Embed Size (px)
Citation preview

Motto:
Navigare necesse est,vivere non est necesse!
– Egy okori hajoskapitany
Navigacio
– Typeset by FoilTEX – 1

Mit is jelent a navigacio?
”Navigation is the process of determining and maintaining a course
or trajectory from one place to another. Processes for estimatingone’s position with respect to the known world are fundamental toit. The known world is composed of the surfaces whose locationsrelative to one another are represented on a map.”
C. R. Gallistel: The Organization of Learning. MIT Press/Bradford Books, MA, 1990
”A navigacio azt a folyamatot jelenti, mely soran meghatarozunk es vegrehaj-
tunk egy cselekvessorozatot, vagy egy palyan eljutunk az egyik helyrol a masikra.
Ehhez szukseges az az alapveto folyamat, amellyel meghatarozzuk helyunket az
ismert vilaghoz kepest. Az ismert vilag pedig olyan feluletekbol all, melyek egy-
mashoz viszonyıtott helyet egy terkepen reprezentaljuk.”
Avagy definialhatjuk pl. az alabbi kerdesekkel:
• Hol vagyok?
• Hol vannak mas helyek hozzam kepest?
• Hogy jutok mas helyekre innen?
T. S. Levitt es D. T. Lawton: Qualitative navigation for mobile robots. Artificial Intelligence 44 305–360, 1990
– Typeset by FoilTEX – 2
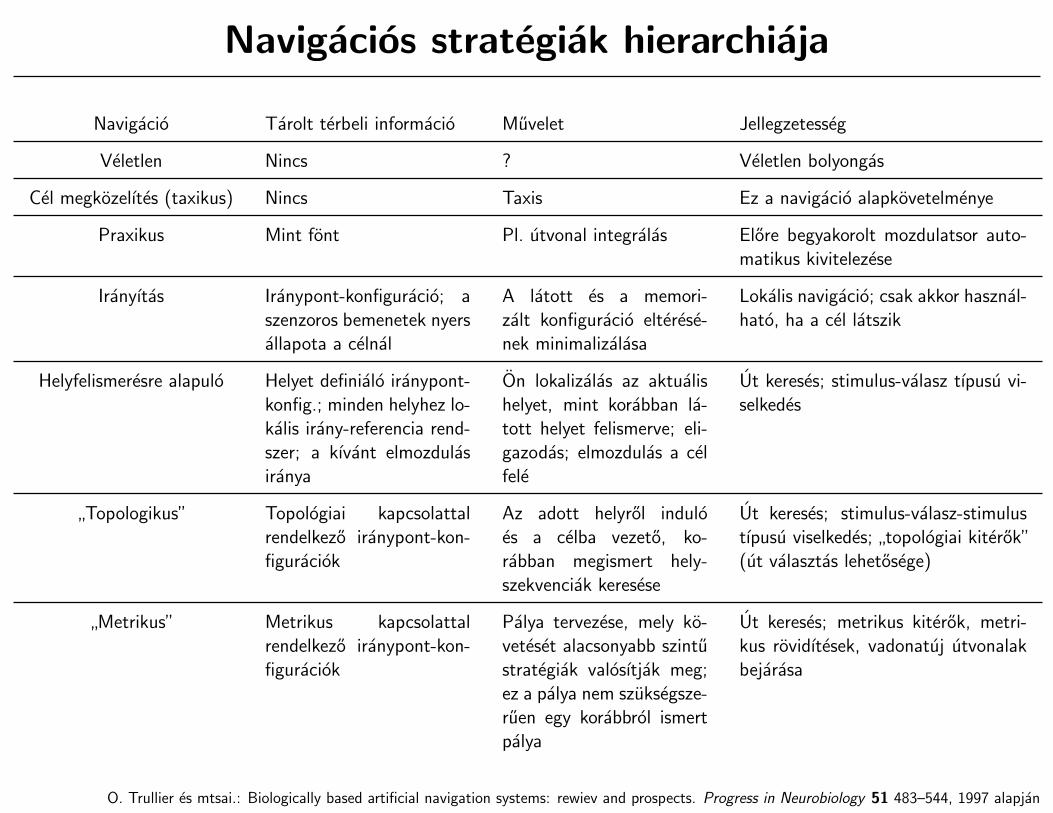
Navigacios strategiak hierarchiaja
Navigacio Tarolt terbeli informacio Muvelet Jellegzetesseg
Veletlen Nincs ? Veletlen bolyongas
Cel megkozelıtes (taxikus) Nincs Taxis Ez a navigacio alapkovetelmenye
Praxikus Mint font Pl. utvonal integralas Elore begyakorolt mozdulatsor auto-matikus kivitelezese
Iranyıtas Iranypont-konfiguracio; aszenzoros bemenetek nyersallapota a celnal
A latott es a memori-zalt konfiguracio elterese-nek minimalizalasa
Lokalis navigacio; csak akkor hasznal-hato, ha a cel latszik
Helyfelismeresre alapulo Helyet definialo iranypont-konfig.; minden helyhez lo-kalis irany-referencia rend-szer; a kıvant elmozdulasiranya
On lokalizalas az aktualishelyet, mint korabban la-tott helyet felismerve; eli-gazodas; elmozdulas a celfele
Ut kereses; stimulus-valasz tıpusu vi-selkedes
”Topologikus” Topologiai kapcsolattal
rendelkezo iranypont-kon-figuraciok
Az adott helyrol induloes a celba vezeto, ko-rabban megismert hely-szekvenciak keresese
Ut kereses; stimulus-valasz-stimulustıpusu viselkedes;
”topologiai kiterok”
(ut valasztas lehetosege)
”Metrikus” Metrikus kapcsolattal
rendelkezo iranypont-kon-figuraciok
Palya tervezese, mely ko-veteset alacsonyabb szintustrategiak valosıtjak meg;ez a palya nem szuksegsze-ruen egy korabbrol ismertpalya
Ut kereses; metrikus kiterok, metri-kus rovidıtesek, vadonatuj utvonalakbejarasa
O. Trullier es mtsai.: Biologically based artificial navigation systems: rewiev and prospects. Progress in Neurobiology 51 483–544, 1997 alapjan
– Typeset by FoilTEX – 3
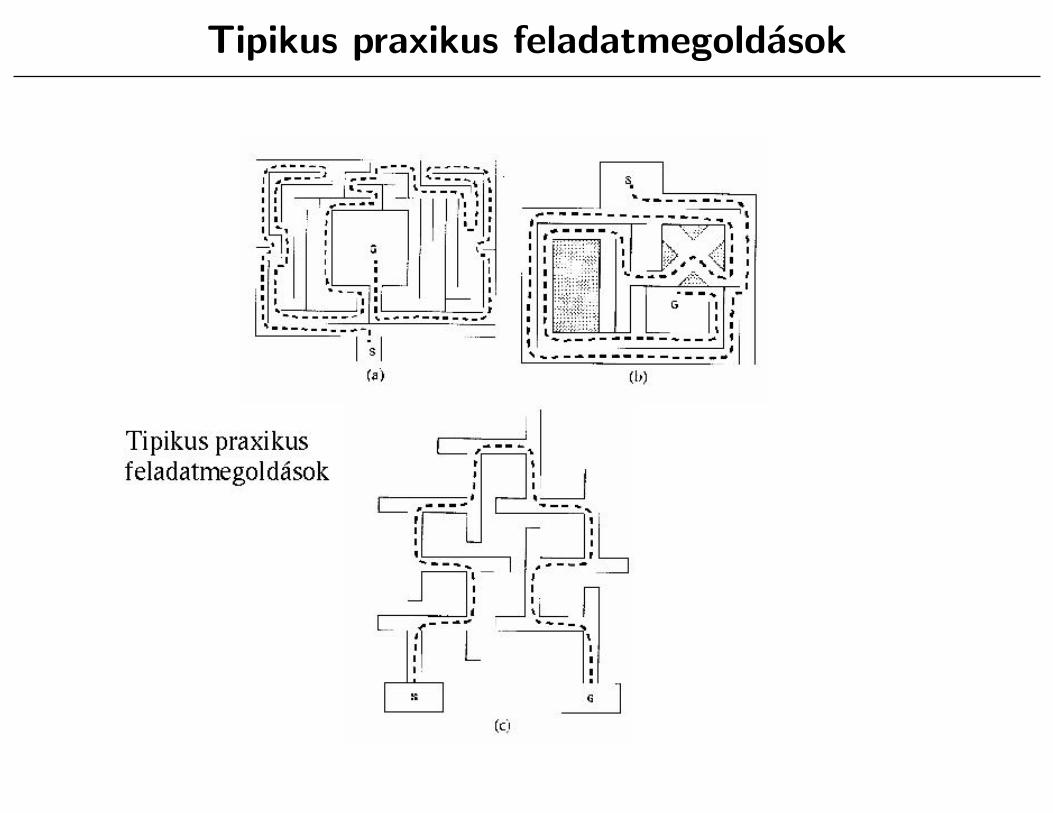
Tipikus praxikus feladatmegoldasok
– Typeset by FoilTEX – 4
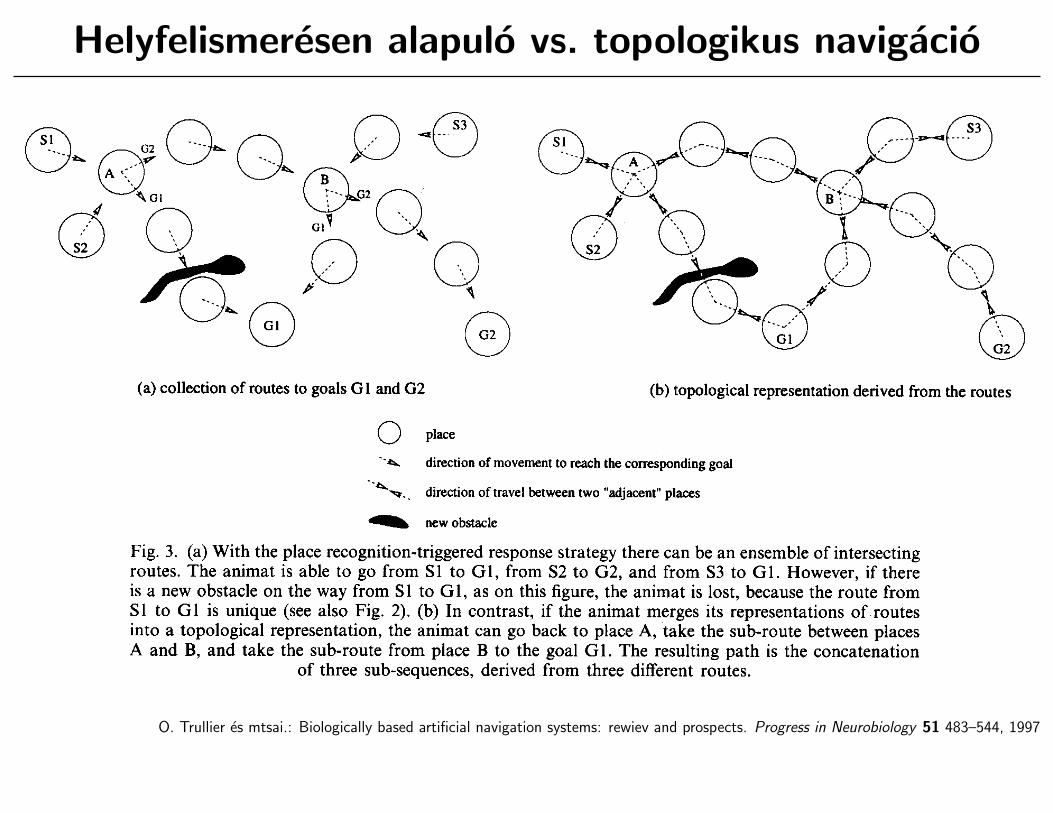
Helyfelismeresen alapulo vs. topologikus navigacio
O. Trullier es mtsai.: Biologically based artificial navigation systems: rewiev and prospects. Progress in Neurobiology 51 483–544, 1997
– Typeset by FoilTEX – 5
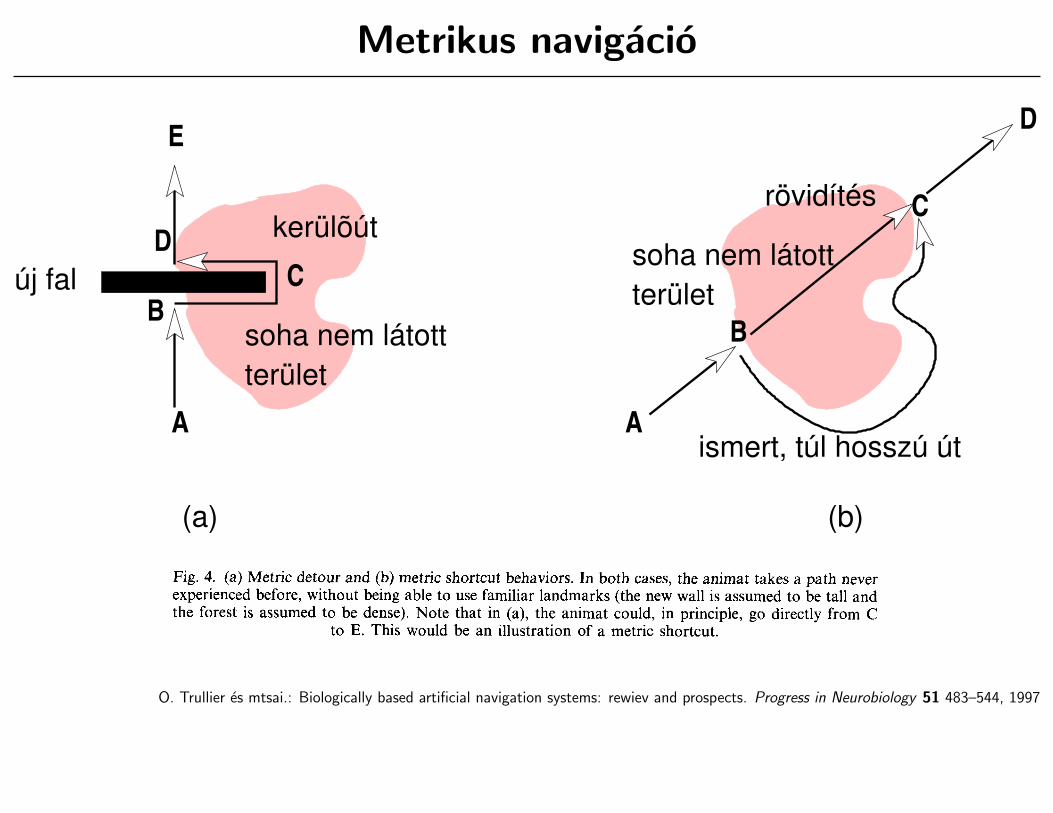
Metrikus navigacio
A
BC
D
E
új fal
kerülõút
A
B
C
D
ismert, túl hosszú út
rövidítés
(a) (b)
soha nem látottterület
soha nem látottterület
O. Trullier es mtsai.: Biologically based artificial navigation systems: rewiev and prospects. Progress in Neurobiology 51 483–544, 1997
– Typeset by FoilTEX – 6

Nehany kıserlet
– Typeset by FoilTEX – 7
