Embed Size (px)
Citation preview

This document is downloaded at: 2018-06-01T14:16:08Z
Title Purification and Characterization of Fimbriae of Vibrio cholerae O1,Classical Biotype, Inaba Serotype
Author(s)Ehara, Masahiko; Ishibashi, Mikako; Iwami, Mamoru; Ichinose, Yoshio;Shimotori, Shoichi; Supawat, Krongkaew; Chiewslip, Dumrong;Tansupousawadikul, Somsit
Citation 熱帯医学 Tropical medicine 31(1). p7-16, 1989
Issue Date 1989-03-31
URL http://hdl.handle.net/10069/4539
Right
NAOSITE: Nagasaki University's Academic Output SITE
http://naosite.lb.nagasaki-u.ac.jp

Trop. Med., 31(1), 7-16, March, 1989 7
Purification and Characterization of Fimbriae of
Vibrio cholerae O1, Classical Biotype, Inaba Serotype
Masahiko EHARA1, Mikako ISHIBASHI1, Mamoru IWAMI1
Yoshio ICHINOSE1, Shoichi SHIMOTORI2
and
Krongkaew SUPAWAT3, Dumrong CHIEWSLIP3,
Somsit TANSUPOUSAWADIKUL4
1Department of Bacteriology, Institute of Tropical Medicine, Nagasaki University,
12-4 Sakamoto-machi, Nagasaki 852, Japan,2Department of Microbiology, School of Health Science, Kyushu University,Higashi-ku, Fukuoka 812, Japan.
3National Institute of Health, Nonthaburi 11000, Thailand.4Bamrasnaradura Infectious Disease Hospital, Nonthaburi 11000, Thailand
Abstract: Fimbrial subunits have been shown to share commonantigenicities between
Vibrio cholerae biotype El Tor and biotype classical (Ehara et al., 1987). The above fact
was confirmed by western blotting and immunoelectron microscopy. Net-work and bun-
dle formations were observed in the fimbrial preparations by immunoelectron microscopy.
Purified fimbriae of Vibrio cholerae O1, classical biotype possessed haemagglutination ac-
tivity (HA) against chicken erythrocytes. This HA was inhibited in the presence of
D-mannose but not by L-fucose. Antigenic determinants of the fimbriae were exposedon the sides of the fiber. The low hydrophobicity of the fimbriae molecule revealed by
the amino acid composition suggests that the fimbriae may interact with epithelial cells
not by hydrophobic interaction. Development of the antibody against fimbrial subunit of
V. cholerae O1 was confirmed in the blood sera obtained from convalescent cholera pa-tients.
Key words: Vibrio cholerae O1, fimbriae, component vaccine
INTRODUCTION
The colonization of enteropathogenic Vibrio cholerae 01 to the intestinal epitheliumis an essential step in the production of watery diarrhoea. Whether V. cholerae 01 produceany colonization factor during the interaction between vibrios and epithelial cells of
Received for Publication, February 3, 1989.Contribution No. 2245 from the Institute of Tropical Medicine, Nagasaki University.

8
small intestine was a controversial problem. The answer to the problem was obtained bythe observation of colonized vibrios after the detachment of the vibrio cells from epithelialcells of small intestine. The detached vibrios were shown to possess several long flexiblefibers (fimbriae) (Ehara et al, 1986).
The purified fimbriae of an El Tor strain was shown to have the haemagglutinatingactivity against human type 0 red blood cells as well as chicken erythrocytes. This HAwas sensitive both to D-mannose and L-fucose (Ehara et al., 1987). Since fimbriae ofclassical vibrios were not characterized in the previous paper, this paper describes thepurification of fimbriae of Vibrio cholerae 01, classical biotype, Inaba serotype and theirpartial characterization.
MATERIALS AND METHODS
Bacteria; Non-motile mutant strain Bgd 17-1 was obtained from the strain Bgd 17(x-23751) by treatment with nitrosoguanidine (50 pg/ml).
Bgd 17 (x-23751) of the classical biotype Inaba serotype was isolated inBangladesh in 1982 and kindly provided by Dr. Y. Takeda.
Non-motile mutant derivatives from Bgd 17 were positive in a rabbit ileal loop testand were confirmed to adhere to the ileal epithelium under scanning electron microscopy.
