Embed Size (px)
Citation preview

Retrovirus e cancerogenesi
HTLV-1 e HTLV-2
www.fisiokinesiterapia.biz

I retrovirus
• 1911: Virus del Sarcoma di Rous
• 1970: trascrittasi inversa
Francis Peyton Rous (1879-1970)
Fonti: La Placa, Principi di Microbiologia Medica, X Edizione. Esculapio Ed.

Rapporto virus-cellula
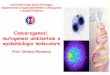
•Il rapporto virus-cellula infetta può sfociare in:
•una infezione citocida (malattia acuta) • latente (malattia recidivante)•persistente (malattia cronica)
Infezione citocidaIn vitro
Degenerazione cellulare
In vivo
Inclusioni cellulari
degenerazione spongiforme
Infezione latenteIl virus realizza un rapporto
di parassitismo controllatocon continua produzione
di antigeni virali.Le lesioni cellulari
sono dovutead azioni lesive
esercitate in tempi più o meno lunghi
dalle risposte immunitariedell’ospite
Infezione latenteIl virus realizza
un rapporto di parassitismocontrollato,
che consente alle cellulenon solo di sopravvivere,ma anche di duplicarsi
Il genoma virale puòessere integrato
con quello della cellula ospite.Il virus permane nella
cellula in forma criptica
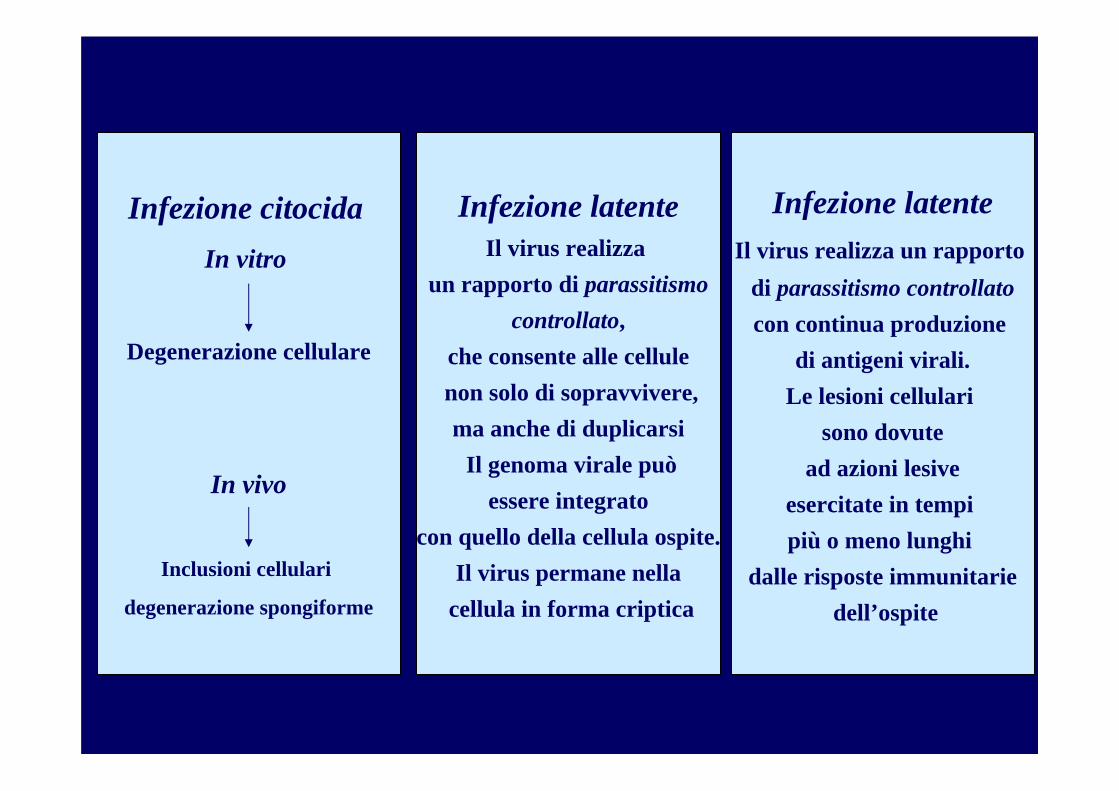
La produzione di malattia è un evento raro
Risposta ospite
Lisi cellulare Decesso
Formazione inclusi virali
Trasformazione cellulare
Disfunzioni cellulari
Malattia
Risposta cellulare
Moltiplicazione virale in assenza di lesioniCiclo abortivo
Esposizione senza penetrazione
Infezione asintomatica
Concetto iceberg
Esposizione senza infezione

Rapporto virus-cellula
•Il rapporto virus-cellula infetta può sfociare in:•una infezione citocida (malattia acuta) • latente (malattia recidivante)•persistente (malattia cronica)
•trasformazione neoplastica
I virus oncogeni hanno la capacità di trasformare (immortalizzare) cellule normali in colture in vitro,
bloccandone la differenziazione e conferendo loro numerose proprietà della cellula neoplastica.

•Virus in grado di indurre o di concorrere alla comparsa di alterazioni del ciclo cellulare.
L’oncogenesi è un processo graduale: non sempre la formazione del tumore rappresenta una inevitabile conseguenza dell’infezione virale.
L’oncogenesi prevede una serie di cambi indipendenti ma irreversibili che insieme contribuiscono alla dis-regolazione della crescita tumorale. L’infezione virale rappresenta uno di questi step. E solo se succedono altri eventi nella stessa cellula, assisteremo allo sviluppo del cancro.
I virus oncogeni

I meccanismi con cui un virus provoca un tumore sono molto divergenti, ma hanno tutti alcune caratteristiche in comune…
a) una singola particella virale è sufficiente per la trasformazione. Non ènecessaria una infezione multipla della stessa cellula.
b) tutto o parte del genoma persiste nella cellula trasformata. Comunque, spesso non c’è produzione di progenie infettante.
c) in tutti i casi di trasformazione indotta del virus, almeno una parte delgenoma viene espresso nelle cellule trasformate
d) la trasformazione è il risultato della rottura dei segnali normali di crescita cellulare.
Oncogenesi virale: quali meccanismi?

