Embed Size (px)
Citation preview

Tierärztliche Hochschule Hannover
UNTERSUCHUNGEN ZUM VORKOMMEN VON SALMONELLA SPP.,
CAMPYLOBACTER SPP. UND YERSINIA ENTEROCOLITICA UND
ZUR DYNAMIK DER SALMONELLENINFEKTION IN SCHWEINE-
MASTBETRIEBEN
Inaugural-Dissertation
zur Erlangung des Grades einer
Doktorin der Veterinärmedizin
(Dr. med. vet.)
durch die Tierärztliche Hochschule Hannover
Vorgelegt von
Christina Nathues
aus Friedberg
Hannover 2011

Wissenschaftliche Betreuung: Univ.-Prof. Dr. Lothar Kreienbrock
Institut für Biometrie, Epidemiologie und Infor-
mationsverarbeitung der Tierärztlichen Hoch-
schule Hannover
Univ.-Prof. Dr. Thomas Blaha
Außenstelle für Epidemiologie der Tierärztlichen
Hochschule Hannover
1. Gutachter: Univ.-Prof. Dr. Lothar Kreienbrock
Univ.-Prof. Dr. Thomas Blaha
2. Gutachter: Univ.-Prof. Dr. Michael Wendt
Tag der mündlichen Prüfung: 09.05.2011
.

Ὕες βορβόρῳ ἥδονται μᾶλλον ἢ καθαρῷ ὕδατι.
Meinem Mann Heiko und meiner Familie


Inhaltsverzeichnis
1. Einleitung ............................................................................................................. 1
2. Literaturübersicht ................................................................................................. 4
2.1 Salmonella spp. ............................................................................................. 4
2.1.1 Taxonomie und Nomenklatur .................................................................. 4
2.1.2 Erregereigenschaften .............................................................................. 5
2.1.3 Nachweismethoden ................................................................................ 7
2.1.4 Salmonellosen beim Menschen ............................................................ 15
2.1.5 Salmonella spp. beim Schwein ............................................................. 19
2.2 Campylobacter spp. ..................................................................................... 29
2.2.1 Erregereigenschaften ............................................................................ 29
2.2.2 Nachweismethoden .............................................................................. 30
2.2.3 Campylobacteriosen beim Menschen ................................................... 34
2.2.4 Campylobacter spp. beim Schwein ....................................................... 37
2.3 Yersinia enterocolitica ................................................................................. 42
2.3.1 Erregereigenschaften ............................................................................ 42
2.3.2 Nachweismethoden .............................................................................. 43
2.3.3 Yersiniosen beim Menschen ................................................................. 46
2.3.4 Y. enterocolitica beim Schwein ............................................................. 50
2.4 Ziele dieser Studie ....................................................................................... 55
3. Material und Methoden ...................................................................................... 57
3.1 Querschnittsuntersuchung zu Salmonella spp., Campylobacter spp. und Y.
enterocolitica ......................................................................................................... 57
3.1.1 Studiendesign und Ziel der Untersuchung ............................................ 57
3.1.2 Auswahl der Bestände .......................................................................... 57

3.1.3 Untersuchung der Bestände ................................................................. 58
3.1.4 Bakteriologische Untersuchung der Proben .......................................... 60
3.1.5 Erhebung der Betriebsdaten ................................................................. 62
3.1.6 Verwaltung und Auswertung der Daten ................................................ 62
3.2 Salmonellenverlaufsuntersuchung .............................................................. 69
3.2.1 Art und Ziel der Untersuchung .............................................................. 69
3.2.2 Auswahl der Bestände .......................................................................... 69
3.2.3 Untersuchung der Bestände ................................................................. 69
3.2.4 Untersuchung der Proben ..................................................................... 72
3.2.5 Eingruppierung der Salmonella-Isolate in klonale Linien ...................... 77
3.2.6 Erhebung von Betriebsdaten................................................................. 77
3.2.7 Verwaltung und Auswertung der Daten ................................................ 78
4. Ergebnisse ........................................................................................................ 82
4.1 Querschnittsuntersuchung ........................................................................... 82
4.1.1 Beschreibung der Ergebnisse auf Probenebene................................... 82
4.1.2 Beschreibung der Ergebnisse auf Lokalisationsebene ......................... 83
4.1.3 Beschreibung der Probenergebnisse auf Betriebsebene ...................... 87
4.1.4 Gemeinsames Auftreten der Erreger .................................................... 92
4.1.5 Risikofaktoren für das Auftreten der Erreger im Betrieb ........................ 96
4.2 Salmonellenverlaufsuntersuchung ............................................................ 109
4.2.1 Beschreibung der Ergebnisse auf Probenebene................................. 111
4.2.2 Beschreibung der Ergebnisse auf Klonebene ..................................... 122
5. Diskussion ....................................................................................................... 136
5.1 Querschnittsuntersuchung ......................................................................... 137
5.1.1 Erregernachweis ................................................................................. 137

5.1.2 Vorkommen der Erreger in der Tierumgebung ................................... 139
5.1.3 Gemeinsames Auftreten der Erreger .................................................. 143
5.1.4 Risikofaktoren ..................................................................................... 148
5.2 Verlaufsuntersuchung ................................................................................ 153
5.2.1 Betriebsübergreifende Analyse ........................................................... 155
5.2.2 Beschreibung der einzelnen betrieblichen Verläufe ............................ 163
5.3 Schlussfolgerungen ................................................................................... 170
6. Zusammenfassung .......................................................................................... 172
7. Summary ......................................................................................................... 175
8. Literaturverzeichnis ......................................................................................... 178
9. Anhang ............................................................................................................ 202

Verzeichnis der Abkürzungen
C Grad Celsius
AFLP amplified fragment length polymorphism
Ag Antigen
bp Basenpaare BPLS Brillantgrün-Phenolrot-Laktose-Saccharose
BPW buffered peptone water (gepuffertes Peptonwasser) C. Campylobacter
ca. circa
CIN Cefsulidon-Irgasan-Neutralrot
DG Durchgang
DIN Deutsches Institut für Normung
DNA desoxyribonuclein acid
EFSA European Food Safety Authority
ELISA enzyme-linked immunosorbent assay
EMB Eosin-Methylenblau
EN ISO Europäische Norm, Internationale Organisation für Normung et al. et alii
etc. et cetera
EU Europäische Union
EWG Europäische Wirtschaftsgemeinschaft
FBI-Zoo Verbundprojekt „Food-borne zoonotic infections“
g Gramm
H2O Wasser
H2S Schwefelwasserstoff
KBR Komplementbindungsreaktion
KG Konfidenzgrenzen
kg Kilogramm
l Liter LB Luria broth
LK Landkreis
mCCD modified charcoal, cefoperazone and deoxycholate
ml Milliliter
ML ST multilocus sequence typing
MLVA multiple locus VNTR analysis
mm Millimeter
MRSV modifiziertes Rappaport-Vassiliadis Medium
NaCl Natriumchlorid
OSCM Oxoid Salmonella chromogenic medium
PCR polymerase chain reaction
PFGE Pulsfeld-Gelelektrophorese pH potentia hydrogenii
pmol picomol
PSB Pepton-Sorbit-Bile

QS Qualität und Sicherheit GmbH
RAPD random amplification of polymorphic DNA
RFLP restriction fragment length polymorphism
rpm rounds per minute
RV Rappaport-Vassiliadis S. Salmonella
SNP single nucleotide polymorphism
spp. species
ssp. subspecies ST Sequenztyp
u. ä. und ähnliche
UK United Kingdom
UPGMA unweighted pair group method with arithmetic mean
USA United States of America
usw. und so weiter
V Volt
VBNC viable but non-culturable
VNTR variable number of tandem repeat
VTP-FBP vancomycin, trimethoprim, polymixin B - ferrous sulphate, sodium bisul-fite, sodium pyruvate
VYE virulent-Y. enterocolitica
WHO World Health Organization
XLD Xylose-Lysin-Desoxycholat XLT4 Xylose-Lysin-Tergitol Y. Yersinia
YOPI young, old, pregnant, immunosuppressed
z. B. zum Beispiel
ZP Zeitpunkt
μg Mikrogramm
μl Mikroliter
μm Mikrometer


Einleitung
1
1. Einleitung
Zoonosen sind „sämtliche Krankheiten und/oder sämtliche Infektionen, die natürli-
cherweise von Tieren auf Menschen übertragen werden können“, so die Definition in
der EU-Zoonose-Richtlinie (92/117/EWG). Dabei stellen die durch Lebensmittel über-
tragbaren Erkrankungen eine wachsende Bedrohung für die öffentliche Gesundheit
dar: Nach Einschätzung der WHO starben 2005 weltweit 1,8 Millionen Menschen an
Durchfallerkrankungen, von denen ein großer Anteil dem Konsum kontaminierter Le-
bensmittel oder Wasser zugeschrieben wird. Dass dies nicht allein ein Problem von
weniger entwickelten Ländern ist, zeigen Daten aus den USA, wo jedes Jahr ge-
schätzte 76 Millionen Fälle von „food-borne diseases“ mit 325.000 Hospitalisierungen
und 5.000 Toten auftreten (ANONYMOUS 2011).
Die drei bedeutendsten bakteriellen Erreger lebensmittelassoziierter Zoonosen in
Europa sind Campylobacter spp., Salmonella spp. und Yersinia (Y.) enterocolitica.
Trotz leicht rückläufiger Tendenzen bei den Fallzahlen ist die Kontrolle dieser wichti-
gen Lebensmittelpathogene derzeit nur von begrenztem Erfolg gezeichnet
(SMULDERS et al. 2008; NEWELL et al. 2010). Während die gegen Salmonellen
implementierten Bekämpfungsprogramme in Teilen Wirkung zeigen (Reduktion von
Salmonellen bei Geflügel und niedrige Prävalenzen beim Schwein in Skandinavien),
ist bei bestimmten Serovaren (S. Typhimurium) ein Anstieg zu verzeichnen. Deutsch-
land gilt nach wie vor als Land mit höherer Prävalenz. Für Campylobacter spp. und Y.
enterocolitica existieren bis dato noch gar keine solche Strategien.
Alle drei Pathogene kommen beim Schwein vor. Dies ist von besonderer Bedeutung,
da Schweinefleisch die am häufigsten konsumierte Fleischart in der europäischen
Union ist. Auch in Deutschland lag Schweinefleisch im Jahr 2008 mit einem Ver-
brauch von 40 kg pro Kopf und Jahr an der Spitze aller Fleischsorten (GURRATH
2008). Bei allen drei Erregern wird ein beachtlicher Anteil humaner Erkrankungsfälle
auf Schweinefleisch bzw. das Reservoir „Schwein“ zurückgeführt. Die besondere
Problematik besteht im meist symptomlosen Auftreten der Pathogene im Bestand.

Einleitung
2
Dies ermöglicht weder eine Identifizierung erregertragender Tiere bei der klassischen
Fleischuntersuchung, noch bei der ante-mortem-Untersuchung auf dem Schlachthof.
Aufgrund dieser fehlenden Möglichkeit der gezielten Eliminierung infizierter Tiere aus
der Lebensmittelkette wird neben Schlachthygienemaßnahmen stattdessen ein risi-
kobasierter Bekämpfungsansatz verfolgt. Dieser zielt auf eine generelle Senkung der
Wahrscheinlichkeit eines Erregereintrages in die Lebensmittelkette ab. Viele Unter-
suchungen belegen, dass die Reduktion der Erregerprävalenzen im Bestand dabei
ein entscheidendes Kriterium ist und zu substantiellen Verringerungen der Kontami-
nation von Schweinefleisch führen kann. Auch der Paradigmenwechsel in der Ge-
setzgebung führt im Zuge des „from-stable-to-table“-Konzeptes zu einer Verlagerung
der Verantwortung für die Lebensmittelsicherheit auf die Primärproduktion (VON
ALTROCK u. WALDMANN 2003). Für eine erfolgreiche Prävention sind allerdings
umfassende Kenntnisse zur Epidemiologie der Erreger im Bestand nötig.
Hier sind auf dem Gebiet von Campylobacter spp. und Y. enterocolitica noch einige
Fragen ungeklärt. Für beide existieren bisher nur wenige Studien zu Risikofaktoren
und Eintragsquellen im Bestand, auch die Rolle der Tierumgebung ist bisher strittig.
Gleiches gilt für Untersuchungen zum gemeinsamen Auftreten aller drei Erreger.
Kenntnisse darüber könnten in der Frage weiterhelfen, ob das Salmonellen-
Monitoring sich auch für eine Risikoabschätzung zum Eintrag der anderen beiden
Pathogene in die Lebensmittelkette eignet, also ein Rückschluss von der Salmonel-
lenkategorie auf die Belastung mit Campylobacter spp. und Y. enterocolitica möglich
ist. Außerdem können diese Kenntnisse eine Aussage über mögliche gemeinsame
Risikofaktoren und somit identische Bekämpfungsstrategien in den Schweinebestän-
den ermöglichen. Dies böte einen großen Vorteil, da Landwirte bereits bei Salmonel-
len aufgrund deren symptomloser Manifestation mitunter wenig Problembewusstsein
entwickeln und die Auferlegung weiterer Maßnahmen möglicherweise als unzumut-
bare Belastung empfunden würde.
Zur Epidemiologie der Salmonellen ist bereits vieles bekannt, und es wurden zahlrei-
che Risikofaktoren und Eintragsquellen identifiziert. Dagegen ist bisher strittig, wel-

Einleitung
3
chen Stellenwert die einzelnen Quellen und Transmissionsrouten im Infektionsge-
schehen einnehmen. In den letzten Jahren ist die Entwicklung molekularbiologischer
Feintypisierungsmethoden weit fortgeschritten, und mit ihr die Möglichkeiten zur
Rückverfolgung einzelner Isolate. Der Einsatz dieser Methoden kann detaillierte Er-
kenntnisse über die genaue Dynamik der Ausbreitung und Verteilung von Salmonel-
len zwischen Tier und Umgebung im Verlauf eines Mastdurchganges ergeben. Diese
könnten den Landwirten gezielte Hinweise liefern, wo sie mit Bekämpfungsmaßnah-
men ansetzen sollen. Das ist deshalb wichtig, da die Landwirte aufgrund der bereits
bestehenden Salmonellen-Bekämpfungsprogramme rechtlich dazu verpflichtet sind,
aber mitunter trotz ernsthafter Bemühungen wenig langfristigen Erfolg verzeichnen.
Daher sollen in dieser Dissertation folgende Fragestellungen untersucht werden:
1. Im Rahmen einer Querschnittserhebung in Schweinemastbeständen soll das Vor-
kommen von Campylobacter spp., Y. enterocolitica und Salmonella spp. in den Tie-
ren und besonders in ihrer Umgebung eruiert werden. Daneben soll untersucht wer-
den, ob und in welchen Kombinationen die Erreger gemeinsam im Betrieb auftreten,
auch unter besonderer Berücksichtigung der Salmonellen-Kategorie. Abschließen
wird dieses Teilprojekt eine Analyse möglicher Einflussfaktoren für das Vorkommen
der Erreger im Bestand.
2. Im Rahmen einer Verlaufsuntersuchung sollen Schweinemastbetriebe über meh-
rere Mastdurchgänge untersucht und das Salmonellenvorkommen bei Tieren und in
der Umgebung von Anfang bis Ende der Mast verfolgt und auf gemeinsame Muster
analysiert werden. Zur besseren Rückverfolgbarkeit der Transmissionsrouten einzel-
ner Isolate werden diese bis auf klonale Ebene feintypisiert.
Die Untersuchungen finden im Weser-Ems-Gebiet in Niedersachsen statt. Die Regi-
on eignet sich im Besonderen dafür, da sich hier das Zentrum der Schweineprodukti-
on Deutschlands befindet und die untersuchten Betriebe somit strukturell repräsenta-
tiv für einen Großteil der deutschen Schweinehaltungen sind.

Literaturübersicht 4
2. Literaturübersicht
2.1 Salmonella spp.
2.1.1 Taxonomie und Nomenklatur
Das Genus Salmonella (Familie der Enterobacteriaceae Ordnung Enterobacterales,
Klasse Gammaproteobacteria, Stamm Proteobacteria) beinhaltet eine Vielzahl von
Krankheitserregern beim Menschen und zahlreichen Tierarten (SELBITZ 2002).
Seit Einführung der Genusbezeichnung Salmonella im Jahr 1900 durch Lignieres zu
Ehren des Entdeckers des Schweinecholera-Bacillus 1885, Salmon, entwickelte sich
das Spezieskonzept der Gattung in verschiedenen Phasen: Zunächst basierte die
Taxonomie auf der klinischen Relevanz bzw. Wirtsbezogenheit der einzelnen Stäm-
me. Nachdem sich einige Stämme als ubiquitär erwiesen, wurden antigenetische Ei-
genschaften Klassifizierungsgrundlage. Nach dem serologischen Schema von Kauff-
mann und White wurden mehrere tausend Serovare unterschieden, die alle eigene
Spezies darstellten. In dieser Zeit begann die Benennung der Stämme nach ihrem
Entdeckungsort. Später erfolgte die Spezieseinteilung anhand biochemischer Cha-
rakteristika; durch KAUFFMANN wurden die Subgenera, die Spezies entsprachen,
mit römischen Zahlen benannt (I - VI) (KAUFFMANN 1960; LE MINOR u. POPOFF
1987). Das den Regeln des Bacteriological Code konforme Schema teilte das Genus
in die Spezies Salmonella (S.) arizonae, S. bongori, S. choleraesuis, S. enteritidis,
S. paratyphi, S. typhi und S. typhimurium. Nach Untersuchungen, die die Zugehörig-
keit dieser Stämme zu einer einzigen DNA-Hybridisierungsgruppe belegten, wurden
sie einer einzigen Spezies zugeordnet. Diese erhielt den Namen Salmonella chole-
raesuis und umfasste sieben Subspezies. Da die Bezeichnung choleraesuis eine
Verwechslungsgefahr mit dem gleichnamigen Serovar birgt, schlugen LE MINOR
und POPOFF (1987) Salmonella enterica als einzige Spezies der Gattung mit den
Subspezies arizonae, bongori, enterica, diarizonae, houtenae, indica und salamae
vor. Diese Einteilung, obwohl nicht offiziell anerkannt, fand schnell weite Verbreitung

Literaturübersicht
5
in der Wissenschaft, während die regelkonformen Bezeichnungen kaum verwendet
wurden. Um diese Diskrepanzen zu beseitigen, wurde im Jahr 2005 von der „Judicial
Commission of the International Committee on Systematics of Prokaryotes“ die Ein-
teilung der Gattung in die beiden Spezies Salmonella bongori (ehemals Ssp. V) und
Salmonella enterica, die die Subspezies Salmonella enterica enterica (Ssp. I), Sal-
monella enterica salamae (Ssp. II) Salmonella enterica arizonae (Ssp. IIIa), Salmo-
nella enterica diarizonae (Ssp. IIIb), Salmonella enterica houtenae (Ssp. IV) und
Salmonella enterica indica (Ssp. VI) umfasst, anerkannt (TINDALL et al. 2005). Im
selben Jahr wurde eine neue Spezies, Salmonella subterranea, publiziert, die aller-
dings eng mit Escherichia hermannii verwandt ist und nicht dem Genus Salmonella
zugerechnet wird (EUZÉBY 2005).
Zur Vereinfachung der Schreibweise wird für Salmonellen der Subspezies I (enterica
enterica) das Serovar, groß und nicht kursiv geschrieben, direkt hinter dem Gat-
tungsnamen aufgeführt, z. B. Salmonella Typhimurium. (LE MINOR u. POPOFF
1987). Serovare der anderen Subspezies erhalten keine Namen, sondern werden
über ihre antigenetische Formel der O- und H-Antigene nach dem Kauffmann-White-
Schema benannt. Insgesamt waren bis zum Jahr 2007 2.610 verschiedene Serovare
registriert (GUIBOURDENCHE et al. 2010): bei S. enterica 2.587 (davon Ssp. enteri-
ca 1.547) und bei S. bongori 23 Serovare.
2.1.2 Erregereigenschaften
Salmonella spp. sind 0,7 - 1,5 x 2,0 - 5,0 μm große gramnegative, fakultativ anaero-
be Stäbchen. Sie sind mit Ausnahme von S. Gallinarum peritrich begeißelt und be-
weglich. Ihr Stoffwechsel ist fermentativ, sie reduzieren Nitrat zu Nitrit, bilden Schwe-
felwasserstoff, nutzen Citrat als alleinige Kohlenstoffquelle und bauen Propylenglycol
ab. Sie sind im Gegensatz zu anderen Enterobakterien laktosenegativ, ausgenom-
men die Subspezies (Ssp.) arizonae und diarizonae (SELBITZ 2002).

Literaturübersicht 6
Salmonellen verfügen über eine hohe Tenazität. Sie vermehren sich bei Temperatu-
ren von 5 C -45 C, überleben auch in gekühlten oder tiefgefrorenen Lebensmitteln,
tolerieren pH-Werte von 4,5 bis 9,0 und Trockenheit und sterben erst unter einem aw-
Wert von unter 0,93 ab (DEDIÉ et al. 1993). Salmonella spp. können bei minimalem
Nährstoffangebot längere Zeit in Kot, Schlamm, Wasser, Staub, Erdboden und Le-
bensmitteln überleben. Für Tierkot wurden Überlebenszeiten von bis zu drei Jahren
beobachtet, für Flüssigmist, Heu und Erdboden ein Jahr, für Staub beinahe vier Jah-
re. Bei Temperaturen ab 60 C sterben sie dagegen innerhalb von fünf bis maximal
25 Minuten ab (BÖHM 1993).
Während die Subspezies I fast ausschließlich bei Warmblütern auftritt, werden Sub-
spezies II und III vorrangig bei poikilothermen Tieren nachgewiesen und sind für
Säugetiere wenig pathogen. Ssp. IV umfasst für Säuger ebenfalls wenig pathogene
Umweltisolate, gleiches gilt für die ehemalige Ssp. V (DEDIÉ et al. 1993).
Anhand ihrer Wirtsspezifität lassen sich drei verschiedene epidemiologisch relevante
Gruppen unterscheiden (BLAHA 1993):
1. Epidemisch vorkommende, speziesadaptierte Serovare wie z. B. S. Typhi und
Paratyphi (Mensch), S. Choleraesuis (Schwein), S. Gallinarum (Huhn),
S. Dublin (Rind): Diese verursachen bei den betroffenen Spezies Erkrankun-
gen, stellen jedoch im Falle der tieradaptierten Serovare keine Gefährdung für
die menschliche Gesundheit dar.
2. Sporadisch vorkommende, nicht speziesadaptierte Serovare wie z. B. S. Ago-
na und S. Infantis: Diese verfügen über zoonotisches Potential, sind aufgrund
ihres sporadischen Auftretens jedoch nicht von großer Relevanz für die
menschliche Gesundheit.
3. Endemisch vorkommende, nicht speziesadaptierte Serovare: S. Typhimurium,
S. Enteritidis: Diese stellen eine ernst zu nehmende Gesundheitsgefährdung
des Menschen dar, weil sie in erheblichen Mengen in die Lebensmittelkette
eingetragen werden können.

Literaturübersicht
7
In einem anderen Einteilungsschema wird nach dem Grad der Wirtsadaptation unter-
schieden: Neben den nicht wirtsadaptierten Serovaren gibt es wirtsspezifische Sero-
vare, die stark mit einer einzelnen Spezies assoziiert sind (S. Typhi, S. Gallinarum
oder S. Typhisuis). Solche, die hauptsächlich bei einer Spezies vorkommen, aber
auch bei anderen Tierarten Krankheiten verursachen können, werden als wirtsadap-
tiert bezeichnet (S. Dublin und S. Choleraesuis) (UZZAU et al. 2000).
2.1.3 Nachweismethoden
2.1.3.1 Kultureller Nachweis
Für den kulturellen Nachweis von Salmonella spp. existieren vermutlich mehr unter-
schiedlichen Medien und Kultivierungsprotokolle als für die meisten anderen Bakte-
rienarten (DAVIES et al. 2000). Direktes Ausstreichen auf Nährböden ist bei nur ge-
ring kontaminiertem Probenmaterial möglich, ansonsten empfiehlt sich eine Voran-
reicherung. Bei sehr geringen vermuteten Keimzahlen oder subletal geschädigten
Erregern sollte eine nicht-selektive Anreicherung mit gepuffertem Peptonwasser
(BPW) für 12 - 24 Stunden bei 37 C erfolgen (DEDIÉ et al. 1993). Eine selektive
Anreicherung kann durch direkte Beimpfung oder nach der Voranreicherung erfolgen
(SELBITZ 2002). Unter den flüssigen Selektivanreicherungsmedien sind die Selenit-
oder die Tetrathionatboullion gebräuchlich. Die ebenfalls weit verbreitete Boullion
nach Rappaport-Vassiliadis (RV) besitzt diesen gegenüber den Vorteil einer Hem-
mung von Proteus. Die parallele Bebrütung bei 37 C und bei 42 C erhöht die Aus-
beute (DEDIÉ et al. 1993). Das halbfeste modifizierte Rappaport-Vassiliadis-Medium
(MRSV) erlaubt, die Fähigkeit des Schwärmens bei Salmonellen zu detektieren
(BUSSE 1995).
Eine Supplementierung der Voranreicherungs- oder Anreicherungsmedien mit Ferri-
oxamin E kann die Erregerausbeute zusätzlich erhöhen (REISSBRODT et al. 1996).

Literaturübersicht 8
Nach 24 Stunden und, wenn nötig, nach 48 - 96 Stunden erfolgt das Ausstreichen
auf zwei verschiedenen festen Nährböden. Da Salmonellen auf Universalnährböden
in der Regel nicht von Kolonien anderer Enterobakterien zu unterscheiden sind, wer-
den je nach vermuteter Salmonella-Spezies und erwarteter Begleitflora verschiedene
Differenzierungs- und Selektivnährböden verwendet. Diese zeigen Salmonella spp.
anhand ihrer Stoffwechselaktivitäten durch Schwärzung (H2S-Bildung) oder Farbum-
schlag (Abbau von Propylenglycol, Fermentation von Saccharose, fehlender Lakto-
seabbau, ß-Galaktosidaseaktivität etc.) an. Zu den selektiv wirkenden Medien zählen
Brillantgrün-Phenolrot-Laktose-Saccharose (BPLS)-Agar, Desoxycholat-Zitratagar,
Wismutsulfitagar, Xylose-Lysin-Desoxycholat (XLD)-Agar und Xylose-Lysin-Tergitol
(XLT4)-Agar. Weniger selektiv aber stärker differenzierend sind beispielsweise
Gassner-Agar, MacConcey-Agar, Eosin-Methylenblau (EMB)-Agar oder Rambach-
Agar. Die Bebrütung erfolgt bei 37 C für 24 - 48 Stunden. Verdächtigte Kulturen
können über eine Objektträgerschnellagglutination vorläufig identifiziert werden, soll-
ten jedoch anschließend in Reinkultur angelegt und durch biochemische und serolo-
gische Methoden zu Spezies, Subspezies- und Serovar differenziert werden (DEDIÉ
et al. 1993; SELBITZ 2002).
Ein internationales Standardverfahren zu Nachweis und Differenzierung beschreibt
die Norm EN ISO 6579:2002 (ANONYMOUS 2002).
2.1.3.2 Phänotypische Differenzierungsmethoden
Die Speziesdifferenzierung erfolgt anhand verschiedener biochemischer Reaktionen,
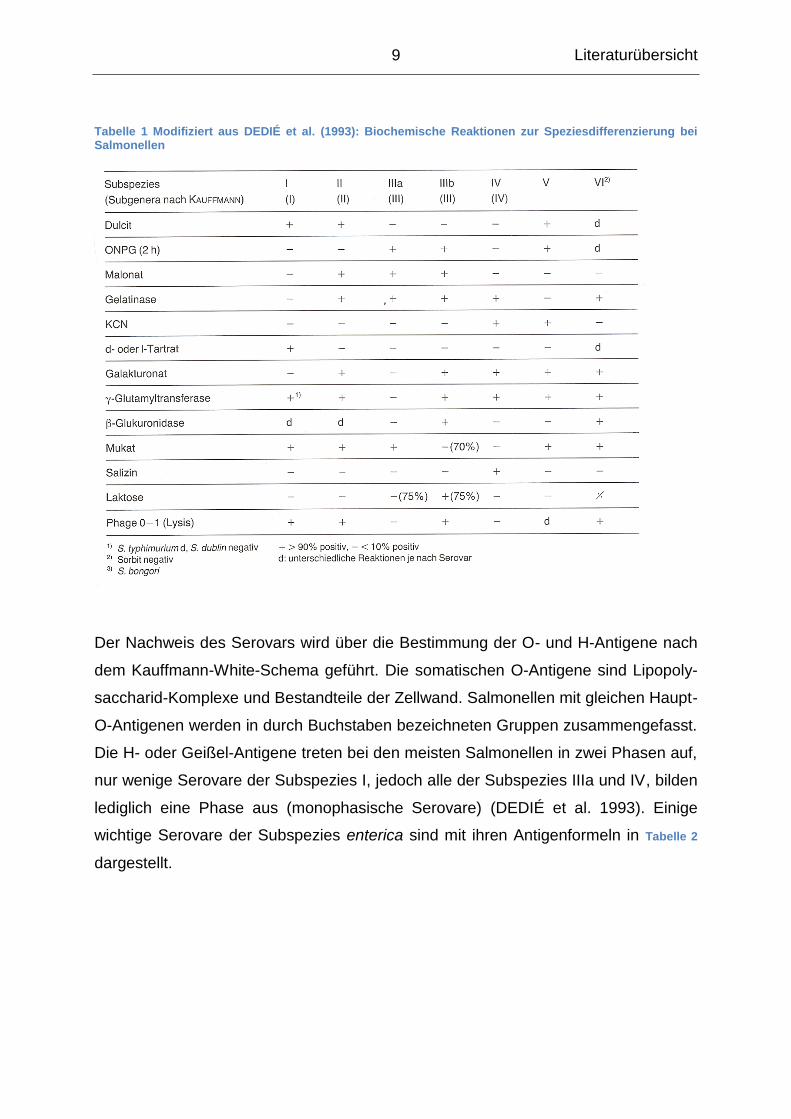
die in Tabelle 1 dargestellt werden. Zudem sind verschiedene kommerzielle Testsys-
teme erhältlich, zum Beispiel das API20 E® (Biomérieux Deutschland, Nürtingen) o-
der das BD BBL Enterotube II® (Beckton Dickinson GmbH, Heidelberg).
Literaturübersicht
9
Tabelle 1 Modifiziert aus DEDIÉ et al. (1993): Biochemische Reaktionen zur Speziesdifferenzierung bei Salmonellen
Der Nachweis des Serovars wird über die Bestimmung der O- und H-Antigene nach
dem Kauffmann-White-Schema geführt. Die somatischen O-Antigene sind Lipopoly-
saccharid-Komplexe und Bestandteile der Zellwand. Salmonellen mit gleichen Haupt-
O-Antigenen werden in durch Buchstaben bezeichneten Gruppen zusammengefasst.
Die H- oder Geißel-Antigene treten bei den meisten Salmonellen in zwei Phasen auf,
nur wenige Serovare der Subspezies I, jedoch alle der Subspezies IIIa und IV, bilden
lediglich eine Phase aus (monophasische Serovare) (DEDIÉ et al. 1993). Einige
wichtige Serovare der Subspezies enterica sind mit ihren Antigenformeln in Tabelle 2
dargestellt.

Literaturübersicht 10
Tabelle 2 Modifiziert aus SELBITZ (2002): Wichtige Salmonella-Serovare mit ihren Antigenformeln nach dem Kauffmann-White-Schema
Die Serovarbestimmung wird mit kommerziell erhältlichen O- und H-Antiseren in der
Objektträgeragglutination durchgeführt.

Literaturübersicht
11
Vervollständigt wird die phänotypische Charakterisierung eines Isolats durch die
Lysotypie, in der das Lyseverhalten bei bestimmten Bakteriophagen überprüft wird,
und durch die Bestimmung von Resistenzmustern (LIEBANA 2002).
Diese phänotypischen Methoden sind traditionell die Basis für epidemiologische Un-
tersuchungen. Die Typisierung eines Erregers ist Grundvoraussetzung dafür, denn
sie ermöglicht die Differenzierung von sporadisch und epidemisch auftretenden
Stämmen und die Rückverfolgung von Infektionsketten und -quellen (WICHELHAUS
et al. 2000). Das Vorherrschen bestimmter Phänotypen in Wirten oder Habitaten so-
wie der zunehmende internationale Handel mit immer leichterer Verbreitung neuer
Salmonella-Stämme erfordern inzwischen die zusätzliche Anwendung von Differen-
zierungsmethoden mit über die Phänotyp-Ebene hinaus gehendem diskriminatori-
schen Potential (LIEBANA 2002)
2.1.3.3 Molekularbiologische Nachweis- und Differenzierungsmethoden
Molekularbiologische (DNA-basierte) Methoden finden auf allen Ebenen von der
Speziesdiagnose bis zur Erreger-Feintypisierung Anwendung. Der molekularbiologi-
sche Erregernachweis bietet gegenüber dem kulturellen den Vorteil eines geringeren
Zeit- und Arbeitsaufwandes. Daher wurden in den vergangenen Jahren verschiedene
PCR-Assays entwickelt, beispielsweise zum Nachweis des Salmonella-spezifischen
invA-Gens, des Plasmid-assoziierten Virulenzgens spvC, oder des hilA-Gens aus Kot,
Organen oder Lebensmitteln (AABO et al. 1995; CHIU u. OU 1996; SCHOLZ et al.
2001; PATHMANATHAN et al. 2003).
DNA-basierte Typisierungsmethoden finden für epidemiologische Untersuchungen
immer breitere Anwendung. In Kombination mit konventionellen Methoden liefern sie
wertvolle Einsichten in das Verständnis der Epidemiologie vieler Infektionskrankhei-
ten. Sie basieren auf dem Prinzip der Klonalität: dass epidemiologisch miteinander in
Bezug stehende Isolate auf die klonale Expansion eines einzigen Vorläufers zurück-
gehen und gleiche genotypische Charakteristika aufweisen („ein Klon - ein epidemio-

Literaturübersicht 12
logischer Prozess“) (LIEBANA 2002). Die Verwendbarkeit eines bestimmten genoty-
pischen Charakteristikums für die Typisierung hängt dabei von seiner Stabilität in-
nerhalb eines Stammes und seiner Diversität innerhalb der Spezies ab (MASLOW et
al. 1993). Der Grad des genomischen Polymorphismus innerhalb einer Population ist
ein entscheidender Einflussfaktor für die molekularbiologische Epidemiologie. Die
starke Klonalität einiger Salmonella-Serotypen wie S. Enteritidis erfordert hoch dis-
kriminatorische Techniken zur Detektion kleinerer Unterschiede im Genotyp der Iso-
late (HELMUTH u. SCHROETER 1994).
Zur Typisierung von Salmonella spp. existieren unterschiedliche Methoden, die grob
in restriktions-basierte, amplifikations-basierte und sequenz-basierte Verfahren unter-
teilt werden können (FOLEY et al. 2009).
Zu den älteren restriktions-basierten Techniken zählen Plasmidanalysen (TIETZE u.
TSCHAPE 1983). Diese schnelle und relativ einfache Methode hat den Nachteil,
dass extrachromosomale DNA ein teils instabiler genetischer Marker ist und besitzt
somit, abhängig vom Serotyp, nur mäßiges diskriminatorisches Potential (HELMUTH
u. SCHROETER 1994) Bei der RFLP (restriction fragment length polymorphism)-
Analyse wird chromosomale DNA mit Restriktionsendonukleasen in unterschiedliche
lange Fragmente geteilt. Je nach verwendetem Restriktionsenzym können die Er-
gebnisse sehr komplex und schwer interpretierbar sein (LIEBANA 2002). Das Ribo-
typing als Unterform der RFLP-Analyse basiert auf Unterschieden in ribosomalen
RNA-Gensequenzen. Der Nutzen dieser Methode für die Salmonella-Typisierung
hängt vom verwendeten Restriktionsenzym, der Gensonde und dem Ursprung des
Isolats ab. Die Pulsfeld-Gel-Elektrophorese (PFGE), bei der im Rahmen einer Mak-
rorestriktionsanalyse weniger Fragmente mit größerer Masse entstehen und mittels
eines alternierenden multidirektionalen Spannungsfeldes aufgetrennt werden, stellt
den klassischen „Goldstandard“ zur Salmonellen-Typisierung dar (FOLEY et al.
2009). Die Enzyme XbaI und BlnI erwiesen sich als am besten geeignet für die Un-
tersuchung von Salmonella-Stämmen. Die Methode, obwohl arbeits- und zeitauf-
wändig, verfügt über ein außerordentlich hohes diskriminatorisches Potential, unter

Literaturübersicht
13
anderem bei S. Typhimurium (MALORNY et al. 2001). Für die einheitliche Interpreta-
tion des Verwandtschaftsgrades zwischen Isolaten wurden Auswertekriterien erstellt
(TENOVER et al. 1995) und später angepasst (BARRETT et al. 2006). Die geneti-
schen Erregerprofile werden im Netzwerk PulseNet (www.pulsenet-europe.org) in
einer Datenbank erfasst und anderen Personen zugänglich gemacht.
Bei den verschiedenen amplifikations-basierten Methoden werden mittels PCR (po-
lymerase chain reaction) DNA-Sequenzen vervielfältigt. Vorteile sind der relativ ge-
ringe Zeitaufwand und die im Vergleich zu restriktions-basierten Methoden geringere
DNA-Menge, die benötigt wird. Während RFLP-PCR und RAPD (random amplificati-
on of polymorphic DNA) sich als zu gering diskriminatorisch erwiesen, wird die AFLP
(amplified fragment length polymorphism) erfolgreich zum Salmonellen-Fingerprinting
eingesetzt (AARTS et al. 1998). In der relativ neuen VNTR (variable number of tan-
dem repeat)-Analyse wird die Anzahl von tandem repeats (sich direkt wiederholende
DNA-Motive) an einem bestimmten Genlocus bestimmt. Die Zahl der Wiederholun-
gen kann selbst innerhalb derselben Spezies hochvariabel sein und ist deshalb gut
für die Differenzierung nicht-klonaler Isolate geeignet. Eine an verschiedenen Loci
durchgeführte VNTR-Analyse wird als MLVA (multiple locus VNTR analysis) be-
zeichnet. Die MLVA bietet verschiedenen Studien zufolge, beispielsweise für S. Ty-
phimurium, eine noch stärkere Diskriminierung als die PFGE und zudem eine größe-
re Schnelligkeit und bessere Interpretierbarkeit der Ergebnisse (LINDSTEDT et al.
2004; TORPDAHL et al. 2007). Ein etabliertes Verfahren zum Fingerprinting von S.
Typhimurium ist die MLVA nach LINDSTEDT et al. (2004), in der in zwei multiplex
PCR´s die Allele von fünf Loci identifiziert werden. Die Methode weist neben ihrer
Schnelligkeit, Robustheit und einfachen Durchführbarkeit als einzige eine sehr gute
Diskriminierung zwischen Isolaten des Phagentyps DT 104 auf.
Eine sequenzbasierte Methode ist die MLST (multilocus sequence typing), bei der
bestimmte Gene im Hinblick auf die Abfolge ihrer Nukleotidbasen untersucht werden
und daraus eine Einteilung der Isolate in Sequenztypen (ST) erfolgt. Sie hat hohes
diskriminatorisches Potential, das jedoch abhängig von den gewählten Genen und

Literaturübersicht 14
von der Anzahl der sequenzierten Loci ist (FOLEY et al. 2009). Nachteilig ist die
recht arbeitsaufwändige Durchführung (WICHELHAUS et al. 2000). Die SNP (single
nucleotide polymorphism)-Analyse, die Nukleotid-Mutationen an spezifischen Loci
zur Isolatdifferenzierung nutzt, ist eine vielversprechende, aber noch in den Anfän-
gen ihrer Entwicklung stehende Methode (FOLEY et al. 2009).
Da jede der Methoden Polymorphismen an unterschiedlichen Stellen des Genoms
detektiert, sollte die Auswahl des jeweils geeigneten Untersuchungsverfahrens an-
hand des Serotyps und des geographischen Ursprungs eines Isolates erfolgen. Der
erfolgversprechendste Ansatz für ein Fingerprinting von Salmonella spp. für epidemi-
ologische Untersuchungen ist laut LIEBANA (2002) die Kombination von mehreren
unterschiedlichen Methoden.
2.1.3.4 Sonstige Nachweismethoden (indirekter Nachweis)
Für Screening-Untersuchungen zur Feststellung des Herdenstatus von Schweinebe-
ständen wie im Rahmen des Salmonellen-Monitorings der „QS Qualität und Sicher-
heit GmbH“ bzw. der Schweine-Salmonellen-Verordnung (ANONYMOUS 2007b)
werden meist serologische Testverfahren eingesetzt. In Deutschland sind verschie-
dene kommerzielle ELISA-Systeme zum Antikörpernachweis aus Serum oder
Fleischsaft erhältlich: der SALMOTYPE® Pig Screen ELISA (Labor Diagnostik Leipzig,
Leipzig), der HerdCheck® Swine Salmonella ELISA (IDEXX Europe B. V., Niederlan-
de) und der PrioCHECK® Salmonella Ab porcine 2.0 (Prionics Deutschland, Planegg-
Martinsried). Ein positives Ergebnis ist ein Indikator dafür, dass die Tiere in der Ver-
gangenheit Erregerkontakt hatten. Die Präsenz des Erregers im Tier zum Zeitpunkt
der Probenentnahme lässt sich aus einem positiven Antikörpernachweis nicht ablei-
ten. Umgekehrt kann eine erst wenige Tage zurück liegende Infektion ein negatives
Testergebnis nach sich ziehen. Ein Vorteil serologischer Untersuchungsverfahren ist,
dass sie nicht von der stark intermittierenden Erregerausscheidung abhängen (LO
FO WONG et al. 2003).

Literaturübersicht
15
2.1.4 Salmonellosen beim Menschen
Die Salmonellose war 2008 laut Community-Report der EFSA mit 131.468 gemelde-
ten Fällen die zweithäufigste bakterielle Zoonose in der Europäischen Union. Die
beiden wichtigsten involvierten Serovare waren S. Enteritidis mit 58,0 % aller huma-
nen Fälle und S. Typhimurium mit 21,9 % aller Fälle. In den letzten Jahren leicht
rückläufige Fallzahlen sind u.a. auf einen Rückgang des hauptsächlich beim Geflügel
auftretenden Serovars S. Enteritidis um 14,0 % gegenüber dem Vorjahr durch bei
Legehennen implementierte Kontrollprogramme zurückzuführen. Demgegenüber
wurde bei S. Typhimurium, dem häufigsten Serovar beim Schwein, ein Anstieg um
27,1 % gegenüber 2007 verzeichnet (ANONYMOUS 2010a).
In Deutschland, wo die Salmonellose wie in anderen EU-Mitgliedsstaaten melde-
pflichtig ist (ANONYMOUS 2000), wurden 2008 42.909 Fälle, entsprechend einer
Erkrankungsrate von 52,2 Fällen auf 100.000 Einwohner, verzeichnet. Die tatsächli-
chen Fallzahlen dürften jedoch um einiges höher sein (ANONYMOUS 2010a).
Als Salmonellosen im engeren Sinne werden Infektionen mit nicht-typhoidalen, meist
nicht-wirtsadaptierten Serovaren bezeichnet, die sich häufig in Form einer Gastroen-
teritis manifestieren (ANONYMOUS 2009b). Demgegenüber stehen die durch die
humanadaptierten S. Typhi und S. Paratyphi verursachten Allgemeinerkrankungen
Typhus und Paratyphus, die heute nur noch in den Entwicklungsländern von Bedeu-
tung sind (ANDREWS-POLYMENIS et al. 2010).
Klinik
Die Infektionsdosis für Erwachsene liegt bei etwa 104 bis 106 Keimen, bei immunge-
schwächten Patienten oder bei Aufnahme sehr fetthaltiger Lebensmittel bei nur 100
Keimen; die Inkubationszeit beträgt je nach Infektionsdosis und Serovar in der Regel
12 - 36 Stunden (ANONYMOUS 2009b). Eine durchschnittliche Ausscheidungsdauer
von einem Monat bei Erwachsenen bis sechs Monaten bei Kindern bedingt eine ver-
hältnismäßig lang andauernde Ansteckungsfähigkeit mit Enteritis-Salmonellen nach
Infektion (BUCHWALD u. BLASER 1984; BÖHME et al. 2009). Die häufigste Form

Literaturübersicht 16
der Salmonellose ist mit etwa 70 % die akute Gastroenteritis mit Schwindel, Übelkeit,
Kopf- und Bauchschmerzen, Durchfall, leichtem Fieber und manchmal Erbrechen.
Die Symptome dauern etwa drei bis sieben Tage an. Daraus kann sich, besonders
bei prädisponierten Personengruppen (sogenannten „YOPIs“: young, old, pregnant,
immunosuppressed) eine Bakteriämie mit dem klinischen Bild einer Sepsis entwi-
ckeln. Diese kann mit metastatischen fokalen Infektionen verschiedener Organsys-
teme einhergehen; meist sind Endothel, Endo- oder Perikard, Gelenke, Meningen,
Knochen, aber auch die Lunge betroffen (DEDIÉ et al. 1993). Während die Hospitali-
sierungsrate in den letzten Jahren in Deutschland bei 24 % der gemeldeten Erkran-
kungen lag (ROSNER et al. 2010), ist die Gesamtletalität mit unter 0,1 % niedrig: Für
das Jahr 2008 vermeldete das Robert-Koch-Institut 33 Todesfälle mit einem Alters-
median von 79 Jahren (ANONYMOUS 2009b).
Therapie
Bei unkomplizierten Verläufen sollte die Therapie sich auf den Ausgleich der Flüssig-
keits- und Elektrolytverluste beschränken, da Antibiotika die Bakterienausscheidung
verlängern und eine Resistenzentwicklung begünstigen können. Bei schweren Ver-
läufen wie einer Sepsis oder bei YOPIs ist eine antimikrobielle Therapie dagegen
unverzichtbar. Diese sollte aufgrund der zunehmenden Resistenzen bei nicht-
typhoidalen Salmonella spp. unbedingt nach einer Sensibilitätsprüfung des Isolates
erfolgen. Als prinzipiell geeignet gelten Cotrimoxazol, Ampicillin, Cephalosporine der
dritten Generation oder Fluorchinolone wie Ciprofloxacin. Besonders im Zusammen-
hang mit den beiden letztgenannten Gruppen wurde jedoch in den vergangenen Jah-
ren über stark angestiegene Resistenzraten berichtet. Eine weitere besorgniserre-
gende Entwicklung ist die Zunahme von Multiresistenzen, besonders bei S. Typhimu-
rium (PARRY u. THRELFALL 2008).
Übertragungswege und involvierte Lebensmittel
Die Salmonellose ist eine klassische Lebensmittelinfektion. Die Erregeraufnahme
erfolgt meist oral über kontaminierte Speisen oder Getränke. Eine Hauptrolle spielen
dabei nicht ausreichend erhitzte Eier und deren Produkte (S. Enteritidis) sowie rohes

Literaturübersicht
17
oder nicht ausreichend erhitztes Fleisch und Fleischprodukte (meist S. Typhimurium).
Durch Kreuzkontamination können auch primär nicht betroffene Lebensmittel eine
Infektionsquelle darstellen (ANONYMOUS 2009b). Dies wird begünstigt durch die
außerordentlich hohe Fähigkeit von Salmonella spp., außerhalb des Wirtsorganismus
zu überleben und sich zu vermehren (DEDIÉ et al. 1993).
Die Anforderungen an Lebensmittel bezüglich Salmonellen (Erregerfreiheit in 25 g)
sind in den EU-Verordnungen (EG) Nr. 2073/2005 und (EG) Nr. 1441/2007 festgelegt
(ANONYMOUS 2005b, 2007a). Nach Putenfleisch (5,6 %) und frischen Broilern
(5,1 %) war Schweinefleisch im Jahr 2008 mit einer europaweit durchschnittlichen
Nachweisrate von 0,7 % in den getesteten Proben laut EFSA Community-Report das
am dritthäufigsten mit Salmonellen kontaminierte Lebensmittel. In Deutschland ver-
stießen 2008 auf Einzelhandelsebene beispielsweise 2,2 % der Proben frischen
Schweinefleischs, 4,2 % der nicht verzehrfertigen Schweinefleischprodukte und
5,7 % der Proben von zum Rohverzehr vorgesehenem Schweinehackfleisch dage-
gen (ANONYMOUS 2010a). In einer Metastudie zu 34 internationalen wissenschaft-
lichen Arbeiten wurde bezüglich Schweinefleisch ein Durchschnittswert von 8,5 %
Salmonella-kontaminierter Rohprodukte und 3,6 % verzehrfertiger Produkte ermittelt
(MATARAGAS et al. 2008). Einer dänischen Studie zufolge stieg die Salmonella-
Nachweisrate in frischem Schweinefleisch aus dem Einzelhandel von 2002 (1,2 %)
bis 2006 (4,2 %) um das 3,5-fache an (HANSEN et al. 2010). Die am häufigsten in
Schweinefleisch nachgewiesenen Serovare waren 2008 in der Europäischen Union
S. Typhimurium (33,1 %), S. Derby (19,8 %), S. Agona (3,9 %), S. Infantis (3,5 %)
und S. London (3,4 %) (ANONYMOUS 2010a).
Salmonellosen treten neben sporadischen Fällen vergleichsweise häufig in Form von
Ausbrüchen auf. (ANONYMOUS 2009b). Europaweit machten Salmonelleninfektio-
nen 2008 mit 35,4 % den größten Anteil an allen 5.332 von der EFSA registrierten
lebensmittelassoziierten Krankheitsausbrüchen aus (ANONYMOUS 2010a).

Literaturübersicht 18
Schweinfleisch war nach Eiern bzw. Eiprodukten und eihaltigen Backwaren die dritt-
häufigste Ursache von verifizierten Salmonella-Ausbrüchen - mit zunehmender Ten-
denz. In der Schweiz ereignete sich beispielsweise ein 150 Fälle umfassender Aus-
bruch durch S. Typhimurium in Schweinefleisch (ANONYMOUS 2010a). Auch im
Rahmen eines länderübergreifenden Ausbruchs durch S. Typhimurium in Dänemark,
Norwegen und Schweden mit 51 Fällen konnte (dänisches) Schweinefleisch als
Quelle identifiziert werden (BRUUN et al. 2009). Der letzte größere Ausbruch in
Deutschland ereignete sich 2007 mit 244 Fällen in einem Krankenhaus in Fulda, ver-
ursacht durch S. Enteritidis in Apfelschnee (ANONYMOUS 2007c).
Aufgrund komplexer Infektketten ist die Ansteckungsquelle, sowohl bei Ausbrüchen
als auch bei sporadischen Salmonellosen, nicht immer identifizierbar. Dennoch, wird
davon ausgegangen, dass insgesamt mindestens 10 - 20 % aller humanen Salmo-
nellosen auf das Reservoir Schwein zurückzuführen sind (ANONYMOUS 2010b).
Neben der Ansteckung über kontaminierte Lebensmittel sind weitere Infektionsrouten
denkbar. So wurde mehrfach eine direkte oder indirekte Übertragung von Mensch zu
Mensch - meist von mehrfach-resistenten Isolaten - bei prädisponierten Personen als
nosokomiale Infektion beschrieben (ANONYMOUS 2007c). Infektionen durch Tier-
kontakt treten vor allem bei Kleinkindern auf, besonders die Heimhaltung von Repti-
lien besitzt ein hohes Gefährdungspotential (BÖHME et al. 2009). Dies bestätigen
die Ergebnisse von zwei US-amerikanischen Fall-Kontroll-Studien an Säuglingen und
Kleinkindern. Die eine Studie (YOUNUS et al. 2010) identifizierte außerdem den
Kontakt mit Katzen, die andere (JONES et al. 2006) das Reisen ins Ausland als Risi-
kofaktor. Eine dänische Fall-Kontroll-Studie stellte zudem die Einnahme von Antibio-
tika einige Wochen vor der Infektion in Zusammenhang mit einem höheren Salmo-
nelloserisiko (GRADEL et al. 2008).
Generell bietet eine gewissenhafte Küchenhygiene den besten individuellen Schutz,
zumal zur Zeit noch keine Möglichkeit zur Impfung gegen nicht-typhoidale Salmonel-
losen besteht (ANONYMOUS 2009c, b).

Literaturübersicht
19
2.1.5 Salmonella spp. beim Schwein
Die unterschiedlichen Serovare kommen bei einer Vielzahl von Tierarten vor und
weisen ein breites Spektrum an Manifestationsformen vom symptomlosen Trägertum
bis zu letalen Verläufen auf (SELBITZ 2002).
Alle beim Schwein vorkommenden Serovare sind als pathogen für den Menschen
einzustufen (BOYEN et al. 2008a). Noch um 1960 dominierte weltweit das wirtsadap-
tierte Serovar S. Choleraesuis, das beim Schwein schwere systemische Erkrankun-
gen verursacht. Heute ist der Erreger in Asien und Nordamerika weit verbreitet, in
Westeuropa dagegen selten. Das wirtsspezifische Serovar S. Typhisuis ruft ebenfalls
häufig tödlich endende Krankheitsverläufe hervor, tritt aber nur äußerst sporadisch
auf (UZZAU et al. 2000). Von den nicht wirtsadaptierten Salmonellen können theore-
tisch viele verschiedene Serovare beim Schwein vorkommen, klar dominierend ist in
Europa jedoch S. Typhimurium (BOYEN et al. 2008a). In anderen Regionen, z. B.
Kanada, wurden teilweise höhere Nachweisraten von S. Derby beschrieben
(WILKINS et al. 2010). S. Typhimurium kann, vor allem bei Vorliegen einer Resis-
tenzminderung durch Belastungsfaktoren, Erkrankungen bei jungen Schweinen, ge-
legentlich auch bei Zuchtschweinen hervorrufen (WALDMANN u. PLONAIT 2004).
Ein symptomloses Trägertum im Magen-Darm-Trakt überwiegt jedoch deutlich.
BLAHA (1993) verglich metaphorisch die klinischen Salmonellosen mit der „Spitze
eines Eisbergs“, während die weitaus größeren Fraktionen von Kontamination, Infek-
tion und Dauerausscheidertum sich unbemerkt „unter Wasser“ befänden.
Bei der Einschätzung von Prävalenzen muss beachtet werden, dass diese stark von
der diagnostischen Methodik abhängen; In verschiedenen Studien wurden stets hö-
here Nachweisraten mit serologischen als mit direkten Nachweismethoden errechnet
(SANCHEZ et al. 2007; FOSSE et al. 2009).
In der europaweiten Baseline-Studie zur Salmonellenprävalenz in Schlachtschwei-
nen der EFSA (ANONYMOUS 2008a) wurde eine Gesamtnachweisrate von 10,3 %
(n = 1.922) aus Ileocaecal-Lymphknoten von insgesamt 18.663 untersuchten

Literaturübersicht 20
Schweinen ermittelt. Deutschland lag mit 10,9 % bei 2.567 untersuchten Schweinen
leicht darüber, Dänemark wies 7,7 % positive Schweine bei 998 untersuchten Tieren
auf, und Schweden, Finnland und Norwegen hatten zusammen unter 0,5 % positive
Tiere von 1221 untersuchten Schweinen (ANONYMOUS 2008a). In einer Auswer-
tung von Daten des deutschen Salmonellen-Monitorings (siehe Punkt 2.1.5.3) von
5.581 Herden wurde für das letzte Quartal 2007 der Anteil von Herden mit einer sero-
logischen Salmonellenprävalenz von über 40 % mit 3,3 %, der Anteil von Herden mit
20 - 40 % Seroprävalenz mit 12,3 % und der Anteil von Herden mit einer Seroprä-
valenz unter 20 % mit 84,4 % angegeben (MERLE et al. 2009). Die bakteriologische
Untersuchung von Rektuminhalt vom Schlachthof in einer großen deutschen Studie
von 2002 an 11.960 Schweinen aus 679 Herden resultierte bei 3,7 % positiven Tie-
ren in einer Herdenprävalenz von 20 %, während bei der Untersuchung der Ileocae-
cal-Lymphknoten 19 % und serologisch 37 % der Herden positiv waren (STEINBACH
et al. 2002). Die Autoren vermuteten jedoch, dass der Anteil tatsächlich positiver
Herden generell höher einzustufen ist als mit diesen Verfahren nachweisbar.
Die laut der Baseline-Studie am häufigsten aus deutschen Schlachtschweinen iso-
lierten Serovare waren S. Typhimurium (55,4 %), S. Derby (8,9 %), S. Enteritidis
(3,1 %), S. Infantis (2,5 %) sowie S. Brandenburg und S. London (je 1,2 %)
(ANONYMOUS 2008a). In einer Studie an 50 nordwestdeutschen Mastschweinebe-
ständen erwiesen sich 79,6 % der Isolate als S. Typhimurium, 18,3 % als S. Derby
und die restlichen 2,1 % als Salmonellen der Serogruppen C und G
(SOMYANONTANAGUL 2009).
2.1.5.1 Infektionswege
Der Hauptübertragungsweg von Salmonella spp. auf Tierebene ist die orale Aufnah-
me nach fäkaler Ausscheidung. Diese kann direkt, d.h. über die Muttersau oder über
andere infizierte Schweine, oder indirekt, über (un)belebte Vektoren bzw. Futtermittel,
geschehen (BLAHA 1993). Die Infektionsdynamik auf Bestandsebene kann drei ver-
schiedenen Mustern folgen: Den Neueintrag durch Einstallung latent infizierter Jung-

Literaturübersicht
21
tiere bezeichnete BLAHA (2008) als vertikale Eintragsroute, die Einschleppung von
außen durch jegliche Art von Vektoren als horizontale Route, die ständige Re-
Infektion jeder neuen Tiergeneration durch im Betrieb verbleibende Salmonella-
Residuen als Zirkulieren der Infektion.
Studien zu den verschiedenen Infektionswegen sind zahlreich: In einigen Studien
wird die Rolle der Sau für die Ansteckung der Ferkel als gering eingeschätzt, da in
Longitudinalstudien die Nachweisraten bei Sauen und Saugferkeln serologisch als
auch bakteriologisch sehr gering waren oder Sauen andere Serovare (z. B. viel häu-
figer S. Derby) als Mastschweine aufwiesen (KRANKER et al. 2003; NOLLET et al.
2005; VONNAHME et al. 2007). Andere Untersucher ermittelten hohe Prävalenzen
bei Sauen und schlossen auf eine hohe Bedeutung der Sau für die Transmission von
Salmonellen zur nachfolgenden Tiergeneration (MERIALDI et al. 2008; VIGO et al.
2009; WILKINS et al. 2010). Aus einer Studie zu Salmonellen in Zuchtschweinen der
EFSA 2008 an 5.117 Betrieben resultierte eine europaweite Prävalenz von 31,8 %
bakteriologisch Salmonella-positiven Beständen (Sammelkot). In Deutschland waren
20,6 % der 46 untersuchten Ferkelerzeuger positiv (ANONYMOUS 2009a). Die Be-
deutung der direkten Tierumgebung als Infektionsquelle wurde unter anderem bei
MAGISTRALI et al. (2008) belegt, wo in einer Longitudinalstudie mit Typisierung
gleiche Isolate im leeren, gereinigten und desinfizierten Abteil wie auch später bei
den dort eingestallten Tieren nachgewiesen wurden, und auch das Transportfahr-
zeug Salmonellen aufwies. In anderen Studien wurden Personenkontakte, kontami-
niertes Equipment, Gülle, Schadnager, Insekten und Wildtiere als Quelle eines Sal-
monelleneintrages identifiziert (LE MOINE et al. 1987; LANGVAD et al. 2006; SKOV
et al. 2008). SOMYANONTANAGUL (2009) isolierte aus 22,3 % der Umweltproben
von serologisch positiven Mastbeständen (Kategorie III, siehe Punkt 2.1.5.3) Salmo-
nella spp., darunter Probenlokalisationen wie Futtertröge, Trennwände, Lüftung,
Gänge, Treibbretter, Tränken, Waagen und Nagerkot. In einer Studie an drei hoch
belasteten Mastbeständen (Kategorie III) in Nordwestdeutschland konnte in der
Tierumgebung eine kulturelle Nachweisrate von 15,2 % der Proben beobachtet wer-
den (BODE 2008).

Literaturübersicht 22
Futtermittel können bei der Übertragung der beim Schwein seltenen Serovare eine
gewisse Rolle spielen, bei den häufig nachgewiesenen ist dies eher nicht der Fall
(BISPING 1993). Dies bestätigen die laut EFSA Community Report sehr niedrigen
Nachweisraten von Salmonella spp. von 0,2 % in Schweinefutterproben in Deutsch-
land und das Nicht-Vorhandensein von S. Typhimurium und S. Derby unter den zehn
am häufigsten nachgewiesenen Serovaren (ANONYMOUS 2010a).
Die Ausscheidung von Salmonellen erfolgt stark intermittierend. KRANKER et al.
(2003) wiesen in einer Verlaufsuntersuchung bei 53 % der 180 untersuchten
Schweine die Ausscheidung zu mindestens einem Zeitpunkt nach, dagegen wurden
nur bei 3,7 % der Tiere mehr als einmal bakteriologisch Salmonellen im Kot nachge-
wiesen. Die Autoren ermittelten keine Ausscheidung bei Ferkeln zum Zeitpunkt des
Absetzens, anschließend einen Ausscheidungspeak im Flatdeck (ca. 9. Lebenswo-
che), danach reduzierte sich die Ausscheidungsrate bis Ende der Mast teils unter die
Nachweisgrenze. Die höchsten Antikörper-Titer konnten dagegen zur Mitte der Mast
nachgewiesen werden.
NOLLET et al. (2005) beschrieben im Gegensatz dazu einen Anstieg der Ausschei-
dung von Salmonellen beim Absetzen, sowohl bei Sauen als auch Ferkeln. Bei VIGO
et al.(2009) stieg die Ausscheidungsrate bis zum Absetzen an und fiel bis Ende der
Mast unter die Nachweisgrenze, während die Anzahl serologisch positiver Schweine
bis zum Flatdeck abnahm, um danach sukzessive zuzunehmen. Dies bestätigt die
Beobachtungen von KRANKER et al. (2003), dass Ausscheidungs-, aber auch sero-
logische Muster sowohl zwischen Beständen als auch innerhalb von Tiergruppen
sehr unterschiedliche Verläufe annehmen können.
Einen entscheidenden Einfluss auf das Ausscheidungsverhalten latent infizierter
Schweine übt Stress aus. Vermutlich induziert die Exkretion von Katecholaminen ei-
ne erhöhte Wachstumsrate und damit Ausscheidung von Salmonellen (METHNER et
al. 2008). Dadurch erklärt sich die häufig festgestellte Erhöhung der Ausscheidungs-
rate bei Schlachtschweinen zwischen Stall und Schlachthof (HURD et al. 2004).

Literaturübersicht
23
Dass auch zu so späten Zeitpunkten wie auf dem Transportfahrzeug zum Schlacht-
hof oder in den dortigen Wartebuchten noch eine Ansteckung bisher nicht infizierter
Tiere möglich ist, belegen mehrere Studien: In diesen wurde eine äußerst schnelle
Ansteckung von Schweinen selbst nach kurzer Exposition von etwa drei Stunden zu
kontaminierter Umwelt oder infizierten Schweinen belegt (FEDORKA-CRAY et al.
1994; HURD et al. 2001).
Der Erreger gelangt auf dem Schlachthof hauptsächlich durch fäkale (Kreuz)-
Kontamination der Karkassen bei der Eviszeration an das Fleisch (HUMPHREY u.
JORGENSEN 2006), obwohl auch dem Verschmutzungsgrad der Hautoberfläche
Bedeutung beigemessen wird (ALBAN u. STARK 2005). In der EFSA-Baseline-
Studie an Schlachtschweinen waren europaweit insgesamt 8,3 % der Karkassen mit
Salmonellen kontaminiert (ANONYMOUS 2008a).
2.1.5.2 Risikofaktoren und Bekämpfungsmöglichkeiten im Bestand
Die Bekämpfung von Salmonellen in Schweinefleisch kann auf Bestandsebene („pre-
harvest“), auf Schlachthofebene- („harvest“) oder auf nachfolgenden Produktionsstu-
fen („post-harvest“) erfolgen (BOYEN et al. 2008a).
ALBAN und STÄRK (2005) ermittelten die Anzahl geschlachteter Herden mit hoher
Salmonellenprävalenz als wichtigsten singulären Einflussfaktor auf die Kontamination
von Karkassen in dänischen Schlachthöfen. In einer Risikofaktorenanalyse der EFSA
war für ein mit Salmonellen infiziertes Schwein die Wahrscheinlichkeit einer Salmo-
nellenkontamination der Karkasse doppelt so hoch wie für ein nicht infiziertes
(ANONYMOUS 2008b). Sie schätzte, dass eine Reduktion von Salmonella spp. auf
Karkassen um 99 % in einer Reduktion der durch Schweinfleisch verursachten hu-
manen Salmonellosen um bis zu 80 % resultiere (ANONYMOUS 2010b). Daher soll-
ten nationale Kontrollprogramme neben Maßnahmen auf dem Schlachthof auch In-
terventionen in der Primärproduktion umfassen.

