Embed Size (px)
Citation preview

TARTU ÜLIKOOL
LOODUS- JA TEHNOLOOGIATEADUSKOND
ÖKOLOOGIA JA MAATEADUSTE INSTITUUT
BOTAANIKA OSAKOND
Kadri Runnel
TORIKSEENED JA EPIKSÜÜLSED SAMBLIKUD
SURNUD SÄILIKPUUDEL
Magistritöö
Juhendajad: Kadi Jairus
Raul Rosenvald
Tartu 2011

1
SISUKORD
1. SISSEJUHATUS ....................................................................................................................................... 2 1.1. Säilikpuude mõiste ja eesmärk ........................................................................................................... 2 1.2. Surnud säilikpuud ja nende elustik ..................................................................................................... 3 1.3. Surnud säilikpuude torikseene- ja samblikuelustikku mõjutada võivad faktorid ............................. 4
1.3.1. Mikroklimaatilised tingimused.................................................................................................... 4 1.3.2. Tüve omadused ........................................................................................................................... 4 1.3.3. Biootilised tegurid ....................................................................................................................... 6 1.3.4. Liikide levimisvõime ................................................................................................................... 7
1.4. Senised uuringud surnud säilikpuude torikseentest ja samblikest ...................................................... 8 1.5. Magistritöö eesmärgid ........................................................................................................................ 9
2. METOODIKA ......................................................................................................................................... 11 2.1. Uurimisalad ja välitööde planeerimine............................................................................................. 11 2.2. Välitööd ............................................................................................................................................ 13
2.2.1. Puidu tiheduse mõõtmised ........................................................................................................ 13 2.2.2. Valgustingimuste hindamine ..................................................................................................... 17 2.2.3. Torikseente ja epiksüülsete samblike koosluste kirjeldamine ................................................... 17
2.3. Torikseente ja samblike määramine ................................................................................................. 19 2.4. Andmeanalüüs .................................................................................................................................. 20
2.4.1. Liikide esinemise ja liigirikkuse analüüsid ............................................................................... 20 2.4.2. Ordinatsioon .............................................................................................................................. 22
3. TULEMUSED ......................................................................................................................................... 24 3.1. Torikseened surnud säilikpuudel ...................................................................................................... 24
3.1.1. Puuliigi ja puu surmast möödunud aja mõju ............................................................................. 24 3.1.2. Liikide arvu tüvel mõjutavad faktorid ....................................................................................... 26 3.1.3. Liigiline koosseis....................................................................................................................... 27
3.2. Epiksüülsed samblikud surnud säilikpuudel .................................................................................... 31 3.2.1. Puuliigi ja puu surmast möödunud aja mõju ............................................................................. 31 3.2.2. Liikide arvu tüvel mõjutavad faktorid ....................................................................................... 32 3.2.3. Liigiline koosseis....................................................................................................................... 34
4. ARUTELU .............................................................................................................................................. 38 4.1. Torikseened surnud säilikpuudel ...................................................................................................... 38
4.1.1. Puuliigi ja puu surmast möödunud aja mõju ............................................................................. 38 4.1.2. Liikide arvu tüvel mõjutavad faktorid ....................................................................................... 41 4.1.3. Liigiline koosseis....................................................................................................................... 42 4.1.4. Looduskaitseliselt huvipakkuvad liigid ..................................................................................... 44
4.2. Epiksüülsed samblikud surnud säilikpuudel .................................................................................... 45 4.2.1. Puuliigi ja puu surmast möödunud aja mõju ............................................................................. 45 4.2.2. Liikide arvu tüvel mõjutavad faktorid ....................................................................................... 47 4.2.3. Liigiline koosseis....................................................................................................................... 47
4.3 Järeldused .......................................................................................................................................... 49 KOKKUVÕTE ............................................................................................................................................ 51 SUMMARY ................................................................................................................................................ 53 TÄNUSÕNAD ............................................................................................................................................ 55 KASUTATUD KIRJANDUS ..................................................................................................................... 56 LISAD ......................................................................................................................................................... 65

2
1. SISSEJUHATUS
1.1. Säilikpuude mõiste ja eesmärk
Viimase mõnekümne aasta jooksul on üleilmselt üha enam hakatud tähelepanu pöörama
inimtegevuse tõttu kiirelt kahaneva elurikkuse säilitamisele, sh intensiivse tööstusliku
metsanduse mõjude leevendamisele. Pidevalt kasvava ressursivajaduse taustal on metsamajandus
viinud laialdase elupaikade vaesumise ja fragmenteerumiseni (Esseen et al. 1992, Siitonen 2001).
Metsaökosüsteemide olukorraga arvestades on arendatud mitmeid uusi, nn säästvaid
metsamajanduse võtteid. Need lähtuvad muuhulgas põhimõttest, et metsanduspraktika peaks
jäljendama looduslike häiringurežiimide (tulekahjude, üleujutuste, tormimurdude) toimet, et
säiliks selle režiimiga kohastunud elustik (Fries et al. 1997, Lindenmayer & Franklin 2002).
Parasvöötme metsade majandamisel laialdaselt kasutatav lageraie erineb oluliselt looduslikest
puistuvahetushäiringutest, sest lageraie tagajärjel tekib üheealine puistu, kus on vähe
jäänukstruktuure (suuri elus ja surnud puid) ning seetõttu puuduvad elutingimused nendega
seotud metsaelustikule. Sellest puudusest lähtuvalt ongi viimase kahekümne aasta jooksul levinud
raiesmikele säilikpuude (ingl k retention trees, Eesti õigusaktides säilikpuud e elustiku
mitmekesisuse tagamiseks jäetavad puud) jätmise praktika.
Säilikpuud on loodusliku häiringu järgselt alles jäänud elusate ja surnud jäänukpuude analoogid,
millel on kolm peamist looduskaitselist ülesannet (Franklin et al. 1997, Kaila et al. 1997,
Lindenmayer & Franklin, 2002): (1) aidata enne raiet puistut asustanud kitsa elupaiga- või
kasvukohanõudlusega ja halva levimisvõimega organismidel üle elada periood järgmise
metsapõlvkonna tekkeni, (2) pakkuda spetsiifilisi mikroelupaiku metsaliikidele järgmises
metsapõlvkonnas, sh häiringuliikidele raiesmikel ja mitmekesistada tekkiva puistu struktuuri,
ning (3) aidata nn astmelaudadena ühendada fragmenteerunud maastikku. Nendest eesmärkidest
lähtuvalt tuleb Eesti Metsaseaduse järgi (§ 13 1–2) raielangil säilitada kasvavaid puid või nende
säilinud püstiseisvaid osi kogumahuga vähemalt 5 m3 ühe hektari kohta, kusjuures tüved
valitakse erinevate puuliikide suurima läbimõõduga puude hulgast, eelistades kõvalehtpuid,

3
mände (Pinus sylvestris) ja haabasid (Populus tremula). Lisaks on eelistatud vanad,
väljakujunenud võra, õõnsuste või muude omapäradega tüved. Paraku suur osa säilikpuudest
sureb mõne raiejärgse aasta jooksul (Rosenvald et al. 2008). Peale surma rikastavad säilikpuud
noort metsa tüügaspuu1 või jämeda lamatüvena, mis on ainuvõimalikud elupaigad mitmetele
kõdupuidust sõltuvatele organismidele (Franklin et al. 1997).
1.2. Surnud säilikpuud ja nende elustik
Säilikpuude bioloogilist mõju uurivad tööd on seni traditsiooniliselt keskendunud elusatele
puudele. Ehkki säilikpuud on olulised tagamaks suuremõõdulise kõdupuidu (ingl k coarse woody
debris, CWD) pidevat lisandumist kasvavas metsas (Vanha-Majamaa & Jalonen 2001), on
säilikmetsanduse mõju kõdupuidust sõltuvatele liikidele uuritud vähe ja ühekülgselt. Surnud
säilikpuid on uuritud peamiselt Põhjamaades, kus aastaks 2010 ilmunud 23-st surnud säilikpuid
käsitlevast tööst 21 keskendus mardikatele jt lülijalgsetele (Gustafsson et al. 2010); sh käsitlevad
nendegi hulgast vaid vähesed oma elutsüklis otseselt kõdupuidust sõltuvaid puidumardikaid.
Ehkki oma elutsükli jooksul täiel määral või osaliselt kõdupuidust sõltuvaid organisme võib
pidada säilikmetsanduse otseseks „sihtgrupiks“, pole selliseid liigirühmi säilikpuudega seoses
kuigivõrd uuritud. Surnud säilikpuud võiksid fragmenteerunud majandusmetsas olla muuhulgas
olulised kahe kõdupuidust oluliselt sõltuva seenerühma: torikseente ja puidul kasvavate
lihheniseerunud seente e epiksüülsete samblike2 puhul.
Suur osa kõdupuidul kasvavatest torikseene- ja epiksüülsete samblike liikidest on sellest otseses
sõltuvuses, st ei saa kasvada ühelgi teisel substraadil. Morfoloogilise rühmana eristatavate
torikseente hulka kuulub maailmas umbes 1200 liiki (Eestis 211 liiki, Niemelä 2008) erinevatest
perekondadest, millest enamikul on ainsaks kasvusubstraadiks puit (Mueller et al. 2007).
1 Kõik vähemalt 1,5 m kõrgused, rinnasdiameetriga vähemalt 14 cm (kuid minimaalne diameeter sõltub uuritavast
liigirühmast) seisvad surnud puud (Lõhmus & Lõhmus 2001, Bunnell et al. 2008, jt).
2 Epiksüülseteks e puidul kasvavateks samblikeks loetakse käesolevas töös ka mõned traditsiooniliselt jalgsamblike e
kalitsioidsete samblike (ingl k calicioid, matseediumiga ja/või jalaga apoteetsiume omavad) hulka arvatud
lihheniseerumata saprotroofsed seened, mis on samblikele süstemaatiliselt lähedased (näiteks liigid perekondadest Mycocalicium ja Chaenothecopsis).

4
Torikseened eelistavad elupaigana suurema niiskustasemega lamapuid (Heilmann-Clausen &
Christensen 2004, Lindhe et al. 2004) ning asustavad oluliste lagundajatena kogu kõdutüve.
Lihheniseerunud seeni kasvab kõdupuidul teadaolevalt umbes 550 liiki, neist 132 liiki on
kõdupuidule spetsialiseerunud ega saa kasvada kusagil mujal. Sellised samblikud on enamasti
koorikja, kasvupinnale liibunud kasvuvormiga ja paljunevad eostega (Spiribille et al. 2008).
Erinevalt saprotroofsetest torikseentest kõdupuidule spetsialiseerunud samblikud enamasti ise
puidu lagundajad ei ole ja kasutavad puitu vaid kinnitumiseks. Kui torikseened on
niiskuslembesed, siis valgus- ja kuivustolerantsematel samblikel on vastavalt konkurentsieelis
tüügaspuudel. Seepärast on kooreta tüügaspuud elupaigaks ainulaadsetele samblikukooslustele,
sh mitmele haruldasele samblikuliigile (Lõhmus & Lõhmus 2001).
1.3. Surnud säilikpuude torikseene- ja samblikuelustikku mõjutada
võivad faktorid
1.3.1. Mikroklimaatilised tingimused
Kuni puurinde taastumiseni erinevad raiesmiku ja vana metsa mikroklimaatilised tingimused
väga palju. See võib tingida mõningate torikseene- ja samblikuliikide eelise teiste ees. Põhiline
erinevus seisneb selles, et puude eemaldamisel valgus intensiivistub ja avatus tuultele suureneb.
Need tegurid tingivad kiirema aurumise, kuivuse ning päikesele avatud pindade kuumenemise
(Chen et al. 1999). Seetõttu võib arvata, et raiesmikel on konkurentsieelis kuivus- ja
valgustolerantsematel ning termofiilsematel sambliku- ja torikseeneliikidel.
1.3.2. Tüve omadused
Torikseente ja samblike liigirikkus ja koosseis surnud säilikpuutüvel võivad sõltuda.
peremeespuu liigist. Kui torikseente puhul on teada paljude liikide spetsiifilised eelistused
peremeespuu suhtes (Niemelä 2008), siis samblike puhul peetakse puuliiki liigilise koosseisu ja
liigirikkuse mõjutajana oluliseks seni, kuni puu on elus või koorega kaetud. Erinevused
puuliikide vahel tingib isesugune puukoore struktuur, pH ja veemahutavus (Kuusinen 1996,

5
Lõhmus & Lõhmus 2001), need erinevused aga vähenevad puu koore eemaldudes – nt
tüügaspuude puidu happesuse tase ei erine märkimisväärselt liigiti ega okaspuude ja lehtpuude
vahel (Lõhmus & Lõhmus 2001). See võib olla põhjuseks, miks kase- (Betula spp.), sanglepa-
(Alnus glutinosa), kuuse- (Picea abies) ja männitüükad osutusid Eestis nii liigirikkuse kui
liigilise kooseisu poolest üsna sarnaseks (Lõhmus & Lõhmus 2001). Samas on siiski tuvastatud
üksikute samblikuliikide eelistusi peremeespuu osas. Näiteks on Soomes ja Rootsis kaitse all
olevat varjusamblike perekonna esindajat Chaenotheca gracillima leitud vaid lepatüügastelt
(Holien 1996).
Lisaks puuliigile võib torikseene- ja samblikuelustik sõltuda ka tüve läbimõõdust. Jämedaid
surnud tüvesid asustab rohkem liike, sh haruldusi (Renvall 1995, Kruys et al. 1999, Humphrey et
al. 2002). Liigirikkuse ja puu läbimõõdu positiivne seos väljendab eelkõige seda, et jämedam tüvi
pakub rohkem mikroelupaiku (Humphrey et al. 2002), suudab hoida stabiilsemaid
niiskustingimusi ja püsib kauem (kõduneb aeglasemalt; Renvall 1995, Botting & DeLong 2009).
Puidul kasvavate samblike puhul pole siiski tuvastatud, et mõne konkreetse liigi esinemist
limiteeriks kasvusubstraadi ebapiisav läbimõõt. Seevastu torikseentel on selliseid seoseid leitud
küll, näiteks peetakse Soome põlismetsade indikaatorliike Skeletocutis stellae (täht-peenpoorik)
ja Antrodia infirma (pehme korgik) sõltuvaks jämedast kõdupuidust (Renvall 1995).
Kõdupuidul kasvavate koosluste liigiline koosseis ja liigirikkus muutuvad tüve kõdunemise
käigus. Nii torikseentel kui puidul kasvavatel samblikel on liigirikkus kõige suurem keskmises
kõdustaadiumis, kus koos kasvavad nn varased ja hilised liigid (Renvall 1995, Crites & Dale
1998, Forsslund & Koffman 1998, Kruys et al. 1999). Mõlema liigirühma puhul hakkab edasise
kõdunemise käigus liigirikkus vähenema, kuid samblike puhul on sellist liigirikkuse muutust seni
kirjeldatud vaid lamatüvedel, mis erinevad tüügaspuudest kõdunemistingimuste ning
konkurentsisuhete poolest (hilisemates kõduastmetes hõivavad puidupinna maapinnal kasvavad
samblad; Botting et al. 2009). Samas, arvestades et näiteks enamik tüügaspuudega seostatud
kalitsioidseid liike üldiselt pehmet puitu ei eelista (Rikkinen 1995), võib sarnane seaduspära
kehtida ka tüügaspuude epiksüülsete samblike koosluste kohta. Torikseente puhul on leitud, et
igal kõdustaadiumil on omad spetsialistliigid, mida ei leidu varasemates ega hilisemates
kõdunemisfaasides (Renvall 1995).

6
Torikseente puhul on leitud ka, et liikide võime mingit kasvukohta asustada võib sõltuda puidu
tihedusest. Majandusvõtete, näiteks harvendusraie tõttu on majandusmetsas loodusmetsadega
võrreldes kiirem puude kasv ja väiksem puidu tihedus (Loehle 1996). See asjaolu mõjutab
torikseente kooslusi: kuna põlismetsa liigid (nt kaitsealune Fomitopsis rosea; roosa pess) võivad
olla kohastunud aeglasekasvulise puu koloniseerimisele, võivad need seetõttu majandusmetsas
konkurentsis alla jääda liikidele, mis ei nõua nii spetsiifilist elupaika (nt Fomitopsis pinicola;
kännupess; Edman et al. 2006).
1.3.3. Biootilised tegurid
Lisaks kõdunemise käigus toimuvatele füüsikalistele ja keemilistele muutustele puidus (Harmon
1982), võib epiksüülsete samblike ja torikseente liigilise koosseisu muutumist kõdupuidul (sh
surnud säilikpuudel) mõjutada liikidevaheline konkurents. Puidul kasvavatest samblikest on
suurem konkurentsivõime mittesuguliselt paljunevatel liikidel (Ellis & Coppins 2007).
Eospaljunejatest puidule spetsialiseerunud samblike osas on sellistest seostest vähe teada.
Üksikutes töödes on küll tuvastatud seaduspärasid, mille järgi võib välja tuua nn varased ja
hilised puidul kasvavad samblikud (Söderström 1988, Botting et al. 2009), kuid pigem võib
suurem esindatus teatud kõdunemise faasis olla tingitud kasvusubstraadi omadustest kui
konkurentsist (Hilmo et al. 2009).
Torikseente puhul saab liikide reeglipärast koosesinemist ja järgnemist lisaks substraadi
omadustele (sama puuliik, sarnane mikrokliima ja kõdunemise staadium; Ovaskainen et al.
2010a) põhjendada ka liikidevaheliste suhetega, mis määravad ühe või teise liigi edu
kasvusubstraadi tüvesisesel hõivamisel ja viljakeha loomisel. Surnud puutüvesid asustab mütseeli
või soikeseisundis olevate levistena3 oluliselt suurem hulk seeneliike kui seda viljakehade järgi
võiks arvata (Ovaskainen et al. 2010b, Rajala et al. 2010). Laborikatsed on näidanud, et need
liigid konkureerivad ruumi ja/või ressursi pärast (Boddy 2000, Heilmann-Clausen & Boddy
2005).
3 Seene-eosed või mütseeli fragmendid, mis leiduvad elusa või surnud puu maltspuidus, kuid ei saa seal ebasobivate tingimuste tõttu arenema hakata (Boddy 2001)

7
Torikseente puhul võib koosluste arengut surnud puudel mõjutada see, milline lagundajaliik on
esimene koloniseerija. Erinevad lagundajaliigid mõjutavad puidu omadusi isesugusel moel
(Heilmann-Clausen & Boddy 2005) ning tingivad seetõttu erineva koosluse arenguraja (Renvall
1995, Holmer et al. 1997). Nii on näiteks kaitsealuse liigi Fomitopsis rosea käivitatud
kõdunemisprotsess sageli hilisemates lagustaadiumites väga liigirikas, sh esinevad sageli
looduskaitseliselt väärtuslikud liigid (nt Phellinus ferrugineofuscus; tumepruun taelik ja
Amylocystis lapponica; poropoorik). Seevastu sagedaste perekonna Trichaptum spp. (kõbjuk)
esindajate käivitatud koosluste arengurada on reeglina oluliselt liigivaesem (Renvall 1995).
Selline järgnevus tähendab, et üht liiki seene vähenemine võib langetada ka mitme teise liigi
arvukust (Renvall 1995, kuid vt ka Komonen et al. 2000 taksoniüleste seoste kohta).
1.3.4. Liikide levimisvõime
Lähtuvalt jäänukstruktuuride (sh surnud säilikpuude) nappusest majandusmetsas võib nende
asustamine oluliselt sõltuda liikide levimisvõimest. Kõdupuit on ajutine elupaik, seetõttu
arvatakse, et sellele kohastunud organismidel on hea levimisvõime (Jonsson et al. 2005). Kuid
edukaks levimiseks peab siiski olema tagatud ka piisav sobiva substraadi hulk (Crites & Dale
1998, Olsson et al. 2011).
Puidul kasvavate samblike puhul on andmed liikide levimisvõime kohta vastuolulised.
Kõdupuidule, sh tüügaspuudele spetsialiseerunud samblikuliigid paljunevad enamasti eoseliselt
(Forsslund & Koffman 1998, Spiribille et al. 2008). Selle paljunemisstrateegia eeliseks on
levikukaugus (eosed on väiksemad ja levivad seetõttu kaugemale kui soreedid või isiidid,
Seymour et al. 2005). Samas ei ole üheselt selge, kui kaugele samblikud (nii eos- kui
vegetatiivselt paljunejad) levivad. Erinevad uurimused on kohati jõudnud ka samade liikide puhul
erisuguste tulemusteni (e.g. Walser et al. 2005 vs Öckinger et al. 2005 Lobaria pulmonaria;
hariliku kopsusambliku kohta). Siiski on teada, et sobivatel tingimustel suudavad samblikud
kasvamiseks ebasobivaid alasid edukalt ületada (Kryus & Jonsson 1997). Näiteks Eestis on
leitud, et esimese põlvkonna metsades esines kõdupuidu spetsialistidele uusi mikroelupaiku küll
vähe, kuid kui vanem metsaala (levikuläte) polnud kaugemal kui paarsada meetrit, oli esimese
põlvkonna metsade kõdupuidu sambliku ja samblakooslus kõigi näitajate poolest vanema

8
metsaga samaväärne, sh oli esindatud isegi enamik hemerofoobsetest liikidest (Lõhmus &
Lõhmus 2008).
Torikseente puhul on küll leidnud kinnitust liikide võime kaugele levida, kuid see ei pruugi
tagada edu kasvusubstraadi asustamisel. Torikseente elujõulised eosed võivad jõuda levikukohast
isegi kaugemale kui 300 km (Risbeth 1959). Samas on dokumenteeritud, et mõningate liikide
levik suuremas maastikumosaiigis võib siiski olla takistatud (Edman et al. 2004a, 2004b).
Enamik eostest langeb seene viljakehast mõne meetri raadiusesse (Edman et al. 2004a). See
tähendab, et levikulätte läheduses asuv kasvusubstraat hõivatakse suurema tõenäosusega, kuid
täpne seos eoste hulga ja asustamise edukuse vahel ei ole siiski teada. Seente levikumustrite
kirjeldamise muudab keerulisemaks ka asjaolu, et lisaks tuullevile võivad seened levida ka
loomade abil (Vasiliauskas & Stenlid 1999, Farris et al. 2004) ning vegetatiivselt mütseeli teel
(Boddy 2001). Samas toimub seente levimine puudesse juba ka enne nende surma ning varem
kohale saabunud ning puu surmani soikeseisundis olevatel levistel võib tüve hilisemal
lagundamisel olla eelisseisund (Boddy 2001).
1.4. Senised uuringud surnud säilikpuude torikseentest ja samblikest
Autorile teadaolevalt on torikseeni seoses säilikpuudega käsitletud kahes uuringus, mõlemad
põhinevad viljakehade määramisel. Lõuna-Rootsis inventeeriti kaheksa aasta jooksul
seenekooslusi katseraiesmikele ja kõrvalolevatesse metsadesse jäetud tüügas- ja lamapuudel
(Lindhe et al. 2004). Leiti, et liigirikkus raiesmikel ja metsades ei erinenud, küll aga tõid erinevad
valgustingimused kaasa mõnevõrra erineva liigilise koosseisu. Soomes läbiviidud säilikpuude
uuringus (Junninen et al. 2006) võrreldi kuni 5 aastat ja 15 aastat kõdunenud säilikhaavatüvesid
metsas ja raiesmikul ning leiti, et keskmiselt kõdunenud puud on raiesmikul liigirikkamad kui
metsas (sh olles kasvupinnaks mitmele looduskaitseliselt olulisele liigile). Järeldati, et mitmed
haavale kui varasuktsessioonilisele puule spetsialiseerunud torikseeneliigid võivad olla ühtlasi
kohastunud häiringualade avatud tingimustele. Kirjeldatud uuringute peamiseks puuduseks on, et
need hõlmavad väga piiratud maa-alasid kaitsealade vahetus läheduses. Seetõttu võib töödes
dokumenteeritud liigirikkus ja liigiline koosseis olla suuresti mõjutatud ümbritseva metsa

9
vanusest ja kõdupuidu rohkusest, sest neist asjaoludest sõltub kohalik levisepank ja seetõttu ka
surnud säilikpuude koloniseerimine torikseeneliikide poolt (Edman 2004b). Lisaks sellele
käsitlevad mõlemad tööd kunstlikult tekitatud kõdupuitu, st raiutud või mürgitatud puid
(Junninen et al. 2006 ühe töötlusena ka loomulikult surnud säilikpuud; kuid töö fookus oli
surmatud puudel), mis on substraadina erinev näiteks parasiitseente poolt surmatud tüvedest. Eri
põhjustel surnud puudele tekivad erinevad seenekooslused ning erinev on ka koosluste
arengurada (Renvall 1995, Heilmann-Clausen & Christensen 2004).
Võrreldes torikseentega on epiksüülsed samblikud, sh ka säilikpuudel kasvavad liigid, suhteliselt
väheuuritud elustikurühm. Seni puuduvad uuringud sellest, kui palju kulub aega eripäraste
epiksüülisete samblike koosluste tekkeks ning millised on selleks vajalikud kasvusubstraadi
omadused. Osaliselt tingib info vähesuse see, et ehkki puidul ja eriti puutüügastel kasvavaid
samblikukooslusi on mitmetes uuringutes nimetatud liigirikkaks ja eripäraseks (e.g. Holien 1996,
Forsslund & Koffman 1998, Humphrey et al. 2002, Lõhmus & Lõhmus 2001, Nascimbene et al.
2008), puudub teaduskirjanduses nende kasvukohtade ühtne käsitlus. Nii leidub uuringuid, kus
epifüütseid samblikke käsitledes ei tehta vahet puukoorel ja puidul kui erinevatel kasvupindadel
(e.g. Moen et al. 2002, Botting et al. 2009); kuid ka selliseid, kus teistest eristatakse küll
erinevatel puidupindadel (nt lamapuude ja tüügaspuude puidul ning kändude lõikepindadel)
kasvavad samblikud, kuid analüüsides vaadeldakse neid ühtse epiksüülsete samblike rühmana
(e.g. Forsslund & Koffman 1998, Kruys et al. 1999).
1.5. Magistritöö eesmärgid
Käesolev uuring keskendub torikseentele ja epiksüülsetele samblikele (nende liigirikkusele ning
vastavate koosluste tekkele ja arengule) kui kõdupuidust sõltuvale elustikurühmadele, mida on
surnud säilikpuudel vähe uuritud. See töö on ühtlasi jätk säilikpuude bioloogilist mõju uurivate
tööde seeriale (Lõhmus et al. 2006, Rosenvald & Lõhmus 2007, Jairus et al. 2009). Uuringu
uudsus tuleneb aegrealisest disainist: koosluste kirjeldamisel lähtutakse teadaolevast surma ajast
kuni 8 aastat surnud säilikpuutüvedel.

10
Töös keskenduti kahele küsimusele:
1. Millised puu omadused (puuliik, tüve läbimõõt, puidu kõdustaadium, valgustatus ja kaugus
metsast kui levikulättest) ja raiesmiku omadused (kõdupuidu hulk) mõjutavad torikseente ja
epiksüülsete samblike esinemist ja liigirikkust (samblikel ka katvust)? Selliste faktorite võimaliku
mõju tundmine lubab esiteks paremini mõista kahe uuritava liigirühma nõudlusi kasvusubstraadi
osas. Teiseks saaks võimalikke mõjusid arvesse võtta metsamajandamise reguleerimisel (näiteks
säilikpuude valiku kohandamisel) ja nii tagada nimetatud rühmade parem kaitse.
2. Kuidas muutuvad surnud säilikpuude seene- ja samblikukooslused ajas? Uuriti, milline on
torikeente ja puidul kasvavate samblike liigiline koosseis ning kuidas see muutub seisvatel ja
lamavatel surnud säilikpuudel 1–8 surmajärgsel aastal. Erinevalt varasematest analoogilistest
uuringutest torikseentel (Lindhe et al. 2004, Junninen et al. 2006), analüüsitakse koosluste tekke
ja liigirikkuse seaduspärasid laiemas maastikumosaiigis, st mitme üksteisest eemal asuva
uuringuala säilikpuudel. Surnud säilikpuid epiksüülsete samblike võimaliku kasvusubstraadina
majandusmetsas pole varem uuritud. Epiksüülsete samblike osas on see uuring teadaolevalt
esimene, mis üldse käsitleb neile vajaliku elupaiga – paljandunud puidupinna – teket ning
samblikukoosluste arengut tüügaspuude puidul.

