Embed Size (px)
Citation preview

Universidad de Costa Rica
Escuela de Tecnología de Alimentos
Trabajo final de graduación presentado a la Escuela de Tecnología de Alimentos para optar
por el título de Licenciatura en Ingeniería de Alimentos
Caracterización de bacterias lácticas en subproductos industriales de piña (Anana
cosmosus L. Merr) y determinación de su capacidad de potencial probiótico en jugo de piña
Elaborado por:
JANNETTE WEN FANG WU WU
Carné: B06978
Ciudad Universitaria Rodrigo Facio,
Agosto, 2016

ii
DEDICATORIA
Para mi papá, mi mamá y mi hermana
Por apoyarme siempre y estar ahí en todo momento de necesidad-
Y a todos los que me apoyaron para escribir y concluir esta tesis,
pues es a ellos a quienes se las debo por su apoyo incondicional.

TRIBUNAL EXAMINADOR
Proyecto de graduacic'Jn presentado a Ia Lscuela de Tecnologia de i\limcntos como requisito
parcial para optar por el grado de l.icenciatura en lngenicria de ;\limcntos .
J:lahorado por:
.!annette Wen hmg Wu '0- 'u
/\probado por:
l'v1.Sc. Carmela Vel{vque/. Carrillo
Presidenta del Tribunal
M.Sc . Natalia l3arb0/.a V~1rgas
Directora del Provecto
/\sesora del Proyecto
Profcsor Designado
Ill

iv
AGRADECIMIENTOS
En primer lugar, agradezco a la Universidad de Costa Rica, la institución en la que
tuve el privilegio de aprender, a la Escuela de Tecnología de Alimentos y su personal, por
velar por el bienestar de sus estudiantes, al Centro Nacional de Ciencia y Tecnología de
Alimentos (CITA) y al Centro de Investigaciones en Biología Molecular y Celular (CIBCM),
por su contribución y apoyo durante el desarrollo del proyecto.
A mi grupo de asesores: Natalia Barboza Vargas, mi directora de tesis, que me
permitió aprender de su amplia experiencia, por apoyarme y aconsejarme durante el
desarrollo del proyecto, que me brindó la oportunidad de ser asistente y participar en una
investigación. A la profesora Jessie Usaga y al profesor Eduardo Thompson, por su guía y
consejos a lo largo de todo el trayecto y su comprensión, apoyo y atención.
Al personal de los laboratorios de química y microbiología del CITA, en especial a
Vanny Mora, Geovanni González y a Luis Morales por estar siempre dispuestos a ayudar y
sus palabras de motivación y guía. A los miembros del Laboratorio 6 del Centro de
Investigación en Biología Celular y Molecular, por aceptarme como un miembro más de la
familia y por guía en el aprendizaje.
A mis compañeros y amigos. A Marce por su compañía y apoyo en las largas jornadas
del laboratorio de microbiología, a Henry por su ayuda en los momentos de necesidad y las
charlas divertidas en el laboratorio, y a Malavasi por sus palabras de motivación durante el
largo periodo de redacción.

v
ÍNDICE GENERAL
DEDICATORIA ................................................................................................................................. ii
TRIBUNAL EXAMINADOR ........................................................................................................... iii
AGRADECIMIENTOS ..................................................................................................................... iv
ÍNDICE DE FIGURAS .................................................................................................................... viii
ÍNDICE DE CUADROS .................................................................................................................... xi
ÍNDICE DE ABREVIATURAS ....................................................................................................... xii
RESUMEN....................................................................................................................................... xiv
I. JUSTIFICACIÓN ............................................................................................................................ 1
II. OBJETIVOS ................................................................................................................................... 4
2.1. Objetivo general ....................................................................................................................... 4
2.2. Objetivo específico .................................................................................................................. 4
III. MARCO TEÓRICO ...................................................................................................................... 5
3.1. Piña y su importancia productiva en Costa Rica ...................................................................... 5
3.1.1. Características generales ................................................................................................... 5
3.1.2. Producción e industrialización del fruto en Costa Rica .................................................... 5
3.1.3. Manejo y aprovechamiento de subproductos industriales de piña .................................... 6
3.2. Bacterias lácticas (BAL) .......................................................................................................... 8
3.2.1. Antecedente histórico de las bacterias lácticas.................................................................. 8
3.2.2. Clasificación de bacterias lácticas ..................................................................................... 9
3.2.3. Metabolismo de bacterias lácticas ..................................................................................... 9
3.3. Perfil del género Lactobacillus spp. ....................................................................................... 10
3.3.1. Características del género Lactobacillus ......................................................................... 10
3.3.2. Heterogeneidad genética de Lactobacillus spp. .............................................................. 11
3.3.3. Lactobacillus fermentum ................................................................................................. 11
3.3.4. Caracterización genómica de Lactobacillus fermentum ................................................. 12
3.4. Probióticos ............................................................................................................................. 13
3.4.1. Definición y características de un probiótico .................................................................. 13
3.4.2. Obtención y almacenamiento de cepas probióticas ......................................................... 15
3.4.3. Criterios de evaluación de la capacidad probiótica ......................................................... 15
3.4.3.1. Pruebas in vitro ............................................................................................................ 15
3. 4. 3. 1. 1. Estabilidad al ácido y a las sales biliares ............................................................... 16

vi
3.4.3.1.2. Adhesión al epitelio intestinal ................................................................................... 17
3.4.3.2. Pruebas in vivo ............................................................................................................. 17
3.5. Aplicación de probióticos en alimentos ................................................................................. 18
3.5.1. Aplicación de probióticos en productos lácteos .............................................................. 18
3.5.2. Aplicación de probióticos en productos no lácteos: jugos .............................................. 18
3.5.2.1. Limitaciones de la sobrevivencia de probióticos en jugos de fruta .............................. 19
3.5.2.2. Mejoramiento de la sobrevivencia de probióticos en jugos ......................................... 20
3.5.2.2.1. Fortificación con prebióticos ..................................................................................... 20
3.5.2.2.2. Adaptación e inducción de resistencia ...................................................................... 21
3.5.2.2.3. Refrigeración y adición de antioxidantes .................................................................. 21
3.5.2.2.4. Microencapsulación .................................................................................................. 21
3.5.3. Características sensoriales de los jugos con probióticos ................................................. 21
3.6. Técnicas de biología molecular para la identificación de bacterias ....................................... 22
3.6.1. Conceptos básicos de las técnicas de biología molecular ............................................... 22
3.6.2. Gel de electroforesis ........................................................................................................ 23
3.6.3. Reacción en cadena de la polimerasa (PCR) ................................................................... 24
3.6.3.1. Aplicaciones del PCR .................................................................................................. 26
3.6.4. Identificación de bacterias ............................................................................................... 26
3.6.5. Identificación por medio del gen del ADNr 16S ............................................................. 27
3.6.6. Ventajas y limitaciones de la técnica de PCR ................................................................. 28
IV.MATERIALES Y MÉTODOS .................................................................................................... 29
4.1. Localización del proyecto ...................................................................................................... 29
4.2. Materias primas ...................................................................................................................... 29
4.3. Aislamiento de los microorganismos ..................................................................................... 29
4.4. Pruebas preliminares: ............................................................................................................. 30
4.5. Identificación molecular de las BAL ..................................................................................... 31
4.5.1. Extracción de ADN ......................................................................................................... 31
4.3.3. Reacción en cadena de la polimerasa (PCR) ................................................................... 32
4.3.4. Análisis filogenético ........................................................................................................ 32
4.5. Caracterización físicoquímica de los jugos de piña inicial y luego de almacenamiento ........ 33
4.6. Ensayo de capacidad de sobrevivencia de una cepa BAL seleccionada ................................ 34
4.6.1. Selección de la bacteria ................................................................................................... 34
4.6.2. Evaluación de la supervivencia de Lactobacilus fermentum en jugo de piña ................. 35

vii
4.6.3. Determinación del conteo de BAL por recuento en placa ............................................... 36
4.7. Análisis estadístico ................................................................................................................. 36
V. RESULTADOS Y DISCUSIÓN .................................................................................................. 38
5.1. Pruebas preliminares de identificación molecular ................................................................. 38
5.1.1. Extracción de ácidos nucleicos totales ...................................................................... 38
5.1.2. Estandarización del protocolo de PCR ............................................................................ 42
5.2. Identificación molecular de las BAL utilizando el ADNr 16 ................................................. 44
5.3. Sobrevivencia de BAL a un pH de 2,0 ................................................................................... 49
5.4. Caracterización fisicoquímica de los jugos de piña durante el almacenamiento ....................... 51
5.5. Evaluación de la sobrevivencia de la cepa de L. fermentum en jugos de piña durante su
almacenamiento a 4 °C. .................................................................................................................... 55
VI. CONCLUSIONES ...................................................................................................................... 60
VII. RECOMENDACIONES ........................................................................................................... 61
VIII. BIBLIOGRAFÍA ...................................................................................................................... 62

viii
ÍNDICE DE FIGURAS
Figura 1. Diferentes rutas metabólicas de las hexosas empleadas por bacterias lácticas
(Kandler, 1983)………………………………………………………………………......10
Figura 2. Diagrama de los ciclos de amplificación del material nucleico por reacción en
cadena de la polimerasa (PCR). …………………………………………………………25
Figura 3. Diagrama de distribución de genes bacterianos………………………………27
Figura 4. (a) ADN de dos extracciones realizadas por duplicado obtenidas utilizando el
protocolo de Fontecha a partir de E. coli no patógena y marcador molecular 100 pb
(Thermo scientific); (b) Productos de dos extracciones de ADN realizadas por duplicado y
obtenidas según el procedimiento de Minipreps, utilizando E. coli no patógena, marcador
molecular 100pb (Thermo Scientific). Geles de agarosa al 1 %, teñidos con azul de carga
con Gel-Red (10 000 X) (Biotium), corridos a 110 V, por 40
minutos………………………………………………………………………………….39
Figura 5. (a) Productos de extracción de ADN obtenidos por protocolo de Fontecha, orden
del gel: KX218443, KX218443, KX218444, KX218444, KX218445, KX218445,
KX218846, KX218846, marcador molecular 100 pb (Thermo Scientific); (b) Productos de
extracción de ADN obtenidos siguiendo el procedimiento de Minipreps, orden del gel:
KX218443, KX218444, KX218445 y KX218446 por duplicado, marcador molecular
100pb (Thermo Scientific). Geles de agarosa al 1 %, teñidos con azul de carga con Gel-
Red (10 000 X) (Biotium), corridos a 110 V, por 40
minutos…………………………………………………………………………………..40
Figura 6. Productos de PCR obtenidos utilizando los iniciadores 27F y 1429R (a)
amplificaciones obtenidas con las extracciones según Fontecha. Orden del gel: marcador
de peso molecular 100 pb (ThermoScientific), KX218443, KX218443, KX218444,
KX218444, blanco, KX218445, KX218445, KX218846, KX218846. (b) Amplificaciones
obtenidas siguiendo el protocolo de Minipreps, orden del gel: KX218443, KX218443,
KX218444, KX218444, KX218445, KX218445, blanco, KX218846, KX218846,
marcador molecular 100 pb (Thermo Scientific). Geles de agarosa al 1 %, teñidos con Gel-

ix
Red (10 000 X) (Biotium), corridos a 110 V, por 40
minutos…………………………………………………………………………………..41
Figura 7. (a) Productos de PCR amplificados con anillamiento de 45 °C, orden del gel:
marcador molecular de 100 pb (Thermo Scientific), KX218443, KX218443, KX218444,
KX218444, KX218445, KX218445, KX218846, KX218846. (b) productos de PCR
amplificados con un anillamiento de 55 °C: KX218443, KX218443, KX218444,
KX218444, KX218445, KX218445, KX218846, KX218846, marcador molecular 100 pb
(Thermo scientific). Geles de agarosa al 1 %, teñidos con azul de carga con Gel-Red
(Biotium) (10 000 X) corridos a 110 V, 40 minutos……………………………………...43
Figura 8. Árbol filogénico para las cepas: KX218445, KX218448, KX218452,
KX218443, KX218444, KX218450, KX218449, KX218453, KX218451, KX218446,
KX218447, KX218454. Los árboles filogenéticos se elaboraron utilizando el programa
Molecular Evolutionary Genetics Analysis (MEGA) versión 3.1 (Kumar et al., 2004). La
robustez de las ramas internas se estimó por medio de análisis de “bootstrap” con 2000
réplicas. Los análisis filogenéticos fueron comparados con los métodos de distancia, el
vecino más cercano (Neighbor Joining), donde las secuencias de este estudio se denotan
con otro color, y solamente se muestran los valores de las ramas mayores al 50 %.............46
Figura 9. Porcentaje de identidad resultantes de la comparación de pares alineados de las
cepas analizadas con individuos de su mismo género y cepas de referencia utilizando un
alineamiento en Muscle y el programa SDT vrsn 1.2 (http://web.cbio.uct.ac.za/SDT;
Muhire et al., 2014)...…………………………………………………………………….48
Figura 10. Pruebas de tolerancia al jugo gástrico por simulación, utilizando medio MRS,
acidificado (pH=2,0) con HCl 1N de las diferentes cepas; las barras de error corresponden
a la desviación estándar (n = 3). Las muestras que no comparten una letra en común
presentan diferencias significativas según la comparación múltiple de
Tuckey…………………………….……………………………………………………..50
Figura 11 Evolución de la población de L. fermentum (a) y variación del pH (b) en tres
jugos de piña comerciales almacenados a 4 °C, durante seis semanas. Las barras de error
representan la desviación estándar con n = 3.

x
*Valores que no comparten la misma letra presentan diferencias significativas (p < 0,05)
basado en el análisis de comparación múltiple por Tuckey. ** El valor Δ corresponde la
variación entre el valor a 0 semanas y el valor a las 6
semanas…………………………………………………..………………………………57

xi
ÍNDICE DE CUADROS
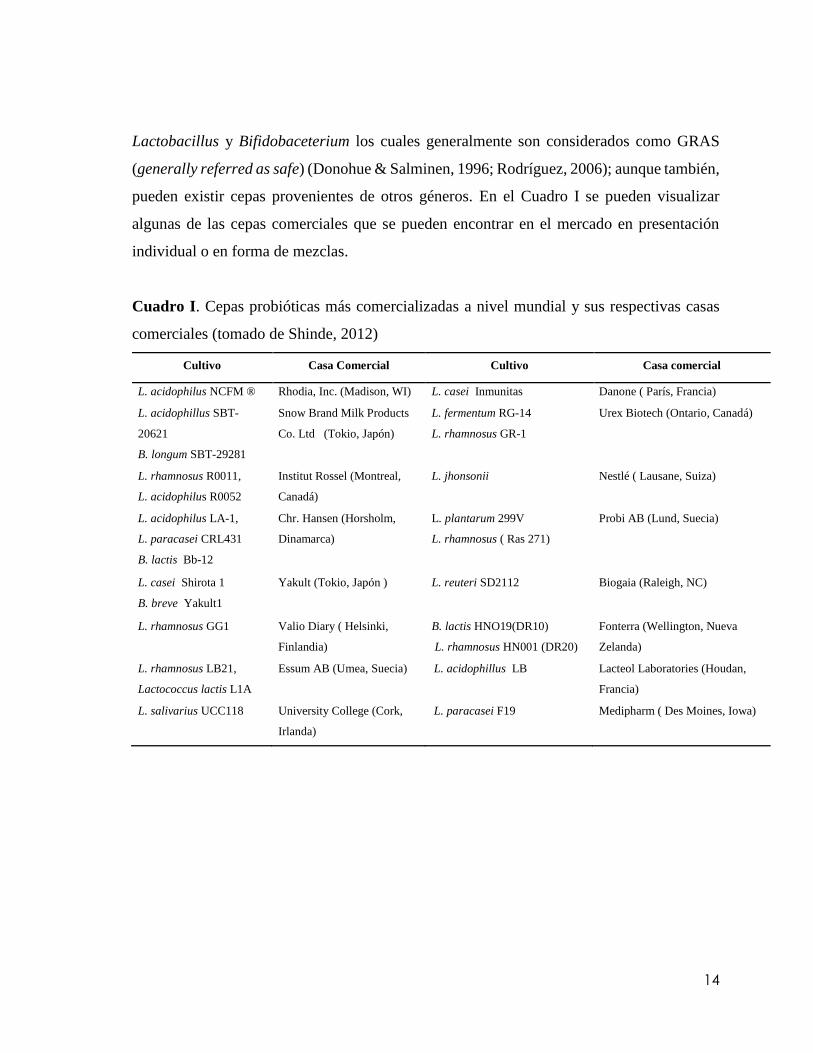
Cuadro I. Cepas probióticas más comercializadas a nivel mundial y sus respectivas casas
comerciales (tomado de Shinde,
2012) …………………………………………………………………………………….14
Cuadro II. Comparación entre el promedio de la concentración del ADN, pureza obtenida
a A260/A280, según las extracciones obtenidas a partir de los protocolos de Fontecha
(2005) y Miniprep (Birnboim & Dolly, 1979) (n = 2)………………………………….41
Cuadro III. Propiedades fisicoquímicas de tres jugos de piña: pulpa, pulpa + cáscara
(80:20) y cáscara evaluados durante su almacenamiento por seis semanas en refrigeración
(4 °C) tras la inoculación con L. fermentum……………………………………………53
xii
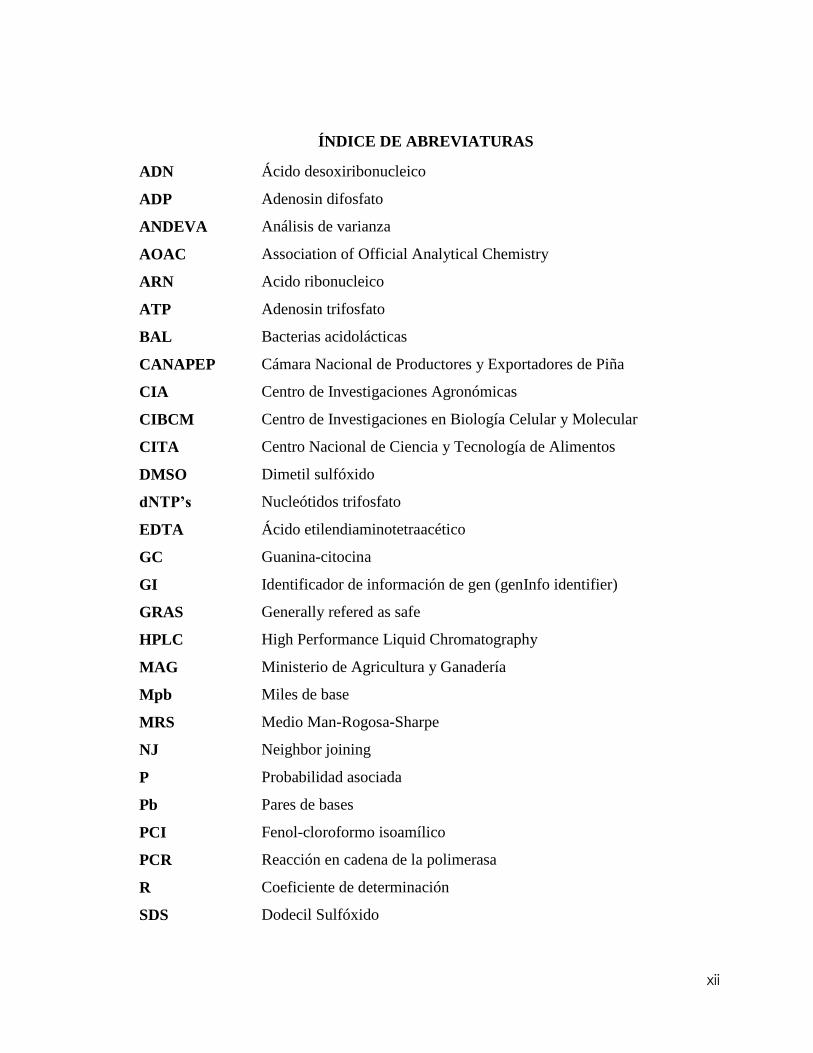
ÍNDICE DE ABREVIATURAS
ADN Ácido desoxiribonucleico
ADP Adenosin difosfato
ANDEVA Análisis de varianza
AOAC Association of Official Analytical Chemistry
ARN Acido ribonucleico
ATP Adenosin trifosfato
BAL Bacterias acidolácticas
CANAPEP Cámara Nacional de Productores y Exportadores de Piña
CIA Centro de Investigaciones Agronómicas
CIBCM Centro de Investigaciones en Biología Celular y Molecular
CITA Centro Nacional de Ciencia y Tecnología de Alimentos
DMSO Dimetil sulfóxido
dNTP’s Nucleótidos trifosfato
EDTA Ácido etilendiaminotetraacético
GC Guanina-citocina
GI Identificador de información de gen (genInfo identifier)
GRAS Generally refered as safe
HPLC High Performance Liquid Chromatography
MAG Ministerio de Agricultura y Ganadería
Mpb Miles de base
MRS Medio Man-Rogosa-Sharpe
NJ Neighbor joining
P Probabilidad asociada
Pb Pares de bases
PCI Fenol-cloroformo isoamílico
PCR Reacción en cadena de la polimerasa
R Coeficiente de determinación
SDS Dodecil Sulfóxido

xiii
ST Sólidos totales
TAE Buffer Tris/Acetato/EDTA
TBE Buffer Tris/ Borato/EDTA
UFC Unidades Formadoras de colonias
UPGMA Unweighted Pair Group Method with Arithmetic Mean

xiv
RESUMEN
Se identificaron, con marcadores moleculares, cepas de bacterias aisladas de silos
fermentados elaborados a partir de subproductos industriales de piña (cáscaras y coronas) por
medio de la amplificación del gen ADNr 16S utilizando los iniciadores 27F y 1429R. Se
registró un total de tres géneros diferentes y doce bacterias lácticas (BAL). Las cepas
identificadas como Lactobacillus spp. (códigos del GenBank: KX218454, KX218452,
KX218453, KX218451, KX218450 KX218449, KX218448) agrupan con Lactobacillus
casei y L. paracasei. La cepa (código del Genbank: KX218446) se agrupó como miembro
del género Weissella fabaria y Weissella ghanensis. Mientras que la cepa GenBank no.
KX218445 agrupa con Fructobacillus troapeoli. Dos cepas (código del GenBank:
KX218443 y KX218444) agruparon con Lactobacillus fermentum. Mientras que la cepa
código del GenBank: KX218447 agrupó con Lactobacillus spicheri. Los iniciadores
utilizados no fueron suficientemente específicos para diferenciar especies de Lactobacillus
spp. que agruparon con L. casei y L. paracasei, lo mismo se observó con los aislamientos
que agruparon con W. fabaria y W. ghanensis. Las BAL identificadas fueron sometidas a
pruebas de tolerancia a un pH de 2,0 para determinar su capacidad de sobrevivencia en el
jugo gástrico de forma in vitro. Cinco de las cepas identificadas mostraron una sobrevivencia
mayor al 90 %, de entre ellas se seleccionó L. fermentum (código del GenBank: KX218443),
que mostró una sobrevivencia del 92,2 % + 0,6 como sujeto de estudio.
La bacteria fue inoculada a una carga aproximada de 108 UFC/mL, en tres tipos de
jugos de piña: pulpa, cáscara y una mezcla de pulpa: cáscara 80:20. Los mismos se
almacenaron a 4 °C durante seis semanas. Los jugos fueron previamente analizados
fisicoquímicamente considerando los parámetros de contenido de azúcares fermentables y
ácidos orgánicos por HPLC, sólidos totales, pH, sólidos solubles y acidez titulable expresada
como ácido cítrico de los jugos en el tiempo 0 y a las 6 semanas de almacenamiento. Los
resultados mostraron interacciones significativas entre el tiempo de muestreo y el tipo de
jugo en cuanto al contenido de sólidos solubles (p < 0,0006), acidez titulable total (p <

xv
0,0001), fructosa (p < 0,0001), glucosa (p < 0,004), sacarosa (p < 0,0001), ácido cítrico (p <
0,0496), ácido málico (p < 0,0001) y ácido láctico (p < 0,0001). El contenido de sólidos
totales no mostró interacción significativa con respecto al tipo de jugo y el tiempo de
muestreo (p > 0,05).
El estudio de la sobrevivencia de la bacteria mostró interacciones significativas (p <
0,0008) entre el tipo de jugo y el tiempo de muestreo sobre el recuento de las bacterias y se
determinó que el jugo de piña elaborado a partir de pulpa de piña presentó las mejores
condiciones para la sobrevivencia de L. fermentum (5,99 LogUFC/mL). Para el caso del jugo
de cáscara y el de pulpa + cáscara, estos no presentaron diferencias significativas en los
recuentos durante el almacenamiento al ser analizados por comparación de medias de Tuckey.
Los resultados del estudio aportan información valiosa sobre el comportamiento de
la cepa de L. fermentum (código del GenBank: KX218443) aislada de los ensilajes de
subproductos de piña y permiten plantear posibles aplicaciones de la bacteria como un
probiótico en bebidas no lácteas.

