Embed Size (px)
Citation preview

The dynamic range of burstingin a network of
respiratory pacemaker cells
Alla Borisyuk
Universityof Utah

Joint work with:
Janet BestJonathan RubinDavid Terman
Martin Wechselberger
Mathematical Biosciences Institute (MBI), OSU
Biological data
Existing model
Previously…
Biological data
Existing model
Numerical simulations
Observations(Predictions)
Previously…
Biological data
Existing model
Numerical simulations
Observations(Predictions)
In this project
Mathematicalstructure
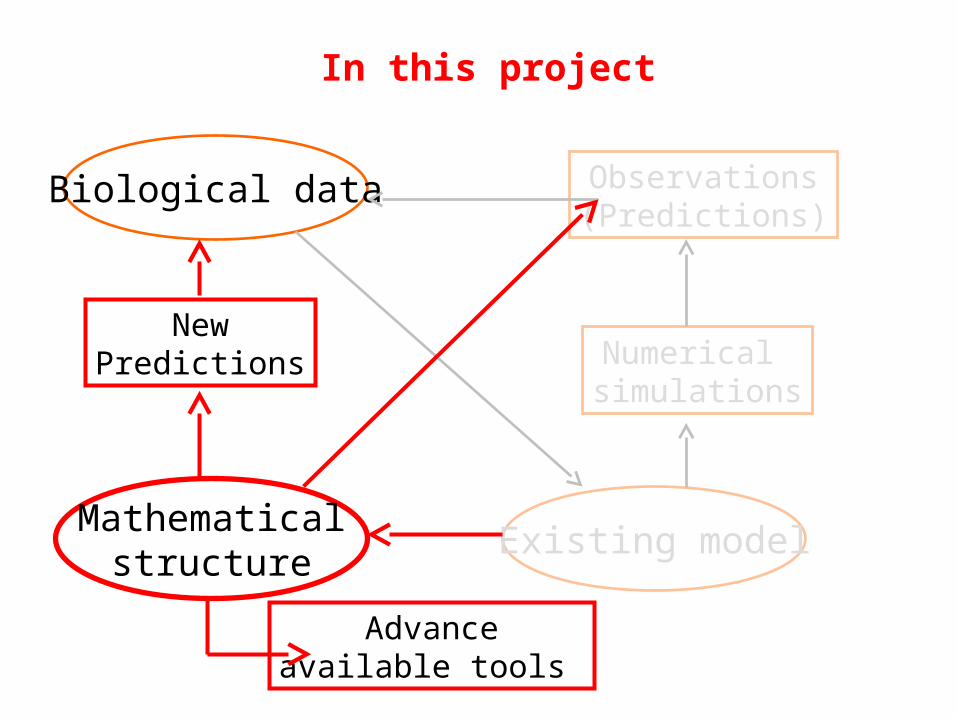
Biological data
Existing model
Numerical simulations
Observations(Predictions)
In this project
Mathematicalstructure
Advanceavailable tools
NewPredictions
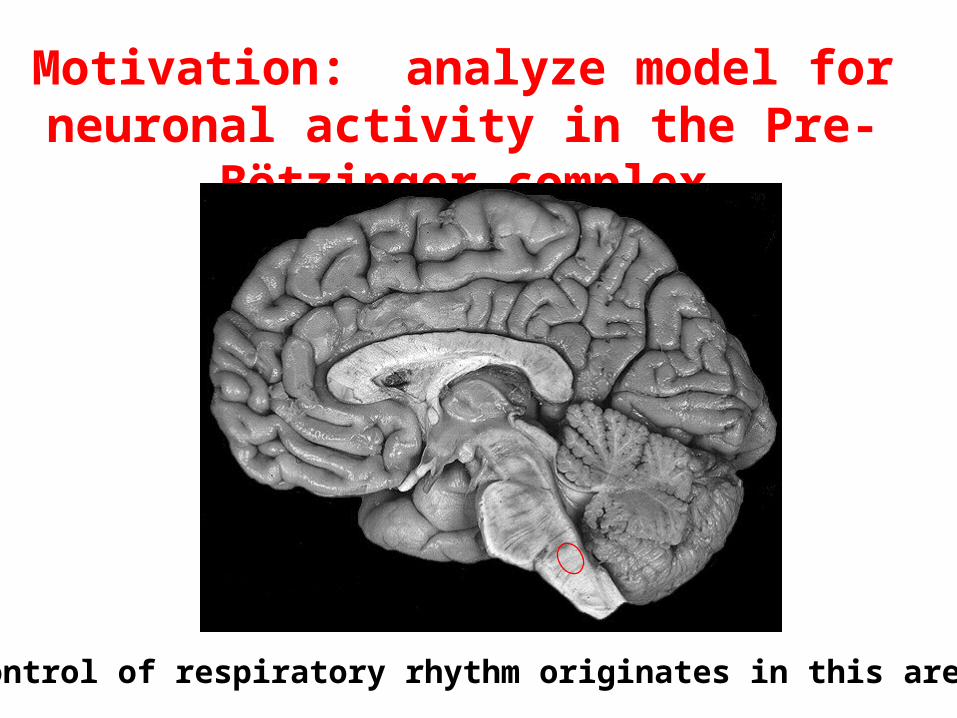
Motivation: analyze model for neuronal activity in the Pre-Bötzinger complex
Control of respiratory rhythm originates in this area
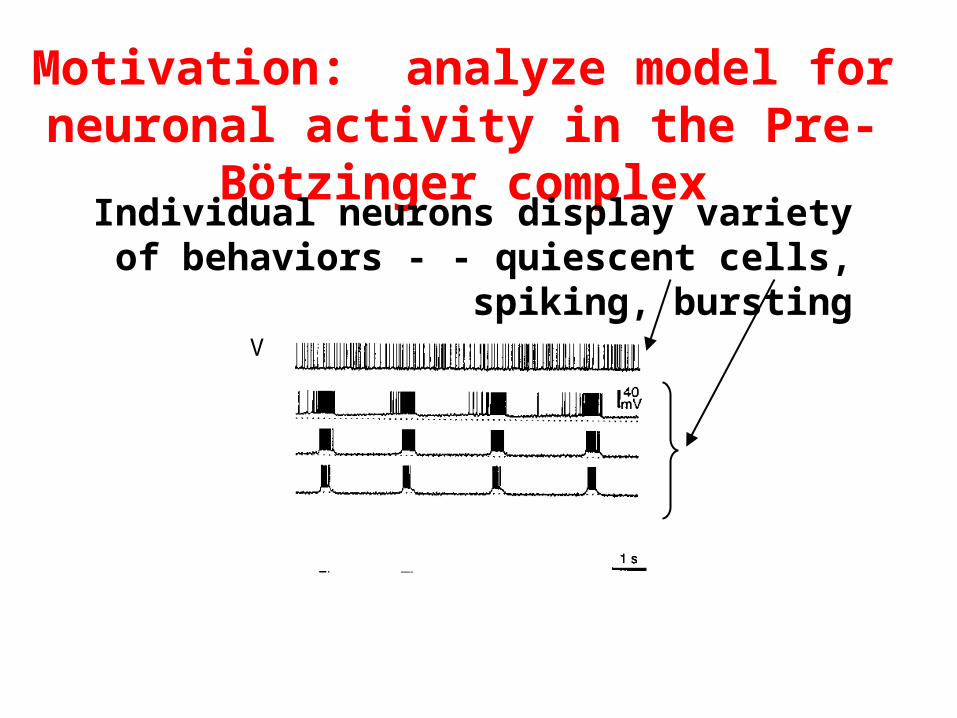
Motivation: analyze model for neuronal activity in the Pre-Bötzinger complex
Individual neurons display variety of behaviors - - quiescent cells, spiking, bursting
V

Motivation: analyze model for neuronal activity in the Pre-Bötzinger complex
Population exhibits synchronous rhythms
figure
Question: How can a synchronous network bursting be supported by heterogeneous (e.g. spiking) cells?
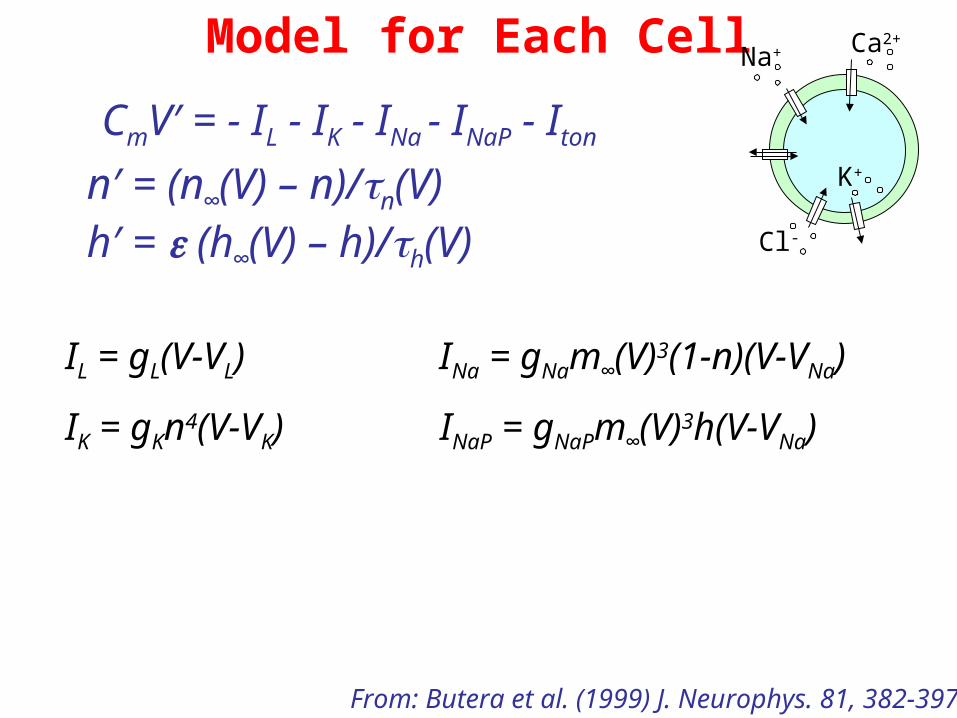
Model for Each Cell
IL = gL(V-VL) INa = gNam∞(V)3(1-n)(V-VNa)
IK = gKn4(V-VK) INaP = gNaPm∞(V)3h(V-VNa)
n′ = (n∞(V) – n)/n(V)h′ = (h∞(V) – h)/h(V)
CmV′ = - IL - IK - INa - INaP - Iton
From: Butera et al. (1999) J. Neurophys. 81, 382-397
Na+ Ca2+
K+
Cl-

Model for Each Cell
IL = gL(V-VL) INa = gNam∞(V)3(1-n)(V-VNa)
IK = gKn4(V-VK) INaP = gNaPm∞(V)3h(V-VNa)
Iton(V) = gton(V-Vsyn) - Input from other brain areas
n′ = (n∞(V) – n)/n(V)h′ = (h∞(V) – h)/h(V)
CmV′ = - IL - IK - INa - INaP - Iton
From: Butera et al. (1999) J. Neurophys. 81, 382-397
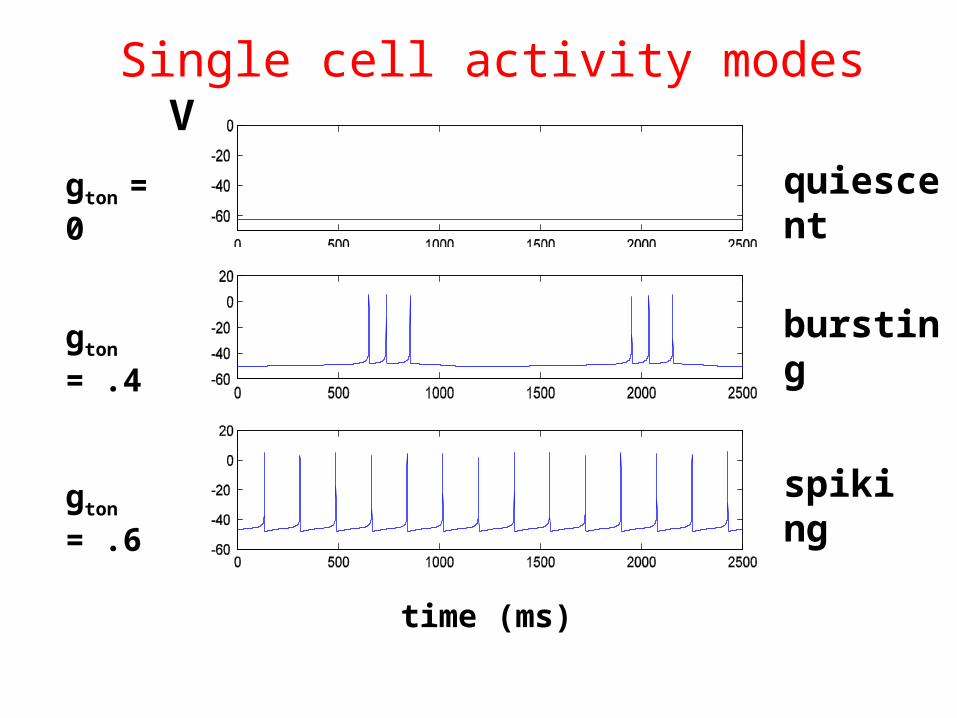
V
time (ms)
quiescent
bursting
spiking
gton = 0
gton = .4
gton = .6
Single cell activity modes

n′ = (n∞(V) – n)/n(V)h′ = (h∞(V) – h)/h(V)si′ = (1-si)H(Vi-)-si
CmV′ = - IL - IK - INa - INaP - Iton - Isyn
Coupling the neurons
Isyn = gsyn( si)(V-Vsyn) - Input from other network cells
From: Butera et al. (1999) J. Neurophys. 81, 382-397
s1
s2

