Embed Size (px)
Citation preview

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 1/141
Univerzitet u Beogradu
Biološki fakultet
mr Saša P. Marić
Evolucijska istorija kompleksa potočne
pastrmke Salmo trutta L. 1758 na područ juRepublike Srbije i značaj za ribarstvo
Doktorska disertacija
Beograd, 2005.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 2/141
Univerzitet u Beogradu
Biološki fakultet
mr Saša P. Marić
Evolucijska istorija kompleksa potočne
pastrmkeSalmo trutta
L. 1758 na područ juRepublike Srbije i značaj za ribarstvo
Mentor:
dr Predrag Simonović – vanredni profesor
Biološki fakultet
Univerzitet u Beogradu
Članovi komisije:
dr Jelka Crnobrnja - Isailović – naučni saradnik
Institut za biološka istraživanja «Siniša Stanković» Beograd
dr Aleš Snoj – naučni saradnik
Biotehnički fakultet
Univerzitet u Ljubljani
Datum odbrane:
Doktorska disertacija
Beograd, 2005.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 3/141
ZahvalnicaIII
Zahvalnica
Zahvaljujem se svima koji su mi na bilo koji nač in olakšali rad na izradidisertacije i koji su svojim zalaganjem i nesebič noš ću doprineli da ona ugleda svetlostdana.
Srdač no se zahvaljujem mentoru i č lanovima Komisije na korektnom i profesionalnom odnosu punom razumevanja.
Zahvaljujem se mentoru Prof. dr Predragu Simonoviću, koji mi je, osim istina izove nauke, pokazao da se autoritet i poštovanje stič u jedino znanjem i korektnim odnosom
prema kandidatu. Posebno se zahvaljujem za ukazano poverenje i neogranič enu slobodu.
Zahvaljujem se dr Alešu Snoju koji je rukovodio laboratorijskim radom i koji me je upoznao sa metodama molekularne genetike. Posebno mu se zahvaljujem na posvećenom vremenu, izuzetnom stpljenju, ukazanom poverenju, ustupljenoj literaturi, savetima, kao i odnosu za koji sigurno mogu reći da prevazilazi granice profesionalnog.
Takođ e se zahvaljujem dr Jelki Crnobrnji – Isailović koja mi je izašla u susretkada je to bilo neophodno, i u najkraćem roku pregledala disertaciju dajući veomakorisne sugestije.
Zahvaljujem se Nenadu Sekuliću iz Zavoda za zaštitu prirode Srbije naustupljenom materijalu sa područ ja Kosova i Metohije, kao i kolegama Prof. dr BožidaruĆ ur č iću, Prof. dr Predragu Simonoviću, Doc. dr Veri Nikolić , dr Ljiljani Tomović ,
Vladanu Bjedovu, Rastku Ajtiću, Nikoli Kolundžiću, Č edomiru Mijoviću i Žiki Milenkoviću, predsedniku Ribarskog područ ja «Južna Morava I» Leskovac, koji su mi narazlič ite nač ine pomogli u prikupljanju analiziranog materijala.
Zahvaljujem se kolegama dr Simoni Sušnik, mr Andreju Raspetu, mr Tamari Jug,mr Brigiti Slavec, mr Poloni Frajman i Vidi Štuhec sa Biotehnič kog fakulteta u Ljubljanikoji su mi pomogli u prevazilaženju svih laboratorijskih problema i koji su svojomljubaznoš ću i gostoprimstvom svakako doprineli da mi deo izrade teze koji se odvijao na
Biotehnickom fakultetu u Ljubljani ostane u prelepom sećanju.
Zahvaljujem se kolegama Prof. dr Milošu Kaleziću, Doc. dr Ani Ivanović , dr
Ljiljani Tomović i mr Imreu Krizmaniću na razumevanju i peuzimanju dela nastavnih idrugih obaveza za vreme izrade disertacije.
Takođ e se zahvaljujem mr Tamari Karan Žnidaršič na korisnim sugestijamavezanim za tehnič ku obradu teksta, a mr Biljani Stojković za pomoć pri tumač enju
pojedinih rezultata.
Posebno bih se zahvalio svojim roditeljima, bratu i naravno supruzi Katarini, za pruženu podršku, moralnu potporu i sva odricanja koja su bila neminovna prilikomizrade disertacije. Najlepše vam hvala.
Saša Marić

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 4/141
IzvodIV
Izvod
Za rasvetljavanje filogeografije populacija potočne pastrmke (Salmo trutta) na
područ ju Republike Srbije, sekvencioniran je 5' kraj kontrolnog regiona mtDNA u dužini
od 561 bp i dobijene sekvence upoređene su sa poznatim sekvencama iz prethodnih
istraživanja na drugim teritorijama. U analizu je uključena 101 jedinka poreklom iz
gornjih tokova reka crnomorskog, egejskog i jadranskog sliva. Sekvencioniranjem je
identifikovano petnaest haplotipova, od kojih se četrnaest smatra autohtonim i oni
pripadaju dunavskoj i jadranskoj liniji, dok samo jedan haplotip (ATcs1), pronađen kod
dve jedinke poreklom iz dve poribljavane reke, pripada atlantskoj liniji koja je
komercijalizovana i najčešće korišćena za poribljavanje. Autohtoni haplotipovi odlikuju
se izrazitom geografskom distribucijom: dunavski haplotipovi su strogo ograničeni na
reke dunavskog sliva, dok jadranski haplotipovi dominiraju u rekama egejskog i
jadranskog sliva; najveći deo ukupne molekularne varijanse (69%) pripisan je upravo
razlikama između slivova. Filogenetskom rekonstrukcijom, koja je dopunjena sa šest
novih haplotipova, po prvi put opisanih u ovom radu, podržano je stanovište o
ancestralnoj poziciji dunavske linije unutar kompleksa potočne pastrmke, kao i naglašeno postojanje posebne farioides (Ad+) klade otkrivene unutar jadransko-mediteranske-
marmoratus filogenetske grupe. Dobijeni rezultati potvrdili su naša očekivanja o
postojanju visokog genetičkog diverziteta balkanskih populacija potočne pastrmke, što
zahteva dalja istraživanja koja bi trebala da obuhvate pastrmske populacije iz celog
regiona.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 5/141

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 6/141
SadržajVI
Sadržaj
Zahvalnica................................................................................................. IIIIzvod ......................................................................................................... IV
Abstract .......................................................................................................V
Sadržaj....................................................................................................... VI
Skraćenice i simboli ................................................................................VIII
1. UVOD......................................................................................................... 1
1.1. Uvodne napomene ...................................................................................... 1
1.2. Pregled lterature .......................................................................................... 3
1.2.1. Uopšteno o Salmonidama ........................................................................... 3
1.2.2. Karakteristike familije Salmonidae............................................................. 4
1.2.3. Klasifikacija i filogenetski odnosi unutar familije Salmonidae.................. 5
1.2.4. Salmo trutta (Linnaeus, 1758) - potočna pastrmka................................... 12
1.2.5. Metode razlikovanja ribljih populacija ..................................................... 21
1.2.5.1. Morfološke metode razlikovanja ribljih populacija .................................. 21
1.2.5.2. Genetičke metode razlikovanja ribljih populacija .................................... 22
1.2.5.2.1. Alozimi ..................................................................................................... 23
1.2.5.2.2. Savremeni genetički markeri .................................................................... 24
1.2.5.2.2.1. Ponavljajuća DNA .................................................................................... 25
1.2.5.2.2.1.1. Mikrosateliti .............................................................................................. 25
1.2.5.2.2.2. Mitohondrijalna DNA............................................................................... 271.2.5.2.2.2.1. Osobine mitohondrijalne DNA ................................................................. 28
1.2.5.2.2.2.2. Mitohondrijska DNA kao genetski marker............................................... 31
1.2.5.2.2.2.3. Upotreba mtDNA za proučavanje vrsta i populacija salmonida............... 32
1.2.6. Ugroženost potočne pastrmke (Salmo trutta) u Evropi ............................ 35
1.3. Ciljevi rada................................................................................................ 38
2. MATERIAL I METODE.......................................................................... 39
2.1. Materijal .................................................................................................... 39
2.1.1. Prikupljanje i čuvanje uzoraka.................................................................. 39
2.1.2. Raspored lokaliteta i brojnost analiziranih uzoraka.................................. 402.1.3. Hemikalije................................................................................................. 42
2.1.3.1. Priprenljeni setovi hemikalija ................................................................... 42
2.1.3.2. Enzimi ....................................................................................................... 42
2.1.3.3. Početni oligonuklotidi (prajmeri).............................................................. 43
2.1.3.4. Markeri...................................................................................................... 43
2.1.3.5. Puferi......................................................................................................... 43
2.1.4. Laboratorijska oprema .............................................................................. 43
2.2. Metode ...................................................................................................... 44
2.2.1. Izolacija DNA ........................................................................................... 44
2.2.1.1. Fenolna ekstrakcija ................................................................................... 44
2.2.1.2. Wizard®
Genomic DNA Purification Kit ................................................. 452.2.1.3. Jet quick tissue DNA spin kit/250 ............................................................ 45

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 7/141
SadržajVII
2.2.2. Provera uspešnosti izolacije DNA ............................................................ 45
2.2.3. Lančana reakcija polimeraze (PCR) ......................................................... 46
2.2.4. Restrikciona analiza kontrolnog regiona mtDNA..................................... 48
2.2.5. Sekvencioniranje mitohondrijalne DNA................................................... 48
2.2.5.1. Izolacija fragmenata DNA iz agaroznog gela ........................................... 48
2.2.5.2. Priprema reakcije za automatski sekvenator............................................. 492.2.5.3. Priprema DNA fragmenata za sekvencioniranje....................................... 50
2.2.5.4. Sekvencioniranje DNA fragmenata .......................................................... 50
2.2.6. Programska analiza kontrolnog regiona mtDNA...................................... 51
3. REZULTATI............................................................................................. 53
3.1. Analiza kontrolnog regiona mtDNA upotrebom RFLP tehnike ............... 53
3.2. Analiza sekvencioniranja kontrolnog regiona mtDNA............................. 54
3.2.1. Upoređivanje sekvenci kontrolnog regiona mtDNA pronađenih
haplotipova na teritoriji Srbije sa već poznatim haplotipovima potočne
pastrmke iz svih filogenetskih linija ......................................................... 63
4. DISKUSIJA .............................................................................................. 71
4.1. Analiza kontrolnog regiona mtDNA......................................................... 72
4.2. Paleozoogeografska razmatranja, sa osvrtom na kolonizaciju Evrope
potočnom pastrmkom ............................................................................... 81
4.3. Konzervacija genetičke raznovrsnosti populacija potočne pastrmke ....... 95
4.3.1. Specifičnosti konzervacije potočne pastrmke ........................................... 95
4.3.2. Status ugroženosti potočne pastrmke u Evropi ......................................... 97
4.3.3. Definisanje konzervacionih jedinica......................................................... 98
4.3.4. Krajnja preporuka u konzervaciji potočne pastrmke ................................ 994.3.5. Konzervacija diverziteta haplotipova u Srbiji i značaj za ribarstvo ....... 100
5. ZAKLJUČCI........................................................................................... 107
6. SUMMARY............................................................................................ 111
7. LITERATURA ....................................................................................... 113

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 8/141
Skraćenice i simboliVIII
Skraćenice i simboli
A adenin
ATP adenozin trifosfat
bp bazni par
C citozin
ddNTP 2’, 3’ – dideoksinukleozid 5’–trifosfat
DNA dezoksiribonukleinska kislina
dNTP 2’ – deoksinukleozid 5’ – trifosfat
EDTA etilen – diamino – tetraosir ćetna kislina
eng. engleski
G guanin
kbp hiljadu baznih parova
mtDNA mitohondrijska DNA
PCR polymerase chain reaction
pH negativni logaritam koncentracije vodonikovih jona
RFLP restriction fragment lenght polymorphismRNA ribonukleinska kislina
rRNA ribozomalna RNA
SDS sodium dodecyl sulphate
T timin
TAE rastvor Trisa, acetata u EDTA
Taq DNA–polimeraza DNA – polimeraza, izolovana iz Thermus aquaticus
TBE rastvor Trisa, borata u EDTA
TEN rastvor Trisa, EDTA u NaCl
Tris 2 – amino – 2 – (hidroksimetil) – 1, 3 – propandiol
tRNA transportna RNA
TSR template suppresion reagent

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 9/141
Uvod
1
1.
UVOD
1.1. UVODNE NAPOMENE
Povoljni prirodni uslovi omogućuju postojanje više hiljada vodotokova na
teritoriji Srbije. Njihova ukupna dužina iznosi 65980 km ili prosečno 747 m/km2. U Srbiji
ima samo 11 reka čija dužina iznosi preko 200 km, što znači da preovlađuju male i
srednje reke dužine ispod 100 km (Gavrilović i Dukić, 2002).
Reke Srbije otiču u tri mora, tako da se može reći da Srbija predstavlja
hidrografski čvor Balkana. Najveća površina teritorije odvodnjava se prema Crnom
(92,46%), a znatno manja površina ka Jadranskom (5,36%) i Egejskom moru (2,18%).
Morski slivovi Srbije razlikuju se ne samo po veličini nego i hidrološki, što je uzrokovano
specifičnim klimatskim prilikama, uslovima oticanja padavina i geološkim sastavom
terena.
Hidrološka raznovrsnost na područ ju Srbije u pogledu broja, veličine, geografskog
položaja i oticanja reka uslovila je i raznovrsnost živog sveta koji je vezan za vodena
staništa. U planinskim vodama koje preovlađuju po broju u odnosu na ravničarske
nalazimo idealne uslove za život pastrmskih vrsta riba. Prirodna nepovezanost morskih
slivova i geotektonski događaji uslovili su da danas na područ ju Srbije žive izolovane,
lokalno specifične populacije potočne pastrmke (Salmo trutta).
U poslednjih 20 godina mnogo je rađeno na utvr đivanju genetičkog identiteta
populacija potočne pastrmke na područ ju Evrope (Ferguson i Fleming, 1983; Krieg i
Guyomard, 1985; Ferguson, 1989; Karakousis i Triantaphyllidis 1990; Phillips i Pleyte,
1991; Bernatchez i sar., 1992; Bernatchez, 1995; 2001; Giuffra i sar., 1994; 1996;
Bernatchez i Osinov, 1995; Riffel i sar., 1995; Apostolidis i sar., 1996; 1996a; 1997;
Garcia-Marin i sar., 1996; 1999; McKay i sar., 1996; Shed'ko i sar., 1996; Oleynik, 1997;
Phillips i Oakley, 1997; Weiss i sar., 2000; 2001; Cortey i García-Marín, 2002; Cortey i
sar., 2004). Na osnovu već poznatih rezultata, predpostavili smo da i populacije na
područ ju Srbije zbog duge izolovanosti i specifičnosti staništa predstavljaju posebne
evolutivne grane.
Snažan razvoj sportskog ribolova, kao i veoma zastupljen krivolov uz narušavanje
prirodnih uslova staništa doveli su do značajnog smanjenja brojnosti populacija potočne
pastrmke, a pojedine populacije su postale ugrožene sa stanovišta opstanka. Kao

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 10/141
Uvod
2
posledica toga dolazi vrlo često do poribljavanja komercijalnim linijama koje su
prilagođene ribnjačkom uzgoju. Najčešće se radi o linijama alohtonog porekla, koje se
nakon ubacivanja u vodotokove nesmetano ukrštaju sa autohtonim populacijama, što
može dovesti do ugrožavanja genetičke specifičnosti autohtone populacije. Unošenje
alohtonih populacija može dovesti i do kompeticije u pogledu osnovnih životnih resursa
što može rezultirati potiskivanjem autohtonih populacija iz njihovih prirodnih staništa.
Pastrmke imaju izrazitu osobinu fenotipske plastičnosti, odnosno odgovora na
promenu uslova staništa što često dovodi do nastanka specifičnih životnih formi, tako da
se genetički različite jedinke fenotipski ne razlikuju i obratno (Bernatchez i sar., 1992).
Fenotipski karakteri mogu biti indikativni (Marić i sar., 2004), ali nisu do kraja pouzdani
za razlikovanje autohtonih od alohtonih populacija, pa je zbog toga neophodno koristiti
precizne molekularno-genetičke metode. Na osnovu rezultata dobijenih uz pomoć ovih
metoda može se sa velikom tačnošću utvrditi autohtonost populacija. Autohtone
populacije su još uvek sačuvane u izvorišnim, nepristupačnim delovima vodotokova koji
su vrlo često odvojeni prirodnim barijerama od nizvodnih delova tokova. Autohtone
populacije su retke ali još uvek postoje, pa, zahvaljujući tome, u budućnosti mogu biti
formirana autohtona matična jata (stvaranje matičnih jata je skupo, nije uvek praktično za
održavanje i isplati se samo za jata zastupljena u ribolovno atraktivnim vodama i gde
postoje dokazi da nije bilo introdukcije). Autohtono matično jato može se koristiti za
repopulaciju na onim lokalitetima na kojima je pastrmka gotovo istrebljena, kao i na
ribolovno atraktivnim vodama pod snažnim ribolovnim pritiskom.
Kao marker za utvr đivanje genetičke varijabilnosti populacija potočne pastrmke u
ovom radu koristili smo kontrolni region mitohondrijalne DNA, koji je, zbog svoje
hipervarijabilnosti, dobar genetički marker. Zbog načina nasleđivanja, mtDNA se koristi
za utvr đivanje filogenetskih odnosa između rodova, vrsta i nižih taksonomskih jedinica.
Ovaj tip istraživanja spada u filogeografske studije pomoću kojih se može utvrditigeografski raspored evolutivnih linija, kao i geografski procesi koji su uticali na taj
raspored, sa posebnim osvrtom na različitost, kako između srodnih vrsta, tako i unutar
vrste (Avise, 1998; Hewitt, 2001).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 11/141
Uvod
3
1.2. PREGLED LITERATURE
1.2.1. UOPŠTENO O SALMONIDAMA
Salmoniformes je relativno brojan red primitivnih Euteleostei koji obuhvata dva
podreda: Osmeroidei i Salmonoidei (Johnson i Patterson, 1996).
U okviru podreda Osmeroidei nalazi se porodica Osmeridae sa šest rodova. Vrste
ove porodice uglavnom su ograničene na severne delove Atlantika i Pacifika. To su ribe
male veličine sa adipoznim perajem. Žive u obalnim zonama odakle mnoge ulaze u reke
gde se mreste. U nekim delovima areala dostižu veliku brojnost, tako da su i ekonomski
važne ribe.
Podred Salmonoidei uključuje anadromne i potamodromne grupe, koje veći deoživotnog ciklusa provode u morima i čije migracije po spektakularnosti nalikuju
migracijama jegulja. Zavičajno ponašanje ovih riba je veoma izraženo. Salmonidnim
ribama pripadaju i grupe koje su isključivo slatkovodne (npr. Hucho, Salmothymus,
Salvethymus). Kod salmonidnih riba iza dorzalnog peraja postoji dobro razvijeno
adipozno peraje. Kod anadromnih salmonidnih riba primarna metamorfoza se događa
tokom nizvodnih migracija. Sekundarna metamorfoza se odvija tokom uzvodnih
migracija pred period mresta. Kod pripadnika roda Salmo i nekih drugih rodova dolazi do"regresivne metamorfoze", gubljenja morfoloških karakteristika nastalih sekundarnom
metamorfozom tokom njihovih nizvodnih migracija posle reprodukcije. Kada naredni put
ove ribe krenu da se mreste, kod njih se ponavlja ciklus morfoloških promena i njihovog
gubljenja (Saunders i Schom, 1985).
Salmonide imaju cirkumpolarno rasprostranjenje i karakteristične su za severnu
Zemljinu hemisferu (Slika 1). Naseljavaju područ ja Severnog ledenog mora i severnog
dela Atlantika i Pacifika, reke i mora Evrope, zapadne Azije i Severne Amerike (Lagler,1977). Zbog njihovog komercijalnog značaja introdukovane su i u vode južne Amerike,
Australije i Novog Zelanda (Wheeler, 1992).
Slika 1. Područ je rasprostranjenja Salmonida (Maitland, 1995) – modifikovano.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 12/141
Uvod
4
Na osnovu palentoloških dokaza utvr đena je pojava salmonidnih vrsta još početkom
tercijara u eocenu, dok su krajem tercijara i početkom kvartara bile široko rasprostranjene
(Mitrović i Pavlović, 1980). Među naučnicima postoje nesuglasice oko pradomovine
salmonida. Neki smatraju da salmonide vode poreklo iz slatkih voda (Neave, 1958;
Vladykov, 1963), ali ima i onih koji veruju da je more njihovo prvobitno stanište (Schmidt,
1947).
1.2.2. K ARAKTERISTIKE FAMILIJE SALMONIDAE
Jedna od osnovnih zajedničkih karakteristika svih vrsta riba iz familije
Salmonidae je postojanje masnog ili adipoznog peraja, koje se nalazi između leđnog i
repnog peraja. Celo telo riba iz ove familije je, izuzev glave, prekriveno sitnim krljuštima.
Bočna linija je jasno uočljiva. Leđno peraje je kratko. Dentalne, gornjevilične i
međuvilične kosti salmonidnih vrsta riba su nazubljene, a od kostiju gornje vilice
najsnažnije je nazubljen vomer. Kod salmonidnih vrsta on je različito nazubljen, pa oblik
i nazubljenost vomera (kao i broj piloričnih nastavaka koji varira između 17 i 210)
predstavlja važan taksonomski karakter, koji služi za identifikaciju riba iz ove familije.
Kako su one izraziti predatori, želudac im je prilagođen varenju hrane životinjskog
porekla - širok je i mišićav.
Leđa i bokovi salmonidnih vrsta su prekriveni crnim i/ili crvenim pegama, a kod
nekih vrsta i raznim tipovima pruga i šara.
Sve salmonidne vrste obitavaju isključivo u hladnim, bistrim i nezagađenim
potocima ili rekama brzog toka, koje su bogate rastvorenim kiseonikom (O2) ili u
planinskim jezerima čije se temperature vode kreću oko 10oC i koje, ni u najtoplije doba
godine, ne prelaze 18oC (maksimalno 20oC). To su sve tzv. reofilne vrste riba,
vretenastog oblika tela, prilagođene životu u brzim rečnim tokovima.
Sve vrste iz familije Salmonidae mreste se isključivo u slatkoj vodi, i to u
izvorišnim delovima potoka ili reka u kojima obitavaju ili pak, u priobalnim delovima
jezera, u kojima žive na peskovitom, šljunkovitom ili kamenitom dnu navedenih vodenih
biotopa. Sazrevanje gonada riba iz ove familije nastupa u kasnim jesenjim danima, od
novembra do januara - ređe od oktobra do februara - izuzev mladice ( Hucho hucho),
mekousne pastrmke (Salmothymus obtusirostris) i lipljana (Thymallus), koje se mreste u
prolećnim mesecima (mart - april). Relativna plodnost (RF – predstavlja broj jaja na 1 kg

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 13/141
Uvod
5
telesne težine) salmonidnih vrsta riba relativno je mala i kreće se od oko 1500 jaja - ikre
(kod potočne pastrmke) do oko 12000 jaja (kod mladice) u odnosu na 1 kg telesne težine.
Veličina riba iz familije Salmonidae je neujednačena. Dok vrste iz roda Salmo,
Salvelinus i Salmothymus u potocima i manjim rečicama narastu otprilike do 30 ili 40 cm
totalne dužine i između 0,30 kg i 1 kg mase tela (nekada i nešto više), one iz većih, dubljih
reka ili jezera narastu znatno više, čak i preko 22 kg, a mladica i preko 50 kg (Aganović,
1979).
1.2.3. K LASIFIKACIJA I FILOGENETSKI ODNOSI UNUTAR FAMILIJE SALMONIDAE
Klasifikacija porodice Salmonidae je često menjana, tako da je vrlo teško dati
precizan broj rodova u okviru podfamilije Salmoninae. Broj rodova, u zavisnosti od
autora, kreće se između 5 i 10 (Nelsen, 1984). Prema klasifikaciji Starley-a i Smith-a
(1993) broj rodova je 8, a prema najnovijoj klasifikaciji salmonida po ITISu (eng.
Integrated Taxonomic Information Sistem, 30. 03. 2004.), broj rodova u okviru
podfamilije Salmoninae je 7.
Superordo: Protacanthopterygii
Ordo: Salmoniformes
Familia: Salmonidae
Subfamilia: Corregoninae (ozimice)
Subfamilia: Thymalinae (lipljani)
Subfamilia: Salmoninae
Genus: Brachymystax, (lenok)
Genus: Hucho, (mladica)
Genus: Oncorhynchus, (pacifički lososi i pacifičke pastrmke)
Genus: Parahucho
Genus: Salmo, (atlantski losos i pastrmka)
Genus: Salvelinus, (zlatovčice)
Genus: Salvethymus

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 14/141
Uvod
6
Porodica Salmonidae obuhvata tri podporodice: Thymallinae, Coregoninae i
Salmoninae (Shaposhnikova, 1975). Ovakva podela porodice Salmonidae na tri
podporodice je opšte prihvaćena. Prema morfologiji rodovi podporodice Salmoninae su
noviji i napredniji oblici u odnosu na rodove podporodica Thymallinae i Coregoninae, za
koje se smatra da su evolutivno stariji (Stearley i Smith, 1993; Slika 2).
Slika 2. Prikaz odnosa među rodovima porodice Salmonidae na osnovu morfoloških
istraživanja (Starley i Smith, 1993).
Mnogo veća neslaganja među autorima možemo sresti pri klasifikaciji rodova
unutar podporodice Salmoninae.
Autori se uglavnom slažu oko filogenetskog statusa najbolje proučenih rodova
( Hucho, Salvelinus, Salmo, Oncorhynchus), za koje se smatra da predstavljaju naprednije
salmonide. To se posebno odnosi na rodove Oncorhynchus i Salmo. Ostali rodovi:
Brachymystax, Salmothymus, Acantholingua i Platysalmo, koji su slabije proučeni,
smatraju se morfološki primitivnijim i njihovo filogenetsko mesto još uvek je potpuno
nejasno (Stearley i Smith, 1993; Osinov, 1999; Phillips i Oakley, 1997).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 15/141
Uvod
7
Slika 3.Prikaz četiri filogenetske hipoteze date na osnovu morfoloških analiza od strane
različitih autora (za odnose između rodova Brachymystax, Hucho i Salvelinus) (1) Norden
(1961), (2) Kendall i Behnke (1984), (3) Dorofeeva (1989) i (4) Stearley i Smith (1993).
(1)
(3)
Prema prvoj hipotezi, Norden (1961) smatra da svaki od rodova ima monofiletsko
poreklo (Slika 3-1). Kendall i Behnke (1984), prema drugoj hipotezi, smatraju da rodovi
Brachymystax, Hucho i Salvelinus vode poreklo od zajedničke monofiletske grupe (Slika
3-2). Dorofeyeva (1989), prema trećoj hipotezi, smatra da rodovi Brachymystax, Hucho i
Parahucho vode poreklo od jedne zajedničke monofiletske grupe, a Salvelinus od druge
(Slika 3-3). Stearley i Smith (1993), prema četvrtoj hipotezi, smatraju da rodovi Hucho i
Salvelinus vode poreklo od jedne monofiletske grupe, Salmothymus i Acantholingua oddruge, a Brachymystax od treće (Slika 3-4).
Intergenerički odnosi u podporodici Salmoninae znatno su izmenjeni nakon
uvođenja molekularnih tehnika (Slika 4). Dobijanjem novih rezultata koji su bili različiti
u odnosu na rezultate dobijene morfološkim analizama, bilo je neophodno izvršiti
reklasifikaciju rodova u okviru podporodice. Dobijeni rezultati bili su pre svega značajni
za utvr đivanje taksonomske pozicije “problematičnih” rodova ( Brachymystax,
Salmothymus, Acantholingua i Platysalmo).
Thymallus
Brachymystax
Hucho
Salvleinus
Salmo
Oncorhynchus
Thymullus
Brachymystax
Hucho
Parahucho
Salvelinus
Salmothymus
Salmo
Oncorhynchus
Thymallus
Brachymystax
Hucho
Salvelinus
Salmo
Oncorhynchus
Thymallus
Brachymystax
Acantholingua
Salmothymus
Hucho
Salvelinus
Salmo
Oncorhynchus
(2)
(4)

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 16/141
Uvod
8
Predstavnici rodova Parahucho i Salvethymus (ITIS, 2004), na osnovu genetičkih
istraživanja, svrstani su u rodove Hucho i Salvelinus (Phillips i Oakley, 1997).
Oakley i Phillips (1999) su sekvencionirali gen hormona rasta (GH2C) i dobili
rezultate na osnovu kojih je rod Brachymystax predstavljen kao izvedeniji predstavnik
roda Hucho. Samim tim je odbačena hipoteza o rodu Brachymystax kao primitivnijem
(arhaičnom) rodu podporodice Salmoninae.
Sistematsko mesto roda Acantholingua je takođe izmenjeno nakon rezultata koje
su dobili Phillips i sar., (2000), sekvencioniranjem gena hormona rasta (GH2C), gena na
ribozomalnoj RNA (ITS1) i citohroma b na mitohondrijalnoj DNA. Na osnovu dobijenih
rezultata, rod Acantholingua filogenetski je najbliži rodu Salmo.
Slika 4. Reklasifikacija rodova podfamilije Salmoninae bazirana na molekularnim
podacima (mtDNA, GH2C i ITS1).
Sistematsko mesto roda Salmothymus takođe je bilo diskutabilno dok nisu urađene
analize nuklearne DNA i sekvencioniranja kontrolnog regiona i citohroma b
mitohondrijalne DNA (Slika 5). Na osnovu dobijenih rezultata, rod Salmothymus, isto kao
i rod Acantholingua, filogenetski je najbliži rodu Salmo (Snoj i sar., 2002).
Thymallus
Brachymystax
Acantholingua
Salmothymus
Hucho
Salvelinus
Salmo
Oncorhynchus
Thymallus
Salmothymus
Hucho
Brachymystax
Salvelinus
Acantholingua
Salmo
Oncorhynchus

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 17/141
Uvod
9
Slika 5. Intergeneričko stablo porodice Salmonidae, urađeno na osnovu analiza nuklearne
DNA i sekvencioniranja kontrolnog regiona i citohroma b mtDNA.
Rodovi Salmothymus, Acantholingua i Salmo čine posebnu grupu u okviru
intergeneričkog stabla, što govori o njihovim srodničkim vezama.
Dalja istraživanja išla su u pravcu utvr đivanja tačnog položaja roda Salmothymus i
Acantholingua unutar grupe Salmo (Slika 6). Istraživanja su vršena ispitivanjem LDH-
C1* i ITS1 gena nuklearne DNA i kontrolnog regiona mtDNA (Snoj i sar., 2002; Snoj,
2003). U analizama se kao veoma bitan pokazao LDH-C1* gen koji je sinapomorfnaodlika rodova Salmothymus, Acantholingua i Salmo, što ga čini validnim genetičkim
markerom za utvr đivanje filogenetskih odnosa ovih rodova.
0.1
Oncorhynchus keta
Oncorhynchus mykiss Salmo
salar
Salmo tru tta
Salmothymus
obtusirostris
Acantholingua
ohridana
Salveli nus fontinalis
Salvelinus alpinus
Coregonus lavaretus
Thymallus thymallus
Thymallus arcticus
Coregonus albul a
Br achymystax lenok

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 18/141
Uvod
10
Slika 6. Filogenetski odnosi unutar grupe Salmo, dobijeni na osnovu sekvencioniranja
LDH-C1* i ITS1 gena nuklearne DNA, kontrolnog regiona i citohroma b mtDNA. Da –
dunavska, At – atlantska, Ma – marmoratus i Ad – jadranska linija.
Na osnovu dobijenih rezultata (Snoj i sar., 2002; Snoj, 2003), autori su zaključili
da unutar grupe Salmo postoje tri podgrupe: Salmo salar , Salmo trutta kompleks koji
uključuje i Salmo marmoratus i treća podgrupa koju čine Acantholingua ohridana i
Salmothymus obtusirostris. Autori takođe smatraju da su vrste iz treće podgrupe
sestrinske i da predstavljaju evolutivni prelaz između Salmo salar i Salmo trutta
kompleksa, pri čemu su znatno bliže kompleksu Salmo trutta.
Autori su na osnovu dobijenih rezultata predložili reklasifikaciju roda
Salmothymus kao zasebnog roda, i predlažu svrstavanje vrsta istog unutar roda Salmo.
Sušnik i sar. (2004) na osnovu rezultata sekvencioniranja mtDNA zaključuju da
vrste Salmo marmoratus i Salmo ( Platysalmo) platycephalus ulaze u sastav filogenetskog
kompleksa Salmo trutta (Slika 7).
Oncorhynchus mykiss
Salmo salar
Salmo obtusir ostr is
Acantholingua ohr idana
Salmo tr utta/ marmoratus
complex
Da
At
Ma
Ad
100
100
94
81
0.01
Salveli nus fonti nali s

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 19/141
Uvod
11
Slika 7. Filogenetski odnosi unutar grupe Salmo, sa posebnim akcentom na položaj vrste
Salmo platycephalus, dobijeni na osnovu sekvencioniranja ITS1 gena nuklearne DNA,
kontrolnog regiona i citohroma b mtDNA (Sušnik i sar., 2004). Ma – marmoratus, Me –
mediteranska, Ad – jadranska, At – atlantska i Da – dunavska linija.
Utvr đivanje preciznih filogenetskih odnosa rodova i vrsta unutar porodice
Salmonidae je veoma bitno zbog njihove zaštite, jer je poznato da su mnoge vrste iz ove
porodice procenjene kao ugrožene (Crivelli, 1996).
Veliki broj autora se slaže da postojeće nedoumice u vezi filogenije rodova i vrsta
porodice Salmonidae mogu biti razrešene samo kombinovanjem rezultata morfoloških i
molekularnih analiza (Delling, 2003; Bernatchez i sar., 1992).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 20/141
Uvod
12
1.2.4. S ALMO TRUTTA (LINNAEUS, 1758) – POTOČ NA PASTRMKA
Pastrmka kao autohtona salmonidna vrsta naseljava vode Evroazije i severne
Afrike. Rasprostranjena je na područ ju od severne Norveške i severoistočnog dela Rusijena severu do planine Atlas u severnoj Africi na jugu, i od Islanda na zapadu do Aralskog
mora na istoku (Behnke, 1986; Elliott, 1994). Tokom prošlog veka pastrmku su naselili u
24 zemlje izvan Evrope, i to u preostali deo Azije i Afrike, zatim u Australiju, Severnu i
Južnu Ameriku (Laikre i sar., 1999).
Kod ove vrste zubi su raspoređeni na vomeru, vilicama, jeziku i na nepcu (Povž i
Sket, 1990). Zubi na vomeru su raspoređeni u dva reda i zadržavaju se tokom celog
života. U gornjoj vilici postoje mnogobrojni zubi usmereni unazad. Ovi zubi služe samo
za pridržavanje plena. Krljušti na telu su sitne, glava je bez krljušti. Branhiospina na
prvom škržnom luku ima od 18 - 24. Piloričnih nastavaka ima od 40 - 100, prosečno
65,38 (Vuković i Ivanović, 1971). Diploidan broj hromozoma iznosi 84, mada je
primećeno znatno variranje među intraspecijskim formama. Po bokovima, leđima,
škržnim poklopcima i leđnom peraju nalaze se brojne tamne, crvene i/ili narandžaste pege
(Povž i sar., 1996). Boja tela veoma varira i zavisi od osobina staništa. Dužina je obično
oko 40 cm, težina do 800 gr.
Potočna pastrmka naseljava hladne vode i to obično gornje tokove reka umereno
kontinentalnog klimata, mada se može naći i u ravničarskim rekama borealne zone, kao i
u jezerima sa čistom i hladnom vodom. Polni dimorfizam je izražen u doba mresta: ženke
imaju zaobljen trbuh i crven nabubreli polni otvor, dok su mužjaci uskog trbuha i nemaju
nabubreli polni otvor, ali se odlikuju veoma razvijenom donjom vilicom u obliku kuke.
Mresti se krajem jeseni i početkom zime - od novembra do januara. RF ženke iznosi do
2000 jaja na kg telesne težine. Ikra se odlaže na kamenitom dnu sa brzim tokom vode.
Prečnik jaja je veliki i iznosi 4.5 do 5 mm. Inkubacioni period traje, zavisno od
temperature vode, od 60 do 90 dana. Larve izvaljene u januaru ili februaru imaju veliku
žumančanu kesicu, koja im omogućuje ishranu, ali otežava kretanje. Aktivan život larvi,
odnosno njihov izlazak iz skrovišta, nastaje sa trošenjem i smanjivanjem žumančane
kesice na 1/3 njene veličine. U prirodnim plodištima mlađ ostaje do početka jeseni, kada
naraste do 10 cm dužine. Ona se kasnije nizvodno seli u dublje i mirnije vode u potrazi za
hranom. Polnu zrelost dostižu u 2 - 3 godini. U prvoj godini jedinke narastu 10 do 14 cm.
Ove ribe imaju juvenilni kolorit, tzv. "mladalačko ruho", istaknuto sa desetak crnih