Literaturübersicht 24
Die große Anzahl von Studien zu Risikofaktoren für Salmonella spp. in Mastbestän-
den in den letzten Jahren führte zu einer großen Anzahl resultierender Einflussgrö-
ßen auf Ausscheidung oder Seroprävalenz:
Haltung und Management
Ein häufig genannter Faktor ist die positiv mit der Ausscheidungsrate assoziierte
Herdengröße, ebenso wie die Belegdichte (GARCIA-FELIZ et al. 2009). In aus-
schließlich Mastschweine haltenden Betrieben wurden bei ROSSEL et al. (2006)
häufiger Salmonellen nachgewiesen als bei Herden im geschlossenen System,
ebenso bei Zusammenstallen verschiedener Tiergruppen. Gleichzeitige Geflügel-
oder Rinderhaltung durch den Schweinehalter oder die Anwesenheit anderer Haus-
tiere stellten in verschiedenen Studien ein Risiko dar (LANGVAD et al. 2006;
ROSSEL et al. 2006). In einer Studie war der Bezug von Ferkeln aus mehr als drei
unterschiedlichen Herkünften mit höherer Seroprävalenz verbunden (LO FO WONG
et al. 2004). Gleiches galt für offene Buchtenabtrennungen, die einen direkten Tier-
kontakt zwischen den Buchten ermöglichen, während in einer anderen Studie
Teilspaltenboden gegenüber Vollspaltenboden mit einem höheren Risiko eines Sal-
monellennachweises behaftet war (NOLLET et al. 2004).
Fütterung
In viele Studien wurde pelletiertes Futter als Risikofaktor benannt (GARCIA-FELIZ et
al. 2009; WILKINS et al. 2010). Dieser Einfluss wurde auch in einer Metastudie
(O'CONNOR et al. 2008) deutlich, die Autoren hegten jedoch aufgrund der Beschaf-
fenheit der ausgewerteten Studien Zweifel an der hohen Aussagekraft dieses Zu-
sammenhangs. Alle weiteren mit der Fütterung assoziierten Parameter waren in die-
ser Studie nicht mit einem höheren Salmonellenrisiko behaftet. Anderen Autoren zu-
folge erwies sich der Vermahlungsgrad des Futters als Einflussfaktor: grob gemahle-
nes Mehl erwies sich im Hinblick auf Salmonellen als vorteilhaft gegenüber fein ge-
mahlenem (PAPENBROCK et al. 2005; VISSCHER et al. 2009). In den letzten Jah-
ren wurden viele Futtermittelzusätze zur Salmonellenbekämpfung, meist organische
Säuren, entwickelt, die sich günstig auf Fermentationsprozesse und den pH-Wert im

Literaturübersicht
25
Magen-Darm-Trakt auswirken sollen (BOYEN et al. 2008a). In einer Studie (BOYEN
et al. 2008b) reduzierte gekapselte Buttersäure die Salmonellenlast im Darm. Bei
ROSSEL et al. (2006) war die Nicht-Ansäuerung von Futter und Trinkwasser ein Ri-
sikofaktor, während in einer anderen Studie (MARTIN-PELAEZ et al. 2010) weder
Laktulose noch Ameisen- oder Milchsäure eine Wirkung zeigten. Als protektiv wur-
den außerdem Mehl- und Flüssigfütterung aufgeführt (HAUTEKIET et al. 2008). Zu-
dem wurde ein positiver Zusammenhang zwischen Nüchterungsdauer vor der
Schlachtung und Salmonellenausscheidung zum Zeitpunkt der Schlachtung belegt
(MARTIN-PELAEZ et al. 2009).
„Biosecurity“
Bei HAUTEKIET et al. 2008 hatten das Vorhandensein einer Hygieneschleuse und
eines Schuhbades, die Belegung im strengen Rein-Raus-Verfahren, eine Abteiltem-
peratur im thermoneutralen Bereich und die Desinfektion der Abteile vor Belegung
einen protektiven Einfluss. Eine Leerstehzeit der Abteile unter drei Tagen und unzu-
reichende Reinigung und Desinfektion dieser wurden als Risikofaktor identifiziert
(BELOEIL et al. 2004), ebenso wie eine Desinfektion der Abteile ohne vorangegan-
gene Reinigung (VAN DER WOLF et al. 2001). Einen weiteren Risikofaktor repräsen-
tierte bei LO FO WONG et al. (2004) eine mangelnde Händereinigung vor dem Um-
gang mit den Tieren und das Fehlen einer Toilette.
Tierbehandlungen
Der metaphylaktische Einsatz von Antibiotika war bei ROSSEL et al. (2006) mit einer
erhöhten Seroprävalenz assoziiert. Der leistungsfördernde Einsatz von z. B. Tylosin,
der bei u.a. VAN DER WOLF et al. (2001) mit erhöhter Seroprävalenz einherging,
spielt heute aufgrund generellen Verbots antibiotischer Wachstumspromotoren keine
Rolle mehr. Der therapeutische Einsatz von Antibiotika bei subklinischen Salmonel-
losen ist kontraindiziert: Die Erregerausscheidung wird nicht verkürzt und die Gefahr
einer Resistenzbildung besteht (MEYER et al. 2004), da die Bakterien aufgrund ihre
intrazellulären Lage häufig nicht erreicht werden (WALDMANN u. PLONAIT 2004).
Zur Immunprophylaxe sind in Deutschland derzeit je ein Lebendimpfstoff gegen S.

Literaturübersicht 26
Choleraesuis (Suisaloral®, IDT Biologika GmbH, Dessau-Roßlau) und einer gegen S.
Typhimurium (Salmoporc®, IDT Biologika GmbH, Dessau-Roßlau) zugelassen. Letz-
terer soll klinische Erkrankungen weitgehend verhindern (SELKE et al. 2007) und
nicht zu erhöhten Antikörper-Titern bei der Untersuchung von Schlachtschweinen
führen.
Andere Erkrankungen
Koinfektionen mit PRRS-Virus und Lawsonia intracellularis erhöhten die Ausschei-
dung von Salmonellen (BELOEIL et al. 2004). Den gleichen Effekt hatte eine hoch-
gradige Infektion mit Ascaris suum (VAN DER WOLF et al. 2001). Auch allgemein
begünstigt eine Immundefizienz im Zuge anderer Infektionen die Empfänglichkeit
gegenüber Salmonellen (BOYEN et al. 2008a).
Saisonalität
In einer Studie von VONNAHME et al. (2007) wurde bei Sauen von einer geringeren
Seroprävalenz im Sommer als im Winter berichtet.
Die Vielzahl möglicher Eintragsquellen, Infektionskreisläufe und Risikofaktoren erfor-
dert eine sehr genaue Analyse der jeweiligen Situation im Bestand (DEDIÉ et al.
1993). Für BLAHA (2008) stellt „ein neues, betriebs- und problemspezifisches, alle
praktischen und theoretischen Rückzugsmöglichkeiten von Salmonellen in Betracht
ziehendes“ Hygiene- und Managementkonzept den Schwerpunkt in der Salmonel-
lenbekämpfung im Bestand dar. WALDMANN und PLONAIT (2004) schränkten ein,
dass eine vollkommene Salmonellenfreiheit in Schweinebeständen nicht möglich sei,
auch nicht durch Medikation, Impfung oder Eradikation erkannter Träger. Dies deckt
sich mit der Einschätzung von BLAHA (1993), dass Salmonellen im Bestand ein
quantitatives Problem darstellten und die Höhe des Infektionsdrucks gesenkt werden
müsse.

Literaturübersicht
27
2.1.5.3 Kontroll- und Bekämpfungsprogramme
Zur Reduktion von Salmonella spp. in der Schweinehaltung existieren in verschiede-
nen Ländern freiwillige oder obligatorische Überwachungs- und Kontrollprogramme.
In Schweden, Norwegen und Finnland wurden bereits vor längerer Zeit Bekämp-
fungssysteme implementiert. Dort werden mittels bakteriologischer Untersuchungen
sämtliche Stufen der Lebensmittelproduktion überwacht und in Interventionen einbe-
zogen. Die Programme erwiesen sich als wirkungsvoll, den drei Ländern wurde von
der EFSA als europaweit einzige der Status „niedrige Salmonellenprävalenz“ zuge-
sprochen (ANONYMOUS 2006b).
Das 1995 eingeführte dänische System basiert auf der serologischen Testung von
Fleischsaftproben auf dem Schlachthof und nachfolgenden Einteilung der Bestände
in verschiedene Risikokategorien. Dänemark erhielt als einziges europäisches Land
den Status „mittlere Salmonellenprävalenz“, während alle übrigen europäischen
Staaten als Länder mit „höherer Salmonellenprävalenz“ gelten (ANONYMOUS
2006b).
Ähnliche serologische Programme werden in Großbritannien, Irland, Belgien und den
Niederlanden durchgeführt (bei letzteren beiden anhand von Blutproben im Bestand)
(BOYEN et al. 2008a).
Auch in Deutschland existiert seit 2002 ein an das dänische System angelehntes
serologisches Salmonellen-Monitoring. Dieses wurde zunächst im freiwilligen Rah-
men von der QS Qualität und Sicherheit GmbH eingeführt. Seit 2007 ist es Teil der
„Schweine-Salmonellen-Verordnung“ (ANONYMOUS 2007b). Darin sind regelmäßi-
ge, obligate Untersuchungen von Fleischsaft oder Blutproben aller Mastbetriebe auf
Salmonellen-Antikörper vorgeschrieben. Jeder Betrieb muss pro Jahr (abhängig von
seiner Größe) bis zu 60 Tiere untersuchen lassen. Anhand der Ergebnisse der letz-
ten zwölf Monate werden alle Betriebe vierteljährlich einer von drei Risikokategorien
zugeordnet: Kategorie I entspricht unter 20 % positiver Proben, Kategorie II
20 - 40 % und Kategorie III über 40 % (MERLE et al. 2007). Dabei wird hier ein hö-

Literaturübersicht 28
herer cut-off für ein positives Ergebnis zugrunde gelegt als im dänischen System.
Betriebe in Salmonellen-Kategorie III sind verpflichtet, bakteriologische und epidemi-
ologische Untersuchungen zur Feststellung der Eintragsquelle und Hygienemaß-
nahmen zur Salmonellenreduktion durchzuführen (ANONYMOUS 2007b). Die Be-
triebe müssen zudem mit Sanktionen durch die Schlachthöfe (Schlachtung nur an
bestimmten Tagen oder ganz zuletzt am Tag etc.) rechnen.

Literaturübersicht
29
2.2 Campylobacter spp.
2.2.1 Erregereigenschaften
Die Familie der Campylobacteraceae (Ordnung Campylobacterales, Klasse Epsilon-
proteobacteria, Stamm Proteobacteria) besteht aus den Genera Arcobacter, Depha-
lospirillum, Sulfurospirillum und Campylobacter. Letztere umfasst zur Zeit mehr als
20 Spezies und einige Subspezies (EUZÉBY 2010), die als Kommensalen oder pa-
rasitär bei einer Vielzahl von Warmblütern vorkommen (URSING et al. 1994). Darun-
ter sind C. avium, C. fetus (ssp. fetus oder ssp. venerealis), C. coli, C. jejuni (ssp.
jejuni oder ssp. doylei), C. lari, C. upsaliensis, C. hyointestinalis, C. cuniculorum und
C. sputorum.
Bakterien der Gattung Campylobacter (griechisch für „gebogener Stab“) sind spiral-
förmige oder gebogene, teils auch gerade, 0,5 – 5,0 μm mal 0,2 – 0,9 μm große
Stäbchen. In älteren Kulturen neigen sie zu sphärischen bis kokkoiden Wuchsformen.
Sie sind gramnegativ, nicht sporenbildend und beweglich durch uni- oder bipolare,
monotriche Begeißelung (VANDAMME et al. 1991).
Ein Bakterienwachstum erfolgt unter mikroaerophilen Bedingungen bei einem spe-
ziesabhängigen Temperaturoptimum zwischen 30 C und 42 C. Zu den thermophi-
len Campylobacter spp. mit einem Wachstumsoptimum von 42 C zählen die zoono-
tisch wichtigsten Spezies C. jejuni, C. coli, C. lari und C. upsaliensis (BUTZLER
2004). Campylobacter spp. produzieren Oxidase, Kohlenhydrate können weder oxi-
dativ noch fermentativ metabolisiert werden. Der Guanin- und Cytosingehalt der DNA
reicht von 28 bis 46 mol-% (VANDAMME et al. 1991). Campylobacter spp. weisen
eine geringe Tenazität auf, ihre Überlebensfähigkeit ist besonders in einer trockenen,
relativ warmen und aeroben Atmosphäre gering (CORRY et al. 1995). Unter subop-
timalen Bedingungen wird der Übergang von Kolonien in so genannte „VBNC“-
Stadien („viable but non-culturable“) beschrieben. Diese sind lebens-, aber nicht
mehr vermehrungsfähig (JONES et al. 1991).

Literaturübersicht 30
2.2.2 Nachweismethoden
2.2.2.1 Kultureller Nachweis
Campylobacter spp. wachsen langsam und stellen hohe Ansprüche an ihre Anzucht.
(ON 1996). Nach Identifizierung als eigene Gattung erfolgte der Nachweis von
Campylobacter spp. zunächst durch Membranfiltration von Kotproben und Anzucht
auf nicht-selektiven Nährmedien. Im Jahr 1977 wurde das erste Selektivmedium
entwickelt (SKIRROW 1977). Seitdem führten methodische Verbesserungen zur
Entwicklung sensitiverer und spezifischer Selektivmedien für die Kultivierung von
Campylobacter spp. (MOORE et al. 2005). Diese beinhalten zur Inhibition der kom-
petitiven Keimflora verschiedene Antibiotika wie z. B. Cephalosporine, Trimethoprim,
Polymixin B, Vancomycin sowie Antimykotika. Des Weiteren enthalten sie zum
Schutz der Bakterien vor toxischen Effekten durch Sauerstoffderivate lysiertes oder
defibriniertes Blut, Kohle oder andere neutralisierende Komponenten. Beispiele für
gängige Selektivmedien sind mCCD (modified Charcoal, Cefoperazone and deoxy-
cholate)-Agar, Karmali-Agar, Preston-Agar, Thioglycolat-Agar, halbfestes Medium
nach Goossens oder Butzler-Agar. Diese ursprünglich für den Nachweis aus Kotpro-
ben konzipierten Medien werden ebenfalls zur Untersuchung von Lebensmittel- oder
Wasserproben verwendet. Als vorgeschalteter Schritt empfiehlt sich eine Anreiche-
rung in einem Flüssigmedium wie Preston-Bouillon, VTP-FBP (vancomycin, trime-
thoprim, polymixin B - ferrous sulphate, sodium bisulfite, sodium pyruvate)-Bouillon
oder Park-and-Sanders-Bouillon (CORRY et al. 1995). Die Standard-Methode zum
Nachweis, nicht nur aus Lebensmittel- und Umweltproben sondern auch aus Milch
und Kotproben, ist laut Mitteilung durch das Friedrich-Loeffler-Institut (PROBST
2010) die Norm DIN ISO 10272-1:2006 (ANONYMOUS 2006).
2.2.2.2 Weiterführende Nachweis- und Differenzierungsmethoden
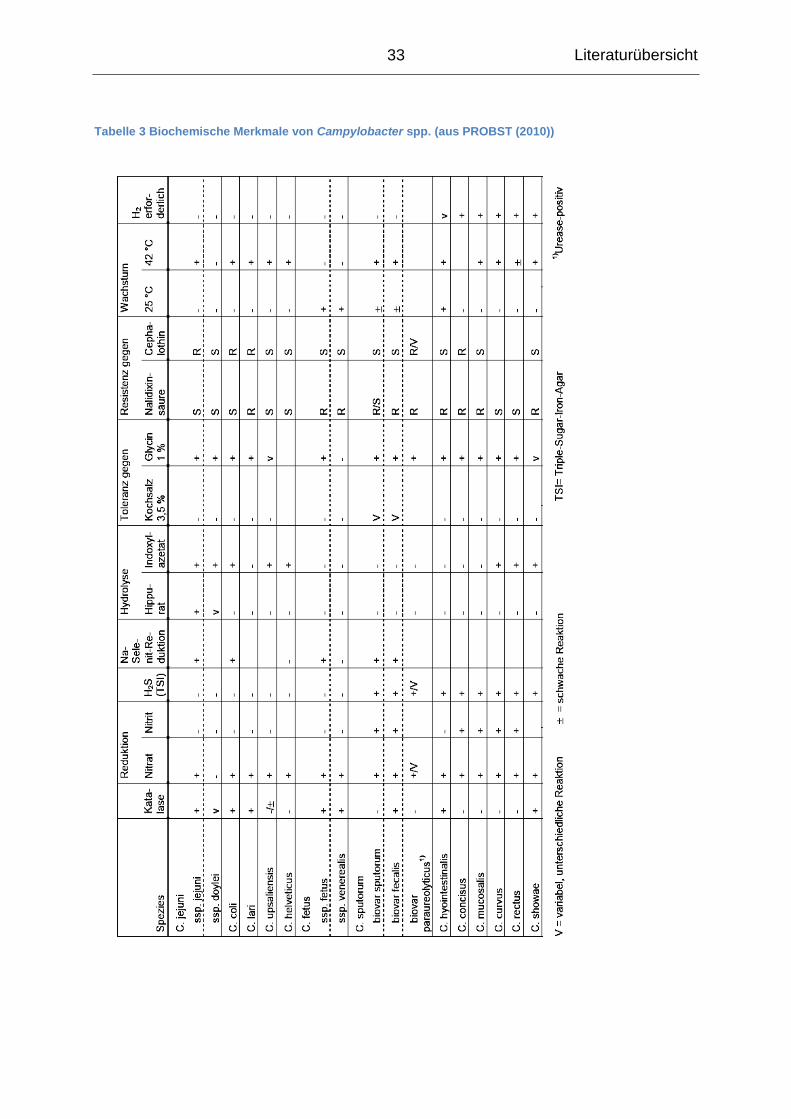
Die klassische phänotypische Speziesdifferenzierung von Campylobacter spp. erfolgt
anhand verschiedener biochemischer Reaktionen, die in Tabelle 3 zusammengefasst
sind (PROBST 2010). Des Weiteren existieren verschiedene Verfahren der Serotypi-

Literaturübersicht
31
sierung, unter anderem die Schemata nach PENNER und LIOR (PENNER u.
HENNESSY 1980; LIOR et al. 1982). Auch Antibiotikaresistenzprofile wurden zur
Speziesdifferenzierung eingesetzt (OWEN et al. 1997). Eine Ergänzung zu den ge-
nannten Methoden kann die Phagentypisierung sein (FROST et al. 1999).
Die sehr anspruchsvolle und zeitaufwändige Kultivierung von Campylobacter spp.,
die je nach Medium ungewollte Selektion bestimmter Spezies (MOORE et al. 2005),
ihre biochemische Inaktivität (ON 1996) und die Variabilität und enorme Diversität
des Bakteriums (HUMPHREY u. JORGENSEN 2006; NEWELL et al. 2010) führte zur
Entwicklung alternativer Methoden:
Zu den antikörperbasierten Nachweismethoden zählen die Latex-Agglutination
(NACHAMKIN u. BARBAGALLO 1990) und ein kommerzieller Antigen-ELISA
(TOLCIN et al. 2000) zur direkten Untersuchung nativer Kotproben.
In den letzten Jahren kommt genombasierten Methoden eine wachsende Bedeutung
zu (HUMPHREY u. JORGENSEN 2006). Die erste PCR (polymerase chain reaction)
zum spezifischen Nachweis von C. jejuni und C. coli in humanen Kotproben wurde
1992 beschrieben (OYOFO et al. 1992). Seitdem wurden unterschiedlichste PCR-
Assays für Speziesnachweis und -differenzierung etabliert (MOORE et al. 2005).
Laut amtlicher Methodensammlung zur Probenahme und Untersuchung von Pro-
benmaterial tierischen Ursprungs für anzeigepflichtige Tierseuchen des Friedrich-
Loeffler-Instituts hat sich eine Multiplex-PCR (DENIS et al. 2001) zur Identifizierung
von C jejuni und C. coli aus Kulturmaterial, Milch- und Kotproben bewährt (PROBST
2010).
Genotypische Differenzierungsmethoden sind das Flagellin Typing, die PFGE, die
RAPD Analyse, das Ribotyping, die AFLP Analyse und die MLST (WASSENAAR u.
NEWELL 2000; ANONYMOUS 2005a).

Literaturübersicht 32
Der indirekte Erregernachweis, also der Nachweis von Antikörpern gegenüber
Campylobacter spp. beim Schwein kann mittels ELISA oder Immunoblot-Methode
(VON ALTROCK et al. 2006) erfolgen. Für den Nachweis aus humanen Stuhlproben
existieren kommerzielle ELISAs (recomWell Campylobacter®, Mikrogen, Neuried,
Deutschland), ebenso für den Nachweis aus Blutproben (Serion ELISA classic
Campylobacter jejuni®, Virion\Serion, Würzburg, Deutschland).
Literaturübersicht
33
Tabelle 3 Biochemische Merkmale von Campylobacter spp. (aus PROBST (2010))

Literaturübersicht 34
2.2.3 Campylobacteriosen beim Menschen
Während viele Campylobacter spp. bei zahlreichen Tierarten ohne krankmachende
Wirkung vorkommen, können sie beim Menschen schwere Erkrankungen auslösen.
Für humane Campylobacteriosen besteht in den meisten EU-Mitgliedsstaaten - in
Deutschland durch das Infektionsschutz-Gesetz (ANONYMOUS 2001) geregelt - ei-
ne Meldepflicht. Infektionen mit Campylobacter spp. standen laut Community-Report
der EFSA mit 190.566 bestätigten humanen Fällen im Jahr 2008 europaweit an der
Spitze der gemeldeten Zoonosen. Insgesamt zeichnete in sich den letzten Jahren in
Europa kein klarer Trend nach oben oder unten ab. In Deutschland wurden 2008
64.731 Fälle gemeldet (ANONYMOUS 2010a).
Die europaweit häufigste beim Menschen nachgewiesene Campylobacter-Spezies
war 2008 C. jejuni (39,5 % gegenüber 44,3 % in 2007), gefolgt von C. coli (2,3 %
gegenüber 2,7 % 2007). Andere Spezies machten zusammen 9,4 % aus, der Anteil
nicht genauer spezifizierter Fälle bzw. solchen mit unbekannter Spezies betrug 49 %
(ANONYMOUS 2010). Es wird vermutet, dass die Inzidenz der Erkrankung unter-
schätzt wird und auf einen gemeldeten Fall etwa neun nicht gemeldete Fälle kom-
men (WHEELER et al. 1999).
Die Krankheit äußert sich nach einer Inkubationszeit von durchschnittlich vier Tagen
durch profuse, häufig blutige Diarrhö, heftige abdominale Schmerzen und Fieber.
Erbrechen tritt nur selten auf. Häufig verläuft die Infektion komplikationslos mit voll-
ständiger Genesung nach etwa einer Woche, in solchen Fällen ist eine antibiotische
Behandlung nicht zwingend notwendig (HUMPHREY et al. 2007). Empfohlene Anti-
biotika sind Erythromycin, Amoxicillin oder Fluorchinolone. Allerdings werden in den
letzten Jahren aufgekommene Resistenzen gegen Makrolide und Fluorchinolone in
vielen Ländern als wachsendes Problem für die öffentliche Gesundheit angesehen
(AARESTRUP u. ENGBERG 2001; ENGBERG et al. 2001). Bei etwa 14 % der Er-
krankten in Deutschland war eine stationäre Behandlung im Krankenhaus erforder-
lich (ROSNER et al. 2010).

Literaturübersicht
35
Eine Hyperexposition kann langfristig eine Immunität verursachen, andererseits er-
höht eine Immunsuppression das Risiko einer schweren Erkrankung. Systemische
Infektionen treten sehr selten auf (0,1 %), meist in Zusammenhang mit C. fetus
(MOORE et al. 2005). Komplikationen einer akuten Infektion umfassen intestinale
Hämorrhagien, mesenteriale Lymphadenitiden, ein toxisches Megakolon und das
hämolytisch-urämische Syndrom (SKIRROW 1977).
Daneben treten zu einem geringen Anteil chronische Verläufe mit Spätfolgen auf.
Erkrankte können eine Urtikaria, eine reaktive Arthritis, ein Erythema nodosum oder
ein Reizdarmsyndrom entwickeln. Als wichtigste Spätfolge einer Campylobacter-
Infektion wird jedoch das Guillain-Barré-Syndrom angesehen. Dabei handelt es sich
um eine akute Demyelinisierung peripherer Neurone, die durch eine aufsteigende
Paralyse gekennzeichnet ist (MOORE et al. 2005). Es entwickelt sich in etwa einem
von tausend Fällen, meist sind bestimmte Serotypen von C. jejuni involviert. Umge-
kehrt wird bei etwa 30 – 40 % der an diesem Symptom Erkrankten eine vorangegan-
gene Campylobacter-Infektion belegt (NACHAMKIN 2002). Die Mortalitätsrate be-
trägt 2 – 3 %, in etwa 20 % der Fälle entwickeln sich gravierende neurologische
Dauerschäden, die übrigen Patienten genesen partiell oder vollständig (MOORE et al.
2005).
Campylobacteriosen können selten Ausbrüchen zugeordnet werden und treten
hauptsächlich als sporadische Erkrankungen auf. Ein prominentes Merkmal ist ihr
saisonales, in den gemäßigten Breiten im späten Frühjahr und Sommer gehäuftes
Vorkommen. Es wird vermutet, dass ein Zusammenhang mit einer vermehrten Aus-
scheidung von Campylobacter spp. bei verschiedenen Tierarten wie Wirtschaftsge-
flügel, wilden Vögeln, Kühen und Hunden sowie dem Auftreten von Insekten in dieser
Jahreszeit besteht. (HUMPHREY et al. 2007).
Die Hauptquelle für Campylobacter spp. – auch durch Kreuzkontamination anderer
Lebensmittel – ist Geflügelfleisch. Dies wurde neben zahlreichen retrospektiven Fall-
Kontroll-Studien (NEIMANN et al. 2003; FRIEDMAN et al. 2004; ADAK et al. 2005)

Literaturübersicht 36
auch durch zwei Ereignisse der letzten Jahre dargelegt: Als in Belgien 1999 aufgrund
eines Dioxin-Skandals Geflügelfleisch und Eier vom Markt genommen wurden, redu-
zierte sich die Zahl der Campylobacter-Erkrankungen um 40 % (VELLINGA u. VAN
LOOCK 2002). In Island erhöhte sich, nachdem ab 1996 neben gefrorenem auch
gekühltes Geflügelfleisch angeboten wurde, die Zahl der Fälle erheblich (STERN et
al. 2003). Frisches Hähnchenfleisch repräsentierte laut EFSA im Jahr 2008 mit EU-
weit 30,1 % positiv getesteten Proben die wichtigste Quelle für Campylobacter spp.,
gefolgt von Fleisch anderer Geflügelspezies (21,9 %) und frischem Putenfleisch
(10,1 %). In frischem Schweinefleisch wurde in 0,5 % der getesteten Proben der Er-
reger nachgewiesen (ANONYMOUS 2010). HUMPHREY und JORGENSEN (2006)
errechneten anhand von Studien aus 25 Ländern einen Mittelwert von 2 % positiven
Proben von rohem Schweinefleisch auf Einzelhandelsebene, während im Mittel 57 %
des beprobten Geflügelfleisches erregerpositiv war. Diese geringere Kontaminations-
rate von „rotem“ Fleisch gegenüber Geflügelfleisch trotz der hohen Prävalenzen bei
Schlachtschweinen wird auf den langsameren Schlachtvorgang mit geringerem Ver-
spritzen von Fäzes und die ausgedehntere Kühlung und Abtrocknung der Karkassen
zurückgeführt. Trotzdem ist „rotes“ Fleisch eine potentielle Gefahrenquelle für die
Ansteckung mit Campylobacter spp., wie Fall-Kontroll-Studien aus Dänemark und
den USA belegen: Dies trifft besonders auf Konsum in der Gastronomie (FRIEDMAN
et al. 2004) und den Verzehr von rotem Fleisch im nicht durchgegarten oder gegrill-
ten Zustand zu (NEIMANN et al. 2003). Letztere geben außerdem den häufigen Ver-
zehr von Schweinefleisch als Risikofaktor an. Weitere, in den zitierten Studien ange-
gebene Risikofaktoren umfassen den Konsum von unpasteurisierter Milch, kontami-
nierten Krustentieren, Meeresfrüchten, rohen Austern und unbehandeltem Wasser.
Die Tatsache, dass Campylobacter spp. sich im Lebensmittel nicht vermehren bzw.
anreichern können, wird durch die sehr geringe minimale Infektionsdosis beim Men-
schen von wenigen hundert Keimen aufgewogen (HUMPHREY und JORGENSEN
2006).

Literaturübersicht
37
Die Campylobacteriose wird auch als „Reisediarrhö“ bezeichnet, da ihr in einer Viel-
zahl der Fälle ein Aufenthalt im Ausland voran geht (HORROCKS et al. 2009).
Als weitere Infektionsquellen wurden die Ansteckung von Mensch zu Mensch (rele-
vant bei kleinen Kindern), der Kontakt mit Haustieren (vor allem mit an Diarrhö er-
krankten jungen Hunden) und mit landwirtschaftlichen Nutztieren identifiziert (BUTZ-
LER 2004). HUMPHREY und JORGENSEN (2006) schätzten sogar die Gefahr einer
direkten Ansteckung bei landwirtschaftlichen Großtieren höher ein als bei Geflügel.
Einen wirksamen Schutz vor Infektionen auf Verbraucherebene stellt FRIEDMAN et
al. (2004) zufolge eine äußerst penible Küchenhygiene – besonders im Umgang mit
rohem Geflügelfleisch – dar.
Auch wenn schon seit einigen Jahren an der Entwicklung von Impfstoffen geforscht
wird (LEE et al. 1999), ist nach Angaben der WHO (World Health Organization) bis
heute noch keiner zugelassen (ANONYMOUS 2009c).
2.2.4 Campylobacter spp. beim Schwein
Campylobacter spp. kommen bei einer Vielzahl von Tierarten vor, darunter Geflügel,
Rinder, Schafe, Heimtiere und Schweine. Bei Rindern und Schafen kann eine Infek-
tion mit klinischen Erkrankungen wie Aborten oder Diarrhö einhergehen; auch bei
Hunden und Katzen können sie Erkrankungen des Verdauungstraktes auslösen
(BUTZLER 2004; ACKE et al. 2010). Beim Geflügel und Schwein dagegen tritt der
Erreger hauptsächlich symptomlos als Kommensale des Magen-Darm-Traktes auf
(MOORE 2005), obwohl auch ein Zusammenhang mit Dysenterie-Erkrankungen
beim Schwein beschrieben wurde (DOYLE 1944). HUMPHREY et al. (2007) mutma-
ßen jedoch, dass Campylobacter spp. keine natürlichen Kommensalen des Magen-
Darm-Trakts wie Escherichia coli sein könnten und dass ihr häufiges Vorkommen bei

Literaturübersicht 38
Rind, Schaf und Schwein eher durch wiederkehrenden Infektionsdruck von außen
und ein Zirkulieren in der Herde bedingt sein könnte.
Die im Hinblick auf die Lebensmittelsicherheit wichtigsten Campylobacter-Spezies
sind C. coli und C. jejuni (HORROCKS et al. 2009). Beide treten beim Schwein auf:
Die EFSA gab in ihrem Report zu Zoonosen in der Europäischen Union 2008 einen
Anteil von 88,8 % C. coli und 2,6 % C. jejuni bei 9,7 % nicht spezifizierten Isolaten an
allen vom Schwein stammenden Campylobacter spp. (aus Kot-, Organ- und Le-
bensmittelproben) an, während beim Geflügel und Rind C. jejuni dominierten (ANO-
NYMOUS 2010). In demselben Bericht wird für Deutschland im Jahr 2008 beim
Schwein eine Herdenprävalenz von 37,3 % in 209 Herden angegeben, im Vorjahr
betrug sie bei 224 Herden 29,5 % (verschiedene, nicht genauer spezifizierte Proben-
herkünfte).
Andere Studien kamen zu teils höheren Ergebnissen: In einer Studie aus Deutsch-
land wurden auf der Basis von Kotproben eine Einzeltierprävalenz für Campylobac-
ter spp. bei 330 untersuchten Endmastschweinen von 79,1 % und eine Herdenprä-
valenz (15 Herden) von 100 % festgestellt (ALTER et al. 2005). VON ALTROCK et al.
(2006) gaben in ihrer Studie an 900 Mastschweinen aus 30 Herden eine Einzeltier-
prävalenz von 69,7 % und eine Herdenprävalenz von 100 % an, WEHEBRINK et al.
(2008) eine Einzeltierprävalenz in der Endmast von 64,7 % (von 354 Tieren). Die se-
rologische Prävalenz betrug bei VON ALTROCK 81,2 % beim Einzeltier und 100 %
auf Herdenlevel.
Bei ALTER et al. (2005) betrug der Anteil von C. coli an allen Campylobacter-Isolaten
aus 1.513 positiven Kotproben 100 %. In anderen Studien (FOSSE et al. 2009) wur-
den aus Kotproben von 215 untersuchten Tieren, davon 100 % Campylobacter spp.
positiv, 95,3 % C. coli und 4,7 % C. jejuni isoliert bzw. bei 800 untersuchten Tieren,
alle Campylobacter spp. positiv, 99,2 % C. coli und 0,2 % C. jejuni. Bei YOUNG et al.
(2000) dagegen betrug der Anteil von C. jejuni an allen Isolaten je nach Tiergruppe
bis zu 89 % (von neun tragenden Sauen, alle Campylobacter spp. positiv). In einem

Literaturübersicht
39
Infektionsversuch einer französischen Arbeitsgruppe wurde beim Schwein jedoch ein
deutlich höheres Kolonisationspotential von C. coli gegenüber C. jejuni belegt
(LEBLANC MARIDOR et al. 2008). Bei FREDRIKSSON-AHOMAA et al. (2009) war
die häufigste Spezies in am Schlachthof entnommenen Kotproben von 50 Tieren
(62 % Campylobacter spp. positiv) C. coli, isoliert an 73 % der positiven Tiere. C. lari,
C. fetus und C. jejuni traten bei 3 %, 9 % und 18 % der positiven Tiere auf.
2.2.4.1 Infektionswege
Das Schwein ist in der Regel symptomloser Träger des Erregers, der in Ingesta und
Geweben des Verdauungstrakts vorkommt und mit dem Kot ausgeschieden wird
(FOSSE et al. 2009).
Die Ansteckung erfolgt häufig schon in der ersten Lebenswoche der Ferkel oro-fäkal
durch den Kot der Sauen, nachdem die Geburt eine erhöhte Ausscheidung bei die-
sen zur Folge hat (WEIJTENS et al. 1997; YOUNG et al. 2000). In diesen Studien
waren die Anteile Campylobacter-ausscheidender Sauen mit 100 % (von 10 Sauen)
bzw. 76 % (von 50 Sauen) hoch, und auch bei mehr als der Hälfte der Ferkel war der
Erreger bereits am ersten Lebenstag nachweisbar. In einer Studie von WEHEBRINK
et. al. (2008), in der nur 33,8 % der 68 untersuchten Sauen den Erreger ausschieden,
wurde dagegen kein Zusammenhang zwischen der Infektion einer Sau und ihrer Fer-
kel gefunden.
In weiteren Studien wird die Ansteckung zu späteren Zeitpunkten, durch andere
Schweine oder Kontamination der Tierumgebung belegt und die Beteiligung von Vek-
toren wie Stiefel, Bekleidung, Schadnager, Insekten oder Vögel vermutet
(WEIJTENS et al. 2000; HORROCKS et al. 2009). Allerdings differierten die Nach-
weisraten aus Umgebungsproben in verschiedenen Studien mit 33,3 % bei von
ALTROCK et al. (2006), 0,7 % bei ALTER et al. (2005) und 0 % bei WEHEBRINK et
al. (2008), und eine Rolle von Wasser und Futter im Infektionsgeschehen konnte
nicht belegt werden.

Literaturübersicht 40
Die Ausscheidung des Bakteriums mit dem Kot erfolgt stark intermittierend und mit
einem Abfall mit zunehmendem Alter der Mastschweine. Bei WEIJTENS et al. (1999)
wurden die höchsten Anzahlen ausgeschiedener Campylobacter spp. am Mastan-
fang und die niedrigsten am Mastende gezählt. Bei WEHEBRINK et al. (2008) wurde
vor dem Absetzen bei 80,9 % (n = 207) der Ferkel, bei 89,2 % (n = 323) der Tiere in
der Anfangs- und nur bei 64,7 % (n = 229) in der Endmast Campylobacter spp.
nachgewiesen. Dies wird einer mit dem Alter der Tiere wachsenden Stabilität der
Magen-Darm-Flora zugeschrieben (WEHEBRINK et al. 2008). Bei ALTER et al.
(2005) stieg dagegen der Anteil positiv getesteter Tiere von 32,8 % (n = 192) nach
dem Absetzen bis auf 66,8 % (n = 394) kurz vor der Schlachtung.
Zum Zeitpunkt der Schlachtung wurde der Erreger in verschiedenen Studien bei zwi-
schen 96 % (24 untersuchte Tiere) und 49 % der Schweine (595 untersuchte Tiere)
aus dem Inhalt verschiedener Darmabschnitte isoliert (NESBAKKEN et al. 2003;
STEINHAUSEROVA et al. 2005). Daneben war das Bakterium auch in verschiede-
nen Geweben des Magen-Darm-Traktes zu finden: Es wurde bei 30 % der Tiere in
den mesenterialen Lymphknoten und bei 46% in den Tonsillen (NESBAKKEN et al.
2003) bzw. bei 50 % von 240 untersuchten Schweinen in der Magenschleimhaut
nachgewiesen (PAYOT et al. 2004). Demgegenüber steht eine wesentlich geringere
Nachweisrate aus den Tonsillen von 6 % bei 50 untersuchten Tieren bei FRED-
RIKSSON-AHOMAA et al. (2009).
Die Kontamination des Fleisches mit Campylobacter spp. erfolgt im Zuge des
Schlachtprozesses, wo durch unsauberes Arbeiten Fäzes bzw. Darminhalt auf die
Karkasse gelangen können (HUMPHREY et al. 2007). Auch das Heraustrennen von
Zunge und Tonsillen zusammen mit dem Geschlinge bietet die Möglichkeit der Ver-
unreinigung der Karkasse mit auf den Tonsillen befindlichen Erregern
(FREDRIKSSON-AHOMAA et al. 2009). In der Studie von NESBAKKEN et al. (2003)
waren 36,5 % der 96 Tupferproben von 24 Karkassen positiv für Campylobacter spp.

Literaturübersicht
41
2.2.4.2 Risikofaktoren und Bekämpfungsmöglichkeiten im Bestand
Grundsätzlich gehen WEIJTENS et al. (1999 und 2000) und Young et al. (2000) da-
von aus, dass die Schaffung Campylobacter-freier Schweinebestände realisierbar ist.
Nach YOUNG et al. (2000) findet eine Bekämpfung aufgrund der sehr frühen Anste-
ckung der Tiere am effektivsten in Zuchtbeständen statt. WEIJTENS et al. (2000)
demonstrierten dies erfolgreich in einem Versuch mit spezifisch pathogen-freien
Sauen.
Des Weiteren konnten bisher jedoch nur sehr wenige Studien Risikofaktoren für Aus-
scheidung und Übertragung von Campylobacter spp. in Schweinebeständen charak-
terisieren (FOSSE et al. 2009). In einer aktuellen Studie wurde eine große Herden-
größe als Risikofaktor identifiziert (FOSSE et al. 2011). Bei WEHEBRINK et al.
(2007) dagegen war die Prävalenz in Herden mit über 1000 Mastplätzen signifikant
geringer als in kleineren Herden. Die gleiche Studie kam zu dem Ergebnis, dass die
Unterbringung der Tiere in separaten Ställen und der präventive Antibiotikaeinsatz zu
Beginn der Mast mit einer geringeren Prävalenz von Campylobacter spp., der Ein-
satz von Anthelmintika und Zukauffutter dagegen mit einer höheren Prävalenz ein-
hergingen. In einer neueren Studie von FOSSE et al. (2011) war dagegen bei Sau-
enherden die Anzahl von Campylobacter spp. im Kot positiv mit der Anzahl der Sau-
en assoziiert, ebenso wie mit der Intervalllänge zwischen Reinigungsmaßnahmen
des Fütterungssystems. Hier hatte auch die Mastgruppe einen signifikanten Effekt
auf die Anzahl der Erreger im Kot.
Der Effekt von Antibiotika auf die Prävalenz des Erregers ist nicht eindeutig determi-
niert, in einer Studie wurden bei Saugferkeln in Betrieben ohne Einsatz von Antibioti-
ka höhere Campylobacter-Prävalenzen festgestellt als bei Betrieben mit Antibiotika-
einsatz. Bei Endmastschweinen war dagegen kein Unterschied erkennbar. (THAKUR
u. GEBREYES 2005). Auch der Transport hatte ALTER et al. (2005) zufolge keinen
signifikanten Einfluss auf die Ausscheidung des Pathogens.

Literaturübersicht 42
2.3 Yersinia enterocolitica
2.3.1 Erregereigenschaften
Die Gattung Yersinia (Familie Enterobacteriaceae, Ordnung Enterobacterales, Klas-
se Gammaproteobacteria, Stamm Proteobacteria) wurde 1944 von VAN LOGHEM
als solche definiert. Namensgeber war der Entdecker des Pesterregers Yersinia (Y.)
pestis im Jahre 1894: A. J. E. Yersin (VAN LOGHEM 1944). Die Gattung umfasst
heute etwa fünfzehn Spezies. Neben Y. pestis sind vor allem der Erreger der Pseu-
dotuberkulose Y. pseudotuberculosis, der Erreger der enteritischen Rotmaulkrankheit
der Forellen Y. ruckeri und Y. enterocolitica, Erreger humaner Darminfektionen, von
Bedeutung. Y. enterocolitica ist in zwei Subspezies unterteilt: Y. enterocolitica
enterocolitica umfasst amerikanische Isolate, Y. enterocolitica palearctica die europä-
ischen Stämme. Andere Spezies wie Y. frederiksenii, Y. kristensii oder Y. intermedia
gelten als apathogen (EUZÉBY 2010).
Yersinia spp. sind gram-negative, nicht sporenbildende, pleomorphe bis kokkoide
Stäbchen mit einer Größe von 0,5 – 0,8 μm mal 1 – 3 μm. Sie sind mit Ausnahme
von Y. pestis bei Vermehrung / Wachstum in einer Umgebungstemperatur von unter
30 C peritrich begeißelt und damit beweglich, bei 37 C dagegen nicht. Sie zeigen
fakultativ anaerobes Wachstum und die Fähigkeit zum oxidativen als auch fermenta-
tiven Metabolismus. Sie sind im Gegensatz zu anderen Enterobacteriaceae
psychrotroph, ihr Wachstumsoptimum beträgt 28 – 30 C (BERCOVIER u.
MOLLARET 1984). Sie sind katalase-positiv, oxidase-negativ und reduzieren Nitrat.
Der Guanin-Cytosin-Gehalt ihrer DNA beträgt 46 mol % bis 48,5 mol % (NEUBAUER
et al. 2001a).
Aufgrund verschiedener biochemischer Eigenschaften werden Y.-enterocolitica-
Isolate in sechs verschiedene Biogruppen, 1A, 1B und 2 – 5, eingeteilt (WAUTERS
et al. 1987). Diese sind mit der Zuordnung zu Serogruppen, mit der ökologischen und
geografischen Verteilung und mit der Humanpathogenität assoziiert: Isolate der Bio-
gruppe 1A kommen hauptsächlich in der Umwelt vor und gelten als apathogen, die

Literaturübersicht
43
Biogruppen 2 –5 umfassen europäische pathogene Isolate, und die Biogruppe 1B
steht für amerikanische pathogene Isolate (BOTTONE 1999).
2.3.2 Nachweismethoden
2.3.2.1 Kultureller Nachweis
Y. enterocolitica wächst, wenn auch langsam, auf den meisten Medien für Darmbak-
terien wie MacConkey-Agar, Hektoen-Enteric-Agar oder Xylose-Lysin-Deoxycholat-
Agar. Nach 24 Stunden bei 37 C zeigen sich stecknadelkopfgroße Kolonien, die
zwischen den größeren Kolonien schneller wachsender Spezies leicht übersehen
werden können (BOTTONE 1999). Daher wurden verschiedene Selektivmedien kon-
zipiert: Der von SCHIEMANN entwickelte Cefsulidon-Irgasan-Neutralrot-Agar (CIN-
Agar) erwies sich als sehr effektiv (SCHIEMANN 1979). Nach 24 – 48 Stunden bei
28 C oder 18 – 24 Stunden bei 32 C zeigen sich Y. enterocolitica als ca. 1 mm gro-
ße glatte Kolonien mit rotem Zentrum. Aufgrund von Ähnlichkeiten in der Kolo-
niemorphologie mit Y. intermedia, Enterobacter, Aeromonas, Proteus oder Citrobac-
ter spp. wurden dem CIN-Medium Josamycin, Oleandomycin und Äskulin hinzuge-
fügt (FUKUSHIMA 1987). Dieses Virulent-Y.-enterocolitica-Agarmedium (VYE) diffe-
renziert nach Inkubation für 24 – 48 Stunden bei 32 C pathogene Stämme (rote Ko-
lonien) von apathogenen (schwarze Kolonien). Ein weiteres gebräuchliches Selek-
tivmedium ist der Salmonella-Shigella-Desoxycholat-Citrat-Agar (NOWAK et al.
2006).
Der Nachweis aus Tonsillen von Schlachtschweinen (VAN DAMME et al. 2010) oder
Proben von klinisch Erkrankten gelingt damit gut, allerdings ist die direkte Isolierung
im Falle von Proben aus Umwelt, Lebensmitteln oder aus Kot asymptomatischer
Träger problematisch: Geringe Keimzahlen, eine dominante Begleitflora und die me-
diumabhängige Hemmung bestimmter pathogener Bioserotypen führen zu niedrigen
Nachweisraten. Daher sollte bei diesen Probenarten der Bebrütung von festen

Literaturübersicht 44
Nährmedien eine Anreicherung in Flüssigmedien vorausgehen (FREDRIKSSON-
AHOMAA u. KORKEALA 2003).
Diese kann unter anderem in Carbencillin-supplementierter Rappaport-Bouillon (22 –
29 C über 48 Stunden) oder Irgasan-Ticarcillin-Kaliumchlorat (ITC)-Bouillon (24 C
über 48 Stunden) erfolgen. Bei Fleisch, Lebensmittelproben, unter Umständen auch
Fäzes, kann eine Behandlung mit 0,25 % Kalilauge die Begleitflora reduzieren. Eine
Kälteanreicherung in PBS für eine bis drei Wochen bei 4 C fördert zwar den ver-
mehrten Wuchs apathogener Stämme, wird aber dennoch für die Diagnostik an Fä-
zes bzw. Stuhlproben empfohlen. Aus dem Anreicherungsmedium erfolgt anschlie-
ßend zur vorläufigen Identifizierung eine Inkubation auf Kligler-Agar für 24 –
48 Stunden bei 30 C (NEUBAUER et al. 2001a; FREDRIKSSON-AHOMAA und
KORKEALA 2003). Andere in der Literatur beschriebene Anreicherungsmedien sind
das Pepton-Sorbit-Bile (PSB)-Medium oder das Ossmer´s Yersinia-Selektiv-Medium
und zur Identifizierung der Urease-Agar (NOWAK et al. 2006). Ein Standardverfahren
zum Nachweis von präsumtiven Y. enterocolitica aus Lebensmittel- und Umweltpro-
ben stellt die Norm ISO 10273:2003 (ANONYMOUS 2003) dar. Für die Diagnostik an
Schweinekotproben existiert jedoch bis heute kein optimales Nachweisverfahren
(LAUKKANEN et al. 2010). Bis zur Entwicklung sensitiverer und selektiverer Metho-
den sollte neben einer Kälteanreicherung immer mehr als ein Isolierungsverfahren
parallel durchgeführt werden.
Zur endgültigen Identifizierung mutmaßlicher Y.-enterocolitica-Isolate steht ein kom-
merzielles Testsystem, das API 20 E® (BioMérieux Deutschland, Nürtingen) zur Ver-
fügung (ARCHER et al. 1987).
2.3.2.2 Weiterführende Differenzierungs- und Nachweismethoden
Die Klassifizierung in sechs Biotypen erfolgt nach einem in Tabelle 4 dargestellten, ver-
schiedene Reaktionen umfassenden Schema nach WAUTERS et al. (1987).
Literaturübersicht
45
Tabelle 4 Biochemische Tests zur Abgrenzung der verschiedenen Biogruppen von Y. enterocolitica (aus: BOTTONE 1999)
Y.-enterocolitica-Stämme lassen sich anhand ihrer somatischen O-Antigene mittels
einer Objektträger-Agglutinationsreaktion serotypisieren (Nomenklatur: O:„Nummer
der Serogruppe„). Dies ist besonders wichtig, da im Gegensatz zu Campylobacter
spp. oder Salmonella spp., wo schon der Speziesnachweis Signifikanz aufweist, der
Nachweis von Y. enterocolitica bei erkrankten Patienten nur im Falle von bestimmten
Bio- und Serovaren Relevanz besitzt (BOTTONE 1997). NEUBAUER et al. (2001a)
zählten 28 verschiedene Serogruppen, von denen nur wenige als humanpathogen
eingestuft sind. Die wichtigsten humanpathogenen Bioserovare sind 1B/O:8,
2/O:5,27, 2/O:9, 3/O:3 und 4/O:3 (FREDRIKSSON-AHOMAA und KORKEALA 2003).
Die Pathogenität eines Y.-enterocolitica-Isolates kann an bestimmten phänotypi-
schen Eigenschaften bestimmt werden, die teilweise bereits bei der Biotypisierung
überprüft werden: Zu den Plasmid-kodierten Virulenzfaktoren gehören unter anderem
ein calcium-abhängiges Wachstum, die Autoagglutination, der Nachweis des V-
Antigens, die Kongorot-Aufnahme und die Kristallviolett-Bindung. Chromosomal ko-

Literaturübersicht 46
diert sind dagegen eine negative Salizin-, Äskulin- und Pyrazinamidasereaktion
(NEUBAUER et al. 2001a). Da diese Tests sehr zeitaufwändig und nicht immer zu-
verlässig sind, wurden verschiedene schnelle und spezifische DNA-basierte Metho-
den zum Nachweis der chromosmalen und plasmoidalen Virulenzgene entwickelt.
(FREDRIKSSON-AHOMAA u. KORKEALA 2003). Inzwischen stehen diverse Kolo-
nie-Hybridisierungs-Assays (JAGOW u. HILL 1986; ROBINS-BROWNE et al. 1989;
IBRAHIM et al. 1992) und PCR-Assays (KWAGA et al. 1992; LANTZ et al. 1998;
TREBESIUS et al. 1998; AARTS et al. 2001) zur Verfügung.
Für den indirekten Erregernachweis existieren verschiedene Methoden. Die human-
medizinisch relevante Serumagglutination erweist sich dabei als problematisch, da
verschiedene Kreuzreaktionen von Y. enterocolitica und bestimmten Serogruppen
mit Morganella morganii, Salmonella spp., Brucella abortus und Rickettsia spp. be-
schrieben sind (CORBEL 1979; SCHLESINGER et al. 1979). In der Veterinärmedizin
wurde in der Vergangenheit üblicherweise die Komplementbindungsreaktion (KBR)
eingesetzt, eine spezifischere Methode stellt der Immunoblot dar (NEUBAUER et al.
2001a). Auch ELISA-basierte Verfahren wurden etabliert, für den Nachweis beim
Schwein ist unter anderem ein kommerzieller ELISA (Pigtype® YOP-Screen, Labor-
diagnostik Leipzig GmbH) erhältlich. Allerdings können auch bei all diesen Methoden
Kreuzreaktionen mit Brucella abortus oder Borrelia burgdorferi auftreten
(KITTELBERGER et al. 1995; RAWLINS et al. 2005).
2.3.3 Yersiniosen beim Menschen
Die Yersiniose ist nach der Campylobacteriose und der Salmonellose die dritthäu-
figste bakterielle Zoonose in Deutschland und in der EU. In Deutschland besteht,
ebenso wie in den meisten weiteren EU-Mitgliedsstaaten, eine Meldepflicht (ANO-
NYMOUS 2001). Seit 2004 sind die Fallzahlen in der EU leicht rückläufig, 2008 wur-
den insgesamt 8.346 gemeldete Fälle verzeichnet, davon 4.354 in Deutschland
(ANONYMOUS 2010). Die tatsächliche Anzahl von Yersiniosen wird jedoch zehnmal
höher eingeschätzt als die Zahl der gemeldeten Fälle (SKJERVE et al. 1998).

Literaturübersicht
47
Y. enterocolitica war mit 91,9 % der Fälle die EU-weit am häufigsten isolierte Spezies,
1,8 % wurden Y. pseudotuberculosis zugerechnet, und 6,3 % der Fälle waren nicht
zuzuordnen (ANONYMOUS 2010).
Die Yersiniose manifestiert sich beim Menschen sehr häufig in Form einer akuten
Gastroenteritis. Die Inkubationszeit nach oraler Infektion beträgt einen bis elf, meist
jedoch ungefähr fünf Tage (ROSNER et al. 2010). Sie ist bei Kindern durch Fieber
und wässrige, mitunter blutige Diarrhö und Vomitus gekennzeichnet (BOTTONE
1997). Bei Erwachsenen dominieren Symptome einer terminalen Ileitis und mesente-
rischen Lymphadenitis mit starken Schmerzen im unteren Bauch, die mit einer Ap-
pendizitis verwechselt werden können (BLACK et al. 1978). Bei protrahierten Verläu-
fen kann sich eine nekrotisierende Enterocolitis mit extensiven Ulzerationen entwi-
ckeln. Diese tritt häufig bei Infektionen mit der Serogruppe O:8 auf, während sich O:3
und O:9 im Gastrointestinaltrakt weniger destruktiv auswirken. Die Krankheitssymp-
tome dauern bei Kindern zwischen drei und 28 Tagen, bei Erwachsenen meist eine
bis zwei Wochen an (BOTTONE 1997).
Daneben kann der Erreger schwere Septikämien mit metastatisch-abszedierenden
Infektionen diverser Organe wie Leber, Milz, Lunge, Herz, Gelenken oder Meningen
bedingen (BOTTONE 1999). Neben der oralen Transmissionsroute tritt diese Er-
scheinungsform oft nach Ansteckung über eine kontaminierte Bluttransfusion auf.
Dabei kommt dem Erreger seine Fähigkeit zur Reproduktion bei niedrigen Tempera-
turen und der Verwertung von Eisen aus zerfallenden Erythrozyten zugute. Des Wei-
teren wird dadurch belegt, dass auch subklinische Bakteriämien ohne apparente Er-
krankung des Trägers möglich sind (ROBINS-BROWNE u. PRPIC 1985).
Zu den immunologischen Spätfolgen, die sich aus einer Infektion mit den Serogrup-
pen O:9 und besonders O:3, Biotyp 4, Phagentyp 8 entwickeln können, zählen die
reaktive Arthritis, das Erythema nodosum, ferner das Reiter´s Syndrom, eine Glome-
rulonephritis sowie eine Myokarditis (LAHESMAA-RANTALA et al. 1987).

Literaturübersicht 48
Die Therapie der Yersiniose kann mit einem wirksamen Antibiotikum erfolgen. In der
Literatur wird Y. enterocolitica als empfindlich gegenüber Chloramphenicol, Ciproflo-
xacin, Gentamicin, Tetrazyklin und Trimethoprim beschrieben, während Resistenzen
gegenüber ß-Laktam-Antibiotika, besonders Ampicillin, aber auch Amoxycillin und
Clavulansäure sowie Cefoxitin und Imipenem vorliegen (BOTTONE 1999). In einer
aktuellen Studie erwiesen sich alle Isolate sensibel gegenüber Ciprofloxacin, Nalidi-
xinsäure, Gentamicin, Ceftiofur, Tetrazyklin, Kanamycin, Cefotaxim und Chlor-
amphenicol, jedoch resistent gegenüber Ampicillin und Sulfamethoxazol (VON
ALTROCK et al. 2010). An der Entwicklung von Impfstoffen gegen Yersiniosen wird
zur Zeit geforscht (BRANGER et al. 2009).
Die Ansteckung mit Y. enterocolitica erfolgt üblicherweise über kontaminierte Le-
bensmittel, kann aber auch durch verunreinigtes Wasser, direkten Tierkontakt oder
Bluttransfusionen geschehen. Als Hauptquelle wird kontaminiertes, rohes oder nicht
durcherhitztes Schweinefleisch - direkt oder durch Kreuzkontamination anderer Le-
bensmittel- angesehen (TAUXE et al. 1987; BOTTONE 1997; ROSNER et al. 2010).
In einer Fall-Kontroll-Studie aus Finnland wurde der Konsum dessen als der Risiko-
faktor mit dem größten Einfluss herausgestellt. Weitere Risikofaktoren waren das
Essen in Restaurants oder Kantinen, der Konsum von importieren Früchten und Rei-
sen ins Ausland (HUOVINEN et al. 2010). In einer schwedischen Fall-Kontroll-Studie
an Kleinkindern erhöhten der Konsum von rohen Schweinefleischprodukten sowie
Würsten, die Verwendung von Schnullern und der Kontakt mit Hunden und Katzen
das Risiko einer Yersiniose signifikant (BOQVIST et al. 2009).
Humane Yersiniosen treten in der Regel sporadisch, d. h. als Einzelfälle, auf. In den
Jahren 2001 bis 2008 waren dies in Deutschland nach Auswertungen des Robert-
Koch-Institutes etwa 98 % der Fälle (ROSNER et al. 2010). Im Gegensatz zu
Campylobacteriosen und Salmonellosen mit Häufungen im Sommer war bei der Yer-
siniose keine ausgeprägte Saisonalität erkennbar. Diesen Daten zufolge erkrankten
Jungen und Männer häufiger als Mädchen und Frauen. Kinder bis fünf Jahre waren
zwölfmal häufiger betroffen als die übrigen Altersgruppen, die höchste Inzidenz lag

Literaturübersicht
49
bei den Einjährigen vor. Bei 17 % der Erkrankten erfolgte eine Hospitalisierung für
mindestens einen Tag, 14 Menschen starben durch die Infektion.
Obwohl Yersiniosen meist sporadisch auftreten, wurden in den vergangenen Jahren
gelegentlich Ausbrüche bekannt. Der erste größere Ausbruch wurde 1976 im US-
Bundesstaat New York dokumentiert, wo 222 Schüler und Angestellte an fünf Schu-
len nach dem Konsum kontaminierten Kakaos erkrankten (BLACK et al. 1978).
Ebenfalls in den USA erkrankten 1982 172 Menschen nach dem Konsum von pas-
teurisierter Milch (TACKET et al. 1984). Die Ursache eines Ausbruches mit 20 Be-
troffenen auf einem kroatischen Öltanker 2002 konnte nicht geklärt werden (BABIC-
ERCEG et al. 2003). Im Jahr 2006 erkrankten in Norwegen elf Personen nach dem
Konsum eines Schweinefleisch-Fertigproduktes (GRAHEK-OGDEN et al. 2007). Der
EFSA wurden im Jahr 2008 EU-weit 22 Ausbrüche (entsprechend 0,4 % aller Aus-
brüche) mit insgesamt 101 Erkrankten gemeldet, davon konnten jedoch nur zwei
Ausbrüche verifiziert werden. Derjenige mit den meisten Betroffenen (50) wurde
durch Y. pseudotuberculosis in rohen Karotten verursacht (ANONYMOUS 2010).
Aufgrund der Bedeutung von pathogenen Yersinia spp. für die menschliche Gesund-
heit ist ihr Nachweis in Lebensmitteln in mehreren EU-Mitgliedsstaaten meldepflichtig,
und in einigen europäischen Staaten bestehen Monitoringsysteme zur Untersuchung
von Lebensmitteln aus dem Handel. Schweinefleisch und -produkte waren 2008 in
der EU die Lebensmittel mit der höchsten Nachweisrate für Y. enterocolitica. Der
Community-Report der EFSA (ANONYMOUS 2010) gibt anhand Daten von sieben
Mitgliedsstaaten eine Nachweisrate von 1,8 % in 4.817 Proben an. Bei den 557 Pro-
ben aus Deutschland waren es 1,9 %, bei den 654 britischen Proben 9,2 %.
Auch andere Studien untersuchten die Kontaminationsraten von Lebensmitteln im
Handel: ANDERSEN et al. (1991) isolierten den Erreger kulturell aus zehn von 33
Schweinehackfleischproben und drei von 24 Rinderhackfleischproben. Letztere
stammten alle aus Metzgereien, in denen auch Schweinehackfleisch positiv war,
wodurch die Autoren auf eine Kreuzkontamination, ausgehend von Schweinefleisch,
schlossen. In der Studie von WEHEBRINK et al. (2008) waren zwei von 86 Schwei-

Literaturübersicht 50
nefleischproben bakteriologisch positiv. Die im Vergleich zu Proben auf dem
Schlachthof niedrigen Nachweisraten von Y. enterocolitica in Lebensmitteln erklären
FREDRIKSSON-AHOMAA und KORKEALA (2003) mit Unzulänglichkeiten der kultu-
rellen Nachweisverfahren. Dies wurde in Studien bestätigt, in denen mittels PCR we-
sentlich höhere Nachweisraten von Y. enterocolitica in Schweinefleisch ermittelt wur-
den: Bei einem Autor (BOYAPALLE et al. 2001) waren dies 80 % von 350 untersuch-
ten Kutteln und 38 % von 350 Schweinemettproben. In einer weiteren Studie
(VISHNUBHATLA et al. 2001) wurde der Erreger in 47 % von 100 Schweinemettpro-
ben detektiert.
2.3.4 Y. enterocolitica beim Schwein
Y. enterocolitica ist im Magen-Darm-Trakt einer Vielzahl von Tierarten und in der
Umwelt, im Wesentlichen in den subtropischen und den gemäßigten Klimazonen,
weit verbreitet. In der Regel ist die Infektion bei Tieren nicht mit einer klinischen
Symptomatik verbunden, wenn auch Berichte zu Allgemeinerkrankungen, Enteritiden
oder Aborten bei Chinchillas, Hunden, Katzen, Wiederkäuern, Kaninchen, Geflügel
und Schweinen beschrieben sind (NEUBAUER et al. 2001b). Da bei gesunden
Schweinen humanpathogene Stämme nachgewiesen wurden und keine Unterschie-
de zwischen porzinen und humanen Isolaten detektierbar sind, gilt das Schwein als
Hauptreservoir der humanpathogenen Isolate (ANDERSEN et al. 1991; BOTTONE
1997; FOSSE et al. 2009). Bei dieser Tierart treten außer der Biogruppe 5 alle Biova-
re und alle wichtigen humanpathogenen Serogruppen (O:3, O:5,27, O:8, O:9) auf
(BOTTONE 1999).
Herden- und Einzeltierprävalenz variieren, auch abhängig von Untersuchungsmateri-
al und Testmethode, in verschiedenen Studien: In einer Studie aus Niedersachsen
an 900 Mastschweinen aus 30 Herden waren 43,3 % der Bestände und 8,4 % der
Einzeltiere in Kotproben bakteriologisch positiv, die Seroprävalenz betrug 83,3 % auf

Literaturübersicht
51
Herdenlevel und 66,8 % bei Einzeltieren (VON ALTROCK et al. 2006). Alle Isolate
gehörten hier der Serogruppe O:3 an. In einer weiteren Studie aus Norddeutschland
(WEHEBRINK et al. 2008) wurde bei 13,3 % der 354 untersuchten Endmastschwei-
ne in Analtupfern Y. enterocolitica nachgewiesen, auch hier ausschließlich Sero-
gruppe O:3; 1,1 % der Schweine wiesen Y. pseudotuberculosis auf. In Kanada wurde
in einer Studie bei 80 % von 20 untersuchten Herden der Erreger im Kot bakteriolo-
gisch nachgewiesen, davon zu 93,5 % (n = 143) O:3 und zu je 1,3 % (n = 2) O:5 und
O:9 (PILON et al. 2000). In einer norwegischen Arbeit waren 86 % von 86 untersuch-
ten Mastbeständen serologisch positiv für Serogruppe O:3 (SKJERVE et al. 1998).
2.3.4.1 Infektionswege
Die Ansteckung der Schweine erfolgt vermutlich hauptsächlich oro-fäkal durch direk-
ten Tier-zu-Tier-Kontakt (PILON et al. 2000). Die Wissenschaftler fanden in kanadi-
schen Mastbeständen jeweils nur einen Genotyp, der über mehrere Mastdurchgänge
nachzuweisen war. Umweltproben waren nur sehr selten, zu 0,6 %, positiv. Daher
messen diese Autoren der Tierumgebung als Infektionsquelle keine große Bedeu-
tung bei. Auch in anderen Studien wurde der Erreger gar nicht (WEHEBRINK et al.
2008) oder mit 3,4 % nur selten (VON ALTROCK et al. 2006) aus Proben der
Tierumgebung nachgewiesen, wodurch diese Autoren zu einem ähnlichen Schluss
kommen. SKJERVE et al. (1998) schreiben der Tierumgebung insofern eine Bedeu-
tung zu, als sie in norwegischen Beständen einen Zusammenhang zwischen dem
Vorkommen von Y. enterocolitica und Katzenhaltung fanden. Andere Autoren erach-
teten die Transmission über die Tierumgebung wichtiger als die von der Sau auf die
Ferkel (WINGSTRAND u. NIELSEN 1996). Da Y. enterocolitica in einer Vielzahl von
Schädlingen wie Nagern oder Fliegen vorkommt, müssen auch diese als Vektoren
Erwägung finden (ALDOVA u. LAZNICKOVA 1979).
Der Zeitpunkt der Ansteckung liegt vermutlich in der Mast: WEHEBRINK et al. (2008)
konnten weder bei Sauen noch bei Saugferkeln Y. enterocolitica kulturell nachweisen.
In der Anfangsmast waren 15,2 % von 362 Tieren und in der Endmast 13,3 % von

Literaturübersicht 52
354 Tieren positiv. Auch in einer Arbeit von KASIMIR (2005) konnte der Erreger im
gesamten Bereich der Ferkelerzeugung (186 Kotproben von Sauen und 1.665 Kot-
proben von Ferkeln) nicht nachgewiesen werden, sondern erst in der Mast: zu Mast-
anfang bei 2 % von 609 Tieren und am Mastende bei 18 % von 578 Tieren.
Die Ausscheidung im Kot erfolgt nur über einen bestimmten Zeitraum nach der Infek-
tion. In einem Infektionsversuch begann die Ausscheidung von Y. enterocolitica im
Kot bei allen Tieren der Versuchsgruppe nach fünf bis 21 Tagen post inoculationem
(NIELSEN et al. 1996). Nach sieben Wochen waren noch 10 % der Tiere im Kot po-
sitiv, nach neun Wochen keines mehr, dagegen blieben alle Tiere bis zu diesem
Zeitpunkt seropositiv, und bei 80 % war der Erreger in den Tonsillen nachweisbar. Zu
einem ähnlichen Ergebnis war bereits eine japanische Studie (FUKUSHIMA et al.
1983) gekommen: Dort dauerte die Ausscheidung mit dem Kot etwa sieben Wochen
an, eine erneute Ausscheidung nach Re-Infektion konnte danach nicht induziert wer-
den.
Die Kontamination des Fleisches kann einerseits durch fäkale Kontamination der
Karkasse am Schlachthof bei der Eviszeration geschehen. Eine dänische Studie
konnte dies anhand des Einflusses verschiedener Eviszerationstechniken belegen
(ANDERSEN et al. 1991). Ein weiteres Indiz dafür ist der Rückgang humaner Yer-
siniosen in Norwegen um 30 - 40 % nach Einführung des Verschlusses des Rektums
mit einem Plastikbeutel vor der Eviszeration (NESBAKKEN et al. 1994).
Eine andere wichtige Kontaminationsquelle sind die Tonsillen, die als Hauptreservoir
des Erregers im Tierkörper gelten. Durch das während der Schlachtprozedur durch-
geführte Heraustrennen der Zunge inklusive der Mandeln mit dem Geschlinge ist ei-
ne Kontamination der Karkasse und des Geschlinges mit auf den Tonsillen befindli-
chen Erregern laut FREDRIKSSON-AHOMAA et al. (2009) leicht möglich. In dieser
Studie wurde bei 62 % der 50 in einem deutschen Schlachthof getesteten Schweine
Y. enterocolitica in den Tonsillen und nur bei 16 % im Kot nachgewiesen. Die Nach-
weisrate von 75 % in den Tonsillen von 24 Schlachtschweinen einer Studie in Nor-

Literaturübersicht
53
wegen war noch höher (NESBAKKEN et al. 2003). Die Autoren bezifferten sie auf
das sechsfache der Nachweisrate aus dem Kot der Schweine und sehen in ihr ein
größeres Kontaminationsrisiko als durch Kot.
Eine weitere Möglichkeit der Keimverschleppung stellt die Inzision der Mandibular-
lymphknoten dar, diese waren in der genannten Studie zu Anteilen von 5,2 % bis
12,5 % mit dem Erreger kontaminiert.
Kontaminationsraten von Karkassen und Innereien auf dem Schlachthof wurden in
verschiedenen Studien bestimmt: Bei einer Untersuchung durch VON ALTROCK et
al. (2010) an 1500 Schweinelebern am Schlachthof wurde bei 4,7 % Y. enterocolitica
nachgewiesen. Bei WEHEBRINK et al. (2008) war nur eine von 122 untersuchten
Karkassen vor der Kühlung bakteriologisch positiv, zwölf Stunden danach keine mehr.
Andere Autoren detektierten kulturell aus 131 Proben in 31 % der Lebern, 69 % der
Nieren und 50 % der Herzen Y. enterocolitica (FREDRIKSSON-AHOMAA et al.
2000). In Norwegen wiesen NESBAKKEN et al. (2003) den Erreger an 3,1 % bis
4,2 % von insgesamt 120 Karkassen nach. Derselbe Autor wies in einer späteren
Studie bei 8,3 % von 60 untersuchten Karkassen sowohl vor als auch nach der Luft-
Sprüh-Kühlung Y. enterocolitica nach und leitete daraus eine geringe Effektivität die-
ses Verfahrens auf die Reduktion des kältetoleranten Erregers auf den Karkassen ab
(NESBAKKEN et al. 2008).
2.3.4.2 Risikofaktoren und Bekämpfungsmöglichkeiten im Bestand
Ebenso wie zu Campylobacter spp., wurden auch zu Y. enterocolitica noch recht we-
nige Daten zu Risikofaktoren für eine Infektion in Schweinebeständen publiziert
(FOSSE et al. 2009).
Während ANDERSEN et al. (1991) keinen Managementfaktor als Einflussfaktor auf
das Vorkommen von Yersinia spp. in Schweineherden ermitteln konnten, fanden

Literaturübersicht 54
TAUXE et al. (1987) einen Zusammenhang zwischen extern erworbenen Futtermit-
teln und erhöhtem Vorkommen des Erregers in Schweineherden.
Bei SKJERVE et al. (1998) stellten Personalhygienemaßnahmen, die Verwendung
eines eigenen Transportfahrzeuges, die häufige Anwesenheit von Katzen am Hof
und die Haltung auf Stroh Risikofaktoren dar, Unterdrucklüftung und manuelle Fütte-
rung waren dagegen protektive Faktoren.
Den wichtigsten protektiven Faktor sahen die Autoren jedoch in der Schweinehaltung
im geschlossenen System gegenüber der Haltung in reinen Mastbetrieben. Das
Kernprinzip zur Bekämpfung von Yersinia war für diese und auch andere Autoren
eine Minimierung der Zahl möglicher Tierkontakte und die strikte Trennung infizierter
und nicht-infizierter Tiere respektive Herden. FUKUSHIMA et al. (1983) waren zu
dem gleichen Ergebnis gekommen, da sie ein stark buchtenweises Übertragungs-
muster in Tiergruppen beobachtet hatten. Bei NOWAK et al. (2006) waren signifikant
mehr auf dem Schlachthof beprobte Tiere aus konventionellen Haltungen molekular-
biologisch positiv als aus alternativen Haltungssystemen. Sie führten dies ebenfalls
auf die größeren Anzahlen verschiedener Tierherkünfte in den konventionellen Hal-
tungen zurück. Norwegische Wissenschaftler beobachteten, dass der Status einer
Herde als spezifiziert „frei von Yersinia“ in geschlossenen Systemen ohne externe
Tierkontakte viel leichter zu erhalten war als in offenen Systemen (NESBAKKEN et al.
2007).
Die Keulung einzelner infizierter Tiere stellte SKJERVE et al. (1998) zufolge keinen
erfolgversprechenden Ansatz zur Herdensanierung dar. Er mutmaßt, dass Maßnah-
men zur Reduktion von Salmonella spp. und anderer enterischer Pathogene im
Schweinebestand auch zur Minderung von Y. enterocolitica dienen können, sieht
aber noch weiteren Forschungsbedarf. ANDERSEN et al. (1991) zweifelten zuvor
aufgrund der weiten Verbreitung von Y. enterocolitica unter Schweinen die Effektivi-
tät von Kontrollmaßnahmen auf Herdenlevel an.

