Embed Size (px)
Citation preview

Reproductive and Metabolic Characteristics of Dairy Cattle Supplemented with/3-Carotene
E. M. B|NDAS, F. C. GWAZDAUSKAS, R. J. A|ELLO, J. H. HERBEIN, M. L. McGILLIARD, and C. E. POLAN
Department of Dairy Science College of Agriculture and Life Sciences
Virginia Agricultural Experiment Station Virginia Polytechnic Institute and State University
Blacksburg 24061--6999
ABSTRACT
Seventy-eight Holstein cows alternately were assigned at calving to receive /3- carotene supplementation or act as controls to determine effects of/3-carotene on reproduction and carotene, luteinizing hormone, progesterone, insulin, glucose, and glucagon concentrations in blood plasma. Cows were fed a corn silage-based complete ration. Biweekly jugular blood samples were collected beginning the week after parturition through 90 days. At day 30, supplemented cows received 600-nag synthetic ~3-carotene daily for 60 days. Plasma carotene reached a peak of 2.45 /.tg/ml compared to 1.50 /lg/ml in controls. Supplementation significantly increased plasma carotene but had no effect on luteinizing hormone, pro- gesterone, insulin, glucose, glucagon, or reproductive measures. Days to first heat, days to first breeding, days open, and services per conception averaged 74, 74, 95, and 1.7 for supplemented cows and 64, 76, 102, and 1.9 for control cows. Progesterone increased as lactation pro- gressed. Somatic cells were not different between supplemented and control cows. Supplementation of /3-carotene did not improve reproductive efficiency or alter luteinizing hormone, progesterone, in- sulin, glucose, or glucagon in blood plasma or affect somatic cells in milk.
INTRODUCTION
As early as 1938 the role of hormones in nutrition and their interrelationships were under discussion (36). During this time /3-
Received September 9, 1983.
carotene was thought to have a biological value equivalent to vitamin A (15), and Converse and Meigs (5) showed that rations of low/3-carotene content were involved with a deficiency that increased calving difficulty. In 1941, Davis and Madsen (7) defined minimum carotene con- centrations of blood plasma needed for birth of normal calves. However, reported recently was that ~3-carotene plays a specific role in re- productive efficiency of cattle apart from that of vitamin A (6, t3, 24). In these studies, lowered services per conception (6, 24), de- creased days open (6, 13), and higher pro- gesterone (24) in blood plasma were observed in cattle fed supplemental ~3-carotene compared to cattle not supplemented. However, Folman et al. (12) did not find significant differences for these reproductive measures when heifers were supplemented with /3-carotene. BremeI et al. (2) found no significant differences for services per conception among supplemented and unsupplemented groups of cows fed either corn silage or cottonseed hulls.
~3-Carotene content of corn silage, a forage that makes up considerable proportions of dairy herd rations, is low (19, 20), and cattle fed high corn silage rations usually have low vitamin A in blood plasma (19, 31). During the 1950's continued feeding of suboptimal carotene rations (<.35 /lg/kg feed) resulted in lower reproductive efficiency and deleterious effects on pituitary, testicular, and ovarian function (3). Ronning et al. (29) found early termination of gestation when cattle were fed less than 40 /lg carotene/kg body weight. Retained placenta and services per conception were increased. Additionally, Kuhlman and Gallup (23) reported that low dietary carotene resulted in more services per conception. Because more recent studies (2, 6, 12, 13, 24) report a conflicting role of carotene in the reproductive process, this study was designed to provide a data base
1984 J Dairy Sci 67:1249--1255 1249

1 2 5 0 BINDAS ET AL.
from which to establish accurate carotene profiles resulting from short-term supple- mentat ion prior to the time of insemination by /3-carotene and to assess the relationship between stresses of the postpartum period, possible carotene deficiency, and reproductive and metabolic variables of dairy cattle.
MATERIALS AND METHODS
Seventy-eight Holstein cattle at Virginia Tech Dairy Cattle Center were assigned ran- domly at calving to a ration supplemented with 3-carotene (n=36) or a control ration (n=42). From day 30 through 90 of lactation, each cow in the 3-carotene group was supple- mented daily with 600 mg synthetic 3-carotene (Roche-Rovimix). Cows were housed either in free stalls or a tie-stall barn and fed during the entire period for ad libitum consumption 16% crude protein ration containing 72% corn silage, 14% high-moisture corn, 12% soybean meal concentrate, 1% molasses, and 1% vitamin/ mineral premix supplying adequate vitamin A (>65,000 IU/day).