Medium: Vibrios were grown on TCG agar (Ehara et al., 1986) (1 % Bactotryptone,0.2 % Yeast extract, 0.5 % NaCl, 0.3 % NaHC03) 0.02 % Thioproline, 0.1 % MonosodiumL-glutamate, 1 mM EGTA, and 2 % Agar) at 30 °C for 48 h.
Purification of fimbriae: Vibrio cells were cultured on TCG agar in 80 Roux bottlescontaining 100 ml medium at 30 °C for 48 h.
After the addition of 10 ml of 0.1 M potassium phosphate buffer pH 6.8 (PPB) con-taining 0.5 mM MgCl2 and 1 mM EGTA, cells were harvested by pipette and centrifugedat 10,000 x^ for 1 h at 4 °C. Whole cells were resuspended in PPB to 0.5 g (wet weight) /ml, then homogenized in a cooled Sorvall Omnimixer at setting 5 for 5 min. Thehomogenate was centrifuged at 12,000 xg" for 30 min.
The resultant supernatants were pooled at 4 °C and the pellet was resuspended withfresh PPB and the homogenization repeated three times. Each supernatant was combinedand kept at 4 °C overnight after the addition of DNase I and RNase, each at 100 pg I ml.The mixture was centrifuged at 48,000 x^ for 1 h at 4 °C to remove cell debris. Thesupernatant was concentrated to 4 ml over an Amicon PM 10 membrane. The concen-trated sample was loaded onto the top of 20-50 % sucrose linear gradient solution andcentrifuged with SW 41 rotor (Beckman) at 200,000 x£- for 16 h at 4 °C. Fractions of 0.5ml were taken from the top of the tube and monitored for purity by electronmicroscopyand sodium dodecyl sulphate polyacryl amide gel electrophoresis. Fimbriae-rich fractionswere combined and loaded onto Sepharose CL-4B column chromatography equilibratedwith 20 mM PPB with 10 % sucrose.Each fraction was monitored for purity as mentioned earlier.

9
Preparation of antiserum to EI Tor fimbriae: Fimbriae purified from K 23 strain
were mixed with an equal volume of Freund's complete adjuvant (0.5 ml fimbnal solution
of 200 jHg/ml and 0.5
the production of high
jected subcutaneously
thigh each week for
substituting incomplete
the last injection. The
ml of the adjuvant) and used to parenterally immunize rabbits for
titre antibody against native EI Tor fimbnae. The rabbits were in-
at two sites in the back and intramuscularly at two sites in the
the first month and every two weeks for the second month
adjuvant for complete adjuvant. They were bled one week after
lgG fraction of the antisera was prepared by DEAE-Sephadex
A-50 column chromatography and was used for immunoblotting and for immunoelectron
microscopy.
SDS-polyacrylamide gel electrophoresis: polyacrylamide gel electrophoresis
(PAGE) was performed in 1.5 mm thick slab gels according to the system of Laemmli
(1970). Fimbrial preparations were heated at 100 ℃ for 5 min in sample buffer (0.05 M
Tns-HCI pH 6.8 containing 2.5 % (w/V) sodium dodecyl sulfate (SDS), 10 % (v./v)
glycerol, 5 % (v/v) 2-mercaptoethanol, and a trace of bromophenol blue as tracking dye).
Samples were applied to the polyacrylamide slab gel (12 %) using a 4 % stacking gel and
were electrophoresed for 7 h at 20 mA constant current per slab. The protein bands were
stained with Coomassie brilliant blue R 250. For molecular weight calibration, the low
molecular weight market kit (Pharmacia Fine Chemicals) was used.
Western blotting: Immunoblotting was performed according to the techniques of
Towbm et at. (1979) and Burnett (1981). Purified fimbriae were subjected to SDS-PAGE
in duplicate on different gels, one of which was stained with Coomassie blue and the other
electroblotted onto nitrocellulose membrane (BIO-RAD) using a BIO-RAD electroblotting
apparatus (30 V for 16 h and 70 V for 1 h). Horseradish-peroxidase-conjugated goat an-
ti-rabbit lgG was from Cappel Laboratories lnc., USA, and 4-chloro-1-naphthol
(BIO-RAD Laboratories) was used in the development of the color reaction.
Haemagglutmation and naemagglutination inhibition tests.
Purified fimbriae of strain Bgd 17-1 were dialysed against 20 mM PPB to remove
sucrose. Techniques for quantification of haemagglutination (HA) and HA inhibition with
sugars were adapted from Jones et al. (1976). Serial two-fold dilution of fimbrial prepara-
tions (200 ng/ml) were prepared in a U-bottomed microtitre plate in 25 //I of 20 mM
PPB. Chicken red blood cells (2 % in 20 mM PPB) were added to 25 〃1, the plate was
tapped to mix the interactions and the erythrocytes were settled at 25 ℃ for 30 min.