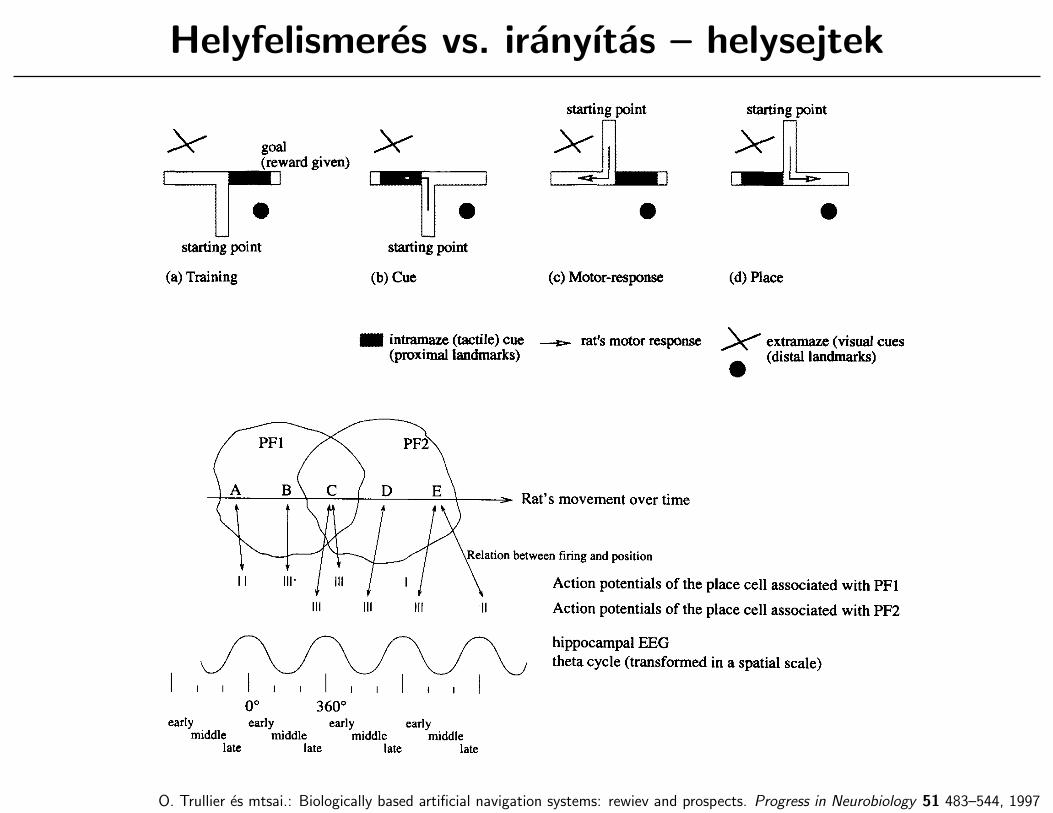
Helyfelismeres vs. iranyıtas – helysejtek
O. Trullier es mtsai.: Biologically based artificial navigation systems: rewiev and prospects. Progress in Neurobiology 51 483–544, 1997
– Typeset by FoilTEX – 8
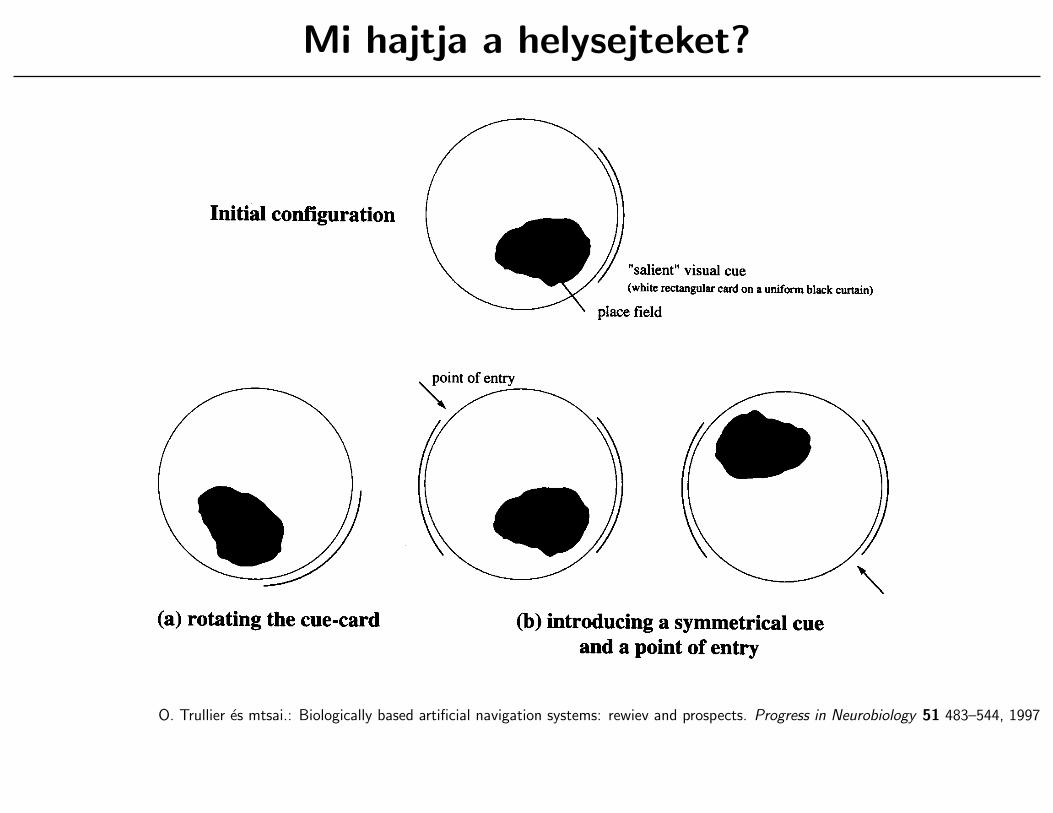
Mi hajtja a helysejteket?
O. Trullier es mtsai.: Biologically based artificial navigation systems: rewiev and prospects. Progress in Neurobiology 51 483–544, 1997
– Typeset by FoilTEX – 9
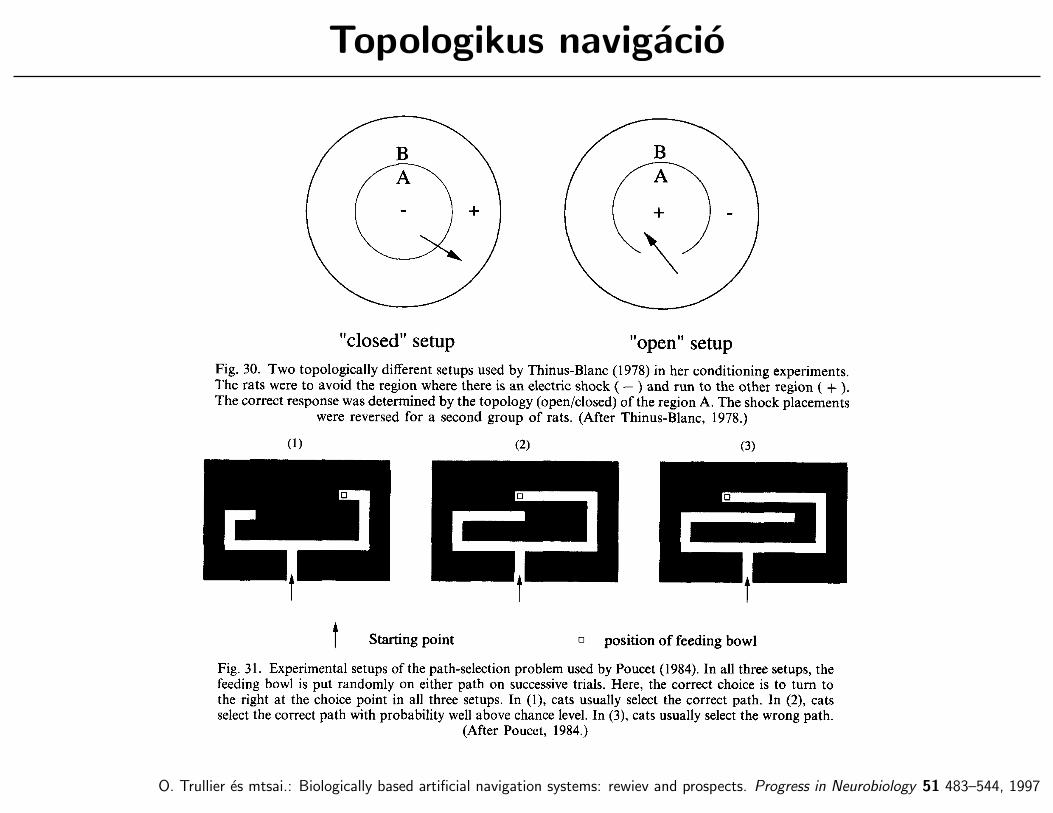
Topologikus navigacio
O. Trullier es mtsai.: Biologically based artificial navigation systems: rewiev and prospects. Progress in Neurobiology 51 483–544, 1997
– Typeset by FoilTEX – 10
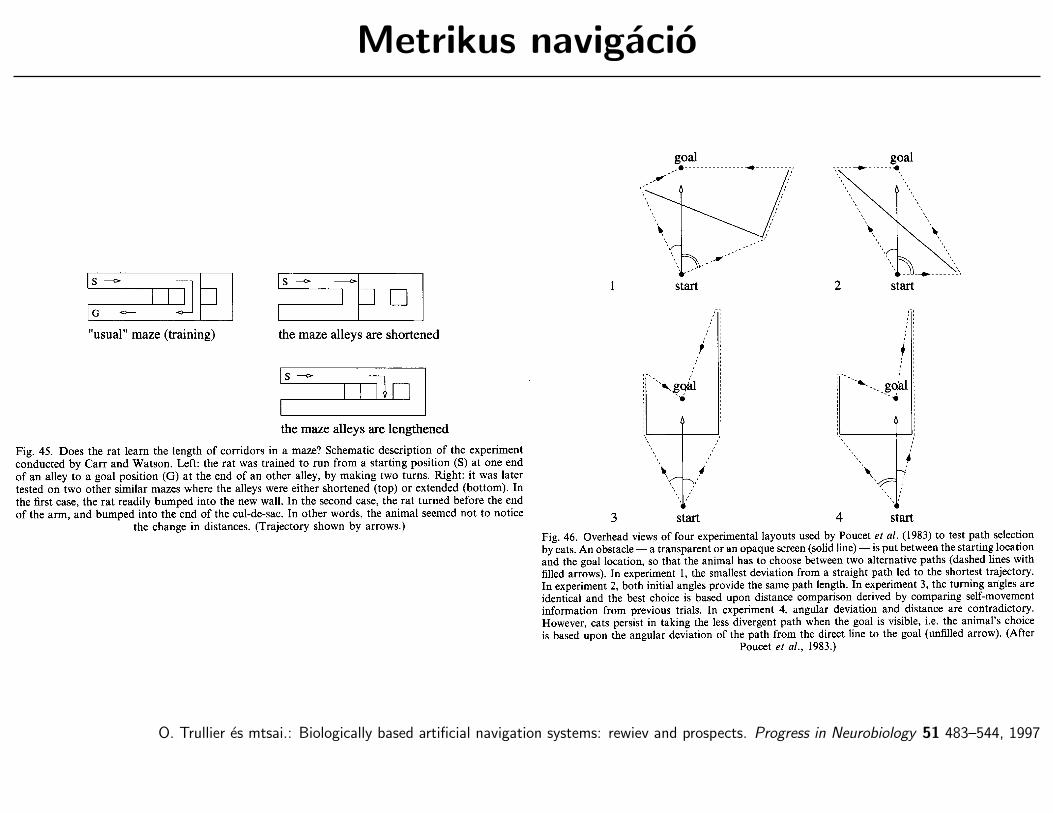
Metrikus navigacio
O. Trullier es mtsai.: Biologically based artificial navigation systems: rewiev and prospects. Progress in Neurobiology 51 483–544, 1997
– Typeset by FoilTEX – 11

Ket modell
– Typeset by FoilTEX – 12
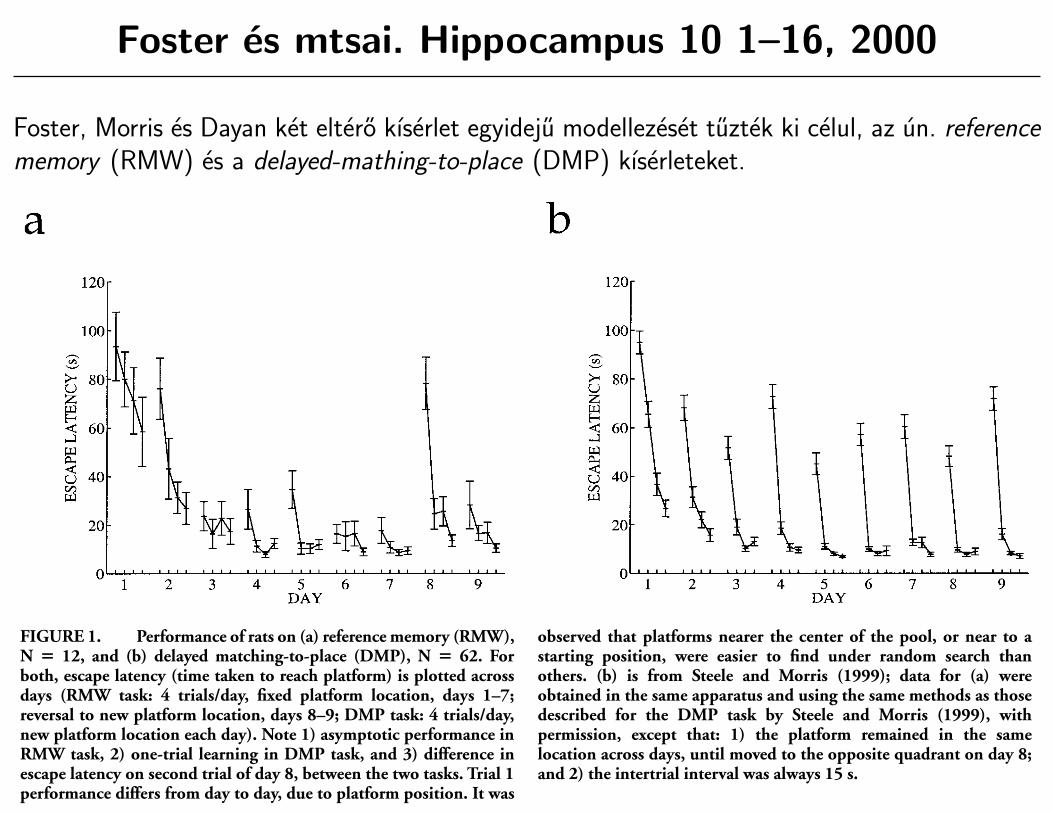
Foster es mtsai. Hippocampus 10 1–16, 2000
Foster, Morris es Dayan ket eltero kıserlet egyideju modellezeset tuztek ki celul, az un. referencememory (RMW) es a delayed-mathing-to-place (DMP) kıserleteket.
THE BEHAVIOR TO BE MODELED
We have chosen to model two behavioral tasks that are highlysensitive to hippocampal lesions and represent the kind ofnavigational problems for which TD learning, used in conjunc-tion with place cells, might provide a solution.
Reference memory in the watermaze (RMW) involves placingrats into a circular tank of water in which there is a hidden escapeplatform towards which they are highly motivated to swim(Morris, 1981). The tank itself affords no local cues as to theposition of the platform, and the use of a different startinglocation on each trial renders useless the strategy of replaying aseries of motor commands that worked previously. The rats mustlearn to navigate to the platform location from any possiblestarting position. Normal rats show more or less direct paths to theplatform after 20 trials, as implied by their short escape latencies(Fig. 1a, days 5–7). If the platform is then moved to a newlocation, performance is disrupted and animals take several trialsbefore they show direct paths to the new platform location (Fig.1a, days 8 and 9).
RMW has been modelled as an instance of conventionalreward-based learning using place cells (Brown and Sharp, 1995).However, the task presents a distal reward problem. We examine asimple TD-learning based ‘‘actor-critic’’ model of learning (Bartoet al., 1983, 1990), in which a set of place cells is associated with arepresentation of reward expectation, and also with a representa-
tion of action choice. Critically, the TD learning rule is used topredict rewards.
Delayed matching-to-place (DMP) is a new protocol for thewatermaze (Steele and Morris, 1999), though similar tasks havebeen explored (Morris, 1983; Panakhova et al., 1984; Whishaw,1985, 1991). As in RMW, rats are given several trials per day witha platform that stays in the same location throughout the day. Thecritical difference is that the platform is at a new and differentlocation on each day. Within each of the first few days, normal ratsshow a gradual decrease in the time taken to reach the platform(see Fig. 1b, days 1–5). A different pattern of escape latenciesemerges by about day 6. Rats by then show ‘‘one-trial learning,’’i.e., near-asymptotic navigational performance on the second trialof the day to a novel platform position.