11
2. METOODIKA
2.1. Uurimisalad ja välitööde planeerimine
Uurimisalad hõlmasid osa Rosenvaldi ja kaasautorite (2008) säilikpuude elumuse uurimise
proovialadest. Need asusid Tartu, Viljandi, Järva ja Pärnu maakonnas (endises Kabala, Kõpu,
Laeva ja Alatskivi metskonnas), vahemikus 58º–59º põhjalaiust ja 25º–27º idapikkust (Joonis 1).
Rosenvaldi ja kaasautorite (2008) uurimus hõlmas nendes metskondades 102 ala, kus toimus raie
aastatel 2001–2002.
Joonis 1. Uurimisalade (48) paiknemine Tartu, Viljandi, Järva ja Pärnu maakonnas
Käesoleva töö planeerimisel lähtuti ülalviidatud uurimuse andmetest, mida on iga-aastaselt
täiendatud ka peale vastava artikli ilmumist. Seega kajastas andmestik 2010. aasta kevadeks
(valimi koostamise aeg) infot säilikpuude kohta kaheksal raiejärgsel aastal. Andmestik sisaldab
kõigi hõlmatud alade säilikpuude surmast möödunud aega (üheaastase täpsusega) ja surmajärgset
asendit (püsti, pikali). Käesoleva töö valim koostati surnud säilikpuid kirjeldavate andmete põhjal

12
eesmärgiga, et ühe puuliigi siseselt oleks kaheksa vaadeldud aasta jooksul võimalikult ühtlaselt
esindatud erinevad tüvede „vanusegrupid“ vastavalt surmast möödunud ajale, (sh nii tüügas- kui
lamapuudena). Sellistele kriteeriumite vastav valim oli võimalik koostada kolme kõige
sagedasema säilikpuuliigi: kase, hariliku männi ja hariliku haava kohta. Väljavalitud puud
jaotusid juhuslikult 48 alale (Joonis 1, Lisa 1), mille keskmine pindala oli 2,9 ha (vahemikus 0,5–
6,9 ha) ja kuhu oli keskmiselt jäetud 12,9 elusat säilikpuud/ha (vahemikus 3–46), peamiselt kask
(38%), harilik mänd (18%), harilik haab (18%) ja harilik saar (Fraxinus excelsior, 7%). Teistest
puuliikidest olid esindatud harilik pärn (Tilia cordata), sanglepp, kuusk, harilik jalakas (Ulmus
glabra), hall lepp (Alnus incana), remmelgas (Salix spp.), harilik tamm (Quercus robur) ja harilik
vaher (Acer platanoides). Aladel oli valdavaks naadi metsakasvukohatüüp (40%), enam esines
veel jänesekapsa-mustika (21%) ja angervaksa (10%) ning vähem karusambla-mustika ja mustika
kasvukohatüüpi kuuluvaid proovialasid (Lisa 1). Ehkki näiteks karusambla-mustika ja mustika
kasvukohatüüpe võiks teistest oluliselt kuivemateks pidada, kasvas sellistel aladel lisaks
mändidele ka kaskesid ja haabasid, mis viitab nende suhteliselt kõrgele niiskustasemele ja
viljakusele, seetõttu peetakse alasid niiskustingimuste poolest üldjoontes omavahel
võrreldavateks.
Valimisse võeti kokku 94 säilikpuudest tekkinud tüügas- ja 132 lamapuud (Tabel 1 ja 2). Valimi
koostamise eesmärk oli puutüvede „vanusegruppide“ ühtlus 8-aastase perioodi lõikes. Suurimaks
kõrvalekaldeks sellisest ühtlasest jaotumisest on hiljuti surnud mändide ja haabade
proportsionaalselt väiksem esindatus (Tabel 1 ja 2). See kõrvalekalle on tingitud säilikpuude
elumuse raiejärgsest dünaamikast. Nimelt on näidatud (Rosenvald et al. 2008), et raiejärgselt
kõrge säilikpuude suremus aja jooksul väheneb, mistõttu selliseid tüvesid katsealadel rohkem
polnud. See valimi probleem lahendati eri moel sõltuvalt analüüsist.
Osa lamapuudest (5 kaske ja 6 haaba) erines teistest kõdunemise ajaloo poolest, olles enne
maapinnale langemist mingi aja vältel ka surnult seisnud. Kirjanduse põhjal on teada, et
(vähemalt viljakehade järgi otsustades) ei ole tüügaspuud enamikule torikseentele eelistatud
kasvusubstraat (e.g. Lindhe et al. 2004) ja seetõttu arvestati liigirikkust puudutavates analüüsides
(ptk 2.4.1.) selliste puude surmast möödunud aega siiski alates maapinnale langemisest. Samas
toimuvad ka seisvates puudes seentest tingitud kõdunemisprotsessid, mis võivad mõjutada

13
torikseente koosluste liigilist koosseisu hilisemal lamatüvel (Renvall 1995, Heilmann-Clausen &
Christensen 2004). Seetõttu eristati kooslusi puudutavates analüüsides sellised tüved ülejäänutest
(ptk 2.4.2.).
2.2. Välitööd
Välitööd viidi läbi 2010. aasta suvel ja sügisel. Välitööde käigus kirjeldati torikseene- ja
samblikukooslusi. Lisaks kasutati lamapuude kirjeldamiseks kuut ning tüügaspuude
kirjeldamiseks seitset tunnust (Tabel 3), sh puidu tiheduse mõõtmiseks digitaalset mikropuuri
(resistograafi) ja valgustingimuste hindamiseks poolsfäärilisi fotosid kalasilm-objektiiviga.
2.2.1. Puidu tiheduse mõõtmised
Tüügaspuudel ja lamatüvedel kasutati erinevat kõdunemuse hindamise metoodikat. Kui
lamatüvedel määrati kõdunemust traditsiooniliselt „noameetodil“, kõduastmete kaudu (Renvall
1995; Tabel 3), siis tüügaspuudel kasutati täpsemat puidu tiheduse mõõtmise tehnikat.
Tüügaspuude puidu tiheduse mõõtmiseks kasutati resistograafi, mis on teadaolevalt täpseim
mõõteseade mittedestruktiivseks puidu tiheduse hindamiseks välitingimustes (Kahl et al. 2009).
Seadme traditsiooniline kasutusvaldkond on pargindus (võimaldab hinnata vanade pargipuude
elumust) ja ehituspuidu analüüs, viimasel ajal on seda tehnoloogiat katsetatud ka kõdupuidu
tiheduse hindamiseks (Creed et al. 2004, Kahl et al. 2009). Tegu on kaasaskantava digitaalse
mikropuuriga, mis läbib puutüve ning võimaldab selle tihedust hinnata puidu poolt puurile
tekitatud vastupanujõu kaudu. Meetodi eeliseks on mõõteseadme kaasaskantavus välitöödel.
Puuduseks on puurimisandmete lokaalsus (Kahl et al. 2009). Käesolevas töös kasutati Inglismaa
firma Sibtec Technology resistograafi DmP (Digital microProbe). See on akuga varustatud seade,
mille puuri diameeter on 1 mm ja millega saab puurida sügavuseni 500 mm. Seadme puur teeb
7000 pööret minutis ja mõõdab puurimise ajal tekkivat vastupanu iga 0,1 mm tagant. Tehtud
puurimiste kohta väljastatud andmed salvestatakse programmi Microsoft Excel failidena
resistograafiga ühendatud ja vastava tarkvaraga varustatud sülearvutisse.

14
Tüügaspuudel olid peamiseks huvipakkuvaks liigirühmaks puidul kasvavad samblikud, kes
erinevalt torikseentest enamasti ise puidu lagundajad ei ole ning kelle hüüfid ulatuvad vaid puidu
pinnakihti (Büdel & Scheidegger 1996). Seetõttu sooviti puidu tihedust kirjeldada eelkõige tüve
pinnakihis. Et vähendada puurimise lokaalsuse mõju (näiteks puuri sattumine puiduprakku)
otsustati pinnakihina käsitleda peamist viit sentimeetrit puutüvest.
Iga tüve puuriti resistograafiga nii jalamilt (0,2 m) kui rinnakõrguselt (1,3 m) kaks kuni neli
korda (Joonis 2). Puurimiste arv sõltus tüve läbimõõdust: puid läbimõõduga alla 20 cm puuriti
mõlemalt kõrguselt kaks korda tüve läbivalt, jämedamaid mõlemal kõrgusel neli korda
hinnanguliselt tüve keskkohani. Fikseeriti ka puu ümbermõõt puurimise kõrgusel ja koore
olemasolu puuri sisenemise kohal. Kui puurimise koht oli koorega kaetud, eemaldati koor
mõõtmisvea vältimiseks.
Tabel 1. Surnud säilikpuudest tekkinud lamatüved (L) ja tüügaspuud (T), millel uuriti torikseeni:
tüvede arv, jaotumine tüve surmast möödunud aja järgi ja torikseentega asustatud tüvede arv
(sulgudes).
Puu Tüvesid kokku 1–2 a 3–4 a 5–6 a 7–8 a L T L T L T L T L T Kask 48(43) 33(33) 11(9) 9(9) 14(13) 7(7) 11(10) 8(8) 12(11) 9(9) Haab 43(41) 35(25) 8(6) 6(5) 11(11) 12(8) 12(12) 8(6) 12(12) 9(6) Mänd 41(31) 26(9) 6(0) 2(0) 7(7) 4(2) 15(12) 8(4) 13(12) 12(3)
ΣΣΣΣ 132(115) 94(67)
Tabel 2. Surnud säilikpuudest tekkinud tüügaspuud, millel uuriti puidul kasvavaid samblikke:
tüvede arv, jaotumine tüve surmast möödunud aja järgi ja puidul kasvavate samblikega asustatud
tüvede arv (sulgudes).
Puu Tüvede arv 1–2 a 3–4 a 5–6 a 7–8 a Kask 33(14) 9(2) 7(2) 8(3) 9(7) Haab 35(24) 6(0) 12(7) 8(8) 9(9) Mänd 26(26) 2(2) 4(4) 8(8) 12(12) ΣΣΣΣ 94(64)

15
Tabel 3. Uuringu tüvesid, raiesmikke ja kooslusi kirjeldavad tunnused
Tunnus Selgitus Tüügaspuid ja lamatüvesid kirjeldavad tunnused Tüve surmast möödunud aeg (a) Kirjeldavatel joonistel ja ordinatsioonanalüüsis
jagatud kaheaastasteks „vanusegruppideks“: 1–2a; 3–4a; 5–6a; 7–8a
Kõduaste „noameetodil“ 1 - nuga pole tüve puitu sisse surutav; 2 - nuga on puitu sisse surutav 1–2 cm ulatuses; 3 - nuga on puitu kergesti sisse surutav 3–5 cm ulatuses; 4 - nuga on puitu üleni sisse surutav; 5 - täielikult lagunenud puit, pudeneb käte vahel laiali
Puidu tihedus 5cm pinnakihis Tüügaspuudel mõõdetud resistograafiga (ptk 2.2.1.)
Rinnasdiameeter või ümbermõõt (cm) Mõõdetud 1,3 m kõrguselt tüve jalamist Kaugus lähimast metsast (m)* Tüügaspuudel ja lamatüvedel Koore katvus (%)**
Tüügaspuudel 2m ulatuses Lamatüvedel kogu tüvel***
Valgustatus Tüügaspuudel mõõdetud poolsfääriliste fotodega (ptk 2.2.2.) Lamatüvedel 1 - varjus; 2 - rohu ja puhmarinne varjavad osaliselt; 3 - päikesele avatud
Raiesmikku kirjeldavad tunnused Lamapuude maht (m3/ha)* Arvestatud vähemalt 14 cm rinnasdiameetriga
säilikpuudest tekkinud lamapuid Vanade tüügaspuude arv raiesmikul (N/ha)* Vähemalt 14 cm rinnasdiameetriga kuivanud
puud ja tüükad (kuivanud juba enne raiet) Torikseene ja samblikukooslusi kirjeldavad tunnused Taksonite arv tüvel Enamasti liik, määramisraskuste puhul perekond
(ptk 2.3.) Torikseente või samblike esinemine tüvel 1/0 Puidul kasvavate samblike suhteline katvus joontransektil
ptk 2.2.3., Joonis 3
Puidul kasvavate samblike katvus liigiti 1 - katab kuni 1 cm2; 2 - katab 1–25 cm2; 3 - katab 25–100 cm2; 4 - katab üle 100 cm2
* mõõdetud eelnevate uuringute käigus (Rosenvald et al 2008) ** paljandunud puidupind = 1– koore katvus *** lamatüvedel on koore katvuse sisse arvestatud ka pind, mis on kaetud samblaga

16
Enne analüüsi tuli resistrograafi andmeid puhastada ning ekstrapoleerida, et vältida andmete
anomaaliaid, mis tekivad puuri sisenemisel ja väljumisel või mõõtesammu suurenemisel
väiksema tihedusega puidu puurimisel. Andmete puhastamiseks (1) kustutati esimene lugem
andmereas (esimene lugem sügavusel 0,0 mm ei näita vastupanu puurimisel, vaid tugevust,
millega suruti puuri otsaga tüve pinnale enne puurima asumist) ja (2) kustutati arvestuslikule
viimasele lugemile järgnevad andmed. Viimase lugemina arvestati jämedamatel puutüvedel tüve
keskkohta (arvutatud ümbermõõdu järgi) ja peenematel kohta, kus puur väljus teisel pool tüve
(ehkki seoses puuri külghõõrdega puuritavas augus jätkub andmete salvestamine ka peale puuri
väljumist, on väljumiskoht andmestikus näha näitude järsu muutumise järgi). Andmete
ekstrapoleerimise vajadus tekkis pehmema puidu puhul, kus resistograafi mõõtesamm võis olla
kuni 10 võrra tavalisest (0,1 mm) suurem. Vea vältimiseks hilisemas analüüsis ühtlustati kõigi
puurimiste mõõtesammud, tuletades puuduvad andmed eelmisest ja järgmisest salvestatud
lugemist.
Joonis 2. Resistograafiga puurimine sõltuvalt tüve läbimõõdust (vasakul <20cm, paremal
>20cm). Klambritega tähistatud tüve pinnakihina arvestatud andmed
Iga puutüve pinnakihi keskmine puidu tihedus arvutati kokku nelja kuni kaheksa puurimise
andmetelt. Selleks eraldati puhastamise ja ekstrapoleerimise järgselt igast (ekstrapoleeritud ja
puhastatud) andmereast info pindmise (väikese diameetriga „läbipuuritud“ tüvede puhul kahelt
poolt pindmise) 5cm kohta. Seejärel arvutati andmeanalüüsiks kõigi antud tüve pinnakihi
puurimisandmete keskmine.

17
2.2.2. Valgustingimuste hindamine
Tüügaspuude valgustingimusi hinnati poolsfäärilistelt fotodelt. Kasutati kalasilm-objektiiviga
Fisheye Converter FC E8 digitaalset fotoaparaati Nikon Coolpix 4500. Igast puust tehti kaks fotot
tüve rinnakõrguselt: üks põhja- ja teine lõunaküljelt. Pildistati horisontaaltasandil, nii et kaamera
ülemine serv oli suunatud põhja. Fotod tehti septembri esimesel või teisel nädalal. Lamapuude
puhul ei olnud sellist valgustatuse hindamise meetodit otstarbekas kasutada, kuna lamatüvede
valgustingimused varieeruvad tüve lõikes oluliselt. Selle asemel hinnati lamatüvedel valgust
klassidena (Tabel 3).
Fotode analüüsiks kasutati WinSCANOPY tarkvara (Regent Instruments Inc., Quebec, Canada),
arvutamaks iga foto üldine kohafaktor ehk otsese ja hajusa kiirguse läbituleku koefitsient (TSF).
Seejärel arvutati kahe tüvest tehtud foto TSF koefitsientide põhjal iga tüve keskmine valgustatus
(keskmine hajusa kiirguse läbituleku koefitsient).
2.2.3. Torikseente ja epiksüülsete samblike koosluste kirjeldamine
Epiksüülsete samblike kooslusi kirjeldati 2010. aasta juunist oktoobrini. Puidul kasvavate
samblike koosluste kirjeldamiseks hinnati esiteks nende üldkatvust. Samblike üldkatvuse
kirjeldamiseks kasutati joontransektide meetodit, mis võimaldab hinnata epiksüülsete samblike
katvust ühtmoodi kõigil tüvedel, sõltumata koorevaba pinna suurusest. Transektide paigutamine
sõltus sellest, kui palju puidupinda oli paljandunud. Juhul kui koor puudus kogu puutüvel või
kattis tüve vaid vähesel määral, määrati esmalt pikitransekt (1,6 m pikkune transekt piki puutüve)
nii tüve põhja kui lõunaküljes. Ristitransektid paiknesid ümber tüve järgmistel kõrgustel: 1,8 m;
1,5 m; 1 m; 0,5 m; 0,2 m (Joonis 3a). Juhul kui puit oli paljandunud vaid osal puutüvest või
laikudena, uuriti iga paljandunud tüve osa (puidulaiku) vastava pikkusega pikitransekti(de) ja
ristitransekti(de)ga. Väikeste puidulaikude puhul paigutati iga laigu keskkohta üks piki ja üks
ristitransekt. „Laiade“ (nt tüve ümbritsevate) puidulaikude puhul paigutati üks pikitransekt tüve
põhja ja teine lõunaküljele nii nagu ülal kirjeldatud. „Pikkade“ puidulaikude puhul paigutati mitu
ristitransekti, järgides samuti ülalkirjeldatud kõrgusi (Joonis 3b). Transektide paigutamisele
järgnevalt mõõdeti nende pikkused, ning joone alla jäävate puidul kasvavate samblikega kaetud
lõikude pikkus. Kui risti või pikitransekti sisse jäi puukoort, lahutati koorega kaetud transekti osa

18
üldpikkusest. Hilisema andmetöötluse käigus arvutati iga tüve suhteline epiksüülsete samblike
katvus, st transektide pikkuse ja epiksüülsete samblikega kaetud transektide osa suhe. Viimaks
vaadati üle kogu puidupind (kuni 1,8m kõrguseni) ning registreeriti tüvelt kõik samblikuliigid
ning nende katvused (tabel 3).
Joonis 3. Tüügaspuude samblikekoosluste kirjeldamisel kasutatud transektide paiknemine (a)
juhul kui puit on paljandunud kogu puutüvel ja (b) laiguti paljandunud puidu korral.
Torikseenekooslusi kirjeldati 2010. aasta sügisel (septembris ja oktoobris). Töös käsitletav
torikseente liikide arv on nn minimaalne liikide arv uuritud puudel, kuna liike otsiti ainult
viljakehade järgi. Igal lamatüvel ja tüügaspuul otsiti seente viljakehasid nii tüvelt kui okstelt,
võimalusel ka tüve või oksi kergitades. Registreeriti vaid viljakehade olemasolu, nende suurust
või arvu ei kirjeldatud. Mitmeaastaste torikseente surnud viljakehasid analüüsides ei arvestatud.
Arvestades, et nii torikseente kui epiksüülsete samblike puhul võis esimestena uuritud raiesmikel
Laeva metskonnas liike vähese leidmiskogemuse tõttu märkamata jääda, vaadati oktoobri alguses
teist korda üle neli sealset prooviala, mida oli külastatud välitööde alguses.

19
2.3. Torikseente ja samblike määramine
Kergesti äratuntavad torikseene- ja samblikuliigid määrati välitööde käigus, samblike puhul
kasutati vajadusel kümnekordselt suurendavat luupi. Raskemini äratuntavad liigid korjati kaasa
hilisemaks määramiseks laboris. Torikseente puhul märgiti eelnevalt üles määramist hõlbustavad
kuid kuivamise käigus kaduda või muutuda võivad viljakeha tunnused (nt pooride arv
millimeetril, värvus ja selle võimalik muutumine puutel, lõhn, maitse ja rabedus). Kaasakorjatud
torikseente viljakehad kuivatati laboris ning pakiti kuivatamise järgselt puhtasse kilekotti. Autori
poolt kaasa kogutud teaduslikku või looduskaitselist väärtust omavaid seene ja samblikuliikide
proove säilitatakse Tartu Ülikooli Loodusmuuseumi seente või samblike herbaariumis (TU).
Määramiseks vaadeldi torikseente ja epiksüülsete samblike morfoloogilisi ja anatoomilisi
tunnuseid stereo- ja/või valgusmikroskoobiga. Samblike puhul kasutati preparaatide
valmistamiseks vett, torikseente puhul sõltuvalt liigist 5% kaaliumhüdroksiidi (KOH) lahust,
50% joodilahust (Meltzeri lahus, IKI) või 5% puuvillasinise (Cotton Blue, CB) lahust. Kasutati
mitmeid määrajaid: Niemelä 2008, Randlane & Saag 2004, Tibell 1999, Trass & Randlane 1994,
Ryvarden & Gilbertson 1–2 1993, võrdlusmaterjalina kasutati ka Tartu Ülikooli lihhenoloogilise
herbaariumi eksemplare. Samblike ja torikseente nomenklatuur järgib Eesti elurikkuse kodulehe
andmeid (eElurikkus 2011).
Mõned kaasakorjatud sambliku ja torikseeneproovid jäid liigini määramata. Töös uuriti varajasi
epiksüülsete samblike kooslusi hiljuti paljandunud tüügaspuude puidul, seetõttu olid mõned
kaasakorjatud proovid juveniilsed ja väljakujunemata liigitunnustega. Samuti polnud sageli
võimalik kaasa võtta piisavalt suurt proovi, et õhukese kihi kromatograafia meetodi (TLC;
Orange et al. 2001) abil sekundaarseid samblikuaineid määrata. Kõige sagemini jäid perekonna
tasemele Lepraria (löövesamblike, 14 16-st proovist), Cladonia (porosamblike, 15 32-st, sh
enamasti vaid protallust sisaldavad) ning Melanohalea (pruunsamblike, 10 11-st) proovid. Lisaks
jäid liigini määramata üksikud proovid perekondadest Calicium (1), Caloplaca (1), Lecanora (1),
Micarea (1) ja Usnea (3). Kahe steriilset pisisamblikku sisaldava proovi puhul polnud võimalik
anda ka perekonna määrangut. Kahte perekonna Cladonia liiki: Cladonia coniocraea (naaskel-
porosamblik) ja C. ochrochlora (tuhk-porosamblik) on väikeste proovide puhul keeruline

20
eristada, seetõttu loeti need üheks koondliigiks. Torikseente puhul jäi liigini määramata 1 ja
perekonnani määramata 6 proovi. Liigini määramata eksemplarid kaasati analüüsidesse
perekonna tasemel. Liigirikkust puudutavatesse analüüsides vaadeldi perekonnana määramata
proove eri liigina, kui tunnuste alusel oli võimalik neid teistest eristada, kuid mitte nimetada.
2.4. Andmeanalüüs
2.4.1. Liikide esinemise ja liigirikkuse analüüsid
Esiteks võrreldi torikseente ja epiksüülsete samblike liigirikkust sõltuvalt kasvusubstraadi
surmast möödunud ajast. Selleks koostati vabavaralise programmi EstimateS 8.2.0. (Gotelli &
Colwell 2001) abil liikide kumuleerumise kõverad. See programm kasutab liikide
kumuleerumiskõvera koostamiseks (traditsioonilise korduva juhusliku ümberjärjestamise alusel
toimiva analüüsi asemel) analüütilist valemit (Mao Tau funktsioon), mis arvutab oodatava liikide
arvu iga olemasolevast väiksema proovide arvu puhul, ning võimaldab seetõttu võrrelda
liigirohkust ka ebatasakaalulise valimi puhul (Colwell et al. 2004).
Teiseks uuriti torikseente ja epiksüülsete samblike esinemist, liigirikkust ning epiksüülsete
samblike katvust sõltuvalt säilikpuude surmast möödunud ajast ning neid määravaid lisafaktoreid.
Selleks kasutati üldiseid ja üldistatud lineaarseid mudeleid tarkvarapaketis Statistica 7. Torikseeni
ja puidul kasvavaid samblikke uuriti andmeanalüüsil eraldi, torikseeni puudutavates analüüsides
käsitleti omakorda eraldi lama- ja tüügaspuid kui erinevate tingimuste ja omadustega
kasvusubstraate.
Uuritavaid sõltuvaid tunnuseid oli viis: (1) torikseente liikide arv lamatüvel; (2) torikseente
liikide arv tüügaspuul; (3) puidul kasvavate samblike esinemine; (4) puidul kasvavate samblike
liikide arv tüügaspuudel ning (5) puidul kasvavate samblike üldkatvus tüügaspuude tüve
paljandunud puidul. Sõltuvate tunnuste analüüsimiseks kasutati üldisi lineaarseid mudeleid
(GLM, tunnused 1, 4 ja 5) ning üldistatud lineaarseid mudeleid (GLZ, tunnused 2 ja 3; vastavalt
Poissoni jaotus ja binoomne jaotus). Tunnuste ja jääkide normaaljaotusi kontrolliti Kolmogorov-
Smirnovi testiga, kusjuures tunnuste puhul kaasati kõik vaadeldud puutüved, jääkide puhul aga

21
kontrolliti normaaljaotust igal sõltuva muutuja tasandil eraldi. Vajadusel normaliseeriti andmeid
juurteisendusega või arkussiinus juurteisendusega.
Selgitava faktorina käsitleti nii torikseeni kui puidul kasvavaid samblike käsitlevates analüüsides
puuliiki. Pideva kirjeldava muutujana kaasati kõikidesse analüüsidesse tüve surmast möödunud
aeg. Lisaks kaasati sõltuvalt analüüsist järgmiseid pidevaid kirjeldavaid tunnuseid: tüve
diameeter, valgustatus, kõduaste (lamatüvedel) või puidu tihedus tüve pinnakihis (tüügaspuudel),
uuritava puutüve kaugus lähimast metsast; vähemalt 10 cm diameetriga lamapuude maht (m3/ha)
uuritaval raiesmikul ja vanade tüügaspuude hulk raiesmikul. Kõiki tunnuseid on lähemalt
kirjeldatud tabelis 3. Tunnuste omavahelisi seoseid uuriti Pearsoni korrelatsoonitestiga.
Tulemuste erinevust võrreldi post hoc Tukey HSD testiga.
Kõikide analüüside puhul kaasati esimeses etapis mitmetunnuselisesse mudelisse kõik uuritavad
kirjeldavad muutujad ja huvipakkuvad koosmõjud. Teises etapis lihtsustati mudelit, eemaldades
ükshaaval ebaolulised tunnused (alustades suurimast p väärtusest).
Torikseente liikide arvu lamatüvedel ja tüügaspuudel (sõltuvad tunnused, GLM ja GLZ, tüüp III)
uurimiseks kaasati analüüsi (lisaks puuliigile ja tüve surmast möödunud ajale) kirjeldavate
pidevate muutujatena tüve diameeter, valgustatus, kõduaste ja uuritava puutüve kaugus metsast.
Juhtudel, kus lamatüvel oli erinev aeg tüve surmast ja mahalangemisest (ptk 2.1.), kaasati
analüüsidesse vaid viimane. Uuriti ka tüve surmast möödunud aja ja puuliigi koosmõju. Pearsoni
korrelatsioonitestis selgus, et kahel tunnusel: tüve surmast möödunud ajal ja kõduastmel on
statistiliselt oluline positiivne seos (N=131; r=0,49; p<0,05), kuid kuna kõduaste kaasati
analüüsidesse vaid esimeses etapis (osutus mitteoluliseks), ei mõjutanud see seos lõplikke
analüüsi tulemusi.
Puidul kasvavate samblike puhul uuriti esmalt nende esinemist või puudumist kaskedel ja
haabadel (mändidel esines puidul kasvavaid samblikke kõigil valimi tüvedel). Seda tehti GLZ
binoomse jaotusega analüüsis, kusjuures analüüs viidi läbi kahel puuliigil eraldi ja samblikuliike
ei eristatud. Analüüsi kaasati kirjeldavate pidevate muutujatena tüve surmast möödunud aeg ja
koore katvus. Edasistest analüüsidest jäeti välja kõik tüved, millel samblikke ei esinenud.
Järgnevalt uuriti (GLM, tüüp III) allesjäänud tüvede (N=64) liikide arvu ja puidul kasvavate