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 21/141
Uvod
13
vertikalnih mrlja na bokovima tela. Sa porastom mlađi nestaju ova obeležja. Totalna
dužina dvogodišnjih jedinki može dostići 20 do 25 cm, a težina 150 do 200 g (Jevtić,
1989). Rastu dosta sporo. U akumulacionom jezeru Lokvara (blizu Delnica), koje pripada
dunavskom slivu, ulovljena je 1968. godine potočna pastrmka dužine 124 cm i težine od
25.5 kg, stara 15 ili 16 godina (Pažur, 1969).
Potočna pastrmka se hrani različitim organizmima: ribama, larvama vodenih
insekata, ikrom drugih riba, insektima koji lete nad površinom vode i padaju na vodu,
račićima i drugim beskičmenjacima (Aganović, 1979).
Od uvođenja binomijalne nomenklature do danas, opisana je, na osnovu
morfologije, 1931 vrsta evropskih salmonida. Ovako veliki broj vrsta javlja se kao
posledica nedovoljnog poznavanja pravila nomenklature i morfologije riba, kao i primene
različitih koncepata vrste. U poslednjih dvesta godina dato je 57 naučnih imena za
potočnu pastrmku Salmo trutta sensu stricto (Kottelat, 1997).
Pastrmka je izuzetno prilagodljiva različitim uslovima staništa spram kojih
pokazuje visok nivo fenotipske plastičnosti. Zbog toga danas postoji veliki broj
geografski specifičnih populacija, sa karakterističnim morfološkim odlikama, i to je
glavni razlog zašto je za jednu vrstu dato toliko naučnih imena.
Postoje tri različita ekološka oblika koje potočna pastrmka razvija u zavisnosti od
uslova staništa:
Salmo trutta forma fario – rečni oblik (Slike 10, 11, 12 i 13)
Salmo trutta forma lacustris – jezerski oblik (Slika 8)
Salmo trutta forma trutta – morski oblik (Slika 9)
Morski i jezerski oblik predstavljaju migratorne populacije, dok rečni oblik uvek
naseljava rečno stanište u okviru koga može preduzimati manje ili veće migratorne pokrete. Morska forma živi i hrani se u moru, kada postane polno zrela migrira u reke na
mrest, a nakon mresta mladi se vraćaju u more. Jezerska forma naseljava jezera, a na
mrest, kao i morska forma, migrira u reke, ili pak u pliće delove jezera (Elliott, 1994).
Migratorne i stalne populacije mogu istovremeno naseljavati neku reku i pojedine studije
pokazuju da se mogu nesmetano ukrštati (Hindar i sar., 1991), mada mehanizam
nasleđivanja migratornosti/stacionarnosti još uvek nije poznat.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 22/141
Uvod
14
Na osnovu rezultata dobijenih sekvencioniranjem mtDNA utvr đeno je da različiti
ekološki oblici nisu genetički uslovljeni, a samim tim ne moraju predstavljati
monofiletske grupe (Ryman 1983; Bernatchez i sar., 1992; Cross i sar., 1992).
Jezerska forma zbog uslova staništa može narasti i preko 20 kg, na bokovima tela
dominiraju crne krupne tačke, crvene su vrlo retke ili ih uopšte nema.
Slika 8. Salmo trutta forma lacustris – jezerski oblik
Slika 9. Salmo trutta forma trutta – morski oblik
Morska forma je veoma slična atlantskom lososu. Po bokovima tela ima veliki
broj crnih piknji nepravilnog oblika. Prvi o njoj je pisao Chiereghini (1818) i dao joj ime
Salmo cenerinus, a posle njega Kolombatović (1890) koji ju je preimenovao u Trutta
adriatica. Snoj i sar. (2002) su, na osnovu rezultata dobijenih sekvencioniranjem mtDNA,
utvrdili da morska forma pripada atlantskoj liniji, pri čemu se smatra da su analizirani
uzorci iz severnog Jadrana alohtonog (ribnjačkog) porekla.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 23/141
Uvod
15
Na osnovu genetičkih analiza (RFLP ( Restriction Fragment Length
Polymorphism), alozimski polimorfizam, sekvenciranje mtDNA) koje su rađene u
poslednjih 20 godina, došlo se do zaključka da pastrmke određenog geografskog područ ja
pokazuju značajne specifične sličnosti i na genetičkom nivou. Utvr đene genetičke
sličnosti se povezuju isključivo sa geografskim područ jima (basenima) i nemaju nikakve
veze sa ekološkim formama (Guyomard i Krieg, 1983; Bernatchez i sar., 1992; Giuffra i
sar., 1996; Largiader i Scholl, 1996; García-Marín i Pla, 1996; García-Marín i sar., 1999;
Vollestad i Hindar, 1997; Berrebi i sar., 2000).
Bernatchez i sar. (1992) su se bavili utvr đivanjem nukleotidnog redosleda unutar
640 bp dugog kontrolnog regiona mtDNA. U analizu su uključili sve tri ekološke forme
(rečnu, jezersku i morsku), glavaticu (Salmo marmoratus), gardsku (Salmo carpio) i
korzikansku pastrmku (Salmo macrostigma). U analizu su uključene, geografski
posmatrano, ukupno 24 populacije pastrmki iz atlantskog, dunavskog, sredozemnog i
jadranskog sliva. Cilj je bio da se na osnovu dobijenih rezultata utvrde filogenetski odnosi
morfološki i geografski različitih populacija pastrmki u Evropi. Autori su utvrdili 21
polimorfno mesto i ukupno 12 haplotipova (haploidni genotip – genetička varijanta) koje
su po obradi podataka razvrstali u pet filogenetskih grupa – linija, koje su se uglavnom
poklapale sa specifičnim geografskim poreklom uzorka:
1. jadranska linija (Ad) (Slika 10)
2. sredozemna (mediteranska) linija (Me) (Slika 11)
3. dunavska linija (Da) (Slika 12)
4. atlantska linija (At) (Slika 13)
5. marmoratus linija (Ma) (Slika 14)
Populacije potočne pastrmke koje pripadaju istoj liniji poseduju neke zajedničkemorfološke osobine.
Karakteristike spoljašnjeg izgleda mediteranske i jadranske linije su veliki broj
sitnih crvenih i crnih tačaka koje su nepravilno raspoređene po celom telu, kao i četiri
široka tamna pojasa, koji kao obruči obavijaju telo i to jedan u nivou glave, dva u nivou
trupa i jedan u nivou repa. Širina pojaseva je individualna (Odak, 2004).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 24/141
Uvod
16
Slika 10. Jadranska linija potočne pastrmke
Slika 11. Sredozemska (mediteranska) linija potočne pastrmke
Dunavska linija ima krupnije crvene i crne tačke podjednako zastupljene i
ravnomerno raspoređene po celom telu.
Slika 12. Dunavska linija potočne pastrmke
Atlantska domestifikovana linija ima karakterističnu crnu pigmentaciju koja
preovladava nad crvenom i nalazi se uglavnom na trupnom delu neposredno iza glave
(Giuffra i sar., 1996).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 25/141
Uvod
17
Slika 13. Atlantska linija potočne pastrmke
Marmoratus linija (glavatica), ima karakteristične mramorne šare po kojima je i
dobila ime. Osnovna boja tela je maslinasto zelena ili braonkasta, a šare se nalaze na
leđima i bokovima tela i nešto su tamnije od osnovne boje. Pojedini primerci mogu imati i
četiri široka tamna pojasa.
Slika 14. Marmoratus linija potočne pastrmke
Prema teoriji molekularnog sata, gde razlika u sekvenci nukleotida od 1% do 2%
predstavlja milion godina (Smith, 1992; Bernatchez, 2001) smatra se da su se pre
približno 10 miliona godina od zajedničkog pretka razdvojile dve vrste, losos (Salmo
salar ) i pastrmka (Salmo trutta). Bernatchez (2001) smatra da se razdvajanje pastrmskih
filogenetskih linija od zajedničke predačke populacije dogodilo se u periodu od pre 0,5 do
2 miliona godina, što ukazuje da su veoma značajnu ulogu pri formiranju filogeografskih
linija imale klimatske promene koje su se desile tokom pleistocenskih glacijacija (700 –
10 (14) hiljada godina). Poslednja velika pomeranja unutar linija desila su se po
poslednjoj glacijaciji (pre oko 20000 – 14000 godina).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 26/141
Uvod
18
Pošto je atlantski basen najduže bio izložen uticaju ledenog doba smatra se da je
atlantska linija imala najskoriju demografsku ekspanziju koja se dogodila pre 26000 –
13000 godina, što se poklapa sa početkom kraja poslednje glacijacije (pre oko 18000
godina). Dunavska linija imala je stariju najznačajniju demografsku ekspanziju mnogo
pre At linije, pošto na nju ledena doba nisu posebno uticala i to pre 300000 – 150000
godina, što se poklapa sa najznačajnjim interkonekcijama (Crno more, Kaspijsko i
Aralsko jezero) i širenjem mora (pre oko 28000 godina). Vreme demografske ekspanzije
unutar jadranske linije najverovatnije se desilo između At i Da linije, pre 77000 – 135000.
Smatra se da ledeno doba nije imalo uticaja na mediteransku i marmoratus liniju
(Bernatchez i sar., 1992; Bernatchez, 2001).
Slika 15. Geografska distribucija pet glavnih mtDNA evolutivnih linija potočne pastrmke
u Evropi (Bernatchez, 2001).
- atlantska linija, - dunavska linija, - jadranska linija
- marmoratus linija- mediteranska linija

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 27/141
Uvod
19
Pet filogenetskih linija koje je opisao Bernatchez i sar. (1992), grupa naučnika koja
je udružena u konzorcijum «TROUTCONCERN» (Laikre i sar., 1999) objedinila je u tri
grupe:
•
pastrmke mediteransko – jadranskog regiona• pastrmke crnomorskog, kaspijskog i aralskog basena
• pastrmke atlantskog regiona
Mediteransko – jadranski region predstavlja oblast u kojoj Salmo trutta – complex
pokazuje najveću fenotipsku raznolikost (Behnke, 1968). Nekoliko oblika potočne
pastrmke sa promenljivim taksonomskim statusom, u zavisnosti od autora, karakteristično
je upravo za ovaj region, a posebno za oblast Balkana i Turske (S. macrostigma, S.
dentex, S peristericus, S. marmoratus, S. carpio, S. obtusirostris, S pelagonicus, S.
farioides, S. macedonicus, S. lumi, S. letnica). Popisi navodnih vrsta ili podvrsta mogu se
pronaći kod Behnke (1965; 1968); Banarescu i sar. (1971); Economidis i Banarescu
(1991); Kottelat (1997); i Dorofeeva (1998). Genetičke studije zasnovane na jedarnoj i
mitohondrijalnoj DNA pokazuju visok nivo genetičke raznovrsnosti (kvantitativno (npr.
veliki broj različitih haplotipova), ali ne i kvaltativno (sa malim međusobnim razlikama))
između populacija potočne pastrmke ovog regiona. Kao rezultat genetičkih analiza
utvr đena su samo dva jasno različita entiteta: mediteranske populacije potočne pastrmke
(Salmo trutta) i glavatica (Salmo marmoratus). Primena novih molekularnih tehnika nije
potvrdila taksonomski položaj populacija fenotipski karakterističnih za Balkansko
poluostrvo, u sistemu klasifikacije zasnovanom na fenotipskim karakteristikama koje su
pod uticajem kompleksa biogeografskih faktora (Karakousis i Triantaphyllidis, 1990;
Apostolidis i sar., 1997). Patarnello i sar. (1994) su na osnovu sekvencioniranja delova
mtDNA otkrili izuzetno nisku stopu genetičkih razlika kod populacija koje su fenotipski
veoma različite (Salmo macrostigma, Salmo carpio i Salmo fibreni).
Crnomorski, kaspijski i aralski basen predstavljaju više od 50% ukupne teritorije
koju naseljava potočna pastrmka u Evropi. Iako se radi o izuzetno velikoj teritoriji, broj
genetički analiziranih populacija je svega desetak i to sa teritorije bivšeg Sovjetskog
Saveza (Bernatchez i sar., 1992; Bernatchez i Osinov, 1995; Riffel i sar., 1995; Largiadèr
i Scholl, 1995; Osinov i Bernatchez, 1996). Sekvencioniranjem mtDNA unutar ovog
regiona pronađene su dve od pet filogenetskih linija: dunavska i atlantska. Dunavska

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 28/141
Uvod
20
linija značajno preovlađuje, dok je svega nekoliko jedinki određeno kao pripadnici
atlantske linije. Autori smatraju da nije moguće sa sigurnošću zaključiti da li je prisustvo
atlantske linije posledica prirodne kolonizacije ili introdukcije (Osinov i Bernatchez,
1996).
Na bazi morfoloških i ekoloških razlika, populacije crnomorskog, kaspijskog i
aralskog basena bile su klasifikovane u posebne taksone (Berg, 1948). Populacije
crnomorskog basena su prepoznate kao Salmo trutta labrax, kaspijskog basena kao Salmo
trutta caspius, a populacije aralskog basena kao Salmo trutta oxianus. Posebna
ekofenotipska forma iz jezera Sevan (kaspijski basen) prepoznata je kao posebna podvrsta
Salmo trutta ischchan. Kombinacijom dobijenih rezultata analize alozima i mtDNA,
Bernatchez i Osinov (1995) i Osinov i Bernatchez (1996), smatraju da se ne može dati
podrška za predhodno iznetu taksonomsku originalnost.
Iako je obrađeno vrlo malo uzoraka dunavske linije iz crnomorskog, kaspijskog i
aralskog basena, utvr đen je visok nivo genetičke diferencijacije populacija (Bernatchez i
Osinov, 1995; Osinov i Bernatchez, 1996).
Atlantska filogeografska linija naseljava reke atlantskog basena od Islanda i
Norveške na severu do Iberijskog poluostrva i Atlas planine u Maroku na jugu, takođe
naseljava reke sliva Baltičkog i Severnog mora. Na osnovu genetičkih rezultata došlo se
do zaključaka da su populacije atlantske linije prilično homogene i da je nivo genetičke
diferencijacije manji u odnosu na mediteranske i dunavske populacije. Utvr đene su
značajne razlike na osnovu analize nuklearne i mtDNA između južnih iberijskih i
severnih populacija (Moran i sar., 1995; Antunes i sar., 1999; Bouza i sar., 1999; García-
Marín i sar., 1999; Weiss i sar., 2000). Utvr đene razlike smatraju se posledicom
neistovremenog naseljavanja severnih delova atlantskog basena nakon poslednje
glacijacije. Na bazi diskontinuiteta u učestalosti alela LDH-C1*(=LDH-5*)100 i *90 Ferguson i Fleming (1983) smatraju da su Velika Britanija i Irska rekolonizovane u
postglacijalnom periodu sa dve «grupe» potočne pastrmke. Prva rekolonizaciona grupa
nazvana je «ancestralna» i odlikovala se prisustvom LDH-C1*100 alela, dok je druga
rekolonizaciona grupa nazvana «moderna» i odlikovala se prisustvom alela LDH-C*90.
Hamilton i sar. (1989), na osnovu dopunskih analiza, proširuju hipotezu o dve nezavisne
rekolonizacije na preostali deo severo-zapadne Evrope potočnom pastrmkom. Filogenija
mtDNA haplotipova i njihov geografski raspored sugerišu da je postglacijalnarekolonizacija severo-zapadne Evrope bila znatno složenija; pretpostavlja se da je bilo

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 29/141
Uvod
21
više od dve rekolonizacije, kako je prethodno predloženo (Hynes i sar., 1996; García-
Marín i sar., 1999). Atlantska linija je pronađena i u dunavskom slivu, ali nema podataka
da li se radi o prirodnom ili veštačkom procesu (Duftner i sar., 2003). Bernatchez i sar.
(1992), smatraju da je moguće da se radi o prirodnoj kolonizaciji, koja se mogla desiti u
periodu interglacijacija. Osim u dunavskom, atlantska linija je pronađena u
mediteranskom basenu, što se smatralo posledicom čovekove aktivnosti (Poteaux i sar.,
1998). Međutim, na Siciliji su nedavno pronađene autohtone populacije atlantske linije
koje najverovatnije vode poreklo iz severne Afrike (usmeno saopštenje Aleš Snoj).
1.2.5. METODE RAZLIKOVANJA RIBLJIH POPULACIJA
Metode razlikovanja ribljih populacija podeljene su u dve osnovne grupe:
1. Morfološke metode
2. Genetičke metode
1.2.5.1. Morfološke metode razlikovanja ribljih populacija
Morfometrija i meristika su najčešće primenjivane morfološke metode. Ove
metode se koriste pre svega zbog lake primene i ekonomskih razloga. Ako se odabere
pravi set karaktera i ako se pravilno izvrše analize, dobijeni rezultati će biti bez sumnje
validni.
Morfometrija jeste oblast bioloških istraživanja u kojoj se kvantitativnim
analizama opisuje forma (oblik i veličina) nekog morfološkog entiteta, bića ili
pojedinačne karakteristike (Oxnard, 1978). Osnovu ovakvih analiza čini statistički pristup
obradi geometrijskih informacija o istraživanom morfološkom objektu. Morfometrijske
karakteristike koje se najčešće koriste u razmatranju veličine i oblika tela jesu rastojanja
između anatomskih tačaka smeštenih na longitudinalnoj osi (opšte dužinske
karakteristike), dorzo-ventralnoj osi (karakteristike visine) i osi koja povezuje levu i
desnu stranu tela (karakteristike širine). U cilju što tačnijeg određenja oblika morfoloških
struktura koriste se i takve karakteristike koje predstavljaju udaljenost između anatomskih
tačaka postavljenih na neortogonalnim (kosim) osama, čiji su uglovi manji ili veći od
pravog u odnosu na pomenute ose (Sttraus i Bookstein, 1982; Bookstein i sar., 1985).
Morfološka istraživanja, kako ona koja se bave spoljašnjom morfologijom, tako i

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 30/141
Uvod
22
osteološka, često su izvor podataka za analizu filogenetskih odnosa pojedinih vrsta riba
(Klykanov, 1975; Shaposhnikova, 1975), bez obzira na nivo klasifikacije. Zadatak takvih
istraživanja je utvr đivanje stanja karakteristika i njihove učestalosti u istraživanim
taksonima. Time se utvr đuje status pojedinih stanja karakteristika u filogenetskom smislu,
tj. jesu li stanja predačka (pleziomorfna) ili izvedena (apomorfna) (Hennig, 1966), a i
rekonstruišu filogenetski odnosi na osnovu ocene stanja tih karakteristika.
Merističke osobine imaju jednostavniju naslednu osnovu u odnosu na
morfometrijske, pa su stoga lakše za analizu ribljih populacija. Meristički karakteri su
diskretni ili brojivi i u merističkim analizama ribljih populacija najčešće se koriste
karakteri kao što su: broj žbica (tvrdih i mekih) u perajima, broj krljušti u bočnoj liniji,
broj piloričnih nastavaka, broj kičmenih pršljenova, broj branhiospina i dr. (Alegria -
Hernandez, 1985)
1.2.5.2. Genetičke metode razlikovanja ribljih populacija
Kao rezultat brzog razvoja molekularne genetike u poslednjih 20 godina, danas
imamo nekoliko laboratorijskih tehnika koje su upotrebljive za akumulaciju genetičkih
podataka na populacijama potočne pastrmke, kao i za druge vrste. Upotrebu molekularnih
metoda pratio je nastanak molekularne sistematike koja se bavi analizom strukture
makromolekula, na osnovu dobijenih rezultata rekonstruiše evoluciju gena i organizama, a
samim tim i objašnjava genetičku raznovrsnost (Moritz i Hillis, 1996). Genetička
raznovrsnost populacija može se utvrditi korišćenjem molekularnih markera koji
predstavljaju sekvencu na molekulu DNA ili proteinu koja se može lako detektovati i čije
je nasleđivanje moguće pratiti (Ford-Lloyd, 1996). Polimorfizam na molekulu DNA javlja
se kao posledica mutacija koje se manifestuju kao supstitucije, a ređe kao insercije ili
delecije, ili pak grešaka prilikom replikacije. Izučavanje polimorfizma je omogućeno
otkrićem DNA tehnika, na prvom mestu lančane reakcije polimeraze, a nakon toga
primene restrikcionih endonukleaza koje seku molekul DNA na fragmente određene
dužine (Ryman i Utter, 1987).
Najčešće korišćene metode molekularne genetike u ispitivanju varijabilnosti
ribljih populacija su alozimi i noviji genetički markeri kao što su ponavljajuće sekvence
jedarne DNA – mikrosateliti i mitohondrijska DNA.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 31/141
Uvod
23
Upotrebom različitih genetičkih markera, kako mitohondrijskih tako i jedarnih,
selektivno neutralnih lokusa (mikrosatelita), kao i lokusa koji su pod uticajem selekcije
(pojedini alozimi), možemo najlakše utvrditi genetičku strukturu populacije. Što više
markera upotrebljavamo, dobijamo potpuniju sliku genetičkog polimorfizama (Berrebi i
sar., 1999).
1.2.5.2.1. Alozimi
Do pred kraj prošloga veka široko primenjivana metoda za utvr đivanje genetičke
raznovrsnosti u molekularnoj genetici bila je elektroforeza alelskih varijanti proteina
(alozima) (Aebersold i sar., 1987; Utter i sar., 1987; Morizot i Schmidt, 1990; May,
1992). Upotreba alozima omogućava utvr đivanje alelskih varijanti, što se može iskoristiti
za ocenjivanje genetičke varijabilnosti prirodnih populacija, praćenje protoka gena,
rešavanje problema hibridizacije, kao i za utvr đivanje filogenetskih odnosa (Ferguson i
Masson, 1981; Murphy i sar., 1996). Mada su danas razvijene mnoge savremenije
tehnike, alozimska elektroforeza je još uvek cenjena i vrlo korisna metoda za utvr đivanje
značajnog nivoa genetičke varijabilnosti na populacijama potočne pastrmke, pre svega
zbog svoje niske cene u poređenju sa drugim savremenim molekularnim metodama i zbog
relativno lake upotrebe(Ferguson, 1989). Posebna prednost je u tome što postoji veliki
broj dostupnih podataka o alozimskoj varijabilnosti, što omogućava upoređivanje uzoraka
iz populacija koje poseduju značajnu vremensku i prostornu udaljenost. Osim prednosti
koje ima upotreba alozimskih markera, postoje i određene mane na koje ukazuju
Gyllensten i Wilson (1987) i Billington i Hebert (1991). Smatra se da su alozimski
markeri pod uticajem selekcije, da zbog degeneracije genetičkog koda ne pokazuju realni
nivo polimorfizma, često se nalaze na vrlo malom kodirajućem delu genoma tako da ih je
vrlo teško detektovati elektroforezom i različite genetičke varijante mogu imati istu
mobilnost na gelu. Lewontin (1974), smatra da je elektroforezom moguće utvrditi samo
trećinu svih aminokiselinskih supstitucija, koje u stvari predstavljaju refleksiju
nukleotidnih supstitucija. Iz svega rečenog zaključuje se da na osnovu rezultata
elektroforetske analize nije uvek pouzdano donositi zaključke o genotipskoj
varijabilnosti. U današnje vreme ovoj metodi pripisuju još jednu veliku zamerku, a to je
žrtvovanje životinja koje je neophodno da bi se došlo do veće količine tkiva koju ova
metoda zahteva.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 32/141
Uvod
24
U poslednjih 30 godina prošloga veka urađen je veliki broj alozimskih analiza na
pastrmskim populacijama iz različitih geografskih područ ja Evrope. Prve analize su
urađene na pastrmskim populacijama atlantskog područ ja, i utvr đen je nizak nivo
genetičke varijabilnosti. Homogenost populacija ovog područ ja objašnjava se veoma
dugim zadržavanjem leda u severnim delovima atlantskog basena za vreme poslednje
glacijacije. Kao rezultat toga dolazi do znatnog sužavanja areala, migracija severnih
populacija ka jugu i ujedinjenja sa tamošnjim populacijama (Allendorf i sar., 1976;
Ryman i sar., 1979; Crozier i Ferguson, 1986; Hansen i sar., 1993). Slične alozimske
analize rađene su na utvr đivanju dijagnostičkih alela između atlantskih i mediteranskih
populacija, (Apostolidis i sar., 1996; García-Marín i sar., 1991) kao i na utvr đivanju
genetičke varijabilnosti unutar mediteranskih pastrmskih populacija (Guyomard i Krieg,
1983). Osinov (1984; 1989) je radio alozimske analize između populacija crnomorskog i
kaspijskog basena i uočio je određeni nivo genetičke varijabilnosti. Veći broj autora je
pokušao da, na osnovu alozimskih analiza, tj utvr đivanja alelskih varijanti koje su
karakteristične samo za ovu vrstu (Giuffra i sar., 1996), odredi sistematski status glavatice
(Salmo marmoratus). Na osnovu dobijenih rezultata autori određuju glavaticu kao
samostalnu, prvobitnu i u odnosu na potočnu pastrmku parapatričnu vrstu (Berrebi i sar.,
2000).
1.2.5.2.2. Savremeni genetički markeri
Relativno skorašnji razvoj novih i visoko varijabilnih genetičkih markera kao što
je materinski nasleđena mitohondrijalna DNA (Avise, 1994) i hipervarijabilni mini- i
mikrosateliti (Estoup i Angers, 1998; Goldstein i Schlötterer, 1998), pružili su nova
saznanja u ispitivanju varijabilnosti populacija potočne pastrmke, koja nisu bila dostupna
primenom samo alozimskih tehnika.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 33/141
Uvod
25
1.2.5.2.2.1. Ponavljajuća DNA
Ponavljajuću DNA nalazimo u dva oblika:
•
raspršena ponavljajuća DNA
• tandemski ponavljajuća DNA
Raspršenu ponavljajuću DNA delimo prema broju ponavljanja na:
• duge rasute elemente (LINE) – koji su duži od 1000 baza
• kratke rasute elemente (SINEs) – koji su kraći od 500 baza
Tandemski ponavljajuća DNA je sinonim za satelite, kod kojih se ponovljivost
tandemskih jedinica kreće između 103 i 107 ponavljanja po lokusu. Kada je broj
ponavljanja tandemskih jedinica znatno manji (do 100 po lokusu), koristimo termineminisateliti i mikrosateliti.
1.2.5.2.2.1.1. Mikrosateliti
Mikrosateliti su otkriveni kod svih do sada proučavanih eukariota, kod kojih su
ravnomerno raspoređeni po celom genomu, ali su uglavnom ograničeni na euhromatin.
Pronađeni su kako u kodirajućim tako i u nekodirajućim regionima, manji broj ih se
nalazi na telomernim i centromernim regijama (Tautz i sar., 1986; Tautz, 1993).
Učestalost pojave mikrosatelita je najmanje jednom na svakih 10 kb. Pronađeni su takođe
kod pojedinih eubakterija i prokariota, ali sa manjom učestalošću (Tautz, 1989).
Mikrosateliti predstavljaju jednostavne nukleotidne nizove koji se sastoje iz ponovljenih
osnovnih motiva, dugih od 1 – 6 baznih parova (bp), koji se unutar lokusa ponavljaju od 5
pa do preko 100 puta (Stallings i sar., 1991). Mikrosateliti kičmenjaka najčešće se javljaju
kao di-, tri- , i tetranukleotidne replike, sastavljene iz svih mogućih kombinacija
nukleotida u osnovnom motivu (Tautz, 1989). Najčešće ponavljajući motiv je CA, a posle
njega AG, AAC, AAG i AAT (Glenn, 1995). Mikrosateliti sadrže veliki broj replika
osnovnog motiva i veoma su podložni mutacijama (10-3 do 10-4 po lokusu u generaciji
kod dinukleotidnih mikrosatelita) koje se javljaju zbog grešaka pri replikaciji DNA, što se
izuzetno odražava u dužinskom polimorfizmu (Amos i sar., 1993). Stopa mutacija
mikrosatelita razlikuje se između replika sa različitim osnovnim motivom, između
različitih lokusa sa replikama istog motiva i između alela istog lokusa (Schlötterer, 1998).
Stopa polimorfizma je uglavnom srazmerna broju replika osnovnog motiva. Kod

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 34/141
Uvod
26
korišćenja mikrosatelita kao markera kod kojih razlikujemo alele na osnovu različitog
broja ponavljanja osnovnog motiva može se javiti problem u neslaganju dužine
mikrosatelitskih fragmenata i njihovog nukleotidnog redosleda. Angeres i sar. (1995) su
analizirali nukleotidni redosled mikrosatelitnih alela jednoga lokusa kod dvanaest
salmonidnih vrsta i zaključili su da pojedini mikrosatelitni aleli kod različitih vrsta mogu
imati jednaku dužinu, ali različit nukleotidni redosled. Autori upozoravaju da se zaključci
o evolutivnim odnosima ne mogu izvoditi isključivo na osnovu dužine mikrosatelitnih
alela, jer različite mutacije vrlo lako mogu rezultirati jednakom konačnom dužinom alela.
Za mikrosatelite se još uvek ne zna način nastanka, odnosno njihova evolucija, zbog čega
se i ne mogu koristiti kao filogenetski markeri.
Mikrosateliti su veoma informativni kao genetički markeri iz više razloga: zbog
visoke stope genetičke varijabilnosti, jednostavnog kodominantnog mendelističkog
nasleđivanja, sačuvanosti mikrosatelitskih lokusa među srodnim vrstama. Spadaju u grupu
neutralnih genetičkih markera jer nisu podložni jakom selekcionom pritisku (Queller i sar.,
1993; Dowling i sar., 1996; Goldstein i Pollock, 1997). Jednostavno se mogu izolovati iz
materijala koji ne zahteva žrtvovanje životinja (krvi, dlake, slina, izmeta (Constable i sar.,
1995), krljušti (Miller i Kapuscinski, 1996), i arheološkog materijala (Bruford i sar., 1998).
Pri analizama možemo upotrebiti veliki broj lokusa, a jednostavna optimizacija lančane
reakcije polimeraze omogućava amplifikaciju više različitih lokusa u jednoj reakciji (Jug,
2002).
Zbog svih predhodno nabrojanih osobina mikrosateliti imaju veoma široku
upotrebu. Koriste se za utvr đivanje genetičke varijabilnosti i populacione strukture u
okviru mnogo kraćih geografskih udaljenosti nego što je to ranije bilo moguće (Estoup i
sar., 1998). Koriste se pri utvr đivanju porekla i srodničkih odnosa između jedinki
(Marshall i sar., 1998; Queller i sar., 1993; Hansen i sar., 1997; Fontaine i Dodson, 1999),
koriste ih pri kartiranju gena (Weissenbach i sar., 1992; Hearne i sar., 1992), kao i pri proceni efektivne veličine populacije (Edwards i sar., 1992). Veoma značajnu ulogu
mikrosateliti imaju pri analizama muzejskog materijala (krljušti), jer na taj način možemo
dobiti podatke o genetičkoj strukturi populacije za dugi vremenski period (Nielsen i sar.,
1997; 1999a; b; Miller i Kapuscinski, 1997; Tessier i Bernatchez, 1999).
Mikrosatelitski markeri počinju intezivnije da se koriste u izučavanju salmonidnih
populacija početkom devedesetih godina prošloga veka. Ovi markeri su se pokazali kao
znatno uspešniji u odnosu na alozimske. Alozimski markeri se mogu koristiti zarazlikovanje populacija između basena (dunavski, mediteranski, atlantski), dok se

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 35/141
Uvod
27
mikrosateliti koriste za utvr đivanje mikrogeografskih diferencijacija između nedavno
razdvojenih populacija. Brunner i sar. (1998) i Angers i sar. (1995) su preko
mikrosatelitskih markera utvrdili visok nivo interpopulacione raznolikosti na izolovanim
populacijama roda Salvelinus. Rezultati ovih ispitivanja se mogu iskoristiti za očuvanje
biološke raznovrsnosti. Mikrosateliti se koriste za utvr đivanje stepena introdukcije
neautohtonih populacija u neki vodotok, kao i za praćenje dinamike obnavljanja
autohtonih populacija (Hansen i sar., 2000; Poteaux i sar., 1999). Estoup i sar. (1993) su
preliminarno objavili pregled polimorfizama mikrosatelitskih lokusa kod geografski
izolovanih populacija potočne pastrmke, a nešto kasnije pronađeni su i mikrosatelitski
markeri za razlikovanje glavatice (Salmo marmoratus) od ostalih pastrmskih linija (Snoj i
sar., 1997; Sušnik i sar., 1997).
1.2.5.2.2.2. Mitohondrijalna DNA
Mitohondrije su semiautonomne ćelijske organele koje su zbog posedovanja
sopstvene DNA (mitohondrijalna DNA – mtDNA), sposobne za autonomnu replikaciju
DNA, kao i za transkripciju i translaciju genetičke informacije u proteine. Mitohondrije
su prisutne u svim ćelijama kičmenjaka osim u eritrocitima sisara. Sličnost između
mtDNA i bakterijske DNA navodi na sumnju da su mitohondrije evolutivni potomci
slobodno živećih bakterija koje su postale subcelularne organele. U procesu integracije u
ćeliju izgubile su mnoge svoje gene ili su ovi prenešeni u jedarni genom. Današnja
mitohondrijalna DNA predstavlja samo deo nekada znatno većeg genoma, čiji su geni
gusto raspoređeni jedan uz drugi bez umetnutih introna, osim u kontrolnom regionu koji
predstavlja jedinu regulatornu sekvencu unutar genoma mtDNA (Harison, 1989).
Mitohondrijalna DNA je prvi put konstatovana kod abnormalnih fibroblasta pileta i u
kinetoplastima tripanozoma. Biofizička istraživanja su pokazala da se mtDNA razlikuje
od nukleusne po mnogim osobinama, pa i po baznom sastavu (Grozdanović-
Radovanović, 2000). Ispitivanja mtDNA na različitim životinjskim taksonima pokazala
su da postoje varijacije duž sekvence, kako u redosledu nukleotida, tako i u veličini gena
(Harrison, 1989).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 36/141
Uvod
28
1.2.5.2.2.2.1. Osobine mitohondrijalne DNA
Mitohondrijalna DNA predstavlja važan genetički marker koji se koristi za
utvr đivanje geneti
čke strukture populacije, taksonomskih razmatranja, filogenetskih
odnosa, ukrštanja, introgresije i praćenja procesa nastanka novih vrsta. Osnovne osobine
mtDNA zbog kojih se i pokazala kao veoma dobar genetički marker su: jednostavna
organizacija, materinsko nasleđivanje, odsustvo rekombinacija i srazmerno visoka stopa
mutacija (Avise i sar., 1987; Avise, 1994).
Mitohondrijska DNA kičmenjaka je zatvoreni kružni molekul, veličine između 14
– 26 kb (Billington i Herbert, 1991), a kod salmonida je utvr đena dužina od 16670 bp
(Berg i Ferris, 1984). mtDNA predstavlja značajan deo genoma koji čini do 1% ukupne
ćelijske mase DNA (Cables, 2001). Najveći deo mitohondrijskog genoma čine kodogene
regije, sastavljene iz ukupno 37 gena, od kojih su dva za kodiranje ribozomalne RNA, 22
gena koji kodiraju tRNA i 13 gena za kodiranje polipeptida angažovanih u transportu
elektrona i oksidativnoj fosforilaciji (Avise, 1994; Curole i Kocher, 1999). Na mtDNA
kodiraju se sledeći proteini: citohrom b, tri podjedinice citohrom oksidaze, dve
podjedinice ATP sintetaze i sedam podjedinica NADH dehidrogenaze (Gillham, 1994;
Montoya i sar., 1983). Pri prevođenju specifičnog genskog koda u mitohondrijske
proteine učestvuju molekuli rRNA i tRNA, kodirani sa mtDNA. Pored 37 gena na
mtDNA nalazi se i kontrolna regija (eng. D-loop region) dužine oko 1000 bp sa
sekvencama koje imaju ulogu u regulaciji replikacije i transkripcije. Procesi replikacije i
translacije su potpuno autonomni u odnosu na iste procese u jedarnoj DNA (Borst i
Grivell, 1981). Ako razmotrimo nukleotidni raspored mtDNA kod različitih kičmenjaka,
možemo utvrditi da su organizacija i položaj gena veoma dobro očuvani tokom evolucije,
kao da postoje razlike u genetskom kodu, što se može iskoristiti pri filogenetskim
studijama (Curole i Kocher, 1999).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 37/141
Uvod
29
Slika 16. Raspored gena na mtDNA, sa prikazom restrikcijskih mesta za pojedine
endonukleaze kod atlantskog lososa (Salmo salar ).
Molekuli mtDNA su prisutni sa oko 1000 kopija u somatskim ćelijama, zrele jajne
ćelije poseduju oko milion a spermatozoidi oko 50 kopija. Mitohondrijska DNA se kod
najvećeg broja životinja nasleđuje po materinskoj liniji, mada ima nekih primera biparentalnog nasleđivanja kod beskičmenjaka (Hoeh i sar., 1991). Kod materinskog
nasleđivanja samo ženski deo populacije prenosi mtDNA na potomstvo, zbog toga što pri
oplođenju jajne ćelije obično dolazi do razgradnje mitohondrija prispelih iz spermatozoida
(Harrison, 1989). Zbog haploidnosti mitohondrijalnog genoma efektivna veličina
populacije za mtDNA iznosi samo jednu četvrtinu jedarne DNA (Birky, 1983). Pošto se
mtDNA jednostrano nasleđuje, između molekula obično ne dolazi do rekombinacija, kao
što je slučaj sa jedarnom DNA, što znači da se mtDNA nepromenjena prenosi sa roditelja
na potomstvo. Pojava kada potomstvo u svim svojim ćelijama ima isti haplotip mtDNA
naziva se homoplazmija, međutim može se dogoditi da u jednom organizmu postoje
različiti haplotipovi mtDNA, gde se ovakva pojava naziva heteroplazmija, vrlo je retka i
prisutna samo u nekim generacijama (Avise, 2000). Heteroplazmija do sada nije zabeležena
kod salmonida, ali kod nekih drugih vrsta riba jeste, i javlja se kao posledica biparentalnog
nasleđivanja ili pak mutacionih promena na delu molekula mtDNA u organizmu
(Lightowlers i sar., 1997).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 38/141
Uvod
30
Mitohondrijsku DNA karakteriše vrlo visok stepen mutacija, što je veoma
značajno pri njenoj upotrebi kao genetskog markera. Utvr đeno je da stepen supstitucija
iznosi 5,7 x 10-8 za pojedinačno nukleotidno mesto za godinu dana, što znači da mtDNA
akumulira 2 – 4% mutacija na milion godina (Brown i sar., 1982; Lewin, 2000). Smatra
se da je uzrok visoke stope mutacija mtDNA njena intenzivnija dinamika i mogućnost
replikacije za 5 – 10 puta brže u odnosu na jedarnu DNA, a samim tim je i stepen zamene
nukleotida za 5 – 10 puta veći nego kod jedarne DNA (Brown i sar., 1979). Na visok
stepen mutacija u mtDNA utiču sledeći razlozi: neefikasnost mehanizama popravke
(Tomkinson i Linn, 1986; Lopez i sar., 1997), visoke koncentracije mutagenih materija
(superoksidni radikali, O-2), koji nastaju pri visokoj metaboličkoj aktivnosti mitohondrija
(Li i Grauer, 1991; Lopez i sar., 1997), nepouzdanost procesa replikacije DNA (Li i
Grauer, 1991), slabiji selekcioni pritisak u određenim regijama, koji zajedno sa
klonalnom selekcijom vrlo brzo dovodi do fiksacije mutacija (Cann i sar., 1984). Između
evolutivno udaljenih linija utvr đene su razlike u stepenu supstitucija istog gena. Kao
najznačajniji razlog za različite stope mutacija su oštećenja DNA koja nastaju kao
posledica sporednih produkata metabolizma (Martin, 1999). Na osnovu rezultata
restrikcije i sekvenciranja mtDNA utvr đeno je da frekvencija mutacija nije ista duž celog
genoma. Najnižu frekvenciju mutacija na genomu mtDNA imaju regije koje kodiraju
rRNA molekule (Ferris i Berg, 1988). Kontrolna regija predstavlja najvarijabilniji deo
mtDNA, koji zbog manjeg funkcionalnog pritiska poseduje najviše mutacija posebno na
3'- i 5'-kraju, a to su regije genoma koji nose zapise za tRNAPro i tRNAPhe.
Najkonzervativniji deo kontrolne regije je centralni deo koji broji oko 300 bp i odgovoran
je za početak replikacije, i ovaj deo poseduje veliki broj citozinskih i guaninskih baza
(Shedlock i sar., 1992). U kontrolnoj regiji roda Salmo javljaju se uglavnom tačkaste
mutacije u obliku supstitucija (tranzicije i transverzije), dok su insercije i delecije vrlo
retke. Nesklad u relativnoj stopi evolucije između mtDNA i jedarne DNA je jasan dokaz
da su genomi u različitim subćelijskim strukturama unutar organizama pod različitom
kontrolom i pod različitim evolutivnim pritiskom, koji se razlikuje između taksona (Brown
i sar., 1979).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 39/141
Uvod
31
1.2.5.2.2.2.2. Mitohondrijska DNA kao genetski marker
Zbog svojih osobina pomenutih u predhodnom poglavlju mtDNA se koristi kao
genetski marker pri filogenetskim studijama, zbog materinskog jednostranog nasleđivanja
koristi se za rekonstrukciju evolutivnih događaja uključujući migracije, introdukcije i
efekat «uskih grla» populacija. mtDNA se pokazala kao dobar marker za praćenje
efikasnosti uvođenja novih linija u akvakulturu, kao i za praćenje delovanja uvedenih
linija na autohtone populacije (Li i Grauer, 1991). Zahvaljujući mogućnosti izolaovanja
mtDNA čak i iz fosilnog materijala, omogućeno nam je upoređivanje izumrlih populacija
sa današnjim. U ovoj studiji koristili smo mtDNA kao marker pomoću koga je moguće
pratiti intraspecijsku genetičku varijabilnost između geografski odvojenih populacija,
odnosno populacija koje pripadaju istom ili različitim morskim slivovima.
Iako je utvr đivanje odnosa među srodnim vrstama na osnovu mtDNA u velikom
broju slučajeva u skladu sa morfološkim, alozimskim i drugim molekularnim metodama,
povremeno dolazi do neslaganja dobijenih rezultata (Harrison, 1989).
Giuffra i sar. (1996) su upoređivali rezultate dobijene upotrebom tri najčešće
korišćena genetska markera: alozimi, mtDNA i mikrosateliti. Na osnovu rezultata
interpopulacionog polimorfizma pastrmki za odgovarajući markerski sistem dobijena su
filogenetska stabla koja su se međusobno razlikovala, pa je i ocena filogenetskih odnosa
među populacijama pastrmki (morfološkim oblicima) različita. Kao razlozi zavisnosti
rezultata od vrste molekularnih podataka navode se: različite evolutivne stope, različite
stope homoplazije kod različitih markera, prisustvo introgresije među lokusima usled
nepotpune reproduktivne izolacije populacija. Različiti načini nasleđivanja i evolucija
mitohondrijskih i jedarnih gena, teoretski može dovesti do nesklada u rezultatima
korišćenjem ova dva markerska sistema (Bernatchez i sar., 1992)
Vrlo je malo studija u kojima su se uporedno pratile morfološke osobine i
genetički polimorfizam. Bernatchez i sar. (1992) i Giuffra i sar. (1994) su pokušali
utvrditi povezanost između haplotipova mtDNA i morfološki diferenciranih populacija
pastrmki. Kao rezultat dobili su da su se morfološki veoma slične populacije
raspoređivale u vrlo različite evolutivne linije. Patarnello i sar. (1994) su, tokom
istraživanja, utvrdili da su relativno male razlike na nivou mtDNA povezane sa relativno
velikim fenotipskim razlikama.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 40/141
Uvod
32
Pored svih pozitivnih osobina koje poseduje mtDNA kao genetički marker,
postoje i neke negativne. Maternalno nasleđivanje mtDNA onemogućava korišćenje ovog
markera pri razlikovanju hibrida u populacijama gde je eventualno došlo do introdukcije.
U tim slučajevima se preporučuje korišćenje informativnijih markera preko kojih
možemo pratiti i maternalno i paternalno nasleđivanje. Takođe, pri analizama vrlo
srodnih jedinki, upotreba mtDNA kao genetičkog markera se ne preporučuje zbog njene
premale varijabilnosti za tu vrstu studija.
Kako se mtDNA nasleđuje kao jedan gen, treba biti posebno oprezan pri
donošenju zaključaka na nivou čitave populacije, jer se evolucija pojedinačnog gena
može razlikovati od evolutivnog proseka celog genoma. Upotreba isključivo mtDNA kao
genetskog markera nosi sa sobom rizik od netačne rekonstrukcije filogenije (Zhang i
Hewitt, 2003).
Većina autora se slaže da je za utvr đivanje što tačnijih filogenetskih odnosa
između taksona potrebno uključiti što više genetičkih sistema, i dobijene rezultate
uporediti sa rezultatima morfoloških analiza.
1.2.5.2.2.2.3. Upotreba mtDNA za proučavanje vrsta i populacija salmonida
Tokom osamdesetih godina prošloga veka većina analiza mtDNA na salmonidama
zasnivala se na utvr đivanju inter- i intraspecijskog polimorfizma korišćenjem
restrikcionih endonukleaza (RFLP – tehnika).
Berg i Ferris (1984) prvi su analizirali mitohondrijalni genom kod salmonida
upotrebom restrikcionih enzima na četiri vrste iz porodice Salmonidae: pacifički losos
(Oncorhynchus tchawitcha), dužičasta pastrmka (Oncorhynchus mykiss), potočna
pastrmka (Salmo trutta) i potočna zlatovčica (Salvelinus fontinalis). Autori su procenili
dužinu genoma mtDNA na 16670 bp, i utvrdili su filogenetske odnose za date vrste, pri
čemu su zaključili da su pacifički losos i dužičasta pastrmka filogenetski bliže u odnosu
na potočnu pastrmku. Autori su utvrdili da se intraspecijski polimorfizam mtDNA kod
salmonida kreće od 0,1 do 2%, što je uočeno i kod većine drugih grupa kičmenjaka, dok
su razlike između vrsta od 3 do 14%.
Gyllesten i Wilson (1988) su radili slična istraživanja kao i Berg i Ferris (1984). U
analizu su uključili populacije potočne pastrmke iz različitih geografskih područ ja,
atlantskog lososa (Salmo salar ), dve vrste iz roda Oncorhynchus (Oncorhynchus mykiss i

