Embed Size (px)
Citation preview

Involvement of glycine receptor a1 subunits in cannabinoid-inducedanalgesia
Jieping Lu a, b, *, 1, Sijia Fan a, 1, Guichang Zou a, Yiwen Hou a, Tao Pan f, Weiwei Guo a,Lei Yao a, Feng Du a, Gregg E. Homanics g, Dan Liu a, f, Li Zhang e, Wei Xiong a, c, d, **
a Hefei National Laboratory for Physical Sciences at the Microscale, School of Life Sciences, University of Science and Technology of China, Hefei 230026,Chinab Department of Neurology, The First Affiliated Hospital of University of Science and Technology of China, Hefei 230001, Chinac Neurodegenerative Disorder Research Center, University of Science and Technology of China, Hefei 230026, Chinad Center for Excellence in Brain Science and Intelligence Technology, Chinese Academy of Sciences, Shanghai 200031, Chinae Laboratory for Integrative Neuroscience, National Institute on Alcohol Abuse and Alcoholism, National Institutes of Health, Bethesda, MD, USAf USTC-Ruida Joint Laboratory for Biomedical Research, Hefei 230000, Chinag Departments of Anesthesiology, Neurobiology, and Pharmacology & Chemical Biology, University of Pittsburgh School of Medicine, Pittsburgh, PA, USA
a r t i c l e i n f o
Article history:Received 11 October 2017Received in revised form24 January 2018Accepted 29 January 2018Available online 1 February 2018
Keywords:Glycine receptorAnalgesiaCannabinoidPotentiationInflammatory pain
a b s t r a c t
Some cannabinoids have been shown to suppress chronic pain by targeting glycine receptors (GlyRs).Although cannabinoid potentiation of a3 GlyRs is thought to contribute to cannabinoid-induced anal-gesia, the role of cannabinoid potentiation of a1 GlyRs in cannabinoid suppression of chronic pain re-mains unclear. Here we report that dehydroxylcannabidiol (DH-CBD), a nonpsychoactive cannabinoid,significantly suppresses chronic inflammatory pain caused by noxious heat stimulation. This effect mayinvolve spinal a1 GlyRs since the expression level of a1 subunits in the spinal cord is positively correlatedwith CFA-induced inflammatory pain and the GlyRs antagonist strychnine blocks the DH-CBD-inducedanalgesia. A point-mutation of S296A in TM3 of a1 GlyRs significantly inhibits DH-CBD potentiation ofglycine currents (IGly) in HEK-293 cells and neurons in lamina I-II of spinal cord slices. To explore thein vivo consequence of DH-CBD potentiation of a1 GlyRs, we generated a GlyRa1S296A knock-in mouseline. We observed that DH-CBD-induced potentiation of IGly and analgesia for inflammatory pain wasabsent in GlyRa1S296A knock-in mice. These findings suggest that spinal a1 GlyR is a potential target forcannabinoid analgesia in chronic inflammatory pain.
© 2018 Elsevier Ltd. All rights reserved.
1. Introduction
Cannabis has been used to relieve chronic pain in human forcenturies (Johnson et al., 2010; Martin-Sanchez et al., 2009; Murrayet al., 2007). Both D9-tetrahydrocannabinol (THC) and cannabidiol(CBD), the major psychoactive and nonpsychoactive components ofcannabis (Wang et al., 2008), are found to produce analgesic effectsin chronic pain (Costa et al., 2007; Johnson et al., 2013). While theprimary action site of THC in brain is cannabinoid receptor type 1
(CB1) (Kawamura et al., 2006), emerging evidence suggests thatsome of THC-induced analgesic effects are independent of CB1 re-ceptors (LedentOV et al., 1999; Costa et al., 2005). For instance, theanalgesic effect of THC remained intact in the tail-flick reflex (TFR)test in CB1 receptor knock-out mice (Wilson and Nicoll, 2002), andthis THC-induced effect was not affected by intrathecal injection ofa selective CB1 receptor antagonist either (Hohmann et al., 2005).In addition, CBD could also induce analgesic effects without acti-vating CB1 receptors (Costa et al., 2007; Izzo et al., 2009; Long et al.,2006). These observations suggest that there are additional targetsother than CB1 receptors contributing to cannabis analgesia.
Emerging evidence has suggested that some cannabinoids candirectly or indirectly act on the glycine receptor (GlyR), an inhibi-tory ion channel, through a CB receptor-independentmechanism inthe central nervous system (Anderson et al., 2009; Hejazi et al.,2006; Lozovaya et al., 2005; Xiong et al., 2011). Molecular cloning
* Corresponding author. School of Life Sciences, University of Science and Tech-nology of China, Hefei 230026, China.** Corresponding author. School of Life Sciences, University of Science and Tech-nology of China, Hefei 230026, China.
E-mail addresses: [email protected] (J. Lu), [email protected] (W. Xiong).1 J.L. and S.F. contributed equally to this work.
Contents lists available at ScienceDirect
Neuropharmacology
journal homepage: www.elsevier .com/locate/neuropharm
https://doi.org/10.1016/j.neuropharm.2018.01.0410028-3908/© 2018 Elsevier Ltd. All rights reserved.
Neuropharmacology 133 (2018) 224e232

has identified four isoforms of the GlyR a subunit (a1-4) and asingle isoform of the b subunit (Betz and Laube, 2006). In general,these GlyRs are functional as pentameric assemblies containinghomomeric a subunits or heteromeric a/b subunits (Lynch, 2009).In adults, a1 and a3 are the most abundant GlyR subunits in thespinal cord (Shiang et al., 1995; Robert Harvey et al., 2004). The a1GlyR is considered to be critical for neuromotor activity sinceseveral naturally occurring point-mutations that cause functionaldeficiency of a1 GlyRs are closely associated with hyperekplexia, anexaggerated startle disease, in human (Zhou and ChillagNigro,2002). Meanwhile, the a3 GlyR, which is abundantly expressed inthe superficial laminae of spinal cord dorsal horn, plays an impor-tant role in the suppression of inflammatory-induced hypersensi-tivity to pain (Robert Harvey et al., 2004).
Previous studies have demonstrated that cannabinoid potenti-ation of GlyR a3 subunits contributes to cannabinoid-inducedanalgesia. For instance, Dehydroxylcannabidiol (DH-CBD), achemically modified cannabinoid, can alleviate acute pain andchronic inflammatory pain by targeting spinal a3 GlyRs (Xionget al., 2012a). Additionally, cannabinoids can also act on GlyR a1subunits (Foadi et al., 2010; Xiong et al., 2012b), which are localizedin brain areas participating in pain transmission such as brainstemand spinal cord (Baer et al., 2009; McDearmid et al., 2006; Piechottaet al., 2001), suggesting that GlyR a1 subunits may also be involvedin cannabinoid analgesia. However, it's very challenging to directlyexplore the role of a1 GlyR in the sensation of pain because of thelack of subunit-specific agonists and antagonists. Geneticallyengineered animal models have been used and shown to be valu-able for identifying the physiological roles of targeted genes in vivo.The previous studies revealed that a serine at 296 (S296) in thethird transmembrane domain of the a1 GlyR is critical for canna-binoid potentiation (Xiong et al., 2011). To investigate the role ofcannabinoid-a1 GlyR interaction in chronic pain, we have gener-ated an GlyRa1S296A mutation knock-in mouse line. Using electro-physiological recordings and behavioral measurements, weexamined the efficacy of DH-CBD potentiation of IGly in spinal slicesof these mutant mice and their sensitivity to DH-CBD-inducedanalgesic effect in chronic pain.