The titre is defined as the reciprocal of the highest dilution in which HA was clear-
ly visible. To test whether the HA reaction was inhibitable by specific monosaccharides,
erythrocytes with 1 % (w,/w) D-mannose or L-fucose were added to the wells containing
serially diluted fimbriae and HA reactions were read after 30 min at 25 ℃.
Ammo acid composition: Amino acid analysis was carried out on the fimbrial
subunit eluted from a preparative SDS-PAGE. The extracted fimbrial subunit solution
was first dialyzed against 0.2 M NaCl to remove glycine in the electrophoresis buffer
before dialysis against distilled water. The sample was hydrolyzed with 6 N HCl at 110℃

10
for 24 h m an evacuated sealed tube. The hydrolysate was analyzed with a JEOL
JLC-200A amino acid analyzer.
Detection of antibody against fimbrial subunit in blood sera from convales-
cent cholera patients: Paired blood sera were taken from cholera patients admitted to
Bamrasnaradura Hospital by one of the authors (S. T.). Detection of the antibody was car-
ried out by western blotting, using crude fimbnal fraction as antigen. Blood sera were
diluted 200-fold with Tns buffered saline.
Electron microscopy: For negative staining, one drop of the sample was placed on a
sheet of PARAFILM and a Formvar-coated copper grid was floated on the drop for 2
min. The excess liquid was washed three times with distilled water each time for 10 s,
then stained with 1 % uranyl acetate for 30 s. The excess stain was removed with the tip
of a filter paper. For immunoelectron microscopy, the copper grids coated with purified
fimbriae or cell-suspension were reacted with the primary antibody (antiーEI Tor fimbriae
antibody) and with the secondary antibody (antトrabbit lgG goat serum conjugated with
15 nm colloidal gold, E. Y. LABS, INC. SANMAEO, CA 94401 USA) before negative
staining. The specimens were examined with a JEM 100 ex electron microscope operated
at 80kV.
RESULTS
Purification of fimbriae: Fimbriae rich fractions (fraction No.12 to 16) obtained bysucrose density gradient centrifugation were combined and loaded onto a Sepharose CL-4
gel : Sepharose CL-4B
column size : 1.5X15(cm)
flow rate : 12 ml/hourBuffer: 20mM PPB with 10% sucrose
Sample: fimbriae rich fractions
o.;
0.2
0 .1
E
c
o
COIN
QO
0 10 20 30
Fraction No.
Fig. 1. Gel filtration profile.

ll
94K67K
43K
30K
20.1K-
14.4K-
Fig. 2. a:western blotb:SDS- PAGE.
a
b