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 41/141
Uvod
33
Oncorhynchus clarki) i potočnu zlatovčicu. Restrikcionom analizom cele mtDNA utvrdili
su bliske srodničke veze između populacija potočne pastrmke i atlantskog lososa na
jednoj strani i vrsta roda Oncorhynchus na drugoj strani.
Taylor i sar. (1999) su vršili ispitivanja na 47 populacija vrste Salvelinus
confluentus, koristeći RFLP – tehniku, i konstatovali su veoma malu varijabilnost
mtDNA, sa divergencijom od 1,2%. Brunner i sar. (1998) su, kao i njihovi prethodnici,
upotrebili RFLP tehniku, na dva regiona mtDNA vrste Salvelinus alpinus iz 12 jezera
alpske regije, i potvrdili su vrlo malu genetičku varijabilnost i nediferenciranost
populacija. Malu varijabilnost su objasnili posledicom nedavne diferencijacije populacija
iz jedne iste grupe, verovatno po završetku ledenog doba.
Početkom devedesetih godina prošloga veka počinje se sa utvr đivanjem redosleda
nukleotida (sekvencioniranjem) vezanim za posebne regione na mtDNA.
Shedlock i sar. (1992) su uradili prva detaljnija istraživanja kontrolnog regiona
kod salmonida. Sekvencionirali su ceo kontrolni region kod 6 vrsta iz roda
Oncorhynchus, atlantskog lososa i arktičkog lipljana (Thymallus arcticus). Zaključili su
da su delovi kontrolnog regiona koji su kod drugih kičmenjaka važili za konzervativne,
takođe očuvani i kod salmonida, kao i da su različiti delovi kontrolnog regiona pod
različitim selekcionim pritiskom.
Bernatchez i sar. (1992) radili su na utvr đivanju nukleotidnog redosleda unutar
640 bp dugog kontrolnog regiona mtDNA (strana 15). U analizu su uključene geografski
posmatrano ukupno 24 populacije pastrmki iz atlantskog, dunavskog, sredozemnog i
jadranskog sliva. Na osnovu dobijenih rezultata autori su utvr đene haplotipove razvrstali
u pet filogenetskih grupa - linija: sredozemna (mediteranska), jadranska, dunavska,
atlantska i marmoratus linija. Autori zaključuju da, na osnovu dobijenih rezultata,
pastrmka predstavlja jednu od najbolje genetički struktuiranih vrsta koju su proučavali.
Giuffra i sar. (1994) su sekvencionirali 5'- deo kontrolnog regiona, fragment genaza citohrom b i gen za ATP-aznu podjedinicu VI, na populacijama pastrmki
severnoitalijanskih voda. Analizirali su devet geografski različitih populacija potočne
pastrmke, osam geografski različitih populacija glavatice i gardsku pastrmku. Na osnovu
dobijenih rezultata potvr đuju pet filogenetskih grupa koje su utvrdili Bernatchez i sar.
(1992).
Apostolidis i sar. (1996a) bavili su se proučavanjem varijabilnosti istih delova
mtDNA na 11 gr čkih populacija potočne pastrmke, na atlantskoj i populaciji ohridske pastrmke. Sekvencioniranjem kontrolnog regiona pronađeno je 10 haplotipova sa

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 42/141
Uvod
34
interpopulacionim polimorfizmom od 0,32% do 2,31%. Autori zaključuju da je
interpopulacioni polimorfizam haplotipova kod pastrmki veoma visok.
Phillips i sar. (2000) su se bavili utvr đivanjem sistematskog položaja vrste
Acantholingua ohridana, između ostalog i sekvencioniranjem gena za citohrom b, dok su
se istim problemima vezanim za vrstu Salmothymus obtusirostris bavili Snoj i sar. (2002),
sekvencioniranjem kontrolnog regiona i citohroma b. Na osnovu dobijenih rezultata
utvr đeno je da obe vrste poseduju specifične mtDNA haplotipove koji ne spadaju u
prethodno pomenute linije.
Weiss i sar. (2000), sekvencioniranjem 5' kraja kontrolnog regiona mtDNA na
sedam portugalskih populacija potočne pastrmke, pronalaze pet novih haplotipova u
okviru At linije, koji su se razlikovali od opšteg severnog At1 haplotipa za dva do tri
mutaciona koraka. Na osnovu dobijenih rezultata predlažu re-evaluaciju postglacijalne
rekolonizacije severne Evrope, za koju se, do objavljivanja ovih rezultata, smatralo da je
rekolonizovana iz dva glavna glacijalna refugijuma (jugozapadnog atlantskog i ponto-
kaspijskog).
Weiss i sar. (2001) sekvencionirali su 5' kraj kontrolnog regiona mtDNA na 27
populacija potočne pastrmke u Austriji. Sve analizirane populacije pripadale su
dunavskom slivu, mada je 44% analiziranih jedinki imalo haplotipove iz At linije.
Pronalazak tako visokog procenta haplotipova At linije u dunavskom slivu otvorio je
pitanje eventualne prirodne kolonizacije, mada je poznato da je u Austriji bilo
poribljavanja upravo materijalom At linije.
Suárez i sar. (2001) sekvencionirali su ceo kontrolni region mtDNA populacija
potočne pastrmke Iberijskog poluostrva i severne Afrike. Na osnovu rezultata identifikuju
četiri glavne grupe haplotipova na Iberijskom poluostrvu: atlantsku, Duero, mediteransku
i andaluzijsku. Na osnovu opisane četiti iberijske grupe, povećavaju broj do tada poznatih
pet glavnih evropskih mtDNA grupa na šest: 1. Atlantska grupa koja uključuje dvarazličita grozda - južno evropski i severno atlantski; 2. Endemična grupa ograničena na
Duero basen Iberijskog poluostrva; 3. Jadransko – Andaluzijska grupa koja uključuje
Jadransko – Jonske populacije u Mediteranu i Andaluzijske populacije na jugu Iberijskog
poluostrva; 4. Mediteranska grupa koja je rasprostranjena od jugo-zapadnog basena
Iberijskog poluostrva do Jonskog basena; 5. Dunavska grupa koja je rasprostranjena u
basenu Crnog mora, Kaspijskog i Aralskog jezera; 6. marmoratus grupa ograničena na
basen Jadranskog mora.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 43/141
Uvod
35
Sušnik i sar. (2004), na osnovu rezultata sekvencioniranja kontrolnog regiona i
citohroma b mtDNA, zaključuju da vrste Salmo marmoratus i Salmo ( Platysalmo)
platycephalus ulaze u sastav filogenetskog kompleksa Salmo trutta, s tim da Salmo
marmoratus spada u liniju marmoratus, što je istakao i Bernatchez i sar. (1992), a Salmo
platycephalus u jadransku liniju.
Cortey i sar. (2004), sekvencioniranjem celog kontrolnog regiona mtDNA, opisuju
filogenetske odnose populacija potočne pastrmke u mediteranskom basenu Iberijskog
poluostrva sa osvrtom na istorijsku biogeografiju. U filogenetsku analizu uključuju svih
pet glavnih mtDNA linija, i zaključuju da je dunavska linija najancestralnija.
Nilsson i sar. (2001) utvr đuju, na osnovu polimorfizma mtDNA, dve glavne linije
atlantskog lososa (Salmo salar ) - severnoameričku i evropsku.
1.2.6. UGROŽENOST POTOČ NE PASTRMKE (S ALMO TRUTTA) U EVROPI
Potočna pastrmka predstavlja jednu od najbolje proučenih slatkovodnih vrsta riba.
Dosadašnje analize pokazuju da je veliki deo intraspecijske varijabilnosti potočne
pastrmke izgubljen, a da je preostali deo veoma ugrožen (Laikre i sar., 1999).
Veliki broj populacija potočne pastrmke ugrožen je razli
čitim tipovima ljudskih
aktivnosti, koje mogu biti podeljene u tri opšte kategorije: degradacija staništa, ribolov i
poribljavanje (Allendorf, 1988; Laikre i Ryman, 1996; Laikre i sar., 1999).
Degradacija staništa može biti direktna i indirektna. Direktna degradacija
podrazumeva fizičke promene staništa, na primer, podizanje hidroelektrana koje mogu
sprečavati migracije pri mrestu, ili pak regulacija rečnog korita. Indirektna degradacija
podrazumeva promenu hemijskih svojstava vode kroz razne vidove zagađenja (kisele
kiše, ispuštanje hemijskih materija u vodotokove i dr.). Degradacija staništa takođe
uključuje i promenu sastava ribljih zajednica (introdukcija egzotičnih ribljih vrsta ili
istrebljenje postojećih vrsta sa kojima je potočna pastrmka prirodno koegzistirala).
Ribolov, kako komercijalni tako i sportski, dovodi do smanjenja brojnosti, do
opadanja intrapopulacionog genetičkog diverziteta, a samim tim i do opadanja vijabilnosti
populacija.
Poribljavanje materijalom iz veštačkog mresta ili prenošenje jedinki sa drugih
lokaliteta sve je češća praksa upravljanja vodama. Do poribljavanja najčešće dolazi zbog
smanjenja brojnosti autohtonih populacija, što je uglavnom rezultat prevelikog ribolovnog

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 44/141
Uvod
36
opterećenja. Poribljavanje predstavlja naročito ozbiljnu pretnju otkako se posmatra kao
blagotvoran način da se pomogne autohtonim populacijama, ali u stvarnosti veoma često
dovodi do izumiranja lokalnih autohtonih genskih fondova (Ryman i Utter, 1987;
Allendorf i Leary, 1988; Ferguson, 1989; Hindar i sar., 1991a; Waples, 1991; Taylor,
1991; Leary i sar., 1993; Hansen i Loeschcke, 1994; Ryman i sar., 1995; Allendorf i
Waples, 1996).
Glavni ugrožavajući faktori po populacije potočne pastrmske u Srbiji su: preveliko
ribolovno opterećenje, degradacija staništa i poribljavanje.
Najznačajniji ugrožavajući faktor predstavlja preveliko ribolovno opterećenje,
koje uključuje ne samo legalan ribolov, već na prvom mestu krivolov. Glavni razlog
izuzetno velikog ribolovnog pritiska je nedovoljno i neprofesionalno sprovođenje
ribolovne kontrole do 2003 godine, kao i neprimenjivanje legislativno regulisanih
zaštitnih mera u ribarstvu.
Drugi po značaju ugrožavajući faktor predstavlja direktna degradacija staništa u
vidu formiranja vodozahvata za potrebe elektroprivrede (Božica, Vrla, Vlasina), za
vodosnabdevanje (Lopatnica, Mlava), za industrijska i rudarska postrojenja (Resava,
sokobanjska Moravica, Lopatnica), kao i izgradnja fabrika flaširane vode (Vrla) – primeri
su samo ilustracija i sigurno su daleko brojniji, ali i o tome nema sređenih i dostupnih
podataka. Socio-ekonomski aspekti korišćenja vode ovih reka kao resursa su nepobitno
važni, ali rešenje problema do kojih to korišćenje dovodi, pre svega problema po
očuvanje autohtonog diverziteta pastrmke u Srbiji, i pored kako-tako zakonom propisane
obaveze (Anonimno 1994; 2004) nije adekvatno u administrativno-upravljačkoj praksi
sektora zaštite životne sredine i zaštite prirode. Indirektna degradacija staništa svodi se na
eventualno zagađenje raznim vrstama hemikalija koje se koriste u poljoprivredi. Zbog sve
manje naseljenosti brdsko-planinskih krajeva kao i ekstenzivne poljoprivrede, ovaj tip
degradacije staništa ne predstavlja posebno značajan ugrožavajući faktor po pastrmske populacije.
Treći po značaju faktor ugrožavanja populacija potočne pastrmke je poribljavanje.
Ova aktivnost još uvek nije uzela previše maha, i do sada su poribljavane samo ribolovno
atraktivne vode, koje trpe veliki ribolovni pritisak. Evidencije o izvršenim
poribljavanjima na teritoriji Srbije uglavnom ne postoje, ali je poznato da je materijal za
poribljavanje nabavljan širom bivše Jugoslavije, ne vodeći računa čak ni o poribljavanju
materijalom koji pripada barem istom slivnom područ ju, a kamoli o poribljavanjumaterijalom autohtonog porekla, poznate genetičke strukture. Postoje negativna iskustva

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 45/141
Uvod
37
u drugim državama (Slovenija) gde se vršilo poribljavanje neautohtonim linijama. U Soču
je unešena potočna pastrmka (za koju se smatra da nije autohtona za sliv pomenute reke)
atlantske i dunavske linije, koja se nesmetano ukrštala sa marmoratus linijom dajući
plodne hibride i preteći da vremenom može doći do potpunog uništenja genetičke
originalnosti marmoratus linije.
Degradacija staništa, ribolov i poribljavanje mogu biti okidač za tri glavna procesa
koji dovode do smanjenja ili gubljenja intraspecijske raznolikosti, a to su: izumiranje
lokalnih populacija, hibridizacija i gubitak genetičke varijabilnosti unutar populacije
(Ryman i sar., 1995a; Laikre i Ryman, 1996). Ovi procesi se mogu odraziti na različite
hijerarhijske nivoe intraspecijske genetičke raznolikosti, na primer, između alela unutar
individue, između individua unutar populacije, između populacija unutar geografskog
regiona. Genetička raznolikost na bilo kom hijerarhijskom nivou biće smanjena ako
specifični aleli, genotipovi ili genski fondovi populacija prestanu da egzistiraju ili ako je
njihov integritet ugrožen hibridizacijom. Značaj genetičke raznolikosti ogleda se u boljem
prilagođavanju na sredinske promene.
Izumiranje lokalnih populacija može rezultirati potpunim gubitkom pojedinih
alela ili alelskih kombinacija. Degradacija staništa je glavni uzrok izumiranja lokalnih
populacija. Preteran izlov i nepromišljeno poribljavanje takođe mogu dovesti do istih
rezultata (Laikre i sar., 1999). Nepromišljeno poribljavanje dovešće, na prvom mestu, do
introgresivne hibidizacije, a na drugom mestu i do kompeticije za osnovne životne
resurse.
Dve najosnovnije preporuke genetičke konzervacije svode se na maksimalno
obraćanje pažnje ka očuvanju ekosistema i na utvr đivanje genetičke strukture populacije,
kao i minimalne vijabilne veličine populacije. Možemo zaključiti da praktično vrlo malo
znamo o ekonomskoj, ekološkoj i evolutivnoj vrednosti pojedinih gena i populacija.
Stoga fokus genetičke konzervacije mora biti usmeren na ispitivanje genetičkogdiverziteta između i unutar populacija unutar svake vrste (Frankel, 1970; 1974; Ryman i
Ståhl, 1980; Utter, 1981; Meffe, 1986; Ryman, 1991).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 46/141
Uvod
38
1.3. CILJEVI RADA
U okviru ovoga rada postavljeni su sledeći ciljevi:
Utvrditi genetičku raznovrsnost populacija potočne pastrmke na teritoriji
Republike Srbije, upotrebom molekularno genetičkih metoda.
Pronađene genetičke varijante (haplotipove) uporediti sa već poznatim i
razvrstati ih u postojeće filogeografske linije (Bernatchez, 1992).
Na osnovu dobijenih rezultata i korišćenjem postojećih, rekonstruisati
evolutivnu istoriju potočne pastrmke na teritoriji Republike Srbije.
Utvrditi ugrožavajuce faktore i efekte njihovog delovanja na autohtone
populacije
Dati predlog mera zaštite autohtonih populacija.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 47/141
Materijal i metode
39
2. MATERIJAL I METODE
2.1. MATERIJAL
2.1.1. PRIKUPLJANJE I ČUVANJE UZORAKA
Uzorci potočne pastrmke koji su analizirani u ovom radu sakupljeni su u periodu
od juna 1997. do juna 2004. godine. Sakupljanje materijala je vršeno na 35 lokaliteta u
okviru sva tri morska sliva (crnomorskog, egejskog i jadranskog) na teritoriji Republike
Srbije. Najveći broj analiziranih jedinki prikupljen je elektroribolovom koji se obavljao
uz pomoć elektroagregata marke Suzuki – Bosch, snage 2100W, izlazne struje 220V DC,
frekvencije 40 Hz i jačine u zavisnosti od osobina vode, maksimalno 8.5 A. Manji brojanaliziranih jedinki prikupljen je udičarskim alatom koji se koristi u rekreativnom
ribolovu.
Nakon izlova, jedinkama je odsečen deo analnog peraja (oko 0.5 cm2) koje ima
mogućnost regeneracije, i sačuvan u 96% etanolu, a jedinke su vraćene nazad u vodu.
Svaki uzorak je sačuvan u posebnoj epruveti sa obeleženim rednim brojem i nazivom
lokaliteta. Delić uzorkovanog peraja dalje je korišćen za molekularne analize.
Slika 17. Način uzimanja uzoraka (odsecanje analnog peraja i odlaganje u etanol).
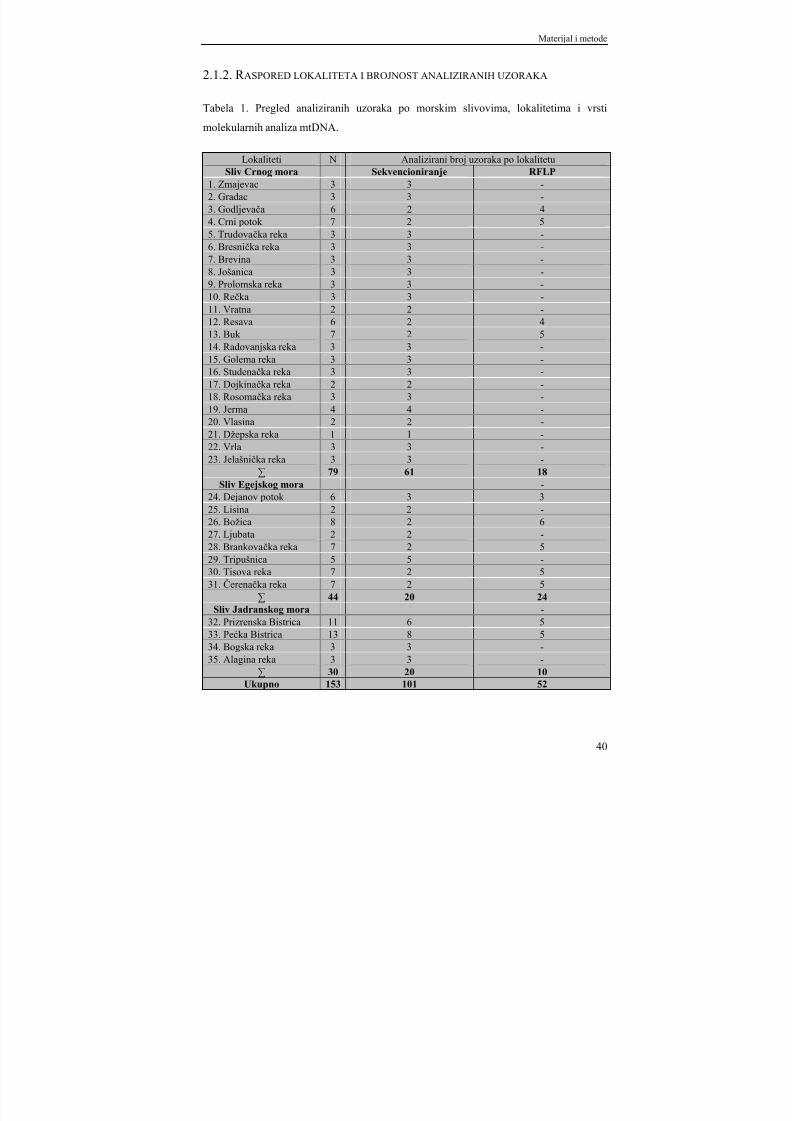
7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 48/141
Materijal i metode
40
2.1.2. R ASPORED LOKALITETA I BROJNOST ANALIZIRANIH UZORAKA
Tabela 1. Pregled analiziranih uzoraka po morskim slivovima, lokalitetima i vrsti
molekularnih analiza mtDNA.
Lokaliteti N Analizirani broj uzoraka po lokalitetuSliv Crnog mora Sekvencioniranje RFLP
1. Zmajevac 3 3 -2. Gradac 3 3 -3. Godljevača 6 2 44. Crni potok 7 2 55. Trudovačka reka 3 3 -6. Bresnička reka 3 3 -7. Brevina 3 3 -
8. Jošanica 3 3 -9. Prolomska reka 3 3 -10. Rečka 3 3 -11. Vratna 2 2 -12. Resava 6 2 413. Buk 7 2 514. Radovanjska reka 3 3 -15. Golema reka 3 3 -16. Studenačka reka 3 3 -17. Dojkinačka reka 2 2 -18. Rosomačka reka 3 3 -19. Jerma 4 4 -20. Vlasina 2 2 -21. Džepska reka 1 1 -22. Vrla 3 3 -23. Jelašnička reka 3 3 -
∑ 79 61 18
Sliv Egejskog mora -24. Dejanov potok 6 3 325. Lisina 2 2 -26. Božica 8 2 627. Ljubata 2 2 -28. Brankovačka reka 7 2 529. Tripušnica 5 5 -30. Tisova reka 7 2 531. Čerenačka reka 7 2 5
∑ 44 20 24
Sliv Jadranskog mora -32. Prizrenska Bistrica 11 6 533. Pećka Bistrica 13 8 534. Bogska reka 3 3 -35. Alagina reka 3 3 -
∑ 30 20 10
Ukupno 153 101 52

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 49/141
Materijal i metode
41
Slika 18. Prostorni raspored analiziranih lokaliteta na teritoriji Republike Srbije (broj i
naziv lokaliteta iz tabele odgovara broju i položaju lokaliteta na karti).
Sliv Crnog mora Sliv Egejskog mora Sliv Jadranskog mora

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 50/141
Materijal i metode
42
2.1.3. HEMIKALIJE
agaroza FMCamonijum acetat Sigma
borna kiselina SigmaEDTA Sigmaetidijum bromid Sigmaetanol 96% i 70% Merckfenol Flukaformamid Applied Biosystemshlorovodonična kiselina Merckizoamil-alkohol Merck - Alkaloidkalijum acetat Kemikakalijum hlorid Kemikasir ćetna kiselina Merck- Alkaloidmagnezijum sulfat Merckmineralno ulje Sigmanatrijum acetat Merck - Alkaloidnatrijum hlorid Merck
Na-dodecil sulfat (SDS) Mercknatrijum hidroksid Merckmagnezijum hlorid Merck - AlkaloidTris-baza GATCTSR Applied Biosystemsvoda za ćelijske kulture Sigma
2.1.3.1. Pripremljeni setovi hemikalija
DNA sequencing Kit, Big DyeTM Terminator Perkin ElmerCycle Sequencing v2.0 Ready Reaction
Wizard® Genomic DNA Purification Kit Perkin ElmerJet quik tissue DNA spin kit/250 Genomed
2.1.3.2. Enzimi
endonukleaze AluI Fermentasrekombinantna Taq DNA-polimeraza Fermentas
proteinaza-K Life TechnologiesRNA-za Fermentas

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 51/141
Materijal i metode
43
2.1.3.3. Početni oligonukleotidi (prajmeri)
Tabela 2. Početni oligonukleotidi koji su korišćeni pri amplifikaciji celog kontrolnog
regiona mtDNA (28Riba i cytR).
Oznaka Nukleotidni redosled Referenca28Riba CACCCTTAACTCCCAAAGCTAAG Snoj i sar ., 2000.cytR GTGTTATGCTTTAGTTAAGC Bernatchez i
Danzmann, 1993.
2.1.3.4. Markeri
100 bp Fermentas sadrži fragmente DNA koji su dugi
80, 100, 200, 300, 400, 500, 600,700, 800, 900 i 1031 bp1 kbp Fermentas sadrži fragmente DNA koji su dugi
250, 500, 700 bp, 1, 1.5, 2, 2.5, 3,3.5, 4, 5, 6, 8 i 10 kbp
2.1.3.5. Puferi
amplifikacijski pufer 0,25% bromfenol plavo0,25% ksilen cianol30% glicerol
50 x pufer TAE 2 M tris acetat0,1 M EDTA (pH 8,0)
10 x pufer TBE 0,5 M Tris baza0,5 M borna kiselina10 mM EDTA (pH 8,3)
pufer TEN 1 M Tris-HCl0,5 M EDTA2 M NaCl
2.1.4. LABORATORIJSKA OPREMA
ABI PRISMTM 310 Perkin Elmer
ABI PRISMTM 3100 Perkin Elmer
automatske pipete (10-1000 µl) Gilsoncentrifuga 5417C Eppendorfvakumski koncentrator EppendorfGel Doc 1000 BioRadinkubator Tehtnica, Eppendorfkadice za elektroforezu Pharmaciamešalica (vortex) Retschmagnetna mešalica Tehtnica
mikroprocesorski vodeni termostat MJ ResearchUV – transiluminator Camag

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 52/141
Materijal i metode
44
2.2. METODE
2.2.1. IZOLACIJA DNA
DNA je izolovana iz tkiva analnog peraja na tri različita načina. Bez obzira na koji
je način vršena izolacija, prvo je odsečeni deo tkiva analnog peraja (oko 2mm2), opran
destilovanom vodom od alkohola i nakon toga je prenet u posudicu (eppendorf) od 1,5
ml.
2.2.1.1. Fenolna ekstrakcija
Nakon pranja i prenošenja dela tkiva analnog peraja u posudicu od 1,5 ml, dodato je 200 µl TEN pufera, 7 µl 10% SDS i 5 µl proteinaze - K. Sadržaj posudice je promešan
obrtanjem, nakon toga posudica je stavljena na inkubiranje preko noći na 37oC.
Nakom inkubacije, dobijeni lizat je prečišćen fenolnom ekstrakcijom. Lizat
(sadržaj bez skeletnih elemenata peraja) prenet je u novu posudicu od 1,5 ml i dodato je
300 µl fenola, hloroforma i izoamil alkohola (25:24:1). Sadržaj posudice je promešan i
centrifugiran 5 min na 10000g.
Posle centrifugiranja, u posudici je uočena gornja i donju frakcija. Gornja frakcijau kojoj je rastopljena DNA odpipetirana je u novu posudicu, vodeći računa da ne dođe do
uzimanja dela belog taloga donje frakcije u kome se nalaze proteini.
U sledećem koraku dodato je 300 µl hloroforma i izoamil alkohola (24:1), sadržaj
je promešan i centrifugiran 5 min na 10000g.
Po centrifugiranju, gornja frakcija je odpipetirana u novu posudicu, u koju je
dodat 2,5 puta veći volumen 96% etanola ohlađenog na – 20oC, da bi došlo do obaranja
genomske DNA. Sadržaj posudice je promešan obrtanjem iste nekoliko puta, dok nisu primećene bele niti DNA.
Uzorak je stavljen da se inkubira 1h na – 20oC. Nakon ovog vremena izvršeno je
centrifugiranje uzorka 10 min na 10000 g.
Po centrifugiranju, iz posudice je odpipetiran supernatant i dobijeni talog DNA
osušen je na vazduhu.
DNA je rastopljena u 20 µl vode za ćelijske kulture (Sigma), i sačuvana ufrižideru na + 4oC.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 53/141
Materijal i metode
45
2.2.1.2. Wizard® Genomic DNA Purification Kit
Druga metoda izolacije DNA je izolacija uz pomoć specijalizovanog kita Wizard®
Genomic DNA Purification Kit. Izolacija je urađena prateći uputstva proizvođača,
koristeći protokol za izolaciju DNA iz mišjeg repa.
2.2.1.3. Jet quick tissue DNA spin kit/250
Treća metoda izolacije DNA je izolacija uz pomoć specijalizovanog kita firme
Genomed, Jet quik tissue DNA spin kit/250. Izolacija je urađena prateći uputstva
proizvođača, koristeći protokol za izolaciju DNA iz životinjskih tkiva.
2.2.2. PROVERA USPEŠNOSTI IZOLACIJE DNA
Provera uspešnosti izolacije DNA, kao i određivanje dužine PCR, RFLP
produkata, te izolacija fragmenata DNA za sekvencioniranje, može se obaviti
elektroforezom na agaroznom gelu, zahvaljujući tome što se molekuli DNA mogu kretati
u električnom polju. Brzina kretanja zavisi od veličine fragmenta DNA, gustine medijuma
i jačine struje.Provera izolacije DNA je izvršena na 1% agaroznom gelu. Agarozni gel je
napravljen od agaroze i 0,5 x TBE pufera. Smeša agaroze i pufera zagrejana je u
mikrotalasnoj pećnici do tačke ključanja, a nakon toga smeša je ohlađena u vodenom
kupatilu na elektromagnetnoj mešalici. Dok se smeša hladila, u nju je dodato 0,5 µl
etidijum bromida. Nakon hlađenja, gel je nasut u modele za elektroforezu i ostavljen oko
30 min da se stegne. Posle stezanja gela, modeli sa gelom preneti su u kadice za
elektroforezu u kojima se nalazio 0,5 x TBE pufer.
Pre nanošenja na gel, uzorku DNA (4 µl) dodat je amplifikacijski pufer (0,25%
brom – fenol plavo, 0,25% ksilen cianol i 30% glicerol). Smeša DNA i pufera
odpipetirana je u rupice na gelu.
U 0,5 x TBE puferu, pri naponu od 120 V, elektroforeza je trajala oko 30 min. Za
to vreme etidijum bromid se interkalirao između baza DNA i fluorescirao pri emitovanoj
svetlosti talasne dužine 302 nm, što je omogućilo vizuelizaciju gelova po elektroforezi
pomoću UV – transiluminatora.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 54/141
Materijal i metode
46
Slika 19. Provera izolacije DNA na agaroznom gelu
Kada je utvr đeno da je koncentracija izolovane DNA dobra, napravljena je radna
koncentracija tako što je osnovna razblažena 10 x dodavanjem Sigma vode. Dobijeno
razblaženje (∼10 - 50 ng/ µl) korišćeno je dalje za pripremu PCR reakcije.
2.2.3. LANČANA REAKCIJA POLIMERAZE (PCR)
Lančana reakcija sinteze DNA pomoću DNA polimeraze u in vitro uslovima je
metoda za amplifikaciju nukleinskih kiselina. PCR (eng. Polymerase Chain Reaction) je
otkrio Karl Mullis 1985 godine (Saiki i sar., 1988). Pomoću ove metode može se
umnožiti željeni segment DNA u velikom broju kopija.
Lančanom reakcijom polimeraze umnožen je kontrolni region (D-loop)
mtDNA dužine oko 1100 bp. Reakcija je tekla u tri koraka na različitim
temperaturama. U prvom koraku, pri visokoj temperaturi dolazi do denaturacije
dvolančanog molekula uzoračke DNA. Snižavanjem temperature u drugom koraku,
početni oligonukleotidi naležu uz komplementarna mesta na jednolančanom molekulu
uzoračke DNA. U trećem koraku uz prisustvo enzima polimeraze sintetizuje se
komplementarni lanac DNA između početnih oligonukleotida.
Tabela 3. Sastav reakcijske smese zapremine od 20 µl:
∑ 20 µl
10,8 µl vode (Sigma)2 µl 10 x pufera PCR (Fermentas)
1,2 µl 2 mM smeše dNTP (dATP, dCTP, dGTP, dTTP)
1 µl 25 mM MgCl2
1 µl 10 pmol/µl početnog oligonukleotida (28Riba)
1 µl 10 pmol/µl početnog oligonukleotida (cytR)
1 µl Taq DNA – polimeraze 5 U/µl
2 µl uzoračke DNA
DNA

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 55/141
Materijal i metode
47
Pripremljena PCR reakcija stavljena je u PCR mašinu MJ Research PTC - 100
(mikroprocesorsko vodeni termostat) na temperaturno - vremenski program 28RCYTR,
koji počinje denaturacijskim korakom (5 min na 95oC) i završava se korakom sinteze
komplementarnog lanca DNA (5 min na 72oC).
Tabela 4. Opis temperaturno vremenskog programa 28RCYTR
Nakon završetka PCR reakcije, izvršena je provera kvaliteta i dužine PCR
produkata na 1,5% agaroznom gelu u 0,5 x TBE puferu. Dužina fragmenata je utvr đena
koristeći markere od 100 bp ili 1kb. (Priprema gela je ista kao u poglavlju 2.2.2.).
Slika 20. Provera PCR produkta na agaroznom gelu
Ime programa Denaturacija Vezivanje početniholigonukleotida
Sinteza komplementarnoglanca
Broj ciklusa
28RCYTR 95oC, 45 s 52oC, 45 s 72oC, 45 s 32

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 56/141
Materijal i metode
48
2.2.4. R ESTRIKCIONA ANALIZA KONTROLNOG REGIONA MTDNA
(TEHNIKA RFLP – R ESTRICTION F RAGMENT L ENGTH P OLYMORPHISM )
U restrikcionoj analizi tretiran je PCR produkt endonukleazom Alu I , koja reže
kontrolni region mtDNA na prepoznatljivoj sekvenci ag/ct, na fragmente karakteristične
dužine za linije potočne pastrmke koji su očekivani na analiziranom područ ju.
U posudici od 0,5 ml pripremljena je restrikciona reakcija zapremine 20 µl koja
sadrži:
5 µl PCR produkt
2 µl odgovarajućeg restrikcionog pufera
0,5 µl (5 U) endonukleaze
12,5 µl vode (Sigma)
Napravljena reakcija je inkubirana 3h na temperaturi od 37oC. Po završenoj
inkubaciji naneti su reakcija (uzorci) i marker (100 bp) na 1,5% agarozni gel u 0,5 x
TBE puferu. Nakon toga otpočeta je elektroforeza 5 min na 80 V, a nakon toga još 10
min na 120 V. Rezultati restrikcije provereni su po elektroforezi pomoću UV –
transiluminatora.
2.2.5. SEKVENCIONIRANJE MITOHONDRIJALNE DNA
Sekvencioniranje je metoda kojom se utvr đuje redosled nuleotida na fragmentu
DNA određene dužine. Za sekvencioniranje i analizu fragmenata mtDNA
upotrebljavani su automatski sekvenatori ABI PRISMTM 310 (sa jednom kapilarom) i
ABI PRISMTM 3100 (sa 16 kapilara).
2.2.5.1. Izolacija fragmenata DNA iz agaroznog gela
Nakon elektroforeze PCR produkata na agaroznom gelu (Slika 20), odmah
ispod fragmenata DNA skalpelom su izrezane rupice pravougaonog oblika, koje su
napunjene 0.5 x TAE puferom. Nastavljena je elektroforeza ali na 200 V u trajanju
od 2 x 1 min. Za to vreme DNA fragmenti su eluirali u pufer, koji je odpipetiran u
posudice od 1,5 ml. Da bi DNA bila oborena, u posudice je dodat 6M NaCl i 96%
etanol ohlađen na -20oC (8 µl 6M NaCl / 50 µl 0,5 x TAE i 2,5 x volumen etanola).
Obaranje je trajalo preko noći na -20 oC ili 1h na -70 oC. Nakon obaranja
(inkubacije) uzorci su centrifugirani 25 min na 20000 g, alkohol je odpipetiran i

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 57/141
Materijal i metode
49
talog DNA osušen na vazduhu. Posle sušenja talog je rastopljen u 20 µl vode
(Sigma).
Svrha izolacije fragmenata DNA iz agaroznog gela je odstranjivanje ostataka
početnih nukleotida i dNTP-a koji se nisu ugradili u PCR produkt, kao i drugih
nečistoća.
Uspešnost izolacije fragmenata iz agaroznog gela može se proveriti
spektrofotometrijski ili na 1,5 % agaroznom gelu u 0,5 x TBE puferu. (Priprema gela je
ista kao u poglavlju 2.2.2.).
Slika 21. Provera izolacije DNA fragmenata iz agaroznog gela
2.2.5.2. Priprema reakcije za automatski sekvenator
Sekvencijska reakcija je imala volumen od 15µl. U specijalne posudice
prilagođene PCR mašini odpipetirano je od 3 – 6 µl prečišćenog PCR produkta iz
agaroznog gela, 4,5 µl razblaženog rastvora BigDye, 0,3 µl početnog oligonukleotida i do
15 µl dosuta je voda (Sigma).
Tabela 5. Nukleotidni redosled početnih nukleotida korišćenih za sekvencioniranje
segmenata kontrolnog regiona.
Oznaka Nukleotidni redosled Referenca28Riba CACCCTTAACTCCCAAAGCTAAG Snoj i sar ., 2000.vHF-F TCCGTCTTTACCCACCAACT Snoj i sar ., 2002.RCSB3-F AAGCCGGGCGTTCTCTTATATGC Snoj i sar ., 2002.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 58/141
Materijal i metode
50
Pripremljena sekvencijska reakcija stavljena je u PCR mašinu MJ Research PTC -
100 (mikroprocesorsko vodeni termostat) na temperaturno - vremenski program HEL2
koji počinje denaturacijskim korakom (5 min na 96oC).
Tabela 6. Opis temperaturno vremenskog programa HEL2
2.2.5.3. Priprema DNA fragmenata za sekvencioniranje
Produkt sekvencijske reakcije (15 µl) odpipetiran je u posudicu od 1,5 ml u kojoj
je predhodno pomešan 2 µl Na – acetata (3 M, pH 4,6) i 50 µl 96% etanola ohlađenog na-20oC. Rastvor je dobro promešan na Vortexu i inkubiran na ledu 10 min. Nakon toga
usledilo je centrifugiranje u trajanju od 25 min na 20800 g. Po centrifugiranju odpipetiran
je supernatant, a talog koji se nalazio uz zid posudice prosušen je na vazduhu. Potom je
talog rastvoren u 15 µl rastvora Template Suppression Reagent, dobro je sve promešano
na Vortexu i nekoliko sekundi centrifugirano. Rastvorena DNA je denaturisana 3 min na
95oC, posle toga je rastvor ohlađen na ledu i prenet u posebne posudice za
sekvencioniranje koje su stavljene u automatski sekvenator na utvr đivanje redosleda
nukleotida.
2.2.5.4. Sekvencioniranje DNA fragmenata
Sekvencijska reakcija u kojoj se nalazi DNA čiji se nukleotidni raspored želi
utvrditi na sekvenatoru, sadrži jedan početni oligonukleotid, smešu nukleotida koji sadrže
i dideoksiribonukleozid – trifosfate (ddNTP) sa fluorescirajućom molekularnom grupom.
Polimeraza takođe ulazi u sastav reakcije, i njena funkcija je da sintetizuje
komplementarni lanac uzoračkoj DNA vezujući iza početnog obične nukleotide sve dok
se ne ugradi prvi ddNTP. Jednolančani molekuli DNA koji sadže (ddNTP) na svome
kraju, putuju kroz električno polje i raspoređuju se po dužini. Svetlost određene talasne
dužine koju emituje laser pada na fluorescirajuće grupe molekula DNA i menja svoju
prvobitnu talasnu dužinu, što beleži detektor. Pošto postoje četiri različite fluorescirajuće
grupe za svaki od četiri molekula ddNTP, možemo na osnovu talasne dužinefluorescirajuće svetlosti utvrditi nukleotidni raspored uzoračke DNA.
Ime programa Denaturacija Vezivanje početniholigonukleotida
Sinteza komplementarnoglanca
Broj ciklusa
HEL2 95oC, 30 s 50oC, 15 s 60oC, 4 min 35