Caratteristiche delle cellule neoplastiche
• Immortalizzazione
• Aumento dell'attività telomerasica: nella cellula normale la telomerasi èrepressa. La lunghezza del telomero è un orologio biologico, che ci dice quanto la cellula può arrivare a vivere. Quando i telomeri sono troppo corti, la cellula non è più in grado di replicarsi. La telomerasi è attiva nelle cellule staminali
• Mancanza della inibizione da contatto• Glicolisi aerobia aumentata (la produzione di ATP è aumentata)• Presentazione di nuovi antigeni (presenti solo nella vita fetale)• Genoma virale totalmente o parzialmente integrato• Produzione di proteine precoci• Corredo cromosomico alterato
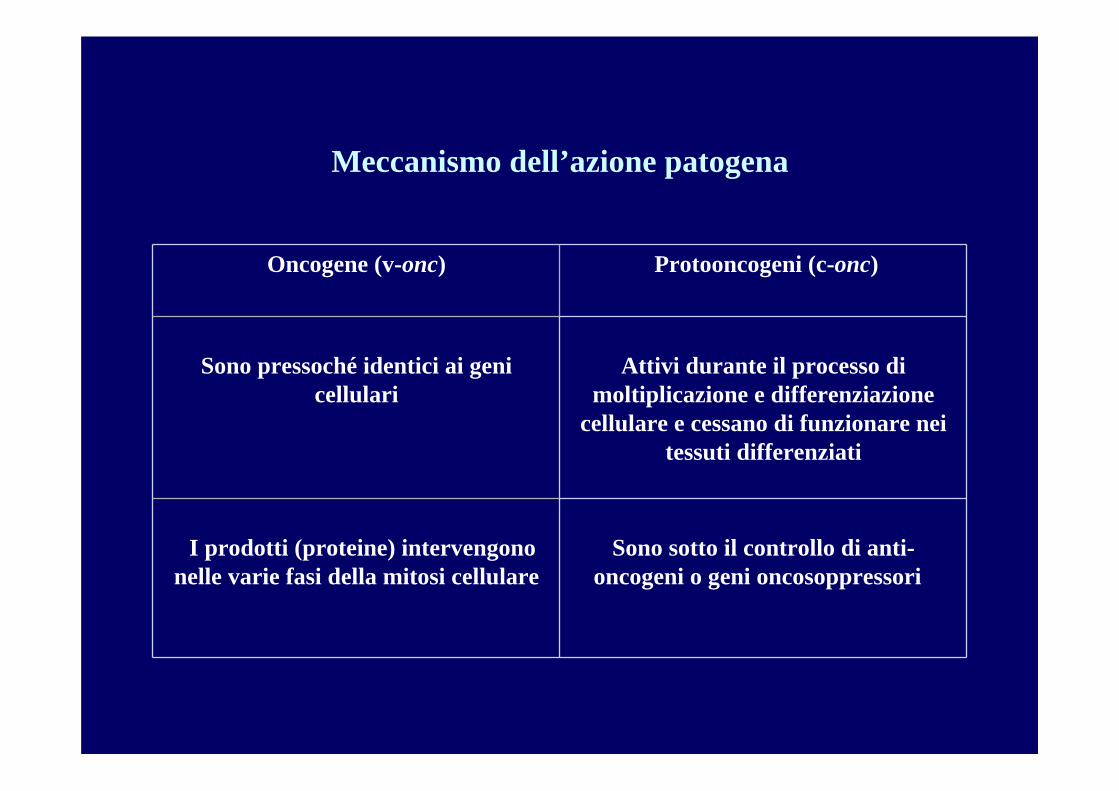
Meccanismo dell’azione patogena
Attivi durante il processo di moltiplicazione e differenziazione
cellulare e cessano di funzionare nei tessuti differenziati
Sono pressoché identici ai geni cellulari
Sono sotto il controllo di anti-oncogeni o geni oncosoppressori
I prodotti (proteine) intervengono nelle varie fasi della mitosi cellulare
Protooncogeni (c-onc)Oncogene (v-onc)
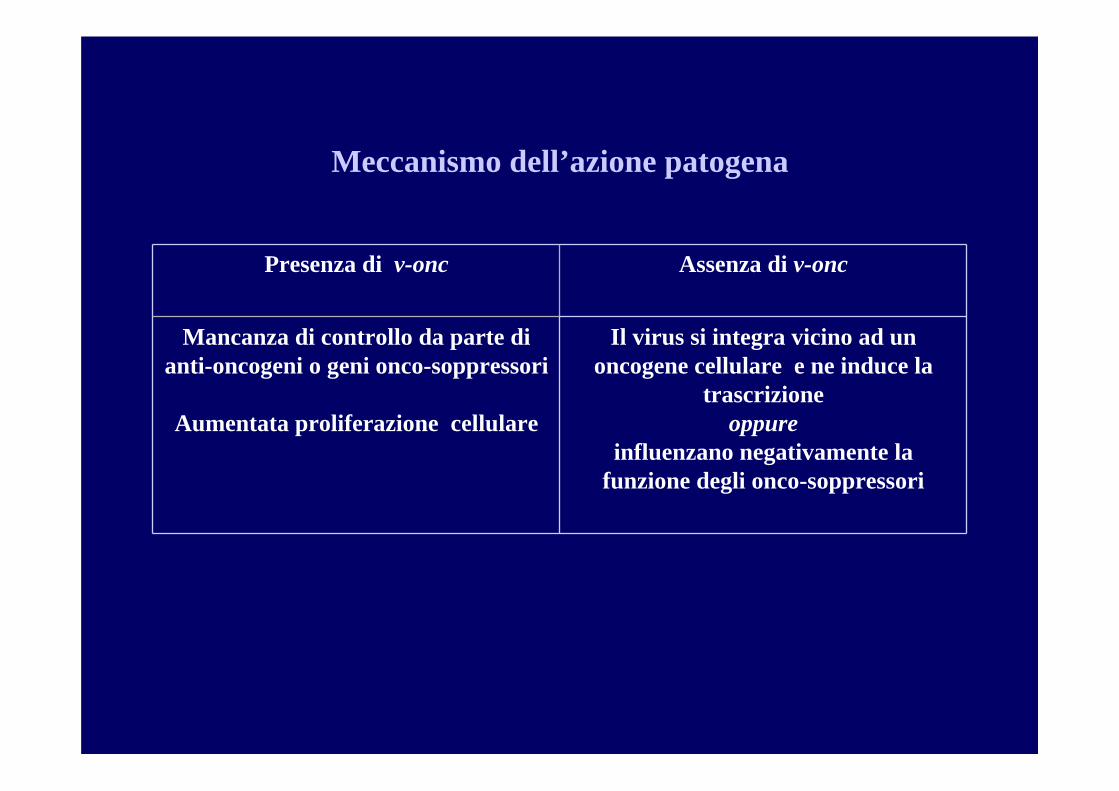
Meccanismo dell’azione patogena
Il virus si integra vicino ad un oncogene cellulare e ne induce la
trascrizione oppure
influenzano negativamente la funzione degli onco-soppressori
Mancanza di controllo da parte di anti-oncogeni o geni onco-soppressori
Aumentata proliferazione cellulare
Assenza di v-oncPresenza di v-onc