B column. Fimbriae were eluted at the void volume fraction (Fig. 1). The fimbrial prepara-tions obtained by the above procedure were analyzed by SDS-PAGE and Western blot.The molecular weight of the structural subunit of fimbriae (fimbrillin) was ca.16,000. Asreported in the previous paper, fimbriae of a classical strain were stained faintly in theLaemmli system, although the same specimen showed numerous fibrils under electronmicroscopy and was densely immunostained in a western blotting (Fig. 2). Electronmicroscopic examination of the purified fimbriae from V. cholerae 01 strain Bgd 17-1showed the presence of numerous, thin, non-channeled fimbriae of external diameter 5-7nm, typical of the fimbriae present on intact, non-homogenized bacteria (Ehara et al.,
a
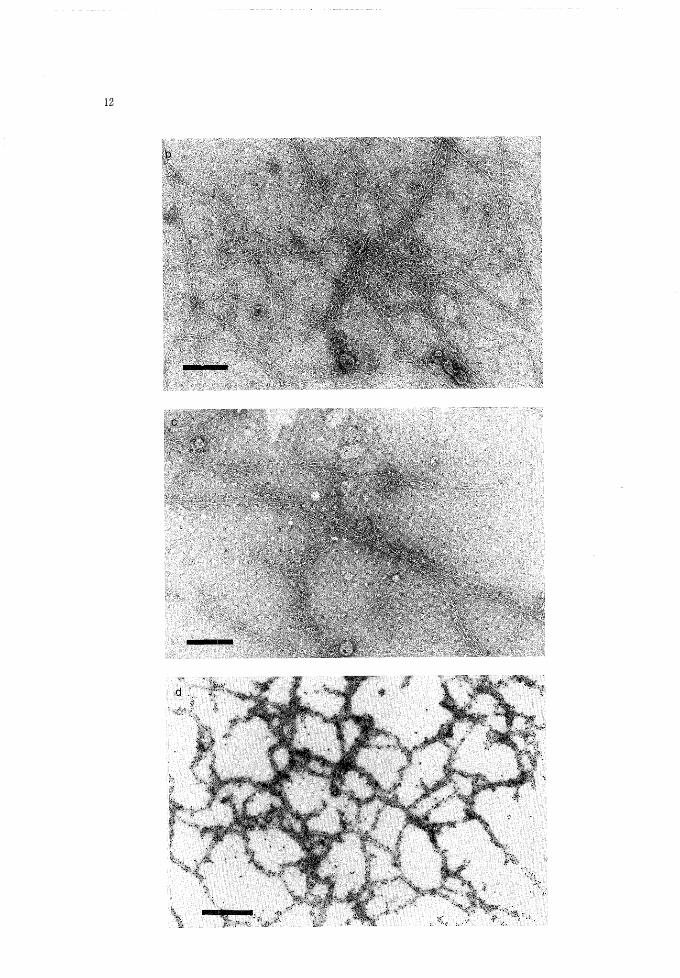
12
i tt'
_c
d

13
Electron micrographs of purified and immunodecorated fimbriae.(a) fimbriae purified through a gel filtration; (b), (c) fimbriae reacted with anti-El Tor fim-briae antibody, (d) fimbriae reacted with anti-El Tor fimbriae antibody for 15 min followingthe reaction with anti-rabbit IgG goat serum conjugated with 15 nm colloidal gold, (e) Bgd17(parent strain) reacted with anti-El Tor fimbriae antibody in the same method as used insample(d).Bars indicate 250 nm in (a)-(c), 500 nm in (d) and 100 nm in (e).
1986, Paris et al., 1982, Tweedy et al, 1968) as shown in Fig. 3 a. When reacted with an-ti-El Tor fimbriae antibody, net-work and bundle formation (Fig. 3 b, c) were observedin the specimens of classical fimbriae. The electron micrographs of classical fimbriaetreated with anti-El Tor fimbriae antibody following the reaction with anti-rabbit IgGgoat serum conjugated with 15 nm colloidal gold were shown in Fig. 3 d and 3 e. The an-tibodies bound to the sides of the fimbrial net-work.
Haemagglutination activity of fimbriae: Fimbriae of classical vibrio had low HAtiter to chicken erythrocytes and HA inhibition was obtained in the presence of D-man-nose but not in the presence of L-fucose as shown in Table 1.
Detection of antibody against fimbrial subunit: The raised antibody against fim-brial subunit was detected in each blood specimen taken 3, 4, 5 and 7 weeks after onset(Fig. 4).
Table. 1. Haemagglutination activity (HA) tochicken erythrocytes
Classical fimbriae (120 fig/ml)
i n the presence ofD -mannoseL-fucose
2
8
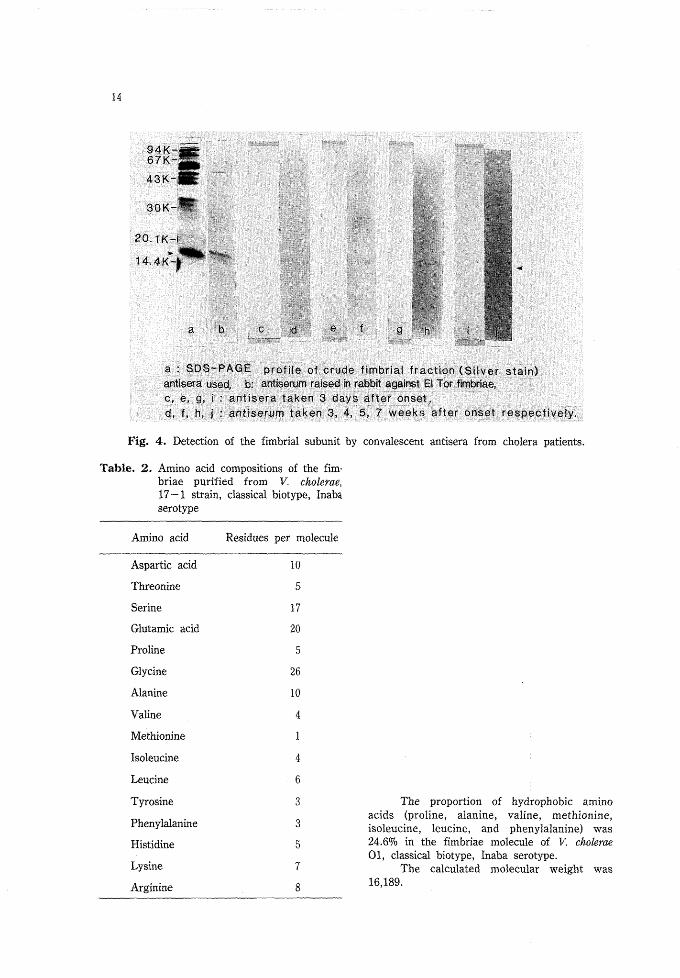
14
20.1K-
14.4K*
a b
c e f
g "- h" i
a : SDS-PAGE profile of;crude fimbriai fraction (Silver stain)antisera used, b: antiserufn raised in rabbit against El Tor fimbriae,c, e, g, I : antisera taken 3 days after onset,d, f, h, j : antfserum taken 3, 4, 5, 7 weeks after onset respectively.