22
samblike suhtelise katvuse sõltumist (lisaks puuliigile ja tüve surmast möödunud ajale)
järgmistest kirjeldavatest tunnustest: tüve kaugus lähimast metsast, läbimõõt, valgustatus, koore
katvus ning puidu tihedus (käsitleti analüüsides puuliigist sõltuvana, nested within). Ka nendes
analüüsides uuriti tüve surmast möödunud aja ja puuliigi koosmõju. Kaasatud muutujatest
korreleerusid oluliselt tüve surmast möödunud aeg ja koore katvus (N=64; r=-0,37; p<0,05), kuid
kuna mõlemad on ökoloogiliselt sisukad tunnused ning nende omavaheline seos ei olnud väga
tugev, kaasati analüüsidesse siiski mõlemad.
Lisaks uuriti, kas torikseente või puidul kasvavate samblike liikide arvu tüvedel võiks mõjutada
ka raiesmikku kirjeldavad tunnused. Torikseente puhul kaasati sellise tunnusena vähemalt 14 cm
diameetriga säilikpuudest tekkinud lamapuude maht (m3/ha) uuritaval raiesmikul ja puidul
kasvavate samblike puhul vanade tüügaspuude arv raiesmikul (N/ha). Et maastikuliste tunnuste
vabadusastmeid mitte üle hinnata, viidi nendega läbi eraldi analüüs (ühetunnuseline regressioon).
Selleks arvutati eelkirjeldatud tüvede liikide arvu selgitava mudeli jääkidest iga raiesmiku
suhteline liigirikkus (võttes arvesse konkreetsel raiesmikul uuritud säilikpuude oodatavat liikide
arvu). Suhteline liigirikkus regresseeriti järgnevalt uuritavate raiesmikku kirjeldavate tunnustega.
2.4.2. Ordinatsioon
Torikseente ja samblike liigilise koosseisu analüüsimiseks erineva surmast möödunud ajaga
säilikpuudel kasutati NMS (Non-metric Multidimensional Scaling) ordinatsiooni
programmipaketis PC-ORD 5.10, mis sobib ökoloogilistele andmetele, mis sageli ei ole
normaaljaotusega ega pidevad (McCune & Mefford 1999). Telgede sobiva arvu valimiseks
kasutati Monte-Carlo testi.
Liikide andmemaatriks sisaldas (1) kõikidel lamapuudel registreeritud torikseene- ja (2) kõigil
tüügaspuudel registreeritud samblikuliikide esinemist-puudumist, va need liigid, mida esines
vähem kui kolmel tüvel (torikseente puhul jäi analüüsidesse 31 liiki 62-st; puidul kasvavate
samblike puhul: 18 liiki 44-st). Tüügaspuude torikseenekooslusi ei ordineeritud, sest paljudel
puudel leidus vaid üks seeneliik. Eelnevalt on teada, et lehtpuid ja okaspuid asustavad erinevad
torikseeneliigid (ptk 1.3.2.), seetõttu vaadeldi torikuseenekoosluste analüüsis mändide liigilist
koosseisu kaskedest ja haabadest eraldi. Seletava kategoorilise tunnusena keskkonnamaatriksis

23
kasutati lama- ja tüügaspuude puuliigi ja puu surmast möödunud aja kombinatsioone, kusjuures
ülevaatlikustamiseks grupeeriti vaadeldud puutüved kaheaastastesse „vanusegruppidesse“
(torikseente puhul 11 ja epiksüülsete samblike puhul 6 vanuse ja puuliigi kombinatsiooni).
Seletavate kvalitatiivsete tunnustena kaasati keskkonnamaatriksisse tüve diameeter ja koore
katvus, epiksüülsete samblike analüüsides lisaks ka tüve valgustatus ning pindmise puidukihi
tihedus. „Vanusegruppide“ vahelisi erinevusi võrreldi mitmese reaktsiooni permutatsiooni
protseduuriga (MRPP; multi-response permutation procedure). Joonistel tähistati eraldi
kõdunemise ajaloo poolest erinevad puutüved (ptk 2.1.).

24
3. TULEMUSED
3.1. Torikseened surnud säilikpuudel
3.1.1. Puuliigi ja puu surmast möödunud aja mõju
Torikseente viljakehi leiti kokku 115 lamatüvelt (87% 132-st) ja 67 tüügaspuult (71% 94-st;
Tabel 1). Kõige sagemini olid viljakehadeta männi lamatüved (10 41-st) ja tüügaspuud (17 26-st),
sh ei leitud torikseeni üheltki hiljuti (1–2 a tagasi) surnud männi lama- (N=6) ega tüügaspuult
(N=2). Veel ei leitud torikseeni viielt kase ega kahelt haava lamatüvelt. Tüügaspuudest polnud
lisaks mändidele viljakehasid ka kümnel haaval. Lisaks leidus haabade hulgas kuus tüügast,
millel esines vaid perekonna Phellinus (taelikud) viljakehasid. See perekond asustab aga juba
elusat puud ja põhjustades sageli ka puu surma, ei iseloomusta tüügaspuud kui torikseente
kasvusubstraati, vaid pigem tüüka tekkimise põhjust.
Kokku leiti surnud säilikpuudelt 475 korral torikseeni 60 erinevast liigist (Lisa 2), sh olid 15 neist
looduskaitseliselt huvipakkuvad (Tabel 5). Kõige rohkem torikseeneliike kasvas haaval (38 liiki),
kasel ja männil oli liike vähem (vastavalt 28 ja 16 liiki). Võrreldes lamatüvede liigirikkust
sõltuvalt puu surmast möödunud ajast, pidi arvestama, et puudelt leitud erinevate liikide arv
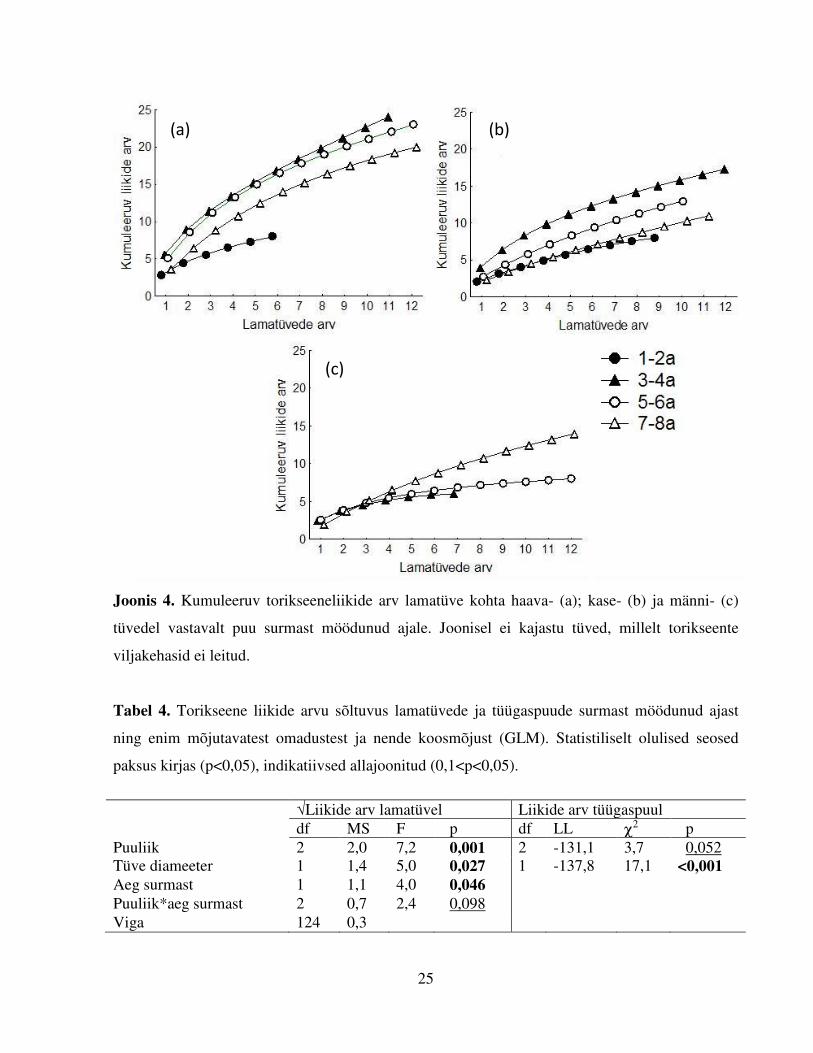
sõltub otseselt ka läbivaadatud tüvede arvust. Liikide kumuleerumise kõverad näitasid, et
haabadel on kõige liigirikkamad 3–6 a tagasi surnud tüved (24 seeneliiki, Joonis 4a) ning
kaskedel 3–4 a tagasi surnud tüved (19 seeneliiki, Joonis 4b), lühemat ja kauemat aega
kõdunenud lehtpuutüvedel oli erinevaid seeneliike vähem. Seevastu mändidest olid kõige
liigirikkamad 7–8 a tagasi surnud tüved (14 seeneliiki, Joonis 4c). Liikide kumuleerumise
kõverad ei saavutanud platood, st iga järgnevalt läbivaadatud tüvega lisandus uusi seeneliike.
Seetõttu võib arvata, et enamate surnud säilikpuude lamatüvede uurimisel oleks leitud ka rohkem
neid asustada suutvaid liike. Tüügaspuudel esines vähem seeneliike kui lamatüvedel (kokku 23
liiki) (Lisa 2).
25
Joonis 4. Kumuleeruv torikseeneliikide arv lamatüve kohta haava- (a); kase- (b) ja männi- (c)
tüvedel vastavalt puu surmast möödunud ajale. Joonisel ei kajastu tüved, millelt torikseente
viljakehasid ei leitud.
Tabel 4. Torikseene liikide arvu sõltuvus lamatüvede ja tüügaspuude surmast möödunud ajast
ning enim mõjutavatest omadustest ja nende koosmõjust (GLM). Statistiliselt olulised seosed
paksus kirjas (p<0,05), indikatiivsed allajoonitud (0,1<p<0,05).
√Liikide arv lamatüvel Liikide arv tüügaspuul df MS F p df LL χ
2 p Puuliik 2 2,0 7,2 0,001 2 -131,1 3,7 0,052 Tüve diameeter 1 1,4 5,0 0,027 1 -137,8 17,1 <0,001 Aeg surmast 1 1,1 4,0 0,046 Puuliik*aeg surmast 2 0,7 2,4 0,098 Viga 124 0,3
(a) (b)
(c)
26
1-2a 3-4a 5-6a 7-8a
Aeg tüve surmast
0
1
2
3
4
5
6
7
8
Liik
ide a
rv tü
vel
Kask
Haab
Mänd
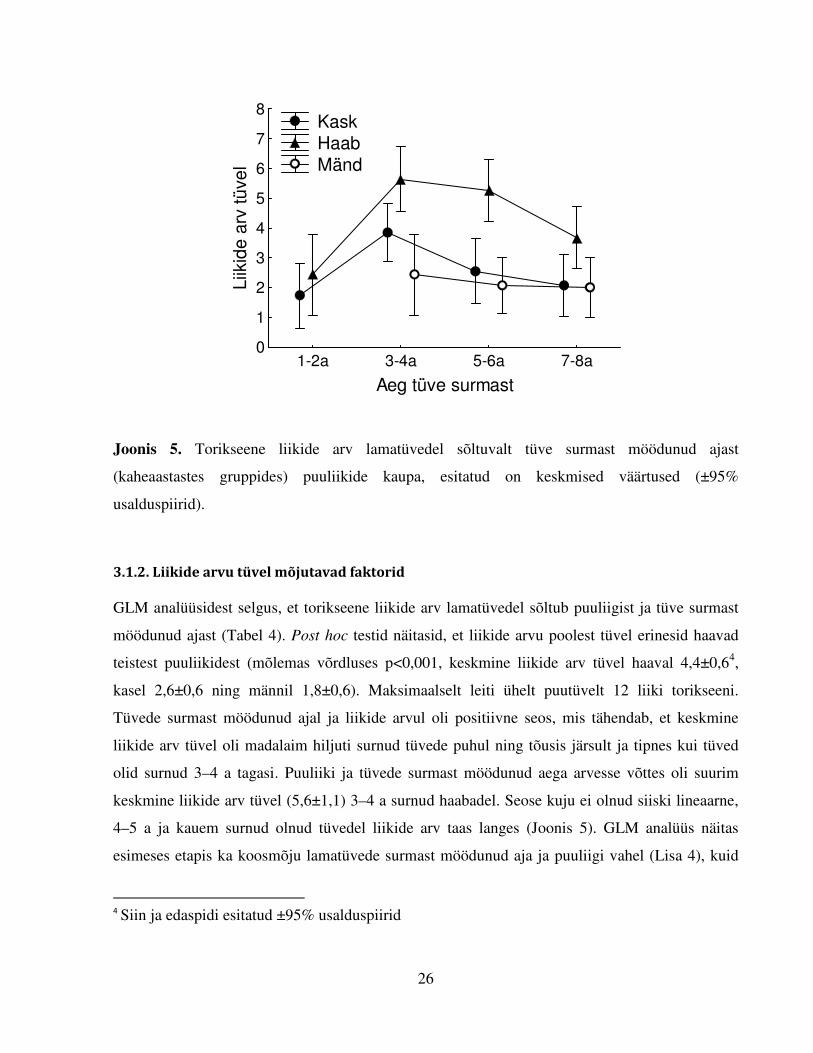
Joonis 5. Torikseene liikide arv lamatüvedel sõltuvalt tüve surmast möödunud ajast
(kaheaastastes gruppides) puuliikide kaupa, esitatud on keskmised väärtused (±95%
usalduspiirid).
3.1.2. Liikide arvu tüvel mõjutavad faktorid
GLM analüüsidest selgus, et torikseene liikide arv lamatüvedel sõltub puuliigist ja tüve surmast
möödunud ajast (Tabel 4). Post hoc testid näitasid, et liikide arvu poolest tüvel erinesid haavad
teistest puuliikidest (mõlemas võrdluses p<0,001, keskmine liikide arv tüvel haaval 4,4±0,64,
kasel 2,6±0,6 ning männil 1,8±0,6). Maksimaalselt leiti ühelt puutüvelt 12 liiki torikseeni.
Tüvede surmast möödunud ajal ja liikide arvul oli positiivne seos, mis tähendab, et keskmine
liikide arv tüvel oli madalaim hiljuti surnud tüvede puhul ning tõusis järsult ja tipnes kui tüved
olid surnud 3–4 a tagasi. Puuliiki ja tüvede surmast möödunud aega arvesse võttes oli suurim
keskmine liikide arv tüvel (5,6±1,1) 3–4 a surnud haabadel. Seose kuju ei olnud siiski lineaarne,
4–5 a ja kauem surnud olnud tüvedel liikide arv taas langes (Joonis 5). GLM analüüs näitas
esimeses etapis ka koosmõju lamatüvede surmast möödunud aja ja puuliigi vahel (Lisa 4), kuid
4 Siin ja edaspidi esitatud ±95% usalduspiirid

27
mudeli lihtsustamisel selle koosmõju olulisus vähenes ning jäi lõplikus mudelis indikatiivse
väärtuse juurde (Tabel 4). Muudest tüve omadustest oli tüve liigirikkusele oluline mõju
läbimõõdul (positiivne seos), mõju ei avaldanud tüve kaugus lähimast metsast, valgustatus ega
kõduaste (Lisa 4).
Ka torikseente arv tüügaspuudel erines puuliigiti (indikatiivne puuliigi mõju, Tabel 4).
Liigivaesemad männid eristusid lehtpuudest (Estimates mõlemas võrdluses p<0,001, keskmine
liikide arv tüvel kasel 1,4±0,3; haaval 1,2±0,2 ning männil 0,4±0,3). Samas ei mõjutanud liikide
arvu tüügaspuudel puu surmast möödunud aeg ega puuliigi ja surmast möödunud aja koosmõju
(Lisa 4 ja 5). GLZ analüüsi esimeses etapis oli tüügaspuude läbimõõdul tüve liikide arvule vaid
indikatiivne mõju (Lisa 4), kuid mudelit lihtsustades selle faktori mõju tugevnes. Uurimisala
kirjeldav tunnus, säilikpuudest tekkinud lamatüvede maht raiesmikul, torikseeneliikide arvu
uuritud lamatüvedel ega tüügaspuudel ei mõjutanud (Lisa 6).
3.1.3. Liigiline koosseis
Surnud säilikpuudelt leitud seeneliikide (N=60) hulka kuulus nii arvukaid kui ka haruldasi5 ja
looduskaitseliselt olulisi liike (Lisa 2; Tabel 5). Tavalisi, arvukaid või sagedasi liike oli 43.
Kaskedel esines kõige sagemini liike Fomes fomentarius (tuletael, 74 leidu), Fomitopsis pinicola
(17 leidu) ja Trametes hirsuta (villtagel, 10 leidu). F. fomentarius oli ülekaalus kõigil
lamatüvedel ja tüügaspuudel, sõltumata nende surmast möödunud ajast. Erineva surma ajaga
puudel levinumate liikide osas võib välja tuua veel, et T. hirsuta oli üks sagedasemaid liike 3–4 a
tagasi surnud kasetüvedel (7 leidu) kuid mitte teistes „vanusegruppides“. Haabadel esines kõige
sagemini liike Trametes ochracea (vööttagel, 37 leidu); Phellinus tremulae (haavataelik, 36
leidu) ja Bjerkandera adusta (harilik suitsik, 26 leidu). Kolm levinumat liiki olid rohkelt
esindatud sõltumata puu surmast möödunud ajast ja olekust. Erineva surmast möödunud ajaga
puudel levinumate seente hulgast võib välja tuua veel liigi Postia alni (sinkjas tümak), mis oli
üks sagedasemaid liike 7–8 a tagasi surnud haabadel (6 leidu), kuid mitte teistel tüvedel.
5 Liigid on jaotatud sagedusklassidesse järgmiselt: väga haruldane (<10 leiukohta Eestis), haruldane (10–20), sage
(20–50), tavaline (50–100), arvukas (üle 100 leiukoha). Kuuluvus sagedusklassi põhineb allikal Parmasto (2004).

28
Mändidelt leiti kõige sagemini liike Trichaptum abietinum (kuusekõbjuk, 26 leidu), Fomitopsis
pinicola (11 leidu) ja Skeletocutis carneogrisea (kõbjuki-peenpoorik, 9 leidu), kusjuures viimane
neist esines enamasti 3–6 a tagasi surnud tüvedel, kuid mitte neil mis olid kauem kõdunenud.
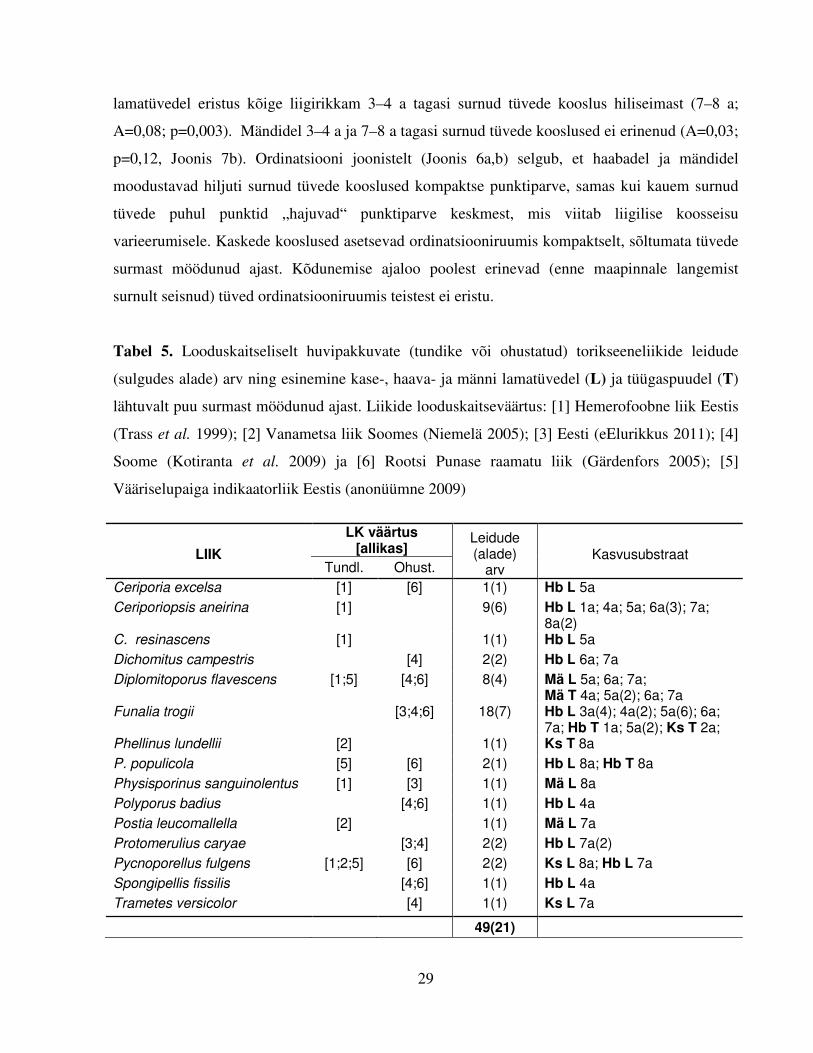
Looduskaitseliselt huvipakkuvaks (tundlikuks või ohustatuks) võib pidada 15 torikseeneliiki
(Tabel 5). Koguni 18 korral (seitsmelt alalt) leiti liiki Funalia trogii (karvane kõbjas), mis kuulub
nii Eestis, Soomes kui Rootsis Punasesse raamatusse. Lisaks leiti haabadelt üheksal korral
hemerofoobseks peetud liiki Ceriporiopsis aneirina (laiapoolne sarvpoorik) ning männitüügastelt
inimpelglikku vääriselupaiga indikaatorit Diplomitoporus flavescens (männikorgik, seitse leidu).
Ülejäänud looduskaitseliselt huvipakkuvate liikide leiud olid ühe- või paarikordsed. Puutüved,
millel esines looduskaitseliselt huvitavaid liike (N=43) ei erinenud teistest läbimõõdu (Mann
Whitney U test, U=3933,5; Z=-1,27; p=0,20), lähima metsa kauguse (U=1443,5; Z=-0,04;
p=0,97) ega valgustatuse poolest (U=3925,0; Z=1,07; p=0,28). Samuti ei erine raiesmikud,
millelt leiti selliseid liike, teistest raiesmikest säilikpuudest tekkinud lamatüvede mahu poolest
(U=179,0; Z=1,26; p=0,20). Küll aga olid säilikpuud, millel looduskaitseväärtusega liike esines,
valimi keskmisest kauem surnud (U=1125,5; Z=2,1; p=0,032) ning seetõttu ka suurema
kõduastmega (U=1008,0; Z=-2,77; p=0,005). Liikide koosesinemismustreid uurides selgus ka, et
haabadel esines looduskaitseliselt väärtuslikke liike suurema tõenäosusega juhul, kui neid
parasiteeris liik Phellinus tremulae (esines selle liigi viljakehi, χ2 test; N=78; χ2=11,55; p<0,001).
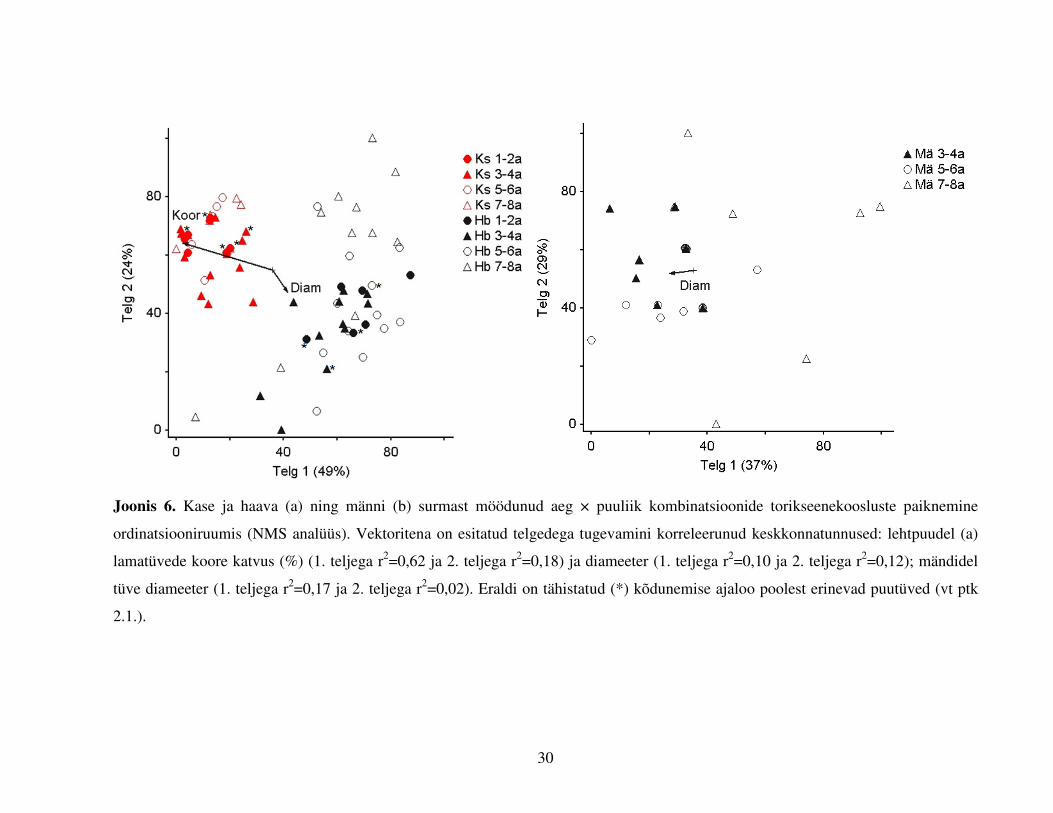
Torikseente liigilise koosseisu kirjeldamiseks lamatüvedel osutus parimaks kahemõõtmeline
lahend, kus lehtpuude puhul (Joonis 6a) seletab esimene telg 49% varieeruvusest, kaks telge koos
aga 73%; männi puhul vastavalt 37% ja 66% (Joonis 6b). Kaskede ja haabade kooslused erinesid
üksteisest kõigi „vanusegruppide“ lõikes (MRPP test, A=-0,01-0,3; p<0,0001 kõigis võrdlustes).
Nende puuliikide lamatüved erinevad üksteisest ka kahe ordinatsiooniruumi vektoritega rohkem
korreleeruva faktori, tüve diameetri ja koore katvuse osas (Studenti T-test t=-4,1; df=88; p<0,001
ja t=14,6; df=88; p<0,001; keskmised diameetrid vastavalt 30±1,7 cm ja 40±3,2 cm ning koore
katvus 93±4% ja 18±9%). Haava lamatüvedel erinesid vaadeldud perioodi lõikes varasemad (1–2
a tagasi surnud tüvede) ja hilisemad (7–8 a tagasi surnud tüvede) seenekooslused (A=0,06;
p=0,008), samuti eristusid varasematest (1–2 a) ja hilisematest (8–9 a) kooslustest (kuid ei
erinenud omavahel) kahe keskmise „vanusegrupi“ kooslused (A=0,03-0,04; p=0,03–0,001). Kase
29
lamatüvedel eristus kõige liigirikkam 3–4 a tagasi surnud tüvede kooslus hiliseimast (7–8 a;
A=0,08; p=0,003). Mändidel 3–4 a ja 7–8 a tagasi surnud tüvede kooslused ei erinenud (A=0,03;
p=0,12, Joonis 7b). Ordinatsiooni joonistelt (Joonis 6a,b) selgub, et haabadel ja mändidel
moodustavad hiljuti surnud tüvede kooslused kompaktse punktiparve, samas kui kauem surnud
tüvede puhul punktid „hajuvad“ punktiparve keskmest, mis viitab liigilise koosseisu
varieerumisele. Kaskede kooslused asetsevad ordinatsiooniruumis kompaktselt, sõltumata tüvede
surmast möödunud ajast. Kõdunemise ajaloo poolest erinevad (enne maapinnale langemist
surnult seisnud) tüved ordinatsiooniruumis teistest ei eristu.
Tabel 5. Looduskaitseliselt huvipakkuvate (tundike või ohustatud) torikseeneliikide leidude
(sulgudes alade) arv ning esinemine kase-, haava- ja männi lamatüvedel (L) ja tüügaspuudel (T)
lähtuvalt puu surmast möödunud ajast. Liikide looduskaitseväärtus: [1] Hemerofoobne liik Eestis
(Trass et al. 1999); [2] Vanametsa liik Soomes (Niemelä 2005); [3] Eesti (eElurikkus 2011); [4]
Soome (Kotiranta et al. 2009) ja [6] Rootsi Punase raamatu liik (Gärdenfors 2005); [5]
Vääriselupaiga indikaatorliik Eestis (anonüümne 2009)
LIIK
LK väärtus [allikas]
Leidude (alade)
arv
Kasvusubstraat
Tundl. Ohust.
Ceriporia excelsa [1] [6] 1(1) Hb L 5a
Ceriporiopsis aneirina [1] 9(6) Hb L 1a; 4a; 5a; 6a(3); 7a; 8a(2)
C. resinascens [1] 1(1) Hb L 5a
Dichomitus campestris [4] 2(2) Hb L 6a; 7a
Diplomitoporus flavescens [1;5] [4;6] 8(4) Mä L 5a; 6a; 7a; Mä T 4a; 5a(2); 6a; 7a
Funalia trogii [3;4;6] 18(7) Hb L 3a(4); 4a(2); 5a(6); 6a; 7a; Hb T 1a; 5a(2); Ks T 2a;
Phellinus lundellii [2] 1(1) Ks T 8a
P. populicola [5] [6] 2(1) Hb L 8a; Hb T 8a
Physisporinus sanguinolentus [1] [3] 1(1) Mä L 8a
Polyporus badius [4;6] 1(1) Hb L 4a
Postia leucomallella [2] 1(1) Mä L 7a
Protomerulius caryae [3;4] 2(2) Hb L 7a(2)
Pycnoporellus fulgens [1;2;5] [6] 2(2) Ks L 8a; Hb L 7a
Spongipellis fissilis [4;6] 1(1) Hb L 4a
Trametes versicolor [4] 1(1) Ks L 7a
49(21)
30
Joonis 6. Kase ja haava (a) ning männi (b) surmast möödunud aeg × puuliik kombinatsioonide torikseenekoosluste paiknemine
ordinatsiooniruumis (NMS analüüs). Vektoritena on esitatud telgedega tugevamini korreleerunud keskkonnatunnused: lehtpuudel (a)
lamatüvede koore katvus (%) (1. teljega r2=0,62 ja 2. teljega r2=0,18) ja diameeter (1. teljega r2=0,10 ja 2. teljega r2=0,12); mändidel
tüve diameeter (1. teljega r2=0,17 ja 2. teljega r2=0,02). Eraldi on tähistatud (*) kõdunemise ajaloo poolest erinevad puutüved (vt ptk
2.1.).
31
3.2. Epiksüülsed samblikud surnud säilikpuudel
3.2.1. Puuliigi ja puu surmast möödunud aja mõju
Tüügaspuude epiksüülsetele samblikele sobiv kasvusubstraat, puidupind, paljandus mändidel ja
haabadel kiiremini ja lausalisemalt kui kaskedel (Joonis 8a). Kokku leiti puidul kasvavaid
samblikke 64 tüügaspuult (68% 94-st), sh kõigil vaadeldud mändidel, sõltumata nende surmast
möödunud ajast (Tabel 2). Valimi tüügaspuudest kattis koor 26-l puul rohkem kui 90% tüve
pinnast, neist enamik (22) olid kased. Seetõttu osutus GLZ analüüsis (Tabel 6) koore katvuse %
epiksüülsete samblike esinemist oluliselt mõjutavaks faktoriks (negatiivne seos) eelkõige
kaskedel. Haabadel seevastu mõjutas puidul kasvavate samblike esinemust eelkõige tüve surmast
möödunud aeg (positiivne seos).
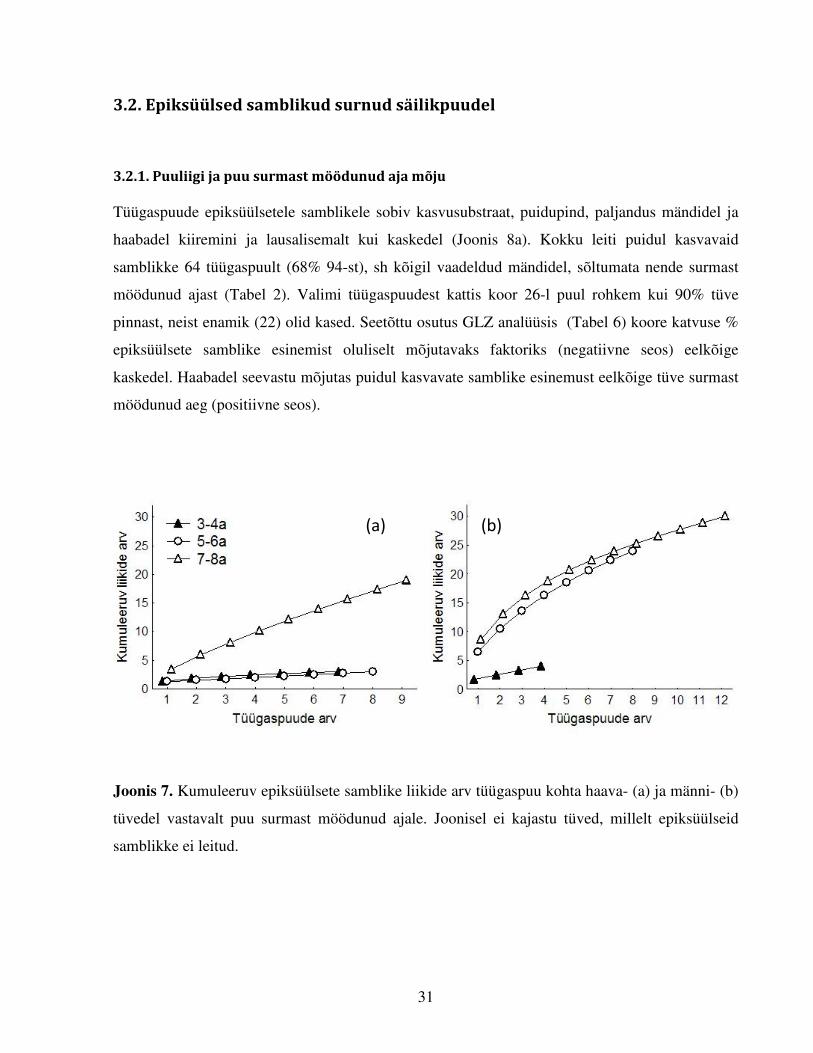
Joonis 7. Kumuleeruv epiksüülsete samblike liikide arv tüügaspuu kohta haava- (a) ja männi- (b)
tüvedel vastavalt puu surmast möödunud ajale. Joonisel ei kajastu tüved, millelt epiksüülseid
samblikke ei leitud.
(a) (b)