DMP is computationally more demanding than RMW. UnlikeRMW, this task involves altering actions after only one trial ofexperience. It does not, however, only involve rapid learning, as isdemanded in a standard delayed match-to-sample task. DMP inthe watermaze is a complex navigation task in which a wholesequence of navigational actions has to be inferred from the singlelearning experience. This suggests that rats learn a representationof space that is goal-independent, which we model as a metriccoordinate system, learned from self-motion information. How-ever, previous attempts at modeling coordinate learning usingself-motion information encountered a global consistency prob-lem (Wan et al., 1994; Redish and Touretzky, 1997). In our
FIGURE 1. Performance of rats on (a) reference memory (RMW),N 5 12, and (b) delayed matching-to-place (DMP), N 5 62. Forboth, escape latency (time taken to reach platform) is plotted acrossdays (RMW task: 4 trials/day, fixed platform location, days 1–7;reversal to new platform location, days 8–9; DMP task: 4 trials/day,new platform location each day). Note 1) asymptotic performance inRMW task, 2) one-trial learning in DMP task, and 3) difference inescape latency on second trial of day 8, between the two tasks. Trial 1performance differs from day to day, due to platform position. It was
observed that platforms nearer the center of the pool, or near to astarting position, were easier to find under random search thanothers. (b) is from Steele and Morris (1999); data for (a) wereobtained in the same apparatus and using the same methods as thosedescribed for the DMP task by Steele and Morris (1999), withpermission, except that: 1) the platform remained in the samelocation across days, until moved to the opposite quadrant on day 8;and 2) the intertrial interval was always 15 s.
___________________________________________________ HIPPOCAMPALLY DEPENDENT NAVIGATION 3
– Typeset by FoilTEX – 13
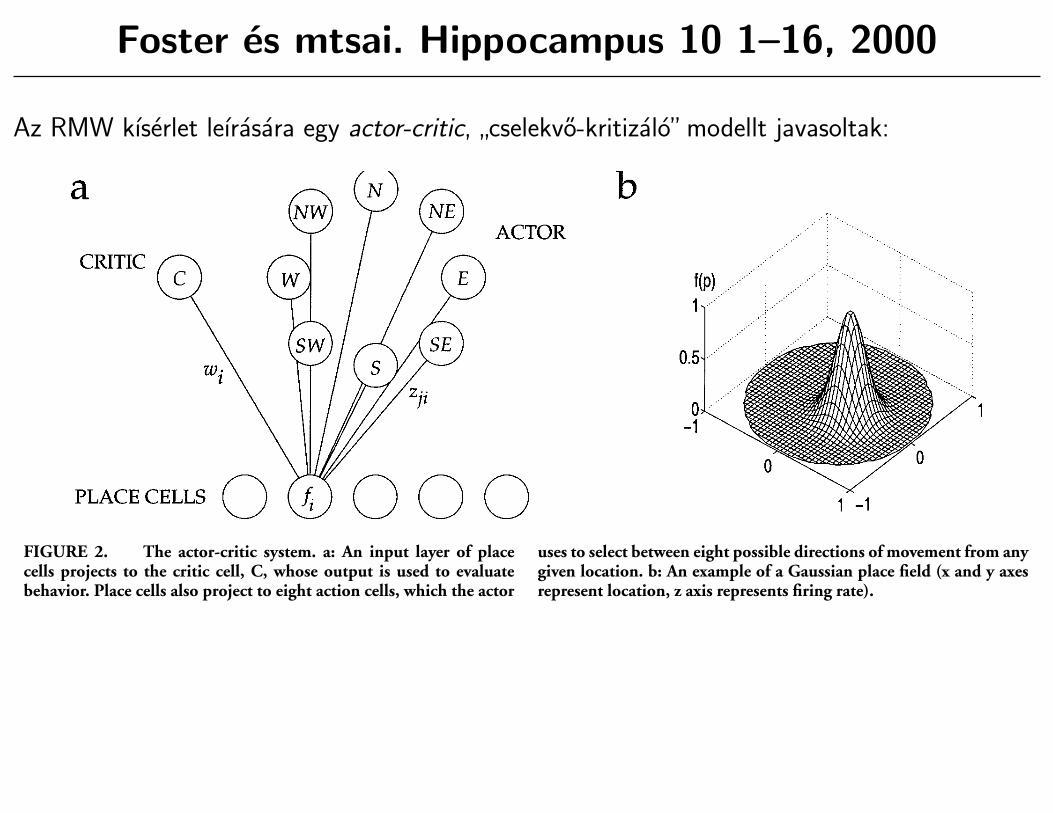
Foster es mtsai. Hippocampus 10 1–16, 2000
Az RMW kıserlet leırasara egy actor-critic,”cselekvo-kritizalo”modellt javasoltak:
model, TD learning is used, in association with a stable place cellrepresentation, to develop consistent coordinates directly.
The paper begins by presenting the reward-based component ofthe model, demonstrating that this component alone capturessome aspects of spatial learning, but not all. In particular, it doesnot capture the flexible way in which rats can learn about novelgoal locations. The second component of the model, the learnedcoordinate system, is then described, along with a simple way inwhich the components can be made to work together. Simulationresults are presented which capture performance in both RMWand DMP tasks. The discussion addresses the role of place cellswithin the model, what can be inferred from the model about thenature of the two tasks, and the relationship of the model toexperimental data and to other models of hippocampal function.Finally, a set of novel experimental predictions is presented.
REWARD-BASED NAVIGATION
Consider a simulated animal in an environment with control ofits own actions. At any given time t, the animal is able to choosean action. Also at any given time t, the environment provides theanimal with a reward Rt. If the animal moves onto the platform (acertain region of the environment) at time t, Rt 5 1; otherwiseRt 5 0. The difficult problem is to learn correct actions given sucha sparse reward signal.