Literaturübersicht
55
BOTTONE et al. (1997) schlugen eine versuchsweise Verfütterung von nichtpatho-
genen Y. enterocolitica an Schweine vor, da diese die pathogenen Stämme durch
kompetitive Verdrängung und Bacteriocin-Produktion hemmen könnten.
2.4 Ziele dieser Studie
Für Salmonella spp, Campylobacter spp. und Y. enterocolitica bestehen auch heute
noch Wissenslücken in Bezug auf eine effektive Bekämpfung ihres Vorkommens in
Schweinebeständen. Besonders zur Epidemiologie von Campylobacter spp. und
Y. enterocolitica im Schweinebestand werden, gerade im Hinblick auf die Rolle der
Umgebung (potentielle Eintragsquellen und Reservoire) detailliertere Kenntnisse be-
nötigt. Studien, die die Tierumgebung untersuchten, taten dies nur am Rande, wäh-
rend ihr Fokus auf den Tieren selbst lag (PILON et al. 2000; VON ALTROCK et al.
2006; WEHEBRINK et al. 2008). Daher soll in der vorliegenden Arbeit der Schwer-
punkt auf einer intensiven Untersuchung der Tierumgebung liegen.
In zahlreichen Studien wurden Prävalenzen der Erreger im Bestand untersucht. Dies
geschah jedoch meist für jedes Pathogen einzeln, ohne auf mögliche Zusammen-
hänge zwischen ihnen einzugehen. VON ALTROCK et al. (2006) untersuchten das
gemeinsame Auftreten von Campylobacter spp. und Y. enterocolitica im Bestand,
ohne deutlichen Hinweis auf Zusammenhänge. In einer weiteren Arbeit (FOSSE et al.
2011) wurden Koinfektionen von Campylobacter spp., Salmonella spp., Clostridium
perfringens, Listeria monocytogenes und Staphylococcus aureus auf Bestandsebene
analysiert. Auch darin konnte für die beiden in der vorliegenden Arbeit thematisierten
Pathogene kein Zusammenhang nachgewiesen werden. In der Dissertation von
DÖHNE (DÖHNE 2010) wurde das gemeinsame Auftreten von Salmonella spp.,
Campylobacter spp. und Y. enterocolitica anhand einer Untersuchung von Blut und
Lymphknoten, beides am Schlachthof entnommen, untersucht. Auch diese schloss
ohne eindeutiges Resultat, allerdings mit Hinweisen auf häufigere Yersinia-

Literaturübersicht 56
Nachweise in Kategorie I-Betrieben. Studien über alle drei Pathogene im Bestand,
auch unter besonderer Berücksichtigung der spezifischen Salmonellen-
Kategorisierung, sind bis dato jedoch nicht bekannt. Kenntnisse darüber zu erlangen,
ist ein Ziel der vorliegenden Arbeit. Dies ist deshalb von Relevanz, da darüber Rück-
schlüsse möglich sind, ob von der Salmonellen-Kategorisierung eine Extrapolation
auf die Risikoeinschätzung der anderen Erreger möglich ist. Dann könnte außerdem
auf ähnliche Risikofaktoren und in der Konsequenz gleiche Bekämpfungsmaßnah-
men geschlossen werden.
Risikofaktoren wurden im Falle von Campylobacter spp. und Y. enterocolitica eben-
falls nur in sehr wenigen Studien charakterisiert (FOSSE et al. 2009), daher soll dies
ein weiterer Schwerpunkt dieser Dissertation sein.
Auf dem Gebiet der Salmonellenbekämpfung wurden in den vergangenen Jahren
intensive Forschung betrieben und neue Erkenntnisse gewonnen: Es wurden einer-
seits zahlreiche Studien zum Vorkommen von Salmonellen und Risikofaktoren in
Mastbeständen durchgeführt. Andererseits wurden enorme Fortschritte in der Ent-
wicklung molekularbiologischer bzw. genombasierter Feintypisierungsmethoden für
Salmonellen erzielt. So führten LANGVAD et al. (2006) ein Fingerprinting von Sal-
monellen in Schweineherden inklusive der Tierumgebung durch. Andere Autoren
(MAGISTRALI et al. 2008; VIGO et al. 2009) initiierten Longitudinalstudien in
Schweineherden mit Feintypisierung der Salmonellen, jedoch nur aus Kotproben.
Bisher wurde in der Literatur keine Studie beschrieben, die eine Longitudinaluntersu-
chung in Mastbeständen mit intensiver Probenentnahme im komplexen Umfeld der
Schweine mit einer hochdiskriminierenden Feintypisierung der Salmonella-Isolate
vereint: Dies soll ein ebenfalls ein Kernpunkt dieser Arbeit sein. Die Ergebnisse kön-
nen dazu dienen, die genaue Dynamik von Salmonellen im Bestand noch dezidierter
abzubilden und betriebsübergreifend spezifische Muster zu identifizieren, die ihrer-
seits den Tierärzten und Landwirten gezielte Hinweise für eine effiziente Bekämp-
fungsstrategie liefern können.

Material und Methoden
57
3. Material und Methoden
3.1 Querschnittsuntersuchung zu Salmonella spp., Campylobacter
spp. und Y. enterocolitica
3.1.1 Studiendesign und Ziel der Untersuchung
Die Untersuchung wurde im Rahmen des durch das Bundesministerium für Bildung
und Forschung geförderten Verbundprojektes „Food-borne zoonotic infections“ (FBI-
Zoo) durchgeführt. Sie erfolgte in Kooperation mit der Außenstelle für Epidemiologie
in Bakum (Ausgangspunkt der Probennahmen) und dem Institut für Mikrobiologie
und Tierseuchen der Stiftung Tierärztliche Hochschule Hannover (bakteriologische
Untersuchungen).
Zweck der Untersuchung war, nähere Erkenntnisse zur Verbreitung und den Über-
tragungswegen im Bestand, dem gemeinsames Auftreten und den Risikofaktoren für
das Vorkommen der drei lebensmittelrechtlich relevanten Erreger Salmonella spp,.
Campylobacter spp. und Y. enterocolitica in Schweinemastbeständen zu erlangen.
Eine weitere Fragestellung betraf etwaige Unterschiede zwischen Betrieben der QS-
Salmonellen-Kategorie I und der Kategorie III. Dazu wurde eine Querschnittsunter-
suchung in 50 niedersächsischen Schweinemastbetrieben mit einer einmaligen Pro-
benentnahme bei den Tieren und in ihrer Umgebung sowie eine Auswertung von mit-
tels Fragebogen erhobenen Betriebsdaten durchgeführt.
3.1.2 Auswahl der Bestände
An der Studie teilnehmen konnten Betriebe mit Standort in Niedersachsen. Der Be-
stand konnte entweder als reiner Mastbestand oder im geschlossen System, welches
neben der Mast auch die Ferkelproduktion umfasst, geführt sein. Die weitere Aus-
wahl erfolgte anhand der Salmonellen-Kategorie: Es wurden je 25 Bestände in Kate-

Material und Methoden 58
gorie I und Kategorie III gesucht. Die Akquise geeigneter und zur Teilnahme bereiter
Betriebe erfolgte durch Vermittlung verschiedener in der Region tätiger Beratungs-
ringe und Tierarztpraxen.
3.1.3 Untersuchung der Bestände
3.1.3.1 Zeitrahmen der Beprobung
Die Untersuchungen der Bestände erfolgten von Juli 2008 bis Februar 2010. In je-
dem Bestand wurde eine einmalige Probenentnahme durchgeführt. Da die Erreger-
last in den Beständen in der Art erfasst werden sollte, dass sie möglichst das Risiko
für eine Erregerübertragung zur nächsten Produktionsstufe abbilden sollte, wurden
Schweine kurz vor ihrer Schlachtung und ihre unmittelbare Umgebung für die Pro-
benentnahme herangezogen: Die ersten Schweine der Stichprobe mussten maximal
zwei Wochen nach der Untersuchung zur Schlachtung abgeliefert werden.
3.1.3.2 Probenarten und -lokalisationen
Die Probenentnahme beinhaltete einerseits Sammelkotproben, andererseits Proben
von verschiedenen Lokalisationen in der Tierumgebung innerhalb und außerhalb des
Abteils sowie Schädlinge. Pro Bestand wurden je vier Sammelkotproben und zehn
Umgebungsproben entnommen. Grundsätzlich wurden - sofern zugänglich -
Futtertröge
Nippeltränken
Spielketten
Buchtentrennwände/-außenwände
Abteilgang
Zentralgang
Fliegen/Insekten
Nagerkot
Treibbretter und
Stiefel untersucht.

Material und Methoden
59
Wenn in einem Betrieb nicht alle aufgeführten Lokalisationen zugänglich waren, wur-
den alternativ
Rohrleitungen
Fenster(-bretter)
Lüftung
Heizung
Arbeitsräume
Tierwaage oder
sonstige Arbeitsmittel untersucht.
3.1.3.3 Durchführung der Probenentnahme
Für die vier Sammelkotproben wurden insgesamt stets so viele Buchten (gleichmä-
ßig über das bzw. die Abteile verteilt) in die Probenentnahme einbezogen, dass in
ihnen insgesamt ca. 140 Tieren eingestallt waren. Im Falle einer Großbucht (mit über
100 Tieren) wurden alle vier Kotproben in dieser einen Bucht entnommen. Wenn der
Stall viele kleine Buchten, die z. B. mit je neun Tieren belegt waren, aufwies, floss
jeweils der Kot von vier Buchten in eine Sammelkotprobe ein (neun Tiere * vier Buch-
ten * vier Sammelkotproben = 144 Tiere). Für eine Sammelkotprobe wurde an ca.
zehn verschiedenen Stellen, gleichmäßig über die berechnete Anzahl der Buchten
verteilt (bei vier Buchten an zwei bis drei Stellen je Bucht), ca. 1 g augenscheinlich
frischer Kot vom Buchtenboden in ein Kotprobengefäß gefüllt, so dass jedes Proben-
gefäß ca. 10 g Kot enthielt.
Zur Untersuchung von Insekten wurden ca. 15 Fliegen, Käfer oder Kakerlaken, wo
möglich, an verschiedenen Stellen im Stall eingefangen bzw. eingesammelt und in
ein Kotprobengefäß gefüllt. Auch Nagerkot wurde, falls vorgefunden, an verschieden
Stellen entnommen und davon 2 – 3 g in ein Kotröhrchen gefüllt.
Für die Entnahme von Umgebungsproben wurden Amies- und Cary-Blair-Tupfer
(Beckton Dickinson Deutschland, Heidelberg) verwendet. Diese wurden zur besseren
Aufnahme der Probenmatrix vor der Probenentnahme mit steriler phosphatgepuffer-
ter Kochsalzlösung benetzt, die sich in mitgeführten Eppendorf-Cups® (Eppendorf,

Material und Methoden 60
Deutschland) befand. Pro Probe wurden ein neuer Cup und ein frisches Paar Hand-
schuhe verwendet.
Je Probe wurden immer zwei Amies- und ein Cary-Blair-Tupfer eingesetzt, die zum
Abwischen der jeweiligen Fläche gleichzeitig dicht nebeneinander in einer Hand ge-
halten wurden.
Es wurden stets ca. acht Nippeltränken, Ketten und Trogflächen von Futterautoma-
ten (gleichmäßig über die Buchten verteilt) in der Art mit einem „Satz“ Tupfer (ein
Cary-Blair-, zwei Amies-Tupfer) abgestrichen, dass jeweils deren komplette für die
Tiere zugängliche Fläche grobflächig einbezogen wurde. Bei Langtrögen, Lüftung,
Rohrleitungen, Buchtentrennwänden, Gang, Zentralgang, Buchtenboden, Decke und
Vorraum wurden an jeweils zehn verschiedenen Stellen, gleichmäßig über die Räum-
lichkeit verteilt, Proben entnommen. Dazu wurde an jeder Lokalisation ein sterilisier-
ter, handelsüblicher Einmach-Gummiring mit innerem Durchmesser von ca. 15 cm
angelegt und die darin liegende Fläche abgestrichen. Treibbretter, Stiefel und Fens-
ter(-bretter) wurden grobflächig komplett abgestrichen.
Die Proben wurden unverzüglich mit der Lokalisation und einer fortlaufenden Num-
mer beschriftet
3.1.4 Bakteriologische Untersuchung der Proben
Nach Ankunft in der Außenstelle wurden die Proben für den Versand verpackt und
per Post an das Institut für Mikrobiologie und Tierseuchen der Stiftung Tierärztliche
Hochschule Hannover versendet. Wenn der Postversand aufgrund später Rückkehr
aus dem Bestand erst am nächsten Tag erfolgen konnte, wurden die Proben bis zum
Versand im Kühlschrank gelagert.

Material und Methoden
61
3.1.4.1 Campylobacter spp.
Die Isolierung erfolgte in Anlehnung an die ISO 10272 (ANONYMOUS 2006a).
Dazu wurden die Tupfer in 10 ml Preston-Bouillon (Oxoid Deutschland, Wesel) über-
führt und bei 42 C für 24 Stunden in mikroaerophiler Atmosphäre bebrütet. Gleiches
geschah mit den vier Kotproben, nachdem der Inhalt jedes Probengefäßes zunächst
durch Rühren homogenisiert und dann jeweils 1 g davon entnommen wurde. Die
Proben wurde nach Ausstreichen auf Karmali-Agar (Oxoid) bei 42 C erneut für 48
Stunden mikroaerophil inkubiert. Verdächtige Kolonien wurden auf Thioglycolatagar
(Oxoid) subkultiviert und am folgenden Tag anhand der Kulturmorphologie und Pha-
senkontrastmikroskopie auf ihre Zugehörigkeit zu Campylobacter spp. untersucht.
Insekten wurden zusammen mit 5 – 6 Glaskugeln und 1 ml 0,9%iger physiologischer
Kochsalzlösung in ein FastPrep®-Röhrchen (MP Biomedicals, Frankreich) gefüllt und
mit dem Bead-Beater® (BioSpec Products, USA) für 40 Sekunden bei Stufe 5,5 ho-
mogenisiert. Das weitere Vorgehen entsprach dem der anderen Probenarten.
3.1.4.2 Y. enterocolitica
Tupfer, Insekten bzw. 1 g Kot (Präparation wie bei Campylobacter spp.) wurden für
insgesamt 3 Wochen bei 4 C in 10 ml PBS inkubiert. Nach 7, 14 und 21 Tagen er-
folgte ein Ausplattieren von PBS auf Cefsulidon-Irgasan-Novobiocin-Agar (Oxoid)
gefolgt von einer Inkubation dieser Platten bei 30 C für 24 – 48 Stunden. Verdächti-
ge Kolonien wurden auf Blut- und Gaßner-Agar für 24 Stunden subkultiviert. An-
schließend erfolgte die Auswertung sowie Differenzierung der Yersinia spp. mittels
API® 20E (BioMérieux).
3.1.4.3 Salmonella spp.
Die Salmonellenisolierung erfolgte nach ISO 6579:2003 (ANONYMOUS 2002). Dazu
wurden Tupfer, Insekten bzw. 1 g Kot (Präparation wie bei Campylobacter spp.) in
9 ml BPW (Oxoid) gegeben und für 20 – 24 Stunden bei 37 C bebrütet. 100 μl dieser

Material und Methoden 62
Suspension wurden in RV-Medium (Oxoid) überführt und 20 – 24 Stunden bei 42 C
inkubiert. Weitere 1 ml des BPW wurden in Müller-Kauffmann-Tetrathionat-
Novobiocin-Medium (VWR / Merck, Deutschland) gegeben und 20 – 24 Stunden bei
37 C bebrütet. Anschließend erfolgte aus beiden Flüssigmedien ein Ausstrich auf
Xylose-Lysin-Desoxycholat-Agar und Brilliantgrün-Phenolrot-Laktose-Saccharose-
Agar, gefolgt von einer Inkubation beider Selektivnährböden für 20 – 24 Stunden bei
37 C. Nach einer Subkultivierung auf Blut- und Gassner-Agar für weitere 20 – 24
Stunden erfolgte die Auswertung der Kulturen durch biochemische Reaktionen wie
Indol-, Citrat- und Ureasereaktion und Agglutinationsreaktion.
3.1.5 Erhebung der Betriebsdaten
Die Betriebsdaten wurden während eines Interviews mit dem Landwirt auf einem
Fragebogen erfasst. Dieser Fragebogen beinhaltete Angaben zu Art und Größe des
Tierbestands, Betriebsmanagement, Haltung, Fütterung sowie Hygienemaßnahmen
im Betrieb (siehe Anhang 1). Diese Daten wurden in eine Access®-Datenbank (Office
2003, www.microsoft.com) eingegeben, die eigens für das Projekt im Institut für Bio-
metrie, Epidemiologie und Informationsverarbeitung entwickelt wurde.
3.1.6 Verwaltung und Auswertung der Daten
Alle Proben wurden mit einem Code für Betrieb und Lokalisation sowie ihrer fortlau-
fenden Nummer in eine standardisierte Excel®-Tabelle (Office 2003,
www.microsoft.com) (Ausschnitt siehe Anhang 2) eingetragen und um die von der
Mikrobiologie übermittelten Untersuchungsergebnisse ergänzt (siehe Abschnitt 3.1.5).
Die Betriebsdaten wurden in einer Access-Datenbank zur Verfügung gestellt. Diese
beiden Datenquellen bildeten die Grundlage für die statistische Auswertung.
Die statistische Auswertung der Daten erfolgte mit der Software Statistical Analysis
System for Windows (SAS® Version 9.1, Cary, USA).

Material und Methoden
63
Das Ziel der Studie war, Kenntnisse über das Vorkommen der Erreger im Bestand
und an bestimmten Lokalisationen, über die Art des gemeinsamen Auftretens und
mögliche betriebliche Einflussfaktoren zu gewinnen. Dabei liegen aufgrund der
Erhebungsstruktur Daten auf unterschiedlichen Hierarchieebenen vor, die bei der
Auswertung berücksichtigt werden müssen. Daher gliederte sich die Auswertung in
folgende Punkte:
Übersicht über die Hierarchieebenen und Unterpunkte
1. Probenebene: Darstellung der bakteriologischen Befunde für alle drei Erreger
(unter Berücksichtigung der Unterschiede zwischen den Kategorien)
2. Lokalisationsebene: Darstellung der bakteriologischen Befunde aller drei Erreger
(mit Unterschieden zwischen den Kategorien) an den einzelnen Lokalisationen
und Lokalisationsgruppen
3. Betriebsebene:
a. Betriebsweise Darstellung der bakteriologischen Befunde aller Erreger
(mit Unterschieden zwischen den Kategorien) allgemein
b. Betriebsweise Darstellung der Befunde für einzelne Lokalisationen und
Lokalisationsgruppen
4. Gemeinsames Auftreten der Erreger
a. allgemein
b. an den einzelnen Lokalisationen
c. im Betrieb

Material und Methoden 64
5. Risikofaktorenanalyse: Verknüpfung der Probenergebnisse jedes Erregers auf
Betriebsebene mit Betriebsmerkmalen aus der Access-Datenbank und
Überprüfung dieser Merkmale auf einen Einfluss auf:
a. das Auftreten der einzelnen Erreger im Betrieb allgemein,
b. das Auftreten der einzelnen Erreger an bestimmten Lokalisationen,
c. das gemeinsame Auftreten von Erregern.
d. Außerdem erfolgte eine multifaktorielle Analyse ausgewählter
Einflussgrößen.
Für alle statistischen Tests wurden aufgrund der gegebenen Stichprobengröße p-
Werte < 0,1 als auffällig und p-Werte < 0,05 als statistisch signifikant betrachtet.
3.1.6.1 Auswertung der Ergebnisse auf Probenebene
Für die Auswertung auf Probenebene wurden Ergebnisse der kulturellen
Untersuchung mit den dichotomen Ausprägungen ‚positiv„ und ‚negativ„ betrachtet.
Neben tabellarischen Darstellungen wurden für den Vergleich des Anteils positiver
Proben je Erreger zwischen den Kategorien Kontingenztafeln (SAS-Prozedur PROC
FREQ) erstellt und der Chi-Quadrat-Homogenitäts-Test (Option CHISQ) angewandt.
Liefert dieser Test einen p-Wert < 0,05, so kann die Nullhypothese auf Homogenität
der beiden Stichproben verworfen werden, und es besteht eine signifikante
Assoziation zwischen der Zeilen- und der Spaltenvariable (KREIENBROCK u.
SCHACH 2005).
3.1.6.2 Auswertung der Ergebnisse auf Lokalisationsebene
Die Auswertung erfolgte analog der Probenebene, mit zusätzlicher Differenzierung
nach Lokalisationen und Lokalisationgruppen. Für alle Vergleiche dichotomer
Merkmale wurde der Fisher-Exakt-Test (PROC FREQ Option FISHER) angewandt.

Material und Methoden
65
Dieser Test wird bei sehr kleinen Stichprobenzahlen anstelle des Chi-Quadrat-Tests
benutzt.
3.1.6.3 Auswertung der Ergebnisse auf Betriebsebene
Für die Analyse der Ergebnisse auf Betriebsebene wurde ein Betrieb insgesamt als
positiv eingestuft, wenn aus mindestens einer in ihm entnommenen Probe ein Erre-
gernachweis erfolgte. Bei Campylobacter spp erfolgte aufgrund der hohen Anzahl
positiver Betriebe eine weitere Unterteilung: Betriebe mit einem bis vier Nachweisen
wurden als „schwach positiv“ definiert, solche mit mindestens fünf positiven Proben
als „hoch positiv“.
Neben dieser Charakterisierung der Betriebe wurden auch die erregerbezogenen
Anzahlen positiver Proben im Betrieb im Sinne eines diskreten Merkmals für
Berechnungen herangezogen.
Vergleiche zwischen Betrieben wurden zunächst in der allgemeinen Darstellung für
jeden Erreger mit dem Fisher-Exakt-Test durchgeführt.
Für die Anzahlen positiver Proben pro Betrieb und Erreger wurden zur Visualisierung
der Verteilungen Boxplots (SAS Modul INSIGHT) erstellt und die Lage der
Verteilungen jeweils zwischen zwei Erregern verglichen. Dies geschah wegen häufig
nicht normaler Verteilungen (visuelle Prüfung mithilfe PROC UNIVARIATE überprüft)
mit dem Wilcoxon-Rangsummen-Test (PROC NPAR1WAY WILCOXON).
Zur genaueren Bestimmung des Einflusses dieser Risikovariablen (Anzahl
erregerpositiver Proben im Betrieb) auf die Kategorie wurde eine logistische
Regression (PROC LOGISTIC) durchgeführt. Das resultierende Odds Ratio setzt bei
dichotomen Einflussmerkmalen die Chance (engl. odds) für den Eintritt eines
Ereignisses bei Exposition mit der Chance des Eintritts ohne Exposition ins
Verhältnis (engl. ratio) (KREIENBROCK u. SCHACH 2005). Bei quantitativer

Material und Methoden 66
Einflussvariable wird damit die Erhöhung der Odds bei Erhöhung dieser
Einflußvariable um den Faktor eins angegeben. Je nach Qualität des Einflusses
(protektiver oder Risikofaktor) kann die Odds Ratio Werte unter bzw. über eins
annehmen.
Anschließend wurden eine Auswertung der Betriebspositivität einzelner
Lokalisationen und Gruppen vorgenommen. Dafür wurde eine Lokalisation oder
Gruppe eines Betriebes analog der allgemeinen Betriebspositivität als positiv
eingestuft, wenn mindestens eine Probe davon positiv war. Anhand dieser Einteilung
wurde überprüft, inwieweit die Ergebnisse von je zwei Lokalisationsgruppen
innerhalb der Betriebe übereinstimmten. Dazu wurde der einfache Kappa-Koeffizient
(PROC FREQ Opt. AGREE) berechnet.
Ergänzend wurde für jeden Erreger mittels logistischer Regression die Stärke des
Einflusses positiver Proben in einer Gruppe auf die Positivität einer anderen Gruppe
sowie auf die Salmonellen-Kategorie geprüft.
3.1.6.4 Auswertung des gemeinsamen Erregerauftretens
Auch hier wurde „von innen nach außen“ vorgegangen und zuerst die
Übereinstimmung von jeweils zwei Erregern in einer Probe und dann auf
Betriebsebene analysiert. Dazu wurde wieder der einfache Kappa-Koeffizient
bestimmt. Für die Lage der Verteilungen der Erregeranzahl im Betrieb je nach
Kategorie wurde erneut der Wilcoxon-Test eingesetzt.
3.1.6.5 Risikofaktorenanalyse
Aus der Fragebogen-Datenbank wurden 17 qualitative Betriebsmerkmale mit jeweils
mehreren Unterpunkten auf ihre Eigenschaft als Risikofaktor überprüft. Die Auswahl
dieser nominalen Variablen erfolgte anhand der Verteilung ihrer einzelnen
Beobachtungen. Es wurden nur Merkmale ausgewertet, bei denen bei mindestens

Material und Methoden
67
zwei der Antwortmöglichkeiten jeweils mindestens fünf Beobachtungen vorlagen.
Lagen bei mehr als zwei Antwortmöglichkeiten solche mit weniger als fünf
Beobachtungen vor, wurden diese mit anderen Antwortmöglichkeiten
zusammengefasst oder nicht berücksichtigt.
Die Analyse erfolgte, für jeden Erreger getrennt, in zwei Varianten: Zum Einen diente
das Vorkommen des Erregers im Betrieb allgemein als Zielvariable. Der Einfluss der
verschiedenen Ausprägungen der Risikovariablen wurde dann mit der logistischen
Regression quantifiziert. Andererseits erfolgten Berechnungen anhand der Anzahlen
positiver Proben im Betrieb als diskretem Merkmal. Hier erfolgten die paarweisen
Vergleiche aufgrund geringer Stichprobenzahl und Nicht-Normalverteilung mit dem
Wilcoxon-Rangsummen-Test. Bei mehr als zwei Merkmalsausprägungen wurden
zunächst der Kruskal-Wallis-Test und anschließend die paarweisen Vergleiche
durchgeführt. Bei diesem multiplen Testen wurde eine Adjustierung des
Signifikanzniveaus für multiples Testen vorgenommen und so eine
versuchsbezogene Irrtumswahrscheinlichkeit (p = 0,05 / Anzahl Vergleiche) gewahrt.
Außerdem wurden fünf quantitative (stetige) Merkmale untersucht, deren Verteilun-
gen in Boxplots dargestellt wurden. Ihre Auswirkungen auf die Anzahlen positiver
Proben eines Betriebs wurden in einer linearen Regression ermittelt (PROC REG).
Um das Testergebnis nicht zu verzerren, wurden die in den Boxplots dargestellten
Ausreißer eines Merkmals aus der Berechnung ausgeschlossen.
Analog den Analysen auf Betriebsebene wurde auch die Positivität bestimmter Loka-
lisationsgruppen und Einzellokalisationen (Sammelkot, tiernahe und tierferne Umge-
bung, Buchtenaußenwände, Buchtentrennwände) im Zusammenhang mit den Ein-
flussvariablen untersucht. Hier kam aufgrund der sehr geringen Stichprobenzahl der
Fisher-Exakt-Test zur Anwendung.
Die Analyse des Einflusses bestimmter Merkmale auf mehrere Erreger gemeinsam
kann anhand eines multivariaten Modells erfolgen. Die statistische Power eines sol-

Material und Methoden 68
chen Modells ist aufgrund der geringen Stichprobengröße jedoch nicht als ausrei-
chend anzusehen. Auch eine Berechnung anhand gebildeter Erregerkombinationen
schied aufgrund der Verteilung dieser (siehe Ergebnisteil, Tabelle 17) aus. Zur Umge-
hung dieser Schwierigkeiten wurde behelfsmäßig ein mehrfaktorielles Modell erstellt,
in dem durch Vertauschen der eigentlichen Zielvariablen (Erregervorkommen) mit
den Einflussfaktoren der Grad der Assoziation einzelner Erreger mit bestimmten Be-
triebsmerkmalen bestimmt wurde. Dafür wurden Betriebsmerkmale ausgewählt, für
die in der einfaktoriellen Analyse ein signifikanter Einfluss auf mindestens zwei Erre-
ger belegt wurde. Dafür wurde, wie bei der multifaktoriellen Analyse beschrieben,
verfahren.
Zur multifaktoriellen Analyse ausgewählter Einflussfaktoren wurde vor Berechnung
des eigentlichen Modells (PROC LOGISTIC) zunächst der Effekt jeder Einflussgröße
einzeln auf die Zielvariable, getrennt für jeden Erreger, getestet. Im zweiten Schritt
schloss sich die Überprüfung eines Zusammenhangs zwischen den einzelnen Ein-
flussgrößen an (beides mittels Chi-Quadrat-Homogenitäts-Test). War dieser sehr
groß, wurden diese Variablen nicht gemeinsam in das Modell aufgenommen. Die
Modelle sollten die Merkmalskomplexe Fütterung, Gerätehygiene, Abteilhygiene und
Schädlingsbekämpfung einschließen.

Material und Methoden
69
3.2 Salmonellenverlaufsuntersuchung
3.2.1 Art und Ziel der Untersuchung
Die Untersuchung wurde im Rahmen des durch das Bundesministerium für Bildung
und Forschung geförderten Verbundprojektes „Food-borne zoonotic infections“ (FBI-
Zoo) durchgeführt. Sie erfolgte in enger Kooperation mit dem Nationalen Referenz-
zentrum für Salmonella spp. des Robert Koch-Instituts in Wernigerode.
Ziel der Untersuchung war, nähere Erkenntnisse über Eintragsquellen und die fol-
gende Entwicklung, Verteilung und Dynamik von Salmonellen in Schweinemastbe-
ständen zu erhalten. Zu diesem Zweck wurde eine Verlaufsuntersuchung in zwölf
Betrieben in Niedersachsen durchgeführt, bei der regelmäßig Kot- und Umgebungs-
proben entnommen und auf Salmonellen untersucht wurden. Diese wurden zur bes-
seren Rückverfolgbarkeit einzelner klonaler Linien, im Folgenden als „Klone“ be-
zeichnet, feintypisiert.
3.2.2 Auswahl der Bestände
Für die Untersuchung ausgewählt wurden Schweinemastbestände in Niedersachsen,
die in Salmonellen-Kategorie III eingestuft waren und bei denen in einer vorange-
gangenen Untersuchung molekularbiologisch Salmonellennachweise erfolgt waren.
Die Bestände mussten in konventioneller Haltung und im Rein-Raus-System wirt-
schaften. Teilnehmen konnten reine Mäster und Bestände im geschlossenen System.
Aus ersterer Gruppe rekrutierten sich elf Bestände, aus letzterer einer.
3.2.3 Untersuchung der Bestände
3.2.3.1 Zeitraum und Frequenz der Probenentnahme
Die Untersuchungen in den Beständen wurden im Zeitraum von Mai 2008 bis No-
vember 2009 durchgeführt. In jedem Bestand wurden regelmäßige Probenentnah-

Material und Methoden 70
men über einen bis maximal drei Mastdurchgänge durchgeführt. Ein Mastdurchgang
(DG), sowohl bei den Mastbetrieben als auch bei dem geschlossenen System, be-
gann mit der Einstallung einer Tiergruppe in das ausgewählte Abteil und endete mit
Ausstallung des letzten Tieres aus diesem. Während jedes DG wurden vier bzw. fünf
Probenentnahmen durchgeführt. Der erste Zeitpunkt der Probenentnahme (ZP 0)
eines jeden Mastdurchgangs war stets die Probenentnahme im leeren, gereinigten
und desinfizierten Abteil. Den nächsten Zeitpunkt (ZP 1) stellte die Ankunft der Ferkel
im Maststall dar. Dafür wurden die Ferkel direkt bei Ankunft auf dem Transportfahr-
zeug bzw. bei geschlossenen Systemen kurz vor Umtrieb in den Maststall im Flat-
deck untersucht. Daran schlossen sich je nach Dauer des Mastdurchgangs drei bis
vier weitere Probenentnahmen im Abstand von vier Wochen an (ZP 2 – 5).
In Betrieben ohne kulturellen Salmonella-Nachweis wurde nur ein Mastdurchgang
untersucht. In Betrieben, die nach zwei Mastdurchgängen maximal fünf Salmonella-
Nachweise aufwiesen, wurden die Probenentnahmen sodann abgeschlossen. In Be-
trieben mit mehr als fünf kulturellen Erregernachweisen nach zwei Durchgängen
wurden die Untersuchungen auf einen dritten Mastdurchgang ausgedehnt.
3.2.3.2 Probenarten und -lokalisationen
Die Probenentnahmen schlossen Sammelkotproben und verschiedene Lokalisatio-
nen in der Tierumgebung, sowohl in den Abteilen als auch außerhalb, ein. Außerdem
wurden im Bestand vorgefundene Schadnager und Insekten in die Untersuchung
einbezogen. Die Probenentnahme erfolgte in jedem Bestand immer in demselben, zu
Beginn festgelegten Abteil und an den gleichen Lokalisationen. Die folgenden Lokali-
sationen wurden untersucht:
Lüftung
Rohrleitungen
Ketten
Nippeltränken
Futtertröge
Buchtentrennwände
Außenwände
Buchtenboden
Sammelkot
Gang
Zentralgang
Treibbrett
Stiefel
Fliegen/Insekten

Material und Methoden
71
Nagerkot
Oberflächentupfer tote Nager
Fenster(-brett)
Heizung
Decke
Futterproben
Boden vor Stall
Waage
Hygiene-/Arbeitsraum
sonst. Arbeitsmittel (Besen u. ä.)
In jedem Betrieb wurden je nach individuellen örtlichen Gegebenheiten 11 bis 23
Proben der genannten Lokalisationen sowie eine bis zwei Sammelkotproben ent-
nommen.
3.2.3.3 Durchführung der Probenentnahme
Die Sammelkotproben wurden mit sogenannten „Sockentupfern“ entnommen. Dazu
wurde ein steriles, ca. 45 cm langes Stück eines Gazeschlauchs (Tubular bandage
tg®, Lohmann und Rauscher, Deutschland) über den Gummistiefel gestülpt und damit
die Buchten im Abteil bzw. das Transportfahrzeug durchschritten. Es wurden immer
so viele Buchten in die Untersuchung einbezogen, dass darin insgesamt ca. 100 Tie-
re eingestallt waren. Für die Kotprobe bei Ankunft der Ferkel wurden eine Ebene des
Transportfahrzeugs und dessen Rampe beschritten.
Die Umgebungsproben wurden mittels „Swab“ Wischtupfer entnommen. Dazu wur-
den etwa 35 cm lange, quadratisch gefaltete und mit gepuffertem Peptonwasser
(BPW®, Oxoid Deutschland, Wesel) getränkte Gazeschlauch-Stücke verwendet. Die-
se waren in der Vorbereitung einzeln in Plastikbeutel eingeschweißt und einer Sterili-
sation unterzogen worden. Die Entnahme jeder Probe erfolgte mit einem frischen
Paar Handschuhe. Anschließend wurde der Tupfer wieder in den zugehörigen Plas-
tikbeutel verbracht.
Für jede Lokalisation wurden bei jeder Probenentnahme in einem Betrieb immer die-
selben Flächen einbezogen.
Bei Nippeltränken, Ketten und Breiautomaten (je nach Betrieb immer vier bis sechs
je Probe) wurde jeweils deren komplette, für die Tiere zugängliche Fläche abge-

Material und Methoden 72
wischt. Bei Lüftung, Rohrleitungen, Buchtentrennwänden, Gang, Zentralgang, Buch-
tenboden, Decke und Vorraum wurden zehn verschiedene Stellen, gleichmäßig über
die Räumlichkeit verteilt, untersucht. Dazu wurde pro Stelle jeweils eine Fläche in der
dreifachen Größe eines komplett auseinander gefalteten „Swabs“ (entsprechend ca.
850 cm2) abgewischt. Treibbretter, Stiefel, Fenster(-bretter), Heizgeräte, Waage und
sonstige Arbeitsmittel wurden komplett abgewischt.
Augenscheinlich kürzlich verstorbene Schadnager (Ratten, Mäuse) wurden durch
Abtupfen ihrer Körperoberfläche beprobt. Zur Untersuchung von Insekten wurden ca.
15 Fliegen, Käfer oder Kakerlaken, wo möglich, an verschiedenen Stellen im Stall
eingefangen bzw. eingesammelt und in ein Kotprobenröhrchen gefüllt. Auch Nager-
kot wurde, falls vorgefunden, an verschieden Stellen entnommen und davon 2 – 3 g
in ein Kotröhrchen gefüllt.
Jede Probe wurde im Betrieb mit einer Identifikationsnummer, die aus einer Kodie-
rung für den Betrieb, den Durchgang, den Zeitpunkt im Durchgang sowie die Lokali-
sation bestand, beschriftet.
3.2.4 Untersuchung der Proben
3.2.4.1 Probenverarbeitung in der Außenstelle für Epidemiologie in Bakum
Nach Rückkehr zur Außenstelle wurden die Proben zusammen mit 225 ml (im Falle
von Sammelkot) bzw. 90 ml (übrige Proben) sterilem gepufferten Peptonwasser
(BPW®, Oxoid Ltd., UK) in Plastikbeutel (Stomacher 400®, Seward Ltd., UK) über-
führt und für 15 Sekunden in einem Stomacher homogenisiert. Bei allen Probenloka-
lisationen, bei denen kein direkter Kontakt mit Tierkot zu erwarten war (Lüftung,
Rohrleitungen etc.), wurden dem Ansatz zusätzlich 120 μl einer Lösung mit 9 μg Fer-
rioxamin in destilliertem Wasser zugefügt, um subletal geschädigte Salmonella spp.
im Wachstum zu unterstützen (Reissbrodt et al., 1995). Im Anschluss wurden die
Proben für 24 Stunden aerob bei 37 C inkubiert und dann je ca. 1,5 ml jeder Probe in

Material und Methoden
73
2-ml-Eppendorf-Cups (Eppendorf®, Deutschland) abgefüllt. Diese wurden zur weite-
ren Bearbeitung und Untersuchung auf Salmonella spp. per Post an das Robert-
Koch-Institut nach Wernigerode versandt.
3.2.4.2 Untersuchung der Proben am Robert-Koch-Institut Wernigerode
3.2.4.2.1 Salmonella-Nachweis und -Isolierung
Anreicherung der Proben
Für die selektive Anreicherung wurden 150 µl der in BPW angereicherten Proben zu
10 ml RV-Anreicherungsbouillon gegeben und 48 h bei 42 °C, 100 rpm und 45 %
Neigung inkubiert. Nach Zentrifugation von 1 ml Kultur für 5 Minuten bei 13.000 rpm
(Biofuge fresco®; Heräus, Hanau) wurde der Überstand mittels Vakuum-
Absaugsystem entfernt, das Pellet in 500 µl sterilem destilliertem Wasser resuspen-
diert und wiederum für 5 Minuten bei 13.000 rpm zentrifugiert. Das Pellet wurde an-
schließend je nach Größe in 50 bzw. 100 µl sterilem destillierten Wasser resuspen-
diert.
Salmonella-Nachweis
Die Identifizierung der Salmonellen erfolgte mittels invA-PCR (CHEN et al. 1997),
modifiziert durch SCHOLZ et al. (SCHOLZ et al. 2001), mit den in Tabelle 5 angegebe-
nen Primern in einem 2720 Thermal Cycler (Applied Biosystems, Deutschland) unter
folgenden Reaktionsbedingungen: 95 °C 10‟; 30 x [94°C 30”; 64°C 30”; 72°C 30”];
72°C 10‟; 4°C. 5 µl des PCR-Produktes wurden in einem 1,5 % Agarosegel bei 150 V
eine Stunde aufgetrennt.
Tabelle 5 Primer der invA-PCR
Primername Richtung Sequenz 5‟-3‟ Endkonzentration
mwg 139 forward 5‟-GTGAAATTATCGCCACGTTCGGGCAA-3‟ 0,5 µM
mwg 141 reverse 5‟-TCATCGCACCGTCAAAGGAACC-3‟ 0,5 µM

Material und Methoden 74
Salmonella-Kultur
Der Ausstrich der in RV-Anreicherungsbouillon gewachsenen Kultur auf Nährmedium
erfolgte je nach Stärke des invA-Signals. Bei starkem Signal wurden 100 µl einer
10-3- und 10-6-Verdünnung auf OSCM-Nährmedium ausplattiert, und die Proben 24
Stunden bei 37 °C inkubiert.
Bei schwachem invA-Signal wurden 100 µl einer 10-3 verdünnten Kultur aus dem RV-
Anreicherungsbouillon auf Rambach-Nährmedium (Oxoid) ausplattiert und 24 Stun-
den bei 37 °C inkubiert. Parallel dazu wurde die übrige Kultur 10 Minuten bei
4000 rpm zentrifugiert, das Pellet in frisches 10 ml RV-Bouillon überführt und die
Probe nochmals 48 Stunden bei 42 °C, 100 rpm und 45° Neigung inkubiert. 100 µl
10-3- Verdünnung wurden anschließend auf OSCM-Nährmedium ausplattiert und 24
Stunden bei 37 °C inkubiert. 20 Salmonella-Kolonien jeder Probe wurden auf OSCM-
Nährmedium überführt. Nach einem Wachstum über 24 Stunden bei 37 °C wurden
die Kolonien nochmals auf OSCM überimpft und bei 37 °C 24 Stunden inkubiert. An-
schließend wurde je eine Einzelkolonie in 100 µl destilliertem Wasser eingerieben.
Screening zur Identifikation von VNTR- Varianten
Zur Detektion von Salmonella-Varianten innerhalb einer Probe wurden die in Tabelle
6aufgeführten drei VNTR-Regionen amplifiziert (LINDSTEDT et al. 2004).
Tabelle 6 VNTR-Regionen der Screening-PCR zur Detektion von Salmonella-Varianten
VNTR-Region Richtung Sequenz 5‟-3‟
STTR 5 forward 5'-ATGGCGAGGCGAGCAGCAGT-3'
reverse 5'-GGTCAGGCCGAATAGCAGGAT-3'
STTR 6 forward 5'-TCGGGCATGCGTTGAAA-3'
reverse 5'-CTGGTGGGGAGAATGACTGG-3'
STTR 9 forward 5'-AGAGGCGCTGCGATTGACGATA-3'
reverse 5'-CATTTTCCACAGCGGCAGTTTTTC-3'
Die PCR erfolgte unter den folgenden Reaktionsbedingungen in einem 2720 Thermal
Cycler (Applied Biosystems):

Material und Methoden
75
Mastermix: 5 µl Hotstar Taq Master Mix 1 µl 2,5 pmol/µl Primer STTR 5 1 µl 2,5 pmol/µl Primer STTR 6 1 µl 2,5 pmol/µl Primer STTR 9
PCR-Ansatz: 7µl Mastermix 3 µl Bakteriensuspension
PCR-Programm: 95°C 15‟; 25 x [94°C 30”; 60°C 1‟30”; 72°C 1‟30”]; 72°C 10‟; 4°C. 2,5 µl Amplifikationsprodukt wurden in einem 2,0 % Agarosegel bei 150 V,
1,5 Stunden aufgetrennt. Anschließend erfolgte die Diskriminierung der MLVA-
Varianten. Von jeder Variante wurde je eine Einzelkolonie zweier unterschiedlicher
Isolate auf LB (Luria Broth)-Nährmedium ausgestrichen und bei 37 °C 24 Stunden
inkubiert. Nach Animpfung von je einer Kolonie in ein Schrägagarröhrchen (8 g/l
Nutrient Broth (Difco, Augsburg, Deutschland), 5 g/l Bacto Pepton (Difco), 2 g/l
Na2PO4 x 2 H2O, 5 g/l NaCl, 15 g/l Agar, pH 7,4) und einer Inkubation bei 37 °C für
24 Stunden wurden die Isolate bis zur weiteren Bearbeitung bei Raumtemperatur im
Dunkeln gelagert. Eine zweite Lagerungsmethode war die Kryokultur. Dazu wurde
eine Einzelkolonie von LB-Nährmedium in 2 ml LB-Flüssigmedium (25 g/l LB (Miller‟s
Broth), ph-Wert 7 +/- 0,2) überführt und bei 37 °C, 100 rpm 6 Stunden inkubiert. An-
schließend wurden 500 µl einer Glycerinlösung (40 % Glycerol in H2O) mit 500 µl der
LB-Flüssigkultur beimpft und diese Proben sodann bei -80 °C gelagert.
3.2.4.2.2 Molekulare Typisierung
An den Nachweis schloss sich zum Zwecke einer Eingruppierung der Isolate in ver-
schiedene klonale Linien die komplexe Typisierung der isolierten Salmonella spp. an.
Diese erfolgte anhand von drei verschiedenen Methoden:
Multiple-Locus Variable-Number of Tandem-Repeats Analysis (MLVA)
Die Amplifikation der VNTR-Orte STTR 3, STTR 5, STTR 6, STTR 9 und STTR 10pl
erfolgte in modifizierter Form nach LINDSTEDT et al. (2004). Die PCR-Produkte
wurden im Genetic Analyser ABI 310® analysiert und mittels GeneMapper Software
Version 4.0 (Applied Biosystems) ausgewertet.

Material und Methoden 76
Die Analyse von elf über das Genom verteilten VNTR-Orten (Repeat units von 50-
200 bp) wurde nach einer laborinternen und noch nicht publizierten Methode durch-
geführt.
Multilocus Sequenz-Typisierung (MLST)
Anschließend wurde eine MLST (KIDGELL et al. 2002) durchgeführt. Die MLST Se-
quenztypen wurden anhand der Webseite http://mlst.ucc.ie/mlst/dbs/Senterica be-
stimmt.
Pulsfeld-Gelelektrophorese (PFGE)
Die PFGE erfolgte nach Verdau der genomischen DNA mit den Restriktionsenzymen
XbaI bzw. BlnI nach dem Puls-Net Protokoll (RIBOT et al. 2006). Die Gele wurden
anschließend mit BioNumerics 5.1 (Applied Maths, Saint-Martins-Latem, Belgien)
analysiert. Die Klusteranalyse erfolgte anhand der DICE-Koeffizienten und einem
UPGMA-Dendrogramm. Das Dendrogramm diente auch zur Definition resp. Diskri-
minierung unterschiedlicher PFGE-Muster.
3.2.4.2.3 Phänotypie
Zur Vervollständigung der Informationen zu den einzelnen Isolaten wurde außerdem
eine klassische Phänotypisierung vorgenommen.
Serotypie
Die serologische Identifizierung der Stämme erfolgte nach dem White-Kauffmann-Le
Minor Schema durch Agglutination von O- und H-Antigen-spezifischen Seren (Sifin
Diagnostica, Berlin, Deutschland).
Lysotypie
Zur Lysotypie (ANDERSON et al. 1977). wurden eine zweistündige LB-Flüssig-Kultur
auf Oxoid-3 Nährmedium ausplattiert und Serovar-spezifische Phagen aufgebracht.

Material und Methoden
77
Nach einer Inkubation von 24 Stunden bei 37 °C erfolgte die mikroskopische Auswer-
tung.
3.2.5 Eingruppierung der Salmonella-Isolate in klonale Linien
Die Salmonella-Isolate wurden im Rahmen eines weiteren Teilprojektes des For-
schungsverbunds FBI-Zoo durch das Robert Koch-Institut feintypisiert und in klonale
Linien, vereinfachend als „Klon“ bezeichnet, eingruppiert.
Dies erfolgte anhand der Übereinstimmung der Ergebnisse der drei unter „3.2.4.2.2
molekulare Typisierung“ beschriebenen Verfahren MLVA, MLST und PFGE: Aus-
schlaggebend waren dabei die Resultate der beiden MLVA-Analysen: Zwei Salmo-
nella-Isolate wurden der gleichen klonalen Linie (in etwa gleichzusetzen mit
„Stamm“) zugeordnet, wenn beide Untersuchungen identische oder nur äußerst ge-
ring abweichende Ergebnisse lieferten. MLST und PFGE wurden zur Bestätigung der
anhand der MLVA vorgenommenen Einteilung der Klone eingesetzt, letztere auch
zur Charakterisierung von Varianten innerhalb einer klonalen Linie.
3.2.6 Erhebung von Betriebsdaten
Zu den Beständen standen betriebliche Informationen in einer Access-Datenbank zur
Verfügung (siehe Querschnittsuntersuchung). Zusätzlich wurden Veränderungen im
Betriebsmanagement (z.B. Futterwechsel, Änderungen bei Reinigung und Desinfek-
tion) während der Untersuchung dokumentiert. Ebenso wurden Bestandserkrankun-
gen und Medikamentengaben in den relevanten Abteilen während des Untersu-
chungszeitraums sowie die Untersuchungsergebnisse bezüglich der Salmonella-
Antikörper erfasst.

Material und Methoden 78
3.2.7 Verwaltung und Auswertung der Daten
Alle Proben wurden mit ihrer Identifikationsnummer fortlaufend in einer standardisier-
ten Excel-Tabelle eingetragen. In dieser wurden die durch das Robert-Koch-Institut
übermittelten Untersuchungsergebnisse aus Kultur und PCR erfasst (siehe Anhang
3). Die Ergebnisse der Feintypisierung wurden durch das Robert-Koch-Institut eben-
falls in Form einer Excel-Tabelle übermittelt (Anhang 4).
Die statistische Auswertung der Daten erfolgte mit (SAS® Version 9.1 (Cary, USA).
Die Auswertung verfolgte das Ziel, typische Verläufe und sich wiederholende Muster
von Salmonelleninfektionen auf räumlicher und zeitlicher Ebene in der Mast zu identi-
fizieren. Der Schwerpunkt der Untersuchung lag aus epidemiologischer Sicht auf der
betriebsübergreifenden Analyse. Dabei wurden folgende Punkte und ihre Kombinati-
onen auf einen möglichen Einfluss auf das Salmonellenvorkommen im Bestand un-
tersucht:
a. Durchgang
b. Zeitpunkt
c. Probenlokalisation
Die Analyse erfolgte auf zwei Ebenen:
1. Darstellung der Ergebnisse auf Ebene der Probe (Salmonella-positiv ja – nein),
und
2. Darstellung der Ergebnisse auf Ebene der durch das Robert-Koch-Institut defi-
nierten klonalen Linie der isolierten Salmonella spp.

Material und Methoden
79
Auswertung auf Probenebene
Die Berechnungen wurden anhand der Ergebnisse der kulturellen Untersuchung
durchgeführt, da diese auch Grundlage aller Analysen auf Klonebene waren (Diffe-
renzierung in der Kultur erhaltener Isolate). Gleichwohl erfolgte ein Abgleich der
Übereinstimmung der kulturellen Ergebnisse mit denen aus der Screening(-invA)-
PCR.
In die Analysen einbezogen wurden alle die Betriebe, in denen kulturell mindestens
ein Salmonella-Nachweis erfolgt war.
Zur Darstellung der zeitlichen Dynamik wurden Trendanalysen mittels des Cochran-
Armitage-Trend-Tests (PROC FREQ Option TREND) durchgeführt. Dieser Test prüft
einen Trend für eine binomial verteilte Variable über die Ausprägungen einer zweiten
kategoriellen Variable.
Des Weiteren wurden die Untersuchungsergebnisse von Proben, die zu verschiede-
nen Zeitpunkten entnommen worden waren, in Gruppen zusammengefasst und in
Kontingenztafeln gegenübergestellt (z. B. leeres - belegtes Abteil).
Zur besseren Auswertung der räumlichen Dynamik wurden alle Einzellokalisationen
einer von vier Gruppen zugeordnet: Die Lokalisationsgruppe „Sammelkot“ beinhalte-
te sowohl Sammelkotproben bei Anlieferung der Schweine, als auch aus dem beleg-
ten Abteil. Die „tiernahe Umgebung“ beinhaltete Spielketten, Tränken, Futtertröge,
Buchtenzwischen- und Buchtenaußenwände. Zur „tierfernen Umgebung“ zählten Lüf-
tung, Rohrleitungen, Abteil- und Zentralgänge, Treibwege, Treibbretter, Stiefel, Fens-
ter und Arbeitsräume. Die „Schädlinge“ inkludierten Nagerkot, Oberflächentupfer von
Nagern und Fliegen.

Material und Methoden 80
Der Vergleich der Untersuchungsergebnisse der Lokalisationsgruppen erfolgte mit-
tels Chi-Quadrat-Test, im Falle von sehr geringen Stichprobenzahlen (Felder mit
weniger als 5 Beobachtungen in der Kontingenztafel) mit dem Fisher-Exakt-Test.
Im Zuge einer kombinierten Analyse räumlicher und zeitlicher Verläufe erfolgten Ver-
gleiche verschiedener Lokalisationsgruppen zu bestimmten Zeitpunkten und Trenda-
nalysen für einzelne Lokalisationsgruppen.
Zur Quantifizierung des Einflusses einer Risikovariablen wurde eine logistische
Regression (PROC LOGISTIC) durchgeführt.
Die gleichen Auswerteschritte erfolgten bei den Vergleichen der einzelnen Betriebe.
Um die Möglichkeit zu überprüfen, dass bei dieser Auswertung eine Zugrundelegung
der PCR-Ergebnisse anstelle der bakteriologischen Befunde andere Resultate gelie-
fert hätte, wurde ein Vergleich beider Verfahren durchgeführt. Dafür wurde die Ab-
weichung der beiden Methoden, sowohl insgesamt, als auch zwischen Betrieben,
Lokalisationsgruppen und Zeitpunkten mittels des Fisher-Exakt-Tests miteinander
verglichen. Der Grad der Übereinstimmung der Gesamtergebnisse wurde zusätzlich
anhand des einfachen Kappa-Koeffizienten überprüft. Außerdem wurde der McNe-
mar-Test auf Symmetrie angewandt.
Auswertung auf Ebene der klonalen Linie („Klonebene“)
Basis der Auswertung war hier die durch das Robert Koch-Institut vorgenommene
Zuordnung aller Salmonella-Isolate zu klonalen Linien. Alle Salmonella-Isolate, die
der gleichen klonalen Linie angehörten, erhielten die gleiche „Klon“-Nummer.
Zum einen wurden die Anzahlen der Nachweise der verschiedenen Klone ausgewer-
tet. Zum anderen wurde für jede mögliche Auswerteeinheit (Betrieb, Durchgang all-

Material und Methoden
81
gemein oder bestimmter Durchgang, Zeitpunkt allgemein oder eines bestimmten
Durchgangs, Lokalisationsgruppe) das allgemeine Vorkommen eines Klons (unab-
hängig von der Anzahl seiner Nachweise) untersucht. Auf diese Weise wurden die
einzelnen Durchgänge, die einzelnen Probenentnahmezeitpunkte innerhalb und zwi-
schen den Durchgängen sowie die Lokalisationen untereinander und in Verbindung
mit Durchgang und Zeitpunkt ausgewertet. Klone, die in verschiedenen Betrieben
auftraten, wurden für jeden Betrieb getrennt berücksichtigt. Bei Auswertung der Zeit-
punkte innerhalb eines Durchgangs allgemein flossen Klone, die in drei Durchgängen
auftraten, auch drei Mal in die Berechnungen ein.
Der Grad der Übereinstimmung des Klonvorkommens zu zwei verschiedenen Zeit-
punkten oder Durchgängen wurde, bei ausreichender Stichprobengröße mittels Kap-
pa-Index und McNemar-Test bestimmt. Die Analyse eines Zusammenhangs zwi-
schen Lokalisationen und dem Vorkommen eines Klons wurde mit dem Chi-Quadrat-
Test oder dem Fisher-Exakt-Test berechnet, das Ausmaß des Zusammenhangs mit-
tels logistischer Regression.
Die Auswertung schloss mit einer kurzen Beschreibung der räumlichen und zeitlichen
Verläufe der Klonnachweise in den einzelnen Betrieben ab. Eine Risikofaktorenana-
lyse konnte aufgrund der geringen Stichprobenzahl der Betriebe und geringer Varia-
tion der Einflussgrößen (Tabelle 23, Ergebnisteil) nicht durchgeführt werden.
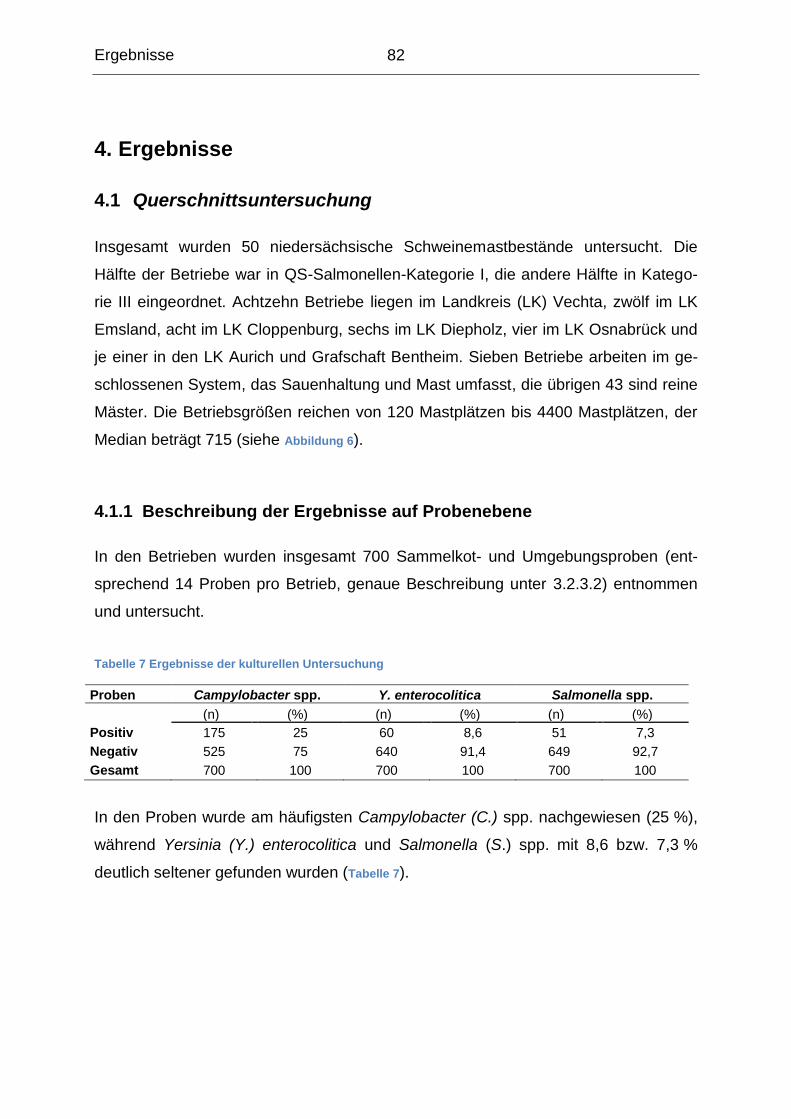
Ergebnisse
82
4. Ergebnisse
4.1 Querschnittsuntersuchung
Insgesamt wurden 50 niedersächsische Schweinemastbestände untersucht. Die
Hälfte der Betriebe war in QS-Salmonellen-Kategorie I, die andere Hälfte in Katego-
rie III eingeordnet. Achtzehn Betriebe liegen im Landkreis (LK) Vechta, zwölf im LK
Emsland, acht im LK Cloppenburg, sechs im LK Diepholz, vier im LK Osnabrück und
je einer in den LK Aurich und Grafschaft Bentheim. Sieben Betriebe arbeiten im ge-
schlossenen System, das Sauenhaltung und Mast umfasst, die übrigen 43 sind reine
Mäster. Die Betriebsgrößen reichen von 120 Mastplätzen bis 4400 Mastplätzen, der
Median beträgt 715 (siehe Abbildung 6).
4.1.1 Beschreibung der Ergebnisse auf Probenebene
In den Betrieben wurden insgesamt 700 Sammelkot- und Umgebungsproben (ent-
sprechend 14 Proben pro Betrieb, genaue Beschreibung unter 3.2.3.2) entnommen
und untersucht.
Tabelle 7 Ergebnisse der kulturellen Untersuchung
Proben Campylobacter spp. Y. enterocolitica Salmonella spp.
(n) (%) (n) (%) (n) (%)
Positiv 175 25 60 8,6 51 7,3
Negativ 525 75 640 91,4 649 92,7
Gesamt 700 100 700 100 700 100
In den Proben wurde am häufigsten Campylobacter (C.) spp. nachgewiesen (25 %),
während Yersinia (Y.) enterocolitica und Salmonella (S.) spp. mit 8,6 bzw. 7,3 %
deutlich seltener gefunden wurden (Tabelle 7).