32
Puidul kasvavaid samblikuliike leiti kokku 43. Kõige rohkem samblikke kasvas mändide puidul
(42 liiki), haaval ja kasel oli liike vähem (vastavalt 23 ja 14 liiki) (Lisa 3). Erineva surma ajaga
tüvede võrdluses olid kõigil puuliikidel kõige liigirikkamad 7–8 a surnud tüved (Lisa 3), kuid
võttes arvesse liikide kumuleerumise kõverate tõuse (Joonis 7), oleks rohkemate tüvede uurimisel
tõenäoliselt 5–6 a tagasi surnud mändidelt leitud sama palju erinevaid liike kui 7–8 a tagasi
surnud tüvedelt (Joonis 7b). Mitmel kasetüvel paljandunud puidupinda ei leidunud, seetõttu
liikide kumuleerumist kaskedel ei kujutatud. 7–8a tagasi surnud tüved eristusid kõigil puuliikidel
suurema epiksüülsete samblikuliikide arvu poolest hiljem surnutest (29 liiki vs haabadel 17 ja
kaskedel 11 liiki). Liikide kumuleerumise kõverad ei saavutanud platood, st iga järgnevalt
läbivaadatud tüvega lisandus uusi samblikuliike. Seetõttu võib arvata, et kui oleks uuritud
enamaid surnud säilikpuudest tekkinud tüügaspuid, oleks leitud ka rohkem neid asustada suutvaid
liike.
3.2.2. Liikide arvu tüvel mõjutavad faktorid
Ka tüügaspuudel, kus epiksüülsetele samblikele sobivad tingimused olid juba tekkinud (leidus
vähemalt üks samblikuliik), sõltus liikide arv tüvel negatiivselt koore katvusest ja positiivselt
surmast möödunud ajast (Tabel 7, Joonis 8c). Maksimaalselt leiti ühelt tüvelt 16 liiki
epiksüülseid samblikke. Esialgses mitmetunnuselises mudelis (GLM) mõjutas tüvede liigirikkust
oluliselt ka puuliik, kuid mudelit lihtsustades vähenes selle muutuja olulisus indikatiivsele
tasemele, post hoc testides eristus mänd liigirikkuse poolest siiski oluliselt lehtpuudest (mõlemas
võrdluses p<0,001; keskmine liikide arv tüvel mändidel 6,3±1,1; haabadel 2,1±1,2 ning kaskedel
2,8±1,6). Analüüsis osutus oluliseks ka puuliigi ja surmast möödunud aja koosmõju. Teistest
tüügaspuude omadusi kirjeldavatest tunnustest (metsa kaugus, tüve läbimõõt, valgustatus, puidu
tihedus) leitud samblike arv ei sõltunud (Lisa 7). Samuti ei sõltunud samblike arv tüvel vanade
tüügaspuude arvust raiesmikul (Lisa 8).
Puidul kasvavate samblike suhtelise katvuse kirjeldamiseks tüügaspuudel koostati kaks
alternatiivset mudelit. Mudeli koostamise etappe järgides (ptk 2.4.1.) leiti esmalt, et pideva
muutuja varieerumist kirjeldavad kõige paremini puuliik, koore katvus (negatiivne seos) ning
puidu tihedus (oluline negatiivne seos haaval ja männil, kasel oli seos ebaoluline, Tabel 8). Post
33
hoc testides selgus taas, et mänd eristub katvuse poolest oluliselt lehtpuudest (mõlemas võrdluses
p<0,001, keskmine epiksüülsete samblike suhteline katvus männil 0,48±0,07, kasel 0,09±0,10 ja
haaval 0,09±0,08). Kuna aga koore katvus on tüve surmast möödunud ajaga korreleeruv tunnus
(ptk 2.4.1.) ning koore katvus üheskoos puidu tihedusega kirjeldasid olulise osa tüve surmast
möödunud aja varieerumisest (GLM; R2=0,22; F=5,5; p<0,001), katsetati lisaks ka nende kahe
tunnuse asendamist tüve surmast möödunud ajaga, kui antud töö kontekstis keskse tunnusega.
Leiti, et ka alternatiivne, vaid tüve surmast möödunud aega ja puuliiki kaasav mudel, kirjeldab
olulise osa puidul kasvavate samblike katvuse varieeruvusest (Tabel 8, Joonis 8b); post hoc
testide tulemused sarnanesid eelmisele mudelile.
Tabel 6. Samblike esinemuse sõltuvus tüve surmast möödunud ajast ning puidupinna suhtelisest
suurusest puuliigiti. Statistiliselt olulised seosed paksus kirjas (p<0,05).
Puidul kasvavate samblike esinemus Kask (N=33) Haab (N=35)
df LL χ2 p df LL χ
2 p Aeg surmast 1 -17,6 1,4 0,230 1 -17,9 20,9 <0,001 Arcsin √koore katvus 1 -19,3 4,9 0,027 1 -7,65 0,5 0,471
Tabel 7. Puidul kasvavate samblike liikide arvu sõltuvus tüügaspuude surmast möödunud ajast,
koore katvusest, puuliigist ning puuliigi ja surmast möödunud aja koosmõjust (GLM).
Statistiliselt olulised seosed paksus kirjas (p<0,05), indikatiivsed allajoonitud (0,1<p<0,05).
√Puidul kasvavate samblikuliikide arv df MS F p Aeg surmast 1 3,9 16,1 <0,001 Arcsin √koore katvus 1 1,6 0,0 0,012 Puuliik 2 0,7 2,9 0,065 Puuliik*aeg surmast 2 1,1 4,8 0,012 Viga 57 0,2

34
Tabel 8. Puidul kasvavate samblike suhtelise katvuse sõltuvus koore katvusest, puuliigist ja
puidu tihedusest (I) ning puuliigist ja surmast möödunud ajast (II). Statistiliselt olulised seosed
paksus kirjas (p<0,05).
Arcsin √Puidul kasvavate samblike suhteline katvus df MS F p I. R2=0,64; F=20,1; p<0,001 Puuliik 2 0,5 12,9 <0,001 Arcsin √koore katvus 1 0,3 7,0 0,011 Puidu tihedus(Puu liik) 3 0,3 6,4 <0,001 Viga 57 0,0 II. R2=0,54; F=25,8; p<0,001 Puuliik 2 2,0 35,7 <0,001 Aeg surmast 1 0,4 6,6 0,01 Viga 60 0,1
3.2.3. Liigiline koosseis
Säilikpuutüügaste paljandunud puidult leitud samblikuliigid olid enamasti fakultatiivsed
epiksüülid6, puiduspetsialiste oli leitud 43 samblikuliigi hulgas kuus. Liigid jaotusid
sagedusklassidesse7 järgnevalt: väga sagedasi liike oli 27, üsna sagedasi kolm ning sagedasi kaks
(Lisa 3). Kõige sagemini esinesid tüügaspuude puidul lihheniseerumata kalitsioid Mycocalicium
subtile (58 leidu), generalist Hypogymnia physodes (harilik hallsamblik, 30 leidu) ja perekond
Lepraria liigid, mis olid enim esindatud sõltumata puuliigist ja ühed sagedasemad ka kõigis
„vanusegruppides“. Leiti ka kaks haruldast liiki (fakultatiivne epiksüül Bacidina chloroticula,
üks leid ja puiduspetsialist Thelocarpon intermediellum, kaks leidu). Fakultatiivselt epiksüülsete
6 Liikide jaotus fakultatiivseteks ja obligatiivseteks epiksüülideks põhineb allikal Lõhmus & Motiejunaite (2008).
7 Liikide sagedusklasside jaotus põhineb allikal Randlane ja Saag (1999): väga haruldane (1–2 leiukohta Eestis),
haruldane (3–5), üsna haruldane (6–10), üsna sage (11–20), sage (21–50) ja väga sage (üle 50 leiukoha). Liigi praegust kuuluvust sagedusklassi kontrolliti Tartu Ülikooli loodusmuuseumi andmebaasi 31. detsember 2010 seisu ning P. Lõhmuse avaldmata andmete põhjal.
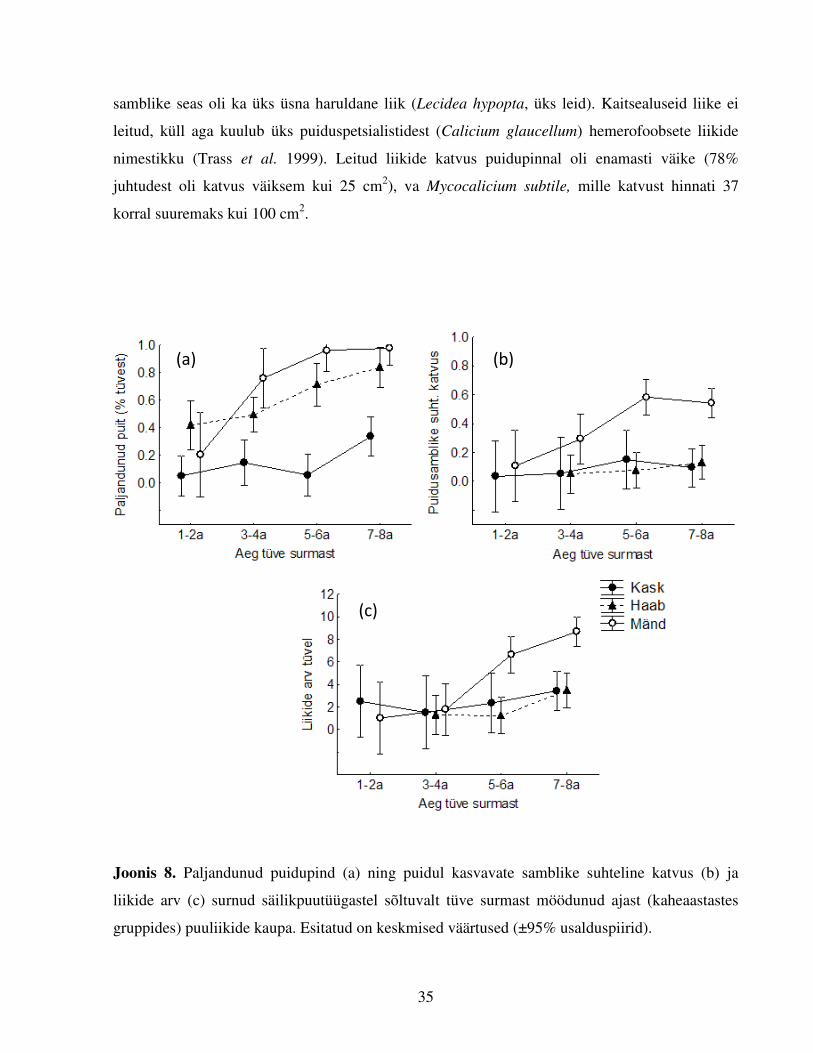
35
samblike seas oli ka üks üsna haruldane liik (Lecidea hypopta, üks leid). Kaitsealuseid liike ei
leitud, küll aga kuulub üks puiduspetsialistidest (Calicium glaucellum) hemerofoobsete liikide
nimestikku (Trass et al. 1999). Leitud liikide katvus puidupinnal oli enamasti väike (78%
juhtudest oli katvus väiksem kui 25 cm2), va Mycocalicium subtile, mille katvust hinnati 37
korral suuremaks kui 100 cm2.
Joonis 8. Paljandunud puidupind (a) ning puidul kasvavate samblike suhteline katvus (b) ja
liikide arv (c) surnud säilikpuutüügastel sõltuvalt tüve surmast möödunud ajast (kaheaastastes
gruppides) puuliikide kaupa. Esitatud on keskmised väärtused (±95% usalduspiirid).
(a) (b)
(c)
36
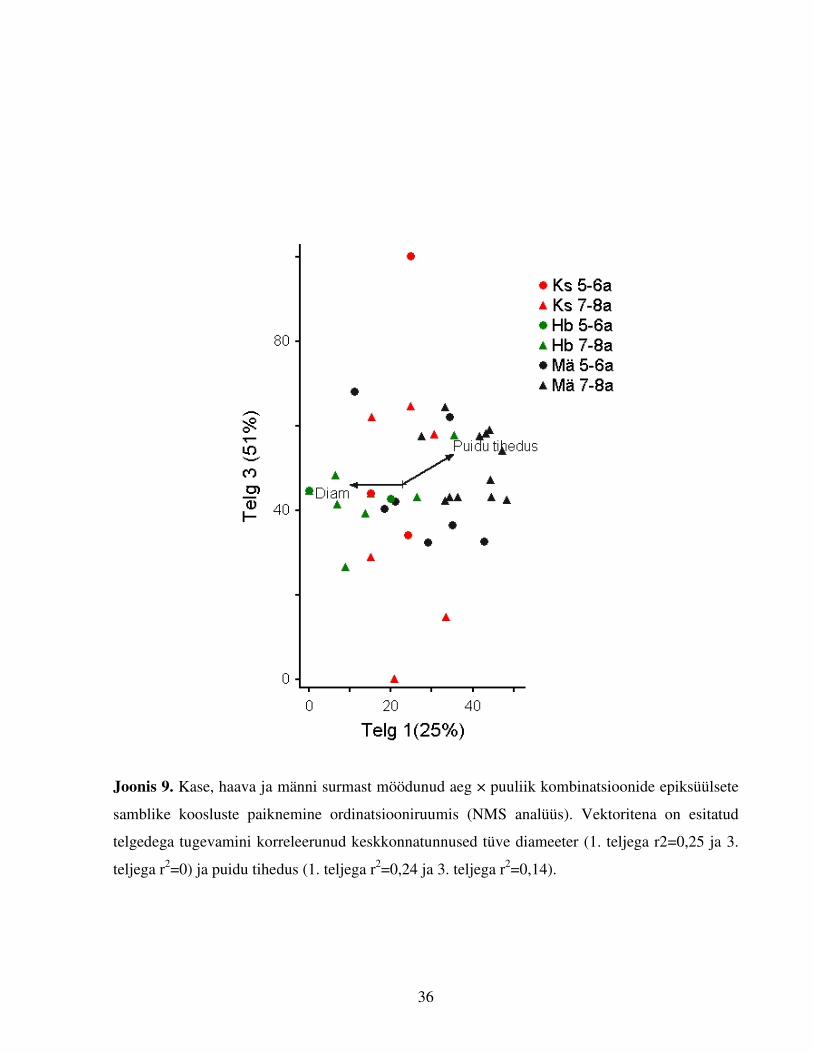
Joonis 9. Kase, haava ja männi surmast möödunud aeg × puuliik kombinatsioonide epiksüülsete
samblike koosluste paiknemine ordinatsiooniruumis (NMS analüüs). Vektoritena on esitatud
telgedega tugevamini korreleerunud keskkonnatunnused tüve diameeter (1. teljega r2=0,25 ja 3.
teljega r2=0) ja puidu tihedus (1. teljega r2=0,24 ja 3. teljega r2=0,14).

37
Puidul kasvavate samblike liigilist koosseisu uuriti vaid 5–8 aastat tagasi surnud puudelt,
rühmitades tüved puuliikide ja kaheaastaste vanusegruppide kaupa (Joonis 9). Lühemat aega
surnud puudel (N=40) enamasti samblikke ei esinenud (58%) või esines vaid üks samblikuliik
(33%). Epiksüülsete samblike liigilise koosseisu kirjeldamiseks tüügaspuudel osutus parimaks
kolmemõõtmeline lahend, kus esimene, teine ja kolmas telg kirjeldasid vastavalt 25%, 7% ja
51%, st kolm telge kokku 83% varieeruvusest. Kaks ordinatsiooniruumi vektoritega rohkem
korreleeruvat faktorit kirjeldavad kase, haava ja männi omavahelisi erinevusi tüve diameetri
(ühefaktoriline ANOVA F2,51=5,1; p=0,009, keskmised vastavalt: 31,6; 41,7 ja 29,3 cm) ning
puidu tiheduse osas (F2,51=6,2; p=0,004, keskmised vastavalt 2,9; 2,5; 4,8 N). Kasetüvede
samblikukooslused paiknevad ordinatsiooniruumis hajusalt (Joonis 9), seetõttu ei avaldu ka
MRPP testides selgeid teiste puuliikidega erinevusi või sarnasusi kirjeldavaid seoseid. Kahe
vaadeldud „vanusegrupi“ tüvede kooslused kaskedel ei erine (A=0,19; p=0,8). Haabade ja
mändide kooslused moodustavad selgesti piiritletavad punktiparved. Puuliikide vahelistes
võrdluses erinevad sama kaua kõdunenud haabade ja mändide (st 5–6 a ja 7–8 a tagasi surnud
tüved puuliikide vahelises võrdluses) samblikukooslused (MRPP test, vastavalt A=0,3; p=0,0003
ja A=0,1; p=0,0003), kuid vanusenihkes (st 7–8 a tagasi surnud haavad ja 5–6 a tagasi surnud
männid) on kooslused sarnased (A=0,03; p=0,1). Puuliigi sisestes võrdlustes on nii haabadel kui
mändidel kahe vaadeldud „vanusegrupi“ kooslused erinevad (vastavalt A=0,11; p=0,018 ja
A=0,05; p=0,016).