1
I. JUSTIFICACIÓN
El mercado mundial de jugos de frutas ha mostrado un importante crecimiento desde
el año 2013 con un aumento de aproximadamente un 1 % de valor y 3 % de volumen (AIJN,
2014). De acuerdo con la Euromonitor (2014), el mercado mundial de jugos, néctares y
bebidas está valorado por cerca de $149 billones con un volumen estimado de 76,5 billones
de litros. Esta tendencia se ha observado principalmente en países en desarrollo donde el
consumo de bebidas carbonatadas ha visto una disminución importante.
Por otra parte, las nuevas tendencias de salud y bienestar han generado un cambio en
los hábitos de consumo aumentando el interés en lo natural y propiedades funcionales
benéficas tales como características antioxidantes o el uso de probióticos (Mintel, 2015).
Los microorganismos probióticos han sido agregados a una variedad de productos
sobre todo de matrices lácteas. Sin embargo, se propone la posibilidad de emplear jugos
frutales como un vehículo para estos microorganismos debido al creciente aumento de
consumidores con respuestas adversas a componentes específicos de las matrices lácteas tales
como reacciones de hipersensibilidad, trastornos metabólicos o bien tendencias de
alimentación (Mintel, 2015); por otro lado, los jugos se han asociado a un sinfín de
propiedades beneficiosas para la salud, y representan una matriz rica en micronutrientes
esenciales (Corbo et al., 2014). Dado a lo anterior, la adición de estos microorganismos a
jugos es un reto y un objetivo deseable ya que permitiría obtener productos con el efecto
nutricional y el valor agregado que otorga la presencia de un probiótico.
Dado el alto consumo de piña y derivados a nivel nacional y mundial (AIJN, 2014),
la adición de probióticos al jugo de esta fruta resulta una muy buena alternativa para
desarrollar bebidas no lácteas con estos microorganismos, ya que para el 2003 un 20 % de la
exportación mundial y un 23 % de la importación mundial de la fruta se efectuó en forma de
jugo (Villegas et al., 2007). Paralelo a esto, el aumento en el consumo de piña conlleva a un
aumento de la producción y esto a su vez está asociado al incremento en generación de
desechos, lo cual se ha convertido en un tema controversial en la actualidad. Por este motivo,
se han desarrollado propuestas para el aprovechamiento alternativo de los subproductos que
usualmente se emplean para la elaboración de silos fermentados destinados a la alimentación

2
animal (Gutiérrez et al., 2003) los cuales son ricos en una alta variedad de microorganismos
que podrían tener un potencial probiótico importante, además de que, debido a las
condiciones usuales en las que se encuentran dentro de ese medio, pueden presentar
características adaptativas que las vuelvan más resistentes a las etapas del procesamiento
industrializado (Perricone et al., 2015). El alto consumo de la fruta lo vuelve un producto
importante para la aplicación del presente estudio; sin embargo, no garantiza que esta matriz
sea la más óptima como un vector de probióticos por lo que resulta interesante estudiar el
comportamiento de una cepa aislada de un entorno “salvaje”, como lo son los silos de
alimentación animal, y determinar su posible aplicabilidad como un probiótico.
En la actualidad, existen diversas opciones de bacterias probióticas que se
comercializan en forma de mezclas sinérgicas debido a los diferentes efectos benéficos que
pueden presentarse según la cepa utilizada; sin embargo, existen otras cepas, aún no
estudiadas, que pueden tener un potencial importante para la industria debido las
características que presentan (Kelanne, 2012; Reid et al., 2003). Dada la gran variabilidad de
microorganismos existentes en la naturaleza, la posibilidad de encontrar nuevas cepas con un
potencial mejorado es muy alta. La microbiota presente en los silos fermentados de
subproductos de piña es rica en bacterias lácticas (Tobía et al., 2003), las cuales, como grupo,
se han asociado previamente a efectos beneficiosos para la salud (Rodríguez, 2006). Sumado
a esto, cabe la posibilidad de que dadas las condiciones en las que se desarrollan, puedan
mostrar características prometedoras como alta tolerancia al ácido en comparación con otras
cepas comerciales aisladas de otras matrices. A su vez, es deseable que estos
microorganismos presenten una serie de características fisiológicas que aseguren su
sobrevivencia a través del tracto digestivo, por lo que es de vital importancia evaluar su
respuesta ante el efecto de pH ácido, presencia de sales biliares, enzimas gástricas,
adhesividad intestinal, entre otras (Gueimonde et al., 2006).
Conforme los estudios en probióticos continúan avanzando, el conocimiento sobre la
aplicación de los beneficios de los microorganismos en jugos específicos, así como las cepas
más aptas para sobrevivir en ellos irá aumentando y generando nuevos productos que tendrán
el potencial de mejorar la calidad de vida de los consumidores. Se estima que cerca de un
75 % de la población mundial pierde su enzima lactasa después del destete (Hertzler et al.,

3
1996) y cerca del 70 % de afroamericanos, 90 % de los asiáticos y 53 % de hispanos presenta
intolerancia a la lactosa (Lomer, 2006). El incremento de la población sensible a productos
derivados de la leche ha generado la necesidad de buscar nuevas alternativas que se puedan
emplear como vehículos para microorganismos probióticos y debido a su amplio consumo,
los jugos de frutas representan una herramienta prometedora. Sin embargo, existen
limitaciones que podrían afectar su uso a nivel industrial como lo es la sobrevivencia de la
bacteria durante el almacenamiento y el impacto de ésta sobre las características sensoriales
y la aceptación por el consumidor (Perricone et al., 2015) que pueden comprometer el éxito
comercial de este tipo de productos (Granato et al., 2010); por lo tanto el estudio del
comportamiento de la bacteria en la matriz y de las variaciones de su composición
fisicoquímica después del almacenamiento es un recurso importante.
El uso de técnicas moleculares como medio de identificación de bacterias se ha vuelto
una técnica práctica y rápida que permite identificar de forma precisa y con una mayor
especificidad los microorganismos presentes en una matriz, en comparación con los métodos
fenotípicos convencionales, ya que se obtiene un perfil genético de las distintas cepas según
el fragmento del gen seleccionado; esto ayuda a mitigar las limitaciones que presentan los
métodos de identificación convencionales.
Debido a lo expuesto con anterioridad, es que en el presente proyecto se identificaron
bacterias aisladas de silos fermentados elaborados a partir de subproductos industriales de
piña utilizando técnicas moleculares, además se estudió el potencial probiótico que estás
podrían tener en matrices de pH ácido como el jugo de piña.

4
II. OBJETIVOS
2.1. Objetivo general
Identificar bacterias ácido lácticas presentes en silos, elaborados con subproductos
industriales de piña y evaluar el efecto de la adición de una cepa potencialmente probiótica
seleccionada (Lactobacillus fermentum) en jugos de piña durante su almacenamiento en
refrigeración para valorar la posible aplicación del microorganismo a nivel industrial.
2.2. Objetivo específico
1. Identificar bacterias lácticas (BAL) presentes en silos para
alimentación animal, elaborados con subproductos industriales de piña, utilizando
marcadores moleculares para la identificación de la presencia de L. fermentum.
2. Realizar la caracterización fisicoquímica de distintos jugos de piña
comerciales producidos a partir de pulpa de fruta, cáscaras y mezcla de pulpa y
cáscara.
3. Determinar la capacidad de sobrevivencia de una cepa de L. fermentum
en jugos de piña durante su almacenamiento en refrigeración (4 °C).

5
III. MARCO TEÓRICO
3.1. Piña y su importancia productiva en Costa Rica
3.1.1. Características generales
La piña (Ananas comosus L. Merr.) es una especie frutal nativa de países tropicales y
subtropicales en los cuales el clima cálido es constante, por lo que su cultivo es común en
América Central, incluido nuestro país, Suramérica y el Caribe (Bartholomew et al., 2003;
Hui et al., 2010). Sus características de aroma y sabor la han convertido en una de las frutas
tropicales y subtropicales no cítricas más populares en el mundo, por lo que su producción y
comercialización se da a gran escala (Hui et al., 2010).
En términos agronómicos, se caracteriza por ser una planta herbácea
monocotiledónea que genera un fruto compuesto formado por 100 a 200 frutículos unidos a
un eje central o corazón (Montero & Cerdas, 2005). Es una especie no climatérica cuya
maduración se alcanza entre los 18 a 20 meses después de que se realizó la siembra de la
planta madre (Murillo, 2014). Dentro de las variedades de piña que se cultivan se encuentran:
Montelirio, Cayena Lisa, Champaca y MD-2. Esta última, también es conocida como piña
Golden o Dorada, fue introducida al país por la compañía Del Monte, y ha adquirido fuerza
en el mercado debido a su fácil propagación, alta productividad, forma y color atractivo, y
dulzor atribuido al alto contenido de azúcares (Vagneron et al., 2009).
3.1.2. Producción e industrialización del fruto en Costa Rica
Se estima que entre 42 000 y 45 000 hectáreas (ha) de territorio nacional están
destinadas a este cultivo, sobre todo en la zona norte del país donde se da la explotación a
mayor escala (CANAPEP, 2012). Según datos de la FAOSTAT (2010), Costa Rica se ubica
entre los primeros lugares en la lista de producción y exportación de piña fresca a nivel
mundial y, para el año 2011, datos de la Promotora de Comercio Exterior (Procomer)

6
declararon un aumento en los ingresos para este producto, de forma que en ese momento el
producto ocupaba un puesto de liderazgo en el sector agrícola nacional (CANAPEP, 2012).
El rendimiento a gran escala puede variar de 80 a 110 ton fruta/ha dependiendo de la
variedad de los cultivares. El rendimiento efectivo oscila entre el 75 y 80 % de producto con
calidad de exportación como fruta fresca; mientras que el restante 15 a 25 %, corresponde a
frutos de rechazo que son destinados al consumo interno o a procesos de industrialización
posteriores (Montero & Cerdas, 2005).
A nivel industrial, la parte de mayor interés de la planta es la fruta, la cual puede ser
comercializada o consumida en su presentación fresca o procesada. Dentro de los productos
elaborados a partir de piña tanto para consumo nacional como para exportación se encuentran:
jugos, productos deshidratados o congelados, jaleas, pulpas y conservas. Como materia prima,
para obtener productos procesados, se utilizan frutas que deben cumplir parámetros de
calidad establecidos (Murillo, 2014) Esto genera, que una fracción importante se descarte
antes de ser procesada. Por otro lado, el porcentaje de rendimiento de la piña lista para
procesar, con respecto a la piña entera, es de un 45 a 55 %. Cerca de un 40 a 50 % constituye
un desecho sólido compuesto principalmente por cáscaras y corazones los cuales pueden
llegar a ser hasta un 65 % del peso total del fruto (Mat & Mat, 2008; Quesada et al., 2005).
A esto, se le suman las enormes cantidades de agua empleada para el lavado de frutas y
equipos que contiene altas concentraciones de materia orgánica y sólidos en suspensión, que
la convierten en un efluente altamente contaminante (Mat & Mat, 2008).
3.1.3. Manejo y aprovechamiento de subproductos industriales de piña
El volumen de desechos generados en el procesamiento de la piña, así como su
producción va en aumento. Esto ha generado un problema de importancia sanitaria,
económica y ecológica, dado a que el manejo y disposición de estos residuos involucra una
fuerte inversión económica y energética (González, 2012). Aunado a lo anterior, un manejo
inadecuado de los desechos orgánicos producidos puede causar malos olores y servir como
sustrato para la reproducción y desarrollo de plagas como la mosca de establo (Stomoxys

7
calcitrans), la cual tiene un fuerte impacto en la ganadería (Taylor & Berkebile, 2006),
además de ser un riesgo para la salud de las personas (MAG, 2013).
Actualmente, los subproductos de piña se emplean sobre todo como sustrato para la
elaboración de ensilajes destinados a la alimentación animal o bien para el consumo directo
de los animales. El ensilaje consiste en la fermentación de carbohidratos solubles del sustrato
en condiciones anaeróbicas que generan un producto final con mayor vida útil (Garcés, 2004).
El mismo, se logra por medio de una fermentación láctica espontánea en condiciones
anaerobias. Las bacterias acidolácticas epífitas fermentan los carbohidratos hidrosolubles del
forraje produciendo ácido láctico y en menor cantidad, ácido acético. Al generarse estos
ácidos el pH del material ensilado baja a un nivel que inhibe la presencia de microorganismos
que inducen la putrefacción (Merry, 1997; Weinberg & Muck, 1996).
Los desechos industriales de piña contienen componentes valiosos que están
constituidos principalmente por sacarosa, glucosa, fructosa, fibra y otros nutrientes en menor
cantidad como proteínas, vitaminas y minerales (Mat & Mat, 2008; Nadzirah et al., 2013).
La composición fisicoquímica de esta matriz hacen que sea una fuente potencial de carbono
para la producción de ácido láctico en sistemas microbianos, por lo que los desechos de piña
han demostrado ser una buena alternativa ecológica y económica como medio de crecimiento
para microorganismos (Pyar et al., 2014). Dado lo anterior, se les ha utilizado como sustratos
de fermentaciones para producir compuestos de alto valor agregado (Abdullah & Mat, 2008).
La flora nativa de los subproductos ensilados es muy variada (Knicky, 2005), por lo
que el estudio de las bacterias que pueden aislarse de este tipo de matrices ha adquirido un
alto interés científico debido a sus aplicaciones potenciales en diferentes áreas de producción
(Tobía et al., 2003; Franco et al., 2011). Las bacterias ácido lácticas (BAL) se caracterizan
por ser uno de los principales grupos de microorganismos que se han aislado de residuos
agroindustriales ensilados (Knicky, 2005).

8
3.2. Bacterias lácticas (BAL)
3.2.1. Antecedente histórico de las bacterias lácticas
Las bacterias ácido lácticas son, quizá, uno de los grupos más antiguos y conocidos a
nivel global. Muchas culturas en diferentes partes del mundo las utilizan en los procesos de
fermentación, para aumentar la vida útil, dar valor nutricional y mejorar las características
sensoriales de alimentos perecederos como leche, vegetales, carnes y cereales desde tiempos
inmemorables, por lo que tienen una función muy importante en la preservación de alimentos
destinados a humanos y animales (Estela et al., 2007; Garcés, 2004; Knicky, 2005). Por otra
parte, algunos tipos de estos microorganismos, son asociados a la producción de
características no deseadas y al deterioro de ciertos alimentos (Blackburn, 2006; Lahtinen et
al., 2012).
La conformación del grupo conocido como “bacterias ácido lácticas” se utilizó en un
inicio para denominar a las bacterias que causaban la fermentación y coagulación de leche
produciendo ácido láctico a partir de lactosa (Desai, 2008); sin embargo, actualmente, el
término engloba un grupo mayor de microorganismos. La base sistemática para la
clasificación de BAL fue elaborado y publicado por Orla (1919) quien les atribuyo el nombre
de familia Lactobacteriaceae, que agrupaba a un grupo fisiológico de bacterias que producían
ácido láctico como metabolito único o en conjunto con ácido acético, alcoholes y dióxido de
carbono (Desai, 2008; Halasz, 2009). A pesar de haber sido examinado varias veces, las
principales características de esta clasificación se han mantenido constantes a través de los
años (Desai, 2008).
Se caracterizan por ser bacterias Gram-positivas, no esporuladas, fermentadoras de
carbohidratos, productoras de ácido láctico, ácido tolerantes, anaerobias y catalasa negativas.
Típicamente, son no motiles y no reducen nitritos. El grupo se reconoce por comprender los
géneros: Streptococcus, Leuconostoc, Pediococcus ptococcus, Leuconostoc, Pediococcus,
Lactobacillus, Enterococcus, Lactococcus, Aerococcus, Carnobacterium, Leucnostoc,
Tetragenococcus y Vagococcus (Desai, 2008). Algunos otros géneros como Fructobacillus

9
y Weissella que originalmente se clasificaban como miembros del género Leuconostoc,
fueron reclasificados por Endo & Okada (2008) y Collins et al. (1993), respectivamente.
3.2.2. Clasificación de bacterias lácticas
De acuerdo con el sistema de clasificación de Orla-Jensen (1919), el género se divide
inicialmente de acuerdo a su morfología en bastones (Lactobacillus y Carnobacterium) y
cocos (todos los demás géneros). El siguiente nivel de clasificación se basa en su forma de
fermentar la glucosa bajo condiciones estandarizadas (sin limitaciones de glucosa, factores
de crecimiento y oxígeno limitado). Bajo estos criterios las bacterias lácticas se clasifican en:
homofermentativas, que convierten, casi cuantitativamente, los azúcares disponibles
exclusivamente en ácido láctico y heterofermentativas, que: producen ácido láctico, etanol,
ácido acético y dióxido de carbono. Otras características como su composición de ácidos
grasos, motilidad y temperaturas de crecimiento también se han empleado como base de
clasificación en algunos casos.
3.2.3. Metabolismo de bacterias lácticas
La principal característica del metabolismo de las bacterias lácticas es la degradación
de carbohidratos y otros derivados de carbono, para la producción de otras sustancias tales
como el ácido láctico y energía en forma de adenosin-tri-fosfato (ATP). El producto
predominante es el ácido láctico; sin embargo, ciertos cambios en la condiciones de
crecimiento puede generar cambios en su vía metabólica. Las hexosas, pentosas y disacáridos
son los principales sustratos metabólicos para estas bacterias. Cada una de éstas sigue una
ruta metabólica específica (Papagianni, 2012).
Las hexosas, como la glucosa y la fructosa, son azúcares recurrentes en la mayoría de
las matrices. El metabolismo de la glucosa sigue una vía homofermentativa; en tanto que,
otras hexosas como la fructuosa, ingresan a una vía de glicólisis después de una
isomerización o fosforilación, las hexosas fosforiladas se rompen en triosafosfatos, y luego,
por oxidación y desfosforilación se convierten en ácido pirúvico que luego se reduce a ácido
láctico. Si se sigue la vía heterofermentativa se obtienen otros productos secundarios
mencionados anteriormente (Papagianni, 2012). Las pentosas, se convierten en ribulosa-5-

10
fosfato o xilulosa-5-fosfato por medio de la epimerasas y se metabolizan por la vía pentosa-
fosfato. Los disacáridos requieren una hidrólisis previa en sus componentes básicos antes de
ser utilizados en alguna de las vías anteriormente descritas. La Figura 1, muestra algunas de
las vías metabólicas con las que se pueden metabolizar los azúcares según Kandler (1983).
Figura 1. Diferentes rutas metabólicas de las hexosas empleadas por bacterias lácticas
(Kandler, 1983).
3.3. Perfil del género Lactobacillus spp.
3.3.1. Características del género Lactobacillus
El género Lactobacillus spp. es el grupo de mayor tamaño en las Lactobacteriaceae,
y contiene al menos 100 especies (Dellaglio et al.., 2005; Satokari et al., 2003). Comprende
bacterias en forma de bastones que se asocian en cadenas cortas, son anaerobias facultativas
o microaerófilas, además de catalasa y citocromo negativas. Pueden poseer motilidad, donde
se mueven ayudados por flagelos peritricos (Estela et al., 2007). Los lactobacilos se
caracterizan por ser auxótrofos quimioorganotróficos, es decir, que necesitan medios
complejos para su crecimiento y que degradan los azúcares para producir lactato, por lo que
se consideran nutricionalmente fastidiosas (Estela et al., 2007; Canchaya et al., 2006). Su
temperatura óptima de crecimiento está entre los 30 y 40 °C. Los lactobacilos son utilizados

11
en la fermentación de materiales vegetales, lácteos y cárnicos, por lo que sus aplicaciones
industriales han sido muy estudiadas (Konings et al., 2000). Es posible aislarlas e
identificarlas en el tracto digestivo como parte de la microbiota intestinal del ser humano,
por lo que algunas especies se han explotado como probióticos (Canchaya et al., 2006). Este
género se caracteriza por ser muy heterogéneo y abarca especies con una amplia variedad de
propiedades fenotípicas, bioquímicas y fisiológicas. Dicha heterogeneidad se ve reflejada por
el contenido de guanina-citocina (GC) del ADN de sus especies (Desai, 2008).
3.3.2. Heterogeneidad genética de Lactobacillus spp.
La agrupación taxonómica de los lactobacilos, al igual que otras bacterias lácticas, se
basa en el metabolismo mayoritario de carbohidratos, que las clasifica en tres grupos:
homofermentativas (grupo metabólico A); heterofermentativas facultativas (grupo
metabólico B) y heterofermentativas estrictas (grupo metabólico C) (Hammes & Vogel, 1995;
Sun et al., 2015); sin embargo, la acumulación de nuevas secuencias del gen ADNr 16s y
otras secuencias genómicas han llevado a la conclusión de que dicha agrupación taxonómica
y filogénica no era concordante (Collins et al., 1991; Canchaya et al., 2006; Kant et al., 2011;
Makarova et al., 2006).
El genoma de los lactobacilos tiene un rango de tamaño que va desde los 1,23 Mpb a
los 4,91 Mpb y su contenido de GC varía considerablemente de 31,93 a 57,02 % (Sun et al.,
2015). Según Canchaya et al. (2006) al comparar los genomas de cinco cepas disponibles de
Lactobacillus; las 354 proteínas nucleares identificadas en estudio no eran específicas para
el género, y todos los árboles construidos mostraron una importante diversidad; del mismo
modo, en estudios posteriores, se ha apoyado dicha afirmación.
3.3.3. Lactobacillus fermentum
Lactobacillus fermentum es un bacilo Gram-positivo que pertenece al género
Lactobacillus spp. Es una bacteria láctica estrictamente heterofermentadora (Foschino et al.,
2001), que se caracteriza por tener una función importante, desde el punto de vista económico,
en la producción y preservación de alimentos fermentados y como cultivo iniciador productor
de ácido. Se ha aislado de una gran variedad de hábitats que incluyen productos fermentados

12
tradicionalmente, productos lácteos, masas fermentadas, vegetales fermentados y frutas,
entre otros (Dan et al., 2015).
El microorganismo se desarrolla a temperaturas que van de 35 a 37 °C. Pruebas
experimentales con aplicación de diferentes pH demostraron que tiene una alta tolerancia
para crecer y sobrevivir después de ser inoculada en matrices modelo con valores de pH de
hasta 3; además, de mostrar una buena tolerancia a la presencia de bilis (LeBlanc et al., 2004;
Pan et al., 2011). Estas características, entre otras, han sido significativas para que sea
considerado como un microorganismo probiótico (Pan et al., 2011). Por otro lado, se ha
demostrado que algunas cepas de L. fermentum son productoras de bacteriocinas que pueden
actuar como agentes antimicrobianos, los cuales se pueden emplear para la inhibición de
bacterias patógenas específicas (Riaz et al., 2010), además de inhibir el crecimiento de
microorganismos de deterioro como bacterias, levaduras y mohos (Fowoyo & Uzoma, 2015)
lo que permite aumentar la vida útil en productos.
3.3.4. Caracterización genómica de Lactobacillus fermentum
El genoma completo de L. fermentum consiste en un cromosoma circular que varía
de 2,0 - 2,2 Mpb, posee un contenido de GC entre 51,77 - 52,08 %, con ausencia de plásmido.
El cromosoma contiene 1109 - 2512 genes codificadores de proteínas predichos, 54 genes
codificantes para ARNt y 20 genes codificantes para ARNr (Jiménez et al., 2010; Kumari et
al., 2015; Illeghems et al., 2015).
El análisis de genoma realizado por Kumari et al. (2015) mostró que L. fermentum
codifica para genes con características para la hidrólisis de sales biliares (Chologlicina
hidrolasa), adhesión celular (unión a fibronectina y proteínas de la superficie de las células),
propiedades antioxidantes (tioredoxina, reductasa y glutatión reductasa), producción de
exopolisacárido (glicosiltransferasa), producción de bacteriocinas (proteína productora de
colicin V) y biosíntesis de folatos (dihidrofolato sintasa). Además, se han identificado genes
para la síntesis de alfa-galactosidasa y beta-galactosidasa que contribuyen a la absorción de
lactosa (Kumari et al., 2015). Estas propiedades pueden presentar variaciones dependiendo
de las cepas, sobre todo en genes codificadores de proteínas, enzimas, aminoácidos, lípidos
y carbohidratos (Jiménez et al., 2010).