To solve this problem we use an ‘‘actor-critic’’ architecture. Acomputational unit called the actor continually produces actions,taking a simulated animal around an environment. While it doesso, a second computational unit called the critic continuallycriticizes the actions taken. The actor adapts its action choicesusing the critic’s information. The critic also adapts in the light ofthe changing actor. The critic’s role is as a go-between, betweenthe actions on one hand, and the reward information on the other,
the latter being too sparse and uninformative to criticize the actordirectly.
Our implementation of the actor-critic has three parts (Fig. 2a):1) an input layer of place cells, 2) a critic network that learnsappropriate weights from the place cells to enable it to outputinformation about the value of particular locations, and 3) anactor network that learns appropriate weights from the place cellswhich enable it to represent the direction in which the rat shouldswim at particular locations.
Hippocampal Place Cells
Following experimental data (O’Keefe and Burgess, 1996), theactivities of place cells are modelled as Gaussian functions oflocation in the maze (Fig. 2b). If the rat is at position p, then theactivity of place cell i 5 1 . . . N is given by:
fi(p) 5 exp 12 \p 2 si \2
2s2 2 (1)
where si is the location in space of the center of cell i’s place field,and s is the breadth of the field, equivalent to the radius of thecircular contour where firing is 61% of the maximal firing rate.We consider an ensemble of place cells (N 5 493) with placefields distributed in an overlapping manner throughout the maze,each with width s 5 0.16 m.
Although clearly idealized, these place cells illustrate thelimitations pointed out in the Introduction: they are not intrinsi-cally informative about spatial or navigational quantities such asdistance or direction from a distant goal. However, such unitsform a basis function representation (e.g., Poggio and Girosi,1990) of location. As such, they would support the representationand learning of functions which vary (usually smoothly) withlocation. This paper explores this hypothesis, that hippocampalplace cells play the limited but nonetheless critical role ofproviding a particular representational substrate.
FIGURE 2. The actor-critic system. a: An input layer of placecells projects to the critic cell, C, whose output is used to evaluatebehavior. Place cells also project to eight action cells, which the actor
uses to select between eight possible directions of movement from anygiven location. b: An example of a Gaussian place field (x and y axesrepresent location, z axis represents firing rate).
4 FOSTER ET AL.
– Typeset by FoilTEX – 14

Foster es mtsai. Hippocampus 10 1–16, 2000
A”kritikus”tuzelesi ratajat a helysejtek szinaptikus sulyokkal sulyozott tuzelesi ratajanak osszege
adja:
C(p) =∑
i
wifi(p),
ahol p az aktualis hely
A”kritikus”feladata, hogy megtanuljon egy minden adott p helyhez tartozo V (p) ertek fuggvenyt,
amely megadja, hogy ha az adott helyen allva koveti a rendszer a”cselekvo” lepeseit, mekkora
jutalmat varhat a jovoben:
V (pt) = 〈Rt + γRt+1 + γ2Rt+2 + . . . 〉 = 〈Rt〉+ γV (pt+1),
ahol Rt a t. idopillanatban kapott jutalom. Ha a”kritikus”ezt a fuggveny teljesen jol megtanulna,
akkor igaz lenne, hogy C(pt) = 〈Rt〉 + γC(pt+1). A szerzok az un. temporal difference (TD)tanulasi szabalyt hasznaljak arra, hogy a
”kritikus”egyre jobban tudja az ertek fuggvenyt:
δt = Rt + γC(pt+1)− C(pt)
∆wi ∝ δtfi(pt),
itt δt-t predikcios hibanak nevezik es C(p) idobeli eltereseibol adodik a fenti keplet szerint.
– Typeset by FoilTEX – 15
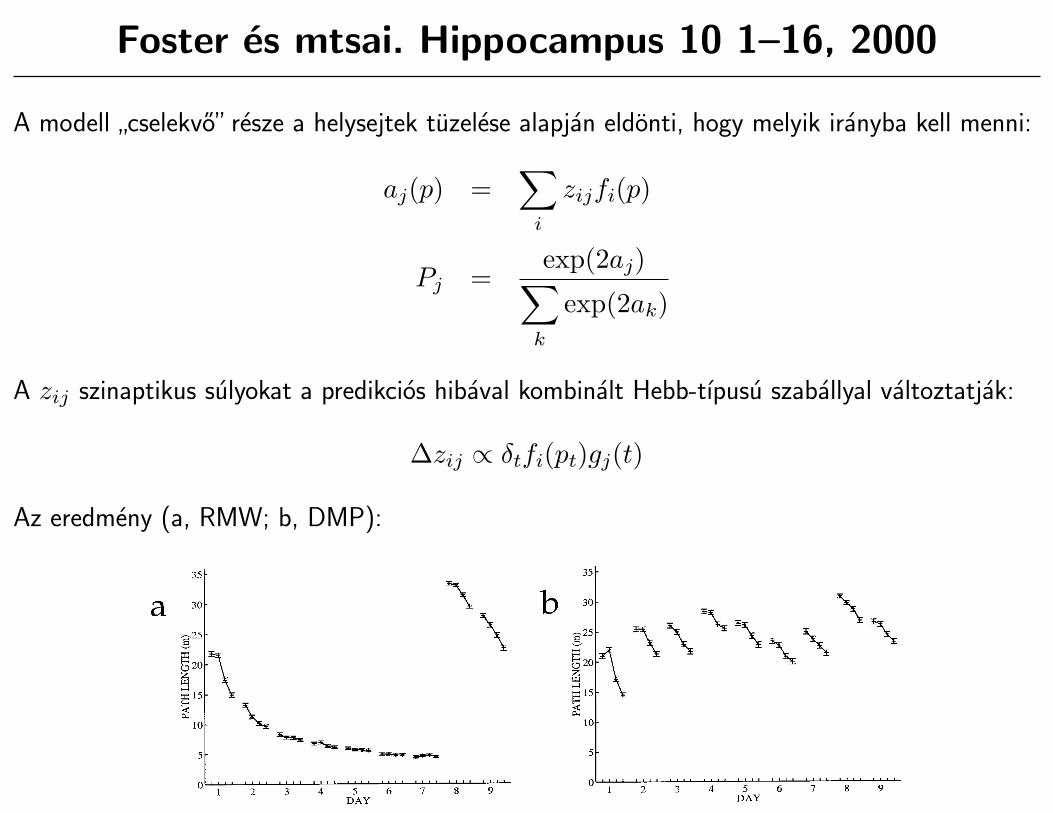
Foster es mtsai. Hippocampus 10 1–16, 2000
A modell”cselekvo” resze a helysejtek tuzelese alapjan eldonti, hogy melyik iranyba kell menni:
aj(p) =∑
i
zijfi(p)
Pj =exp(2aj)∑
k
exp(2ak)
A zij szinaptikus sulyokat a predikcios hibaval kombinalt Hebb-tıpusu szaballyal valtoztatjak:
∆zij ∝ δtfi(pt)gj(t)
Az eredmeny (a, RMW; b, DMP):
provides no mechanism by which the experience of previous dayscan provide any help with learning a new platform position.
One-trial learning by rats on DMP reveals that rats sufferneither of these limitations. Under appropriate training condi-tions, rats can not only avoid interference between training onsuccessive days, but can also generalize from experience on early
days to help performance on later days. To make this clear incomputational terms, consider trial 2 on day 6 of training (Fig.1b). The starting position may be in an area of the environmentnot explored on trial 1 of that day; nevertheless, the rat swimsimmediately to the platform. Clearly, knowledge from previousdays is being used.
FIGURE 3. Learning in the actor-critic system in RMW. For eachtrial, the critic’s value function C(p) is shown in the upper,three-dimensional plot; at lower left, the preferred actions at variouslocations are shown (the length of each arrow is related to theprobability that the particular action shown is taken by a logarithmicscale); at lower right is a sample path. Trial 2: After a timed-out firsttrial, the critic’s value function remains zero everywhere, the actions
point randomly in different directions, and a long and tortuous pathis taken to the platform. Trial 7: The critic’s value function havingpeaked in the northeast quadrant of the pool, the preferred actionsare correct for locations close to the platform, but not for locationsfurther away. Trial 22: The critic’s value function has spread acrossthe whole pool and the preferred actions are close to correct in mostlocations, and so the actor takes a direct route to the platform.
FIGURE 4. Performance of the actor-critic model. For each datapoint, the mean and standard error in the mean are obtained from1,000 simulation runs. (a) RMW task, in which the platformoccupies the same location. The actor-critic captures acquisition,producing direct paths after around 10 trials. For the last eight trials,however (days 8 and 9), the platform is moved to a different position(reversal), and the model fails to adapt rapidly enough. Thesesimulation results can be compared to Figure 1a. (b) DMP task, in
which the platform remains in the same position within a day, butoccupies a novel position on each new day. The actor-critic modelcaptures acquisition for the four trials of day 1, for which the task isindistinguishable from RMW. However, as a soon as the platform ismoved, the actor-critic not only fails to generalize to the new goallocation, but suffers from interference from the previous days’ goallocations. Rats suffer neither of these limitations (Fig. 1b).
___________________________________________________ HIPPOCAMPALLY DEPENDENT NAVIGATION 7
– Typeset by FoilTEX – 16
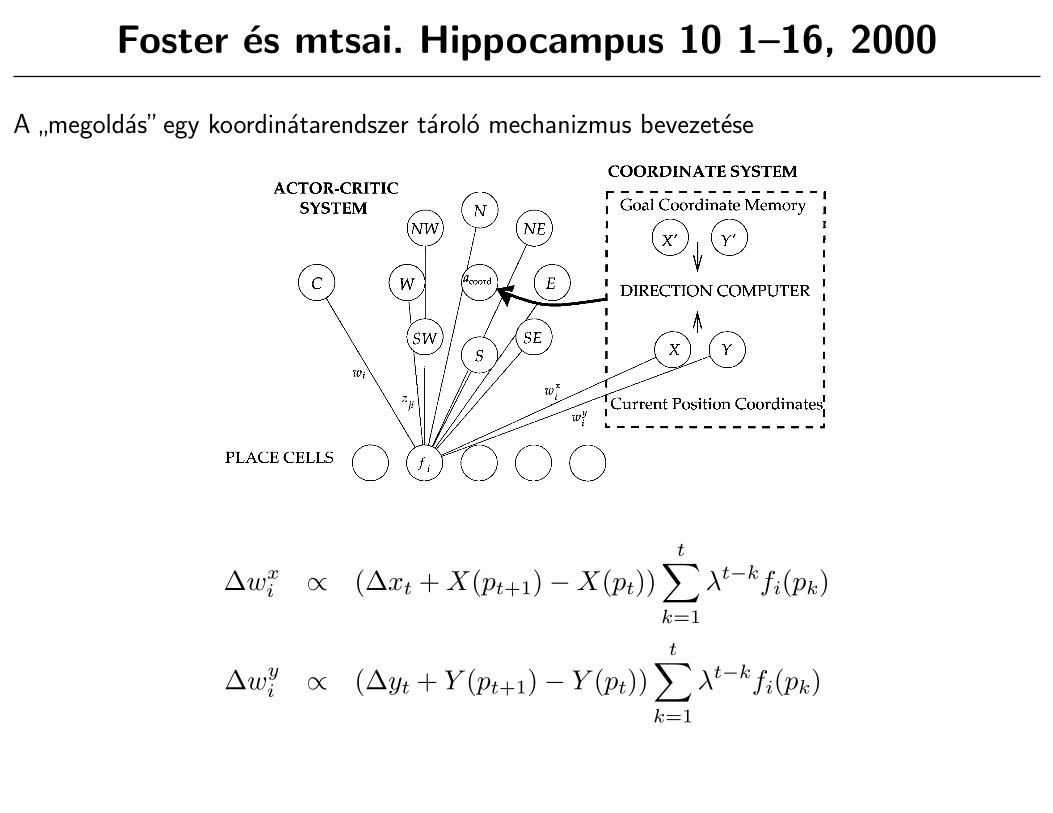
Foster es mtsai. Hippocampus 10 1–16, 2000
A”megoldas”egy koordinatarendszer tarolo mechanizmus bevezetese
Our model of coordinate learning is based on the observationthat the computations involved in the dead-reckoning abilities ofanimals could subserve an all-to-all navigation system for openspaces like a watermaze, if only the dead-reckoning coordinatescould be made to be consistent across separate trials, i.e., tied to anallocentric representation of the environment. In effect, weconsider making a dead-reckoning system hippocampal-depen-dent, i.e., dependent on input from the place cell system, andshow how such a system can be used to account for one-triallearning in the DMP task.