n′ = (n∞(V) – n)/n(V)h′ = (h∞(V) – h)/h(V)si′ = (1-si)H(Vi-)-si
CmV′ = - IL - IK - INa - INaP - Iton - Isyn
Coupling the neurons
From: Butera et al. (1999) J. Neurophys. 81, 382-397
gsyn =0 individual cells
Isyn = gsyn( si)(V-Vsyn) - Input from other network cells

n′ = (n∞(V) – n)/n(V)h′ = (h∞(V) – h)/h(V)si′ = (1-si)H(Vi-)-si
CmV′ = - IL - IK - INa - INaP - Iton - Isyn
Full system
Iton = gton(V-Vsyn) Isyn = gsyn( si)(V-Vsyn)
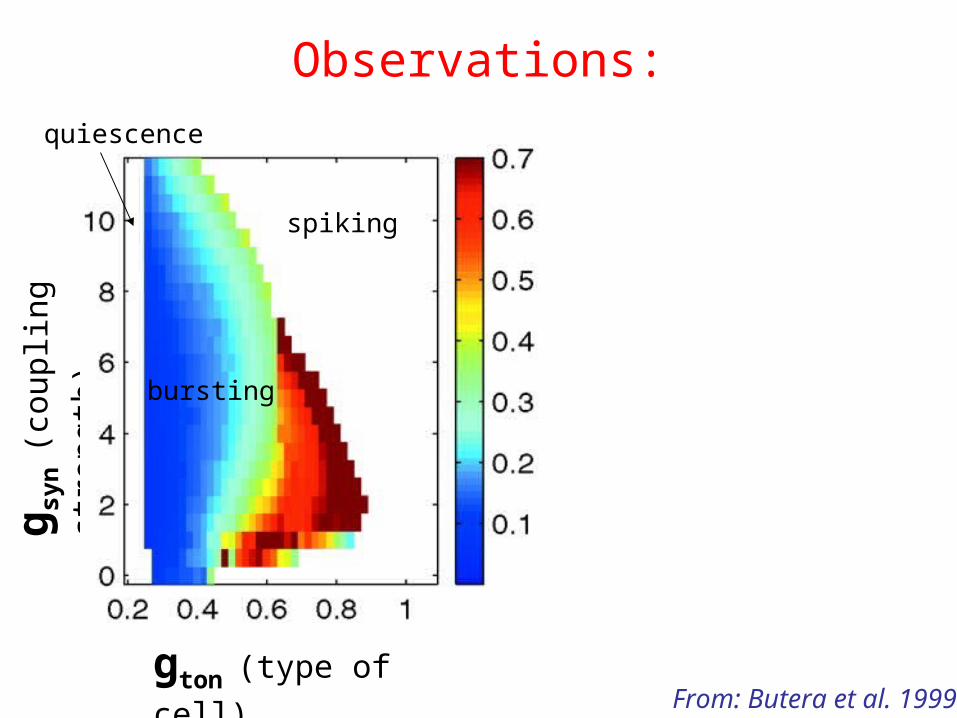
Observations:
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
bursting
spiking
quiescence
From: Butera et al. 1999
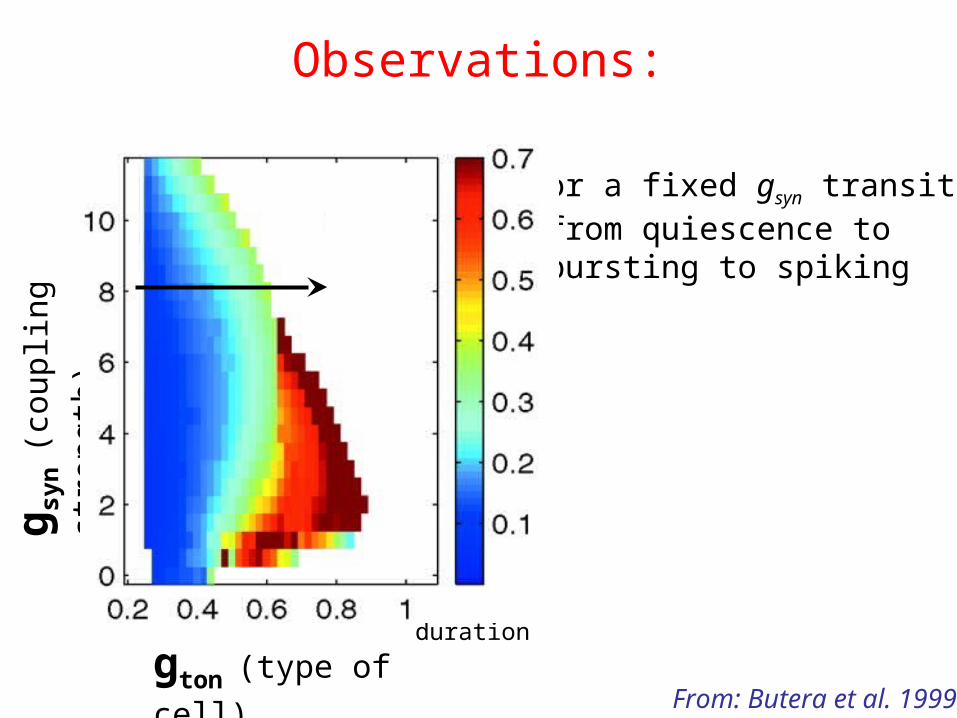
Observations:
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
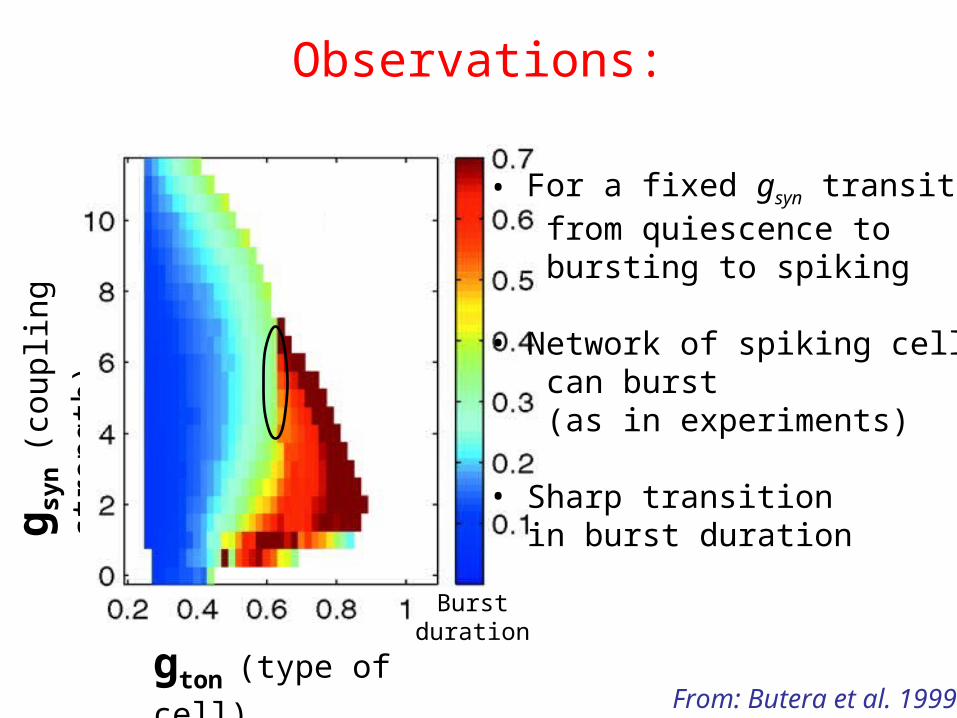
• For a fixed gsyn transitions from quiescence to bursting to spiking
Burstduration
From: Butera et al. 1999
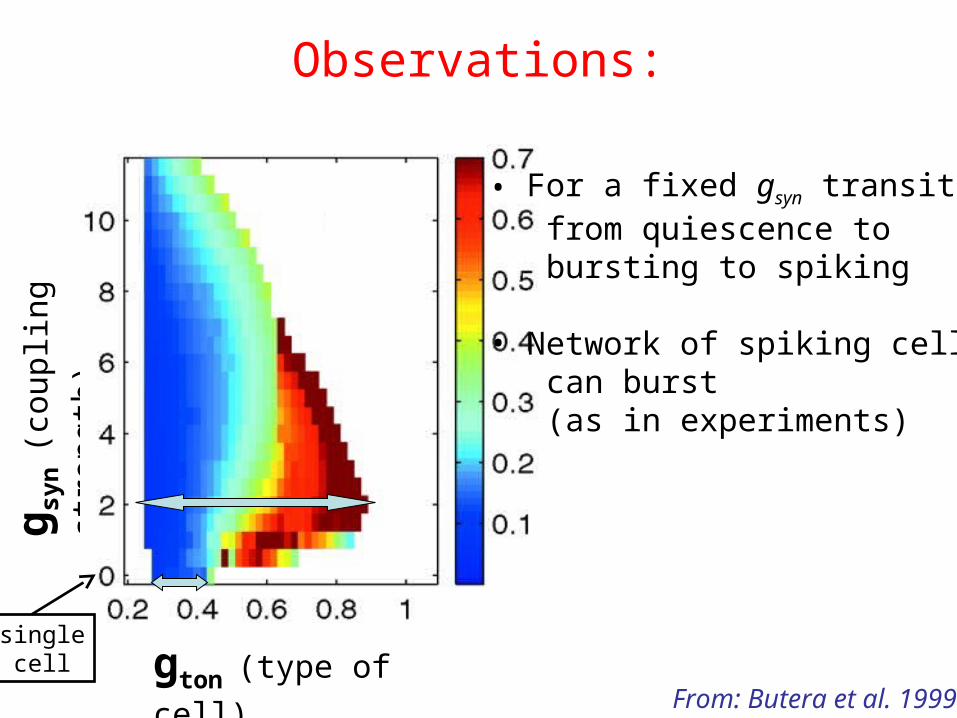
Observations:
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
From: Butera et al. 1999
• For a fixed gsyn transitions from quiescence to bursting to spiking
• Network of spiking cells can burst (as in experiments)
singlecell
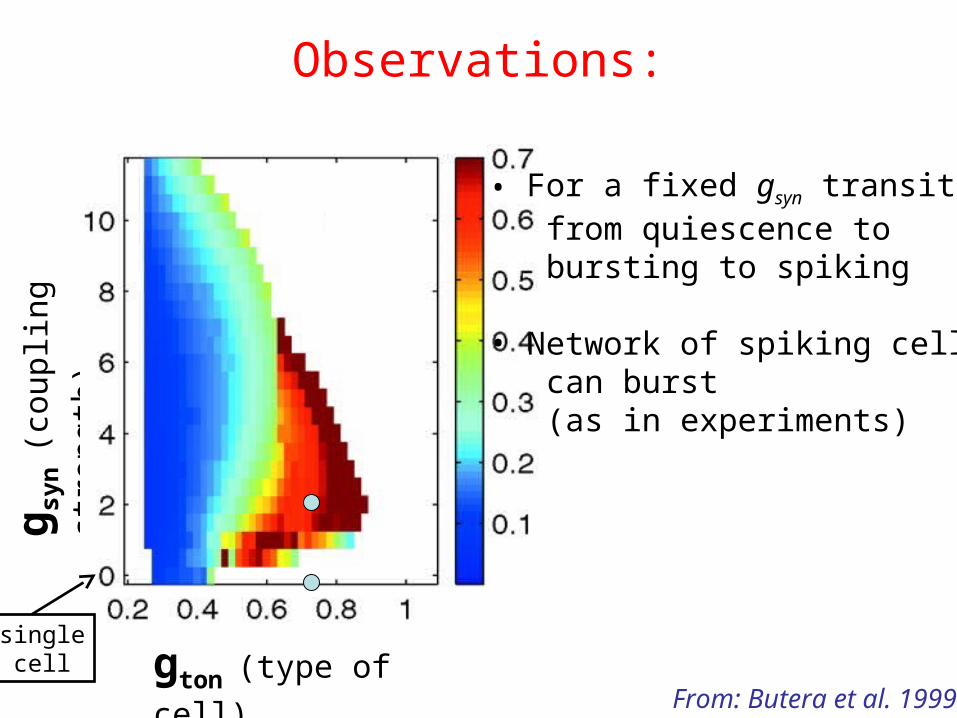
Observations:
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
From: Butera et al. 1999
• For a fixed gsyn transitions from quiescence to bursting to spiking
• Network of spiking cells can burst (as in experiments)
singlecell
Observations:
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
Burstduration
From: Butera et al. 1999
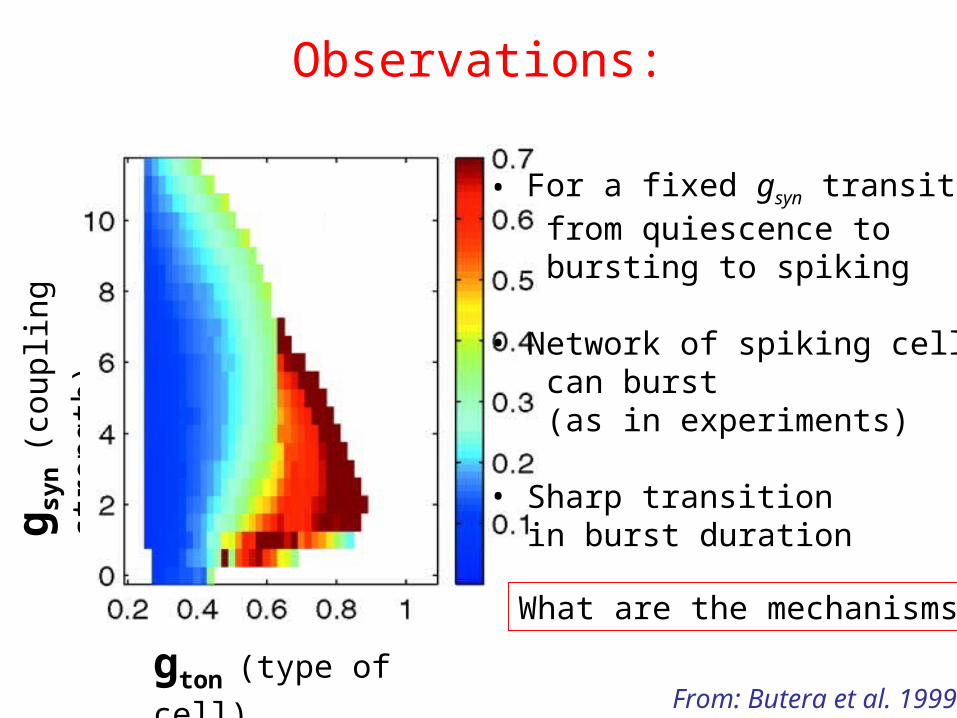
• For a fixed gsyn transitions from quiescence to bursting to spiking
• Network of spiking cells can burst (as in experiments)
• Sharp transition in burst duration
Observations:
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
From: Butera et al. 1999
What are the mechanisms?
• For a fixed gsyn transitions from quiescence to bursting to spiking
• Network of spiking cells can burst (as in experiments)
• Sharp transition in burst duration