2. Methods
2.1. Animals
C57BL/6 female mice used in the experiments were obtainedfrom Vital River Laboratory Animal Technology Co., Ltd. (Beijing,China) at the age of 8 weeks. A Strain 129SvJ mouse genomic DNAbased gene targeting construct (see Fig. 5) was used to convertserine at position 296 to alanine and to introduce a silent mutationthat adds a NdeI restriction site for genotyping. This construct waslinearized and transfected into R1 mouse embryonic stem cells(Nagy et al., 1993) as previously described (Homanics et al., 1997).G418 resistant embryonic stem cell clones were analyzed for genetargeting by Southern blot analysis (details available from G.E.H.).Correctly targeted clones were microinjected into C57BL/6J (Jack-son Laboratory, Bar Harbor, ME, USA) blastocysts to producegermline competent chimeric founder mice. Chimeric males werebred to C57BL/6J females to produce mice that harbored the Aneoallele (see Fig. 5B). Aneo animals were subsequently crossed toC57BL/6J FLPe expressing transgenic mice (Rodriguez et al., 2002)to remove the drug selectable neomycin cassette. The FLPe trans-gene was removed from the pedigree by backcrossing to C57BL/6J.Heterozygous (S/A) breeders were intercrossed to produce wildtype (S/S), heterozygotes, and homozygous knock-in (A/A) animals.Genotyping was performed on tail snips using following primers:forward:50-GAATCTTCCAGGCAACATTTCAG-30, reverse:50-
AGTATCCCACCAAGCCAGTCTTT-3’. After weaning, mice werehoused in an animal room with ad libitum access to rodent chowand water with 12-h light/dark cycles (lights on at 8:00 a.m.). Allmice were between 8 and 12 weeks of age for behavioral experi-ments. Each mouse was used for only one experiment, and all micewere DH-CBD-naïve at the start of each experiment. Only femalemice were used for experiments. Male mice are not applicable inhot-plate test because that the skin of scrotum of male mice is moresensitive than the paw to thermal pain (B€olcskei et al., 2010). Allexperiments were approved by the Institutional Animal Care andUse Committee at each university and conducted in accordancewith the University of Science and Technology of China guidelineswith regard to the use of animals in research.
2.2. Hot-plate test
Experiments were conducted during the light cycle (8 a.m.e8p.m.) in a separated and air conditioned (22 !C) testing room. Thehot-plate test was performed by using an electronically controlled25 * 37 cm hotplate analgesia meter (Hot/Cold Plate; Ugo Basile,Italy) heated to 52 !C (±0.1 !C). The female mice were individuallyplaced on the hot plate apparatus, and the latency of hind paw-flutter, -lift or -lick was taken as the index of nociceptivethreshold. This was simultaneously registered by two observers.The animals were taken off immediately after the first reaction orafter a maximum time of 30 s. After each test the arenawas cleanedwith 75% ethanol.
2.3. CFA-induced inflammation
First, animals were tested for nociceptive threshold in the hot-plate test. Following the establishment of baseline latencies allmice received a subplantar injection of 20 mL CFA (1:4 in saline) orvehicle controls (saline). Mice were re-tested for changes in noci-ceptive threshold before and 4 h, 24 h and 72 h after subplantarinjection.
2.4. Startle reflex test
The mice were placed in the chamber of the Med AssociatesStartle Reflex System (Med Associates Inc.) and allowed to habit-uate for 5min. Then the mice were given a test to measure theirstartle responses using 30 pseudorandom white noise startlestimuli (10 per intensity, 85 dB, 90 dB or 95 dB) with a variable58e63s inter-trial interval (ITI).
2.5. Rotarod test
To test motor coordination and equilibrium, animals wereplaced on the rotating rod of the rotarod apparatus (XR1514, Xin-ruan Inc., Shanghai, China) on two successive days. Before the firsttraining sessions, the mice were habituated staying on the sta-tionary rod for 2min. A total six trials of the rotarod test werecarried out using an accelerating protocol from 4 to 60 rpm in 300swith 20min ITI. Three training sessions every day were performedfor 2 days. The latency (sec) to slip off the rod was automaticallyrecorded (maximal trial duration: 300 s). After falling, the micewere immediately placed back into the home cage. When finished,the apparatus and testing area were cleaned with 75% ethanol.
2.6. Western blotting
The spinal cord tissues of lumbar level L3 to L5 from mice withCFA inflammation or vehicle controls were homogenized andsonicated followed by centrifugation at 10000g, 4 !C for 30min. The
J. Lu et al. / Neuropharmacology 133 (2018) 224e232 225

supernatants were diluted 1:1 with Laemmli 2X sample buffer anddenatured at 99 !C for 10min. The samples were separated on 12%SDS/PAGE gels. After electro-transferring to a 0.45 mm PVDFmembrane, the membrane was blocked in 5% BSA and blotted withfollowing antibodies: Anti-GlyRa1 (NB300-113, Novus, 1:1000) andanti-GAPDH (Millipore, 1:40000). Horseradish peroxidase (HRP)-conjugated secondary antibody (SA00001-2, proteintech, 1:5000)and chemiluminescent substrate (Thermo Fisher) were used todetect signals.
2.7. HEK 293 cell transfection and recording
HEK-293 cells were cultured at 37 !C in a humidified 5% CO2incubator in culture medium (Dulbecco's Modified Eagle Medium(DMEM) with penicillin (100 U/mL), streptomycin (100 mg/mL) and10% fetal bovine serum). Plasmids coding GlyR a1 subunits andgreen fluorescent protein (GFP) were co-transfected into HEK-293 cells in a 1:1 ratio with Lipofectamine™ 2000 transfection re-agent (Thermo Fisher) for 24e48 h. Transfected cells were thentreated with 0.25% trypsin 2 h before recording. Cells were liftedand recorded with external solution containing (in mM) 140 NaCl, 5KCl, 2.0 CaCl2, 1.0 MgCl2, 10 glucose and 10 HEPES (pH 7.4 withNaOH, ~320 mOsm with sucrose). Patch pipettes (3e5MU) werefilled with intracellular solution contained (in mM) 140 CsCl, 4MgCl2, 10 EGTA, 10 HEPES, 0.5 Na-GTP and 2Mg-ATP (pH 7.2 withCsOH, ~280 mOsm). Membrane currents were recorded in thewhole-cell configuration using an Axopatch 200B amplifier (AxonInstruments, Foster City, CA) at room temperature. Cells were heldat " 60mV unless otherwise indicated. Data were acquired usingpClamp 10.4 software (Molecular Devices, Sunnyvale, CA). Drugswere applied using a Warner fast-step stepper motoredriven sys-tem. The solution exchange time constants were~4ms for an openpipette tip and 4e12ms for whole-cell recording.
2.8. Spinal cord slices preparation and recording
Spinal cord slices at the L3-L5 level were prepared from wild-type (WT) and GlyRa1S296A mutant mice aged from P21 to P30.The spinal cord was quickly removed after decapitation andimmersed in preoxygenated cutting solution containing (in mM)194 sucrose, 30 NaCl, 26 NaHCO3, 1 MgCl2, 10 glucose, 4.5 KCl, and1.2 NaH2PO4, pH 7.4. Spinal cords were sliced (200 mM) using aVibratome (VT 1200S) and continually incubated with artificialcerebrospinal fluid (aCSF) containing (inmM) 124NaCl, 26 NaHCO3,4.5 KCl, 1.2 NaH2PO4, 1 MgCl2, 2 CaCl2, and 10 glucose (final pH, 7.4;
osmolality, 315 mOsm kg, bubbled with 95% O2/5% CO2). The sliceswere incubated at 32 !C for approximately 30min and then cooledto room temperature (21e23 !C) at least 30min before theexperiment.
Slices were transferred into the recording chamber andcontinuously perfused at 3e4mL/min with aCSF at 28 !C. Neuronsof the spinal cord lamina I-II were visualized using a fixed-stagemicroscope (BX50WI; Olympus) with differential interferencecontrast/infrared illumination. We obtained all whole-cell voltageclamp mode recordings using borosilicate glass pipettes (5e7MU)filled with internal solutions. The internal solution (pH 7.2) forwhole cell current recording contained (in mM) 140 CsCl, 4 MgCl2,10 HEPES, 2Mg-adenosine triphosphate (ATP), 0.5 Na-guanosine5antriphosphate (GTP), and 10 ethylene glycol-bis (b-aminoethylether)-N,N,N0,N'-tetraacetic acid (EGTA). GlyR-mediated currentswere recorded at a holding potential of " 60mV.
2.9. The synthesis of DH-CBD
DH-CBD was synthesized according to the procedure asdescribed previously (Wilkinson et al., 2013). A solution of 1S,4R-4-isopropenyl-1-methyl-cyclohex-2-en-1-ol (100.5mg, 0.66mM), m-pentylphenol (120.0mg, 0.73mM) and anhydrous magnesiumsulfate (100mg) in 6mL freshly distilled DCM was purged withnitrogen and cooled to " 78 !C. Boron trifluoride-diethyl etherate(7.4 mL, 0.059mM) was added slowly and the mixture warmedto " 10 !C over 2 h. After adding anhydrous sodium bicarbonate(30mg), the solution was filtered and the filtrate was condensed.The crude residue was purified by flash chromatography (60:1,petroleum spirits (40e60 !C): ethyl acetate) to yield DH-CBD(74.8mg, 38% yield, 99.66% purity from HPLC) as a colorless oil.[a]20D ¼ " 128.6! (c, 0.783, CHCl3); 1H NMR (400MHz, CDCl3):d 6.86 (1H, d), 6.63 (2H, m), 5.51 (1H, s, br), 5.44 (1H, s), 4.66 (1H, t),4.56 (1H, s), 3.42e3.37 (1H, m), 2.51 (2H, t), 2.31 (1H, m), 2.20 (1H,m), 2.06 (1H, dt), 1.84e1.68 (5H, m), 1.61e1.54 (5H, m), 1.35e1.25(4H, m), 0.88 (3H, t). m/z: HRMS (ESI) calculated for C21H31O([Mþ H]þ , 100%): 299.2375; Found, 299.2371 (Figs. S1e2). DH-CBDwas diluted directly in ethanol (30mg/mL) before further dilu-tion. The final ethanol concentration was <8mM, which did notsignificantly affect IGly. In behavioral tests, DH-CBD was furtherdiluted to a proper concentration in saline.