38
4. ARUTELU
Käesoleva töö tulemusena näidati, et kaheksa aasta jooksul asustab surnud säilikpuid 60 liiki
torikseeni ja 43 liiki puidul kasvavaid samblikke. Torikseentega olid (viljakehade järgi
otsustades) asustatud enamik vaadeldud kase, haava ja männi lamatüvesid ning tüügaspuid,
teistest puuliikidest eristus haab suurema liigirikkuse, suurema tüve keskmise liikide arvu ja
mitmete looduskaitselist huvi pakkuvate liikide esinemise poolest. Surnud säilikpuude
omadustest mõjutas seeneliikide arvu puul positiivselt tüve läbimõõt. Vaadeldud kaheksa-aastase
arenguetapi lõikes domineerivad liigid seenekooslustes ei vahetunud. Paljandunud puit kui
epiksüülsetele samblikele sobiv kasvupind tekkis säilikpuutüügastel sõltuvalt puuliigist isesuguse
kiirusega, nii polnud paljudel uuritud kaskedel veel puidul kasvavatele samblikele kohta. Haavad
ja männid kaotasid kiiresti suure osa koorest, kuid männipuidu asustasid epiksüülsed samblikud
varem. Männid eristusidki kaskedest ja haabadest suurema puidul kasvavate samblike liigirikkuse
ja seetõttu omanäolisema koosluse poolest. Domineerivad samblikuliigid olid kõigil puuliikidel
samad, nende seas olid enamuses fakultatiivsed epiksüülsed samblikud, puiduspetsialiste leidus
nii varastes kooslustes vähe.
4.1. Torikseened surnud säilikpuudel
4.1.1. Puuliigi ja puu surmast möödunud aja mõju
Kas uuringu säilikpuudest tekkinud lamatüvede (N=132) ja tüügaspuude (N=94) seeneelustikku
võib pidada liigirikkaks? Torikseente osas on võrdlusmaterjali keeruline leida, sest metoodika
poolest sarnaseid uuringuid napib – maastikuinventuurides ei tooda enamasti välja konkreetset
uuritud tüvede arvu. Arvestades mitmete selliste inventuuride ulatust ja leitud liikide arvu, võib
käesoleva uuringu säiliktüvede seenekooslust siiski pidada liigirikkaks. Näiteks Soomes leiti 60
erinevat torikseeneliiki uuringus, mis hõlmas 32 ala ja viis kümnemeetrise raadiusega katseruutu
igal alal (Sippola et al. 2004) ning sama palju seeneliike ka teises, 41 ala ja kümmet

39
kümnemeetrise raadiusega katseruutu hõlmavas seeneinventuuris (Junninen et al. 2006).
Mõlemad uuringud viidi läbi okaspuuenamusega segametsades, kus olid esindatud ka käesolevas
töös uuritud puuliigid, sh paiknes osa viimase uuringu katsealadest vanades loodusmetsades.
Pisut lihtsam on käesoleva töö tulemusi võrrelda kolmanda Soome uuringuga, kus oluliselt
suurema valimi juures (570 lamatüve, peamiselt männid, kuid ka lehtpuud) leiti vaid 37 liiki
torikseeni (Sippola & Renvall 1999). Seetõttu võib säilikpuudest tekkinud kõdupuitu pidada
torikseente jaoks oluliseks kasvupinnaks.
Surnud säilikpuude torikseenekoosluste liigirikkust selgitab osaliselt metsa varane arenguetapp.
Ka varasemates töödes on täheldatud, et puistuvahetushäiringu (ka inimtekkelise, st raie) järgset
aega noores metsas iseloomustab suur torikseente liigirikkus (Lõhmus 2011a; b, Junninen et al.
2006, Berglund et al. 2011a). Loodusliku häiringu järgselt on see tingitud kõdupuidu rohkusest –
sellisel puhul jääb metsa suures hulgas erineva mahuga surnud puitu. Raiejärgses metsas tagavad
torikseente liigirikkuse eelkõige raiejäätmed (peen kõdupuit e puidufraktsioonid, mille läbimõõt
ei ületa 9,9 cm-t) ja kännud, mis samuti sobivad elupaigaks mitmele pioneerliigile (Junninen et
al. 2006). Kui võrrelda liikide arvu samas mahuühikus peenel ja jämedal kõdupuidul, on esimene
neist liigirikkamgi (Nordèn et al. 2004, Heilmann-Clausen & Christensen 2004). Lisaks
raiejäätmetele ja kändudele jääb sageli raiejärgselt metsa ka eelmise metsapõlvkonna hilis-
suktsessiooniliste puuliikide (nt kuuse) lamatüvesid (Sippola & Renvall 1999). Kõik need seente
kasvusubstraadid tingivad häiringujärgse metsa suure levisepanga, mis omakorda aitab kaasa
kiirele uute tekkivate kasvupindade (nt mõne aasta jooksul peale raiet surevate säilikpuude)
koloniseerimisele. Arvestades samas sellega, et peen kõdupuit ja kännud on torikseente
kasvusubstraadina lühikese kestvusega (Sippola & Renvall 1999, Lõhmus 2011b) ja kõigile
torikseeneliikidele elupaigaks ei sobi (Berglund et al. 2011b), on surnud säilikpuutüvede vajadus
raiesmikul ilmselge.
Antud uuringus erinesid vaadeldud puuliigid torikseente liigirikkuse osas. Haavad erinesid
kaskedest ja mändidest nii suurema torikseeneliikide ja looduskaitselist väärtust omavate liikide
hulga kui keskmise liikide arvu poolest tüvel (Joonis 4; 5). Surnud haabade liigirikast
seeneelustikku majandusmetsas on näidatud ka varasemates töödes (Junninen et al. 2007,
Lõhmus 2011a). See on selgitatav haava ökoloogilise nišiga metsaökosüsteemis. Haab on

40
hemiboreaalsetes ja boreaalsetes metsades häiringujärgne pioneerpuuliik (Linder et al. 1997).
Seetõttu võivad haaval elavad liigid olla kohanenud häiringujärgse avatud elupaiga
mikroklimaatilistele tingimustele, mida on lisaks torikseentele (Junninen et al. 2007), täheldatud
ka puidumardikate (Martikainen et al. 2001, ja epifüütsete samblike puhul (Hazell & Gustafsson
1999).
Kase- ja männitüvede väiksema liigirikkuse tingivad tõenäoliselt siiski pigem nende puuliikide
eripärad torikseente kasvusubstraadina kui raiesmike ebasobiv mikrokliima. Nimelt on teada, et
häiringust taastuva metsa mikroklimaatiliste oludega tulevad lisaks haavale toime ka peamiselt
teistel puuliikidel elavad torikseened (Junninen et al. 2006, Berglund et al. 2011b). Seetõttu võib
pigem arvata, et: (1) väiksem liikide arv kasel on tingitud sellest, et Eestis ongi vähem sellel
puuliigil elavaid seeni (nt haavalt on Eestis leitud 78, Eesti kõige levinumalt lehtpuult kaselt aga
68 torikseeneliiki, Parmasto 2004); teisest küljest on vähene liikide vaheldumine kõdunemise
käigus kaskede seenekooslustele iseloomulik (vt arutlust ptk 4.1.4.); (2) väiksemat liikide arvu
männil selgitab selle puuliigi keemilistest ja füüsikalistest omadustest tulenev aeglasem
kõdunemine (Harmon 1986, Sippola et al. 1998). Seetõttu polnud valimi männid tõenäoliselt veel
mitmetele seentele sobivat lagustaadiumi saavutanud. See selgitab ka tüve surmast möödunud aja
ja puuliigi koosmõju tüve liikide arvu kirjeldavas mudelis – männi lamatüvedel ei leidunud
erinevalt teistest puuliikidest torikseeni enne kui tüve surmast oli möödunud kolm aastat.
Käesolevas töös kirjeldatud liigirikkuse dünaamika kõdunemise vältel erineb eelnevatest
uuringutest. Varasemad tööd on näidanud, et seente liigirikkus kasvab kõdunemise käigus, tipneb
keskmise kõdunemuse juures („noameetodil“ hinnatud 3. kõduaste; Renvall 1995, Kruys et al.
1999) ja langeb alles puidu edasise lagunemise ja ammendumise käigus. Käesolevas töös aga
tuvastati, et lehtpuu lamatüvedel oli torikseente liigirikkus ja tüvelt leitud liikide arv kõige
suurem 3–4 aastat (haabadel 3–6 aastat) peale surma ning langes seejärel (Joonis 4; 5). Samas
saavutasid käesoleva töö lamatüved „soodsaima“ 3. kõduastme enamasti alles 7.–8. surmajärgsel
aastal – siis oli liigirikkus võrreldes varasemaga juba langenud. Seega ei ole langus liigirikkuses
tingitud varasemates töödes kirjeldatud seentele soodsamate kõdustaadiumite ületamisest.
Tõenäoliselt võib erinevusi käesoleva ja varasemate tööde tulemuste vahel selgitada
võrdlusandmete ebatäpsusega. Varane liigirikkuse tipp ilmnes ka Rootsi uuringus (Lindhe et al.

41
2004, vt ka ptk 1.4.), seal põhjendati erinevust eelnevatest töödest halvasti võrreldavate
andmetega. Nimelt vaadeldi ka selles töös liigirikkuse muutumist kõdutüvedel üheaastase
täpsusega, seevastu enamasti on torikseeni käsitlevates uuringutes ainsaks infoks tüvede
kõdunemisaja kohta kõduaste (e.g. Junninen et al. 2007, Heilmann-Clausen & Christensen 2005).
„Noameetodil“ hinnatud kõduaste annab puidu lagunemuse kohta ebatäpse hinnangu (Kahl et al.
2009, Larjavaara & Muller-Landau 2010): nt võib üleminek teisest kõduastmest kolmandasse
lehtpuude puhul kesta üle viie ja okaspuude puhul üle kümne aasta (Sholohov & Sholohova
2001). Seetõttu on võimalik, et selliselt kõdupuidu vanust hinnates on varane liigirikkuse tipp
neis uuringutes lihtsalt märkamata jäänud. Keskmist kõdunemust on kõige liigirikkamaks peetud
seetõttu, et selles faasis leidub kooslustes veel pioneerliike, aga juba ka hilisemaid lagundajaliike
(Renvall 1995, Boddy 2001). Käesoleva töö tüvedel tuvastati sellise üleminekukoosluse
tunnuseid alles 7–8 aastat tagasi surnud haabadel ja mändidel (vt arutlust ptk 4.1.4.), mistõttu
võib oletada, et üleminekukooslusega kaasnev liigirikkuse tipu saavutamiseks peavad surnud
säilikpuud kõdunema kauem kui käesolevas uuringu periood.
4.1.2. Liikide arvu tüvel mõjutavad faktorid
Metsade looduslähedase majandamise seisukohast rakenduslikult olulise tulemusena leiti antud
uurimuses, et suurema läbimõõduga säilikpuutüvedel on rohkem torikseeneliike (Tabel 4).
Sarnasele järeldusele on jõutud ka mitmes varasemas uurimuses (e.g. Renvall 1995, Lindhe et al.
2004), seetõttu oli see tulemus ootuspärane. Samas tuleks surnud säilikpuude puhul tüve suure
diameetri olulisust seeneelustikule eriti esile tõsta raiesmiku heitlike mikroklimaatiliste
tingimuste tõttu. Lamatüve kontakt maapinnaga aitab vähendada ümbritsevate valgus- ja
niiskustingimuste mõju seentele (Heilmann-Clausen & Christensen 2003). See puhverdav mõju
on raiesmiku karmistunud tingimustes tõenäoliselt olulisem kui metsapuude varjus ning seda
suurem, mida mahukam on puutüvi. Teiseks kõduneb suur tüvi aeglaselt ja kestab elustikule
kauem (Renvall 1995). Kui küpses metsas (isegi üheealises majandusmetsas) tekib pidevalt
kõdupuitu juurde, siis raiejärgses puittaimede taastumise faasis on säilikpuud aastakümneteks
ainsaks noore metsa suuremõõdulise kõdupuidu allikaks (Vanha-Majamaa & Jalonen 2001). Seda
arvestades on säilikpuust tekkinud kõdutüvede kestmine ja järelikult ka mõõtmed seeneelustikule
eriti olulised.

42
Käesolevas töös ei tuvastatud, et surnud säilikpuudest tekkinud lamatüvede maht raiesmikul
oleks mõjutanud liikide arvu uuritud kõdutüvedel (Lisa 6). Samas varasemates töödes on
kõdupuidu mahtu maastikul peetud üheks olulisemaks liigirikkust ennustavaks faktoriks (e.g.
Penttilä et al. 2004, Rolstad et al. 2004, Hottola et al. 2009). Seose puudumist saab seletada
sellega, et uuritud kõdutüved asusid aladel, kus toimus raie 8–9 aastat tagasi. Tõenäoliselt ei
olnud nii lühikese ajaga veel raie tulemusena tekkinud (raiejäätmed ja kännud); ega eelmise
metsapõlvkonna pärandina alles jäänud kõdupuit torikseente kasvusubstraadina ammendunud.
Uuringud näitavad, et lageraiest taastuvates metsades ammenduvad need kasvusubstraadid
torikseentele umbes 20 aastat peale raiet (Sippola & Renvall 1999, Lõhmus 2011b). Seetõttu võib
arvata, et muu päritoluga kõdupuidu olemasolu raiesmikel varjutas surnud säilikpuude mahu
efekti vaadeldud puutüvede liigirikkusele. Nende teadmiste põhjal võib järeldada, et surnud
säilikpuud muutuvad torikseentele oluliseks eelkõige muu kõdupuidu ammendudes. Samas
mitmed torikseeneliigid kände ja raiejäätmeid ei asusta (Berglund et al. 2011b), sellistele liikidele
on surnud säilikpuutüvede olemasolu oluline juba varem.
4.1.3. Liigiline koosseis
Häiringujärgse metsa varase arenguetapi seenekooslused erinevad liigilise kooseisu poolest
oluliselt kõigist hilisematest. Varaseid seenekooslusi iseloomustab peenikest kõdupuitu asustada
suutvate, valgemädanikku tekitavate seeneliikide domineerimine (Sippola & Renvall 1999),
siiski võivad täiesti puududa mitmed hilisemates kooslustes tavalised seeneperekonnad (Lõhmus
2011b). Samas leidub varastes kooslustes tõenäoliselt ka just neile kohastunud liike (Lõhmus
2011b).
Käesolevas töös uuritud säilikpuudest tekkinud lamatüvedel ja tüügaspuudel enam esinevad
torikseeneliigid on ka teiste tööde varastes seenekooslustes tavalised (e.g. Sippola & Renvall
1999, Lindhe et al. 2004, Junninen et al. 2006). Kuus sagedasemat liiki – kaskedel Fomes
fomentarius ja Trametes hirsuta; haabadel Trametes ochracea ja Bjerkandera adusta ning
mändidel Trichaptum abietinum ja Skeletocutis carneogrisea – on Eesti metsades arvukalt
levinud (Parmasto 2004), suudavad asustada erinevaid kasvusubstraate (Parmasto 2004, Niemelä
2008, Lõhmus 2011b), omavad seetõttu ilmselt Eesti maastikus laialdast levisepanka ning sellest
tulenevalt konkurentsieelist võrreldes mitmete vähem levinud liikidega (Berglund 2011b). Lisaks

43
ülalnimetatutele leidus nii kase- kui männitüvedel tihti liigi Fomitopsis pinicola viljakehasid. F.
pinicola on mitme teise seene jaoks oluline puitu ettevalmistav seen (Niemelä et al. 1995) ning
on Eestis levinud kõigis metsatüüpides (Parmasto 2004). Põhjamaades on aga leitud, et see liik
sõltub jämedatest puutüvedest (Renvall 1995, Kruys et al. 1999, Lindhe et al. 2004) ning võib
nende puudumisel piirkonniti kaduda (Olsson et al. 2011). Seetõttu võivad surnud säilikpuutüved
sellele liigile olla eriti oluline kasvupind raiejärgses metsas.
Torikseente varast liigilist koosseisu mõjutab nii substraadi (Olsson et al. 2011) kui ala tasemel
(Junninen et al. 2006) ümbritseva piirkonna majandatuse aste, millest sõltub kõdupuidu hulk
maastikus ja kohalik liigibaas. Käesoleva töö lamatüvede kooslused esimestel surmast möödunud
aastatel asetsesid ordinatsiooniruumis kompaktselt, see viitab uuringualade taustsüsteemi
suhtelisele homogeensusele. Varasemates töödes on näidatud, et arengu käigus muutub
torikseente koosluste struktuur üha raskemini ennustatavaks (Renvall 1995, Høiland & Bendiksen
1996, Heilmann-Clausen 2001), liigilist koosseisu mõjutavaid asjaolusid on mitmeid (vt arutlust
ptk 1.3.3. ja 1.3.4.). Nende teadmistega on kooskõlas käesoleva töö koosluste „hajumine“
ordinatsiooniruumis tüvede surmast möödunud aja kasvades (Joonis 6). Koosluste muutumine
kaheksa surmajärgse aasta jooksul oli kõige selgem kiirekasvulisel ja pehme puiduga (Tamm
2000) haaval, mille 7–8 a tagasi surnud tüved erinesid koosluste poolest MRPP testis
varasematest, hõlmates veel ka esimesi pioneerliike, kuid juba ka hilisemat koosluse arengut
iseloomustavaid seeni (nt Ganoderma applanatum, jänesvaabik (Lindhe et al. 2004)). Üsna
sarnane koosluste muutumise dünaamika oli märgata ka männi puhul – kuid ehkki ka 7–8 a tagasi
surnud männitüvedel esines hilisematele koosluste arenguetappidele iseloomulikke liike (nt
pruunmädanikku tekitavad liigid perekondadest Oligoporus ja Postia pole enamasti esimesed
lagundajad, Olsson et al. 2011), MRPP test erinevust 7–8 a tagasi ja hiljem surnud tüvede vahel
ei tuvastanud – see viitab männipuidu omadustest tulenevale aeglasemale kõdunemisele (Harmon
1986, Sippola et al. 1998, Yatskov et al. 2003) ja on ootuspärane.
Erinevalt haabadest ja mändidest olid kaskede seenekooslused sõltumata puu surmast möödunud
ajast üsna sarnased, seda kinnitab ka ordinatsioonanalüüs (Joonis 6). Teistsuguse seenekoosluste
arengu kase lamatüvedel tingib asjaolu, et need võivad kõdunemise lõppfaasini olla koorega
kaetud (Krankina et al. 1999). Lehtpuudel domineerivad kõdunemise alguses sageli tüves juba
enne puu surma soikeseisundis kohal olnud seened (Boddy 2001), nt Fomes fomentarius (Baum

44
et al. 2003). Puukoor tagab selliste liikide konkurentsieelise ka edasises koosluse arengus, seda
kahel moel: (1) luues tüves stabiilsema mikrokliima, mis annab eelise väljast tulevate, tüüpiliselt
stressitolerantsete, liikide ees ning (2) takistab väljast tulevate liikide ligipääsu (Heilman-Clausen
2001). Seetõttu võib kooslus muutumatuna püsida kuni tüve hiliste lagustaadiumiteni (Lange
1992, Heilmann-Clausen 2001), nagu näitab ka käesolev töö.
Tasub siiski silmas pidada, et käesolevas töös tuvastati liikide esinemist surnud säilikpuudel vaid
viljakehade põhjal, mis ei peegelda täpselt seente tüvesisest arengut. Erineva elustrateegiaga
(sensu Boddy & Heilmann-Clausen 2008) torikseened moodustavad viljakehasid erineva
kiirusega, mistõttu võivad hilisemad viljakeha moodustajad tüve asustada varemgi kui varasemad
(Stenlid et al. 2008). Viljakeha ei iseloomusta ka tingimata mütseeli suurt kohasust
kasvusubstraadi sees, vaid võib viidata hoopis seene „põgenemiskatsele“ ebasobivate biootiliste
või abiootiliste keskkonnatingimuste eest (Boddy 2001). Neil põhjustel tasuks viljakehade põhjal
järelduste tegemisel jääda ettevaatlikuks.
4.1.4. Looduskaitseliselt huvipakkuvad liigid
Surnud säilikpuudel kasvas mitu Eestis, Soomes ja Rootsis kaitsealust või hemerofoobset
torikseeneliiki (Tabel 5). Enamasti leidus sellised liike haaval, kuid mõned ka männil ja kasel.
Säilikhaabade puhul leiti ka Soomes, et need võivad looduskaitseväärtust omavatele liikidele
oluliseks kasvupinnaks olla – kuid seda peeti sealset uuringuala ümbritseva piirkonna mõjuks
(Junninen et al. 2007). Käesolevas töös uuritud tüved asuvad aga 48 erineval alal, (sh haavad 18
raiesmikul) ja on seetõttu vähem mõjutatud konkreetsest taustametsast ning ilmestavad sellest
tulenevalt säilikpuude väärtust torikseente kasvusubstraadina paremini. Kaks ilmselgelt
säilikpuudest kasu saavat, peamiselt haavapuitu kasutavat kaitsealust torikseeneliiki (Funalia
trogii ja Ceriporiopsis aneirina, vt ka Junninen et al. 2007 ja Lõhmus 2011b) esinesid siiski väga
agregeeritult (mitu leidu sama raiesmiku säilikpuudelt). Sellest võib järeldada, et säilikpuude
edukus looduskaitseliselt väärtuslike liikide kasvupinnana sõltub ümbritseva maastiku
omadustest, mida käesolevas töös ei uuritud: liikide esinemisest, fragmenteeritusest, kõdupuidu
hulgast või lättepopulatsiooni suurusest.

45
Varasemates uuringutes on leitud, et erinevalt enamikust teistest liikidest, eelistavad kaitsealused
liigid üldiselt hilisemas kõdunemise faasis puitu (Renvall 1995, Heilmann-Clausen &
Christensen 2005). Käesolevas töös uuritud tüved üldiselt keskmist kõdunemust ei ületanud, kuid
sarnane trend esines siingi: looduskaitseliselt väärtuslikud liigid esinesid pigem valimi rohkem
kõdunenud puudel (surma ajast möödas 5 aastat ja enam). Ilmselt viitab hilisemate kõduastmete
eelistusele ka see, et väärtuslikke haaval kasvavaid seeneliike oli tõenäolisem leida liigi Phellinus
tremulae parasiteeritud tüvedelt. Parasiitseene tingitud lagundamisprotsesside tõttu on sellised
puud sageli juba surres seest õõnsad (Niemelä 2008), kõdunevad maapinnale langedes kiiremini
(Tamm 2000) ning muutuvad seetõttu tervetest tüvedest varem hilisemaid laguastmeid
eelistavatele liikidele sobivaks kasvukohaks. Samas ammenduvad sellised tüved kiire
kõdunemise tõttu torikseente jaoks varem kui parasiteerimata puud. Seetõttu oleks soodsas
kõdustaadiumis haavapuidu kättesaadavus seentele metsa taastumise faasis kõige ühtlasem, kui
säilikpuudeks valitaks sama ala piires nii liigi P. tremulae parasiteeritud kui terveid haavapuid.
4.2. Epiksüülsed samblikud surnud säilikpuudel
4.2.1. Puuliigi ja puu surmast möödunud aja mõju
Selleks, et surnud säilikpuudest tekkinud puutüügastel saaksid arenema hakata epiksüülsete
samblike kooslused, peab tüügaspuudel leiduma sellistele samblikele sobivat kasvupinda.
Paljandunud puidupind tekkis uuritud puuliikidest haabadel ja mändidel peatselt peale surma
(Joonis 8a). Kasetüvedest seevastu olid mitmed koorega kaetud ka surmast kaheksa aasta
möödudes. Varasemad uuringud ongi näidanud, et kaskedel pole koore eemaldumise ja puu
surmast möödunud aja vahel seost (Yatskov et al. 2003), seetõttu ei sobi käesoleva uuringu
aegrealine disain ka kaskede epiksüülsete samblikukoosluste tekke kirjeldamiseks. Siiski ei saa
säilikpuudest tekkinud kasetüügaste olulisust puidul kasvavatele samblikele alahinnata: uuringud
on näidanud, et maastikus harvadel kooreta kasetüügastel on liigirikas epiksüülsete samblike
elustik, sh on mõnda samblikuliiki leitud vaid kaskede paljandunud puidult (Lõhmus & Lõhmus
2001).

46
Varaseid epiksüülsete samblike kooslusi kirjeldades tuleb arvestada samblike kolonisatsiooniks ja
kasvuks kuluva ajaga. Samblikud on võrreldes torikseente ja soontaimedega tuntud kui aeglase
kolonisatsiooniga organismid (Chlebicki et al. 1996, Nash 1996). Asustamise järgselt määrab
nende tuvastamise puidult kasvuks kuluv aeg – ja ehkki erinevate liikide kasvukiirus võib olla
isesugune, on samblikud üldiselt ka aeglase kasvuga organismid (Nash 1996). Nii ei leitud
Rootsis kuusekändude lõikepindadelt puidul kasvavaid samblikke enne, kui raiest oli möödas viis
aastat (Caruso et al. 2010). Seega on ootuspärane, et käesolevas töös ei tuvastatud osal hiljuti
paljandunud puiduga puutüvedest veel ühtegi samblikuliiki. Ka käesoleva uuringu tüügaspuudelt
leitud samblikuliikide arv jääb alla varasemates uuringutes leitule (e.g. Forsslund & Koffman
1998, Lõhmus & Lõhmus 2001), kuid nendes uuriti kauem kõdunenud tüügaspuid. Seega on
väiksem liikide arv käesoleva uuringu tüügaste tekkest möödunud nappi aega arvestades
loomulik. Ka uuritud tüügaste valim oli väike, selle suurendamine oleks tõenäoliselt mitmeid
liike lisanud (liikide kumuleerumise kõverad ei saavutanud platood). Leitud liikide arv oli
tõenäoliselt ka mõnevõrra suurem, sest mitmed proovid jäid materjali vähesuse või
väljakujunemata tunnuste tõttu liigini määramata (ptk 2.3.).
Käesoleva uuringu tulemusena leiti, et varases koosluste tekkefaasis iseloomustab männitüükaid
haabadest suurem puidul kasvavate samblikuliikide arv tüvel ja samblike katvus (Joonis 8b; c);
ning männipuidu asustamiseks kulub samblikel ka vähem aega (puu surmast möödunud aja ja
puuliigi koosmõju kõigis samblikke puudutavates analüüsides). Seevastu üldiselt ei arvata, et
puuliik oleks epiksüülsete samblike liigirikkuse koha pealt määrav. Enamasti ei peeta puidul
kasvavate samblike uuringutes vajalikuks puuliiki eristadagi (Cieśliński et al. 1996, Chleblicki et
al. 1996, Forsslund & Koffman 1998). Varasemas Eesti tüükauuringus osutasid väikesed
erinevused liigirikkuses pigem lehtpuupuidu soodsale mõjule (Lõhmus & Lõhmus 2001), kuid
see töö ei käsitlenud haabasid. Eesti epiksüülsete ja epifüütsete samblike liigirikkust analüüsivas
uuringus (Jüriado et al. 2003), tuvastati küll püstisurnud mändidel haabadega võrreldes suurem
epiksüülsete samblike liigirikkus, kuid selle võis tingida uuritud männitüügaste suurem vanus
ning esindatus maastikus. Seega pole taoline erinevus mändide ja haabade vahel varem kinnitust
leidnud. Haabade aeglasem koloniseerimine võib olla tingitud koore ebatäielikust eemaldumisest.
Nimelt võib vastpaljandunud haavapuitu esimestel aastatel katta korba niinekiudude mass (isiklik
tähelepanek), mis kauem kõdunenud tüvedel puudub – seetõttu võib haaval, erinevalt männist,

47
koore eemaldumise ja võimaliku epiksüülsete samblikega asustamise vahel olla ajanihe.
Erinevale koloniseerimiskiirusele võivad kaasa aidata ka puidu füüsikalised ja keemilised
omadused, kuid see vajaks edasist uurimist.
4.2.2. Liikide arvu tüvel mõjutavad faktorid
Puu surmast möödunud aeg ja puuliik olid olulisimad epiksüülsete samblikuliikide arvu ja
katvuse varieerumist kirjeldavad faktorid (Tabel 6; 7). Puidupinna omadustest peetakse samblike
liigirikkuse ja liigilise koosseisu koha pealt üldiselt oluliseks ka puidu kõdunemust (e.g.
Söderström 1988, Forsslund & Koffmann 1998; Botting & DeLong 2009). Samas ei ole uuritud,
kas võrreldes vähem kõdunenud puiduga tingib suurema liikide arvu keskmise kõdunemuse
juures kolonisatsiooniks ja kasvuks kuluv aeg või vastav nõudlus kasvusubstraadi osas.
Käesolevas töös oli võimalik neid kahte muutujat eristada, kuna lisaks teadaolevale puu surma
ajale fikseeriti täpselt puidu lagunemus (puidupinna tiheduse muutumise kaudu). Kui
analüüsidesse kaasati mõlemad, omas olulist mõju vaid puu surmast möödunud aeg.
Liikide arvu ja suhtelist katvust surnud säilikpuudel mõjutas tüve omadust ka puidupinna osakaal
(1 – koore katvus, Tabel 6; 7). See tulemus ei ole tüügaspuude eripära, suure pindala positiivset
mõju on rõhutatud ka teistes epiksüülsete samblike kasvupindade liigirikkust käsitlevates töödes
(e.g. Humphrey et al. 2002, Botting & DeLong 2009, Lõhmus et al. 2010) – kui puidupind on
suurem, on ka suurem tõenäosus selle asustamiseks erinevate liikide poolt. Käesolevas töös
vaadeldi hiljuti surnud puutüvesid, kus puidupinna suuruse määras pigem koore eemaldumine kui
teine ökoloogiliselt sarnase sisuga tüve tunnus: diameeter. See on tõenäoliselt põhjuseks, miks
see varasemates puidul kasvavaid samblikke käsitlenud töödes (e.g. Söderström 1988, Forsslund
& Koffmann 1998) oluliseks osutunud tüve omadus surnud säilikpuude puhul mõju ei omanud.
4.2.3. Liigiline koosseis
Kooreta tüügaspuude samblikukooslused muudab võrreldes teiste kõdupuidu tüüpidega
eripäraseks suhteliselt suur jalgsamblike e kalitsioidide hulk (Lõhmus & Lõhmus 2001,
Nascimbene et al. 2008). Kalitsioidsed liigid on halvad konkureerijad, mistõttu potentsiaalselt
substraadilt – lamapuidult – tõrjutakse need sammalde ja porosamblike poolt välja (Holien 1996).