Mathematical analysis
• Self-coupled cell - single cell - synchronous network
• Two cell network - strong coupling - weaker coupling
Mathematical analysis
• Self-coupled cell - single cell - synchronous network
• Two cell network - strong coupling - weaker coupling
• Transitions mechanism quiescence bursting spiking
Questions
• Why network is more bursty than a single cell (shape of bursting border)
• Sharp transition in burst duration

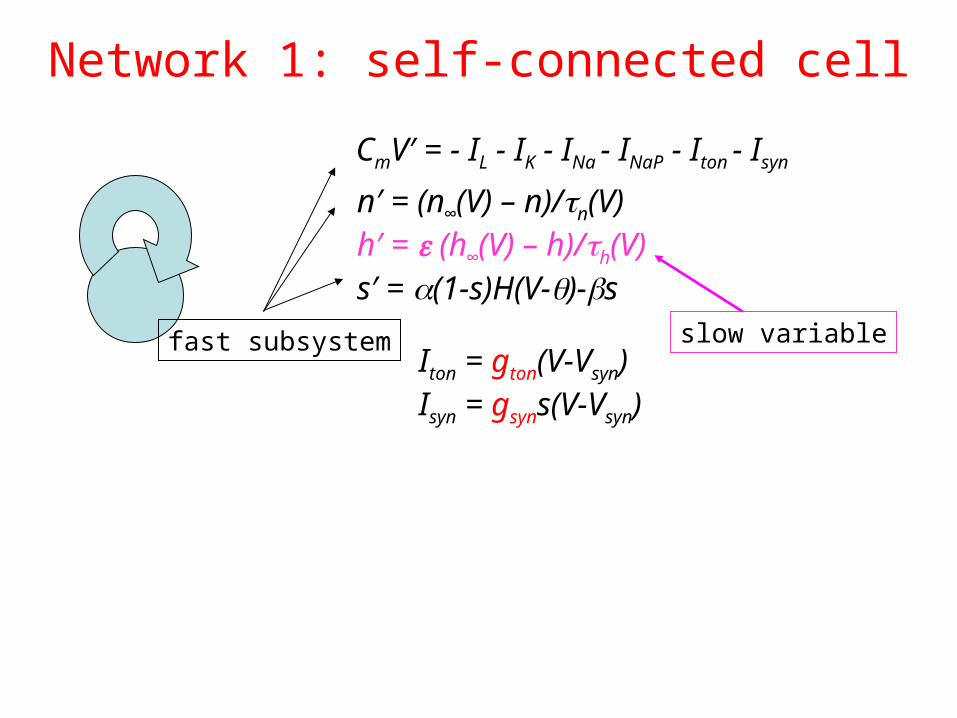
Network 1: self-connected cell
n′ = (n∞(V) – n)/n(V)h′ = (h∞(V) – h)/h(V)s′ = (1-s)H(V-)-s
CmV′ = - IL - IK - INa - INaP - Iton - Isyn
Iton = gton(V-Vsyn) Isyn = gsyns(V-Vsyn)