Dead-reckoning abilities have been documented in (at least)ants, bees, wasps, geese, gerbils, pigeons, rats, and humans(Gallistel, 1990). These abilities are based on the availability ofinstantaneous estimates of the animal’s self-motion, which can beintegrated in order to calculate the direction back to a startingpoint. We will make use of this self-motion information, and thesimple geometrical processing required to calculate a headingfrom the current position; however, we will not make use of pathintegration. Instead, we will use place cell responses and apredictive TD-based learning rule to acquire a coordinate systemin the maze which is defined allocentrically, i.e., independent ofthe animal’s point of origin.
It is hard to acquire an appropriate coordinate system usingpath integration information alone because of the problem ofconsistency. When the rat is put in the maze in a new place, thereis no way of ensuring that the dead reckoning coordinates itassigns are automatically consistent with those it has assigned
elsewhere in previous traversals of the maze. The essential task forthe model is learning this consistency (see also Wan et al., 1994).Note that the problem of having a consistent report of headdirection (implicitly required in the model) is quite similar.However, head direction generalizes over a much greater spatialextent than does dead reckoning, and, in the experiments beingmodeled, vestibular disorientation or other manipulations of thehead direction system were not used.
The problem for the rat is therefore to learn globally consistentcoordinates based only on local relative self-motion. The keyobservation is that for every move that the rat makes, thedifference between its estimates of coordinates at the ending andstarting locations should be exactly the relative self-motion duringthe move. This consistency condition can be used as the basis for aTD learning rule for learning coordinates.
Figure 5 shows a simple model of learning and using coordi-nates. The coordinate system consists of two networks, one whichlearns X coordinates (as X(p) 5 Siwi
X fi(p)), and one which learnsY coordinates (as Y(p) 5 Siwi
Y fi(p)), both using inputs from placecells which act in exactly the same way as in the actor-critic model,each producing a firing rate fi(p) as a function of location p. Thechoice of X and Y coordinates, or even just two orthogonaldirections, is of course arbitrary, but the basic problem of makingcoordinates consistent will exist whatever particular coordinatesystem is used. The X and Y coordinates have been chosen forsimplicity, and to illustrate clearly the learning problem.
FIGURE 5. The combined coordinate and actor-critic modelincorporates both the actor-critic system and a coordinate system.The coordinate system consists of three components: 1) a coordinaterepresentation of current position made up of two cells X and Y, thefiring of which is a function of place cell input; 2) a goal coordinatememory consisting of two cells, X8 and Y8, whose firing reflects the
coordinate location of the last place at which the platform was found;and 3) a mechanism which computes the direction in which to swimto get from the current position to the goal. The output directionfrom the coordinate system is integrated with that from the actor-critic through the ‘‘abstract action,’’ marked acoord, which receivesreinforcement depending on its performance.
8 FOSTER ET AL.
∆wxi ∝ (∆xt + X(pt+1)−X(pt))
t∑k=1
λt−kfi(pk)
∆wyi ∝ (∆yt + Y (pt+1)− Y (pt))
t∑k=1
λt−kfi(pk)
– Typeset by FoilTEX – 17
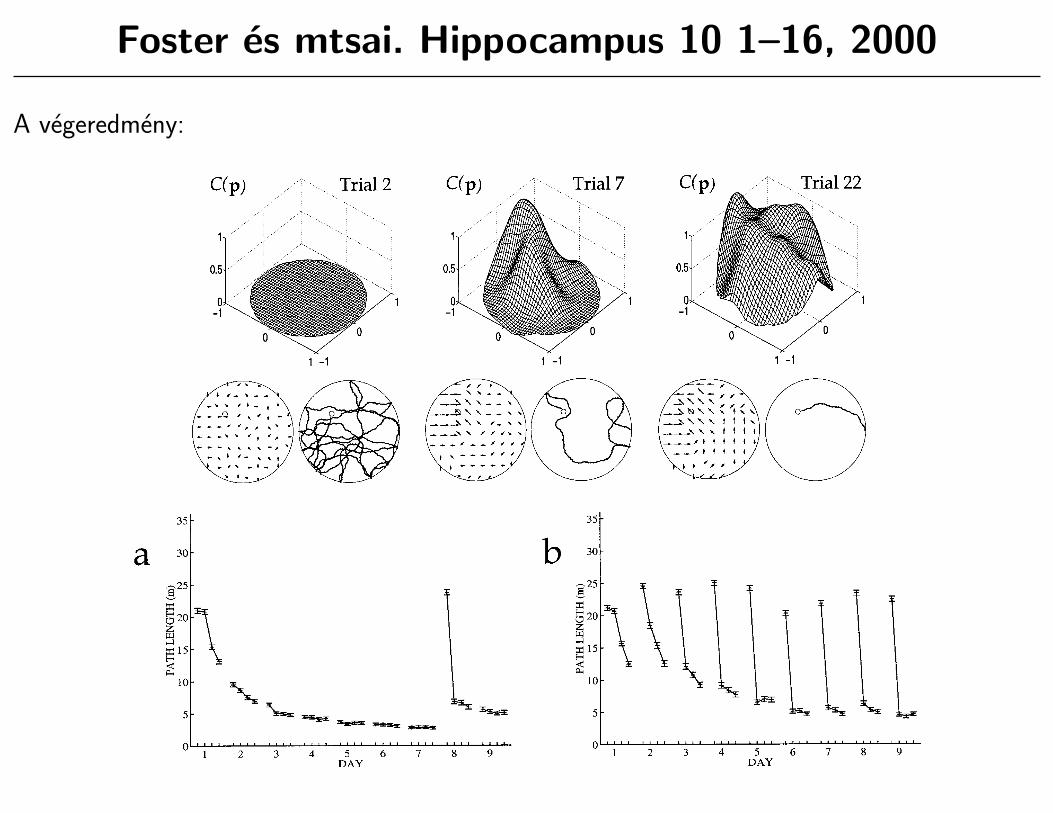
Foster es mtsai. Hippocampus 10 1–16, 2000
A vegeredmeny:
provides no mechanism by which the experience of previous dayscan provide any help with learning a new platform position.
One-trial learning by rats on DMP reveals that rats sufferneither of these limitations. Under appropriate training condi-tions, rats can not only avoid interference between training onsuccessive days, but can also generalize from experience on early
days to help performance on later days. To make this clear incomputational terms, consider trial 2 on day 6 of training (Fig.1b). The starting position may be in an area of the environmentnot explored on trial 1 of that day; nevertheless, the rat swimsimmediately to the platform. Clearly, knowledge from previousdays is being used.
FIGURE 3. Learning in the actor-critic system in RMW. For eachtrial, the critic’s value function C(p) is shown in the upper,three-dimensional plot; at lower left, the preferred actions at variouslocations are shown (the length of each arrow is related to theprobability that the particular action shown is taken by a logarithmicscale); at lower right is a sample path. Trial 2: After a timed-out firsttrial, the critic’s value function remains zero everywhere, the actions
point randomly in different directions, and a long and tortuous pathis taken to the platform. Trial 7: The critic’s value function havingpeaked in the northeast quadrant of the pool, the preferred actionsare correct for locations close to the platform, but not for locationsfurther away. Trial 22: The critic’s value function has spread acrossthe whole pool and the preferred actions are close to correct in mostlocations, and so the actor takes a direct route to the platform.
FIGURE 4. Performance of the actor-critic model. For each datapoint, the mean and standard error in the mean are obtained from1,000 simulation runs. (a) RMW task, in which the platformoccupies the same location. The actor-critic captures acquisition,producing direct paths after around 10 trials. For the last eight trials,however (days 8 and 9), the platform is moved to a different position(reversal), and the model fails to adapt rapidly enough. Thesesimulation results can be compared to Figure 1a. (b) DMP task, in
which the platform remains in the same position within a day, butoccupies a novel position on each new day. The actor-critic modelcaptures acquisition for the four trials of day 1, for which the task isindistinguishable from RMW. However, as a soon as the platform ismoved, the actor-critic not only fails to generalize to the new goallocation, but suffers from interference from the previous days’ goallocations. Rats suffer neither of these limitations (Fig. 1b).
___________________________________________________ HIPPOCAMPALLY DEPENDENT NAVIGATION 7
loops are possible. This difficulty motivates the combination ofthe coordinate control with the actor-critic, allowing the conven-tional actions of the actor-critic to dominate early on, butenabling coordinate control to come to dominate as its actionsprove more reliable than the conventional ones. This transfer ofcontrol happens rapidly during the DMP task (Fig. 6e).
Figure 8a shows the performance of the combined model in theRMW task. Like the actor-critic model discussed above, thecombined coordinate and actor-critic model successfully capturesthe acquisition of this task. Moreover, this model can also accountfor the rapid learning to the novel platform during the reversalphase, as seen in Figure 1a. Figure 8b shows the performance ofthe combined model in the DMP task. Just as in Figure 1b,acquisition during early days is gradual, while by day 6, one-triallearning is evident in the difference in performance between trials1 and 2.
DISCUSSION
A model of hippocampally dependent navigation has beenpresented that uses place cells as a representational substrate forlearning three different functions of position in an environment.The actor-critic component of the model learns the temporalproximity of locations to a single escape platform and alsoappropriate actions that get there quickly. By itself, the actor-criticmodel captures initial acquisition performance in RMW. How-ever, its performance diverges from that of rats the moment theplatform is moved, failing to account for the good reversalperformance shown by rats, or for the even more striking one-triallearning in DMP. A further component of the model learns X andY coordinates, a goal-independent representation of the environ-ment, and this provides the flexibility necessary for DMP by
FIGURE 7. Gradient of the coordinate functions. The gradient isa very sensitive measure of smoothness. On trial 4, coordinates arestill not at all smooth; navigation based on these functions alonewould be prone to catastrophic loops, i.e., would never reach theplatform. By comparison, the actor-critic scheme develops effective
values and actions for control by trial 4 (Fig. 3), and it is this controlthat allows the rat to move through the environment, and so improveits coordinate functions. By trial 36, coordinates are smoother andthe gradients reflect the X and Y directions.
FIGURE 8. Performance of the combined coordinate and actor-critic model. For each data point, the mean and standard error in themean are obtained from 1,000 simulation runs. (a) RMW task, inwhich the platform occupies the same location. The combined modelcaptures both acquisition, producing direct paths after around 10trials, and reversal, producing rapid adaptation to the change in
platform position on day 8 (see Fig. 1a). (b) DMP task, in which theplatform remains in the same position within a day, but occupies anovel position on each new day. The combined model captures theacquisition of one-trial learning: the improvement within each day isgradual early in training, but becomes a one-trial improvement byday 6. The model provides a good match to the data (Fig. 1b).
__________________________________________________ HIPPOCAMPALLY DEPENDENT NAVIGATION 11
– Typeset by FoilTEX – 18

Trullier es Meyer Biol. Cybern. 83 271–285, 2000
• Truller es Meyer modelljeben a hippokampusz a kornyezet”kognitıv grafjat” tanulja meg,
illetve hasznalja kesobb a navigaciohoz. Ez felfoghato ugy is, hogy a hippokampusz helyek(vagy esemenyek) szekvenciajat tanulja meg.
• A modellben meglehetos biologiai realisztikussaggal hasznalnak helysejteket, fej-irany sejteketes
”cel sejteket”. A navigacios modszerekhez altalaban szukseg van a pozıcio es az irany
reprezentalasara a rendszerben (helysejtek, fej-irany sejtek), illetve a”cel sejtekre”, melyek a
szinaptikus sulyok valtoztatasahoz, a cel adott helyhez viszonyitott iranyanak megtanulasahozkellenek.
• A modellben nincsenek komplikalt grafkereso algoritmusok.
• A helyfelismeresen alapulo navigaciot implementalo modellek korebe tartozik.
• A helysejtek fazis precessziojat mestersegesen allıtjak elo es a helymezobeli pontos helyetkodoljak vele, pontosabban a tanulasi algoritmusban hasznaljak.
• A modellben kihasznaljak a tuzelesi idoktol fuggo tanulasi mechanizmusokat: ha ket helysejta hippokampalis theta egymast koveto fazisaban tuzel, a koztuk levo kapcsolat erosodhet, hacsak az egyik tuzel, a kapcsolat gyengul.