Individual cow records included milk weights every 5th day during the supplementation period, days to first heat, days to first breeding, days open, and services per conception. Cows were observed daily for estrus in an exercise lot and as they were moved to and from the milking parlor. Cows were inseminated on first estrus after 60 days postpartum.
Biweekly jugular blood samples (20 ml) were taken beginning the 1st wk after calving between 0800 and 1000 h or approximately 3 to 4 h prior to milking. Feed was available at all times, and in most cases additional feed was added 1 to 2 h before bleeding. Plasma was obtained and frozen in aluminum foil-covered tubes (to prevent destruction of 3-carotene by exposure to light) until quantification of hormones and carotene. Plasma was analyzed for luteinizing hormone (LH) (1), progesterone (8), insulin (22), and glucagon (16) by radioimmunoassay and glucose (11) and carotene (21) by spectro- photometry , lntra- and interassay coefficients of variation were 11.5 and 8.9 for carotene and 7.5 and 10.7% for progesterone. Luteinizing hormone, insulin, and glucagon were quantified in single assays, and coefficients of variation within assay were 6.3, 2.7, and 6.1%. Somatic cell counts were obtained from Dairy Herd
Improvement records for the months cows were on trial.
Statistical models were: Yijk = /l + Ti + C(T)ij + Pk + (TP)ik + eijk where Yijk = hormone or metaboli te concentration and p = parametric mean; i = 1, 2 treatments (T); j = 1, 2 . . . n cows (C) per T; k = 1, 2 . . . 5 periods (P); (TP)ik = interaction of T and P; and eij k = residual deviation; Yi" =/ l+Ti + L" + (TL) i' + e i" where J . 'J J l . Yij = reproductive variable and/a = paramemc mean; i = 1, 2 t reatments (T); j -= 1, 2 . . . n lactation numbers (L); (TL)ij = interaction of T and L; and eij = residual deviation; and Yijkl = a
2 b n M(ijk)l n + + Ti + Nj + bS(ijk)l + 2;n= 1 2 (T*N)i j + 23n= 1 bni M(ijk)lnTi + e(ijk)l where
Yijkl = somatic cell count and a = Y intercept; i = 1, 2 treatments (T); j = 1 , 2 . . . n lactation numbers (N); S(ijk)l = initial somatic cell count (S) prior to t reatment; 1 variation between months (M) during the study; b = regression of Yijkl on S(ijk)l; bn = linear and quadratic regressions (n=l or 2) of Yij.kl on M(ijk)l; bni = linear and quadratic regressions (n= 1 or 2) of Yijkl on M(ijk)l within Ti expressed as a deviation from bn; (T*N)ij = interaction of T and N; and eijkl residual deviation.
RESULTS A N D DISCUSSION
Analysis of variance for plasma carotene is in Table 1. Treatment, cow, bleeding period, and interaction of t reatment by bleeding period were significant (P<.01). Variation among cows also was significant in studies by Sutton et al. (33), McGillivray (25), and David and Madsen (7).