Human pathogenic retroviruses
Lentivirinae
Human immunodeficiency virus type 1
Human immunodeficiency virus type 2
Oncovirinae
Human T-lymphotropic virus type 1 (HTLV-1)
Human T-lymphotropic virus type 2 (HTLV-2)
Human T-lymphotropic virus type 3 (HTLV-3) simian origin
Human T-lymphotropic virus type 4 (HTLV-4) no known primate counterpart
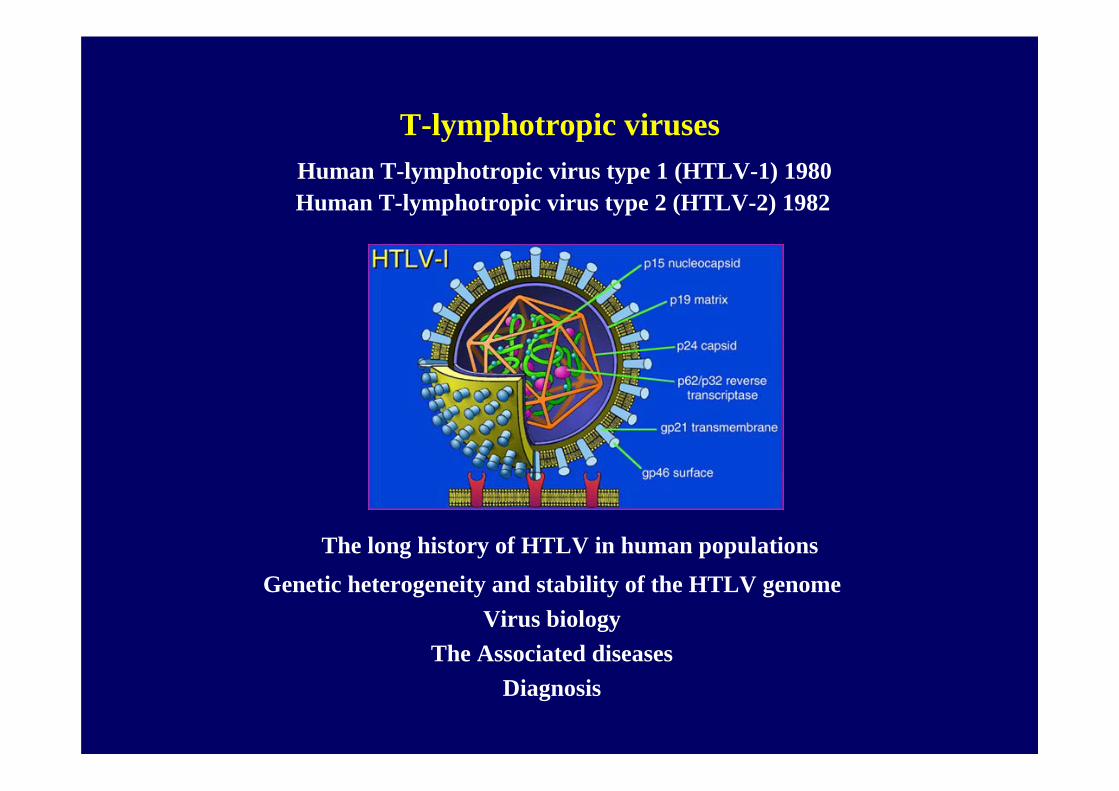
T-lymphotropic virusesHuman T-lymphotropic virus type 1 (HTLV-1) 1980Human T-lymphotropic virus type 2 (HTLV-2) 1982
The long history of HTLV in human populationsGenetic heterogeneity and stability of the HTLV genome
Virus biologyThe Associated diseases
Diagnosis
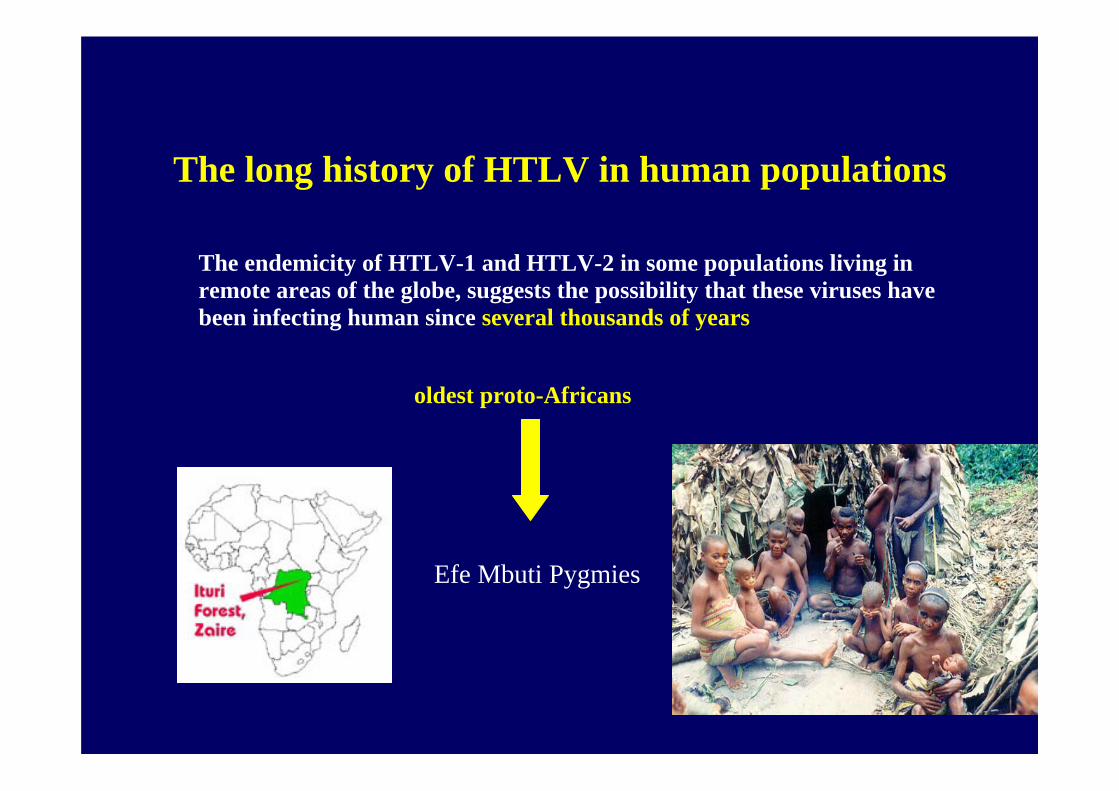
The long history of HTLV in human populations
The endemicity of HTLV-1 and HTLV-2 in some populations living in remote areas of the globe, suggests the possibility that these viruses have been infecting human since several thousands of years
oldest proto-Africans
Efe Mbuti Pygmies

The long history of HTLV in human populations
HTLV-1 and HTLV-2 infections among Pygmies are
ancient origin result of more recent interspecies transmission of STLVs:
jumping from primates to humans
STLV-1 infects most primate species in Africa and Asia and
has been associated with lymphoma and leukemia
PTLV-I: HTLV-1 and STLV-IPTLV-II : HTLV-2/STLV-II

Stability of the HTLV genome
RNA viruses evolve at the rate of 10-3 to 10-5 nucleotide substitution per site per year
but
HTLV-1 and HTLV-2 have a very stable genomeWhy?
• HTLVs are slowly replicating viruses• HTLV mutation rates are unusually low, due to a high fidelity reverse
transcriptase• After the integration in the host genome, HTLVs predominantly propagate as
proviral DNA during cellular mitosis using the host cell machinery.

