Embed Size (px)
Citation preview

UNIVERSIDADE DO ALGARVE
FACULDADE DE CIÊNCIAS E TECNOLOGIA
APONTAMENTOS DE TÓPICOS DE FÍSICA CONTEMPORÂNEA –
MÓDULO DE BIOFÍSICA/FÍSICA MÉDICA
Curso: 1º ciclo em Física
2007/2008 - 1º Semestre, 2º módulo, 1ª semana
Docente: Carla Silva

1
RESUMO
A Biofísica e a Física Médica são áreas interdisciplinares que correspondem à aplicação de conceitos, modelos ou equipamentos desenvolvidos em Física que sejam utilizados para a compreensão dos sistemas biológicos e no desenvolvimento de diferentes valências na prática clínica. Uma vez que são tópicos que devem a sua especificidade ao objecto de estudo e não tanto ao formalismo que utilizam,1 exigemum conhecimento abrangente de diversas matérias de Física (Clássica e Moderna) e uma especial aptidão para as áreas relacionadas com a Biologia e as Ciências da Saúde. Em especial no que toca à Física Médica, a fisiologia humana toma particular relevância, uma vez que o seu principal objectivo é o desenvolvimento de ferramentas que permitam ajudar nos diagnósticos, terapias e todo o tipo de avaliação clínica.
Sendo este um módulo de uma disciplina ministrada ao nível do primeiro ano de um 1º ciclo em Física, assume-se que os estudantes possuem conhecimentos em Física e em Fisiologia apenas ao nível do ensino secundário, evitando-se o recurso a formalismos matemáticos complexos. A abordagem será feita, fundamentalmente, na perspectiva de como os conceitos desenvolvidos pela Física podem ser aplicados em áreas de impacto social tão importante como o da saúde e, nestes apontamentos, a profundidade a que a maioria dos temas são desenvolvidos é um pouco superior àquela que é exigida na disciplina. Pensa-se, desta forma, permitir ao aluno interessado em ampliar os seus conhecimentos em alguns dos domínios deste módulo a oportunidade de o fazer.
Este curso está organizado em quatro capítulos, que pretendem fornecer alguma informação sobre a função que um Físico poderá ocupar no âmbito das Ciências da Saúde, cobrir algumas técnicas de imagem utilizadas em Medicina eintroduzir a área de trabalho da docente responsável por este módulo, por forma a ilustrar um possível domínio de investigação.
Num primeiro capítulo serão abordadas, de uma forma muito superficial,algumas das áreas da Biologia e da Medicina em que o papel dos Físicos tem sido de especial relevância, enfatizando essa contribuição e reconhecendo alguns assuntos cujo desenvolvimento continua em aberto. Algumas dessas áreas serão: o estudo da difusão de substâncias nos tecidos biológicos; a dinâmica de proteínas; a construção de sensores; a utilização da radioactividade no âmbito da oncologia ou as aplicações dos lasers à Medicina.
O segundo capítulo será inteiramente dedicado às técnicas de diagnóstico que envolvem imagem. Num primeiro momento serão explicados alguns dos métodos de obtenção de imagens médicas, como sejam: o uso dos raios-X (tanto em radiografia, como em Tomografia Axial Computadorizada - TAC); o uso de marcadores nucleares (Cintigrafia, Tomografia de Fotão Único – SPECT e Tomografia de Emissão de Positrões - TEP); a ecografia (tradicional e de efeito de Doppler) e a Ressonância Magnética Nuclear – RMN (referente a imagens anatómicas e funcionais). Neste passo, será feita a distinção entre imagens estruturais e imagens funcionais e serão
1 A Mecânica, a Termodinâmica, o Electromagnetismo, a Mecânica Quântica, por exemplo, são áreas da Física que se distinguem pelo formalismo a que cada uma delas recorre, ao contrário da Biofísica ou da Física Médica que aplicam qualquer destes formalismos a diferentes sistemas biológicos ou tecnologias associadas à saúde.

2
exploradas com maior detalhe as imagens obtidas através de ultrassons e da técnica de Ressonância Magnética Nuclear2.
No terceiro e quarto capítulos serão referidas duas das mais conhecidas técnicas de medição de potenciais eléctricos no corpo humano: a electrocardiografia (ECG) e a electroencefalografia (EEG). Neles são discutidos não só alguns aspectos ao nível da instrumentação, como também do processamento posterior dos sinais medidos, dando especial relevância ao tratamento de registos correspondentes à actividade espontânea e evocada do EEG, passando pela análise dos ritmos do sono e o tipo de informação que poderá ser explorada a partir dos traçados da actividade eléctrica cardíaca.
2 As técnicas de imagem que usam radiação ionizante, como sejam a radiografia, as TAC, as SPECT e as PET serão objecto de estudo num outro módulo desta disciplina.