2.10. Data analysis
Statistical analysis of concentration-response data were
BA
0 10 20 30 40 50 605
10
15
20
time (min)
PWL
(s)
100 mg/kg
vehicle
50 mg/kg15 mg/kg
*****
i.p. DH-CBD
Day 0 Day 3
i.p. DH-CBD
0 20 40 6010
15
20
25
time (min)
PWL
(s)
ns***
intraplantar CFA
Fig. 1. Effects of DH-CBD on acute pain and inflammatory pain in the hot-plate test in mice. (A) The effects of various doses of DH-CBD (5e100mg/kg) on acute pain caused by hotplate during the 60min following i.p. injection. (B) The dose-dependent analgesic effects of DH-CBD (i.p., 5e100mg/kg) in mice three days after 20 mL CFA (1:4 in saline) pawinjection. PWL, paw withdrawal latency. Data are mean ± S.E.M. from 6 to 10 mice. **P < 0.01 (15 mg/kg vs vehicle), ***P < 0.001 (50 mg/kg vs vehicle, 100 mg/kg vs vehicle)compared with vehicle group based on two-way analysis of variance (ANOVA). ns, not significant (5 mg/kg vs vehicle, P ¼ 0.405).
J. Lu et al. / Neuropharmacology 133 (2018) 224e232226
performed with the use of the nonlinear curve-fitting programPrism (Graph-Pad Software). Data were fitted using the Hill equa-tion I/Imax ¼ Bottom þ (Top - Bottom)/((1 þ 10 (̂LogEC50 - Log[Agonist] % HillSlope)), where I is the current amplitude activatedby a given concentration of agonist, Imax is the maximum responseof the cell, and EC50 is the concentration eliciting a half-maximalresponse.
A one-way repeated analysis of variance (ANOVA) was used tocompare the data between time points in each group. Data fromdifferent drug groups were compared using a two-way mixedmodel ANOVA. The Tukey's honestly significant difference post-hoc test was used to compare specific data points in ANOVA.STATISTICA 6.0 software (StatSoft, Inc.) was used for analysis, anddata are expressed as mean± S.E.M.. P < 0.05 was consideredsignificant.
3. Results
3.1. DH-CBD suppresses chronic inflammatory pain
As a small molecule chemically modified from THC, DH-CBD canremarkably potentiate a1 GlyRs, but neither activate CB1 receptorsnor induce psychoactive side effects in animals (Xiong et al., 2014).This feature makes DH-CBD an appropriate tool to study thefunction of a1 GlyR in vivo. First, we examined the effect of DH-CBDon acute pain and CFA-induced chronic inflammatory pain in mice.DH-CBD did not affect acute thermal pain in the hot-plate test(Fig. 1A). Three days after subplantar injection, CFA caused pro-longed hypersensitivity to thermal pain in mice, reflected by asignificant decrease in paw withdrawal latency (PWL) upon expo-sure to 52 !C hot plate (Fig. 1B). Intraperitoneal (i.p.) administrationof DH-CBD caused a significant increase in the PWL. The magnitudeof DH-CBD-induced analgesic effect reached maximum 20min af-ter injection and vanished in 1 h. This analgesic effect was dose-
GlyR α1
CBA
36kD
48kD
0 4 24 720
5
10
15
20
time (hour)
PWL
(s)
*******
0 4 24 720.0
0.5
1.0
1.5
2.0
2.5
time (hour)
norm
aliz
ed
***
Vehicle
CFA4h 24h 72h
GAPDH
****
n=20
n=9
n=14
n=14
n=14
n=12
n=12
n=9
-10 20 50 80
0
20
40
60
changes in spinal α1 GlyRs expression (%)
cha
nges
in P
WL
(%)
r 2= 0.71
0h
4h
24h72hP < 0.0001
D
0 4 24 720.0
0.5
1.0
1.5
norm
aliz
ed
time (hour)
n=4
n=4
n=4
n=4
4h 24h 72h
pre
pre
0 4 24 720
5
10
15
time (hour)
n=6
n=6
n=6
n=5
PWL
(s)
36kD
48kD
GAPDH
GFE
GlyR α1
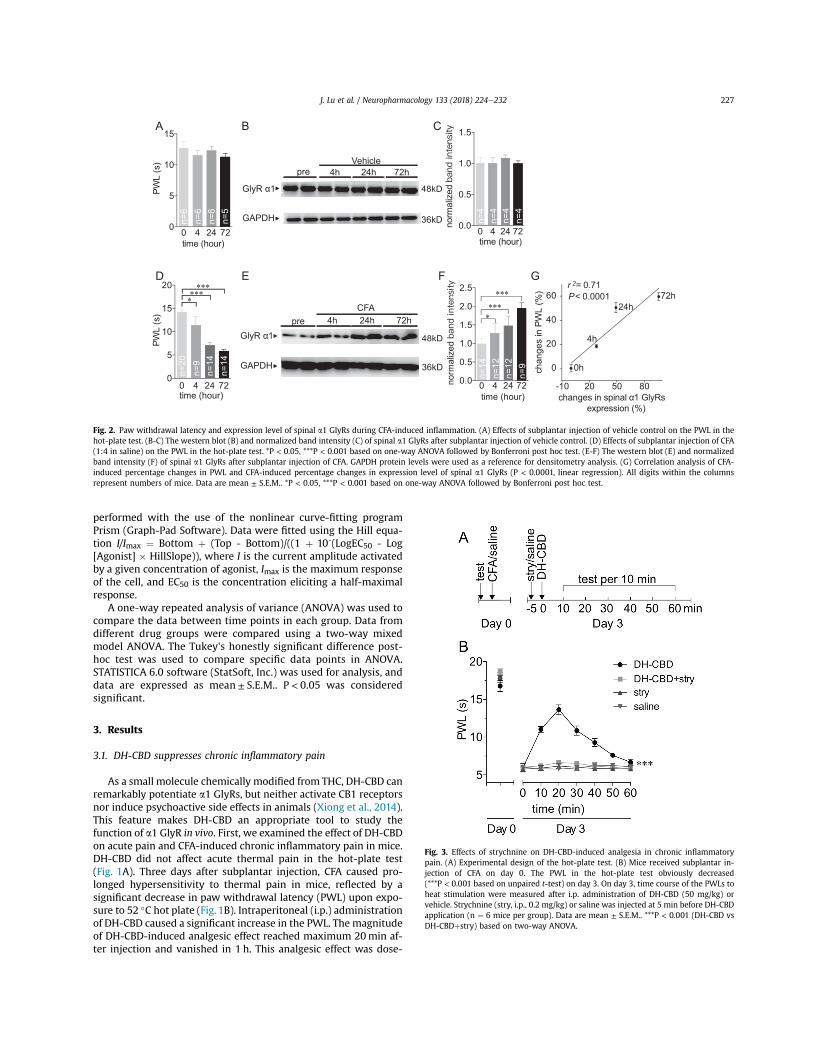
Fig. 2. Paw withdrawal latency and expression level of spinal a1 GlyRs during CFA-induced inflammation. (A) Effects of subplantar injection of vehicle control on the PWL in thehot-plate test. (B-C) The western blot (B) and normalized band intensity (C) of spinal a1 GlyRs after subplantar injection of vehicle control. (D) Effects of subplantar injection of CFA(1:4 in saline) on the PWL in the hot-plate test. *P < 0.05, ***P < 0.001 based on one-way ANOVA followed by Bonferroni post hoc test. (E-F) The western blot (E) and normalizedband intensity (F) of spinal a1 GlyRs after subplantar injection of CFA. GAPDH protein levels were used as a reference for densitometry analysis. (G) Correlation analysis of CFA-induced percentage changes in PWL and CFA-induced percentage changes in expression level of spinal a1 GlyRs (P < 0.0001, linear regression). All digits within the columnsrepresent numbers of mice. Data are mean ± S.E.M.. *P < 0.05, ***P < 0.001 based on one-way ANOVA followed by Bonferroni post hoc test.