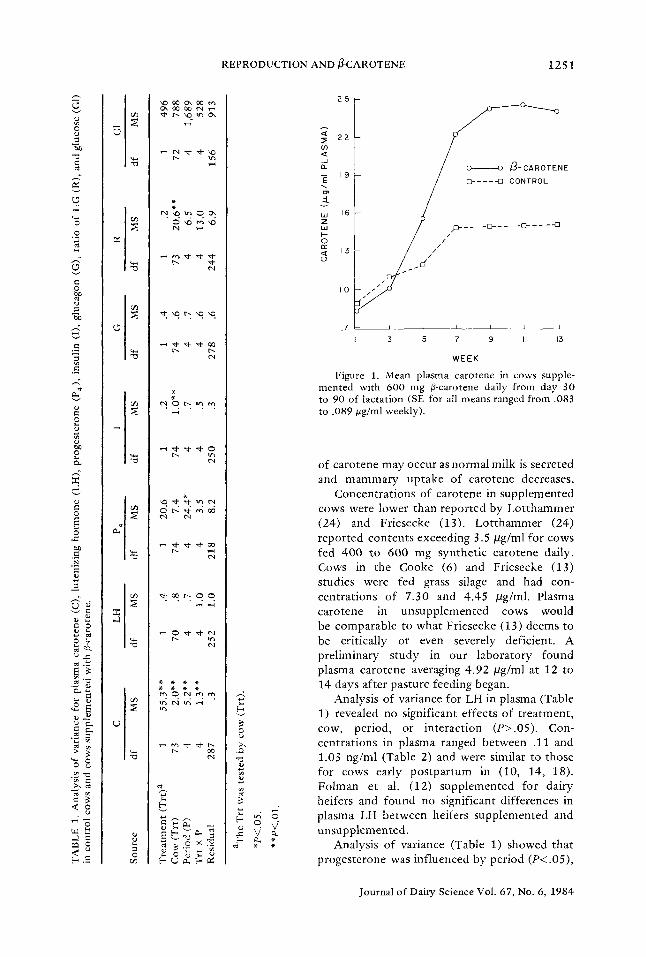
Figure 1 shows means of plasma carotene in both t reatment groups during the seven biweekly bleeding periods. Plasma carotene was increased significantly by 1 wk following initiation of supplementation (wk 5), and supplementation increased plasma carotene to maximum con- centrations of approximately 2.45 /~g/ml by 9 and 11 wk postpartum. Plasma carotene in- creased in control cows after parturit ion, despite no carotene fed to a maximum con- centration of 1.50 /lg/ml by wk 7 and then plateaued. Postpartum increases of carotene have been reported by (30, 34, 35). The mam- mary gland absorbs large amounts of carotene near parturi t ion to supply newborn calves with vitamin A via colostrum. Postpartum increases
Journal of Dairy Science Vol. 67, No. 6, 1984
REPRODUCTION AND ~-CAROTENE 1251
r ~ v
O
.--.7
v
O
. - Z
v
.o
o
, - Z
O
O e~
_=
< o
e ~
I ~ o E
N N ~ M ~
m
m ~ m m ~ M M M '
g
o
0
25~-
2 2 O0
g
Ld I'- 0 I1: o< J3
I 0
.7
.d// -- I I I I I l
5 5 7 9 II 15
WEEK
Figure 1. Mean plasma carotene in c o w s supple- mented with 6 0 0 m g fl-carotene daily from day 30 to 90 o f lactat ion (SE for all means ranged from , 0 8 3
to . 089 #g /ml w e e k l y ) .
of carotene may occur as normal milk is secreted and mammary uptake of carotene decreases.
Concentrations of carotene in supplemented cows were lower than reported by Lotthammer (24) and Friesecke (13). Lotthammer (24) reported contents exceeding 3.5 #g/ml for cows fed 400 to 600 mg synthetic carotene daily. Cows in the Cooke (6) and Friesecke (13) studies were fed grass silage and had con- centrations of 7.30 and 4.45 ~g/ml. Plasma carotene in unsupplemented cows would be comparable to what Friesecke (13) deems to be critically or even severely deficient. A preliminary study in our laboratory found plasma carotene averaging 4 .92 / l g /ml at 12 to 14 days after pasture feeding began.
Analysis of variance for LH in plasma (Table 1) revealed no significant effects of treatment, cow, period, or interaction (/)>.05). Con- centrations in plasma ranged between .11 and 1.03 ng/ml (Table 2) and were similar to those for cows early postpartum in (10, 14, 18). Folman et at. (12) supplemented for dairy heifers and found no significant differences in plasma LH between heifers supplemented and unsupplemented.
Analysis of variance (Table 1) showed that progesterone was influenced by period (P<.05),
Journal of Dairy Science Vol. 67, No. 6, 1984

1 2 5 2 BINDAS ET AL.
TABLE 2. Least squares means and SE for luteinizing ho rmone (ng/ml), glucagon (ng/ml), ratio of insulin :glucagon, and glucose (mg/dl) in cows supplemented with 600 mg B carotene during early lactation.