HTLV-1 and 2 subtypes
• HTLV-1a, the Cosmopolitan subtype, joins strains from different geographic regions
• HTLV-1b, also known as Central African subtype, joins strains isolated from Central Africa populations
• HTLV-1c is the Melanesian subtype to which belong the divergent strains isolated in Papua New Guinea and from Australian aboriginals
• HTLV-1d has been recently described as a new distinct molecular subtype isolated from Cameroonian pygmies and from an infected Gabonese individual
• HTLV-2 isolates: prevalent in Southeastern Europe
• HTLV-2a subtypes: prevalent in Northern European IDUs, such as in UK, Ireland and Sweden
• HTLV-2b subtypes: isolated in Florida, Panama, Colombia, Argentina, Paraguay, Chile
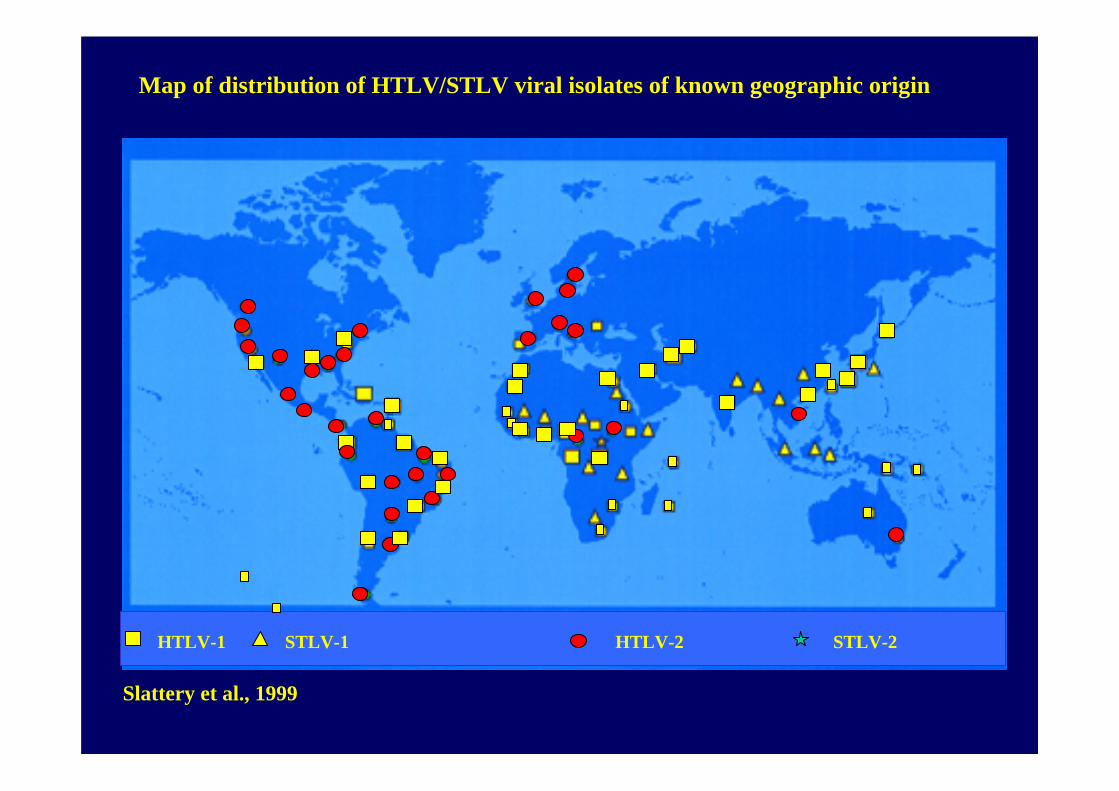
HTLV-1 STLV-1 HTLV-2 STLV-2
Slattery et al., 1999
Map of distribution of HTLV/STLV viral isolates of known geographic origin

HTLVs Biology
HTLVs have a central icosahedral nucleocapsid surrounded by a round shaped envelope and the virions are seen as spherical
particles of about 100 nm in diameter
HTLV 1 and 2: same genomic organization and up to 65-70% of nucleotide similarity
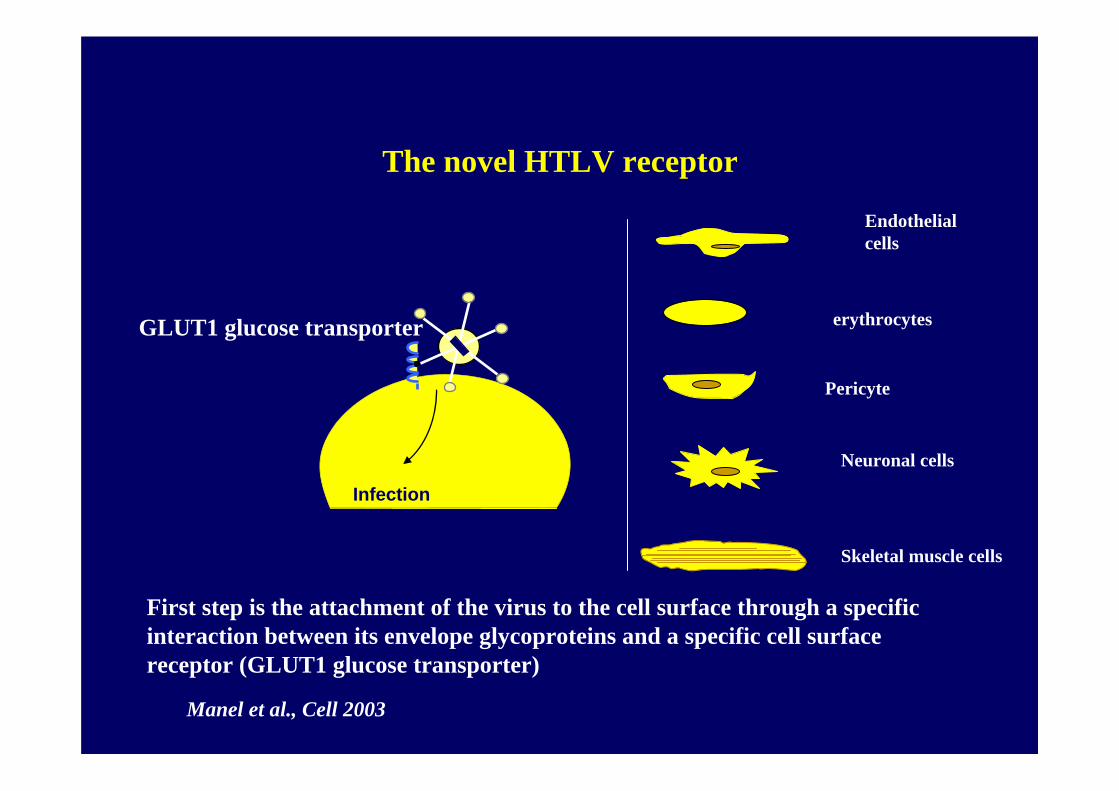
GLUT1 glucose transporter
First step is the attachment of the virus to the cell surface through a specific interaction between its envelope glycoproteins and a specific cell surface receptor (GLUT1 glucose transporter)
The novel HTLV receptor
Infection
Endothelial cells
erythrocytes
Pericyte
Skeletal muscle cells
Neuronal cells
Manel et al., Cell 2003