Fig. 3. Effects of strychnine on DH-CBD-induced analgesia in chronic inflammatorypain. (A) Experimental design of the hot-plate test. (B) Mice received subplantar in-jection of CFA on day 0. The PWL in the hot-plate test obviously decreased(***P < 0.001 based on unpaired t-test) on day 3. On day 3, time course of the PWLs toheat stimulation were measured after i.p. administration of DH-CBD (50 mg/kg) orvehicle. Strychnine (stry, i.p., 0.2 mg/kg) or saline was injected at 5 min before DH-CBDapplication (n ¼ 6 mice per group). Data are mean ± S.E.M.. ***P < 0.001 (DH-CBD vsDH-CBDþ stry) based on two-way ANOVA.
J. Lu et al. / Neuropharmacology 133 (2018) 224e232 227
dependent when DH-CBD was injected at concentrations from 5 to100mg/kg (Fig. 1B). These results indicate that the analgesic effectof DH-CBD is specific to chronic inflammatory pain but not acutepain.
3.2. CFA-induced inflammatory pain promotes an up-regulation ofspinal a1 GlyRs
Numerous lines of evidences have indicated that there isremarkable alteration of protein expression levels in the spinal cordduring chronic inflammatory pain, including ion channels, re-ceptors, and enzymes (Clifford and Woolf, 2000; Luo et al., 2004;Honore et al., 2000; Raghavendra et al., 2004). Next we examinedwhether the protein expression level of a1 GlyR in spinal cordchanges during CFA-induced inflammation. First, the mice receivedan injection of 20 mL CFA (1:4 in saline) or vehicle control in thepaw. Then the hot-plate tests were performed each day for threedays. Our results showed that subplantar injection of vehicle con-trol affected neither pain threshold (Fig. 2A) nor expression levelsof spinal GlyR a1 subunits (Fig. 2B-C, Fig. S3) in mice. However, CFAinduced significant nociception after subplantar injection in a time-dependent manner (Fig. 2D). Consistently, western blot analysisrevealed that expression of a1 GlyRs in the spinal L3-L5 alsoincreased after CFA administration in a similar time-dependentmanner (Fig. 2E-F, Fig. S3). There was a strong correlation be-tween the CFA-induced nociception (% decrease in PWL) and theCFA-induced upregulation of spinal a1 GlyRs (% increase inexpression level) (Fig. 2G; r2¼ 0.71, P< 0.0001, linear regression),suggesting an involvement of spinal a1 GlyRs in the process of in-flammatory pain.
3.3. Strychnine abolishes the analgesic effect of DH-CBD oninflammatory pain
To examine the role of increased spinal GlyRs in DH-CBD-induced analgesic effect on inflammatory pain, we used strych-nine, a specific antagonist of GlyRs which can cross the blood-brainbarrier (Du et al., 2015; Huang et al., 2015; Schmieden et al., 1989).Three days after subplantar injection of CFA, mice received i.p.administration of strychnine (0.2mg/kg) followed by DH-CBD (i.p.,50mg/kg) 5min later (Fig. 3A). The hot-plate test indicated that thestrychnine completely abolished the antinociception induced byDH-CBD (Fig. 3B), suggesting that GlyR might be involved in theanalgesic effect of DH-CBD on inflammatory pain.
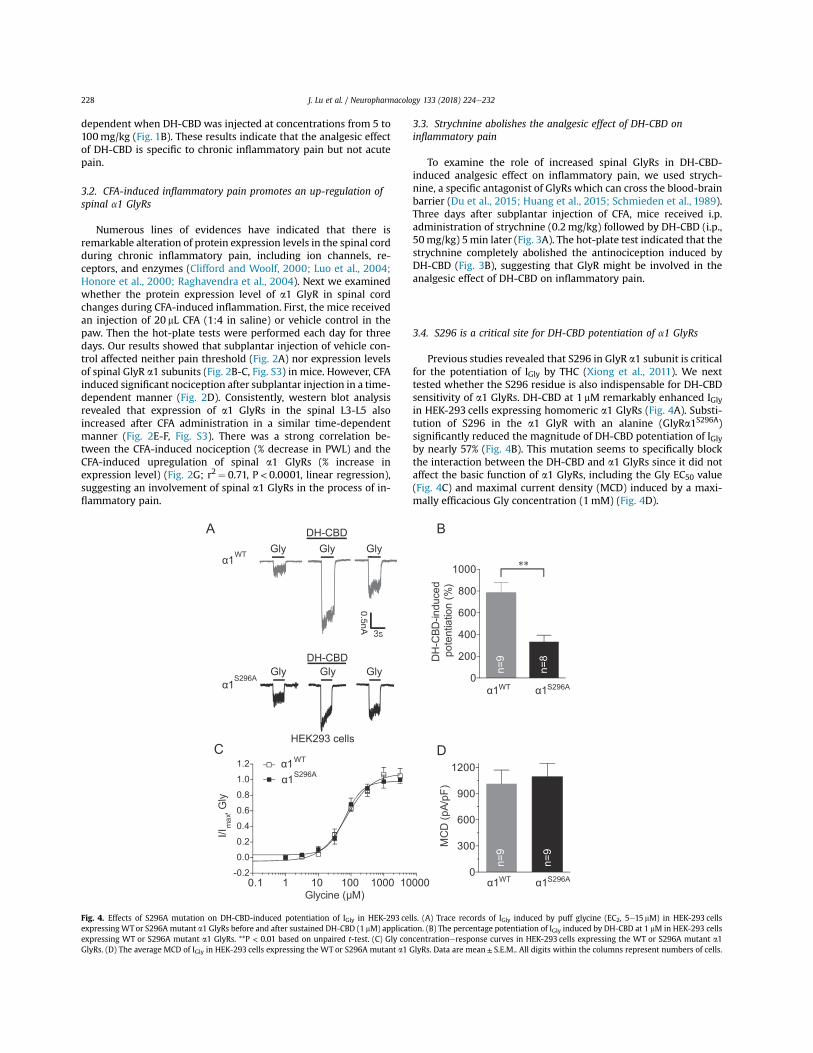
3.4. S296 is a critical site for DH-CBD potentiation of a1 GlyRs
Previous studies revealed that S296 in GlyR a1 subunit is criticalfor the potentiation of IGly by THC (Xiong et al., 2011). We nexttested whether the S296 residue is also indispensable for DH-CBDsensitivity of a1 GlyRs. DH-CBD at 1 mM remarkably enhanced IGlyin HEK-293 cells expressing homomeric a1 GlyRs (Fig. 4A). Substi-tution of S296 in the a1 GlyR with an alanine (GlyRa1S296A)significantly reduced the magnitude of DH-CBD potentiation of IGlyby nearly 57% (Fig. 4B). This mutation seems to specifically blockthe interaction between the DH-CBD and a1 GlyRs since it did notaffect the basic function of a1 GlyRs, including the Gly EC50 value(Fig. 4C) and maximal current density (MCD) induced by a maxi-mally efficacious Gly concentration (1mM) (Fig. 4D).
0
300
600
900
1200
MC
D (p
A/pF
)
HEK293 cells
S296Aα1
S296A
WTα1
WT
α1
α1
S296Aα1WTα1
S296Aα1WTα10
200
400
600
800
1000
DH
-CBD
-indu
ced
n=8
n=9
9=n9=n
pote
ntia
tion
(%)
**
C D
A B
0.1 1 10 100 1000 10000-0.2
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Glycine (μM)
I/Im
ax, G
ly
Gly Gly Gly
Gly Gly Gly
0.5nA
3s
DH-CBD
DH-CBD
0.5nA
3s
Fig. 4. Effects of S296A mutation on DH-CBD-induced potentiation of IGly in HEK-293 cells. (A) Trace records of IGly induced by puff glycine (EC2, 5e15 mM) in HEK-293 cellsexpressing WT or S296A mutant a1 GlyRs before and after sustained DH-CBD (1 mM) application. (B) The percentage potentiation of IGly induced by DH-CBD at 1 mM in HEK-293 cellsexpressing WT or S296A mutant a1 GlyRs. **P < 0.01 based on unpaired t-test. (C) Gly concentrationeresponse curves in HEK-293 cells expressing the WT or S296A mutant a1GlyRs. (D) The average MCD of IGly in HEK-293 cells expressing the WT or S296A mutant a1 GlyRs. Data are mean± S.E.M.. All digits within the columns represent numbers of cells.