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 59/141
Materijal i metode
51
2.2.6. PROGRAMSKA ANALIZA KONTROLNOG REGIONA MTDNA
Za utvr đivanje grešaka i polimorfnih mesta na 5`- kraju kontrolnog regiona
mtDNA (561 bp) upotrebljen je program Chromas 2.23 ©Technelysium Pty. Ltd..Upoređivanje dobijenih sekvenci sa sekvencama koje postoje u bazi podataka “GenBank”
urađeno je u programu Clustal_X (Thompson i sar., 1997).
Filogenetska stabla nacrtana su koristeći dve vrste metoda za obradu
genetičkih podataka. “Neighbour-Joining” (NJ) spada u nediskretne metode (metode
distanci) koje za oblikovanje filogenetskog stabla koriste matrice parnih distanci.
Parne distance između pojedinih haplotipova izračunate su koristeći Kimura-2
parametarsku metodu. “Neighbour-Joining” (NJ) se drugačije naziva metoda
pridruživanja suseda, koja za formiranje filogenetskog stabla koristi uzastopno
pridruživanje operacionalno taksonomskih jedinica (suseda) u parove, tako da
smanjuje ukupnu dužinu grana za svaki nivo grupisanja. Dužina grananja se utvr đuje
po dodavanju pojedinih operacionalno taksonomskih jedinica. Metoda NJ korišćena
je iz genetičkog programskog paketa MEGA3 (Kumar i sar., 2004) sa potporom
formiranja čvorišta od 10000 bootstrap ponavljanja i programskog paketa PAUP
(Swofford, 2000) sa 1000 bootstrap ponavljanja.
“Maximum parsimony” (MP) – metoda najveće štedljivosti - spada u grupu
diskretnih metoda (“Character State” metode) koje formiraju filogenetsko stablo ne
uzimajući u obzir distance između polimorfnih mesta već ostale promene na
sekvencama tipa supstitucija, insercija i delecija. Kod ove metode primenjen je
heuristički princip sa TBR (Tree Branch Replications) zamenom grana kako bi se
pronašao put do najkraćeg stabla. Drugim rečima, “Maximum parsimony” je
karakter zasnovana metoda koja izvodi konačnu sliku filogenetskog stabla
minimizacijom ukupnog broja evolutivnih koraka potrebnih za objašnjavanje
dotičnog seta podataka. Ova metoda u prvoj fazi formira konsenzus stablo sa
čvorištima koja se javljaju u najmanje 50% slučajeva (Majority Rule 50% Consensus
Tree), a u drugoj fazi dodaje bootstrap vrednosti nakon 1000 ponavljanja. Metoda
MP korišćena je iz programskog paketa PAUP (Swofford, 2000).
Upotrebom statističkog “parsimony” kriterijuma u programu TCS 1.3 (Clement i
sar., 2000) slikovito su prikazani, u obliku mreže, osnovni filogenetski odnosi
analiziranih haplotipova.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 60/141
Materijal i metode
52
Genetička varijabilnost utvr đena je korišćenjem analize molekularne varijanse
(AMOVA testa) iz programskog paketa “ARLEQUIN” (Schneider i sar., 2000) na tri
hijerarhijska nivoa: između slivova, između populacija unutar slivova i unitar populacija.
Značajnost komponenti varijanse i Φ-statistike testirana je višestrukim permutacijama
(1000×) originalnog seta podataka.
Da bi se dobila šira filogenetska slika, haplotipovi pronađeni u Srbiji analizirani
su skupa sa nekoliko već pronađenih haplotipova sa drugih teritorija iz različitih mtDNA
linija.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 61/141
Rezultati
53
3. REZULTATI
3.1. ANALIZA KONTROLNOG REGIONA mtDNA UPOTREBOM RFLP
TEHNIKE (Restriction Fragment Length Polimorphism)
Upotrebom RFLP tehnike odnosno restrikcionog enzima Alu I dobijeni su
preliminarni rezultati zastupljenosti osnovnih mtDNA linija potočne pastrmke unutar sva
tri sliva na teritoriji Srbije. Restrikcioni enzim Alu I rezao je kontrolni region mtDNA
dužine oko 1100 bp na fragmente karakteristične dužine za dva različita restrikciona
profila.
Kontrolni region uzoraka iz crnomorskog sliva restrikcioni enzim Alu I je rezao na
četiri mesta formirajući pet fragmenata dužine 464, 311, 252, 37 i 4 bp (Slika 22a), dok je
uzorke iz egejskog i jadranskog sliva isti enzim rezao na tri mesta formirajući četiri
fragmenta dužine 563, 464, 37 i 4 bp (Slika 22b). Fragmenti od 4 i 37 bp vrlo su kratki i
ne mogu se uočiti na gelu.
Slika 22. Restrikcija kontrolnog regiona mtDNA sa endonukleazom Alu I . Uzorci sa
lokaliteta crnomorskog sliva označeni su sledećim brojevima: 1. Resava, 2.
Godljevača, 3. Buk, 4. Crni potok, 5. Brevina i 9. Gradac. Uzorak broj 6. Božica,
pripada egejskom slivu, a uzorci 7. Pećka Bistrica i 8. Prizrenska Bistrica pripadaju
jadranskom slivu. Poslednja kolona desno na obe slike predstavlja marker od 1 kbp.
a) b)

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 62/141
Rezultati
54
3.2. ANALIZA SEKVENCIONIRANJA KONTROLNOG REGIONAmtDNA
Analiza sekvencioniranja kontrolnog regiona mtDNA urađena je na ukupno 101
uzorku iz sva tri sliva, pri čemu je iz crnomorskg sliva sekvencioniran 61 uzorak, a izegejskog i jadranskog sliva po 20 uzoraka (Tabela 1). Sekvencioniranjem je utvrđen
nukleotidni redosled kontrolnog regiona mtDNA u dužini od 561 bp, počevši od 5’ kraja
do mesta “poli T” koje se nalazi oko polovine ukupne dužine kontrolnog regiona.
Na osnovu dobijenih rezultata sekvencioniranja identifikovano je ukupno 18
polimorfnih mesta (Tabela 10) na osnovu kojih je u sva tri sliva pronađeno 15 različitih
haplotipova koji pripadaju Da – dunavskoj (8 haplotipova), «južnoj»1 (6 haplotipova) i At
– atlantskoj filogenetskoj grupi (1 haplotip) (Tabela 9). Od 15 pronađenih haplotipova 6 je novih za nauku, i to 3 dunavska i 3 jadranska (dva iz egejskog i jedan iz jadranskog
sliva). Haplotip Ad+Prz koji je pronađen u Prizrenskoj Bistrici kao autohtoni, zajedno sa
naknadno analiziranim haplotipovima iz Rijeke Crnojevića Ad+RC i iz Neretve Ad+N
formira potpuno novu, do sada nepoznatu liniju, kojoj je dodeljena oznaka Ad+ i ime
farioides. Novo otkrivena linija je geografski ograničena na reke jadranskog sliva jugo-
zapadnog Balkana.
Pronađeni dunavski (Da) i «južni» jadransko – egejski (Ad i Ad+) haplotipovi
pokazuju izrazitu geografsku pravilnost u distribuciji. Da haplotipovi bili su striktno
ograničeni na dunavski sliv, dok su «južni» haplotipovi bili dominantni u jadransko –
egejskom slivu, sa izuzetkom pet jedinki koje su bile pronađene u dunavskom slivu (4
jedinke su pronađene u Jermi i 1 u Vrli, i sve pripadaju haplotipu ADcs1). Genetička
segmentacija primećena je i između «južnih» haplotipova. Prostorni raspored 5 od 6
pronađenih haplotipova bio je striktno povezan sa jadranskim ili egejskim slivom, samo
je haplotip Ad+Prz bio ravnomerno raspoređen u oba sliva (Tabela 9).
Radi kompletnije i preciznije slike o evolutivnoj istoriji 15 pronađenih
haplotipova potočne pastrmke na teritoriji Srbije, analizirano je još 8 haplotipova, pre
svega iz linija koje nisu autohtone za istraživano područje (At, Ma, Me – po dva haplotipa
za svaku liniju), kao i dva Ad+ haplotipa (Crna Gora i Bosna i Hercegovina) (Tabela 13).
1 «Južna» grupa obuhvata haplotipove 1Ad i novo opisane 2Ad+ linije (1, 2 pronađeni su na analiziranom području), kao i haplotpove koji su naknadno uključeni u analizu iz Ma i Me linije.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 63/141
Tabela 9. Prikaz pronađenih haplotipova po lokalitetima i njihova frekvenca. Haplotipovi koji su obeleženi zvezdicom (*) obeležen znakom + pripada novo opisanoj Ad+ ( farioides) liniji.
Dunavski haplotipovi »Južni« haplotipovi (AdLokaliteti N
Da1 Da2 Da23b Da*Vl Da*Dž Da22 Da-s6 Da*Vr ADcs1 ADcs11 Ad*Pe Ad*Ti A
Dunavski sliv Σ 611. Zmajevac 3 1 22. Gradac 3 1 1 3. Godljevača 2 2
4. Crni potok 2 25. Trudovačka reka 3 36. Bresnička reka 3 37. Brevina 3 38. Jošanica 3 39. Prolomska reka 3 310. Rečka 3 2 111. Vratna 2 2 212. Resava 2 1 113. Buk 2 214. Radovanjska reka 3 1 215. Golema reka 3 316. Studenačka reka 3 317. Dojkinačka reka 2 218. Rosomačka reka 3 2 119. Jerma 4 4
20. Vlasina 2 221. Džepska reka 1 122. Vrla 3 2 123. Jelašnička reka 3 3
% hapl. unutar sliva 59 8 8 3 2 2 5 3 8 Egejski sliv Σ 20
24. Dejanov potok 3 1 25. Lisina 2 1 26. Božica 2 27. Ljubata 2 28. Brankovačka reka 2 1 29. Tripušnica 5 30. Tisova reka 2 231. Čerenačka reka 2 2
% hapl. unutar sliva 15 20
Jadranski sliv Σ 2032. Prizrenska Bistrica 6 33. Pećka Bistrica 8 4 434. Bogska reka 3 335. Alagina reka 3 3
% hapl. unutar sliva 50 20 Ukupno 101 36 5 5 2 1 1 3 2 8 10 4 4
% hapl. u ukupnom uzorku 35 5 5 2 1 1 3 2 8 10 4 4

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 64/141
Rezultati
56
Iz prethodne tabele može se videti da je sa 35 lokaliteta iz sva tri sliva
identifikovano ukupno 15 haplotipova. Na 23 lokaliteta crnomorskog sliva pronađeno je
10 haplotipova od kojih je 8 autohtono za crnomorski sliv. Od tih 8, tri su nova za nauku:
Da*Vl (Vlasina), Da*Dž (Džepska) i Da*Vr (Vrla)), pet je bilo već poznato od ranije
Da1, Da2, Da-s6, Da23b i Da22, a dva pronađena haplotipa ADcs1 (jadranski) i At1
(atlantski) nisu autohtona za ovo slivno područje.
Slika 23. Brojčana i procentualna zastupljenost haplotipova crnomorskog sliva.
36, 59%
5, 8%
2, 3%
2, 3%
1, 2%
1, 2%
3, 5%
5, 8%
5, 8%
1, 2%
Da1
Da2
Da*Vr
Da*Vl
Da*Dž
Da22
Da-s6
Da23b
ADcs1
At1
Brojčano i procentualno najzastupljeniji haplotip crnomorskog sliva je Da1, koji
je identifikovan na 16 od 23 analizirana lokaliteta i predstavlja 59% zastupljenosti svih
pronađenih dunavskih haplotipova. Posle Da1 haplotipa, sa po 8% u ukupnoj
zastupljenosti učestvuju haplotipovi Da2 (pronađen na 4 lokaliteta), Da23b (pronađen na3 lokaliteta) i ADcs1 (pronađen na 2 lokaliteta). Sa 5% zastupljen je haplotip Da-s6
(pronađen na 1 lokalitetu), sa po 3% haplotipovi Da23b i Da*Vl (pronađeni na po 1
lokalitetu). Sa po 2% zastupljeni su haplotipovi Da*Dž, Da22 i At1 (svaki je pronađen na
posebnom lokalitetu).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 65/141
Rezultati
57
U egejskom slivu, na 8 analiziranih lokaliteta pronađeno je 5 haplotipova od kojih
su dva: Ad*Bož (sliv Božice) i Ad*Ti (Tisova i Čerenačka reka) nova za nauku, dok su
haplotipovi ADcs1, Ad+Prz, At1poznati od ranije.
Slika 24. Brojčana i procentualna zastupljenost haplotipova egejskog sliva.
3, 15%
5, 25%
4, 20%
7, 35%
1, 5%ADcs1
Ad+Prz
Ad*Ti
Ad*Bož
At1
Brojčano i procentualno najzastupljeniji haplotip egejskog sliva je Ad*Bož, koji je
idetifikovan na 4 od 5 analiziranih lokaliteta u slivu reke Dragovištice i predstavlja 35%
zastupljenosti svih haplotipova pronađenih na ovom slivnom području. U slivu iste reke
sa 15% zastupljen je haplotip ADcs1 (pronađen na 3 lokaliteta) i haplotip At1 sa 5%
(pronađen na 1 lokalitetu). U slivu reke Vardar koja pripada istom slivnom području
obrađene su leve i desne pritoke. U levoj pritoci pronađen je haplotip Ad+Prz (pronađen
na jednom lokalitetu - Pčinja), koji predstavlja 25% ukupne zastupljenosti haplotipova
ovog sliva, a od desnih pritoka obrađena su dva lokaliteta (Tisova i Čerenačka reka) na
kojima je pronađen haplotip Ad*Ti koji učestvuje sa 20% u ukupnoj zastupljenosti.
U jadranskom slivu na 4 analizirana lokaliteta pronađena su 3 haplotipa, od kojih
je jedan novi za nauku Ad*Pe (Pećka Bistrica), a dva su već poznata od ranije Ad+Prz i
ADcs11.
Slika 25. Brojčana i procentualna zastupljenost haplotipova jadranskog sliva.
10, 50%
6, 30%
4, 20%
ADcs11
Ad+Prz
Ad*Pe
Brojčano i procentualno najzastupljeniji haplotip jadranskog sliva je ADcs11, koji
je identifikovan na sva 3 obrađena lokaliteta u slivu Pećke Bistrice i zastupljen je sa 50%.
Novi haplotip Ad*Pe koji je pronađen samo u Pećkoj Bistrici zastupljen je sa 20%, a

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 66/141
Rezultati
58
haplotip Ad+Prz koji je pronađen u Prizrenskoj Bistrici sa 30% u ukupnom broju svih
haplotipova na slivnom području.
Slika 26. Brojčana i procentualna zastupljenost svih pronađenih haplotipova na
analiziranom području.
36, 35%
5, 5%
5, 5%
2, 2%
1, 1%
1, 1%
3, 3%
2, 2%
8, 8%
10, 10%
11, 11%
4, 4%
7, 7%
4, 4%
2, 2%
Da1
Da2
Da23b
Da*Vl
Da*Dž
Da22
Da-s6
Da*Vr
ADcs1
ADcs11
Ad+Prz
Ad*Ti
Ad*Bož
Ad*Pe
At1
Procentualne vrednosti zastupljenosti haplotipova u ukupnom uzorku uglavnom seslažu sa brojčanim vrednostima zbog broja analiziranih jedinki (101 jedinka). Po
brojnosti, u ukupnom uzorku dominira haplotip Da1 (36 jedinki), a prate ga Ad+Prz (11
jedinki) i ADcs11 (10 jedinki), ADcs1 (8 jedinki), Ad*Bož (7 jedinki), dok su ostali
haplotipovi zastupljeni sa pet i manje jedinki.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 67/141
Slika 27. Sekvenca 5’ kraja kontrolnog regiona mtDNA (561bp), najbrojnijeg haplotipa Da1 na analiziranom
sekvence pokazuju varijabilna nukleotidna mesta između analiziranih haplotipova (pronađenih i naknadno
(Bernatchez i sar., 1992).
2 26 61
* * *
ACTTTTCAGC TATGTACAAT AACAAATGTT GTACCTTGCT AACCCAATGT TATACTACAT CTATGT
113 126 14
* * *
TATGTATTTA CCCATATATA TAATATAGCA TG-TGAGTAG TACATCATAT GTATTATCAA CATTAG
178 196
* *
CATACATCAG CACTAACTCA AGGTTTACAT AAAGCAAAAC ACGTGATAAT AACCAACTAA GTTGTC
243 262 263
* **
TGTTATATCA ATAAAACTCC AGCTAACACG GGCTCCGTCT TTACCCACCA ACTTTCAGCA TCAGTC
387
AAGAACCGAC CAACGATATA TCAGTAGGCA TACTCTTATT GATGGTCAGG GACAGATATC GTATTA
403
*
AATTATTCCT GGCATTTGGT TCCTATATCA AGGGCTATCC TTAAGAAACC ACCCCCTGAA AGCCGA
484 530 542 543
* * **
GTTAATGGTG TCAATCTTAT TGCCCGTTAC CCACCAAGCC GGGCGTTCTC TTATATGCAT AACGTT
TT

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 68/141
Rezultati
60
Tabela 10. Polimorfna mesta na 5’ kraju kontrolnog regiona mtDNA za 15 haplotipova
pronađenih na teritoriji Srbije. Polimorfna mesta analiziranih haplotipova predstavljena su
u odnosu na haplotip At1, gde crtica ( - ) predstavlja ponovljeni nukleotid iz pvog reda, a
kosa crta ( / ) deleciju.
Kod mutacija haplotipova pronađenih u Srbiji tranzicije su zabeležene u 12
slučajeva (polimorfna mesta obeležena crnom bojom), transverzije u 4 (polimorfna mesta
obeležena crvenom bojom), a na 2 polimorfna mesta (obeležena plavom bojom), zbogspecifičnih mutacija na obeleženim pozicijama, a karakterističnih za pojedine haplotipove,
može se reći da postoji «kombinacija tranzicija i transverzija» za dato mesto. Haplotip Da-
s6 ima tranziciju na poziciji broj 26, u odnosu na sve ostale dunavske haplotipove (A→G).
Na istoj poziciji postoji transverzija između dunavskih i ostalih haplotipova (At i «južnih»).
Na poziciji broj 235 postoji očigledna tranzicija između prva tri dunavska (Da2, Da23b i
Da*Vl) i svih ostalih haplotipova (G→A), osim u slučaju haplotipa Da22 koji ima
transverziju na toj poziciji u odnosu na sve druge haplotipove (Tabela 10).
U Da liniji pronađena su tri nova haplotipa (Da*Vl, Da*Dž and Da*Vr), koji se
karakterišu mutacionim događajima na sledećim pozicijama: Da*Vl 390 (T), haplotip
Da*Dž 61 (T), 530 (T), 542 (G), i 548 (C), haplotip Da*Vr 228 (G), i 543 (G), dok su
polimorfizmi na pozicijama 530, 542, i 548 isti kao i kod haplotipa Da*Dž.
U Ad liniji prvi put su opisana tri haplotipa Ad*Bož, Ad*Pe i Ad*Ti koja se
karakterišu mutacionim događajima na sledećim pozicijama: Ad*Bož 26 (T), Ad*Pe 387
(A) i Ad*Ti 543 (C) (Tabela 10).
Polimorfna mestaHaplotipovi 2 26 61 80 126 146 228 235 236 262 387 389 390 403 530 542 543 548
At1 T T C T C G T A T G G C T T T G G CDa2 C A - - - - - G G - - T C - C A C TDa23b C A - - - A - G G - - T C - C A C TDa*Vl C A - - - - - G G - - T - - C A C TDa22 C A - - - - - T G - - T C - C A C TDa1 C A - - - - - - G - - T C - C A C TDa-s6 C G - - - - - - G - - T C - C A C TDa*Dž C A T - - - - - G - - T C - - - C -
Da*Vr C A - - - - G - G - - T C - - - - -ADcs1 - C - - - - - - - C - T C C - - - -ADcs11 - C - C - - - - - C - T C C - - - -Ad*Pe - C - - - - - - - C A T C C - - - -Ad*Ti - C - - - - - - - C - T C C - - C -Ad*Bož - - - - - - - - - C - T C C - - - -Ad+Prz - C - - T - - - - T - T C C - - - -

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 69/141
Rezultati
61
Tabela 11. Procentualne vrednosti divergencija između parova haplotipova (ispoddijagonale), i broj polimorfnih mesta između njih na analiziranom fragmentu kontrolnogregiona mtDNA(iznad dijagonale).
Srodnički odnosi između pojedinih parova haplotipova određivani su
upoređivanjem haplotipova putem dvoparametarskog Kimura modela sa odnosom
tranzicije/transverzije = 2. Procentualne vrednosti distanci za haplotipove pronađene u
Srbiji kreću se od 0,18% do 2,12% ( X= 1.2 SD = ±0.68 ). Što se tiče razlika u
polimorfnim mestima, haplotipovi se među sobom razlikuju za od 1 do 12 supstitucija.
Genetička divergencija unutar Da linije je znatno izraženija i za oko tri puta veća (0,6%
od unutar grupne srednje genetičke distance) u poređenju sa divergencijom unutar
«južne» grupe linija (Ad i Ad+).
Tabela 12. Rezultati AMOVA testa.
Vrednosti Φ statistike AMOVA testa pokazuju da je genetička divergencija
povezana sa geografskim rasporedom odnosno hidrološkim paternom (68.80% od ukupne
varijabilnosti između slivova, Φ CT = 0.68798, p < 0.001). Rezultati ukazuju da je visoknivo populacione strukturiranosti (Φ ST = 0. 91565, p < 0.001) u ukupnom uzorku (sve
Haplotipovi
D a
2
D a
2 3 b
D a
* V l
D a
2 2
D a
1
D a
- s 6
D a
* D ž
D a
* V r
A t 1
A D
c s 1
A D
c s 1 1
A d
* P e
A d
* T i
A d
* B o ž
A d
+ P r z
Da2 1 1 1 1 2 5 6 10 10 11 11 9 10 11Da23b 0,18 2 2 2 3 6 7 11 11 12 12 10 11 12Da*Vl 0,18 0,35 2 2 3 6 7 9 11 12 12 10 11 12Da22 0,18 0,35 0,35 1 2 5 6 10 10 11 11 9 10 11Da1 0,18 0,35 0,35 0,18 1 4 5 9 9 10 10 8 9 10
Da-s6 0,35 0,53 0,53 0,35 0,18 5 6 9 9 10 10 8 9 10Da*Dž 0,88 1,05 1,05 0,88 0,70 0,88 3 7 7 8 8 6 7 8Da*Vr 1,05 1,23 1,23 1,05 0,88 1,05 0,53 6 6 7 7 7 6 7
At1 1,76 1,94 1,59 1,76 1,59 1,59 1,23 1,05 5 6 6 6 4 6ADcs1 1,76 1,94 1,94 1,76 1,59 1,59 1,23 1,05 0,88 1 1 1 1 2
ADcs11 1,94 2,12 2,12 1,94 1,76 1,76 1,41 1,23 1,05 0,18 2 2 2 3Ad*Pe 1,94 2,12 2,12 1,94 1,76 1,76 1,41 1,23 1,05 0,18 0,35 2 2 3Ad*Ti 1,59 1,76 1,76 1,59 1,41 1,41 1,05 1,23 1,05 0,18 0,35 0,35 2 3
Ad*Bož 1,76 1,94 1,94 1,76 1,59 1,59 1,23 1,05 0,70 0,18 0,35 0,35 0,35 3Ad+Prz 1,94 2,12 2,12 1,94 1,76 1,76 1,41 1,23 1,05 0,35 0,53 0,53 0,53 0,53
Izvori
varijabilnosti
dfSume
kvadrata
Komponente
varijanse
Procenat
varijabilnosti
φφφφ - statistika
Između slivova 2 165.779 2.80100 68.80 0.72965 (φ sc)Između populacijaunutar slivova
32 93.851 0.92690 22.77 0.91565 (φ st)
Unutar populacija 66 22.667 0.34343 8.43 0.68798 (φ ct)Ukupno 100 282.297 4.07133 100

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 70/141
Rezultati
62
lokalne populacije) posledica velike različitosti haplotipova između populacija unutar
različitih slivova. Svaka lokalna populacija predstavljena je najčešće sa jednim ili retko sa
dva ili tri haplotipa, što je rezultiralo visokim vrednostima za oba Φ ST i Φ SC statistička
parametra (Φ SC = 0.72965, p < 0.001).
Utvrđivanje stepena srodnosti između pronađenih haplotipova na teritoriji Srbije
predstavljeno je filogenetskim stablom koje je urađeno po metodi "Neighbour-Joining"
(NJ), koja spada u metode distanci. Filogenetsko stablo napravljeno je i po metodi
"Minimum evolution" (ME), ali se ono nije bitno razlikovalo od stabla napravljenog
predhodnom (NJ) metodom.
Slika 28. Filogenetsko stablo haplotipova pronađenih na teritoriji Srbije, urađeno na
osnovu sekvenci 5' kraja kontrolnog regiona mtDNA metodom "Neighbour-Joining" (NJ)
u programu "MEGA 3". Brojevi uz čvorišta predstavljaju procentualne vrednosti
ponavljanja grananja u 10000 ponavljanja. Kao "outgroup" uključene su dve vrste Salmo
ohridanus i Salmo salar .
Na osnovu dobijenog stabla evidentno je da su dunavski i «južni» (Ad i Ad+)
egejski i jadranski haplotipovi monofiletske grupe, a da se između njih nalazi At1
haplotip koji pripada atlantskoj liniji.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 71/141
Rezultati
63
3.2.1. UPOREĐIVANJE SEKVENCI KONTROLNOG REGIONA mtDNA PRONAĐENIH
HAPLOTIPOVA NA TERITORIJI SRBIJE SA VEĆ POZNATIM
HAPLOTIPOVIMA POTOČNE PASTRMKE IZ SVIH FILOGENETSKIH LINIJA
Da bi se dobila kompletnija slika filogenetskih odnosa, u dalju analizu, osimhaplotipova pronađenih u Srbiji, uključeno je još osam haplotipova iz četiri filogenetske
linije: atlantske, mediteranske, marmoratus i farioides (Ad+). Iz dunavske linije nisu
uključeni dodatni haplotipovi jer je pronađen veliki broj različitih haplotipova na teritoriji
Srbije.
Iz atlantske linije analizirana su još dva haplotipa iz Austrije čije su sekvence za
kontrolni region mtDNA preuzete iz Banke gena: At-11a (AY185 578) i At-10 (AY185
577). Iz farioides linije analizirana su još dva haplotipa čije sekvence nisu do sadaobjavljene u Banci gena (Ad+N – Neretva i Ad+RC-Rijeka Crnojevića; usmeno
saopštenje Aleš Snoj). Iz mediteranske linije analizirana su dva haplotipa čije su sekvence
zavedene u Banci gena: MEcs1 (AY 836350) i MEcs7 (AY 836356) (Španija). Iz
marmoratus linije analizirana su dva haplotipa iz Slovenije: MAcs1 (AY 836365) i Ma-s2
čija cela sekvenca još nije objavljena (usmeno saopštenje Aleš Snoj).
Kao "outgroup" analizirane su dve vrste čije su sekvence zavedene u Banci gena
Salmo ohridanus (AY260 512) i Salmo salar (U12143).
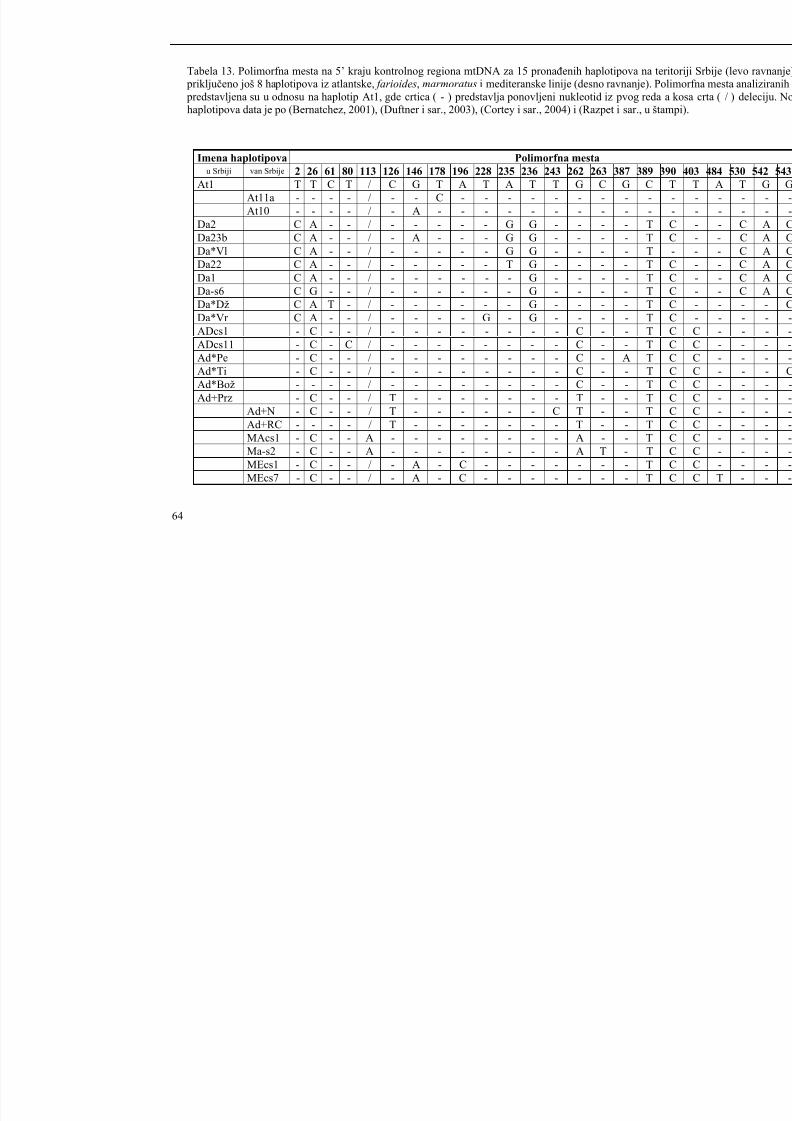
7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 72/141
64
Imena haplotipova Polimorfna mestau Srbiji van Srbije 2 26 61 80 113 126 146 178 196 228 235 236 243 262 263 387 389 39
At1 T T C T / C G T A T A T T G C G C TAt11a - - - - / - - C - - - - - - - - - -At10 - - - - / - A - - - - - - - - - - -
Da2 C A - - / - - - - - G G - - - - T CDa23b C A - - / - A - - - G G - - - - T CDa*Vl C A - - / - - - - - G G - - - - T -Da22 C A - - / - - - - - T G - - - - T C
Da1 C A - - / - - - - - - G - - - - T CDa-s6 C G - - / - - - - - - G - - - - T CDa*Dž C A T - / - - - - - - G - - - - T CDa*Vr C A - - / - - - - G - G - - - - T CADcs1 - C - - / - - - - - - - - C - - T CADcs11 - C - C / - - - - - - - - C - - T CAd*Pe - C - - / - - - - - - - - C - A T CAd*Ti - C - - / - - - - - - - - C - - T CAd*Bož - - - - / - - - - - - - - C - - T CAd+Prz - C - - / T - - - - - - - T - - T C
Ad+N - C - - / T - - - - - - C T - - T CAd+RC - - - - / T - - - - - - - T - - T CMAcs1 - C - - A - - - - - - - - A - - T CMa-s2 - C - - A - - - - - - - - A T - T CMEcs1 - C - - / - A - C - - - - - - - T CMEcs7 - C - - / - A - C - - - - - - - T C
Tabela 13. Polimorfna mesta na 5’ kraju kontrolnog regiona mtDNA za 15 pronađenih haplotipova na terit priključeno još 8 haplotipova iz atlantske, farioides, marmoratus i mediteranske linije (desno ravnanje). Polim predstavljena su u odnosu na haplotip At1, gde crtica ( - ) predstavlja ponovljeni nukleotid iz pvog reda a khaplotipova data je po (Bernatchez, 2001), (Duftner i sar., 2003), (Cortey i sar., 2004) i (Razpet i sar., u štam

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 73/141
Rezultati
65
U prethodnoj tabeli upoređena su polimorfna mesta na 5’ kraju kontrolnog regiona
mtDNA haplotipova pronađenih u Srbiji sa još osam haplotipova koji pripadaju
atlantskoj, farioides, mediteranskoj i marmoratus liniji, a nisu pronađeni na analiziranom
području. Iz tabele se može videti da haplotipovi svake linije poseduju nukleotidne
sinapomorfizme na određenim pozicijama.
Analizirani haplotipovi atlantske linije poseduju dva sinapomorfizma na
pozicijama 389 (C) i 390 (T).
Haplotipovi dunavske linije poseduju veći broj sinapomorfizama i to na
pozicijama: 2 (C), 26 (A) sa izuzetkom Da-s6 koji poseduje tranziciju (A→G), pozicija
236 (G), pozicija 530 (C) sa izuzecima Da*Dž i Da*Vr koji poseduju tranziciju (C→T),
pozicija 542 (A) sa izuzecima Da*Dž i Da*Vr koji poseduju tranziciju (A→G), pozicija
543 (C) sa izuzetkom Da*Vr koji poseduje transverziju (C→G) i pozicija 548 (T) sa
izuzecima Da*Dž i Da*Vr koji poseduju tranziciju (T→C). Poslednja četiri polimorfizma
kod haplotipa Da*Dž i Da*Vr zajednička su za najveći broj haplotipova koji pripadaju
drugim linijama (At, Ad, Ad+ Ma i Me) (Tabela 13).
Haplotipovi Ad i Ad+ linije poseduju takve sinapomorfizme da se mogu jasno
razlikovati. Zajednički sinapomorfizam za najveći broj haplotipova obe grupe je prisustvo
(C) na poziciji broj 26, a to su svi prikazani haplotipovi izuzev Ad*Bož i Ad+RC koji na
istoj poziciji imaju tranziciju (C→T). Sinapomorfizam bitan za prvu grupu (Ad) je na
poziciji 262 (C). U drugu grupu (Ad+) spadaju haplotipovi koji na poziciji 126 i 262
imaju (T), a to su Ad+Priz, Ad+N i Ad+RC.
Haplotipovi linije marmoratus (Ma) poseduju sinapomorfizam koji se ogleda u
prisustvu insercije na poziciji 113 (A) i prisustvu (A) na poziciji 262.
Haplotipovi mediteranske (Me) linije poseduju sinapomorfizme na pozicijama 146
(A) i 196 (C).
Haplotipovi iz Ad+ grupe, i iz Ma i Me linije pokazuju po dva interkladnasinapomorfizma po kojima se razlikuju, što ukazuje na sličnu dubinu divergencije ovih
klada (Tabela 13).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 74/141
Rezultati
66
Slika 29. Filogenetsko stablo haplotipova pronađenih na teritoriji Srbije uz dodatak 8
haplotipova iz atlantske, farioides, marmoratus i mediteranske linije, sa dva "outgroup"
haplotipa Salmo ohridanus i Salmo salar , urađeno na osnovu sekvenci 5' kraja kontrolnog
regiona mtDNA metodom "Neighbour-Joining" (NJ) u programu "MEGA 3". Brojevi uz
čvorišta predstavljaju procentualne vrednosti ponavljanja grananja u 10000 ponavljanja.
Na osnovu dobijenog stabla evidentno je da su haplotipovi grupisani u već
poznate linije (dunavsku, atlantsku, mediteransku, marmoratus i jadransku) i novu Ad+
( farioides) liniju, pri čemu su haplotipovi Da i At linije posebno grupisani u odnosu na
ostale «južno» – evropske haplotipove (Me, Ma, Ad i Ad+). U okviru Ad linije grupisani
su sledeći haplotipovi: Ad*Bož, Ad*Pe, Ad*Ti, ADcs1 i Adcs11, dok Ad+ grupu čine tri
haplotipa: Ad+Prz, Ad+N i Ad+RC.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 75/141
Rezultati
67
Slika 30. Filogenetsko stablo haplotipova pronađenih na teritoriji Srbije uz dodatak 8
haplotipova iz atlantske, farioides, marmoratus i mediteranske linije, sa dva "outgroup"
haplotipa Salmo ohridanus i Salmo salar . Stablo je urađeno u programu "PAUP 4" na
osnovu sekvenci 5' kraja kontrolnog regiona mtDNA metodom "Maximum parsimony
(MP) Majority role consensus 50%". Brojevi uz čvorišta predstavljaju procentualne
vrednosti ("bootstrap") ponavljanja grananja u 1000 ponavljanja. Vrednosti iznad grana
date su za "MP Majority role consensus 50%", prva vrednost ispod grana predstavlja MP
"bootstrap" i druga vrednost ispod grana predstavlja NJ "bootstrap".