Fig. 4. Detection of the fimbrial subunit by convalescent antisera from cholera patients.
Table. 2. Amino acid compositions of the fim-briae purified from V. cholerae,17-1 strain, classical biotype, Inabaserotype
Arainoacid Residues per molecule
Aspartic acid
Threonine
S erine
Glutamic acid
Prolin e
Glycine
Alanine
Valine
Methionine
Isoleucine
Leucine
Tyrosine
Phenylalanine
Histidine
Lysine
Arginine
10
5
17
20
5
26
10
4
1
4
6
3
3
5
7
The proportion of hydrophobia aminoacids (proline, alanine, valine, methionine,isoleucine, leucine, and phenylalanine) was24.6% in the fimbriae molecule of V. cholerae01, classical biotype, Inaba serotype.
The calculated molecular weight was16,189.

15
Amino acid composition: A quantitative amino acid analysis was performed on thefimbrial subunit purified by a preparative SDS-PAGE (Table 2). The amino acid composi-tion was characterized by low concentrations of hydrophobic amino acids. Values of Serand Gly residues may be overestimated due to contamination during the preparativeSDS-PAGE.
DISCUSSION
It is well known both from epidemiological studies of cholera in endemic areas(Ehara et al, 1985, Glass et al, 1982, Mosley, 1969, Woodward, 1971) and from volunteerstudies (Cash et al., 1974, Levine et al., 1981) that a natural infection with V. cholerae 01resulting in diarrhoea gives rise to long-lasting protective immunity against a second at-tack of cholera. A recent study by Lycke et al. (1986) using the RITARD model has alsodescribed a state of protective immunity which extends to strains of unrelated serotypeand biotype. From the data obtained by a western blotting (Fig. 2 a) and by immunoelec-tron microscopy (Fig. 3 d) it is apparent that classical fimbrial subunit has a molecularweight of 16,000 and shares antigenicity independent of biotype and serotype. This studyshows that the major antigenic determinants of fimbriae of classical vibrios are exposed onthe sides of the fimbriae and easily detected by the antibody. It is noteworthy that the an-tibody against fimbriae was detected in the blood sera obtained from convalescent cholera
patients. Rabbits parenterally immunized with El Tor fimbriae were protected from col-onization of vibrios unrelated serotype and biotype (Ehara et al., 1988). Our data mention-ed as above suggest that the fimbriae of V. cholerae is one of colonization factors andhighly antigenic, independent of biotype and serotype. Therefore, the fimbrial antigen ofV. cholerae may be a suitable antigen to be analyzed in the blood sera of convalescentcholera patients and of healthy carriers in cholera endemic areas.
The relatively low hydrophobicity of the fimbriae molecule of classical vibrio maysuggest that this protein interacts with epithelial cells by ionic interaction or with specificaffinity to certain receptor and not by hydrophobic interaction.