Glucose Transporter GLUT-1 and the paradox
The apparently restricted tropism of HTLV to T lymphocytes in infected patients contrasts with the ability of the viral-encoded envelope glycoprotein (Env) to bind to and direct entry into all vertebrate cell types tested in vitro
In vitroSeveral different cell types can be infected
In vivoHTLV-1 and 2 can infect CD4+ and CD8+ T-cells
HTLV-1 primarily has a CD4+ tropism HTLV-2 primarily has CD8+ tropism.
…….what happened during the first phase of infection?
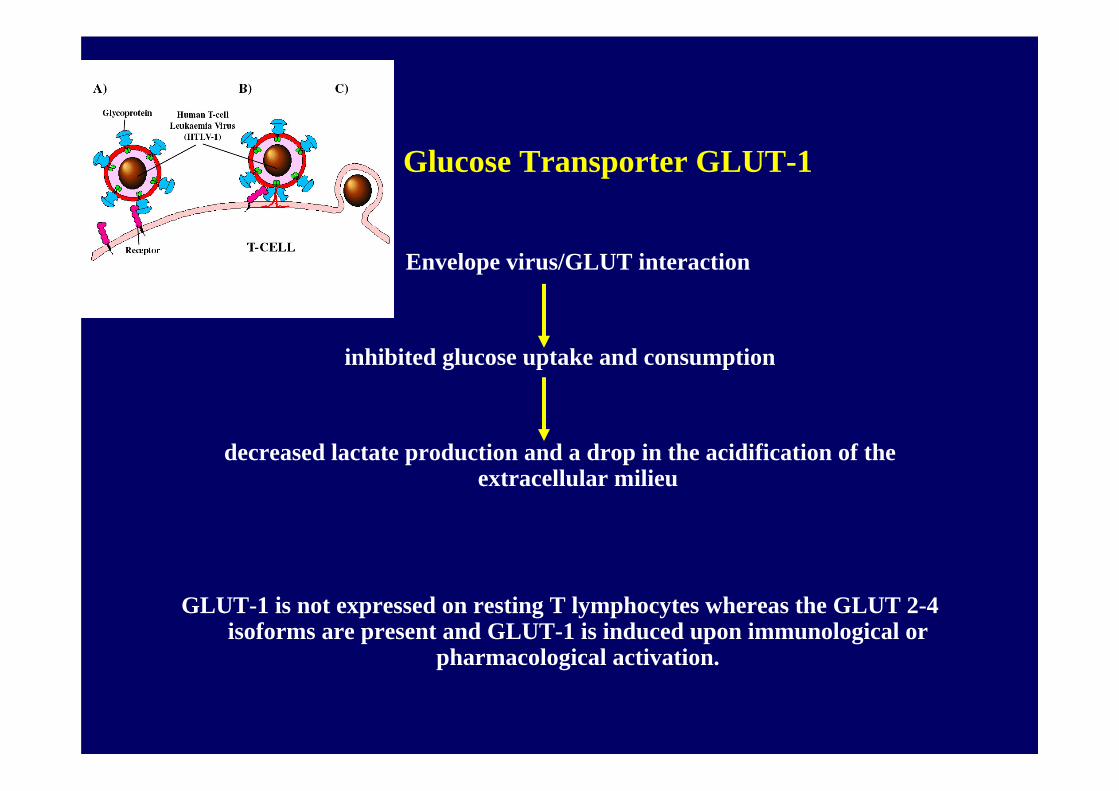
Glucose Transporter GLUT-1
Envelope virus/GLUT interaction
inhibited glucose uptake and consumption
decreased lactate production and a drop in the acidification of the extracellular milieu
GLUT-1 is not expressed on resting T lymphocytes whereas the GLUT 2-4 isoforms are present and GLUT-1 is induced upon immunological or
pharmacological activation.
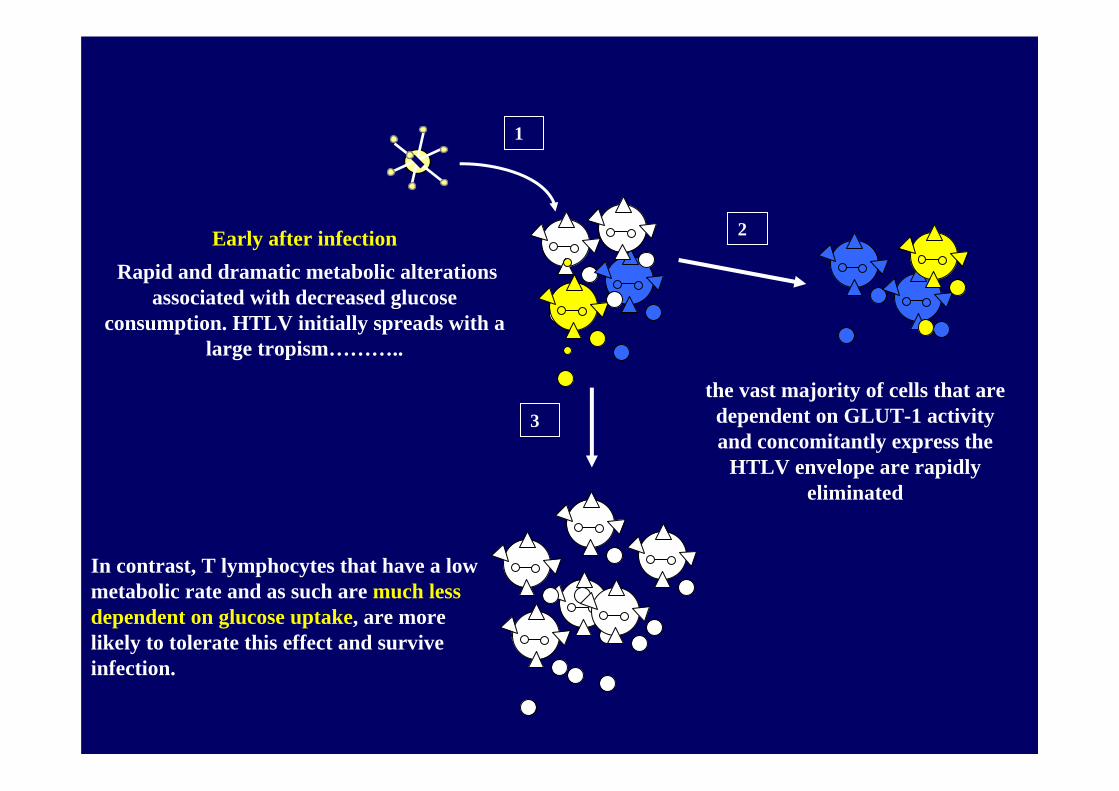
Early after infectionRapid and dramatic metabolic alterations
associated with decreased glucose consumption. HTLV initially spreads with a
large tropism………..
the vast majority of cells that are dependent on GLUT-1 activity and concomitantly express the
HTLV envelope are rapidly eliminated
In contrast, T lymphocytes that have a low metabolic rate and as such are much less dependent on glucose uptake, are more likely to tolerate this effect and survive infection.