13
3.4. Probióticos
3.4.1. Definición y características de un probiótico
Las bacterias ácido lácticas (BAL) son utilizadas en diversos tipos de alimentos
fermentados donde contribuyen con su sabor, textura y vida útil (Horn et al., 2005). Muchos
de estos microorganismos tienen efectos probióticos a través de su interacción con el intestino
humano y su microflora intestinal (Fiora et al., 2003). Los probióticos se definen
internacionalmente como microorganismos que, al ser administrados en cantidades
adecuadas por un tiempo sostenido, confieren ciertos beneficios al consumidor (Sanders,
2008). Dichos microorganismos deben estar presentes en una concentración mínima de 106
UFC/g para poder tener el efecto benéfico (Shah, 2000; Gueimonde & Sánchez, 2012).
Entre las características esperadas de un microorganismo para poder ser considerado
como probiótico según Tomasik & Tomasik (2003) y Shinde et al. (2012) se destacan:
Sobrevivencia a través del tracto digestivo, a un pH bajo y acción de
la bilis.
Adhesión a las células del epitelio intestinal.
Estabilidad de la microflora intestinal.
Cepa no patogénica ni tóxica.
Sobrevivencia en alimentos y posibilidad para la producción de
preparaciones liofilizadas.
Multiplicación rápida con permanente o temporaria colonización del
tracto gastrointestinal.
El consumo de los probióticos ha aumentado con la aparición de los hábitos de
consumo más saludables que promueven los efectos benéficos que éstos generan en el
organismo; sin embargo, su uso se ha limitado principalmente a productos de origen lácteo y
cereales (Pérez & Ramírez, 2007).
Los probióticos pueden contener una o más cepas bacterianas, algunas de ellas se
caracterizan por ser lactobacilos, bifidobacterias, estreptococos, enterococos, levaduras entre
otras. Las bacterias probióticas más comunes provienen principalmente de dos grupos:
14
Lactobacillus y Bifidobaceterium los cuales generalmente son considerados como GRAS
(generally referred as safe) (Donohue & Salminen, 1996; Rodríguez, 2006); aunque también,
pueden existir cepas provenientes de otros géneros. En el Cuadro I se pueden visualizar
algunas de las cepas comerciales que se pueden encontrar en el mercado en presentación
individual o en forma de mezclas.
Cuadro I. Cepas probióticas más comercializadas a nivel mundial y sus respectivas casas
comerciales (tomado de Shinde, 2012)
Cultivo Casa Comercial Cultivo Casa comercial
L. acidophilus NCFM ® Rhodia, Inc. (Madison, WI) L. casei Inmunitas Danone ( París, Francia)
L. acidophillus SBT-
20621
B. longum SBT-29281
Snow Brand Milk Products
Co. Ltd (Tokio, Japón)
L. fermentum RG-14
L. rhamnosus GR-1
Urex Biotech (Ontario, Canadá)
L. rhamnosus R0011,
L. acidophilus R0052
Institut Rossel (Montreal,
Canadá)
L. jhonsonii Nestlé ( Lausane, Suiza)
L. acidophilus LA-1,
L. paracasei CRL431
B. lactis Bb-12
Chr. Hansen (Horsholm,
Dinamarca)
L. plantarum 299V
L. rhamnosus ( Ras 271)
Probi AB (Lund, Suecia)
L. casei Shirota 1
B. breve Yakult1
Yakult (Tokio, Japón ) L. reuteri SD2112 Biogaia (Raleigh, NC)
L. rhamnosus GG1 Valio Diary ( Helsinki,
Finlandia)
B. lactis HNO19(DR10)
L. rhamnosus HN001 (DR20)
Fonterra (Wellington, Nueva
Zelanda)
L. rhamnosus LB21,
Lactococcus lactis L1A
Essum AB (Umea, Suecia) L. acidophillus LB Lacteol Laboratories (Houdan,
Francia)
L. salivarius UCC118 University College (Cork,
Irlanda)
L. paracasei F19 Medipharm ( Des Moines, Iowa)

15
3.4.2. Obtención y almacenamiento de cepas probióticas
Cuando se pretende estudiar cepas nuevas con capacidad probiótica es importante
hacer un muestreo de éstas en matrices en las cuales se encuentre el microorganismo de
acuerdo a su ecología y metabolismo. Una vez aisladas, deben ser identificadas
apropiadamente por medio de pruebas bioquímicas o por métodos moleculares (Ortiz et al.,
2008). Las cepas seleccionadas deben ser almacenadas a una temperatura mínima entre -
20 °C a -80 °C en crioviales que contienen una alícuota de la bacteria y un 20 a 30 % de
glicerol o dimetil sulfóxido (DMSO) al 5 %, de modo que la viabilidad del microorganismo
pueda mantenerse entre 3 a 5 años (Morales et al., 2010); en tanto que, otros métodos de
almacenamiento puede involucrar la liofilización de las cepas.
3.4.3. Criterios de evaluación de la capacidad probiótica
Los criterios utilizados para la selección de probióticos definen el control de calidad
óptimo de las cepas. Cambios en la matriz inoculada a lo largo de los procesos industriales
pueden afectar la estabilidad del probiótico, y por otra parte, las condiciones del sistema
gastrointestinal pueden ser sumamente estresantes, por lo que la cepas deben ser capaces de
mantenerse viables y activas aún bajo estos ambientes hostiles por lo que se han establecido
pruebas in vitro e in vivo para evaluarlas (Valerio, 2006).
3.4.3.1. Pruebas in vitro
Las pruebas in vitro permiten estudiar las características fisiológicas de las cepas
utilizado los perfiles de actividad fermentativa y enzimática para su evaluación; en estos
casos, es de vital importancia seleccionar el sustrato específico y la actividad enzimática
relevante a la funcionalidad esperada de la cepa. Este tipo de pruebas permiten evaluar la
resistencia del microorganismo a condiciones características del tracto gastrointestinal
(Valerio, 2006)
Dichas pruebas pueden ser cualitativas o cuantitativas (Kumura et al., 2004), y
permiten relacionar la actividad de las cepas con su capacidad probiótica in vivo, y las más
aplicadas son su estabilidad a un pH bajo, la acción de sales biliares y la adhesión celular.
Muchas de estas pruebas se aplican para la identificación de cepas con potencial probiótico
(Tuomola et al., 2001).

16
3. 4. 3. 1. 1. Estabilidad al ácido y a las sales biliares
Los microorganismos probióticos deben ser capaces de sobrevivir las condiciones que
se dan a lo largo de su trayecto por el tracto digestivo, lo cual implica que deben superar dos
barreras biológicas importantes: el ambiente de alta acidez del estómago y la actividad de la
bilis secretada por el duodeno (Shinde, 2012). Es esencial que el microorganismo sea capaz
de sobrevivir el ácido gástrico para poder alcanzar el intestino delgado y colonizarlo, de modo
que logre generar los beneficios asociados a ellas; por ende, se espera que un microorganismo
potencialmente probiótico pueda tolerar al menos un pH de 3,0 puesto que el pH del
estómago puede llegar a ser de 1,0 a 2,0 (Fernández et al., 2003). Por ello, la característica
de tolerancia ácida es una de las primeras y más importantes propiedades evaluadas al elegir
cepas (Tuomola et al., 2001; Desai, 2008). La tolerancia a sales biliares también es una de
las características que se puede tomar como base para la discriminación y selección de cepas
potencialmente beneficiosas.
El efecto de las condiciones gastrointestinales en la sobrevivencia de probióticos se
ha evaluado con diferentes métodos sobre diferentes cepas comerciales ya identificadas
como probióticas, presentando como resultado diferentes variaciones en los niveles de
tolerancia; por lo que se han desarrollado muchas técnicas para dichas evaluaciones, sin
embargo, no existe un protocolo estándar para la evaluación de la tolerancia al ácido y a las
sales biliares (Tuomola et al., 2001; Noriega et al., 2004). Se ha mostrado que estas
propiedades dependen de la cepa, por lo que se debe procurar tener cuidado cuando se
seleccionan las bacterias bajo este criterio. Se han identificado cepas de especies tales como
L. acidophilus y Bifidobacterium spp. que no presentan una buena tolerancia al ácido y a las
sales biliares (Lankaputhra & Shah, 1995; Desai, 2008); por lo que estas características no
son generalizables para todos los probióticos; sin embargo, se consideran propiedades válidas
para la selección de cepas potenciales.

17
3.4.3.1.2. Adhesión al epitelio intestinal
La evaluación de la capacidad de adhesión a la mucosa intestinal es otro criterio para
la selección de un probiótico. Las bacterias con una buena capacidad de adhesión se
consideran mejores probióticos debido a que esta característica puede aumentar el tiempo de
retención en el intestino así como su contacto con las células epiteliales lo que favorece la
colonización (Shinde, 2012). Al igual que en el caso anterior, se han desarrollado diferentes
métodos y modelos que incluyen la adhesión a la mucosa y la adhesión a las células
epiteliales (Gueimonde et al., 2006; Gueimonde et al., 2005).
Los resultados de las pruebas in vitro demuestran que se pueden presentar variaciones
dentro de la mismas cepas. Tuomola (2001) presentó, utilizando células de carcinoma del
colon (Caco-2) y glicoproteínas ileostómicas humanas, que solamente dos de seis cepas
evaluadas presentaron adhesión in vitro al epitelio intestinal y a la mucosa, en ambos modelos,
por lo que esta característica depende de las propiedades de la superficie de las células.
3.4.3.2. Pruebas in vivo
Las pruebas in vivo utilizan estudios clínicos con animales de laboratorio y personas
a los cuales se les suministra una dieta fortificada con el probiótico y posteriormente se hacen
evaluaciones de heces y biopsias del tejido gástrico (Zoetendal et al., 2002; Bron et al., 2004).
Las pruebas con animales son por lo general más complejas, y los resultados no son
siempre extrapolables a seres humanos. Dado lo anterior, estos resultados no pueden
utilizarse directamente para predecir la eficacia del probiótico en los seres humanos. Es
necesario realizar ensayos clínicos con personas; sin embargo, este tipo de pruebas
involucran una gran dificultad de aleatorización, control, inversión económica y están ligadas
a la ética profesional (Goktepe et al., 2006). Es importante evaluar la actividad probiótica in
situ; dentro de este contexto, el desarrollo de la biología molecular ha propuesto nuevos
métodos indirectos para la evaluación de la viabilidad y crecimiento del probiótico como lo
son la medición de metabolitos asociados a la muerte o del crecimiento celular (Bron et al.,
2004).

18
3.5. Aplicación de probióticos en alimentos
3.5.1. Aplicación de probióticos en productos lácteos
El sector lácteo está fuertemente ligado al uso de los probióticos y conforma el
mercado mayoritario de alimentos funcionales con un aproximado de 33 % del mercado
(Leatherhead Food International, 2006); a su vez, el consumo de leches fermentadas como el
yogurt ha mostrado un crecimiento importante en los últimos años (Granato et al., 2010). Los
principales probióticos asociados a lácteos son L. acidophillus, Lactobacillus casei y las
bifidobacterias que no solo dan un aporte funcional, sino que también, proporcionan
propiedades sensoriales deseables en los productos (Granato, 2010).
La leche y sus derivados son un medio ideal como vehículo de probióticos ya que la
mayoría de estas bacterias utilizan la latosa como fuente de energía; además, las proteínas
presentes en la leche proveen a los microorganismos de una protección importante durante
su paso a través del estómago (Charteris et al., 1998). Sumado a esto, el pH base de la leche
(5,5 – 6,8) resulta óptimo para la sobrevivencia del probiótico durante el almacenamiento.
Estudios realizados con algunas cepas específicas durante el almacenamiento de leche
inoculada y de yogurt por 28 días en refrigeración, ha mostrado una alta estabilidad y una
menor pérdida de viabilidad (Gilliland, 2002).
Las bebidas lácteas fermentadas generan una importante contribución a la dieta
humana en muchos países ya que el proceso fermentativo es una tecnología económica que
permite preservar y mejorar la calidad nutricional y sensorial (Gadaga et al., 1999). La
estabilidad comercial de este tipo de productos permite la expansión y la cobertura requerida
en diferentes categorías de consumidores.
3.5.2. Aplicación de probióticos en productos no lácteos: jugos
Los productos probióticos son usualmente comercializados en forma de leches
fermentadas y yogurt; sin embargo, con el aumento de consumidores veganos y vegetarianos
alrededor mundo, así como las personas intolerantes a la lactosa y los consumidores
preocupados por el contenido de colesterol, ha generado la necesidad de buscar nuevas
alternativas no lácteas para la elaboración de bebidas con probióticos (Prado et al., 2007).

19
Existe una amplia gama de bebidas no lácteas fermentadas y tradicionales en de diversas
partes del mundo, donde la mayoría consiste en bebidas no alcohólicas elaboradas
principalmente a partir de cereales (Prado et al., 2007).
Recientemente, las bebidas a base de frutas, vegetales, cereales y soya se han
propuesto como vehículos para probióticos. Los jugos de frutas, en particular, se han
reportado como medio novedoso y apropiado para los probióticos debido a su alto contenido
de nutrientes esenciales, y porque es reconocido como un alimento saludable (Luckow et al.,
2006). Sin embargo, el bajo pH de la mayoría de los jugos representa la mayor limitante para
mantener la viabilidad y la actividad de las bacterias probióticas durante la vida útil del
alimento (Perricone et al., 2015).
3.5.2.1. Limitaciones de la sobrevivencia de probióticos en jugos de fruta
Como se mencionó anteriormente, las propiedades beneficiosas de los probióticos se
basan estrictamente en su concentración en el alimento, y por su capacidad de sobrevivencia
a través del tracto digestivo; por lo tanto, la concentración de las bacterias en el producto al
final de su vida útil debe ser entre 106 a 107 UFC/mL (Gueimonde et al., 2012; Perricone et
al., 2015).
La composición fisicoquímica de los jugos los convierte en matrices ricas en
nutrientes esenciales como azúcares, minerales, vitaminas, fibra y antioxidantes; sin embargo,
existen ciertos factores importantes que pueden limitar la sobrevivencia de las bacterias en
jugos. Tripathi & Ciri (2014) agrupan dichos factores como:
(1) Parámetros del alimento: pH, acidez titulable, oxígeno molecular, actividad de
agua, presencia de sales, azúcares y compuestos químicos como peróxido de
hidrógeno, bacteriocinas, saborizantes y colorantes.
(2) Parámetros del proceso: tratamientos térmicos, temperaturas de incubación, razón
de enfriamiento, material de empaque, métodos y condiciones de almacenamiento,
nivel de oxígeno y volumen.
(3) Parámetros microbiológicos: cepas, razón y proporción del inóculo.

20
El pH es uno de los factores más importantes que afectan la sobrevivencia del
probiótico. Los jugos poseen un alto contenido de ácidos orgánicos, y un valor de pH bajo
aumenta la concentración de la forma no disociada de la molécula, por lo que se presume que
existe un acción combinada de las condiciones acidas y el efecto antimicrobiano intrínseco
sobre las bacterias (Perricone et al., 2015). Las especies del genero Lactobacillus son,
generalmente, resistentes y sobreviven en jugos con valores de pH que oscilan entre 3,7 y
4,3. Por el contrario, las bifidobacterias son menos tolerantes al ácido y un pH de 4,6 es
perjudicial para su sobrevivencia (Tripathi & Giri, 2014). En general, el pH genera un efecto
perjudicial, pero las proteínas y la fibra dietética pueden funcionar como protectores de las
células ante el estrés ácido. Por otro lado, la función del ácido cítrico y del ácido málico se
considera controversial ya que estos muestran cierta capacidad de protección de las bacterias,
mientras que los fenoles pueden causar una pérdida importante de viabilidad (Nualkaekul &
Charalampopoulos, 2011; Perricone et al., 2015).
A pesar de que el pH representa una limitante de sobrevivencia importante, se ha
considerado que la incorporación de bacterias lácticas en jugos de alta acidez puede mejorar
la resistencia de las mismas a condiciones estresantes de acidez subsecuentes como las que
se dan en el tracto gastrointestinal (Ranadheera et al., 2014).
3.5.2.2. Mejoramiento de la sobrevivencia de probióticos en jugos
Muchos autores han propuesto distintas estrategias para mejorar la tasa de
sobrevivencia de probióticos en jugos de frutas y en esta sección se describirán brevemente
algunas de estas estrategias.
3.5.2.2.1. Fortificación con prebióticos
Los prebióticos son sustancias fermentables que favorecen el crecimiento y la
actividad de los microorganismos probióticos, de forma que se potencian los efectos
benéficos de estos sobre su hospedero (Gibson et al., 2004). La adición de este tipo de
productos puede generar un efecto de protección y además mejora la estabilidad de las
bacterias.

21
3.5.2.2.2. Adaptación e inducción de resistencia
La exposición de los probióticos a condiciones sub-letales de estrés puede inducir una
cierta resistencia o a una respuesta adaptativa al estrés que ayudan a mejorar sus
características (Gobbetti et al., 2010; Perricone et al., 2014). Otros métodos pueden
involucrar una mutagénesis inducida de la especie para crear una variante más tolerante a los
factores deseados (Saarela et al., 2011).
3.5.2.2.3. Refrigeración y adición de antioxidantes
La viabilidad del probiótico está relacionada de forma negativa con la temperatura de
almacenamiento; de forma que bajas temperaturas pueden extender la sobrevivencia,
mientras que, temperaturas más altas han mostrado efectos perjudiciales (Perricone et al.,
2015).
Por otro lado, el nivel de oxígeno dentro del envase durante el almacenamiento
debería ser lo más bajo posible para prevenir la toxicidad y muerte, debidas a la inducción
del daño oxidativo por la formación de especies reactivas de oxígeno como el peróxido de
hidrógeno (Perricone et al., 2015).
3.5.2.2.4. Microencapsulación
Se han desarrollado y diseñado tecnologías de microencapsulación con diversos
materiales como hidrocoloides que genera una barrera protectora alrededor de la bacteria
impidiendo cualquier daño causado por el entorno en el que se encuentra, especialmente las
condiciones de alta acidez (Anal & Singh, 2007). Varios estudios reportan que la aplicación
de la microencapsulación genera un ambiente anaérobico favorable para bacterias
especialmente sensibles (Ding & Shah, 2008).
3.5.3. Características sensoriales de los jugos con probióticos
Uno de los principales retos del desarrollo de productos no lácteos con probióticos es
la aceptación por parte de los consumidores. El éxito comercial de este tipo de productos
depende directamente del sabor, apariencia, precio y del efecto benéfico que se presenta a los

22
consumidores; a esto, además, se suman los aspectos técnicos como la estabilidad
fisicoquímica, tipo de empaque y vida útil, entre otras (Granato et al., 2010).
Los efectos de los probióticos en las características sensoriales de los jugos dependen
directamente del microorganismo empleado, las condiciones de almacenamiento, y de la
adición de otros componentes (Perricone et al., 2015). El empleo de algunas cepas no han
afectado la aceptación general de los jugos como es el caso del jugo de piña con Lactobacillus
reuteri (Perricone et al., 2014). Sin embargo, este no es el caso de muchos otros
microorganismos, dado a que el efecto fermentativo que desarrollan durante el
almacenamiento pueden generar aromas y sabores indeseados que podrían asociarse al
deterioro debido a la formación de sustancias como el ácido láctico y a otras alteraciones en
las propiedades fisicoquímicas del jugo (Blackburn, 2006).
Una posible solución para los jugos en los cuales la presencia del probiótico tiene un
efecto negativo en la aceptación general del producto es el enmascaramiento por medio de la
adición de aromas o bien por el reforzamiento de las características aromáticas intrínsecas de
jugo en sí (Luckow et al., 2006; Ranadheera et al., 2014).
3.6. Técnicas de biología molecular para la identificación de bacterias
El ácido desoxirribonucleico (ADN) y ribonucleico (ARN) son moléculas que
almacenan la información genética de todo organismo metabólicamente activo; su química
única permite que la información sea copiada y heredada a la descendencia, lo cual ha
permitido el desarrollo de ensayos innovadores a nivel molecular (Clark, 2005).
3.6.1. Conceptos básicos de las técnicas de biología molecular
La biología molecular es el estudio de los organismos vivos y virus a nivel de las
moléculas que los controla y les permite existir. A diferencia de la biología tradicional que
estudia los organismos y su interacción dentro de una población, la biología molecular se
centra en estudiar los organismos a partir de sus componentes (Clark, 2005) y la bioquímica
que conecta los procesos de la vida por medio de técnica moleculares especializadas.

23
Las técnicas moleculares son herramientas para el análisis de los microorganismos de
alimentos y de otras sustancias biológicas; y además, proveen formas de diagnóstico para un
amplio rango de agentes por medio de una única prueba (Babalola, 2003). Las técnicas varían
según su poder de discriminación, reproductibilidad, facilidad de uso e interpretación (Lasker,
2002). Entre las más utilizadas pueden citarse, la electroforesis, la reacción en cadena de la
polimerasa (PCR), la digestión restrictiva, la ligación, el blotting y la clonación.
3.6.2. Gel de electroforesis
Esta técnica permite separar ADN, ARN y las proteínas según su tamaño y carga
eléctrica (Kryndushkin et al., 2003). El principio, consiste en exponer una mezcla de
biomoléculas cargadas a un campo eléctrico de una fuerza E, por lo que las mismas se
movilizan hacia el electrodo de carga opuesta; sin embargo, las diferentes moléculas se
mueven a un ritmo distinto dependiendo de las propiedades individuales de cada una
(Rabindra & Raju, 2012), donde esta separación se conoce como tamizado (Sambrook &
Russel, 2001).
La velocidad del movimiento (v) depende de la carga de la molécula dentro del campo
eléctrico, según las variables descritas por la ecuación (1):
𝑣 = 𝐸.𝑞
𝑓 (𝟏)
En donde f es el coeficiente de fricción y q la carga neta sobre la molécula. El
coeficiente de fricción describe la resistencia al movimiento y depende de factores como la
masa, su nivel de compactación, la viscosidad del buffer y la porosidad de la matriz, mientras
que la carga neta está determinada por las cargas de la molécula (Rabindra & Raju, 2012).
Los geles más utilizados en esta técnica son los geles de agarosa y de poliacrilamida;
cada uno de estos es óptimo para diferentes tipos y tamaños del analito. Los geles de
poliacrilamida son utilizados para proteínas y se caracterizan por tener un alto poder de
resolución para fragmentos muy pequeños de ADN (5 - 500 pb); los geles de agarosa, por el
contrario, tienen un poder de resolución más bajo, pero un mayor rango de separación por lo
que son adecuados para fragmentos que van de los 50 - 20 Kpb (Maniatis et al., 1989;

24
Rabindra & Raju, 2012). El sistema se completa con adición de un “buffer” (solución tampón)
que provee los iones que promueven el flujo de electrones y mantienen el pH a un valor
relativamente estable, donde los más utilizados son el Tris/Acetato/ácido
etilendiaminotetraacético (EDTA) (TAE) y el Tris/Borato/EDTA (TBE).
La electroforesis en gel se utiliza para evaluar la amplificación del ADN por PCR;
sin embargo, también puede emplearse como una técnica preparativa previa al uso de otros
métodos como la clonación, la secuenciación y el blotting.
3.6.3. Reacción en cadena de la polimerasa (PCR)
Los métodos de PCR se han descrito con detalle por Hoelzel & Green (1998); esta
técnica consiste en amplificar pequeñas cantidades de ADN a un volumen que puede ser
empleado para otro análisis. Es una herramienta básica en biología molecular que se basa en
una ciclación térmica que consiste en ciclos repetitivos de tres pasos que requieren distintas
temperaturas. Cada uno de dichos pasos está dirigido a procesos que conllevan a la
generación de copias del gen seleccionado en presencia de primers o iniciadores que
contienen la secuencia complementaria a la región blanco y la enzima ADN polimerasa que
permite la amplificación selectiva y repetitiva. A medida que el PCR progresa, el ADN
generado se utiliza como el molde de replicación, lo cual desencadena una reacción en cadena
exponencial (Ishmael & Stellato, 2008).

25
El primer paso consiste en la separación del ADN de doble banda por medio de un
calentamiento de la muestra a aproximadamente 90 °C; luego, la muestra enfría permitiendo
el anillamiento (apareamiento de bases específicas) de los iniciadores a sus bandas
complementarias del ADN (Wittwer et al., 1997). En el tercer paso, la mezcla se calienta
hasta 72 °C la cual es la temperatura óptima para la actividad de la Taq ADN polimerasa, y
se inicia la síntesis de las hebras de ADN utilizando los iniciadores como puntos de partida
y los nucleótidos trifosfato (dNTPs) presentes en la mezcla para generar la hebra
complementaria específica en un proceso denominado elongación (Figura 2). La repetición
de estos tres pasos resulta en la duplicación del número de copias por cada ciclo, es decir que
el número de copias equivale a 2n, donde n representa al número de ciclo; por lo tanto, la
generación de productos en el PCR sigue un patrón exponencial que alcanza un fase constante
Figura 2. Diagrama de los ciclos de amplificación del material nucleico por reacción en cadena
de la polimerasa (PCR).

26
después de 30 - 40 ciclos con lo que se ha consumido la mayoría de los reactivos (Ishmael &
Stellato, 2008).
3.6.3.1. Aplicaciones del PCR
Dado a que el PCR ayuda con la amplificación y manipulación del ADN, esta técnica
ha permitido expandir la aplicación de la biología molecular a otros campos de investigación,
ya que un gen de interés puede ser selectivamente amplificado utilizando los iniciadores
específicos, e introducidos a otros plásmidos o vectores para un estudio a mayor profundidad
(Ishmael & Stellato, 2008).
Actualmente, la técnica se aplica para una gran diversidad de análisis que incluyen
estudios genéticos de mutación, recombinación y mejoramiento, así como expresión de
características deseadas; por otra parte, el PCR también se emplea para el diagnóstico de
enfermedades inducidas por anormalidades celulares, males genéticos y cáncer (Hensel et al.,
1991; Hentemann et al., 1990; Saiki et al., 1985). La técnica en si se ha empleado
ampliamente en la medicina forense, y actualmente, se aplica para la identificación de
patógenos como virus y bacterias.
3.6.4. Identificación de bacterias
Las bacterias se clasifican e identifican con el objetivo de permitir la distinción de un
organismo de otro, y a su vez, agrupar organismos similares según criterios de interés
microbiológicos o científicos. Dentro de los diversos niveles de clasificación posibles, uno
de los más importantes es la especie (Clark, 2005).
Los miembros de una misma especie suelen compartir características en común, ya
que dentro de la misma pueden existir cepas y subgrupos con propiedades que difieren y las
identifican. Originalmente, las especies se creaban con base a criterios de comparación de
una morfología y de descripción fenotípica precisa de la muestra con respecto a lo reportado
para una especie conocida (Clarridge, 2004). La verificación de la identificación debe ser
corroborada con pruebas bioquímicas o genéticas (Baron, 1996).