J. Lu et al. / Neuropharmacology 133 (2018) 224e232228

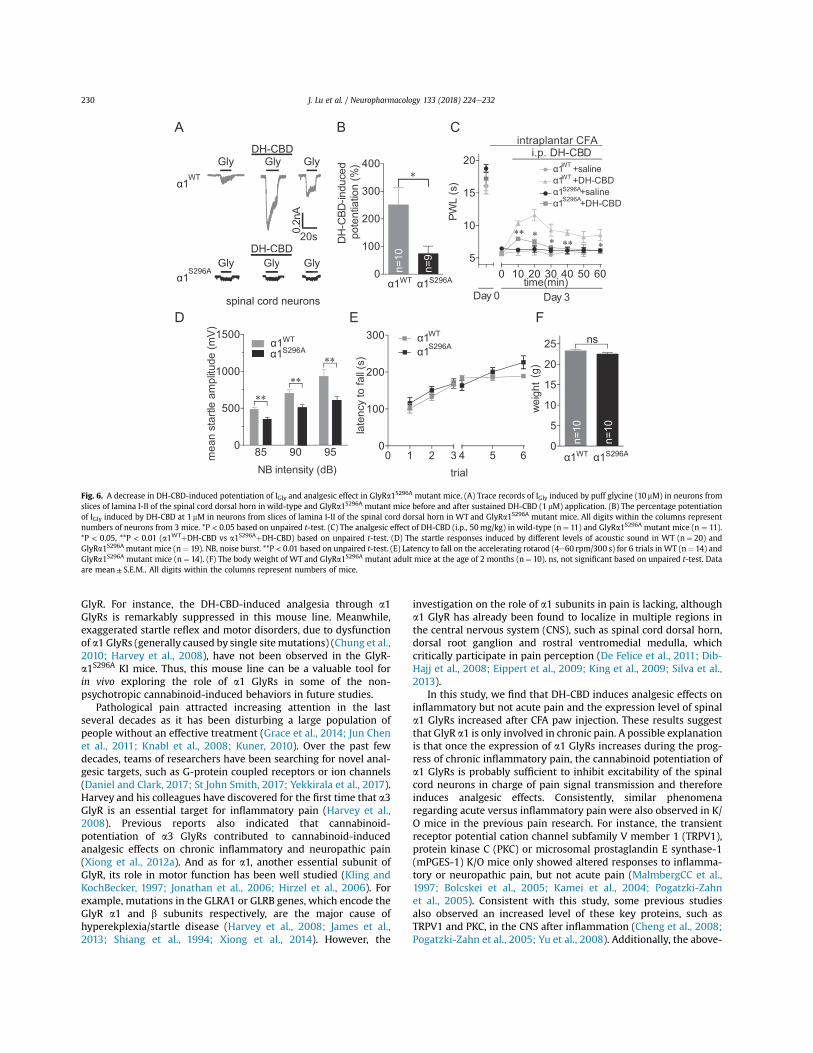
3.5. GlyR a1 S296A mutation reduces DH-CBD potentiation of IGly inspinal neurons and DH-CBD analgesia
To test whether DH-CBD-induced analgesic effects on inflam-matory pain are mediated by DH-CBD-induced potentiation of a1GlyRs, we generated a genetically engineered mouse line thatcarries the a1 GlyR S296A point mutation (Fig. 5).
First, we examined the effects of DH-CBD on GlyRs in spinalneurons of the GlyRa1S296A mutant mice. Consistently, DH-CBD at1 mM induced obvious potentiation of IGly evoked by puffing glycine(10 mM) in neurons from the slices of the lamina I-II of the spinalcord dorsal horn in the WT mice. This DH-CBD-induced potentia-tion was significantly suppressed in the GlyRa1S296A mutant mice(Fig. 6A-B), suggesting that this mutation indeed interrupted theinteraction between DH-CBD and spinal a1 GlyRs.
The hot-plate test indicated that the DH-CBD induced analgesiceffect was also significantly suppressed in the GlyRa1S296A mutantmice compared to the WT littermates (Fig. 6C). This effect is notlikely due to motor deficiency induced by a1 GlyR mutationbecause the mutant mice did not show an exaggerated startleresponse, a typical motor disorder generally detected in the micecarrying sitemutation of a1 GlyR (Zhou and ChillagNigro, 2002). Onthe contrary, the startle response of GlyRa1S296A KI mice was evenslightly lower than that of WT mice (Fig. 6D). Moreover, there was
no difference between GlyRa1S296A mutant and WT mice in themotor coordination ability measured by accelerating rotarod test(Fig. 6E). The body weight of these mutant mice appeared normalas compared to the WTmice (Fig. 6F). Overall, these results suggestthat a1 GlyRs are involved in cannabinoid-induced analgesia forchronic inflammatory pain.
4. Discussion
Our study provides evidence that GlyR a1 subunit is involved inDH-CBD-induced analgesia for inflammatory pain. First, DH-CBDsuppresses persistent inflammatory pain induced by subplantarinjection of CFA in mice. Such a suppression by DH-CBD is blockedby the GlyR antagonist strychnine. Second, the expression level ofspinal cord a1 GlyRs increases after CFA administration, suggestingan involvement of spinal a1 GlyRs during the progress of inflam-matory pain. Third, the analgesic effect of DH-CBD is suppressed inthe transgenic mice carrying GlyR a1 S296A mutation, which alsoblocks DH-CBD-induced potentiation of IGly in neurons of the spinalcord dorsal horn lamina I-II. Collectively, all these results suggestthat DH-CBD relieves inflammatory pain by targeting a1 GlyRs.
Additionally, this study develops a novel mouse line (GlyR-a1S296A) that specifically interrupts the interaction between DH-CBD and a1 GlyRs, but does not affect the basic function of a1
A
B
Fig. 5. Overview of gene targeting to produce GlyRa1S296A knock-in mice. (A) Partial DNA sequence of Exon 8 of WT (S296) and knock-in (A296) genes. S296 was converted to A296by a single nucleotide substitution (T to G). Also note that a silent T to A substitution that introduces a NdeI restriction endonuclease recognition site but does not change theisoleucine encoded at position 284 was also created for genotyping. (B) Gene targeting strategy used to modify the Glra1 locus in mouse embryonic stem cells. Exon 8 correspondsto nucleotides 913 to 1059 (nucleotide 1 corresponds to the start of translation) of the Glra1 cDNA (Ensembl Transcript ID: ENSMUST00000075603.4). The targeting vector was usedto create the Aneo allele in mouse embryonic stem cells. Following germline transmission of the Aneo allele, FLPe transgenic mice were used to remove the neomycin (NEO)resistance cassette from the genome and create the A296 knock-in allele.
J. Lu et al. / Neuropharmacology 133 (2018) 224e232 229
GlyR. For instance, the DH-CBD-induced analgesia through a1GlyRs is remarkably suppressed in this mouse line. Meanwhile,exaggerated startle reflex and motor disorders, due to dysfunctionof a1 GlyRs (generally caused by single sitemutations) (Chung et al.,2010; Harvey et al., 2008), have not been observed in the GlyR-a1S296A KI mice. Thus, this mouse line can be a valuable tool forin vivo exploring the role of a1 GlyRs in some of the non-psychotropic cannabinoid-induced behaviors in future studies.
Pathological pain attracted increasing attention in the lastseveral decades as it has been disturbing a large population ofpeople without an effective treatment (Grace et al., 2014; Jun Chenet al., 2011; Knabl et al., 2008; Kuner, 2010). Over the past fewdecades, teams of researchers have been searching for novel anal-gesic targets, such as G-protein coupled receptors or ion channels(Daniel and Clark, 2017; St John Smith, 2017; Yekkirala et al., 2017).Harvey and his colleagues have discovered for the first time that a3GlyR is an essential target for inflammatory pain (Harvey et al.,2008). Previous reports also indicated that cannabinoid-potentiation of a3 GlyRs contributed to cannabinoid-inducedanalgesic effects on chronic inflammatory and neuropathic pain(Xiong et al., 2012a). And as for a1, another essential subunit ofGlyR, its role in motor function has been well studied (Kling andKochBecker, 1997; Jonathan et al., 2006; Hirzel et al., 2006). Forexample, mutations in the GLRA1 or GLRB genes, which encode theGlyR a1 and b subunits respectively, are the major cause ofhyperekplexia/startle disease (Harvey et al., 2008; James et al.,2013; Shiang et al., 1994; Xiong et al., 2014). However, the
investigation on the role of a1 subunits in pain is lacking, althougha1 GlyR has already been found to localize in multiple regions inthe central nervous system (CNS), such as spinal cord dorsal horn,dorsal root ganglion and rostral ventromedial medulla, whichcritically participate in pain perception (De Felice et al., 2011; Dib-Hajj et al., 2008; Eippert et al., 2009; King et al., 2009; Silva et al.,2013).