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 76/141
Rezultati
68
Stablo koje je dobijeno metodom "MP Majority rule consensus 50%", vrlo je
slično stablima koja su dobijena korišćenjem metoda MP i NJ u programu "PAUP 4"‚
zato je upotrebljeno jedno stablo da se prikažu rezultati za sve tri metode. Dobijeno
stablo, prema položaju haplotipova i rasporedu u filogenetske linije, veoma je slično
stablu dobijenom u programu "MEGA 3" metodom "Neighbour-Joining" (NJ) (slika 29).
Konsensus stablo takođe prikazuje jasno razdvajanje dve glavne klade (Da i At klada plus
«južno» – evropska klada). Uključivanjem dva "outgroup" haplotipa Salmo ohridanus i
Salmo salar omogućeno je utvrđivanje statističke značajnosti prvog grananja koje se
pokazalo kao vrlo značajno. Unutrašnja topologija u okviru dve glavne grupe (Da i At
plus «južno» – evropska) bila je podržana nešto nižim "bootstrap" vrednostima, što
posebno važi za haplotip Ad*Bož, koji na ovom stablu zauzima ancestralni položaj u
odnosu na ostale «južno» – evropske haplotipove, ali bez podrške za MP analizu i sa
niskom "bootstrap"vrednošću za NJ analizu. Grupisanje u okviru «južno» – evropske
grupe haplotipova u Ad, Ma i Me i dopunsku Ad+ grupu (Ad+Prz, Ad+N i Ad+RC) isto
je kao i pri NJ analizi u programu "MEGA 3" (Slika, 29). Haplotipovi Da*Dž i Da*Vr
zauzimaju na svim filogenetskim stablima bazalni položaj unutar Da linije, što je i
statistički podržano.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 77/141
Rezultati
69
Slika 31. Kladogram (mreža) haplotipova pronađenih na
lokalitetima sva tri sliva na teritoriji Srbije, uz dodatak
haplotipova koji su naknadno uključeni u analizu (crveni
krugovi iste veličine). Veličina krugova haplotipova
pronađenih na teritoriji Srbije odgovara njihovoj
frekvenci, a dvobojnost označava da je haplotip pronađen
na dva slivna područja. Haplotipovi pronađeni u
dunavskom slivu označeni su različitim nijansama zelene
boje, u egejskom nijansama žute i u jadranskom različitim
nijansama plave boje. Svaka linija koja učestvuje u
povezivanju haplotipova predstavlja jedan mutacionikorak po algoritmu programa «TCS», a crni krugovi
predstavljaju teoretski nedostajuće haplotipove.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 78/141
Rezultati
70
Na osnovu kladograma (mreže) haplotipova (Slika 31) uočava se grupisanje
haplotipova u odgovarajuće linije. Preko broja algoritamskih mutacionih koraka (spojnih
linija) moguće je utvrditi tačan broj polimorfizama (mutacija) između haplotipova koji
pripadaju istoj liniji.
Na osnovu kladograma mogu se utvrditi genealoški odnosi između haplotipova.
Posmatranjem kladograma u celini, centralnu poziciju zauzima haplotip Da*Vr koji je
približno podjednako udaljen od Da (≥ 5 mutacionih koraka), At (≥ 5 mutacionih koraka)i
«južne» klade (≥ 6 mutacionih koraka). Unutar Da klade centralnu poziciju zauzima
haplotip Da1 koji se od hapotipova Da-s6, Da22 i Da2 razlikuje za jedan mutacioni korak,
dok se od haplotipova Da23b i Da*Vl razlikuje za dva mutaciona koraka. Najudaljeniji
haplotipovi u okviru Da linije su Da*Dž i Da*Vr koji se razlikuju od centralnog haplotipa
za 4 odnosno 5 mutacionih koraka. Unutar linija koje pripadaju južnoj kladi (Ad, Ad+,
Ma i Me) haplotip ADcs1 zauzima centralno mesto. Ostali Ad haplotipovi razlikuju se od
centralnog ADcs1 haplotipa za jedan mutacioni korak, dok se haplotipovi iz Ma, Me i
Ad+Prz linija razlikuju od centralnog za ≥2 mutaciona koraka.
Na osnovu rezultata dobijenih analizama isključivo na haplotipovima koji su
pronađeni na teritoriji Srbije (Slike 22b i 28) nije se moglo uočiti jasno razdvajanje unutar
jadranske linije. Međutim, dopunskim uključivanjem u filogenetsku analizu haplotipova
Ad+N i Ad+RC, koji su sa haplotipom Ad+Prz formirali na filogenetskim stablima novu
grupu, statistički visoko značajnu (Slika 29 i 30), kao i posebnu grupu na kladogramu
haplotipova (Slika 31), omogućen je uvid u razdvajanje prvobitne Ad linije i novo
otkrivene Ad+ ( farioides) linije.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 79/141
Diskusija
71
4.
DISKUSIJA
Bernatchez i sar. (1992) smatraju da, bez obzira na izuzetno veliki broj rezultata
koji su dobijeni u poslednjih 20 godina, za rasvetljavanje filogeografske strukture populacija potočne pastrmke još uvek nedostaju rezultati sa vrlo značajnih geografskih
područ ja kao što su severna Afrika i istočna Evropa. Kottelat (1997) naglašava da je
veliki problem u rešavanju taksonomije pastrmskih populacija Balkanskog poluostrva
upravo nedostatak informacija sa područ ja republika bivše Jugoslavije. Poseban značaj u
izučavanju balkanskih pastrmskih populacija je upravo u tome što se smatra da je
Balkansko poluostrvo, pored Apeninskog i Iberijskog, bilo refugijalni centar, a samim tim
i centar diverziteta za vreme glacijacija (Hewitt, 1996; 1999).U najbližoj okolini Srbije samo na teritoriji Austrije, Slovenije i Gr čke utvr đena je
genetička diferenciranost pastrmskh populacija (Duftner i sar., 2003; Snoj i sar., 2004;
Apostolidis i sar., 1996; 1996a; 1997). Poslednjih godina sakupljan je materijal iz Bosne i
Hercegovine, Hrvatske i Crne Gore, ali rezultati još nisu objavljeni.
Ako se uporede rezultati istraživanja sa pomenutih teritorija, kao i rezultati koje je
objavio Bernatchez (1992), sa rezultatima koji su dobijeni na teritoriji Srbije (101
jedinka/15 haplotipova), uočava se vrlo sličan diverzitet haplotipova. Bernatchez i sar.
(1992) pronašli su, analizom 310 bp kontrolnog regiona mtDNA, 12 haplotipova na 154
analizirane jedinke iz 24 populacije zapadne Evrope. Duftner i sar. (2003) su na teritoriji
Austrije opisali 16 haplotipova iz uzorka od 123 jedinke sa 20 lokaliteta (crnomorski
sliv), analizom kompletnog kontrolnog regiona mtDNA, gde 7 haplotipova pripada
atlantskoj liniji, čiju autohtonost na analiziranom područ ju tek treba utvrditi. Apostolidis i
sar. (1997) su za teritoriju Gr čke opisali 10 haplotipova iz uzorka od 76 jedinki sa 12
lokaliteta (egejski, jonski i jadranski sliv), analizom kontrolnog regiona mtDNA u dužini
od 310 bp. Snoj i sar. (2004) su na teritoriji Slovenije opisali 12 haplotipova iz uzorka od
364 jedinke sa 20 lokaliteta (crnomorski i jadranski sliv), analizom kontrolnog regiona
mtDNA u dužini od 420 bp.
Analizom celog kontrolnog regiona mtDNA i obradom većeg broja uzoraka sa
lokaliteta na teritoriji Kosova i Metohije (razvođe slivova), verovatno bi genetički
diverzitet pastrmskih populacija na teritoriji Srbije bio veći od prikazanog.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 80/141
Diskusija
72
4.1. ANALIZA KONTROLNOG REGIONA mtDNA
Na osnovu rezultata RFLP tehnike, korišćenjem restrikcionog enzima Alu I
dobijena je prva informacija o zastupljenosti mtDNA linija potočne pastrmke na
analiziranom područ ju. Upotrebom enzima Alu I dobijene su dve različite dužine
fragmenata, koje su odgovarale haplotipovima Da i Ad linije. Da haplotipovi pronađeni
su na analiziranim lokalitetima crnomorskog sliva, dok su Ad haplotipovi pronađeni na
lokalitetima jadranskog i egejskog sliva. Ranija istraživanja su pokazala da su u egejskom
slivu najzastupljeniji haplotipovi Ad linije (Apostolidis i sar., 1996a; 1997). Enzim Alu I
ne može razdvojiti haplotipove Da i At, kao ni Ad i novo opisane Ad+ ( farioides) linije,
jer imaju potpuno ista restrikciona mesta.
Sekvenciranjem dela kontrolnog regiona mtDNA dobijen je precizan pregled (koji
restrikciona analiza nije mogla pružiti) pronađenih haplotipova na analiziranom područ ju,
koji je omogućio detekciju genetičkog diverziteta potočne pastrmke u okviru tri već
poznate mtDNA linije (Da, Ad i At) kao i četvrte nove Ad+, koje naseljavaju gornje
tokove reka određenih slivova na teritoriji Srbije. Haplotipovi Da, i «južnih» linija su
generalno autohtoni za lokalitete crnomorskog odnosno jadranskog i egejskog sliva, dok
je At haplotip alohtonog porekla za analizirano područ je.
Ako se uporede rezultati istraživanja iz crnomorskog sliva na teritoriji Srbije sa
dobijenim rezultatima iz uzvodnog dela istog sliva (Slovenija i Austrija) uočava se da je
diverzitet haplotipova veoma sličan. U crnomorskom slivu Srbije pronađeno je 8
haplotipova naspram 7 Da haplotipova iz Slovenije (Snoj i sar., 2004) i 8 Da haplotipova
iz Austrije (od 5' kraja do poli T bloka kontrolnog regiona mtDNA) (Duftner i sar., 2003).
Najučestaliji haplotip u okviru sva tri analizirana područ ja je Da1. Od zajedničkih
haplotipova koji se sreću u Srbiji i u uzvodnim delovima crnomorskog sliva, osim
haplotipa Da1 pronađeni su Da2, Da22, Da23b haplotipovi sa vrlo sličnim učestalostima
u okviru analiziranih područ ja. Haplotipovi Da22 i Da23b su vrlo retki i svaki je, osim u
Srbiji, pronađen na malom broju lokaliteta (Da22 u Sloveniji i u Austriji (na po 2
lokaliteta), Da23b u Austriji (1 lokalitet/1 primerak)). Haplotip Da22 je jedini u Da liniji
koji na poziciji 235 poseduje transverziju G→T, dok haplotip Da23b poseduje tranziciju
G→A na poziciji 146, što je karakteristika najvećeg broja mediteranskih i vrlo malog
broja atlantskih haplotipova. Da23b haplotip pronađen je na tri lokaliteta u Istočnoj Srbiji

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 81/141
Diskusija
73
(Rečka, Vratna i Radovanjska reka - pritoka Timoka), a to su reke koje imaju neposrednu
komunikaciju sa Dunavom. Isti haplotip pronađen je u Austriji na lokalitetu Lohnbach
koji je takođe u neposrednoj vezi sa Dunavom.
Od haplotipova koji su pronađeni nizvodno i istočnije (basen Crnog mora,
Kaspijskog i Aralskog jezera), sa visokom učestalošću se takođe javlja haplotip Da1,
nešto ređe Da2, i Da-s6 (jedini Da haplotip koji na poziciji 26 poseduje tranziciju A→G).
Da-s6 haplotip je veoma redak i pripada posebnim ekofenotipskim formama iz jezera
Sevan (Jermenija, sliv - Kaspijskog jezera), prepoznatim kao posebna vrsta Salmo
ischchan. Behnke (1986) je dao hipotezu da se radi o pastrmci izvedenoj od primitivnog
pretka svih ostalih populacija potočne pastrmke. Bernatchez i Osinov (1995) i Osinov i
Bernatchez (1996) odbacuju pomenutu hipotezu na osnovu kombinovanih rezultata
alozima i mitohondrijalnih markera i zaključuju da Salmo ischchan predstavlja
morfološki i ekološki jedinstvenu formu pastrmki koja se razvila relativno skoro i pripada
istoj evolutivnoj liniji kao i ostale pastrmke iz basena Crnog mora, Kaspijskog i Aralskog
jezera. Pronalazak haplotipa Da-s6 na teritoriji Srbije u Studenačkoj reci (sliv Visočice i
Nišave) predstavlja prvi nalaz ovog haplotipa van jezera Sevan, što daje potvrdu gore
iznešenim tvrdnjama Bernatchez i Osinov (1995) i Osinov i Bernatchez (1996).
Slika 32. Pastrmke kod kojih je pronađen haplotip Da-s6.
Salmo ischchan - jezero Sevan (Jermenija)
Salmo trutta – Studenačka reka (Srbija)

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 82/141
Diskusija
74
Tri nova Da haplotipa (Da*Vl, Da*Dž, Da*Vr) pronađena su na razvođu
crnomorskog – dunavskog sliva (Južna Morava) i egejskog (Dragovištica/Struma) sliva
(Slika 1). Da*Vl je prvi dunavski haplotip koji na poziciji 390 poseduje tranziciju C→T,
što je odlika haplotipova At linije i jednog do sada pronađenog haplotipa Ad linije
(ADcs3) (Cortey i sar., 2004). Haplotip Da*Dž poseduje novo polimorfno mesto na
poziciji 61 (tranzicija, C→T), dok haplotip Da*Vr poseduje novi polimorfizam na
poziciji 228 (transverzija, G→T). Haplotipovi Da*Dž i Da*Vr se značajno odvajaju od
ostale grupe Da haplotipova (Slika 31) zbog polimorfizama na pozicijama 530 (tranzicija,
C→T), 542 (tranzicija, A→G), 543 (transverzija, C→G, važi samo za Da*Vr, dok Da*Dž
ima C kao i ostali Da haplotipovi) i 548 (tranzicija, T→C) (Tabela 13). Oba haplotipa
imaju na pozicijama 530, 542, 543 i 548 raspored baza karakterističan za pojedine
haplotipove ostalih linija. Za pomenute pozicije postoje sinapomorfizmi između Da*Dž i
Ad*Ti kao i između Da*Vr i najvećeg broja haplotipova svih ostalih linija (At, Ad, Ad+,
Me i Ma). Prema filogenetskom položaju klada za haplotipove Da*Dž i Da*Vr, može se
videti da je njihov položaj ancestralni u okviru Da linije (Slika 30) i istovremeno da su to
Da haplotipovi koji su najbliži haplotipovima svih ostalih linija (Slika 30 i 31). Ovakav
položaj pomenutih Da haplotipova podržava tvrdnju Cortey i sar . (2004) da, analizom
celog kontrolnog regiona mtDNA, topološki Da haplotipovi zauzimaju ancestralni položaj
u odnosu na ostale linije, što se slaže sa stanovištem Bernatchez i sar. (1992), Bianco
(1990) i Durand i sar. (2003) da recentna slatkovodna fauna Evrope ima istočnjačko
poreklo (Azija).
Na lokalitetima Jerma (br. 19) i Vrla (br. 22) koji se nalaze na razvođu dunavskog
i egejskog sliva, pronađen je haplotip ADcs1 koji je u Jermi jedini haplotip, a u Vrli je
pronađen zajedno sa novim dunavskim haplotipom Da*Vr. Objašnjenje za prisustvo
haplotipa ADcs1 u Jermi je njeno poribljavanje u gornjem delu toka kroz Bugarsku, kao i
njenih desnih pritoka koje izviru takođe na teritoriji Bugarske (usmeno saopštenje SašaPanić, Pirot). Poribljavanje je izvršeno matičnim materijalom iz sliva Strume (egejski
sliv) koji pripada Ad liniji. Upravo haplotip ADcs1 pronađen je u rekama sliva
Dragovištice/Struma (Tabela 9), što ide u prilog pomenutom usmenom saopštenju.
Prevođenje voda iz sliva Dragovištice u sliv Južne Morave za potrebe akumuliranja vode
radi proizvodnje električne energije (Ocokoljić, 1987), smatra se uzrokom prisustva
haplotipa ADcs1 u Vrli. Prevođenje vode sistemom cevi najverovatnije je omogućilo
prenošenje oplođene ikre ili čak mlade ribe iz jednog slivnog područ ja u drugo. Iz sveganavedenog, najverovatnije da je haplotip ADcs1 alohtonog porekla za oba lokaliteta.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 83/141
Diskusija
75
Na lokalitetu Gradac (br. 2) pronađen je haplotip At linije (At1), koji je alohtonog
porekla za deo crnomorskog sliva na teritoriji Srbije. Reka Gradac je više puta
poribljavana jer trpi veliki ribolovni pritisak, a jedno od poribljavanja vršeno je i
materijalom sa Vrela Bune gde se uzgajala potočna pastrmka At linije (usmeno saopštenje
Sreten Đor đević). Materijal koji se koristi za poribljavanje vrlo često je alohtonog porekla
i pripada komercijalnim linijama kao što je At, pre svega zbog prilagodljivosti
ribnjačkom uzgoju. U uzvodnom delu sliva Dunava u zoogeografskoj kontakt zoni
crnomorskog i atlantskog sliva pronađeni su haplotipovi At linije. Poznato je da je u
vodama Austrije, Švajcarske i Nemačke bilo masovnog poribljavanja upravo materijalom
At linije, ali pojedini autori smatraju da mogućnost post-glacijalne kolonizacije gornjih
delova sliva Dunava u oblasti kontakt zone od strane At haplotipova ne može biti u
potpunosti odbačena (Duftner i sar., 2003).
Bernatchez i Osinov (1995) i Osinov i Bernatchez (1996) smatraju da populacije
Da linije unutar basena Crnog mora i Kaspijskog i Aralskog jezera odlikuje prisutnost
posebnih alela ili mtDNA genotipova, kao i visok nivo genetičke diferenciranosti što
treba uzeti u obzir prilikom programa konzervacije. Rezultati dobijeni na pastrmskim
populacijama crnomorskog sliva iz Slovenije, Austrije i Srbije u potpunosti podržavaju
stavove pomenutih autora.
Apostolidis i sar. (1997) podelili su vode slivova Gr čke u dve zone Južno –
Jadransko Jonsku zonu i Ponto – Egejsku zonu. Analizirani deo jadranskog sliva (Beli
Drim) na teritoriji Srbije pripada po pomenutoj podeli severnom delu Južno – Jadransko
Jonske zone. Na ovom delu analizirane teritorije, od tri pronađena haplotipa Ad*Pe je
novi, a dva su već od ranije poznata: ADcs11 i Ad+Prz (Slika 1; Tabela 10). Haplotip
Ad*Pe je jedinstven po tranziciji (G→A) na 387 mestu, koja do sada nije bila opisana i
pronađen je samo u Pećkoj Bistrici. Haplotip ADcs11 (Cortey i sar., 2004) ili AD-s7 poBernatchez (2001), pronađen je u Pećkoj Bistrici ali i njenim pritokama (Tabela 10) i
prvobitno je bio opisan od strane Apostolidis i sar. (1997) u slivu Jonskog mora kao
ADs7r9. To je jedini haplotip koji na poziciji 80 poseduje tranziciju T→C.
Haplotip Ad+Prz pronađen je u Mrtvici (pritoka Morače) (usmeno saopštenje Aleš
Snoj), ali nije još zvanično objavljen. Na teritoriji Srbije pronađen je na dva lokaliteta u
jadranskom (Prizrenska Bistrica; br. lok. 32/Beli Drim) i egejskom slivu (Tripušnica; br.
lok. 29/Vardar). Haplotip Ad+Prz poseduje tranziciju C→T na poziciji 126 i 262 što jekarakteristika samo još dva poznata haplotipa iz Ad linije, jedan je iz Neretve (Ad+N) a

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 84/141
Diskusija
76
drugi je iz Rijeke Crnojevića (Ad+RC) (usmeno saopštenje Aleš Snoj). Haplotip AdN
jedino poseduje C na poziciji 243, dok AdRC haplotip poseduje T na poziciji 26, i to su
dva mesta po kojima se ovi haplotipovi razlikuju od Ad+Prz.
Zbog specifičnih sinapomorfizama (pozicije 126 i 262), haplotipovi Ad+Prz,
Ad+N i Ad+RC formiraju posebnu grupu sa visokom bootstrap potporom za sve tri
urađene analize. Ova grupa je pozicionirana kao sestrinska u odnosu na ostale jadranske,
mediteranske i marmoratus haplotipove (Slika 30), pa je zato i označena kao nova Ad+
linija. Pošto sva tri haplotipa naseljavaju reke u slivu južnog Jadrana, i jasno se grupišu
kao i ostale linije sa visokim bootstrap vrednostima, možda možemo predpostaviti da se
radi o Salmo fariodes (Karaman, 1937), za koju je opisano da naseljava pritoke srednjeg i
južnog Jadrana (Krka, Neretva, Beli Drim, pritoke Ohridskog i Skadarskog jezera).
Međutim, nalaz istog haplotipa u slivu Vardara (Tripušnica) izvan je areala u kome je
opisan, što je najverovatnije posledica poribljavanja.
U Ponto – Egejskoj zoni na teritoriji Srbije analizirali smo izvorišne delove reka u
slivovima Vardara i Strume.
Na analiziranim lokalitetima pronađeno je pet haplotipa, od kojih su dva nova
(Ad*Ti i Ad*Bož), a tri su već bila poznata (ADcs1, Ad+Prz i At1) (Slika 1; Tabela 10).
Na lokalitetima Tisova i Čerenačka reka (br. lok. 30 i 31/desne pritoka Vardara),
pronađen je novi haplotip Ad*Ti koji se za samo jednu transverziju (G→C) na poziciji
543 razlikuje od haplotipa ADcs1. U okviru iste zone, samo u levoj pritoci Vardara
(Tripušnica) ponađen je haplotip Ad+Prz koji je predhodno pomenut u jadranskom slivu.
Prisustvo Ad+Prz haplotipa u Tripušnici najverovatnije je posledica poribljavanja koje je
izvedeno 1978. godine putem mlađi koja je nabavljena u Konjicu (Neretva – jadranski
sliv). Pošto je haplotip Ad+Prz pronađen u slivu Belog Drima i Skadarskog jezera,
najverovatnije ga ima i u slivu Neretve, odakle je prenet u Tripušnicu, u kojoj do tada
pastrmke nije bilo (usmeno saopštenje «Ribolovačko društvo Trgovište»).U slivu Strume identifikovan je ranije pomenuti haplotip ADcs1 (Cortey i sar.,
2004) ili AD-s1 po Bernatchezu (2001), koji je već pronađen na više lokaliteta u okviru
egejskog sliva u Gr čkoj (Apostolidis i sar., 1997), ali i u slivovima Jadranskog, Jonskog i
Sredozemnog mora, kao i Atlantskog okeana (Bernatchez, 2001). Za ADcs1 haplotip
može se reći da je jedan od Ad haplotipa sa najširim rasprostranjenjem, što se može
objasniti njegovom izuzetnom sposobnošću kolonizacije. U istom slivu pronađen je novi
haplotip Ad*Bož koji na pozicijama 26 ima timin kao i haplotipovi ADcs16 i Ad+RC, alise od prvog razlikuje po citozinu na poziciji 262, a od drugog po citozinu na poziciji 126 i

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 85/141
Diskusija
77
262. Pošto se u istom slivu nalaze haplotipovi ADcs1 i Ad*Bož, najverovatnije su oni
naseljeni tokom dve nezavisne kolonizacije, pri čemu je haplotip Ad*Bož verovatno
stariji, pre svega zbog njegove topologije na filogenetskom stablu (Slika 30). Prema
neobjavljenim rezultatima najskorijih ispitivanja kraćih sekvenci (390 bp) kontrolnog
regiona mtDNA uzoraka iz Rilske reke (sliv Strume) i Arde (sliv Marice), utvr đeno je
prisustvo haplotipa koji do 390 bp odgovara haplotipu Ad*Bož. Ako ostatak sekvence
bude identičan kao kod haplotipa Ad*Bož, ovaj haplotip biće karakterističan za egejski
sliv. Međutim, ista sekvenca kao za Rilsku reku i Ardu pronađena je analizom uzoraka iz
Krke (jadranski sliv), koja je verovatno različit haplotip od Ad*Bož, što će se,
predpostavljamo, utvrditi budućim sekvencioniranjem još nedostajućih 170 bp do poli T
bloka. Podudarnost između sekvenci egejskog i jadranskog sliva verovatno je posledica
paralelizma (paralelne slučajne mutacije) na poziciji 26 (Tabela 10) po kojoj se ovaj
haplotip i razlikuje od najrasprostranjenijeg haplotipa jadranske linije ADcs1 (usmeno
saopštenje Aleš Snoj).
Ad*Bož haplotip zauzima ancestralni položaj u odnosu na sve ostale haplotipove
“južnih” – mediteranskih linija (Ad, Ad+ ( farioides), Me, i Ma) (Slika 30). Haplotip
Ad*Bož se razlikuje od haplotipova Me linije po četiri mutacije, od farioides i Ma
haplotipova po tri, i od svih Ad haplotipova po dve mutacije, izuzev od ADcs1 od koga se
razlikuje po samo jednoj mutaciji (Slika 31). Ancestralni položaj haplotipa Ad*Bož treba
dokazati i rezultatima drugih DNA analza, ali i analizom drugih tipova kararaktera
(morfološki), zbog nedetektabilno niskih “bootstraping” verovatnoća za NJ i MP analize,
i zbog toga što postoje tri druga haplotipa (ADcs11, AD*Pe and Ad*Ti) koja su barem
podjednako udaljena od Ma, Me i drugih Ad haplotipova po broju mutacija (Slika 31).
Vrlo je teško dodeliti taksonomski status nekom od pronađenih egejskih
haplotipova. Karaman (1924) je opisao samo jednu vrstu, Salmo macedonicus, u reci
Treski (sliv Vardara), čije ime je kasnije primenjeno i na potočnu pastrmku iz rekeStrume. Posebnu poteškoću pri rešavanju taksonomskih problema predstavlja postojanje
dva haplotipa u slivu Strume (ADcs1 i Ad*Bož) koji se fenotipski razlikuju, što pravi
dodatnu nomenklaturnu komplikaciju.
U slivu Strume na lokalitetu Brankovačka reka (br. 28) pronađen je haplotip At
linije (At1), isti kao i na lokalitetu Gradac u crnomorskom slivu. Haplotipovi At linije
nisu autohtoni za egejski sliv i njihovo prisustvo u Brankovačkoj reci najverovatnije je
posledica poribljavanja Dragovištice u delu toka kroz Bugarsku, mada se ne isključuju i

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 86/141
Diskusija
78
poribljavanja u Srbiji o kojima nema ni pismenih ni usmenih dokaza, zbog loše
organizovanog administrativno-ribarstvenog upravljanja.
Rezultati istraživanja populacija potočne pastrmke na obrađenim lokalitetima
egejskog i jadranskog sliva u potpunosti su u skladu sa dosadašnjim saznanjima da
mediteransko – jadranska oblast predstavlja poseban region Evrope u kome Salmo trutta
kompleks ispoljava najviši fenotipski diverzitet (Behnke 1968), što je praćeno takođe
visokom genetičkom raznovrsnošću, koja je utvr đena studijama na jedarnoj i
mitohondrijalnoj DNA (Laikre i sar., 1999). Najveći broj ranijih taksonomskih
klasifikacija bio je zasnovan na fenotipskoj varijabilnosti, što je rezultiralo kompleksnom
biogeografskom strukturom posebno populacija Balkanskog poluostrva, a što nije
potvr đeno rezultatima dobijenim upotrebom savremenih molekularnih tehnika
(Karakousis i Triantaphyllidis 1990, Apostolidis i sar., 1997).
Generalno, može se reći da je očekivano velika varijabilnost linija potočne pastrmke
Bernatchez (1992) između slivova na teritoriji Srbije (Tabela 12) praćena veoma značajnom
međupopulacionom varijabilnošću unutar slivova, što je najverovatnije posledica skorašnje
izolacije i lokalne diferencijacije unutar svake od linija.
Evropske populacije potočne pastrmke pokazuju biogeografska razgraničenja koja
su poslužila pri definisanju rečnih basena i slivova kao diskretnih jedinica (Apostolidis i
sar., 1997; Bernatchez i sar., 1992; Giuffra i sar., 1994). Pripadnost haplotipova
pojedinim linijama (što je uglavnom praćeno i pripadnošću određenim rečnim basenima),
može se uvideti preko filogenetskih stabala (Slika 29 i 30) urađenih različitim metodama
(NJ, MP, Majority role consensus 50%), analizom haplotipova iz svih mtDNA linija
(Bernatchez i sar., 1992). Rezultati koji se mogu izdvojiti kao posebno značajni su:
•
diferenciranost haplotipova u linije
• razdvajanje Da i At klade od klade ostalih «južnih» linija (Ad, Ad+ Ma i Me)
• ancestralni položaj haplotipova Da*Dž i Da*Vr u Da kladi
• formiranje posebne “ farioides” (Ad+) grupe (linije), koja ima potpuno istu značajnost
kao i grupe ostalih južno evropskih linija

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 87/141
Diskusija
79
U oba slučaja (Slika 29 i 30), dobijeni rezultati su posledica već pomenutih
sinapomorfizama (Tabela 13). Međutim, striktna interpretacija filogenetskih stabala je da
Da-At klada zauzima sestrinski položaj u odnosu na ostale «južno» evropske linije.
Upotrebom pomenutih metoda do sada nije dobijena ovakva topologija stabla, ali može se
reći da se ona slaže sa predloženim evolutivnim modelom Salmo trutta – kompleksa, po
kome su najstarija filogeografska razdvajanja povezana sa alopatričkom fragmentacijom
u gornjem i srednjem pleistocenu između tri glavna evropska basena: atlantskog (At
linija), ponto – kaspijskog (Da linija) i mediteranskog (Ad, Ma i Me linija). Kasnija
divergencija ili subsegmentacija u mediteranskom basenu dovela je do nastanka Ad, Ma i
Me linija (Bernatchez i sar., 1992; Patarnello i sar., 1994; Snoj i sar., 2002), a rezultati
ove studije promovišu još jednu liniju ( farioides ili Ad+) unutar ovog basena.
Pripadnost haplotipova određenim linjama kao i broj polimorfizama između njih
može se utvrditi i preko kladograma (mreže) (Slika 31), s tim što utvr đivanje broja
mutacija između haplotipova koji pripadaju različitim linijama preko broja algoritamskih
mutacionih koraka nije potpuno precizno. Rezultati dobijeni na osnovu kladograma
uglavnom se podudaraju sa rezultatima filogenetskih analiza (Slika 29 i 30). Jedina
značajnija razlika je što su Da haplotipovi na mreži zauzeli ancestralni položaj u odnosu
na sve ostale haplotipove, dok na filogenetskim stablima (Slika 29 i 30) nije moguće
utvrditi koja je linija u okviru Salmo trutta kompleksa ancestralna. Da i At haplotipovi
čine sestrinsku kladu u odnosu na kladu «južnih» haplotipova, s tim što je u okviru Da i
At klade, At linija započela nešto kasniju diferencijaciju na haplotipove. U «južnoj» kladi
sve linije (Ad, Ad+, Me i Ma) pokazuju sličnu starost (vreme diferencijacije), koje je pri
dobijenoj rezoluciji stabla nešto između vremena diferencijacije Da i At haplotipova, što
se podudara i sa tvrdnjom Bernatcheza (2001). Izrazita ancestralnost haplotipa Aa*Bož u
okviru «južne» klade, kao što je već rečeno, tek mora biti ispitana.
Cortey i sar. (2004) sekvencioniranjem celog kontrolnog regiona mtDNA iupotrebom istih filogenetskih analiza dobija takvu rezoluciju stabla pri kojoj Da linija
zauzima izrazito bazalni položaj u odnosu na ostale linije, kao što je to slučaj i na našem
kladogramu (Slika 31). Smatramo da bi i naša filogenetska stabla potvrdila ancestralniji
položaj Da linije sekvencioniranjem celog kontrolnog regiona mtDNA analiziranih uzoraka.
Na osnovu rezultata sekvencioniranja, osim detekcije haplotipova, utvr đena je i
varijabilnost unutar 5' kraja nekodirajućeg kontrolnog regiona mtDNA. Unutar fragmenta
kontrolnog regiona dužine 561 bp pronađeno je 18 polimorfnih mesta, što je u proseku jedno polimorfno mesto na svakih 31,2 nukleotida. Utvr đena varijabilnost unutar

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 88/141
Diskusija
80
kontrolnog regiona mtDNA analiziranih uzoraka nešto je manja u odnosu na rezultate
Bernatchez i sar. (1992), koji su analizom fragmenta dužine 310 bp pronašli 13
polimorfnih mesta, što je u proseku jedno polimorfno mesto na 23,8 nukleotida. Razlog
za manju varijabilnost uzoraka koji su analizirani je neravnomeran raspored polimorfnih
mesta. Najveći broj polimorfnih mesta se upravo nalazi u prvoj trećini 5' kraja kontrolnog
regiona, dok se dalje prema sredini njihov broj smanjuje.
Salmonide odlikuje približno jednaka varijabilnost kodogenih i nekodogenih
regiona molekula mtDNA (Giuffra i sar., 1994), što je razlika u odnosu na ostale
kičmenjake, kod kojih je varijabilnost kontrolnog regiona 5 do 15 puta veća od
varijabilnosti u kodirajućim regionima (Aquadro i Greenberg, 1983).
U kontrolnom regionu mtDNA najčešće se javljaju tačkaste mutacije, koje
nastaju kao rezultat supstitucija, delecija ili insercija. Do pojave tačkastih mutacija
najčešće dolazi usled pogrešnog sparivanja nukleotida prilikom replikacije, ili kao
rezultat spontanih mutacija koje nisu vezane sa replikacijom, i koje nastaju najčešće
zbog nukleotidne depurinizacije i/ili deaminacije (Alberts i sar., 1994). Pogrešno
spareni nukleotidi mogu biti uklonjeni pomoću posebnog mehanizma koji vrši
popravku na DNA, na taj način što prepoznaje i izrezuje pogrešno sparene nukleotide
prilikom polimerizacije i ligacije novog lanca (Lewin, 2000). Mehanizmi za
uklanjanje grešaka su efikasni ali nisu stoprocentno sigurni, tako da se neke mutacije
mogu fiksirati i dalje prenositi na potomstvo, što dovodi do genetskih promena
narednih generacija (Snoj, 1997).
U kontrolnom regionu mtDNA kao najčešće greške pri sparivanju nukleotida
javljaju se supstitucije, dok su delecije i insercije znatno ređe. Među supstitucijama,
tranzicije su mnogo češće u odnosu na transverzije, a među tranzicijama dominiraju
supstitucije C→T (Li i Grauer, 1991), što su i rezultati ovog rada pokazali (Tabela 10).
Transverzije G↔C su vrlo retke među supstitucijama, i zbog toga imaju veoma
veliki značaj pri utvr đivanju srodnosti između različitih sekvenci. Zbog težine
nastanka takve supstitucije, najverovatnije da je većina takvih zamena posledica
višestrukih supstitucija na istom mestu (npr.: G→A→C). Takav tip supstitucija javlja
se i u rezultatima ovoga rada na tri polimorfna mesta (26, 262, 543). Prema dobijenim
rezultatima, odnos tranzicija i transverzija je 14:7, a broj transverzija je nešto povećan
zbog polimorfne pozicije (26) na kojoj se javljaju dve transverzije (Tabela 10).
Bernatchez i sar . (1992) dobili su odnos tranzicija i transverzija (17:6) na 12

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 89/141
Diskusija
81
haplotipova potočne pastrmke, dok je odnos tranzicija i transverzija na lipljanu
identičan sa rezultatima ovoga rada - 14:7 (Sušnik, 2001). Za razliku od Salmonida,
kod većine drugih kičmenjaka na intraspecijskom nivou preovlađuju tranzicije nad
transverzijama (10 do 32 puta), kako u kodirajućim tako i u nekodirajućim regionima
mtDNA (Moritz i sar., 1987).
Snoj (1997) smatra, da što je stepen divergencije između dve nukleotidne
sekvence manji, mogućnost da na određenom mestu dođe do više od jedne supstitucije
gotovo je zanemarljiva i broj uočenih supstitucija između takve dve sekvence trebalo bi
da koincidira sa stvarnim brojem supstitucija. U suprotnom slučaju, kada postoji velika
razlika između sekvenci, uočeni broj razlika je manji od stvarnog, što je posledica
višestrukih supstitucija na istom mestu.
4.2. PALEOZOOGEOGRAFSKA RAZMATRANJA, SA OSVRTOM NAKOLONIZACIJU EVROPE POTOČ NOM PASTRMKOM
Da bi se dobio uvid u današnje rasprostranjenje i poreklo slatkovodnih ribaEvrope, samim tim i potočne pastrmke, neophodno je izvršiti rekonstrukciju
paleogeografskih događaja koji su imali veoma značajan uticaj na hidrografiju Evrope i
Mediterana tokom srednjeg i donjeg miocena, pliocena i pleistocena.
Tokom jure i krede Tetis je bio otvoreno ekvatorijalno more koje je razdvajalo
Evroaziju i Afriku, i spajalo prvobitni Atlantik sa Indopacifičkim okeanom. Pomeranjem
Evroazijske i Afričke ploče morski put između pomenutih okeana i Tetisa bio je
redukovan ili prekinut, a kao rezultat toga, tokom gornjeg i srednjeg miocena, formirala
su se dva glavna unutrašnja basena: Mediteran i Paratetis (Hsü i sar., 1977).
Za vreme srednjeg miocena (pre 15 miliona godina) dolazi do orogeneze Alpskog,
Dinarskog, Balkanskog i Pindskog planinskog masiva, što dovodi do formiranja izrazite
barijere između Mediterana i Paratetisa. Kao rezultat formiranja ovakve barijere, dolazi
do preusmeravanja velikog broja bivših centralno evropskih rečnih slivova u Paratetis,
koji takođe prima slatku vodu sa šireg područ ja centralnog Palearktika. Višak vode koji se
javljao u Paratetisu rezultirao je pozitivnim vodnim bilansom što je dovelo do prelivanja
vode u susedna mora Mediterana preko transalpijske, panonske i transegejske veze (Slika

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 90/141
Diskusija
82
33). Analizom sedimenata dna na područ ju današnjeg Crnog mora, a predačkog
Paratetisa, navodi se da je u periodu 15 – 5 miliona godina Paratetis bio oligosalino
(bočatno) ili čak slatkovodno «more». U istom periodu Mediteran je posedovao slanu
vodu i funkcionisao je kao otvoreno more, usled postojanja široke veze sa Atlantskim
okeanom (Hsü, 1978). Pored formiranja prirodnih barijera u vidu pomenutih planinskih
masiva ipak je između Paratetisa i Mediterana postojala komunikacija do kraja miocena.
Slika 33. Rekonstrukcija geografije Mediterana i Paratetisa tokom srednjeg miocena.
Moguće morske veze: A i B - veze Mediterana i Atlantika; C – Transalpijska brazda; D –
Jadransko - Panonska veza; E – Transegejska veza; F – Paratetis Baltička veza; G -
Paratetis Arktička veza (Hsü, 1978; modifikovano).
Najveći broj fosilnih ostataka riba iz srednjeg miocena pronađenih u Evropi
predpostavlja se da potiče iz Sibira i istočne Azije (Banarescu, 1960; 1977). Smatra se da
je kolonizacija Evrope izvršena preko mreže rečnih veza između basena Lene, Jeniseja iOba sa Paratetisom, odakle je preko Tauranskih vrata omogućena kolonizacija istočne
Evrope, a opadanje saliniteta u Paratetisu praćeno je daljom kolonizacijom ka zapadnoj
Evropi i Engleskoj (Bianco, 1990).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 91/141
Diskusija
83
Krajem miocena (6,1 do 5,1 miliona godina), u tzv. mesinskom periodu, desila su
se dva veoma važna geološka događaja:
1. Značajno povećanje zapadno Antarktičke ledene mase, koje je praćeno
opadanjem nivoa svetskog mora.
2. Približavanje afričke i evropske ploče, u kombinaciji sa opadanjem nivoa mora,
rezultira izolacijom Mediterana od Atlantika.
Kao posledica navedenih događaja, Mediteran postaje evaporativni basen
(McKenzie i Oberhansli, 1985; Cita i McKenzie, 1986), koji zbog izdizanja planinskih
lanaca Alpskog, Dinarskog, Balkanskog i Pindskog masiva gubi vezu i sa Paratetisom
(Slika 34). Od momenta izolacije Mediterana od Paratetisa svaki od basena prolazi kroz
različitu istoriju koja je uglavnom izazvana različitim hidrološkim statusom. Isušivanje
Mediterana poznato je kao «mesinska kriza saliniteta», koja je trajala oko milion godina
(Slika 35).
Slika 34. Događaji koji su doveli do krize saliniteta: razdvajanje Mediterana od Atlantika
i od Paratetisa (Hsü, 1978; modifikovano).
Za vreme mesinskog perioda Paratetis je bio bočatna ili slatkovodna sredina, čiji
se nivo vode povećavao posebno u vlažnim periodima, što je uzrokovalo povremeno
povezivanje evropskih rečnih sistema, a samim tim i mogućnost za širenje ili pakizolaciju slatkovodnih riba.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 92/141
Diskusija
84
Slika 35. Mesinska kriza saliniteta, uzrok isušivanja Mediterana. (Hsü, 1978).
Krajem mesinskog perioda, slatka voda iz Paratetisa prodire u Mediteran koji je
do tada bio suv, pri čemu se basen Paratetisa redukuje na mrežu jezera (panonski,
dakijski, pontski i kaspijski basen), a voda koja je dospela u Mediteran ispunjava
depresije, takođe formirajući mrežu jezera (Hsü i sar., 1977; Rögl i Steininger, 1983).
Ovaj događaj poznat je kao «Lago Mare» faza Mediterana i Paratetisa (Slika 36).
Mediteran je bio ispunjen slatkom vodom oko 100 000 godina, do pred sam početak
pliocena, kada je došlo do otvaranja Gibraltarskog moreuza i ulaska slane vode iz
Atlantika u Mediteran.
Slika 36. Mesinska «Lago Mare» faza Mediterana i Paratetisa (Hsü, 1978; mod.).
Period «Lago Mare» nesumnjivo je odigrao veoma važnu ulogu u ranom prolasku
primarno slatkovodne faune riba iz Paratetisa u Mediteran, kao i u disrtibuciji pomenute
faune u peri-mediteranske rečne sisteme. Potvrdu ovakvom scenariju daju brojni fosilniostaci koji potiču iz mesinskog perioda.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 93/141
Diskusija
85
Morska voda koja iz Atlantika ulazi u Mediteran postepeno počinje da se preliva i
u Paratetis, posebno u njegov zapadni deo (oblast današnjeg Crnog mora, Kaspijskog i
Aralskog jezera), što dovodi do katastrofalnih posledica po specijalizovane zajednice
slatkovodnih riba (Slika 37) (Bianco, 1990).
Slika 37. Ponovno uspostavljanje saliniteta (morskih uslova) u basenima Mediterana i
Paratetisa. Crvene tačke predstavljaju hipotetičke prvobitne reke u kojima je mogla
preživeti primarno slatkovodna fauna riba Paratetisa (Hsü, 1978; modifikovano).
Najstariji fosilni ostaci roda Salmo iz područ ja Paratetisa pronađeni su u Hrvatskoj
u blizini Samobora (Samoborska Gora). Predpostavlja se da se radi o vrsti Salmo
immigratus (Gorjanović – Kramberger, 1891) koja je zauzimala stratigrafsku poziciju
donjeg Sarmata u okviru centralno-panonskog basena (Anđelković, 1989). Na osnovu
ovog nalaza može se zaključiti da je rod Salmo naseljavao Paratetis još u srednjem
miocenu, pre oko 13 miliona godina, a verovatno i ranije. Banarescu (1973) smatra da su
predstavnici familije Salmonidae kolonizirali evropske prostore iz voda Sibira, što može
biti vrlo verovatno pošto je Paratetis tokom srednjeg i donjeg miocena bio ispunjen bočatnom ili slatkom vodom i povezan sa istočno-sibirskim rekama (Jenisej, Lena i Ob)
(Bianco, 1990). Kasnije širenje Salmonida iz Paratetisa omogućeno je ka severu preko
veza sa Baltikom i ka zapadu preko veze sa Rajnom.
Fosilni nalaz vrste Salmo immigratus u panonskom basenu Paratetisa iz srednjeg
miocena može imati ključnu ulogu za razrešenje porekla roda Salmo u evropskim
vodama. Najverovatnije da je Salmo immigratus bio predstavnik istočno-azijskih
salmonida koji se naselio u Paratetisu. Kako je postojala miocenska veza Paratetisa iBaltika, verovatno je deo populacije Salmo immigratus koji je migrirao ka severu bio

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 94/141
Diskusija
86
predački za atlantskog lososa (Salmo salar ), dok je populacija koja je ostala u Paratetisu i
njegovim pritokama bila predačka za evropske pastrmke. Nakon davanja predačke osnove
za dve grupe roda Salmo (losos i pastrmka), Salmo immigratus izumire.
Atlantski losos danas naseljava isključivo severna mora i obale istočnog
Atlantika, i uopšte ga nema u današnjim basenima bivšeg Paratetisa (Crno more,
Kaspijsko i Aralsko jezero), što ukazuje na nastanak lososa upravo u morima severne i
zapadne Evrope. Činjenice da je Patatetis u Saramatskom katu bio ispunjen uglavnom
bočatnom vodom koja po svojim osobinama jako nalikuje vodi današnjih severnih
mora, kao i da su ga naseljavale ribe sa jako širokom ekološkom valencom u odnosu
na salinitet (Anđelković, 1989), uz pojavu anadromije («zavičajno ponašanje») koja je
karakteristična za lososa, mogu predstavljati dodatne argumente koji idu u prilog
iznetoj hipotezi da je predak lososa (Salmo immigratus) živeo u Paratetisu odakle je
migrirao ka severnim morima za koja predpostavljamo da su i bila mesto njegovog
nastanka, nakon prekida veze preko basena Rone i/ili Rajne sa basenom Atlantika i
njegovih priobalnih mora.
Najstariji fosilni ostaci potočne pastrmke pronađeni su na Kavkazu, datiraju sa
početka pleistocena i procenjuje se da su stari oko 2 miliona godina (Osinov i
Bernatchez, 1996), s tim što se smatra da je potočna pastrmka kao vrsta znatno starija.
Pošto se radi o fosilnom nalazu iz basena bivšeg Paratetisa, može se predpostaviti da
pastrmka vodi poreklo od Salmo immigratus kao jedine vrste roda Salmo koja je
naseljavala Paratetis u doba srednjeg i donjeg miocena. Derzhavin (1934) i Vladimirov
(1944; 1948) smatraju upravo populacije potočne pastrmke sa područ ja Kavkaza
najprimitivnijim formama, od kojih su širenjem svoga areala nastale sve ostale
populacije pastrmki koje naseljavaju južna mora. Antunes i sar. (2002) analizom
geneologije gena za transferin dolaze do zaključka da najancestralnije populacije potočne pastrmke naseljavaju pritoke Crnog mora, Kaspijskog i Aralskog jezera. Cortey
i sar. (2004) navode da korišćenjem filogenetskih metoda NJ i MP, haplotipovi
dunavske linije zauzimaju ancestralni položaj u odnosu na haplotipove ostalih linija. Iz
svega navedenog može se uvideti da postoje argumenti koji idu u prilog napred iznetoj
hipotezi o poreklu evropskih pastrmki od pretka koji je naseljavao Paratetis i njegove
pritoke u doba miocena.
Tokom pliocena (5 - 1.8 miliona godina), Paratetis prolazi kroz fazu redukcije svojeveličine, prvo raspadom na manje pojedinačne međusobno povezane basene, da bi kasnije