Week pos tpar tum ,,
1 3 a 5 7 9 11 13 Overall SE
Luteinizing ho rmone B-Carotene .11 .62 .99 .68 1.03 .65 .69 .74 .15 Control .59 .80 .74 .56 .71 .95 .74 .73 .!5
Glucagon B-Carotene .24 .36 .36 .35 .31 .33 .38 .33 .33 Control .30 .59 .75 .34 .35 .31 .33 .42 .42
Insulin: Glucagon B-Carotene 2.39 1.43 2.53 1.70 3.21 2.14 2.87 2.33 .46 Control 1.58 1.14 1.96 2.24 1.93 2.79 2.89 2.08 .46
Glucose B-Carotene 66.8 60.7 71.7 67.4 60.3 55.6 69.4 64.6 2.6 Control 60.7 58.1 70.3 53.5 54.6 61.8 65.6 60.7 2.7
aSupplementa t ion began after wk 3.
but there was no significant effect of t reatment (P>.O5). Lot thammer (24) found significantly higher progesterone in plasma of cows fed /3-carotene compared to unsupplemented cows. However, Folman et al. (12) did not find differences between heifers supplemented and unsupplemented.
Figure 2 shows mean progesterone in plasma in both t reatment groups. Plasma progesterone increased in both groups throughout the study, which concurs with studies reporting postpar tum increases in progesterone as the bovine estrous cycle slowly returned to a normal ovulatory pat tern following parturi t ion (9, 37). By 5 wk postpartum, progesterone concentrations in- dicated that cows in both t reatment groups had begun to cycle. Thereafter, plasma concen- trations were comparable to bovine luteal phase. However, because of the biweekly sampling protocol , all progesterone concen- trations were not luteal concentrations. Some cow may have been near estrus, and some after 9 wk were pregnant. Between 60 to 90 days postpartum, 15 of 36 (42%)cows had conceived in the supplemented group, and 15 of 42 (36%) had conceived in the control group. Thus, progesterone averages also included concentra- tions during pregnancy maintenance.
Least squares means for reproductive vari- ables are in Table 3. No differences were significant between these means. For days to first heat, at least one heat period had been
missed or a silent ovulation had occurred prior to first detectable heat. Bremel et al. (2) observed similar days to first heat in a recent study of/3-carotene supplementation. Incidence of silent heats in early postpar tum cows is frequent and could be due to lack of signs of estrus or unobserved signs of estrus. Whitemore et al. (38) reported 43% of 386 first ovulations
47
4.2 // \" ///f~D
-~:o22"" z, / 5 ; • ¢c[~2:222 /' ENE uJu) 17 , , /
0 ~e t2 a .
3 5 7 9 II 13
WEEK
Figure 2. Mean plasma progesterone in cows supplemented with 600 mg B-carotene daily f rom day 30 to 90 o f lactation (SE for all means ranged from .42 to .49 ng/ml weekly).
Journal o f Dairy Science Vol. 67, No. 6, 1984

REPRODUCTION AND ~CAROTENE 1253
TABLE 3. Least square means of reproductive variables measured in control cows and cows fed 3-carotene.
3-carotene Control Measure X SE (n) X SE (n)
Days to first heat 74 7 (35) 64 11 (40) Days to first breeding 74 4 (32) 76 4 (37) Days open 95 11 (28) 102 8 (35) Services per conception 1.7 .3 (28) 1.9 .2 (35)
occurred without observation of estrus behavior. In study of California dairy herds, Pelissier (26) found that 42% of the cows' showed first heat after 60 days postpartum, which suggested failure to detect first postpartum estrus.
Supplementation with Ê-carotene of cows in Europe (6, 13, 24) reduced days open. Our results indicate similar and optimal days open for both groups. Perhaps if a herd with obvious reproductive problems had been used, im- provement of days open may have been notice- able for cows supplemented with fl-carotene. Services per conception were similar to those in other supplementation studies (6, 12, 13, 24). There were no significant differences for services per conception between treatment groups in our study or in (12). This is in contrast to (6, 13).
i.oo
P i
! O- . . . . ~ CONTROL /
~" 0 0 ,~-CAROTENE / ~ ,
d o .