27
La clasificación molecular más temprana de las bacterias utilizó el contenido de GC,
el perfil de plásmidos y la compatibilidad a la transformación genética (Barghouthi, 2011).
En la década de los años 80´s se desarrolló un nuevo estándar para la identificación de
bacterias que utiliza la comparación de un fragmento estable del código genético (Clarridge,
2004; Woese et al., 1985; Woese, 1987). Los fragmentos candidatos en bacterias incluían
genes que codificaban para el ADNr 5S, 16S y el 23S, además de las regiones intergénicas
transcritas (ITS, por sus siglas en inglés) (Figura 3); sin embargo, el fragmento más utilizado
para la identificación taxonómica de bacterias es gen ARNr 16S, también llamado ADNr 16s
(Barghouthi, 2011; González et al., 2006).
3.6.5. Identificación por medio del gen del ADNr 16S
El gen del ADNr 16S se caracteriza por ser una región muy preservada con pequeños
cambios que pueden ser característicos de diferentes especies (González, et al., 2006), forma
parte de la subunidad ribosomal 30S; y está constituida por aproximadamente 1500 pb que
codifican para un ARN catalítico y su funcionalidad constante asegura su validez como
cronómetro molecular.
Este gen se encuentra presente en todas las células procariotas y contiene regiones de
secuencias conservadas y variables que evoluciona a ritmos distintos, lo cual es crítico para
la amplificación universal concurrente y la medición de relaciones filogenéticas cercanas y
distantes. Se ha demostrado que los genes del ADNr 16S bacteriano muestran regiones
hipervariables que presentan diversidad de secuencias entre distintas bacterias que permiten
una diferenciación por medio de la amplificación de secuencias específicas utilizando de
iniciadores universales (McCabe et al., 1999, Srinivasan et al., 2015; Van de Peer et al.,
Figura 3. Diagrama de distribución de genes bacterianos.

28
1996); lo anterior, permite utilizar esta región para establecer relaciones cercanas en géneros
(Clarridge, 2004) e inclusive especies (Newton & Roeseler, 2012). Este gen puede ser
comparado con el de todas las bacterias, además con el ADNr 16S de las arqueobacterias y
el ADNr 18S de los eucariotas.
3.6.6. Ventajas y limitaciones de la técnica de PCR
La identificación precisa de las bacterias es importante, y muchos se han cuestionado
si los métodos convencionales que utilizan el cultivo celular permiten la distinción correcta
entre los aislados. Dada la infinidad de especies que comparten fenotipos morfológicos y
bioquímicos similares, la identificación errónea de los individuos es muy común si se basa
únicamente en estos criterios como parámetros de evaluación (Clarridge, 2004), por lo que a
pesar de que los métodos convencionales son más económicos, en muchos casos no logran
los resultados más precisos.
Actualmente, la identificación taxonómica que utiliza el ADNr 16S se realiza por
comparación de pares alineados, con secuencias recopiladas en bases de datos que
corresponden a bacterias conocidas. Su utilidad depende de que las secuencias presenten
diferencias significativas interespecies y pequeñas diferencias intraespecie. Una de sus
principales limitantes, es que cuando dos bacterias poseen una secuencia casi idéntica del
gen, la técnica no es suficiente para distinguir entre ellas, por lo que es vital hacer un análisis
a profundidad de los resultados antes de llegar a concluir la identidad de la muestra (Woo et
al., 2008). Otra de las limitaciones, es que las secuencias analizadas deben tener el largo
completo o cercano al gen, dado que el uso de genes incompletos o muy cortos limita la
resolución taxonómica (Huse et al., 2008; Klindworth et al., 2013).

29
IV.MATERIALES Y MÉTODOS
4.1. Localización del proyecto
El desarrollo de la identificación molecular de las BAL aisladas de los ensilados se
realizó en las instalaciones del Centro de Investigaciones de Biología Celular y Molecular
(CIBCM) de la Universidad de Costa Rica, sede Rodrigo Facio. Para efecto de las pruebas
de sobrevivencia, caracterización fisicoquímica de los jugos y el recuento de L. fermentum,
los análisis se efectuaron en los laboratorios de química y microbiología del Centro Nacional
de Ciencia y Tecnología de Alimentos (CITA) y en el laboratorio de Análisis Químico de la
Escuela de Tecnología de Alimentos de la Universidad de Costa Rica.
4.2. Materias primas
Se seleccionaron tres tipos de matrices para realizar la evaluación de la sobrevivencia
de una cepa de bacteria láctica previamente identificada. Las matrices consistieron en tres
jugos de piña comercialmente estériles y provistos en empaques asépticos, procedentes de la
empresa nacional Florida Products S.A ubicada en la zona industrial de Barreal de Heredia.
Los jugos evaluados fueron los siguientes:
a) Jugo obtenido de la pulpa de piña procesada.
b) Jugo concentrado obtenido a partir del prensado de las cáscaras de piña.
c) Jugo concentrado obtenido de una mezcla de los jugos indicados en (a)
y (b), en una proporción 80:20, respectivamente.
Para efecto de análisis, los jugos (b) y (c) fueron reconstituidos hasta alcanzar los
13 °Brix y sometidos a un proceso de llenado en caliente para garantizar la esterilidad
comercial. El pH de los jugos no fue ajustado.
4.3. Aislamiento de los microorganismos
Las 12 cepas estudiadas fueron aisladas por colaboradores del Centro de
Investigaciones Agronómicas (CIA), a partir de subproductos del procesamiento industrial

30
de la piña (cáscaras y coronas), destinados a la elaboración de ensilajes para alimentación
animal provenientes de Florida Products S.A. Las cepas de las BAL fueron almacenadas en
refrigeración a 4 °C hasta su respectiva extracción de ADN.
4.4. Pruebas preliminares:
Se evaluaron tres protocolos de extracción de ADN para determinar el método óptimo
para la obtención de ácidos nucleicos de buena calidad: extracción de bacterias utilizando
Chelex® (BioRad), extracción de ADN ribosomal de bacterias (Fontecha, 2005) y el
extracción de plásmido MiniPrep (Birnboim & Doly, 1979) empleando ARNasa A a 10
ng/uL (Thermo Scientific) y utilizando como sujeto de ensayo una cepa de Escherichia coli
no patógena. Según los resultados obtenidos en esta prueba preliminar, se probaron dos de
los protocolos que mejor funcionaron con cinco BAL previamente aisladas de los silos de
piña, donde el ADN de las mismas fue extraído por duplicado.
4.4.1. Estandarización del protocolo de extracción de ADN para las BAL
Las diferentes extracciones se corrieron en geles de agarosa al 1 % a 100 V por 30
min. La tinción se realizó utilizando Gel-red (10 000X) (Biotium, Estados Unidos) como
tintura de los ácidos nucleicos para determinar la integridad del material. El ADN fue
cuantificado por densidad óptica utilizando un espectrofotómetro UV-Vis NanoDrop 2000c
(Thermo Scientific, Estados Unidos) a una longitud de onda de 260 nm y se determinó su
pureza usando como referencia el factor A260/A280.
4.4.2 Estandarización del protocolo de amplificación para el ADNr16S
Las extracciones de mejor calidad y pureza previamente cuantificadas, fueron
diluidas a concentraciones de 20, 30 y 50 ng/uL. La estandarización de las reacciones de PCR
se realizó utilizando los iniciadores 1492R (5´-GGTTACCTTGTTACGACTT-3´) y 27F (5´-
AGAGTTTGATCCTGGCT CAG-3´); además, se utilizó un rango de temperaturas de
anillamiento de 45-56 °C. Los productos de PCR obtenidos se corrieron en geles de agarosa
al 1 %, utilizando 110 V y 40 min, luego fueron teñidos utilizando azul de carga y Gel-red
(10 000X) (Biotium, Estados Unidos) como tintura de los ácidos nucleicos.

31
4.5. Identificación molecular de las BAL
4.5.1. Extracción de ADN
Las cepas de bacterias lácticas (BAL) aisladas de los silos, se crecieron en medio
Man-Rogosa-Sharpe (MRS) líquido (Oxoid, Thermo Scientific Fisher, Reino Unido). Fueron
incubadas en agitación a 225 rpm por al menos 12 h a 35 ± 1 °C. La extracción de los ácidos
nucleicos totales (ANt) se realizó siguiendo el protocolo de extracción de ADN de plásmido
Miniprep por método convencional con fenol (Birnboim & Doly, 1970) para el cual se tomó
1 mL del cultivo bacteriano y se centrifugó a 13 000 rpm por 2 min en una microcentrífuga
modelo 5415D (Eppendorf, Alemania). Se descartó el sobrenadante, y el botón obtenido fue
resuspendido en 100 µL de solución I (25mM Tris-HCL pH de 8,0; 10 mM de ácido
etilendiaminotetraacético (EDTA), 50 mM D-Glucosa. La mezcla fue incubada a temperatura
ambiente por 5 min; posteriormente, se adicionaron 200 µL de solución II (0,2 N de
Hidróxido de Sodio (NaOH) y 1 % de dodecilsulfato sódico (SDS), se mezcló por inversión
y se incubo por 5 min a 4 °C. Finalmente, se adicionaron 150 µL de solución III (3 M acetato
de potasio y 2 M de ácido acético pH de 4,8) que se mantuvo a 4 °C. La mezcla se incubó
por otros 5 min a 4 °C. Luego, se realizó una centrifugación a 13 000 rpm por 10 min. Un
volumen de 450 µL del sobrenadante se transfirió a un tubo limpio y se adicionó 450 µL de
fenol/cloroformo (1:1 v/v). La mezcla se agitó por al menos 2 min y se centrifugó a 13 000
rpm por 10 min. La fase acuosa fue transferida a un tubo limpio y se adicionó 800 µL de
isopropanol. La mezcla se realizó por inversión e incubando por 2 min a temperatura
ambiente. Luego, fue centrifugada por 10 min a 12 000 rpm, se descartó el sobrenadante, y
el botón obtenido se lavó con 500 µL de etanol 70 % y se dejó secar.

32
4.3.3. Reacción en cadena de la polimerasa (PCR)
Los ácidos nucleicos totales (ANt) fueron resuspendidos en 50 μL de agua bidestilada
estéril. Para la reacción de PCR se utilizaron los iniciadores 1492R (5´-
GGTTACCTTGTTACGACTT-3´) y 27F (5´-AGAGTTTGATCCTGGCT CAG-3´). El
volumen final fue de 25 µl, que contenía 1 X de buffer (ThermoScientific, Estados Unidos),
2,5 µL de dNTPs (10 µM), 0,4 µM de cada iniciador, 1 U Taq ADN polimerasa (Thermo
Scientific, Estados Unidos) y 20 ng de ADN. El programa de amplificación consistió en una
desnaturalización inicial a 94 °C por 1 min, seguido por 30 ciclos de la siguiente manera:
94 °C a 30 s; 55 °C a 30 s y una elongación de 72 °C por 1,30 min; finalmente, se da una
elongación terminal de 72 °C a 10 min. Los productos de PCR se corrieron en geles de
agarosa al 1 % utilizando azul de carga y Gel-red (10 000 X) (Biotium, Estados Unidos)
como tintura de los ácidos nucleicos a 100V por al menos 40 min. Se utilizó un marcador de
100 pb (Thermo Scientific, Estados Unidos). Los productos de PCR obtenidos fueron
purificados utilizando el kit NucleoSpin (Machery Nagel, Alemania). Las muestras fueron
enviadas para realizar un análisis posterior de sus secuencias a Macrogen Inc. (Corea).
4.3.4. Análisis filogenético
Las secuencias obtenidas fueron ensambladas utilizando el programa Staden 4.0
(Bonfield et al., 1995). El contig obtenido fue comparado con las secuencias de otras
bacterias lácticas obtenidas de la base de datos públicas (Altschul et al., 1990), se realizó una
comparación de la identidad de las secuencias utilizando la herramienta BlastN
(www.ncbi.com). Los análisis de filogenia se realizaron utilizando todas las secuencias
obtenidas de las 12 BAL estudiadas, y se incluyeron secuencias de otras cepas de colección
disponibles en el GenBank (http://www.ncbi.nlm.nih.gov/). Las secuencias fueron alineadas
utilizando el programa MUSCLE (Mega 6.0) (Tamura et al., 2013). Los árboles filogenéticos
se elaboraron utilizando el programa Molecular Evolutionary Genetics Analysis (MEGA)
versión 3.1 (Kumar et al., 2004). La robustez de las ramas internas se estimó por medio de
análisis de “bootstrap” con 2000 réplicas. Los análisis filogenéticos fueron comparados con
los métodos de distancia, el vecino más cercano (Neighbor Joining) y el Unweighted Pair

33
Group Method with Arithmetic Mean (UPGMA) , las secuencias de este estudio se denotan
con otro color, y solamente se muestran los valores de las ramas mayores al 50 %.
La matriz de colores para realizar la demarcación de las especies fue elaborada
utilizado la herramienta SDTv1.2 (Muhire et al., 2014). Todas las secuencias de las BAL
fueron depositadas en el GenBank (http://www.ncbi.nlm.nih.gov) y comparadas con las
cepas más cercanas disponibles en esta base de datos y el porcentaje de identidad fue obtenido
utilizando la herramienta Pairwise Alignment (http://www.ncbi.nlm.nih.gov).
4.5. Caracterización físicoquímica de los jugos de piña inicial y luego de
almacenamiento
Los jugos de piña comerciales empleados para el estudio se sometieron a análisis
fisicoquímicos en el tiempo inicial (0 semanas) y transcurrido el período de almacenamiento
(6 semanas) en refrigeración a 4 °C, luego de ser inoculados con una alícuota de una
concentración de aproximada de 108 UFC/mL de una cepa L. fermentum aislada de los silos
de subproducto industrial de piña. Los análisis realizados consistieron en:
Sólidos totales: Se determinó el contenido de humedad basado en la medida
gravimétrica de pérdida de masa por evaporación de agua en una estufa de convección al
vacío (National Appliance Co., Estados Unidos) según el método P-SA-MQ-002 (CITA,
2009d) basado en los métodos oficiales AOAC 920.151 y 925.45 (2005e, 2005h). El
contenido de sólidos se obtiene por diferencia respecto al contenido de humedad.
pH: se midió utilizando un pH-metro modelo 450 (Corning, Estados Unidos)
previamente calibrado con buffers con pH de 4,00 y pH de 7,00 siguiendo el procedimiento
establecido en el documento P-SA-MQ-012 (CITA, 2009c) el cual se basa en el método
AOAC 981.12 (2005f).
Acidez total titulable: se realizó la determinación de acidez titulable por medio de una
valoración siguiendo el método establecido por el método P-SA-MQ-011 (CITA, 2009a), el
anterior se basa en el método AOAC 942.15 (2005a). Las valoraciones se realizaran con el
titulador automático Tiamo, 836 Titrando (Methrohm, Suiza) y se expresó en términos de
gramos de ácido cítrico por cada 100 mL de jugo.

34
Sólidos solubles totales (SST): Se determinó por medio de un refractómetro de Abbé
1T (Atago, Japón) colocando una gota de cada jugo sobre el prisma. Los sólidos solubles
fueron determinados por lectura directa en el equipo y expresados como ºBrix corregidos a
20 °C según lo estipulado en el método AOAC 932.12 (2005 g).
Azúcares fermentables: El contenido de azúcares fermentables se determinó por
medio de cromatografía líquida de alto rendimiento (HPLC, por su siglas en inglés)
utilizando como fase móvil acetonitrilo y agua purificada a una razón de 75:25 v/v con flujo
isocrático de 1,0 mL/min a temperatura ambiente y empleando estándares de fructosa,
glucosa y sacarosa según lo establece el método P-SA-MQ-006 (CITA, 2011). Los azúcares
fueron cuantificados mediante la comparación de las áreas de cada pico con respecto a las
del patrón (Nadzirah et al., 2013).
Ácidos orgánicos totales: la determinación del contenido de ácidos orgánicos totales
se realizó por HPLC según lo descrito por el método P-SA-MQ-051 (CITA, 2016) usando
como fase móvil ácido sulfúrico 0,00225 mol/L con una temperatura de 60 °C, a una longitud
de onda de 210 nm y un flujo de 0,50 mL/min.
4.6. Ensayo de capacidad de sobrevivencia de una cepa BAL seleccionada
4.6.1. Selección de la bacteria
Los 12 morfotipos de BAL aislados se sometieron a ensayos de tolerancia a un pH de
2,0 para determinar su resistencia a una acidez similar a la del jugo gástrico. El ensayo se
realizó siguiendo el procedimiento descrito por Ramos et al. (2013). Las células bacterianas
fueron crecidas en MRS líquido (Oxoid, Reino Unido) a 35 °C, durante 24 h y se
centrifugaron (5000 rpm por 5 min a 24 °C). El botón fue lavado dos veces en agua peptonada
0,1 % m/v, pH de 7,0. Las células fueron resuspendidas en agua peptonada (densidad óptica
0,2 a 600 nm) con una concentración estimada de aproximadamente 108 UFC/mL. Se
inocularon tubos de MRS líquido acidificado con ácido clorhídrico 1 N a un pH de 2 y a una
concentración de 106 UFC/mL. Se realizó un recuento por vaciado a los tiempos 0 y 3 h. Las
placas se incubaron en condiciones anaerobias a 35 ± 1 °C por 72 ± 3 h según lo establece el

35
procedimiento P-SA-MM-006 del CITA (2012) basado en los métodos de Pouch & Ito (2001)
y Vanderzant & Splittstoesser (1992).
Para efectos de este estudio, se determinó la tolerancia al pH ácido según el porcentaje
de sobrevivencia de la cepa seleccionada transcurrido el tiempo del ensayo usando la
ecuación (2):
𝑃𝑜𝑟𝑐𝑒𝑛𝑡𝑎𝑗𝑒 𝑑𝑒 𝑠𝑜𝑏𝑟𝑒𝑣𝑖𝑣𝑒𝑛𝑐𝑖𝑎(%) =𝑅𝑒𝑐𝑢𝑒𝑛𝑡𝑜 𝑓𝑖𝑛𝑎𝑙
𝑅𝑒𝑐𝑢𝑒𝑛𝑡𝑜 𝑖𝑛𝑖𝑐𝑖𝑎𝑙𝑥100 (𝟐)
4.6.2. Evaluación de la supervivencia de Lactobacilus fermentum en jugo de piña
Para la evaluación de la capacidad de sobrevivencia de la bacteria L. fermentum se
emplearon los jugos de piña comercialmente estériles descritos en la sección 2. Para
comprobar su esterilidad comercial al inicio del estudio, se montó un recuento total aerobio
de mesófilos según el procedimiento CITA P-SA-MM-001, el cual se basa en los métodos
descritos por Maturin & Peeler (2001), Pouch & Ito (2001) y Vanderzant & Splittstoesser
(1992). Además, se realizó un recuento de mohos y levaduras por esparcimiento según el
procedimiento del CITA, AMCITA-M007, basado en el método descrito por Pouch & Ito
(2001) y la Norma Oficial Mexicana de Bienes y Servicios: Métodos para la cuenta de mohos
y levaduras (Secretaría de Salud, 1994).
El inóculo inicial de L. fermentum se preparó siguiendo el protocolo descrito en la
Guía para la Evaluación del Desempeño de Métodos de Prueba Microbiológicos para el
Análisis de Alimentos CCAYAC-P062 (Secretaría de Salud, 2012). El microorganismo se
incubó a 35 ± 2 °C por 24 ± 2h en MRS líquido. Después, 1 mL fue inoculado en 25 mL de
MRS líquido, y se mantuvo en agitación a 35 ± 2 °C por 24 ± 2 h. Luego se procedió a
preparar crioviales en tubos Eppendorf de 1,5 mL empleando 1 mL de medio y 30 % de
glicerol, y se almacenaron a -20 °C. Para corroborar el recuento del vial en el inóculo inicial
se montó un recuento de BAL por triplicado, y se obtuvo una concentración estimada de 109
UFC/mL.
Para evaluar la sobrevivencia de la BAL seleccionada, se siguió el procedimiento
descrito por Nualkaekul & Charalampopoulos (2011). Se prepararon botellas de 100 mL de

36
los distintos jugos y se inocularon con 0,025 mL del microorganismo seleccionado, para
obtener una concentración de aproximadamente 108 UFC/mL (Lourens & Viljoen, 2001).
Los tubos de cada muestra se almacenaron a una temperatura de refrigeración (4 °C) por un
periodo de seis semanas basándose en el estudio de Nualkaekul & Charalampopoulos (2011).
Todas las semanas se realizaron mediciones de pH y de sólidos solubles del jugo; así como
del recuento total de bacterias lácticas. Para verificar que no se produjo contaminación del
jugo durante el periodo experimental, se realizó un recuento total aerobio mesófilo y de
mohos y levaduras por triplicado a muestras de cada uno de los jugos reservadas como
controles negativos durante el periodo de almacenamiento. Todas las pruebas aplicadas en el
estudio de sobrevivencia fueron realizadas por triplicado.
4.6.3. Determinación del conteo de BAL por recuento en placa
La determinación de carga celular se realizó por recuento en placa durante las seis
semanas de almacenamiento del jugo. Para ello, se prepararon diluciones seriadas de los
jugos utilizando 1 mL de muestra en 9 mL de agua peptonada estéril al 1 % y se inocularon,
por duplicado, 0,1 mL en placas con MRS sólido (Difco, Bekton & Dikinson) y se
esparcieron sobre la superficie del agar con asas de Drigalski. Las placas fueron incubadas
en condiciones anaerobias a 35 ± 1 °C por 72 ± 3 h según lo establece el procedimiento P-
SA-MM-006 del CITA (2012) basado en los métodos de Pouch & Ito (2001) y Vanderzantt
& Splittstoesser (1992).
4.7. Análisis estadístico
Para la prueba de sobrevivencia de L. fermentum en los jugos estudiados se efectuó
un análisis de varianza (ANDEVA) de dos vías para determinar el efecto de tres niveles de
jugo (pulpa, cáscara y pulpa + cáscara) y dos niveles de tiempo de almacenamiento (0
semanas y 6 semanas) sobre el recuento de L. fermentum y el pH del jugo.
Para la evaluación de las propiedades fisicoquímica se aplicó un ANDEVA de dos
vías para determinar el efecto de tres niveles de jugo (pulpa, cáscara y pulpa + cáscara) y dos
niveles de tiempo de almacenamiento (0 semanas y 6 semanas) sobre las siguientes
propiedades: sólidos totales, sólidos solubles, acidez titulable total, azúcares fermentables

37
(fructosa, glucosa y sacarosa) y ácidos orgánicos (cítrico, málico y láctico). Las diferencias
entre las medias para ambos análisis, se efectuaron por medio de una prueba de comparación
múltiple de Tuckey. Para ambos análisis se utilizó el programa JMP®Pro versión 11.0.0
(SAS Institute Inc., Cary, NC) considerando como valores significativos un valor p < 0,05.