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 95/141
Diskusija
87
došlo do potpunog isčezavanja panonskog i dakijskog basena, čime se uspostavio današnji
sliv Dunava (Hsü, 1978). Po završetku pliocena, od Paratetisa ostaju samo Crno more,
Kaspijsko i Aralsko jezero (Rabrenović i sar., 2003), što vodi ka značajnoj izolaciji
pastrmskih populacija Paratetisa.
U drugoj polovini pliocena (3,1 miliona godina) desile su se značajne
klimatske oscilacije. Globalno pan-okeansko postepeno opadanje temperature
započelo je pre oko 2,4 miliona godina, što je nagoveštavalo veliko zahlađenje tokom
pleistocena (Rage i Roček, 2003).
Početkom pleistocena dolazi do formiranja ledenog pokrivača koji se povremeno
širio (glacijacija) ili povlačio (interglacijacija). U poslednjih 0,9 miliona godina, glacijacije
su se javljale u ciklusima od oko 100 000 godina (Roy i sar., 1996). Analizom genetičkih
podataka životinjskih populacija Evrope i Severne Amerike, došlo se do zaključka da
genetička diferencijacija današnjih populacija predstavlja, između ostalog, odraz uslova za
vreme pleistocenskih glacijacija, kao i kolonizacija koje su pratile povlačenje ledenog
pokrivača (Avise, 1992; Hewitt, 1996; Bernatchez i Wilson, 1998; Taberlet i sar ., 1998).
Slatkovodne i anadromne ribe predstavljaju važnu grupu organizama u zoogeografskim
studijama, na osnovu kojih je sugerisano da postoji značajan glacijalni uticaj na
geografsku distribuciju intraspecijskog genetičkog polimorfizma.
Bernatchez (2001) smatra da svih pet mtDNA linija potočne pastrmke (Bernatchez,
1992) imaju monofiletsko poreklo, a da su se razvile nezavisno kao rezultat predačke
alopatričke fragmentacije. Na osnovu širokog opsega divergencije (1 – 2% na milion
godina), pretpostavlja se da je do razdvajanja linija od predačke populacije došlo u periodu
između 0,5 i 2 miliona godina, što ukazuje da je formiranje glavnih filogeografskih grupa
povezano sa klimatskim promenama, kao i promenama staništa, što se podudara sa
pleistocenskim glacijacijama. Poseban akcenat je stavljen na klimatske promene u
poslednjih 700 000 godina (Anderson i Borns, 1994) koje su verovatno imale značajanuticaj u izolaciji nekoliko velikih rečnih sistema (danas slivova različitih basena: Rona,
Rajna i Dunav) (Villinger, 1986). Najstarije razdvajanje na linije potočne pastrmke
povezano je sa alopatričkom fragmentacijom između tri glavna basena: atlantskog (At –
linija), ponto – kaspijskog (Da – linija) i mediteranskog, s tim da je u mediteranskom
basenu verovatno kasnije došlo do istovremenih fragmentacija koje su dovele do
razdvajanja Me, Ma i Ad linije (Bernatchez, 2001).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 96/141
Diskusija
88
Razmatranjem pleistocenskih dešavanja i globalne geografske slike distribucije i
genetičkog diverziteta za svaku od linija, moguće je izvesti zaključak o hipotetičkim
centrima porekla za mtDNA linije potočne pastrmke.
Reke u slivu atlantskog basena predstavljaju centar porekla At linije. Međutim,
pošto je severni deo atlantskog basena bio pod jakim uticajem širenja ledene kape tokom
pleistocenskih glacijacija, smatra se da je ancestralni centar porekla At linije bio više južno,
verovatno u obalnim pritokama Iberijskog poluostrva ili čak severne Afrike (Bernatchez,
2001). Utvr đivanje centra porekla Da linije je prilično problematično zbog veoma brojnih
interkonekcija, kao i povećanja i smanjenja nivoa Crnog mora, Kaspijskog i Aralskog
jezera za vreme pleistocenskih glacijacija, koje ipak nisu imale toliko izrazit uticaj na
gubitak staništa kao u slučaju At linije (Arkhipov i sar., 1995). Na osnovu kavkaskih
fosilnih ostataka potočne pastrmke, smatra se da je ancestralna populacija od koje su
nastale sve ostale populacije Da linije naseljavala pritoke Crnog mora (Bernatchez, 2001).
Karakteristična slika geografske distribucije tri preostale linije (Me, Ma i Ad) značajno
potkrepljuje realnost poznatih mediteranskih refugijuma: jugozapadni (iberijsko –
mediteranski), centralni (jadransko – mediteranski) i istočni (balkansko – anadolijski)
(Keith, 1998). Me linija je pretežno pronalažena u slivu reka zapadnog Mediterana, zbog
čega se pretpostavlja da vodi poreklo upravo iz istog regiona. Ma linija je uglavnom
ograničena na sliv reke Po, ali naseljava i reke jadranskog sliva Slovenije i Bosne i
Hercegovine koje su mogle biti povezane za vreme faza maksimalnih interglacijacija
(Bianco, 1990). Alozimske studije takođe su potvrdile severno jadransko poreklo Ma linije
(Giffura i sar., 1996). Ad linija je predominantna u pritokama istočnog Mediterana u
odnosu na Ma i Me liniju, što ukazuje da verovatno vodi porelo iz Balkansko –
Anadolijskog refugijuma (Bernatchez, 2001), dok Ad+ linija najverovatnije vodi poreklo iz
refugijuma jugozapadnog Balkana, gde je i danas rasprostranjena.
Na osnovu alozimskih istraživanja utvr đeno je da je slika geografske distribucijelinija povezana sa postglacijalnim geološkim događajima, pri čemu je predstavljen model
rekolonizacije deglacijalnih područ ja severo-zapadne Evrope od tri preglacijalne linije (I, II
i III). Za vreme povlačenja leda smatra se da je došlo do rekolonizacije uglavnom u
graničnim područ jima i to iz tri smera: 1. severo – zapadna migracija iz mediteransko –
kaspijskog refugijuma (linija III), 2. migracija ka severu iz refugijuma atlantskog sliva
(Iberijsko poluostrvo i južna Francuska - linija II) i 3. severno i istočno širenje iz
refugijuma sa središtem u blizini Engleskog Kanala (Lamanš - linija I). Rekolonizacijadeglacijalnih područ ja severo-zapadne Evrope iz četvrtog mediteranskog refugijuma,

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 97/141
Diskusija
89
smatra se da nije vršena (linija IV) (García - Marín i sar., 1999), (Slika 38). Linije I, II i III
odlikuju se različitim kombinacijama fiksiranih genotipova na lokusima LDH-C* i CK-A1,
koji su danas prisutni u pastrmskim populacijama deglacijalnih područ ja, dok linija IV
poseduje alel LDH - A2*100QL, koji odsustvuje kod populacija iz postglacijalno
kolonizovanih regiona. Sadašnje populacije koje naseljavaju deglacijalna područ ja, smatra
se da predstavljaju mešovite ili čiste potomke navedene tri migratorne grupe.
Slika 38. Distribucija predpostavljenih glavnih linija (I, II, III i IV) i pravci migracija
populacija potočne pastrmke za vreme postglacijalne rekolonizacije za linije I, II i III.
Isprekidana linija predstavlja granicu leda za vreme poslednje glacijacije (pre oko 13 000
godina), (García - Marín i sar., 1999 - modifikovano).
García - Marín i sar. (1999) i Bernatchez (2001) predlažu sličan model
rekolonizacije koji se zasniva na brzom, istovremenom širenju populacija potočne pastrmke
iz glacijalnih refugijuma, kao što je sugerisao i Hewitt (1999) za druge organizme. Do
rekolonizacije bi došlo širenjem «prednjeg ruba» populacija migranata na veće distance
koje bi ustanovile «kolonije» daleko ispred glavnog talasa populacije. Ovi migranti bi senaglo širili i zauzimali staništa pre ostalih, čime bi njihovi geni postali dominantni u
genofondu novouspostavljene populacije. Ovakav tip rekolonizacije može dovesti do
pojave velikog stepena homozigotnosti, kao i efekta «uskog grla» (eng. bottleneck ) koji
može usloviti gubitak genetičkog diverziteta (Nei i sar., 1975). Severno atlantski basen
koji naseljava At linija potočne pastrmke odlikuje se malim genetičkim diverzitetom što
bi se moglo objasniti ovakvom hipotezom o načinu rekolonizacije faune. Prema scenariju,
populacije potočne pastrmke koje su ve
ć prisutne u refugijumu datog basena ili
geografskog prostora, bile bi prve koje bi se proširile i zauzele dostupne ekološke niše,

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 98/141
Diskusija
90
ostavljajući malu verovatnoću za osnivanje populacijama koje su preživele glacijacije u
udaljenijim delovima refugijuma. Ovaj proces bi bio još izrazitiji ako bi vreme
geografske izolacije (i/ili selekcije u različitim sredinama) bilo prilično značajno za
akumulaciju genetičkih i ekoloških razlika (Bernatchez, 2001). Naseljavanjem novog
područ ja populacijama sa «prednjeg ruba», populacije koje se nalaze iza njih vrlo malo
mogu uticati na genetičku strukturu populacije na «pionirskom» područ ju - u pitanju je
jednostavno gustina populacije. Kao rezultat ovog načina rekolonizacije desilo bi se da
populacije koje se nalaze «iza» populacija sa severne granice širenja (u centru ili na jugu
refugijuma) neće biti u mogućnosti da brzo napreduju, što znači da se one moraju
povlačiti na druga staništa ili opstati gde jesu. Pomenuti proces, uz fizičku izolaciju i
biološke faktore, može biti «generator» procesa nastanka specifičnih genoma i podvrsta u
južnim refugijumima (Hewitt, 1993).
Raspored kopna i mora kroz pleistocen postepeno se približava današnjem
rasporedu. Međutim, u starijem pleistocenu ima velikih odstupanja od današnjeg izgleda
obalskih linija. Takođe su velika odstupanja u odnosu na današnje stanje bila za vreme
velikih zahlađenja (glacijacija), uključujući i ona u mlađem pleistocenu (virm), kada je
usled povećanja kontinentalnih i planinskih lednika smanjena količina vode u okeanima i
snižen nivo mora (Rabrenović i sar., 2003).
Razmena primarno slatkovodne faune, a samim tim i pastrmskih populacija,
između reka različitih oblasti, bila je verovatno omogućena opadanjem nivoa mora za
vreme glacijacija (Bianco, 1990). Procene opadanja nivoa mora u Mediteranu su
različite za određene faze glacijacija. Za vreme Virm glacijalne faze smatra se da je
nivo Mediterana bio niži za maksimalno 130 m, dok je za vreme Mindel i Ris
glacijalnih faza nivo mora bio za oko 200 m niži u odnosu na današnji nivo (Bianco,
1990; Pielou, 1979). Postoje mišljenja da opadanje nivoa mora u Mediteranu za oko200 m pre oko 250 000 godina, za vreme Ris faze glacijacije (Brown i Gibson, 1983)
uslovljava sužavanje Gibraltarskog moreuza na 1/5 današnje širine. Kao posledica
ovih dešavanja dolazi do smanjenog dotoka vode u Mediteran iz Atlantika, što izaziva
sekundarnu krizu saliniteta i pad nivoa vode u Mediteranu za oko 500 m, za šta je bilo
potrebno oko 500 godina. Opadanjem nivoa vode u Mediteranu za 500 m može se
objasniti prisustvo slatkovodnih riba na Rodosu koje su porekom iz Turske, kao i
razmena slatkovodne riblje faune između severo – zapadne Afrike i Španije (Bianco,1990). Ovom pojavom se može objasniti i izuzetna širina areala Ad i Me linije u

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 99/141
Diskusija
91
Mediteranu, koja se najverovatnije javlja kao posledica uspostavljanja veza između
rečnih slivova različitih oblasti. Takođe treba pomenuti i rasprostranjenje At linije u
Mediteranu koja je pronađena u istočnoj Španiji, na severo – zapadu Afrike i na
Siciliji (usmeno saopštenje Aleš Snoj).
Slika 39. Narandžasta i crvena linija duž jadranske, jonske i egejske obale prikazuju
sniženje mora od 100 m (Virm faza) i 200 m (Ris i Mindel faza), kao i posledice tih
dešavanja u različitim regionima (1 – 7) (Bianco, 1990 – modifikovano).
U regionu 1 (Slika 39), kao posledica povlačenja nivoa mora, dolazi do značajnog
proširenja reke Po, u regionu 2 dolazi do uspostavljanja veza između dalmatinskih reka, u
regionu 3 dolazi do uspostavljanja transjadranske komunikacije (opadanjem nivoa mora
od najmanje 160 m), u regionu 4 dolazi do uspostavljanja veza između albanskih reka
(Drim ↔ Vjose), u regionu 5 dolazi do uspostavljanja veza između reka jonskog sliva(Taimis ↔ Aheron), u regionu 6 dolazi do uspostavljanja veza između reka sa suprotnih
strana Patras - korintskog zaliva, i u regionu 7 dolazi do uspostavljanja slatkovodne veze
između gornjeg dela Egejskog i Crnog mora.
Osim brojnih rečnih veza koje su se uspostavile opadanjem nivoa mora za vreme
glacijacija i koje su omogućile migriranje pastrmskih populacija, treba pomenuti i brojne
međurečne veze, ledene ustave i slatkovodna jezera koje su uspostavljene u doba
interglacijacija otapanjem ledenog pokrivača u zapadnom Sibiru i istočnoj Evropi. Kao posledica otapanja lednika dolazi do akumulacije i prelivanja vode iz jednog u drugi

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 100/141
Diskusija
92
basen bivšeg Paratetisa (Aralsko → Kaspijsko jezero → Crno more), koji je preko Crnog
mora komunicirao sa gornjim delom Egejskog mora. Ceo pomenuti sistem, uključujući i
gornji deo Egejskog mora, najverovatnije je posle svake glacijacije – interglacijacije
posedovao slatku ili bočatnu vodu (Slika 39), (Thunell i Williams, 1983). Kao potvrdu
veze između ponto – kaspijskog i mediteranskog basena, imamo nalaz Da haplotipova, za
koje se smatra da su autohtonog porekla u egejskom slivu Gr čke (Apostolidis i sar.,
1997), i Turske (Bernatchez, 2001). Takođe je postojala komunikacija između ponto –
kaspijskog basena i basena Tigrisa i Eufrata (Persijski zaliv), što potvr đuje pronalazak
dunavskog haplotipa Da25 (Sušnik i sar., 2005). Osim pomenutih veza između ponto –
kaspijskog basena sa mediteranskim i basenom Indijskog okeana (Persijski zaliv),
postojala je, za vreme različitih faza glacijacija, veza ponto – kaspijskog i atlantskog
basena (Bernatchez, 2001). Veze Kaspijsko jezero – Baltik i Kaspijsko jezero – Arktik
(Ekman, 1957), kao i veza između gornjeg Dunava i gornje Rajne (Balon, 1968;
Englbrecht i sar., 2000) potvr đena je i prisustvom At haplotipova u slivu gornjeg Dunava,
za koje se sumnja da su autohtonog porekla (Weiss i sar., 2001), kao i nalaz jedne
populacije potočne pastrmke u pritoci gornje Volge (kaspijski basen), (Osinov i
Bernatchez, 1996). Današnje filogeografske studije (Durand i sar., 1999; Nesbo i sar.,
1999), kao i tradicionalne biogeografske studije (Banarescu, 1990) potvr đuju ponto –
kaspijsko poreklo drugih slatkovodnih riba u atlantskom basenu koje su tu dospele za
vreme pleistocenskih glacijalnih događaja.
Kako su postojale pleistocenske veze između basena Crnog mora, Kaspijskog i
Aralskog jezera sa Atlantikom na jednoj i sa Mediteranom i Indijskim okeanom na drugoj
i trećoj strani, naša predpostavka da su populacije potočne pastrmke iz basena bivšeg
Paratetisa ancestralne za sve ostale populacije koje naseljavaju severna i južna mora
postaje realna. Realnost ove hipoteze ogleda se u postojanju haplotipova Da*Vr i Da*Dž
koji su pronađeni u slivu Južne Morave (Dunav → Crno more), koji zauzimajuancestralni položaj u okviru Da klade na filogenetskim stablima (Slika 29 i 30), kao i
haplotipa Da24 koji je pronađen u reci Waldaist u Austriji (dunavski sliv), (Duftner i sar.,
2003). Haplotipovi Da*Vr, Da*Dž i Da24 imaju poslednja četiri polimorfna mesta 530,
542, 543 i 548 do poly «T» regiona sinapomorfna sa haplotipovima iz Ad, Ad+ At, Ma i
Ma linije. Kao posledica pomenutih sinapomorfizama, haplotipovi Da*Vr, Da*Dž
zauzimaju centralni položaj na kladogramu – mreži (Slika 31), (posebno haplotip Da*Vr)
u odnosu na sve haplotipove iz ostalih linija. Haplotip Da*Vr zauzima centralni položaj i po procentualnim vrednostima divergencije između haplotipova različitih linija (Tabela

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 101/141
Diskusija
93
11). Između haplotipa Da*Vr i najbližih haplotipova «južne» i At linije distance se kreću
od 1,05 do 1,23%, dok je vrednost distanci između Da*Vr i Da haplotipova između 0,88 i
1,23%, izuzev haplotipa Da*Dž koji mu je najbliži i distanca između njih je 0,53%. Na
osnovu iznetih činjenica može se zaključiti da haplotip Da*Vr (kao i njemu najbliži
haplotipovi Da24 i Da*Dž) spadaju u grupu ancestralnih haplotipova ne samo za ostale
Da haplotipove, već i za haplotipove ostalih linija.
Unutar Da linije prikazana je duboka genetička divergencija, koja prema
genetičkoj distanci skoro tri puta pravazilazi genetičku divergenciju između
haplotipova Ad, Me i Ma linija. Duboka divergencija unutar Da linije (Tabela 11), kao
i njena bazalna pozicija na filogenetskom stablu (Cortey i sar., 2004) i na kladogramu
(mreži) haplotipova (Slika 31), sugerišu da Da populacije predstavljaju najstarije
odvojenu liniju iz Salmo trutta kompleksa, a samim tim i ancestralnu za druge
haplotipove (linije) unutar kompleksa.
Haplotipovi koji su pronađeni u slivu Neretve i Skadarskog jezera (Ad+N i
Ad+RC), a koji zajedno sa Ad+Prz formiraju jugo-zapadno balkansku farioides (Ad+)
liniju, verovatno predstavljaju ostatke ancestralnog polimorfizma «južne» klade.
Nakon prihvatanja stanovišta o postojanju pet glavnih evolutivnih linija (Da, At,
Ad, Me i Ma) potočne pastrmke (Bernatchez i sar., 1992), sumnjalo se u mogućnost
postojanja novih linija slične dubine divergencije, jer je smatrano da je distribuciono
područ je Salmo trutta kompleksa bar u pogledu pronalaska novih linija adekvatno
istraženo (Bernatchez, 2001). Međutim, ispostavilo se da pojedini regioni nisu dovoljno
istraženi, i da još uvek ne postoje podaci za pojedina područ ja u okviru prirodnog areala
potočne pastrmke. Najnovija detaljna istraživanja populacija potočne pastrmke na
Iberijskom poluostrvu otkrila su postojanje bitnih polimorfizama unutar At klade (Suárez
i sar., 2001; Weiss i sar., 2000) na osnovu kojih je predloženo opisivanje nove evolutivne
linije «Duero» (Suárez i sar., 2001; Cortey i Garcia-Marin, 2002). Pošto Balkansko poluostrvo, kao i Iberijsko, predstavlja «vruću tačku» biodiverziteta (Kryštufek i sar.,
2004), veoma bitnu za post-glacijalnu evoluciju faune i kolonizaciju Evrope, a pri tome je
slabo istraženo, pronalazak Salmo farioides (Ad+) linije u rekama jadranskog sliva jugo-
zapadnog Balkana, s obzirom na sve okolnosti područ ja i nedostatka informacija, postaje
sasvim realan.
Dobijeni rezultati nukleotidnih divergencija na kontrolnom regionu mtDNA od
0,18% do 2,12% (Tabela 11), uglavnom se slažu sa ocenom Gyllestena i Wilsona (1988)da je intraspecijska nukleotidna divergencija mtDNA kod Salmonida između 0.1% i 2 %.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 102/141
Diskusija
94
Primena molekularnog sata na kontrolni region mtDNA veoma je problematična zbog
neprecizne kalibracije koja se po različitim autorima kreće od 0,5% do 2% na milion
godina. Smith (1992) procenjuje stopu mutacija za mtDNA od 1% na milion godina za
Salmonide. Ako bi se vrednosti distanci za haplotipove pronađene na teritoriji Srbije
obračunale po mutacionoj stopi Smita od 1% na milion godina, dobile bi se vrednosti
divergencije između 180 000 i 2,12 miliona godina, što korespondira sa geološkim
epohama kraja pliocena i gotovo celog pleistocena. Iz ovoga proizilazi da su
pleistocenske glacijacije odigrale veoma važnu ulogu u oblikovanju genetičkog
diverziteta populacija potočne pastrmke. Međutim, treba pomenuti i procenu Apostolidis i
sar. (1997) koji smatraju da se divergencija glavnih mtDNA linija desila od zajedničkog
pretka u slično geološko vreme i to u doba mesinske periode i ranog pliocena, u periodu
od pre 2,5 – 6 miliona godina. U ovom periodu desila su se dva veoma važna geološka
događaja: 1. dolazi do izolacije Mediterana od Atlantika, kao i od basena Paratetisa (Slika
34), 2. krajem mesinskog perioda Mediteran i Paratetis bivaju redukovani na mrežu
povezanih jezera poznatu pod nazivom «Lago Mare» faza (Slika 36), (Bianco, 1990).
Pomenuti autori smatraju da je «Lago Mare» faza verovatno odigrala veoma važnu ulogu
u izolaciji pastrmki Mediterana, kao inicijalnih panmiktičnih populacija koje su bile
razdvojene u posebnim jezerima, iz kojih su se kasnije širile u peri-mediteranske rečne
sisteme (Slika 37). Međutim, postoje mišljenja koja odbacuju hipotezu Apostolidis i sar.
(1997) iz sledećeg razloga: pošto je u periodu krajem miocena i tokom prvog dela
pliocena klima postala toplija i humidnija, u Mediteranu su vladali tropsko - subtropski
uslovi (Rage, 1997), koji su verovatno sprečavali mediteransku ekspanziju salmonidnih
riba sve do pojave zahlađenja krajem pliocena i u pleistocenu (pre oko 2,5 miliona
godina) koje je zahvatilo i ovaj region (Cortey i sar., 2004), što i rezultati ovog rada u
potpunosti podržavaju.
Tokom pleistocena javljaju se orogeni pokreti na severnom obodu Mediterana
(Tirensko, Jonsko, Jadransko, Egejsko more, Makedonija, Trakija, Anadolija), što dovodi
do rasedanja u oblasti Makedonije, Skadarskog basena i na južnom obodu panonskog
basena (Stevanović, 1982). Pleistocenski orogeni pokreti, uz uticaj glacijalnih i
interglacijalnih kolebanja nivoa mora, koja su praćena uspostavljanjem rečnih veza, imaju
veoma značajan uticaj na rasprostranjenje i diverzitet pastrmskih populacija na područ ju
Balkanskog poluostrva. Jasno geografsko i filogenetsko razdvajanje Da haplotipova kojisu pronađeni isključivo u crnomorskom slivu, od Ad i Ad+ haplotipova koji su kao

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 103/141
Diskusija
95
autohtoni pronađeni u egejskom i jadranskom slivu (u zavisnosti od haplotipa) na teritoriji
Srbije, uz analizu molekularnog sata i geološke istorije šireg područ ja Balkana,
objašnjava razvoj pomenutih linija najverovatnije alopatričkom fragmentacijom za vreme
Pleistocena. Veliki diverzitet pronađenih haplotipova u okviru linija na analiziranom
područ ju, korenspondira sa fizičkom izolacijom kao i biološkim faktorima koji su
doprineli njihovom ograničenom rasprostranjenju.
4.3. KONZERVACIJA GENETIČKE RAZNOVRSNOSTI POPULACIJAPOTOČ NE PASTRMKE
4.3.1. SPECIFIČ NOSTI KONZERVACIJE POTO
Č NE PASTRMKE
Potočna pastrmaka, zbog veoma važnog socio-ekonomskog značaja, koji se
ogleda ne samo u ekološkoj vrednosti vrste već i u značaju za sportski i komercijalni
ribolov, predstavlja veoma ugroženu vrstu sa gledišta gubitka intraspecijske genetičke
varijabilnosti, najčešće kao posledice čovekovih aktivnosti (degradacija staništa,
prekomeran izlov, poribljavanje), (Laikre i Ryman 1996).
Specifičnost konzervacije potočne pastrmke ogleda se u sledećem:
Opsežnim genetičkim istraživanjima u poslednjih 20 godina na taksonu
koji je poznat kao «potočna pastrmka» otkriven je veoma značajan nivo
genetičke raznovrsnosti, koji se ogleda u postojanju više evolutivnih linija
(Bernatchez, 1992), kao i daljoj genetičkoj diferencijaciji između lokalnih
populacija unutar svake od linija. Veoma značajan deo genetičke
varijabilnosti predstavljen je na među-populacionom nivou, i to je glavni
razlog što se konzervacione mere, ribarstveno i konzervaciono upravljanje
ovom vrstom, moraju fokusirati na lokalne populacije (Frankel, 1970;
1974; Ryman i Ståhl, 1980; Utter, 1981; Meffe, 1986; Ryman, 1991).
Degradacija staništa dovodi do istrebljenja ili redukcije brojnosti lokalnih
populacija, a samim tim i do gubitka značajnog intraspecijskog diverziteta.
Najveći broj preostalih, pre svega ribolovno atraktivnih, populacija
potočne pastrmke veoma su ugrožene zbog svoga ekonomskog značaja,
pre svega zbog aktivnosti kao što su: prekomeran izlov i poribljavanje.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 104/141
Diskusija
96
Ove aktivnosti se smatraju bezopasnim ili čak korisnim, ali su najčešće
katastrofalne po genofond autohtonih populacija (Laikre i sar., 1999).
Osnovni faktori ugroženosti populacija potočne pastrmke koji dovode do gubitka
genetičke varijabilnosti obrazloženi su u poglavlju Uvod (1.2.6.), a mogu se videti i iz
tabele 14.
Tabela 14. Prikaz tri glavne kategorije aktivnosti koje ugrožavaju intraspecijsku
varijabilnost potočne pastrmke, kao i njihov odnos sa tri glavna procesa koja dovode do
gubitka genetičkog diverziteta (Laikre i sar., 1999).
Aktivnost Izumiranje Hibridizacija Gubitakvarijabilnosti
unutarpopulacija
Ribolov X X
Poribljavanje X X X
Degradacijastaništa
X (X) X
Aktivnosti koje utiču na degradaciju staništa kao i na prekomeran izlov,
nesumljivo prouzrokuju i treći vid aktivnosti, a to je poribljavanje. Poribljavanje se
uglavnom smatra glavnom ribarstvenom merom za povećavanje brojnosti pastrmskih
populacija. Odgovarajuće izvedeno poribljavanje može biti razmatrano kao značajna
mera pružanja «prve pomoći» populacijama koje su u neposrednoj opasnosti odizumiranja prouzrokovanog demografskim faktorima. Samo dva tipa poribljavanja mogu
omogućiti dugoročna pozitivna rešenja:
Poribljavanje iz konzervacionih razloga
Poribljavanje radi ponovnog uspostavljanja populacije (reintrodukcija),
s tim sto se mora imati u vidu da pomenuta poribljavanja mogu biti pozitivna samo uz
utvr đivanje i otklanjanje faktora koji su doveli do redukovanja ili izumiranja
populacije. U ostalim slučajevima (naj
češće radi poboljšanja ribolova), poribljavanje
predstavlja glavnu pretnju genetičkom integritetu populacije (Hindar i sar., 1991a).
Riblja mlađ koja se koristi za poribljavanje najčešće je genetički različita od
autohtonih populacija, i predstavlja uglavnom domestifikovane - ribnjačke populacije.
Rezultati do kojih su došli Apostolidis i sar. (1996a, 1997) pokazuju da je
poribljavanje reke Nestos (egejski sliv) neautohtonim materijalom iz reke Aheloos
(Jonski sliv), radi poboljšanja sportskog ribolova, imalo veoma mali doprinos u
poboljšanju ribolova. Preko mtDNA markera procenjeno je da je oko 75% autohtone
populacije bilo zamenjeno poribljenom pastrmkom.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 105/141
Diskusija
97
4.3.2. STATUS UGROŽENOSTI POTOČ NE PASTRMKE U EVROPI
Značajan problem koji se javlja pri konzervaciji potočne pastrmke vezan je upravo za
taksonomsku konfuziju unutar pomenutog taksona (pogledati Uvod 1. 2. 4). Glavni razlog pojave taksonomskih problema su nedoumice oko klasifikovanja različitih morfoloških i/ili
genetičkih grupa, koje se prema nekim autorima smatraju vrstama, a prema drugim podvrstama
ili specifičnim genetičkim formama unutar vrste (Laikre i sar., 1999). Međutim, taksonomska
konfuzija ne sme imati presudan uticaj na određivanje konzervacionih mera i samog
gazdovanja ovim taksonom. Efikasna konzervacija potočne pastrmke može se realizovati samo
na jedan način, a to je posvećivanje pažnje genetičkim razlikama između populacija, bez obzira
da li ove populacije nazivamo vrste, podvrste ili lokalne populacije.
Eksperti «TROUTCONCERN» mreže složni su u stavu da se trenutna ugroženost
potočne pastrmke ne shvata dovoljno ozbiljno, kao i da je sadašnji konzervacioni napor
fokusiran samo na nivo vrste, ne uzimajući pri tome u obzir genetički diverzitet unutar vrste. U
slučaju da se ovakav trend nastavi, veoma brzo će doći do gubitka dragocenih bioloških resursa
u vidu lokalnih populacija potočne pastrmke, koje će izumirati neslućenom brzinom (Laikre i
sar., 1999).
Pored toga što trenutni klasifikacioni sistem ne uzima u obzir intraspecijsku genetičku
strukturu, veoma veliki nedostak koji onemogućava konzervaciju ne samo potočne pastrmke
već i ostalih vrsta riba, jeste što je generalno konzervacioni status do sada ustanovljen za
relativno mali broj taksona riba. Upavo pomenuta dva nedostatka predstavljaju glavnu manu
raznih lista ugroženih životinjskih vrsta (Ryman i sar., 1995a; Leidy i Moyle, 1998). Potočna
pastrmka nije uvrštena na listu Bernske konvencije, kao ni na Crvenu Listu (IUCN) ugroženih
životinjskih vrsta na internacionalnom nivou. Na Crvenoj Listi status ugroženih taksona dobili
su: Salmo carpio, Salmo letnica, i Salmo platycephalus, dok su dva druga taksona Salmo
marmoratus i Salmo dentex uvrštena u kategoriju vrsta sa nedovoljno podataka. Taksoni Salmo
marmoratus i Salmo macrostigma uključeni su na listu «EU Habitat Directive» kao vrste od
opšteg interesa koje zahtevaju primenu posebnih konzervacionih mera. Eksperti
«TROUTCONCERN» mreže saglasni su da sadašnja Crvena Lista u najvećem broju slučajeva
neadekvatno procenjuje ugroženost potočne pastrmke, iz razloga što se glavni ugrožavajući
faktori ne uzimaju sa dovoljnom ozbiljnošću.
Allendorf (1988) smatra da Crvena Lista ugroženih riba u velikoj meri reflektuje
geografsku distribuciju ihtiologa, a ne ugroženih vrsta.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 106/141
Diskusija
98
4.3.3. DEFINISANJE KONZERVACIONIH JEDINICA
Pri svakoj konzervacionoj aktivnosti postavlja se osnovno i krucijalno pitanje: Šta
treba da bude predmet biološke konzervacije? Iz prethodno iznešenih stavova jasno je dakonzervacija vrste predstavlja zastareli princip, i da svakako nije dovoljna jer ne uzima u
obzir genetičku varijabilnost između i unutar populacija, koja takođe mora biti sačuvana
(Frankel i Soulé 1981). U poslednjih 20 godina vođene su debate na temu utvr đivanja
odgovarajuće jedinice za konzervaciju. Najveći broj autora složio se da konzervacioni
napor treba usmeriti ka nivou evolutivnih linija unutar vrste (Ryder, 1986; Waples,
1991a; 1995; Moritz, 1994; 1995; 1999). Evolutivne linije unutar vrste nazvane su
«Evolutionary Significant Units» ili ESU. Ovaj princip je prvo bio primenjen na
pacifičkom lososu (Waples, 1991a), ali je sugerisano da je primenjiv za bilo koju vrstu
(Ryder, 1986).
Waples (1991a) definiše «Evolutionary Significant Units» kao populaciju ili
grupu populacija koje su suštinski reproduktivno izolovane od drugih konspecijskih
populacionih jedinica, i koje predstavljaju važnu komponentu evolutivnog nasleđa vrste.
Evolutivno nasleđe vrste čini genetička varijabilnost kao produkt prošlih evolucionih
događaja, koja predstavlja osnovu od koje će zavisiti buduća evolucija vrste.
U pogledu nivoa minimalne značajnosti koja bi bila neophodna za dodeljivanje
ESU ne postoji određeni standard (Nielsen, 1995), tako da odluka šta može biti smatrano
kao biološki – evolutivno značajno mora biti rešavana od slučaja do slučaja (Waples,
1995). Metode za utvr đivanje ESU uključuju sakupljanje genetičkih i ekoloških podataka,
kao i njihovu analizu fenetičkim i kladističkim tehnikama radi utvr đivanja hijerarhijskih
genetičko – ekoloških odnosa između populacija (Waples, 1991a; 1995; Nielsen, 1995).
Kada se odnosi između populacija utvrde, neophodno je doneti odluku o nivou na koji će
biti fokusiran konzervacioni napor (Dodson i sar., 1998), a to je onaj nivo koji nosi
značajnu komponentu evolutivnog nasleđa vrste.
Rešavanje pitanja konzervacije od slučaja do slučaja, uvođenjem koncepta ESU,
predstavlja pokušaj da konzervacija ignoriše filogenetski (pragmatični) koncept vrste
(Cracraft, 1987), kako bi preskočila taksonomske diskusije.
Trenutno u Evropi nije zauzet jasan stav oko definisanja ESU za potočnu
pastrmku.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 107/141
Diskusija
99
U velikom broju realnih situacija odluka o konzervacionom naporu neće biti
zasnovana isključivo na biološkim aspektima, iako su oni primarnog značaja, jer
izražavaju ugroženost ESU kao osnovne - najniže jedinice biološke raznovrsnosti.
Ekonomski, socijalni i pravni aspekti su veoma značajni faktori u ovom procesu. U
slučaju postojanja izrazitih socio-ekonomskih implikacija, biološki aspekt sam po sebi
nije dovoljan za definisanje ESU. Koncept «Operational Conservation Units» (OCUs)
uveden je da opiše jedinicu konzervacije koja bi bila rezultat interakcije između bioloških
i socio-ekonomskih faktora (Dodson i sar., 1998). Biološki zahtevi su u velikoj meri
ustanovljeni unutar ESU. Prema tome, OCU predstavlja konstitutivni činilac ESU i
njegovu interakciju sa socio-ekonomskim pitanjima.
Privremena operativna konzervaciona jedinica može biti definisana u slučaju kada
je neophodno brzo izvršiti konzervacione aktivnosti, a još uvek ne postoji dovoljno
bioloških podataka za opisivanje preciznih evolutivnih odnosa između populacija.
4.3.4. K RAJNJA PREPORUKA U KONZERVACIJI POTOČ NE PASTRMKE
Potočna pastrmka predstavlja ekološki, ekonomski, estetski i naučno izuzetno
vrednu riblju vrstu čiji nezadovoljavajući konzervacioni status u evropskim zemljama
poziva na posvećivanje dodatne pažnje i preduzimanje konkretnih mera zaštite. Jedan
od glavnih razloga neadekvatne zaštite i konzervacije ove vrste u Evropi je generalna
neobaveštenost o aspektu biološkog diverziteta koji predstavlja genetička
varijabilnost unutar vrste. Zaštita isključivo vrste nije dovoljna, već se mora obratiti
pažnja na postojanje razlika na genetičkom nivou unutar i između lokalnih populacija,
koje takođe moraju biti konzervirane. Opšti cilj konzervacije biološkog diverziteta na
svim nivoima, od gena do ekosistema, ustanovljen je internacionalno i primenjuje se u
svim državama (među kojima je i SCG) koje imaju ratifikovanu Konvenciju o
Biološkom Diverzitetu (Rio de Janeiro 1992). Međutim, pomenute obaveze zaštite na
genetičkom nivou u najvećem broju zemalja još nisu transformisane u adekvatne
akcije. Prepoznavanje važnosti konzervacije genetičkog diverziteta je posebno važno
za vrste kao što je potočna pastrmka, koje se karakterišu velikom genetičkom
diferenciranošću lokalnih populacija. Gubitak lokalnih populacija potočne pastrmke
može rezultirati u iskorenjivanju relativno velikog dela genetičke varijabilnosti unutar
vrste (Laikre i sar., 1999).

