soc. Gen. Microbiol. vol. 8.
17. Nikaido, H. 1976. Outer membrane of Salinone//l tvyp/linlurilml:transmembrane diffusion of some hydrophobic substances. Bio-chim. Biophys. Acta 433:118-132.
18. Nikaido, H., and T. Nakae. 1979. The outer membrane ofgram-negative bacteria. Adv. Microb. Physiol. 20:163-250.
19. Nikaido, H., and M. Vaara. 1985. Molecular basis of bacterialouter membrane permeability. Microbiol. Rev. 49:1-32.
20. Nishikawa, K., S. Oi, and T. Yamamoto. 1979. A bacteriumresistant to benzalkonium chloride. Agric. Biol. Chem. 43:2473-2478.
21. Okamoto, S., and Y. Suzuki. 1965. Chloramphenicol, dihydros-treptomycin, and kanamycin-inactivating enzymes from multi-ple-resistant Escherichia coli carrying episome 'R." Nature(London) 5017:1301-1303.
22. Ordal, E. J., and A. F. Borg. 1942. Effect of surface activeagents on oxidations of lactate by bacteria. Proc. Soc. Exp.Biol. Med. 50:332-336.
23. Osborne, M. J., and H. C. P. Wu. 1980. Proteins of the outermembrane of gram-negative bacteria. Annu. Rev. Micr-obiol.34:369-422.
24. Pollock, M. R. 1960. In K. Myrback, H. Lardy, and P. D. Boyer(ed.), The enzymes, 2nd ed. Academic Press, Inc., New York.
25. Putnam, F. W. 1948. The interaction of protein and syntheticdetergents. Adv. Protein Chem. 4:79-89.
26. Richards, R. M. E., and R. H. Cavill. 1976. Electron microscope
study of effect of benzalkonium chloride and edetate disodiumon cell envelope of Pseuldoinionias (eleruginiosai. J. Pharm. Sci.65:76-80.
27. Russell, A. D. 1986. Mechanisms of bacterial resistance to
antibacterial agents. Pharm. Int. 7:300-308.28. Russell, A. D., and G. W. Gould. 1988. Resistance of enterobac-
teriaceae to preservatives and disinfectants. J. Appl. Bacteriol.64: 167S-195S.
29. Scharif, T. G., and W. C. Maupin. 1960. Correlation of themetabolic effects of benzalkonium chloride with its membraineeffects in yeast. Biochem. Pharmacol. 5:79-86.
30. Shaw, W. V. 1967. The enzymatic acetylation of chloramphen-icol by extracts of R factor-resistant Escherichia coli. J. Biol.Chem. 25:687-693.
31. Shigeta, S., H. Okamura, K. Honzumi, Y. Yasunaga, R. Ku-mada, and S. Endo. 1976. Cerebritis after the operation byAc(hronobactervx!/loso.idlans which occurred frequently on thepatients of V-P shunt detention. Diagn. Treatment 64:1872-1877.
32. Stewart, T. 1965. The penicillin group of drugs. Elsevier Bio-medical Press, Amsterdam.
33. Stickland, L. M. 1956. The Pasteur effect in normal yeast and itsinhibition by various agents. Biochem. J. 64:503-515.
34. Suzuki, Y., and S. Okamoto. 1967. The enzymatic acetylation ofchloi-amphenicol by the multiple drug-resistant Escherichia (olicarrying R factor. J. Biol. Chem. 25:4722-4730.
35. Tennent, J. M., B. R. Lyon, M. T. Gillespie, J. W. May, andR. A. Skurray. 1985. Cloning and expression of Staphl/ococcsls(aurcIis plasmid-mediated quaternary ammonium resistance inEsce/irichia (o/i. Antimicrob. Agents Chemother. 27:79-83.
36. Umezawa, H., M. Okanishi, R. Utahara, K. Maeda, and S.Kondo. 1967. Isolation and structure of kanamycin inactivatedby a cell free system of kanamycin-resistant E. co/i. J. Antibiot.20:136-141.
37. Umezawa, H., S. Umezawa, T. Tsuchiya, and Y. Okazaki. 1971.3'.4'-dideoxy-kanamycin B active against kanamycin-resistantEschericlija coli and PseuICloinona1is aieruilginlosai. J. Antibiot.24:485-486.
38. Umezawa, Y., M. Yagisawa, T. Sawa, T. Takeuchi, H. Umezawa,H. Matsumoto, and T. Tazaki. 1975. Aminoglycoside 3'-phos-photransferase 111. a new phosphotransferase. Resistancemechanism. J. Antibiot. 28:845-853.
39. Uyeda, M., M. Sakata, M. Shibata, and T. Hitoshi. 1982. Someproperties of chlorhexidine-resistant stirains of Serrauia inlarces-cens. Chemotherapy (Tokyo) 30:286-290.
APPL. ENVIRON. MICROBIOL.
on January 15, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from





