In this study, we find that DH-CBD induces analgesic effects oninflammatory but not acute pain and the expression level of spinala1 GlyRs increased after CFA paw injection. These results suggestthat GlyR a1 is only involved in chronic pain. A possible explanationis that once the expression of a1 GlyRs increases during the prog-ress of chronic inflammatory pain, the cannabinoid potentiation ofa1 GlyRs is probably sufficient to inhibit excitability of the spinalcord neurons in charge of pain signal transmission and thereforeinduces analgesic effects. Consistently, similar phenomenaregarding acute versus inflammatory pain were also observed in K/O mice in the previous pain research. For instance, the transientreceptor potential cation channel subfamily V member 1 (TRPV1),protein kinase C (PKC) or microsomal prostaglandin E synthase-1(mPGES-1) K/O mice only showed altered responses to inflamma-tory or neuropathic pain, but not acute pain (MalmbergCC et al.,1997; Bolcskei et al., 2005; Kamei et al., 2004; Pogatzki-Zahnet al., 2005). Consistent with this study, some previous studiesalso observed an increased level of these key proteins, such asTRPV1 and PKC, in the CNS after inflammation (Cheng et al., 2008;Pogatzki-Zahn et al., 2005; Yu et al., 2008). Additionally, the above-
C
FD E
A B
Gly Gly GlyDH-CBD
Gly Gly GlyDH-CBD
DH
-CBD
-indu
ced
pote
ntia
tion
(%)
PWL
(s) α1 +DH-CBD
α1 +DH-CBDα1 +saline
Day 0 Day 3
85 90 950
500
1000
1500
NB intensity (dB)
mea
n st
artle
am
plitu
de (m
V)
trial
late
ncy
to fa
ll (s)
α1α1
S296A
WT
α1
α1
0
100
200
300
400*
intraplantar CFAi.p. DH-CBD
n=10
n=9
0
5
10
15
20
25
wei
ght
(g)
n=10
n=10
S296Aα1WTα1
S296Aα1WTα1
WTα1S296Aα1
WT
S296AS296A
WTS296A
spinal cord neurons
****
**
ns
0 10 20 30 40 50 605
10
15
20
time(min)
** ** ** *
α1 +salineWT
0 1 2 3 4 5 60
100
200
300
Gly Gly GDH-CBD
A
0.2n
A
20s
Fig. 6. A decrease in DH-CBD-induced potentiation of IGly and analgesic effect in GlyRa1S296A mutant mice. (A) Trace records of IGly induced by puff glycine (10 mM) in neurons fromslices of lamina I-II of the spinal cord dorsal horn in wild-type and GlyRa1S296A mutant mice before and after sustained DH-CBD (1 mM) application. (B) The percentage potentiationof IGly induced by DH-CBD at 1 mM in neurons from slices of lamina I-II of the spinal cord dorsal horn in WT and GlyRa1S296A mutant mice. All digits within the columns representnumbers of neurons from 3 mice. *P < 0.05 based on unpaired t-test. (C) The analgesic effect of DH-CBD (i.p., 50mg/kg) in wild-type (n¼ 11) and GlyRa1S296A mutant mice (n ¼ 11).*P < 0.05, **P < 0.01 (a1WTþ DH-CBD vs a1S296Aþ DH-CBD) based on unpaired t-test. (D) The startle responses induced by different levels of acoustic sound in WT (n¼ 20) andGlyRa1S296A mutant mice (n ¼ 19). NB, noise burst. **P < 0.01 based on unpaired t-test. (E) Latency to fall on the accelerating rotarod (4e60 rpm/300 s) for 6 trials in WT (n¼ 14) andGlyRa1S296A mutant mice (n¼ 14). (F) The body weight of WT and GlyRa1S296A mutant adult mice at the age of 2 months (n¼ 10). ns, not significant based on unpaired t-test. Dataare mean± S.E.M.. All digits within the columns represent numbers of mice.
J. Lu et al. / Neuropharmacology 133 (2018) 224e232230

mentioned feature of DH-CBD makes it a competitive candidateanalgesic as it can specifically relieve the chronic inflammatorypain without impairing the normal transmission of sensoryinformation.
Overall, the data presented in this study strongly suggest that inaddition to GlyR a3 subunit, the a1 subunit is also involved incannabinoid-induced antinociception against chronic inflamma-tory pain. The novel mechanism and therapeutic potential of DH-CBD-GlyR interaction revealed in this study open a new avenuefor developing analgesic agents by targeting GlyR a1 subunits.
Conflicts of interest
The authors state that there are no financial or other arrange-ments that might lead to a conflict of interest for this study.
Acknowledgements
We acknowledge support from the National Key Research andDevelopment Program of China (2016YFC1300500-2), NationalNatural Science Foundation of China (Grants 31471014, 91432103,91649121), the Strategic Priority Research Program of the ChineseAcademy of Sciences (Grant XDB02010000), the FundamentalResearch Funds for the Central Universities, the Major/InnovativeProgram of Development Foundation of Hefei Center for PhysicalScience and Technology, and Recruitment Program of Global Ex-perts, and the National Institutes of Health (Grant AA10422).
Appendix A. Supplementary data
Supplementary data related to this article can be found athttps://doi.org/10.1016/j.neuropharm.2018.01.041.
References
Anderson, W.B., Graham, B.A., Beveridge, N.J., Tooney, P.A., Brichta, A.M.,Callister, R.J., 2009. Different forms of glycine- and GABA(A)-receptor mediatedinhibitory synaptic transmission in mouse superficial and deep dorsal hornneurons. Mol. Pain 5, 65.
Baer, K., Waldvogel, H.J., Faull, R.L., Rees, M.I., 2009. Localization of glycine receptorsin the human forebrain, brainstem, and cervical spinal cord: an immunohis-tochemical review. Front. Mol. Neurosci. 2, 25.
Betz, H., Laube, B., 2006. Glycine receptors: recent insights into their structuralorganization and functional diversity. J. Neurochem. 97 (6), 1600e1610.
Bolcskei, K., Helyes, Z., Szabo, A., Sandor, K., Elekes, K., Nemeth, J., Almasi, R.,Pinter, E., Petho, G., Szolcsanyi, J., 2005. Investigation of the role of TRPV1 re-ceptors in acute and chronic nociceptive processes using gene-deficient mice.Pain 117 (3), 368e376.
B€olcskei, K., Peth}o, G., Szolcs"anyi, J., 2010. Noxious heat threshold measured withslowly increasing temperatures: novel rat thermal hyperalgesia models, 617,57e66.
Cheng, H.T., Suzuki, M., Hegarty, D.M., Xu, Q., Weyerbacher, A.R., South, S.M.,Ohata, M., Inturrisi, C.E., 2008. Inflammatory pain-induced signaling eventsfollowing a conditional deletion of the N-methyl-D-aspartate receptor in spinalcord dorsal horn. Neuroscience 155 (3), 948e958.
Chung, S.K., Vanbellinghen, J.F., Mullins, J.G., Robinson, A., Hantke, J.,Hammond, C.L., Gilbert, D.F., Freilinger, M., Ryan, M., Kruer, M.C., Masri, A.,Gurses, C., Ferrie, C., Harvey, K., Shiang, R., Christodoulou, J., Andermann, F.,Andermann, E., Thomas, R.H., Harvey, R.J., Lynch, J.W., Rees, M.I., 2010. Patho-physiological mechanisms of dominant and recessive GLRA1 mutations inhyperekplexia. J. Neurosci. 30 (28), 9612e9620.
Clifford, J., Woolf, M.W.S., 2000. Neuronal plasticity: increasing the gain in pain.Science 288 (5472), 1765e1768.
Costa, B., Trovato, A.E., Colleoni, M., Giagnoni, G., Zarini, E., Croci, T., 2005. Effect ofthe cannabinoid CB1 receptor antagonist, SR141716, on nociceptive responseand nerve demyelination in rodents with chronic constriction injury of thesciatic nerve. Pain 116 (1e2), 52e61.