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 108/141
Diskusija
100
4.3.5. K ONZERVACIJA DIVERZITETA HAPLOTIPOVA U SRBIJI I ZNAČAJ ZA RIBARSTVO
Problem konzervacije fonda potočne pastrmke veoma je slojevit i stoga veoma
složen, što se može zaključiti iz prethodno iznetih činjenica.
Tabela 15. Primeri glavnih aktuelnih pretnji po populacije potočne pastrmke u nekim
zemljama Evrope (Laikre i sar., 1999).
Država Degradacija staništa Ribolov Poribljavanje
BraneRegulacija
rekaZagađenje
Širenje
bolesti
i
parazita
Prekomeran
ribolovLovokrađa
Puštanje
domestifikovanih
i/ili ne
autohtonih
populacija
Danska X X X X
Estonija X X X X X
Finska X X X
Francuska X X X
Nemačka X X
Gr čka X X X X X X
Republika
Irska X X X X X
Norveška X X X X
Portugal X X X X X
Rusija X X X
Španija X X X X
Švedska X X X
Švajcarska X X X X
Velika
BritanijaX X X X X
U različitim zemljama Evrope, na osnovu molekularno-genetičkih studija,
ukazano je na postojanje različitog broja glavnih evolutivnih linija (Bernatchez, 1992),
kao i preostalih autohtonih populacija, što sugeriše da su pojedini prostori mogli sačuvati
veći deo preostalog genetičkog diverziteta vrste. U zemljama Evope konzervacija
genetičkog diverziteta potočne pastrmke veoma varira, pre svega iz razloga što su
evolutivni faktori rezultirali u različitoj količini i strukturi genetičkog diverziteta vrste u pojedinim zemljama.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 109/141
Diskusija
101
Ako iz tabele 15. uporedimo glavne ugrožavajuće faktore po evropske
populacije potočne pastrmke, kao i populacije u Srbiji (pogledati Uvod), najviše
sličnosti po vrsti i značaju ugrožavajućih aktivnosti naći ćemo između Gr čke i
Srbije. U Gr čkoj najozbiljniju pretnju po populacije potočne pastrmke ima preveliko
ribolovno opterećenje, koje uključuje i sve vidove krivolova (eksplozivna sredstva,
otrovi i različite klopke). Uništenje staništa je drugi veoma značajan razlog
ugroženosti, i obuhvata izgradnju brana, regulaciju tokova i izgradnju sistema
kanala, kao i zagađenje poljoprivrednim i kućnim otpadnim produktima. Na trećem
mestu nalazi se poribljavanje, mada, kao i u najvećem broju drugih zemalja, ne
postoji sistematična informacija o stepenu poribljavanja kao i o njegovim efektima
na autohtone populacije (Laikre i sar ., 1999). Redosled i značajnost ugrožavajućih
faktora na pastrmske populacije Gr čke u potpunosti koincidira sa trenutnim stanjem
ugroženosti populacija potočne pastrmke u Srbiji.
Kada su nam poznati ugrožavajući faktori koji predstavljaju potencijalnu,
ako ne i realnu opasnost da vremenom dođe do istrebljenja jedinstvenih pastrmskih
populacija na teritoriji Srbije, neophodno je izvršiti uklanjanje negativnih
(ugrožavajućih) aktivnosti, kao i administrativno upravljački odrediti konzervacionu
jedinicu na koju će biti usmeren konzervacioni napor. Ratifikacijom Konvencije o
Biološkom Diverzitetu naša zemlja se obavezala da će primenjivati konzervacione
mere na svim nivoima, od gena do ekosistema, što se u potpunosti slaže sa dve
glavne preporuke genetičke konzervacije:
1. sačuvati što je više moguće prirodnih ekosistema
2. utvrditi genetičku strukturu populacije
Rezultati ove teze započeli su identifikaciju genetičkog diverziteta pastrmskih populacija, predstavljenog sa 15 mtDNA haplotipova u okviru tri evolutivne linije
(Bernatchez i sar., 1992), uzorkovanjem na 35 lokaliteta u okviru sva tri slivna područ ja
Republike Srbije. Na osnovu dobijenih rezultata haplotip se može odrediti za
konzervacionu jedinicu na koju će biti usmeren konzervacioni napor.
Na osnovu saznanja dobijenih na terenu prilikom prikupljanja materijala može
se reći da je ugroženost populacija potočne pastrmke izuzetno velika, i da je
neophodna hitna primena konzervacionih mera u vidu dodeljivanja konzervacionihstatusa, kako staništu, tako i haplotipovima. Posebna pažnja mora biti posvećena

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 110/141
Diskusija
102
haplotipovima koji su pronađeni po prvi put u okviru areala vrste. Populacije
novopronađenih haplotipova dunavskog sliva (Da*Vl, Da*Vr i Da*Dž) izuzetno su
malobrojne. U izvorišnom delu reke Vlasine na uzorkovanih 100 m toka pronađene su
samo dve ribe i obe su posedovale haplotip Da*Vl, dok je donji deo reke ve ć
poribljavan neautohtonom pastrmkom. U gornjem delu reke Vrle pronađen je haplotip
Da*Vr koji već deli stanište sa introdukovanim haplotipom ADcs1, a samo stanište je
ugroženo izgradnjom fabrike vode na obali reke. Haplotip Da*Dž pronađen je na 1
primerku u Džepskoj reci analizom oko 100 m toka. Novi haplotip jadranskog sliva
Ad*Pe, kao i ostali koji su pronađeni u okviru istog slivnog područ ja (Ad+Prz i
ADcs11), kao i dela egejskog sliva Ad*Ti (Lepenac sa pritokama), trenutno nisu pod
našim uvidom zbog nemogućnosti pristupa pomenutom područ ju. Novopronađeni
haplotip Ad*Bož uzorkovan je u slivu Dragovištice i, u odnosu na ostale nove
haplotipove, njegova brojnost je na zavidnom nivou. Međutim, i on je ugrožen
poribljavanjem Dragovištice u donjem delu toka kroz Bugarsku materijalom At linije.
Posebno je bitno pomenuti da je na lokalietima Vlasina, Vrla i Božica prisutna
izrazita degradacija staništa, zbog formiranja akumulacionih jezera, dok se iz
Ljubatske reke značajan deo vode prevodi u sliv Južne Morave. Takođe, izuzetno
mala brojnost primećena je na ostalim uzorkovanim lokalitetima, izuzev lokaliteta gde
je uzorkovan haplotip Da1.
U Srbiji je do sada, na nacionalnom nivou, jedino populacijama potočne pastrmke
u slivu Dragovištice bio dodeljen konzervacioni status na bazi IUCN kriterijuma. Još pre
utvr đivanja novog mtDNA haplotipa (Ad*Bož), iz literaturnih podataka bilo je poznato
da egejski sliv jugo-istočne Srbije naseljava vrsta Salmo macedonicus (Karaman, 1924).
Ispitivanjima koja su obavili Marić i sar. (2004), utvr đena je i biomasa pastrmskih
populacija sliva Dragovištice koja je iznosila 8,8 kg km-1, što je bilo za oko četiri puta
manje u odnosu na biomasu populacija potočne pastrmke u reci Mlavi (istočna Srbija).Populacije potočne pastrmke u slivu Dragovištice zbog njihove taksonomske
specifičnosti, kao i veoma malog areala na teritoriji Srbije koji naseljavaju, uz utvr đenu
malu biomasu, dobile su operativni status Lower Risk (conservation dependent).
Trenutno, ugroženost potočne pastrmke pokušava se legislativno regulisati
određenim zaštitnim merama u ribarstvu, kao što su minimalna dozvoljena dužina ulova,
period lovostaja u vreme mresta, strogo propisan režim ribolova u pogledu mogućnosti i
količine ulovljenih pastrmki, broja ribolovnih dana tokom sezone, itd. Sve pobrojane

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 111/141
Diskusija
103
mere su veoma dobre, ali one do sada nisu dale pozitivne rezultate u zaštiti, pre svega
zbog njihovog nepotpunog sprovođenja.
Za potpunu zaštitu pastrmskih populacija neophodno je primeniti sledeće
preporuke:
1. Utvr đivanje stanja populacije
2. Utvr đivanje i otklanjanje ugrožavajućih faktora koji su doveli do smanjenja
brojnosti populacije
3. Utvr đivanje genetičke strukture populacije
4. U slučaju pronalaska novih genetičkih varijanti izvršiti dodelu konzervacionog
statusa populaciji i staništu
Na osnovu dobijenih rezultata svi lokaliteti na kojima su pronađeni novi
haplotipovi, kao i lokaliteti na kojima su pronađeni vrlo retki haplotipovi (Tabela
9), trebalo bi da dobiju poseban konzervacioni status kao stanište (određenu
kategoriju i stepen zaštite), uz dodelu konzervacionog statusa na nacionalnom
nivou haplotipovima koji su na datim lokalitetima pronađeni. Iako potočna
pastrmka nije uvrštena na Crvenu Listu (IUCN) ugroženih životinja na
internacionalnom nivou, pojedine države su joj dodelile status ugroženih vrsta na
nacionalnom nivou (Gr čka - Endangered, Španija – Vulnerable i Švedska - Lower
risk, near threatened). Pošto su ugrožavajući faktori u Gr čkoj i kod nas vrlo slični,
logično bi bilo da i mi sledimo primer Gr čke i da bar našim jedinstvenim
populacijama (teritorija uže Srbije) dodelimo istu kategoriju ugroženosti. Pri
dodeljivanju nacionalnog konzervacionog statusa moramo uzeti u obzir i ribolovnu
atraktivnost pomenutih voda, kao i njihov potencijalni socio-ekonomski značaj.
Vodama koje nisu posebno ribolovno atraktivne u izvorišnim delovima (Vrla,Vlasina i Džepska), a koje predstavljaju staništa jedinstvenih genetičkih formi
(Da*Vr, Da*Vl i Da*Dž), treba kao staništu dodeliti I stepen zaštite (od izvorišta u
dužini od 4 km), a haplotipovima status ugroženosti (Endangered). Reke u slivu
Dragovištice u kojima je pronađen haplotip Ad*Bož (Lisina, Božica, Ljubata i
Brankovačka reka (potencijalno)) koje su ribolovno atraktivne, a samim tim mogu
imati i ekonomski značaj, moraju dobiti specijalni režim zaštite koji bi omogućio
očuvanje genetičke jedinstvenosti pastrmskih populacija, kao i održivi ribolov kao jedan od izvora prihoda lokalnoj zajednici. Pastrmske populacije koje naseljavaju

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 112/141
Diskusija
104
pomenute reke sliva Dragovištice dobile bi konzervacioni status Lower risk, near
threatened. Izvorišni delovi navedenih reka u dužini od 4 km morali bi dobiti kao
stanište I stepen zaštite, dok bi ostali deo toka pomenutih reka do ušća u
Dragovišticu, kao i sama Dragovištica, bio pod II stepenom zaštite. Dodela
navedenog konzervacionog statusa uz primenu strogih mera kontrole i upravljanja
(lov po pravilu “uhvati-i-pusti” uz mogućnost nošenja jedne ribe preko 40 cm)
obezbedio bi stalnost korišćenja, posebno ukoliko bi se adekvatnim poribljavanjem
kroz neki period restaurirala genetička struktura populacije približna originalnoj.
Perspektive haplotipa Ad*Bož u pogledu opstanka biće vrlo optimistične, naročito
ako se obistine sumnje da je areal ovog haplotipa vezan i za ostale reke egejskog
sliva, što će pokazati rezultati budućih istraživanja (Diskusija str. 79).
Rekreativni («sportski») ribolov, koji se u velikom delu areala pastrmke
smatra najelitnijim vidom rekreativnog ribolova, upražnjava se intenzivno. Intenzitet
ribolova (a kod nas se pod ovim mora obuhvatiti i krivolov) je veliki, jer je cena
takvog ribolova znatna i donosi upravljačima ili vlasnicima ribolovnih voda na
kojima se vrši velike prihode u svetu, a ponegde i kod nas. Ovaj ribarstveno –
komercijalni pritisak je pod uslovima kada nije bilo moguće zaokruženo odrediti i
sprovoditi adekvatne konzervacione mere u većem delu areala pastrmke, a posebno
kod nas i u bližem okruženju, snažan ugrožavajući faktor po pastrmski (geno)fond.
Kao posledica velikog ribolovnog pritiska, najčešće se pribegava poribljavanju kao
meri upravljanja pastrmskim fondom, što je uglavnom kontraproduktivno po
originalnost pastrmskih populacija. Kod nas se poribljavanje najčešće vršilo i vrši
onim materijalom koji je u datom trenutku dostupan, bez obzira na činjenicu koliko
je odgovarajući, bez skoro bilo kakvih ograničenja i kontrole. Za područ je stare
SFRJ (izuzev Slovenije), a posebno za Srbiju, ne postoji precizno vođena evidencija
o poribljavanju pastrmskih reka, osim u nekoliko pojedinačnih slučajeva. Zna se da je materijal za poribljavanje nabavljan iz mrestilišta u Ivanjici, Konjicu, Vrelu Bune
i Blagaju, kao i iz Slovenije i sa Ohrida, i da su poribljavanja vršena širom Srbije.
Srećom po pastrmski fond kod nas, ta poribljavanja su uglavnom vršena na
ribolovno atraktivnim vodama: Mlavi, Krupaji, Gradcu, Moravici, Drini i ponegde
drugde, ali su još preostale vode (koje su bile u fokusu uzorkovanja ove teze) koje
su i / ili ribolovno neatraktivne, i / ili fizičkim preprekama odvojene od delova,
obično nizvodnih, koji su ribolovno atraktivni svojom lakom dostupnošću iliveličinom pastrmke u njima (Marić, 2002). Prepoznavanje genetičkog identiteta

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 113/141
Diskusija
105
pastrmki omogućava uspostavljanje sistema konzervacije uključenog u mere
održivog ribolovnog korišćenja pastrmskog fonda i po obimu te aktivnosti, i po
dubini implementacije. Ovakav vid ribarstvenog korišćenja pastrmskog fonda biće
od značaja tek kada budemo imali s jedne strane potrebne parametre (stanje
populacije, ribolovno opterećenje, genetičku strukturu), a sa druge uređen sistem
proizvodnje autohtonog materijala za poribljavanje, u slučajevima kada je ova mera
neophodna. Da bi se proizvodnja autohtonog materijala za poribljavanje mogla
realizovati, za početak je neophodno ustanoviti referentnu laboratoriju koja bi se
bavila utvr đivanjem genetičke strukture populacija, na osnovu koje bi se za
specifične i ugrožene populacije formiralo matično jato koje bi služilo za dobijanje
materijala za poribljavanje. Pošto je formiranje referentne laboratorije neophodan
uslov za proveru genetičke strukture populacije, a samim tim i za konzervaciju
retkih i ugroženih (autohtonih) populacija, cela procedura morala bi biti i zakonski
regulisana. Ako se u ovom smislu u dogledno vreme ne promeni zakonska
regulativa, i ne omogući utvr đivanje genetičke strukture populacija, najbolje bi bilo
ne vršiti poribljavanja genetički neproverenim materijalom, već posebnu pažnju
obratiti na zaštitu ugroženih populacija uz strogu zakonsku primenu zaštitnih mera u
ribarstvu, ili pak, ako je neophodno, zabraniti izlov, pa u krajnjem slučaju i ribolov
u periodu neophodnom za oporavak populacije.
Kako oba aspekta konzervacionih i ribarstvenih aktivnosti zaslužuju najveću
pažnju, u sadašnjim uslovima, u kojima se ribarstvo kao privredna delatnost kod
nas nalazi (a u okviru nje i rekreativni ribolov), zahtevaju korenite promene i u
zakonskom smislu i u pogledu dosledne implementacije u budućoj ribarstvenoj i
konzervacionoj praksi. Sa zakonske strane, potreba za harmonizacijom naših
ribolovnih propisa i konzervacione prakse sa istima u zemljama EU doprineće
uvođenju pravila primerenijih vremenu i stanju u oblasti koja bi se time regulisala,dok bi, sa ribarstvene strane, uređenje celog sistema višestruko povećalo
produktivnost pastrmskog ribolova i omogućilo upravljačima pastrmskih ribolovnih
voda obavljanje delatnosti koja bi po profitabilnosti bila približna onoj u zemljama
u kojima je to odgovarajuće regulisano. Time bi se jasno pomogli interesi nadležnih
državnih organa iz oblasti zaštite životne sredine i ribarstva s jedne strane (kao
propisivača, kontrolora i implementatora zakona) i najrazličitijeg opsega
ribarstvenih subjekata kao upravljača pastrmskih ribolovnih voda, što otvara i putka odgovoru na pitanje subjekata koji bi trebalo da se oko ove teme (i drugih, šire

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 114/141
Diskusija
106
posmatrano) angažuju. Pri tome, uzimajući u obzir broj pastrmskih voda i ribolovni
prtisak, sasvim je izvesno da bi se konzervacione aktivnosti vezane za pastrmski
fond u Srbiji odvijale niskim intenzitetom, ali obavezno konstantno, što bi trebalo
da bude finansijski održiva aktivnost u sklopu ribarstveno-finansijskog obima, dok
bi ribarstvene aktivnosti, pre svega one vezane za zaštitu ribljeg fonda u ribarstvu,
trebalo da budu višestruko intenzivirane u praksi. Ovo je pre svega značajno stoga
sto bi pozitivne efekte ovako uređene održive ribarstvene i konzervacione prakse
osetili stanovnici ruralnih, trenutno slabo razvijenih i raznovrsnošću delatnosti
ograničenih područ ja Srbije i što bi, u sklopu ostalih vidova konzervaciono zavisnih
aktivnosti, to moglo predstavljati zamajac njihovom održivom privrednom razvoju
u celini.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 115/141
Zaključci
107
5. ZAKLJUČCI
Na osnovu rezul tat a ove stud ije utvr đeno je postojanje tri glavne
filogeografske grupe potočne pastrmke na teritoriji Srbije: dunavske (Da),
«južne» (Ad; Ad+) i atlantske (At). U okviru sve tri grupe identifikovano je,
na osnovu 18 polimorfnih mesta, ukupno 15 haplotipova, od kojih je šest po
prvi put op isano untar areala vrste. Od šest novo opisanih haplot ipova, tr i su
iz dunavske linije i pronađena su u slivu Južne Morave (Da*Vl (Vlasina),
Da*Vr (Vrla) i Da*Dž (Džepska reka)), dok su dva od preostala tri koja
pr ipadaju Ad lin iji, pronađena u egejskom slivu (Ad*Bož (sliv Dragovištice) i
Ad*Ti (sliv Lepenca), a jedan je pronađen u jadranskom slivu Ad*Pe (Pećka
Bistrica).
Rekonstrukcijom filogenije NJ i MP metodama dobijena je vrlo slična
topologija stabla, pri kojoj haplotipovi pokazuju pripadnost filogenetskim
linijama potočne pastrmke. «Majority role consensus 50% tree» otkriva jasno
razdvajanje sa visokom potporom na dve glavne klade (Da i At klada + «južna
klada» (Ad, Ad+, Ma i Me)). U «južnoj» kladi haplotip Ad*Bož zauzima
ancestralnu poziciju (Slika 30), ali zbog niskih “bootstraping” verovatnoća za
NJ i MP analize njegovu ancest ralnos t tek treba dokaza ti rezul tat ima drug ih
DNA analiza, ali i analiziranjem drugih tipova kararaktera (morfoloških). U
«južnoj» kladi, pored Ad haplotipova (ADcs1, ADcs11, Ad*Pe i Ad*Ti) koji
nisu jasno grupisani u kladu, javljaju se dve već poznate podgrupe
(mediteranska i marmoratus) i treća, dopunska, koja je prvi put identifikovana
(Ad+). Članovi Me, Ma i Ad+ klade karakterišu se istim brojem intra- kladnih
sinapomorfizama (pozicije 113 i 262 za Ma, 146 i 196 za Me, 126 i 262 za
Ad+; Tabela 13) što sugeriše isti nivo divergencije ovih klada. Novu (Ad+)
kladu čine haplotipovi Ad+Prz, Ad+N i Ad+RC, i ona je statistički poduprta
istim nivoom verovatnoće kao i već ustanovljene podgrupe (klade Me i Ma).
Haplotip Ad+Prz pronađen je u Prizrenskoj Bistrici, Mrtvici i Tripušnici (u
koju je, kako pretpostavljamo, introdukovan iz Neretve), Ad+N je pronađen u
Nere tv i, a Ad+RC u Ri jeci Crno jevića. Geografska distribucija pomenutih
haplotipova vezana je za reke jadranskog sliva jugo-zapadnog Balkana i

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 116/141
Zaključci
108
poklapa se sa di str ibucijom vrste Salmo farioides (Karaman, 1937), što i
navodi na zaključak da Ad+ klada ustvari predstavlja Salmo farioides.
Unutar Da klade uočen je veliki nivo genetičke divergencije (Tabela
11). Ovaj podatak, kao i sinapomorfne pozicije (Tabela 13) najbazalnijih
haplotipova Da klade (Da*Vr i Da*Dž) (Slika 30) i haplotipova iz svih ostalih
linija, ide u prilog stavu da dunavske populacije predstavljaju najstariju
fragmentiranu liniju Salmo trutta kompleksa. Ancestralna pozicija Da linije
pr ikazana je i sekvencioni ranjem celog kontrolnog regiona mtDNA (Cor tey i
sar., 2004; Templeton, 2004). Ovakvi rezultati navode na zaključak da je
potočna pastrmka, najverovatnije, poreklom iz Paratetisa, odakle se, preko
pleis tocenskih veza , širi la iz basena Crnog mora, Kaspi jskog i Aralskog jezera
prema At lant iku na jednoj i Medi teranu na drugoj st ran i. Pr ilog ovoj hipotezi
preds tavl ja pronalazak ekstremno divergentnih haplot ipova kao što su Da*Vr i
Da*Dž, koji zauzimaju intermedijarni položaj na mreži (Slika 31) između Da,
At i «južne grupe» (Ad, Ad+, Ma i Me) haplotipova, što je posebno
karakteristično za haplotip Da*Vr, čija se centralna pozicija može uočiti i
preko vrednost i genetičkih distanci između pomenutih grupa. Sve navedeno
dodatno potvr đuje dugo postojanje i ancestralno poreklo Da linije u evoluciji
potočne pastrmke.
Na analizi ranom područ ju, u zav isnos ti od sliva, bi lo je očekivano
pr isus tvo autoh tonih haplot ipova dunavske i Ad lini je. Međutim, analizom je
utvr đeno i prisustvo haplotipa At1 koji pripada atlantskoj liniji. Nalaz
pomenutog haplot ipa na dva lokalit eta , Gradac i Brankovačka reka, posledica
je poribl javanja, jer se radi o haplot ipu atlant ske lin ije ko ji preovladava u
ribnjačkoj proizvodnji mlađi potočne pastrmke. Takođe, bitno je pomenuti i
nalaz haplotipa Ad linije ADcs1 na dva lokaliteta unutar dunavskog sliva
(Jerma i Vrla), za koga smatramo da je na ove lokalitete dospeo antropogenom
aktivnošću iz sliva Dragovištice (Jerma – poribljavanje, Vrla – ispumpavanje
vode u sliv Južne Morave). Između egejskog i jadranskog sliva utvr đena je
ja sna gene tička raspodela haplotipova, koju jedino narušava haplotip Ad+Prz
pronađen u Tripušnici (egejski sliv), koji je u ovaj sliv dospeo kao posledica
poribl javanja mate rij alom iz sl iva Neretve .

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 117/141
Zaključci
109
Visok nivo genetičkog diverziteta koji je otkriven opisivanjem nekoliko
novih polimorfizama unutar različitih linija potočne pastrmke, izrazita
divergencija unutar dunavskih populacija na teritoriji Srbije, kao i
pretpostavka postojanja evo lu tivno zasebne lin ije (Ad+; Salmo farioides) u
jadranskom slivu, suger išu da bi u na rednom periodu tr ebalo preduzet i
opsežne analize pastrmskih populacija na preostalom delu Balkana (Crna
Gora, Makedonija, Albanija, Bosna i Hercegovina, Bugarska). Pošto
analizirani prostor Srbije predstavlja samo mali deo Balkanskog poluostrva, za
očekivati je da je genetički polimorfizam potočne pastrmke unutar celog
regiona znatno veći.
Ova studija je pokazala da autohtone populacije još uvek postoje u
Srbiji, barem u izvorišnim delovima reka. Ova informacija je od posebne
važnosti za pravilno gazdovanje u cilju održanja autohtonog genetičkog
diverziteta potočne pastrmke u Srbiji. Gazdovanje mora prepoznati biološku
realnost postojanja genetičkog diverziteta i pronaći odgovarajuće strategije na
bazi ove realnosti za efikasnu konzervaciju genetičke varijabilnosti (Ryman,
1991).
Pošto su novo pronađeni haplotipovi na teritoriji Srbije malobrojni i
pod st alnom su pretn jom ugrožava jućih aktivnosti, neophodno im je, na
nacionalnom nivou, dodeliti odgovarajuće konzervacione statuse.
Novopronađenim haplotipovima (Da*Vr, Da*Vl i Da*Dž) u vodama koje nisu
ribolovno atraktivne, u izvorišnim delovima (Vrla, Vlasina, Džepska),
predložena je dodela st atusa ugroženost i (Endangered), dok on ima koji se
nalaze u ribolovno atraktivnim vodama (reke sliva Dragovištice) u kojima je
pronađen haplotip Ad*Bož predlaže se dodela konzervacionog statusa Lower
risk, near threatened, koji bi trebao da zadovolji očuvanje genetičke
jedins tvenost i, sa jedne str ane , a sa druge održ ivi ribolov. Takođe, neophodno
je dode lit i stani št ima na kojima su pronađeni novi haplotipovi, a to su gornji
tokovi reka (Vrla, Vlasina, Džepska i reke sliva Dragovištice), odgovaraju ću
kategoriju i I stepen zaštite u prva 4 km toka.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 118/141
Zaključci
110
Oba aspekta konzervacionih i ribarstvenih aktivnosti na populacijama
potočne pastrmke zahtevaju vrlo brze i korenite promene, kako u zakonskom
smislu, tako i u doslednoj primeni u budućoj konzervacionoj i ribarstvenoj
praks i.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 119/141
Summary
111
6. SUMMARY
Upon results of this study, the occurrence of three main phylogeographic groups of the brown
trout was confirmed at the territory of Serbia: Danube (Da), “southern” (Ad; Ad+) and Atlantic (At).
Within all groups, 15 haplotypes based on 18 polymorphic places were identified; six of them were
recorded for the first time. Three new haplotypes belong to Danube group and were found in Južna Morava
drainage (Da*Vl (Vlasina), Da*Vr (Vrla) i Da*Dž (Džepska River)), while within three others, belonging
to Ad group, two were found in Aegean Sea basin (Ad*Bož (Dragovištica River drainage) and Ad*Ti
(Lepenac River drainage); one was found in Adriatic Sea basin Ad*Pe (Pećka Bistrica River).
Phylogeny reconstruction based on NJ and MP showed very similar tree topology. «50%
Majority role consensus tree» revealed clear division of three main clades with high support level (Da and
At clade + «southern clade» (Ad, Ad+, Ma and Me). Haplotype Ad*Bož have the ancestral position in the
«southern» clade (Fig 30), but because of the low “bootstrapping” probabilities for NJ i MP analyses, this
ancestral state should be additionaly proved with other DNA analyses and analyses of morphologic
characters. Within «southern» clade, in addition to Ad haplotypes (ADcs1, ADcs11, Ad*Pe i Ad*Ti) that
were not apparently grouped together, the presence of two well known subdivisions was confirmed
(Mediterranean and marmoratus); the third additional subdivision Ad+ was identified for the first time. The
members of the Me, Ma i Ad+ clades are characterized with the same numbers of intra-clade
synapomorphies (positions 113 and 262 for Ma, 146 and 196 for Me, 126 and 262 for Ad+; Table 13); this
suggest the same level of divergence of these clades. The new clade (Ad+) includes haplotypes Ad+Prz,
Ad+N and Ad+RC, and has the same level of support as other (previously known) subdivisions (clades Me
and Ma). Haplotype Ad+Prz was found in Prizrenska Bistrica River, Mrtvica River and Tripušnica River
(we think that this haplotype was introduced from the Neretva River into Tripušnica River); Ad+N was
found in Neretva River and Ad+RC in Rijeka Crnojevića River. Geographic distribution of those
haplotypes is connected with the rivers of the Adriatic basin of the south-western Balkan peninsula and
corresponds with the distribution of Salmo farioides (Karaman, 1937); this suggests that Ad+ clade
represents Salmo farioides, and could belong to the new line within the “southern” group.
Strong genetic divergence was found within the Da clade, that was based on the genetic distances
(Table 11). Strong divergence, synapomorphic positions (Table 13) of the most basal haplotypes belonging
to Da clade (Da*Vr and Da*Dž) (Fig 30), as well as position of the haplotypes of all other lineages, suggest
that Danube populations represent the oldest fragmental line of Salmo trutta complex. Ancestral position of
Da line was also shown by sequencing of the whole mtDNA control region (Cortey at al., 2004;Templeton, 2004). These results suggest that brown trout originated from the Parathetis and spred from the

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 120/141
Summary
112
Paratethis through the Pleistocene connections from the Black See basin, Caspian and Aral lakes toward the
Atlantic Ocean and Mediterranean See. Extremely divergent haplotypes like Da*Vr and Da*Dž have
intermediate positions in the network (Fig 31) between Da, At and «southern group» (Ad, Ad+, Ma i Me)
of haplotypes. That specially accounts for the Da*Vr haplotype, that holds central position judging from its
genetic distances to the rest of haplotypes. All these facts confirm the long term presence and ancestral
origin of Da lineage in evolution of the brown trout.
In spite of expectation that only wild haplotypes of Ad and Da lineages occur in Serbia, the
analyses confirmed presence of haplotype At1 which belongs to Atlantic line. The finding of this haplotype
in two localities, Gradac River and Brankovačka River, is a fisheries-related impact that predomintes in
hatcheries’ production of brown trout fry for stocking. The occurrence of haplotype ADcs1 of the Ad
lineage in two localities in the Danube River basin (Jerma and Vrla Rivers) could be a consequence of
sotcking and transfer of water from the Dragovistica River drainage, respectively. The clear genetic
dispersion of haplotypes was recorded between basins of Aegean and Adriatic Seas. It was disturbed only
by Ad+Prz haplotype, which occured in Tripusnica River (Aegean Sea basin) after the stocking from the
Neretva River drainage.
Since the territory of Serbia represents only the small part of the Balkan Peninsula, the higher lever
of genetic polymorphism of the brown trout from the whole region of the Balkans (Montenegro,
Macedonia, Albania, Bosnia and Herzegovina, Bulgaria) could be expected, considering the recorded high
level of genetic diversity, finding of new polymorphic places within different lines of the brown trout,
strong divergence within Danube populations in the territory of Serbia, as well as suspected presence of
evolutionary divergent line (Ad+; Salmo farioides) in the Adriatic basin.
This study showed that aboriginal, i.e., populations still exist in Serbia, at least in spring sections of
streams and rivers. This is very important for the conservation of autochthonous genetic diversity of the
brown trout in Serbia. The efficient conservation of existing genetic variability, i.e., diversity, is to be taken
into account in the strategy of fisheries management (Ryman, 1991).
Since the new haplotypes are not very frequent in Serbia and are under the constant threat, proper
conservation status in national legislation is needed. The conservation status of three new haplotypes from
Da group (Da*Vr, Da*Vl and Da*Dž) in the headwaters of Vrla, Vlasina and Džepska Rivers should be
set as EN, whereas for AD*Bož haplotype it should be set as LRnt in particular waters of the Dragovistica
River drainage where some brown trout, attractive for fishing, occur. That would enable both maintenance
of existing genetic uniqueness and sustainable fisheries utilization. However, the first four kilometers of
each of those rivers should be assigned in the highest protection category. Both conservational and fisheries
practice concerning the brown trout are in strong and urgent necessity for changes in legislation, as well as
in consistent and harmonized application.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 121/141
Literatura
113
7. LITERATURA
Aebersold, P. B., Winans, G. A., Teel, D. J., Milner, G. B., Utter, F. M. 1987. Manual forstarch gel electrophoresis: a method for the detection of genetic variation. NOAATechnical Report NMFS 61, U.S. Department of Commerce, National Oceanicand Atmospheric Administration, National Marine Fisheries Service, U.S.A.
Aganović, M. 1979. Salmonidne vrste riba i njihov izgoj. IGKRO "Svijetlost", Sarajevo.
Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K., Watson, J. D. 1994. DNA repair.V: Molecular biology of the cell. New York, Gerland Publishing, pp. 242-251.
Alegria Hernandez V. 1985. A contribution to the study of heterogeneity of the Adriatic
sardine (Sardina pilchardus Walb.) population. Acta Adriatica, 26: 109-122.
Allendorf, F. W. 1988. Conservation biology of fishes. Conservation Biology, 2: 145-148.
Allendorf, F. W., Leary, R. F. 1988. Conservation and distribution of genetic variation ina polytypic species, the cutthroat trout. Conservation Biology, 2: 170-184.
Allendorf, F. W., Ryman, N., Stennek, A., Stähl, G. 1976. Genetic variation inScandinavian brown trout (Salmo trutta L.): evidence of distinct sympatric
populations. Hereditas, 83: 73-82.
Allendorf, F. W., Waples, R. S. 1996. Conservation and Genetics of Salmonid Fishes. In:Avise, J. C. & Hamrick, J. L. (eds.) Conservation Genetics. Case Stories from
Nature, Chapman & Hall, New York, pp. 238-280.
Amos, B., Schlöterer, C., Tautz, D. 1993. Social structure of pilot whales revealed byanalytical DNA profiling. Science, 260: 670-672.
Anderson, B. G., Borns, H. W. Jr. 1994. The Ice Age world. Scandinavian Univ. Press,Oslo, Norway.
Anđelković, S. J. 1989. Tercijarne ribe Jugoslavije stratigrafsko – paleontološko
paleoekološka studija. Paleontologica Jugoslavica. Jugoslovenska AkademijaZnanosti i Umjetnosti Zagreb, 38: 1-121.
Angeres, B., Bernatchez, L., Angers, A., Desgroseillers, L. 1995. Specific microsatelliteloci for brook charr reveal strong population subdivision on a micrographic scale.Journal of Fish Biology, 47 (Suppl. A): 177-185.
Anonimni autori. 1994. «Zakon o ribarstvu». Službeni glasnik Republike Srbije, 35, pp.2342-2346.
Anonimni autori. 2004. «Zakon o zaštiti životne sredine». Službeni glasnik RepublikeSrbije, 135, pp. 29-43.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 122/141
Literatura
114
Antunes, A., Alexandrino, P., Ferrand, N. 1999. Genetic characterization of Portuguese brown trout (Salmo trutta L.) and comparison with other European populations.Ecology of Freshwater Fish, 8: 194-200.
Antunes, A., Templeton, R. A., Guyomard, R., Alexandrino, P. 2002. The role of Nuclear
Genes in Intraspecific Evolutionary Inference: Geneology of the transferin Genein the Brown Trout. Molecular Biology and Evolution, 19 (8): 1272-1287.
Apostolidis, A., Karakousis, Y., Triantaphyllidis, C. 1996. Genetic divergence and phylogenetic relationships among Salmo trutta L. (brown trout) populations fromGreece and other European countries. Heredity, 76: 551-560.
Apostolidis, A., Karakousis, Y., Triantaphyllidis, C. 1996a. Genetic differentiation and phylogenetic relationships among Greek Salmo trutta L. (brown trout) populationsas revealed by RFLP analysis of PCR amplified mitochondrial DNA segments.Heredity, 77: 608-618.
Apostolidis, A. P., Triantaphyllidis, C., Kouvatsi, A., Economidis, P.S. 1997.Mitochondrial DNA sequence variation and phylogeography among Salmo trutta L (Greek brown trout) populations. Molecular Ecology, 6:531-542.
Aquadro, C. F., Greenberg, B. D. 1983. Human mitochondrial DNA variation andevolution: analysis of nucleotide sequences from seven individuals. Genetics, 103:287-312.
Arkhipov, S. A., Ehlers, J., Johnson R. G., Wright H. E. Jr. 1995. Glacial drainagestowards the Mediterranean during middle and late Pleistocene. Boreas, 24: 196-206.
Avise, J. C. 1992. Molecular population structure and the biogeographic history of aregional fauna: a case history with lessons for conservation biology. Oikos, 63:62-76.
Avise, J. C. 1994. Molecular tools. V: Molecular markers, natural history and evolution.Chapman & Hall, New York, pp. 44 - 91.
Avise, J. C. 1998. The history of phylogeography: a personal reflection. Molecular
Ecology, 7: 371-379.
Avise, J. C. 2000. Phylogeography. The history and Formation of Species. HarvardUniversity Press, Cambridge, MA.
Avise, J. C., Arnold, J., Ball, R. M., Bermingham, E., Lamb, T., Neigel J. E. Reeb, C. A.Saunders, R. C. 1987. Intraspecific phylogeography: The mitochondrial DNA and
bridge between population genetics and systematics. Annual Review of Ecologyand Systematics, 18: 489-522.
Balon, K. E. 1968. Notes to origin and evolution of trouts and salmons with special
reference to the Danubian trouts. Acta Societatis Zoologicae Bohemoslovacae, 32(1): 1-21.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 123/141
Literatura
115
Banarescu, P. 1960. Einige Fragen zur Herkunft und Verbreitung derSüsswasserfishfauna der europäisch – mediterranen Unterregion. Archiv fuerHydrobiologie, 57: 16-134.
Banarescu, P. 1973. Origin and affinities of the freshwater fish fauna of Europe.
Ichthyologia, Beograd 5: 1-8.
Banarescu, P. 1977. Position zoogéographique de l'ichthyo – faune d'eau douce d'Asieoccidentale. Cybium, 2: 35-55.
Banarescu, P. 1990. Zoogeography of freshwater: general distribution and dispersal offreshwater animals. Aula Verlag ed. Wiesbaden, 1: 1-511.
Banarescu, P., Blanc, M., Gaudet, J. L., Mureau, J. C. 1971. European inland water fish,multilingual catalogue. Fish News Books, London, pp. 170.
Behnke, R. J. 1965. A systematic study of the family Salmonidae with special referenceto the genus Salmo. Ph.D. thesis, University of Berkley, California, U.S.A., pp.273.
Behnke, R. J. 1968. A new subgenus and species of trout, Salmo (Platysalmo) platycephalus from south central Turkey, with comments on the classification ofthe subfamily Salmonidae. Mitteilungen Hamburgisches Zoologisches Museumund Institut, 66: 1-15.
Behnke, R. J. 1986. Brown trout. Trout, 27: 42-47.
Berg, L. S. 1948. Freshwater fishes of the USSR and adjacent countries. ZoologicalInstitute Akademy Nauk Moscow USSR 1(27), volume 1. In Russian, Englishtranslation, 1962: Office of Technical Services, Department of Commerce,Washington, DC.
Berg, W. J., Ferris, S. D. 1984. Restriction endonuclease analysis of salmonidmitochondrial DNA. Canadian Journal of Fisheries and Aquatic Sciences, 41:1041-1047.
Bernatchez, L. 1995. A role for molecular systematics in defining significant evolutionary
units. In Nielsen, J.L. and Powers, D.A. (eds.). Evolution and the aquaticecosystem: defining unique units in population conservation. American FisheriesSociety Symposium 17, American Fisheries Society, Bethesda, Maryland, USA,
pp. 114-132.
Bernatchez, L. 2001. The evolutionary history of brown trout (Salmo trutta L.) interredfrom phylogenetic, nested clade, and mismatch analyses of mitochondrial DNAvariation. Evolution, 55 (2): 351-379.
Bernatchez, L., Danzmann, R. G. 1993. Congrugence in control - region sequence andrestriction – site variation in mitochondrial DNA of brook charr (Salvelinus
fontinalis Mitchill). Molecular Biology and Evolution, 10: 1002-1014.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 124/141
Literatura
116
Bernatchez, L., Guyomard, R., Bonhomme, F. 1992. DNA sequence variation of themitochondrial control region among geographically and morphologically remoteEuropean brown trout Salmo trutta populations. Molecular Ecology, 1: 161-173.
Bernatchez, L., Osinov, A. G. 1995. Genetic diversity of trout (genus Salmo) from its
most eastern native range based on mitochondrial DNA and nuclear genevariation. Molecular Ecology, 4: 285-297.
Bernatchez, L., Wilson, C. C. 1998. Comparative phylogeography of Nearctic andPalearctic fishes. Molecular Ecology, 7: 431-452.
Berrebi, P., Povž, M., Jesenšek, D., Cattaneo – Berrebi, G., Crivelli, A. J. 2000. Thegenetic diversity of native, stocked, and hybrid populations of marble trout inSoča river, Slovenia. Heredity, 85 (3): 277-287.
Bianco, P. G. 1990. Potential role of paleohistory of the Mediterranean and Parathetys
basins on the early dispersal of Euro-Mediterranean freshwater fishes.Ichthyological Exploration of Freshwaters, 1: 167-184.
Billington, B., Herbert, P. D. N. 1991. Mitochondrial DNA diversity in fishes and itsimplications for introductions. Canadian Journal of Fisheries and AquaticSciences, 48: 80-94.
Birky, C. W. 1983. Relaxed cellular controls and organelle heredity. Science, 222: 468-475.
Bookstein, F. L., Chernoff, B. L., Humphries, J. M., Smith, G. R., Strauss, E. R. 1985.Morphometrics in evolutionary biology. The Academy of Natural Sciences ofPhiladelphia, Special Publication, 15 pp. 277.
Borst, P., Grivell, L. A. 1981. Small is beautiful-porterait of a mitochondrial genome. Nature, 290: 443-444.
Bouza, C., Arias, J., Castro, J., Sánchez, L., Martínez, P. 1999. Genetic structure of brown trout, Salmo trutta L., at the southern limit of the distribution range of theanadromous form. Molecular Ecology, 8: 1991-2002.
Brown, W. M., George, M., Wilson, A. C. 1979. Rapid evolution of animal mitochondrialDNA. Proceedings of the National Academy of Sciences USA, 76: 1967-1971.
Brown, J. H., Gibson, A. C. 1983. Biogeography. Mosby, St Louis, Toronto & London,643 pp.
Brown, W. M., Prager, E. M., Wang, A., Wilson, A. C. 1982. Mitochondrial DNAsequences of primates: tempo and mode of evolution. Journal of MolecularEvolution, 18: 225-239.
Bruford, M. W., Ciofi, C., Funk, S. M. 1998. Characteristics of Microsatellites. U:
Molecular Tools for Screening Biodiversity. Chapman & Hall, New York, pp.202-205.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 125/141
Literatura
117
Brunner, P. C., Douglas, M. R., Bernatchez, L. 1998. Microsatellite and mitochondrialDNA assessment of population structure and stocking effects in Arctic charrSalvelinus alpinus (Teleostei: Salmonidae) from central Alpine lakes. MolecularEcology, 7: 209-223.
Cables, L. 2001. Mitochondria. Horsepower.http://homepages.ihug.co.nz/~lcables/mitochondria.htm
Cann, R. L., Brown, W. M., Wilson, A. C. 1984. Polymorphic sites and the mechanismsof evolution in human mitochondrial DNA. Genetics, 106: 479-499.
Chiereghini, A. S. 1818. Descrizoine de’testacei e de’pesci she abitano le lagune e golfoVento: pp 128.
Cita, M. B., McKenzie, J. A. 1986. The terminal Miocene event. International GeologicalCorrelation Programme, Messinian Project, 96: 123-143.
Clement, M. Posada, D. Crandall, K. 2000. TCS: A computer program to estimate genegenealogies. Molecular Ecology, 9(10): 1657-1660
Constable, J. J., Packer, C., Collins, D. A., Pusey, A. E. 1995. Nuclear DNA from primatedung. Nature, pp. 373-393.
Cortey M., García-Marín J. L. 2002. Evidence for phylogeographically informativeseqence variation in the mitochondral control region of Atlantic brown trout.Journal of Fish Biology, 60: 1058-1063.
Cortey, M., Pla, C., García-Marín, J. L. 2004. Historical biogeography of Mediterraneantrout. Molecular Fhylogenetics and Evolution, 33: 831-844.
Cracraft, J. 1987. Species concept and the analogy of evolution. Biology and Phylosophy,3: 329-346.
Crivelli, A. J. 1996. The freshwater fish endemic to the northern Mediterranean region.Le Sambuc, Arles, France.
Cross, T. F., Mills, C. P. R., Courcy Williams, M. 1992. An intensive study of allozyme
variation in freshwater resident and anadromous trout, Salmo trutta L., in WesternIreland. Journal of Fish Biology, 40: 25-32.
Crozier, W. W., Ferguson, A. 1986. Electrophoretic examination of the populationstructure of brown trout, Salmo trutta L., from the Lough Neagh catchment,
Northern Irland. Journal of Fish Biology, 28: 459-477.
Curole J. P., Kocher, T. D. 1999. Mitogenomics: digging deeper with completemitochondrial genomes. TREE, 14: 394-398.
Delling, B. 2003. Species diversity and phylogeny of Salmo with emphasis on southerntrouts (Teleostei, Salmonidae). Department of Zoology, Stockholm University: pp.142.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 126/141
Literatura
118
Derzhavin, A. N. 1934. Freshwater fishes from the south coast of the caspian.Transactions Azerb. Otd. Zakavkaz. Filiala AN SSR, 7: 91-126.
Dodson, J. J., Gibson, R. J., Cunjak, R. A., Friedland, K. D., Garcia de Leaniz, C., Gross,M. R., Newburym, R., Nielsen, J. L., Power, M. E., Roy, S. 1998. Elements in the
development of conservation plans for Atlantic salmon (Salmo salar ). CanadianJournal of Fisheries and Aquatic Sciences, 55: 312-323.
Dorofeeva, E. A. 1989. The basic principles of classification and phylogeny of thesalmonid fishes (Salmoniformes, Salmonoidei, Salmonidae). Biology andPhylogeny of Fishes, (Korovina, V. M., ed.). St. Petersburg: Proceedings ofZoologikal Institute, USSR Academy of Sciences (in Russian), pp. 5-16.
Dorofeeva, E. D. 1998. Systematics and distribution history of European Salmonid fishesof the genus Salmo. Journal of Ichthyology, 38: 419-429.
Dowling, T. E., Moritz, C., Palmer, J. D., Riesberg, L. H. 1996. Nucleic AcidsIII:Analysis of fragments and restriction sites. V. Molecular systematics. Sunderland,Sinauer, pp. 249-320.
Duftner, N., Weiss, S., Medgyesy, N., Sturmbauer, C. 2003. Enhanced phylogeographicinformation about Austrian brown trout populations derived from completemitochondrial control region secuences. Journal of Fish Biology, 62: 427-435.
Durand, J. D., Bianco, P. G., Laroche, J., Gilles, A. 2003. Insight into the origin ofendemic Mediterranean ichthyofauna: Phylogeography of Chondrostoma Genus(Teleostei: Cyprinidae). The Journal of Heredity, 94: 315-328.
Durand, J. J., Persat, H., Bouvet, Y. 1999. Phylogeography and postglacial dispersion ofthe chub ( Leuciscus cephalus) in Europe. Molecular Ecology., 8: 989-998.
Economidis, P. S., Banarescu, P. M. 1991. The distribution and origins of freshwaterfishes in the Balkan peninsula, especially in Greece. Internationale revue dergesamten hydrobiologie, 76: 257-283.
Edwards, A., Hammond, H. A., Jin, L., Caskey, C. T., Chakraborty, R. 1992. Geneticvariation at five trimeric and tetrameric tandem repeat loci in four human
population groups. Genomics, 12: 241-253.
Ekman, S. 1957. Zoogeography of the sea. Sidgwic & Jackson, London, pp. 417.
Elliott, J. M. 1994. Quantitative ecology and the brown trout. Oxford Series in Ecologyand Evolution, Oxford University Press, Oxford, Great Britain, 286 pp.
Englbrecht, C. C., Freyhof, J., Nolte, A., Rassmann, K., Schliewen, U., Tautz, D. 2000.Phylogeography of the bullhead Cottus gobio (Pisces: Teleostei: Cottidae)suggests a pre-Pleistocene origin of the major central European populations.Molecular Ecology, 9: 709-722.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 127/141
Literatura
119
Estoup, A., Angers, B. 1998. Microsatellites and minisatellites for molecular ecology:Theoretical and empirical considerations. In Carvalho, G.R. (ed.). Advances inMolecular Ecology. IOS Press, Amsterdam, pp. 55-86.
Estoup, A., Presa, P., Krieg, F., Vaiman, D., Guyomard, R. 1993. (CT)n and (GT)n
microsatellite: a new class of genetic markers for Salmo trutta L. (brown trout).Heredity, 71: 488-496.
Estoup, A., Rousset, F., Michalakis, Y., Cornuet, J.-M., Adriamanga, M., Guyomard, R.1998. Comparative analysis of microsatellite and allozyme markers: a case studyinvestigating microgeographic differentiation in brown trout (Salmo trutta).Molecular Ecology, 7: 339-353.
Ferguson, A. 1989. Genetic differences among brown trout, Salmo trutta L., stocks andtheir importance for the conservation and menagment of the species. FreshwaterBiology, 21: 35 - 46.
Ferguson, A., Fleming, C. C. 1983. Evolutionary and taxonomic significance of proteinvariation in the brown trout (Salmo trutta L.) and other salmonid fishes. In ProteinPolymorphism: Adaptive and Taxonomic Significance, Vol. 24 (Oxvord, G. S. &Rollinson, D., eds) London: Academic Press, pp. 84-99.
Ferguson, A., Mason F. M. 1981. Allozyme evidence for reproductively isolatedsympatric populations of brown trout Salmo trutta L. in Lough Melvin, Ireland.Journal of Fish Biology, 18: 629-642.
Ferris, S. D., Berg, W. J. 1988. The utility of mitochondrial DNA in fish genetics andfishery managment. V: Population genetics and fishery management. Seattle,Washington press, pp. 277-299.
Fontaine, P. M., Dodson, J. J. 1999. An analysis of the distribution of juvenile Atlanticsalmon (Salmo salar ) in nature as a function of relatedness using microsatellites.Molecular Ecology, 8: 189-198.
Ford-Lloyd, G. 1996. Measuring genetic variation using molecular markers. University ofBrimingham, Kevin Painting, IPGRI, Rome.
Frankel, O. H. 1970. Sir William Macleay memorial lecture 1970. Variation - the essenceof life. Proceedings of the Linneana Society of New South Wales 95: 158-169.
Frankel, O. H. 1974. Genetic conservation: our evolutionary responsibility. Genetics, 78:53-65.
Frankel, O. H., Soulé, M. E. 1981. Conservation and evolution. Cambridge UniversityPress, Cambridge, pp. 327.
Garcia-Marin, J. L., Jorde, P. E., Ryman, N., Utter, F., Pla, C. 1991. Managementimplications of genetic differentiation between native and hatchery populations of
brown trout (Salmo trutta) in Spain. Aquaculture, 95: 235-249.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 128/141
Literatura
120
Garcia-Marin, J. L., Pla C. 1996. Origins and relationships of native populations of browntrout (Salmo trutta) in Spain. Heredity, 76: 313-323.
Garcia-Marin, J. L., Utter, F. M., Pla, C. 1999. Postglacial colonization of brown trout inEurope based on distribution of allozyme variants. Heredity, 82: 46-56.
Gavrilović, LJ., Dukić, D. 2002. Reke Srbije. Zavod za uđžbenike i nastavna sredstva.Beograd.
Gillham, W. N. 1994. Organelle genome organization and gene content. V: Organellegenes and genomes. New York, Oxford University Press: 50-91.
Giuffra, E., Bernatchez, L., Guyomard, R. 1994. Mitochondrial control region and proteincoding genes sequence variation among phenotypic forms of brown trout Salmo
trutta from Northern Italy. Molecular Ecology, 3: 161-172.
Giuffra, E., Guyomard, R., Forneris, G. 1996. Phylogenetic relationships andintrogression patterns between incipient parapatric species of Italian brown trout(Salmo trutta L. complex). Molecular Ecology, 5: 207-220.
Glenn, T. C. 1995. Microsatellite manual, Version 6, Unpubished manuscript, FTP:onyx.si.edu/protocols/msatmanV#rtf
Goldstein, D. B., Pollock, D. D. 1997. Launching microsatellites: A review of mutation processes and methods of phylogenetic inference. The Journal of Heredity, 88:335-342.
Goldstein, D. B., Schlötterer, C. 1998. Microsatellites. Evolution and Applications.Oxford University Press, Oxford, pp. 352.
Gorjanović – Kramberger, D. 1891. Paleoihtioložki pilozi III. Rad JugoslovenskeAkademije Znanosti i Umjetnosti, Zagreb, 106: 59-129.
Grozdanović - Radovanović, J. 2000. Citologija. Zavod za udžbenike i nastavna sredstva,Beograd.
Guyomard, R., Krieg, F. 1983. Electrophoretic variation in six populations of brown trout
(Salmo trutta L.). Canadian Journal of Genetics and Cytology, 25: 403-413.
Gyllensten, U., Wilson, A. C. 1988. Mitochondrial DNA of salmonids. V: Populationgenetics and fishery management. Seattle, Washington press, pp. 301-317.
Hamilton, K. E., Ferguson, A., Taggart, J. B., Tomasson, T., Walker, A., Fahy, E. 1989.Post-glacial colonization of brown trout, Salmo trutta L.: Ldh-5 as a
phylogeographical marker locus. Journal of Fish Biology, 35: 651-664.
Hansen, M. M., Loeschcke, V. 1994. Effects of releasing hatchery-reared brown trout towild trout populations. In Loeschcke, V., Tomiuk, J. & Jain, S. K. (eds.).
Conservation Genetics. Birkhäuser Verlag, Basel. pp. 273-289