38
V. RESULTADOS Y DISCUSIÓN
5.1. Pruebas preliminares de identificación molecular
5.1.1. Extracción de ácidos nucleicos totales
Se realizó una comparación entre los tres protocolos de extracción a los cuáles se les
aplicó RNAsa A 10 ng/ul (Thermo Scientific) utilizando como sujeto de prueba E. coli no
patógena por triplicado y se obtuvieron diferencias significativas (p < 0,0001) en la
concentración y la pureza de las extracciones. Las amplificaciones realizadas con los
iniciadores del ADNr 16 y el procedimiento descrito por Fontecha (2005) generaron bandas
muy tenues cuando se visualizaron en los geles de agarosa las extracciones (Figura 4a); las
mismas fueron cuantificadas y se determinó su pureza. Se obtuvieron concentraciones de
104,8 ± 2 ng/µL (en adelante + representa la desviación estándar) con una pureza de 1,32 ±
0,06. Se realizó un PCR de prueba; sin embargo, algunas de las muestras no amplificaron y
las bandas obtenidas fueron tenues y de muy baja calidad.
En el caso de las muestras extraídas por el protocolo de Chelex®-100 (BioRad,
Estados Unidos) no mostraron bandas distinguibles, el ADN fue cuantificado y se obtuvieron
valores de 73 ± 2 ng/uL, con una pureza de 1,30 ± 0,05; al igual que con el protocolo anterior,
se realizó un PCR de prueba, en este caso no se observó ninguna amplificación (figura no
disponible).
El tratamiento con el método de Minipreps (Birnboim & Dolly, 1979) permitió
obtener extracciones que mostraron la presencia de ADN de mayor integridad cuando fueron
visualizados en los geles de agarosa (Figura 4b). En la cuantificación se obtuvieron valores
de 332 ± 3 ng/uL, con una pureza de 1,98 ± 0,03, lo cual se vio reflejado como bandas únicas
y de mayor intensidad. El PCR mostró una amplificación en todas las muestras y se
obtuvieron bandas del tamaño esperado y algunas bandas inespecíficas.
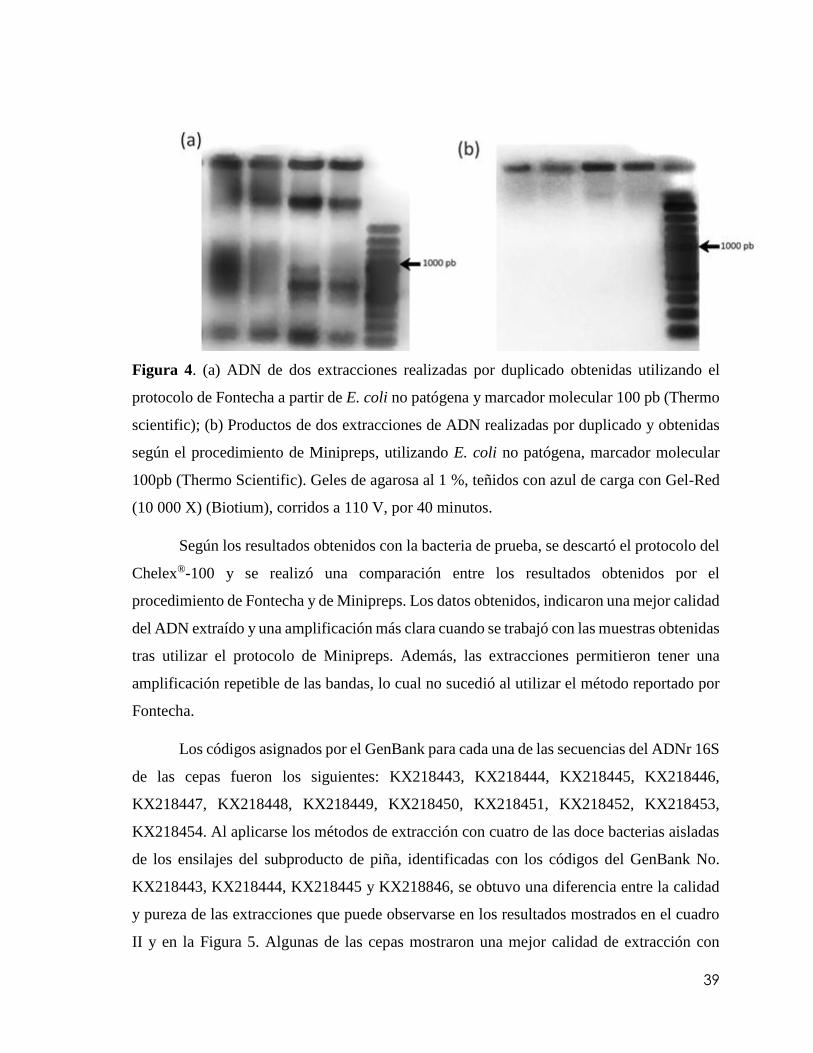
39
Figura 4. (a) ADN de dos extracciones realizadas por duplicado obtenidas utilizando el
protocolo de Fontecha a partir de E. coli no patógena y marcador molecular 100 pb (Thermo
scientific); (b) Productos de dos extracciones de ADN realizadas por duplicado y obtenidas
según el procedimiento de Minipreps, utilizando E. coli no patógena, marcador molecular
100pb (Thermo Scientific). Geles de agarosa al 1 %, teñidos con azul de carga con Gel-Red
(10 000 X) (Biotium), corridos a 110 V, por 40 minutos.
Según los resultados obtenidos con la bacteria de prueba, se descartó el protocolo del
Chelex®-100 y se realizó una comparación entre los resultados obtenidos por el
procedimiento de Fontecha y de Minipreps. Los datos obtenidos, indicaron una mejor calidad
del ADN extraído y una amplificación más clara cuando se trabajó con las muestras obtenidas
tras utilizar el protocolo de Minipreps. Además, las extracciones permitieron tener una
amplificación repetible de las bandas, lo cual no sucedió al utilizar el método reportado por
Fontecha.
Los códigos asignados por el GenBank para cada una de las secuencias del ADNr 16S
de las cepas fueron los siguientes: KX218443, KX218444, KX218445, KX218446,
KX218447, KX218448, KX218449, KX218450, KX218451, KX218452, KX218453,
KX218454. Al aplicarse los métodos de extracción con cuatro de las doce bacterias aisladas
de los ensilajes del subproducto de piña, identificadas con los códigos del GenBank No.
KX218443, KX218444, KX218445 y KX218846, se obtuvo una diferencia entre la calidad
y pureza de las extracciones que puede observarse en los resultados mostrados en el cuadro
II y en la Figura 5. Algunas de las cepas mostraron una mejor calidad de extracción con

40
respecto al material genético de las demás, que puede deberse a la variabilidad de estructura
y composición de las membranas celulares, lo cual influye en la capacidad y eficacia del
método para lograr la ruptura y liberación del material genético (Pesvner, 2009), y por ello,
se obtuvieron mejores resultados al realizar la amplificación de las bandas (Figura 6); sin
embargo, sí se observaron bandas inespecíficas lo cual implica que se requiere de una
optimización de las condiciones de PCR.
Figura 5. (a) Productos de extracción de ADN obtenidos por protocolo de Fontecha, orden
del gel: cepas KX218443, KX218443, KX218444, KX218444, KX218445, KX218445,
KX218846, KX218846, marcador molecular 100 pb (Thermo scientific); (b) Productos de
extracción obtenidos por protocolo de Minipreps, orden del gel: KX218443, KX218444,
KX218445 y KX218446 por duplicado, marcador molecular 100pb (Thermo Scientific).
Geles de agarosa al 1 %, teñidos con azul de carga con Gel-Red (10 000 X) (Biotium),
corridos a 110 V, por 40 minutos.
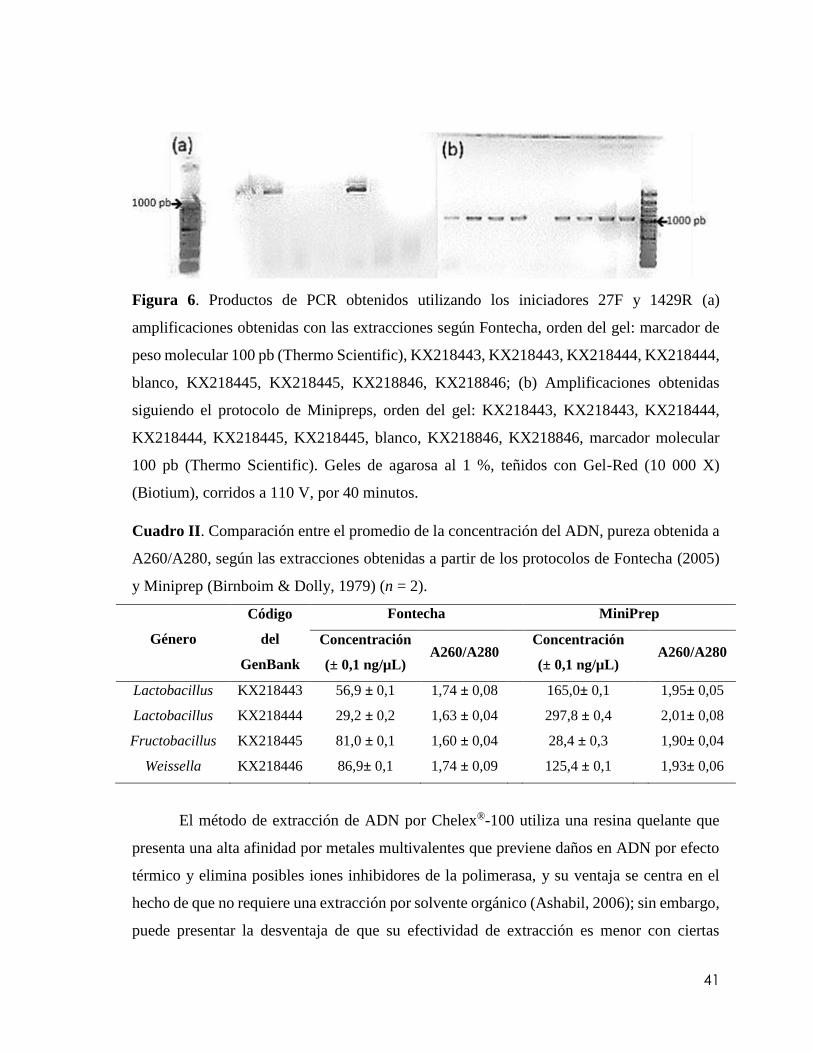
41
Figura 6. Productos de PCR obtenidos utilizando los iniciadores 27F y 1429R (a)
amplificaciones obtenidas con las extracciones según Fontecha, orden del gel: marcador de
peso molecular 100 pb (Thermo Scientific), KX218443, KX218443, KX218444, KX218444,
blanco, KX218445, KX218445, KX218846, KX218846; (b) Amplificaciones obtenidas
siguiendo el protocolo de Minipreps, orden del gel: KX218443, KX218443, KX218444,
KX218444, KX218445, KX218445, blanco, KX218846, KX218846, marcador molecular
100 pb (Thermo Scientific). Geles de agarosa al 1 %, teñidos con Gel-Red (10 000 X)
(Biotium), corridos a 110 V, por 40 minutos.
Cuadro II. Comparación entre el promedio de la concentración del ADN, pureza obtenida a
A260/A280, según las extracciones obtenidas a partir de los protocolos de Fontecha (2005)
y Miniprep (Birnboim & Dolly, 1979) (n = 2).
Género
Código
del
GenBank
Fontecha MiniPrep
Concentración
(± 0,1 ng/µL) A260/A280
Concentración
(± 0,1 ng/µL) A260/A280
Lactobacillus KX218443 56,9 ± 0,1 1,74 ± 0,08 165,0± 0,1 1,95± 0,05
Lactobacillus KX218444 29,2 ± 0,2 1,63 ± 0,04 297,8 ± 0,4 2,01± 0,08
Fructobacillus KX218445 81,0 ± 0,1 1,60 ± 0,04 28,4 ± 0,3 1,90± 0,04
Weissella KX218446 86,9± 0,1 1,74 ± 0,09 125,4 ± 0,1 1,93± 0,06
El método de extracción de ADN por Chelex®-100 utiliza una resina quelante que
presenta una alta afinidad por metales multivalentes que previene daños en ADN por efecto
térmico y elimina posibles iones inhibidores de la polimerasa, y su ventaja se centra en el
hecho de que no requiere una extracción por solvente orgánico (Ashabil, 2006); sin embargo,
puede presentar la desventaja de que su efectividad de extracción es menor con ciertas

42
muestras y los remanentes de la resina tienen cierto efecto inhibitorio en el PCR (NFSTC,
2015), lo cual podría ser la causa de que las muestras extraídas por el este método no
presentaran amplificación alguna en el PCR. Sumado a esto, es posible que el método no
fuera apto para las bacterias estudiadas, lo cual generó una extracción de ADN de muy baja
calidad.
El protocolo de Fontecha (2005) se basa en un método de extracción por fenol-
cloroformo isoamílico (PCI) el cual es un método tradicional para la extracción del material
nuclear; en este caso la lisis se realiza con la aplicación de un detergente (SDS al 1 %) y
lisozima que favorece la ruptura de las paredes celulares y el uso de proteinasa k que ayuda
en la degradación de enzimas y proteínas presentes obteniéndose un ADN de buena calidad
(Moore et al., 2004). La lisozima y la proteinasa K son las enzimas más empleadas
comúnmente; sin embargo, su efectividad en la ruptura del tejido celular no es generalizada,
por lo que puede no ser efectiva para cualquier organismo (Moore et al., 2004). Esto podría
explicar las diferencias en los resultados de extracción de ADN.
Por otra parte, el protocolo de Minipreps se basa una lisis básica con hidróxido de
sodio (NaOH) y dodecilsulfato sódico (SDS) que permite la desnaturalización del ADN
cromosomal, plásmido y proteínas respectivamente (Engebrecht et al., 1991), que genera un
ADN de muy buena calidad. El uso del medio alcalino favorece una mejor ruptura del tejido
celular, por lo que el material obtenido presentó una mejor calidad que los obtenidos por
otros métodos.
5.1.2. Estandarización del protocolo de PCR
Para las pruebas de selección del protocolo de extracción se trabajó con las
condiciones de PCR descritas según Fontecha (2005), donde se refieren a los iniciadores para
ADNr 16S; sin embargo, es importante recalcar que no todas las condiciones son aplicables
para toda reacción de PCR, y se debe de ajustar su concentración, temperatura y tiempos para
mejorar la eficiencia de la amplificación (Applied Biosystems, 2006), lo cual debe lograrse
mediante pruebas de optimización. Dado lo anterior, se realizaron pruebas iniciales para la
estandarización del PCR utilizando los iniciadores 27 F y 1429 R (Thomas, 2004). Se trabajó
un volumen final de 50 µL por reacción, y una concentración de 50 ng/µL de ADN. El PCR

43
fue desarrollado utilizando un anillamiento a 45 °C. Sin embargo, no fue posible observar
ninguna amplificación. Dado lo anterior se realizaron diferentes variaciones a las condiciones
del PCR, utilizando un rango de 20-50 ng/µL de ADN. Los resultados obtenidos indican una
banda de mayor intensidad y claridad al adicionar 20 ng/µL de ADN, por lo que se estableció
dicha concentración como la elegida para trabajar; a su vez, se ajustaron los volúmenes de
reacción para tener un volumen de 25 µL por reacción.
Para terminar de optimizar el protocolo de amplificación, se procedió a evaluar la
temperatura de anillamiento. Se utilizó un gradiente de temperaturas entre 45 °C y 58 °C. Tal
y como se observa en la Figura 7b, las mejores amplificaciones se obtuvieron utilizando una
temperatura de 55 °C en la cual se observó la amplificación de bandas más nítidas, concretas
y únicas a aproximadamente 1200 pb, en comparación con las bandas observadas en la Figura
7a. La temperatura de anillamiento se basa en la temperatura de fusión de los
oligonucleótidos o iniciadores seleccionados para la amplificación; la presencia de bandas
no deseadas implica una inespecificidad en las condiciones aplicadas, por lo que se
recomienda el aumento de temperatura ya que el aumento de ésta implica una mayor
especificidad (Applied Biosystems, 2006).
Figura 7. (a) Productos de PCR amplificados con anillamiento de 45 °C, orden del gel:
marcador molecular de 100 pb (Thermo Scientific), KX218443, KX218443, KX218444,
KX218444, KX218445, KX218445, KX218846, KX218846. (b) productos de PCR
amplificados con un anillamiento de 55 °C: KX218443, KX218443, KX218444, KX218444,
KX218445, KX218445, KX218846, KX218846, marcador molecular 100 pb (Thermo

44
Scientific). Geles de agarosa al 1%, teñidos con azul de carga con Gel-Red (Biotium) (10
000 X) corridos a 110 V, 40 minutos.
5.2. Identificación molecular de las BAL utilizando el ADNr 16
Se identificaron un total de tres géneros diferentes, considerando los 12 aislamientos
de BAL: Lactobacillus (10), Fructobacillus (1) y Weissella (1). El árbol filogenético
construido con NJ (Figura 8), muestra que las cepas con los número de accesión del
GenBank: KX218447, KX218448, KX218449, KX218451, KX218453, KX218452 y
KX218450 fueron identificadas como individuos pertenecientes al género Lactobacillus
spp. y mostraron relación con las cepas GI: 1848053; GI: 48843512; GI: 28194135; GI:
1848052T, identificadas como L. casei y las cepas GI: 886849520; GI: 1848056; GI:
1808583, previamente descritas como L. paracasei. Mientras que la cepa KX218446
agrupó con especies identificadas como Weissella fabaria (GI: 259052530) y W.
ghanensis (GI: 157100985) respectivamente. La cepa GenBank no. KX218445 agrupó
con miembros de las cepas Fructobacillus troapeoli (GI: 296785023). A su vez, las cepas
con los números de accesión KX218443 y KX218444 están claramente relacionadas al
grupo de L. fermentum (GI: 157907360, GI: 57907348, GI: 94490347 y GI: 32487282),
mientras que la cepa con el número de accesión KX218447 agrupa con las cepas de L.
spicheri (GI: 329025383; GI: 38602388; GI: 268527436 y GI: 38602388). Como se
observa en la Figura 8, el marcador del ADNr 16 no permitió una clara diferenciación
entre especies tanto en el caso de L. casei y L. paracasei como en el caso e W. fabaria y
W. ghanensis.
Una de las principales limitantes del método, es que cuando dos bacterias poseen una
secuencia casi idéntica del gen, la técnica no es suficiente para distinguir entre ellas (Woo
et al., 2008); además, las regiones hipervariables del ADNr 16s exhiben diferentes grados
de variación y por ende una única región no basta para la distinción de todas las bacterias;
esto puede ser lo observado para L. casei y L. paracasei, para las cuales, el segmento del
ADNr 16s no es suficiente para lograr una distinción clara entre las especies. Dicho
inconveniente puede solventarse por medio de la selección de otro segmento más

45
específico del ADN, utilizando marcadores como los MLST (Multilocus sequence typing)
que permiten aislar especies microbianas a partir de la secuenciación de fragmentos de
genes seleccionados. Esto ha permitido una identificación del microorganismo con
genotipos altamente relacionados, lo que implica que es posible identificar todas las
variaciones con una mayor especificidad (Larsen et al., 2012; Maiden et al., 2013
Vázquez & Berrón, 2004). Otra alternativa, es utilizar el gen de Girasa (gyraA o gyraB)
el cual codifica para una subunidad de proteína del ADN girasa con lo cual es posible
determinar diferencias entre especies muy cercanamente relacionadas como es el caso de
L. casei y L. paracasei. (Wang et al., 2007; Balas, 2001; Bonasera et al., 2014).
Las bacterias encontradas coinciden con varias de las cepas reportadas en la
literatura como flora normal de la piña y de otras frutas y vegetales inclusive. Los
microorganismos pertenecientes al género Lactobacillus spp. corresponden al grupo
mayoritario de las BAL y se han reportado muchas especies, entre ellas, las encontradas
en este estudio, en productos derivados de las frutas, incluida la piña (DiCagno et al.,
2010; Uyen, 2013); respecto a la presencia de Fructobacillus tropaeoli, se encuentra que
estas bacterias son comunes en matrices en las cuales se tiene un alto contenido de
fructosa como es el caso de flores, frutas y fermentados de fruta; también, se han
identificado en sistemas digestivos de organismos que consumen fructosa, lo cual sugiere
que se encuentran presentes en cualquier medio en el que el contenido de este azúcar sea
importante, lo cual coincide con el perfil de la piña (Endo, 2012); por otro lado, Weissella
spp. es un género poco común en frutas, y es más comúnmente aislado en vegetales como
zanahorias, coles, entre otras, aunque también se han identificado en frutas como el
banano (Torino et al. 2015). Originalmente se le consideró un miembro del género
Leuconostoc spp. y al igual que estas se caracteriza por la capacidad de producir
dextranos (Collins et al., 1993; Srinivas & Padma, 2015); dado a que la piña presenta un
contenido importante de sacarosa, no es raro encontrarla en esta matriz.
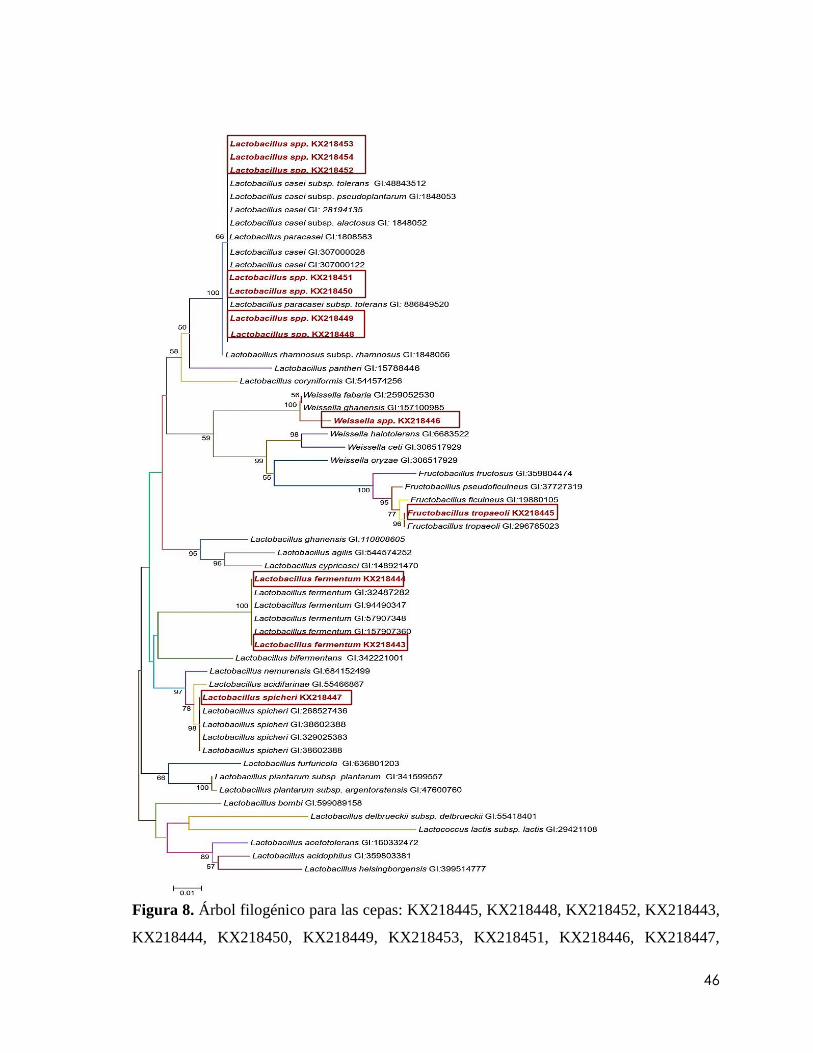
46
Figura 8. Árbol filogénico para las cepas: KX218445, KX218448, KX218452, KX218443,
KX218444, KX218450, KX218449, KX218453, KX218451, KX218446, KX218447,

47
KX218454. Los árboles filogenéticos se elaboraron utilizando el programa
Molecular Evolutionary Genetics Analysis (MEGA) versión 3.1 (Kumar et al., 2004). La
robustez de las ramas internas se estimó por medio de análisis de “bootstrap” con 2000
réplicas. Los análisis filogenéticos fueron comparados con los métodos de distancia, el
vecino más cercano (Neighbor Joining), donde las secuencias de este estudio se denotan con
otro color, y solamente se muestran los valores de las ramas mayores al 50 %.
Las secuencias de las cepas identificadas se compararon por una alineación de pares
(Pairwise Alignment) el cual consiste en alinear dos secuencias para obtener el máximo nivel
de identidad de forma que sea posible evaluar el grado de identidad y homología entre dos
moléculas (Pesvner, 2009). Al realizar la comparación entre cepas de la misma especie y
género aisladas en este estudio y cepas de referencias más cercanas, se determinó que las
cepas identificadas con los códigos del GenBank no. KX218448, KX218449, KX218451, y
KX218450 muestran un 99 % de identidad al comparar pares entre ellas y un 99 % cuando
se comparan con los individuos L. casei (GI: 307000122) y L. paracasei subsp. tolerans (GI:
886849520 y GI: 48843512), por otra parte, la cepa KX218452 mostró un porcentaje de
identidad de 97 % ; lo cual podría implicar que dicha cepa es un individuo distinto de L. casei
y L. paracasei, sin embargo es necesario corroborar con marcadores más específicos para
poder hacer dicha afirmación. La cepa KX218446 muestra un 99 % de identidad con ambas
especies de Weissella sp: W. fabaria (GI: 259052530) y W. ghanensis (GI: 157100985),
respectivamente. Las cepas de L. fermentum KX218443 y KX218444 presentan un 99 % de
identidad entre ellas y el mismo valor con respecto a la cepa de referencia L. fermentum (GI:
32487282). Del mismo modo, la cepa KX218445 presenta un 99 % de identidad con
Fructobacillus tropaeoli (GI: 296785023). La cepa KX218447 muestra un 99 % de identidad
con respecto a L. spicheri (GI: 38602388). Las relaciones entre pares de individuos se pueden
observar con mayor claridad en la matriz (Figura 9) en la cual se identifican por color los
porcentajes de identidad entre los pares de cepas comparados.
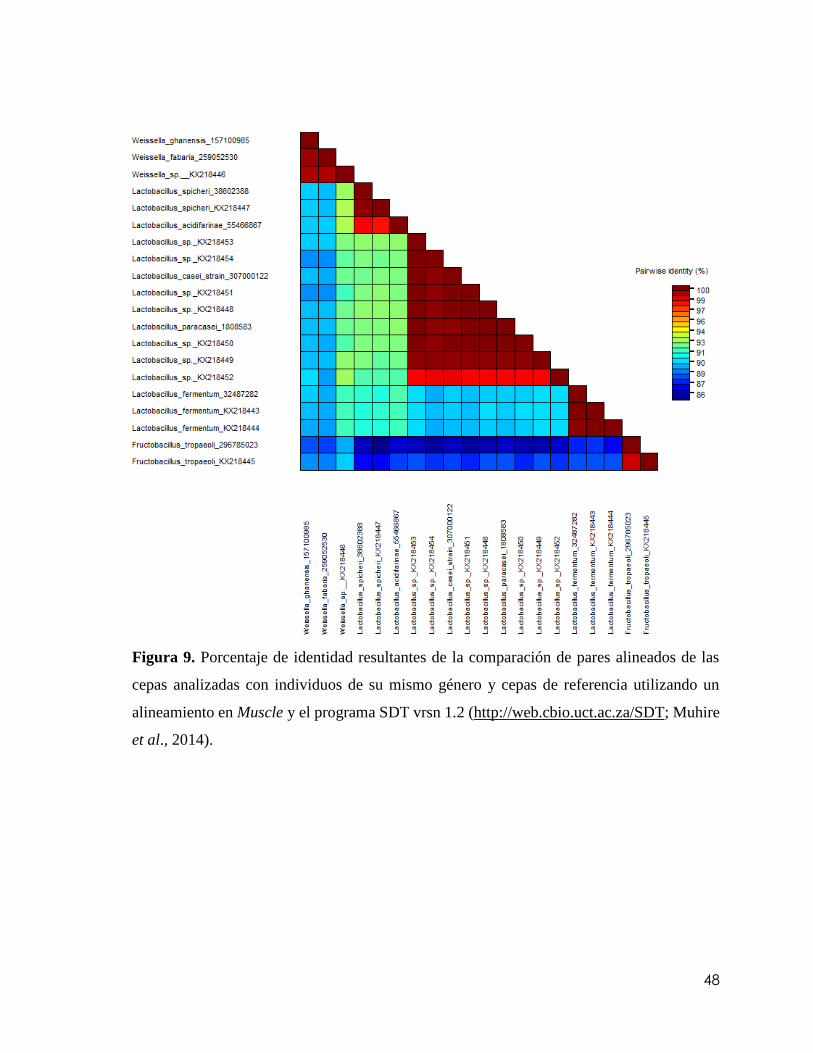
48
Figura 9. Porcentaje de identidad resultantes de la comparación de pares alineados de las
cepas analizadas con individuos de su mismo género y cepas de referencia utilizando un
alineamiento en Muscle y el programa SDT vrsn 1.2 (http://web.cbio.uct.ac.za/SDT; Muhire
et al., 2014).