Network 1: self-connected cell
n′ = (n∞(V) – n)/n(V)h′ = (h∞(V) – h)/h(V)s′ = (1-s)H(V-)-s
CmV′ = - IL - IK - INa - INaP - Iton - Isyn
Iton = gton(V-Vsyn) Isyn = gsyns(V-Vsyn)
Why is this an interesting case?
• Includes individual neuron case (gsyn= 0)• Equivalent to a fully synchronized network• One slow variable (h) /h(V) ≪ 1/n(V)
h is slower than V
Network 1: self-connected cell
n′ = (n∞(V) – n)/n(V)h′ = (h∞(V) – h)/h(V)s′ = (1-s)H(V-)-s
CmV′ = - IL - IK - INa - INaP - Iton - Isyn
Iton = gton(V-Vsyn) Isyn = gsyns(V-Vsyn)
fast subsystem slow variable
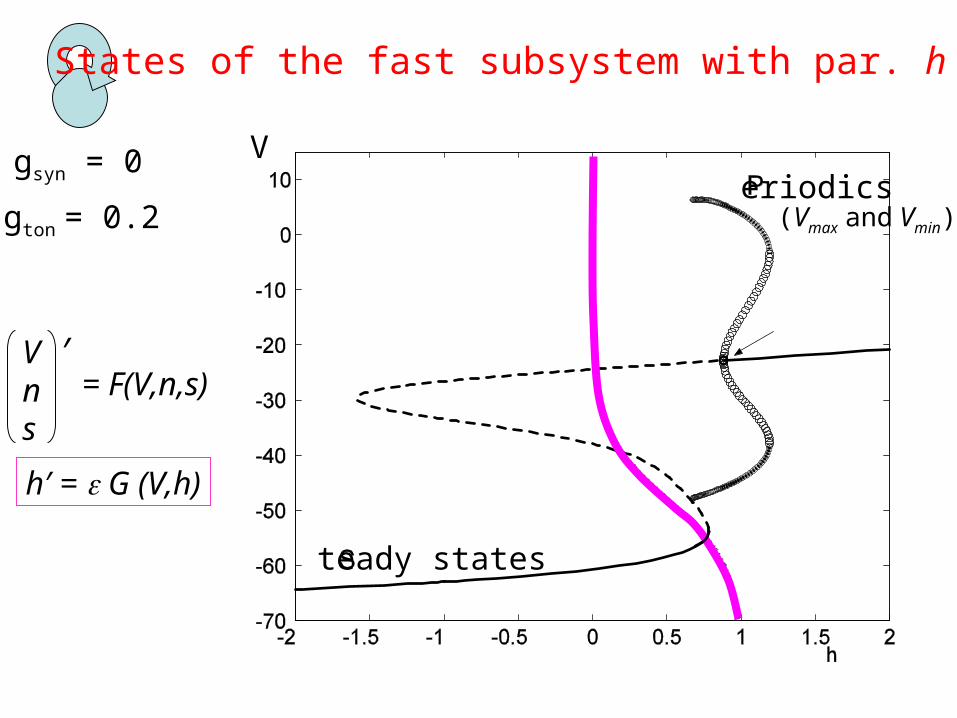
gsyn = 0
States of the fast subsystem with par. h
gton = 0.2
V
teady states
eriodics(Vmax and Vmin)
Vns
′= F(V,n,s)
h′ = G (V,h)
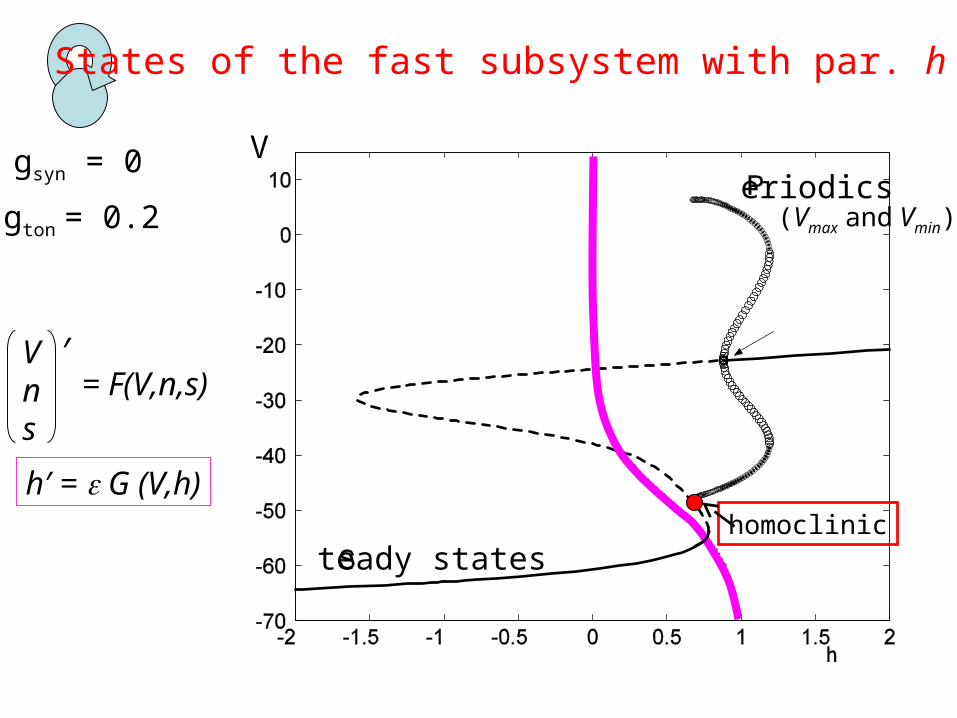
gsyn = 0
States of the fast subsystem with par. h
gton = 0.2
V
teady states
eriodics(Vmax and Vmin)
Vns
′= F(V,n,s)
h′ = G (V,h)homoclinic
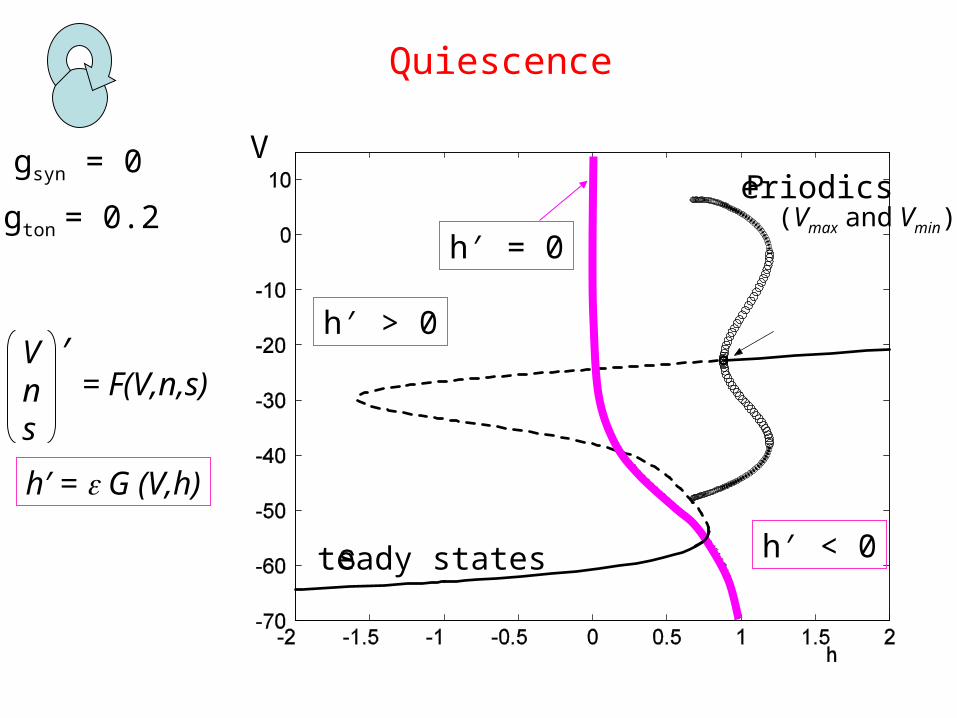
gsyn = 0
Quiescence
gton = 0.2
V
teady states
eriodics
h′ = 0
h′ < 0
h′ > 0
(Vmax and Vmin)
Vns
′= F(V,n,s)
h′ = G (V,h)
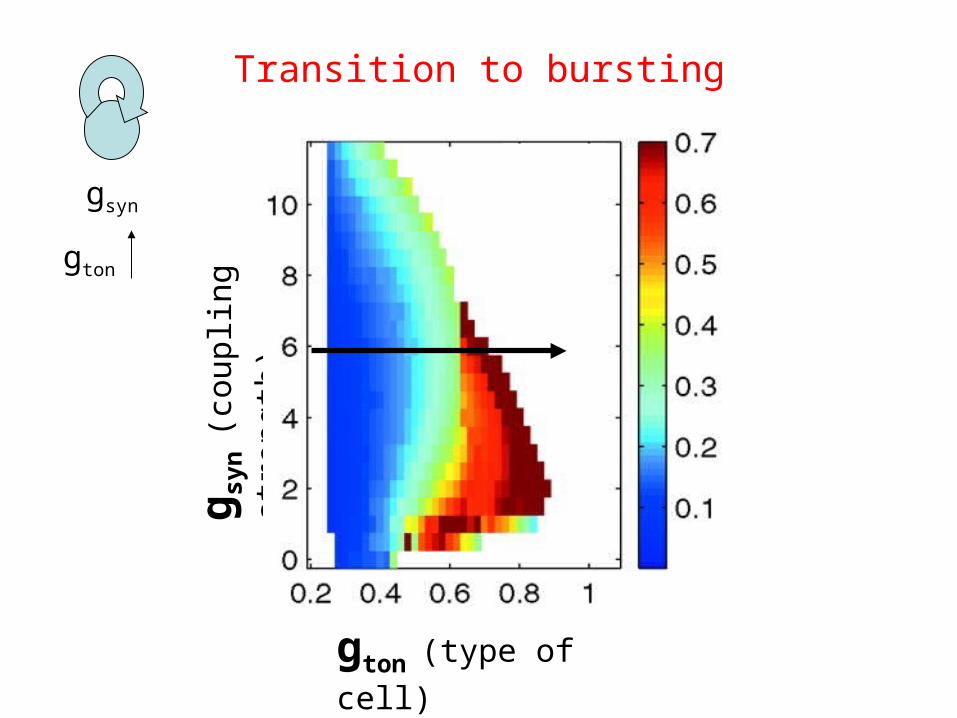
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
gsyn
Transition to bursting
gton
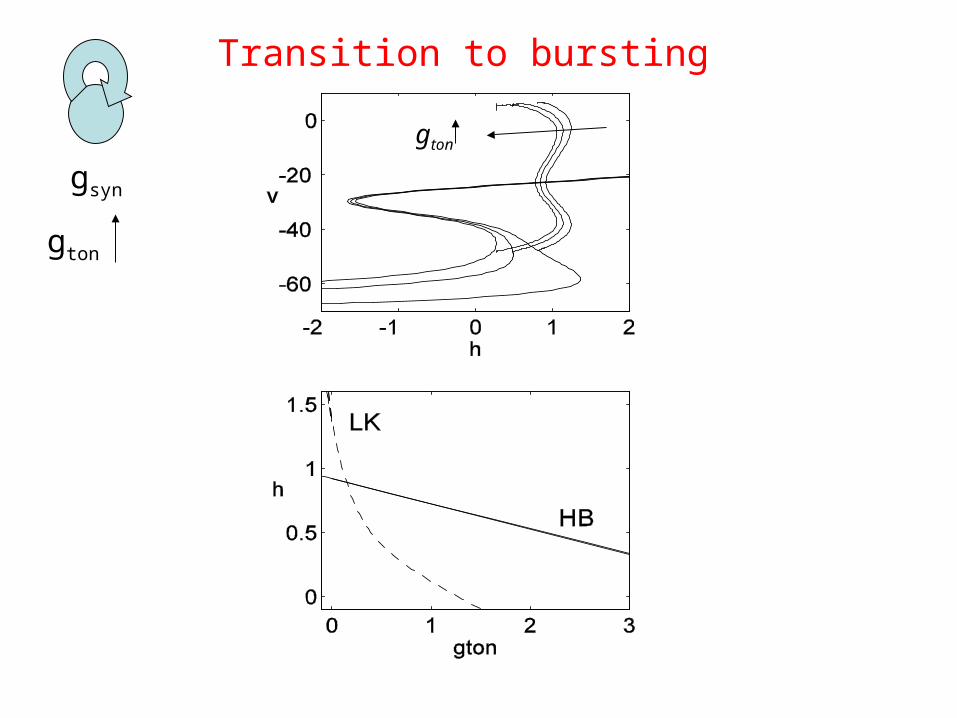
gsyn
gton
Transition to bursting
gton
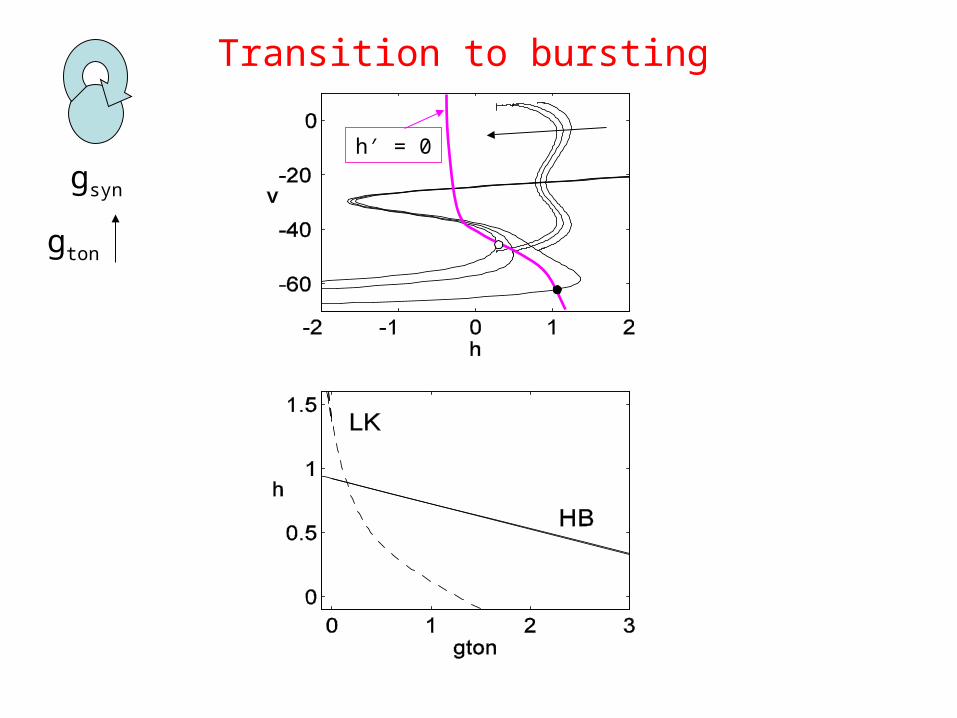
gsyn
h′ = 0
gton
Transition to bursting
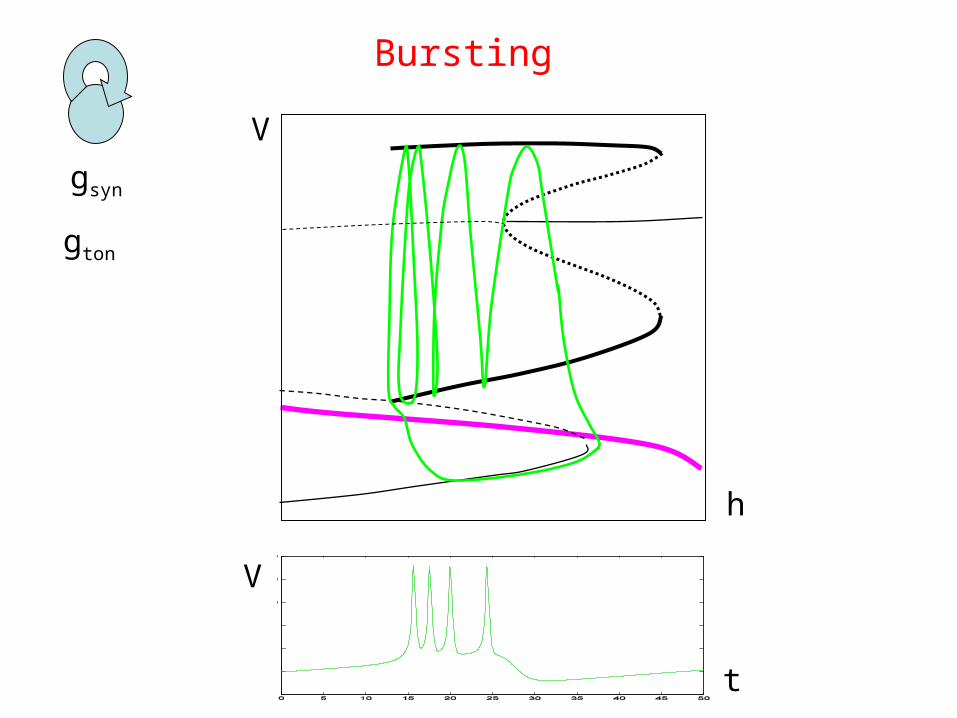
gsyn
h
V
Bursting
gton
t
V
gsyn
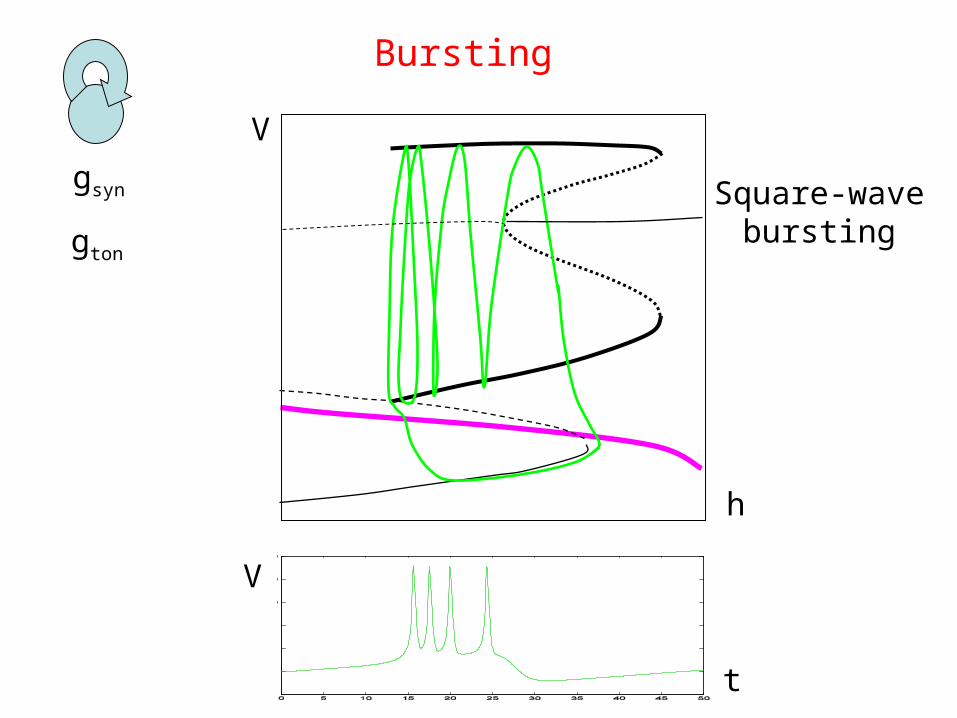
h
V
Bursting
gton
t
V
Square-wavebursting
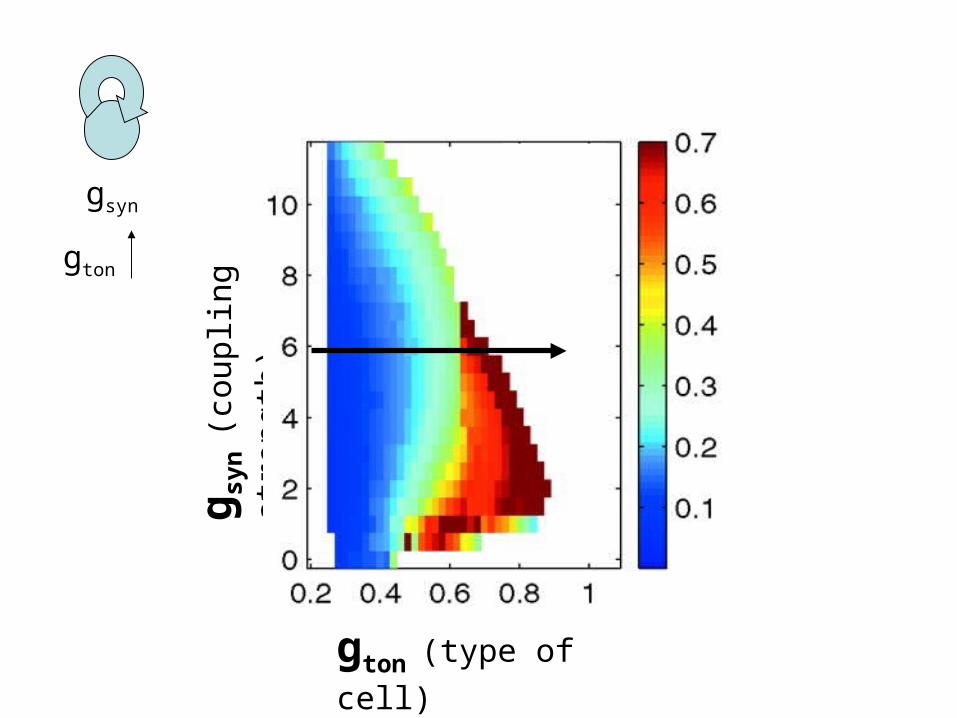
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
gsyn
gton
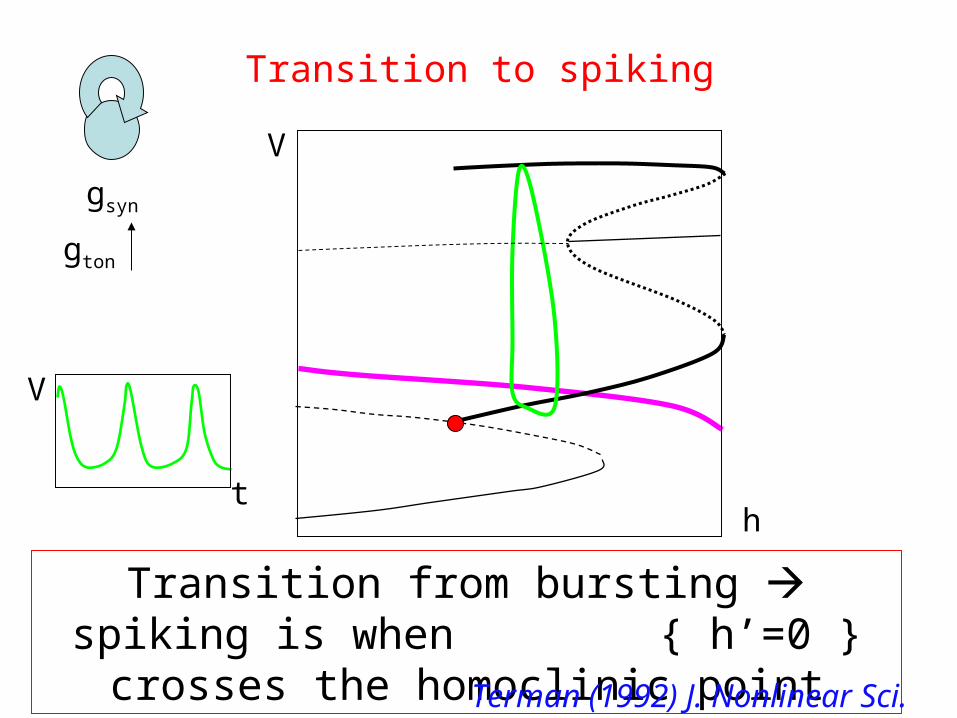
gsyn
Transition to spiking
gton
h
V
Transition from bursting spiking is when { h’=0 } crosses the homoclinic point
t
V
Terman (1992) J. Nonlinear Sci.
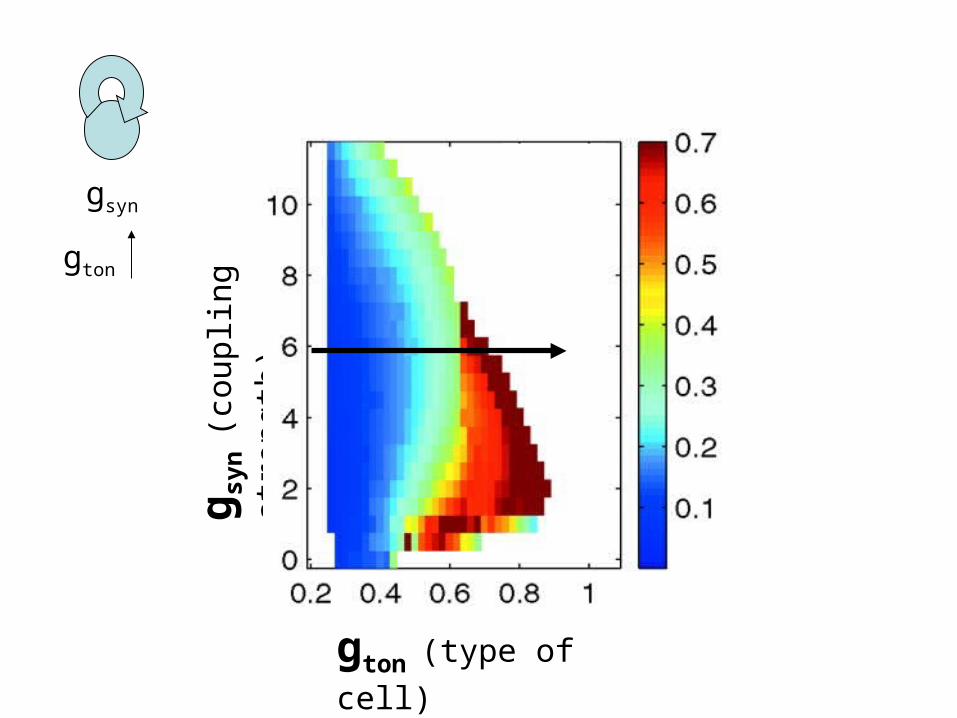
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
gsyn
gton
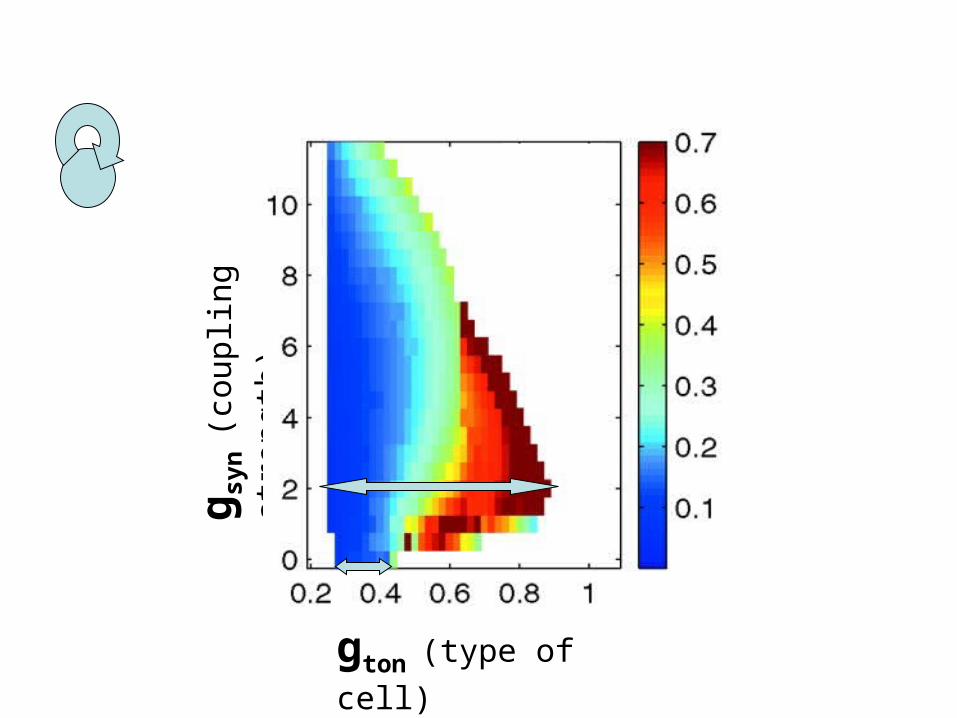
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
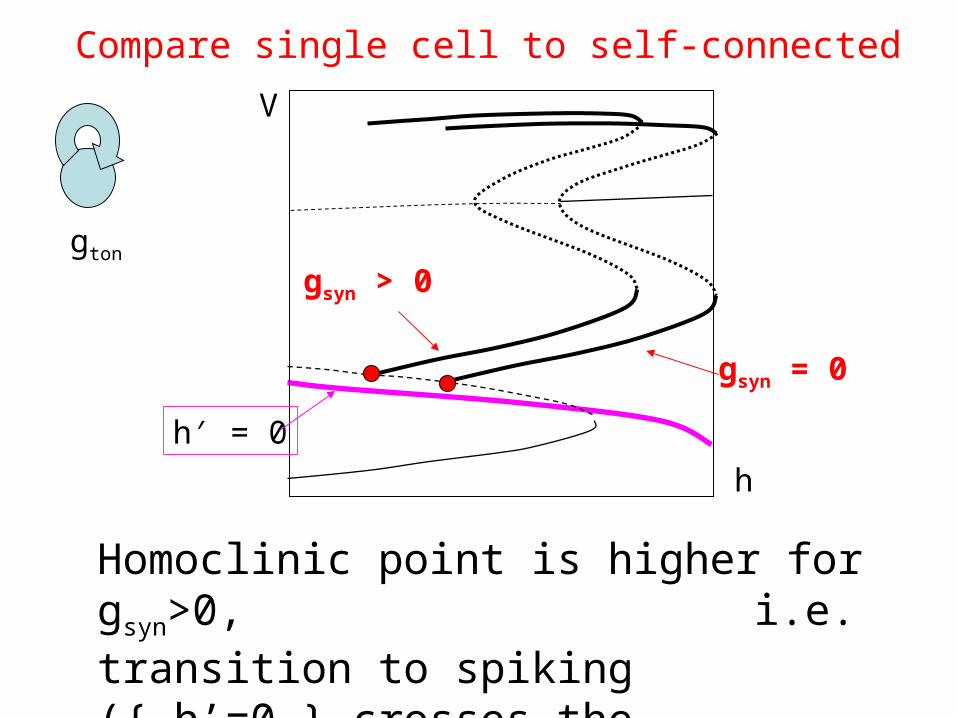
Compare single cell to self-connected
gton
h
V
gsyn = 0
gsyn > 0
h′ = 0
Homoclinic point is higher for gsyn>0, i.e. transition to spiking ({ h’=0 } crosses the homoclinic point) will happen for larger gton
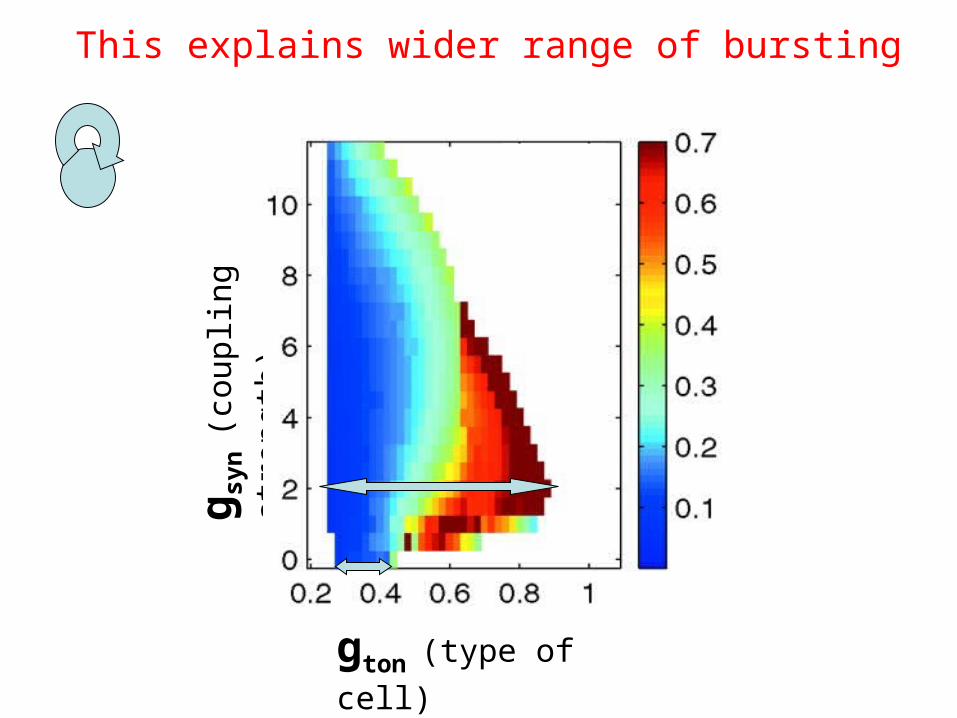
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
This explains wider range of bursting
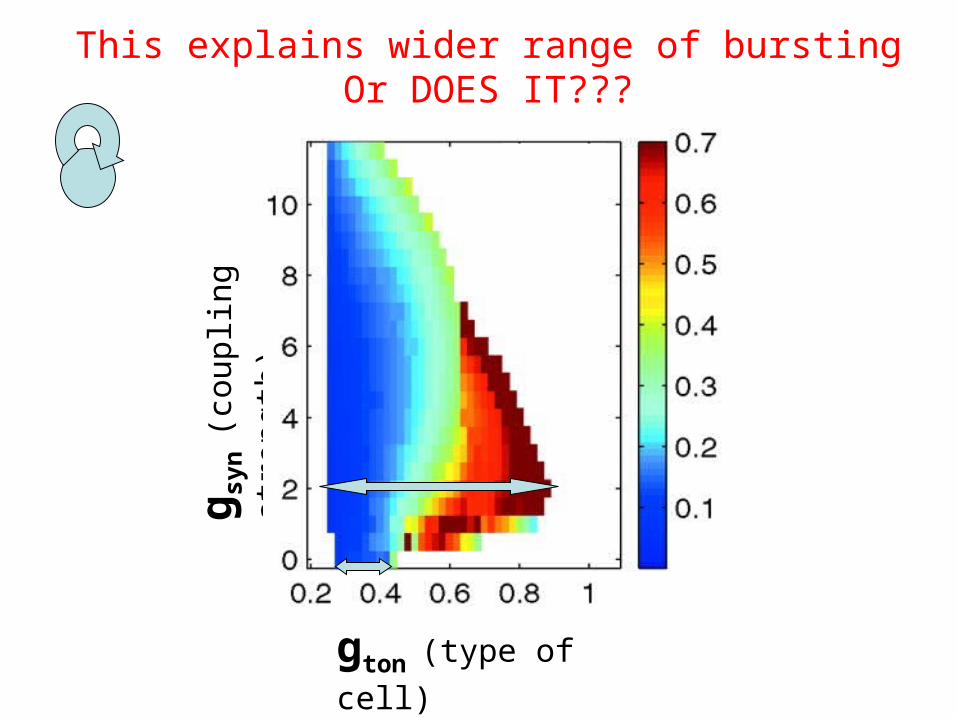
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
This explains wider range of burstingOr DOES IT???
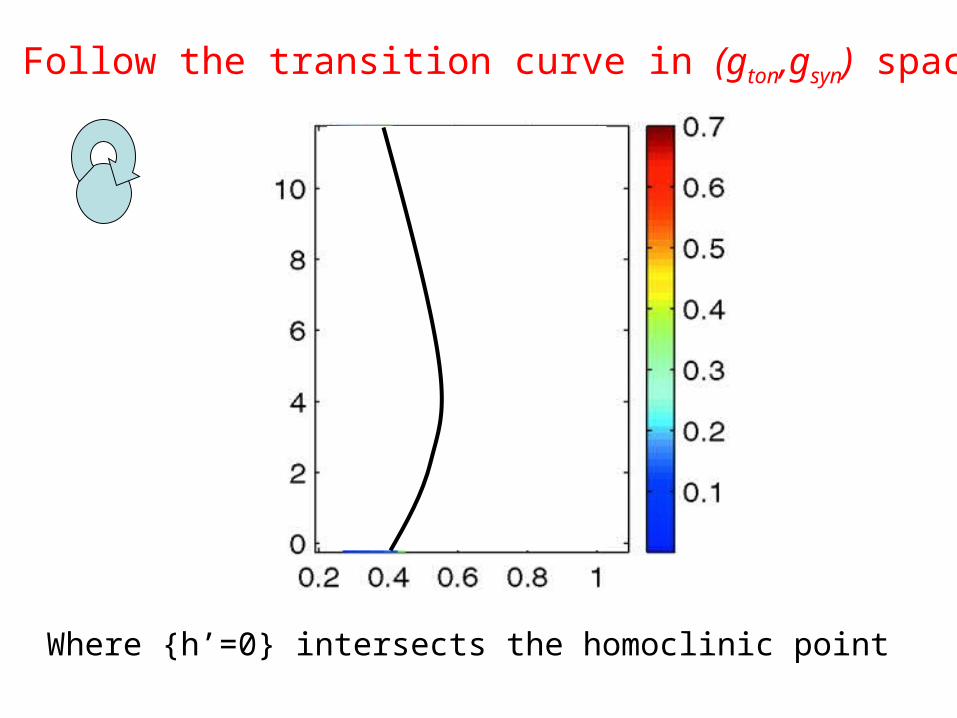
Follow the transition curve in (gton,gsyn) space
Where {h’=0} intersects the homoclinic point
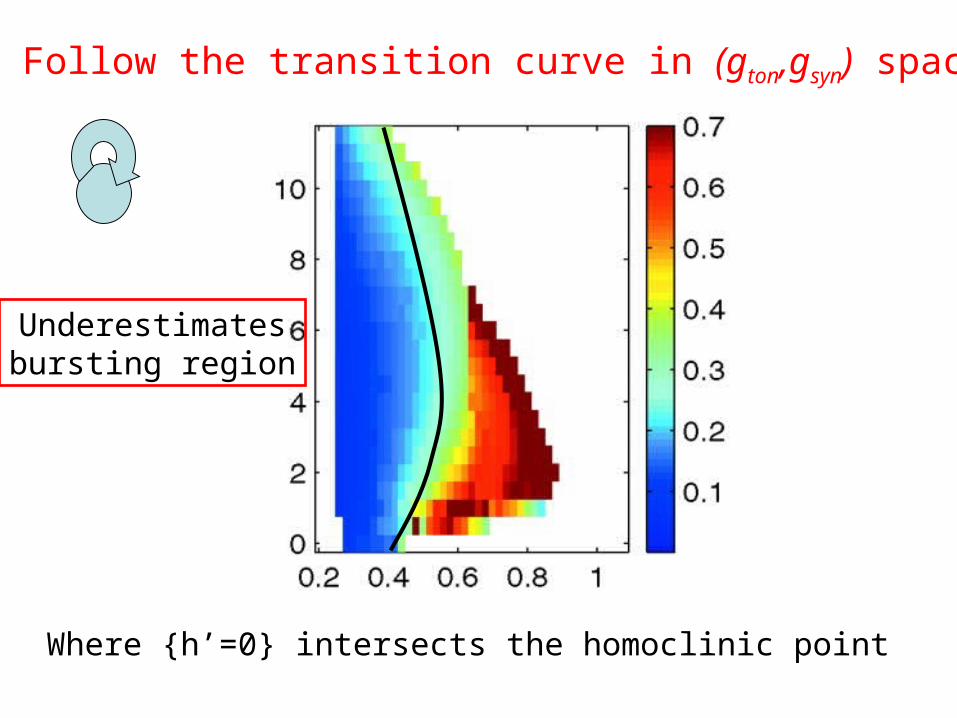
Follow the transition curve in (gton,gsyn) space
Where {h’=0} intersects the homoclinic point
Underestimatesbursting region
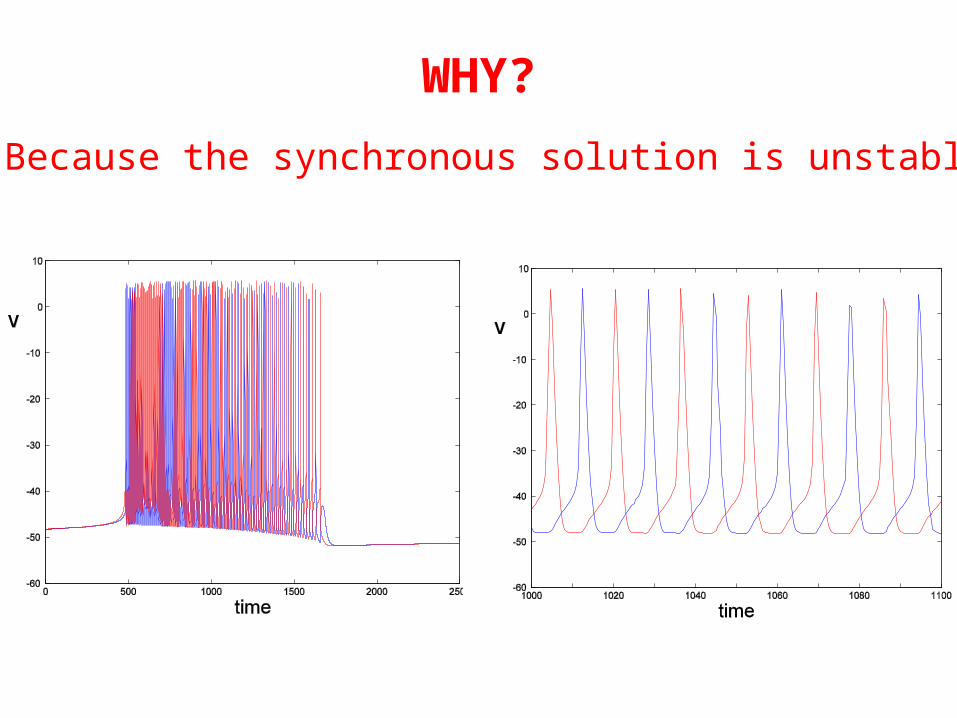
WHY?
Because the synchronous solution is unstable