The cell-associated haemagglutinin of classical vibrios has been reported to be sen-sitive to L-fucose and not to D-mannose (Hanne and Finkelstein). Our data show thatpurified fimbriae of classical vibrios are D-mannose-sensitive haemagglutinin (MSHA)and are not affected in the presence of L-fucose. This discrepancy may suggest thepresence of different type of cell-associated haemagglutinin in classical vibrios or thepresence of one type of filamentous haemagglutinin with different chemical modificationsdue to culture conditions. Furthermore, the low HA titer of purified fimbriae of a classicalstrain may suggest that native cell-associated haemagglutinins have different structureand function from purified ones.
REFERENCES
1) Burnett, W. N. (1981): Western blotting. : electrophoretic transfer of proteins from

16
SDS-polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody
and radioiodinated protein A. Anal. Biochem. 112, 195-203.
2) Cash, R. A., Music, S. E., Libonati, J. P., Snyder, M. J., Menzel, R. P. & Hornick, R. B. (1974):
Response of man to infection with Vibrio cholerae. II. Protection from illness afforded by previousdisease and vaccine. J. Infect. Dis. 130, 325-333.
3) Ehara, M., Ishibashi, M., Ichinose, Y., Iwanaga, M., Shimotori, S. & Naito, T. (1988): Fimbriae
of Vibrio cholerae 01. pp89-104. In Ohtomo, N. & Sack, R. B. (ed.). ADVANCES IN
RESEARCH ON CHOLERA AND RELATED DIARRHEAS. KTK Scientific Publishers, Tokyo.
4) Ehara, M., Ishibashi, M., Ichinose, Y., Iwanaga, M., Shimotori, S., & Naito, T. (1987): Purifica-
tion and partial characterization of fimbriae of Vibrio cholerae 01. Vaccine 5 (4), 283-288.
5) Ehara, M., Ishibashi, M., Watanabe, S., Iwanaga, M., Shimotori, S., & Naito, T. (1986): Fimbriae
of Vibrio cholerae 01: Observation of fimbriae on the organisms adherent to the intestinal
epithelium and development of a new medium to enhance fimbriae production. Trop. Med. 28 (1),21-33.
6) Ehara, M., Watanabe, S., Ichinose, Y., Shimotori, S., T. K. Arap Siongok, Muriith. K. A. A., &
Sang, F. C. (1985): Epidemiological observation of cholera in rural Kenya in 1983. Trop. Med. 27
(4), 229-240.
7) Paris, A., Lindahl, M., & Wadstrom, T. (1982): High surface hydrophobicity of haemagglutinating
Vibrio cholerae and other vibrios. Curr. Microbiol. 7, 357-362.
8) Glass, R. I., Becker, S., Hug, I., et al. (1982): Endemic cholera in rural Bangladesh. Am. J.
Epidemiol. 116, 959-969.
9) Hanne, L. F., & Finkelstein, R. A. (1982): Characterization and distribution of the hemagglutinins
produced by Vibrio cholerae. Infect. Immun. 36, 209-214.
10) Jones, G. W., & Freter, R. (1976): Adhesive propertoes of Vibrio cholerae : nature of the interac-
tion with isolated rabbit brush border membranes and human erythrocytes. Infect. Immun. 14,
240-245.
ll) Laemmli, U. K. (1970): Cleavage of structural proteins during the assembly of the head of
bacteriophase T 4. Nature (London) 227, 680-685.
12) Levine, M. M., Black R. E., Clements, M. L., Cisneros, L., Nalin, D. R., & Young, C. R. J.
(1981): Duration of infection derived immunity to cholera. J. Infect. Dis. 143, 818-820.
13) Lycke, N., Svennerholm, A. M., & Holmgren, J. (1986): Strong biotype and serotype
cross-protective antibacterial and antitoxic immunity in rabbits after cholera infection. J. Infect.
Dis. 1, 361-371.
14) Mosley, W. H. (1969): Vaccines and somatic antigens. The role of immunity in cholera. A review
of epidemiological and serological studies. Tex. Rep. Biol. Med. 27 (suppl), 227-241.
15) Towbin, H., Stahelin, T., & Gordon, J. (1979): Electrophoretic transfer of proteins from
polyacrylamide gels to nitrocellulose sheets. Proc. Nate. Acad. Sci. USA 76, 4350-4354.
16) Tweedy, J. M., Park, R. W. A., & Hodgkiss, W. (1968): Evidence for the presence of fimbriae
(pili) on vibrio species. J. Gen. Microbiol. 51, 235-244.
17) Woodward, W. E. (1971): Cholera reinfection in man. J. Infect. Dis. 123, 61-66.





