1
2
3
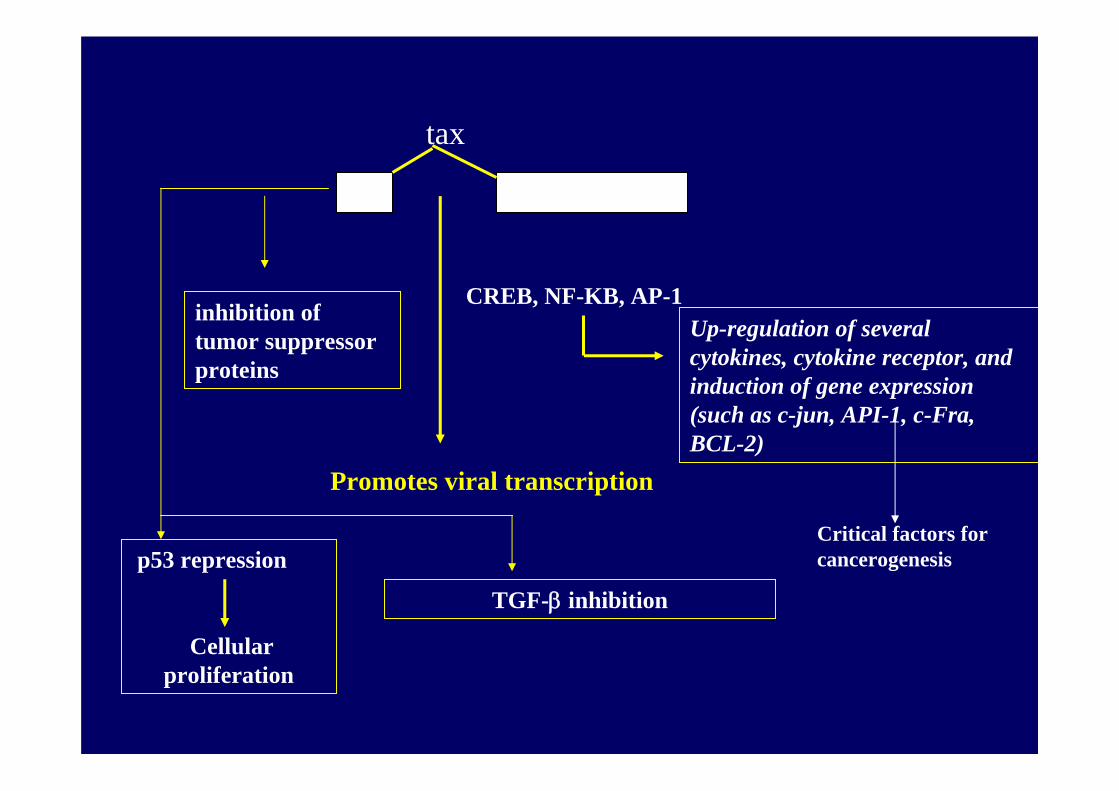
tax
CREB, NF-KB, AP-1
Promotes viral transcription
Up-regulation of several cytokines, cytokine receptor, and induction of gene expression (such as c-jun, API-1, c-Fra, BCL-2)
p53 repression
Cellular proliferation
TGF-β inhibition
Critical factors for cancerogenesis
inhibition of tumor suppressor proteins
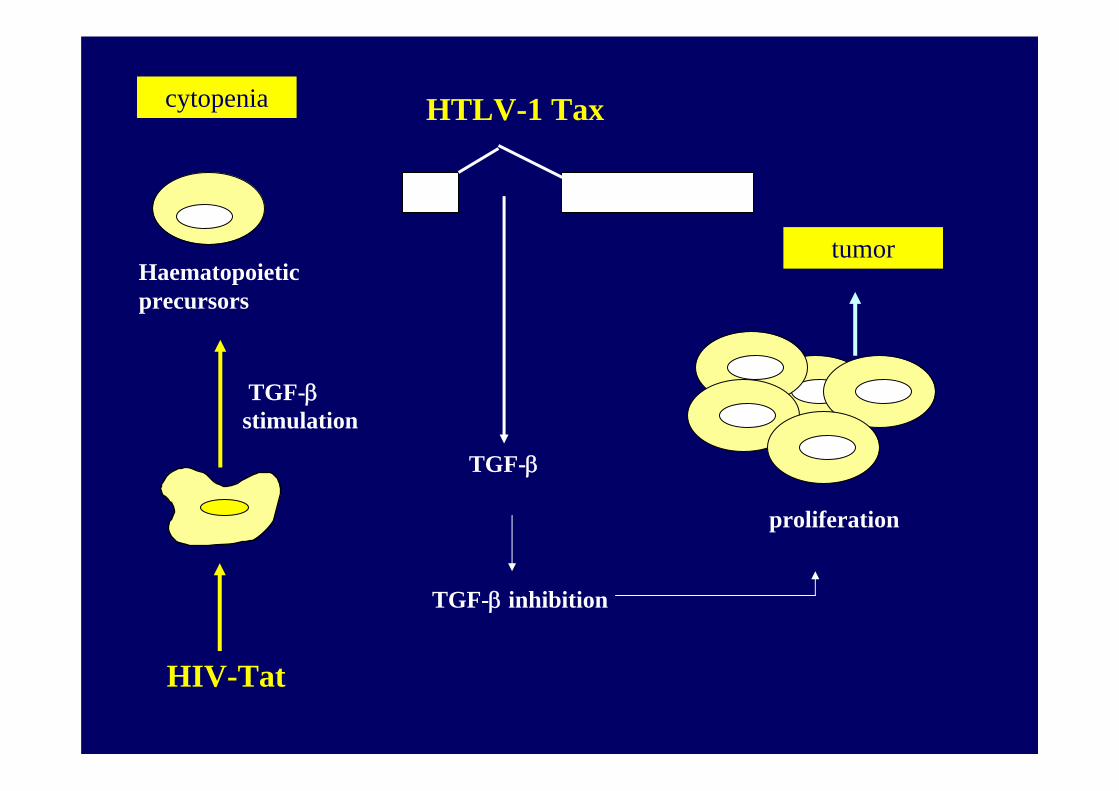
HTLV-1 Tax
TGF-β inhibition
TGF-β
Haematopoieticprecursors
proliferation
HIV-Tat
cytopenia
tumor
TGF-β stimulation
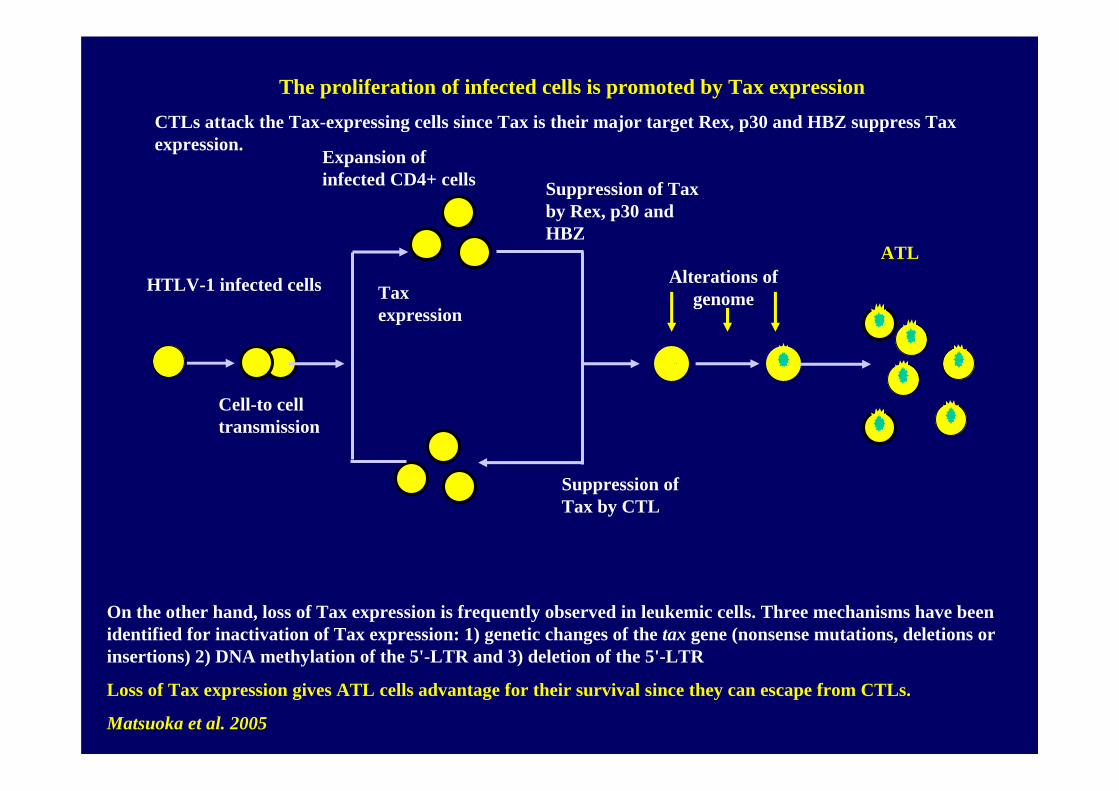
Cell-to cell transmission
Expansion of infected CD4+ cells Suppression of Tax
by Rex, p30 and HBZ
Suppression of Tax by CTL
Alterations of genome
ATL
HTLV-1 infected cells
On the other hand, loss of Tax expression is frequently observed in leukemic cells. Three mechanisms have been identified for inactivation of Tax expression: 1) genetic changes of the tax gene (nonsense mutations, deletions or insertions) 2) DNA methylation of the 5'-LTR and 3) deletion of the 5'-LTR
Loss of Tax expression gives ATL cells advantage for their survival since they can escape from CTLs.
Matsuoka et al. 2005
The proliferation of infected cells is promoted by Tax expressionCTLs attack the Tax-expressing cells since Tax is their major target Rex, p30 and HBZ suppress Tax expression.
Tax expression