Costa, B., Trovato, A.E., Comelli, F., Giagnoni, G., Colleoni, M., 2007. The non-psychoactive cannabis constituent cannabidiol is an orally effective therapeu-tic agent in rat chronic inflammatory and neuropathic pain. Eur. J. Pharmacol.556 (1e3), 75e83.
Daniel, J.T., Clark, R.J., 2017. G-protein coupled receptors targeted by analgesicvenom peptides. Toxins (Basel) 9 (11).
De Felice, M., Sanoja, R., Wang, R., Vera-Portocarrero, L., Oyarzo, J., King, T.,
Ossipov, M.H., Vanderah, T.W., Lai, J., Dussor, G.O., Fields, H.L., Price, T.J.,Porreca, F., 2011. Engagement of descending inhibition from the rostralventromedial medulla protects against chronic neuropathic pain. Pain 152 (12),2701e2709.
Dib-Hajj, S.D., Estacion, M., Jarecki, B.W., Tyrrell, L., Fischer, T.Z., Lawden, M.,Cummins, T.R., Waxman, S.G., 2008. Paroxysmal extreme pain disorder M1627Kmutation in human Nav1.7 renders DRG neurons hyperexcitable. Mol. Pain 4, 37.
Du, J., Lu, W., Wu, S., Cheng, Y., Gouaux, E., 2015. Glycine receptor mechanismelucidated by electron cryo-microscopy. Nature 526 (7572), 224e229.
Eippert, F., Finsterbusch, J., Bingel, U., Buchel, C., 2009. Direct evidence for spinalcord involvement in placebo analgesia. Science 326 (5951), 404.
Foadi, N., Leuwer, M., Demir, R., Dengler, R., Buchholz, V., de la Roche, J., Karst, M.,Haeseler, G., Ahrens, J., 2010. Lack of positive allosteric modulation of mutatedalpha(1)S267I glycine receptors by cannabinoids. Naunyn-Schmiedeberg’s Arch.Pharmacol. 381 (5), 477e482.
Grace, P.M., Hutchinson, M.R., Maier, S.F., Watkins, L.R., 2014. Pathological pain andthe neuroimmune interface. Nat. Rev. Immunol. 14 (4), 217e231.
Harvey, R.J., Topf, M., Harvey, K., Rees, M.I., 2008. The genetics of hyperekplexia:more than startle! Trends Genet. 24 (9), 439e447.
Hejazi, N., Zhou, C., Oz, M., Sun, H., Ye, J.H., Zhang, L., 2006. Delta9-tetrahydrocannabinol and endogenous cannabinoid anandamide directlypotentiate the function of glycine receptors. Mol. Pharmacol. 69 (3), 991e997.
Hirzel, Klaus, Muller, Ulrike, Latal, A. Tobias, Hulsmann, Swen, Grudzinska, Joanna,Seeliger, Mathias W., HB, Laube, aB., 2006. Hyperekplexia phenotype of Glycinereceptor a1 subunit mutant mice identifies Zn2þ as an essential endogenousmodulator of Glycinergic neurotransmission. Neuron 52 (4), 572e574.
Hohmann, A.G., Suplita, R.L., Bolton, N.M., Neely, M.H., Fegley, D., Mangieri, R.,Krey, J.F., Walker, J.M., Holmes, P.V., Crystal, J.D., Duranti, A., Tontini, A., Mor, M.,Tarzia, G., Piomelli, D., 2005. An endocannabinoid mechanism for stress-induced analgesia. Nature 435 (7045), 1108e1112.
Homanics, G.E., Ferguson, C., Quinlan, J.J., Daggett, J., Snyder, K., Lagenaur, C., Mi, Z.P.,Wang, X.H., Grayson, D.R., Firestone, L.L., 1997. Gene knockout of the alpha6subunit of the gamma-aminobutyric acid type A receptor: lack of effect onresponses to ethanol, pentobarbital, and general anesthetics. Mol. Pharmacol.51 (4), 588e596.
Honore, P., Rogers, S.D., Schwei, M.J., Salak-Johnson, J.L., NML, Sabino, M.C.,Clohisy, D.R., Mantyh, P.W., 2000. Murine models of inflammatory, neuropathicand cancer pain each generates a unique set of neurochemical changes in thespinal cord and sensory neurons. Neuroscience 98 (3), 585e598.
Huang, X., Chen, H., Michelsen, K., Schneider, S., Shaffer, P.L., 2015. Crystal structureof human glycine receptor-alpha3 bound to antagonist strychnine. Nature 526(7572), 277e280.
Izzo, A.A., Borrelli, F., Capasso, R., Di Marzo, V., Mechoulam, R., 2009. Non-psy-chotropic plant cannabinoids: new therapeutic opportunities from an ancientherb. Trends Pharmacol. Sci. 30 (10), 515e527.
James, V.M., Bode, A., Chung, S.K., Gill, J.L., Nielsen, M., Cowan, F.M., Vujic, M.,Thomas, R.H., Rees, M.I., Harvey, K., Keramidas, A., Topf, M., Ginjaar, I.,Lynch, J.W., Harvey, R.J., 2013. Novel missense mutations in the glycine receptorbeta subunit gene (GLRB) in startle disease. Neurobiol. Dis. 52, 137e149.
Johnson, J.R., Burnell-Nugent, M., Lossignol, D., Ganae-Motan, E.D., Potts, R.,Fallon, M.T., 2010. Multicenter, double-blind, randomized, placebo-controlled,parallel-group study of the efficacy, safety, and tolerability of THC: CBDextract and THC extract in patients with intractable cancer-related pain. J. PainSymptom Manag. 39 (2), 167e179.
Johnson, J.R., Lossignol, D., Burnell-Nugent, M., Fallon, M.T., 2013. An open-labelextension study to investigate the long-term safety and tolerability of THC/CBD oromucosal spray and oromucosal THC spray in patients with terminalcancer-related pain refractory to strong opioid analgesics. J. Pain SymptomManag. 46 (2), 207e218.
Jonathan, R., McDearmid, M.L., Drapeau, Pierre, 2006. Glycine receptors regulateinterneuron differentiation during spinal network development. PNAS 103 (25),9679e9684.
Jun Chen, S.K.J., DiDomenico, Stanley, Perner, Richard J., Mikusa, Joe P.,Gauvin, Donna M., Segreti, Jason A., Han, Ping, Zhang, Xu-Feng,Niforatos, Wende, Bianchi, Bruce R., Baker, Scott J., Zhong, Chengmin, GHS,McDonald, Heath A., Schmidt, Robert G., McGaraughty, Steve P., Chu, KatharineL., Faltynek, Connie R., Kort, Michael E., Reilly, Regina M., Kym, Philip R., 2011.Selective blockade of TRPA1 channel attenuates pathological pain withoutaltering noxious cold sensation or body temperature regulation. Pain 152 (5),1165e1172.
Kamei, D., Yamakawa, K., Takegoshi, Y., Mikami-Nakanishi, M., Nakatani, Y., Oh-Ishi, S., Yasui, H., Azuma, Y., Hirasawa, N., Ohuchi, K., Kawaguchi, H., Ishikawa, Y.,Ishii, T., Uematsu, S., Akira, S., Murakami, M., Kudo, I., 2004. Reduced pain hy-persensitivity and inflammation in mice lacking microsomal prostaglandin esynthase-1. J. Biol. Chem. 279 (32), 33684e33695.
Kawamura, Y., Fukaya, M., Maejima, T., Yoshida, T., Miura, E., Watanabe, M., Ohno-Shosaku, T., Kano, M., 2006. The CB1 cannabinoid receptor is the majorcannabinoid receptor at excitatory presynaptic sites in the hippocampus andcerebellum. J. Neurosci. 26 (11), 2991e3001.
King, T., Vera-Portocarrero, L., Gutierrez, T., Vanderah, T.W., Dussor, G., Lai, J.,Fields, H.L., Porreca, F., 2009. Unmasking the tonic-aversive state in neuropathicpain. Nat. Neurosci. 12 (11), 1364e1366.
Kling, C., Koch, M., Becker, BSaCM., 1997. The frameshift mutation oscillator(Glra1spd-ot) produces a complete loss of glycine receptor a1-polypeptide inmouse central nervous system. Neuroscience 78 (2), 411e417.
J. Lu et al. / Neuropharmacology 133 (2018) 224e232 231

Knabl, J., Witschi, R., Hosl, K., Reinold, H., Zeilhofer, U.B., Ahmadi, S., Brockhaus, J.,Sergejeva, M., Hess, A., Brune, K., Fritschy, J.M., Rudolph, U., Mohler, H.,Zeilhofer, H.U., 2008. Reversal of pathological pain through specific spinalGABAA receptor subtypes. Nature 451 (7176), 330e334.