– Typeset by FoilTEX – 19
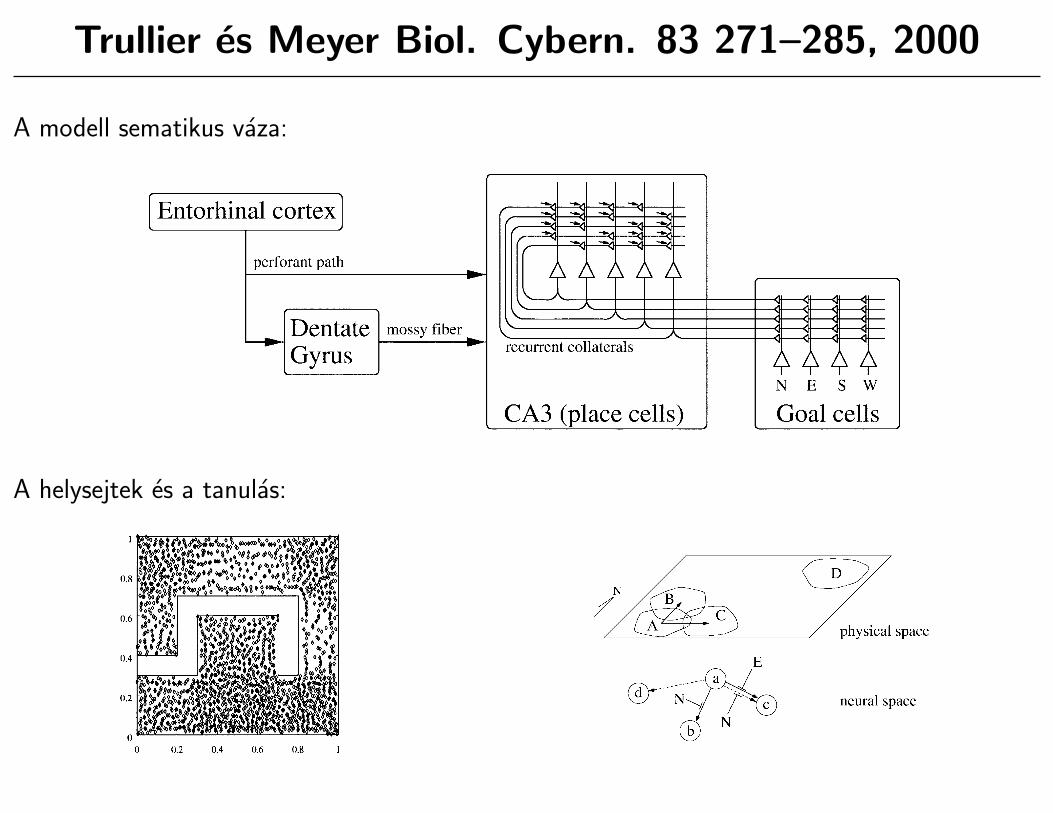
Trullier es Meyer Biol. Cybern. 83 271–285, 2000
A modell sematikus vaza:
�� �� ����� ������ ���� ��� �� �� ��� ��� �� ����� �� ����� � ������ ����� >� ��� � ���� �� ���� � ������ $���� ���� � ���� ��� � (��� �� ���������� �� � ���� ����� ��� $�.� � ������ �� ������ ���� �� �� ���� ���� >� �� $�� ����� �� �� ���������� ��� ��� (��� �� ������ ��� � � � �� �� ��� ��� $� ��.� ��� ���� �� ������� ���� ����������� ������ � ��� �� ���� ���� ��� �� $� ��������� ���� ��� �� �� ��� � �� ������� �������� (F��� C�.� I��� ������� ������� ���� ������ � ���� � �� ������� � ���� ��� � � ������� (�� �� �� � � � %����� � �� ',,C.�>� � ���� ���� � � �� � �� � ���� ���� �
�������� �� ����� �� ������ � �� ������ ��� ����� ����� � �� �� � ������ � �� ���� �����9��� � � �������� ��� ����� �� � ������ �� E ���(',,&. ��� ������ ������ ������ ��� �� �������� �� � ���7�� ��� 8���� (',,<.� >� ���� ����� ���� ��� ���� �� � ��� � ������� � ��� ����������� �� �� ��� ��� ���! $��� ����� ���� �� ������ ��� �� � ���� ��
�������� ��� ����� ��� ��� ��� ��� � ���� ������������ �� �� ���� ����� �� ������ �� �� � �� ���7 ��� ����������� E�� ��� ��������� � ����� � �� �������� � � ������ ����� � ������������ >� �� ������� ��� � ��� �� � ������������ � ������ �?
��� ��
�
��'
�� ���� � �
���:
�:
� �
��� � �� �� ����� � �������� ��� ����� ������ ��� �� �� �� �� ������ �� � �� ������� ������ ������ � ���� � � �� ������ ��� ������� �(�� � ��(������ �������. � ���� 3 � �� � '.� �
���� ��
��� ������ �� ��� � � �� ������� ���� �� ���� ���� �� ��� � ��� �� ������� ��� ������� ����� � �� � ������� ���� �� ��� ������� �� ��7 ��� ������� ��� $��� !�� � ������� �� � � �������� �� ���������� �� � �3�'� �� ��� �� ���� ����������� � � ���� ����� � �� ��� � �� � ��� � ������� �� ���� � �� $� � �� �������� ��� ��� >��������� �� � ���� �� �� ������� � �� ������������ >+� ��� �� ��� ������� ������� �� ��������� �� ��� � >+� � ���� ����� � �� �������������� � �� ���� ���� ������� �������� � ���������� � ���� ��!�� ��� �� ��� ���� $� ��� �� ���� ���
������� �� � �� ����� �� � �� ��� � �� �������� ���� ������ �� ������ ���� � (F��� A.� ������ ��� � �������� �� �������� ���� �� ��������� �� ����� ���������0 ��6��� �� ��� ����� ��� � ��� ���� ��
�� � �� ���� �� ��6� �� �� ��� ��� �� ��������� (F��� <.� #� � � ������� � �� �����
!��� "� #��������� � �� �� � �� � �� ��! ����� ���� ���� ����� � ��� ���� ��� ����� � �� �� �� ������ � �$������ � ��������� � �� ����� ����� ��� � �� �#& ��� � ��� ��� ����� ���� ����� �� ����� ������� �� �� � ���� ��� ����� � �� ��� ������ �� ��� �� �� � ���� ��� �� ��� � ���� ����� � �� ��� �� ����� � ������ ���� (� � ������� ������� �� ���������.� ���� ��� ����� � �� � ��� �� �� �#& �������� �� (���� �������. ��� ��� �� ��� �� � ��� $���� ��� �#& ��� �� ������ �� "��� �� (�������� ��� ��� �#' ��� ��������. �
������� � � ��� ����� ������� �� ������ ������� ��� ���������� � � � ������� ������� ���� ��� ������� �� ���� ������������ � ��� ����� �� � �!�� �� ��� � �� �� � ��� #� ��� %� �� �� �� ������ 0 ��� (��� .� ��� � # ��� ��� � %$� �� ��� (��� � ����� �������.� #� �� ��� ���� �� � ����� ��� � ��� � # � ��� � %� ���� �� 0 ��� ���������� ���� ����� � (����� ����� ������ �� � ������� ����. �� ���� ����(0 ��� �� ��������� ��� ���� � � ����� ������ � ������ �� ���������� � ��������� � ������.
:-C
A helysejtek es a tanulas:
!��� #�$�� #������� � � � ����� � ��� ��� � � ���� � ���� � � �� ������ ��� ��������������� I��� ������ ������� �� ������� ���� � � � ��� � (� ���� � � ����������� �.� � #� ��� ������� � L ' ��� L :� $� ��� �� (��� �������. ��� � �������� $� �� �� �� � � � ����� �� ������� ���� �� �� � ��� � �� ������ � ���� � � �������� (����.� �� �� ������ �� � � �������� �;��� ������ ��� �� ��"����� � ���� �� ������$� ��� $� ������ � ��� �������� ����� (������� �� � �� ����� �����. �� ��� ��������� ����$� ����� � $���� (������ ������ �������.� �� ���� ���������� ������ �� � & � '� � 9������� ������ � � ����� �� ������ ���� ��� � ���� �� �������� ����� ��������� �� � � � ����� �� ���� ����� ��� �� ����� �� � ��� �� ����� ��������
!��� %� 8�������� � �� ��� � ��� ��� ������� �� ���� �������������� !�� ��� �� � � ��� ����� ��� �� � ������ �������� ���� � (��� ���� $� ��� �� ��� ���� ��������� �� �� ������ ����� �.� � �� ��� � �� �������� ���� �� ��� ��������� �� �������� (� �� �����. ���� �� ������ � ���� � � �������� (���� � . ��� ��� � �� ����� �������� (���� ������.
!��� &� � ������ � � �� ��� ���� �� ������� �� �� �� � ��� !�� E�� $�� �� ���� ��� ���������� �� �� ���� ������������ � � ���� ������� "�� E�� �� �� ������� �� �������� �� �� �� ���� ����� #� �� �� � �� �� ���� �� ���$�� �� ���������� �� �� ���� ���� ���� � ���� �� ���� � ��� ������ � �� ���� ���� ( ������.� ���� ��� � ��� ���������� �� �����
:-A
�������� �� $���� � ���� ����� � �� ����� �� �� � �� �� �� ������� � �� ������#� � ��������� �� ���� ���� ��� � ��������� ��
��� �� � �������� � ����� �������� ��� � � � ����������� � � ��� �� � �� � ������� �� �������� �� �� �� ���� ���� ��� �������� � � ��������� ��������� � ����� �� � �� �� � ������� � �� ���� ����� #� � � ������� � 6 ����� ��� � ��� ��� ��� ��� �� ��� ���� �� ��� ��������� � �� ������� �������� � �� �������� � ���� �� � ��� ���� �� ����� � ��� ��� ����� �� � �� � � �� 5 ���� � ��� ���� ������� � �������� �� � ��� � ��� � ��������$���� � � �������� �� �� � � �� ����������
��# ������� �������� � ��
�� �#& ��� � � �� ���� ������ ��� � ���������� � ��������� ���� ������ ������� (8 �',,A.� G�� 9�� M � �� ��� � � (',,A.� � ��������� ��� ������� ��� � ���� �� ���������� ���� ����� � ��� ���� �� �� ������ � � �������� �� � ������������ � � � � ����������� �� �� � ���������� � ��$���� � � �� � ����� �� ���� �� ���� ������ �� ����� �� �� ������ � � ���� ����� � �� ������ ���������� � ��� ����� �� ����� �� �������� �� �� ��� ���� ����� ������� � ���������� �� � ���� ����� � ��� $�� �� ������� ��� ��$�� 9������� ������ �� ������ ( ������� ������� � � G�E. �� � �� ��� ��� � ����������� ���� ����� �� �������� ����� (F��� &� � �� �� F��� C�.���� �� ������ ( ������ ������ � � G�+. ������ �� ��� � �� � ����������� � �� ����� ��� �� ��� �� ����� #�� ������� ��� �� ������� ������7�� � ��� ���� �N��� �� ���� ���� ��� $��� ���� ��� $�� �� ����� �� �� ���� ����� ������ ��F � �������� �� � ����� � ���� �� �� ��� ������������ ����� � ���� ��� � � ��� ��� � � ��F��� - ��� �� ������ ������ �� ���� � ����� �������� ��� �� � ��� $� #� � ���� ����� � �� � ��� $� %� � ���� ����� � �� �� �� ��� � ����
� �������� 9������� �� � ����� � ���� �� ������������� ��� ����� � ���� ��� � � ��� ���� � ��� �� ������� 5 ���� �� � ����� � ������ �� ��� ��������� ��� ����� � ���� � ��� � ����� �� � � �� ������ (�� �� ��� �� � �� � ������� ������ ������� ��� ��� �� �������� ��� ����� ��� $�� # ��� � ��� ������ � ���. ��� �� ����� �� ���� ��� � � ��� ��� � � ����� ������ ����� ��� $�� # ��� + �� � �������� �� � ��� ����>� �� ��� �� � ����� � � �#& ��� �� ����
���� ������� ������� ��� � ���� �� � ������������ ��� � �� �� ���� ��� ���� ������ ������ ���� � �� �� ��"���� / ���� ���� ���� ��� ������������ � ��� ����� �� >� ��� � ���� �� �� � � �� ���� ��������� � � �� ���� ���� (F��� 1�.�
��� $�� �������� � ��
9��� � � � ���� ��������� � ��� � ��� � �������� ���� ���� ��� � ����� �� � �� ������ ������� � ���� �� � ������ ������ �� E� ������ � ��������� ��� ��� � � � ���� ����� �� � �������� ������� �� ��� (��� ������B ',,' ��� 9����"�� ������� ',,:. � � ���� �������� 5 ���� �� �������� ����� ���� �� ����� �� ������ ���� �� � ��� �� ����� ����� ���� ����� � �������� ����������� ��� � ���� ������� �� � � ��� (����� ���� � � � �� ��� ��� ����� � �������� ����.� ��� ����� �� ������ � ����� �� � �� ������ ������� ��� ���� � � �������� � ���� ���� �� � ���� ��� �� ����� �� �� ������� � (� ��������� � ����� ��� �� �� � �������� ���� ��.�:
#��������� %����� � �� (',,C. ��� ����7� �������� � � ���� � � ��� � ������� � ������ ������� ���� � �� � � � � �� �������� � ���� ����� ����� � �� � �� ���� � �� ��� ����� �� ���� �� � �� ��� ��� F � �������� �� ��I������ � � � �� $� ��� �� ������ �� � �� ��� � ��� �� >� �� � � ��� �� � ��� %����� � �� ������� � ���� � ���� �� �� � ����� �� �� � ����$���� �� � �� � � ��� � ��� � ����$� ���� � ���� �� � ����� � ��� � (� � � ���� �.� ����������� �� � $����� ���� � ���� �� �� �� �� �� ������� ��� >� ��� � ��� ������ ��� � ��$���� � ��� � ����� �� ���� ��� �� ��� �� � ����� ����� � � � �� � ��� � ���� �� �� �� � ������ ���� #� � � ������� �� �� ��� �� ����$� �� � �� ����� ���� ��� ��� �� ���� ��� ���$�� ���� �� �� �� � �� ������� �� ��� �� �� ���� ���� �� ����� $��� � � � � � ��� ��� ��$�� �� �� ��� � �� ��� $�� � �� ��� �������� �� / �� �� � ����� �� � ����� �� ��� ������ ��� / ��� ���� �� �� �� ������ � �� ������ ��������� ���� ����� � �� � � ���� �� 5 ���� ��� � ��� ��� �� ���� $� �� � �� ���� ���� ���
!��� '� �� � ��$� � ����� � ���� �� ��� �� �� �������� � ���� ��� � �� ����� ���� �� � ���� ����� ��� $�� ������� �� ��� ���� �� ��� $� � �� � ����������� � �� �� �������� � � ���� ����� � �� ���������� � ���� � ����� ��� ����� �� ���� ����� � �� ��� $� � �� ����������� �
:F � � �������� � � �� �� ���� ���������� � ��� � ����� � ����� �� �� �� � ���� �� ���� �� ����� � �������� �� (',,-�. ��� � ����� � �� (',,-.�
:-<
– Typeset by FoilTEX – 20

Trullier es Meyer Biol. Cybern. 83 271–285, 2000
Felfedezes, a cel iranyanak hozzarendelese a helyhez es mozgas a cel fele
���� ��� $�� � �������� �� � � ���� � (������� ������ �� ������ � ��� � �� $�� �� �������� �� ������ �� � ���� ����� ��� $�.� >� ���� ���� �� ����� $���� � � � �� ���� � ���� �� ����� ������� �� � �� � � ����� �� ��7 � ���$�� �� ������ #� � � ������� �������� � ��������� ����� ������ �� � �� ��7 � �� ��� $����� �� ���� ���� ��� � � ��� � ���� ���� ������� �� � � � %����� � �� � � � � �� � ��������� ���� ����� ���� ���� ��� ���� � ������� �� ���� ���� � ���� � �� ������� F������ ��� �������� � � ����� ���� ������ �� �� � ���>� ���� � ��� � �� � � � � ���� �� ��������
� ����� �� ������ � ���� �� ������ � � � ��� #��� �� � � � %����� � �� (',,C.� �� ������� �� �$����� � � � ���� �� �� �� �� �� � ������ ��� >���� � ��� ������ ��� �� �������� � �� ��� ���������� �� � � ���� � / ��� �� ������ �������� �� / �� �� ������ ������ �� �#& ��� ������ ��� �� � ����� �� ���� �� ���� �� �� ���������������� � ��� ����� � ��� � � ����� �� �������������� �� �� � � ������ ����� �� �� ��� � ������ � ����� ���� �������� ������ ���� ��� �� � ���$� ��� ��� �� � �� � � ��� ������ �� � � ������� ��� ���� (F��� 1�� F��� ,.� %�� �� ��� �� ���������� ���� �� �� �� ����� �� �� ����� �� ������� ���� ��������� ��� ������� �� �� ����� ��� ���� ����� ��� � ���� ����� � �� � � (!�� ��F��� 1�.� ������� � ������� �� � �� � �� �� � ������ �� �� � ��� �������� ��� �� � �� �������� � � �� ������ ��� ��� � 5����� ������ ����� ��� ������ � � �� �� ������ � ����� #� � � �������� �� �������� ����� ���� � ���� ��� ���� � ���� � � � �� �� ����� ������ � � �� �������� �� � �� � � � �� ������ ���� � � �� ����� # ������ � �� � ����� ��� ����� � �� ���� ������� �� � �� F � �������� �� F��� ,� �� �� ������ ����� ���� &1 ���� (�� �� &,�� ���� � ��� � ���������� ��� � �� �������.� ����� �� � ����� ���� � �� ��� �� ��������� �� � � ���� � ����� ��I����� � � �� �� ������ &1 ����� ��� ��� ����� �� �� � �� ��� �� ��������� �� 0 ����I��� � ��� � �� ���� ���� �� ������ �� �� � �����
! ��� ���� �� �������� ��� ��� �� � ����� ��� ����� �� � �� � � � ��� �� � ��$�� �������� �� ���� �� � � �� 9��������� ��� � ����� ����� ��� �� �� �� �� � ������ �� $���� ��� � � �� >� ��� � ���� �� $���� � �� ��� �� ���� ����� � �� ���� � � �� ��� �� ������� ������ ����� � � �� ���� �� ������ ��� �� ������� �� ���� ����� � �� ���� � �� F � ��������F��� 1� �� �� � ��� � ���� ��� �� ��� ����� �������� ���� ��������� �� ��9 ����� ��� �� ��9 ����!����
!��� (�$�� 9������ � �� �������� � �� � �� �� ������� ���� �� �� �� ��� ��� ������ � ���� ����� � �� ���� � (�.���� ����� � ���� � � � ���� � (�.� ��� �� ����� � (�.� � #� �������� � �� ���� ��� � ����� �� ���� ��� �� �� � ��$�� �N�� �� � � ���� �����7��� � � �� � ���� ����� ���$��� +����� � ��� ����� � �� �� ������� � #� � � �� ����� ����� � � ���� ����� � �� ������ ��� �� �� ������ ��� ����� ���������� � ���� �������� # �� ��� �� ���� �� � ����$������� � ���� ����� � �� ������ � � ���� � �� ������� ����� ��� ��� ����� ���� � � � �� � I��� ��� � ������� ��$���� � �� ���� ����� � � ��� ��� � � �� � �� �� �� ����� �� �� ������ � � � � � ����� � �� � �� (! ��� �������� � �� � � �������� � ���� ��� #� � � ���� ��� �� ��� � � � ��������� �����7� �� �� ��� ���� ������� ��� ������ � � ��������� �� � �� �� ��������� � �� ����� ����� ���.
�
:--
���� ��� $�� � �������� �� � � ���� � (������� ������ �� ������ � ��� � �� $�� �� �������� �� ������ �� � ���� ����� ��� $�.� >� ���� ���� �� ����� $���� � � � �� ���� � ���� �� ����� ������� �� � �� � � ����� �� ��7 � ���$�� �� ������ #� � � ������� �������� � ��������� ����� ������ �� � �� ��7 � �� ��� $����� �� ���� ���� ��� � � ��� � ���� ���� ������� �� � � � %����� � �� � � � � �� � ��������� ���� ����� ���� ���� ��� ���� � ������� �� ���� ���� � ���� � �� ������� F������ ��� �������� � � ����� ���� ������ �� �� � ���>� ���� � ��� � �� � � � � ���� �� ��������
� ����� �� ������ � ���� �� ������ � � � ��� #��� �� � � � %����� � �� (',,C.� �� ������� �� �$����� � � � ���� �� �� �� �� �� � ������ ��� >���� � ��� ������ ��� �� �������� � �� ��� ���������� �� � � ���� � / ��� �� ������ �������� �� / �� �� ������ ������ �� �#& ��� ������ ��� �� � ����� �� ���� �� ���� �� �� ���������������� � ��� ����� � ��� � � ����� �� �������������� �� �� � � ������ ����� �� �� ��� � ������ � ����� ���� �������� ������ ���� ��� �� � ���$� ��� ��� �� � �� � � ��� ������ �� � � ������� ��� ���� (F��� 1�� F��� ,.� %�� �� ��� �� ���������� ���� �� �� �� ����� �� �� ����� �� ������� ���� ��������� ��� ������� �� �� ����� ��� ���� ����� ��� � ���� ����� � �� � � (!�� ��F��� 1�.� ������� � ������� �� � �� � �� �� � ������ �� �� � ��� �������� ��� �� � �� �������� � � �� ������ ��� ��� � 5����� ������ ����� ��� ������ � � �� �� ������ � ����� #� � � �������� �� �������� ����� ���� � ���� ��� ���� � ���� � � � �� �� ����� ������ � � �� �������� �� � �� � � � �� ������ ���� � � �� ����� # ������ � �� � ����� ��� ����� � �� ���� ������� �� � �� F � �������� �� F��� ,� �� �� ������ ����� ���� &1 ���� (�� �� &,�� ���� � ��� � ���������� ��� � �� �������.� ����� �� � ����� ���� � �� ��� �� ��������� �� � � ���� � ����� ��I����� � � �� �� ������ &1 ����� ��� ��� ����� �� �� � �� ��� �� ��������� �� 0 ����I��� � ��� � �� ���� ���� �� ������ �� �� � �����
! ��� ���� �� �������� ��� ��� �� � ����� ��� ����� �� � �� � � � ��� �� � ��$�� �������� �� ���� �� � � �� 9��������� ��� � ����� ����� ��� �� �� �� �� � ������ �� $���� ��� � � �� >� ��� � ���� �� $���� � �� ��� �� ���� ����� � �� ���� � � �� ��� �� ������� ������ ����� � � �� ���� �� ������ ��� �� ������� �� ���� ����� � �� ���� � �� F � ��������F��� 1� �� �� � ��� � ���� ��� �� ��� ����� �������� ���� ��������� �� ��9 ����� ��� �� ��9 ����!����
!��� (�$�� 9������ � �� �������� � �� � �� �� ������� ���� �� �� �� ��� ��� ������ � ���� ����� � �� ���� � (�.���� ����� � ���� � � � ���� � (�.� ��� �� ����� � (�.� � #� �������� � �� ���� ��� � ����� �� ���� ��� �� �� � ��$�� �N�� �� � � ���� �����7��� � � �� � ���� ����� ���$��� +����� � ��� ����� � �� �� ������� � #� � � �� ����� ����� � � ���� ����� � �� ������ ��� �� �� ������ ��� ����� ���������� � ���� �������� # �� ��� �� ���� �� � ����$������� � ���� ����� � �� ������ � � ���� � �� ������� ����� ��� ��� ����� ���� � � � �� � I��� ��� � ������� ��$���� � �� ���� ����� � � ��� ��� � � �� � �� �� �� ����� �� �� ������ � � � � � ����� � �� � �� (! ��� �������� � �� � � �������� � ���� ��� #� � � ���� ��� �� ��� � � � ��������� �����7� �� �� ��� ���� ������� ��� ������ � � ��������� �� � �� �� ��������� � �� ����� ����� ���.