Network 2: two connected cells
ni′ = (n∞(Vi) – ni)/n(Vi)si′ = (1-si)H(Vi-)-si
2 slow variables: hi′ = (h∞(Vi) – hi)/h(Vi)
CmVi′ = - IL - IK - INa - INaP - Iton - Isyn
Iton = gton(Vi-Vsyn) Isyn = gsynsj(Vi-Vsyn) i∈{1,2}, j=3-i
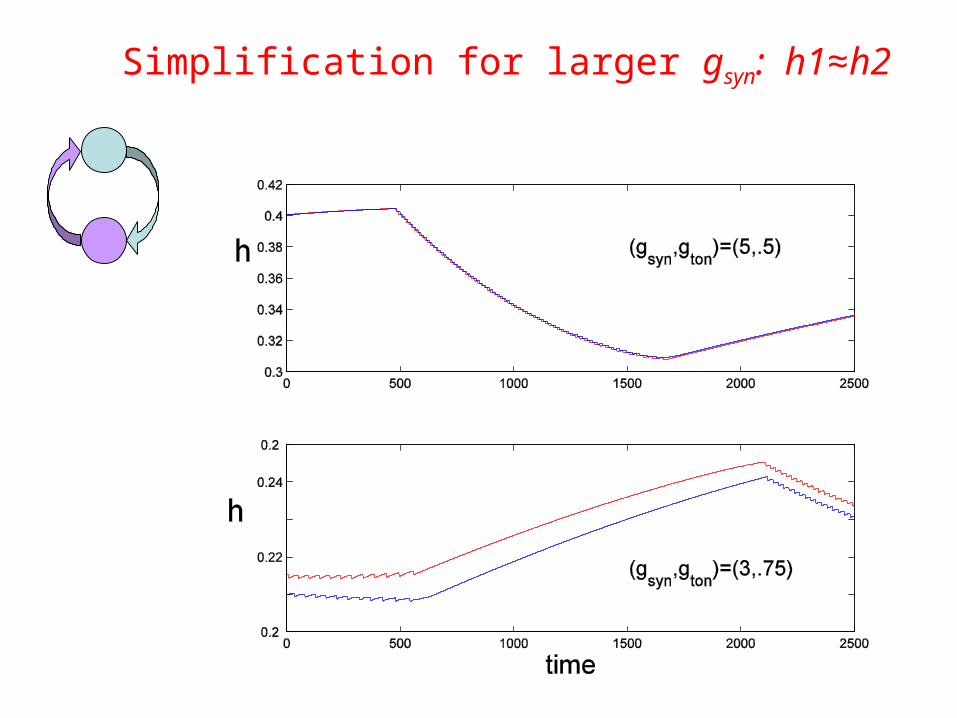
Simplification for larger gsyn: h1≈h2
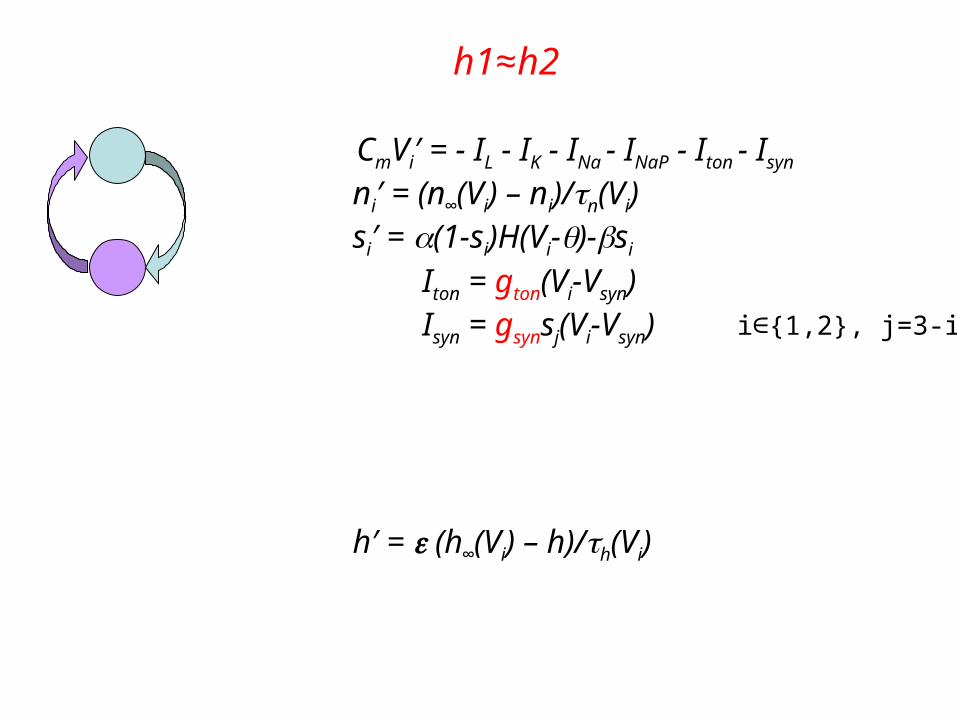
ni′ = (n∞(Vi) – ni)/n(Vi)si′ = (1-si)H(Vi-)-si
h′ = (h∞(Vi) – h)/h(Vi)
CmVi′ = - IL - IK - INa - INaP - Iton - Isyn
Iton = gton(Vi-Vsyn) Isyn = gsynsj(Vi-Vsyn) i∈{1,2}, j=3-i
h1≈h2
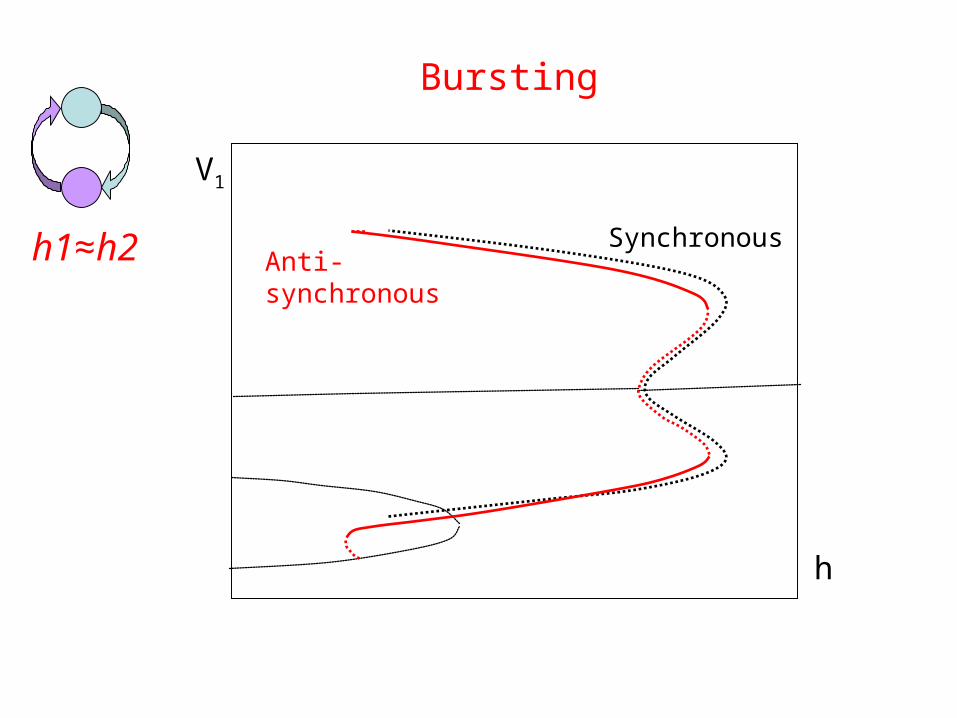
SynchronousAnti-synchronous
h
V1
Bursting
h1≈h2
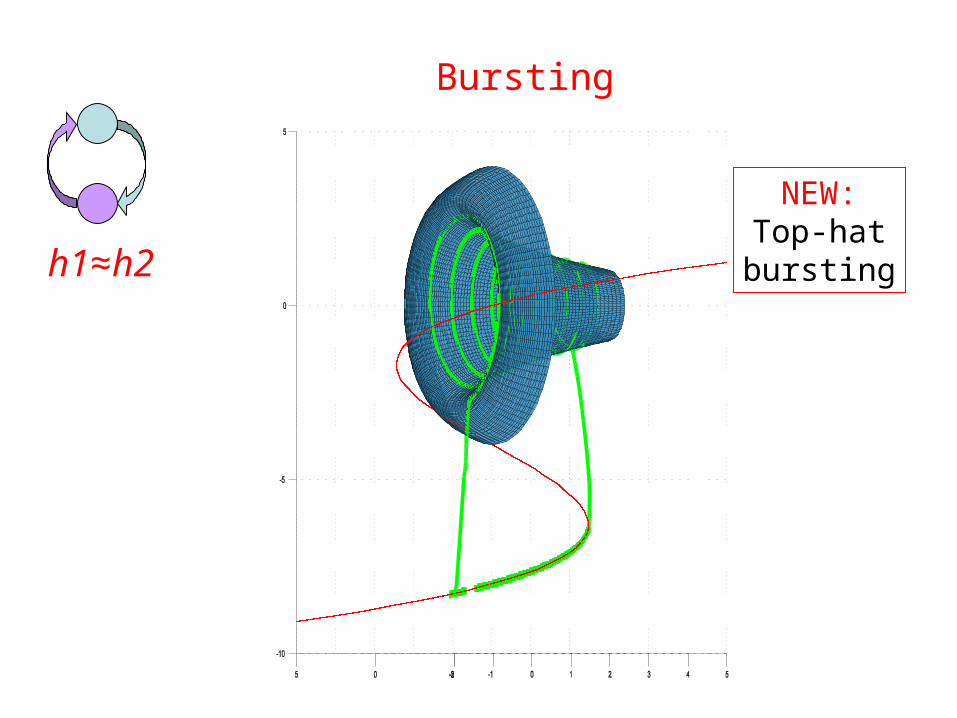
Bursting
h1≈h2
NEW:Top-hatbursting