49
5.3. Sobrevivencia de BAL a un pH de 2,0
Las 12 BAL aisladas e identificadas fueron sometidas a un ensayo de tolerancia ácida
a un pH de 2,0, que simula el ambiente gástrico durante el proceso de digestión, y que se
realiza como una prueba preliminar de evaluación de su potencial como microorganismo
probiótico. En general, se observa una reducción en el recuento de los diferentes
microorganismos durante el periodo de incubación en el medio acidificado. En la muestras
control (pH de 6,0) no se observa una variación importante en los recuentos a las distintas
horas. Por otro lado, como se muestra en la Figura 10, se observó que la cepa Fructobacillus
tropaeoli (KX218445) presentó el mayor porcentaje de sobrevivencia con un valor de 95,8 ±
0,4 %, seguido por dos cepas de Lactobacillus spp.: KX218448 y KX218452 que presentaron
valores de 95,5 ± 0,9 %, y 93,8 ± 0,1%, respectivamente. Mientras que L. fermentum
(KX218443; KX218444) presentaron un porcentaje de sobrevivencia de 92,2 ± 0,6 % y 90,2
± 0,6 %. Las cepas que mostraron una menor resistencia se identificaron como Lactobacillus
spicheri (KX218447), con un valor de 70,3 ± 0,7 %, seguida por Weisella fabaria
(KX218446) con una sobrevivencia de 77,5 ± 2,7 % y Lactobacillus spp. (KX218454) con
una sobrevivencia de solo el 45,0 ± 0,3 %. Esta última, fue la que presentó la mayor reducción
logarítmica después de las tres horas de incubación a un pH de 2,0.
En general, se observa una sobrevivencia mayor al 80,0 % en las cepas estudiadas en
el ensayo, con excepción de las cepas KX218446, KX218447 y KX218454. Con algunas
excepciones, los mejores resultados se obtuvieron con individuos que previamente se
identificaron como pertenecientes los géneros Lactobacillus spp. y Fructobacillus tropaeoli,
lo cual, concuerda con lo reportado en la literatura (Tripathi & Giri, 2014; Kelanne, 2012).
Estos resultados son determinantes para la selección del microorganismo que se emplearía
para las pruebas de sobrevivencia durante el almacenamiento, ya que es un indicativo de su
capacidad de sobrevivencia a pH similares al que se presenta en el ambiente gástrico (Lin et
al., 2006). La cepa que se seleccionó para inocular los tres tipos de jugos de piña, con base a
los resultados obtenidos en las pruebas de identificación molecular y de resistencia a un pH
de 2,0, fue L. fermentum. Los miembros del género Lactobacillus son usualmente
consideradas bacterias intrínsecamente tolerantes al ácido (Tannock, 2004).
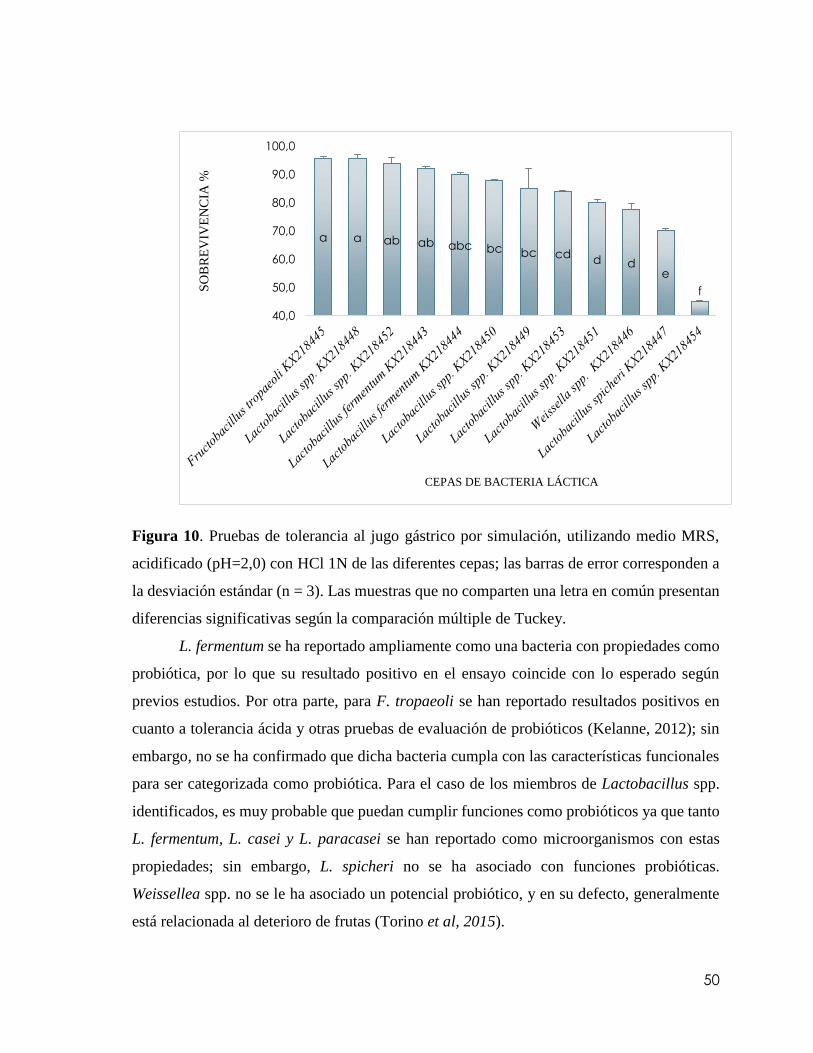
50
Figura 10. Pruebas de tolerancia al jugo gástrico por simulación, utilizando medio MRS,
acidificado (pH=2,0) con HCl 1N de las diferentes cepas; las barras de error corresponden a
la desviación estándar (n = 3). Las muestras que no comparten una letra en común presentan
diferencias significativas según la comparación múltiple de Tuckey.
L. fermentum se ha reportado ampliamente como una bacteria con propiedades como
probiótica, por lo que su resultado positivo en el ensayo coincide con lo esperado según
previos estudios. Por otra parte, para F. tropaeoli se han reportado resultados positivos en
cuanto a tolerancia ácida y otras pruebas de evaluación de probióticos (Kelanne, 2012); sin
embargo, no se ha confirmado que dicha bacteria cumpla con las características funcionales
para ser categorizada como probiótica. Para el caso de los miembros de Lactobacillus spp.
identificados, es muy probable que puedan cumplir funciones como probióticos ya que tanto
L. fermentum, L. casei y L. paracasei se han reportado como microorganismos con estas
propiedades; sin embargo, L. spicheri no se ha asociado con funciones probióticas.
Weissellea spp. no se le ha asociado un potencial probiótico, y en su defecto, generalmente
está relacionada al deterioro de frutas (Torino et al, 2015).
a a ab ab abc bc bc cdd d
e
f
40,0
50,0
60,0
70,0
80,0
90,0
100,0
SO
BR
EV
IVE
NC
IA %
CEPAS DE BACTERIA LÁCTICA

51
5.4. Caracterización fisicoquímica de los jugos de piña durante el almacenamiento
Para evaluar el efecto del tiempo de almacenamiento (6 semanas en total) en
refrigeración a 4°C sobre la las características fisicoquímicas de tres jugos de piña inoculados
con L. fermentum, se aplicó un diseño factorial completo con tres niveles para la variable
“tipo de jugo” (pulpa, pulpa + cáscara y cáscara) y dos niveles para el tiempo de
almacenamiento (semana 0 y semana 6). Se realizaron las determinaciones de las propiedades
fisicoquímicas para los tres tipos de jugo de piña: de pulpa, mezcla de pulpa y cáscara a una
razón 80:20 y de cáscara tomando una muestra al inicio del experimento y luego del periodo
de las seis semanas de almacenamiento a 4 °C tras la inoculación. El análisis de varianza
aplicado a los resultados muestra que los jugos presentaron diferencias significativas (p <
0,05) en sus propiedades fisicoquímicas después del período almacenamiento, con excepción
del contenido de sólidos, y que estas diferencias resultaron dependientes del tipo de jugo
inoculado (Cuadro III).
El contenido de sólidos totales no mostró variaciones significativas (p > 0,05) entre
los jugos ni entre los tiempos de muestreo. En general, para los tres jugos y tiempos
analizados, los valores se mantuvieron entre 12 y 14 %. Este resultado es consistente con lo
esperado según lo reportado en la literatura para los jugos de piña, ya que se estima un
contenido de sólidos totales entre 12 y 19 % (Acevedo et al., 2009). Es importante mencionar,
que para el porcentaje de sólidos totales, de un 2 a un 3 % corresponde a fibra soluble; a su
vez, los mismos valores pueden presentar variaciones dependiendo de la naturaleza de la
materia prima (Acevedo et al., 2009). Sin embargo, se observó que la variación en este
parámetro, no se vio afectada de forma significativa (p > 0,005) por el tipo del jugo ni el
tiempo de almacenamiento evaluado posterior a la inoculación de las muestras.
Con respecto a la acidez titulable total, expresada como ácido cítrico, se determinó
que el contenido presenta diferencias significativas dependiendo del tipo de jugo (p < 0,0001),
donde la comparación de medias indica que la acidez de los jugos de pulpa + cáscara y pulpa
pura no es significativamente distinta entre ellos, pero sí difiere con respecto a la acidez del
jugo de cáscara. Según lo reportado en la literatura, los valores de acidez titulable esperables
para el jugo de piña rondan entre los 0,7 - 1,6 g de ácido cítrico/100 mL de jugo (HANNA
Instruments, 2015); sin embargo, estos valores pueden ser menores (Frazier et al., 1995)

52
dependiendo de la materia prima dado a que están sujetos a la localidad, clima, temporada y
variedad y grado de madurez de la fruta (Zambrano & Castellano, 1997).
La cuantificación de los sólidos solubles (°Brix) para los tres jugos mostró una
interacción significativa entre el tiempo de almacenamiento y tipo de jugo (p = 0,006). El
análisis de comparación de medias indica que los promedios distintos corresponden a los
obtenidos para pulpa y cáscara después de las seis semanas de almacenamiento en el cual se
observó una disminución del valor con respecto al inicial; este resultado puede explicarse
debido a la presencia de microorganismos metabólicamente activos, presentes en la muestra.
Asimismo, al analizar el perfil de azúcares fermentables, el análisis estadístico determinó que
se presentan interacciones significativas entre los tipos de jugo y el tiempo de
almacenamiento para los resultados de contenido de fructuosa (p < 0,0001), glucosa (p =
0,004) y la sacarosa (p < 0,0001) (Cuadro III). Los resultados de la cuantificación de azúcares
para los distintos jugos mostraron diferencias significativas entre los tipos de jugos; a su vez,
los valores obtenidos concuerdan con las cifras estimadas para la piña (Haruenkit, 2004; Siti
et al., 2013). El azúcar predominante corresponde a la sacarosa, seguido por un fracción
relativamente equivalente de fructosa y glucosa tanto para los jugos elaborados a partir de
pulpa como los de cáscara lo cual coincide con los resultados obtenidos por Haruenkit (2004)
y Siti et al. (2013).
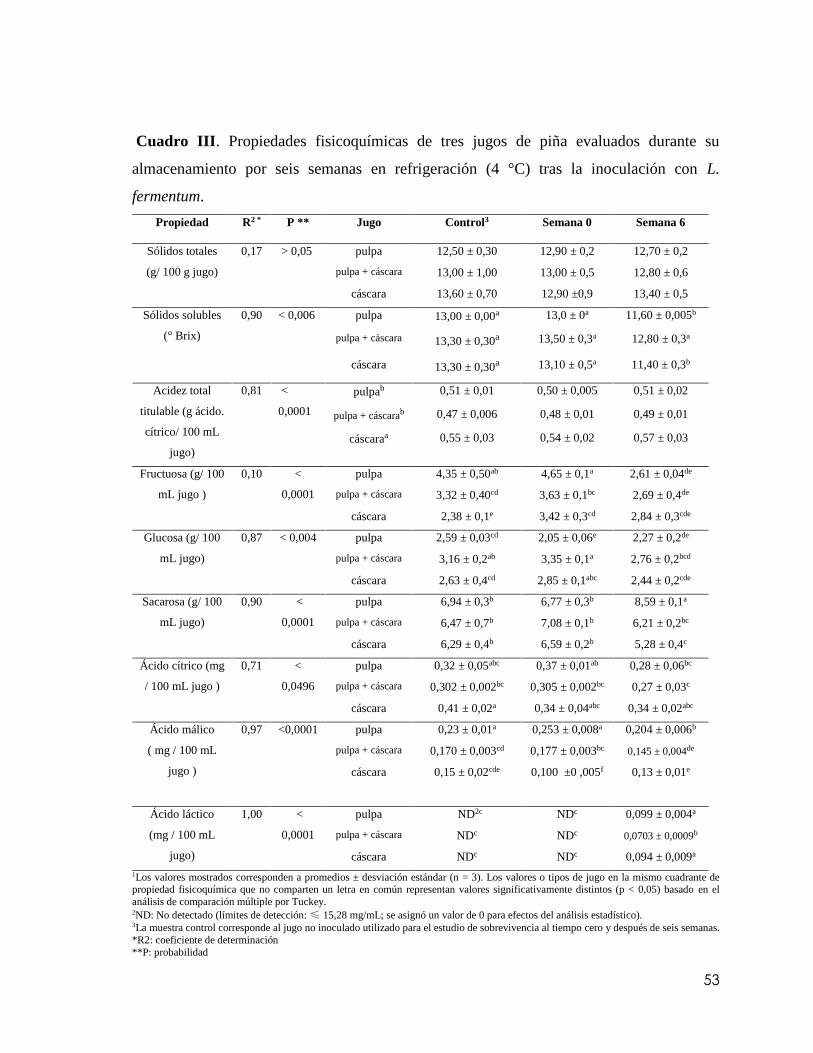
53
Cuadro III. Propiedades fisicoquímicas de tres jugos de piña evaluados durante su
almacenamiento por seis semanas en refrigeración (4 °C) tras la inoculación con L.
fermentum.
Propiedad R2 * P ** Jugo Control3 Semana 0 Semana 6
Sólidos totales
(g/ 100 g jugo)
0,17 > 0,05 pulpa 12,50 ± 0,30 12,90 ± 0,2 12,70 ± 0,2
pulpa + cáscara 13,00 ± 1,00 13,00 ± 0,5 12,80 ± 0,6
cáscara 13,60 ± 0,70 12,90 ±0,9 13,40 ± 0,5
Sólidos solubles
(° Brix)
0,90 < 0,006 pulpa 13,00 ± 0,00a 13,0 ± 0a 11,60 ± 0,005b
pulpa + cáscara 13,30 ± 0,30a 13,50 ± 0,3a 12,80 ± 0,3a
cáscara 13,30 ± 0,30a 13,10 ± 0,5a 11,40 ± 0,3b
Acidez total
titulable (g ácido.
cítrico/ 100 mL
jugo)
0,81 <
0,0001
pulpab 0,51 ± 0,01 0,50 ± 0,005 0,51 ± 0,02
pulpa + cáscarab 0,47 ± 0,006 0,48 ± 0,01 0,49 ± 0,01
cáscaraa 0,55 ± 0,03 0,54 ± 0,02 0,57 ± 0,03
Fructuosa (g/ 100
mL jugo )
0,10 <
0,0001
pulpa 4,35 ± 0,50ab 4,65 ± 0,1a 2,61 ± 0,04de
pulpa + cáscara 3,32 ± 0,40cd 3,63 ± 0,1bc 2,69 ± 0,4de
cáscara 2,38 ± 0,1e 3,42 ± 0,3cd 2,84 ± 0,3cde
Glucosa (g/ 100
mL jugo)
0,87 < 0,004 pulpa 2,59 ± 0,03cd 2,05 ± 0,06e 2,27 ± 0,2de
pulpa + cáscara 3,16 ± 0,2ab 3,35 ± 0,1a 2,76 ± 0,2bcd
cáscara 2,63 ± 0,4cd 2,85 ± 0,1abc 2,44 ± 0,2cde
Sacarosa (g/ 100
mL jugo)
0,90 <
0,0001
pulpa 6,94 ± 0,3b 6,77 ± 0,3b 8,59 ± 0,1a
pulpa + cáscara 6,47 ± 0,7b 7,08 ± 0,1b 6,21 ± 0,2bc
cáscara 6,29 ± 0,4b 6,59 ± 0,2b 5,28 ± 0,4c
Ácido cítrico (mg
/ 100 mL jugo )
0,71 <
0,0496
pulpa 0,32 ± 0,05abc 0,37 ± 0,01ab 0,28 ± 0,06bc
pulpa + cáscara 0,302 ± 0,002bc 0,305 ± 0,002bc 0,27 ± 0,03c
cáscara 0,41 ± 0,02a 0,34 ± 0,04abc 0,34 ± 0,02abc
Ácido málico
( mg / 100 mL
jugo )
0,97 <0,0001 pulpa 0,23 ± 0,01a 0,253 ± 0,008a 0,204 ± 0,006b
pulpa + cáscara 0,170 ± 0,003cd 0,177 ± 0,003bc 0,145 ± 0,004de
cáscara 0,15 ± 0,02cde
0,100 ±0 ,005f 0,13 ± 0,01e
Ácido láctico
(mg / 100 mL
jugo)
1,00 <
0,0001
pulpa ND2c NDc 0,099 ± 0,004a
pulpa + cáscara NDc NDc 0,0703 ± 0,0009b
cáscara NDc NDc 0,094 ± 0,009a
1Los valores mostrados corresponden a promedios ± desviación estándar (n = 3). Los valores o tipos de jugo en la mismo cuadrante de propiedad fisicoquímica que no comparten un letra en común representan valores significativamente distintos (p < 0,05) basado en el
análisis de comparación múltiple por Tuckey. 2ND: No detectado (límites de detección: ≤ 15,28 mg/mL; se asignó un valor de 0 para efectos del análisis estadístico). 3La muestra control corresponde al jugo no inoculado utilizado para el estudio de sobrevivencia al tiempo cero y después de seis semanas.
*R2: coeficiente de determinación
**P: probabilidad

54
Los resultados tras el periodo de almacenamiento muestran una tendencia general a
la disminución del contenido de azúcares luego de las seis semanas de almacenamiento. Para
la fructuosa, se presentaron diferencias significativas al comparar el tiempo cero con las seis
semanas luego de inoculado con L. fermentum. Esta diferencia fue mayor entre el jugo de
pulpa y el jugo de pulpa + cáscara. Para el caso del jugo de cáscara puro no se observó
diferencia entre los dos tiempos comparados. Los resultados de la glucosa, también presentan
diferencias significativas entre tiempo cero y seis semanas; en este caso, se observa que las
diferencias significativas debido al período de almacenamiento sólo se dan para el jugo de
cáscara + pulpa; lo que puede indicar que en esta matriz, el microorganismo metabolizó dicho
azúcar en mayor medida. Por su parte, para la sacarosa se observan diferencias significativas
entre los jugos de pulpa y el jugo de cáscara. Nótese que para el caso del jugo de pulpa, el
resultado obtenido no coincide con lo esperado, puesto que el contenido de sacarosa mostró
un leve aumento en lugar de disminuir. En condiciones normales, se esperaría una reducción
del contenido del azúcar durante el almacenamiento debido a la actividad metabólica del
microorganismo inoculado; sin embargo, la variación de dicho análisis bien pudo deberse a
errores ocurridos durante el análisis de este parámetro o bien por la variabilidad propia del
ensayo dado a que la tendencia normal que se ha encontrado en otros estudios es la reducción
(Adil et al., 2014).
El perfil de ácidos orgánicos presenta variaciones para los tres ácidos analizados
(cítrico, málico y láctico). En estos tres parámetros, se encontraron interacciones
significativas entre el tipo de jugo y el tiempo de almacenamiento: cítrico (p = 0,0496),
málico (p < 0,0001) y láctico (p < 0,0001). El ácido cítrico, en la semana cero, no presentó
diferencias significativas entre los tres jugos; sin embargo, para la semana seis, este ácido en
el jugo de cáscara presentó diferencias significativas, cuando se comparó con el presente en
el jugo de pulpa y el jugo de pulpa + cáscara. Los dos primeros mostraron una leve reducción
del contenido inicial de 0,09 ± 0,07 mg/ 100 mL para el jugo de pulpa y 0,03 ± 0,03 mg/ 100
mL en el jugo de pulpa y cáscara, mientras que el tercero se mantuvo prácticamente constante;
dicha disminución puede sugerir que el microorganismo consumió el ácido cítrico en su
metabolismo lo cual coincide con lo reportado por Pardo (2003). El ácido málico, presentó
diferencias significativas entre tiempo cero y seis semanas en los tres jugos. Los dos jugos

55
con contenido de pulpa, presentaron una disminución importante del ácido con respecto a la
cuantificación inicial (semana 0), mientras que para el jugo de cáscara la disminución se dio
en menor medida. Es importante mencionar, que la medición del perfil de ácidos orgánicos
es utilizada como índice de autenticidad en productos frutales, debido a su menor
susceptibilidad, con algunas excepciones, al cambio durante el procesamiento y
almacenamiento en comparación con otros componentes de la fruta (Cámara et al., 1994;
Sun, 2008). Sin embargo, en este estudio sí se observaron cambios significativos dentro del
perfil de ácidos orgánicos de los jugos evaluados, lo cual puede ser atribuido a la presencia
del microorganismo inoculado, que puede degradar ácido cítrico y málico dado a que al igual
que otros miembros del género Lactobacillus spp., L. fermentum pueden metabolizarlos por
medio de la formación de oxalacetato y por la enzima maloláctica, respectivamente (Pardo,
2003). En este caso, la disminución significativa se presenta para los tipos de jugo y el tiempo
de almacenamiento. La ausencia de ácido láctico al inicio (semana cero) coincide con lo
esperado dado que de acuerdo a lo reportado en la literatura, los ácidos orgánicos que se
encuentra normalmente en la piña corresponden únicamente al ácido cítrico en mayor medida
y al málico en un menor porcentaje (Haruenkit, 2004). Por lo tanto, otra evidencia de la
activad metabólica de L. fermentum, es la aparición de ácido láctico en los jugos analizados
en la semana seis. El ácido láctico es un metabolito típico de L. fermentum (Kandler, 1983)
por lo que su presencia es un indicador importante de la actividad de la bacteria. En general,
los tres jugos presentan una cantidad bastante pequeña de ácido láctico, y no se encontraron
diferencias significativas entre el contenido de ácido láctico en jugo de pulpa y cáscara en la
semana seis; sin embargo, sí se encuentra, de forma significativa, un menor contenido de este
ácido en el jugo de pulpa + cáscara.
5.5. Evaluación de la sobrevivencia de la cepa de L. fermentum en jugos de piña durante
su almacenamiento a 4 °C.
Para evaluar el efecto del tiempo de almacenamiento en refrigeración sobre la
sobrevivencia de L. fermentum, se aplicó un diseño factorial completo con tres niveles para
tipo de jugo (pulpa, pulpa + cáscara y cáscara) y siete niveles para el tiempo de
almacenamiento (de semana 0 a semana 6). La población del microorganismo inoculado