48
Eriti oluliseks elupaigaks on tüügaspuud kalitsioididest mittelihheniseerunud saprotroofsetele
seentele perekondadest Chaenothecopsis, Mycocalicium ja Microcalicium, mis on
lihheniseerunud liikidest kasvusubstraadi suhtes suurema tundlikkusega (Rikkinen 1995).
Käesoleva töö surnud säilikpuudelt leiti ainult neli kalitsioidset liiki (Calicium glaucellum,
Chaenothecopsis pusilla, C. savonica ja Mycocalicium subtile), mida on võrreldes varasemate
uuringutega vähe (Holien 1996, Forsslund & Koffmann 1998, Lõhmus & Lõhmus 2001). Kõik
mainitud liigid on Eestis väga sagedased, mistõttu leidub metsamaastikul tõenäoliselt rohkelt
nende leviseid ning seeläbi on tagatud nende efektiivsem koloniseerimine haruldasemate
kalitsioidsete liikidega võrreldes. Viimase omaduse poolest paistab käesoleva töö põhjal eriti
silma Mycocalicium subtile, mis esines enamikul käesolevas töös uuritud puutüvedest. See liik
ongi tuntud valgusrohkete kasvukohtade eelistamise poolest (Lõhmus 2004).
Uuritud tüügaspuude puidul kasvavate samblike liigiline koosseis tervikuna peegeldab koosluste
varast arengujärku. Kõdupuitu asustavate epiksüülsete samblike koosluste tekkimist on varem
uuritud kändudel (Caruso & Rudolphi 2009) ja lamatüvedel (Söderström 1988, Botting et al.
2009). Mõlema puitsubstraadi puhul on täheldatud, et kõigepealt asustavad uue kasvupinna
fakultatiivsed epiksüülid – liigid, mis saavad kasvada ka mujal kui puidul. Edasise kõdunemise
käigus lisanduvad puiduspetsialistid, kellel põhineb tüügaspuude koosluste eripära (Forsslund &
Koffman 1998, Humphrey et al. 2002, Lõhmus & Lõhmus 2001, Nascimbene et al. 2008). Rootsi
segametsade uuringus olid kõigist erinevas kõdunemuses tüügaspuude puidul kasvavatest
liikidest 28% spetsialistid (Forsslund & Koffman 1998). Neile teadmistele tuginedes on liigiline
koosseis käesolevas töö hiljuti paljandunud puiduga tüügaspuudel ootuspärane: enamik leitud
samblikuliikidest asustavad metsas teisigi substraate (elusad puud, maapind), kuid surmast
möödunud ajaga lisandus ka puiduspetsialiste. Viimased moodustasid vaadeldud varastes
kooslustes siiski vaid väikese osa leitud liikidest. Koosluste muutumisele puu surmast möödunud
ajaga viitasid aga ka mõned fakultatiivsed epiksüülid: näiteks liik Dimerella pineti, mida peetakse
eelkõige rohkem kõdunenud kasvupindade asustajaks (Selva 1994, Lõhmus et al. 2010). See liik
leiti kasetüüka puidulaigult; leid viitab, et suuremat kõdunemust eelistavatele liikidele võivad
koore all kiiresti kõdunevad kased (Yatskov et al. 2003) haabadest ja mändidest varem sobivat
elupaika pakkuda.

49
4.3 Järeldused
Antud töö olulisemate tulemustena selgus, et:
1. Säilikpuud pakuvad juba surmast mõne aasta möödudes elupaika liigirikkale
torikseenekooslusele, sh mitmele looduskaitselist väärtust omavale liigile. Puidul kasvavate
samblike koosluste teke seevastu on aeganõudev, ning uuritud (1–8 aastat surnud) tüvedel alles
algusjärgus. Varastes torikseene-kooslustes eristuvad teistest puuliikidest liigirikkuse ja
looduskaitseväärtust omavate liikide arvu poolest haava lamatüved. Epiksüülsete samblike
kooslused on liigirikkamad männi tüügaspuudel.
2. Suurema diameetriga säilikpuudel leidub (viljakehade järgi) suurem arv torikseeneliike. Ka
epiksüülsete samblike puhul tagab kasvupinna suurus (paljandunud puidu osakaal tüvel) elupaiga
enamatele liikidele.
3. Torikseene-elustik ja koosluste arengumuster on kolmel uuritud puuliigil erinev. Puidul
kasvavate samblike liigiline koosseis kaskedel, haabadel ja mändidel kattub, kuid erinevatel
puuliikidel on isesugune koore eemaldumise ja kõdunemise dünaamika. Lisaks asustavad
epiksüülsed samblikud männipuidu haavapuidust rutem – kuid samas kõdunevad haab ja kask
kiiremini ning pakuvad varem elupaika suuremat kõdunemust eelistavatele liikidele. Seetõttu
peaksid nii torikseentest kui puidul kasvavatest samblikest lähtuvalt raiesmikul olema esindatud
erinevat liiki säilikpuud.
4. Kaheksa puu surmast möödunud aastaga leidub torikseenekooslustes esimeste pioneerliikide
kõrval ka „hilisemate“ lagundajaliikide viljakehasid. Looduskaitselist väärtust omavad liigid
eelistavad kõdunenumat puitu, millele viitas ka haaval kasvavate liikide puhul nende sagedasem
esinemine parasiitseene Phellinus tremulae asustatud tüvedel. Puidul kasvavate samblike
koosluseid iseloomustab vaadeldud varases arengujärgus väheste puidule spetsialiseerunud
samblikuliikide esinemine.

50
Käesoleva töö koostamise käigus ilmnes mitmeid küsimusi, mis vajaksid täiendavat uurimist:
1. Looduskaitseliselt väärtuslikud torikseeneliigid esinesid uurimisaladel agregeeritult, sõltudes
arvatavasti liigi esinemisest taustsüsteemis. Seetõttu oleks vaja uurida, milline peaks olema
lättepopulatsiooni suurus, taustaalade pidevus ja kvaliteet (nt kõdupuidu hulga osas), et
looduskaitseväärtust omavad torikseeneliigid saaksid surnud säilikpuud kasvusubstraadina
kasutusele võtta.
2. Töös selgus, et vastpaljandunud männipuitu asustavad epiksüülsed samblikud kiiremini kui
haavapuitu. See erinevus võib olla tingitud koore eemaldumise iseärasustest, kuid ka männi ja
haavapuidu erinevatest füüsikalistest ja keemilistest omadustest. Täpsemate põhjuste edasine
uurimine võimaldaks paremini mõista puidul kasvavate samblike koloniseerimise ja koosluste
moodustumise mehhanisme.
3. Töö käsitles vaid raiejärgse metsa taastumisperioodi algfaasi. Mõistmaks paremini säilikpuude
rolli torikseente ja puidul kasvavate samblike „pagulana“ majandusmetsas, tuleks koosluseid
uurida ka kauem kõdunenud säilikpuudel hilisemates metsasuktsessiooni faasides.

51
KOKKUVÕTE
Säilitamaks majandatavas metsamaastikus elurikkust jäetakse raiesmikele säilikpuid. Säilikpuud
on metsaelustikule olulised ka peale surma, kõdupuiduna, ent nende ökoloogilist rolli
kõdupuidust sõltuvatele organismidele on uuritud väga napilt. Käesolev töö keskendub kahele
sellisele liigirühmale: torikseentele ja epiksüülsetele samblikele. Varasematest analoogilistest
uuringutest eristub töö aegrealise disaini poolest: koosluste kirjeldamisel lähtutakse teadaolevast
puu surmast möödunud ajast. Töö eesmärgiks on uurida (1) millised surnud säilikpuude
omadused mõjutavad torikseente ja epiksüülsete samblike esinemist ja liigirikkust ning (2) kuidas
muutuvad surnud säilikpuude seenekooslused uuritud perioodil, 1–8 surmajärgsel aastal.
Tuginedes eelnevale uurimusele (Rosenvald et al. 2008), kirjeldati 46 alal Tartu-, Pärnu-,
Viljandi- ja Järvamaal säilikpuudest tekkinud 132 lamatüve ning 94 tüügaspuu (kased, haavad ja
männid) torikseenekooslusi ning tüügaspuudel ka epiksüülsete samblike kooslusi. Lisaks mõõdeti
uuritavaid liigirühmasid mõjutada võivaid kasvusubstraadi omadusi (tüve läbimõõt, puidu
kõdustaadium, valgustatus ja kaugus metsast) ning kaasati liikide leviku seisukohast olulisi
raiesmiku tunnuseid (nt lamapuude maht raiesmikul).
Surnud säilikpuudelt leiti 60 liiki torikseeni (sh mitu looduskaitseliselt huvipakkuvat liiki) ja 43
liiki puidul kasvavaid samblikke. Säilikpuud pakuvad juba surmast mõne aasta möödudes
elupaika liigirikkale torikseenekooslusele. Seevastu aeganõudvam epiksüülsete samblike
koosluste kujunemine oli uuritud tüvedel alles algusjärgus. Puuliikidest eristusid liigirikkuse
poolest torikseente puhul haava lamatüved (millel oli sh ka suhteliselt palju looduskaitseliselt
huvipakkuvaid liike) ja epiksüülsete samblike puhul männi tüügaspuud. Torikseene-elustik oli
kasel, haaval ja männil erinev, samas epiksüülsete samblike liigiline koosseis uuritud puuliikidel
kattus. Erinevatel puuliikidel oli aga isesugune koore eemaldumise ja kõdunemise dünaamika.
Leiti, et epiksüülsed samblikud asustavad männipuitu haavapuidust rutem, samas kõdunevad
haab ja kask kiiremini ning pakuvad seetõttu varem elupaika suuremat lagunemust eelistavatele
liikidele. Mõlemal uuritud rühmal mõjutas ootuspäraselt liikide arvu tüvel kasvupinna suurus:

52
suurema läbimõõduga säilikpuudel leidus (viljakehade järgi) rohkem torikseeneliike ning
suurema paljandunud puidu osakaaluga tüvedel oli enam epiksüülseid samblikuliike.
Kaheksa puu surmast möödunud aastaga leidus torikseenekooslustes esimeste pioneerliikide
kõrval ka „hilisemate“ lagundajaliikide viljakehasid. Looduskaitseliselt huvipakkuvad liigid
eelistasid kõdunenumat puitu, millele viitas haava puhul ka selliste liikide sagedasem esinemine
parasiitseene Phellinus tremulae asustatud tüvedel. Puidul kasvavate samblike kooslustes oli
vaadeldud varases arengujärgus vähe puidule spetsialiseerunud samblikuliike.
Edaspidised surnud säilikpuude torikseene- ja epiksüülsete samblike kooslusi käsitlevad tööd
võiksid (1) uurida ka ajaliselt kauem kõdunenud säilikpuid hilisemates metsasuktsessiooni
faasides ja (2) kaasata ümbritsevate alade liikide leviku seisukohast olulisi omadusi, uurimaks,
milline peaks olema taustaalade kvaliteet võimaldamaks liikidel surnud säilikpuud
kasvusubstraadina kasutusele võtta.

53
Polypore fungi and epixylic lichens on dead retention trees
Kadri Runnel
SUMMARY
In order to preserve biodiversity in woodlands, retention trees are left on cut areas. These trees
are essential even after their death, but their role in ecosystems as sustainers of organisms
depending on dead wood has not been studied much. This paper focuses on two of such species
groups – polypore fungi and epixylic lichens. The difference from earlier similar studies lies in
the chronosequence design of the study – the period from the death of each tree was known and
used for describing the communities. The aim of the work is to study (1) which characteristics of
retention trees influence the occurrence and species-richness of polypore fungi and epixylic
lichens, and (2) how the mycobiota on retention trees changes from 1 to 8 years after the death of
the tree.
Polypore fungal communities were described on 132 fallen trunks and 94 snags (birch, aspen, or
pine), epixylic lichen communities only on snags within 46 areas in Tartu, Pärnu, Viljandi and
Järva Counties. In addition, characteristics of the dead retention trees that may have an impact on
fungal communities (diameter of the trunk, decay stage, light conditions and distance from forest)
were measured, and some characteristics of the cut areas essential for fungi (such as the volume
of fallen retention trees on the cut area) were included.
The number of species found on dead retention trees was 60 for polypore fungi (incl. several
species of conservation concern), and 43 for epixylic lichens. Within even a few years after the
death of the tree the dead wood becomes a habitat for a species-rich polypore fungal community.
The formation of epixylic lichen communities takes more time, and on the tree trunks studied it
was still in the formative stage. Of tree species studied, the number of polypore fungi was the
highest on fallen aspen trunks (where several species of conservation concern were also found),
while the number of epixylic lichens was highest on pine snags. The communities of polypore

54
fungi varied by tree species, but epixylic lichen communities were more or less similar. However,
birches, pines and aspens have their own characteristic decortication and decomposing dynamics.
It was found that epixylic lichens colonise pine trunks faster than aspen trunks, but as the decay
processes of aspen and birch trunks are quicker, those trees offer a suitable environment for
species preferring a higher degree of decay sooner. As expected, both studied species groups
were influenced by the substrate area – trees with larger diameters had more species of polypore
fungi (counted by their fruit bodies), and the trunks with larger areas of debarked wood had a
larger number of epixylic lichen species.
Within eight years after the death of the tree, the first fruit bodies of so-called later decomposers
also appeared in polypore fungal communities. The species of conservation concern preferred
higher level of decay, which was also indicated by the fact that on aspen, these species occurred
more frequently on trunks colonised by the parasitic fungus Phellinus tremulae. In the formative
stage of lichen communities, the number of true epixylic lichen species was small.
Further studies on polypore fungi and epixylic lichens on dead retention trees should
(1) include retention trees in later decomposition phases and during the more advanced stages of
forest succession, and (2) incorporate characteristics of surrounding areas essential to the
spreading of these species, in order to determine the qualitative parameters of background areas
needed to enable these species to use retention trees as their growing substrate.

55
TÄNUSÕNAD
Selle töö valmimisel oli mul palju hindamatuid abilisi. Kogu tööprotsessi käigus andsid häid
nõuandeid juhendajad Kadi Jairus ja Raul Rosenvald. TÜ Ökofüsioloogia töörühma liikmed
laenasid fototehnikat ning Kalle Pilt EMÜst resistograafi. Välitöödel olid suureks abiks lisaks
(sääsehulgast sõltumatult heatujulisele) Raul Rosenvaldile ka Marianne Pitk ja Annika Karusion.
TÜ looduskaitsebioloogia töörühma sõbralik seltskond mõtles seminarides kaasa ja jagas oma
kogemusi. Eraldi soovin tänada Piret Lõhmust, kes juhendas asjatundlikult samblike
tundmaõppimisel ja määramisel ning Asko Lõhmust, kes tutvustas torikseentega, andis nõu siis
kui teised enam ei osanud – ning oli heaks eeskujuks.
Tänan ka abikaasa Tõnu ja poeg Tõnist, minu väikest vahvat peret, kes magistritöö kirjutamisele
kaasa elas.
Töö valmimist toetasid rahaliselt Eesti Teadusfond (projekt 7402) ning Haridus- ja
teadusministeerium (sihtfinantseeritav teadusteema 0180012s09), mõlemil vastutav täitja Asko
Lõhmus.

56
KASUTATUD KIRJANDUS
Baum S, Sieber TN, Schwarze FWMR, Fink S. 2003. Latent infections of Fomes fomentarius
in the xylem of European beech (Fagus sylvatica). Mycological Progress 2: 141–148.
Berglund H, Hottola J, Penttilä R, Siitonen J. 2011b. Linking substrate and habitat
requirements of wood-inhabiting fungi to their regional extinction vulnerability. Ecography, in
press.
Berglund H, Jönsson MT, Penttilä R, Vanha-Majamaa I. 2011a. The effects of burning and
dead-wood creation on the diversity of pioneer wood-inhabiting fungi in managed boreal spruce
forests. Forest Ecology and Management 261: 1293–1305.
Boddy L. 2000. Interspecific combative interactions between wood-decaying basidiomycetes.
FEMS Microbiology Ecology 31: 185–194.
Boddy L. 2001. Fungal community ecology and wood decomposition processes in angiosperms:
from standing tree to complete decay of coarse woody debris. Ecological Bulletins 49: 43–56.
Boddy L, Heilmann-Clausen J. 2008. Basidiomycete community development in temperate
angiosperm wood. In: Boddy L, Frankland JC, van West P, eds. Ecology of saprotrophic
basidiomycetes. Eastbourne, United Kingdom: Academic Press. 211–238.
Botting R, DeLong C. 2009. Macrolichen and bryophyte responses to coarse woody debris
characteristics in sub-boreal spruce forest. Forest Ecology and Management 258: 85–94.
Bunnell FL, Spiribille T, Houde I, Goward T, Björk C. 2008. Lichens on down wood in
logged and unlogged forest stands. Canadian Journal of Forest Research 38: 1033–1041.
Büdel B, Scheidegger C. 1996. Thallus morphology and anatomy. In: Nash TH III, ed. Lichen
biology. Cambridge, United Kingdom: Cambridge University Press, 37–64.
Caruso A, Rudolphi J. 2009. Influence of substrate age and quality on species diversity of
lichens and bryophytes on stumps. Bryologist 112: 520–531.
Caruso A, Thor G, Snäll T. 2010. Colonization-extinction dynamics of epixylic lichens along a
decay gradient in a dynamic landscape. Oikos 119: 1947–1953.
Chen J, Saunders SC, Crow TR, Naiman RJ, Brosofske KD, Mroz GD, Brookshire BL,
Franklin JF. 1999. Microclimate in forest ecosystem and landscape ecology. BioScience 49:
288–297.

57
Chlebicki A, Żarnowiec J, Cieśliński S, Klama K, Bujakiewicz A, Załuski T, 1996.
Epixylites, lignicolous fungi and their links with different kinds of wood. Phytocoenosis 8: 75–
109.
Cieśliński S, Czyżewska K, Klama H, Żarnowiec J, 1996. Epiphytes and epiphytism.
Phytocoenosis 8: 15–35.
Colwell RK, Mao CX, Chang J. 2004. Interpolating, extrapolating, and comparing incidence-
based species accumulation curves. Ecology 85: 2717–2727.
Creed IF, Morrison DL, Nicholas MS. 2004. Is coarse woody debris a net sink or source of
nitrogen in the red spruce - fraser fir forest of the southern appalachians, U.S.A.? Canadian
Journal of Forest Research 34: 716-727.
Crites S, Dale MRT. 1998. Diversity and abundance of bryophytes, lichens, and fungi in relation
to woody substrate and successional stage in aspen mixedwood boreal forests. Canadian Journal
of Botany 76: 641–651
Edman M, Gustafsson M, Stenlid J, Ericson L. 2004a. Abundance and viability of fungal
spores along a forestry gradient – responses to habitat loss and isolation? Oikos 104: 35–42.
Edman M, Gustafsson M, Stenlid J, Jonsson BG, Ericson L. 2004b. Spore deposition of
wood-decaying fungi: importance of landscape composition. Ecography 27: 103–111.
Edman M, Möller R, Ericson L. 2006. Effects of enhanced tree growth rate on the decay
capacities of three saprotrophic wood-fungi. Forest Ecology and Management 232: 12–18.
Ellis CJ, Coppins BJ. 2007. Reproductive strategy and the compositional dynamics of crustose
lichen communities on aspen (Populus tremula L.) in Scotland. The Lichenologist 39: 377–391.
Esseen P-A, Ehnström B, Ericson L, Sjöberg K. 1992. Boreal forests – the focal habitats of
Fennoscandia. In: Hansson L, ed. Ecological Principles of Nature Conservation. Elsevier, London
and New York, 252–325.
Farris KL, Huss MJ, Zack S. 2004. The role of foraging woodpeckers in the decomposition of
ponderosa pine snags. Condor 106: 50–59.
Franklin JF, Berg DR, Thornburgh DA, Tappeiner JC. 1997. Alternative silvicultural
approaches to timber harvesting: variable retention harvest systems. In: Kohm KA, Franklin JF,
eds. Creating a Forestry for the 21st Century: The Science of Ecosystem Management.
Washington, USA: Island Press, 111–139.

58
Fries C, Johansson O, Pettersson B, Simonsson P. 1997. Silvicultural models to maintain and
restore natural stand structures in swedish boreal forests. Forest Ecology and Management 94:
89-103.
Forsslund A, Koffman A. 1998. Species diversity of lichens on decaying wood – a comparison
between old-growth and managed forest. Växtekologi 2: 1–40.
Gotelli NJ, Colwell RK. 2001. Quantifying biodiversity: Procedures and pitfalls in the
measurement and comparison of species richness. Ecology Letters 4: 379–391.
Gustafsson L, Kouki J, Sverdrup-Thygeson A. 2010. Tree retention as a conservation measure
in clear-cut forests of northern Europe: a review of ecological consequences. Scandinavian
Journal of Forest Research 25: 295–308.
Gärdenfors U. 2005. The 2005 red list of Swedish species. Uppsala: ArtDatabanken, SLU.
Harmon ME. 1982. Decomposition of standing dead trees in the southern Appalachian
mountains. Oecologia 52: 214–215.
Hazell P, Gustafsson L. 1999. Retention of trees at final harvest. Evaluation of a conservation
technique using epiphytic bryophyte and lichen transplants. Biological Conservation 90: 133–
142.
Heilmann-Clausen J. 2001. A gradient analysis of communities of macrofungi and slime
moulds on decaying beech logs. Mycological Research 105: 575–596.
Heilmann-Clausen J, Boddy L. 2005. Inhibition and simulation effects in communities of wood
decay fungi: Exudates from colonized wood influence growth by other species. Microbal Ecology
49: 1–8.
Heilmann-Clausen J, Christensen M. 2003. Fungal diversity on decaying beech logs –
implications for sustainable forestry. Biodiversity and Conservation 12: 953–973.
Heilmann-Clausen J, Christensen M. 2004. Does size matter? on the importance of various
dead wood fractions for fungal diversity in danish beech forests. Forest Ecology and
Management 201: 105–117.
Heilmann-Clausen J, Christensen M. 2005. Wood-inhabiting macrofungi in Danish beech-
forests. Conflicting diversity and red list species. Biological Conservation 122: 633–642.
Hilmo O, Holien H, Hytteborn H, Ely-Aalstrup H. 2009. Richness of epiphytic lichens in
differently aged Picea abies plantations situated in the oceanic region of Central Norway. The
Lichenologist 41: 97–108.

59
Holien H. 1996. Influence of site and stand factors on the distribution of crustose lichens of the
Caliciales in a suboceanic spruce forest area in Central Norway. Lichenologist 28: 315–330.
Holmer L, Stenlid J, Renvall P. 1997. Selective replace ment between species of wood-rotting
basidiomycetes, a lab oratory study. Mycology Research 101: 714-720.
Hottola J, Ovaskainen O, Hanski I. 2009. A unified measure of the number, volume and
diversity of dead trees and the response of fungal communities. Journal of Ecology 97: 1320–
1328.
Høiland K, Bendiksen E. 1997. Biodiversity of wood-inhabiting fungi in a boreal coniferous
forest in Sør-Trondelag County, Central Norway. Nordic Journal of Botany 16: 643–659.
Humphrey JW, Davey S, Peace AJ, Ferris R, Harding K. 2002. Lichens and bryophyte
communities communities of planted and semi-natural forests in Britain: the influence of site
type, stand structure and deadwood. Biological Conservation 107: 165–180.
Jairus K, Lõhmus A, Lõhmus P. 2009. Lichen acclimatization on retention trees: A
conservation physiology lesson. Journal of Applied Ecology 46: 930–936.
Jonsson BG, Kruys N, Ranius T. 2005. Ecology of species living on dead wood – Lessons for
dead wood management. Silva Fennica 39: 289–309.
Junninen K, Similä M, Kouki J, Kotiranta H. 2006. Assemblages of wood-inhabiting fungi
along the gradients of succession and naturalness in boreal pine-dominated forests in
Fennoscandia. Ecography 29: 75–83.
Junninen K, Penttilä R, Martikainen P. 2007. Fallen retention aspen trees on clear-cuts can be
important habitats for red-listed polypores: A case study in Finland. Biodiversity and
Conservation 16: 475–490.
Jüriado I, Paal J, Liira J. 2003. Epiphytic and epixylic lichen species diversity in Estonian
natural forest. Biodiversity and Conservation 12: 1587–1607.
Kahl T, Wirth C, Mund M, Böhnisch G, Schulze ED. 2009. Using drill resistance to quantify
the density in coarse woody debris of norway spruce. European Journal of Forest Research 128:
467–473.
Kaila L, Martikainen P, Punttila P. 1997. Dead trees left in clear-cuts benefit saproxylic
coleoptera adapted to natural disturbances in boreal forest. Biodiversity and Conservation 6: 1-
18.

60
Komonen A, Penttilä R, Lindgren M, Hanski I. 2000. Forest fragmentation truncates a food
chain based on an old-growth forest bracket fungus. Oikos 90: 119–126.
Kotiranta H, Saarenoksa R, Kytövuori I. 2009. Aphyllophoroid fungi of Finland. A check-list
with ecology, distribution, and threat categories. Norrlinia 19: 1–223.
Krankina ON, Harmon ME, Griazkin AV. 1999. Nutrient stores and dynamics of woody
detritus in a boreal forest: modeling potential implications at the stand level. Canadian Journal of
Forest Research 29: 20–32.
Kruys N, Fries C, Jonsson BG, Lämås T, Ståhl G. 1999. Wood inhabiting cryptogams on dead
Norway spruce (Picea abies) trees in managed Swedish boreal forests. Canadian Journal of
Forest Research 29: 178–186.
Kruys N, Jonsson G. 1997. Insular patterns of calicioid lichens in a boreal old-growth forest-
wetland mosaic. Ecography 20: 605–613.
Kuusinen M. 1996. Epiphyte flora and diversity on basal trunks of six old-growth forest tree
species in southern and middle boreal Finland. Lichenologist 28: 443–463.
Larjavaara M, Muller-Landau H. 2010. Comparison of decay classification, knife test, and two
penetrometers for estimating wood density of coarse woody debris. Canadian Journal of Forest
Research 40: 2313–2321.
Lange, M. 1992. Sequence of Macromycetes on decaying beech logs. Persoonia 14: 449–456.
Lindhe A, Asenblad N, Toresson HG. 2004. Cut logs and high stumps of spruce, birch, aspen
and oak – nine years of saproxylic fungi succession. Biological Conservation 119: 443–454.
Lindenmayer DB, Franklin JF. 2002. Conserving forest biodiversity: A comprehensive
multiscaled approach. Washington, USA: Island Press.
Linder P, Elfving B, Zackrisson O. 1997. Stand structure and successional trends in virgin
boreal forest reserves in Sweden. Forest Ecology and Management 98: 17–33.
Loehle C. 1996. Optimal defensive investments in plants. Oikos 75: 299–302.
Lõhmus, A. 2011a. Aspen-inhabiting Aphyllophoroid fungi in a managed forest landscape in
Estonia. Scandinavian Journal of Forest Research 26: 212–220.
Lõhmus, A. 2011b. Silviculture as a disturbance regime: the effects of clear-cutting, planting and
thinning on polypore communities in mixed forests. Journal of Forest Research: in press.
Lõhmus A, Lõhmus P. 2008. First-generation forests are not necessarily worse than long-term
managed forests for lichens and bryophytes. Restoration Ecology 16: 231–239.