Ergebnisse
83
Tabelle 8 Anzahl positiver Proben für alle Erreger, aufgeteilt nach QS-Salmonellen-Kategorie
Positive Proben Kat. I Kat. III gesamt Chi-Quadrat
(n) (%) (n) (%) (n) p-Wert
Campylobacter spp. 109 31,1 66 18,9 175 0,0002
Y. enterocolitica 45 12,9 15 4,3 60 <0,0001
Salmonella spp. 21 6,0 30 8,6 51 0,1906
allgemein (≥ 1 Erreger) 141 40,3 94 26,9 235 0,0002
In der QS-Salmonellen-Kategorie I war der Anteil positiver Nachweise von Campy-
lobacter spp. und Y. enterocolitica signifikant höher als in Kategorie III. Die Nach-
weisrate für Salmonella spp. lag in Proben aus Betrieben der Kategorie III nur gering-
fügig höher als in Kategorie 1 (Tabelle 8).
4.1.2 Beschreibung der Ergebnisse auf Lokalisationsebene
Die 700 Proben verteilten sich auf 4 Lokalisationsgruppen: Es wurden 205 Sammel-
kotproben untersucht. Aus der tiernahen Umgebung wurden 260 Proben untersucht,
dazu zählten Spielketten, Tränken, Futtertröge, Buchtenzwischen- und Buchtenau-
ßenwände. Die tierferne Umgebung wurde insgesamt 170 Mal beprobt, dies beinhal-
tete Lüftung, Rohrleitungen, Abteil- und Zentralgänge, Treibwege, Treibbretter, Stie-
fel, Fenster und Arbeitsräume. Schädlinge in Form von Nagerkot, Oberflächentupfern
von Schadnagern und Fliegen wurden 65 Mal untersucht (Tabelle 9).
Tabelle 9 Positive Proben nach Probenlokalisationsgruppe und Erreger
entnomme-ne Proben
davon positiv für
Campylobacter spp. Y. enterocolitica Salmonella spp.
Lokalisationsgruppe (n) (n) (%) (n) (%) (n) (%)
Sammelkot 205 78 38,05 35 17,07 23 11,22
tiernahe Umgebung 260 85 32,69 21 8,08 20 7,69
tierferne Umgebung 170 9 5,29 2 1,18 7 4,12
Schädlinge 65 3 4,62 2 3,08 1 1,54
gesamt 700 175 25,00 60 8,57 51 7,29
Bei Campylobacter spp. waren über ein Drittel der Sammelkotproben positiv, dies
waren 45 % aller campylobacter-positiven Proben. Aus der tiernahen Umgebung wa-

Ergebnisse
84
ren fast ebenso viele Proben positiv, dies war sogar fast die Hälfte aller campylobac-
ter-positiven Proben. Der Anteil der positiven Proben der tierfernen Umgebung be-
trug 5 %, ebenso wie der Anteil an Campylobacter-Nachweisen. Schädlinge waren
zu 4,6 % positiv, dies machte 1,7 % aller campylobacter-positiven Proben aus. Bei
den Vergleichen von Sammelkot als Referenzgruppe mit je einer anderen Gruppe
ergab sich ein signifikanter Unterschied zu tierferner Umgebung und Schädlingen
(Fisher-Test: p < 0,0001). Auch die Nachweishäufigkeiten von tiernaher und tierfer-
ner Umgebung wichen signifikant voneinander ab (Fisher-Test: p < 0,0001).
Auch bei Y. enterocolitica war Sammelkot mit 17 % die am häufigsten positive Grup-
pe, sein Anteil an allen Yersinia-positiven Proben war knapp 60 %. Aus der tiernahen
Umgebung, zu 8,1 % positiv, stammte ein Drittel aller Yersinia-positiven Proben, tier-
ferne Umgebung und Schädlinge waren mit je 3 % vertreten. Es bestand ein signifi-
kanter Unterschied von Sammelkot zu tiernaher Umgebung, zu Schädlingen (Fisher-
Test jeweils p = 0,003) und zu tierferner Umgebung (Fisher-Test: p = 0,0001). Auch
tiernahe und tierferne Umgebung wichen voneinander ab (Fisher-Test: p = 0,002)
Bei Salmonella spp. waren noch weniger Sammelkotproben positiv (11 %), knapp die
Hälfte aller Salmonella-positiven Proben. Die tiernahe Umgebung war zu 7,7 % posi-
tiv, somit war der Anteil an allen Salmonella-positiven Proben beinahe 40 %. Aus der
tierfernen Umgebung waren 4 % der Proben positiv, diese Gruppe hatte damit im
Vergleich zu den anderen Erregern einen höheren Anteil an allen Salmonella-
positiven Proben (13,7 %). Aus Schädlingen wurden Salmonella spp. dagegen nur
einmal isoliert. Sowohl tierferne Umgebung als auch Schädlinge unterschieden sich
signifikant von Sammelkot (Fisher-Test: p = 0,013).
Bei den paarweisen Vergleichen der positiven Proben pro Erreger innerhalb einer
Gruppe ergaben sich nur signifikante Unterschiede bei Sammelkotproben und tier-
naher Umgebung. Diese bestanden jeweils zwischen Campylobacter spp. und Y.
enterocolitica sowie Campylobacter spp. und Salmonella spp. (Fisher-Test: alle
p < 0,0001).
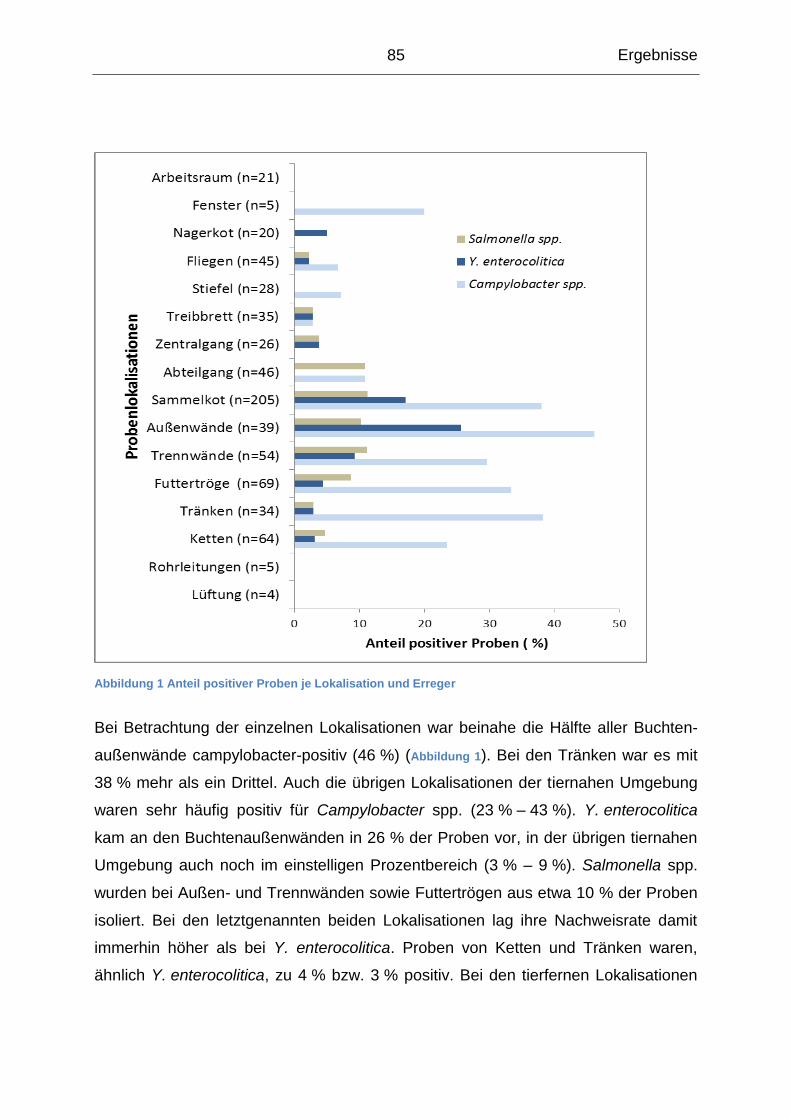
Ergebnisse
85
Abbildung 1 Anteil positiver Proben je Lokalisation und Erreger
Bei Betrachtung der einzelnen Lokalisationen war beinahe die Hälfte aller Buchten-
außenwände campylobacter-positiv (46 %) (Abbildung 1). Bei den Tränken war es mit
38 % mehr als ein Drittel. Auch die übrigen Lokalisationen der tiernahen Umgebung
waren sehr häufig positiv für Campylobacter spp. (23 % – 43 %). Y. enterocolitica
kam an den Buchtenaußenwänden in 26 % der Proben vor, in der übrigen tiernahen
Umgebung auch noch im einstelligen Prozentbereich (3 % – 9 %). Salmonella spp.
wurden bei Außen- und Trennwänden sowie Futtertrögen aus etwa 10 % der Proben
isoliert. Bei den letztgenannten beiden Lokalisationen lag ihre Nachweisrate damit
immerhin höher als bei Y. enterocolitica. Proben von Ketten und Tränken waren,
ähnlich Y. enterocolitica, zu 4 % bzw. 3 % positiv. Bei den tierfernen Lokalisationen

Ergebnisse
86
waren die Unterschiede nicht signifikant. Die Nachweisraten lagen fast alle unter
10 %. Während in Zentralgängen Y. enterocolitica und Salmonella spp. nachgewie-
sen wurde, gelang in den Abteilgängen der Nachweis von Salmonella spp. und
Campylobacter spp. Dieser wurde sogar an Stiefeln nachgewiesen, in Nagerkot dafür
Y. enterocolitica. Von Treibbrettern und Fliegen gelang bei allen drei Erregern der
Nachweis, aus dem Arbeitsraum bei keinem.
Ein Vergleich der Nachweisraten an den einzelnen Lokalisationen und Gruppen zwi-
schen Kategorie I und III zeigte für Campylobacter spp. eine im Allgemeinen höhere
Nachweisrate in Proben aus Beständen der Kategorie I (Tabelle 10). Bei tiernaher
Umgebung und Ketten war dieser Unterschied statistisch signifikant. Y. enterocolitica
kam in der tiernahen Umgebung und den zugehörigen Lokalisationen tendenziell
häufiger in Kategorie I vor, Signifikanz war jedoch nur bei Sammelkot mit höherer
Nachweisrate in Kategorie I gegeben. Salmonella spp. wurden bei den meisten Loka-
lisationen und -gruppen häufiger aus Proben der Kategorie-III-Betriebe isoliert; dieser
Unterschied war jedoch statistisch nicht signifikant.
Tabelle 10 Vergleich der Nachweise zwischen Proben aus Kat. I und Kat. III-Betrieben (inkl. p-Werte des Fisher-Exakt-Tests) für ausgewählte Lokalisationen und -gruppen je Erreger
Anzahl Nachweise je nach Kategorie
Campylobacter spp. Y. enterocolitica Salmonella spp.
Lokalisation (-gruppe)
Kat. I (n)
Kat. III (n)
p-Wert
Kat. I (n)
Kat. III (n)
p-Wert
Kat. I (n)
Kat. III (n)
p-Wert
Sammelkot 44 34 0,196 27 8 0,001 9 14 0,277
tiernahe Umgebung 56 29 0,012 15 6 0,114 9 11 0,485
tierferne Umgebung 6 3 0,304 2 0 0,209 2 5 0,458
Schädlinge 3 0 0,075 1 1 1 1 0 0,431
Außenwände 9 9 1 6 4 0,716 2 2 1
Trennwände 9 7 0,766 5 0 0,051 2 4 0,669
Futtertröge 14 9 0,309 2 1 1 3 3 1
Tränken 11 2 0,075 1 0 1 0 1 0,353
Ketten 13 2 0,015 1 1 1 2 1 1
Gang 4 1 0,346 0 0 - 2 3 1
Stiefel 2 0 0,482 0 0 - 0 0 -
Treibbrett 0 1 0,486 1 0 1 0 1 0,486
Fliegen 3 0 0,080 1 0 0,444 1 0 0,444
Nagerkot 0 0 - 0 1 1 0 0 -
Zentralgang 0 0 - 1 0 0,461 0 1 1

Ergebnisse
87
4.1.3 Beschreibung der Probenergebnisse auf Betriebsebene
4.1.3.1 Allgemeine Darstellung
Ein Betrieb wurde als positiv eingestuft, wenn aus mindestens einer in ihm entnom-
menen Probe ein Erregernachweis erfolgte. Etwa die Hälfte aller Betriebe war positiv
für Y. enterocolitica (Tabelle 11). Im Fisher-Exakt-Test wurde ein signifikanter Unter-
schied zwischen dem Anteil positiver Betriebe in Kategorie I und dem Anteil positiver
Betriebe in Kat. III festgestellt. In einem Drittel der Betriebe wurden Salmonella spp.
nachgewiesen. Der Anteil positiver Betriebe war in Kategorie III geringfügig höher als
in Kategorie I.
Tabelle 11 Vergleich positiver Betriebe nach Erreger und Kategorie
Anzahl positiver Betriebe
Kat. I Kat. III gesamt Fisher-Test
Erreger (n) (%) (n) (%) (n) (%) p-Wert
Campylobacter spp. 22 88,0 18 72,0 40 80,0 0,2890
Y. enterocolitica 19 76,0 5 20,0 24 48,0 <0,0001
Salmonella spp. 7 28,0 9 36,0 16 32,0 0,7624
allgemein (≥ 1 Erreger) 24 92,3 23 88,5 27 90,4
Dagegen waren 80 % der Betriebe positiv für Campylobacter spp., die Abweichun-
gen zwischen den Kategorien waren nicht signifikant.
Die campylobacter-positiven Betriebe wurden zusätzlich in „schwach positiv“ (1 – 4
Proben) und „hoch positiv“ (ab fünf Proben positiv)unterteilt (Tabelle 12).
Tabelle 12 Darstellung schwach und hoch campylobacter-positiver Betriebe nach Kategorie
Anzahl Campylobacter-positiver Betriebe
Kat. I Kat. III gesamt
(n) (%) (n) (%) (n) (%)
negativ 3 12,0 7 28,0 10 20,0
schwach pos. (1 - 4 Proben) 10 40,0 13 52,0 23 46,0
hoch pos. (≥ 5 Proben) 12 48,0 5 20,0 17 34,0
positiv insgesamt 22 88,0 18 72,0 40 80,0

Ergebnisse
88
Bei dem Vergleich der lediglich „hoch positiven“ Betriebe mit den negativen wurde
ein beinahe signifikanter Unterschied zwischen den beiden Kategorien festgestellt (p-
Wert = 0,0568 im Fisher-Exakt-Test): Während in Kategorie I zwölf Betriebe hoch
positiv waren, wurden in Kategorie III nur fünf Betriebe als hoch positiv eingestuft.
Auch die Anzahlen positiver Erregernachweise je Betrieb unterscheiden sich zwi-
schen den einzelnen Erregern.
Bei den positiven Betrieben war die mittlere Anzahl positiver Proben für Campylobac-
ter spp. am höchsten und für Y. enterocolitica am niedrigsten (Tabelle 13, Abbildung 2).
Die Lage der Verteilungen unterschied sich bei den paarweisen Erregervergleichen
im Wilcoxon-Test jedoch nicht signifikant.
Tabelle 13 Ausgewählte statistische Kenngrößen und Vergleich der positiven Proben im positiven Betrieb der einzelnen Erreger
Positive Proben im positiven Betrieb
Erreger Betriebe
(n) arithm. Mittelwert Median Std.-
Abweich. Min. Max.
Campylobacter spp. 40 4,375 4 2,993 1 12
Y. enterocolitica 24 2,500 2 1,285 1 5
Salmonella spp. 16 3,188 2,5 2,400 1 9
Abbildung 2 Box-Plot der Anzahlen positiver Proben je Betrieb
0 1
3
6
12
0 0 0
2
5
0 0 0 1
2
0
5
10
Positiv
e P
robe
n
Y. enterocolitica Salmonella spp. Campylobacter spp.
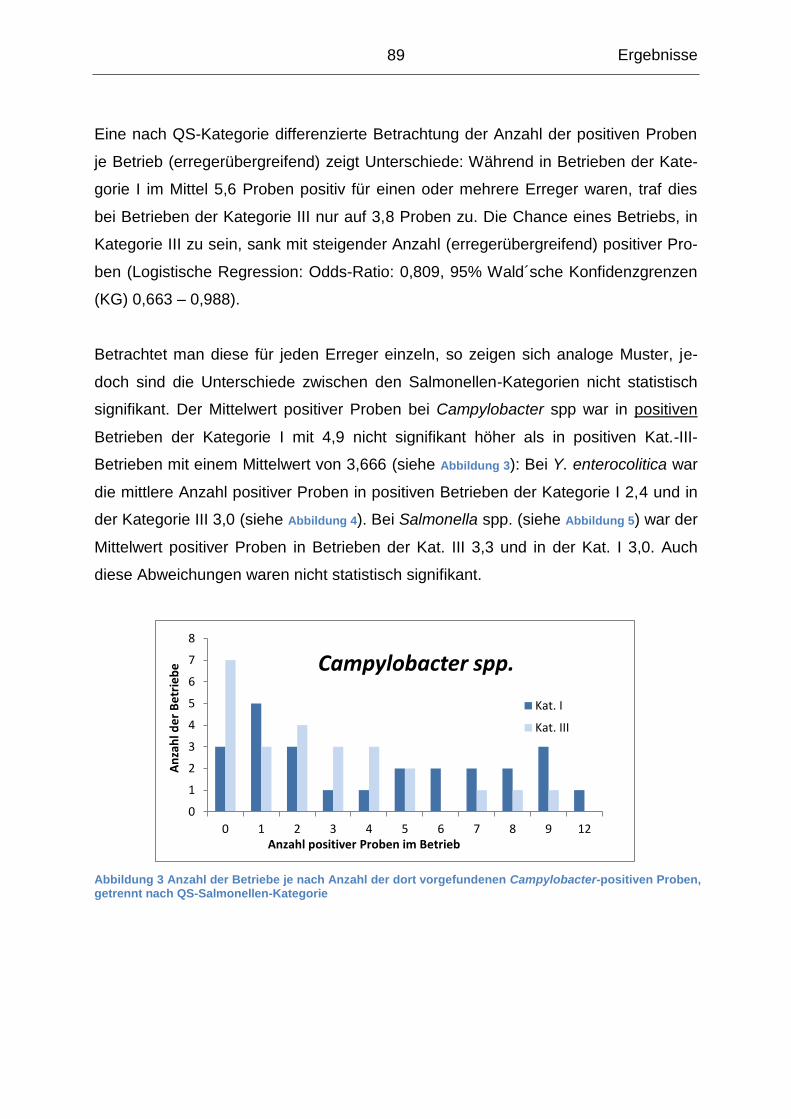
Ergebnisse
89
Eine nach QS-Kategorie differenzierte Betrachtung der Anzahl der positiven Proben
je Betrieb (erregerübergreifend) zeigt Unterschiede: Während in Betrieben der Kate-
gorie I im Mittel 5,6 Proben positiv für einen oder mehrere Erreger waren, traf dies
bei Betrieben der Kategorie III nur auf 3,8 Proben zu. Die Chance eines Betriebs, in
Kategorie III zu sein, sank mit steigender Anzahl (erregerübergreifend) positiver Pro-
ben (Logistische Regression: Odds-Ratio: 0,809, 95% Wald´sche Konfidenzgrenzen
(KG) 0,663 – 0,988).
Betrachtet man diese für jeden Erreger einzeln, so zeigen sich analoge Muster, je-
doch sind die Unterschiede zwischen den Salmonellen-Kategorien nicht statistisch
signifikant. Der Mittelwert positiver Proben bei Campylobacter spp war in positiven
Betrieben der Kategorie I mit 4,9 nicht signifikant höher als in positiven Kat.-III-
Betrieben mit einem Mittelwert von 3,666 (siehe Abbildung 3): Bei Y. enterocolitica war
die mittlere Anzahl positiver Proben in positiven Betrieben der Kategorie I 2,4 und in
der Kategorie III 3,0 (siehe Abbildung 4). Bei Salmonella spp. (siehe Abbildung 5) war der
Mittelwert positiver Proben in Betrieben der Kat. III 3,3 und in der Kat. I 3,0. Auch
diese Abweichungen waren nicht statistisch signifikant.
Abbildung 3 Anzahl der Betriebe je nach Anzahl der dort vorgefundenen Campylobacter-positiven Proben, getrennt nach QS-Salmonellen-Kategorie
0
1
2
3
4
5
6
7
8
0 1 2 3 4 5 6 7 8 9 12
An
zah
l de
r B
etr
ieb
e
Anzahl positiver Proben im Betrieb
Campylobacter spp.
Kat. I
Kat. III
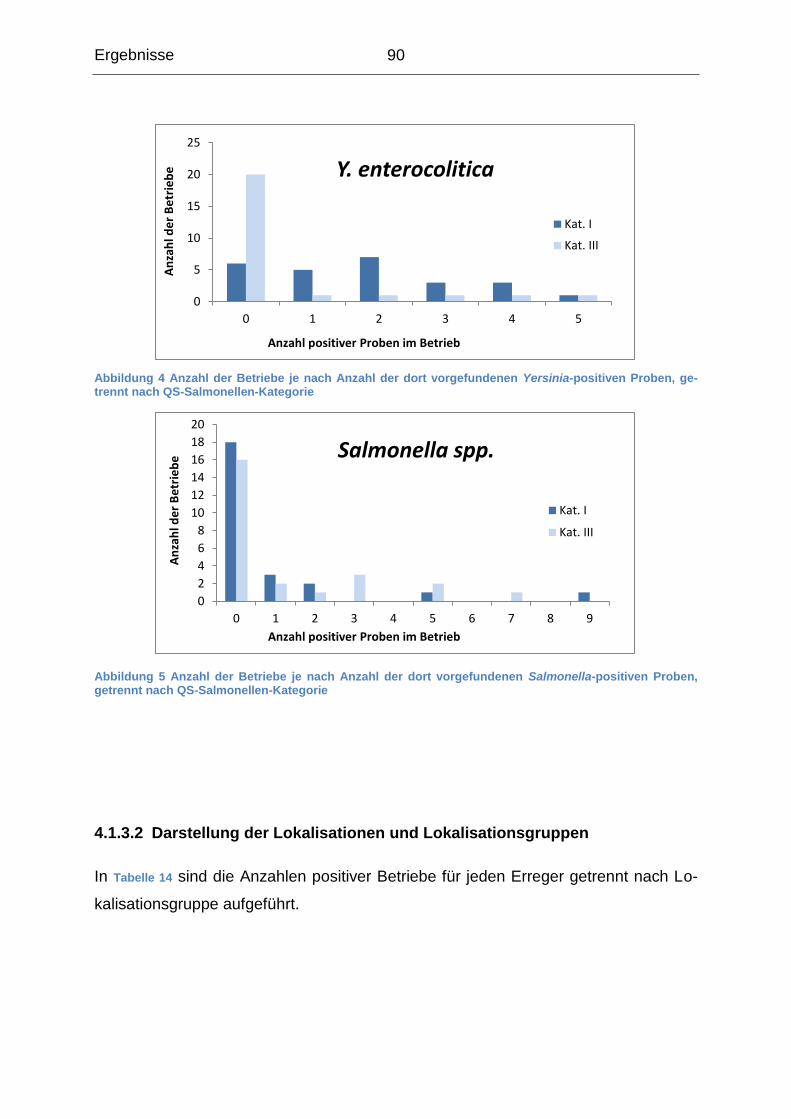
Ergebnisse
90
Abbildung 4 Anzahl der Betriebe je nach Anzahl der dort vorgefundenen Yersinia-positiven Proben, ge-trennt nach QS-Salmonellen-Kategorie
Abbildung 5 Anzahl der Betriebe je nach Anzahl der dort vorgefundenen Salmonella-positiven Proben, getrennt nach QS-Salmonellen-Kategorie
4.1.3.2 Darstellung der Lokalisationen und Lokalisationsgruppen
In Tabelle 14 sind die Anzahlen positiver Betriebe für jeden Erreger getrennt nach Lo-
kalisationsgruppe aufgeführt.
0
5
10
15
20
25
0 1 2 3 4 5
An
zah
l de
r B
etr
ieb
e
Anzahl positiver Proben im Betrieb
Y. enterocolitica
Kat. I
Kat. III
0
2
4
6
8
10
12
14
16
18
20
0 1 2 3 4 5 6 7 8 9
An
zah
l de
r B
etr
ieb
e
Anzahl positiver Proben im Betrieb
Salmonella spp.
Kat. I
Kat. III

Ergebnisse
91
Tabelle 14 Anzahl positiver Betriebe für jede Lokalisationsgruppe und jeden Erreger
untersuchte Betriebe
positive Betriebe
Campylobacter spp. Y. enterocolitica Salmonella spp.
Probengruppe (n) (n) (%) (n) (%) (n) (%)
Sammelkot 50 32 64,00 21 42,00 13 26,00
tiernahe Umgebung 50 31 62,00 15 30,00 10 20,00
tierferne Umgebung 48 8 16,67 2 4,17 6 12,50
Schädlinge 42 3 7,14 2 4,76 1 2,38
Bei jeweils über 60 % der Betriebe wurde Campylobacter spp. in Sammelkot bzw.
tiernaher Umgebung nachgewiesen.
Bei Überprüfung der Übereinstimmung der beiden Gruppen in einem Betrieb wurden
24 Betriebe gezählt, in denen Sammelkot und tiernahe Umgebung gleichzeitig positiv
waren. Dies ergab einen Kappa-Index von 0,358. In einer logistischen Regression
wurde die Chance eines Betriebs, positive Proben in der tierfernen Umgebung zu
aufzuweisen, je nach Anzahl positiver Sammelkotproben, überprüft. Sie erhöhte sich
bei steigender Anzahl positiver Sammelkotproben signifikant (OR: 4,714, KG: 1,364
– 16,295).
In der tierfernen Umgebung wurde bei acht Betrieben Campylobacter spp. nachge-
wiesen. Bei Analyse der Übereinstimmung mit anderen Gruppen zeigte sich, dass
diese Betriebe allesamt ebenfalls positive Nachweise in tiernaher Umgebung und
Sammelkot aufwiesen.
In drei Betrieben gelang der Nachweis aus Schädlingen, jeweils zwei davon waren
auch gleichzeitig positiv in Sammelkot bzw. tiernaher Umgebung, keiner davon in der
tierfernen Umgebung.
Auch bei Y. enterocolitica gelang bei den meisten Betrieben (21) der Nachweis aus
Sammelkot, bei einem Drittel in der tiernahen Umgebung. Bei 13 Betrieben wurden
Erreger in diesen beiden Gruppen gleichzeitig gefunden (Kappa-Index = 0,572; Odds
Ratio: 21,937, KG: 4,069 – 118,272).

Ergebnisse
92
Nachweise aus tierferner Umgebung und Schädlingen erfolgten in jeweils zwei Be-
trieben. Die Prüfung der Übereinstimmung der Gruppen ergab jedoch, dass es sich
nicht um dieselben Betriebe handelte. Die in der tierfernen Umgebung positiven Be-
triebe wiesen beide auch erregerhaltigen Sammelkot auf, von den beiden Betrieben
mit positiven Schädlingen war dies nur bei einem der Fall. Dagegen war sowohl bei
den „tierfern“ als auch den bei Schädlingen positiv getesteten Betrieben nur je einer
auch positiv in der tiernahen Umgebung.
Bei einem Viertel aller Betriebe wurden Salmonella spp. im Sammelkot nachgewie-
sen, bei 20 % in der tiernahen Umgebung. Beide Gruppen gleichzeitig positiv waren
in sieben Betrieben (Kappa-Index = 0,4944; Odds Ratio: 13,222, KG: 2,651 – 65,951).
Bei sechs Betrieben erfolgte der Nachweis in der tierfernen Umgebung Alle davon
hatten ebenfalls im tiernahen Bereich positive Nachweise, und vier davon auch im
Sammelkot. In dem einen Betrieb mit Nachweis von Salmonella spp. in Schädlingen
wurde der Erreger auch in allen anderen Lokalisationsgruppen nachgewiesen.
Des Weiteren wurde untersucht, ob ein Erregervorkommen in einer bestimmten Lo-
kalisationsgruppe einen Einfluss auf die QS-Kategorie der Betriebe hat. So wurde für
die Chance eines Betriebs, in Kategorie III zu sein bei steigender Anzahl Yersinia-
positiver Sammelkotproben, ein Odds Ratio von 0,37 (KG 0,171 - 0,799) errechnet.
Analog betrug das Odds Ratio bei steigender Anzahl positiver Umgebungsproben für
Campylobacter spp. 0,776 (KG 0,629 – 0,958) sowie für Y. enterocolitica 0,440 (KG
0,261 – 0,742).
4.1.4 Gemeinsames Auftreten der Erreger
4.1.4.1 Probenweise Darstellung
In der probenweisen Auswertung konnte in 235 Proben mindestens einer der unter-
suchten Erreger nachgewiesen werden. Davon wiesen 186 genau einen Erreger auf,

Ergebnisse
93
in 47 wurden zwei Erreger gleichzeitig nachgewiesen, und in zwei Proben wurden
alle drei Erreger detektiert. Sowohl einfach als auch doppelt positive Proben wurden
häufiger in Kategorie I festgestellt als in Kategorie III (Tabelle 15).
Tabelle 15 Anzahl verschiedener Erreger je Probe, getrennt nach QS-Kategorie
Anzahl Erreger je Probe nach Kategorie
Kategorie
Summe I III Anzahl Erreger (n) (Spalten-%) (n) (Spalten-%) (n)
0 209 59,71 256 73,1 465 1 108 30,9 78 22,3 186 2 32 9,1 15 4,3 47 3 1 10,3 1 0,3 2
Summe 350 350 700
Bei den doppelt positiven Proben wurde 28 Mal die Kombination Campylobacter spp.
und Y. enterocolitica nachgewiesen, 16 Mal Campylobacter spp. und Salmonella
spp., und dreimal trat Y. enterocolitica gemeinsam mit Salmonella spp. auf (Tabelle 16).
Zur Überprüfung des Grades der Übereinstimmung der Probenergebnisse für die
einzelnen Erreger wurde der Kappa-Index ermittelt. Die nur seltene Übereinstim-
mung der Kulturergebnisse von Proben ergab niedrige Kappa-Koeffizienten von
0,0123 für Y. enterocolitica und Salmonella spp., 0,0524 für Campylobacter spp. und
Salmonella spp. und 0,14638 für Campylobacter spp. und Y. enterocolitica.
Bei Betrachtung der Lokalisationen und -gruppen waren die zweifach positiven Pro-
ben häufig aus der tiernahen Umgebung: Ketten und Tränken waren je einmal betrof-
fen, Futtertröge dreimal, Trenn- und Außenwände sogar jeweils siebenmal (13 %
bzw. 18 % aller dort entnommenen Proben repräsentierend) (Tabelle 16). Auch in
Sammelkotproben wurden 27 Mal zwei Erreger nachgewiesen (13 % aller Sammel-
kotproben). In Fliegen wurden einmal zwei verschiedene Erreger nachgewiesen. Alle
drei Erreger gleichzeitig wurden einmal aus Sammelkot und einmal von den Außen-
wänden isoliert. Aus der tierfernen Umgebung dagegen gelang nie der Nachweis von
zwei oder drei Erregern gleichzeitig.
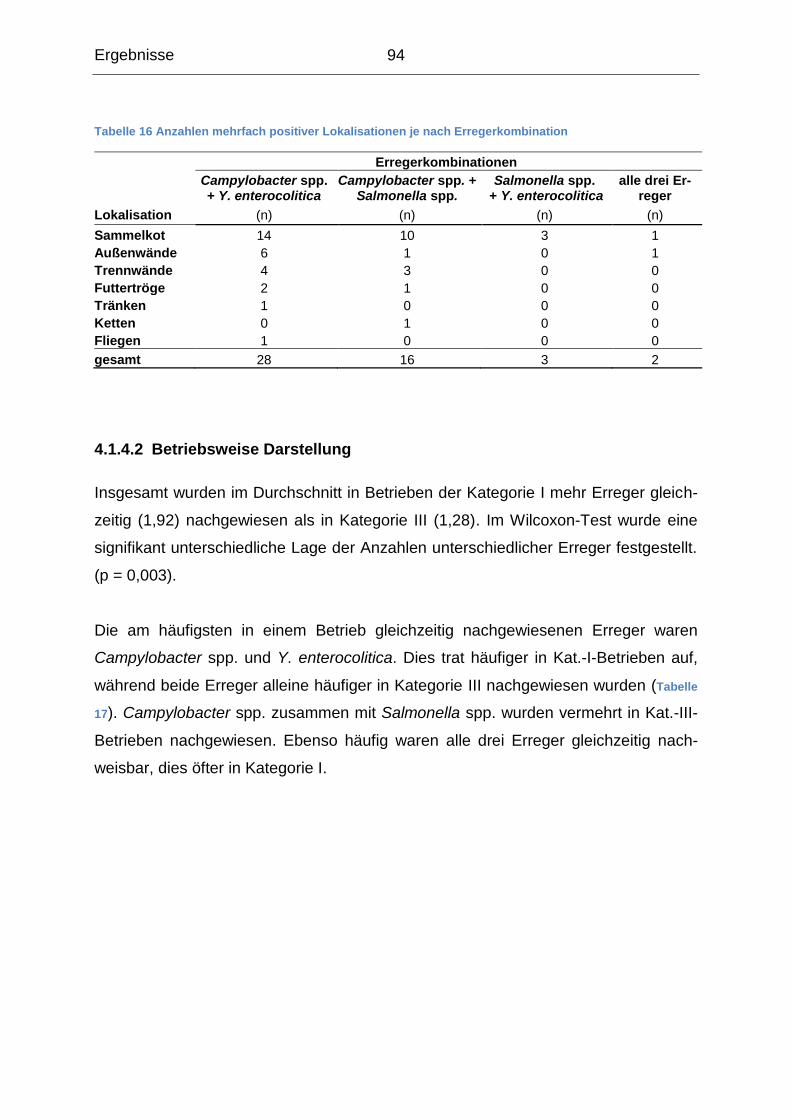
Ergebnisse
94
Tabelle 16 Anzahlen mehrfach positiver Lokalisationen je nach Erregerkombination
Erregerkombinationen
Campylobacter spp. + Y. enterocolitica
Campylobacter spp. + Salmonella spp.
Salmonella spp. + Y. enterocolitica
alle drei Er-reger
Lokalisation (n) (n) (n) (n)
Sammelkot 14 10 3 1
Außenwände 6 1 0 1
Trennwände 4 3 0 0
Futtertröge 2 1 0 0
Tränken 1 0 0 0
Ketten 0 1 0 0
Fliegen 1 0 0 0
gesamt 28 16 3 2
4.1.4.2 Betriebsweise Darstellung
Insgesamt wurden im Durchschnitt in Betrieben der Kategorie I mehr Erreger gleich-
zeitig (1,92) nachgewiesen als in Kategorie III (1,28). Im Wilcoxon-Test wurde eine
signifikant unterschiedliche Lage der Anzahlen unterschiedlicher Erreger festgestellt.
(p = 0,003).
Die am häufigsten in einem Betrieb gleichzeitig nachgewiesenen Erreger waren
Campylobacter spp. und Y. enterocolitica. Dies trat häufiger in Kat.-I-Betrieben auf,
während beide Erreger alleine häufiger in Kategorie III nachgewiesen wurden (Tabelle
17). Campylobacter spp. zusammen mit Salmonella spp. wurden vermehrt in Kat.-III-
Betrieben nachgewiesen. Ebenso häufig waren alle drei Erreger gleichzeitig nach-
weisbar, dies öfter in Kategorie I.
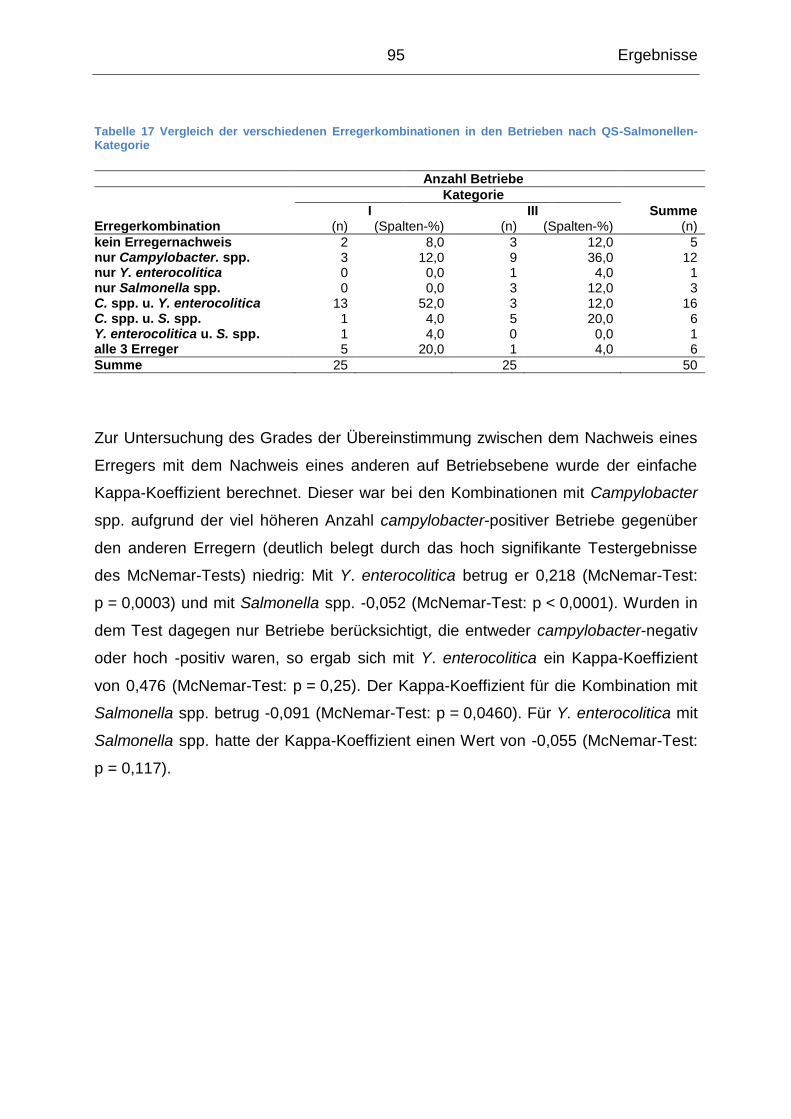
Ergebnisse
95
Tabelle 17 Vergleich der verschiedenen Erregerkombinationen in den Betrieben nach QS-Salmonellen-Kategorie
Anzahl Betriebe
Kategorie
I III Summe Erregerkombination (n) (Spalten-%) (n) (Spalten-%) (n)
kein Erregernachweis 2 8,0 3 12,0 5 nur Campylobacter. spp. 3 12,0 9 36,0 12 nur Y. enterocolitica 0 0,0 1 4,0 1 nur Salmonella spp. 0 0,0 3 12,0 3 C. spp. u. Y. enterocolitica 13 52,0 3 12,0 16 C. spp. u. S. spp. 1 4,0 5 20,0 6 Y. enterocolitica u. S. spp. 1 4,0 0 0,0 1 alle 3 Erreger 5 20,0 1 4,0 6
Summe 25 25 50
Zur Untersuchung des Grades der Übereinstimmung zwischen dem Nachweis eines
Erregers mit dem Nachweis eines anderen auf Betriebsebene wurde der einfache
Kappa-Koeffizient berechnet. Dieser war bei den Kombinationen mit Campylobacter
spp. aufgrund der viel höheren Anzahl campylobacter-positiver Betriebe gegenüber
den anderen Erregern (deutlich belegt durch das hoch signifikante Testergebnisse
des McNemar-Tests) niedrig: Mit Y. enterocolitica betrug er 0,218 (McNemar-Test:
p = 0,0003) und mit Salmonella spp. -0,052 (McNemar-Test: p < 0,0001). Wurden in
dem Test dagegen nur Betriebe berücksichtigt, die entweder campylobacter-negativ
oder hoch -positiv waren, so ergab sich mit Y. enterocolitica ein Kappa-Koeffizient
von 0,476 (McNemar-Test: p = 0,25). Der Kappa-Koeffizient für die Kombination mit
Salmonella spp. betrug -0,091 (McNemar-Test: p = 0,0460). Für Y. enterocolitica mit
Salmonella spp. hatte der Kappa-Koeffizient einen Wert von -0,055 (McNemar-Test:
p = 0,117).

Ergebnisse
96
4.1.5 Risikofaktoren für das Auftreten der Erreger im Betrieb
4.1.5.1 Nominale und dichotome Einflussvariablen
Aus dem Fragebogen wurden verschiedene Merkmale der Fütterungs- und
Haltungsbedingungen in den Betrieben auf einen potentiellen Einfluss auf das
Auftreten der Erreger im Betrieb untersucht. Die nominalen bzw. dichotomen
Risikovariablen sind, für jeden Erreger getrennt, in Tabelle 18 (Campylobacter spp.),
Tabelle 19 (Y. enterocolitica) und Tabelle 20 (Salmonella spp.) dargestellt.
Auftreten von Campylobacter spp. im Betrieb
Die Wahrscheinlichkeiten eines Campylobacter-Nachweises im Betrieb je nach
Ausprägung verschiedener Einflussvariablen sind in Tabelle 18 dargestellt.
Tabelle 18 Campylobacter spp.: Verteilungen Einflussvariablen und Ergebnisse der logistischen Regression
Einflussfaktor Ausprägungen Verteilung
der Betriebe
Anteil positiver Betriebe
OR 95 % Kon-
fidenz-intervall
p-Wert
(n) (%) (%)
Betriebssystem geschlossenes System 7 14,0 71,0 1
reiner Mäster 43 86,0 81,0 1,75 0,286 10,700 0,616
Ferkelherkunft keine Zukäufe 7 14,0 71,0 n.b.
aus einer Herkunft (H.) 33 66,0 94,0 1
mehrere wechselnde H. 10 20,0 40,0 0,533 0,364 0,781 0,001
Generelles Umstallen in der Mast
nein 36 72,0 78,0 1
ja 14 28,0 86,0 1,714 0,316 9,296 0,53
Vereinzeltes Umstallen in der Mast
nein 28 56,0 86,0 1 ja 22 44,0 73,0 0,444 0,108 1,829 0,26
Umgang mit kranken Tieren Bucht 28 56,0 75,0 1
Krankenbucht 19 38,0 84,0 1,778 0,396 7,974 0,45
Krankenstall 3 6,0 100,0 n.b.
Restbestände bei Neueinstel-lung
nein 26 52,0 69,0 1 ja 24 48,0 92,0 4,889 0,920 25,97 0,06
Zusammenstallen versch. Altersgruppen
nein 44 88,0 80,0 1
ja 6 12,0 83,0 1,286 0,133 12,43 0,82
Bodenart Vollspalten 37 74,0 89,0 1
Teilspalten 13 26,0 54,0 0,141 0,031 0,637 0,011
Oberfläche der Außenwände glatt 6 12,0 67,0 1
viele Risse, Unebenheiten 44 88,0 82,0 1,500 0,591 3,806 0,39
Oberfläche der Trennwände glatt 29 60,4 86,0 1
viele Risse, Unebenheiten 19 39,6 74,0 0,669 0,321 1,395 0,28
Zeitpunkt der Gülleentleerung nur nach Ausstallung 8 16,0 75,0 1
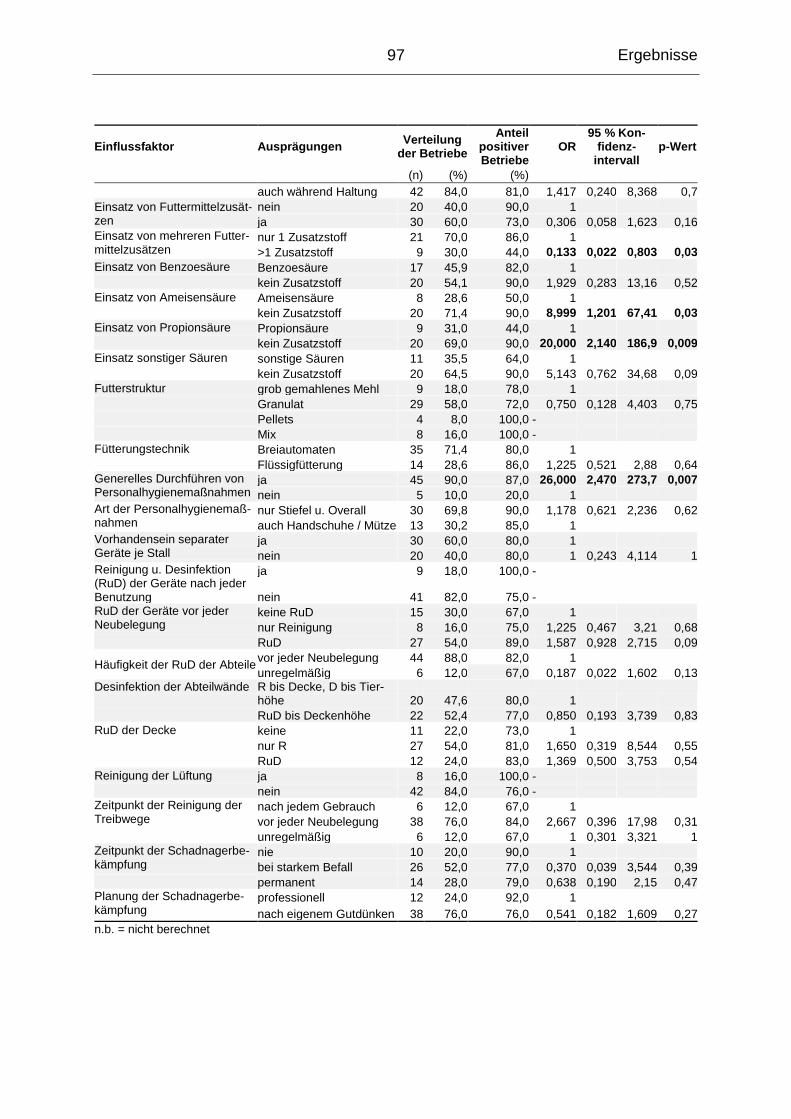
Ergebnisse
97
Einflussfaktor Ausprägungen Verteilung
der Betriebe
Anteil positiver Betriebe
OR 95 % Kon-
fidenz-intervall
p-Wert
(n) (%) (%)
auch während Haltung 42 84,0 81,0 1,417 0,240 8,368 0,7
Einsatz von Futtermittelzusät-zen
nein 20 40,0 90,0 1
ja 30 60,0 73,0 0,306 0,058 1,623 0,16 Einsatz von mehreren Futter-mittelzusätzen
nur 1 Zusatzstoff 21 70,0 86,0 1 >1 Zusatzstoff 9 30,0 44,0 0,133 0,022 0,803 0,03
Einsatz von Benzoesäure Benzoesäure 17 45,9 82,0 1 kein Zusatzstoff 20 54,1 90,0 1,929 0,283 13,16 0,52
Einsatz von Ameisensäure Ameisensäure 8 28,6 50,0 1
kein Zusatzstoff 20 71,4 90,0 8,999 1,201 67,41 0,03
Einsatz von Propionsäure Propionsäure 9 31,0 44,0 1
kein Zusatzstoff 20 69,0 90,0 20,000 2,140 186,9 0,009
Einsatz sonstiger Säuren sonstige Säuren 11 35,5 64,0 1
kein Zusatzstoff 20 64,5 90,0 5,143 0,762 34,68 0,09
Futterstruktur grob gemahlenes Mehl 9 18,0 78,0 1
Granulat 29 58,0 72,0 0,750 0,128 4,403 0,75
Pellets 4 8,0 100,0 -
Mix 8 16,0 100,0 -
Fütterungstechnik Breiautomaten 35 71,4 80,0 1
Flüssigfütterung 14 28,6 86,0 1,225 0,521 2,88 0,64
Generelles Durchführen von Personalhygienemaßnahmen
ja 45 90,0 87,0 26,000 2,470 273,7 0,007
nein 5 10,0 20,0 1 Art der Personalhygienemaß-nahmen
nur Stiefel u. Overall 30 69,8 90,0 1,178 0,621 2,236 0,62
auch Handschuhe / Mütze 13 30,2 85,0 1 Vorhandensein separater
Geräte je Stall ja 30 60,0 80,0 1
nein 20 40,0 80,0 1 0,243 4,114 1 Reinigung u. Desinfektion (RuD) der Geräte nach jeder Benutzung
ja 9 18,0 100,0 -
nein 41 82,0 75,0 - RuD der Geräte vor jeder
Neubelegung keine RuD 15 30,0 67,0 1
nur Reinigung 8 16,0 75,0 1,225 0,467 3,21 0,68
RuD 27 54,0 89,0 1,587 0,928 2,715 0,09
Häufigkeit der RuD der Abteile vor jeder Neubelegung 44 88,0 82,0 1
unregelmäßig 6 12,0 67,0 0,187 0,022 1,602 0,13 Desinfektion der Abteilwände R bis Decke, D bis Tier-
höhe 20 47,6 80,0 1
RuD bis Deckenhöhe 22 52,4 77,0 0,850 0,193 3,739 0,83
RuD der Decke keine 11 22,0 73,0 1
nur R 27 54,0 81,0 1,650 0,319 8,544 0,55
RuD 12 24,0 83,0 1,369 0,500 3,753 0,54
Reinigung der Lüftung ja 8 16,0 100,0 -
nein 42 84,0 76,0 - Zeitpunkt der Reinigung der Treibwege
nach jedem Gebrauch 6 12,0 67,0 1 vor jeder Neubelegung 38 76,0 84,0 2,667 0,396 17,98 0,31
unregelmäßig 6 12,0 67,0 1 0,301 3,321 1
Zeitpunkt der Schadnagerbe-kämpfung
nie 10 20,0 90,0 1
bei starkem Befall 26 52,0 77,0 0,370 0,039 3,544 0,39
permanent 14 28,0 79,0 0,638 0,190 2,15 0,47 Planung der Schadnagerbe-kämpfung
professionell 12 24,0 92,0 1 nach eigenem Gutdünken 38 76,0 76,0 0,541 0,182 1,609 0,27
n.b. = nicht berechnet

Ergebnisse
98
Die Wahrscheinlichkeit, Campylobacter spp. nachzuweisen, war für Betriebe mit
mehreren, wechselnden Ferkelherkünften geringer als in Betrieben, die ihre Ferkel
stets aus dem selben Herkunftsbetrieb bezogen (Tabelle 18).
Für Betriebe mit Teilspaltenboden war die Wahrscheinlichkeit eines Campylobacter-
Nachweises geringer als bei Betrieben mit Vollspaltenboden. Auch die Anzahlen
positiver Proben im Betrieb unterschieden sich. Die mittlere Anzahl positiver Proben
betrug bei Vollspaltenboden 4,162 im Gegensatz zu 1,615 bei Teilspaltenboden. Die
Lage der Verteilungen der Merkmalsausprägungen stellte sich im Wilcoxon-Test
signifikant verschieden dar (p-Wert = 0,0053).
Betriebe, die mehr als einen Futtermittelzusatzstoff einsetzten, wiesen eine
geringere Wahrscheinlichkeit für Campylobacter spp. auf als Betriebe, die nur einen
Futtermittelzusatzstoff einsetzten. Die mittlere Anzahl positiver Proben war in diesen
Betrieben geringer (1,667 gegenüber 4,143). Auch die Lage Merkmalsausprägungen
auf der Zahlengeraden unterschied sich signifikant (Wilcoxon-Test: p = 0,046)
Wurde dem Futter Ameisensäure zugesetzt, war die Chance eines Betriebes, den
Erreger aufzuweisen, geringer als ohne diesen Zusatzstoff. Das gleiche galt für
Propionsäure.
In Betrieben, die Personalhygienemaßnahmen durchführten (Anziehen von Stiefeln
und / oder Schutzkleidung), war die Chance eines Campylobacter-Nachweises im
Betrieb höher als in Betrieben, die keine derartigen Maßnahmen realisierten.
Hinsichtlich der Futterstruktur wurden Unterschiede in den Anzahlen positiver
Proben je Betrieb festgestellt. Die Mittelwerte positiver Proben im Betrieb waren für
Eigenmischung 7,6, Pellets 6,3, grob gemahlenes Mehl 3,0 und für Granulat 2,1. Im
Kruskal-Wallis-Test zum Vergleich der Lage aller vier Ausprägungen ergab sich ein
p-Wert von 0,0006. Bei den paarweisen Vergleichen unterschieden sich unter
Wahrung der versuchsbezogenen Irrtumswahscheinlichkeit (α = 0,05 / 6 Vergleiche)

Ergebnisse
99
Betriebe mit Verfütterung von Granulat signifinant von solchen mit Pelletfütterung
und von Betrieben mit Eigenmischung (Wilcoxon-Tests: p = 0,0040 und p = 0,0004).
Auftreten von Y. enterocolitica im Betrieb
Die Wahrscheinlichkeiten eines Yersinia-Nachweises im Betrieb je nach Ausprägung
verschiedener Einflussvariablen sind in Tabelle 19 dargestellt.
Tabelle 19 Y. enterocolitica: Verteilungen Einflussvariablen und Ergebnisse der logistischen Regression
Einflussfaktor Ausprägungen Verteilung
der Betriebe
Anteil positiver Betriebe
OR 95 % Kon-
fidenz-intervall
p-Wert
(n) (%) (%)
Betriebssystem geschlossenes System 7 14,0 26,0 1
reiner Mäster 43 66,0 51,0 2,619 0,475 15,000 0,42
Ferkelherkunft keine Zukäufe 7 14,0 26,0 n.b.
aus einer Herkunft (H.) 33 66,0 61,0 1
mehrere wechselnde H. 10 20,0 20,0 0,695 0,495 0,977 0,04
Generelles Umstallen in der Mast
nein 36 72,0 50,0 1 ja 14 28,0 43,0 0,75 0,216 2,602 0,65
Vereinzeltes Umstallen in der Mast
nein 28 56,0 61,0 1 ja 22 44,0 32,0 0,302 0,093 0,978 0,046
Umgang mit kranken Tieren Bucht 28 56,0 46,0 1
Krankenbucht 19 38,0 47,0 1,038 0,323 3,337 0,95
Krankenstall 3 6,0 67,0 n.b.
Restbestände bei Neuein-stellung
nein 26 52,0 42,0 1 ja 24 48,0 54,0 1,612 0,527 4,93 0,4
Zusammenstallen versch. Altersgruppen
nein 44 88,0 50,0 1 ja 6 12,0 33,0 0,500 0,083 3,017 0,45
Bodenart Vollspalten 37 74,0 54,0 1
Teilspalten 13 26,0 31,0 0,378 0,099 1,448 0,16
Oberfläche der Außenwände glatt 6 12,0 33,0 1
viele Risse, Unebenhei-ten
44 88,0 50,0 1,414 0,576 3,473 0,45
Oberfläche der Trennwände glatt 29 60,4 52,0 1
viele Risse, Unebenhei-ten
19 39,6 37,0 0,738 0,408 1,333 0,31
Zeitpunkt der Gülleentlee-rung
nur nach Ausstallung 8 16,0 38,0 1
auch während Haltung 42 84,0 50,0 1,667 0,352 7,883 0,52
Einsatz von Futtermittelzu-sätzen
nein 20 40,0 60,0 1
ja 30 60,0 40,0 0,444 0,14 1,411 0,17
Einsatz von mehreren Fut-termittelzusätzen
nur 1 Zusatzstoff 21 70,0 52,0 1
>1 Zusatzstoff 9 30,0 11,0 0,114 0,012 1,076 0,058
Einsatz von Benzoesäure kein Zusatzstoff 20 54,1 60,0 1 Benzoesäure, 17 45,9 41,0 0,467 0,125 1,741 0,26
Einsatz von Ameisensäure Ameisensäure 8 28,6 13,0 1
kein Zusatzstoff 20 71,4 60,0 10,500 1,076 102,5 0,04
Einsatz von Propionsäure Propionsäure 9 31,0 33,0 1
kein Zusatzstoff 20 69,0 60,0 5,999 0,598 60,14 0,13
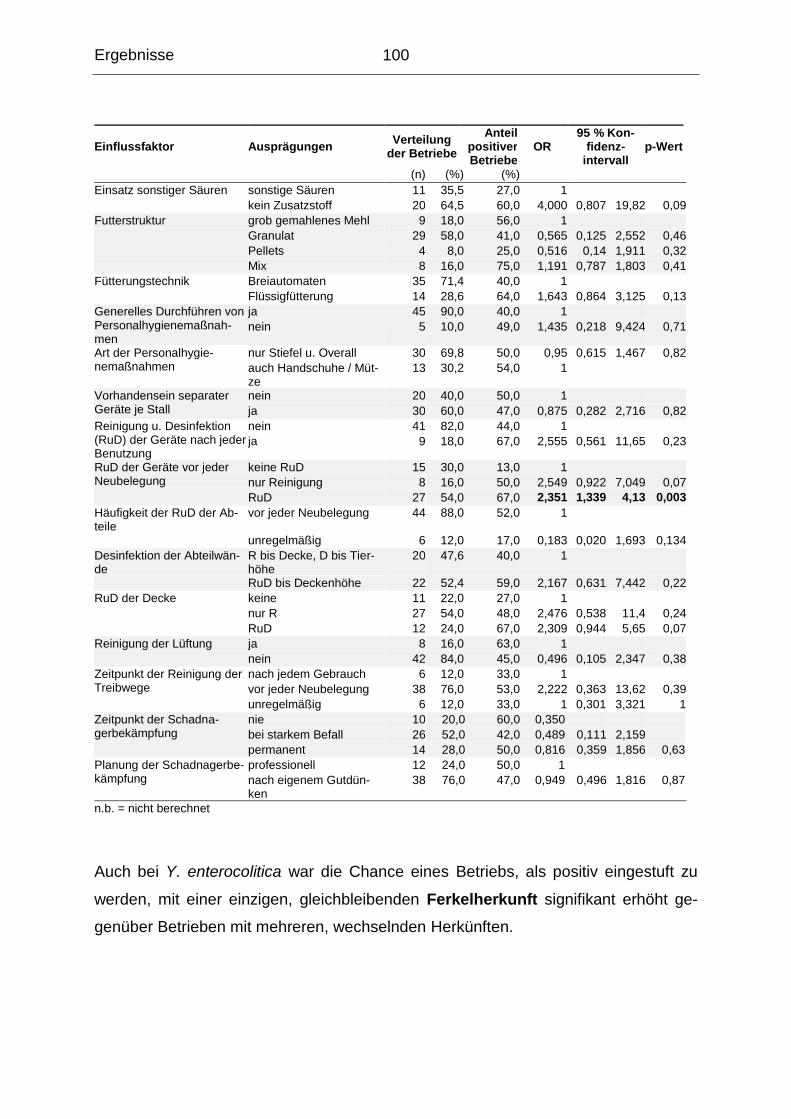
Ergebnisse
100
Einflussfaktor Ausprägungen Verteilung
der Betriebe
Anteil positiver Betriebe
OR 95 % Kon-
fidenz-intervall
p-Wert
(n) (%) (%)
Einsatz sonstiger Säuren sonstige Säuren 11 35,5 27,0 1
kein Zusatzstoff 20 64,5 60,0 4,000 0,807 19,82 0,09
Futterstruktur grob gemahlenes Mehl 9 18,0 56,0 1
Granulat 29 58,0 41,0 0,565 0,125 2,552 0,46
Pellets 4 8,0 25,0 0,516 0,14 1,911 0,32
Mix 8 16,0 75,0 1,191 0,787 1,803 0,41
Fütterungstechnik Breiautomaten 35 71,4 40,0 1
Flüssigfütterung 14 28,6 64,0 1,643 0,864 3,125 0,13
Generelles Durchführen von Personalhygienemaßnah-men
ja 45 90,0 40,0 1
nein 5 10,0 49,0 1,435 0,218 9,424 0,71
Art der Personalhygie-nemaßnahmen
nur Stiefel u. Overall 30 69,8 50,0 0,95 0,615 1,467 0,82
auch Handschuhe / Müt-ze
13 30,2 54,0 1
Vorhandensein separater Geräte je Stall
nein 20 40,0 50,0 1 ja 30 60,0 47,0 0,875 0,282 2,716 0,82
Reinigung u. Desinfektion (RuD) der Geräte nach jeder Benutzung
nein 41 82,0 44,0 1 ja 9 18,0 67,0 2,555 0,561 11,65 0,23
RuD der Geräte vor jeder Neubelegung
keine RuD 15 30,0 13,0 1
nur Reinigung 8 16,0 50,0 2,549 0,922 7,049 0,07
RuD 27 54,0 67,0 2,351 1,339 4,13 0,003
Häufigkeit der RuD der Ab-teile
vor jeder Neubelegung 44 88,0 52,0 1
unregelmäßig 6 12,0 17,0 0,183 0,020 1,693 0,134
Desinfektion der Abteilwän-de
R bis Decke, D bis Tier-höhe
20 47,6 40,0 1
RuD bis Deckenhöhe 22 52,4 59,0 2,167 0,631 7,442 0,22
RuD der Decke keine 11 22,0 27,0 1
nur R 27 54,0 48,0 2,476 0,538 11,4 0,24
RuD 12 24,0 67,0 2,309 0,944 5,65 0,07
Reinigung der Lüftung ja 8 16,0 63,0 1
nein 42 84,0 45,0 0,496 0,105 2,347 0,38
Zeitpunkt der Reinigung der Treibwege
nach jedem Gebrauch 6 12,0 33,0 1
vor jeder Neubelegung 38 76,0 53,0 2,222 0,363 13,62 0,39
unregelmäßig 6 12,0 33,0 1 0,301 3,321 1
Zeitpunkt der Schadna-gerbekämpfung
nie 10 20,0 60,0 0,350
bei starkem Befall 26 52,0 42,0 0,489 0,111 2,159
permanent 14 28,0 50,0 0,816 0,359 1,856 0,63
Planung der Schadnagerbe-kämpfung
professionell 12 24,0 50,0 1
nach eigenem Gutdün-ken
38 76,0 47,0 0,949 0,496 1,816 0,87
n.b. = nicht berechnet
Auch bei Y. enterocolitica war die Chance eines Betriebs, als positiv eingestuft zu
werden, mit einer einzigen, gleichbleibenden Ferkelherkunft signifikant erhöht ge-
genüber Betrieben mit mehreren, wechselnden Herkünften.

Ergebnisse
101
In Betrieben, die vereinzelt Tiere von einer Mastgruppe in eine andere umstallten,
war die Wahrscheinlichkeit eines Erregernachweises geringer als in Betrieben, die
nie Tiere umstallten.
Betriebe, die Ameisensäure im Futter einsetzten, wiesen eine geringere Wahr-
scheinlichkeit, yersinia-positiv zu sein, auf als Betriebe ohne Ameisensäure im Futter.
Die Anzahlen positiver Proben im Betrieb unterschieden sich ebenso. Betriebe mit
Ameisensäureeinsatz hatten niedrigere Mittelwerte positiver Proben (1,4 zu 0,125).
Der Wilcoxon-Test zur Überprüfung der gleichen Lage der Verteilungen war ebenfalls
signifikant: p = 0,0182.
In Betrieben, die im Stall verwendete Geräte vor Neubelegung stets reinigten und
desinfizierten, lag eine höhere Wahrscheinlichkeit mindestens einer positiven Probe
vor, als in Betrieben, die Geräte vor Neubelegung weder reinigten noch desinfizierten.
Der Mittelwert positiver Proben im Betrieb betrug bei Reinigung und Desinfektion
(RuD) 1,6667, bei Reinigung ohne Desinfektion 1,3750, und wenn nichts erfolgte,
0,2667. Die Lage der Merkmalsverteilungen unterschied sich im Wilcoxon-Test zwi-
schen RuD und Verzicht auf RuD: p = 0,0016).
Bei Einsatz von mehreren Futtermittelzusatzstoffen wies ein Betrieb eine geringe-
re Anzahl positiver Proben auf als bei Einsatz nur eines Zusatzstoffes. Der Mittelwert
war bei letzteren signifikant höher (1,4762) als bei ersteren (0,1111). Im Wilcoxon-
Test wich die Lage der Verteilungen signifikant voneinander ab (p = 0,027).
Auftreten von Salmonella spp. im Betrieb
Die Wahrscheinlichkeiten eines Salmonella-Nachweises im Betrieb je nach
Ausprägung verschiedener Einflussvariablen sind in Tabelle 20 dargestellt.

Ergebnisse
102
Tabelle 20 Salmonella spp.: Verteilungen Einflussvariablen und Ergebnisse der logistischen Regression
Einflussfaktor Ausprägungen Verteilung der Betrie-
be
Anteil positiver Betriebe
OR 95 % Kon-
fidenz- intervall
p-Wert
(n) (%) (%) Betriebssystem geschlossenes System 7 14,0 29,0 1
reiner Mäster 43 86,0 32,0 1,207 0,208 7,012 1
Ferkelherkunft keine Zukäufe 7 14,0 29,0 n.b.
aus einer Herkunft (H.) 33 66,0 33,0 1
mehrere wechselnde H. 10 20,0 30,0 0,970 0,713 1,318 0,84
Generelles Umstallen in der Mast
ja 14 28,0 43,0 1,950 0,539 7,052 0,31
nein 36 72,0 28,0 1
Vereinzeltes Umstallen in der Mast
ja 22 44,0 32,0 0,985 0,297 3,263 0,98
nein 28 56,0 32,0 1
Umgang mit kranken Tieren Bucht 28 56,0 39,0 1
Krankenbucht 19 38,0 21,0 0,412 0,108 1,572 0,19
Krankenstall 3 6,0 33,0 n.b.
Restbestände bei Neueinstel-lung
ja 24 48,0 25,0 0,533 0,158 1,799 0,31
nein 26 52,0 38,0 1
Zusammenstallen versch. Al-tersgruppen
ja 6 12,0 33,0 1,071 0,175 6,56 0,94
nein 44 88,0 32,0 1
Bodenart Vollspalten 37 74,0 30,0 1
Teilspalten 13 26,0 38,0 1,477 0,394 5,537 0,56
Oberfläche der Außenwände glatt 6 12,0 33,0 1
viele Risse, Unebenheiten 44 88,0 32,0 0,966 0,390 2,391 0,94
Oberfläche der Trennwände glatt 29 60,4 24,0 1
viele Risse , Unebenhei-ten
19 39,6 42,0 1,512 0,811 2,819 0,19
Zeitpunkt der Gülleentleerung nur nach Ausstallung 8 16,0 25,0 1
auch während Haltung 42 84,0 33,0 1,500 0,267 8,41 0,64
Einsatz von Futtermittelzusätzen nein 20 40,0 30,0 1
ja 30 60,0 33,0 1,167 0,344 3,956 0,8
Einsatz von mehreren Futtermit-telzusätzen
nur 1 Zusatzstoff 21 70,0 29,0 1
>1 Zusatzstoff 9 30,0 44,0 2,000 0,396 10,108 0,4
Einsatz von Benzoesäure Benzoesäure, 17 45,9 29,0 0,972 0,236 4,003 0,97
kein Zusatzstoff 20 54,1 30,0 1
Einsatz von Ameisensäure Ameisensäure 8 28,6 50,0 1
kein Zusatzstoff 20 71,4 30,0 0,429 0,08 2,308 0,32
Einsatz von Propionsäure Propionsäure 9 31,0 55,6 1
kein Zusatzstoff 20 69,0 30,0 0,188 0,027 1,304 0,09
Einsatz sonstiger Säuren sonstige Säuren 11 35,5 36,0 1
kein Zusatzstoff 20 64,5 30,0 0,750 0,158 3,558 0,72
Futterstruktur grob gemahlenes Mehl 9 18,0 44,0 1
Granulat 29 58,0 28,0 0,476 0,101 2,236 0,38
Pellets 4 8,0 75,0 1,936 0,523 7,168 0,32
Mix 8 16,0 13,0 0,709 0,432 1,162 0,17
Fütterungstechnik Breiautomaten 35 71,4 31,0 1
Flüssigfütterung 14 28,6 29,0 0,934 0,473 1,845 0,84
Generelles Durchführen von Personalhygienemaßnahmen
ja 45 90,0 60,0 1
nein 5 10,0 29,0 0,271 0,04 1,814 0,18
Art der Personalhygienemaß-nahmen
auch Handschuhe / Mütze 13 30,2 31,0 1
nur Stiefel u. Overall 30 69,8 30,0 0,988 0,617 1,582 0,96
Vorhandensein separater Gerä-te je Stall
ja 30 60,0 23,0 0,372 0,11 1,262 0,11
nein 20 40,0 45,0 1

Ergebnisse
103
Einflussfaktor Ausprägungen Verteilung der Betrie-
be
Anteil positiver Betriebe
OR 95 % Kon-
fidenz- intervall
p-Wert
(n) (%) (%) Reinigung u. Desinfektion (RuD) der Geräte nach jeder Benut-zung
nein 41 82,0 32,0 1 ja 9 18,0 33,0 1,077 0,232 4,994 0,92
RuD der Geräte vor jeder Neu-belegung
keine RuD 15 30,0 40,0 1
nur Reinigung 8 16,0 50,0 1,225 0,516 2,907 0,65
RuD 27 54,0 22,0 0,754 0,477 1,192 0,23
Häufigkeit der RuD der Abteile vor jeder Neubelegung 44 88,0 27,0 1
unregelmäßig 6 12,0 67,0 5,333 0,862 32,997 0,07
Desinfektion der Abteilwände R bis Decke, D bis Tier-höhe
20 47,6 35,0 1
RuD bis Deckenhöhe 22 52,4 27,0 0,696 0,187 2,589 0,6
RuD der Decke keine 11 22,0 27,0 1
nur R 27 54,0 37,0 1,569 0,336 7,314 0,57
RuD 12 24,0 25,0 0,943 0,372 2,392 0,9
Reinigung der Lüftung ja 8 16,0 63,0 1
nein 42 84,0 26,0 0,213 0,044 1,042 0,06
Zeitpunkt der Reinigung der Treibwege
nach jedem Gebrauch 6 12,0 33,0 1
vor jeder Neubelegung 38 76,0 29,0 0,815 0,13 5,112 0,83
unregelmäßig 6 12,0 50,0 1,414 0,44 4,54 0,56
Zeitpunkt der Schadnagerbe-kämpfung
nie 10 20,0 20,0 1 1,000
bei starkem Befall 26 52,0 31,0 1,778 0,306 10,324 0,52
permanent 14 28,0 43,0 1,732 0,678 4,426 0,25
Planung der Schadnagerbe-kämpfung
professionell 12 24,0 25,0 1
nach eigenem Gutdünken 38 76,0 34,0 1,249 0,599 2,603 0,55
n.b. = nicht berechnet
In der logistischen Regression konnte für keinen der getesteten Faktoren eine signifi-
kante Erhöhung oder Erniedrigung der Wahrscheinlichkeit eines Salmonella-
Nachweises im Betrieb ermittelt werden.
In Betrieben, die vor jeder Neubelegung eine Reinigung und Desinfektion des
Abteils durchführten, war der Mittelwert positiver Proben kleiner als in Betrieben, die
diese nicht regelmäßig vor Neubelegung durchführten. (0,841 gegenüber 2,333) Im
Wilcoxon-Test wurde eine inhomogene Lage der Verteilungen festgestellt
(p = 0,0397).