56
corresponde a la variable respuesta. Se aplicó un análisis de variancia de dos vías a los
resultados obtenidos, y el modelo factorial resultante presentó un coeficiente de
determinación (R2) de 0,97, mientras que la interacción entre los factores nominales tipo de
jugo y tiempo de almacenamiento resultó significativa (p < 0,0008). El valor de coeficiente
de determinación indica que el modelo presentó un ajuste relativamente bueno y por ende se
determinó que el comportamiento la población de L. fermentum depende de la interacción
entre el tipo de jugo y el tiempo de almacenamiento (Figura 11a).
Las curvas poblacionales en los distintos jugos muestran una tendencia general de
reducción a través del tiempo. Puede observarse que a partir de la primera semana de
almacenamiento hay una menor sobrevivencia de la bacteria en los jugos de cáscara y pulpa-
cáscara (Figura 11a). En el caso del jugo de pulpa, a partir de la semana tres, se observa una
ligera estabilización de los recuentos para este jugo, y ya para la semana seis, la población
viable aún se mantenía en el orden de los seis logaritmos. Este resultado es muy importante,
pues la concentración de la cepa seleccionada se encuentra dentro del valor logarítmico
aceptado, para que un microorganismo sea considerado como probiótico (Lourens & Viljoen,
2001).
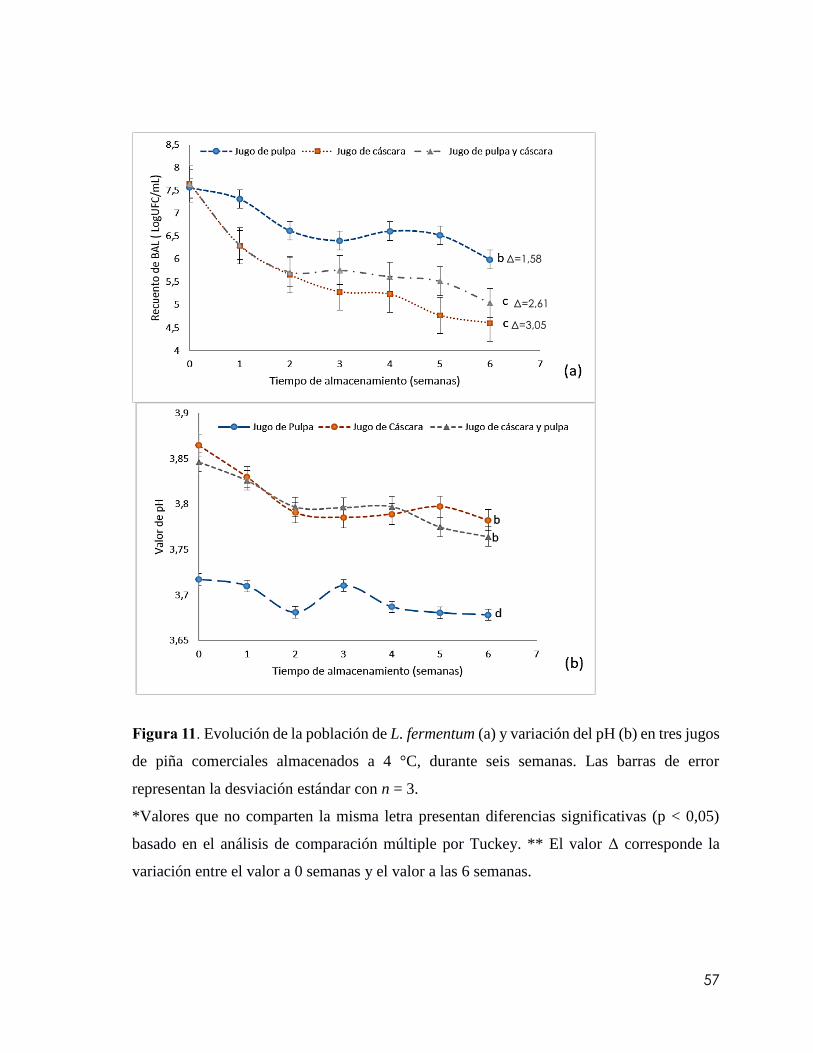
57
Figura 11. Evolución de la población de L. fermentum (a) y variación del pH (b) en tres jugos
de piña comerciales almacenados a 4 °C, durante seis semanas. Las barras de error
representan la desviación estándar con n = 3.
*Valores que no comparten la misma letra presentan diferencias significativas (p < 0,05)
basado en el análisis de comparación múltiple por Tuckey. ** El valor Δ corresponde la
variación entre el valor a 0 semanas y el valor a las 6 semanas.
Δ=2,61
Δ=3,05
Δ=1,58

58
El comportamiento de la bacteria en los jugos de cáscara y pulpa-cáscara, durante el
tiempo de almacenamiento es muy similar entre ellos (comparación realizada con la
población obtenida en la semana 6 del estudio de sobrevivencia), ya que no se presentan
diferencias significativas (p > 0,05) al comparar las poblaciones de la bacteria entre ambos
jugos. Es importante mencionar que, en ambos jugos, los recuentos fueron menores a los seis
logaritmos después de la segunda semana en almacenamiento a 4 °C, que según la literatura
corresponde a la población mínima del microorganismo requerida para ser considerado un
producto con probiótico. Dado lo anterior, la cepa evaluada no puede utilizarse
industrialmente en jugo de cáscara de piña y jugo de cáscara + pulpa en proporción 20:80
como microorganismo probiótico después de 6 semanas de almacenamiento en refrigeración.
En el caso del jugo de pulpa, el microorganismo puede considerarse un candidato como cepa
probiótica durante las primeras seis semanas de almacenamiento. Sin embargo, para
corroborar esto, se debe realizar un estudio de vida útil para determinar si alguna de las
propiedades físicoquímicas, sensoriales o nutricionales del jugo se ven comprometidas de
forma negativa durante este tiempo de almacenamiento en refrigeración, además de que hay
que analizar si el producto es aceptable sensorialmente para el consumidor.
El análisis anterior muestra que el tipo de jugo utilizado, tiene un efecto importante
sobre la sobrevivencia de la bacteria, y que este fenómeno depende del tiempo de
almacenamiento. Estas diferencias podrían explicarse por las características fisicoquímicas
de los jugos (Cuadro III). Es importante mencionar, que el sustrato principal del metabolismo
de las bacterias lácticas son los azúcares fermentables: fructosa, glucosa y sacarosa, y de
forma secundaria puede utilizar otros compuestos de carbono como los ácidos orgánicos
(Foschino et al., 2001; Dan et al., 2015); lo cual implica que su sobrevivencia se ve limitada
por el contenido de estas fuentes de carbono en la matriz donde son inoculadas. De acuerdo
con lo determinado para los tres jugos, el jugo de pulpa presenta el mayor contenido de
azúcares fermentables, seguido por el de pulpa + cáscara y, por último, el jugo de cáscara
puro, que se caracterizó por ser la matriz más limitada en azúcares. Nótese que dicho orden
se refleja en la sobrevivencia de la bacteria. Esto implica que el contenido de azúcares fueron
determinantes para la sobrevivencia de la misma. Por tanto, la adición de ingredientes con

59
propiedades prebióticas como algunas fibras, los fructooligosacáridos y la inulina (Slavin,
2013) al jugo para obtener una bebida podría ser una opción para la introducción del
microorganismo con propiedades probióticas en un producto derivado de la producción
piñera.
El modelo factorial utilizado para evaluar la variación del pH durante el experimento
de almacenamiento de seis semanas en refrigeración, presentó un coeficiente de
determinación (R2) de 0,97 y una interacción significativa entre el tipo de jugo y el tiempo
de almacenamiento (p = 0,0195) (Figura 11b). Se observa una tendencia a la reducción del
pH dependiendo del tiempo de almacenamiento, la cual resultó más pronunciada a partir de
la segunda semana de almacenamiento, momento a partir del cual muestra una leve
estabilidad en el caso del jugo de pulpa + cáscara y el jugo de cáscara; en el caso del jugo de
pulpa se observaron ciertas variaciones que bien pudieron deberse a la variabilidad del equipo
de medición. Estos resultados, podrían sugerir que se dio una producción importante del
ácido láctico en las primeras dos semanas de almacenamiento; sin embargo, su producción
se vio disminuida durante las siguientes cuatro semanas de almacenamiento. Estos resultados,
son concordantes con las curvas de sobrevivencia de la población (Figura 11b), lo cual podría
señalar que la menor variación de pH mostrada a partir de la semana seis podría ser
consecuencia de la muerte de la bacteria. De acuerdo con Ramírez et al (2011), la presencia
de L. fermentum activa y viable implica que la misma cumple con su ciclo metabólico natural,
que conlleva a una mayor producción de ácidos orgánicos, principalmente el ácido láctico,
que causan la caída del pH. Lo anterior tiene coherencia con las determinaciones de ácidos
orgánicos (Cuadro III), donde se identificó una concentración mínima en la producción de
ácido láctico en la sexta semana de almacenamiento.

60
VI. CONCLUSIONES
Las técnicas moleculares permitieron la identificación de un total de tres géneros de
bacterias lácticas aisladas de silos elaborados a partir de subproductos de piña, entre
ellas: Lactobacillus spp., Fructobacillus spp. y Weissella spp.
Se identificaron efectivamente diferentes especies de Lactobacillus sp., Weissella sp.
y Fructobacillus tropaeoli por medio de marcadores moleculares que amplificaron la
región del ADNr 16S; sin embargo, en alguno de los casos, las técnicas no fueron
suficientemente específicas para lograr la distinción clara entre especies.
Se determinó que nueve de las cepas estudiadas presentan una buena tolerancia a un
medio con un pH de 2,0 (sobrevivencia mayor al 90 %), por lo que podrían ser
consideradas como microorganismos potencialmente probióticos.
Los tres jugos analizados presentan características fisicoquímicas significativamente
distintas (sólidos solubles, acidez total titulable, perfil de azúcares fermentables y
perfil de ácidos orgánicos) entre ellos.
El jugo de pulpa presentó las mejores condiciones fisicoquímicas para el desarrollo
de la bacteria ya que presento un perfil de azúcares fermentables más alto y una acidez
total menor.
El jugo de pulpa mostró las mejores condiciones para la sobrevivencia de L.
fermentum KX218443 durante las seis semanas de almacenamiento.
No se observaron diferencias significativas en el comportamiento de las poblaciones
a lo largo de las seis semanas de almacenamiento, entre los jugos de cáscara y pulpa
+ cáscara. En ambos caso se observó una tendencia a la muerte celular.

61
VII. RECOMENDACIONES
Emplear otro tipo de marcadores como los MLSTs y el de Girasa para identificación
molecular más específica de las bacterias que no pudieron ser separadas por especie
utilizando el gen ADNr 16S.
Efectuar pruebas adicionales como tolerancia a la lisozima, bilis y adherencia a la
mucosa y epitelio intestinal o inclusive pruebas in vivo para evaluar el potencial
probiótico de las bacterias estudiadas.
Realizar este estudio de sobrevivencia con jugos estandarizados (pH y ° Brix) y
evaluando la posibilidad de la adición de prebióticos o el uso de cepas en
combinaciones sinérgicas como las que se emplean comercialmente.
Realizar un estudio de vida útil para confirmar si las propiedades físicoquímicas,
sensoriales o nutricionales del jugo se ven negativamente comprometidas durante el
de almacenamiento en refrigeración y para determinar la aceptación del jugo por parte
de consumidores potenciales.

62
VIII. BIBLIOGRAFÍA
ABDULLAH, M. & MAT, H. 2008. Characterization of solid and liquid pineapple waste.
Reaktor 12 (1): 48 – 52.
ACEVEDO, I. GARCÍA, O. CONTRERAS, J & ACEVEDO, I. 2009. Elaboración y evaluación
de las características sensoriales de un yogurt de leche caprina con jalea semifluida de
piña. Revista UDO Agrícola 9 (2): 442-448.
ADIL, M. RAFIQ, M. KAMRAN, M. AHMAD, S & SHAH, F. 2014. Study on the storage
stability of fruits concentrates. Polish Journal of Food and Nutrition Sciences 24 (2): 101-
107.
AIJN. 2014. 2014 Liquid Fruit Market Report. William Press, Bélgica.
ALTSCHUL, S. GISH, W. MILLER, W. MYERS, E & LIPMAN, D. 1990. Basic local
alignment search tool. Journal of Molecular Biology 215: 403- 410.
ANAL, A & SINGH, H. 2007. Recent advances in microencapsulation of probiotics for
industrial applications and targeted delivery. Trends in Food Science 18: 240-251.
AOAC. 2005a. AOAC Official Method 942.15: Acidity (Titratable) of Fruit Products. Official
Methods of Analysis of AOAC International. 18 ed., Association of Official Analytical
Chemists. Estados Unidos.
AOAC. 2005e. AOAC Official Method 925.45: Loss on Drying (Moisture) in Sugars. Official
Methods of Analysis of AOAC International. 18 ed., Association of Official Analytical
Chemists. Estados Unidos
AOAC. 2005f. AOAC Official Method 981.12: pH of Acidified Foods. Official Methods of
Analysis of AOAC International. 18 ed., Association of Official Analytical Chemists.
Washington. Estados Unidos
AOAC. 2005g. AOAC Official Method 932.12: Solids (Soluble) in Fruits and Fruit Products
(Refractometer Method). Official Methods of Analysis of AOAC International. 18 ed.,
Association of Official Analytical Chem. Estados Unidos
AOAC. 2005h. AOAC Official Method 920.151: Solids (Total) in Fruits and Fruit Products.
Official Methods of Analysis of AOAC International. 18 ed., Association of Official
Analytical Chemists. Estados Unidos.

63
APPLIED BIOSYSTEMS. 2006. PCR optimization: reaction conditions and components.
INTERNET: https://www3.appliedbiosystems.com. Recuperado el 5 abril, 2016.
ASHABIL, A. 2006. Nucleic acid extraction from clinical specimens for PCR applications.
Turkish Journal of Biology 30: 107- 120.
BABALOLA, O. 2003. Molecular techniques: an overview of methods for the detection of
bacteria. African Journal of Biotechnology 2: 728-731.
BALAS, D. 2004. Caracterización del gen gyrA de Streptococcus pneumoniae y regulación de
expresión. Universidad Complutense, España.
BARGHOUTHI, S. 2011. A Universal Method for the Identification of Bacteria Based on
General PCR Primers. Indian Journal of Microbiology 51 (4): 430–444.
BARON, E. 1996. Classification. In: Baron S, editor. 1996. Medical Microbiology. 4th ed.
University of Texas Medical Branch. Galveston,
BARTHOLOMEW, D.; PAULL, R. & ROHRBACH, K. 2003. The pineapple: botany,
production and uses. Cabi Publishing, Reino Unido.
BIRNBOIM, H.C. AND DOLY, J. 1979. A rapid alkaline procedure for screening recombinant
plasmid DNA. Nucleic Acids Research 7: 1513–1523.
BLACKBURN, C. 2006. Food spoilage microorganisms. CRC Press, Inglaterra.
BONASERA, J. ASSELIN, J & BEER, S. 2014. Identification of bacteria pathogenic to or
associated with onion (Allium cepa) based on sequence differences in a portion of the
conserved gyrase B gene. Journal of Microbiological Methods 103: 138- 143.
BONFIELD, J. SMITH, K. & STADEN, R. 1995. A new DNA sequence assembly program.
Nucleic Acids Research 23 (24): 4992--4999.
BRON, P. GRANGETTE, C. MERCENIER, A. DE VOS, W & KLEEREBEZEM, M. 2004.
Identification of Lactobacillus plantarum genes that are induced in the gastrointestinal
tract of mice. Journal of Bacteriology 186 (17): 5721- 5729.
CÁMARA, M. DÍEZ, C. TORIJA, M & CANO, M. 1994. HPLC determination of organic acids
in pineapple juices and nectar. Zeitschrift für Lebensmittel- Untersuchnunh und –
Forschung 198 (1): 52-56.
CÁMARA NACIONAL DE PRODUCTORES Y EXPORTADORES DE PIÑA (CANAPEP).
2012. Piña: Líder en generación de divisas. Piña de Costa Rica, 8: 42-44.

64
CANCHAYA, C. CLAESSON, M. FITZGERALD, G. SINDEREN, D & O’TOOLE, P. 2006.
Diversity of the Genus Lactobacillus revealead by comparative genomics of five species.
Microbiology 152: 3185-3196.
CHARTERIS, W. KELLY, P. MORELLI & COLLIN, L. 1998. Development and application
of an in vitro methodology to determine the transit tolerance of potentially probiotic
Lactobacillus & Bifidobacterium species in the upper human gastrointestinal tract.
Journal of Applied Microbiology 84: 759-768.
CITA. 2009a. Acidez Titulable. Método Analítico P-SA-MQ-011. Emisión 3. Laboratorio de
Química del CITA. Centro Nacional de Ciencia y Tecnología de Alimentos, Costa Rica.
CITA. 2009c. Determinación del pH. Método Analítico P-SA-MQ-012. Emisión 1. Laboratorio
de Química del CITA. Centro Nacional de Ciencia y Tecnología de Alimentos, Costa
Rica.
CITA. 2009d. Humedad. Método Analítico P-SA-MQ-002. Emisión 1. Laboratorio de Química
del CITA. Centro Nacional de Ciencia y Tecnología de Alimentos, Costa Rica.
CITA. 2011. Azúcares por HPLC. Método Analítico P-SA-MQ-024. Emisión 11. Laboratorio
de Química del CITA. Centro Nacional de Ciencia y Tecnología de Alimentos, Costa
Rica
CITA. 2012. Recuento de bacterias ácido lácticas. Método de análisis microbiológico P-SA-
MM-006. Emisión 1. Laboratorio de Microbiología del CITA. Centro Nacional de
Ciencia y Tecnología de Alimentos, Costa Rica.
CITA. 2016. P-SA-MQ-051: Determinación de Ácidos orgánicos. Laboratorio de Química del
CITA. Centro Nacional de Ciencia y Tecnología de Alimentos, Costa Rica.
CLARK, P. 2005. Molecular Biology. Academic Press, Estados Unidos.
CLARRIDGE, J. 2004. Impact of 16S rRNA Gene Sequence Analysis for Identification of
Bacteria on Clinical Microbiology and Infectious Diseases. Clinical Microbiology
Reviews 17(4): 840–862.
COLLINS M. D. SAMELIS J. METAXOPOULOS J & WALLBANKS S. 1993. Taxonomic
studies on some Leuconostoc−like organisms from fermented sausages: description of a
new genus Weissella for the Leuconostoc paramesenteroides group of species. Journal
of Applied Microbiology 75: 595–603.

65
COLLINS, M. RODRIGUES, U. ASH, C. AGUIRRE, M. FARROW, J. MARTINEZ-MURCIA,
A. PHILLIPS, B. WILLIAMS, A & WALLBANKS, S.1991. Phylogenetic analysis of
the genus Lactobacillus and related lactic-acid bacteria as determined by reverse-
transcriptase sequencing of 16s ribosomal-RNA. FEMS Microbiology Letters 77: 5–12.
CORBO, M. BEVILACQUA, A. PETRUZZI, L. CASANOVA, F.P. & SINIGAGLIA, M. 2014.
Functional beverages: The emerging side of functional foods commercial trends, research,
and health implications. Comprehensive Reviews in Food Science and Food Safety 13:
1192–1206.
DAN, T. LIU, W. SONG, Y. XU, H. MENGHE, B. ZHANG, H & SUN, Z. 2015.The evolution
and population structure of Lactobacillus fermentum from different naturally fermented
products as determined by multilocus sequence typing (MLST). BMC Microbiology 15:
107.
DELLAGLIO, F & FELIS, G. 2005. Taxonomy of lactobacilli and bifidobacteria. En:
TANNOCK, G (Ed). 2005. Probiotic and prebiotics: Scientific aspects. Academic Press,
Reino Unido.
DESAI, A. 2008. Strain identification, viability and probiotics properties of Lactobacillus casei.
Universidad de Victoria, Australia.
DICAGNO, R. CARDINALO, G. MINERVINI, G. ANTONIELLI, L. RIZZELLO, C.
RICCIUTI, P & GOBBETTI, M. 2010. axonomic structure of the yeasts and lactic acid
bacteria microbiota of pineapple (Ananas comosus L. Merr.) and use of autochthonous
starters for minimally processing. Food Microbiology 27 (3): 381- 389.
DING, W & SHAH, N. 2008. Survival of free and microencapsulated probiotic bacteria in
orange and apple juices. International Food Research Journal 15: 219–232
DONOHUE, D. & SALMINEN, S. 1996. Safety of probiotic bacteria. Asia Pacific Journal of
Clinical Nutrition 5: 25-28.
ENDO A. & OKADA S. 2008. Reclassification of the genus Leuconostoc and proposals of
Fructobacillus fructosus gen. nov., comb. nov., Fructobacillus durionis comb. nov.,
Fructobacillus ficulneus comb. nov. and Fructobacillus pseudoficulneus comb. nov.
International journal of systematic bacteriology 58: 2195–2205.

66
ENDO, A. 2012. Fructophilic lactic acid bacteria inhabit fructose-rich niches in nature.
Microbial Ecology in Health 23: 6-9.
ENGEBRECHT, J. BRENT, R & KADERBHAI, M. 1991. Minipreps of plasmid DNA. En:
AUSUBEL, F. BRENT, R. KINGSTON, R. MOORE, D. SEIDMAN, J. SMITH, J &
STRUHL, K (Eds). 1991. Current protocols in molecular biology. John Wiley & Sons,
Estados Unidos.
ESTELA, W. RYCHTERA, M. MELZOCH, K. QUILLAMA, E & EGOAVIL, E. 2007.
Producción de ácido láctico por Lactobacillus plantarum L10 en cultivos batch y
continuo. Revista Perú Biología 14 (2): 271-275.
EUROMONITOR. 2014. J. Food World Juice Conference. España.
FAOSTAT. 2010. FAOSTAT Pineapple exports. FAO, Roma. INTERNET:
http://www.faostat.fao,org. Recuperado el 1 marzo, 2015.
FIORA, J. THEODOROU, V & BUENO, L. 2003. Probiotics: what are they? What are their
effects on gut physiology? Best Practice & Research Clinical Gastroenterology 17: 711–
724.
FONTECHA, G. 2005. Protocolo MOCAT para la extracción de ADN ribosomal de bacterias.
Laboratorio Centro de Investigaciones en Biología Celular y Molecular. Universidad de
Costa Rica, Costa Rica.
FOSCHINO, R., PICOZZI, C& GALLI, A. 2001. Comparative study of nine Lactobacillus
fermentum bacteriophagues. Journal of Applied Microbiology 91 (3): 394-403.
FOWOYO, P & UZOMA, E. 2015. Studies on the antifungal activity of Lactobacillus plantarum
and Lactobacillus fermentum on spoilage fungi of tomato fruit. Journal of Microbiology
Research 5 (3): 95- 100.
FRANCO, T. GARICÍA, S. HIROOKA, E. ONO, Y & SANTOS, J. 2011. Lactic acid bacteria
in the inhibition of Fusarium graminearum and deozynivalenol detoxication. Journal of
Applied Microbiology 111: 739- 748.
FRAZIER, W. & WESTHOFF, D. 1995. Food Microbiology 4th Edition, McGraw Hill. New
Delhi.

67
GADAGA, T. MUTUKUMIRA, A. NARVHUS, J & FERESU, S. 1999. A review of traditional
feremented foods and beverages of Zimbabwe. International Journal of Food
Microbiology 53: 1-11.
GARCÉS, A. RUÍZ, S. BUILES, A. SERNA, J & BERRIO, L. 2004. Ensilaje como fuente de
alimentación para el ganado. Revista Lasallista de Investigación 1 (1): 68-71.
GIBSON, G. PROBERT, H. LOO, J. RASTALL, R & ROBERFROID, M. 2004. Dietary
modulation of human colonic microbiota: updating the concept of prebiotics. Nutrition
Research Reviews 17 (2): 259- 275.
GILLILAND, S. 2002. Technological & Commercial Applications of Lactic Acid Bacteria;
Health & Nutritional Benefits in Dairy Products. FAO, Estados Unidos.
GOBBETTI, M. DI CAGNO, R & DE ANGELIS, M. 2010. Functional microorganisms for
functional food quality. Critical Reviews in Food Science and Nutrition 50: 716–727.
GOKTEPE, I. JUNEJA, V & AHMEDNA, M. 2006. Probiotics in food safety and human health.
CRC Press, Estados Unidos.
GONZÁLEZ, A. GUILLAMÓN, J. MAS, A &POBLET, M. 2006. Application of molecular
methods for routine identification of acetic acid bacteria. International Journal of Food
Microbiology 108 (1): 141-146.
GONZÁLEZ, L. 2012. Manual técnico para el manejo de rastrojos en el cultivo de piña. Servicio
Fitosanitario del Estado- Ministerio de agricultura y Ganadería, Costa Rica.
GRANATO, D. BRANCO, D. GOMES, A. FONSECA, J & SHAH, N. 2010. Probiotic dairy
products as functional foods. Comprehensive Reviews in Food Science and Food Safety
9 (5): 455–470.
GUEIMONDE, M. DE LOS REYES, C & SÁNCHEZ, B. Stability of Lactic Acid Bacteria. En:
GUEIMONDE, M. JALONEN, L. HE, F. HIRAMATSU, M & SALMINEN, S. 2006.
Adhesion and competitive inhibition and displacement of human enteropathogens by
selected lactobacilli. Food Research International 39 (4): 467-471.
GUEIMONDE, M. NORIEGA, L. MARGOLLES, A. DE LOS REYES-GAVILÁN, C &
SALMINEN, S. 2005. Ability of Bifidobacterium strains with acquired resistance to bile
to adhere to human intestinal mucus. International Journal of Food Microbiology 101:
341−346.