61
Lõhmus P. 2004. Perekond Mycocalicium Vain. (1890). In: Randlane T, Saag A, eds. Eesti
pisisamblikud. Tartu, Estonia: Tartu University Press.388–389.
Lõhmus P, Lõhmus A. 2001. Snags and their lichen flora in old Estonian peatland forests.
Annales Botanici Fennici 38: 265–280.
Lõhmus P, Motiejunaite J. 2008. Epixylic lichens in Fennoscandia, Estonia and Lithuania:
patterns of specificity. In: Saar I, Suija, A, eds. Abstracts of XVII Symposium of Baltic
Mycologists and Lichenologists: XVII Symposium of Baltic Mycologists and Lichenologists.
Tartu, Estonia: Tartu University Press.
Lõhmus P, Rosenvald R, Lõhmus A. 2006. Effectiveness of solitary retention trees for
conserving epiphytes: differential short-term responses of bryophytes and lichens. Canadian
Journal of Forest Research 36: 1319–1330.
Lõhmus P, Turja K, Lõhmus A. 2010. Lichen communities on treefall mounds depend more on
root-plate than stand characteristics. Forest Ecology and Management 260: 1754–1761.
Martikainen P. 2001. Conservation of threatened saproxylic beetles: significance of retained
aspen Populus tremula on clearcut areas. Ecological Bulletins 49: 205–218.
McCune B, Mefford MJ. 1999. PC-ORD. Multivariate Analysis of Ecological Data. USA,
Oregon: MjM Software.
Moen J, Jonsson BG. 2003. Edge effects on liverworts and lichens in forest patches in a mosaic
of boreal forest and wetland. Conservation Biology 17: 380-388.
Mueller GM, Schmit JP, Leacock PR, Buyck B, Cifuentes J, Desjardin DE, Halling RE,
Hjortstam K, Iturriaga T, Larsson KH, Lodge DJ, May TW, Minter D, Rajchenberg M,
Redhead SA, Ryvarden L, Trappe JM, Watling R, Wu Q. 2007. Global diversity and
distribution of macrofungi. Biodiversity and Conservation 16: 37–48.
Nascimbene J. Marini L, Motta R, Nimis PL. 2008. Lichen diversity of coarse woody habitats
in a Pinus-Larix stand in the Italian Alps. Lichenologist 40: 153–163.
Nash TH III. 1996. Phothosynthesis, respiration, productivity and growth. In: Nash TH III, ed.
Lichen Biology. Cambridge, United Kingdom: Cambridge University Press. 88–120.
Niemelä T. 2008. Torikseened Soomes ja Eestis. Tartu, Estonia: Eesti Loodusfoto.
Niemelä T, Renvall P, Penttila R. 1995. Interactions of fungi at late stages of wood
decomposition. Annales Botanici Fennici 32: 141–152.

62
Nitare J, ed. 2000. Indicator species for assessing the nature conservation value of woodlands
sites — a flora of selected cryptogams. Jönköping, Sweden: Skogsstyrelsen.
Nordén B, Ryberg M, Go ̈tmark F, Olausson B. 2004. Relative importance of coarse and fine
woody debris for the diversity of wood-inhabiting fungi in temperate broadleaf forests. Biological
Conservation 117: 1–10.
Olsson J, Jonsson BG, Hjältén J, Ericson L. 2011. Addition of coarse woody debris - the early
fungal succession on picea abies logs in managed forests and reserves. Biological Conservation
144: 1100–1110.
Orange A, James PW, White FJ. 2001. Microchemical methods for the identification of lichens.
London, United Kingdom: British Lichen Society.
Ovaskainen O, Hottola J, Sihtonen J. 2010a. Modeling species co-occurrence by multivariate
logistic regression generates new hypotheses on fungal interactions. Ecology 91: 2514-2521.
Ovaskainen O, Nokso-Koivisto J, Hottola J, Rajala T, Pennanen T, Ali-Kovero H,
Miettinen O, Oinonen P, Auvinen P, Paulin L, Larsson KH, Mäkipää, R. 2010b. Identifying
wood-inhabiting fungi with 454 sequencing: what is the probability that BLAST gives the correct
species? Fungal Ecology 3: 274-283.
Parmasto E. 2004. Eesti seente levikuatlas. III. Torikseened. Tartu, Estonia: EPMÜ Zooloogia ja
Botaanika Instituut.
Penttilä R, Siitonen J, Kuusinen M. 2004. Polypore diversity in managed and oldgrowth boreal
Picea abies forests in southern Finland. Biological Conservation 117: 271–283.
Rajala T, Peltoniemi M, Pennanen T, Mäkipää R. 2010. Relationship between wood-
inhabiting fungi determined by molecular analysis (denaturing gradient gel electrophoresis) and
quality of decaying logs. Canadian Journal of Forest Research 40: 2384-2397.
Randlane T, Saag A, eds. 2004. Eesti pisisamblikud. Tartu, Estonia: Tartu University Press.
Renvall P. 1995. Community structure and dynamics of wood-rotting fungi on decomposing
conifer trunks in northern Finland. Karstenia 35: 1–51.
Rikkinen J. 1995. Boreal Caliciales – Life in a cheimophotophytic habitat. IN: Rikkinen J.
What’s behind the pretty colours? A study on the photobiology of lichens. Bryobrothera 4: 145–
163.
Risbeth J. 1959. Dispersal of Fomes annosus and Peniophora gigantea. Transactions of the
British Mycological Society 42: 243–260.

63
Rolstad J, Sætersdal M, Gjerde I, Storaunet KO. 2004. Wood-decaying fungi in boreal forest:
are species richness and abundance influenced by small-scale spatiotemporal distribution of dead
wood? Biological Conservation 177: 539–555.
Rosenvald R, Lõhmus A. 2007. Breeding birds in hemiboreal clear-cuts: Tree-retention effects
in relation to site type. Forestry 80: 503-516.
Rosenvald R, Lõhmus A. 2008. For what, when, and where is green-tree retention better than
clear-cutting? A review of the biodiversity aspects. Forest Ecology and Management 255: 1–15.
Rosenvald R, Lõhmus A, Kiviste A. 2008. Preadaptation and spatial effects on retention-tree
survival in cut areas in Estonia. Canadian Journal of Forest Research 38: 2616–2625.
Ryvarden L, Gilbertson RL. 1993–94. European Polypores, Vol. 1–2. Oslo, Norway:
Fungiflora
Shorohov EV, Shorohova AA. 2001. Coarse woody debris dynamics and stores in a boreal
virgin spruce forest. Ecological Bulletins 49: 129–135.
Selva SB, 1994. Lichen diversity and stand continuity in the northern hardwoods and spruce-fir
forests of northern New England and western New Brunswick. The Bryologist 97: 424–429.
Siitonen J. 2001. Forest management, coarse woody debris and saproxylic organisms:
Fennoscandian boreal forests as an example. Ecological Bulletins 49: 11–41.
Sippola A-L, Siitonen J, Kallio R. 1998. Amount and quality of coarse woody debris in natural
and managed coniferous forests near the timberline in Finnish Lapland. Scandinavian Journal of
Forest Research 13: 204–214.
Sippola AL, Renvall P. 1999. Wood-decomposing fungi and seed-tree cutting: a 40 year
perspective. Forest Ecology and Management 115: 183–201.
Sippola AL, Similä M, Mönkkönen M, Jokimäki J. 2004. Diversity of polyporous fungi
(Polyporaceae) in northern boreal forests: effects of forest site type and logging intensity.
Scandinavian Journal of Forest Research 19: 152–163.
Spribille T, Thor G, Bunnell FL, Goward T, Björk CR. 2008. Lichens on dead wood: species
substrate relationships in the epiphytic lichen floras of the Pacific Northwest and Fennoscandia.
Ecography 31: 741–750
Stenlid J, Penttilä R, Dahlberg A. 2008. Wood-decay basidiomycetes in boreal forests:
distribution and community development. In: Boddy L, Frankland JC, van West P, eds. Ecology
of saprotrophic basidiomycetes. Eastbourne, United Kingdom: Academic Press. 211–238.

64
Sullivan TP, Sullivan DS, Lindgren PMF. 2001. Influence of variable retention harvests on
forest ecosystems. I. Diversity of stand structure. Journal of Applied Ecology 38: 1221–1233.
Söderström L. 1988. Sequense of bryophytes and lichens in relation to substrate variables of
decaying coniferous wood in Northern Sweden. Nordic Journal of Botany 8: 89–97.
Tamm, Ü. 2000. Haab Eestis. Tallinn, Estonia: Eesti Loodusfoto.
Tibell L. 1999. Calicioid lichens and fungi – Nordic Lichen Flora 1: 20–94.
Trass H, Randlane T, eds. 1994. Eesti suursamblikud. Tartu, Estonia: Tartu University Press.
Trass H, Vellak K, Ingerpuu N. 1999. Floristical and ecological properties for identifying of
primeval forests in Estonia. Annales Botanici Fennici 36: 67–80.
Vanha-Majamaa I, Jalonen J. 2001. Green tree retention in Fennoscandian forestry.
Scandinavian Journal of Forest Research Suppl 3: 79–90.
Vasiliauskas R, Stenlid J. 1999. Vegetative compatibility groups of Amylostereum areolatum
and A. chailletii from Sweden and Lithuania. Mycological Research 102: 824–829.
Walser JC, Holderegger F, Gugerli S, Hoebee E, Scheidegger C. 2005. Microsatellites reveal
regional population differentiation and isolation in Lobaria pulmonaria, an epiphytic lichen.
Molecular Ecology 14: 457–467.
Öckinger E, Niklasson M, Nilsson SG. 2005. Is local distribution of the epiphytic lichen
Lobaria pulmonaria limited by dispersal capasity or habitat quality? Biodiversity and
Conservation 14: 759–773.
Yatskov M, Harmon ME, Krankina ON. 2003. A chronosequence of wood decomposition in
the boreal forests of Russia. Canadian Journal of Forest Research 33: 1211–1226.
Internetilehekülg
eElurikkus 2011. Eesti ohustatud liikide punane nimestik
[http://elurikkus.ut.ee/prmt.php?lang=est]. Vaadatud 10.02.2011

65
LISAD
Lisa 1. Uurimisalade (48) asukoht, peamised parameetrid ja uuritud surnud säilikpuud.
Lisa 2. Surnud säilikpuudelt (lamatüvedelt ja tüügaspuudelt) leitud torikseeneliigid ja nende
esinemissagedus.
Lisa 3. Surnud säilikpuudelt (tüügaspuudelt) leitud epiksüülsed samblikuliigid ja nende
esinemissagedus.
Lisa 4. Torikseene liikide arvu sõltuvus lamatüvede ja tüügaspuude surmast möödunud ajast ning omadustest.
Lisa 5. Torikseene liikide arv tüügaspuudel sõltuvalt tüve surmast möödunud ajast.
Lisa 6. Torikseene suhtelise liikide arvu sõltuvus surnud säilikpuude mahust raiesmikul.
Lisa 7. Epiksüülsete samblike liikide arvu ja suhtelise katvuse sõltuvus tüügaspuude surmast möödunud ajast ning omadustest.
Lisa 8. Epiksüülsete samblike suhtelise liikide arvu sõltuvus vanade tüügaspuude arvust raiesmikul.
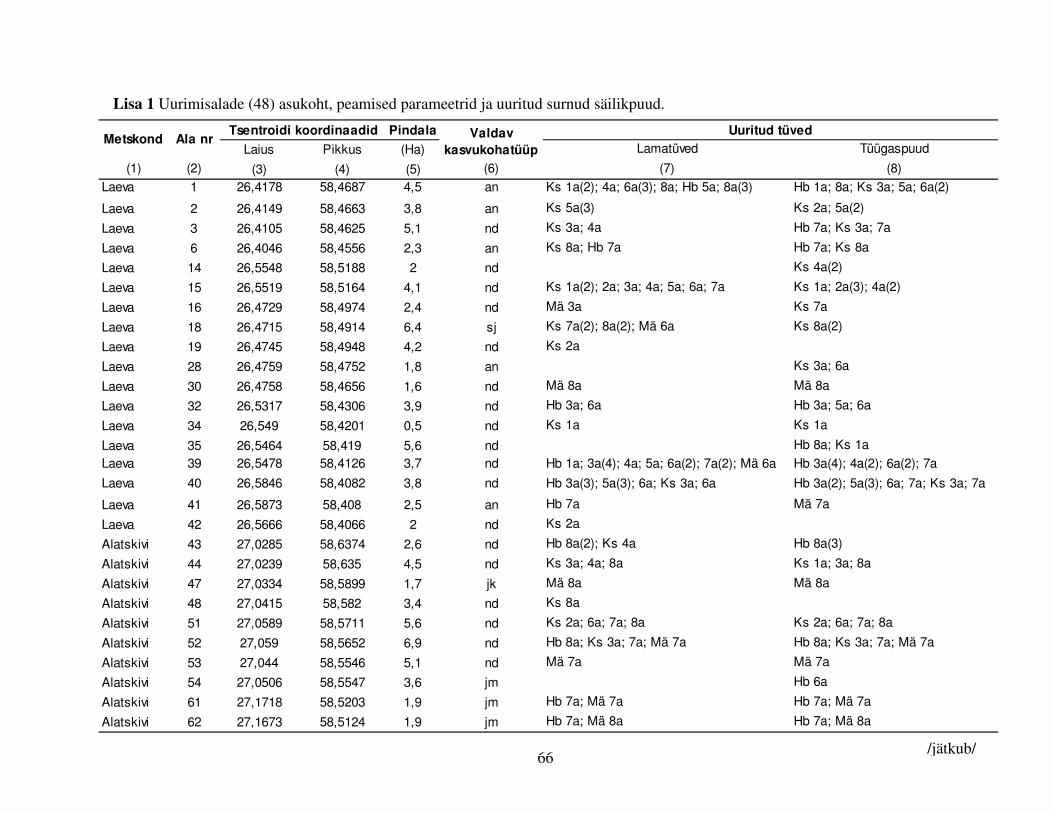
66
Lisa 1 Uurimisalade (48) asukoht, peamised parameetrid ja uuritud surnud säilikpuud.
Pindala
Laius Pikkus (Ha) Lamatüved Tüügaspuud
(1) (2) (3) (4) (5) (6) (7) (8)
Laeva 1 26,4178 58,4687 4,5 an Ks 1a(2); 4a; 6a(3); 8a; Hb 5a; 8a(3) Hb 1a; 8a; Ks 3a; 5a; 6a(2)
Laeva 2 26,4149 58,4663 3,8 an Ks 5a(3) Ks 2a; 5a(2)
Laeva 3 26,4105 58,4625 5,1 nd Ks 3a; 4a Hb 7a; Ks 3a; 7a
Laeva 6 26,4046 58,4556 2,3 an Ks 8a; Hb 7a Hb 7a; Ks 8a
Laeva 14 26,5548 58,5188 2 nd Ks 4a(2)
Laeva 15 26,5519 58,5164 4,1 nd Ks 1a(2); 2a; 3a; 4a; 5a; 6a; 7a Ks 1a; 2a(3); 4a(2)
Laeva 16 26,4729 58,4974 2,4 nd Mä 3a Ks 7a
Laeva 18 26,4715 58,4914 6,4 sj Ks 7a(2); 8a(2); Mä 6a Ks 8a(2)
Laeva 19 26,4745 58,4948 4,2 nd Ks 2a
Laeva 28 26,4759 58,4752 1,8 an Ks 3a; 6a
Laeva 30 26,4758 58,4656 1,6 nd Mä 8a Mä 8a
Laeva 32 26,5317 58,4306 3,9 nd Hb 3a; 6a Hb 3a; 5a; 6a
Laeva 34 26,549 58,4201 0,5 nd Ks 1a Ks 1a
Laeva 35 26,5464 58,419 5,6 nd Hb 8a; Ks 1a
Laeva 39 26,5478 58,4126 3,7 nd Hb 1a; 3a(4); 4a; 5a; 6a(2); 7a(2); Mä 6a Hb 3a(4); 4a(2); 6a(2); 7a
Laeva 40 26,5846 58,4082 3,8 nd Hb 3a(3); 5a(3); 6a; Ks 3a; 6a Hb 3a(2); 5a(3); 6a; 7a; Ks 3a; 7a
Laeva 41 26,5873 58,408 2,5 an Hb 7a Mä 7a
Laeva 42 26,5666 58,4066 2 nd Ks 2a
Alatskivi 43 27,0285 58,6374 2,6 nd Hb 8a(2); Ks 4a Hb 8a(3)
Alatskivi 44 27,0239 58,635 4,5 nd Ks 3a; 4a; 8a Ks 1a; 3a; 8a
Alatskivi 47 27,0334 58,5899 1,7 jk Mä 8a Mä 8a
Alatskivi 48 27,0415 58,582 3,4 nd Ks 8a
Alatskivi 51 27,0589 58,5711 5,6 nd Ks 2a; 6a; 7a; 8a Ks 2a; 6a; 7a; 8a
Alatskivi 52 27,059 58,5652 6,9 nd Hb 8a; Ks 3a; 7a; Mä 7a Hb 8a; Ks 3a; 7a; Mä 7a
Alatskivi 53 27,044 58,5546 5,1 nd Mä 7a Mä 7a
Alatskivi 54 27,0506 58,5547 3,6 jm Hb 6a
Alatskivi 61 27,1718 58,5203 1,9 jm Hb 7a; Mä 7a Hb 7a; Mä 7a
Alatskivi 62 27,1673 58,5124 1,9 jm Hb 7a; Mä 8a Hb 7a; Mä 8a
Metskond Ala nrTsentroidi koordinaadid Valdav
kasvukohatüüp
Uuritud tüved
/jätkub/
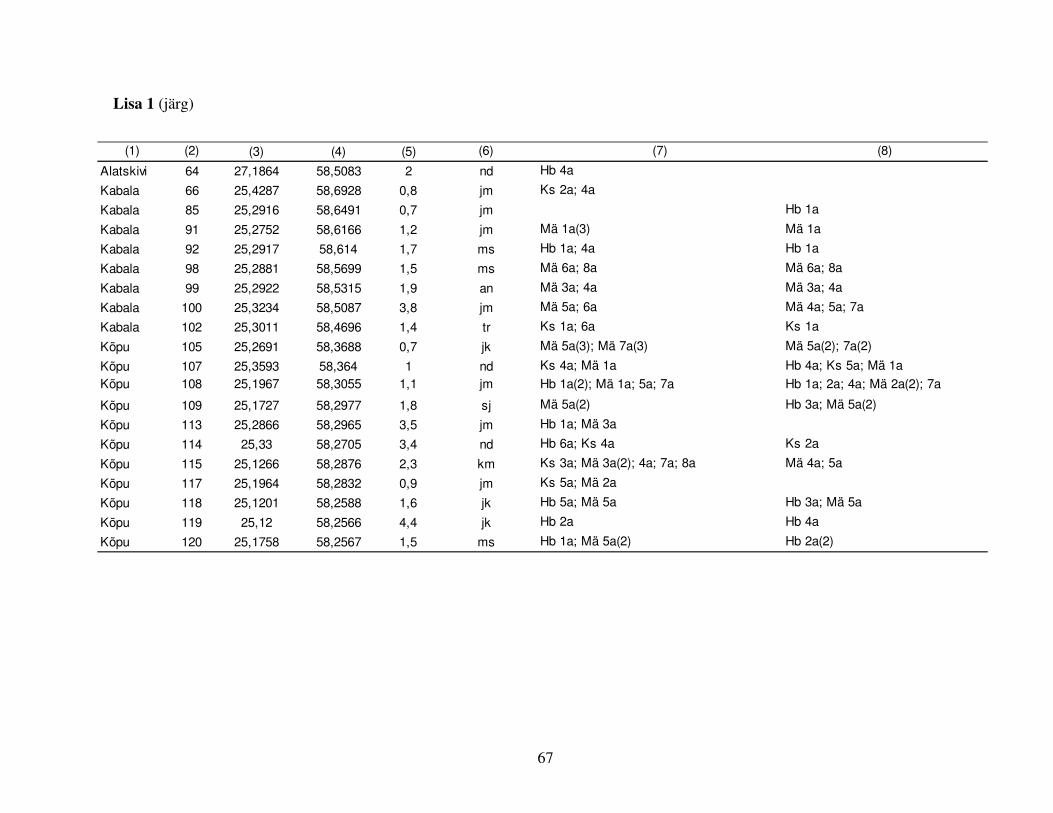
67
Lisa 1 (järg)
(1) (2) (3) (4) (5) (6) (7) (8)
Alatskivi 64 27,1864 58,5083 2 nd Hb 4a
Kabala 66 25,4287 58,6928 0,8 jm Ks 2a; 4a
Kabala 85 25,2916 58,6491 0,7 jm Hb 1a
Kabala 91 25,2752 58,6166 1,2 jm Mä 1a(3) Mä 1a
Kabala 92 25,2917 58,614 1,7 ms Hb 1a; 4a Hb 1a
Kabala 98 25,2881 58,5699 1,5 ms Mä 6a; 8a Mä 6a; 8a
Kabala 99 25,2922 58,5315 1,9 an Mä 3a; 4a Mä 3a; 4a
Kabala 100 25,3234 58,5087 3,8 jm Mä 5a; 6a Mä 4a; 5a; 7a
Kabala 102 25,3011 58,4696 1,4 tr Ks 1a; 6a Ks 1a
Kõpu 105 25,2691 58,3688 0,7 jk Mä 5a(3); Mä 7a(3) Mä 5a(2); 7a(2)
Kõpu 107 25,3593 58,364 1 nd Ks 4a; Mä 1a Hb 4a; Ks 5a; Mä 1a
Kõpu 108 25,1967 58,3055 1,1 jm Hb 1a(2); Mä 1a; 5a; 7a Hb 1a; 2a; 4a; Mä 2a(2); 7a
Kõpu 109 25,1727 58,2977 1,8 sj Mä 5a(2) Hb 3a; Mä 5a(2)
Kõpu 113 25,2866 58,2965 3,5 jm Hb 1a; Mä 3a
Kõpu 114 25,33 58,2705 3,4 nd Hb 6a; Ks 4a Ks 2a
Kõpu 115 25,1266 58,2876 2,3 km Ks 3a; Mä 3a(2); 4a; 7a; 8a Mä 4a; 5a
Kõpu 117 25,1964 58,2832 0,9 jm Ks 5a; Mä 2a
Kõpu 118 25,1201 58,2588 1,6 jk Hb 5a; Mä 5a Hb 3a; Mä 5a
Kõpu 119 25,12 58,2566 4,4 jk Hb 2a Hb 4a
Kõpu 120 25,1758 58,2567 1,5 ms Hb 1a; Mä 5a(2) Hb 2a(2)
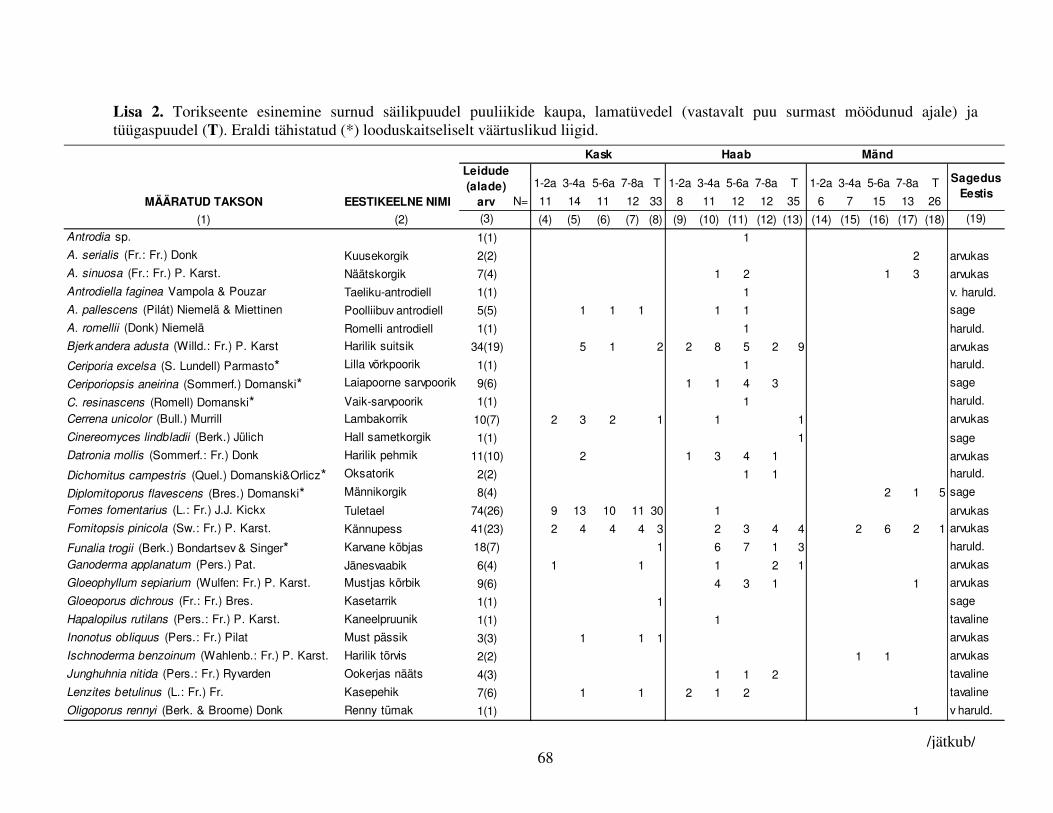
68
Lisa 2. Torikseente esinemine surnud säilikpuudel puuliikide kaupa, lamatüvedel (vastavalt puu surmast möödunud ajale) ja tüügaspuudel (T). Eraldi tähistatud (*) looduskaitseliselt väärtuslikud liigid.
1-2a 3-4a 5-6a 7-8a T 1-2a 3-4a 5-6a 7-8a T 1-2a 3-4a 5-6a 7-8a T
MÄÄRATUD TAKSON EESTIKEELNE NIMI N= 11 14 11 12 33 8 11 12 12 35 6 7 15 13 26
(1) (2) (3) (4) (5) (6) (7) (8) (9) (10) (11) (12) (13) (14) (15) (16) (17) (18) (19)
Antrodia sp. 1(1) 1
A. serialis (Fr.: Fr.) Donk Kuusekorgik 2(2) 2 arvukas
A. sinuosa (Fr.: Fr.) P. Karst. Näätskorgik 7(4) 1 2 1 3 arvukas
Antrodiella faginea Vampola & Pouzar Taeliku-antrodiell 1(1) 1 v. haruld.
A. pallescens (Pilát) Niemelä & Miettinen Poolliibuv antrodiell 5(5) 1 1 1 1 1 sage
A. romellii (Donk) Niemelä Romelli antrodiell 1(1) 1 haruld.
Bjerkandera adusta (Willd.: Fr.) P. Karst Harilik suitsik 34(19) 5 1 2 2 8 5 2 9 arvukas
Ceriporia excelsa (S. Lundell) Parmasto* Lilla võrkpoorik 1(1) 1 haruld.
Ceriporiopsis aneirina (Sommerf.) Domanski* Laiapoorne sarvpoorik 9(6) 1 1 4 3 sage
C. resinascens (Romell) Domanski* Vaik-sarvpoorik 1(1) 1 haruld.
Cerrena unicolor (Bull.) Murrill Lambakorrik 10(7) 2 3 2 1 1 1 arvukas
Cinereomyces lindbladii (Berk.) Jülich Hall sametkorgik 1(1) 1 sage
Datronia mollis (Sommerf.: Fr.) Donk Harilik pehmik 11(10) 2 1 3 4 1 arvukas
Dichomitus campestris (Quel.) Domanski&Orlicz* Oksatorik 2(2) 1 1 haruld.
Diplomitoporus flavescens (Bres.) Domanski* Männikorgik 8(4) 2 1 5 sage
Fomes fomentarius (L.: Fr.) J.J. Kickx Tuletael 74(26) 9 13 10 11 30 1 arvukas
Fomitopsis pinicola (Sw.: Fr.) P. Karst. Kännupess 41(23) 2 4 4 4 3 2 3 4 4 2 6 2 1 arvukas
Funalia trogii (Berk.) Bondartsev & Singer* Karvane kõbjas 18(7) 1 6 7 1 3 haruld.
Ganoderma applanatum (Pers.) Pat. Jänesvaabik 6(4) 1 1 1 2 1 arvukas
Gloeophyllum sepiarium (Wulfen: Fr.) P. Karst. Mustjas kõrbik 9(6) 4 3 1 1 arvukas
Gloeoporus dichrous (Fr.: Fr.) Bres. Kasetarrik 1(1) 1 sage
Hapalopilus rutilans (Pers.: Fr.) P. Karst. Kaneelpruunik 1(1) 1 tavaline
Inonotus obliquus (Pers.: Fr.) Pilat Must pässik 3(3) 1 1 1 arvukas
Ischnoderma benzoinum (Wahlenb.: Fr.) P. Karst. Harilik tõrvis 2(2) 1 1 arvukas
Junghuhnia nitida (Pers.: Fr.) Ryvarden Ookerjas nääts 4(3) 1 1 2 tavaline
Lenzites betulinus (L.: Fr.) Fr. Kasepehik 7(6) 1 1 2 1 2 tavaline
Oligoporus rennyi (Berk. & Broome) Donk Renny tümak 1(1) 1 v haruld.
Kask Haab MändLeidude (alade)
arv
Sagedus Eestis
/jätkub/