�
:--
���� ��� $�� � �������� �� � � ���� � (������� ������ �� ������ � ��� � �� $�� �� �������� �� ������ �� � ���� ����� ��� $�.� >� ���� ���� �� ����� $���� � � � �� ���� � ���� �� ����� ������� �� � �� � � ����� �� ��7 � ���$�� �� ������ #� � � ������� �������� � ��������� ����� ������ �� � �� ��7 � �� ��� $����� �� ���� ���� ��� � � ��� � ���� ���� ������� �� � � � %����� � �� � � � � �� � ��������� ���� ����� ���� ���� ��� ���� � ������� �� ���� ���� � ���� � �� ������� F������ ��� �������� � � ����� ���� ������ �� �� � ���>� ���� � ��� � �� � � � � ���� �� ��������
� ����� �� ������ � ���� �� ������ � � � ��� #��� �� � � � %����� � �� (',,C.� �� ������� �� �$����� � � � ���� �� �� �� �� �� � ������ ��� >���� � ��� ������ ��� �� �������� � �� ��� ���������� �� � � ���� � / ��� �� ������ �������� �� / �� �� ������ ������ �� �#& ��� ������ ��� �� � ����� �� ���� �� ���� �� �� ���������������� � ��� ����� � ��� � � ����� �� �������������� �� �� � � ������ ����� �� �� ��� � ������ � ����� ���� �������� ������ ���� ��� �� � ���$� ��� ��� �� � �� � � ��� ������ �� � � ������� ��� ���� (F��� 1�� F��� ,.� %�� �� ��� �� ���������� ���� �� �� �� ����� �� �� ����� �� ������� ���� ��������� ��� ������� �� �� ����� ��� ���� ����� ��� � ���� ����� � �� � � (!�� ��F��� 1�.� ������� � ������� �� � �� � �� �� � ������ �� �� � ��� �������� ��� �� � �� �������� � � �� ������ ��� ��� � 5����� ������ ����� ��� ������ � � �� �� ������ � ����� #� � � �������� �� �������� ����� ���� � ���� ��� ���� � ���� � � � �� �� ����� ������ � � �� �������� �� � �� � � � �� ������ ���� � � �� ����� # ������ � �� � ����� ��� ����� � �� ���� ������� �� � �� F � �������� �� F��� ,� �� �� ������ ����� ���� &1 ���� (�� �� &,�� ���� � ��� � ���������� ��� � �� �������.� ����� �� � ����� ���� � �� ��� �� ��������� �� � � ���� � ����� ��I����� � � �� �� ������ &1 ����� ��� ��� ����� �� �� � �� ��� �� ��������� �� 0 ����I��� � ��� � �� ���� ���� �� ������ �� �� � �����
! ��� ���� �� �������� ��� ��� �� � ����� ��� ����� �� � �� � � � ��� �� � ��$�� �������� �� ���� �� � � �� 9��������� ��� � ����� ����� ��� �� �� �� �� � ������ �� $���� ��� � � �� >� ��� � ���� �� $���� � �� ��� �� ���� ����� � �� ���� � � �� ��� �� ������� ������ ����� � � �� ���� �� ������ ��� �� ������� �� ���� ����� � �� ���� � �� F � ��������F��� 1� �� �� � ��� � ���� ��� �� ��� ����� �������� ���� ��������� �� ��9 ����� ��� �� ��9 ����!����
!��� (�$�� 9������ � �� �������� � �� � �� �� ������� ���� �� �� �� ��� ��� ������ � ���� ����� � �� ���� � (�.���� ����� � ���� � � � ���� � (�.� ��� �� ����� � (�.� � #� �������� � �� ���� ��� � ����� �� ���� ��� �� �� � ��$�� �N�� �� � � ���� �����7��� � � �� � ���� ����� ���$��� +����� � ��� ����� � �� �� ������� � #� � � �� ����� ����� � � ���� ����� � �� ������ ��� �� �� ������ ��� ����� ���������� � ���� �������� # �� ��� �� ���� �� � ����$������� � ���� ����� � �� ������ � � ���� � �� ������� ����� ��� ��� ����� ���� � � � �� � I��� ��� � ������� ��$���� � �� ���� ����� � � ��� ��� � � �� � �� �� �� ����� �� �� ������ � � � � � ����� � �� � �� (! ��� �������� � �� � � �������� � ���� ��� #� � � ���� ��� �� ��� � � � ��������� �����7� �� �� ��� ���� ������� ��� ������ � � ��������� �� � �� �� ��������� � �� ����� ����� ���.
�
:--
– Typeset by FoilTEX – 21
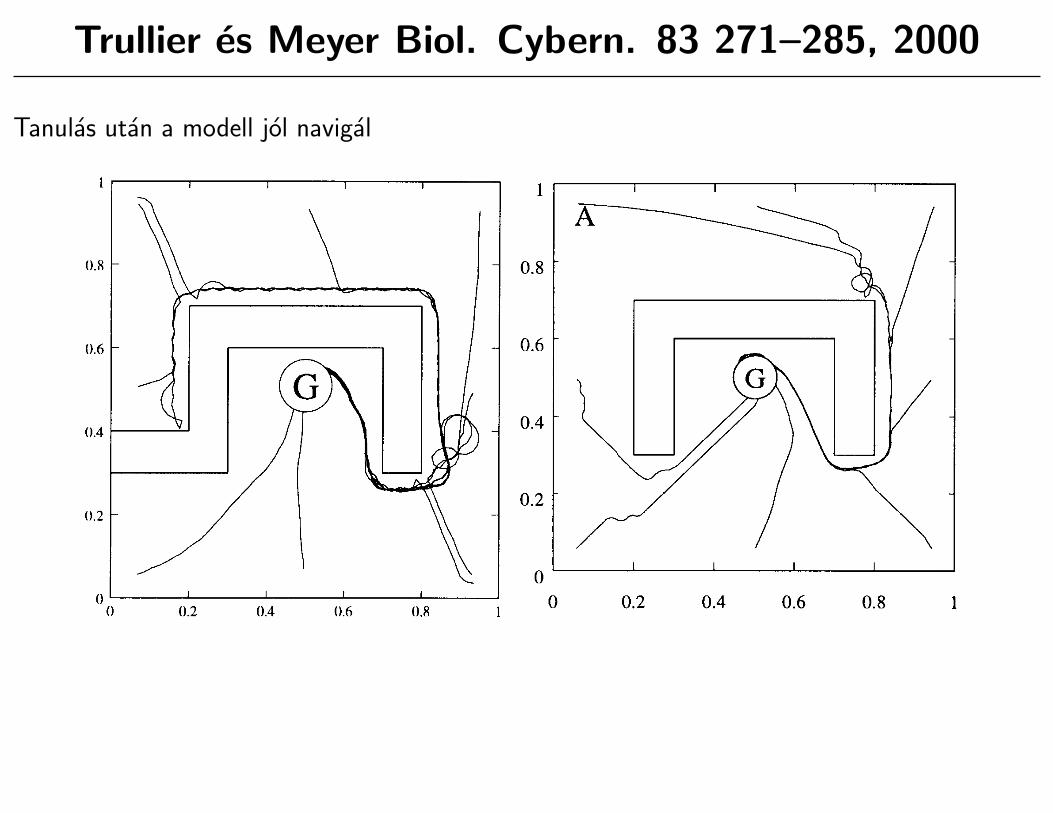
Trullier es Meyer Biol. Cybern. 83 271–285, 2000
Tanulas utan a modell jol navigal
��� ������ ������ ����� �� ��� � � ���� ���� � ���� � � � ����� �� �������� ����� ��������� ����� � �� ���� � ����������������� (*�����',1:.� ���� � ����������� �� ���� ��� � � ���7����� 8���� (',,<.� ����� ����� ���� ��� �� �� �������� � � �������� � ������ ������ 5 ���� �� � �� � ��� ���� �� �� ��� � ��������� ��� ���� �� ������ �� �� ��� ���� ����� ! �� � ��������� �� �� �� �� � �� �� 9��� � ���� ��� �0����� � (',,-. ��� ����� � ����� � ����� ������ ��� �� �� �$�� � ��� �� (������. � ������ � ����� � �������� � ��� �� ��� �� ����� �� ���� � � ��� � �� ���� � ���� �� �������� ����� (���� �������� �.� >� ���� ���� ���� ����� �� ��� ����� � ��� ����� ���� ��� ��� ������ �� � � � � ���� ������ ���$�� ��������������� ��! ��� ���� �� ���� ������ ������� ���
���� �� ������?
'� �� ���� ������ ���� �� ��6��� ������ ����� �� � � �$������ � (� ������� � � �� ��� �. ��� �����$� ��� � � ����$� ���
:� �� ��� �� �� � �� ������ ���� ������ ������6��� � �$������ ��
�� � � ������ �� � ���� � �� �� �� ��������� �� $��� ����� ��� ������ ��� ���� ��� ��� ���������� �� ����� ��� �� �� ������� �������� ���� ��
&�� ' ��� ���� � ���� ����� � �� ����������
9������ ��� ������� � ��� ��������� � ��� ������� ��������� � (���� ������ ���� ����.����� � � ����� �� �� ���� � ��� � ������� ��� �� � �� �� ���� ��� ����� ��� %������� ���� �� ���� �� � �� ���� ���� � ����� �� ������� ��� ����� ��� �� �������� �� � ������ ��� �� ����� ��� �� � � � ��$� �� � �������� �� �� ���� ��� �� � � ���� ���� ������ ��� �� �� ���� ���� (F��� '<.� >� ���� � ������� �� �� ��� �� � �� �� �� ���� ���� ���� ���� � �� ������� �� ���� ��� �� (�� �� ���� ���� ������.� �� �� ���� ���� ��� � ����� � ���
!��� %� ���"�� ��� ������ ��� ����� � ������ �� ��� ����� ����� (�� �. ��� ��������� � ���� �� � �� �������� ����� �� (�� .� ��� �� ����� ������� ��� ��� ���� ����� �� � (���.� ��� (�� ��� �� ��. �� ������ ������ 0 � ���� �� ���� �� ���� ���� � ���������� ������ Q������ �������� ������ ��� ������� � ������� � � � �� (J� '� :���� &.� �� ��� � ��� ������ � ����� � �� ' ������� ����� � � �� ���� ��� ��� � � ��� �� �� ����� � � � �� J� #� �� ������� � �� ����� �� :��� & �� ��� ����� ���� ����� � '� !�� �� ������ �� �� ��0 ����!�� � ��� � �� ����� ���� (#.� �� � �� � ���������� � & ��� ��� � ����� �'� ������ � ����� � �� �������� ��� ���� � �� ��� ��� ��� ������ >���� ��� ������������ ��������� � ������������ �' �& �� �� ��� �����' �:� ���� ������� ��� ������ � �� ���� ����� ��� �� ������ �� ������ � ������ �� ���� ������ � ����� � � �� ���� ����� � ��� �� ��� ������� � (� ��� � � ����.
:1:
��� ������ ������ ����� �� ��� � � ���� ���� � ���� � � � ����� �� �������� ����� ��������� ����� � �� ���� � ����������������� (*�����',1:.� ���� � ����������� �� ���� ��� � � ���7����� 8���� (',,<.� ����� ����� ���� ��� �� �� �������� � � �������� � ������ ������ 5 ���� �� � �� � ��� ���� �� �� ��� � ��������� ��� ���� �� ������ �� �� ��� ���� ����� ! �� � ��������� �� �� �� �� � �� �� 9��� � ���� ��� �0����� � (',,-. ��� ����� � ����� � ����� ������ ��� �� �� �$�� � ��� �� (������. � ������ � ����� � �������� � ��� �� ��� �� ����� �� ���� � � ��� � �� ���� � ���� �� �������� ����� (���� �������� �.� >� ���� ���� ���� ����� �� ��� ����� � ��� ����� ���� ��� ��� ������ �� � � � � ���� ������ ���$�� ��������������� ��! ��� ���� �� ���� ������ ������� ���
���� �� ������?
'� �� ���� ������ ���� �� ��6��� ������ ����� �� � � �$������ � (� ������� � � �� ��� �. ��� �����$� ��� � � ����$� ���
:� �� ��� �� �� � �� ������ ���� ������ ������6��� � �$������ ��
�� � � ������ �� � ���� � �� �� �� ��������� �� $��� ����� ��� ������ ��� ���� ��� ��� ���������� �� ����� ��� �� �� ������� �������� ���� ��
&�� ' ��� ���� � ���� ����� � �� ����������
9������ ��� ������� � ��� ��������� � ��� ������� ��������� � (���� ������ ���� ����.����� � � ����� �� �� ���� � ��� � ������� ��� �� � �� �� ���� ��� ����� ��� %������� ���� �� ���� �� � �� ���� ���� � ����� �� ������� ��� ����� ��� �� �������� �� � ������ ��� �� ����� ��� �� � � � ��$� �� � �������� �� �� ���� ��� �� � � ���� ���� ������ ��� �� �� ���� ���� (F��� '<.� >� ���� � ������� �� �� ��� �� � �� �� �� ���� ���� ���� ���� � �� ������� �� ���� ��� �� (�� �� ���� ���� ������.� �� �� ���� ���� ��� � ����� � ���
!��� %� ���"�� ��� ������ ��� ����� � ������ �� ��� ����� ����� (�� �. ��� ��������� � ���� �� � �� �������� ����� �� (�� .� ��� �� ����� ������� ��� ��� ���� ����� �� � (���.� ��� (�� ��� �� ��. �� ������ ������ 0 � ���� �� ���� �� ���� ���� � ���������� ������ Q������ �������� ������ ��� ������� � ������� � � � �� (J� '� :���� &.� �� ��� � ��� ������ � ����� � �� ' ������� ����� � � �� ���� ��� ��� � � ��� �� �� ����� � � � �� J� #� �� ������� � �� ����� �� :��� & �� ��� ����� ���� ����� � '� !�� �� ������ �� �� ��0 ����!�� � ��� � �� ����� ���� (#.� �� � �� � ���������� � & ��� ��� � ����� �'� ������ � ����� � �� �������� ��� ���� � �� ��� ��� ��� ������ >���� ��� ������������ ��������� � ������������ �' �& �� �� ��� �����' �:� ���� ������� ��� ������ � �� ���� ����� ��� �� ������ �� ������ � ������ �� ���� ������ � ����� � � �� ���� ����� � ��� �� ��� ������� � (� ��� � � ����.
:1:
– Typeset by FoilTEX – 22





