HTLV-1 and associated diseases
• Adult T-cell Leukemia (ATL)
• The lifetime risk of developing ALT in HTLV-1 seropositive individuals in about 0.1% to 5%
• The latency period is usually more than 20 years
ATL is classified in four clinical stagesLymphoma
SmoulderingChronic
Acute type

HTLV-1 and associated diseases
Tropical spastic paraparesis/HTLV associated myelopathy (TSP/HAM)
• A slowly progressive myelopathy with spastic paraparesis of lower extremities
• Moreover…. variable peripheral sensory abnormalities, hyperreflexia, bladder and bowel dysfunction, male impotence.
• The lifetime risk of developing the disease in HTLV-1 seropositivepatients is less than 1%, with a mean age of onset of about 40 years
• The incubation time can greatly vary and it can be as short as a few weeks in patients who get the infection from blood transfusions.

HTLV-2 associated diseases
Firstly isolated from a patient with hairy-cell leukaemia ………………….
Most of the HTLV-2 infected individuals are asymptomatic during their entire life.
Recently there have been increasing evidences that the infectionmay be associated with neurological disorders
HTLV-2 RNA is found in HAM-like lesions of the central nervous system
Moreover, an increased prevalence of infectious diseases, such as pneumonia, minor fungal infection or kidney infection, has been associated with the presence of HTLV-2
HTLV-2 is present at significant rate among injecting drug users (IDUs) co-infected by HIV-1
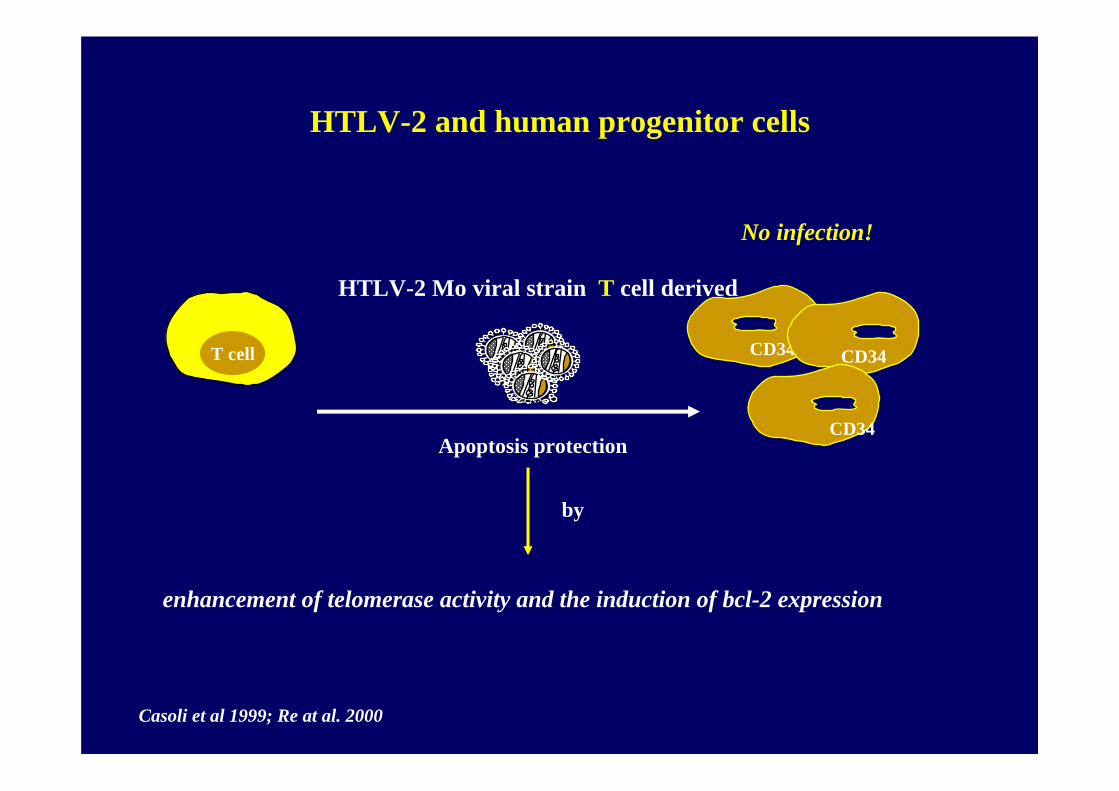
No infection!
HTLV-2 Mo viral strain T cell derived
CD34T cell CD34
Apoptosis protectionCD34
Casoli et al 1999; Re at al. 2000
enhancement of telomerase activity and the induction of bcl-2 expression
CD34
by
HTLV-2 and human progenitor cells