69
Lisa 2 (järg)
(1) (2) (3) (4) (5) (6) (7) (8) (9) (10) (11) (12) (13) (14) (15) (16) (17) (18) (19)
Phellinus alni (Bondartsev) Parmasto Lepataelik 1(1) 1 arvukas
P. cinereus s.l. (Niemelä) M. Fischer Hall taelik 2(1) 1 1 arvukas
P. lundellii Niemelä* Lundelli taelik 1(1) 1 sage
P. populicola Niemelä* Haava-tuletaelik 2(1) 1 1 arvukas
P. tremulae (Bondartsev) Bondartsev & Borisov Haavataelik 36(15) 5 7 9 4 11 arvukas
Physisporinus sanguinolentus (Alb.&Schwein.)Pilat* Punav lodupoorik 1(1) 1 v haruld.
Piptoporus betulinus (Bull.: Fr.) P. Karst. Kasekäsn 4(4) 1 2 1 arvukas
Polyporus badius (Pers.) Schwein. Läiktorik 1(1) 1 sage
P. varius (Pers.) Fr. Kännutorik 7(5) 3 1 1 2 arvukas
Postia alni Niemelä & Vampola Sinkjas tümak 12(7) 2 1 2 6 1 arvukas
P. lactea (Fr.) P. Karst. Valge tümak 3(3) 1 2 sage
P. leucomallella (Murrill) Jülich* Karune tümak 1(1) 1 arvukas
P. tephroleuca (Fr.: Fr.) Jülich Kuusetümak 7(6) 1 1 2 2 1 tavaline
Protomerulius caryae (Schwein.) Ryvarden* Tardpoorik 2(2) 2 sage
Pycnoporellus fulgens (Fr.) Donk* Roostetorik 2(2) 1 1 arvukas
Pycnoporus cinnabarinus (Jacq.: Fr.) P. Karst. Punapoorik 5(4) 4 1 tavaline
Rigidoporus corticola (Fr.) Pouzar Haavatarjak 12(5) 1 4 3 3 1 arvukas
R. populinus (Schumach.) Pouzar Vahtratarjak 1(1) 1 arvukas
Skeletocutis amorpha (Fr.: Fr.) Kotl. & Pouzar Punatarrik 7(5) 2 4 1 arvukas
S. biguttulata (Romell) Niemelä Tavatarrik 1(1) 1 tavaline
S. carneogrisea A. David Kõbjukitarrik 9(6) 4 4 1 tavaline
S. nivea (Jungh.) Jean Keller Piimvalge peenpoorik 1(1) 1 arvukas
Spongipellis fissilis (Berk. & M.A. Curtis) Murrill* Rasune konttümak 1(1) 1 haruld.
Spongiporus undosus (Peck) A. David Looktümak 1(1) 1 haruld.
Trametes hirsuta (Fr.: Fr.) Vill-tagel 13(12) 1 7 2 1 1 1 arvukas
T. ochracea (Pers.) Gilb. & Ryvarden Vööt-tagel 37(22) 2 4 1 1 4 10 6 5 4 arvukas
T. pubescens (Schumach.: Fr.) Pilat Pehme tagel 1(1) 1 tavaline
T. velutina (Pers.) G.H. Cunn Samettagel 3(3) 1 1 1 haruld.
T. versicolor (L.: Fr.) Pilat* Libliktagel 1(1) 1 arvukas
/jätkub/
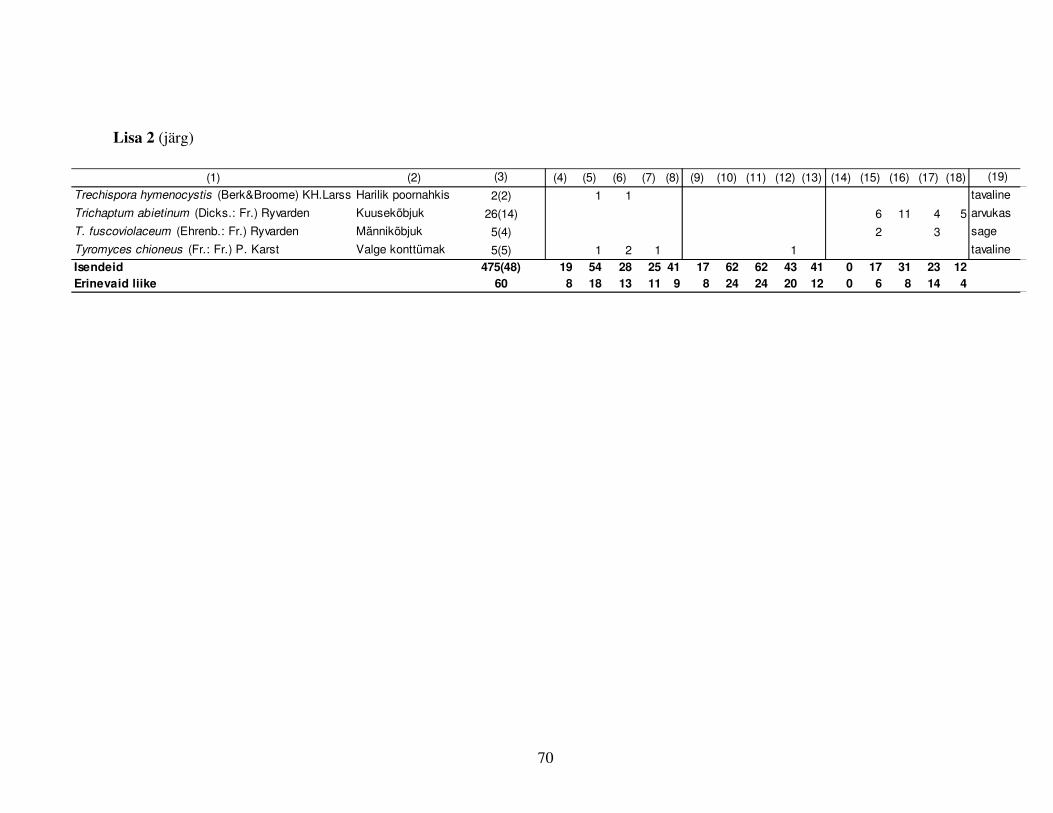
70
Lisa 2 (järg)
rg (1) (2) (3) (4) (5) (6) (7) (8) (9) (10) (11) (12) (13) (14) (15) (16) (17) (18) (19)
Trechispora hymenocystis (Berk&Broome) KH.Larss Harilik poornahkis 2(2) 1 1 tavaline
Trichaptum abietinum (Dicks.: Fr.) Ryvarden Kuusekõbjuk 26(14) 6 11 4 5 arvukas
T. fuscoviolaceum (Ehrenb.: Fr.) Ryvarden Männikõbjuk 5(4) 2 3 sage
Tyromyces chioneus (Fr.: Fr.) P. Karst Valge konttümak 5(5) 1 2 1 1 tavaline
Isendeid 475(48) 19 54 28 25 41 17 62 62 43 41 0 17 31 23 12Erinevaid liike 60 8 18 13 11 9 8 24 24 20 12 0 6 8 14 4
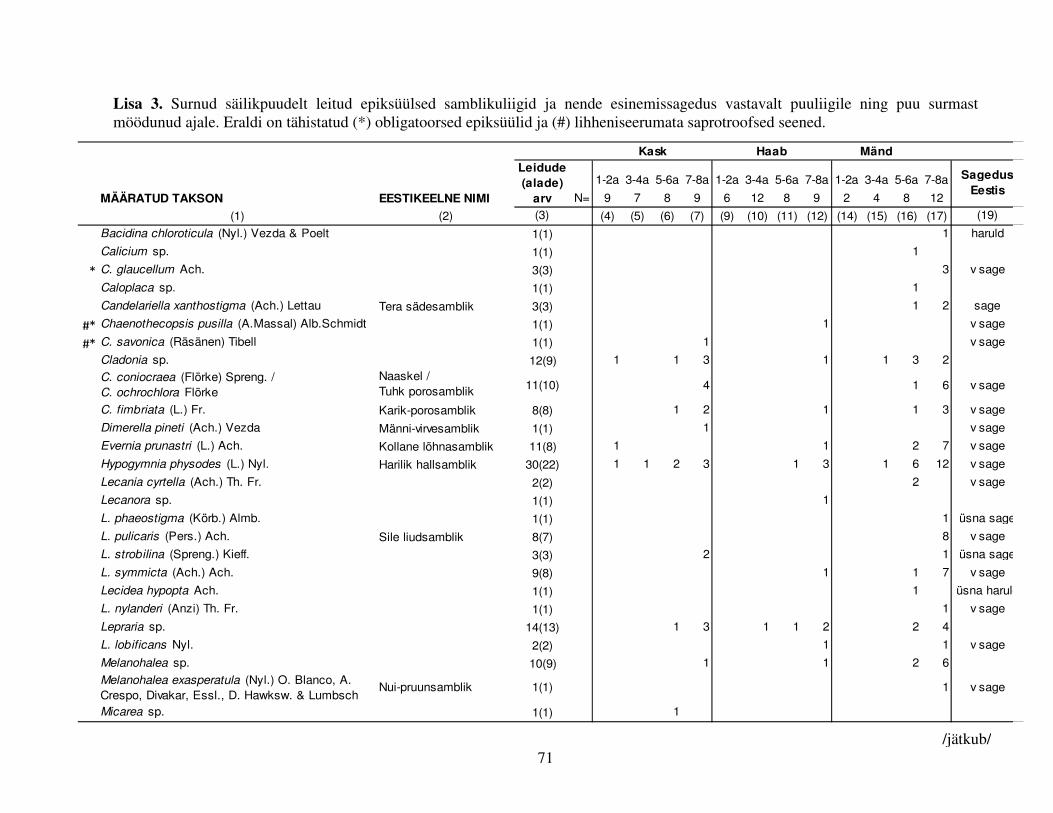
71
Lisa 3. Surnud säilikpuudelt leitud epiksüülsed samblikuliigid ja nende esinemissagedus vastavalt puuliigile ning puu surmast möödunud ajale. Eraldi on tähistatud (*) obligatoorsed epiksüülid ja (#) lihheniseerumata saprotroofsed seened.
1-2a 3-4a 5-6a 7-8a 1-2a 3-4a 5-6a 7-8a 1-2a 3-4a 5-6a 7-8a
MÄÄRATUD TAKSON EESTIKEELNE NIMI N= 9 7 8 9 6 12 8 9 2 4 8 12
(1) (2) (3) (4) (5) (6) (7) (9) (10) (11) (12) (14) (15) (16) (17) (19)
Bacidina chloroticula (Nyl.) Vezda & Poelt 1(1) 1 haruld
Calicium sp. 1(1) 1
* C. glaucellum Ach. 3(3) 3 v sage
Caloplaca sp. 1(1) 1
Candelariella xanthostigma (Ach.) Lettau Tera sädesamblik 3(3) 1 2 sage
#* Chaenothecopsis pusilla (A.Massal) Alb.Schmidt 1(1) 1 v sage
#* C. savonica (Räsänen) Tibell 1(1) 1 v sage
Cladonia sp. 12(9) 1 1 3 1 1 3 2
C. coniocraea (Flörke) Spreng. /
C. ochrochlora Flörke
Naaskel /
Tuhk porosamblik11(10) 4 1 6 v sage
C. fimbriata (L.) Fr. Karik-porosamblik 8(8) 1 2 1 1 3 v sage
Dimerella pineti (Ach.) Vezda Männi-virvesamblik 1(1) 1 v sage
Evernia prunastri (L.) Ach. Kollane lõhnasamblik 11(8) 1 1 2 7 v sage
Hypogymnia physodes (L.) Nyl. Harilik hallsamblik 30(22) 1 1 2 3 1 3 1 6 12 v sage
Lecania cyrtella (Ach.) Th. Fr. 2(2) 2 v sage
Lecanora sp. 1(1) 1
L. phaeostigma (Körb.) Almb. 1(1) 1 üsna sage
L. pulicaris (Pers.) Ach. Sile liudsamblik 8(7) 8 v sage
L. strobilina (Spreng.) Kieff. 3(3) 2 1 üsna sage
L. symmicta (Ach.) Ach. 9(8) 1 1 7 v sage
Lecidea hypopta Ach. 1(1) 1 üsna haruld
L. nylanderi (Anzi) Th. Fr. 1(1) 1 v sage
Lepraria sp. 14(13) 1 3 1 1 2 2 4
L. lobificans Nyl. 2(2) 1 1 v sage
Melanohalea sp. 10(9) 1 1 2 6
Melanohalea exasperatula (Nyl.) O. Blanco, A.
Crespo, Divakar, Essl., D. Hawksw. & LumbschNui-pruunsamblik 1(1) 1 v sage
Micarea sp. 1(1) 1
Leidude (alade)
arv
Sagedus Eestis
Kask Haab Mänd
/jätkub/
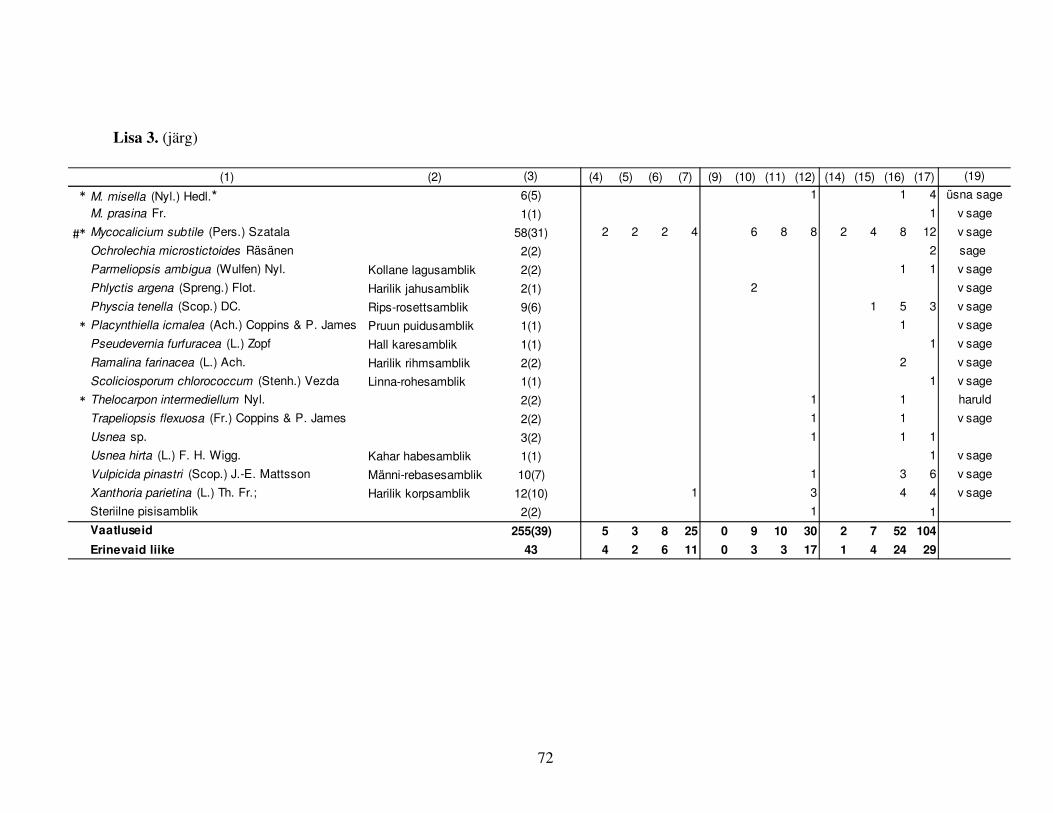
72
Lisa 3. (järg)
(1) (2) (3) (4) (5) (6) (7) (9) (10) (11) (12) (14) (15) (16) (17) (19)
* M. misella (Nyl.) Hedl.* 6(5) 1 1 4 üsna sage
M. prasina Fr. 1(1) 1 v sage
#* Mycocalicium subtile (Pers.) Szatala 58(31) 2 2 2 4 6 8 8 2 4 8 12 v sage
Ochrolechia microstictoides Räsänen 2(2) 2 sage
Parmeliopsis ambigua (Wulfen) Nyl. Kollane lagusamblik 2(2) 1 1 v sage
Phlyctis argena (Spreng.) Flot. Harilik jahusamblik 2(1) 2 v sage
Physcia tenella (Scop.) DC. Rips-rosettsamblik 9(6) 1 5 3 v sage
* Placynthiella icmalea (Ach.) Coppins & P. James Pruun puidusamblik 1(1) 1 v sage
Pseudevernia furfuracea (L.) Zopf Hall karesamblik 1(1) 1 v sage
Ramalina farinacea (L.) Ach. Harilik rihmsamblik 2(2) 2 v sage
Scoliciosporum chlorococcum (Stenh.) Vezda Linna-rohesamblik 1(1) 1 v sage
* Thelocarpon intermediellum Nyl. 2(2) 1 1 haruld
Trapeliopsis flexuosa (Fr.) Coppins & P. James 2(2) 1 1 v sage
Usnea sp. 3(2) 1 1 1
Usnea hirta (L.) F. H. Wigg. Kahar habesamblik 1(1) 1 v sage
Vulpicida pinastri (Scop.) J.-E. Mattsson Männi-rebasesamblik 10(7) 1 3 6 v sage
Xanthoria parietina (L.) Th. Fr.; Harilik korpsamblik 12(10) 1 3 4 4 v sage
Steriilne pisisamblik 2(2) 1 1
Vaatluseid 255(39) 5 3 8 25 0 9 10 30 2 7 52 104
Erinevaid liike 43 4 2 6 11 0 3 3 17 1 4 24 29
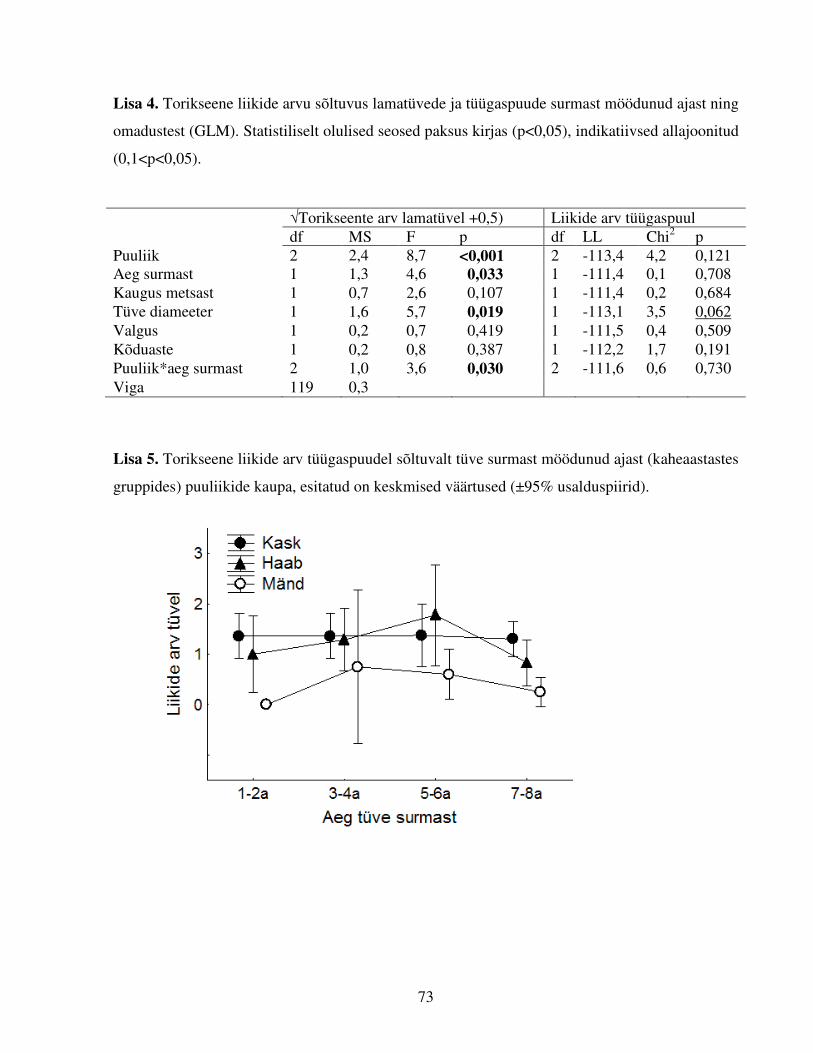
73
Lisa 4. Torikseene liikide arvu sõltuvus lamatüvede ja tüügaspuude surmast möödunud ajast ning
omadustest (GLM). Statistiliselt olulised seosed paksus kirjas (p<0,05), indikatiivsed allajoonitud
(0,1<p<0,05).
√Torikseente arv lamatüvel +0,5) Liikide arv tüügaspuul df MS F p df LL Chi2 p Puuliik 2 2,4 8,7 <0,001 2 -113,4 4,2 0,121 Aeg surmast 1 1,3 4,6 0,033 1 -111,4 0,1 0,708 Kaugus metsast 1 0,7 2,6 0,107 1 -111,4 0,2 0,684 Tüve diameeter 1 1,6 5,7 0,019 1 -113,1 3,5 0,062 Valgus 1 0,2 0,7 0,419 1 -111,5 0,4 0,509 Kõduaste 1 0,2 0,8 0,387 1 -112,2 1,7 0,191 Puuliik*aeg surmast 2 1,0 3,6 0,030 2 -111,6 0,6 0,730 Viga 119 0,3
Lisa 5. Torikseene liikide arv tüügaspuudel sõltuvalt tüve surmast möödunud ajast (kaheaastastes
gruppides) puuliikide kaupa, esitatud on keskmised väärtused (±95% usalduspiirid).
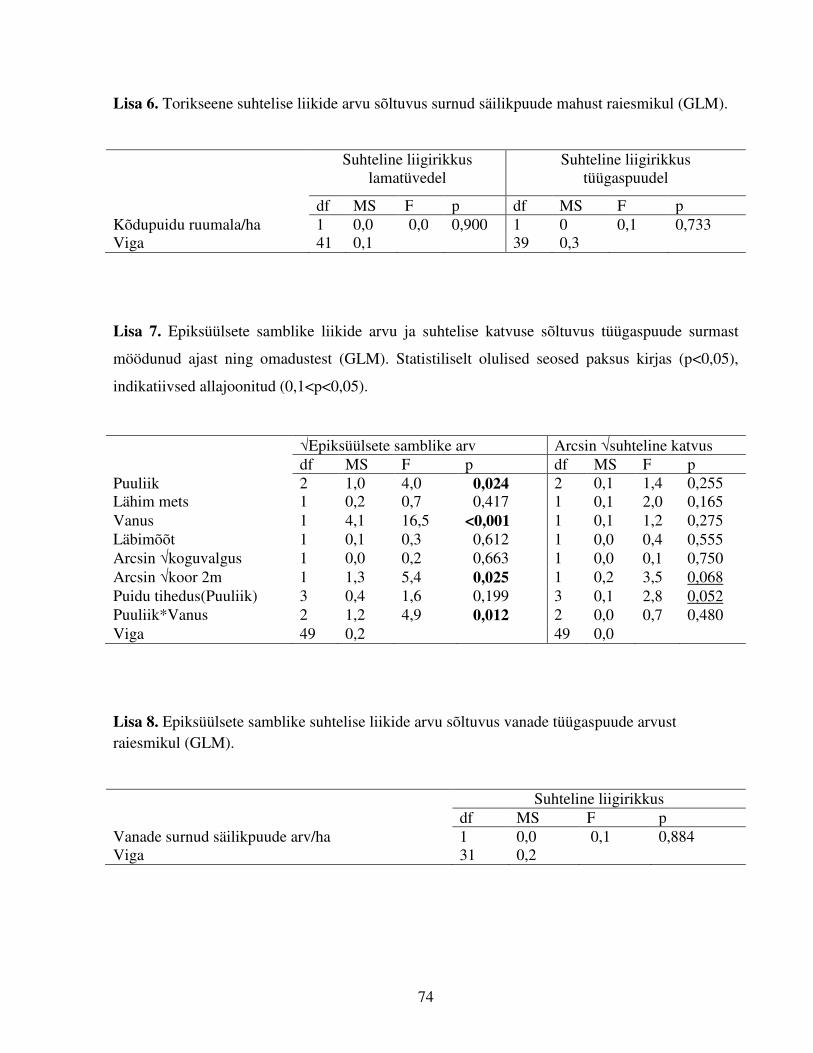
74
Lisa 6. Torikseene suhtelise liikide arvu sõltuvus surnud säilikpuude mahust raiesmikul (GLM).
Suhteline liigirikkus lamatüvedel
Suhteline liigirikkus tüügaspuudel
df MS F p df MS F p Kõdupuidu ruumala/ha 1 0,0 0,0 0,900 1 0 0,1 0,733 Viga 41 0,1 39 0,3
Lisa 7. Epiksüülsete samblike liikide arvu ja suhtelise katvuse sõltuvus tüügaspuude surmast
möödunud ajast ning omadustest (GLM). Statistiliselt olulised seosed paksus kirjas (p<0,05),
indikatiivsed allajoonitud (0,1<p<0,05).
√Epiksüülsete samblike arv Arcsin √suhteline katvus df MS F p df MS F p Puuliik 2 1,0 4,0 0,024 2 0,1 1,4 0,255 Lähim mets 1 0,2 0,7 0,417 1 0,1 2,0 0,165 Vanus 1 4,1 16,5 <0,001 1 0,1 1,2 0,275 Läbimõõt 1 0,1 0,3 0,612 1 0,0 0,4 0,555 Arcsin √koguvalgus 1 0,0 0,2 0,663 1 0,0 0,1 0,750 Arcsin √koor 2m 1 1,3 5,4 0,025 1 0,2 3,5 0,068 Puidu tihedus(Puuliik) 3 0,4 1,6 0,199 3 0,1 2,8 0,052 Puuliik*Vanus 2 1,2 4,9 0,012 2 0,0 0,7 0,480 Viga 49 0,2 49 0,0
Lisa 8. Epiksüülsete samblike suhtelise liikide arvu sõltuvus vanade tüügaspuude arvust raiesmikul (GLM).
Suhteline liigirikkus df MS F p Vanade surnud säilikpuude arv/ha 1 0,0 0,1 0,884 Viga 31 0,2

75





