Features of top-hat bursting:
h1≈h2
• Square wave bursters, when coupled, can generate top hat bursting
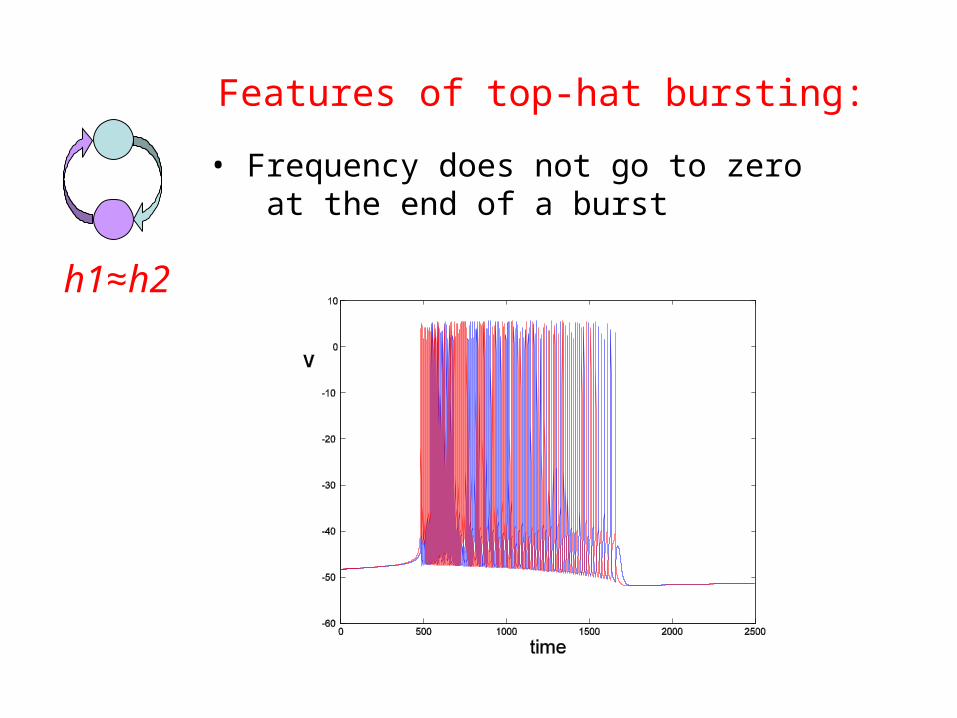
Features of top-hat bursting:
h1≈h2
• Frequency does not go to zero at the end of a burst