Kuner, R., 2010. Central mechanisms of pathological pain. Nat. Med. 16 (11),1258e1266.
Ledent, Catherine, OV, Cossu, Gregorio, Petitet, François, Aubert, Jean-François,Beslot, Françoise, B€ohme, Georg A., Imperato, Assunta, Pedrazzini, Thierry,Roques, Bernard P., Vassart, Gilbert, Fratta, Walter, Parmentier, Marc, 1999.Unresponsiveness to cannabinoids and reduced addictive effects of opiates inCB1 receptor knockout mice. Science 283 (5400), 401e404.
Long, L.E., Malone, D.T., Taylor, D.A., 2006. Cannabidiol reverses MK-801-induceddisruption of prepulse inhibition in mice. Neuropsychopharmacology 31 (4),795e803.
Lozovaya, N., Yatsenko, N., Beketov, A., Tsintsadze, T., Burnashev, N., 2005. Glycinereceptors in CNS neurons as a target for nonretrograde action of cannabinoids.J. Neurosci. 25 (33), 7499e7506.
Luo, H., Cheng, J., Han, J.-S., Wan, Y., 2004. Change of vanilloid receptor 1 expressionin dorsal root ganglion and spinal dorsal horn during inflammatory nociceptioninduced by complete Freund's adjuvant in rats. Neuroreport 15 (4), 655e658.
Lynch, J.W., 2009. Native glycine receptor subtypes and their physiological roles.Neuropharmacology 56 (1), 303e309.
Malmberg, Annika B., CC, Tonegawa, Susumu, Basbaum, Allan I., 1997. Preservedacute pain and reduced neuropathic pain in mice lacking PKC. Science 278.
Martin-Sanchez, E., Furukawa, T.A., Taylor, J., Martin, J.L., 2009. Systematic reviewand meta-analysis of cannabis treatment for chronic pain. Pain Med. 10 (8),1353e1368.
McDearmid, J.R., Liao, M., Drapeau, P., 2006. Glycine receptors regulate interneurondifferentiation during spinal network development. Proc. Natl. Acad. Sci. U. S. A.103 (25), 9679e9684.
Murray, R.M., Morrison, P.D., Henquet, C., Di Forti, M., 2007. Cannabis, the mind andsociety: the hash realities. Nat. Rev. Neurosci. 8 (11), 885e895.
Nagy, A., Rossant, J., Nagy, R., Abramow-Newerly, W., Roder, J.C., 1993. Derivation ofcompletely cell culture-derived mice from early-passage embryonic stem cells.Proc. Natl. Acad. Sci. U. S. A. 90 (18), 8424e8428.
Piechotta, K., Weth, F., Harvey, R.J., Friauf, E., 2001. Localization of rat glycine re-ceptor alpha1 and alpha2 subunit transcripts in the developing auditorybrainstem. J. Comp. Neurol. 438 (3), 336e352.
Pogatzki-Zahn, E.M., Shimizu, I., Caterina, M., Raja, S.N., 2005. Heat hyperalgesiaafter incision requires TRPV1 and is distinct from pure inflammatory pain. Pain115 (3), 296e307.
Raghavendra, V., Tanga, F.Y., DeLeo, J.A., 2004. Complete Freunds adjuvant-inducedperipheral inflammation evokes glial activation and proinflammatory cytokineexpression in the CNS. Eur. J. Neurosci. 20 (2), 467e473.
Robert Harvey, U.D., W€assle, Heinz, Ahmadi, Seifollah, Heindl, Cornelia,Reinold, Heiko, Smart, Trevor, Harvey, Kirsten, Schütz, Burkhard, Abo-Salem, Osama, Zimmer, Andreas, Poisbeau, Pierrick, Welzl, Hans, Wolfer, David,Betz, Heinrich, Müller, Hanns, 2004. GlyR a3: an essential target for spinal
PGE2-mediated inflammatory pain sensitization. Science 304 (5672), 884e887.Rodriguez, C.I., Buchholz, F., Galloway, J., Sequerra, R., Kasper, J., Ayala, R.,
Stewart, A.F., Dymecki, S.M., 2002. High-efficiency deleter mice show that FLPeis an alternative to Cre- loxP. Nat. Genet. 25, 139e140.
Schmieden, V., GG, Schofield, P.R., Betz, H., 1989. Functional expression in Xenopusoocytes of the strychnine binding 48 kd subunit of the glycine receptor. EMBO J.8 (3), 695e700.
Shiang, R., Ryan, S.G., Zhu, Y.Z., Wasmuth, J.J., 1994. Mutations in the alpha 1 subunitof the inhibitory glycine receptor cause the dominant neurologic disorder,hyperekplexia. Nat. Genet. 5 (4), 351e358.
Shiang, Rita, Ryan, Stephen G., Zhu, Ya-Zhen, Fielder, Thomas J., Allen, Richard J.,Fryer, Alan, Yamashita, Sumimasa, O'Connell, Peter, Wasmuth, J.J., 1995. Muta-tional analysis of familial and sporadic hyperekplexia. Ann. Neurol. 38 (1),85e91.
Silva, M., Amorim, D., Almeida, A., Tavares, I., Pinto-Ribeiro, F., Morgado, C., 2013.Pronociceptive changes in the activity of rostroventromedial medulla (RVM)pain modulatory cells in the streptozotocin-diabetic rat. Brain Res. Bull. 96,39e44.
St John Smith, E., 2017. Advances in understanding nociception and neuropathicpain. J. Neurol. 1e8. https://doi.org/10.1007/s00415-017-8641-6.
Wang, T., Collet, J.P., Shapiro, S., Ware, M.A., 2008. Adverse effects of medical can-nabinoids: a systematic review. CMAJ 178 (13), 1669e1678.
Wilkinson, S.M., Price, J., Kassiou, M., 2013. Improved accessibility to the desoxyanalogues of D9-tetrahydrocannabinol and cannabidiol. Tetrahedron Lett. 54(1), 52e54.
Wilson, R.I., Nicoll, R.A., 2002. Endocannabinoid signaling in the brain. Science 296(5568), 678e682.
Xiong, W., Cheng, K., Cui, T., Godlewski, G., Rice, K.C., Xu, Y., Zhang, L., 2011.Cannabinoid potentiation of glycine receptors contributes to cannabis-inducedanalgesia. Nat. Chem. Biol. 7 (5), 296e303.
Xiong, W., Cui, T., Cheng, K., Yang, F., Chen, S.R., Willenbring, D., Guan, Y., Pan, H.L.,Ren, K., Xu, Y., Zhang, L., 2012. Cannabinoids suppress inflammatory andneuropathic pain by targeting alpha3 glycine receptors. J. Exp. Med. 209 (6),1121e1134.
Xiong, W., Wu, X., Li, F., Cheng, K., Rice, K.C., Lovinger, D.M., Zhang, L., 2012.A common molecular basis for exogenous and endogenous cannabinoidpotentiation of glycine receptors. J. Neurosci. 32 (15), 5200e5208.
Xiong, W., Chen, S.R., He, L., Cheng, K., Zhao, Y.L., Chen, H., Li, D.P., Homanics, G.E.,Peever, J., Rice, K.C., Wu, L.G., Pan, H.L., Zhang, L., 2014. Presynaptic glycinereceptors as a potential therapeutic target for hyperekplexia disease. Nat.Neurosci. 17 (2), 232e239.
Yekkirala, A.S., Roberson, D.P., Bean, B.P., Woolf, C.J., 2017. Breaking barriers to novelanalgesic drug development. Nat. Rev. Drug Discov. 16 (11), 810.
Yu, L., Yang, F., Luo, H., Liu, F.Y., Han, J.S., Xing, G.G., Wan, Y., 2008. The role of TRPV1in different subtypes of dorsal root ganglion neurons in rat chronic inflam-matory nociception induced by complete Freund's adjuvant. Mol. Pain 4, 61.
Zhou, Lan, Chillag, Kipp L., Nigro, M.A., 2002. Hyperekplexia: a treatable neuro-genetic disease. Brain Dev. 24 (7), 669e674.
J. Lu et al. / Neuropharmacology 133 (2018) 224e232232







![OPEN ACCESS International Journal of Molecular Sciences...hair growth [2].Platelet-derived growth factor (PDGF) isoforms reportedlyinduce and maintain theanagen phase of the murine](https://img.pdfslide.tips/doc/110x75/60f85444d7faee31306fdb0e/open-access-international-journal-of-molecular-sciences-hair-growth-2platelet-derived.jpg)