Ergebnisse
104
4.1.5.2 Stetige Variablen
Zusätzlich wurden fünf stetige Merkmale untersucht, deren Verteilungen in den fol-
genden Boxplots dargestellt sind (Abbildung 6). Bei der Anzahl der Mastplätze und der
Belegdichte waren unteres und oberes Quartil (Box) etwa gleich groß, daneben la-
gen Ausreißer (einzelne Punkte) nach oben vor, bei letzterer auch nach unten. Bei
der Tierzahl im Abteil lag eine rechtsschiefe Verteilung vor, mit einigen oberen Ext-
remwerten. Bei der Tierzahl in der Bucht und der Buchtengröße waren Box und
„Whiskers“ sehr klein, jedoch mit extremen Ausreißern nach oben.
Abbildung 6 Box-Plots der stetigen Variablen Mastplätze im Bestand, Tierzahl im Abteil, Tierzahl in der Bucht, Buchtengröße (m
2), Belegdichte (m
2 / Tier)
Diese stetigen Merkmale wurden mittels einer linearen Regressionsanalyse auf einen
potentiellen Einfluss auf die Anzahl positiver Proben im Betrieb untersucht. Nach
120 380 715 1000
1584
Mastplätze
1000
2000
4000
3000
40 102 137 210
353
200
400
600
800
Tierzahl im Abteil
8 12 16 25 43
100
200
300
400
Tierzahl in der Bucht
6 9 11 19 32
100
200
300
Buchtengröße in m2
0.62 0.69 0.76 0.82
0.95
0.6
0.8
1.0
1.2
Belegdichte in m2
/Tier
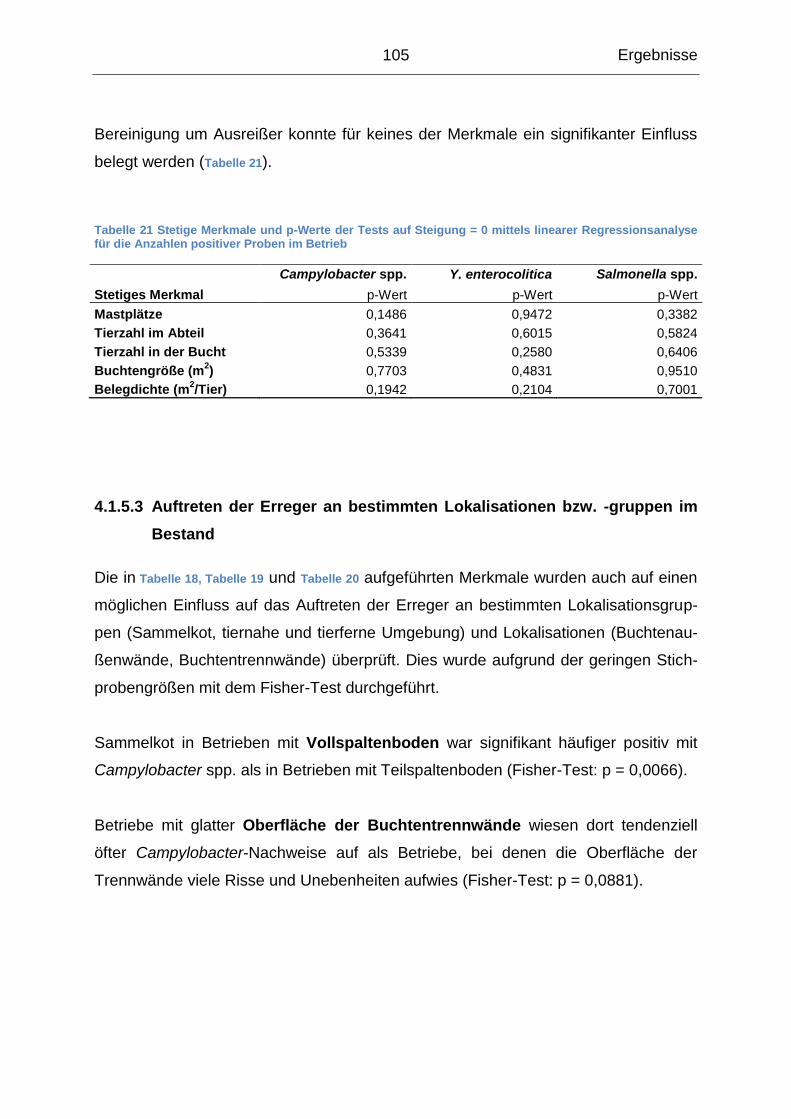
Ergebnisse
105
Bereinigung um Ausreißer konnte für keines der Merkmale ein signifikanter Einfluss
belegt werden (Tabelle 21).
Tabelle 21 Stetige Merkmale und p-Werte der Tests auf Steigung = 0 mittels linearer Regressionsanalyse für die Anzahlen positiver Proben im Betrieb
Campylobacter spp. Y. enterocolitica Salmonella spp.
Stetiges Merkmal p-Wert p-Wert p-Wert
Mastplätze 0,1486 0,9472 0,3382
Tierzahl im Abteil 0,3641 0,6015 0,5824
Tierzahl in der Bucht 0,5339 0,2580 0,6406
Buchtengröße (m2) 0,7703 0,4831 0,9510
Belegdichte (m2/Tier) 0,1942 0,2104 0,7001
4.1.5.3 Auftreten der Erreger an bestimmten Lokalisationen bzw. -gruppen im
Bestand
Die in Tabelle 18, Tabelle 19 und Tabelle 20 aufgeführten Merkmale wurden auch auf einen
möglichen Einfluss auf das Auftreten der Erreger an bestimmten Lokalisationsgrup-
pen (Sammelkot, tiernahe und tierferne Umgebung) und Lokalisationen (Buchtenau-
ßenwände, Buchtentrennwände) überprüft. Dies wurde aufgrund der geringen Stich-
probengrößen mit dem Fisher-Test durchgeführt.
Sammelkot in Betrieben mit Vollspaltenboden war signifikant häufiger positiv mit
Campylobacter spp. als in Betrieben mit Teilspaltenboden (Fisher-Test: p = 0,0066).
Betriebe mit glatter Oberfläche der Buchtentrennwände wiesen dort tendenziell
öfter Campylobacter-Nachweise auf als Betriebe, bei denen die Oberfläche der
Trennwände viele Risse und Unebenheiten aufwies (Fisher-Test: p = 0,0881).

Ergebnisse
106
Aus dem Sammelkot von Betrieben mit Benzoesäure als Futterzusatz wurden ten-
denziell häufiger Campylobacter spp. isoliert als von Betrieben, bei denen andere
Säuren zum Einsatz kamen (Fisher-Test: p = 0,061).
Betriebe, die die Buchtenwände nur bis in Tierhöhe desinfizierten, waren in der
tiernahen Umgebung seltener positiv für Y. enterocolitica als Betriebe, die dies bis in
Deckenhöhe taten (Fisher-Test: 0,0017). Gleiches galt für die Einzellokalisation der
Außenwände (Fisher-Test: p = 0,0207). Dagegen waren Betriebe, die beides bis in
Deckenhöhe durchführten, hier seltener positiv für Salmonella spp. (Fisher-Test:
p = 0,0318).
In der tierfernen Umgebung von Betrieben ohne Schadnagerbekämpfung wurden
häufiger Salmonella spp. nachgewiesen als in Betrieben mit Schadnagerbekämpfung
(Fisher-Test: p = 0,0399).
4.1.5.4 Assoziation sämtlicher Erreger mit Einflussfaktoren
Wie in Abschnitt 3.1.6 erläutert, kann der Zusammenhang der Einflussfaktoren auf
das Auftreten der Erreger auch entgegen der eigentlichen Kausalitätsrichtung model-
liert werden. So wurden verschiedene Betriebsmerkmale wie die Salmonellen-
Kategorie, die Anzahl der Herkünfte, das generelle und das mehrfache Verfüttern
von Futtermittel-Zusätzen, die einzelnen Säurezusätze und die Fütterungstechnik in
Abhängigkeit vom gemeinsamen Auftreten aller drei Erreger in einem logistischen
Regressionsmodell betrachtet.
Betrachtete man die Kategorie als abhängige Variable, so zeigten sich unter Berück-
sichtigung des Auftretens aller Erreger in einem Betrieb signifikante Unterschiede
(p = 0,0080 Wald‟sches Chi-Quadrat). Bezogen auf die einzelnen Erreger konnte
jedoch – analog zu den einfaktoriellen Analysen – nur für das Vorkommen von Y.
enterocolitica ein signifikanter Unterschied festgestellt werden (OR für Kat. III gegen-
über Kat. I: 0,094, KG 0,023 – 0,382).
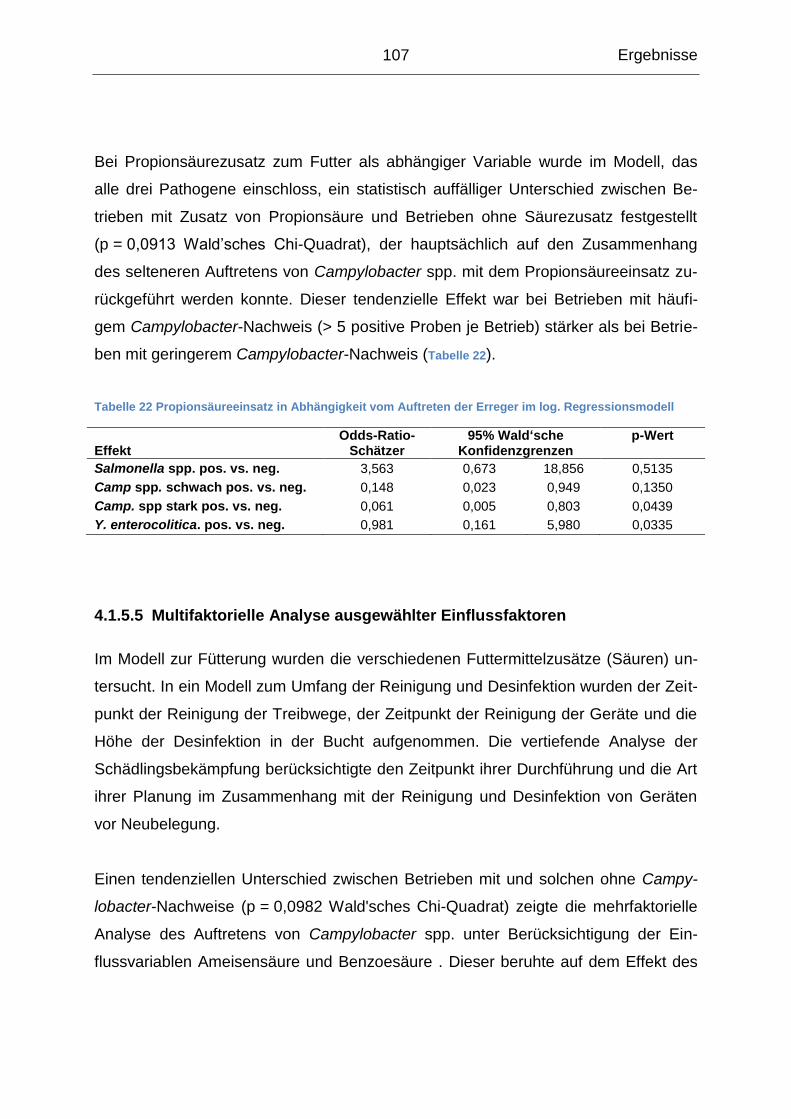
Ergebnisse
107
Bei Propionsäurezusatz zum Futter als abhängiger Variable wurde im Modell, das
alle drei Pathogene einschloss, ein statistisch auffälliger Unterschied zwischen Be-
trieben mit Zusatz von Propionsäure und Betrieben ohne Säurezusatz festgestellt
(p = 0,0913 Wald‟sches Chi-Quadrat), der hauptsächlich auf den Zusammenhang
des selteneren Auftretens von Campylobacter spp. mit dem Propionsäureeinsatz zu-
rückgeführt werden konnte. Dieser tendenzielle Effekt war bei Betrieben mit häufi-
gem Campylobacter-Nachweis (> 5 positive Proben je Betrieb) stärker als bei Betrie-
ben mit geringerem Campylobacter-Nachweis (Tabelle 22).
Tabelle 22 Propionsäureeinsatz in Abhängigkeit vom Auftreten der Erreger im log. Regressionsmodell
Effekt Odds-Ratio-
Schätzer 95% Wald‘sche
Konfidenzgrenzen p-Wert
Salmonella spp. pos. vs. neg. 3,563 0,673 18,856 0,5135
Camp spp. schwach pos. vs. neg. 0,148 0,023 0,949 0,1350
Camp. spp stark pos. vs. neg. 0,061 0,005 0,803 0,0439
Y. enterocolitica. pos. vs. neg. 0,981 0,161 5,980 0,0335
4.1.5.5 Multifaktorielle Analyse ausgewählter Einflussfaktoren
Im Modell zur Fütterung wurden die verschiedenen Futtermittelzusätze (Säuren) un-
tersucht. In ein Modell zum Umfang der Reinigung und Desinfektion wurden der Zeit-
punkt der Reinigung der Treibwege, der Zeitpunkt der Reinigung der Geräte und die
Höhe der Desinfektion in der Bucht aufgenommen. Die vertiefende Analyse der
Schädlingsbekämpfung berücksichtigte den Zeitpunkt ihrer Durchführung und die Art
ihrer Planung im Zusammenhang mit der Reinigung und Desinfektion von Geräten
vor Neubelegung.
Einen tendenziellen Unterschied zwischen Betrieben mit und solchen ohne Campy-
lobacter-Nachweise (p = 0,0982 Wald'sches Chi-Quadrat) zeigte die mehrfaktorielle
Analyse des Auftretens von Campylobacter spp. unter Berücksichtigung der Ein-
flussvariablen Ameisensäure und Benzoesäure . Dieser beruhte auf dem Effekt des

Ergebnisse
108
Ameisensäureeinsatzes (OR: 6,244, KG: 1,145 – 34,060). Da Propionsäure und
sonstige Säuren meist gemeinsam mit Ameisensäure eingesetzt werden, wurden
diese beiden Variablen im Modell nicht berücksichtigt. Der beobachtete Effekt des
Ameisensäureeinsatzes kann jedoch auch aus dem Einsatz von Propionsäure oder
sonstigen Säuren resultieren.
Das Modell, das gleichzeitig das Vorhandensein von eigenen Arbeitsgeräten für je-
den Stall und die Durchführung von Reinigung und Desinfektion der Arbeitsgeräte
vor jeder Neubelegung in Bezug auf Y. enterocolitica im Betrieb berechnete, ergab
einen signifikanten Unterschied (p = 0,0174 Wald´sches Chi-Quadrat). Dieser konnte,
ebenso wie in der einfaktoriellen Analyse, auf den positiven Zusammenhang mit der
Reinigung und Desinfektion der Geräte vor jeder Neubelegung zurückgeführt werden
(OR: 0,145, KG: 0,038 – 0,548).

Ergebnisse
109
4.2 Salmonellenverlaufsuntersuchung
Das Ziel der Untersuchung war, sich wiederholende Muster im Salmonellen- bzw.
Klonvorkommen zu identifizieren. Dafür wurden in zwölf Mastschweine haltenden
Betrieben Proben verschiedener Lokalisationen entnommen und kulturell sowie mo-
lekularbiologisch auf Salmonella enterica untersucht und die Isolate feintypisiert. Von
den Betrieben lagen je fünf in den Landkreisen Vechta und Emsland, zwei befanden
sich im Landkreis Cloppenburg. Unter ihnen waren elf reine Mastbestände und eine
Haltung im geschlossenen System. Weitere Betriebsmerkmale sind in Tabelle 23 dar-
gestellt.
Tabelle 23 Betriebsmerkmale der zwölf Betriebe der Verlaufsuntersuchung
Einflussfaktor Ausprägungen A B C D E F G H I J K L
Anzahl der Mast-schweine
(n) 145 1000 460 500 1100 880 300 850 4400 1250 1350 1350
Tierzahl im Abteil (n) 145 128 353 58 135 80 117 98 95 347 148 199
Tierzahl in der Bucht (n) 13 13 11 12 17 40 28 14 24 11 37 25
Buchtengröße in m² (n) 8 10 9 9 12 40 21 9 16 9 24 19
Belegdichte in m²/Tier (n) 0,62 0,77 0,82 0,75 0,71 0,78 0,75 0,64 0,66 0,82 0,65 0,76
Ferkelherkunft
keine Zukäufe
x
aus einer Herkunft x x
x x
x
x x x
mehrere wechselnde H.
x
x
x
Generelles Umstallen in der Mast
nein x x x x
x x x x x x
ja
x
x
Vereinzeltes Um-stallen in der Mast
nein
x
x
x
x
ja x x
x x x
x
x
x
Umgang mit kranken Tieren
Bucht x
x x
x
x x
Krankenbucht
x
x x
x
Krankenstall
x x
Restbestände bei Neueinstellung
nein x
x x
x
x x
ja
x
x x
x x
x
Zusammenstallen versch. Altersgruppen
nein x x x x x x x x x x x
ja
x
Bodenart Vollspalten
x
x x x x
x
x x
Teilspalten x
x
x
x
Oberfläche der Au-ßenwände
glatt
x
x
viele Risse, Uneben-heiten
x x x x x x x x x
x
Oberfläche der Trennwände
glatt
x
x x x
x x x x
viele Risse , Uneben-heiten
x
x x
x
Zeitpunkt der Gülleent-leerung
nur nach Ausstallung x
x x
auch während Haltung
x x x x x x x x
x
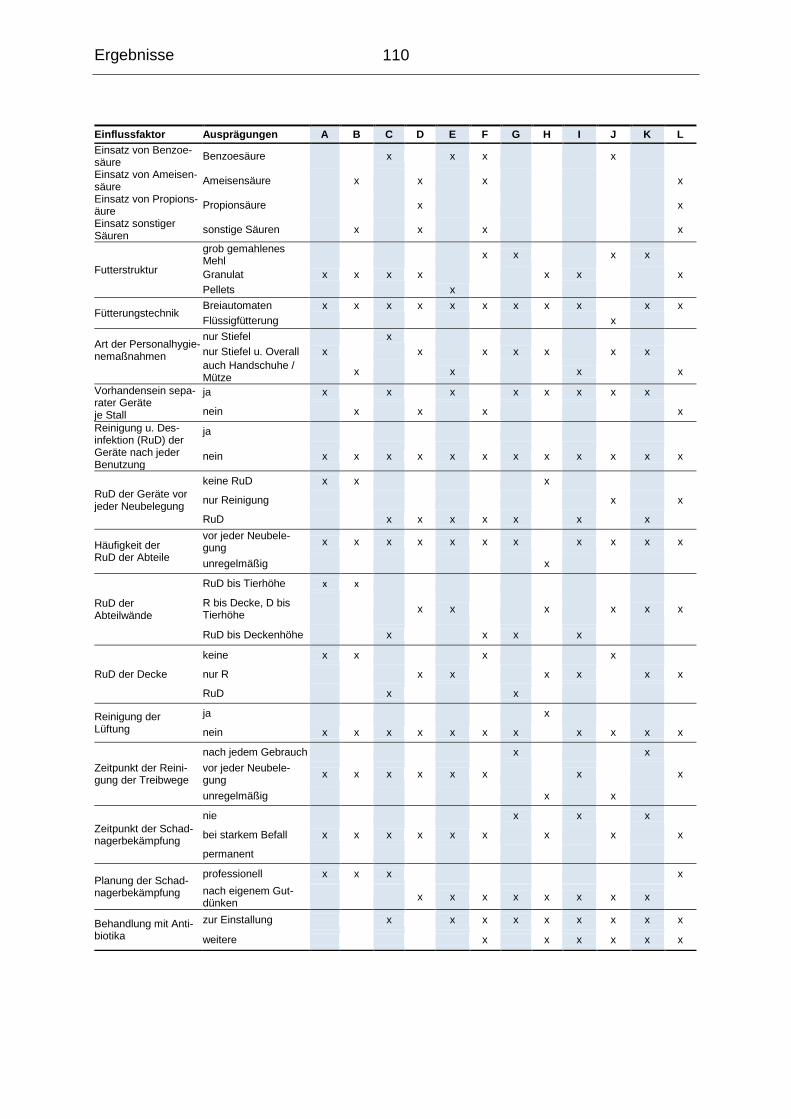
Ergebnisse
110
Einflussfaktor Ausprägungen A B C D E F G H I J K L
Einsatz von Benzoe-säure
Benzoesäure
x
x x
x
Einsatz von Ameisen-säure
Ameisensäure
x
x
x
x
Einsatz von Propions-äure
Propionsäure
x
x
Einsatz sonstiger Säuren
sonstige Säuren
x
x
x
x
Futterstruktur
grob gemahlenes Mehl
x x
x x
Granulat x x x x
x x
x
Pellets
x
Fütterungstechnik Breiautomaten x x x x x x x x x
x x
Flüssigfütterung
x
Art der Personalhygie-nemaßnahmen
nur Stiefel
x
nur Stiefel u. Overall x
x
x x x
x x
auch Handschuhe / Mütze
x
x
x
x
Vorhandensein sepa-rater Geräte je Stall
ja x
x
x
x x x x x
nein
x
x
x
x
Reinigung u. Des-infektion (RuD) der Geräte nach jeder Benutzung
ja
nein x x x x x x x x x x x x
RuD der Geräte vor jeder Neubelegung
keine RuD x x
x
nur Reinigung
x
x
RuD
x x x x x
x
x
Häufigkeit der RuD der Abteile
vor jeder Neubele-gung
x x x x x x x
x x x x
unregelmäßig
x
RuD der Abteilwände
RuD bis Tierhöhe x x
R bis Decke, D bis Tierhöhe
x x
x
x x x
RuD bis Deckenhöhe
x
x x
x
RuD der Decke
keine x x
x
x
nur R
x x
x x
x x
RuD
x
x
Reinigung der Lüftung
ja
x
nein x x x x x x x
x x x x
Zeitpunkt der Reini-gung der Treibwege
nach jedem Gebrauch
x
x
vor jeder Neubele-gung
x x x x x x
x
x
unregelmäßig
x
x
Zeitpunkt der Schad-nagerbekämpfung
nie
x
x
x
bei starkem Befall x x x x x x
x
x
x
permanent
Planung der Schad-nagerbekämpfung
professionell x x x
x
nach eigenem Gut-dünken
x x x x x x x x
Behandlung mit Anti-biotika
zur Einstallung x x x x x x x x x
weitere x x x x x x

Ergebnisse
111
Bei drei Betrieben wurden die Probennahmen nach einem Mastdurchgang aufgrund
fehlender kultureller Nachweise von Salmonella spp. beendet. Ein weiterer Betrieb
wurde zwei Mastdurchgänge lang mit negativem Ergebnis untersucht. In diesen vier
Betrieben wurden insgesamt 262 Proben entnommen. In den übrigen acht Betrieben
wurden in jeweils drei Durchgängen insgesamt 1410 Proben entnommen.
4.2.1 Beschreibung der Ergebnisse auf Probenebene
4.2.1.1 Ergebnisse der kulturellen Untersuchung
In 144 Proben wurden kulturell Salmonella spp. nachgewiesen.
Beinahe alle Isolate gehörten dem Serovar S. Typhimurium an; nur ein Isolat wurde
als Serovar S. Derby klassifiziert, vier Isolate ließen sich keinem Serovar zuordnen.
Je 36 % der Salmonella spp. gehörten den Phagentypen DT 120 und DT 193 an,
11 % waren PTU 302, knapp 6 % zählten zu DT 104. Jeweils 4 % waren PTU 311
bzw. nicht typisierbar und je einmal kamen die Phagentypen DT 170 und DY PT 108
vor.
Die Gesamtzahlen entnommener Proben je Betrieb und die jeweiligen Anteile positi-
ver Proben sind in Tabelle 24 dargestellt.
Tabelle 24 Kulturergebnisse der Proben je Betrieb
Betrieb
Kulturergebnis A B C D E F G H I J K L insg.
positiv (n) 0 1 0 4 6 48 0 12 14 20 0 39 144
(Spalten-%) 0 1,7 0 3,5 5,1 14,0 0 6,5 7,5 10,5 0 18,3 8,6
negativ (n) 62 59 55 111 111 295 59 173 173 170 86 174 1528
(Spalten-%) 100 98,3 100 96,5 94,9 86,0 100 93,5 92,5 89,5 100 81,7 91,4
Summe 62 60 55 115 117 343 59 185 187 190 86 213 1672

Ergebnisse
112
4.2.1.1.1 Betriebsübergreifende Analyse
In alle folgenden Berechnungen wurden nur die Betriebe mit mindestens einem Sal-
monellennachweis einbezogen.
Vergleich der Durchgänge
Die Anteile positiver Proben unterschieden sich zwischen den Durchgängen. Den
größten Anteil positiver Proben verzeichnete mit 15 % der erste Durchgang, den ge-
ringsten mit 7 % der zweite, im dritten waren 9 % aller Proben positiv (Abbildung 7).
Abbildung 7 Darstellung der Probenergebnisse je Durchgang
Vergleich der Zeitpunkte
Die Anteile positiver Proben im Abteil (Probenentnahmezeitpunkte (ZP) 0, 2 - 5, Ab-
bildung 8) ließen im Verlauf des Mastdurchgangs keinen klar gerichteten Trend erken-
nen (Cochran-Armitage-Trend-Test: p = 0,797). Dies galt auch für den Vergleich der
Untersuchungszeitpunkte im belegten Abteil (ZP 2 – 5) (Cochran-Armitage-Trend-
Test: p = 0,345). Nur die Sammelkotproben bei Anlieferung der Ferkel (ZP 1) waren
mit 29,6 % häufiger positiv als die anderen Zeitpunkte. Es konnten bereits Salmonel-
la spp. vor Einstallung der Ferkel im Abteil nachgewiesen werden. Zudem trugen
auch die Ferkel den Erreger in die Mast ein.

Ergebnisse
113
Abbildung 8 Vergleich der Probenergebnisse nach Beprobungszeitpunkt (ohne ZP 1, LKW-Sammelkot)
Um zu untersuchen, wie sich Salmonellenfunde vor Einstallung (im leeren Abteil
(ZP 0) und bei den Ferkeln auf dem Transportfahrzeug (ZP 1)) auf das Erregervor-
kommen während der Mast (ZP 2 bis 5) auswirken, wurden diese in Tabelle 25 gegen-
übergestellt: Dort sind alle Proben aus belegten Abteilen getrennt aufgeführt, je
nachdem, ob im zugehörigen Mastdurchgang und Betrieb schon vor Einstallung
Salmonellennachweise erfolgt waren.
Tabelle 25 Vergleich der Proben aus dem belegten Abteil, je nachdem, ob in dem Betrieb und Durchgang, aus dem sie stammen, ZP 0 und/oder ZP 1 positiv waren
Kulturergebnis
negativ positiv Summe Proben der ZP 2 bis 5 (n) (%) (n) (%)
ZP 0 negativ, ZP 1 negativ 470 95,14 24 4,86 494 ZP 0 positiv, ZP 1 negativ 162 85,71 27 14,29 189 ZP 0 negativ, ZP 1 positiv 31 91,18 3 8,82 34 ZP 0 positiv ZP 1 positiv 301 84,31 56 15,69 357
Summe 964 110 1074
Von den 1074 im belegten Abteil entnommenen Proben entstammten 494 (46 %)
einem Betrieb und Durchgang, in dem vor Einstallung kein Salmonellennachweis er-
folgt war. Ihr Anteil an allen positiven Proben aus belegtem Abteil betrug dagegen
8,41 10,39
11,96 9,6
7,98
0
5
10
15
20
25
30
35
40
45
50
0 (leeresgereinigtes,desinfiziertes Abteil)
2 (4 Wochen nachEinstallung)
3 (8 Wochen nachEinstallung)
4 (12 Wochen nachEinstallung)
5 (16 Wochen nachEinstallung)
An
teil
po
siti
ver
Pro
be
n (
%)
Zeitpunkt
positive Proben

Ergebnisse
114
nur 21,8 %. Bei 189 (17,6 %) der Proben war im zugehörigen Betrieb und Durchgang
der Erreger im leeren Abteil nachgewiesen worden, sie stellten knapp ein Viertel aller
positiven Proben im belegten Abteil. Nur 34 (3,2 %) der Proben stammten aus einem
Betrieb und Durchgang, wo nur der LKW-Sammelkot positiv gewesen war. Der Anteil
der positiven Proben dieser Gruppe an allen positiven Proben betrug 2,7 %. Bei ei-
nem Drittel (357) aller Proben war im gleichen Durchgang und Betrieb zuvor bereits
sowohl das leere Abteil, als auch der LKW-Sammelkot positiv gewesen. Aus dieser
Gruppe rührte die Hälfte aller positiven Proben des belegten Abteils. Für die Chance
einer Probe, Salmonella-positiv zu sein, je nachdem, ob im laufenden Durchgang vor
Einstallung entweder weder das leere Abteil noch der LKW-Sammelkot, nur eines
der beiden oder beide positiv waren, wurde ein Odds Ratio von 2,62 (KG 1,795 –
3,825) ermittelt.
Zusammenfassend zeigten sich fast alle Durchgänge, in denen bereits vor Einstal-
lung Salmonella spp. nachweisbar waren, auch während der Mast positiv: Von den
insgesamt 20 untersuchten Durchgängen wurden bei drei schon im gereinigten und
desinfizierten leeren Abteil Salmonellen nachgewiesen, in einem Durchgang war nur
der LKW-Kot positiv. In sechs Durchgängen wurde Salmonella spp. sowohl im LKW-
Sammelkot als auch im leeren Abteil detektiert, nur einer davon blieb danach ohne
Nachweise. In zehn Durchgängen wurde der Erreger weder im leeren Abteil noch im
LKW-Sammelkot gefunden. Zwei davon blieben auch im weiteren Verlauf negativ.
Vergleich der Lokalisationen
Die am häufigsten positive Probenlokalisation war LKW-Sammelkot, die zweithäu-
figste der Zentralgang, an dritter Stelle folgten Oberflächentupfer von Schadnagern.
An Fenster, Heizung, Decke, Arbeitsraum und sonstigen Arbeitsmitteln wurden da-
gegen nie Salmonella spp. nachgewiesen (Tabelle 26).

Ergebnisse
115
Tabelle 26 Anteil positiver der Kulturergebnisse nach Lokalisation
Kulturergebnis Summe
negativ positiv
Lokalisation n Zeilen-% n Zeilen-% n
LKW-Sammelkot 19 70,4 8 29,6 27 Zentralgang 68 76,4 21 23,6 89 Oberfläche Nager 15 83,3 3 16,7 18 Gang 116 84,7 21 15,3 137 Sammelkot 77 85,6 13 14,4 90 Treibbrett 63 87,5 9 12,5 72 Trennwände 103 88,0 14 12,0 117 Buchtenboden 23 88,5 3 11,5 26 Nagerkot 31 88,6 4 11,4 35 Außenwände 99 89,2 12 10,8 111 Tränken 68 91,9 6 8,1 74 Rohrleitungen 103 92,0 9 8,0 112 Futtertröge 105 92,1 9 7,9 114 Ketten 108 94,7 6 5,3 114 Lüftung 115 96,6 4 3,4 119 Stiefel 38 97,4 1 2,6 39 Fliegen 70 98,6 1 1,4 71 Fenster 14 100,0 0 0,0 14 Heizung 4 100,0 0 0,0 4 Decke 9 100,0 0 0,0 9 Waage 14 100,0 0 0,0 14 Arbeitsraum 2 100,0 0 0,0 2 sonstige Arbeitsmittel 2 100,0 0 0,0 2
Summe 1266 89,8 144 10,2 1410
Bei Betrachtung der Lokalisationsgruppen war Sammelkot mit 18,0 % am häufigsten
positiv, danach folgte die tierferne Umgebung (Lüftung, Rohrleitungen, Gang,
Zentralgang, Treibbrett, Stiefel, Fenster, Heizung, Decke, Waage, Arbeitsraum und
sonstige Arbeitsmittel) mit 10,6 %. Etwas seltener positiv war mit 9,1 % die tiernahe
Umgebung (Ketten, Tränken, Futtertröge, Buchtentrenn- und Buchtenaußenwände),
in Schädlingen (Fliegen, Nagerkot und Oberflächentupfer von Nagern) wurden mit
6,5 % am wenigsten Salmonella spp. nachgewiesen (Chi-Quadrat-Test der globalen
Unabhängigkeitshypothese: p = 0,0598).
Die Darstellung der Probenergebnisse im Verlauf eines Mastdurchgangs zeigte für
Sammelkot eine Tendenz zur Abnahme von der Untersuchung bei Anlieferung bis
zum Untersuchungszeitpunkt 3 (Abbildung 9). Der Trend konnte jedoch nicht statistisch
belegt werden (Cochran-Armitage-Trend-Test: p = 0,099).

Ergebnisse
116
Abbildung 9 Darstellung der Probenergebnisse je Beprobungszeitpunkt für Sammelkot, tiernahe, tierferne Umgebung und Schädlinge
In der tiernahen Umgebung waren im leeren, gereinigten und desinfizierten Abteil
etwa 5 % der Proben positiv, zu den folgenden Zeitpunkten lagen die Anteile mit
Ausnahme des letzten Zeitpunkts höher (Cochran-Armitage-Trend-Test: p = 0,589).
Die tierferne Umgebung unterschied sich kaum zwischen den Zeitpunkten (Cochran-
Armitage-Trend-Test: p = 0,746), hier war der Anteil positiver Proben im leeren Abteil
mit 12,2 % sogar am höchsten. Auch bei den Schädlingen war im Verlauf der Mast
kein Trend nach oben oder unten erkennbar, die Anteile lagen zwischen 3,5 % und
9,7 % (Cochran-Armitage-Trend-Test: p = 0,701).
Bei Vergleich der Lokalisationsgruppen zum ZP 0 unterschieden sich tiernahe und
tierferne Umgebung signifikant (Chi-Quadrat-Test: p = 0,0405) voneinander. Im be-
legten Abteil (ZP 2 – 5) unterschieden sich die Lokalisationsgruppen nicht signifikant.
Die am häufigsten positiven Lokalisationen nach Reinigung und Desinfektion (ZP 0)
waren Treibbretter und Gänge (je viermal), Rohrleitungen, Trennwände, Buchtenbo-
den und Arbeitsräume waren je dreimal positiv. Jeweils einmal wurde der Erreger
0
5
10
15
20
25
30
35
1 2 3 4 5
An
teil
Pro
be
n in
%
Beprobungszeitpunkt
Sammelkot tiernahe Umgebung tierferne Umgebung Schädlinge

Ergebnisse
117
von Lüftungseinrichtungen, Futtertrögen, Außenwänden, und dem Boden des
Zentralgangs isoliert.
4.2.1.1.2 Zwischenbetriebliche Analyse
Die Anzahlen positiver Proben für jeden Betrieb sind in Tabelle 24 dargestellt. Eine in-
tensive Untersuchung der Befunde innerhalb eines Betriebes wurde nur in ausge-
wählten Betrieben vorgenommen, bei denen eine größere Anzahl Salmonella-
Nachweise gelang.
Der Vergleich der Durchgänge von allen Betrieben mit drei Mastdurchgängen zeigte
für die Betriebe F (Cochran-Armitage-Test: p = 0,0003), I (p < 0,0001) und J (p =
0,0008) eine Abnahme der Anteile positiver Proben von Durchgang 1 zu Durchgang
3 (Abbildung 10). Betrieb H blieb über alle Durchgänge auf einem ähnlichen, niedrigen
Nachweisniveau (p = 0,84), in Betrieb L waren in Durchgang 2 weitaus weniger Pro-
ben positiv als in den, mit vielen positiven Proben untersuchten Durchgängen 1 und
3.
Abbildung 10 Darstellung der positiven Proben an den 3 Durchgängen für die Betriebe 6, 8, 9, 10 und 12
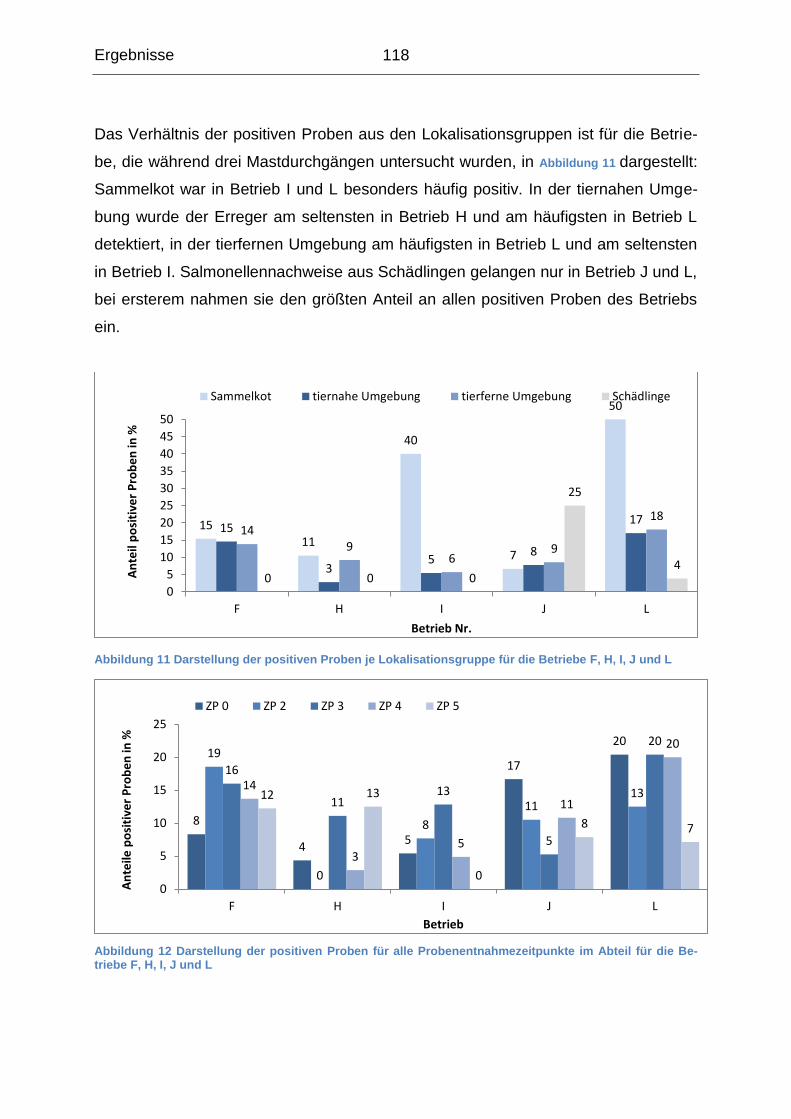
Ergebnisse
118
Das Verhältnis der positiven Proben aus den Lokalisationsgruppen ist für die Betrie-
be, die während drei Mastdurchgängen untersucht wurden, in Abbildung 11 dargestellt:
Sammelkot war in Betrieb I und L besonders häufig positiv. In der tiernahen Umge-
bung wurde der Erreger am seltensten in Betrieb H und am häufigsten in Betrieb L
detektiert, in der tierfernen Umgebung am häufigsten in Betrieb L und am seltensten
in Betrieb I. Salmonellennachweise aus Schädlingen gelangen nur in Betrieb J und L,
bei ersterem nahmen sie den größten Anteil an allen positiven Proben des Betriebs
ein.
Abbildung 11 Darstellung der positiven Proben je Lokalisationsgruppe für die Betriebe F, H, I, J und L
Abbildung 12 Darstellung der positiven Proben für alle Probenentnahmezeitpunkte im Abteil für die Be-triebe F, H, I, J und L
15
11
40
7
50
15
3 5
8
17 14
9 6
9
18
0 0 0
25
4
0
5
10
15
20
25
30
35
40
45
50
F H I J L
An
teil
po
siti
ver
Pro
be
n in
%
Betrieb Nr.
Sammelkot tiernahe Umgebung tierferne Umgebung Schädlinge
8
4 5
17
20 19
0
8
11 13
16
11 13
5
20
14
3 5
11
20
12 13
0
8 7
0
5
10
15
20
25
F H I J L
An
teile
po
siti
ver
Pro
be
n in
%
Betrieb
ZP 0 ZP 2 ZP 3 ZP 4 ZP 5

Ergebnisse
119
Bei Vergleich der einzelnen Probenentnahmezeitpunkte im Abteil waren die Verläufe
innerhalb der Betriebe unterschiedlich (Abbildung 12). Gemeinsam war allen Betrieben,
dass bereits im leeren, gereinigten und desinfizierten Abteil Salmonella spp. nach-
gewiesen wurden, am häufigsten in Betrieb L (20,4 %). Bei keinem Betrieb war ein
signifikanter Unterschied zwischen den Anteilen positiver Proben im leeren Abteil
und im belegten Abteil feststellbar.
Abbildung 13 Anzahlen der LKW-Sammelkotproben (ZP 1) für die Betriebe 6, 8, 9, 10 und 12
In jedem der über drei Mastdurchgänge untersuchten Betriebe war mindestens eine
Sammelkotprobe bei Anlieferung der Ferkel positiv (Abbildung 13).
Bei den Betrieben mit nur geringer Nachweisrate (Betriebe B, D und E, nicht darge-
stellt), waren dagegen ausschließlich Proben im belegten Abteil positiv. Es wurden
hier weder Salmonellen im gereinigten und desinfizierten leeren Abteil, noch im
Sammelkot der Ferkel bei Anlieferung nachgewiesen.

Ergebnisse
120
4.2.1.2 Ergebnisse aus der molekularbiologischen Untersuchung und Ver-
gleich mit den Ergebnissen der kulturellen Untersuchung
Nach der Beschreibung der kulturellen Untersuchungsergebnisse soll kurz auf die
Ergebnisse der molekularbiologischen Untersuchung auf Salmonella spp. (invA-
PCR) eingegangen werden. Diese war der kulturellen Untersuchung zum Zwecke
eines Vor-Screenings auf Salmonella-verdächtige Proben vorgeschaltet. (Anmer-
kung: In den folgenden Berechnungen sind 21 Proben enthalten, die aufgrund ihrer
in der Verlaufsuntersuchung nicht auswertbaren Lokalisation (kein regelmäßiger Zu-
gang zu Futterproben, Umgebung des Flatdecks) aus allen sonstigen Analysen aus-
geschlossen wurden.) Tabelle 27 stellt diese Ergebnisse den kulturellen Ergebnissen
gegenüber.
Tabelle 27 Darstellung der invA-PCR-Ergebnisse, vergleichend mit den Kultur-Ergebnissen
Vergleich PCR- und Kulturergebnis
Kultur
negativ positiv Summe PCR (n) (%) (n) (%) (n) (%)
negativ 1401 82,75 0 0 1401 82,75 schwach positiv 103 6,08 4 0,24 107 6,32 positiv 39 2,30 146 8,62 185 10,93
Summe 1543 91,14 150 8,86 1693 100,00
Alle Proben, die in der PCR negativ getestet wurden, erwiesen sich auch in der Kul-
tur als negativ. Umgekehrt konnte aus etwa einem Fünftel der Proben, in denen Ge-
nomfragmente nachgewiesen worden waren, keine Salmonella isoliert werden. Bei
6 % der Proben lautete das PCR-Ergebnis schwach positiv. Der Großteil davon war
in der Kultur negativ, aus vier Proben gelang jedoch kulturell ein Erregernachweis.
Bei der Mehrzahl der Betriebe (A, B, G und K (fast) keine Nachweise, F, I, J und L
mit hoher Nachweisrate) wichen die Ergebnisse beider Verfahren nicht voneinander
ab. In zwei Betrieben waren deutlich weniger Proben kulturell als molekularbiologisch
positiv (C: 0 % zu 22 %, D:4 % zu 17 %). In zwei weiteren Betrieben wurden in der
PCR überdurchschnittlich viele Proben als schwach positiv ausgewiesen (E: 20 %, H:
15 %).

Ergebnisse
121
Bei Vergleich der einzelnen Lokalisationsgruppen im Hinblick auf die Höhe der Ab-
weichung zwischen PCR- und Kulturergebnis ergaben sich keine signifikanten Unter-
schiede im Fisher-Exakt-Test. Bei Sammelkot waren 27 % der in der PCR positiven
Proben kulturell negativ, bei tiernaher und tierferner Umgebung 21 % und bei den
Schädlingen 20 %. Auch schwach positive PCR-Ergebnisse traten gleichmäßig in
allen vier Gruppen auf: in Sammelkotproben und der tiernahen Umgebung zu 6 %
(drei Proben davon in der Kultur positiv), in der tierfernen Umgebung (eine Probe in
der Kultur positiv) zu 7 % und bei Schädlingen zu 8 %.
Die Probenentnahmezeitpunkte unterschieden sich hinsichtlich der Anteile der Ab-
weichungen zwischen Kultur und PCR ebenfalls nicht signifikant (Fisher-Exakt-Test).
Allerdings waren im belegten Abteil (ZP 2 – 5) mehr Proben (8 %) schwach positiv
als vor Einstallung (ZP 0: 3 %, ZP 1: 0 %).
Zusammenfassend ergibt sich (bei Nichtberücksichtigung der nicht eindeutig zuord-
enbaren schwach positiven Proben) mit 97,6 % konkordanten Paaren eine sehr hohe
Übereinstimmung der beiden Testverfahren. Dies bestätigt der Kappa-Index von 0,87.
Der McNemar-Test mit p < 0,0001 belegt allerdings eine nicht vollständige Identität,
sondern Asymmetrie der beiden Testverfahren, die durch die höhere Anzahl positiver
Proben in der PCR im Vergleich zur Kultur hervorgerufen wird.
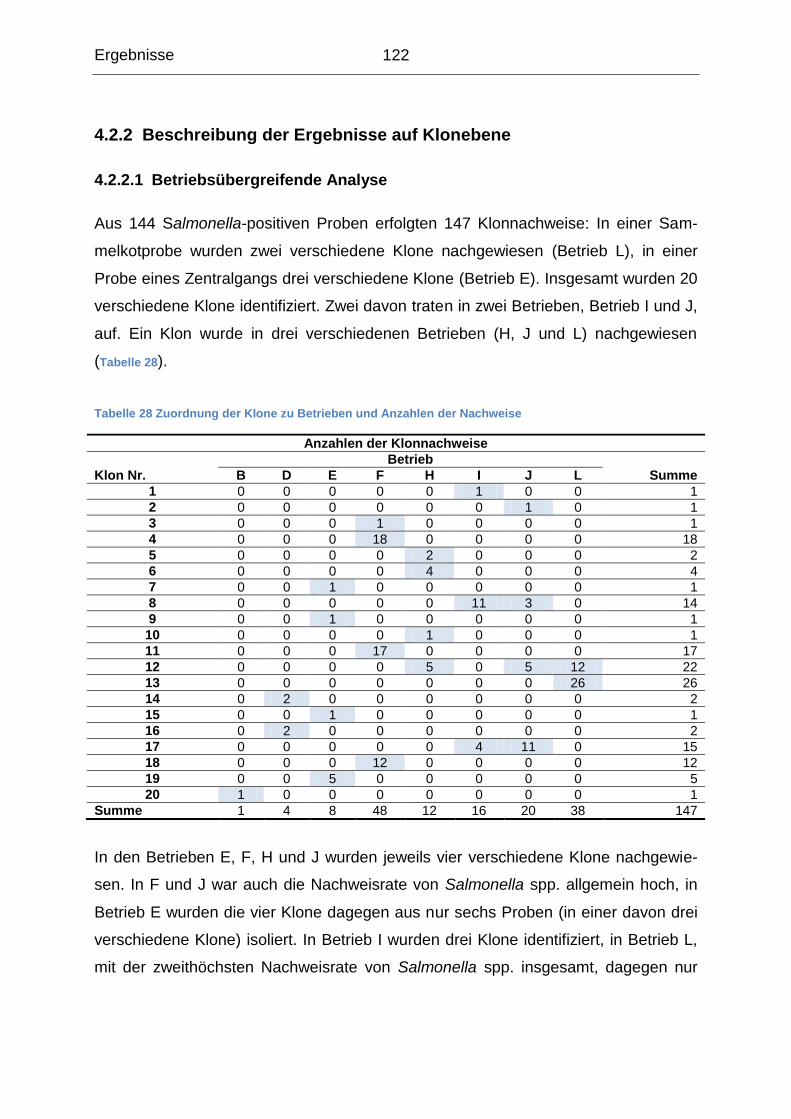
Ergebnisse
122
4.2.2 Beschreibung der Ergebnisse auf Klonebene
4.2.2.1 Betriebsübergreifende Analyse
Aus 144 Salmonella-positiven Proben erfolgten 147 Klonnachweise: In einer Sam-
melkotprobe wurden zwei verschiedene Klone nachgewiesen (Betrieb L), in einer
Probe eines Zentralgangs drei verschiedene Klone (Betrieb E). Insgesamt wurden 20
verschiedene Klone identifiziert. Zwei davon traten in zwei Betrieben, Betrieb I und J,
auf. Ein Klon wurde in drei verschiedenen Betrieben (H, J und L) nachgewiesen
(Tabelle 28).
Tabelle 28 Zuordnung der Klone zu Betrieben und Anzahlen der Nachweise
Anzahlen der Klonnachweise
Betrieb
Klon Nr. B D E F H I J L Summe
1 0 0 0 0 0 1 0 0 1
2 0 0 0 0 0 0 1 0 1
3 0 0 0 1 0 0 0 0 1
4 0 0 0 18 0 0 0 0 18
5 0 0 0 0 2 0 0 0 2
6 0 0 0 0 4 0 0 0 4
7 0 0 1 0 0 0 0 0 1
8 0 0 0 0 0 11 3 0 14
9 0 0 1 0 0 0 0 0 1
10 0 0 0 0 1 0 0 0 1
11 0 0 0 17 0 0 0 0 17
12 0 0 0 0 5 0 5 12 22
13 0 0 0 0 0 0 0 26 26
14 0 2 0 0 0 0 0 0 2
15 0 0 1 0 0 0 0 0 1
16 0 2 0 0 0 0 0 0 2
17 0 0 0 0 0 4 11 0 15
18 0 0 0 12 0 0 0 0 12
19 0 0 5 0 0 0 0 0 5
20 1 0 0 0 0 0 0 0 1
Summe 1 4 8 48 12 16 20 38 147
In den Betrieben E, F, H und J wurden jeweils vier verschiedene Klone nachgewie-
sen. In F und J war auch die Nachweisrate von Salmonella spp. allgemein hoch, in
Betrieb E wurden die vier Klone dagegen aus nur sechs Proben (in einer davon drei
verschiedene Klone) isoliert. In Betrieb I wurden drei Klone identifiziert, in Betrieb L,
mit der zweithöchsten Nachweisrate von Salmonella spp. insgesamt, dagegen nur
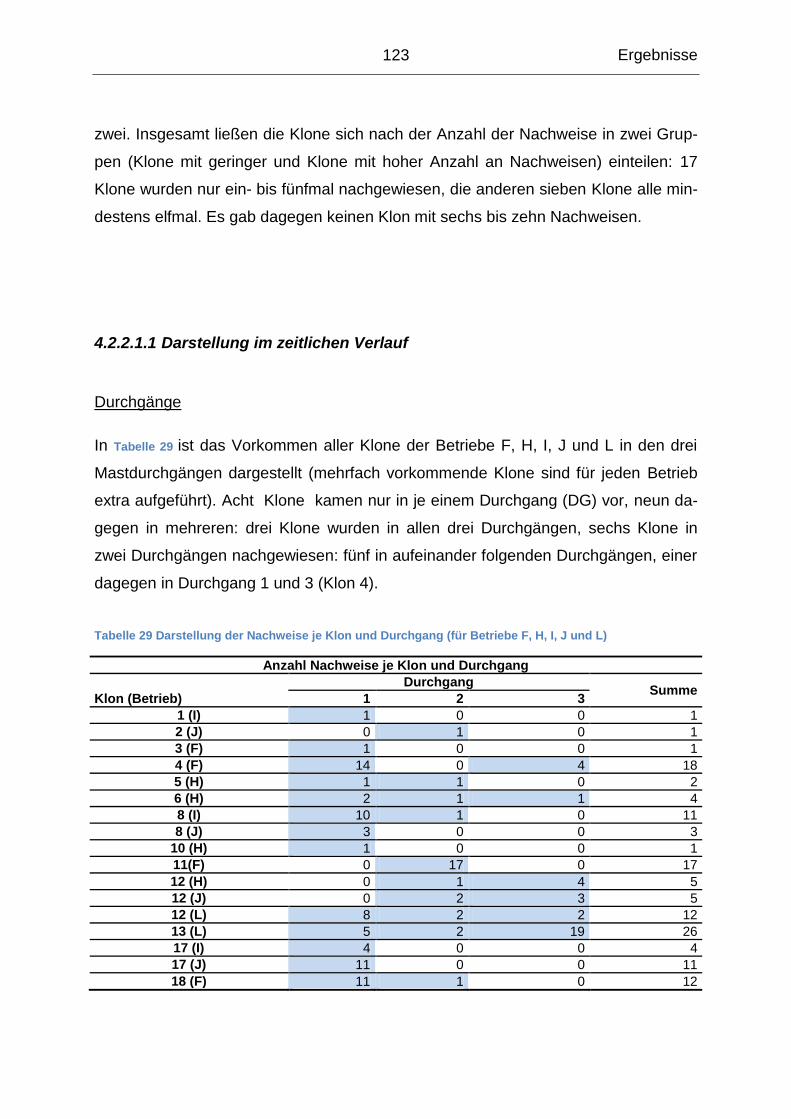
Ergebnisse
123
zwei. Insgesamt ließen die Klone sich nach der Anzahl der Nachweise in zwei Grup-
pen (Klone mit geringer und Klone mit hoher Anzahl an Nachweisen) einteilen: 17
Klone wurden nur ein- bis fünfmal nachgewiesen, die anderen sieben Klone alle min-
destens elfmal. Es gab dagegen keinen Klon mit sechs bis zehn Nachweisen.
4.2.2.1.1 Darstellung im zeitlichen Verlauf
Durchgänge
In Tabelle 29 ist das Vorkommen aller Klone der Betriebe F, H, I, J und L in den drei
Mastdurchgängen dargestellt (mehrfach vorkommende Klone sind für jeden Betrieb
extra aufgeführt). Acht Klone kamen nur in je einem Durchgang (DG) vor, neun da-
gegen in mehreren: drei Klone wurden in allen drei Durchgängen, sechs Klone in
zwei Durchgängen nachgewiesen: fünf in aufeinander folgenden Durchgängen, einer
dagegen in Durchgang 1 und 3 (Klon 4).
Tabelle 29 Darstellung der Nachweise je Klon und Durchgang (für Betriebe F, H, I, J und L)
Anzahl Nachweise je Klon und Durchgang
Durchgang Summe
Klon (Betrieb) 1 2 3
1 (I) 1 0 0 1
2 (J) 0 1 0 1
3 (F) 1 0 0 1
4 (F) 14 0 4 18
5 (H) 1 1 0 2
6 (H) 2 1 1 4
8 (I) 10 1 0 11
8 (J) 3 0 0 3
10 (H) 1 0 0 1
11(F) 0 17 0 17
12 (H) 0 1 4 5
12 (J) 0 2 3 5
12 (L) 8 2 2 12
13 (L) 5 2 19 26
17 (I) 4 0 0 4
17 (J) 11 0 0 11
18 (F) 11 1 0 12
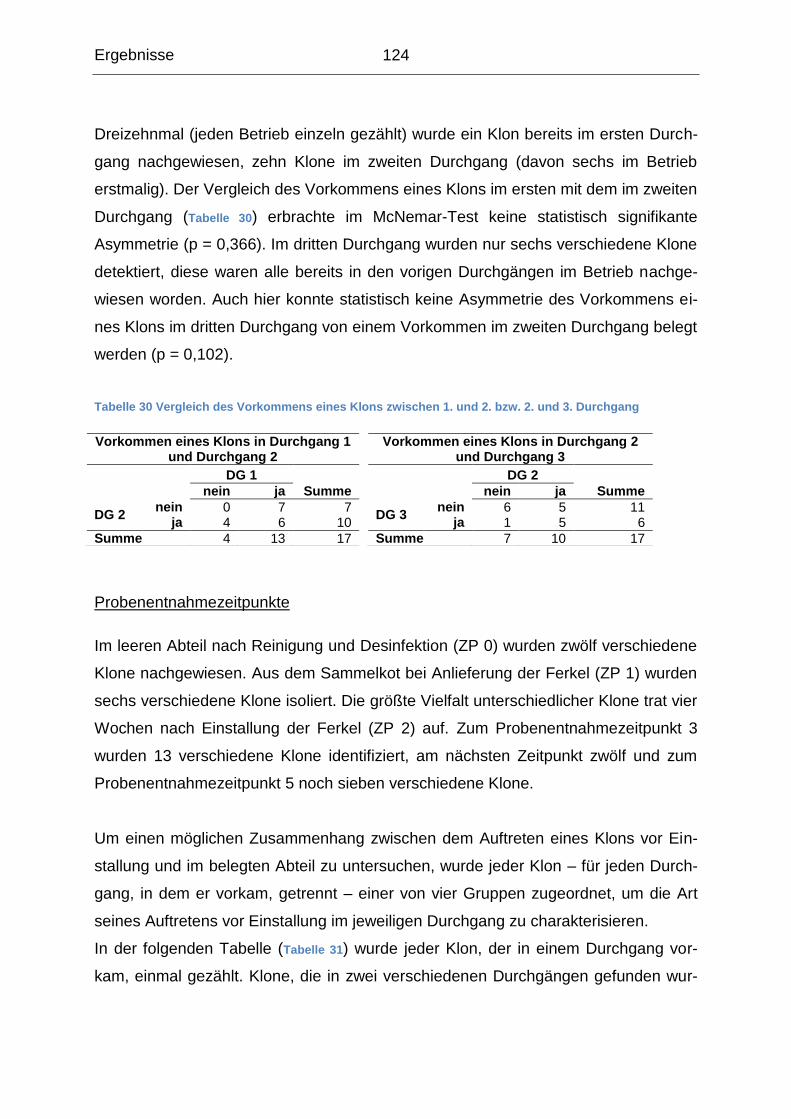
Ergebnisse
124
Dreizehnmal (jeden Betrieb einzeln gezählt) wurde ein Klon bereits im ersten Durch-
gang nachgewiesen, zehn Klone im zweiten Durchgang (davon sechs im Betrieb
erstmalig). Der Vergleich des Vorkommens eines Klons im ersten mit dem im zweiten
Durchgang (Tabelle 30) erbrachte im McNemar-Test keine statistisch signifikante
Asymmetrie (p = 0,366). Im dritten Durchgang wurden nur sechs verschiedene Klone
detektiert, diese waren alle bereits in den vorigen Durchgängen im Betrieb nachge-
wiesen worden. Auch hier konnte statistisch keine Asymmetrie des Vorkommens ei-
nes Klons im dritten Durchgang von einem Vorkommen im zweiten Durchgang belegt
werden (p = 0,102).
Tabelle 30 Vergleich des Vorkommens eines Klons zwischen 1. und 2. bzw. 2. und 3. Durchgang
Probenentnahmezeitpunkte
Im leeren Abteil nach Reinigung und Desinfektion (ZP 0) wurden zwölf verschiedene
Klone nachgewiesen. Aus dem Sammelkot bei Anlieferung der Ferkel (ZP 1) wurden
sechs verschiedene Klone isoliert. Die größte Vielfalt unterschiedlicher Klone trat vier
Wochen nach Einstallung der Ferkel (ZP 2) auf. Zum Probenentnahmezeitpunkt 3
wurden 13 verschiedene Klone identifiziert, am nächsten Zeitpunkt zwölf und zum
Probenentnahmezeitpunkt 5 noch sieben verschiedene Klone.
Um einen möglichen Zusammenhang zwischen dem Auftreten eines Klons vor Ein-
stallung und im belegten Abteil zu untersuchen, wurde jeder Klon – für jeden Durch-
gang, in dem er vorkam, getrennt – einer von vier Gruppen zugeordnet, um die Art
seines Auftretens vor Einstallung im jeweiligen Durchgang zu charakterisieren.
In der folgenden Tabelle (Tabelle 31) wurde jeder Klon, der in einem Durchgang vor-
kam, einmal gezählt. Klone, die in zwei verschiedenen Durchgängen gefunden wur-
Vorkommen eines Klons in Durchgang 1 und Durchgang 2
DG 1
Summe nein ja
DG 2 nein 0 7 7
ja 4 6 10
Summe 4 13 17
Vorkommen eines Klons in Durchgang 2 und Durchgang 3
DG 2
Summe nein ja
DG 3 nein 6 5 11
ja 1 5 6
Summe 7 10 17

Ergebnisse
125
den, sind in dieser Tabelle auch zwei Mal berücksichtigt. 13 Klone kamen im jeweili-
gen Durchgang im leeren, gereinigten und desinfizierten Abteil vor, davon kamen
sechs Klone im gleichen Durchgang später nicht mehr vor, sieben traten auch zu
späteren ZP wieder auf. Von den fünf Klonen, die in einem Durchgang im Sammelkot
bei Ferkelanlieferung nachgewiesen wurden, waren drei auch im belegten Abteil
nachweisbar. Die zwei Klone, die in einem Mastdurchgang sowohl im leeren Abteil
als auch im LKW-Sammelkot identifiziert wurden, wurden beide auch während der
Mast wieder nachgewiesen. 17 Klone wurden während eines Durchgangs nur im be-
legten Abteil und vorher nicht nachgewiesen. Insgesamt zwölf Mal wurde demnach
ein Klon im jeweiligen Durchgang sowohl vor Einstallung (also im leeren Abteil und /
oder im LKW-Sammelkot) als auch im belegten Abteil nachgewiesen, acht Mal nur
vor Einstallung und 17 Mal nur im belegten Abteil. Die Odds Ratio für das Vorkom-
men eines Klons im belegten Abteil je nach seinem Vorkommen vor Einstallung (gar
nicht - ZP 0 oder 1 - beides) war gerade nicht signifikant (OR: 1,945, KG 0,951 –
3,977).
Tabelle 31 Anzahl verschiedener Klone im belegten Abteil eines DG je nach Auftreten dieser vor Einstal-lung (aus allen 16 positiven DG)
Anzahl Klone im belegten Abteil je nach Vorkommen vor Einstallung
Vorkommen an ZP 2 - ZP 5 (belegtes Abt.)
nein (n) ja (n) Summe
Vorkommen vor Einstallung
ZP 0 positiv ZP 1 negativ 6 7 13
ZP 0 negativ ZP 1 positiv 2 3 5
ZP 0 und ZP 1 positiv 0 2 2
ZP 0 und ZP 1 negativ - 17 17
Summe 8 29 37
Der Test auf Übereinstimmung zwischen dem Vorkommen eines Klons im leeren und
im belegten Abteil (Tabelle 32) ergab eine leichte Übereinstimmung (Kappa = 0,19),
während der McNemar-Test auf Symmetrie nicht signifikant war (p = 0,07).

Ergebnisse
126
Tabelle 32 Übereinstimmung des Vorkommens eines Klons im leeren und im belegten Abteil
Übereinstimmung Klonvorkommen in leerem und belegtem Abteil
Vorkommen am ZP 0 Summe
Anzahl Klone (n) Nein Ja
Vorkommen an ZP 2 - 5
Nein 27 8 35
Ja 17 12 29
Summe 44 20 64
Um das Auftreten eines Klons im Durchgang nicht nur qualitativ (im Sinne einer Ja-
Nein-Entscheidung) zu untersuchen, sondern auch die Häufigkeit des Auftretens zu
berücksichtigen, wurde im Folgenden die Anzahl der Klonnachweise dargestellt: Ta-
belle 33 teilt alle Klone nach ihrem ersten Nachweis im Durchgang in Gruppen ein. Die
Anzahlen der Nachweise des oder der Klone vor Einstallung werden denen nach
Einstallung gegenüber gestellt. Während 21 Nachweise von Klonen, die im Durch-
gang im leeren Abteil auftraten, gelangen, wurden diese zu den folgenden Zeitpunk-
ten insgesamt 33 Mal wieder nachgewiesen. Das Verhältnis betrug 1 : 1,6. Den neun
Nachweisen von Klonen, die im LKW-Kot (aber nicht im leeren Abteil) auftraten,
konnten 24 Nachweise dieser im Verlauf des Durchgangs gegenübergestellt werden.
Hier war das Verhältnis 1 : 2,7. Viermal wurden Klone, die gleichzeitig im leeren Ab-
teil und im LKW-Kot auftraten, detektiert. Zehnmal wurden solche Klone zu den ZP 2
bis 5 erneut isoliert. Somit betrug das Verhältnis 1 : 2,5. Klone, die im Durchgang erst
im belegten Abteil auftraten, wurden insgesamt 46 Mal nachgewiesen.
Tabelle 33 Anzahl der Nachweise von Klonen der einzelnen Erstnachweis-Gruppen vor u. nach Belegung
Klonnachweise im Durchgang
Anzahl der Klonnachweise (n)
Summe vor Einstallung belegtes Abteil
Art des ersten Auftretens
ZP 0 positiv ZP 1 negativ 21 33 54
ZP 0 negativ ZP 1 positiv 9 24 33
ZP 0 und ZP 1 positiv 4 10 14
P 0 und ZP 1 negativ - 46 46
Summe 34 113 147

Ergebnisse
127
Verknüpfung von Zeitpunkt und Durchgang
Daneben sollte auch der Einfluss des Zeitpunktes des Erstnachweises eines Klons in
einem Durchgang auf sein Auftreten im Folgenden untersucht werden. Dieses Ver-
hältnis ist anhand der Klonanzahlen in den beiden folgenden Tabellen visualisiert:
Abbildung 14 Darstellung des Vorkommens eines Klons im 1. Mastdurchgang, aufgeteilt nach seinem Vorkommen im 2. Durchgang (Betriebe D, E, F, H, I, J, L)
Die Abbildung 14 schließt die DG 1 und 2 aller sieben Betriebe mit mindestens zwei
untersuchten Mastdurchgängen ein. Insgesamt wurde etwas weniger als die Hälfte
der Klone des ersten Durchgangs im zweiten reisoliert. Zwei Klone, die im leeren Ab-
teil detektiert wurden, wurden im nächsten DG erneut isoliert, fünf dagegen nicht.
Drei Klone, die erst im belegten Abteil nachgewiesen wurden, fanden sich auch in
DG 2 wieder, vier nicht. Bei den anderen beiden Gruppen gelang je einmal der
nochmalige Nachweis, einmal nicht. Fünf Klone wurden in DG 2 erstmalig identifiziert.
In Abbildung 15 flossen die fünf Betriebe mit drei beprobten Mastdurchgängen ein:
2 1 1
3
5
5
1 1
4
0
1
2
3
4
5
6
7
8
ErstnachweisZP 0
ErstnachweisZP 1
ErstnachweisZP 0 & 1
Erstnachweis ZP 2 – 5
kein Nachweis
An
zah
l Klo
ne
Erstnachweis in DG 1
Klone in DG 1
kein Nachweis im 2. DG
Nachweis im 2. DG

Ergebnisse
128
Abbildung 15 Darstellung des Vorkommens eines Klons im 2. Mastdurchgang, aufgeteilt nach seinem Vorkommen im 3. Durchgang (Betriebe F, H, I, J, L)
Im dritten Durchgang wurde knapp die Hälfte der Klone (5) des vorigen Durchgangs
reisoliert. Ein Klon wurde ohne vorherigen Nachweis im zweiten Durchgang im dritten
Durchgang nachgewiesen (die übrigen sechs wurden nur im ersten nachgewiesen).
Umgekehrt konnten bei Vergleich des Auftretens eines Klons vor Einstallung des
zweiten Durchgangs je nach Vorkommen im belegten Abteil des ersten (Tabelle 34)
fünf Klone wiederholt isoliert werden (alle sieben Betriebe mit zwei Durchgängen
einbezogen): Alle vier im leeren Abteil nachweisbaren Klone waren bereits während
des vorigen Mastdurchgangs detektiert worden. Ein Klon, der im vorigen Durchgang
während der Mast auftrat, wurde nun bei den Ferkeln vor Einstallung nachgewiesen.
Tabelle 34 Darstellung des Vorkommens eines Klons vor Einstallung des 2. Durchgangs je nach Auftreten im belegten Abteil des 1. DG (alle Betriebe mit zwei DG)
Anzahl der Klone vor Einstallung im 2. DG je nach ZP 2 - 5 des 1. DG
ZP 2 – 5 des 1. DG
Summe negativ (n) positiv (n)
vor Einstallung des 2. DG
ZP 0 positiv 0 4 4
ZP 1 positiv 1 1 2
Negativ 7 10 17
Summe 8 15 23
2 2 1 1
2 3
6
0
1
2
3
4
5
6
7
8
ErstnachweisZP 0
ErstnachweisZP 1
ErstnachweisZP 0 & 1
Erstnachweis ZP 2 – 5
kein Nachweis
An
zah
l Klo
ne
Erstnachweis in DG 2
Klone in DG 2
kein Nachweis im 3. DG
Nachweis im 3. DG

Ergebnisse
129
Die entsprechende Auswertung der Durchgänge 2 und 3 zeigte ebenfalls, dass die
Klone, die im leeren Abteil gefunden wurden, bereits im vorherigen Durchgang nach-
gewiesen worden waren. (Tabelle 35). Drei Klone des zweiten Mastdurchgangs wurden
im 3. Durchgang vor Einstallung nicht reisoliert.
Tabelle 35 Darstellung des Vorkommens eines Klons vor Einstallung des 3. Durchgangs je nach Auftreten im belegten Abteil des 2. DG (Betriebe F, H, I, J, L)
Anzahl der Klone vor Einstallung im 3. DG je nach ZP 2 - 5 des 2. DG
ZP 2 – 5 des 2. DG
Summe negativ (n) positiv (n)
vor Einstallung des 3. DG
ZP 0 positiv 0 2 2
ZP 1 positiv 0 1 1
Negativ 11 3 14
Summe 11 6 17
4.2.2.1.2 Darstellung nach Lokalisationen und -gruppen
In Abbildung 16 sind für jeden Klon die Anzahlen seiner Nachweise in den vier Lokali-
sationsgruppen (über alle Durchgänge hinweg, für jeden Betrieb getrennt) dargestellt.
Die höchste Anzahl von Nachweisen erfolgte in der tierfernen Umgebung. Alle Klone
bis auf einen wurden dort nachgewiesen. In der tiernahen Umgebung kamen zwölf
verschiedene Klone vor. Im Sammelkot wurden zehn verschiedene Klone nachge-
wiesen: vier davon nur bei Ferkelanlieferung, vier nur während der Mast. Lediglich
zwei Klone traten sowohl in Sammelkotproben der Ferkelanlieferung als auch des
laufenden Mastdurchgangs auf. Aus Schädlingen wurden nur drei verschiedene Klo-
ne isoliert, ein Klon von der Oberfläche toter Schadnager, ein Klon zusätzlich aus
dem Nagerkot, einer aus Fliegen.
Bei den Klonen mit geringer Anzahl von Nachweisen (1 - 5) war fast immer haupt-
sächlich die tierferne Umgebung betroffen: bei neun Klonen ausschließlich, bei vier

Ergebnisse
130
Klonen in Kombination mit einer anderen Gruppe. Zwei Klone kamen zusätzlich in
zwei weiteren Gruppen vor. Nur ein Klon wurde nur aus dem LKW-Sammelkot isoliert.
Die sieben Klone mit vielen (mindestens elf) Nachweisen traten immer in drei oder
sogar allen vier Gruppen auf.
Abbildung 16 Darstellung der der Klone mit Anzahl der Nachweise in den einzelnen Lokalisationsgruppen
Vergleich der Fundorte zwischen zwei Durchgängen
Bei Vergleich von zwei aufeinander folgenden Durchgängen (DG 1 mit DG 2 bzw.
DG 2 mit DG 3) wurden zwölf Klone in beiden Durchgängen nachgewiesen (Tabelle 36).
Fünf davon waren im Durchgang davor nur an einer einzigen Lokalisationsgruppe
präsent, vier an zwei verschiedenen bzw. drei verschiedenen Gruppen. Somit wur-

Ergebnisse
131
den aus allen Lokalisationsgruppen bzw. -kombinationen, in denen Klone im vorigen
Durchgang vorlagen, auch wieder Klone im nächsten Durchgang nachgewiesen. Von
den sieben Klonen, die im ersten von zwei aufeinander folgenden Durchgängen nur
in der tierfernen Umgebung nachweisbar waren, fanden sich drei im folgenden
Durchgang wieder. Bei Klonen der Kombination Sammelkot - tiernahe - tierferne
Umgebung traten zwei von fünf im folgenden Durchgang auf. In der tiernahen Umge-
bung bzw. in der Kombination tierferne Umgebung und Schädlinge trat jeweils nur
ein Klon in einem der ersten beiden Durchgänge auf, beide wurden auch im jeweils
folgenden Durchgang isoliert. Bei genauerer Betrachtung der Lokalisationsgruppen,
an denen die Klone dann im folgenden Durchgang auftraten, konnte kein regelmäßi-
ger Zusammenhang mit dem Ort des Auftretens im Durchgang zuvor gefunden wer-
den (Tabelle 36).
Vergleich der Fundorte vor und nach Einstallung eines Durchgangs
Bei Vergleich der Fundorte von Klonen vor und nach Einstallung (Tabelle 37) zeigte
sich kein signifikanter Unterschied der Wiederfindungsrate zwischen den Lokalisati-
onsgruppen. Vor Einstallung traten Klone am häufigsten nur in der tierfernen Umge-
bung auf. Von diesen sieben wurden später drei im belegten Abteil nachgewiesen.
Von den vier Klonen aus tiernaher und tierferner Umgebung wurden ebenfalls drei
auch im belegten Abteil reisoliert. Die Hälfte der vier Klone, die vor Einstallung aus-
schließlich im Sammelkot detektiert wurden, fand sich auch während des Mastdurch-
gangs wieder. Drei Klone wurden zusätzlich auch im leeren Abteil (in tierferner oder
tiernaher Umgebung) vorgefunden. Jedes Mal wurden diese Klone im belegten Abteil
erneut nachgewiesen. Auch hier konnte kein einheitliches Verteilungsmuster identifi-
ziert werden. Genau ein Klon trat im belegten Abteil in genau den gleichen Lokalisa-
tionsgruppen auf wie vor Einstallung.