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 129/141
Literatura
121
Hansen, M. M., Loeschcke, V., Rasmussen, G., Simonsen, V. 1993. Geneticdifferentiation among Danish brown trout (Salmo trutta) populations. Hereditas,118: 177-185.
Hansen, M. M., Nielsen, E. E., Mensberg, K.-L.D. 1997. The problem of sampling
families rather than populations: Relatedness among individuals in samples of juvenile brown trout (Salmo trutta L.). Molecular Ecology, 6: 469-474.
Hansen, M. M., Ruzzante, D. E., Nielsen, E. E., Mensberg, K-L. D. 2000. Microsatelliteand mitochondrial DNA polymorphism reveals life – history dependentinterbreeding between hatchery and wild brown trout (Salmo trutta L.) MolecularEcology., 9: 583-594.
Harrison, R. G. 1989. Animal mitochondrial DNA as a genetic marker in population andevotutionary biology. Trends in Ecology and Evolution, 4: 6-11.
Hearne, C. M., Ghosh, S., Todd, J. A. 1992. Microsatellites for linkage of genetic analysisof genetic traits. Trends in Genetics, 8: 288-294.
Hennig, W. 1966. Phylogenetic systematics. Univ. Illinois Pres, Urbana, pp. 263.
Hewitt, G. M. 1993. Post - glacial distribution and species substructure: lessons from pollen, insects and hybrid zones. In: Evolutionary Patterns and Processes, 97-123.Linnean Society Symposium Series 14. Lees, D. R. & Edwards, D. (Eds.).Academic Press, London.
Hewitt, G. M. 1996. Some genetic consequences of ice ages, and their role in divergenceand speciation. Biological Journal of Linnean Society, 58: 274-276.
Hewitt, G. M. 1999. Post-glacial re-colonization of European biota. Biological Journal ofLinnean Society, 68: 87-112.
Hewitt, G. M. 2001. Speciation, hybrid zones and phylogeography – or seeing genes inspace and time. Molecular Ecology, 10: 537-549.
Hindar, K., Jonsson, B., Ryman, N., Ståhl, G. 1991. Genetic relationships amonglandlocked, resident, and anadromous brown trout, Salmo trutta L. Heredity, 66:
83-91.
Hindar, K., Ryman, N., Utter, F. M. 1991a. Genetic effects of cultured fish on natural fish populations. Canadian Journal of Fisheries and Aquatic Science, 48: 945-957.
Hoeh, W.R., Blakley, K. H., Brown, W. M. 1991. Heteroplasmy suggests limited biparental inheritance of Mytilis mitochondrial DNA. Science, 251: 1488-1490.
Hsü, K. 1978. When the Black sea was drained. Scientific American, 238: 52-63.
Hsü, K., Montadert, L., Bernouilli. D., Cita, M. B., Erikson, A., Garrison, R. E., Kidd, R.
B., Melieres, F., Müller, C., Wright, R. 1977. History of the Mediterraneansalinity crisis. Nature, 267: 399-403.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 130/141
Literatura
122
Hynes, R. A., Ferguson, A., McCann, M. A. 1996. Variation in mitochondrial DNA and post-glacial colonisation of north-west Europe by brown trout (Salmo trutta L.).Journal of Fish Biology, 48: 4-67.
ITIS, 2004. Integrated taxonomic information system. Smithsonian Institution,
Washington D. C. www.itis.usda.gov
Jevtić, J. 1989. Ribarstvo. Praktikum. Naučna knjiga, Beograd.
Johnson, G. D. C., & Patterson, C. 1996. Relationshipses of lower euteleostean fishes. In:Interrelations of Fishes. Stiassny , M. L. J., L. R. Parenti, G. D. Johnson (eds.).Academic Press, San Diego pp. 251-332.
Jug, T. 2002. Genetska raznolikost soške postrvi (Salmo marmoratus) v Sloveniji.Magistarsko delo, Medicinska fakulteta, Univerza v Ljubljani.
Karakousis, Y., Triantaphyllidis, C. 1990. Genetic structure and differentiation amongGreek brown trout (Salmo trutta L.) populations. Heredity, 64: 297-304.
Karaman, S. 1924. Pisces Macedoniae. Derzeit am institut Z. Erforschung undBeksampfung D. Malaria, Trogir (Dalmatien) Split.
Karaman, S. 1937. Beitrag zur Kenntnis der Süsswasserfische Jugoslaviens. GlasnikSkopskog naučnog društva, 18: 131-139.
Keith, P. 1998. Evolution des peuplements ichtyologiques de France et stratégies deconservation. Ph. D. diss., Université de Rennes I, France.
Kendall, A. W., Behnke, R. J. 1984. Salmonidae: developments and relationships. InOntogeny and Systematics of Fishes (Moser, H. G., ed.) American Society ofIchthiologysts and Herpetologists Spetial Publikation, 1: 142-149.
Klykanov, V. A. 1975. Systematic interrelations of the smelts of genera Osmerus Hypomesus of family Osmeridae. Ichthyologia, 7 (I): 31-40.
Kolombatović, G. 1890. Notizie ittiologiche. Glasnik prirodnjačkog društva, Zagreb.
Kottelat, M. 1997. European freshwater fishes. An heuristic checklist of the freshwaterfishes of Europe (exclusive of former USSR), with an introduction for non-systematists and comments on nomenclature and conservation. Biologia, SectionZoology 52 Suppl. 5: 1-271.
Krieg, F., Guyomard, R. 1985. Population genetics of French brown trout (Salmo trutta L.): large geographical differentiation of wild populations and high similarity ofdomesticated stocks. Génétique, Sélection et Evolution, 17: 225-242.
Kryštufek B., Reed J. M. 2004. Pattern andProcess in Balakn Biodiversity - an Overview,in:. Griffiths H.I., Kryštufek B., Reed J.M. (Ed), Balkan Biodiversity, Pattern and
Process in the European Hotspot, Kluwer Academic Publishers, Dordrecht, pp.203-217.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 131/141
Literatura
123
Kumar, S. Tamura, K. Nei, M. 2004. MEGA3: Integrated software for MolecularEvolutionary Genetics Analysis and sequence alignment. Briefings inBioinformatics, 5: 2 150-164.
Lagler, K. F. 1977. Freshwater fishery biology. Dubuque, WM. C. Brown Company
Publishers, pp.19-35.
Laikre, L., Antunes, A., Apostolidis, A., Berrebi, P., Duguid, A., Ferguson, A., Garcia –Marin, J. L., Guyomard, R., Hansen, M. M., Hindar, K., Koljonen, M. – L.,Largiader, C., Martinez, P., Nielsen, E. E., Palm, S., Ruzzante, D. E., Ryman, N.,Trianthaphyllidis, C. 1999. Conservation genetic management of brown trout(Salmo trutta) in Europe. Report by the concerted action on identification,management and exploitation of genetic resources in the brown trout (Salmo
trutta), "TROUTCONCERT"; EU FAIR CT97-3882. Silkeborg, Danmarksfiskeriundersrgelser: pp. 91.
Laikre, L., Ryman, N. 1996. Effects on intraspecific biodiversity from harvesting andenhancing natural populations. Ambio, 25: 504-509.
Largiadèr, C. R., Scholl, A. 1996. Genetic introgression between native and introduced brown trout (Salmo trutta L.) populations in The Rhône River Basin. MolecularEcology, 5: 417-426.
Leary, R. F., Allendorf, F. W., Forbes, S. H. 1993. Conservation genetics and bull trout inthe Columbia and Klamath River drainages. Conservation Biology, 7: 856-865.
Leidy, R. A., Moyle, P. B. 1998. Conservation status of the world's fish fauna: anoverview. In Fiedler, P.L. and Kareiva, P.M. (eds.). Conservation Biology for thecoming decade. Second edition. Chapman & Hall, International ThomsonPublishing, USA, pp. 187-227.
Lewin, B. 2000. Genus VII. Oxsfod University Press, University of Oxsfod, Oxsfod,United Kindom.
Lewontin, R. C. 1974. The genetic basis of evolutionary change. Columbia UniversityPress, New York.
Li, W. H., Grauer, D. 1991. Fundamentals of Molecular Evolution. Sinauer Associates,Sunderland, MA.
Lightowlers, R. N., Chinnera, P. F., Turnbull, D. M., Howell, N. 1997. Mammalianmitochondrial genetics: heredity, heteroplasmy and disease. Trends in genetics,13: 450-455.
Lopez, J. V., Culver, M., Stephens, J. C., Johnson, W. E., O'Brien, S. J. 1997. Rates ofnuclear and cytoplasmic mitochondrial DNA sequence divergence in mammals.Molecular Biology and Evolution., 14: 277-286.
Maitland, P. S. 1995. The conservation of freshwater fish: past and present experience.Biological Conservation, 72: 259-270.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 132/141
Literatura
124
Marić, S. 2002. Morfološka varijabilnost pastrmki (subfamilia Salmoninae) – značaj za biološku konzervaciju. Magistarski rad, Biološki fakultet, Univerziteta uBeogradu.
Marić, S., Hegediš, A., Nikolić, V., Simonović, P. 2004. Conservation status of two
eastern Balkan endemic fish species in Serbia and a proposal for their protection.Acta Zoologica Bulgarica 56 (2): 213-222.
Marshall, T. C., Slate, J., Kruuk, L. B., Pemberton, J. B. 1998. Statistical confidence forlikelihood-based paternity inference in natural populations. Molecular Ecology, 7:639-655.
Martin, A. P. 1999. Substitution rates of organelle and nuclear genes in sharks:implicating metabolic rate. Molecular Biology and Evolution., 16: 996-1002.
May, B. 1992. Starch gel electrophoresis of allozymes. In Hoelzel, A.R. (ed.). Molecular
Genetic Analysis of Populations. A Practical Approach. IRL Press, Oxford, pp. 1-27.
McKay, S. J., Devlin, R. H., Smith, M. J. 1996. Phylogeny of Pacific salmon and trout based on growth hormone type-2 and mitohondrial NADH dehydrogenase subunit3 DNA sequences. Canadian Journal of Fisheries and Aqatic Sciences, 53: 1156-1176.
McKenzie, J. A., Oberhansli, H. 1985. Paleoceanography expressions of the Messiniansalinity crisis, p. 99 – 123. In Hsü, K., Weissert, H. J. (eds), South AtlanticPaleoceanography. Cambridge University Press, Cambridge.
Meffe, G. K. 1986. Conservation genetics and the management of endangered fishes.Fisheries, 11: 14-23.
Miller, L. M., Kapuscinski, A. R. 1996. Microsatellite DNA Markers Reveal New Levelsof Genetic Variatio in Northen Pike. Transactions of the American FisheriesSociety, 125: 971-977.
Miller, L. M., Kapuscinski, A. R. 1997. Historical analysis of genetic variation revealslow effective population size in a northern pike ( Esox lucius) population.
Genetics, 147: 1249-1258.
Mitrović, J., Pavlović, M. 1980. Paleozoogeorafija. Rudarsko geološki fakultet – OOURGrupa za regionalnu geologiju i paleontologiju. Beograd.
Montoya, J., Gaines, G. L., Attardi, G. 1983. The pattern of transcription of the humanmitochondrial rRNA genes reveals two overlapping transcription units. Cell, 34:151-159.
Morán, P., Pendás, A. M., García-Vásquez, E., Izquierdo, J. I., Lobon-Cervia, J. 1995.Estimates of gene flow among neighbouring populations of brown trout. Journal
of Fish Biology, 46: 93-602.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 133/141
Literatura
125
Moritz, C. 1994. Applications of mitochondrial DNA analysis in the conservation: acritical review. Molecular Ecology, 3: 401-411.
Moritz, C. 1995. Uses of molecular phylogenies for conservation. PhilosophicalTransactions of the Royal Society of London. Series B, Biological sciences, 349:
113-118.
Moritz, C. 1999. Conservation units and translocations: strategies for conservingevolutionary processes. Hereditas, 130: 217-228.
Moritz, C., Dowling, T. E., Brown, W. M. (1987). Evolution of animal mitochondrialDNA: relevance for population biology and systematics. Annual Review ofEcology and Systematics, 18: 269-292.
Moritz, C., Hillis, D. M. 1996. Molecular Systematics: Context and Controversis. V:Molecular systematics. Sunderland, Sinauer, pp. 1-13.
Morizot, D. C., Schmidt, M. E. 1990. Starch gel electrophoresis and histochemicalvisualization of proteins. In Whitmore, D.H. (ed.). Electrophoretic and IsoelectricFocusing Techniques in Fishery Management. CRC Press, Boca Raton, AnnArbor, pp. 23-80.
Murphy, R. W., Sites, J. W., Buth, D. G., Haufler, C. H. 1996. Proteins: Isozymeelectrophoresis. V: Molecular systematics. Sunderland, Sinauer, pp. 51-120.
Neave, F. 1958. The origin and speciation of Oncorhynchus. Transaction of the RoyalSociety of Canada, Series 3, 52, (5): 25-49.
Nei, M., Maruyama, T., Chakraborty, R. 1975. The bottleneck effect and geneticvariability in populations. Evolution, 29: 1-10.
Nelsen, J. S. 1984. Fishes of the world. Wiley, New York.
Nesbo, C. L., Fossheim, T., Vollestad, L. A., Jakobsen, K. S. 1999. Genetic divergenceand phylogeographic relationships among European perch ( Perca fluviatilis)
populations reflect glacial refugia and postglcial colonization. Molecular Ecology,8: 1387-1404.
Nielsen, E. E., Hansen, M. M., Loeschcke, V. 1997. Analysis of microsatellite DNA fromold scale samples of Atlantic salmon: A comparison of genetic composition oversixty years. Molecular Ecology, 6: 487-492.
Nielsen, E. E., Hansen, M. M., Loeschcke, V. 1999a. Genetic variation in time and space:Microsatellite analysis of extinct and extant populations of Atlantic salmon.Evolution, 53: 261-268.
Nielsen, E. E., Hansen, M. M., Loeschcke, V. 1999b. Analysis of DNA from old scalesamples: Technical aspects, applications and perspectives for conservation.
Hereditas, 130: 265-276.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 134/141
Literatura
126
Nielsen, J. L. (ed.). 1995. Evolution and the aquatic ecosystem: defining unique units in population conservation. American Fisheries Society Symposium 17, pp. 435.
Nilsson, J., Gross, R., Asplund, T., Dove, O., Jansson, H., Kelloniemi, J., Kohlmann, K.,Löytynoja, A., Nielsen, E. E., Paaver, T., Primmer, C. R. Titov, S., Vasemägi, A.,
Veselov, A., Öst, T., Lumme, J. 2001. Matrilinear phylogeography of atlanticsalmon (Salmo salar L.) in Europe and postglacial colonization of Baltic sea area.Molecular Ecology, 10: 89-102.
Norden, C. R. 1961. Comparative osteology of representative salmonide fish with particular reference to the grayling (Thymallus arcticus) and phylogeny. Journalof the Fisheries Research Board of Canada, 18: 679-791.
Oakley, T. H., Phillips, R. B. 1999. Phylogeny of salmonine fishes based on growthhormone inrons: Atlantic (Salmo) and Pacific (Oncorhynchus) salmon are notsister taxa. Molecular Phylogenetics and Evolution, 11: 381-393.
Ocokoljić, M. 1987. Visinsko zoniraje voda u slivu Velike Morave i neki aspekti njihovezaštite. Posebna izdanja Srpskog geografskog društva. Sveska 64, Beograd.
Odak, T. 2004. Molekularno – biološka obilježja endemske mekousn pastrve(Salmothymus obtusirostris salontiana). Magistarski rad, Agronomski fakultet,Sveučilišta u Zagrebu.
Oleynik, A. G. 1997 Molecular phylogeny of salmonid fishes: concordance of resultsfrom nuclear and mitohondrial DNA analyses. Genetika, 33: 229-234 (in Russian).
Osinov, A. 1984. Zoogeographical origins of brown trout , Salmo trutta (Salmonidae):data from biochemical genetic markers. Journal of Ichtyology., 24: 10-23.
Osinov, A. 1989. Brown trout (Salmo trutta L., Salmonidae) in basins of the Black andCaspian Seas: A population genetic analysis. Genetika, 24: 1523-1534.
Osinov, A. G. 1999. Salmonid fishes of the genera Salmo, Parasalmo, andOncorhynchus: genetic divergence, phylogeny, and classification. Journal ofIchtyology, 39: 571-578.
Osinov, A., Bernatchez, L. 1996. Atlantic and Danubean phylogenetic groupings of brown trout (Salmo trutta L.) complex: genetic divergence, evolution, andconservation. Journal of Ichthyology, 36: 762-786.
Oxnard, C. E. 1978. One biologist’s view of morphometrics. Annual Review of Ecologyand Systematics, 9: 219-241.
Patarnello, T., Bargelloni, L., Caldara, F., Colombo, L. 1994. Cytohrome b and 16 rRNAsequence variation in the Salmo trutta (Salmonidae, Teleostei) species coplex.Molecular Phylogenetics and Evolution, 3: 69-74.
Pažur, K. 1969. Divovske pastrve u jezeru Lokvare. Ribarstvo Jugoslavije, God. XXIV, 2,Zagreb.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 135/141
Literatura
127
Phillips, R. B., Matsuoka, M. P., Konon, I., Reed, K. M. 2000. Phylogenetic analysis ofmitochondrial and nuclear sequences supports inclusion of Acantholingua
ochridana in the genus Salmo. Copeia, 2: 546-550.
Phillips, R. B., Oakley, T. H. 1997. Phylogenetics relationships among the Salmoninae
based on nuklear and mitochondrial DNA sequences. In Molecular Systemaaticsof Fishes (Kocher, T. D. & Stepien, C. A.; eds), Academic Prees, San Diego pp.145-162.
Phillips, R. B., Plate, K. A. 1991. Nuklear DNA and salmonid phylogenetics. Journal ofFish Biology, 39 (Suppl. A): 259-275.
Pielou, E. C. 1979. Biogeography. Wiley, New York, pp. 351.
Poteaux, C., Beaudou, D., Berrebi, P. 1998. Temporal variations of genetic introgressionin stocked brown trout populations. Journal of Fish Biology 53: 701-713.
Poteaux, C., Bonhomme, F., Berrebi, P. 1999. Microsatellite polymorphism and geneticimpact of restocking in Mediterranean brown trout (Salmo trutta L.). Heredity, 82:645-653.
Povž, M., Jesenšek, D., Berrebi, P., Crivelli, A. J. 1996. Soška postrv Salmo trutta
marmoratus, Cuvier 1817, v poreč ju Soče v Sloveniji. Arles, Tour du Valat, pp.65.
Povž, M., Sket, B. 1990. Naše sladkovodne ribe. Mladinska knjiga, Ljubljana, pp 85-86.
Queller, D. C., Strassmann, J. E., Hughes, C. R. 1993. Microsatellites and kinship. Trendsin Ecology and Evolution., 8: 258-288.
Rabrenović, D., Knežević, S., Rundić, Lj. 2003. Istorijska geologija. Rudarsko – Geološkifakultet Univerziteta u Beogradu, Beograd.
Rage, J. C. 1997. Palaeobiological and palaeogeographical background of the Europeanherpetofauna. In: Atlas of Amphibians and Reptiles in Europe. Gasc, J. - P.,Cabela, A., Crnobrnja - Isailović, J., Dolmen, D., Grossenbacher, K., Haffner, P.,Lescure, J., Martens, H., Martinez Rica, J. P., Maurin, H., Oliveira, M. E.,
Sofianidou, T. S., Veith, M. & Zuiderwijk, A. (Eds.). Paris: Societas EuropaeaHerpetologica and Museum National d'Histoire Naturelle, pp. 23-27.
Rage, J. C., Roček, Z. 2003. Evolution of Anuran assemblages in the Tertiary andQuaternary of Europe, in the context of palaeoclimate and palaeogeography.Amphibia-Reptilia, 24: 133-167.
Riffel, M., Storch, V., Schreiber, A. 1995. Allozyme variability of brown trout (Salmo
trutta L.) populations across the Rhenanian-Danubian watershed in southwestGermany. Heredity, 74: 241-249.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 136/141
Literatura
128
Rögl, F., Steininger, F. F. 1983. Vom Zerfall der Tethys zu Mediterran und Paratethys. Dieneogene Paläogeographie und Palinspastik der zirkum - mediterranen Raumes.Annalen des Naturhistorischen Museum Wien A 85, 135-163.
Roy, K., Valentine, J. W., Jablonski, D., Kidwell, S. M. 1996. Scales of climatic
variability and time averaging in Pleistocene biotas: implications for ecology andevolution. Trends in Ecology and Evolution, 11: 458-453.
Ryder, O. A. 1986. Species conservation and systematics: the dilemma of subspecies.Trends in Ecology and Evolution, 1: 9-10.
Ryman, N. 1983. Patterns of distribution of biochemical genetic variation in salmonids:differences between species. Aquaculture, 33: 1-21.
Ryman, N. 1991. Conservation genetics considerations in fishery management. Journal ofFish Biology, 39 (Supplement A): 211 - 224.
Ryman, N., Allendorf, F. W., Stähl, G. 1979. Reproductive isolation with little geneticdivergence in sympatric populations of brown trout (Salmo trutta). Genetics, 92:247-262.
Ryman, N., Jorde, P. E., Laikre, L. 1995. Supportive breeding and variance effective population size. Conservation Biology, 9: 1619-1628.
Ryman, N., Ståhl, G. 1980. Genetic changes in hatchery stocks of brown trout ( Salmo trutta). Canadian Journal of Fisheries and Aquatic Sciences, 37: 82-87.
Ryman, N., Utter, F. (eds.). 1987. Population Genetics and Fishery Management.Washington Sea Grant Publications/University of Washington Press, Seattle andLondon, pp. 420.
Ryman, N., Utter, F., Laikre, L. 1995a. Protection of intraspecific biodiversity ofexploited fishes. Reviews in Fish Biology and Fisheries, 5: 417-446.
Saiki, R., Scharf, S., Faloona, F., Mullis, K. B., Horn, G. T., Erlich, H. A., Arnheim, N.1988. Enzymatic amplification of b – globin genomic sequences and restrictionsite analysa for diagnosis of sickle cell anemia. Science, 4: 1350-1354.
Saunders, R. L., Schom, C. B. 1985. Importance of the variation in the life history parameters of atlantic salmon (Salmo salar). Canadian Journal of Fisheries andAquatic Sciencis, 42: 625-618.
Schlötterer, C. 1998. Genome evolution: Are microsatellites really simple sequences?.Current Biology, 8: 132-134
Schmidt, D. Y. 1947. Migrations of Fishes. USSR Academy of Sciences, Leningrad.
Schneider, S. Roessli, D. Excoffier, L. 2000. Arlequin: A software for population genetics
data analysis. Ver 2.000. Genetics and Biometry Lab, Dept. of Anthropology,University of Geneva.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 137/141
Literatura
129
Shaposhnikova, G. Ch. 1975. Systematic relation of some representatives of the familySalmonidae. - Ichthyologia, Vol. 7, No. 1, 61-70.
Shed'ko, S. V., Ginatulina, L. K., Parpura, I. Z., Ermolenko, A. V. 1996. Evolutionary andtaxonomic relationships among Far-Eastern salmonid fishes inferred from
mitichondrial DNA divergence. Journal of Fish Biology, 49: 815-829.
Shedlock, A. M., Parker, J. D., Crispin, D. A., pietsch, T. W., Burmer, G. C. 1992.Evolution of the salmonid mitochondrial control region. Molecular Phylogeneticsand Evolution, 1: 179-192.
Smith, S. R. 1992. Introgression in fishes: significance for paleontology, cladistics, andevolutionary rates. Systematic Biology, 41: 41-57.
Snoj, A. 1997. Molekularno biološka karakterizacija soške postrvi (Salmo marmoratus,Cuvier 1817). Doktorska disertacija, Biotehniška fakulteta, Univerza v Ljubljani.
Snoj, A. 2003. Molecular phylogeny of «arhaic trouts» and their taxonomic classification.3rd congress of the genetics society of Slovenija, Bled. Zbornik sažetaka, pp. 73-74.
Snoj, A. 2004. Filogenetska struktura postrvi (Salmo trutta L.) v Sloveniji. Ribič, GlasiloSlovenskog ribištva. Ribiška sveza Slovenije, 10: 239-243.
Snoj, A., Jug, T., Melkič,. E., Sušnik, S., Pohar, J., Dovč,. P., Budihna, N. 2000.Mitochondrial and mirosatellite DNA analysis of marble trout in Slovenia. Journalof Fish Biology (Quaderni ETP), 29: 5-11.
Snoj, A., Melkič,. E., Sušnik, S., Muhamedagić, S., Dovč, P. 2002. DNA phylogenysupports revised classification of Salmothymus obtusirostris. Biological Journal ofthe Linnean Society, 77: 399-411.
Snoj, A., Pohar, J., Dovč, P. 1997. The first microsatellite DNA marker for marble trout,BFRO001. Journal of Animal Science, 75: 1983.
Stalling, R. L., Ford, A. F., Nelson, D., Torney, D. C., Hildebrand, C. E., Moyzis, R. K.1991. Evolution and distribution of (GT)n repetitive sequences in Mammalian
genomes. Genomics, 10: 807-815.
Stearley, R. F., Smith, G. R. 1993. Phylogeny of the Pacific trouts and salmons(Oncorhynchus) and genera of the family Salmonidae. Transactions of theAmerican Fisheries Society, 122: 122 - 133.
Stevanović, M. P. 1982. Istorijska geologija. Rudarsko – Geološki fakultet Univerziteta uBeogradu, Beograd.
Strauss, R. E., Bookstein F. L. 1982. The truss: body form reconstructions inmorphometrics. Systematic Zoology, 31: 113-135.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 138/141
Literatura
130
Suárez J., Bautista J. M., Almodóvar A., Machordom A. 2001. Evolution ofmitochondrial control region in Palaeartic brown trout (Salmo trutta) populations:the biogeographical role of the Iberian Peninsula. Heredity, 87: 198-206.
Sušnik, S. 2001. Polimorfizem kromosomske in mitohondrijske DNA lipana (Thymallus
thymallus) in filogenetski odnosi med njegovimi geografsko ločenimi populacijami. Doktorska disertacija, Medicinska fakulteta, Univerza v Ljubljani.
Sušnik, S., Schöffman, J., Snoj, A. 2004. Phylogenetic position of Salmo ( Platisalmo) platicephalus Behnke 1968 from south – central Turkey, evidenced by geneticdata. Journal of Fish Biology, 64: 947-960.
Sušnik, S., Schöffman, J., Weiss, S. 2005. Genetic verification of native brown trout fomthe Persian Gulf (Catak Cay River, Tigris basin). Journal of Fish Biology, 66: 1-6.
Sušnik, S., Snoj, A., Pohar, J., Dovč, P. 1997 The microsatellite marker (BFRO 002)
characteristic for different geographically remote brown trout, Salmo trutta L., population. Animal Genetics, 28, 5: 372.
Swofford, D. L. 2000. PAUP*, b-VERSION 4.0. Sunderland, MA: Sinauer.
Taberlet, P., Fumagalli, L., Wust - Saucy, A. - G., Cosson, J. - F. 1998. Comparative phylogeography and postglacial colonization routes in Europe. MolecularEcology, 7: 453-464.
Tautz, D. 1989. Hypervariability of simple sequences as a general source for polymorphicDNA markers. Nucleic Acid Research, 17: 6463-6471.
Tautz, D. 1993. Notes on the definition and nomenclature of tandemly repetitive DNAsequences. V: DNA fingerprinting: State of the science. Basel, Birkhäuser Verlag,
pp. 21-28.
Tautz, D., Trick, M., Dover, G. A. 1986. Cryptic simplicicity in DNA is a major source ofgenetic variation. Nature, 322: 652-656.
Taylor, E. B. 1991. A review of local adaptation in Salmonidae, with particular referenceto Pacific and Atlantic salmon. Aquaculture, 98: 185-207.
Taylor, E. B., Pollard, S., Louie, D. 1999. Mitochondrial DNA variation in bull trout(Salvelinus confluentus) from northwestern North America: implications forzoogeography and conservation. Molecular Ecology, 8: 1155-1170.
Templeton, A. R. 2004. Using haplotype trees for phylogeographic and species inferencein fish populations. Environmental Biology of Fishes, 69: 7-20.
Tessier, N., Bernatchez, L. 1999. Stability of population structure and genetic diversityacross generations assessed by microsatellites among sympatric populations oflandlocked Atlantic salmon (Salmo salar L.). Molecular Ecology, 8: 169-179.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 139/141
Literatura
131
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., Higgins, D. G. 1997. TheClustal_X windows interface: flexibile strategies for multiple sequence aligmentaided by quality analysis tools. Nucleic Acid Research, 25, 24: 4876-4882.
Thunell, R. C., Williams, D. F. 1983. Paleotemperature and paleosalinity history of the
eastern Mediterranean during the Late Quaternary. Palaeogeography,Palaeoclimatology, Palaeoecology, 44: 23-39.
Tomkinson, A. E., Linn, S. 1986. Purification and properties of a strand specificendonuclease from mouse cell mitochondria. Nucleic Acid Research, 14: 9579-9593.
Utter, F. M. 1981. Biological criteria for definition of species and distinct intraspecific populations and salmonids under the U.S. Endangered Species Act of 1973.Canadian Journal of Fisheries and Aquatic Sciences, 38: 1626-1635.
Utter, F., Aebersold, P., Winans, G. 1987. Interpreting genetic variation detected by
electrophoresis. In Ryman, N. and Utter, F. (eds.) Population Genetics and FisheryManagement. Washington Sea Grant Program/University of Washington Press,Seattle, USA.
Villinger, E. 1986. Untersuchungen zur Flussgeschichte von Aare-Donau/Alpenrhein undzur Entwicklung des Malm-Karts in Su¨dwestdeutschland. Germany.
Vladimirov, V. I. 1944. Brook from of Sevan trout: Salmo ischchan Kessler morphaalabalach nova. Izv Akademy Nauk Arm. SSR, 3: 61-72.
Vladimirov, V. I. 1948. Brook trout in Armenia and its relationship to otherrepresentatives of the Salmo genus. Transactions sevan Gigrobiology Stantsii., 10:87-178.
Vladykov, V. D. 1963. A review of salmonid genera and their broad geographicaldistribution. Transaction of the Royal Society of Canada, Series IV, Section III, 1:459-504.
Vollestad, L. A., Hindar, K. 1997. Developmental stability and environmental stress inSalmo salar (Atlantic salmon). Heredity, 78: 215-222.
Vuković, T., Ivanović, B. 1971. Slatkovodne ribe Jugoslavije. Zemaljski muzej BIH,
Sarajevo.
Waples, R. S. 1991. Genetic interactions between hatchery and wild salmonids - lessonsfrom the Pacific Northwest. Canadian Journal of Fisheries and Aquatic Sciences,48 (Suppl. 1): 124-133.
Waples, R. S. 1991a. Pacific salmon, Oncorhynchus spp., and the definition of "species"under the Endangered species Act. US National Marien Fisheries Service, MarineFisheries Review, 53: 11-22.
Waples, R. S. 1995. Evolutionary significant units and the conservation of biological
diversity under the endangered species act. American Fisheries SocietySymposium, 17: 8-27.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 140/141
Literatura
132
Weiss, S., Antunes, A., Schlötterer, C., Alexandrino, P. 2000. Mitochondrial haplotypediversity among Portuguese brown trout Salmo trutta L. populations: relevance tothe post-Pleistocene recolonization of northern Europe. Molecular Ecology, 9:691-698.
Weiss, S., Schlötterer, C., Waidbacher, H., Jungwirth, M. 2001. Haplotype (mtDNA)diversity of brown trout Salmo trutta in tributaries of the Austrian Danube:massive introgresion of Atlantic basin fish – by man or nature. MolecularEcology, 10: 1241-1246.
Weissenbach, J., Gypapy, G., Dib, C., Vignal, A., Morissette, J., Millasseau, P., Vaysseix,G., Lathrop, M. 1992. A second-generation linkage map of the human genome.
Nature, 359: 794-801.
Wheeler, A. 1992. Freshwater fishes of Britain and Europe. Rainbow Books, London, pp.54-55.
Zhang, D.-X., Hewitt, G. M. 2003. Nuclear DNA analyses in genetic studies of populations: practice, problems and prospects. Molecular Ecology, 12: 563-584.

7/24/2019 Razlikovanje Evolutivnih Linija Potocne Pastrmke
http://slidepdf.com/reader/full/razlikovanje-evolutivnih-linija-potocne-pastrmke 141/141