68
GUTIÉRREZ, F. ROJAS, A. DORMOND, H. POORE, M & WING- CHING, R. 2003.
Características nutricionales y fermentativas de mezclas enziladas de desechos de piña y
avícolas. Agronomía Costarricense 27 (1): 79-89.
HALASZ, A. 2009. Lactic Acid Bacteria. En: Food Quaity and Standards, Vol.III. EOLSS,
Estados Unidos.
HAMMES, W & VOGEL, R. 1995. The genus Lactobacillus.En: WOOD, B & HOLZAPFEL,
W(Eds). 1995. The lactic acid bacteria, Vol.II. Blackie Academic and Profesional, Reino
Unido.
HANNA INSTRUMENTS. 2015. HI84432 Automatic miniTitrator and pH Meter for titratable
Acidity determination in fruit juices. Estados Unidos.
HARUENKIT, R. 2004 Analysis of sugars and organic acids in pineapple, papaya and star fruit
by HPLC using an aminex HPX-87 H column. King Mongkut's Agricultural Journal 22
(3): 11-18.
HENSEL, C. XIANG, R. SAKAGUCHI, A & NAYLOR, S. 1991. Use of the single strand
conformation polymorphism technique and PCR to detect p53 gene mutations in small
cell lung cancer. Oncogene 6 (6): 1067–1071
HENTEMANN, M. REISS, J. WAGNER, M & COOPER, D. 1990 .Rapid detection of deletions
in the Duchenne muscular dystrophy gene by PCR amplification of deletion-prone exon
sequences. Human Genetics 84: 228–232
HERTZLER, S. HUYNH, B & SAVAIANO, D. 1996. How much lactose is low lactose? Journal
of the American Dietetic Association 96:243-246.
HOELZEL, A & GREEN, A. 1998. PCR protocols and population analysis by direct DNA
sequencing and PCR-based DNA fingerprinting. En: HOELZEL, A (Eds). 1998. 2da Ed.
Oxford University Press, Inglaterra.
HORN, S. ASPMO, S. & EIJSINK, V. 2005. Growth of Lactobacillus plantarumin media
containing hydrolysates of fish viscera. Journal of Applied Microbiology 99 (5): 1082-
1089.
HUI, Y. CHEN, F & NOLLET, L. 2010. Handbook of fruit and vegetable flavors. Wiley, Estados
Unidos.

69
HUSE, S. DETHLEFSEN, L. HUBER, J. MARK, D & RELMAN, D. 2008. Exploring microbial
diversity and taxonomy using SSU rRNA hypervariable tag sequencing. PLoS Genet 4:
e1000255. doi: 10.1371/journal.pgen.1000255. pmid:19023400
ILLEGHEMS, K. DE VUYST, L & WECKX, S. 2015. Comparative genome analysis of the
candidate functional starter culture strains Lactobacillus fermentum 222 and
Lactobacillus plantarum 80 for controlled cocoa bean fermentation processes. BMC
Genomics 16 (1): 766.
ISHMAEL, F & STELLATO, C. 2008. Principles and aplications of polymerase chain reaction:
basic science for the practicing physician. Annal of Allergy, Asthma & Immunology 101
(4): 437- 443.
JIMÉNEZ, E. LANGA, S. MARTIN, V. ARROYO, R. MARTÍN, R. FERNÁNDEZ, L &
RODRÍGUEZ, J. 2010. Complete Genome Sequence of Lactobacillus fermentum CECT
5716, a Probiotic Strain Isolated from Human Milk. Journal of bacteriology 192 (18):
4800.
KANDLER, O. 1983. Carbohydrate metabolism in lactic acid bacteria. Antonie van
Leeuwenhoek Journal of Microbiology and Serology 49: 209-224.
KANT, R. BLOM, J. PALVA, A. SIEXEN, R & DE VOS, W. 2011. Comparative genomics of
Lactobacillus. Microbial biotechnology 4 (3): 323-332.
KELANNE, N. 2012. Aplicability of fructophilic lactic acid bacteria in food industry. Turku
University of Applied Sciences, Finlandia.
KLINDWORTH, A. PRUESSE, E. SCHWEER, T. PEPLIES, J & QUAST, C. 2013. Evaluation
of general 16S ribosomal RNA gene PCR primers for classical and next-generation
sequencing-based diversity studies. Nucleic Acids Research 41: e1. doi:
10.1093/nar/gks808. pmid: 22933715
KNICKY, M. 2005. Possibilities to improve Silage Conservation: Effects of crop, ensiling
technology and additives. Swedish University of Agricultural Sciences, Suiza. pp
KONINGS, W. KOK, J. KUIPERS, O & POOLMAN, B. 2000. Lactic acid bacteria: the bugs
of the new millennium. Current Opinion in Microbiology 3: 276–282.

70
KRYNDUSHKIN, D. ALEXANDROV, I. TER-AVANESYAN, M & KUSHNIROV, V. 2003.
Yeast [PSI+] prion aggregates are formed by small Sup35 polymers fragmented by
Hsp104. Journal of Biological Chemistry 278 (49): 49636–49643.
KUMAR, S. TAMURA, K. & NEI, M. 2004. MEGA3: Integrated software for Molecular
Evolutionary Genetics Analysis and sequence alignment. Briefings in Bioinformatics
5:150-163.
KUMARI, M. KUMAR-SWARNKAR, M. KUMAR, S. KUMAR- SIGH, A & GUPTA, M.
2015. Genome sequence of potential probiotic strain, Lactobacillus fermentum HFB3,
isolated from human gut. Genome Announcements 3 (6): e01296-15.
KUMURA, H. TANQUE, Y. TSUKARA, M. TANAKA, T & SHIMAZAKI, K. 2004.
Screening of dairy yeast strains for probiotic applications. Journal of Dairy Science 87:
4050- 4056.
LAHTINEN, S. OUWEHAND, A. SALMINEN, S & VON WRIGHT, A. 2012. Lactic acid
bacteria: microbiological and functional aspects. 4ed. CRC Press, Boca Ratón.
LANKAPUTHRA, W & SHAH, N. 1995. Survival of Lacotbacillus acidophilus and
Bifidobacterium spp in the presence of acid and bile salts. Cult. Dairy Products Journal
30 (3): 2-7.
LARSEN, M. COSENTINO, S. RASMUSSEN, S. FRIIS, C. HASMAN, H. MARVIG, R.
JELSBAK, L. SICHERITZ, T. USSERY, D. AARESTRUP, F & LUND, O. 2012.
Multilocus sequence typing of total genome sequenced bacteria. Journal of Clinical
Microbiology 50 (4): 1355 -1361.
LASKER, B. 2002. Evaluation of performance of four genotypic methods for studying the
genetic epidemiology of Aspergillus fumihatus isolates. Journal of Clinical Microbiology
40 (8): 2886-2892.
LEATHERHEAD FOOD INTERNATIONAL. 2006. The international market for functional
foods. En: Functional Food Market Report. Reino Unido.
LEBLANC, J. GARRO, M & SAVOY, G. 2004. Effect of pH on Lactobacillus fermentum
growth, raffinose removal, α-galactosidase activity and fermentation products. Applied
Microbiology and Biotechnology 65: 119-123.

71
LIN, W. HWANG, C. CHEN, L & TSEN, H. 2006. Viable counts, characteristic evaluation for
commercial lactic acid bacteria products. Food Microbiology 23: 74-81.
LOMER. 2006. Lactose intolerance and lactase deficiency. The Dairy Council Text Book, Reino
Unido.
LOURENS, A. & VILJOEN, B. 2001. Yogurt as probiotic carrier food. International Dairy
Journal. 11:1- 17.
LUCKOW, T. SHEEHAN, V. FITZGERALD, G & DELAHUNTY, C. 2006. Exposure, health
information and flavour-masking strategies for improving the sensory quality of probiotic
juice. Appetite 47: 315–323.
MAIDEN, M. JANSEN, M. BRAY, J. EARLE, S. FORD, S. JOLLEY, K & MCCARTHY, N.
2013. MLST revisited: the gene by gene approach to bacterial genomics. Nature Reviews
Microbiology 11 (10): 728- 736.
MAKAROVA, K. SLESAREV, A. WOLF, Y. SOROKIN, A. MIRKIN, B. KOONIN, E.
PAVLOV, A. PAVLOVA, N. KARAMYCHEV, V. POLOUCHINE, N. SHAKHOVA,
V. GRIGORIEV, I. LOU, Y. ROHKSAR, D. LUCAS, S. HUANG K. GOODSTEIN DM.
HAWKINS T. PLENGVIDHYA V. WELKER D. HUGHES J. GOH Y. BENSON A.
BALDWIN, K. LEE, J. DÍAZ-MUÑIZ, I. DOSTI, B. SMEIANOV, V. WECHTER, W.
BARABOTE, R. LORCA, G. ALTERMANN, E. BARRANGOU, R. GANESAN, B.
XIE, Y. RAWSTHORNE, H. TAMIR, D. PARKER, C. BREIDT, F. BROADBENT, J.
HUTKINS, R. O'SULLIVAN, D. STEELE, J. UNLU, G. SAIER, M. KLAENHAMMER,
T. RICHARDSON, P. KOZYAVKIN, S. WEIMER, B & MILLS, D. 2006. Comparative
genomics of the lactic acid bacteria. Proceedings of the National Academy of Sciences.
103:15611–15616.
MANIATIS, T. FRITSCH, E. & SAMBROOK, J. 1989. Molecular Cloning, a Laboratory
Manual. Cold Spring Harbor, NY. Cold Spring Harbor Laboratory Press.
MATURIN, L & PEELER, T. 2001. Aerobic Plate Count. En: Food and Drug Administration
(FDA). Bacteriological Analytical Manual Online, 8va Ed. Silver Spring, Berlín.
MAT, A. & MAT, H. 2008. Characterization of solid and liquid pineapple waste. Reaktor 12 (1):
48-58.

72
MCCABE, K. ZHANG, Y. HUANG, B. WAGAR, E & MCCABE, E. 1999- Bacterial species
identification after DNA amplification with universal primer pair. Molecular Genetics
and Metabolism 66: 205- 211.
MERRY, R. J. 1997. Current and future approaches to biocontrol in silage. En: Proceedings of
the 8th International Symposium on Forage Conservation 8: 17-27.
MINISTERIO DE GANADERIA Y AGRICULTURA (MAG). 2013. Manual de
Recomendaciones del manejo de Mosca de Establo. PIITA-PIÑA, Costa Rica.
MINTEL. 2015. Informe Global sobre Tendencias en Alimentos y Bebidas para 2016.
INTERNET: http://es.mintel.com/. Recuperado el 15 de abril, 2016.
MONTERO, M. & CERDAS, M. 2005. Guías técnicas del Manejo Postcosecha de piña para el
mercado fresco. Ministerio de Agricultura y Ganadería, Costa Rica.
MOORE, W. ARNSCHEIDT, A. KRUGER, A. STROMPL, C & MAU, M. 2004. Simplified
protocols for the preparation of genomic DNA from bacterial cultures. Molecular
Microbial Ecology Manual 101: 3 -18.
MORALES, Y. DUQUE. E. RODRIGUEZ, O. TORRE, J. MARTÍNEZ, R. PÉREZ, R. &
MUÑOZ, J. 2010. Bacterias preservadas, una fuente importante de recursos
biotecnológicos. Biotecnología 14 (2): 11-29.
MUHIRE, B. VARSANI, A & MARTIN, D. 2014. SDT: A Virus Classification Tool Based on
Pairwise Sequence Alignment and Identity Calculation. PLoS ONE 9(9): e108277.
doi:10.1371/journal.pone.0108277
MURILLO, O. 2014. Ficha Técnica de industrialización de Piña. Centro Nacional de
Producción. Costa Rica.
NADZIRAH, K. ZAINAL, S. NORIHAM, A. NORMAH, I. SITIROHA, A & NADYA, H.
2013. Physico-chemical properties of pineapple variety N36. Harvested and stored at
different maturity stages. International Food Research Journal 20 (1): 225-231.
NATIONAL FORENSIC SCIENCE TECHNOLOGY CENTER (NFSTC). 2015. Chelex ® 100
Extraction Process. INTERNET: http://projects.nfstc.org. Recuperado el 10 de mayo,
2016.
NEWTON, I & ROESELERS, G. 2012. The effect of training set on the classification of honey
bee gut microbiota using the Naive Bayesian Classifier. BMC Microbiology 12: 221

73
NORIEGA, L. GUEIMONDE, M. SÁNCHEZ, B. MARGOLLES, A & DE LOS REYES-
GAVILÁN, C. 2004. Effect of the adaptation to high bile salts concentrations on
glycosidic activity, survival at low pH and cross-resistance to bile salts in
Bifidobacterium. International Journal of Food Microbiology 94:79−86.
NUALKAEKUL, S. & CHARALAMPOPOULOS, D. 2011. Survival of Lactobacillus
plantarum in model solutions and fruit juices. International Journal of Food Microbiology
146: 111-117.
ORLA, S. 1919. The lactic acid bacteria. Host and Son, Copenhagen.
ORTIZ, A. REUTO, J. FAJARDO, E. SARMIENTO, S. AGUIRRE, A. ARBELÁEZ,G.
GÓMEZ, D & QUEVEDO, B. 2008. Evaluación de la capacidad probiótica in vitro de
una cepa nativa de Saccharomyces cerevisiae. Universitas scientiarum 13 (2): 138- 148.
PAN, DAO D., XIA Q. ZENG, & TIAN, Y. 2011. Characterization of Lactobacillus fermentum
SM-7 isolated from koumiss, a potential probiotic bacterium with cholesterol-lowering
effects. Journal of the Science of Food and Agriculture 91(3): 512-518.
PAPAGIANNI, M. 2012. Metabolic engineering of lactic acid bacteria for the production of
industrially important compounds. Computational and Structural Biotechnology Journal.
http://doi.org/10.5936/csbj.201210003J
PARDO, I. 2003. Metabolismo de sustratos del mosto y vino por bacterias lácticas y sus
implicaciones en la calidad del vino. ACE Revista de Enología. INTERNET:
http://www.acenologia.com/. Recuperada el 12 de abril, 2016.
PÉREZ, M & RAMÍREZ, C. 2007. Utilización de bacterias acido lácticas termorresistentes
como probióticos en embutidos cárnicos conocidos. NACMEH 1 (2): 87- 96.
PERRICONE, M. BEVILACQUA, A. ALTIERI, C. SINIGAGLIA, M & CORBO, M. 2015.
Challenges for the production of probiotic fruit juices. Beverages 1: 95- 103.
PERRICONE, M. CORBO, M. SINIGAGLIA, M. SPERANZA, B & BEVILACQUA, A. 2014.
Viability of Lactobacillus reuteri in fruit juices. Journal of Functional Foods 10: 421–
426.
PESVNER, J. 2009. Bioinformatics and functional genomics. 2da ed. John Wiley & Sons Inc.,
Estados Unidos.

74
POUCH, F & ITO, K. 2001. Compendium of methods for the microbiological examination of
foods. 4ta Ed. American Public Health Association, Estados Unidos.
PRADO, F. PARADA, J. PANDEY, A & SOCCOL, C. 2007. Trends in non-dairy probiotic
beverages. Food Research International 41: 111- 123.
PYAR, H. LIONG, M. & PEH, K. 2014. Potentials of pineapple waste as growth medium for
Lactobacillus species. International Journal of Pharmacy and Pharmaceutical Sciences 6
(1): 142-145.
QUESADA, K., ALVARADO, P., SIBAJA, R. & VEGA, J. 2005. Utilización de las fibras del
rastrojo de piña (Ananas comosus, variedad champaka) como material de refuerzo en
resinas de poliéster. Revista Iberoamericana de Polímeros 6 (2): 158 – 179.
RABINDRA, P & RAJU, N. 2012. Gel-Electrophoresis and Its Applications, Gel
Electrophoresis. En: SAMEH MAGDELDIN Ed. 2012. Principles and Basics. DOI:
10.5772/38479.
RAMÍREZ, J. ROSAS, P. VELÁZQUES, M. ULLOA, J & ARCE, F. 2011. Bacterias lácticas:
Importancia en alimentos y sus efectos en la salud. Revista Fuente 2 (7): 3-15.
RAMOS, C. THORSEN, L & FREITAS, R. 2013. Strain-specific probiotics properties of
Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus brevis isolates
from Brazilian food products. Food Microbiology 36 (1): 22- 29.
RANADHEERA, C. PRASANNA, P & VIDANARACHCHI, J. 2014. Fruit juice as probiotic
carriers. En: Fruit Juices: ELDER, K. Ed. 2014. Types, Nutritional Composition and
Health Benefits. Nova Science Publishers: Hauppauge, Estados Unidos.
REID, G. SNADERS, M. GASKINS, R. GIBSON, G. MERCENIER, A. RASTALL, R.
ROBERFROID, M. ROWLAND, I- CHERBUT, C & KLAENHAMMER, T. 2003. New
scientific paradigms for probiotics and prebiotics. Journal of Clinical gastroenterology
37 (2): 105-118.
RIAZ, A. NAWAZ, S & HASNAIN, S. 2010. Bacteriocins produced by L. fermentum and L.
acidophilus can inhibit cephalosporin resistant E.coli. Brazilian Journal of microbiology
41: 643-648.
RODRÍGUEZ, J. 2006. Microorganismos y salud: bacterias lácticas y bifidobacterias
probióticas. Editorial Complutense, España.

75
SAARELA, M. ALAKOMI, H.L. MÄTTÖ, J. AHONEN, A & TYNKKYNEN, S. 2011. Acid
tolerant mutants of Bifidobacterium animalis subsp. lactis with improved stability in fruit
juice. Food Science and Technology 44: 1012–1018.
SAIKI, R. SCHARF, S & FALOONA, F. 1985. Enzymatic amplification of β-globin genomic
sequences and restriction site analysis for diagnosis of sickle cell anemia. Science 230:
1350–1354.
SAMBROOK, J & RUSSEL, D. 2001. Molecular Cloning: A Laboratory Manual 3rd ed. Cold
Spring Harbor Laboratory Press. Cold Spring Harbor, NY.
SANDERS, M. 2008. Probiotics: Definition, Sources, Selection, and Uses. Clinical Infections
Diseases 46 (2): 558-561.
SATOKARI, R. VAUGHAN, E. SMIDT, H. SAARELA, M. MATTO, J & DE VOS, W. 2003.
Molecular approaches for the detection and identification of bifidobacteria and
lactobacilli in the human gastrointestinal tract. Systematic and Applied Microbiology 26:
572 -584.
SECRETARÍA DE SALUD. 1994. Norma oficial mexicana NOM-111-SSA1-1994: Bienes y
servicios. Métodos para la cuenta de mohos y levaduras en alimentos. México.
SECRETARÍA DE SALUD. 2012. Guía para la Evaluación del Desempeño de Métodos de
Prueba Microbiológicos para el Análisis de Alimentos CCAYAC-P062. México.
SHAH, N. 2000. Probiotic Bacteria: Selective Enumeration and Survival in Dairy Foods. Journal
of Dairy Science 83(4):894-907.
SHINDE, P. 2012. Probiotic: an overview for selection and evaluation. International Journal of
Pharmacy and pharmaceutical sciences 4 (2): 14-21.
SITI, A. ZAINAL, S. NORIHAM, A & NADZIRAH, K. 2013. Determination of sugar content
in pineapple waste variety N36. International Food Research Journal 20 (4): 1941- 1943.
SLAVIN, J. 2013. Fiber and prebiotics: Mechanisms and Health Benefits. Nutrients 5 (4): 1417
-1435.
SRINIVAS, B & PADMA, N. 2015. Effect of Process Parameters on Dextran Production by
Weissella confuse. International Journal for Research in Applied Science & Engineering
Technology 3 (9): 373- 377.

76
SRINIVASAN, R. KARAO, U. VOLEGOVA, M. MACKICHAN, J. KATO-MAEDA, M &
MILLER, S. 2015. Use of 16S rRNA Gene for Identification of a Broad Range of
Clinically Relevant Bacterial Pathogens. PLoS ONE 10 (2): e0117617.
doi:10.1371/journal. pone.0117617
SUN, D. 2008. Modern Techniques for food authentication. Elsevier, Irlanda.
SUN, Z. HARRIS, H. MCCANN, A. GUO, C. ARGIMÓN, S. ZHANG, W. YANG, X.
JEFFERY, I. COONEY, J. KAGAWA, T. LIU, W. SONG, W. SALVETTI, E.
WROBEL, A. RASINKANGAS, A. PARKHILL, J. REA, M. O’SULLIVAN, O.
RITARI, J. DOUILLARD, F. ROSS, R. YANG, R. BRINER, A. FELIS, G. VOS, W.
BARRANGOU, R. KLAENHAMMER, T. CAUFIELD, P. CUI, Y. ZHANG, H. &
O’TOOLE, P. 2015. Expanding the biotechnology potential of lactobacilli through
comparative genomics of 213 strains and associated genera. Nature Communication 6:
1-13.
TAMURA, K. STECHER, G. PETERSON, D. FILIPSKI, A & KUMAR, S. 2013. MEGA 6:
Molecular evolutionary genetics analysis version 6.0. Molecular Biology Evolution 30:
2725- 2729.
TANNOCK, G. 2004. A special fondness for lactobacilli. Applied Enviromental Microbiology
70:3189-3194.
TAYLOR, D. & BERKEBILE, D. 2006. Comparative efficiency of six stable fly (Diptera:
Muscidade) traps. Journal of Economic Entomology 90 (4): 1414-1419.
THOMAS, P. 2004. Isolation of Bacillus pumilus from in vitro grapes as a long- term alcohol
survivinf and rhizogenesis inducing covert endophyte. Journal of Applied Microbiology
97: 114 -123.
TOBÍA, C. URIBE, L. VILLALOBOS, E. SOTO, H & FERRIS, I. 2003. Aislamiento, selección
y caracterización de bacterias ácido lácticas en ensilajes de soya. Agronomía
Costarricense 27 (2): 21- 27.
TOMASIK, P & TOMASIK, P. 2003. Probiotics and prebiotics. Cereal Chemistry 80 (2): 113-
117.
TORINO, M. FONT, G & MOZZI, F. 2015. Biolpolymers from lactic acid bacteria: novel
applications in foods and beverages. Frontiers in microbiology 6:1 -9.

77
TRIPATHI, M & GIRI, S. 2014. Probiotic functional foods: Survival of probiotics during
processing and storage. Journal Functional Foods 9: 225–241.
TUOMOLA, E. CRITTENDEN, R. PLAYNE, M. ISOLAURI, E & SALMINEN S. 2001.
Quality assurance criteria for probiotic bacteria. American Journal of Clinical Nutrition
73: 393S–398S.
UYEN, M. 2013. Fermentation of pineapple juice by lactic acid bacteria. Can Tho University,
Vietnam.
VAGNERON, I. FAURE, G & LOEILLET, D. 2009. Is there a pilot in the chain? Identifying
the key drivers of change in the fresh pineapple sector. Food Policy 34: 437 – 446.
VALERIO, F. DE BELLIS, P. LONIGRO, S. MORELLI, L. VISCONTI, A &
LAVERMICOCCAIN, P. 2006. In vitro and in vivo survival and transit tolerance of
potentially probiotic strains carried by artichokes in the gastrointestinal tract. Applied
and Environmental Microbiology 72: 3042-3045.
VAN DE PEER, Y. CHAPELLE, S & WACHTER, R. 1996. A quantitative map of nucleotide
substitution rates in bacterial rRNA. Nucleic Acids Research 24: 3381- 3391.
VANDERZANT, C & SPLITTSTOESSER, D. 1992. Compedium of methods for the
microbiological examination of foods. 3er Ed. American Public Health Association,
Estados Unidos.
VÁZQUEZ, J & BERRÓN, S. 2004. Multilocus sequence typing: el marcador molecular de la
era de Internet. Enfermedades infecciosas y microbiología clínica 22: 113- 120.
VILLEGAS, O. VARGAS, F. PÉREZ, J. GARCÍA, R. PORRAS, S. MENESES, D. QUESADA,
A. DELGADO, G. ALPÍZAR, D. MORA, B. LEÓN, R & ALFARO, D. 2007.
Caracterización y plan de acción para el desarrollo de la agrocadena de piña en la región
Huétar Norte. Ministerio de Agricultura y Ganadería, Costa Rica.
WANG, J. LEE, L. LAI, H. WANG. S. JAN, I. YU, C. HSUEH, P & YANG, P. 2007.
Fluoroquinolone resistance in Mycobacterium tuberculosis isolates: associated genetic
mutations and relationship to antimicrobial exposure. Journal Antimicrobiology
Chemother 59: 860–865.
WEINBERG, Z & MUCK, R. 1996. New trends and opportunities in the development and use
of inoculants for silage. FEMS Microbiology Reviews 19 (1): 53-68.

78
WITTWER, C. HERRMANN, M. MOSS, A & RASMUSSEN, R. 1997. Continuous
fluorescence monitoring of rapid cycle DNA amplification. Biotechniques 22: 130–131;
134–138.
WOESE, C. 1987. Bacterial Evolution. Microbiological Reviews 51 (2): 221-271.
WOESE, C. STACKEBRANDT, E. MACKE, T & FOX, G. 1985. A phylogenetic definition of
the major eubacterial taxa. Systematic and Applied Microbiology 6: 143-51.
WOO, P. LAU, S. TENG, J. TSE, H & YUEN, K. 2008. Then and now: use of 16S rDNA gene
sequencing for bacterial identification and discovery of novel bacteria in clinical
microbiology laboratories. Clinical Microbiology and Infection 14 (10): 908-934.
ZAMBRANO, J & CASTELLANO, E. 1997. Características físico-químicas en tres variedades
de piña (Ananas comosus) durante el período postcosecha. Revista de Facultad de
Agronomía 14: 849-857.
ZOETENDAL, E. WRIGHT, A. VILPPONEN-SALMELA, T. BEN-AMOR, K.
AKKERMANS, A & DE VOS, W. 2002. Mucosa associated bacteria in the human
gastrointestinal tract are uniformly distributed along the colon and differ from the
community recovered from the faeces. Applied Enviromental Microbiology 68:3
401−3407.





