HTLV-2 and HIV-1. Does an interaction exist?
• HTLV-II can interfere with the replicative potential of HIV-1 by up-regulating viral suppressive CC-chemokines and, in particular, MIP-1alpha.
• HTLV-II can influence HIV replication, at least in vitro, via up-regulation of HIV-suppressive chemokines
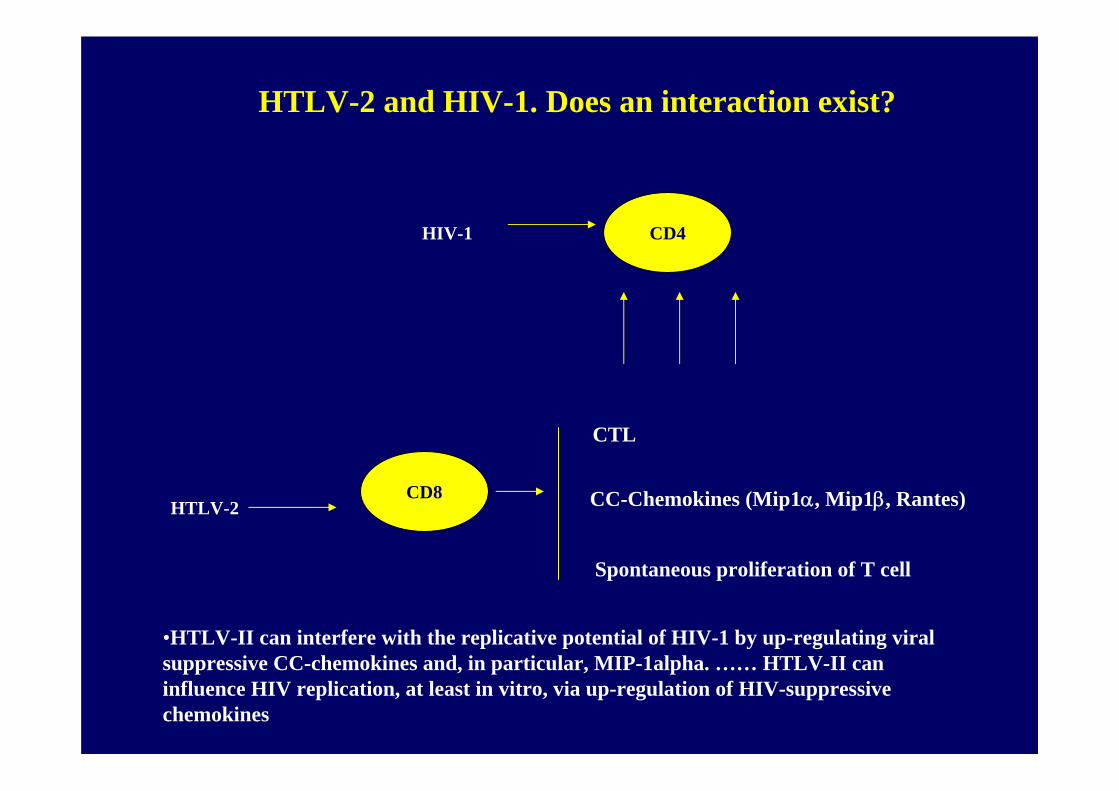
CD4
CD8
HIV-1
HTLV-2
Spontaneous proliferation of T cell
CC-Chemokines (Mip1α, Mip1β, Rantes)
CTL
HTLV-2 and HIV-1. Does an interaction exist?
•HTLV-II can interfere with the replicative potential of HIV-1 by up-regulating viral suppressive CC-chemokines and, in particular, MIP-1alpha. …… HTLV-II can influence HIV replication, at least in vitro, via up-regulation of HIV-suppressive chemokines
HTLV-1 and 2: diagnosis of infection
• Immunoenzymatic assay• Immunoblotting• Provirus detection (qualitative and/or quantitative PCR)

HTLV-1 and 2: diagnosis of infection
• Immunoblotting: The first antibodies, which appeared within 2 months after exposure, were directed against rgp21 and were followed by antibodies to p24 and p19.
• the Public Health Service Working Group has recommended that a serum should be considered as HTLV-antibody positive when reactive to p24 and to gp46 or gp61/68

HTLV-1 and 2: diagnosis of infection
Blood donor screening for HTLV was introduced in:
• 1980s: first in Japan• 1988 - 1989 in the United States and Canada • 1991 in France
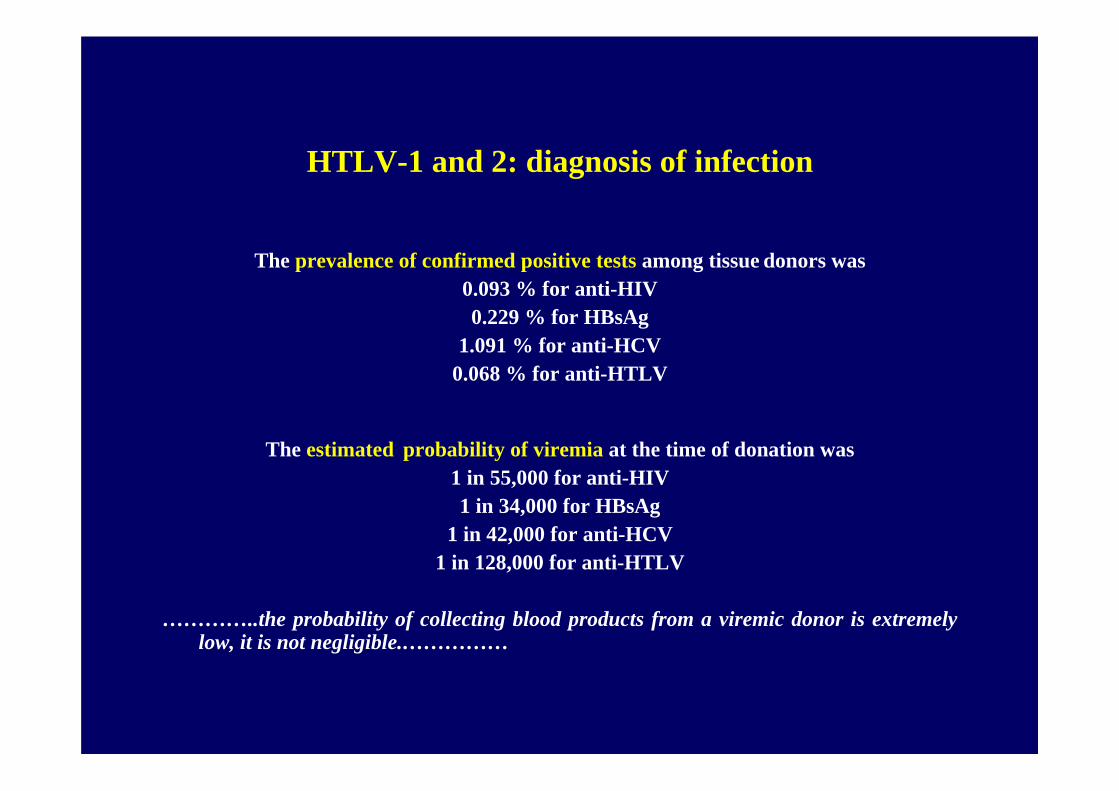
HTLV-1 and 2: diagnosis of infection
The prevalence of confirmed positive tests among tissue donors was 0.093 % for anti-HIV0.229 % for HBsAg
1.091 % for anti-HCV0.068 % for anti-HTLV
The estimated probability of viremia at the time of donation was 1 in 55,000 for anti-HIV1 in 34,000 for HBsAg
1 in 42,000 for anti-HCV1 in 128,000 for anti-HTLV
…………..the probability of collecting blood products from a viremic donor is extremely low, it is not negligible.……………

An imported case of adult T cell leukemia in a HTLV-I-infected patient, in Italy
•From an epidemiological point of view, it is important to report a case of imported ATL in a non-endemic area, as it may explain the otherwise untraceable origin of some rare and apparently autochthonous cases of ATL in non-endemic areas ………………… Re et al. 2003 and 2004