E 7 0
_z 60 ,,~ . . . . ~ / . . . . d" .J
_z 50 i I
3 5 7 9 I I 13
WEEK
Figure 3. Mean plasma insulin in cows supplemented with 600 mg 3-carotene daily from day 30 to 90 of lactation (SE for all means ranged from .09 to .13 ng/ml weekly).
Insulin and glucose in blood plasma were not affected by fl-carotene treatment (Table 1). Insulin concentrations tended to increase in both treatment groups (Figure 3), although period was not significant, and glucose did not change during the sampling period (Table 2). Several studies reported low plasma insulin following parturition and subsequent increasing concentrations as lactation proceeded (17, 22, 27, 32). Mobilization of stored energy and gluconeogenic processes would be enhanced during early lactation when insulin concen- trations are low. It is believed that insulin inhibits lipolysis and gluconeogenesis, which would prevent fulfillment of metabolic demands during peak production (28).
Glucagon in blood plasma was not changed significantly by 13-carotene (Tables 1 and 2). Little information has been reported concerning control of glucagon concentrations or even the glucagon profile itself in early lactation cows.
There was a trend for ratio of insulin to glucagon (Table 2) to increase over time al- though the increase by period was not signifi- cant. The change of ratio could be explained by declining milk production (Table 4) and,
TABLE 4. Average biweekly milk production (kg/day) of control cows and cows fed 3 carotene between 30 and 90 days of lactation.
3-Carotene Control Week X SE X SE
1 25.7 .9 25.2 .9 3 28.1 .9 27.6 .9 5 29.8 .9 30.1 1.0 7 29.9 .9 29.6 1.0 9 29.2 .8 28.9 1.1
11 28.6 .8 28.2 1.0 13 28.2 .8 28.0 1.0 Overall 28.5 .9 28.2 1.0
Journal of Dairy Science Vol. 67, No. 6, 1984

1254 BINDAS ET AL.
the re fore , r e t u r n to posi t ive energy balance. Smi th et al. (32) r epo r t ed insul in c o n c e n t r a t i o n s were posi t ively cor re la ted w i th days in to l ac ta t ion and feed in take and negat ively cor- re la ted wi th mi lk yield.
Mean somat ic cell c o u n t s were 113 -+ 15 (x 103 ) for s u p p l e m e n t e d cows c o m p a r e d to 126 -+ 14 (x 103 ) cel ls /ml for con t r o l cows. The on ly sources of var ia t ion t h a t were s ignif icant were cow and l ac t a t ion n u m b e r (P< .01) . Chew et al. (4), however , f o u n d s igni f icant d i f fe rences in v i t amin A and /3-carotene for cows where mas t i t i s was eva lua ted wi th Cal i forn ia Mast i t is Tes t scores. Cows wi th lower /3-carotene and v i t amin A in b lood p lasma had h igher scores t h a n cows showing h igher c o n c e n t r a t i o n s of ca ro t ene and v i t amin A. Effects of v i t amin A def ic iency, i.e., epi thel ia l degrada t ion , m a y n o t have been m a n i f e s t e d if v i tamin A was suff ic ient .
In conc lus ion , /3-carotene s u p p l e m e n t a t i o n did n o t improve r e p r o d u c t i o n n o r did it a l ter LH, p roges te rone , insulin, glucose, or g lucagon in b l o o d p lasma for dairy ca t t le in early l ac ta t ion . S u p p l e m e n t a t i o n did increase c o n c e n t r a t i o n s of /3-carotene in plasma. No p a t t e r n was d iscern ib le for LH in plasma, pe rhaps because of sampl ing f r equency . Proges te rone increased as n o r m a l r ep roduc t ive cycl ic i ty resumed. Insul in t e n d e d to increase as mi lk p r o d u c t i o n decl ined, and energy ba lance b e c a m e posit ive. Glucagon and glucose were n o t d i f fe ren t over t ime. Supple- m e n t a t i o n did n o t a f fec t somat ic cell coun t .
Thus, these resul ts suggest t h a t t3-carotene s u p p l e m e n t a t i o n is n o t w a r r a n t e d for Hols te in cows based on t he ra t ions fed and measures in this s tudy.