Features of top-hat bursting:
h1≈h2
• Different mechanism of (bursting spiking)

Features of top-hat bursting:
h1≈h2
• Different mechanism of (bursting spiking)
- Reduce full system to equations for slow variables:

Features of top-hat bursting:
h1≈h2
• Different mechanism of (bursting spiking)
- Reduce full system to equations for slow variables:
active phase
hL hR
For hL < h < hR let:
(Vi(t,h),ni(t,h),si(t,h)) - periodic orbitT(h) - period
When 0
h′ = (1/T(h))∫ (h∞(Vi(t,h))-h)/h(Vi(t,h)) dt ≡ a(h)0
T(h)
Bursting: a(h) < 0 for hL < h < hR

Features of top-hat bursting:
h1≈h2
• Different mechanism of (bursting spiking)
- Reduce full system to equations for slow variables:
active phase
hL hR
For hL < h < hR let:
(Vi(t,h),ni(t,h),si(t,h)) - periodic orbitT(h) - period
When 0
h′ = (1/T(h))∫ (h∞(Vi(t,h))-h)/h(Vi(t,h)) dt ≡ a(h)0
T(h)
Spiking: a(hR) < 0, a(hL) > 0

Features of top-hat bursting:
h1≈h2
• Different mechanism of (bursting spiking)
- Reduce full system to equations for slow variables:
active phase
hL hR
For hL < h < hR let:
(Vi(t,h),ni(t,h),si(t,h)) - periodic orbitT(h) - period
When 0
h′ = (1/T(h))∫ (h∞(Vi(t,h))-h)/h(Vi(t,h)) dt ≡ a(h)0
T(h)
Transition: a(hL) = 0
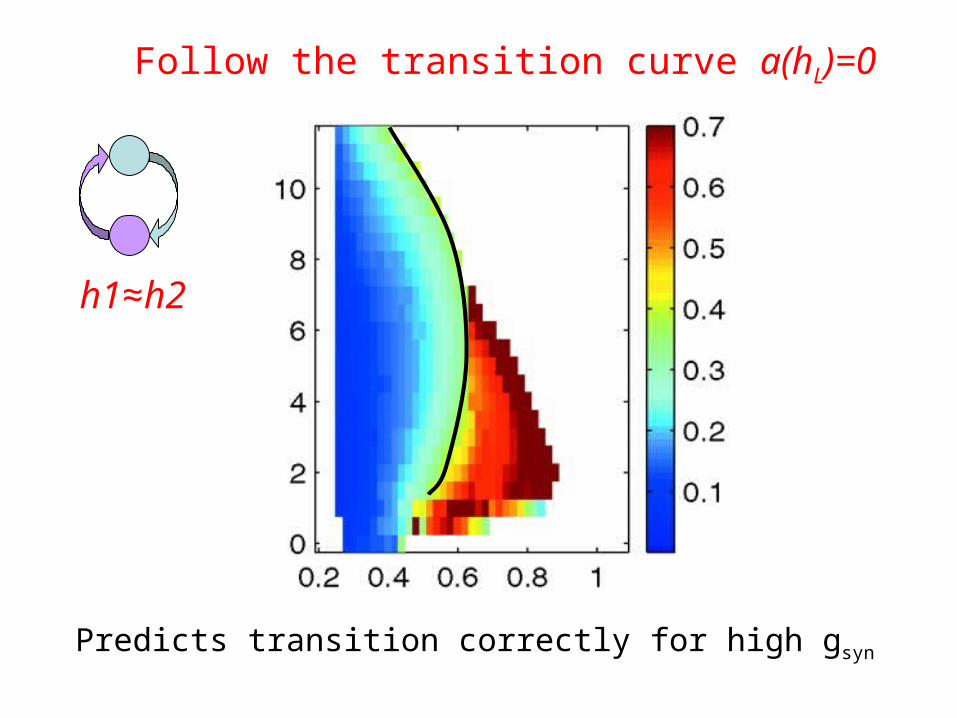
Follow the transition curve a(hL)=0
Predicts transition correctly for high gsyn
h1≈h2

Small and moderate gsyn: h1h2
• Define region R in (h1, h2) space such that fast subsystemsupports oscillations• Reduce full system: for R in (h1, h2)
• Transition (bursting spiking) can be understood by analyzing the phase planes of this system
h1′ = (1/T(h1,h2))∫ G(V1,p(t,h1,h2),h1) dt ≡ a1(h1,h2)0
T(h1,h2)
h2′ = (1/T(h1,h2))∫ G(V2,p(t,h1,h2),h2) dt ≡ a2(h1,h2)0
T(h1,h2)
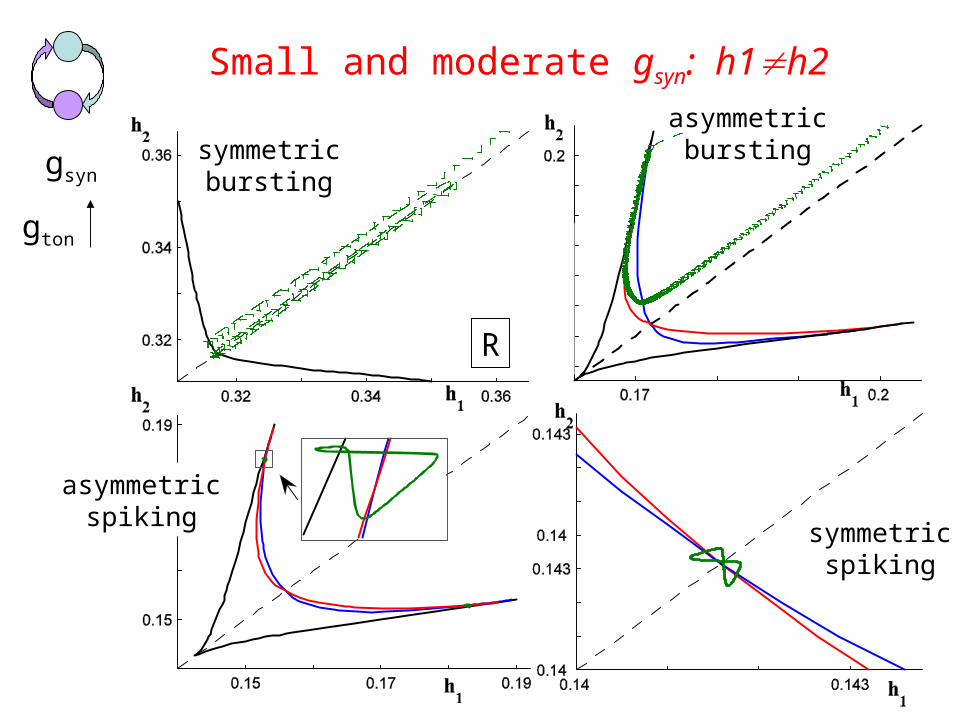
Small and moderate gsyn: h1h2
gsyn
gton
R
symmetricbursting
asymmetricbursting
asymmetricspiking
symmetricspiking
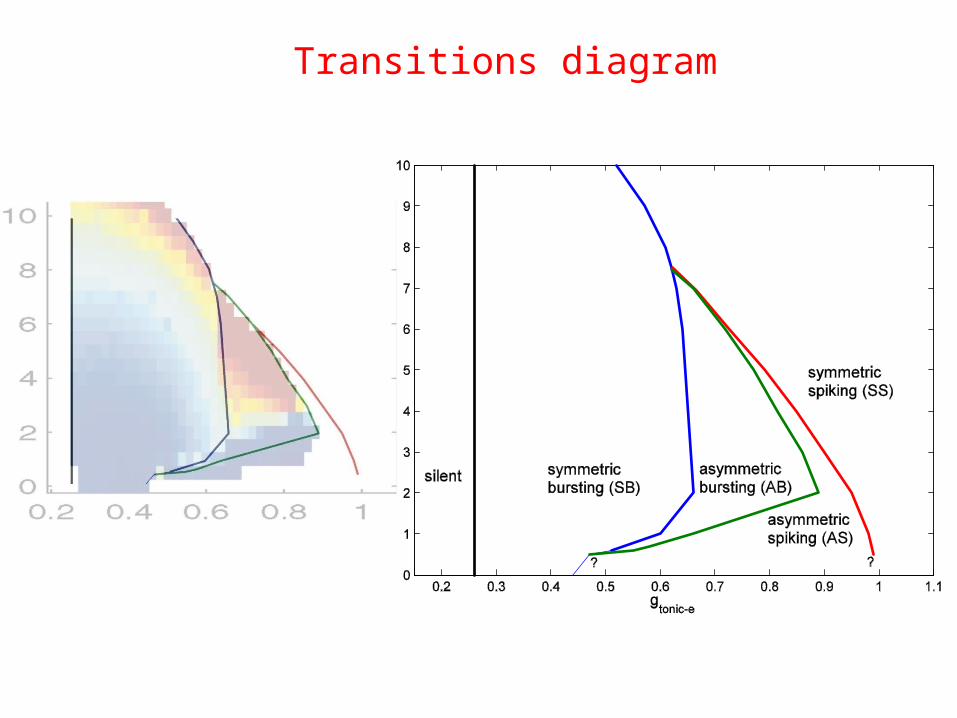
Transitions diagram
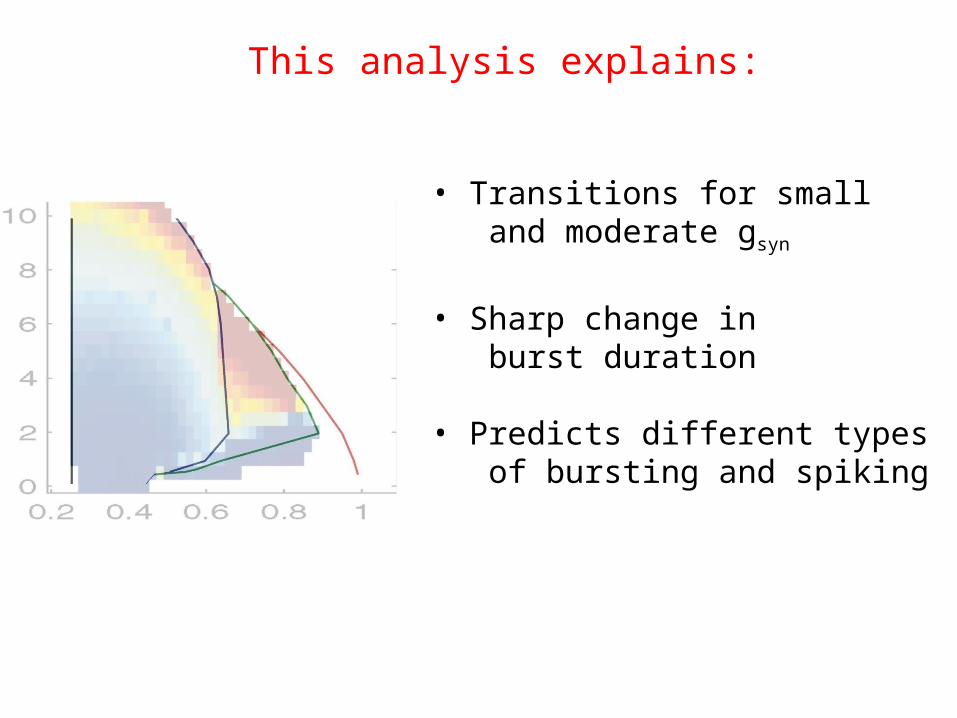
• Transitions for small and moderate gsyn
• Sharp change in burst duration
• Predicts different types of bursting and spiking
This analysis explains:

Conclusions
New in networks of bursting cells:
• Coupled square-wave bursters can generate top-hat bursting• Activity modes of coupled bursters can be characterized by considering phase space of averaged slow-variable equations
New predictions for experiments:
• Isolated cell has infrequent spikes at the end of a burst, but a cell in the network does not• In a pair of cells there can be two different types of bursting and two different types of spiking. Transitions can be made by changing gton
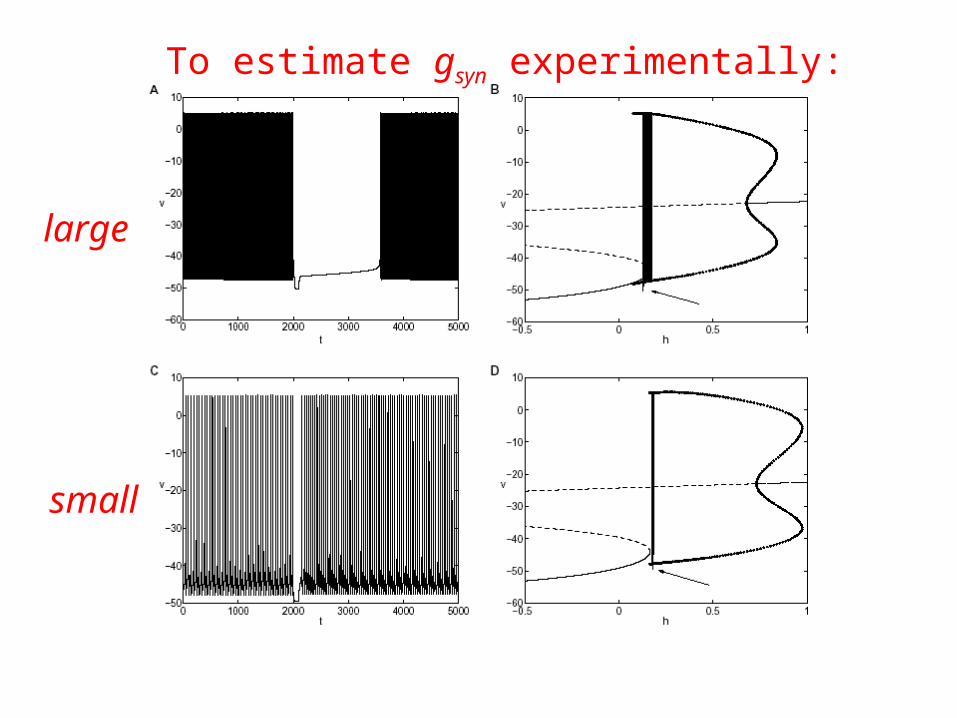
To estimate gsyn experimentally:
large
small

• J. Best, J. Rubin, D. Terman, M. Wechselberger• Supported by NSF (agreement No. 0112050) through Mathematical Biosciences Institute (MBI), OSU
Acknowledgments


- Motivation (Pre-Botz)- Butera et al. model and some results (dynamic range? Freq jump?)-What is the mathematical structure that underlies it + uncoupled cells: square wave bursting, transition to spiking + coupled cells: what is the correct reduced model? If we had a self-coupled cell, transition to spiking would happen when h’=0 crosses homoclinic point. But is this a valid reduced model? Compute: anti-phase + Numerically: anti-phase solution is a top-hat burster. Transition to spiking is when the average at the saddle-node of periodics is zero – incorrect. + Next: h1 ~=h2 - It is good to know mathematical structure, also good for bio (predictions)
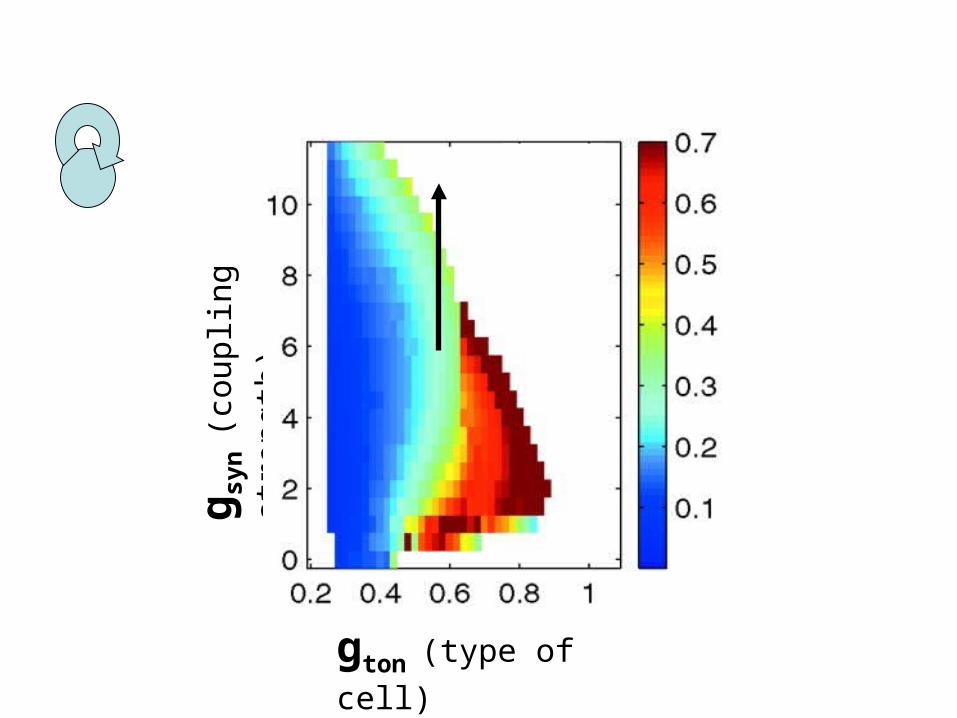
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
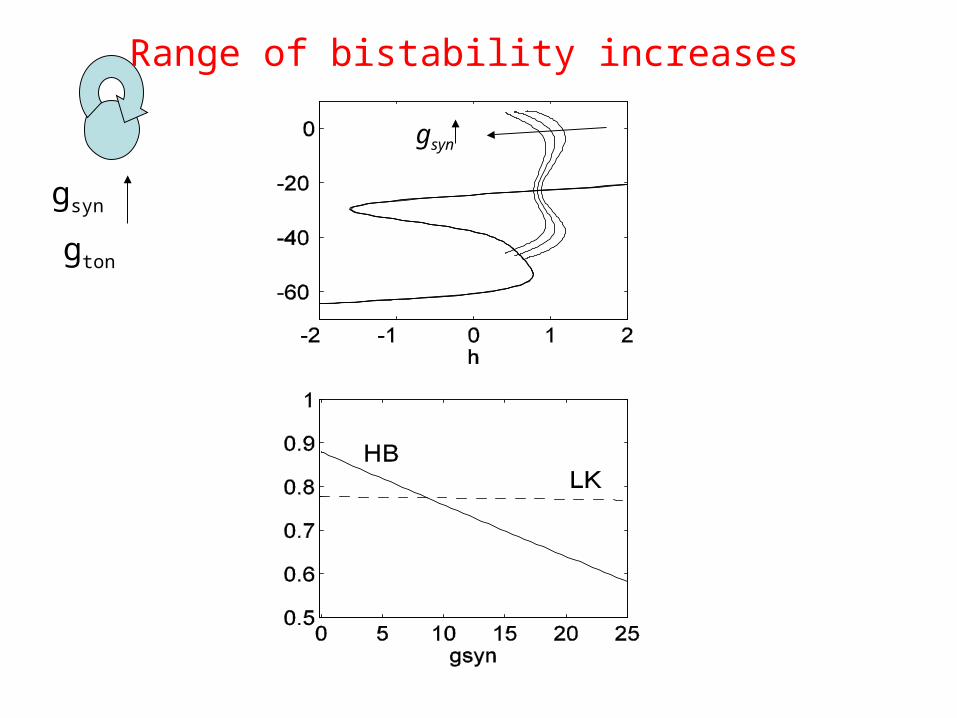
gsyn
gton
gsyn
Range of bistability increases
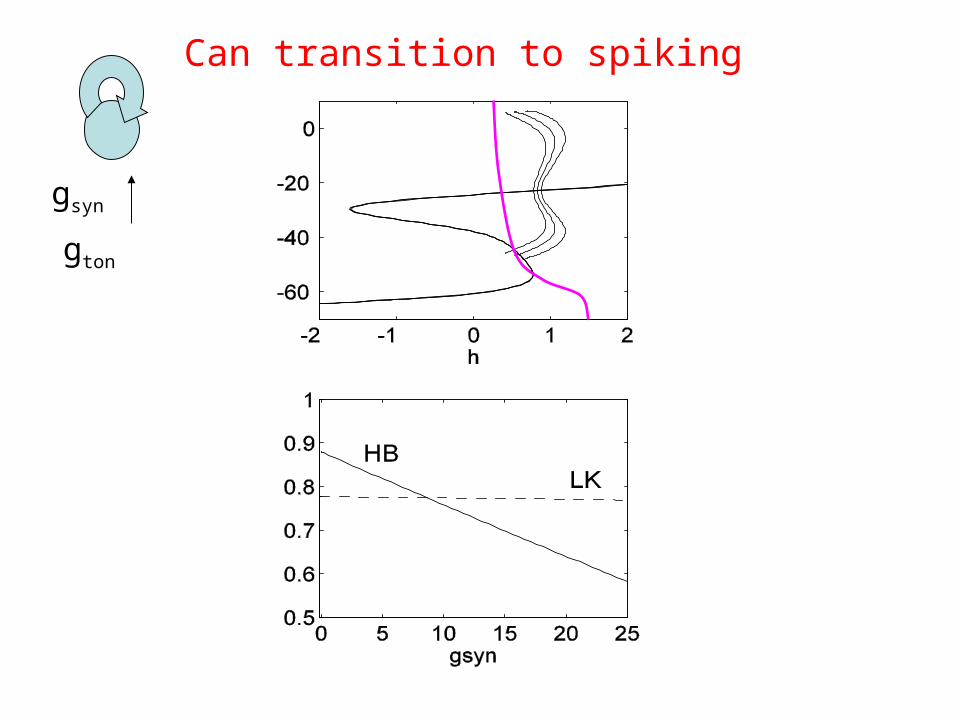
gsyn
gton
Can transition to spiking
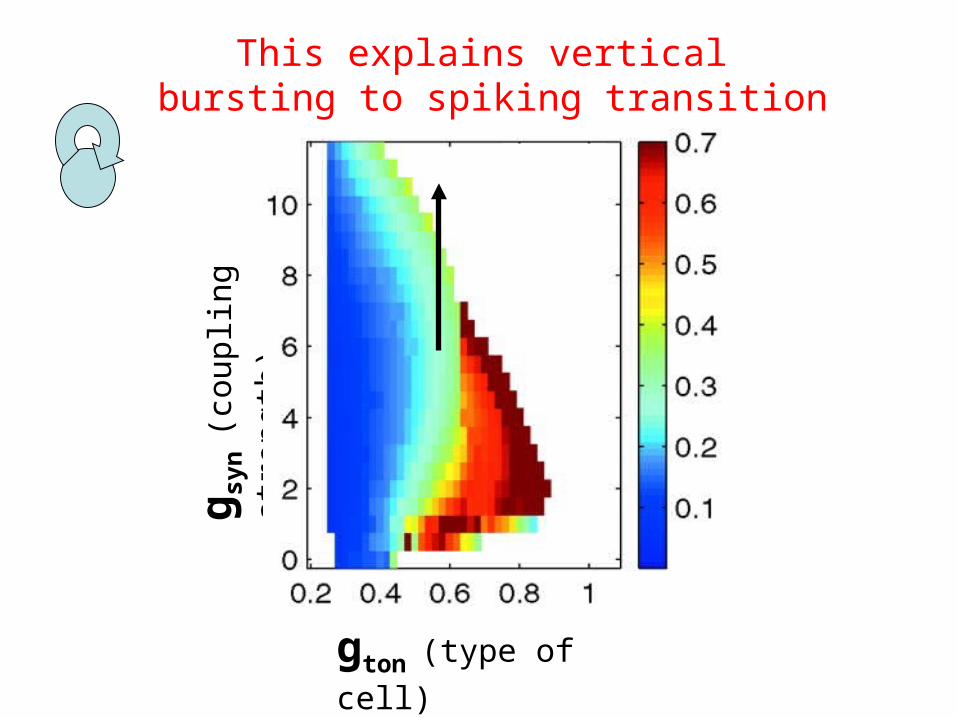
gton (type of cell)
g syn
(cou
plin
g st
reng
th)
This explains vertical bursting to spiking transition

2 experimental figuresDiff figure from butera et al for burst durationSlide for H functionsCorrect figure with h’sDifferent figure from JanetInsert schematic of h1-h2 plane





