Ergebnisse
132
Tabelle 36 Vergleich der Lokalisationsgruppe(n), an denen ein Klon auftritt, zwischen zwei aufeinander folgenden Durchgängen (alle beprobten DG in positiven Betrieben)
Anzahl Klone
Lokalisationsgruppe im folgenden Durchgang
Gruppen im vori-gen Durchgang
kein Nachweis
Sammel-kot (SK)
tier- nahe U.
tier- ferne U.
SK u. tierfer- ne U.
tiernahe u. tierfer-
ne U.
tierferne U. u. Schädlinge
SK, tiernahe u. tierferne U.
Summe
kein Nachweis 6 1 0 2 0 1 1 1 12
Sammelkot 2 0 0 1 0 0 0 0 3
tiernahe U. 0 0 1 0 0 0 0 0 1
tierferne U. 7 0 1 2 0 0 0 0 10
Sammelkot u. tierferne U.
1 0 0 0 0 1 0 1 3
tiernahe u. tierfer-ne U.
2 0 0 0 1 0 0 0 3
tierferne U. u. Schädlinge
0 0 0 0 0 1 0 0 1
SK, tiernahe u. tierferne U.
3 1 0 1 0 0 0 0 5
tiernahe, tierferne U. u. Schädlinge
1 0 0 0 1 0 0 0 2
Summe 22 2 2 6 2 3 1 2 40
Tabelle 37 Vergleich der Lokalisationsgruppe(n), an denen ein Klon auftritt, vor Einstallung und im beleg-ten Abteil (alle positiven DG)
Anzahl Klone
Lokalisationsgruppen im belegten Abteil
kein
Nach-weis
Sam-melkot
(SK)
tier-nahe U.
tierfer-ne U.
SK u. tierferne
U.
tiernahe u. tierfer-
ne U.
tiernahe U. u.
Schädl.
tierfer- ne U. u. Schädl.
SK, tier-nahe u.
tierferne U.
tiernahe, tierferne U. u. Schädl.
Summe Gruppen vor Ein-stallung
kein Nachweis 27 1 1 10 0 3 0 1 1 0 44
Sammelkot 2 0 0 1 0 0 0 0 1 0 4
tiernahe U. 1 0 0 0 0 0 0 0 0 0 1
tierferne U. 4 1 0 0 1 0 0 0 0 1 7
SK u. tiernahe U. 0 0 0 0 0 1 0 0 0 0 1
Sammelkot u. tierferne U.
0 0 1 0 0 0 0 0 1 0 2
tiernahe u. tierfer-ne U.
1 0 0 1 1 1 0 0 0 0 4
tiernahe, tierferne U. u. Schädlinge
0 0 0 0 0 0 1 0 0 0 1
Summe 35 2 2 12 2 5 1 1 3 1 64

Ergebnisse
133
4.2.2.2 Innerbetriebliche Analyse
Im Folgenden soll kurz auf die Salmonellenvorkommen und -verläufe in den einzel-
nen Betrieben eingegangen werden:
In Betrieb B wurden einmalig am ZP 2 Salmonella spp. am Treibbrett nachgewiesen.
In Betrieb D wurde in DG 1 am ZP 2 ein Klon an Futtertrögen und Zentralgang de-
tektiert, ein weiterer an den Trennwänden. Dieser fand sich vier Wochen später an
den Stiefeln des Landwirts wieder. In DG 2 gelangen keine Nachweise.
In Betrieb E wurden vier verschiedene Klone in fünf Proben nachgewiesen: im ers-
ten DG am ZP 2 im Zentralgang drei Klone, vier Wochen später nur noch einer da-
von; im nächsten Durchgang, ZP 3, wurde im Zentralgang ein „neuer“ Klon identifi-
ziert, vier Wochen später wies das Treibbrett den in DG 1 zweimal nachgewiesenen
Klon auf.
In Betrieb F wurden in DG 1 bereits im leeren Abteil alle drei Klone des Durchgangs
nachgewiesen. Der nur einmalig detektierte Klon 3 wurde von den Rohrleitungen iso-
liert. Der häufige Klon 4 trat nach RuD ebenfalls an Rohrleitungen, aber auch Lüftung
und Buchtenboden auf. Er wurde zu allen weiteren ZP des DG reisoliert, meist in der
tierfernen Umgebung, nur gegen Ende zweimal aus Sammelkot. Klon 18 kam an den
Trennwänden nach RuD vor, wurde aber auch durch die Tiere eingetragen (ZP 1). Im
weiteren Verlauf war er ausschließlich im Abteil und nicht mehr im Kot zu finden, er
verlagerte sich von der tiernahen immer mehr in die tierferne Umgebung. In DG 2 trat
Klon 18 noch ein einziges Mal auf: zum ZP 0 auf dem Treibbrett. Im belegten Abteil
wurde nur noch der „neue“ Klon 11 nachgewiesen, an den ZP 2 - 4 hauptsächlich in
der tiernahen Umgebung, zum ZP 5 tierfern und im Sammelkot. In DG 3 wurde wie-
der und ausschließlich Klon 4 nachgewiesen, allerdings erst im belegten Abteil. Zum
ZP 2 trat er in den Buchten und im Gang auf, danach nur noch einmal im Gang.

Ergebnisse
134
In Betrieb H wiesen zwölf Proben vier unterschiedliche Klone auf. In DG 1 wurde ein
Klon aus dem LKW-Sammelkot isoliert (ZP 1), der jedoch danach nie wieder nach-
gewiesen wurde. Zwei Klone traten Mitte bis Ende der Mast sporadisch in der tierfer-
nen Umgebung (Treibbrett und Gang) auf. In DG 2 wurden zwei Klone des vorigen
DG nach RuD von Buchtenboden und Gang isoliert, im LKW-Sammelkot trat ein
„neuer“ Klon (auch in Betrieben J und L nachgewiesen!) auf. Keiner dieser Klone
fand sich im weiteren DG wieder. In DG 3 wurden zunächst keine Salmonellen nach-
gewiesen, ab Mitte der Mast dann Klon 12 in Gang und Arbeitsraum. Am ZP 5 trat
wieder einer der beiden bereits in DG 1 und 2 nachgewiesenen Klone in der tierna-
hen Umgebung auf.
In Betrieb I wurden drei verschiedene Klone nachgewiesen, zwei davon (8, 17) auch
in Betrieb J. Ein Klon wurde vor Einstallung des ersten DG im Gang nachgewiesen,
danach nie mehr, ein weiterer (17) im Arbeitsraum. Die Ferkel trugen bei Anlieferung
Klon 8 in die Mast ein, dieser dominierte im weiteren Mastverlauf, zunächst in Sam-
melkot, Gang und tiernaher Umgebung, später im Sammelkot. Der anfangs im Ar-
beitsraum detektierte Klon (17) trat Ende der Mast erst im Gang, dann zusätzlich im
Sammelkot auf. Im zweiten DG trat nur zum ZP 2 einmal Klon 8 aus dem ersten DG
auf. Der dritte DG wies keine Salmonellenfunde auf.
Betrieb J wies vier verschiedene Klone (8, 17 wie Betrieb I, 12 wie L) auf. In DG 1
wurden vor Einstallung an mehreren Lokalisationen Klone nachgewiesen: Klon 17
sowohl in tiernaher, tierferner Umgebung und auf Schadnagern, Klon 8 im Gang und
bei den ankommenden Ferkeln. Klon 17 wurde im Mastverlauf zu jedem ZP, in der
tiernahen Umgebung und auf Nagern nachgewiesen, Klon 8 nur zum ZP 2 an
Trennwänden. Im folgenden DG traten zwei völlig andere Klone auf: gegen Masten-
de Klon 12 in tierferner Umgebung und auf Schadnagern und Klon 2 einmalig im Ar-
beitsraum. In DG 3 wurde Klon 12 nach RuD erneut an Trennwänden und im Arbeits-
raum nachgewiesen. Nach drei negativen Beprobungen wurde er zum ZP 5 erneut
im Arbeitsraum isoliert.

Ergebnisse
135
In Betrieb L wurden trotz 35 positiver Proben nur zwei verschiedene Klone detektiert.
Das Abteil präsentierte sich in DG 1 bereits vor Einstallung mehrfach positiv in tier-
naher (Klon 13) und tierferner Umgebung (12 und 13). Beide Klone waren auch im
weiteren Verlauf in der Umgebung, Klon 12 auch in Fliegen präsent. Im nächsten DG
wiesen vor Einstallung das Treibbrett Klon 12, die Ferkel Klon 13 auf. Im Folgenden
gelangen nur wenige Klonnachweise (beide Klone je einmal im Sammelkot und 13 im
Zentralgang). In DG 3 wurden beide erneut im leeren Abteil, Klon 13 auch bei den
Ferkeln nachgewiesen, allein dieser Klon fand sich im weiteren Mastdurchgang in
Umgebung und Sammelkot.

Diskussion
136
5. Diskussion
Die vorliegende Arbeit umfasst Untersuchungen zu Salmonella spp., Campylobacter
spp. und Y. enterocolitica in nordwestdeutschen Schweinemastbeständen. Diese
Erreger repräsentieren die drei häufigsten Ursachen bakteriell bedingter, lebensmit-
telassoziierter Enteritiden beim Menschen in der Europäischen Union. Das Schwein
spielt als Reservoir aller drei Pathogene eine wichtige Rolle, da Schweinefleisch der
europaweit höchste Pro-Kopf-Konsum von allen Fleischarten zugeschrieben wird.
Daher wurden im Rahmen des Forschungsverbundes „Food-borne zoonotic infec-
tions“ in Kooperation mit dem Robert Koch-Institut Wernigerode und dem Institut für
Mikrobiologie und Tierseuchen der Stiftung Tierärztliche Hochschule Hannover zwei
Teilprojekte bearbeitet: eine Querschnittsuntersuchung zu Vorkommen und Risiko-
faktoren der drei Erreger und eine Verlaufsuntersuchung zur Dynamik von Salmonel-
len in Schweinemastbeständen.
In der Querschnittsuntersuchung wurden in 25 Betrieben der Salmonellen-Kategorie I
und 25 Betrieben der Salmonellen-Kategorie III insgesamt 700 Kot- und Umge-
bungsproben entnommen und bakteriologisch auf die drei Pathogene untersucht.
Zudem wurde eine epidemiologische Erhebung und Auswertung von Betriebsdaten
durchgeführt.
.
In der Verlaufsuntersuchung wurden in zwölf Betrieben der Kategorie III während
einem bis drei Mastdurchgängen 1672 Kot- und Umgebungsproben entnommen.
Diese wurden bakteriologisch und molekularbiologisch auf das Vorhandensein von
Salmonella spp. analysiert und die Isolate feintypisiert.

Diskussion
137
5.1 Querschnittsuntersuchung
5.1.1 Erregernachweis
In den Betrieben wurden jeweils vier Sammelkotproben und diverse Umgebungspro-
ben entnommen. Für den Erregernachweis wurde die kulturelle Untersuchung ge-
wählt, da sie für alle drei Erreger bei günstigem Kosten-Nutzen-Verhältnis eine sensi-
tive und spezifische Methode darstellt und bereits in vielen vorangegangenen Stu-
dien angewandt wurde. Zudem besitzt sie gegenüber molekularbiologischen Metho-
den den Vorteil, dass ein Erregernachweis gleichzeitig ein Nachweis seiner Vermeh-
rungsfähigkeit und damit seiner potentiellen Infektiosität darstellt. Auf eine Spezies-
differenzierung wurde wegen geringer Relevanz für diese Studie verzichtet. Für die
Untersuchung wurde ein Zeitpunkt kurz vor Schlachtung gewählt, auch wenn einigen
Studien zufolge das Ausscheidungsmaximum der Erreger mit dem Kot meist früher in
der Mast terminiert ist (WEIJTENS et al. 1999; KASIMIR 2005; WEHEBRINK et al.
2008). Dies erfolgte deshalb, weil das Erregervorkommen möglichst kurz vor der
Schlachtung abgebildet werden sollte, da hier ein viel engerer Zusammenhang mit
einem möglichen Eintrag in die Lebensmittelkette konstatiert wird.
Die Gesamtnachweisrate über alle Proben betrug für Campylobacter spp. 25,0 %,
Y. enterocolitica 8,6 % und Salmonella spp. 7,3 %. Da es sich um ein Probenkollektiv
unterschiedlicher Lokalisationen handelte, sind diese Zahlen a priori nicht mit denen
anderer Studien vergleichbar. Hierfür müssen zur Beurteilung auf Tierebene die Er-
gebnisse der Kotproben herangezogen werden: Zur Untersuchung der Tiere wurden
je Betrieb vier Sammelkotproben entnommen. In diesen betrug die Nachweisrate
insgesamt für Campylobacter spp. 38,1 %, für Y. enterocolitica 17,1 % und für Sal-
monella spp. 11 %.
Damit liegt die Nachweisrate im Falle von Campylobacter spp. niedriger als bei AL-
TER et al. (2005), der eine Einzeltierprävalenz von 79,1 % angab. Auch VON ALT-
ROCK et al. (2006) und WEHEBRINK et al. (2008) ermittelten mit 69,7 % bzw.
64,7 % höhere bakteriologische Einzeltierprävalenzen. Dabei sind jedoch auch diese

Diskussion
138
Angaben nicht unmittelbar miteinander vergleichbar, da in diesen Arbeiten Einzeltier-
kotproben untersucht wurden, wohingegen in der vorliegenden Studie der Fokus auf
der Tierumgebung lag. Daher wurde, um bei überschaubarer Probenzahl ein mög-
lichst größeres Tierkollektiv abzubilden, auf Sammelkot zurückgegriffen. Obwohl die
Entnahme möglichst frischen Kots anvisiert wurde, könnte dies aufgrund der geringe-
ren Tenazität von Campylobacter spp. außerhalb des Wirtes einen Einfluss auf die
Überlebensfähigkeit des Erregers gehabt haben. In diesem Zusammenhang dürfte
auch der Transport der Kotproben ohne Transportmedium nachteilig gewesen sein.
In den zitierten Studien wurden stets Tupfer mit Cary-Blair-Medium verwandt. Zudem
unterschieden sich die Untersuchungsprotokolle zum kulturellen Nachweis des Erre-
gers zwischen den Studien.
Der Anteil von Y. enterocolitica lag dagegen höher als bei VON ALTROCK et al.
(8,4 %) und WEHEBRINK et al. (12,2 %). Bei diesem weniger empfindlichen Erreger
könnte sich die Untersuchung einer größeren Tierzahl durch gepoolte Kotproben von
Vorteil erwiesen haben, denn die genannten Studien untersuchten ausschließlich
Einzeltiere. Auch die Untersuchungsprotokolle für den kulturellen Nachweis von
Y. enterocolitica wichen voneinander ab. Dies ist für die Beurteilung von Yersinia-
Nachweisen besonders relevant, weil gerade der Nachweis dieses Erregers sehr
stark je nach verwendeter Labormethodik variiert (FREDRIKSSON-AHOMAA u.
KORKEALA 2003) In der vorliegenden Arbeit wurde im Gegensatz zu den anderen
beiden Studien eine Kälteanreicherung über drei Wochen durchgeführt. Diese wird
besonders für Kotproben empfohlen, da sie das Wachstum der Begleitflora unter-
drückt (LAUKKANEN et al. 2010).
Für Salmonella spp. bewegte sich die Nachweisrate innerhalb der durch das Litera-
turstudium generierten Erwartungen. In einer deutschen Studie (STEINBACH et al.
2002) erwiesen sich bei individueller kultureller Untersuchung von Rektuminhalt auf
dem Schlachthof 3,7 % der Tiere als positiv. Die höhere kulturelle Nachweisrate von
16 % aus Poolproben bei SOMYANONTANAGUL (2009) erklärt sich aus der aus-
schließlichen Rekrutierung von Kategorie III-Betrieben. LO FO WONG et al. (2003)

Diskussion
139
ermittelten in Sammelkotproben eine recht ähnliche Nachweisrate von 9,3 %. Dass
Poolproben sich für den Nachweis von Salmonella spp. besser eignen als Individual-
proben, wird in mehreren Studien belegt (SANCHEZ et al. 2007; FOSSE et al. 2009;
WILKINS et al. 2010).
Auf Herdenlevel stimmten die Ergebnisse weitgehend mit den Angaben in der Litera-
tur überein: Genau 80 % der Betriebe wurden bei Berücksichtigung aller Proben als
campylobacter-positiv eingestuft. Dies ist weniger als die von VON ALTROCK et al.
(2006) und ALTER et al. (2005) angegebenen 100 %, ist jedoch durch die geringere
Anzahl von Sammelkotproben und stärkeren Gewichtung der (in der Regel seltener
positiven) Tierumgebung begründbar. Die 48 % yersinia-positiver Herden entspra-
chen recht genau den von VON ALTROCK et al. (2006) ermittelten 43,3 % aus Ein-
zelkotproben. Der Anteil von 32 % als salmonella-positiv eingeordneten Beständen
liegt genau zwischen den von LO FO WONG et al. (2003) angegebenen 41,6 %
(Poolproben) und den durch STEINBACH et al. (2002) berechneten 20 % (Einzel-
proben).
5.1.2 Vorkommen der Erreger in der Tierumgebung
Eine Zielsetzung der Arbeit war, genauere Kenntnisse über die Rolle der Tierumge-
bung in der Übertragung der drei Erreger in Mastbeständen zu erlangen. Während
diese Kenntnisse für Salmonella spp. bereits in ausreichendem Maß vorhanden sind,
herrschen im Hinblick auf die anderen beiden Pathogene noch Defizite. Daher wurde
in dieser Studie der Schwerpunkt auf eine detaillierte Untersuchung der direkten und
indirekten Tierumgebung gelegt. Die Zusammenfassung aller Lokalisationen zu vier
Lokalisationsgruppen erwies sich dabei als vorteilhaft für einen besseren Überblick
bei der Bewertung der Ergebnisse.
Die Studie konnte zeigen, dass der Tierumgebung für alle drei Erreger eine nicht zu
unterschätzende Bedeutung in der Übertragung im Bestand zukommt. Alle drei Erre-
ger waren in allen vier Lokalisationsgruppen nachweisbar. Auch die Nachweisraten

Diskussion
140
lagen über denen einiger anderer Studien, z. B. 0,7 % campylobacter-positiver Um-
gebungsproben bei ALTER et al. (2005), 0 % für Campylobacter spp. und Y.
enterocolitica bei WEHEBRINK et al. (2008), 3,4 % für Y. enterocolitica bei VON
ALTROCK et al. (2006) und 0,6 % für Y. enterocolitica bei PILON et al. (2000). Die
Arbeit belegt, dass die Erreger nicht nur im oder in der unmittelbaren Nähe des Tie-
res, sondern auch an weiter entfernten Orten ohne direkten Tierkontakt überleben
und von dort weiterverbreitet werden können.
5.1.2.1 Campylobacter spp.
Dieser Erreger wurde neben Abteilgängen auch an Treibbrettern und Stiefeln des
Landwirts nachgewiesen. Dies ist umso interessanter, als es sich dabei um einge-
trocknete Verschmutzungen handelte, wo aufgrund der Empfindlichkeit von Campy-
lobacter spp. gegenüber Trockenheit (CORRY et al. 1995) ein Nachweis eher nicht
zu erwarten war. Lokalisationen mit direktem Tierkontakt, allen voran Buchtenwände
und Nippeltränken, aber auch Ketten und Futtertröge, waren sogar fast ebenso häu-
fig positiv wie Sammelkot. Dies weist auf eine generell erhebliche Kontamination der
unmittelbaren Tierumgebung mit Kot in den üblichen Haltungsformen hin und unter-
streicht deren Rolle bei der Übertragung aller möglichen mit dem Kot ausgeschiede-
nen Pathogene. Diese Erkenntnisse bestätigen die Einschätzung von VON ALT-
ROCK et al. (2006), die zumindest für Campylobacter spp. der fäkalen Kontamination
der Umgebung für die Erregerausbreitung den gleichen Stellenwert wie der direkten
fäkal-oralen Übertragung beimessen. Auch auf Betriebsebene hatten ebenso viele
Betriebe Nachweise in Sammelkot wie in der tiernahen Umgebung. Zwar erhöhte
sich die Chance auf eine kontaminierte tiernahe Umgebung signifikant mit steigender
Anzahl positiver Sammelkotproben, dennoch war die Übereinstimmung zwischen
den beiden Lokalisationsgruppen nur mäßig: Ein Viertel der Betriebe wies nur Funde
in einer der beiden Gruppen auf. Es gab folglich einige Betriebe, die bei ausschließli-
cher Untersuchung der Tiere selbst als falsch negativ klassifiziert worden wären.
Dies unterstreicht - bedingt durch die intermittierende und zum Mastende generell
niedrigere Ausscheidungsrate - die Notwendigkeit, dass zur Einschätzung des

Diskussion
141
Campylobacter-Herdenstatus eines Bestands nicht ausschließlich nur Kotproben
verwendet werden sollten (WEIJTENS et al. 1999; WEHEBRINK et al. 2008). Die
tierferne Umgebung war dagegen nur positiv, wenn auch Erregernachweise in tier-
naher Umgebung und Sammelkot erfolgten. Dies lässt vermuten, dass diese kein
abgegrenztes Reservoir repräsentiert, sondern in engem Zusammenhang zu einer
Positivität der Tiere selbst und der unmittelbaren Umgebung steht.
Gleichwohl trat der Erreger ebenso wie bei ALTER et al. (2005) in Fliegen auf, so
dass auch die u.a. von WEIJTENS et al. (1997) aufgestellte These der möglichen
Übertragung durch belebte Vektoren nicht ausgeschlossen werden kann.
5.1.2.2 Y. enterocolitica
Auch Y. enterocolitica wurde, wenn auch seltener als Campylobacter spp., sowohl in
tiernaher als auch tierferner Umgebung nachgewiesen. Die Nachweisrate war höher
als in anderen Untersuchungen. Der Erreger wurde zu immerhin 8 % aus der tierna-
hen Umgebung isoliert, die Buchtenwände waren sogar zu einem Viertel und damit
häufiger positiv als der Tierkot selbst. Hier war die Übereinstimmung beider Gruppen
auf Betriebsebene etwas höher, trotzdem gab es auch hier Betriebe mit Nachweisen
nur in der tiernahen Umgebung. Dies unterstreicht einerseits die Schlussfolgerung
von VON ALTROCK et al. (2006), dass die alleinige kulturelle Untersuchung der Tie-
re selbst aufgrund der intermittierenden Ausscheidung wegen sodann falsch negati-
ver Ergebnisse keine zuverlässige Einschätzung des Herdenstatus zulässt. Sie zeigt
jedoch auch, dass die Tierumgebung entgegen der Hinweise aus jener und anderen
Untersuchungen (PILON et al. 2000; WEHEBRINK et al. 2008) möglicherweise doch
eine größere Rolle bei der Übertragung spielt als angenommen. Dies entspricht der
Haltung von WINGSTRAND und NIELSEN (1996), wonach die Übertragung durch
die Umgebung wichtiger als die von der Sau auf die Ferkel scheint. Diese Prä-
sumtion wird gestützt durch Erregernachweise auch vom Boden des Zentralgangs
und von Treibbrettern, wonach auch diese weiter vom Tier entfernten Lokalisationen
als Ansteckungsquellen fungieren können. PILON et al. (2000) vermuteten aufgrund

Diskussion
142
des potentiellen Vorkommens von Yersinien in vielen belebten Vektoren (ALDOVA u.
LAZNICKOVA 1979) eine Beteiligung dieser am Infektionsgeschehen. Sie konnten in
ihrer Studie allerdings in keinem der Betriebe Yersinien in Schädlingen detektieren.
Dieser Nachweis gelang in der vorliegenden Arbeit sowohl aus Nagerkot, als auch
aus Fliegen, und dient somit der Erhärtung dieser Hypothese. Ein Grund für die un-
terschiedlichen Erregerfunde zwischen den Studien könnte in der Methodik der Pro-
benentnahme und Labordiagnostik liegen: In der vorliegenden Arbeit wurden für die
Untersuchung mehr Fliegen je Probe und statt Darm(-inhalt) einzelner, außerhalb der
Gebäude gefangener Nager in den Gebäuden vorgefundener (Sammel-)Kot verwen-
det. Zudem wurde auch bei diesen Proben im Gegensatz zu der zitierten Untersu-
chung eine Kälteanreicherung durchgeführt.
5.1.2.3 Salmonella spp.
Salmonella spp. wurden generell seltener nachgewiesen als die beiden anderen Pa-
thogene. Dieser Unterschied war besonders deutlich beim Sammelkot, während die
tiernahe Umgebung genauso oft positiv war wie bei Y. enterocolitica, nämlich zu
7,7 %. Dies kongruiert mit den Ergebnissen anderer Studien, in denen der Erreger
ebenfalls regelmäßig an Futtertrögen, Wänden, Ketten und Tränken, nachweisbar
war (BODE 2008; SOMYANONTANAGUL 2009). Die im Vergleich zu den anderen
beiden Erregern stärker ausgeprägte Besiedelung der tierfernen Umgebung und so-
mit größere Bedeutung als Reservoir für Salmonella spp. konnte in dieser Studie nur
bedingt belegt werden. Im Vergleich zu den anderen Pathogenen gab es mehr Be-
triebe, in denen der Erreger in der tiernahen Umgebung detektiert wurde, und es
konnte eine relativ häufige Kontamination der Lauf- und Treibwege nachgewiesen
werden. Dagegen gelang kein Nachweis von Salmonella spp. von einigen tierfernen
Lokalisationen wie Stiefeln, Arbeitsräumen, Rohrleitungen oder Lüftern, die in der
Verlaufsuntersuchung und Studien anderer Autoren (BODE 2008;
SOMYANONTANAGUL 2009) als wichtiges Reservoir identifiziert wurden. Im Falle
der beiden letztgenannten Lokalisationen lag dies daran, dass diese nur im Ausnah-
mefall untersucht wurden, da dort für anderen Erreger kaum Nachweise zu erwarten

Diskussion
143
waren. Weitere Gründe dafür könnten in der Labormethode oder der Probenentnah-
metechnik liegen, die für die Umweltproben mittels Tupfer in Medium erfolgte. Dies
wurde aufgrund der geringeren Tenazität der anderen Erreger in der Umwelt
(CORRY et al. 1995) für alle drei Pathogene einheitlich praktiziert. Der Nachteil liegt
in einer recht geringen Aufnahmekapazität kontaminierten Materials. Dies würde die
insgesamt recht niedrige Nachweisrate erklären, denn in der Untersuchung von SO-
MYANONTANAGUL (2009), der die gleichen Lokalisationen mittels Swabs (größere
Materialaufnahmekapazität) untersuchte, wurden in 20,7 % aller Proben kulturell
Salmonellen nachgewiesen. Dass die Nachweisrate von Salmonella spp. in der eige-
nen Verlaufsuntersuchung trotz Verwendung von Swabs auch nur bei 8,6 % lag,
spricht nicht dagegen, da dort auch Zeitpunkte und Lokalisationen mit sehr geringen
erwarteten Nachweisraten untersucht wurden.
5.1.3 Gemeinsames Auftreten der Erreger
5.1.3.1 Erregernachweise
Ein weiteres Anliegen der Studie war es, das gemeinsame Auftreten der drei Erreger
zu untersuchen. Dazu existieren bis dato nur ganz wenige Studien: VON ALTROCK
et al. (2006) analysierten Campylobacter spp. und Y. enterocolitica auf Einzeltier-
ebene. und FOSSE et al. (2011) Campylobacter und Salmonella spp. im Bestand.
Das Auftreten aller drei Bakterien wurde bei DÖHNE (2010) untersucht, allerdings
durch Probenentnahme auf dem Schlachthof und nicht im Bestand.
Das gemeinsame Auftreten von Erregern ist besonders deshalb von Interesse, da ein
unmittelbarer Rückschluss vom Salmonellenvorkommen auf das der anderen Erreger
eine Kontrolle dieser stark vereinfachen würde. Dies würde bedeuten, dass man kei-
ne eigenen Überwachungsprogramme für die beiden Erreger implementieren müsste,
sondern eine indirekte Einschätzung anhand des Salmonellen-Monitorings möglich
wäre. Zudem könnte auch auf gleiche Risikofaktoren und entsprechend gleiche Be-

Diskussion
144
kämpfungsstrategien spekuliert werden. Dann würden auch Einschätzungen bestä-
tigt, denen zufolge umfassende Maßnahmen zur Salmonellenbekämpfung auch ei-
nen positiven Effekt auf anderen zoonotische Keimbelastungen hätten (SKJERVE et
al. 1998; KAMPHAUSEN u. JAEGER 2009).
Insgesamt 40 Betriebe wiesen Campylobacter spp., 24 Betriebe Y. enterocolitica und
16 Betriebe Salmonella spp. auf. Die Analyse ergab, dass sechs Betriebe alle drei
Erreger gleichzeitig aufwiesen. Campylobacter spp. traten 16 Mal gemeinsam mit Y.
enterocolitica auf und sechsmal gemeinsam mit Salmonella spp. Ein Betrieb wurde
als positiv für Y. enterocolitica und Salmonella spp. eingestuft. Immerhin drei Betrie-
be wiesen nur Salmonella spp. auf, während das singuläre Auftreten von
Y. enterocolitica trotz deren generell häufigeren Auftretens lediglich bei einem Be-
trieb nachgewiesen wurde. Die Überprüfung des Grades der Übereinstimmung mit-
tels Kappa-Koeffizient ergab für Campylobacter spp. und Y. enterocolitica eine mittle-
re Übereinstimmung. Dies ist mehr als bei VON ALTROCK et al. (2006), die weder
anhand der serologischen, noch der kulturellen Ergebnisse einen Zusammenhang
feststellen konnten. Allerdings testeten diese auf Einzeltierebene, womit die geringe-
re Übereinstimmung erklärbar ist. DÖHNE (2010) konnte serologisch und kulturell
weder auf Einzeltier- noch auf Betriebsebene einen klaren Zusammenhang identifi-
zieren. FOSSE et al. (2011) erkannten ebenfalls keinen Zusammenhang.
In der vorliegenden Arbeit zeigten die Kombinationen mit Salmonella spp. keine we-
sentlich über den Zufall hinausgehende Kon- oder Diskordanz.
Bei diesen Berechnungen muss jedoch generell berücksichtigt werden, dass das Er-
gebnis dieses statistischen Tests stark von den Randverteilungen der zu prüfenden
Stichproben beeinflusst wird. Sobald in der Kontingenztafel auch nur eine Zelle jen-
seits der Diagonalen gefüllt ist, hängt der maximale Übereinstimmungswert von den
Randverteilungen ab. Er wird umso geringer, je weiter sich die Randverteilungen von
einer Gleichverteilung entfernen (BRENNAN u. PREDIGER 1981), hier also die An-
zahlen positiver und negativer Betriebe. So wird der niedrige Kappa-Koeffizient der

Diskussion
145
Kombinationen mit Salmonella spp. bereits durch die stark unterschiedlichen Anzah-
len positiver Betriebe verursacht.
Daher ergibt sich anhand dieser Berechnungen zwar ein Hinweis auf eine gewisse
Übereinstimmung zwischen Campylobacter spp. und Y. enterocolitica, jedoch keine
zuverlässige Aussage über die Art des gemeinsamen Erregerauftretens bei Salmo-
nella spp. Deshalb wurde zur weitergehenden Erörterung der Thematik die Salmo-
nellen-Kategorie der Betriebe betrachtet:
5.1.3.2 Kategorie
Die Salmonellen-Kategorisierung erlaubt, auch wenn keine Aussage über den Status
präsens des Erregers möglich ist, eine gute Einschätzung der allgemeinen Belastung
bzw. des Risikos eines Bestands für Salmonellen. Ein grundsätzlicher Vorteil ist zu-
dem, dass diese Daten generell für alle Betriebe erhoben werden und daher direkt
(ohne zusätzliche Untersuchungen) verfügbar sind.
Bei Salmonella spp. war aufgrund des direkten Kausalzusammenhangs eine positive
Abhängigkeit mit der Kategorisierung zu erwarten. Im Ergebnis waren zwar sowohl
mehr Proben als auch Betriebe der Kategorie III positiv (8,6 % zu 6 % Nachweisrate
und 9 zu 7 Betriebe), die Differenzen fielen hier jedoch wesentlich geringer und nicht
signifikant aus.
Dies liegt vermutlich im System der Kategorisierung begründet: Diese wird anhand
serologischer Untersuchungsbefunde rückwirkend für die vergangenen zwölf Monate
berechnet und vierteljährlich angepasst. Sie besagt also nur, dass in den letzten
zwölf Monaten ein bestimmter Anteil der untersuchten Schweine Erregerkontakt hat-
te, und nichts über die tatsächliche Belastung des Bestands mit Salmonellen am Un-
tersuchungstag. Daher ist es aus verschiedenen Gründen durchaus möglich, dass
die am Untersuchungstag festgestellte Salmonellenbelastung eines Bestands nicht
mit der Salmonellen-Kategorie übereinstimmt. Dies wird bestärkt durch den mit der

Diskussion
146
intermittierenden Ausscheidung einhergehenden gewissen „Zufallscharakter“ eines
bakteriologischen Befunds (KRANKER et al. 2003).
In Bezug auf die anderen beiden Erreger wurde ein deutlicherer Zusammenhang mit
der Kategorie festgestellt: Es fiel auf, dass in Kategorie I-Betrieben häufiger Y.
enterocolitica und Campylobacter spp. nachgewiesen wurden als in Kategorie III-
Betrieben. Auf Probenebene war dieser Unterschied sowohl bei Campylobacter spp.
(31,1 % zu 18,9 % Nachweise) als auch bei Y. enterocolitica (12,9 % zu 4,3 %
Nachweise) statistisch signifikant. Auch bei Betrachtung der Betriebsebene, bei der
eine Einstufung als positiver Betrieb bei mindestens einem Nachweis erfolgte, be-
stand dieser Unterschied fort. Er lag allerdings nur bei Y. enterocolitica innerhalb des
Signifikanzniveaus (19 zu 5 positive Betriebe). Bei Campylobacter spp. war das Ver-
hältnis 22 zu 18 Betriebe.
Ein saisonaler Zusammenhang konnte als unwahrscheinlich angenommen werden:
Die Untersuchungen der Kategorie I- und der Kategorie III-Betriebe verteilten sich
jeweils gleichmäßig über das ganze Jahr.
Die häufigeren Erregerfunde von Campylobacter spp. und Y. enterocolitica in Kate-
gorie I-Betrieben könnten folgende Ursachen haben:
Zunächst besteht bei der Stichprobengröße von 50 Betrieben stets ein Risiko, dass
die Unterschiede zwischen den Kategorien bei Campylobacter spp. und Y. enteroco-
litica zufällig bedingt sind. Ein Hinweis dagegen ist jedoch, dass auch DÖHNE (2010)
in ihrer Arbeit serologisch signifikant mehr Yersinien-Infektionen in Kategorie I-
Betrieben nachwies als in Kategorie III-Betrieben. Daher kann vermutet werden, dass
tatsächlich Unterschiede zwischen den Kategorien und den Betrieben bestehen. Dies
könnte, wie DÖHNE (2010) mutmaßt, durch unterschiedliche Risikofaktoren bedingt
sein, oder aber durch eine unterschiedliche Stärke ihres Einflusses.

Diskussion
147
Landwirte, deren Betrieb in Kategorie III eingestuft wurde, haben daraufhin häufig
bereits Maßnahmen zur Salmonellenbekämpfung, z. B. Verbesserungen im Hygie-
nemanagement oder einen Wechsel der Ferkelherkunft, eingeleitet. (Dieser „selec-
tion bias“ ergab sich dadurch, dass die Studie auf freiwilliger Teilnahme der Betriebe
basierte. Möglicherweise erklärten sich gerade in Kategorie III eher Landwirte dazu
bereit, die die Problematik ernst nahmen und Maßnahmen gegenüber offen stan-
den.) Diese Maßnahmen könnten sich möglicherweise unmittelbarer auf Y. enteroco-
litica und Campylobacter spp. auswirken als auf Salmonella spp., da jene Erreger
„näher am Tier“ verbleiben und eine geringere Anzahl potentieller Habitate in der
Tierumgebung aufweisen. Salmonellen, die eine viel höhere Überlebensfähigkeit au-
ßerhalb des Wirts aufweisen, können in einer viel größeren Anzahl unterschiedlicher
Reservoire auftreten, was viel komplexere und langwierigere Maßnahmen erfordert
(BLAHA 2008; BODE 2008).
Umgekehrt könnte dies ein Grund für die höheren Yersinia- und Campylobacter-
Nachweise in Kategorie I-Betrieben sein, die sich aufgrund der nicht vorhandenen
Salmonellenproblematik nicht zu gesonderten Anstrengungen im Hygienemanage-
ment etc. veranlasst sehen. Dies wird zusätzlich gestützt durch die Beobachtungen
bei Analyse der einzelnen Lokalisationsgruppen: In Betrieben der Kategorie I schie-
den nicht nur die Schweine häufiger Campylobacter spp. und Y. enterocolitica aus,
ein noch größerer Einfluss auf die Kategorie wurde für den Kontaminationsgrad der
Umgebung berechnet. Neben der tiernahen waren besonders die tierferne Umge-
bung und Schädlinge signifikant häufiger positiv. Dies betraf besonders die „neuralgi-
schen“ Lokalisationen Treibbrett, Stiefel, Gang und Zentralgang.
Eine weitere denkbare Möglichkeit besteht in einer kompetitiven Verdrängung zwi-
schen Salmonellen und den anderen Erregern im Bestand. Die Abklärung eines sol-
chen Zusammenhangs ist im Rahmen des vorliegenden Studientyps jedoch nicht
möglich, dies müsste in Infektionsversuchen o. ä. erfolgen.

Diskussion
148
5.1.4 Risikofaktoren
Für die Analyse potentieller Risikofaktoren wurden die Betriebe anhand ihrer Erre-
gernachweise einerseits in „positiv“ und „negativ“ eingeteilt, andererseits wurden ihre
jeweiligen Anzahlen der Erregernachweise als Zielgrößen betrachtet. Für die Be-
rechnungen wurden nur Faktoren herangezogen, die sich hinsichtlich ihrer Verteilung
dafür eigneten. Daher mussten einige Merkmale aus der Analyse ausgeschlossen
werden. Auf eine Einbeziehung antibiotischer Behandlungen in die Risikofaktoren-
analyse musste verzichtet werden. Für eine aussagekräftige Analyse wäre eine Viel-
zahl von Zusatzinformationen nötig gewesen wie Art und Anzahl der Antibiotika,
Dauer und Häufigkeit der Behandlung, Dosis, Indikation usw. Eine solch detaillierte
Aufteilung hätte bei der vorliegenden Stichprobengröße zu für eine statistische Aus-
wertung nicht ausreichenden Gruppengrößen geführt. Daher hätte eine starke Ver-
einfachung der Daten, z. B. nur „Antibiotika ja - nein“, erfolgen müssen, was aufgrund
des hohen Informationsverlustes eine nur geringe Aussagekraft dieses Vergleichs
erwarten ließ. Außerdem musste seitens der Landwirte teilweise mit ungenauen bis
unzutreffenden Angaben zu Behandlungen gerechnet werden, was die Zuverlässig-
keit einer solchen Auswertung zusätzlich in Frage gestellt hätte. Ein möglicher Ein-
fluss von Erkrankungen der Tiere zum Untersuchungszeitpunkt wurde durch Befra-
gung des Landwirtes zum aktuellen Gesundheitsstatus zu reduzieren versucht (nur
laut Aussagen des Landwirtes gesunde Bestände wurden untersucht).
Für Salmonella spp. ließ sich bei kaum einem Bestandsmerkmal ein signifikanter Ein-
fluss auf das Erregervorkommen nachweisen. So konnte für das Betriebssystem
(reine Mast oder geschlossenes System) kein Effekt belegt werden. Die wenigen
identifizierten Einflussfaktoren stimmten jedoch mit den Angaben in der Literatur
überein. Der protektive Einfluss einer konsequenten Reinigung und Desinfektion vor
jeder Neubelegung des Abteils konnte zumindest bedingt (im Wilcoxon-Test) bekräf-
tigt werden (BLAHA 2008). Auch zeigten Betriebe, die ihre Buchten bis in Deckenhö-
he reinigten und desinfizierten, weniger Erregernachweise. Der Einsatz von Propion-
säure wies dagegen zumindest eine Tendenz zu erhöhten Salmonella-Nachweisen
auf. Die häufigeren Erregernachweise in der tierfernen Umgebung in Betrieben ohne

Diskussion
149
Schadnagerbekämpfung untermauern die Rolle dieser als Überträger (LE MOINE et
al. 1987). Insgesamt kann nicht ausgeschlossen werden, dass vorrangegangene an-
tibiotische Behandlungen, die in der vorliegenden Untersuchung aus oben genannten
Gründen nicht berücksichtigt werden konnten, einen Einfluss auf die Studienergeb-
nisse gehabt haben könnten, z. B. im Hinblick auf den berechneten Einfluss von Ri-
sikofaktoren (dies gilt ebenso für die anderen Erreger).
Zwischen Campylobacter spp. und Y. enterocolitica ergaben sich, wie aufgrund des
häufiger gemeinsamen Auftretens zu erwarten war, einige Übereinstimmungen. Für
beide war das Risiko eines Erregernachweises bei wechselnden Ferkelherkünften
verringert, Dies steht zumindest für Y. enterocolitica im Gegensatz zu SKJERVE et al.
(1998), der dafür im Vergleich zu geschlossenen Systemen ein erhöhtes Risiko er-
rechnete. Für Campylobacter spp. verlief die Literaturrecherche dazu erfolglos. Im
Weiteren wiesen Betriebe, die mehrere Futtermittelzusatzstoffe bzw. Ameisensäure
einsetzten, für beide Pathogene geringere Nachweisraten auf. Diverse Hygienemaß-
nahmen waren erstaunlicherweise bei beiden Erregern mit höheren Nachweisraten
assoziiert. Campylobacter spp. im Betrieb traten außerdem häufiger bei Pelletfütte-
rung und Vollspaltenboden auf, Y. enterocolitica seltener bei Umstallen einzelner
Tiere zwischen den Mastgruppen.
Insgesamt fällt auf, dass für manche der Faktoren, die mit erhöhten Nachweisraten
für Campylobacter spp. und Y. enterocolitica einhergingen, in der Literatur eine pro-
tektive Wirkung gegenüber Salmonellen beschrieben wird. Dies ist konsistent zu der
Tatsache, dass die serologisch geringer Salmonellen-belasteten Kategorie I-Betriebe
häufiger Campylobacter spp. und Y. enterocolitica aufwiesen und kann wie oben be-
gründet, u.a. Effekt einer generellen Selektion von Betrieben in die Studienpopulation
sein (KREIENBROCK u. SCHACH 2005).
Das gilt für den negativen Effekt wechselnder Ferkelherkünfte (LO FO WONG et al.
2004), des Teilspaltenbodens und des Umstallens zwischen Mastgruppen (ROSSEL
et al. 2006), sowie den protektiven Effekt verschiedener Hygienemaßnahmen
(BELOEIL et al. 2004; HAUTEKIET et al. 2008). Hier könnten also für die Erreger

Diskussion
150
unterschiedliche Einflussfaktoren vorliegen. Andererseits könnte auch das von VAN
DER WOLF et al. (2001) bezüglich Salmonella spp. Gesagte auf Campylobacter spp.
und Y. enterocolitica übertragbar sein: Sie fanden heraus, dass eine ausschließliche
Reinigung der Buchten eine geringere Erregerprävalenz zur Folge hatte, als wenn im
Anschluss desinfiziert wurde. Dies könnte durch die dann nachlässigere Ausführung
beider Arbeitsschritte im Vergleich zu einer sehr gründlichen Reinigung liegen. Da-
neben könnte durch die gleichzeitige Abtötung der Pathogene samt der kompetitiven
Flora eine verstärkte Vermehrung ersterer bei Neueintrag stattfinden (MEYER et al.
2004).
Zudem geht eine Assoziation nicht zwingend mit einer Kausalität einher. Es kann
auch ein Scheinzusammenhang vorliegen („confounding bias“), der durch einen nicht
berücksichtigen bzw. nicht erkannten Drittfaktor („confounder“) bedingt ist
(KREIENBROCK u. SCHACH 2005). Dies könnte beispielsweise auf den Einflussfak-
tor der Personalhygienemaßnahmen (Anziehen von Stiefeln / Schutzkleidung etc.)
zutreffen, insbesondere, wenn man die praktische Umsetzung dieser Maßnahmen
abbilden will (Compliance). Die Nicht-Durchführung jeglicher Personalhygienemaß-
nahmen erwies sich in Bezug auf Campylobacter spp. als protektiv. So ist z.B. denk-
bar, dass Landwirte, die Stiefel / Schutzkleidung verwendeten, diese so jedoch selten
säuberten / wechselten, dass damit entgegen des gewünschten Effekts eher zu einer
Erhöhung der Erregerlast beitrugen. Zudem halten Landwirte ohne jegliche Schutz-
kleidung sich möglicherweise weniger lange im Stall und seltener in sehr schmutzi-
gen Bereichen (z.B. in den Buchten) auf und könnten dadurch weniger zur Erreger-
übertragung beitragen. Der confounder besteht hier also im Verhalten des Landwirts.
Eine weitere Möglichkeit, warum Studienergebnisse fehlerhaft sein können, ist die
Informationsverzerrung („information bias“). Die Betriebsdaten wurden durch Befra-
gung der Landwirte erhoben. Daher kann der Wahrheitsgehalt ihrer Aussagen in vie-
len Punkten nicht verifiziert werden, so dass den Berechnungen entweder unbeab-
sichtigt oder beabsichtigt falsche Informationen zugrunde liegen können. Ersteres
kann durch abweichende Definitionen einer Frage und deren Antwortmöglichkeiten

Diskussion
151
entstehen. So können Landwirte, die „bei starkem Befall“ eine Schadnagerbekämp-
fung durchführen, sehr unterschiedlicher Auffassung über das Vorhandensein eines
„starken Befalls“ sein. Während die sporadische Sichtung eines Nagers im Stall für
den einen Landwirt eine „normale“ Befallstärke darstellt, kann dies für den anderen
bereits ein stark erhöhtes Aufkommen bedeuten. Letzteres - eine absichtliche Fal-
schinformation - könnte möglicherweise bei sensiblen Fragestellungen eintreten. Hier
muss berücksichtigt werden, dass Schweine haltende Landwirte regelmäßig im
Rahmen von QS-Audits zu ihrem Betrieb befragt werden und somit auf die „richti-
ge“ Beantwortung bestimmter Fragen „trainiert“ sein könnten. Manche Landwirte ge-
ben möglicherweise nur ungern zu, dass sie nicht im konsequenten Rein-Raus-
Verfahren ihre Ställe belegen, jedes Mal eine komplette Reinigung und Desinfektion
der Abteile vor Einstallung vornehmen oder auf keinen Fall jemals Tiere umstallen.
In beiden Fällen wäre somit eine nicht-differentielle Fehlklassifikation durch die Be-
fragung denkbar, die zur einer Verzerrung führt, die bei nicht erfolgten Hygienemaß-
nahmen zu einem schützenden Effekt hinsichtlich des Campylobacter- bzw. Yersinia-
Befundes führen.
Unter diesen Punkt („information bias“) fällt auch die Sensitivität der Nachweisme-
thode: Gerade im Hinblick auf Salmonella spp. könnte unter Umständen bei einigen
Betrieben eine Fehlklassifikation bezüglich ihres Erregerstatus vorgelegen haben, die
auf der bei Kotproben stets vorhandenen Problematik der intermittierenden Aus-
scheidung, verbunden mit der nicht optimalen Entnahmetechnik bei Umgebungspro-
ben (Tupfer mit geringer Materialaufnahme) begründet ist (falsch-negative Klassifika-
tion). Dieser Prozess des nicht Erkennens einer Salmonellenbelastung dürfte aber
unabhängig von den untersuchten betrieblichen Einflussfaktoren sein. Damit besteht
die Möglichkeit der sogenannten nicht-differentiellen Fehlklassifikation, bei der die
Quoten der Klassifikationsfehler für beide zu vergleichende Gruppen gleich sind. Ein
solcher Fehler führt aber zu einer nach unten verzerrten Effektschätzung, also Unter-
schätzung von Odds-Ratios. So könnten betriebliche Einflussfaktoren, für die in die-
ser Studie kein Einfluss auf den Salmonellen-Status berechnet wurde (z. B. Zusam-

Diskussion
152
menstallen verschiedener Altersgruppen: OR = 1,071, KG: 0,175 - 6,56), in Wahrheit
ein höheres Odds-Ratio und somit einen Effekt aufweisen, der durch die Fehlklassifi-
kation allerdings verwischt wurde. Ebenso könnte der Effekt, den die Kategorie auf
den Salmonellenstatus eines Betriebes hat, unterschätzt worden sein.
Insgesamt konnte die Studie zeigen, dass Y. enterocolitica und Campylobacter spp.
zwar in sehr vielen Betrieben vorkommen, aber durchaus systematische Unterschie-
de vorliegen, wie die je nach Kategorie unterschiedlichen Ergebnisse andeuten. Dies
weist auf die generelle Möglichkeit einer Kontrolle der Erreger hin. WEIJTENS et al.
(1997) konnten die Nachweisraten in Mastbeständen durch die Verwendung von
spezifisch pathogen freien Tieren zwar nicht auf null absenken, jedoch beträchtlich
reduzieren. Voraussetzung dafür war ein striktes Hygienemanagement. Sie gingen
von einer Übertragung durch den Tierhalter selbst, Schadnager oder Wildvögel aus.
ALTER et al. (2005) wiesen in der Mast andere Campylobacter-Genotypen nach als
im Flatdeck. Zudem konnten sie Campylobacter spp. im gereinigten und desinfizier-
ten leeren Abteil vor Einstallung nachweisen. Auch dies belegt die Infektion während
der Mastperiode und damit die Möglichkeit, hier in der Bekämpfung anzusetzen.
NESBAKKEN et al. (2007) hielten zwar eine Campylobacter-Freiheit in Schweine-
herden für nicht erreichbar, eine Freiheit von Y. enterocolitica dagegen schon.
SKJERVE et al. (1998) stellten in geschlossenen Systemen geringere Nachweisraten
als in reinen Mastbetrieben fest. Sie hielten die Kontrolle von Y. enterocolitica durch
strikte Trennung infizierter und nicht-infizierter Tiere für möglich. WEHEBRINK et al.
(2008) und KASIMIR (2005) siedelten den Ansteckungszeitpunkt in der Mast an. Für
WEIJTENS et al. (2000) stellten Futter, Wasser und lebende Vektoren einschließlich
des Menschen permanente Risikofaktoren für Ferkel dar. Dies deutet darauf hin,
dass die Mast Angriffspunkte zur Bekämpfung der Erreger bietet. Die vorliegende
Studie zeigt einige Ansätze dazu auf.
Die Erwartungen, dass ein Auftreten dieser beiden Erreger im Bestand positiv mit
dem von Salmonella spp. assoziiert sei und gleiche Risikofaktoren vorlägen, konnten
mit den Untersuchungen in dieser Studie nicht bestätigt werden. Damit konnten die

Diskussion
153
Vermutungen einiger Autoren (SKJERVE et al. 1998; KAMPHAUSEN u. JAEGER
2009), wonach Sanierungsmaßnahmen z. B. für Salmonella spp. sich auch auf ande-
re zoonotische Erreger auswirkten, nicht belegt werden. Dies ist ein sehr wichtiger
Aspekt vor dem Hintergrund, dass im Zuge des Verbraucherschutzes umfassende
Monitoringsysteme gefordert sin, die sich nicht nur auf die Kontrolle einzelner Erreger
wie zur Zeit Salmonellen beschränken, sondern auf andere Zoonoseerreger erwei-
terbar sein müssen (VON ALTROCK u. WALDMANN 2003).
5.2 Verlaufsuntersuchung
In dieser Untersuchung sollte das Salmonellenvorkommen in der Schweinemast im
zeitlichen und räumlichen Verlauf genauer untersucht werden. Dazu wurde nicht nur
die Dynamik von Salmonella-Nachweisen allgemein analysiert. Zusätzlich wurde
auch von den immer weiter entwickelten Möglichkeiten der Feintypisierung von Erre-
gern Gebrauch gemacht, so wie es im Zuge einer modernen Aufarbeitung von Infek-
tionsgeschehen vermehrt gefordert wird (LIEBANA 2002). Dazu wurden alle Salmo-
nellen mithilfe eines innovativen molekularen Fingerprintings bis auf klonale Ebene
feintypisiert. So konnte auch das Auftreten und Verhalten einzelner Stämme unter-
sucht werden. Daraus wurden vertiefende Kenntnisse und Rückschlüsse über den
Stellenwert einzelner Eintragsquellen und Schwachstellen im Betriebsmanagement
erwartet, die der Aufdeckung von Ansatzmöglichkeiten einer noch effektiveren Be-
kämpfung im Bestand dienen sollten.
Es wurden zwölf Bestände ausgewählt, die in ihrem Aufbau und Management als
typisch für die Region Nordwestdeutschland angesehen wurden.
Die Auswahl der Bestände erfolgte anhand der Salmonellen-Kategorie. Wichtig war,
dass auch bei den letzten Untersuchungen des Kategorisierungszeitraumes Salmo-
nellennachweise vorlagen.

Diskussion
154
In vier Betrieben verliefen die Untersuchungen auf Salmonellen erfolglos, in drei Be-
trieben gelangen nur ganz wenige (1 - 6) Salmonella-Nachweise, bei zwei weiteren
Betrieben lag die Nachweisrate bei 7 %, bei den übrigen drei über 10 %. Dies zeigt,
dass selbst nach wiederholter Beprobung erheblich variierende Nachweisraten zwi-
schen den Betrieben vorliegen können. Insgesamt lag der Anteil positiver Proben mit
8,6 % in der gleichen Größenordnung wie bei den Kategorie-III-Betrieben in der
Querschnittsuntersuchung. Dabei muss berücksichtigt werden, dass hier auch Loka-
lisationen und Zeitpunkte untersucht wurden, an denen erfahrungsgemäß eher selte-
ner Salmonellen zu erwarten sind (z. B. gereinigtes und desinfiziertes Abteil).
Für die Auswertung herangezogen wurden die Ergebnisse der kulturellen Untersu-
chung. Zuvor war geprüft worden, ob mit der PCR andere Resultate erzielt worden
wären: Mit der Auswertung konnte gezeigt werden, dass die beiden Methoden bei
geringfügig höherer Sensitivität der PCR eine sehr gute Übereinstimmung aufwiesen.
Auch die zwischen den einzelnen Lokalisationsgruppen differenzierende Analyse
ergab, dass die Probenherkunft keinen signifikanten Einfluss auf die Abweichungen
zwischen den beiden Verfahren hatte. Daher wurde im Zuge der besseren Aussage-
kraft über die Vermehrungsfähigkeit der Erreger die kulturelle Methode favorisiert.
Der Schwerpunkt der Untersuchung lag auf der Identifizierung und Bewertung von
Mustern und Gemeinsamkeiten von Salmonelleninfektionen im Bestand, ohne Be-
rücksichtigung der Verhältnisse in einzelnen Betrieben. Dazu wurden in einer be-
triebsübergreifenden Analyse zunächst alle Salmonellennachweise auf Speziesebe-
ne ausgewertet, dies wurde durch eine Feintypisierung der Isolate komplementiert.
Für alle Analysen, sowohl auf Proben- als auch auf Klonebene, muss grundsätzlich
berücksichtigt werden, dass nur der Nachweis Aussagekraft hat, nicht jedoch ein ne-
gatives Ergebnis. Zum einen liegt dies in der intermittierenden Ausscheidung der
Salmonellen durch den Kot begründet. Andererseits kann auch bei gewissenhafter
Probenentnahmetechnik nicht garantiert werden, dass zu jedem Zeitpunkt stets iden-
tische Flächen beprobt wurden. Manche Stallgeräte waren nicht zu jeder Untersu-
chung auffindbar oder könnten zwischenzeitlich gesäubert worden sein.

Diskussion
155
5.2.1 Betriebsübergreifende Analyse
5.2.1.1 Eintragsquellen und zeitliche Verläufe
Die Nachweisraten waren im ersten Durchgang im Mittel höher als in den weiteren.
Dies zeigt eine betriebsübergreifend abnehmende Tendenz im Verlauf mehrerer
Durchgänge, was womöglich darin begründet ist, dass der Beginn der Untersuchun-
gen genau zu einem Zeitpunkt stattfand, an dem das Problem für die Landwirte ob-
jektiv oder auch subjektiv akut war. Dass Änderungen im Betriebsmanagement wäh-
rend der Durchgänge für den Rückgang der Nachweishäufigkeit verantwortlich waren,
konnte selten festgestellt werden. Teilweise wurden von den Landwirten gar keine
Umstellungen vorgenommen bzw. wenigstens nicht angegeben. Möglicherweise
könnte es an einer unbewusst gewissenhafter ausgeführten Stallhygiene gelegen
haben. Einzelheiten dazu werden bei einzelnen Betrieben diskutiert.
Statistisch konnte kein signifikanter Unterschied bei den Nachweisen im leeren zu
denen im belegten Abteil nachgewiesen werden. Das bedeutet, dass der Kontamina-
tionsgrad bereits im leeren, gereinigten und desinfizierten Abteil ebenso hoch war
wie während der ganzen Zeit der Anwesenheit der Schweine. Dieser Umstand be-
weist ein in vielen Fällen defizitäres Hygienemanagement in den untersuchten Be-
trieben und unterstreicht die Bedeutung der Zirkulation von Salmonellen im Bestand
für die Infektionsdynamik, wie bereits von BLAHA (2008) und BODE (2008) be-
schrieben. Das Ergebnis bestätigt, dass auf dem Gebiet der Hygiene noch große
Verbesserungen nötig sind und die Landwirte weiterer Unterstützung und Aufklärung
bedürfen.
Bei den Sammelkotproben war dagegen ein zeitlicher Verlauf erkennbar: Höher als
zu allen anderen Probenentnahmezeitpunkten fiel das Ergebnis der Probenentnah-
me von den Ferkeln auf dem Transport-LKW bei Anlieferung aus: Fast ein Drittel die-
ser Sammelkotproben war bakteriologisch positiv. Dafür können unterschiedliche
Gründe infrage kommen: Manche Studien bemessen der Übertragung von der Sau
auf die Ferkel kaum Bedeutung zu (KRANKER et al. 2003; NOLLET et al. 2005),

Diskussion
156
andere sehr wohl (MERIALDI et al. 2008; VIGO et al. 2009; WILKINS et al. 2010). In
jedem Falle dürfte der Transportstress die erhöhte Ausscheidung induziert haben,
wie METHNER et al. (2008) feststellten. Immerhin ermittelte die EFSA 2008 eine
Herdenprävalenz von über 20 % für deutsche Ferkelerzeugerbestände. Möglich wäre
auch eine horizontale Ansteckung während der Flatdeckphase, ferner könnte auch
das Transportfahrzeug kontaminiert gewesen sein. Die Frage, wo die Tiere sich an-
gesteckt hatten, kann im Rahmen dieser Studie nicht beantwortet werden. Für das
Stichprobenkollektiv kann jedoch festgestellt werden, dass ein beträchtlicher Anteil
der Ferkelpartien mit dem Erreger infiziert war und diesen in die Mast eintrug. Dies
stützt die Erkenntnisse der EFSA im Rahmen einer quantitativen Risikoabschätzung
zu Salmonellen für Zucht- und Mastschweine (ANONYMOUS 2010b): Diese schätzt
für den Fall der Salmonellenfreiheit bei Zuchtschweinen eine Reduktion in der Prä-
valenz von Schlachtschweinen um 70 - 80 % für „high prevalence“-Länder. Damit
misst sie der vertikalen Übertragung einen sehr hohen Stellenwert bei. Der Abfall der
Nachweise im Verlauf der Mast könnte des Weiteren auch auf die Wirkung von Anti-
biotika im Zuge der in den meisten Betrieben durchgeführten metaphylaktischen Be-
handlung zur Einstallung zurück zu führen sein.
Um Hinweise über die Höhe des jeweiligen Einflusses dieser Transmissionsrouten zu
erlangen, wurde untersucht, bei welchem Szenario im belegten Abteil die meisten
Salmonella-Nachweise resultierten: Bei der Hälfte der 20 Mastdurchgänge gelang ein
Salmonellennachweis erst im belegten Abteil. Aus diesen Durchgängen stammten
jedoch nur 22 % der positiven Proben. Dies könnte bedeuten, dass der schon vorher
vorhandene Erreger zufällig nicht entdeckt wurde. Diese Gefahr besteht selbst bei
intensiver Beprobung immer, daher sind nur positive Ergebnisse beweisend, wäh-
rend negative keine definitive Aussage erlauben. Andererseits könnte ein Neueintrag
von außerhalb oder ein Kontakt mit einem Salmonellenreservoir erfolgt sein. Um ge-
naueren Aufschluss über die Zusammenhänge zu erhalten, wurde eine Analyse der
Lokalisationen der einzelnen Klone vorgenommen:

Diskussion
157
Diese Auswertung ergab, dass in den entsprechenden Durchgängen sehr häufig nur
die tierferne Umgebung positiv war. Ähnliches spiegelte sich auch bei Betrachtung
des Auftretens bestimmter Klone innerhalb der Durchgänge wider, wo beinahe alle
Klone mit Erstnachweis im belegten Abteil dort nur in der tierfernen Umgebung auf-
traten, und hier besonders an prädestinierten Lokalisationen wie Gängen, Stiefeln
oder Treibbrettern. Diesen Lokalisationen ist gemein, dass sich hier „Übertragungs-
wege kreuzen“ und sie für indirekte Kontaktmöglichkeiten zwischen einzelnen Sal-
monellenreservoiren sorgen.
Diese Ergebnisse könnten ein Indiz dafür sein, dass bereits an anderen Stellen im
Betrieb, z. B. in einer anderen Mastgruppe vorhandene Salmonellen weiterverbreitet
wurden. Dies belegt das immense Gefahrenpotential, das von diesen unbelebten
Vektoren für eine Verteilung der Erreger im Bestand ausgeht. Es verdeutlicht, wie
wichtig eine gute Hygiene auch während der Mast ist, um die Zirkulation von Salmo-
nellen zu unterbinden. Es betont darüber hinaus die Wichtigkeit einer strikten Tren-
nung zwischen verschiedenen Tier- oder Altersgruppen.
In einem Durchgang war zusätzlich ein Nager mit einem Klon kontaminiert, der vor-
her noch nie in diesem Bestand nachgewiesen worden war. Folglich könnte dies ein
Hinweis auf einen tatsächlichen Neueintrag von außen sein.
In insgesamt neun Durchgängen wiesen die Ferkel auf dem LKW und in sieben
Durchgängen das leere Abteil Salmonellen auf:
Davon wurden in drei Durchgängen Salmonellen lediglich im leeren Abteil detektiert,
aus diesen stammte ein Viertel aller positiven Proben aus belegten Abteilen. Hier
waren nach Reinigung und Desinfektion immer die tierferne und zweimal auch die
tiernahe Umgebung betroffen, später dann verschiedene Gruppen. Meist stimmten
auch die Klone überein, nur ein Klon eines Treibbretts wurde später im belegten Ab-
teil nicht mehr nachgewiesen.