A C K N O W L E D G M E N T S
The au tho r s wish to express apprec i a t ion to C. N. Miller L. I. Eckler, D. L. Aalse th , and E. Read for s u p p o r t dur ing various aspects of the s tudy, H o f f m a n - L a R o c h e for provid ing the supp l emen ta l /3-carotene, and Vi rg in ia -Nor th Caro l ina select sires for par t ia l f inancia l suppor t .
REFERENCES
1 Barnes, M. A., G. W. Kazmer, G. P. Birrenkott, and L. W. Grimes. 1983. Induced gonadotropin release in adrenocorticotropin-treated bulls and steers. J. Anita. Sci. 56:155.
2 Bremel, D. H., R. W. Hemken, G. Heersche, Jr., L. A. Edgerton, and D. Olds. 1982. Effects of /3- carotene on metabolic and reproductive parameters
in lactating dairy cows. J. Dairy Sci. 65 (Suppl. 1): 178. (Abstr.)
3 Byers, J. H., I. R. Jones, and J. F. Bone. 1956. Carotene in the ration of dairy cattle. II. The influence of suboptimal levels of carotene intake upon the microscopic aspect of selected organs. J. Dairy Sci. 39:1556.
4 Chew, B. P., L. L. Hollen, J. K. Hillers, and M. L. Herlugson. 1982. Relationship between vitamin A and i3-carotene in blood plasma and milk and mastitis in Holsteins. J. Dairy Sci. 65:2111.
5 Converse, H. T., and E. B. Meigs. 1938. The carotene requirements for normal reproduction. J. Dairy Sci. 21:114.
6 Cooke, B. C. 1978. A study of the relationship between beta-carotene and fertility problems in dairy cows. Roche Symp., London.
7 Davis, R. E., and L. L. Madsen. 1941. Carotene and vitamin A in cattle blood plasma with observations on reproductive performance at restricted levels of carotene intake. J. Nutr. 21 : 135.
8 DeSilva, A.W.M.V., G. W. Anderson, F. C. Gwaz- dauskas, M. L. McGilliard, and J. A. Lineweaver. 1981. Interrelationships with estrous behavior and conception in dairy cattle. J. Dairy Sci. 64:2409.
9 Edgerton, L. A., and H. D. Hafs. 1973. Serum luteinizing hormone, prolactin, glucocorticoid, and progestin in dairy cows from calving to gestation. J. Dairy Sci. 56:451.
10 Fernandes, L. C., W. W. Thatcher, C. J. Wilcox, and E. P. Call. 1978. LH release in response to GnRH during the postpartum period of dairy cows. J. Anita. Sci. 46:443.
11 Feteris, W. A. 1965. A serum glucose method without protein precipitation. Am. J. Med. Technol. 31:17.
12 Folman, Y., I. Ascarelli, Z. Herz, M. Rosenberg, M. Davidson, and A. Halevi. 1979. Fertiity of dairy heifers given a commercial diet free of beta carotene. J. Nutr. 41:353.
13 Friesecke, H. 1978. The significance of beta carotene in ruminant nutrition: Field results - Europe. Roche Symp., London.
14 Goodale, W. S., H. A. Garverick, D. J. Kesler, C. J. Bierschwal, R. G. Elmore, and R. S. Youngquist. 1978. Transitory changes of hormones in plasma of postpartum dairy cows. J. Dairy Sci. 61:740.
15 Guilbert, H. R. 1936. Determination of carotene as a means of estimating the vitamin A value of forage. J. Dairy Sci. 19:55.
16 Harris, V., G. R. Fallona, and R. H. Unger. 1979. Glucagon. Pages 643 656 in Methods of hormone radioimmunoassay. 2nd ed. B. M. Jaffe and H. R. Behrman, ed., Academic Press, New York, NY.
17 Hart, I. C., J. A. Bines, S. V. Morant, and J. L. Ridley. 1978. Endocrine control of energy me- tabolism in the cow: Comparison of the levels of hormones and metabolites in the plasma of high- and low-yielding cattle at various stages of lactation. J. Endocrinol. 77:333.
18 lngalls, W. G., E. M. Convey, and H. D. Hafs. 1973. Bovine serum LH, GH, and prolactin during late pregnancy, parturition and early lactation. Proc. Soc. Exp. Biol. Med. 143:161.
Journal of Dairy Science Vol. 67, No. 6, 1984

REPRODUCTION AND fl-CAROTENE 1 2 5 5
19 Jordan, H. A., A. L. Neumann , G. S. Smith, J. E. Z immerman, and R. J. Vat thauer . 1961. Vitamin A status of steers fed "high ni t ra te" corn silages, and a s tudy of subsequent effects upon carotene utilization. J. Anim. Sci. 20:937.
20 Kane, E. A., and C. A. Cary. 1935. Effect of the condit ion of corn plant at cut t ing upon the carotene con ten t o f silage. J. Dairy Sci. 18:437.
21 Kimble, M. S., 1939. The photocolor imetr ic determinat ion o f vi tamin A and carotene in h u m a n plasma. J. Lab. Clin. Med. 24:1055.
22 Koprowski, J. A., and H. A. Tucker. 1973. Bovine serum growth hormone , corticoids and insulin during lactation. Endocrinology 93:645.
23 Kuhlman, A. H., and W. S. Gallup. 1942. Carotene (Provitamin A) requirements of dairy cattle for conception. J. Dairy Sci. 25:688.
24 Lot thammer , K. H. 1978. Impor tance and rope of beta carotene for bovine fertility original re- searches. Roche Symp., London.
25 McGillivray, W. A. 1960. Carotenoids and vitamin A in the blood of the bovine. N. Z. J. Agric. Res. 3:934.
26 Pelissier, C. L. 1976. Dairy cattle breeding problems and their consequences. Theriogenology 6:575.
27 Phillips, R. W., and V. N. Athanasiou. 1978. Stability of plasma metabol i tes and hormones in lactating dairy cows. Am. J. Vet. Res. 39:949.
28 Prior, R. L., and S. B. Smith. 1982. Hormonal effects on parti t ioning of nutr ients for tissue ~oxvth: Role of insulin. Fed. Proc. 40:2545.
29 Ronning, M., E. R. Berousek, A. H. Kuhlman, and W. D. Gallup. 1953. The carotene requirements for reproduct ion in Guernsey cattle. J. Dairy Sci.
36:52. 30 Semb, J., C. A. Baumann, and H. Steenbock. 1934.
The carotene and vitamin A conten t of colostrum. J. Biol. Chem. 107:697.
31 Smith, A. G., A. L. Neumann, W. G. Huber, H. A. Jordan, and O. B. Ross. 1961. Avitaminosis in cattle fed silage rat ions supplemented with vitamin A. J. Anita. Sci. 20:952.
32 Smith, R. D., W. Hansel, and C. E. Coppock. 1976. Plasma growth ho rmone and insulin during early lactation in cows fed silage based diets. J. Dairy Sci. 59:248.
33 Sut ton, T. S., H. E. Kaeser, and P. A. Soldner. 1945. Changes in the level of vitamin A and carotene in the blood plasma of dairy cows as- sociated with parturi t ion and beginning lactation. J. Dairy Sci. 28:933.
34 Sutton, T. S.. and P. A. Soldner. 1943. Carotene and vitamin A in the blood plasma of dairy cows. J. Dairy Sci. 26:740.
35 Thompson , S. Y., and W. A. McGillivray. 1956. Changes in vitamin A and carotenoid content of the blood plasma of cows and heifers near parturi- tion in relation to secretion of these substances in the milk fat. Dairy Res. 24:108.
36 Turner, C. W. 1938. Relation of nutr i t ion to the hormones. J. Dairy Sci. 21:99.
37 Webb, R., G. E. Lamming, N. B. Haynes, and G. R. Foxcroft . 1980. Plasma progesterone and gonado- tropin concentra t ions and ovarian activity in post -par tum dairy cows. J. Reprod. Fertil. 59:133.
38 Whitanore, H. L., W. J. Tyler, and L. E. Casida. 1974. Effects of early pos tpa r tum breeding in dairy cattle. J. Anim. Sci. 38:339.
Journal o f Dairy Science Vol. 67, No. 6, 1984





