Diskussion
158
In einem Durchgang war nur der Sammelkot der angelieferten Ferkel positiv, dieser
Durchgang trug mit 2,7 % zu allen positiven Proben belegter Abteile bei. Dort waren
im Anschluss nur tierferne Lokalisationen (Gang und Treibbrett) sporadisch positiv
und es handelte sich nicht um denselben Klon.
In einem Drittel aller Durchgänge waren sowohl die Ferkel als auch das leere Abteil
vor Anlieferung positiv. Aus diesen Durchgängen stammten überproportional viele
positive Proben, die während der anschließenden Mast entnommen wurden: über die
Hälfte. In diesen Durchgängen waren vor Einstallung meist Proben der tiernahen
Umgebung positiv, während der Mast stets eine Vielzahl unterschiedlicher Lokalisati-
onen. Auch die Klone, sowohl aus LKW-Kot als auch dem Abteil, fanden sich meist
wieder, nur bei zwei Klonen aus der tierfernen Umgebung war dies nicht der Fall.
Dies zeigt, dass eine defizitäre Reinigung und Desinfektion vor Einstallung sich stark
auf das Salmonellenaufkommen während der Mast auswirkt. Sie hatte dort stets ho-
he Gesamtnachweise zur Folge und die Klone hatten sich in Tierumgebung und den
Tieren selbst verbreitet. Dies stimmt mit den Beobachtungen von MAGISTRALI et al.
(2008) überein, wo in einer Longitudinalstudie mit Typisierung gleiche Isolate im lee-
ren gereinigten und desinfizierten Abteil wie auch später bei den dort eingestallten
Tieren nachgewiesen wurden.
Der alleinige Eintrag mit den Ferkeln beeinflusste dagegen weniger stark die Ge-
samtbelastung in der laufenden Mastperiode.
Über die Gründe, warum dies überhaupt nur in einem Durchgang der Fall war, an-
sonsten das kontaminierte leere Abteil stets in Kombination mit positiven Ferkeln auf-
trat, kann spekuliert werden. Es könnte bedeuten, dass häufig der primäre Eintrag in
einen Bestand durch die Ferkel stattfindet und dieser Keim fortan persistiert. So wur-
de in drei verschiedenen Durchgängen ein identischer Klon sowohl bei den Ferkeln
vor Anlieferung als auch im leeren Abteil detektiert. Daraufhin kann sich die Salmo-
nellenbelastung bei mangelnder Hygiene in den folgenden Durchgängen langsam

Diskussion
159
aber stetig erhöhen (Zirkulation). Diese Hypothese wird durch die Beobachtung ge-
stützt, dass der stärkste Effekt, d.h. die meisten Salmonellennachweise, in Durch-
gängen festgestellt wurde, in denen Salmonellen über mehrere Wege in die Mast
gelangten.
Diese Schlussfolgerungen zu Hygienemanagement, Eintragsquellen und Zirkulation
im Bestand konnten bei genauerer Betrachtung einzelne Klone bestätigt werden:
So war ein weiterer Hinweis für die länger andauernde Persistenz von Salmonellen
im Bestand, dass die Mehrheit der Klone in mehreren Durchgängen je Betrieb vor-
kam, drei sogar in allen drei Mastdurchgängen. Dabei war nicht ausschlaggebend,
wo und wann der Klon im ersten Durchgang zuerst auftrat. Auffällig war aber, dass
die wenigen Klone mit Erstnachweis im LKW-Sammelkot im nächsten Durchgang
besonders weit verbreitet waren. Klone, die nach Reinigung und Desinfektion oder im
belegten Abteil erstmalig auftraten, wurden dagegen im folgenden Durchgang spora-
discher und eher tierfern nachgewiesen. Das gleiche galt für Klone, die ganz neu,
also ohne Nachweis in vorigen Durchgängen auftraten. Ein Klon „übersprang“ einen
Durchgang, er wurde im ersten Durchgang sehr häufig und im dritten sporadisch an
Gang und Buchtenwänden nachgewiesen.
Umgekehrt waren alle Klone aus den Reinigungs- und Desinfektions-Proben bereits
im vorigen Durchgang nachweisbar vorhanden, aber auch zwei Klone, die bei den
Ferkeln vor Anlieferung nachweisbar waren, waren bereits im vorigen Mastdurch-
gang präsent gewesen.
Ähnliches wurde auch bei Betrachtung der Klone innerhalb eines Durchgangs deut-
lich:
Sehr häufig wurden Klone im Durchgang übereinstimmend vor und nach Einstallung
nachgewiesen. Wenn dies nicht der Fall war, traten sie während der Mast häufig an
Lokalisationen wie Gängen, Stiefeln, Treibbrettern oder Schädlingen auf.

Diskussion
160
Gemessen an den Anzahlen der Nachweise wurden Klone aus dem LKW-Kot wäh-
rend der Mast mit sehr hohen nachgewiesenen Anteilen isoliert, Klone aus dem lee-
ren Abteil eher sporadisch.
Diese Ergebnisse bestätigen die Annahme des von Salmonella-Residuen ausgehen-
den Zirkulierens. Es zeigt sich, dass Salmonellen, sobald erst einmal eingetragen,
häufig über mehrere Durchgänge im Bestand persistieren. Außerdem bestätigt es die
Bedeutung eines Primäreintrags durch die Ferkel, wonach gerade diese neuen Klone
sich erst langsam, aber dann sehr vehement im Bestand ausbreiten, während resi-
dente Keime eher sporadisch auftreten. Im Übrigen wurde dadurch demonstriert,
dass Ferkel häufig immer wieder dieselben Klone eintragen.
Zusammenfassend lässt sich feststellen, dass der Eintrag von Salmonellen durch die
Ferkel zwar auf die Gesamtanzahl positiver Proben im gleichen Mastdurchgang kei-
ne starke Wirkung hat. Die Klone allerdings kamen in den folgenden Durchgängen
sehr vehement vor. Umgekehrt führten Salmonellennachweise im leeren, gereinigten
und desinfizierten Abteil zu allgemein höherem Salmonellenvorkommen im gleichen
Mastdurchgang. Die einzelnen Klone jedoch traten in den folgenden Durchgängen
eher sporadisch auf.
Die beschriebenen Beobachtungen wurden bei genauerer Analyse der involvierten
Lokalisationen untermauert:
5.2.1.2 Rolle der Umgebung
Die Studie konnte die große Bedeutung der Tierumgebung als Reservoir für Salmo-
nellen (BLAHA 1993; BÖHM 1993) untermauern. Die am häufigsten positive Proben-
art war der Sammelkot, dahinter folgten Proben der tierfernen Umgebung. Hier wa-
ren mehr Proben positiv als in der tiernahen Umgebung. Dies unterstreicht die wich-
tige Rolle von Salmonellenresiduen, die nicht in regelmäßigem Kontakt zu den Tieren
stehen und daher von den Landwirten regelmäßig unterschätzt werden. Die hinter

Diskussion
161
dem LKW-Sammelkot mit Abstand am häufigsten positive Lokalisation war der
Zentralgang. Dies unterstreicht das Gefahrenpotential, bei mangelnder Hygiene Sal-
monellen von dort weiterzutragen. Das Gleiche gilt für Treibbretter und Stiefel, wo
ebenfalls mehrfach Salmonellen nachgewiesen wurden. Auch an Lüftern und Rohr-
leitungen im Abteil wurden regelmäßig Salmonella spp. nachgewiesen. Diese Ge-
genstände finden bei der Reinigung und Desinfektion des Abteils selten Berücksich-
tigung, da die Tierhalter hier keine Kontamination vermuten. Sie stehen zwar nicht in
direkten Kontakt zu den Tieren, befinden sich aber im Abteil, so dass der Erreger
durch von dort herunterfallendem Staub zu den Tieren gelangen kann. Zusätzlich
können Schädlinge eine Verbreitung des Erregers von dort bewirken.
Bei Betrachtung des zeitlichen Verlaufes wird ähnliches deutlich: Die Studie konnte
zeigen, dass in der tierfernen Umgebung vor Einstallung signifikant mehr Nachweise
gelangen als in der tiernahen, während in der Mast keine Unterschiede belegbar wa-
ren. Die am häufigsten positiven Lokalisationen nach Reinigung und Desinfektion
waren Treibbretter und Gänge (je viermal); Rohrleitungen, Trennwände, Buchtenbo-
den und Arbeitsräume waren je dreimal positiv. Jeweils einmal wurde der Erreger
von Lüftung, Futtertrögen, Außenwänden, und Zentralgang isoliert.
Zudem waren tiernahe Lokalisationen vor Mastbeginn statistisch seltener positiv als
währenddessen. Hier hatten die Landwirte offenbar eine ordnungsgemäße Reinigung
und Desinfektion durchgeführt. In der tierfernen Umgebung wurden dagegen keine
signifikanten Unterschiede in der Nachweisrate vor und nach Einstallung berechnet,
ihr Anteil war im leeren Abteil sogar am höchsten. Dies bestätigt die Vernachlässi-
gung dieser Lokalisationen und Gegenstände im Hygienemanagement zusätzlich.
Sehr häufig wiesen auch Nager Salmonellen auf. Da diese Tiere sich meist frei zwi-
schen den Ställen und Abteilen bewegen können, stellen auch sie ein ernstzuneh-
mendes Risiko für die Übertragung von Salmonellen im Bestand dar (LE MOINE et al.
1987; SKOV et al. 2008). Dies betont die Bedeutung einer wirksamen Schadna-
gerbekämpfung. Meist wurde der Erreger bei den Tieren im belegten Abteil nachge-

Diskussion
162
wiesen und die Klone waren schon vorher an anderen Lokalisationen präsent. Ein-
mal wurde jedoch ein völlig neuer Klon dort zum ersten Mal nachgewiesen. Dies
könnte für einen Neueintrag von außen sprechen.
Bei Betrachtung der Klondiversität war diese in der tierfernen Umgebung am höchs-
ten. Fast alle auftretenden Klone dieser Studie wurden dort nachgewiesen. Auch ge-
rade Klone mit nur wenigen Nachweisen traten fast ausnahmslos hier auf, während
sehr häufige Klone sich gleichmäßig über alle Gruppen verteilten. In der tiernahen
Umgebung traten etwas weniger verschiedene Klone auf, im Sammelkot kam nur die
Hälfte aller Klone vor (auch wenn dies der intermittierenden Ausscheidung geschul-
det sein könnte). Dies ist ein weiterer Hinweis auf die wichtige Bedeutung der
Tierumgebung als Reservoir. Bei Schädlingen war die Diversität gering: Schadnager
wiesen zwei Klone auf, Fliegen einen.
Die Analyse der Lokalisationen und -gruppen, an denen die Klone vor und nach Ein-
stallung bzw. von einem zum anderen Durchgang nachgewiesen wurden, zeigte kei-
ne einheitlichen Verteilungsmuster. Vielmehr ergaben sich alle möglichen „Vorher-
Nachher“-Kombinationen: Manche Klone verblieben an denselben Lokalisationen,
andere wurden an völlig anderen Orten nachgewiesen. Beispielsweise fanden sich
Klone der tierfernen Umgebung später an unterschiedlichsten Orten inklusive Sam-
melkot und Schädlingen wieder.
Interessanterweise wurden vier Klone aus dem Sammelkot nur vor Einstallung der
Ferkel nachgewiesen, aber dort zu späteren Zeitpunkten nicht wieder isoliert. Umge-
kehrt traten vier Klone des Sammelkots nur während der Mast auf. Lediglich zwei
Klone wurden übereinstimmend vor und nach Einstallung bei den Tieren nachgewie-
sen. Damit wird die Feststellung einer stark intermittierenden Ausscheidung durch
KRANKER et al. (2003) bestätigt, bei denen in einer Verlaufsuntersuchung nur 3,7 %
der Tiere mehr als einmal bakteriologisch Salmonellen im Kot aufwiesen. Ein weite-
rer Grund dafür könnte die in den meisten Betrieben durchgeführte metaphylaktische
Antibiotikagabe zur Einstallung sein, die möglicherweise die Salmonellen-

Diskussion
163
Ausscheidung zeitweise unterdrückte. Ausgehend von Sammelkot wurden die Klone
dafür später sehr oft in der Umgebung nachgewiesen. Insgesamt fand also eine star-
ke Verteilung der Erreger in Abteil und Bestand statt.
Eine Einbeziehung von betrieblichen Risikofaktoren bei der Analyse der Infektions-
dynamiken wurde aufgrund der geringen Stichprobenzahl, verbunden mit geringer
Variabilität in den Merkmalen (Tabelle 23), nicht durchgeführt. Auch bei Betrachtung
der einzelnen Betriebe konnte nicht immer ein Zusammenhang zwischen Infektions-
dynamik und betrieblichen Einflussfaktoren hergestellt werden, in manchen Fällen
ergaben sich jedoch Hinweise darauf:
5.2.2 Beschreibung der einzelnen betrieblichen Verläufe
In den einzelnen Betrieben zeigten sich recht unterschiedliche Infektionsmuster und -
verläufe.
In vier Betrieben gelang trotz deren Einordnung in Kategorie III während eines bzw.
zwei Durchgängen kein kultureller -Nachweis von Salmonella spp.. Bei allen zeigte
sich nach Abschluss der Beprobungen, dass sie sich in der Kategorie verbessert hat-
ten. Drei Betriebe waren mittlerweile mit niedrigen Werten in Kategorie II, einer sogar
in Kategorie I eingestuft. Die Gründe dafür sind unklar. Es waren keine Unterschiede
in Management und Haltung oder Gesundheitsstatus zu anderen Betrieben erkenn-
bar, und die Landwirte gaben an, innerhalb des Mastdurchganges keine Verände-
rungen vorgenommen zu haben. Möglicherweise hatten sie ihre Reinigung und Des-
infektion intensiviert und konnten dadurch die Infektionskette von einem zum nächs-
ten Mastdurchgang unterbrechen. Es belegt allerdings, dass zwischen Salmonellen-
vorkommen im Bestand und der Salmonellen-Kategorie in der Tat ein Zusammen-
hang besteht. Damit wird jedoch auch gezeigt, dass eine gewisse „Trägheit im Sys-
tem“ besteht. Da die Kategorisierung retrospektiv über zwölf Monate erfolgt, reagiert
sie erst verzögert auf eine Reduktion der Salmonellenbelastung. So werden wieder-
holt negative Probenentnahmen in präsumtiv hoch belasteten Beständen erklärbar.

Diskussion
164
Dies könnte zur Erklärung der relativ wenigen Nachweise in den Kategorie III-
Betrieben der Querschnittsuntersuchung beitragen.
Zwischen den Betrieben mit Nachweisen von Salmonellen gab es ebenfalls Unter-
schiede in der Höhe des Salmonellenvorkommens, auch hier stimmte die Entwick-
lung der Kategorie jeweils zeitversetzt mit den Verläufen der bakteriologische Befun-
de überein.
Ein Betrieb (B) verzeichnete lediglich einen Nachweis, zum Anfang der Mast am
Treibbrett. Der Erreger verbreitete sich von dort nicht, obwohl der Betrieb nach eige-
nen Angaben weder für jeden Stall eigene Geräte hatte, noch diese nach jeder Be-
nutzung oder vor Neubelegung reinigte und desinfizierte. Der Betrieb war nach Ende
der Untersuchung in Kategorie II.
Ein weiterer Betrieb (D), in dem vier Nachweise von zwei Klonen im ersten Durch-
gang gelangen, war im zweiten Durchgang komplett negativ. Hier waren im ersten
Durchgang eklatante Mängel in der Sauberkeit des Abteils nach Reinigung und Des-
infektion und starke Schadnagerpräsenz festgestellt worden, auch wenn bei beidem
keine Salmonellen detektiert wurden. Im zweiten Durchgang waren immer noch Na-
ger vorhanden, jedoch wurde die Sauberkeit subjektiv als erheblich verbessert emp-
funden. Auch dieser Betrieb war Ende des ersten Durchgangs in Kategorie III, nach
Ende des zweiten Durchgangs in Kategorie II eingestuft.
Betrieb E wies ebenfalls wenige Salmonellenfunde auf, hier fiel jedoch eine starke
Diversität (4 verschiedene Klone in 5 positiven Proben, alle im Zentralgang) auf. Der
Betrieb hatte in beiden Durchgängen aufgrund von Atemwegserkrankungen und Di-
arrhö mehrfach Amoxycillin, Lincomycin und Tylosin verabreicht. Ihr Einfluss auf die
Salmonellen im Betrieb bleibt unklar. Die Diversität der Klone auf den Treibwegen
könnte auf die Größe des Stalls, die große Anzahl verschiedener Altersgruppen und
die Bauart, wonach alle Abteile auf den Zentralgang führten, zurückzuführen sein.

Diskussion
165
Dennoch war die Nachweisrate gering und der Betrieb verbesserte sich nach Ende
des zweiten Durchgangs zur Kategorie II.
Betrieb F wies im ersten Durchgang sehr viele Salmonellen auf, im dritten dagegen
nur noch wenige. Ende des ersten Durchgangs gelangte der Betrieb in Kategorie II,
Ende des zweiten Durchgangs war er bereits in Kategorie I. Der Bestand wies im
ersten Durchgang bereits im leeren Abteil, auf dem LK und anschließend während
der Mast häufig Salmonellen auf. Zu Anfang der Mast wurden Tetrazyklin und Sul-
fadiazin-Trimethoprim, später wegen einer Atemwegserkrankung Amoxycillin verab-
reicht. Dies Behandlung könnte zum hohen Salmonellenaufkommen beigetragen
werden, da eine Immunsuppression vorlag, Antibiotika die Salmonellenausscheidung
verlängern können (WALDMANN u. PLONAIT 2004) und einige der Isolate Resisten-
zen gegenüber Ampicillin und Tetrazyklin aufwiesen. Im zweiten Durchgang, indem
etwas weniger Nachweise gelangen, waren die Tiere weder krank, noch erhielten sie
Antibiotika. Auch hatte sich gegenüber dem ersten Durchgang subjektiv die Reini-
gung und Desinfektion stark verbessert, so dass nur noch das Treibbrett Salmonellen
aufwies. Im dritten Durchgang, in dem die Tiere zu Mastbeginn eine Metaphylaxe mit
Tetrazyklin und Sulfadiazin-Trimethoprim erhielten, aber im weiteren Verlauf keine
Krankheiten aufwiesen, traten nur sporadisch Salmonellen auf.
Betrieb H, eher sporadisch Salmonella-positiv, bewegte sich von Kategorie II Ende
des ersten über Kategorie I Anfang des dritten zu Kategorie II Ende des dritten Mast-
durchgangs. Trotz eines Nachweises im Ferkelkot bei Anlieferung trat dieser Klon im
weiteren Verlauf des ersten Durchgangs nicht mehr auf und nur die tierferne Umge-
bung war ganz selten positiv. Im zweiten Durchgang trat ebenfalls ein Klon einmal im
LKW und danach im Durchgang nicht wieder auf. Überhaupt war der zweite Durch-
gang trotz Salmonellennachweisen im leeren Abteil während der Mast nicht positiv.
Der Klon aus LKW-Sammelkot des zweiten Durchgangs (Klon Nr. 12) trat dann im
belegten Abteil des dritten Durchgangs einige Male in der tierfernen Umgebung auf.

Diskussion
166
Genau dieser Klon (12) wurde gleich zu Beginn der Untersuchungen im Betrieb L
nach Reinigung und Desinfektion nachgewiesen und trat dort auch in den folgenden
Durchgängen regelmäßig, sogar bei den Ferkeln vor Einstallung, auf. Der Zusam-
menhang war zunächst unklar, da keine Gemeinsamkeiten zwischen den Betrieben
(z. B selber Futterlieferant, Tierarzt etc.) gefunden wurden. Eine Übertragung durch
die Untersucherin selbst war unwahrscheinlich, da die Betriebe meist nicht am glei-
chen Tag untersucht wurden, stets neue Schutzkleidung verwendet wurde und die
Proben getrennt bearbeitet wurden. Durch Nachforschungen wurde bekannt, dass
Betrieb L, der einzige Betrieb im geschlossenen System, überschüssige Ferkel an
einen Ferkelhändler verkaufte. Über eben diesen Händler bezog Betrieb H seine
Ferkel, die generell aus wechselnden Herkünften stammten. Möglicherweise wurden
Betrieb H Ferkel von Betrieb L geliefert oder auf einem Transportfahrzeug war eine
Kreuzkontamination zwischen Ferkeln erfolgt. In Betrieb L wurde bei hoher Nach-
weisrate eine generell geringe Klondiversität festgestellt. Hier wurden nur zwei ver-
schiedene Klone nachgewiesen. Dies zeugt von einer stärkeren Stabilität im Erreger-
vorkommen in geschlossenen Systemen. Bereits die Ferkel auf dem Flatdeck waren
nachweislich mit dem Erreger infiziert. Der Tierhalter betrieb nur geringen Hygiene-
aufwand im Betrieb: keine regelmäßige Reinigung und Desinfektion von Geräten,
häufiges Umstallen zwischen den Mastgruppen und Vorhandensein von Restbestän-
den, kein aus der Bucht Verbringen kranker Tiere, starker Fliegenbefall. Daher konn-
ten die Erreger sich während der Mast ungehindert ausbreiten. Betrieb L verbesserte
sich allmählich von Kategorie III zu Kategorie I ab Ende des zweiten Durchganges, in
dem im Vergleich zu den anderen beiden nur sehr wenige Nachweise erfolgten. In
Durchgang 3 stieg die Nachweisrate wiederum an, was sich vier Monate nach Been-
digung in einer erneuten Eingruppierung in Kategorie III widerspiegelte.
Klon Nr. 12 wurde noch in einem weiteren Betrieb identifiziert: Ende des zweiten
Durchgangs wurde er erstmals in Betrieb J nachgewiesen. Dieser Betrieb liegt zwar
nicht im selben Ort wie Betrieb H, aber dennoch räumlich nur wenige Kilometer ent-
fernt. Der Klon wurde im Erstnachweis auf einem Schadnager identifiziert, der für
den Neueintrag verantwortlich gewesen sein könnte. Der Klon dominierte die nur sel-

Diskussion
167
ten positiven Proben in den Durchgänge 2 und 3. Möglicherweise übertrug der Nager
(eine Ratte) den Klon von einem zum anderen Betrieb. Andererseits könnte es sich
auch um ein Salmonellenreservoir innerhalb dieser Tierpopulation handeln, das den
Primäreintrag in beide Betriebe verursachte. Beide Möglichkeiten wurden in der Lite-
ratur beschrieben (LE MOINE et al. 1987; LANGVAD et al. 2006; SKOV et al. 2008).
In den vorigen Durchgängen waren bereits zwei andere Klone aufgetreten, die zeit-
gleich in einem weiteren Betrieb nachgewiesen wurden: Ein Klon (8) war im ersten
Durchgang vor Einstallung sowohl im Abteil als auch bei den Ferkeln präsent.
Dieser Klon wurde auch im LKW-Kot von Betrieb I nachgewiesen. Es stellte sich her-
aus, dass beide Betriebe ihre Ferkel über die gleiche Erzeugergenossenschaft bezo-
gen. Auch der zweite Klon (17) wurde bei beiden Betrieben im leeren Abteil detektiert,
was auf einen Eintrag in den vorigen Durchgängen hindeutet, möglicherweise eben-
falls durch die Ferkel. Eine andere Option könnte die Übertragung durch Schadnager
sein. Beide Betriebe liegen in demselben Ort. Der Klon wurde in Betrieb J, der eben-
so wie Betrieb I trotz Bekämpfungsmaßnahmen einen extrem hohen Befall aufwies,
mehrfach in Nagerkot und auf der Körperoberfläche dieser nachgewiesen. Insgesamt
reduzierte sich jedoch in beiden Betrieben die Salmonellenbelastung. In Betrieb I
erfolgte im zweiten Durchgang nur ein Nachweis, im dritten keiner. Der Bestand hatte
in allen drei Durchgängen Gesundheitsprobleme (Atemwegserkrankungen, Diarrhö)
worauf die Tiere mehrfach mit Tetrazyklin, Trimethoprim-Sulfadiazin, Colistin, Sulfa-
merazin, Sulfadimidin, Doxycyclin und Tylosin behandelt wurden. Eine Überprüfung
der Sensibilität der Isolate gegenüber verschiedenen Antibiotika ergab keine Hinwei-
se auf Resistenzen. Es kann nicht ausgeschlossen werden, dass in diesem Fall die
massive Anwendung von Antibiotika zu einer Reduktion der Salmonellen führte, die
ohnehin meist in Kot und tiernaher Umgebung auftraten. Der Betrieb verbesserte
sich im dritten Durchgang von Kategorie III zu II.
Betrieb J hatte vor Beginn des zweiten Durchgangs mit dem Einsatz von Benzoesäu-
re über das Futter begonnen, die Reinigung und Desinfektion sowie die Schadna-
gerbekämpfung einer Fremdfirma überlassen. Letzteres könnte eine Reduktion des

Diskussion
168
Salmonellenaufkommens bewirkt haben, da im ersten, durch sehr viele Erreger-
nachweise gekennzeichneten Durchgang sehr oft Schadnager kontaminiert waren.
Der an diesen gefundene Klon trat im zweiten Durchgang nicht mehr auf. Insgesamt
war dieser Durchgang, wie auch der dritte, nur recht selten „positiv“, wenn auch im-
mer noch Nager angetroffen wurden. Welche der Maßnahmen zum Erfolg führte,
kann nicht abschließend beurteilt werden. Der Betrieb war ab Ende des zweiten
Durchgangs in Kategorie II eingestuft.
Diese unterschiedlichen Verläufe legen dar, wie divers die Infektionswege und Ver-
teilungsmuster in den einzelnen Betrieben sein können. Die Resultate unterstreichen,
wie akribisch bei der Suche nach Eintragsquellen und Übertragungsrouten vorge-
gangen werden muss. Es konnte gezeigt werden, dass für jeden Betrieb individuelle,
teilweise sehr komplexe Bekämpfungsmaßnahmen zur Salmonellenreduktion erfor-
derlich sind. Dies wurde bereits von BLAHA (2008) konstatiert, der nicht eine aus-
schließliche Intensivierung der heute üblichen Reinigung und Desinfektion fordert,
sondern ein betriebs- und problemspezifisches, alle praktischen und theoretischen
Rückzugsmöglichkeiten von Salmonellen einbeziehendes Bekämpfungskonzept.
Dies dürfe nicht nur einmalig Anwendung finden, sondern müsse als permanente
Maßnahme fest in das Qualitätsmanagement integriert werden.
Für diese Auswertung erwies sich die Feintypisierung mittels Fingerprinting als äu-
ßerst wertvolles Werkzeug, um auch auf den ersten Blick nicht ersichtliche epidemio-
logische Zusammenhänge aufzudecken und den Tierhaltern Schwachstellen und
damit Bekämpfungsansätze aufzuzeigen.
Trotz der teilweise unterschiedlichen Situationen in den Betrieben konnte die Studie
einige in allen Betrieben ähnliche Muster in der Dynamik von Salmonelleninfektionen
im Bestand charakterisieren. Es handelt sich zwar um eine exemplarische Untersu-
chung einer limitierten Anzahl von Betrieben, und daher wurden eher Beobachtungen
als allgemeingültige „Gesetzmäßigkeiten“ formuliert. Dennoch sind sicherlich viele
Feststellungen aus dieser Untersuchung auf andere Betriebe übertragbar und verhel-

Diskussion
169
fen zu einem noch besseren Verständnis von Salmonelleninfektionen und Bekämp-
fungsmöglichkeiten im Bestand.

Diskussion
170
5.3 Schlussfolgerungen
Diese Untersuchung kam zu dem Ergebnis, dass die wichtigste Route zum Neuein-
trag von Salmonellen von außerhalb in die untersuchten Mastbestände durch Ferkel
selbst darstellten, während bei Schadnagern und anderen Vektoren ein echter Neu-
eintrag von außen selten untermauert werden konnte. Dies bestätigt die Ausführun-
gen der EFSA, die für „Hoch-Prävalenz“ Länder die mit 70 - 80 % die höchste Reduk-
tion von Salmonellen bei Schlachtschweinen durch Freiheit von Zuchtschweinen an-
nimmt, während durch Verhinderung der Einträge via externe Quellen (Schadnager
etc.) eine Verminderung um 10 -20 % erwartet wird.
Nach erfolgtem Neueintrag verbreiten sich diese Salmonellen sukzessive im Bestand
und persistieren während der folgenden Mastdurchgänge an unterschiedlichsten Or-
ten im Sinne eines Stallhospitalismus. Dies belegt den Zusammenhang mit einer
mangelhaften Reinigung und Desinfektion des Stalls vor Neubelegung, wie sie durch
Salmonella-Nachweise zu diesem Zeitpunkt in vielen Betrieben festgestellt wurde.
Zudem wird ersichtlich, dass eine penible Stallhygiene auch während der Mast und
möglichst strikte Trennung zwischen verschiedenen Tier- bzw. Mastgruppen (auch
über getrennte Arbeitsgeräte, -kleidung, Treibwege usw.) nötig ist, um ein Zirkulieren
der Infektion im Bestand zu unterbinden.
Landwirte müssen also für eine nachhaltige Bekämpfung von Salmonellen im Mast-
bestand an mehreren Ebenen ansetzen, dazu gehört der Zukauf Salmonella-
unverdächtiger Ferkel ebenso wie das Hygienemanagement im eigenen Betrieb.
Die wichtige Rolle der Tierumgebung in der Erregerübertragung konnte nicht nur für
Salmonella spp., sondern auch für Campylobacter spp. und Y. enterocolitica hervor-
gehoben werden. Für diese Erreger wurde der Infektion über die Umgebung bisher
eine eher untergeordnete Bedeutung beigemessen. Allerdings ließen sich zwischen
Campylobacter spp. und Y. enterocolitica einerseits und Salmonella spp. anderer-
seits sehr wohl Differenzen im Verteilungsmuster erkennen. Während Salmonellen

Diskussion
171
an sehr vielen unterschiedlichen Lokalisationen überall im Stall auftreten, wurden die
anderen beiden Pathogene tendenziell näher am Tier vorgefunden.
Daraus kann gefolgert werden, dass sowohl Unterschiede in den Übertragungswe-
gen, als auch in den möglichen Risikofaktoren bestehen, wie auch die Analyse mög-
licher Einflussfaktoren zeigte. Daher dürften vermutlich keine gänzlich identischen
Bekämpfungsstrategien für alle drei Erreger anwendbar sein, obwohl grundsätzlich
eine Kontrolle von Campylobacter spp. und Y. enterocolitica in der Schweinepopula-
tion möglich scheint.
Dies schlug sich im Erregervorkommen in den einzelnen Betrieben nieder: Während
Campylobacter spp. und Y. enterocolitica häufig gemeinsam auftraten, konnte kein
Zusammenhang mit dem Vorkommen von Salmonella spp. hergestellt werden. Aller-
dings war dieser indirekt bei Betrachtung der Kategorie wahrnehmbar. Betriebe in
Kategorie I wiesen häufiger Campylobacter spp. und Y. enterocolitica auf als Betrie-
be in Kategorie III. Möglicherweise ging dies auf eine mit dem Wissen um die
„schlechte“ Kategorisierung assoziierte Sensibilisierung der Landwirte gegenüber
einer guten Betriebshygiene zurück.
Die Mutmaßung, dass Hygienemaßnahmen sich womöglich schneller auf Campy-
lobacter spp. und Y. enterocolitica als auf Salmonellen auswirkten, spiegelte sich in
der Entwicklung der Kategorie in den Betrieben wider. Die Kategorisierung bildet,
über einen ausreichenden Zeitraum betrachtet, durchaus die Entwicklung des Sal-
monellenvorkommens im Bestand ab. Sie reagiert jedoch mit einer gewissen Zeitver-
zögerung auf Veränderungen, so dass von ihr, z. B. bei stichpunktartiger Betrachtung,
nicht in jedem Falle Rückschlüsse auf die aktuelle Belastung zulässig sind.
Da die Untersuchungen jeweils an einer relativ geringen Anzahl von Betrieben
durchgeführt wurden, sind zur Überprüfung der Allgemeingültigkeit der getätigten
Schlussfolgerungen weitere Untersuchungen mit größeren Stichprobenumfängen
erforderlich.

Zusammenfassung
172
6. Zusammenfassung
Christina Nathues
Untersuchungen zum Vorkommen von Salmonella spp., Campylobacter spp.
und Yersinia enterocolitica und zur Dynamik der Salmonelleninfektion in
Schweinemastbetrieben
Campylobacter spp., Yersinia enterocolitica und Salmonella spp. repräsentieren die
drei häufigsten bakteriellen Ursachen lebensmittelassoziierter Enteritiden beim Men-
schen. Alle drei Zoonoseerreger kommen beim Schwein vor, dessen Fleisch der
höchste Pro-Kopf Konsum in der gesamten Europäischen Union beigemessen wird.
Daher war das Ziel dieser Arbeit, in zwei Teilprojekten das Vorkommen der drei Pa-
thogene in der Schweinemast genauer zu untersuchen. Dadurch wurden neue Er-
kenntnisse zu Übertragungswegen und (gemeinsamen) Risikofaktoren der Erreger
erwartet, die zu einer besseren Bekämpfung der Pathogene im Bestand führen sollen.
Im Rahmen einer Querschnittsuntersuchung wurden 50 niedersächsische Schwei-
nemastbetriebe, je zur Hälfte in QS-Salmonellen-Kategorie I und III, auf das Vor-
kommen der drei Erreger untersucht. Dazu wurden eine Befragung zu Betriebsfakto-
ren durchgeführt und Sammelkotproben, Umgebungsproben sowie Proben von
Schädlingen entnommen und kulturell untersucht. Anhand dessen wurden das Vor-
kommen aller Erreger im Tier und an den verschiedenen Lokalisationen und ihr ge-
meinsames Auftreten auf Proben- und Betriebsebene - auch differenziert nach Kate-
gorie - analysiert. Zudem wurden betriebliche Faktoren im Hinblick auf einen mögli-
chen Einfluss auf den in der kulturellen Untersuchung ermittelten Herdenstatus un-
tersucht.
In einer Verlaufsuntersuchung wurden zwölf Mastbetriebe der QS-Salmonellen-
Kategorie III über einen bis drei Mastdurchgänge regelmäßig auf Salmonella spp.
untersucht. Die Probenentnahmen von Sammelkot, Umgebung und Schädlingen

Zusammenfassung
173
fanden regelmäßig im vierwöchigen Rhythmus, beginnend kurz vor Einstallung der
Ferkel bis zum Ende des Mastdurchgangs, statt. Die Auswertung der zeitlichen und
räumlichen Entwicklung der Salmonelleninfektion im Mastverlauf erfolgte zum einen
auf Speziesebene, zum anderen anhand einer auf einer komplexen Feintypisierung
basierenden Einteilung der Salmonellen in klonale Linien.
In der Querschnittsuntersuchung konnte gezeigt werden, dass neben Salmonella (S.)
spp. auch Campylobacter (C.) spp. und Y. enterocolitica häufig in der Tierumgebung
vorkommen. So wurden in insgesamt 33 % aller Proben aus der direkten Tierumge-
bung (Buchtenwände, Tränken etc.) Campylobacter spp. nachgewiesen, fast ebenso
häufig wie im Sammelkot selbst. In 8 % dieser Umgebungsproben waren
Y. enterocolitica nachweisbar. Auch an Lokalisationen ohne oder mit nur sporadi-
schem Tierkontakt (tierferne Umgebung) wurden beide Erreger nachgewiesen: So
wurden C. spp. von knapp 10 % aller Stiefel und 12 % aller Abteilgänge nachgewie-
sen, Y. enterocolitica von je 4 % aller Treibbretter und Zentralgänge isoliert, Beide
Erreger wurden auch in Fliegen bzw. Nagerkot nachgewiesen. Dies ist ein Hinweis,
dass die Rolle der Tierumgebung bei der Übertragung dieser Erreger bisher mög-
licherweise unterschätzt wurde. Salmonella spp. wurden seltener als die anderen
Pathogene nachgewiesen, nur in der tierfernen Umgebung ähnlich häufig wie C. spp.
Sowohl Campylobacter spp. als auch Y. enterocolitica traten häufiger in Betrieben
der Kategorie I als in Kategorie III auf (C. spp.: 88 % zu 72 %, Y. enterocolitica: 76 %
zu 20 %). Des Weiteren konnten keine gemeinsamen Risikofaktoren für alle drei Er-
reger charakterisiert werden, so dass es auch weiterhin unklar ist, ob identischen
Bekämpfungsstrategien für alle drei Erreger im Bestand möglich sein werden. Sal-
monella spp. traten nur wenig öfter in Betrieben der Kategorie III als denen Kategorie
I auf (36 % der Betriebe zu 28 % der Betriebe), so dass die Salmonellen-
Kategorisierung nicht in jedem Fall Rückschlüsse auf die momentan aktuelle Belas-
tung eines Betriebs mit Salmonellen zulässt.
In der Verlaufsuntersuchung wurden bei acht der zwölf Kategorie-III-Betriebe kulturell
Salmonellen nachgewiesen, der Anteil positiver Proben betrug insgesamt 8,6 %. Die

Zusammenfassung
174
mit 30 % am häufigsten positive Probenart war der Kot der Ferkel vor Anlieferung,
Bei neun von 23 Durchgängen wurden hier Salmonella spp. nachgewiesen. Somit
erfolgte der Neueintrag von Salmonellen in die Mast am häufigsten mit den Ferkeln.
Bei sieben Durchgängen konnte der Erreger bereits im leeren, gereinigten und desin-
fizierten Abteil detektiert werden. Zudem unterschieden sich die Anteile positiver
Proben aus dem leeren und dem belegten Abteil kaum (8 % zu 10 %). Dies zeigte,
dass die in den Betrieben durchgeführte Reinigung und Desinfektion nicht zu einer
Minderung der Salmonella-Belastung beitrug und so zu einer Übertragung des Erre-
gers von einer zur nächsten Mastgruppe führte. Die tierferne Umgebung war die am
zweithäufigsten positive Lokalisationsgruppe (11 % positive Proben). Dies weist auf
ihre wichtige Bedeutung als permanentes Erregerreservoir hin. Anhand der komple-
xen Feintypisierung konnten alle Salmonellenfunde einer von 20 klonalen Linien
(Klone) zugeordnet werden. Die Mehrheit der Klone trat in mehreren Mastdurchgän-
gen eines Betriebs auf, was auf eine länger andauernde Persistenz im Bestand hin-
deutet. Die höchste Klondiversität fand sich in der tierfernen Umgebung, fast alle klo-
nalen Linien wurden dort nachgewiesen. Zudem konnten anhand einiger in mehreren
Betrieben aufgetretener Klone mögliche Übertragungswege zwischen den Beständen,
zweimal durch Nager, einmal durch zugekaufte Ferkel rekonstruiert werden, wodurch
das Potential einer Erreger-Feintypisierung für epidemiologische Untersuchungen
unterstrichen wird.

Summary
175
7. Summary
Christina Nathues
Research on the prevalences of Salmonella spp., Campylobacter spp. and Yer-
sinia enterocolitica and the dynamics of Salmonella spp. infections in fattening
pig herds
Campylobacter spp. Yersinia enterocolitica and Salmonella spp. represent the most
common bacterial causes of food-borne intestinal disease in humans in Europe. Pigs
are a reservoir of all three agents, with pork being the meat species with the highest
per-capita consumption in the EU. Therefore the aim of this study was to assess - in
two sub-projects - the occurrence of the three pathogens in fattening pig herds. Thus
it was expected to gain more detailed knowledge concerning transmission routes and
(common) risk factors of the three zoonotic agents that might lead to a more efficient
control of these pathogens on the pre-harvest level.
In a cross-sectional study 50 fattening pig herds, located in Lower-Saxony, were in-
vestigated for the presence of the three pathogens, half of them being categorized in
QS Salmonella category I and category III respectively, Therefore a survey on speci-
fic management factors of the farms via questionnaire was carried out, supplemented
by the collection and cultural examination of faecal and environmental samples and
samples of pests. This was followed by a statistical analysis of the occurrence of the
pathogens in the animals and their environment, for each alone and also for all three
together, on sample as well as herd level. Additionally, a risk factor analysis of cer-
tain management factors, based on the culturally assessed pathogens status of each
farm, was performed.
Furthermore, a longitudinal study was conducted in twelve fattening pigs herds in
Salmonella category III over the course of one up to three fattening periods. The
samplings of faeces, environment and pests took place at regular intervals of four

Summary
176
weeks, starting before the housing in of the piglets until the end of the fattening peri-
od. The analysis of the dynamics of Salmonella infections in fattening periods, in the
course of time as well as in its spatial expansion, was done on species level and ad-
ditionally on the level of the results of a molecular fingerprinting (clone level).
In the cross-sectional study it could be shown that apart from Salmonella (S.) spp.
also Campylobacter (C.) spp. and Y. enterocolitica were found frequently in the pig‟s
environment. C. spp. were detected in 33% of all samples taken from the direct envi-
ronment of the pigs (pen walls, nipple drinkers etc.), almost as much as from faeces.
From 8% of these samples Y. enterocolitica were isolated. Even at locations without
or with only sporadic animal contact (indirect environment) both pathogens were de-
tected: C. spp. from 10% of the farmer‟s boots and 12% of the passage ways, Y. en-
terocolitica on 4% of guide boards and passage ways. Both were also found in flies /
rodents. This indicates that the importance of the pig‟s environment might have been
underestimated until present. Salmonella spp. were detected less frequently than the
other pathogens with the exception of the indirect environment where it was quite as
frequent as C. spp. Campylobacter spp. as well as Y. enterocolitica were isolated
more often in farms of Salmonella category I than category III (C. spp. 88% vs. 72%,
Y. enterocolitica 76% vs. 20%). Furthermore there couldn‟t be characterized any
common risk factor for all the three agents, leaving the question whether the imple-
mentation of one single control strategy for all three bacteria on farm level might be
possible unresolved. Salmonella spp. were found only slightly more often in category
I herds than category III herds (36% vs. 28% positive herds). This indicated that from
the QS Salmonella category one cannot always conclude directly on the actual bac-
terial load of the herd.
In the longitudinal study Salmonella spp. were detected culturally in eight out of
twelve farms, the overall detection rate was 8.6%. The sample type offering the high-
est proportion of positive samples (30%) was the piglet‟s faeces before housing them
in. Here, in nine out of 23 examined fattening periods the pathogen was detected.
This leads to the assumption that S. spp. were brought into the farm most often via

Summary
177
the piglets themselves. In seven fattening periods the agent was detected already in
the empty cleaned and disinfected compartment. Besides, there were no statistical
differences in the proportions of positive samples from the empty compartment and
during the fattening period (8% vs. 10%). This shows that the cleaning an disinfection
of the compartment, carried out on the farms, did not lead to a considerable reduction
of the Salmonella load in the unit and could not prevent the transmission of the pa-
thogen from one to the following fattening period. The indirect environment was the
group of locations second most frequently positive (11% of these samples), sug-
gesting its high importance as a permanent reservoir of Salmonella. With the help of
the molecular fingerprinting all isolates could be assigned to one of 20 different clonal
lineages. Almost all different clonal lineages were present in more than one fattening
period within each farm, indicating their long term presence in the herd. The highest
diversity of clones was found in the indirect environment; nearly all of the 20 clonal
lineages were detected here. Besides, by revealing the presence of some identical
clonal lineages in different herds, the transmission routes of Salmonella spp. be-
tween the farms could be traced back (piglets in one, rodents in two cases), sug-
gesting a high potential of a molecular fingerprinting of the isolates as a valuable tool
in epidemiological investigations.

Literaturverzeichnis
178
8. Literaturverzeichnis
Anonyme Schriften ANONYMOUS (2002): EN ISO 6579:2002+A1:2007 Mikrobiologie von Lebensmitteln und Futtermitteln - Horizontales Verfahren zum Nachweis von Salmonella spp. Beuth Verlag, Berlin ANONYMOUS (2005a): Advisory Committee on the Microbiological Safety of Food: Second report on Campylobacter. www.food.gov.uk/multimedia/pdfs/acmsfcampyloreport.pdf Abrufdatum: 01 Nov 2011 ANONYMOUS (2006a): ISO 10272-1:2006 Horizontales Verfahren zum Nachweis und zur Zählung von Campylobacter spp. - Teil 1: Nachweisverfahren. Beuth Verlag, Berlin ANONYMOUS (2006b): Opinion of the Scientific Panel on Biological Hazards on the request from the Commission related to "Risk assessment and mitigation options of Salmonella in pig production". EFSA J. 341, 1-131 ANONYMOUS (2007c): Zu einem nosokomialen Ausbruch durch S. Enteritidis in Fulda, zu einem Ausbruch in einem Wolfsburger Klinikum, zu einem Ausbruch am Klinikum Dortmund. http://www.gpk.de/downloadp/STIKO_2007_Bulletin48_071130_Salmonella_Enteritidis.pdf Abrufdatum: 19 Jan 2011 ANONYMOUS (2008a): Report of the Task Force on Zoonoses Data Collection on the analysis of the baseline survey on the prevalence of Salmonella in slaughter pigs, Part A: Salmonella Prevalence estimates. EFSA J. 135, 1-111 ANONYMOUS (2008b): Report of the Task Force on Zoonoses Data Collection on the analysis of the baseline survey on the prevalence of Salmonella in slaughter pigs, Part B: factors associated with Salmonella infection in lymph nodes, Salmonella surface contamination of carcasses, and the distribution of Salmonella serovars. EFSA J. 206, 1-111

Literaturverzeichnis
179
ANONYMOUS (2009a): Analysis of the baseline survey on the prevalence of Salmonella in holdings with breeding pigs, in the EU, 2008, Part A: Salmonella prevalence estimates. EFSA J. 7, 1-93 ANONYMOUS (2009b): RKI: Epidemiologisches Bulletin 13/2009, RKI-Ratgeger Infektionskrankheiten - Merkblätter für Ärzte Salmonellose. http://www.rki.de/cln_169/nn_504488/DE/Content/Infekt/EpidBull/Merkblaetter/Ratgeber__Mbl__Salmonellose.html Abrufdatum: 17.01.2011 ANONYMOUS (2009c): World Health Organization Initiative for Vaccine Research - Diarrhoeal Diseases (Updated February 2009). http://www.who.int/vaccine_research/diseases/diarrhoeal/en/index2.html Abrufdatum: 11 Dec 2010 ANONYMOUS (2010a): The Community Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and food-borne outbreaks in the European Union in 2008. EFSA J. 8(1), 1-311 ANONYMOUS (2010b): Scientific Opinion on a Quantitative Microbiological Risk Assessment of Salmonella in slaughter and breeder pigs. EFSA J. 8, 1-80 ANONYMOUS (2011): World Health Organization: Food safety and foodborne illness. http://www.who.int/mediacentre/factsheets/fs237/en/index.html Abrufdatum: 29 Jan 2011
AABO, S., J. E. OLSEN, E. J. THRELFALL u. D. J. BROWN (1995): Characterization of non-virulence plasmids with homology to the virulence plasmid of Salmonella dublin. Res. Microbiol. 146, 751-759 AARESTRUP, F. M., u. J. ENGBERG (2001): Antimicrobial resistance of thermophilic Campylobacter. Vet. Res. 32, 311-321

Literaturverzeichnis
180
AARTS, H. J., R. G. JOOSTEN, M. H. HENKENS, H. STEGEMAN u. A. H. VAN HOEK (2001): Rapid duplex PCR assay for the detection of pathogenic Yersinia enterocolitica strains. J. Microbiol. Meth. 47, 209-217 AARTS, H. J., L. A. VAN LITH u. J. KEIJER (1998): High-resolution genotyping of Salmonella strains by AFLP-fingerprinting. Let. Appl. Microbiol. 26, 131-135 ACKE, E., K. MCGILL, A. LAWLOR, B. R. JONES, S. FANNING u. P. WHYTE (2010): Genetic diversity among Campylobacter jejuni isolates from pets in Ireland. Vet. Rec. 166, 102-106 ADAK, G. K., S. M. MEAKINS, H. YIP, B. A. LOPMAN u. S. J. O'BRIEN (2005): Disease risks from foods, England and Wales, 1996-2000. Emerg. Infect. Dis. 11, 365-372 ALBAN, L., u. K. D. STARK (2005): Where should the effort be put to reduce the Salmonella prevalence in the slaughtered swine carcass effectively? Prev. vet. med. 68, 63-79 ALDOVA, E., u. K. LAZNICKOVA (1979): Comments on the ecology and epidemiology of Yersinia enterocolitica in Czechoslovakia. Contrib. Microbiol. Immunol. 5, 122-131 ALTER, T., F. GAULL, S. KASIMIR, M. GURTLER, H. MIELKE, M. LINNEBUR u. K. FEHLHABER (2005): Prevalences and transmission routes of Campylobacter spp. strains within multiple pig farms. Vet. Microbiol. 108, 251-261 ANDERSEN, J. K., R. SORENSEN u. M. GLENSBJERG (1991): Aspects of the epidemiology of Yersinia enterocolitica: a review. Int. J. Food Microbiol. 13, 231-237 ANDERSON, E. S., L. R. WARD, M. J. SAXE u. J. D. DE SA (1977): Bacteriophage-typing designations of Salmonella typhimurium. J. Hyg. (London) 78, 297-300 ANDREWS-POLYMENIS, H. L., A. J. BAUMLER, B. A. MCCORMICK u. F. C. FANG (2010): Taming the elephant: Salmonella biology, pathogenesis, and prevention. Infect. Immun. 78, 2356-2369

Literaturverzeichnis
181
ARCHER, J. R., R. F. SCHELL, D. R. PENNELL u. P. D. WICK (1987): Identification of Yersinia spp. with the API 20E system. J. Clin. Microbiol. 25, 2398-2399 BABIC-ERCEG, A., Z. KLISMANIC, M. ERCEG, D. TANDARA u. M. SMOLJANOVIC (2003): An outbreak of Yersinia enterocolitica O:3 infections on an oil tanker. Eur. J. Epidemiol. 18, 1159-1161 BARRETT, T. J., P. GERNER-SMIDT u. B. SWAMINATHAN (2006): Interpretation of pulsed-field gel electrophoresis patterns in foodborne disease investigations and surveillance. Foodborne Pathog. Dis. 3, 20-31 BELOEIL, P. A., P. FRAVALO, C. FABLET, J. P. JOLLY, E. EVENO, Y. HASCOET, C. CHAUVIN, G. SALVAT u. F. MADEC (2004): Risk factors for Salmonella enterica subsp. enterica shedding by market-age pigs in French farrow-to-finish herds. Prev. vet. med. 63, 103-120 BERCOVIER, H., u. H. H. MOLLARET (1984): Genus XIV. In: D. H. BERGEY, J. G. HOLT und N. R. KRIEG (Hrsg.): Bergey´s Manual Vol. 1 Williams & Wilkins, Baltimore, S. 498-506 BISPING, W. (1993): Salmonellen in Futtermitteln. Dtsch. tierärztl. Wochenschr. 100, 262-263 BLACK, R. E., R. J. JACKSON, T. TSAI, M. MEDVESKY, M. SHAYEGANI, J. C. FEELEY, K. I. MACLEOD u. A. M. WAKELEE (1978): Epidemic Yersinia enterocolitica infection due to contaminated chocolate milk. New Engl. J. Med. 298, 76-79 BLAHA, T. (1993): Die Ausbreitungsdynamik von Salmonellen in Tierbestanden. Dtsch. tierärztl. Wochenschr. 100, 278-280 BLAHA, T. (2008): Salmonellenbekämpfung in der EU und in Deutschland unter besonderer Berücksichtigung der Ökologischen Tierhaltung. In: G. RAHMANN und U. SCHUMACHER (Hrsg.): Landbauforschung Johann Heinrich von Thünen-Institut, Braunschweig, S. 45-51

Literaturverzeichnis
182
BODE, K. (2008): Serologische und epidemiologische Untersuchungen zur Salmonellendynamik in Schweinebeständen für die Optimierung des Salmonellenmonitorings beim Schwein. Hannover, Tierärztl. Hochsch., Diss. BÖHM, R. (1993): Verhalten ausgewahlter Salmonellen in der Umwelt. Dtsch. tierärztl. Wochenschr. 100, 275-278 BÖHME, H., A. FRUTH u. W. RABSCH (2009): Reptilien-assoziierte Salmonelleninfektionen bei Sauglingen und Kleinkindern in Deutschland. Klin. Pädiatr. 221, 60-64 BOQVIST, S., H. PETTERSSON, A. SVENSSON u. Y. ANDERSSON (2009): Sources of sporadic Yersinia enterocolitica infection in children in Sweden, 2004: a case-control study. Epidemiol. Infect. 137, 897-905 BOTTONE, E. J. (1997): Yersinia enterocolitica: the charisma continues. Clin. Microbiol. Rev. 10, 257-276 BOTTONE, E. J. (1999): Yersinia enterocolitica: overview and epidemiologic correlates. Microbes Infect. 1, 323-333 BOYAPALLE, S., I. V. WESLEY, H. S. HURD u. P. G. REDDY (2001): Comparison of culture, multiplex, and 5' nuclease polymerase chain reaction assays for the rapid detection of Yersinia enterocolitica in swine and pork products. J. Food Prot. 64, 1352-1361 BOYEN, F., F. HAESEBROUCK, D. MAES, F. VAN IMMERSEEL, R. DUCATELLE u. F. PASMANS (2008a): Non-typhoidal Salmonella infections in pigs: a closer look at epidemiology, pathogenesis and control. Vet. Microbiol. 130, 1-19 BOYEN, F., F. HAESEBROUCK, A. VANPARYS, J. VOLF, M. MAHU, F. VAN IMMERSEEL, I. RYCHLIK, J. DEWULF, R. DUCATELLE u. F. PASMANS (2008b): Coated fatty acids alter virulence properties of Salmonella Typhimurium and decrease intestinal colonization of pigs. Vet. Microbiol. 132, 319-327

Literaturverzeichnis
183
BRANGER, C. G., A. TORRES-ESCOBAR, W. SUN, R. PERRY, J. FETHERSTON, K. L. ROLAND u. R. CURTISS, 3RD (2009): Oral vaccination with LcrV from Yersinia pestis KIM delivered by live attenuated Salmonella enterica serovar Typhimurium elicits a protective immune response against challenge with Yersinia pseudotuberculosis and Yersinia enterocolitica. Vaccine 27, 5363-5370 BRENNAN, R., u. D. PREDIGER (1981): Coefficient Kappa: Some Uses, Misuses, and Alternatices. Educ. Psychol. Meas. 41, 687-699 BRUUN, T., G. SORENSEN, L. P. FORSHELL, T. JENSEN, K. NYGARD, G. KAPPERUD, B. A. LINDSTEDT, T. BERGLUND, A. WINGSTRAND, R. F. PETERSEN, L. MULLER, C. KJELSO, S. IVARSSON, M. HJERTQVIST, S. LOFDAHL u. S. ETHELBERG (2009): An outbreak of Salmonella Typhimurium infections in Denmark, Norway and Sweden, 2008. Euro. Surveill. 14, 19147 BUCHWALD, D. S., u. M. J. BLASER (1984): A review of human salmonellosis: II. Duration of excretion following infection with nontyphi Salmonella. Rev. Infect. Dis. 6, 345-356 BUSSE, M. (1995): Media for Salmonella. Int. J. Food Microbiol. 26, 117-131 BUTZLER, J. P. (2004): Campylobacter, from obscurity to celebrity. Clin. Microbiol. Infect. 10, 868-876 CHEN, S., A. YEE, M. GRIFFITHS, C. LARKIN, C. T. YAMASHIRO, R. BEHARI, C. PASZKO-KOLVA, K. RAHN u. S. A. DE GRANDIS (1997): The evaluation of a fluorogenic polymerase chain reaction assay for the detection of Salmonella species in food commodities. Int. J. Food Microbiol. 35, 239-250 CHIU, C. H., u. J. T. OU (1996): Rapid identification of Salmonella serovars in feces by specific detection of virulence genes, invA and spvC, by an enrichment broth culture-multiplex PCR combination assay. J. Clin. Microbiol. 34, 2619-2622

Literaturverzeichnis
184
CORBEL, M. J. (1979): The relationship between the protective and cross-reacting antigens of Brucella spp., Yersinia enterocolitica 0:9 and Salmonella serotypes of Kauffmann-White group N. Contrib. Microbiol. Immun. 5, 50-63 CORRY, J. E., D. E. POST, P. COLIN u. M. J. LAISNEY (1995): Culture media for the isolation of campylobacters. Int. J. Food Microbiol. 26, 43-76 DAVIES, P. R., P. K. TURKSON, J. A. FUNK, M. A. NICHOLS, S. R. LADELY u. P. J. FEDORKA-CRAY (2000): Comparison of methods for isolating Salmonella bacteria from faeces of naturally infected pigs. J. Appl. Microbiol. 89, 169-177 DEDIÉ, K., J. BOCKEMÜHL, H. KÜHN, K.-J. VOLKMER u. T. WEINKE (1993): Bakterielle Zoonosen bei Tier und Mensch: Epidemiologie, Pathologie, Klinik, Diagnostik und Bekämpfung. Enke, Stuttgart DENIS, M., J. REFREGIER-PETTON, M. J. LAISNEY, G. ERMEL u. G. SALVAT (2001): Campylobacter contamination in French chicken production from farm to consumers. Use of a PCR assay for detection and identification of Campylobacter jejuni and Camp. coli. J. Appl. Microbiol. 91, 255-267 DÖHNE, S. (2010): Untersuchungen zum Auftreten verschiedener bakterieller Zoonoseerreger und zu den Risikofaktoren in norddeutschen Schweinemastbeständen. Hannover, Tierärztl. Hochsch., Diss. ENGBERG, J., F. M. AARESTRUP, D. E. TAYLOR, P. GERNER-SMIDT u. I. NACHAMKIN (2001): Quinolone and macrolide resistance in Campylobacter jejuni and C. coli: resistance mechanisms and trends in human isolates. Emerg. Infect. Dis. 7, 24-34 FEDORKA-CRAY, P. J., S. C. WHIPP, R. E. ISAACSON, N. NORD u. K. LAGER (1994): Transmission of Salmonella typhimurium to swine. Vet. Microbiol. 41, 333-344 FOLEY, S. L., A. M. LYNNE u. R. NAYAK (2009): Molecular typing methodologies for microbial source tracking and epidemiological investigations of Gram-negative bacterial foodborne pathogens. Infect. Genet. Evol. 9, 430-440

Literaturverzeichnis
185
FOSSE, J., M. LAROCHE, N. OUDOT, H. SEEGERS u. C. MAGRAS (2011): On-farm multi-contamination of pigs by food-borne bacterial zoonotic hazards: an exploratory study. Vet. Microbiol. 147, 209-213 FOSSE, J., H. SEEGERS u. C. MAGRAS (2009): Prevalence and risk factors for bacterial food-borne zoonotic hazards in slaughter pigs: a review. Zoonoses Public Health 56, 429-454 FREDRIKSSON-AHOMAA, M., M. GERHARDT u. A. STOLLE (2009): High bacterial contamination of pig tonsils at slaughter. Meat Sci. 832, 334-336 FREDRIKSSON-AHOMAA, M., u. H. KORKEALA (2003): Low occurrence of pathogenic Yersinia enterocolitica in clinical, food, and environmental samples: a methodological problem. Clin. Microbiol. Rev. 16, 220-229 FREDRIKSSON-AHOMAA, M., T. KORTE u. H. KORKEALA (2000): Contamination of carcasses, offals, and the environment with yadA-positive Yersinia enterocolitica in a pig slaughterhouse. J. Food Prot. 63, 31-35 FRIEDMAN, C. R., R. M. HOEKSTRA, M. SAMUEL, R. MARCUS, J. BENDER, B. SHIFERAW, S. REDDY, S. D. AHUJA, D. L. HELFRICK, F. HARDNETT, M. CARTER, B. ANDERSON u. R. V. TAUXE (2004): Risk factors for sporadic Campylobacter infection in the United States: A case-control study in FoodNet sites. Clin. Infect. Dis. 38 Suppl 3, S285-296 FROST, J. A., J. M. KRAMER u. S. A. GILLANDERS (1999): Phage typing of Campylobacter jejuni and Campylobacter coli and its use as an adjunct to serotyping. Epidemiol. Infect. 123, 47-55 FUKUSHIMA, H. (1987): New selective agar medium for isolation of virulent Yersinia enterocolitica. J. Clin. Microbiol. 25, 1068-1073 FUKUSHIMA, H., R. NAKAMURA, Y. ITO, K. SAITO, M. TSUBOKURA u. K. OTSUKI (1983): Ecological studies of Yersinia enterocolitica. I. Dissemination of Y. enterocolitica in pigs. Vet. Microbiol. 8, 469-483

Literaturverzeichnis
186
GARCIA-FELIZ, C., A. CARVAJAL, J. A. COLLAZOS u. P. RUBIO (2009): Herd-level risk factors for faecal shedding of Salmonella enterica in Spanish fattening pigs. Prev. vet. med. 91, 130-136 GRADEL, K. O., C. DETHLEFSEN, T. EJLERTSEN, H. C. SCHONHEYDER u. H. NIELSEN (2008): Increased prescription rate of antibiotics prior to non-typhoid Salmonella infections: a one-year nested case-control study. Scand. J. Infect. Dis. 40, 635-641 GRAHEK-OGDEN, D., B. SCHIMMER, K. S. CUDJOE, K. NYGARD u. G. KAPPERUD (2007): Outbreak of Yersinia enterocolitica serogroup O:9 infection and processed pork, Norway. Emerg. Infect. Dis. 13, 754-756 GUIBOURDENCHE, M., P. ROGGENTIN, M. MIKOLEIT, P. I. FIELDS, J. BOCKEMUHL, P. A. GRIMONT u. F. X. WEILL (2010): Supplement 2003-2007 (No. 47) to the White-Kauffmann-Le Minor scheme. Res. Microbiol. 161, 26-29 GURRATH, P. (2008): Vom Erzeuger zum Verbraucher Fleischversorgung in Deutschland Ausgabe 2008. Statistisches Bundesamt, Wiesbaden HANSEN, T. B., B. B. CHRISTENSEN u. S. AABO (2010): Salmonella in pork cuttings in supermarkets and butchers' shops in Denmark in 2002 and 2006. Zoonoses Public Health 57 Suppl 1, 23-29 HAUTEKIET, V., V. GEERT, V. MARC u. G. RONY (2008): Development of a sanitary risk index for Salmonella seroprevalence in Belgian pig farms. Prev. vet. med. 86, 75-92 HELMUTH, R., u. A. SCHROETER (1994): Molecular typing methods for S. enteritidis. Int. J. Food Microbiol. 21, 69-77 HORROCKS, S. M., R. C. ANDERSON, D. J. NISBET u. S. C. RICKE (2009): Incidence and ecology of Campylobacter jejuni and coli in animals. Anaerobe 15, 18-25

Literaturverzeichnis
187
HUMPHREY, T., u. F. JORGENSEN (2006): Pathogens on meat and infection in animals - Establishing a relationship using campylobacter and salmonella as examples. Meat Sci. 74, 89-97 HUMPHREY, T., S. O'BRIEN u. M. MADSEN (2007): Campylobacters as zoonotic pathogens: a food production perspective. Int. J. Food Microbiol. 117, 237-257 HUOVINEN, E., L. M. SIHVONEN, M. J. VIRTANEN, K. HAUKKA, A. SIITONEN u. M. KUUSI (2010): Symptoms and sources of Yersinia enterocolitica-infection: a case-control study. BMC Infect. Dis. 10, 122 HURD, H. S., J. K. GAILEY, J. D. MCKEAN u. M. H. ROSTAGNO (2001): Experimental rapid infection in market swine following exposure to a Salmonella contaminated environment. Berl. Münch. tierärztl. Wochenschr. 114, 382-384 HURD, H. S., J. D. MCKEAN, R. D. GRIFFITH u. M. H. ROSTAGNO (2004): Estimation of the Salmonella enterica prevalence in finishing swine. Epidemiol. Infect. 132, 127-135 IBRAHIM, A., W. LIESACK u. E. STACKEBRANDT (1992): Differentiation between pathogenic and non-pathogenic Yersinia enterocolitica strains by colony hybridization with a PCR-mediated digoxigenin-dUTP-labelled probe. Mol. Cell. Probes 6, 163-171 JAGOW, J., u. W. E. HILL (1986): Enumeration by DNA colony hybridization of virulent Yersinia enterocolitica colonies in artificially contaminated food. Appl. Environ. Microbiol. 51, 441-443 JONES, L. A. INGRAM, K. E. FULLERTON, R. MARCUS, B. J. ANDERSON, P. V. MCCARTHY, D. VUGIA, B. SHIFERAW, N. HAUBERT, S. WEDEL u. F. J. ANGULO (2006): A case-control study of the epidemiology of sporadic Salmonella infection in infants. Pediatrics 118, 2380-2387 JONES, T., E. M. SUTCLIFFE u. A. CURRY (1991): Recovery of viable but non-culturable Campylobacter jejuni. J. Gen. Microbiol. 137, 2477-2482

Literaturverzeichnis
188
KAMPHAUSEN, R., u. F. JAEGER (2009): Prävalenzsenkungen im Hinblick auf eine integrierte Strategie zur Reduzierung des Zoonoseerregereintrags in die Lebensmittelkette. in: Bundesinstitut für Risikobewertung (Hrsg.): Zoonosen und Lebensmittelsicherheit. BfR-Hausdruckerei Dahlem, Berlin; S. 13-14 KASIMIR, S. (2005): Verlaufsuntersuchungen zum Vorkommen potentiell humanpathogener Yersinia enterocolitica und Campylobacter spp. in Schweinebeständen von der Geburt bis zur Schlachtung sowie Genotypisierung ausgewählter Isolate. Leipzig, Univ., Veterinärmed. Fak., Diss. KAUFFMANN, F. (1960): Two biochemical sub-divisions of the genus Salmonella. Acta Pathol. Microbiol. Scand. 49, 393-396 KIDGELL, C., U. REICHARD, J. WAIN, B. LINZ, M. TORPDAHL, G. DOUGAN u. M. ACHTMAN (2002): Salmonella typhi, the causative agent of typhoid fever, is approximately 50,000 years old. Infect. Genet. Evol. 2, 39-45 KITTELBERGER, R., F. HILBINK, M. F. HANSEN, G. P. ROSS, M. A. JOYCE, S. FENWICK, J. HEESEMANN, H. WOLF-WATZ u. K. NIELSEN (1995): Serological crossreactivity between Brucella abortus and Yersinia enterocolitica 0:9 II the use of Yersinia outer proteins for the specific detection of Yersinia enterocolitica infections in ruminants. Vet. Microbiol. 47, 271-280 KRANKER, S., L. ALBAN, J. BOES u. J. DAHL (2003): Longitudinal study of Salmonella enterica aerotype Typhimurium infection in three Danish farrow-to-finish swine herds. J. Clin. Microbiol. 41, 2282-2288 KREIENBROCK, L., u. S. SCHACH (2005): Epidemiologische Methoden. Spektrum Akademischer Verlag, Heidelberg Berlin KWAGA, J., J. O. IVERSEN u. V. MISRA (1992): Detection of pathogenic Yersinia enterocolitica by polymerase chain reaction and digoxigenin-labeled polynucleotide probes. J. Clin. Microbiol. 30, 2668-2673

Literaturverzeichnis
189
LAHESMAA-RANTALA, R., K. GRANFORS, H. ISOMAKI u. A. TOIVANEN (1987): Yersinia specific immune complexes in the synovial fluid of patients with yersinia triggered reactive arthritis. Ann. Rheum. Dis. 46, 510-514 LANGVAD, B., M. N. SKOV, E. RATTENBORG, J. E. OLSEN u. D. L. BAGGESEN (2006): Transmission routes of Salmonella Typhimurium DT 104 between 14 cattle and pig herds in Denmark demonstrated by molecular fingerprinting. J. Appl. Microbiol. 101, 883-890 LANTZ, P. G., R. KNUTSSON, Y. BLIXT, W. A. AL SOUD, E. BORCH u. P. RADSTROM (1998): Detection of pathogenic Yersinia enterocolitica in enrichment media and pork by a multiplex PCR: a study of sample preparation and PCR-inhibitory components. Int. J Food Microbiol. 45, 93-105 LAUKKANEN, R., M. HAKKINEN, J. LUNDEN, M. FREDRIKSSON-AHOMAA, T. JOHANSSON u. H. KORKEALA (2010): Evaluation of isolation methods for pathogenic Yersinia enterocolitica from pig intestinal content. J. Appl. Microbiol. 108, 956-964 LE MINOR, L., u. M. Y. POPOFF (1987): Request for an Opinion - Designation of Salmonella enterica s. nov., nom. rev., as the Type and Only Species of the Genus Salmonella. Int. J Syst. Bacteriol. 37, 465-468 LE MOINE, V., P. VANNIER u. A. JESTIN (1987): Microbiological studies of wild rodents in farms as carriers of pig infectious agents. Prev. vet. med. 4, 399-408 LEBLANC MARIDOR, M., M. DENIS, F. LALANDE, B. BEAUREPAIRE, R. CARIOLET, P. FRAVALO, M. FEDERIGHI, H. SEEGERS u. C. BELLOC (2008): Experimental infection of specific pathogen-free pigs with Campylobacter: excretion in faeces and transmission to non-inoculated pigs. Vet. Microbiol. 131, 309-317 LEE, L. H., E. BURG, 3RD, S. BAQAR, A. L. BOURGEOIS, D. H. BURR, C. P. EWING, T. J. TRUST u. P. GUERRY (1999): Evaluation of a truncated recombinant flagellin subunit vaccine against Campylobacter jejuni. Infect. Immun. 67, 5799-5805

Literaturverzeichnis
190
LIEBANA, E. (2002): Molecular tools for epidemiological investigations of S. enterica subspecies enterica infections. Res. Vet. Sci. 72, 169-175 LINDSTEDT, B. A., T. VARDUND, L. AAS u. G. KAPPERUD (2004): Multiple-locus variable-number tandem-repeats analysis of Salmonella enterica subsp. enterica serovar Typhimurium using PCR multiplexing and multicolor capillary electrophoresis. J. Microbiol. Meth. 59, 163-172 LIOR, H., D. L. WOODWARD, J. A. EDGAR, L. J. LAROCHE u. P. GILL (1982): Serotyping of Campylobacter jejuni by slide agglutination based on heat-labile antigenic factors. J. Clin. Microbiol. 15, 761-768 LO FO WONG, D. M., J. DAHL, H. STEGE, P. J. VAN DER WOLF, L. LEONTIDES, A. VON ALTROCK u. B. M. THORBERG (2004): Herd-level risk factors for subclinical Salmonella infection in European finishing-pig herds. Prev. vet. med. 62, 253-266 LO FO WONG, D. M., J. DAHL, P. J. VAN DER WOLF, A. WINGSTRAND, L. LEONTIDES u. A. VON ALTROCK (2003): Recovery of Salmonella enterica from seropositive finishing pig herds. Vet. Microbiol. 97, 201-214 MAGISTRALI, C., A. M. DIONISI, P. DE CURTIS, L. CUCCO, O. VISCHI, S. SCUOTA, A. ZICAVO u. G. PEZZOTTI (2008): Contamination of Salmonella spp. in a pig finishing herd, from the arrival of the animals to the slaughterhouse. Res. Vet. Sci. 85, 204-207 MALORNY, B., A. SCHROETER, C. BUNGE, B. HOOG, A. STEINBECK u. R. HELMUTH (2001): Evaluation of molecular typing methods for Salmonella enterica serovar Typhimurium DT104 isolated in Germany from healthy pigs. Vet. Res. 32, 119-129 MARTIN-PELAEZ, S., A. COSTABILE, L. HOYLES, R. A. RASTALL, G. R. GIBSON, R. M. LA RAGIONE, M. J. WOODWARD, E. MATEU u. S. M. MARTIN-ORUE (2010): Evaluation of the inclusion of a mixture of organic acids or lactulose into the feed of pigs experimentally challenged with Salmonella Typhimurium. Vet. Microbiol. 142, 337-345

Literaturverzeichnis
191
MARTIN-PELAEZ, S., B. PERALTA, E. CREUS, A. DALMAU, A. VELARDE, J. F. PEREZ, E. MATEU u. S. M. MARTIN-ORUE (2009): Different feed withdrawal times before slaughter influence caecal fermentation and faecal Salmonella shedding in pigs. Vet. J 182, 469-473 MASLOW, J. N., M. E. MULLIGAN u. R. D. ARBEIT (1993): Molecular epidemiology: application of contemporary techniques to the typing of microorganisms. Clin. Infect. Dis. 17, 153-162; quiz 163-154 MATARAGAS, M., P. N. SKANDAMIS u. E. H. DROSINOS (2008): Risk profiles of pork and poultry meat and risk ratings of various pathogen/product combinations. Int. J. Food Microbiol. 126, 1-12 MERIALDI, G., G. BARIGAZZI, P. BONILAURI, C. TITTARELLI, M. BONCI, M. D'INCAU u. M. DOTTORI (2008): Longitudinal study of Salmonella infection in Italian farrow-to-finish swine herds. Zoonoses Public Health 55, 222-226 MERLE, R., S. KÖSTERS, U. PORTSCH, T. MAY u. L. KREIENBROCK (2009): The serological Salmonella monitoring in German pork production: How can progress be observed? In: 14. Congr. Int. Society An. Hyg., Vechta 2009 Vol. 2, S. 759-761 MERLE, R., B. SCHNEIDER, B. FRANZ, U. PORTSCH, T. MAY, T. BLAHA u. L. KREIENBROCK (2007): The serological Salmonella Monitoring in German pork production: the structure of the central database and preliminary results of a basic epidemiological report In: Safepork 7. Int. Symp. on epidemiol. & contr. of foodborne pathog. in pork, Verona 2007 Vol. 2, S. 140-144 METHNER, U., W. RABSCH, R. REISSBRODT u. P. H. WILLIAMS (2008): Effect of norepinephrine on colonisation and systemic spread of Salmonella enterica in infected animals: role of catecholate siderophore precursors and degradation products. Int. J. Med. Microbiol. 298, 429-439 MEYER, C., E. GROSSE BEILAGE u. J. KRIETER (2004): Salmonellen in der Schweineproduktion - Risikofaktoren und Ansätze zur Bekämpfung. Züchtungskunde 76, 354-366

Literaturverzeichnis
192
MOORE, J. E., D. CORCORAN, J. S. DOOLEY, S. FANNING, B. LUCEY, M. MATSUDA, D. A. MCDOWELL, F. MEGRAUD, B. C. MILLAR, R. O'MAHONY, L. O'RIORDAN, M. O'ROURKE, J. R. RAO, P. J. ROONEY, A. SAILS u. P. WHYTE (2005): Campylobacter. Vet. Res. 36, 351-382 NACHAMKIN, I. (2002): Chronic effects of Campylobacter infection. Microbes Infect. 4, 399-403 NACHAMKIN, I., u. S. BARBAGALLO (1990): Culture confirmation of Campylobacter spp. by latex agglutination. J. Clin. Microbiol. 28, 817-818 NEIMANN, J., J. ENGBERG, K. MOLBAK u. H. C. WEGENER (2003): A case-control study of risk factors for sporadic campylobacter infections in Denmark. Epidemiol. Infect. 130, 353-366 NESBAKKEN, T., K. ECKNER, H. K. HOIDAL u. O. J. ROTTERUD (2003): Occurrence of Yersinia enterocolitica and Campylobacter spp. in slaughter pigs and consequences for meat inspection, slaughtering, and dressing procedures. Int. J Food Microbiol. 80, 231-240 NESBAKKEN, T., K. ECKNER u. O. J. ROTTERUD (2008): The effect of blast chilling on occurrence of human pathogenic Yersinia enterocolitica compared to Campylobacter spp. and numbers of hygienic indicators on pig carcasses. Int. J Food Microbiol. 123, 130-133 NESBAKKEN, T., T. IVERSEN u. B. LIUM (2007): Pig herds free from human pathogenic Yersinia enterocolitica. Emerg. Infect. Dis 13, 1860-1864 NESBAKKEN, T., E. NERBRINK, O. J. ROTTERUD u. E. BORCH (1994): Reduction of Yersinia enterocolitica and Listeria spp. on pig carcasses by enclosure of the rectum during slaughter. Int. J Food Microbiol. 23, 197-208 NEUBAUER, H., L. D. SPRAGUE, H. SCHOLZ u. A. HENSEL (2001a): Die Diagnostik der Yersinia enterocolitica-Infektionen: Eine Übersicht uber klassische Nachweistechniken und neue molekularbiologische Methoden. Berl. Münch. tierärztl. Wochenschr. 114, 1-7 NEUBAUER, H., L. D. SPRAGUE, H. SCHOLZ u. A. HENSEL (2001b): Yersinia-enterocolitica-Infektionen: 2. Bedeutung bei Menschen. Berl. Münch. tierärztl. Wochenschr. 114, 81-87

Literaturverzeichnis
193
NEWELL, D. G., M. KOOPMANS, L. VERHOEF, E. DUIZER, A. AIDARA-KANE, H. SPRONG, M. OPSTEEGH, M. LANGELAAR, J. THREFALL, F. SCHEUTZ, J. VAN DER GIESSEN u. H. KRUSE (2010): Food-borne diseases - the challenges of 20 years ago still persist while new ones continue to emerge. Int. J Food Microbiol. 139 Suppl 1, S3-15 NIELSEN, B., C. HEISEL u. A. WINGSTRAND (1996): Time course of the serological response to Yersinia enterocolitica O:3 in experimentally infected pigs. Vet. Microbiol. 48, 293-303 NOLLET, N., K. HOUF, J. DEWULF, L. DUCHATEAU, L. DE ZUTTER, A. DE KRUIF u. D. MAES (2005): Distribution of salmonella strains in farrow-to-finish pig herds: a longitudinal study. J Food Prot. 68, 2012-2021 NOLLET, N., D. MAES, L. DE ZUTTER, L. DUCHATEAU, K. HOUF, K. HUYSMANS, H. IMBERECHTS, R. GEERS, A. DE KRUIF u. J. VAN HOOF (2004): Risk factors for the herd-level bacteriologic prevalence of Salmonella in Belgian slaughter pigs. Prev. vet. med. 65, 63-75 NOWAK, B., T. V. MUEFFLING, K. CASPARI u. J. HARTUNG (2006): Validation of a method for the detection of virulent Yersinia enterocolitica and their distribution in slaughter pigs from conventional and alternative housing systems. Vet. Microbiol. 117, 219-228 O'CONNOR, A. M., T. DENAGAMAGE, J. M. SARGEANT, A. RAJIC u. J. MCKEAN (2008): Feeding management practices and feed characteristics associated with Salmonella prevalence in live and slaughtered market-weight finisher swine: a systematic review and summation of evidence from 1950 to 2005. Prev. vet. med. 87, 213-228 ON, S. L. (1996): Identification methods for campylobacters, helicobacters, and related organisms. Clin. Microbiol. Rev: 9, 405-422 OWEN, R. J., E. LORENZ u. J. GIBSON (1997): Application of the Mast resistotyping scheme to Campylobacter jejuni and C. coli. J. Med. Microbiol. 46, 34-38

Literaturverzeichnis
194
OYOFO, B. A., S. A. THORNTON, D. H. BURR, T. J. TRUST, O. R. PAVLOVSKIS u. P. GUERRY (1992): Specific detection of Campylobacter jejuni and Campylobacter coli by using polymerase chain reaction. J. Clin Microbiol. 30, 2613-2619 PAPENBROCK, S., K. STEMME, G. AMTSBERG, J. VERSPOHL u. J. KAMPHUES (2005): Investigations on prophylactic effects of coarse feed structure and/or potassium diformate on the microflora in the digestive tract of weaned piglets experimentally infected with Salmonella Derby. J. Anim. Physiol. Anim. Nutr. (Berlin) 89, 84-87 PARRY, C. M., u. E. J. THRELFALL (2008): Antimicrobial resistance in typhoidal and nontyphoidal salmonellae. Curr. Opin. Infect. Dis. 21, 531-538 PATHMANATHAN, S. G., N. CARDONA-CASTRO, M. M. SANCHEZ-JIMENEZ, M. M. CORREA-OCHOA, S. D. PUTHUCHEARY u. K. L. THONG (2003): Simple and rapid detection of Salmonella strains by direct PCR amplification of the hilA gene. J. Med. Microbiol. 52, 773-776 PAYOT, S., S. DRIDI, M. LAROCHE, M. FEDERIGHI u. C. MAGRAS (2004): Prevalence and antimicrobial resistance of Campylobacter coli isolated from fattening pigs in France. Vet. Microbiol. 101, 91-99 PENNER, J. L., u. J. N. HENNESSY (1980): Passive hemagglutination technique for serotyping Campylobacter fetus subsp. jejuni on the basis of soluble heat-stable antigens. J. Clin. Microbiol. 12, 732-737 PILON, J., R. HIGGINS u. S. QUESSY (2000): Epidemiological study of Yersinia enterocolitica in swine herds in Quebec. Can. Vet. J. 41, 383-387 RAWLINS, M. L., C. GERSTNER, H. R. HILL u. C. M. LITWIN (2005): Evaluation of a western blot method for the detection of Yersinia antibodies: evidence of serological cross-reactivity between Yersinia outer membrane proteins and Borrelia burgdorferi. Clin. Diagn. Lab. Immunol. 12, 1269-1274

Literaturverzeichnis
195
REISSBRODT, R., E. VIELITZ, E. KORMANN, W. RABSCH u. H. KUHN (1996): Ferrioxamine E-supplemented pre-enrichment and enrichment media improve various isolation methods for Salmonella. Int. J. Food Microbiol. 29, 81-91 RIBOT, E. M., M. A. FAIR, R. GAUTOM, D. N. CAMERON, S. B. HUNTER, B. SWAMINATHAN u. T. J. BARRETT (2006): Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 3, 59-67 ROBINS-BROWNE, R. M., M. D. MILIOTIS, S. CIANCIOSI, V. L. MILLER, S. FALKOW u. J. G. MORRIS, JR. (1989): Evaluation of DNA colony hybridization and other techniques for detection of virulence in Yersinia species. J. Clin. Microbiol. 27, 644-650 ROBINS-BROWNE, R. M., u. J. K. PRPIC (1985): Effects of iron and desferrioxamine on infections with Yersinia enterocolitica. Infect. Immun. 47, 774-779 ROSNER, B. M., K. STARK u. D. WERBER (2010): Epidemiology of reported Yersinia enterocolitica infections in Germany, 2001-2008. BMC Public Health 10, 337 ROSSEL, R., J. ROULLIER, P.-A. BELOEIL, C. CHAUVIN, F. BASTA, J. P. CRABOS u. S. THEAU-AUDIN (2006): Salmonella en élevage de porcs du Sud-Ouest de la France: séroprévalence en fin d'engraissement et facteurs de risque associés. J. Rech. Porc. 38, 371-378 SANCHEZ, J., I. R. DOHOO, J. CHRISTENSEN u. A. RAJIC (2007): Factors influencing the prevalence of Salmonella spp. in swine farms: a meta-analysis approach. Prev. vet. med. 81, 148-177 SCHIEMANN, D. A. (1979): Synthesis of a selective agar medium for Yersinia enterocolitica. Can. J. Microbiol. 25, 1298-1304 SCHLESINGER, M., S. POLLAK u. P. A. VARDY (1979): Cross-antigenicity between Yersinia and Rickettsia. Isr. J. Med. Sci. 15, 612-613

Literaturverzeichnis
196
SCHOLZ, H. C., T. ARNOLD, H. MARG, U. RÖSLER u. A. HENSEL (2001): Improvement of an invA-based PCR for the specific detection of Salmonella typhimurium in organs of pigs. Berl. Münch. tierärztl. Wochenschr. 114, 401-403 SELBITZ, H.-J. (2002): Bakterielle Krankheiten der Tiere. In: A. MAYR (Hrsg.): Medizinische Mikrobiologie, Infektions- und Seuchenlehre 7. Aufl., Enke Verlag, Stuttgart, S. 462-478 SELKE, M., J. MEENS, S. SPRINGER, R. FRANK u. G. F. GERLACH (2007): Immunization of pigs to prevent disease in humans: construction and protective efficacy of a Salmonella enterica serovar Typhimurium live negative-marker vaccine. Infect. Immun. 75, 2476-2483 SKIRROW, M. B. (1977): Campylobacter enteritis: a "new" disease. Br. Med. J. 2, 9-11 SKJERVE, E., B. LIUM, B. NIELSEN u. T. NESBAKKEN (1998): Control of Yersinia enterocolitica in pigs at herd level. Int. J. Food Microbiol. 45, 195-203 SKOV, M. N., J. J. MADSEN, C. RAHBEK, J. LODAL, J. B. JESPERSEN, J. C. JORGENSEN, H. H. DIETZ, M. CHRIEL u. D. L. BAGGESEN (2008): Transmission of Salmonella between wildlife and meat-production animals in Denmark. J. Appl. Microbiol. 105, 1558-1568 SMULDERS, F. J., I. VAGSHOLM u. H. KORKEALA (2008): Food-borne zoonoses, the EU zoonosis legislation and the prospects for food safety and consumer protection during primary animal production. Wien. Klin. Wochenschr. 120, 587-598 SOMYANONTANAGUL, N. (2009): Comparison between detecting Salmonella spp. by bacteriological method and Real-Time PCR assay in samples from pig herds. Hannover, Tierärztl. Hochsch., Diss. STEINBACH, G., T. BLAHA u. U. METHNER (2002): Estimating the prevalence of Salmonella spp. in swine herds--influence of sensitivity and specificity of Salmonella detection. J. Vet. Med. B. Infect. Dis. Vet. Public Health 49, 438-444

Literaturverzeichnis
197
STEINHAUSEROVA, I., M. NEBOLA u. M. MIKULICOVA (2005): Prevalence of thermophilic Campylobacter spp. in slaughtered pigs in the Czech Republic, 2001 - 2003. Vet. Med. Czech. 50, 171-174 STERN, N. J., K. L. HIETT, G. A. ALFREDSSON, K. G. KRISTINSSON, J. REIERSEN, H. HARDARDOTTIR, H. BRIEM, E. GUNNARSSON, F. GEORGSSON, R. LOWMAN, E. BERNDTSON, A. M. LAMMERDING, G. M. PAOLI u. M. T. MUSGROVE (2003): Campylobacter spp. in Icelandic poultry operations and human disease. Epidemiol. Infect. 130, 23-32 TACKET, C. O., J. P. NARAIN, R. SATTIN, J. P. LOFGREN, C. KONIGSBERG, JR., R. C. RENDTORFF, A. RAUSA, B. R. DAVIS u. M. L. COHEN (1984): A multistate outbreak of infections caused by Yersinia enterocolitica transmitted by pasteurized milk. JAMA 251, 483-486 TAUXE, R. V., J. VANDEPITTE, G. WAUTERS, S. M. MARTIN, V. GOOSSENS, P. DE MOL, R. VAN NOYEN u. G. THIERS (1987): Yersinia enterocolitica infections and pork: the missing link. Lancet 1, 1129-1132 TENOVER, F. C., R. D. ARBEIT, R. V. GOERING, P. A. MICKELSEN, B. E. MURRAY, D. H. PERSING u. B. SWAMINATHAN (1995): Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol. 33, 2233-2239 THAKUR, S., u. W. A. GEBREYES (2005): Prevalence and antimicrobial resistance of Campylobacter in antimicrobial-free and conventional pig production systems. J. Food Prot. 68, 2402-2410 TIETZE, E., u. H. TSCHÄPE (1983): Plasmid pattern analysis of natural bacterial isolates and its epidemiological implication. J. Hyg. (London) 90, 475-488 TINDALL, B. J., P. A. GRIMONT, G. M. GARRITY u. J. P. EUZEBY (2005): Nomenclature and taxonomy of the genus Salmonella. Int. J. Syst. Evol. Microbiol. 55, 521-524 TOLCIN, R., M. M. LASALVIA, B. A. KIRKLEY, E. A. VETTER, F. R. COCKERILL, 3RD u. G. W. PROCOP (2000): Evaluation of the Alexon-trend ProSpecT Campylobacter microplate assay. J. Clin. Microbiol. 38, 3853-3855

Literaturverzeichnis
198
TORPDAHL, M., G. SORENSEN, B. A. LINDSTEDT u. E. M. NIELSEN (2007): Tandem repeat analysis for surveillance of human Salmonella Typhimurium infections. Emerg. Infect. Dis. 13, 388-395 TREBESIUS, K., D. HARMSEN, A. RAKIN, J. SCHMELZ u. J. HEESEMANN (1998): Development of rRNA-targeted PCR and in situ hybridization with fluorescently labelled oligonucleotides for detection of Yersinia species. J. Clin. Microbiol. 36, 2557-2564 URSING, J. B., H. LIOR u. R. J. OWEN (1994): Proposal of minimal standards for describing new species of the family Campylobacteraceae. Int. J. Syst. Evol. Microbiol. 44, 842-845 UZZAU, S., D. J. BROWN, T. WALLIS, S. RUBINO, G. LEORI, S. BERNARD, J. CASADESUS, D. J. PLATT u. J. E. OLSEN (2000): Host adapted serotypes of Salmonella enterica. Epidemiol. Infect. 125, 229-255 VAN DAMME, I., I. HABIB u. L. DE ZUTTER (2010): Yersinia enterocolitica in slaughter pig tonsils: enumeration and detection by enrichment versus direct plating culture. Food. Microbiol. 27, 158-161 VAN DER WOLF, P. J., W. B. WOLBERS, A. R. ELBERS, H. M. VAN DER HEIJDEN, J. M. KOPPEN, W. A. HUNNEMAN, F. W. VAN SCHIE u. M. J. TIELEN (2001): Herd level husbandry factors associated with the serological Salmonella prevalence in finishing pig herds in The Netherlands. Vet. Microbiol. 78, 205-219 VAN LOGHEM, J. J. (1944): The Classification of the Plague-Bacillus. Antonie van Leeuwenhoek J. Microbiol. Serol. 10, 15-16 VANDAMME, P., E. FALSEN, R. ROSSAU, B. HOSTE, P. SEGERS, R. TYTGAT u. J. DE LEY (1991): Revision of Campylobacter, Helicobacter, and Wolinella taxonomy: emendation of generic descriptions and proposal of Arcobacter gen. nov. Int. J. Syst. Evol. Microbiol. 41, 88-103 VELLINGA, A., u. F. VAN LOOCK (2002): The dioxin crisis as experiment to determine poultry-related campylobacter enteritis. Emerg. Infect. Dis. 8, 19-22

Literaturverzeichnis
199
VIGO, G. B., J. A. CAPPUCCIO, P. E. PINEYRO, A. SALVE, M. A. MACHUCA, M. A. QUIROGA, F. MOREDO, G. GIACOBONI, J. L. CANCER, I. G. CAFFER, N. BINSZTEIN, M. PICHEL u. C. J. PERFUMO (2009): Salmonella enterica subclinical infection: bacteriological, serological, pulsed-field gel electrophoresis, and antimicrobial resistance profiles--longitudinal study in a three-site farrow-to-finish farm. Foodborne Pathog. Dis. 6, 965-972 VISHNUBHATLA, A., R. D. OBERST, D. Y. FUNG, W. WONGLUMSOM, M. P. HAYS u. T. G. NAGARAJA (2001): Evaluation of a 5'-nuclease (TaqMan) assay for the detection of virulent strains of Yersinia enterocolitica in raw meat and tofu samples. J. Food Prot. 64, 355-360 VISSCHER, C. F., P. WINTER, J. VERSPOHL, J. STRATMANN-SELKE, M. UPMANN, M. BEYERBACH u. J. KAMPHUES (2009): Effects of feed particle size at dietary presence of added organic acids on caecal parameters and the prevalence of Salmonella in fattening pigs on farm and at slaughter. J. Anim. Physiol. Anim. Nutr. (Berlin) 93, 423-430 VON ALTROCK, A., A. L. LOUIS, U. ROSLER, T. ALTER, M. BEYERBACH, L. KREIENBROCKS u. K. H. WALDMANN (2006): [The bacteriological and serological prevalence of Campylobacter spp. and Yersinia enterocolitica in fattening pig herds in Lower Saxony]. Berl. Münch. tierärztl. Wochenschr. 119, 391-399 VON ALTROCK, A., U. ROESLER, R. MERLE u. K. H. WALDMANN (2010): Prevalence of pathogenic Yersinia enterocolitica strains on liver surfaces of pigs and their antimicrobial susceptibility. J. Food Prot. 73, 1680-1683 VON ALTROCK, A., u. K. H. WALDMANN (2003): [Outlook for control of current zoonoses in swine--an issue on consumer health protection]. Dtsch. tierärztl. Wochenschr. 110, 354-357 VONNAHME, J., L. KREIENBROCK u. E. BEILAGE (2007): [Seasonal variation for Salmonella seroprevalences in breeding gilt rearing herds]. Berl. Münch. tierärztl. Wochenschr. 120, 61-66 WALDMANN, K. H., u. H. PLONAIT (2004): Erkrankungen der Verdauungsorgane und des Abdomens. In: K. H. WALDMANN und M. WENDT (Hrsg.): Lehrbuch der Schweinekrankheiten 4. Aufl., Parey Verlag, Stuttgart, S. 307-386

Literaturverzeichnis
200
WASSENAAR, T. M. u. D. G. NEWELL (2000): Genotyping of Campylobacter spp. Appl. Environ. Microbiol. 66, 1-9 WAUTERS, G., K. KANDOLO u. M. JANSSENS (1987): Revised biogrouping scheme of Yersinia enterocolitica. Contrib. Microbiol. Immunol. 9, 14-21 WEHEBRINK, T., N. KEMPER, E. GROSSE BEILAGE u. J. KRIETER (2008): Prevalence of Campylobacter spp. and Yersinia spp. in the pig production. Berl. Münch. tierärztl. Wochenschr. 121, 27-32 WEIJTENS, M. J., R. D. REINDERS, H. A. URLINGS u. J. VAN DER PLAS (1999): Campylobacter infections in fattening pigs; excretion pattern and genetic diversity. J. Appl. Microbiol. 86, 63-70 WEIJTENS, M. J., H. A. URLINGS u. J. VAN DER PLAS (2000): Establishing a campylobacter-free pig population through a top-down approach. Let. Appl. Microbiol. 30, 479-484 WEIJTENS, M. J., J. VAN DER PLAS, P. G. BIJKER, H. A. URLINGS, D. KOSTER, J. G. VAN LOGTESTIJN u. J. H. HUIS IN'T VELD (1997): The transmission of campylobacter in piggeries; an epidemiological study. J. Appl. Microbiol. 83, 693-698 WHEELER, J. G., D. SETHI, J. M. COWDEN, P. G. WALL, L. C. RODRIGUES, D. S. TOMPKINS, M. J. HUDSON u. P. J. RODERICK (1999): Study of infectious intestinal disease in England: rates in the community, presenting to general practice, and reported to national surveillance. The Infectious Intestinal Disease Study Executive. Br. med. J. 318, 1046-1050 WICHELHAUS, T. A., V. SCHÄFER u. V. BRADE (2000): Typisierungsverfahren in der Infektionsepidemiologie. Chemother. J. 2000-2, 93-98 WILKINS, W., A. RAJIC, C. WALDNER, M. MCFALL, E. CHOW, A. MUCKLE u. L. ROSENGREN (2010): Distribution of Salmonella serovars in breeding, nursery, and grow-to-finish pigs, and risk factors for shedding in ten farrow-to-finish swine farms in Alberta and Saskatchewan. Can. J. Vet. Res. 74, 81-90 WINGSTRAND, A., u. B. NIELSEN (1996): Cross-sectional investigation of pig herds for Yersinia enterocolitica. In: 14. Congr. Int. Pig Vet. Society, Bologna 1996, Vol. 1, S. 320

Literaturverzeichnis
201
YOUNG, C. R., R. HARVEY, R. ANDERSON, D. NISBET u. L. H. STANKER (2000): Enteric colonisation following natural exposure to Campylobacter in pigs. Res. Vet. Sci. 68, 75-78 YOUNUS, M., M. J. WILKINS, H. D. DAVIES, M. H. RAHBAR, J. FUNK, C. NGUYEN, A. E. SIDDIQI, S. CHO u. M. SAEED (2010): Case-control study of disease determinants for non-typhoidal Salmonella infections among Michigan children. BMC Res. Notes 3, 105
Gesetze und Verordnungen
2000 Gesetz zur Verhütung und Bekämpfung von Infektionskrankheiten beim Menschen (Infektionsschutzgesetz). 20 Jul 2000 Bundesgesetzbl. I 2000, 1045
2005 Verordnung (EG) Nr. 2073/2005 der Kommission vom 15. November 2005 über mikrobiologische Kriterien für Lebensmittel. 15 Nov 2005 Amtsbl. der EU 2005, 1 2007a Verordnung (EG) Nr. 1441/2007 der Kommission vom 5. Dezember 2007 zur Änderung der Verordnung (EG) Nr. 2073/2005 über mikrobiologische Kriterien für Lebensmittel. Vom 5 Dec 2007 Amtsbl. der EU 2007a, 12 2007b Verordnung zur Verminderung der Salmonellenverbreitung durch Schlachtschweine (Schweine-Salmonellen-Verordnung). Vom 17 Mar 2007 Bundesgesetzbl. I 2007b, 322

Anhang
202
9. Anhang
Anhang 1: Fragebogen zu Betriebsdaten
Fragebogen zur Ermittlung von Risikofaktoren in Schweine haltenden Betrieben be-züglich hoher Salmonellenseroprävalenzen bei Schlachtschweinen A. Allgemeine Angaben zum Betrieb A.1. Lage: H ______ ; R ______ Landkreis: _____ Anlage 5 ausfüllen! A.2. Tierbestand
A.3. Standorte
A.4. Teilnahme am QS seit __________ Bündeler: ________________ A.5. Personenverkehr 1. Welche Personen haben Zutritt zum Bestand?
Betriebspersonal einschließlich Tierarzt, Berater
Tiergruppe Plätze
Sauen
Jungsauen
Eber
Saugferkel
Absatzferkel
Aufzuchtschweine
Mastschweine
Stall Abteile Lage Art Anlage 1!
Bezeichnung Tiergruppe Tierzahl am Hof einzeln Fremder Hof
Frei-land
Aus- lauf

Anhang
203
Familienmitglieder weitere Personen
2. Werden von Personen, die Zutritt zum Bestand haben, andere Tierhaltungen betreut?
nein (Ausnahme: behandelnder Tierarzt/Berater) ja, aber nicht am selben Tag ja, am selben Tag nicht bekannt
A.6. mögliche Tierkontakte 1. Werden folgende Tierarten auf dem Betrieb gehalten? Falls ja – besteht eine Kontaktmöglichkeit der Tiere zu den Schweinen?
nein ja
mit Kontakt zu den Schweinen
nein ja
mit Kontakt zu den Schweinen
nein ja ? nein ja ?
Rinder Hunde
Geflügel Katzen
Schafe Kaninchen
Ziegen Tauben
Equiden Ziervögel
Wildtiere sonstige
2. Haben folgende Tiere der Umgebung Zutritt zu den Ställen?
nein ja ?
Wildvögel
Mäuse
Ratten
sonstige
A.7. Umgebung 1. Gibt es einen Mist- oder Komposthaufen in Stallnähe?
nein ja
B. Bereich Sauen / Ferkel B.1. Herkünfte 1. Werden Schweine zugekauft?
Jungsauen Eber sonstige nein nein nein ja ja ja, nämlich __________
2. Aus wie vielen Betrieben stammen die Schweine im Bestand? Jungsauen aus ____ Betrieb(en) Eber aus __ __ Betrieb(en) Sonstige aus Betrieb(en) 3.Salmonellenstatus der Herkunftsbetriebe:
Alle Zukäufe stammen aus Kategorie – I – Betrieben.

Anhang
204
Es werden auch Tiere aus Betrieben der Kategorie II oder III zugekauft. Der Salmonellenstatus ist unbekannt.
4. Wie viele Tiere wurden im letzten Jahr zugekauft? _____________ 5. Wie viele Jungsauenlieferungen haben Sie pro Jahr bezogen? ___________ B.2. Eingliederung
1. Erfolgt die Eingliederung von Schweinen nach Quarantäne?
(Jung)sauen Eber Sonstige
nein nein nein
ja, ohne Ausnahmen Anlage 2 !
ja, ohne Ausnahmen Anlage 2!
ja, ohne Ausnahmen Anlage 2!
unterschiedlich unterschiedlich unterschiedlich
B.3. Stallbelegung 1. Wie werden die folgenden Stallbereiche in der Regel belegt?
kontinuierlich Rein - Raus Entfällt wegen…
abteil stall arbeitsteiliger Fer-kelproduktion
Keine räumliche Tren-nung
Eroscenter
Wartestall
Abferkelbereich
Flatdeck
Aufzuchtbereich
Mastbereich
2. Wie ist der Abferkelrhythmus?
Wöchentlich 3 Wochen 2 Wochen sonstiges
3. Absetzalter: Tage 4. Wann kommen die Sauen in die Abferkelbucht? Tage vor dem Abferkeltermin 5. Erfolgen Reinigung und Desinfektion der Abferkelabteile vor Neubelegung?
Reinigung: nie nicht jedes Mal immer Desinfektion: nie nicht jedes Mal immer
6. Erfolgen Reinigung und Desinfektion der Flatdecks vor Neubelegung?
Reinigung: nie nicht jedes Mal immer Desinfektion: nie nicht jedes Mal immer
7. Wie häufig wird in den Abferkelbuchten der Sauenkot entfernt?
täglich wöchentlich unregelmäßig nach dem Ausstallen

Anhang
205
sonstiges: __________________________ B.4. Betreuung des Zuchtbestandes 1. Wie viele Personen haben täglichen Umgang mit den Tieren im Zuchtbereich? ____ Personen (ohne vertretungsweise Aushilfen) [Anzahl] 2. Wie viel Zeit wenden Sie einschließlich aller Mitarbeiter für die tägliche Betreuung des Sauenbe-standes auf? Betreuung der Tiere: ca. ____ Stunden. Servicearbeiten, Schreibarbeiten, Organisation: ca. ____ Stunden.
C. Management im Mastbereich C.1. Herkunft 1. Woher stammen die Mastschweine?
keine Tierzukäufe im Mastbereich
aus einem Ferkelerzeugerbetrieb
aus einem Ferkelaufzuchtbetrieb mit einer Herkunft
aus einem Ferkelaufzuchtbetrieb mit mehreren Herkünften
aus _____ gleich bleibenden Ferkelerzeugerbetrieben [Anzahl]
aus gleich bleibenden Ferkelaufzuchtbetrieben [Anzahl]
aus mehreren, wechselnden Ferkelerzeugerbetrieben
aus mehreren, wechselnden Ferkelaufzuchtbetrieben C.2. Tierbewegung 1. Wird während der Flatdeckphase (d.h. vom Absetzen bis zum Umsetzen in den Maststall) umge-stallt?
nein ja nicht zutreffend Werden während dieser Phase einzelne Schweine umgesetzt?
nein ja nicht zutreffend 2. Wird während der Mast (d.h. vom Ende der Flatdeckphase bis zum Abtransport zum Schlachthof) umgestallt?
nein ja nicht zutreffend Werden während dieser Phase einzelne Schweine umgesetzt?
nein ja nicht zutreffend 3. Wo verbleiben üblicherweise (Ü) und in Ausnahmen (A) kranke Schweine und Kümmerer? (mehrere Antworten möglich) Ü A
Alle kranken Schweine und Kümmerer bleiben in der Bucht. In einer Krankenbucht In einem Krankenstall, -abteil Euthanasie, Notschlachtung Rückstallung zu jüngeren Tieren Sonstiges __________________

Anhang
206
4. Wo verbleiben Restbestände? Restbestände bleiben bei Neueinstallung von Ferkeln im Abteil. Restbestände werden bei bereits im Bestand stehenden Mastschweinen untergebracht. Restbestände werden in einen gesonderten Stall verbracht. Bei Neueinstallung sind nie Restbestände vorhanden.
D. Haltungs- und Fütterungsmerkmale homogener Tiergruppen
gemeinsame Merkmale
Tier-gruppe
Standort
Anzahl Abteile
r-r zusammen A
Futter B
Schutzkleidung C
Geräte D
Stall E
Hygieneschl. F
G
D.1. Gebäude 1. Sind die Stalltüren geschlossen?
nein ja es gibt keine Türen
2.Bleiben die Fenster immer geschlossen?
nein ja es gibt keine Fenster
3. Angaben zur Ermittlung von Belegdichte, Tier-Fressplatz-Verhältnis, Tier-Tränke-Verhältnis:
Standort A B C D E F G H
Tierzahl pro Abteil
Anzahl Buchten pro Abteil
Tierzahl pro Bucht
Buchtengröße in m²
Anzahl Tränken pro Bucht
Troglänge in m
Mast / Aufzucht: 4. Sind Schweine unterschiedlicher Herkunft in einem Abteil untergebracht?
nein ja es gibt keine Abteile 5. Sind Schweine unterschiedlicher Herkunft in einer Bucht untergebracht?
nein ja es gibt keine Buchten 6. Sind verschiedene Altersgruppen im selben Abteil untergebracht?
nein ja es gibt keine Abteile 7. Bodenart:
Vollspaltenboden Tiefstreu/ Tretmistverfahren
Teilspaltenboden Erdboden
Vollsp. m. reduziertem Schlitzanteil Sonstige
Boden plan befestigt

Anhang
207
8. Ist beim befestigten Boden die Bodenoberfläche rissig?
nein ja nicht bekannt
9. Verschmutzungsgrad des Bodes im Tierbereich:
Kotansammlungen auf den Spalten sichtbar Kotansammlungen auf den Liegeflächen sichtbar Durchfallkot sichtbar Flüssigkeitsansammlungen sichtbar Der Tierbereich ist sauber.
10. Verschmutzungsgrad des Bodens im Gang:
Staub- und Futterablagerung sichtbar Flüssigkeitsansammlungen sichtbar Der Gang ist sauber.
11. Oberflächenbeschaffenheit der Decke
Glatte Oberfläche Oberfläche weist viele Unebenheiten und/oder Risse auf Holzbalken Rieseldecke aus Schilf oder ähnlichem Material
12. Oberflächenbeschaffenheit der (Trenn-)/Wände
Glatte Oberfläche Holz Oberfläche weist viele Unebenheiten und/oder Risse auf
13.Stroh
Einstreu Beschäftigungsmaterial Kein Stroheinsatz
Herkunft: eigen fremd Lagerung: innen außen D.2. Gülle- und Festmistbeseitigungsmanagement: 1. Lagerung der Gülle
Im Abteil ► Entleerung Nach Ausstallung
Nach Bedarf auch während der Haltung
►Maximaler Füllstand ≥ 10 cm Abstand zu Spalten
≤ 10 cm Abstand zu Spalten
Gülle fließt kontinuierlich aus dem Abteil ab
2. Falls Festmist anfällt, wie wird dieser gelagert?
auf einer betonierten Platte auf dem Erdboden sonstiges kein Festmist
D.3. Fütterung und Tränke 1. Herkunft des Futters: Betriebseigenes gemischt

Anhang
208
Zukaufsfutter 2. Welches Futter wird zusätzlich zum Grundfutter gefüttert?
Frisches Gras Obst, Gemüse Heu, Stroh sonstiges Getreideschlempe kein zusätzliches Futter
3. Enthält das Futter folgende Futtermittelzusätze?
Laktobazillen Benzoesäure Laktulose Zitronensäure Kaliumdiformiat Propionsäure Ameisensäure sonstige keine Futtermittelzusätze nicht bekannt
4. Struktur des Futtermittels:
fein gemahlenes Mehl CCM grob gemahlenes Mehl fermentiertes Futter Granulat Mix Pellets sonstiges
5. Wie hoch ist der Anteil von nicht wärmebehandelten Futter?
weniger als 15% mehr als 15% nicht bekannt 6. Ist der Anteil von Gerste im Futter höher als der Anteil von Weizen?
nein ja nicht bekannt 7.Fütterungstechnik:
Trogfütterung per Hand Flüssigfütterung, pH-Wert Breiautomaten Sensorfütterung Trockenfutterautomat mit Quertrog sonstiges
8. Wie häufig werden Futterreste am Freßplatz entfernt?
täglich wöchentlich nach Ausstallung unregelmäßig nie keine Futterreste
9. Wie häufig werden Futterleitungen und Behälter der Fütterungsanlage gereinigt?
täglich wöchentlich monatlich nach Ausstallung unregelmäßig nie
10.Futterlagerung:
innen außen offen geschlossen
11. Wird das Futterlager vor Neubefüllen desinfiziert oder gereinigt?
Reinigung nein ja
Desinfektion nein ja
12. Haben Vögel, Nagetiere oder Wild Zugang zum Futterlager?
nein ja

Anhang
209
nicht bekannt 13. Herkunft des Tränkwassers:
Stadtwasser Brunnen – die chemische Zusammensetzung wurde untersucht und entspricht Trinkwasserqualität Brunnen – die mikrobiologische Zusammensetzung wurde und entspricht Trinkwasserqualität Brunnen- die chemische Zusammensetzung wurde untersucht und entspricht nicht Trinkwasser-
qualität Brunnen – die mikrobiologische Zusammensetzung wurde und entspricht nicht Trinkwasserqualität Brunnen – zur Wasserqualität gibt es kein Untersuchungsergebnis
14. Erhält das Tränkwasser einen Säurezusatz?
nein ja nicht bekannt Wenn ja, welchen?
Benzoesäure Ameisensäure sonstigen
15.Tränken:
Nippeltränken am Freßplatz außerhalb des Freßplatzes Napftränken am Freßplatz außerhalb des Freßplatzes Trog
16. Gibt es einen Vorlaufbehälter?
nein ja 17. Wie oft erfolgt die Reinigung der Tränkwasserleitungen und -Behälter?
täglich wöchentlich monatlich nach Ausstallung unregelmäßig nie
18. Herkunft des Reinigungswassers:
Stadtwasser Brunnen – die chemische Zusammensetzung wurde untersucht und entspricht Trinkwasserqualität Brunnen – die mikrobiologische Zusammensetzung wurde und entspricht Trinkwasserqualität Brunnen – die chemische Zusammensetzung wurde untersucht und entspricht nicht Trinkwasser-
qualität Brunnen – die mikrobiologische Zusammensetzung wurde und entspricht nicht Trinkwasserqualität Brunnen – zur Wasserqualität gibt es kein Untersuchungsergebnis.
E. Hygiene E.1. Hygieneeinrichtungen 1. auf dem Betriebsgelände
Nein Ja
a)Ist das Gehöft vollständig umzäunt?
b) Ist die Fläche im Einfahrtbereich und Stallumgebung befestigt?
c) Ist eine Desinfektionsdurchfahrwanne im Gebrauch?
d) Müssen Fahrzeuge des Futtermittellieferanten den Hof befahren?
e) Müssen Fahrzeuge der TKBA den Hof befahren?
l) Ist eine Verladerampe vorhanden?
2. Kadaverentsorgung

Anhang
210
Nein Ja
a) Werden die Kadaver in Stallnähe gelagert?
b) Ist der Kadaverlagerungsplatz befestigt?
c) Wird der Kadaverlagerungsplatz nach jeder Abholung gereinigt?
d) Wird der Kadaverlagerungsplatz nach jeder Abholung desinfiziert?
e) Werden die Kadaver in einem geschlossenen Behälter gelagert?
f) Wird dieser Behälter nach jeder Abholung gereinigt?
g) Wird dieser Behälter nach jeder Abholung desinfiziert?
3. Verladen
Nein Ja
a)Ist eine Verladerampe vorhanden?
b) Wird diese nach jeder Benutzung gereinigt?
c) Wird diese nach jeder Benutzung desinfiziert?
E.2 Hygieneschleuse 1. Welche Art von Hygieneschleuse ist vorhanden? Anlage 3!
es gibt keine Hygieneschleuse. es gibt einen Umkleideraum. es gibt einen Umkleideraum mit Handwaschbecken. es gibt einen Umkleideraum mit Handwaschbecken und Dusche.
2. Wie ist der Zutritt zu den Ställen geregelt?
alle Ställe können nur durch eine Hygieneschleuse betreten werden. jeder Stall hat seine eigene Hygieneschleuse. jeder Stall hat seine eigene Hygieneschleuse und weitere Zugänge. alle Ställe sind frei zugänglich.
E.2. Personalhygiene 1. Schutzkleidung
Nein Ja
a) Sind saubere Overalls vorhanden?
b) Sind saubere Stiefel vorhanden?
c) Gibt es für jede Tiergruppe(Sauen/Läufer) separate Overalls?
d) Gibt es für jede Tiergruppe (Sauen/Läufer) separate Stiefel?
e) Werden die Stiefel vor jedem Betreten des Stalles desinfiziert?
f) Tragen Betriebsangehörige dieselbe Schutzkleidung in mehreren Ställen?
g)Wird Schutzkleidung, die in den Ställen getragen wird, auch außerhalb des Be-triebsgeländes getragen?
2. Welche Maßnahmen der Personalhygiene werden durchgeführt?
Das Personal zieht Stiefel an. Das Personal zieht Overalls an. Das Personal duscht sich vor Betreten des Stalles. Das Personal wäscht sich vor der Tierbetreuung die Hände. Das Personal trägt Handschuhe. Das Personal trägt eine Kopfbedeckung. Es werden keine Maßnahmen durchgeführt.
3. Besucher

Anhang
211
Nein Ja
a) Gelten für Besucher dieselben Anforderungen an die Personalhygiene?
b) Sind für Besucher separate Stiefel vorhanden?
c) Sind für Besucher separate Overalls vorhanden?
d) Tragen Besucher dieselbe Schutzkleidung in mehreren Ställen des Be-triebs?
e) Wird Besucherkleidung nach jedem Besuch gewaschen oder entsorgt?
E.3. Gerätehygiene 1. Reinigungsgeräte (Besen, Hochdruckreiniger),Treibbretter, Beschäftigungsmaterial, Injektionssprit-zen, …
nein ja
a) Ist jeder Stall mit separaten Geräten ausgestattet?
b) Sind für jede Tiergruppe (Sauen/Läufer) getrennte Geräte vorhanden?
c) Erfolgt eine Reinigung der Geräte nach jeder Einsatz mit Tierkontakt?
d) Erfolgt eine Desinfektion der Geräte nach jedem Einsatz mit Tierkontakt?
e) Erfolgt die Gerätereinigung vor Neubelegung?
f) Erfolgt eine Desinfektion der Geräte vor Neubelegung?
2. Tierwaage
Nein Ja
a) Gibt es für jede Tiergruppe eine separate Waage?
b) Wird die Waage nach jedem Gebrauch gereinigt?
c) Wird die Waage nach jedem Gebrauch desinfiziert?
d) Wird die Waage vor Neueinstallungen gereinigt?
e) Wird die Waage vor Neueinstallungen desinfiziert?
3. Viehtransporter
Nein Ja
a) Gibt es für jede Tiergruppe einen separaten Transporter?
b) Wird der Transporter nach jedem Gebrauch gereinigt?
c) Wird der Transporter nach jedem Gebrauch desinfiziert?
E.4. Gebäudehygiene 1. Gibt es Wannen mit Desinfektionsmittel vor den Ställen/Abteilen?
nein ja Wenn ja, wie oft wird das Desinfektionsmittel gewechselt?
täglich wöchentlich bei Bedarf nach dem Ausstallen unregelmäßig nie
2. Wann erfolgen Reinigung und Desinfektion der Abteile?
Reinigung vor jeder Neubele-gung
unregelmäßig nie
Desinfektion vor jeder Neubele-gung
unregelmäßig nie
3. Was schließt die Reinigung [R] und Desinfektion [D] der Abteile ein? R D R D
Boden Lüftungsschächte Buchtenabtrennungen Futtertröge / -automaten

Anhang
212
Wände in Tierhöhe Tränken Wände bis zur Decke Beschäftigungsmaterial Decke Gerätschaften Sonstiges __________
4. Wann erfolgt die Reinigung?
nach jedem Gebrauch
vor jeder Neu-belegung
unregel-mäßig
nie nicht be-kannt
Treibwege Ladezone Nebenräume
5. Wann erfolgt die Desinfektion?
nach jedem Gebrauch
vor jeder Neu-belegung
unregel-mäßig
nie nicht be-kannt
Treibwege Ladezone Nebenräume
6. Welche Nebenräume gibt es?
Abstellraum Büro Steuerungszentrale sonstige Keine Nebenräume
7. Werden die Räume und Flachen nach Reinigung und Desinfektion ausschließlich mit sauberen Kleidung und desinfiziertem Schuhwerk betreten?
nein ja nicht bekannt E.5. Schadnager 1. In welchem Umfang kommen Schadnager in den Betriebsgebäuden vor?
kein Hinweis auf Schadnagerbesatz erkennbar Nagerkot, Nagespuren (an Türen, Verkleidungen), Tritt- und Schleifspuren der Schwänze (auf stau-
bigen Oberflächen), Fraßspuren im Futter Ratten / Mäuse anwesend
2. Wie häufig erfolgt die Schadnagerbekämpfung?
nie bei starkem Befall bei geringem Befall permanent
3. Wer plant die Schadnagerbekämpfung und wer führt sie durch?
Ein professioneller IHK-geprüfter Schädlingsbekämpfer führt die Schadnagerbekämpfung durch. Die Schadnagerbekämpfung wird nach einem professionell erstellten Bekämpfungsplan selbst
durchgeführt. Es gibt keinen Schadnagerbekämpfungsplan, die Bekämpfung wird selbst durchgeführt.
4. Welche Mittel kommen bei der Bekämpfung von Mäusen [M] und Ratten [R] zum Einsatz?
M R
Fraßgift
Kontaktgift
Sonstiges

Anhang
213
E.6. Insektenbekämpfung 1. Fliegen – Befallsintensität
starker Fliegenbefall unauffällig nicht zutreffend 2. Wann werden Fliegen bekämpft?
permanent bei starkem Befall nie
3. Mit welchen Mittel werden Fliegen bekämpft?
Bekämpfung von Fliegenlarven mit Larvizid. Insektizidanstrich an bevorzugten Sitzflächen der Fliegen Leimfliegenfänger, Lichtfallen sonstiges: _____________________________________
4. Gibt es Schaben im Stall?
nein jaF. Salmonellen 1. Wurden aus folgenden Bereichen Proben auf Salmonellen untersucht?
nein ja Positiv Negativ
a) Futterproben aus Lager
b) Futterproben am Freßplatz
c) Futterreste aus dem Silo
d) Futterreste in den Behältern
e) Futterreste in den Leitungen
f)Tränkewasser
g) Lüftung
h) Wände/ Abtrennungen
i) Treibwege
j) Geräte (z.B. Treibbretter)
k) sonstiges
2. Erhalten die Schweine im Herkunftsbetrieb eine Salmonellenimpfung?
Impfung der Sauen Impfung der Ferkel nein nicht bekannt
3. Motivation
nein ja
Betrachten Sie Salmonellen in Ihrem Betrieb als ein Problem?
Sehen Sie einen Zusammenhang zwischen Salmonellenstatus und Produkti-onsdaten in Ihrem Betrieb?
Wurden in Ihrem Betrieb bereits Maßnahmen zur Salmonellenreduzierung durchgeführt?
Sind in Ihrem Betrieb Maßnahmen zur Salmonellenreduzierung geplant?
Nutzen Sie Ihren EDV- Zugang (Passwort), um die Daten aus der QS-Salmonellen-Datenbank einzusehen?
Wurden im Bestand in den letzten 3 Monaten Salmonellen bakteriologisch im Kot nachgewiesen?
Sind Sie dazu bereit, an weiterführenden Untersuchungen teilzunehmen?
Anlage 1 Schweinehaltung im Freien

Anhang
214
Auslaufhaltung 1. Größe der Auslauffläche(gesamt) : m
2
2. Größe des Auslaufs für eine begrenzte Tierzahl: m
2
3. Anzahl der Tiere pro Auslauf: Schweine 4. Ist der Boden komplett befestigt?
nein ja 5. Hat die Auslauffläche eine komplette Überdachung?
nein ja 6. Wird die Auslauffläche von verschiedenen Tiergruppen (Sauen/Mastschweine) gleichzeitig benutzt?
nein ja 7. Gibt es Vogelschutznetze?
Vogelschutznetze schützen alle Auslaufflächen teilweise sind Vogelschutznetze vorhanden Vogelschutznetze sind nicht vorhanden
8. Wo findet die Fütterung statt?
außen innen teilweise im Freien, teilweise im Gebäude
9. Woher kommt das Tränkewasser?
Stadtwasser Brunnen – die chemische Zusammensetzung wurde untersucht und entspricht Trinkwasserqualität Brunnen – die mikrobiologische Zusammensetzung wurde und entspricht Trinkwasserqualität Brunnen – die chemische Zusammensetzung wurde untersucht und entspricht nicht Trinkwasser-
qualität Brunnen – die mikrobiologische Zusammensetzung wurde und entspricht nicht Trinkwasserqualität Brunnen – zur Wasserqualität gibt es kein Untersuchungsergebnis.
10. Habe die Tiere ansonsten noch Zugang zu Wasser (z.B. Bach/Teich)?
nein ja nicht bekannt 11. Gibt es eine Suhle, Pfützen oder sumpfiges Gelände im Auslauf?
nein (nie) ja (immer oder zeitweise) Freilandhaltung 1. Größe der Fläche(gesamt) : m2 2. Größe der Fläche für eine begrenzte Tierzahl: m2 3. Anzahl der Tiere pro begrenzte Fläche: 4. Wird die Fläche von verschiedenen Tiergruppen (Sauen/Mastschweine) gleichzeitig genutzt?
nein ja 5. Gibt es eine Suhle, Pfützen oder sumpfiges Gelände auf der Fläche?
Anhang
215
nein (nie) ja (Immer oder zeitweise) 6. Wo findet die Fütterung statt?
außen innen teilweise im Freien, teilweise im Gebäude
7. Woher kommt das Tränkewasser?
Stadtwasser Brunnen – die chemische Zusammensetzung wurde untersucht und entspricht Trinkwasserqualität Brunnen – die mikrobiologische Zusammensetzung wurde und entspricht Trinkwasserqualität Brunnen – die chemische Zusammensetzung wurde untersucht und entspricht nicht Trinkwasser-
qualität Brunnen – die mikrobiologische Zusammensetzung wurde und entspricht nicht Trinkwasserqualität Brunnen – zur Wasserqualität gibt es kein Untersuchungsergebnis.
8. Habe die Tiere ansonsten noch Zugang zu Wasser (z. B. Bach/Teich)?
nein ja nicht bekannt
Anlage 2 Quarantäne
Anlage 4 Arbeitsschritte bei Reinigung und Desinfektion
Arbeitsschritt dabei nein ja n. b.
1 alle Tiere aus dem Abteil entfernt
Erh
eb
un
g n
ur
wen
n
ein
e Q
ua
ran
tän
e d
urc
hg
efü
hrt
wir
d:
Jung(sauen) Eber sonstige
a) Dauer der Quarantäne: _____Tage ______Tage Tage
b) Erfolgt die Quarantäne in einem separa-ten Gebäude?
nein ja, ohne Ausnahmen
nein ja, ohne Ausnahmen
nein ja, ohne Ausnahmen
c) Werden Arbeitsgeräte, die im Quaran-tänestall eingesetzt werden, auch in anderen Betriebsbereichen benutzt?
nein, ohne Ausnah-men
ja
nein, ohne Ausnah-men
ja
nein, ohne Ausnah-men
ja
d) Wird beim Betreten und Verlassen des Quarantänestalls die Kleidung gewech-selt?
nein ja, ohne Ausnahmen
nein ja, ohne Ausnahmen
nein ja, ohne Ausnahmen
e) Werden während der Quarantäne Tiere in den Quarantänestall zugestellt?
nein, ohne Ausnah-men
ja
nein, ohne Ausnah-men
ja
nein, ohne Ausnah-men
ja
f) Erhalten die Tiere während der Quaran-täne eine Salmonellen-schutzimpfung?
nein ja unterschiedlich
nein ja unterschiedlich
nein ja unterschiedlich
g) Werden die Tiere in der Quarantäne mit Antibiotika behandelt?
nein ja, regelmäßig unterschiedlich
nein ja, regelmäßig unterschiedlich
nein ja, regelmäßig unterschiedlich
h) Werden während der Quarantäne Sal-monellenuntersuchungen durchgeführt?
nein Kotproben bei An-
kunft Kotproben zum Ende Blutproben bei An-
kunft Blutproben zum Ende unterschiedlich
nein Kotproben bei An-
kunft Kotproben zum Ende Blutproben bei An-
kunft Blutproben zum
Ende unterschiedlich
nein Kotproben bei An-
kunft Kotproben bei Ende Blutproben bei An-
kunft Blutproben zum
Ende unterschiedlich
Anhang
216
2 Mechanische Reinigung besen-rein
Fäkalien entfernt
Futterreste entfernt
Staub und Spinnweben von Wänden und Decke entfernt
Wasser aus Behältern des Tränkesystems abgelassen
3 Einweichen der Oberflächen
warmes Wasser
Hochdruck 100-120 bar
Reinigungsmittel
4 Spülen
5 Trocknen, Lüften
6 Aufheizen auf Raumtemperatur
7 Ansatz des Desinfektionsmittels vor jedem Einsatz frisch
Handwarmes Wasser
Konzentration nach Herstel-lerangabe
Ansatz im Fass (200 l)
automatische Beimengung
Funktionsfähigkeit des Dosie-rers
8 Desinfektion
Mittel aus DVG-Liste
verschiedene Mittel
Einwirkdauer nach Herstel-lerangabe
angegebene Menge eingehal-ten
9 Abspülen der Desinfektionsmittel-reste
10 Dokumentation
Anlage 5: Geographische Aspekte
im Umkreis von
r =1 km r = 2 km r = 5 km Tierarten
Anzahl Schweinezuchtbestände
Anzahl Schweinemastbestände
Schweineanzahl
Anzahl Geflügelhaltungen
Geflügelanzahl
Anzahl der Haltungen von Rindern, Schafen, Ziegen, Pferden
Wild- und Zootierhaltungen
Gewässer mit Wasservögeln
Kläranlagen
Schlachtbetriebe
Verarbeitungsbetriebe
TKBA
Mülldeponien, Kompostierungsanlagen

Anhang
217
Biogasanlagen
bearbeitet wurden: A. Allgemeiner Teil
B. Sauen und Ferkel
C. Management im Mastbereich
__ x D. Haltungs- und Fütterungsmerkmale homogener Tiergruppen
E. Hygiene
F. Salmonellen
Anlage 1 Schweinehaltung im Freien
Anlage 2 Quarantäne
Anlage 3 Hygieneschleuse
Anlage 4 Vorgehensweise bei Reinigung und Desinfektion
Anlage 5 geographische Aspekte
Datum der Befragung: Interviewer-ID: Besonderheiten des Betriebes: EDV-Eingabe durch: EDV-Eingabe am: Berechnungen durch: Berechnungen am:

Anhang
218
Anhang 2: Ausschnitt aus der Excel-Tabelle zur Erfassung der Probenergebnisse der
Querschnittsuntersuchung
ID_Probe Datum ID_Betr Ort Camp Yers Salm Kat Gruppe
379 3.11.09 28 13 0 0 0 1 3
380 3.11.09 28 7 0 0 0 1 2
381 3.11.09 28 5 1 0 0 1 2
382 3.11.09 28 4 1 0 0 1 2
383 3.11.09 28 8 1 0 0 1 2
384 3.11.09 28 12 0 0 0 1 3
385 3.11.09 28 11 0 0 0 1 3
386 3.11.09 28 6 1 1 0 1 2
387 3.11.09 28 15 1 0 0 1 4
388 3.11.09 28 16 0 0 0 1 4
389 3.11.09 28 10 1 1 0 1 1
390 3.11.09 28 10 0 0 0 1 1
391 3.11.09 28 10 0 0 0 1 1
Anhang 3: Ausschnitt aus der Excel-Tabelle zur Erfassung der Kultur- und PCR-
Ergebnisse der Verlaufsuntersuchung
ID_Probe Datum ID_Betr Durchgang Zeitpunkt Nr_Probe Ort InvA Kultur Gruppe
1 15.05.08 1 1 1 1 1 0 0 1
2 15.05.08 1 1 1 2 1 0 0 1
3 15.05.08 1 1 1 3 1 0 0 1
4 15.05.08 1 1 0 1 2 0 0 3
5 15.05.08 1 1 0 2 19 0 0 3
6 15.05.08 1 1 0 3 4 0 0 2
7 15.05.08 1 1 0 4 19 0 0 3
8 15.05.08 1 1 0 5 2 0 0 3
9 15.05.08 1 1 0 6 11 0 0 3
10 15.05.08 1 1 0 7 15 0 0 4
11 15.05.08 1 1 0 8 6 0 0 2
12 15.05.08 1 1 0 9 9 0 0 2
13 15.05.08 1 1 0 10 3 0 0 3
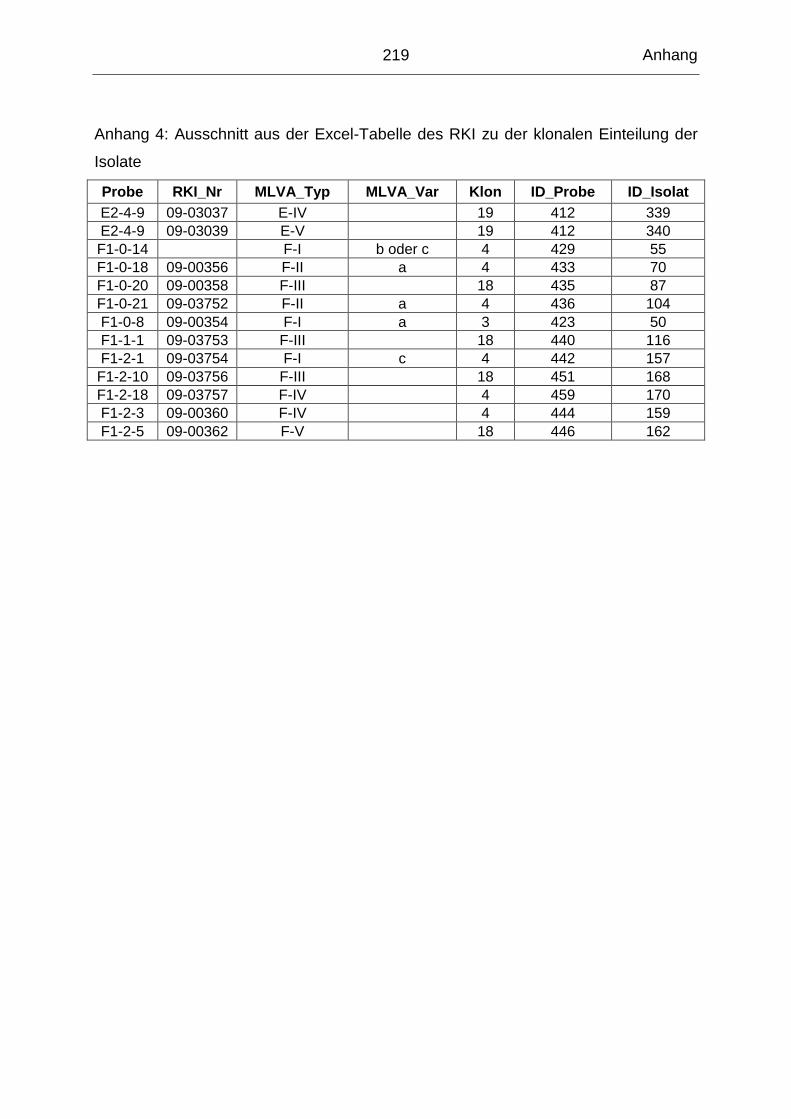
Anhang
219
Anhang 4: Ausschnitt aus der Excel-Tabelle des RKI zu der klonalen Einteilung der
Isolate
Probe RKI_Nr MLVA_Typ MLVA_Var Klon ID_Probe ID_Isolat
E2-4-9 09-03037 E-IV
19 412 339
E2-4-9 09-03039 E-V
19 412 340
F1-0-14
F-I b oder c 4 429 55
F1-0-18 09-00356 F-II a 4 433 70
F1-0-20 09-00358 F-III
18 435 87
F1-0-21 09-03752 F-II a 4 436 104
F1-0-8 09-00354 F-I a 3 423 50
F1-1-1 09-03753 F-III
18 440 116
F1-2-1 09-03754 F-I c 4 442 157
F1-2-10 09-03756 F-III
18 451 168
F1-2-18 09-03757 F-IV
4 459 170
F1-2-3 09-00360 F-IV
4 444 159
F1-2-5 09-00362 F-V
18 446 162

220
Danksagung
Mein herzlicher Dank gilt all denen, die mich bei der Anfertigung meiner Doktorarbeit
fachlich, seelisch und moralisch unterstützt haben:
Meinen beiden Doktorvätern Prof. Dr. Lothar Kreienbrock und Prof. Dr. Thomas
Blaha für die Überlassung des spannenden und abwechslungsreichen Themas und
die jederzeit freundliche, hilfsbereite und kompetente Betreuung.
Meiner Betreuerin Dr. Roswitha Merle für die exzellente fachliche Unterstützung bei
der Bearbeitung der Studien, aber auch für ihre freundliche und sonnige Art, die die
Zusammenarbeit stets zur Freude machte.
Frau Verena Gotter für die gute Kooperation und Hilfe bei den Probenentnahmen in
den Betrieben und die Überlassung der Betriebsdaten.
Allen meinen Kolleginnen und Kollegen, sowohl der Außenstelle für Epidemiologie
als auch des Instituts für Biometrie, Epidemiologie und Informationsverarbeitung, die
mir immer mit Rat und Tat zur Seite standen und ein offenes Ohr für alle Belange
hatten.
Den Mitarbeitern des Robert-Koch-Instituts Wernigerode, Herrn Dr. Eberhard Tietze,
Frau Kathrin Blank, Frau Franka Hebner und Frau Elena Kempf, für die Untersu-
chung der Proben der Verlaufsuntersuchung.
Den Mitarbeitern des Instituts für Mikrobiologie, Frau Dr. Petra Grüning und Frau
Tanja Bauer, für die Untersuchung der Proben aus der Querschnittsuntersuchung.
Allen Landwirten, die mir die Untersuchung ihrer Betriebe ermöglichten.

221
Meiner Familie, die mir in guten wie in schlechten Zeiten immer ganz viel Rückhalt
bot und mit Wort und Tat zur Seite stand.
Und natürlich meinem Mann Heiko für Alles!
Danke!





