3
1. INTRODUÇÃO
Muitas são as áreas relacionadas com a Biologia e as Ciências da Saúde em que a aplicação da Física tem um papel preponderante ao nível da sua compreensão e desenvolvimento. Para além daquelas que iremos detalhar um pouco mais adiante em capítulos subsequentes, ao nível desta introdução pretende-se apenas enumerar alguns tópicos em que esse papel se tem mostrado particularmente interessante. Longe de elaborar uma lista exaustiva, aprofundada, ou até mesmo que foque os exemplos mais importantes, teve-se apenas como objectivo apresentar alguns temas que possam ilustrar a forma como a Física fornece abordagens, muitas vezes criativas e originais, no estudo dos sistemas biológicos.
1.1 Membranas e difusão de substâncias
Como é fácil compreender, a forma como as substâncias se movimentam nos tecidos reveste-se de particular interesse na compreensão do funcionamento do corpo humano. Por este motivo, é comum os estudiosos desta matéria recorrerem ao formalismo associado ao transporte de substâncias através de meios que poderão ser homogéneos (difusão livre) ou heterogéneos (através de membranas, por exemplo).
Relativamente à difusão livre, a experiência diz-nos que se introduzirmos uma solução colorida no interior de um líquido se observa e a cor da solução a espraiar-se gradualmente através do líquido. A este processo, em que as moléculas da solução se movimentam das regiões de maior concentração para as regiões de menor concentração, dá-se o nome de difusão.
A difusão deve-se ao caminho aparentemente aleatório que as moléculas percorrem nas suas constantes colisões. Sabe-se, com base na teoria cinética dos gases que as moléculas não só possuem uma velocidade média, como colidiam incessantemente umas com as outras. Seja L a distância média que as moléculas percorrem sem colidir, também denominado o livre percurso médio das partículas. Verifica-se, através de cálculos estatísticos, que a distância a que uma molécula se encontra do ponto de partida, após colidir N vezes é dada por:
NLS .
Repare-se que a dependência na raiz quadrada de N, em vez de uma dependência linear (como acontece em percursos lineares), deve-se ao facto de o percurso das moléculas ser essencialmente errático.
Através de cálculos muito simples e com base na expressão anterior é possível determinar o tempo necessário para uma determinada substância difundir-se através de uma distância S. Comece-se, pois, por calcular o espaço percorrido pela partícula:
L
S
L
SLLN
2
2
2
percorridoespaço .
Se a velocidade média das partículas for v, então o tempo necessário para percorrer S, será:

4
Lv
S
vt
2percorridoespaço .
Atendendo a que as molécula num líquido como a água estão muito próximas (L é muito pequeno, da ordem de 10-8 cm) e que a sua velocidade média é, à temperatura ambiente, cerca de 102 m s-1. Então o tempo necessário para uma molécula de água percorrer 1 cm, por exemplo, será:
s40m46h2s10
1010
10 4210
222
Lv
St .
Repare-se, no entanto, que, se a distância de difusão for da ordem de 10-3 cm (que é a dimensão típica de uma célula tecidular) a difusão é extremamente rápida (da ordem de 10-2 s). Por este motivo, embora a difusão macroscópica num líquido seja um processo relativamente lento, este mecanismo aplicado à troca de oxigénio, nutrientes e detritos ao nível celular é bastante eficiente.
Note-se, ainda, que o facto de nos gases as moléculas estarem mais afastadas, implica que o livre percurso médio seja maior do que nos líquidos (cerca de 3 ordens de grandeza nos gases à pressão atmosférica: cm10 5L ). Pelo que uma molécula de gás, nessas condições, demora apenas cerca de 10 s para percorrer uma distância de 1 cm.
A abordagem sugerida nos últimos parágrafos é, evidentemente, muito simples, porém, deve ter-se em atenção que os modelos utilizados para compreender o mecanismo de difusão ao nível dos sistemas biológicos pode ser, evidentemente, muito sofisticado, sendo possível ter em conta as diferentes concentrações do soluto que se difunde; diferentes pressões; várias geometrias das regiões por onde as partículas se difundem; presença de várias partículas de diferentes naturezas, etc.
Para complicar ainda mais este tipo de estudos deve ainda ponderar-se um outro factor que se prende com o facto de, parte do transporte de substâncias ao nível dos sistemas biológicos, ocorrer através de membranas. Neste caso é necessário ter em conta a permeabilidade das membranas, a qual depende, essencialmente, dos canais existentes nas mesmas, os quais são, na maioria dos casos, selectivas, ou seja, permitem a passagem de algumas substâncias, mas evitam a passagem de outras. Em particular, esta situação verifica-se com muita frequência no que respeita à água3. A este efeito selectivo associado à água dá-se o nome de osmose. Suponha que se divide um recipiente em dois através de uma membrana colocada no seu interior que só permita a passagem da água (Figura 1). Numa das divisões coloca-se água pura e na outra uma solução de água com açúcar, por exemplo. Após deixar o sistema evoluir, verifica-se que a situação de equilíbrio corresponde a uma parte da água na primeira divisória ter sido transferida para a segunda. A explicação microscópica deste fenómeno prende-se com o facto de as moléculas de água da solução terem maior dificuldade em chegar às paredes da membrana, uma vez que encontram pelo caminho as moléculas de açúcar. Por sua vez a água pura atinge as paredes da membrana com mais facilidade, o que implica que o fluxo de moléculas de água da primeira para a segunda divisória seja maior do que em sentido contrário.
3 A água é constituída por moléculas muito pequenas cuja circulação raramente é barrada pelas membranas que evitam a passagem de moléculas maiores, mas são completamente permeáveis à água.

5
início equilíbrio
Figura 1- Ilustração do fenómeno de osmose. (Adap. de J.B Marion e W.F. Hornyak, 1985).
Devido aos motivos anteriormente expressos, verifica-se que, no equilíbrio, a solução apresenta uma altura superior à da água pura (Figura 1). À quantidade ρgh(onde h é a diferença de alturas, g a aceleração da gravidade e ρ a densidade da solução) dá-se o nome de pressão osmótica da solução e representa-se pela letra grega Π. Uma forma de compreender o significado físico da pressão osmótica é entender que esta é a pressão adicional que se teria que exercer na água para que esta não atravessasse a membrana. Um resultado bastante interessante foi o encontrado por J.H. van’t Hoff ao perceber que a pressão osmótica é directamente proporcional à concentração da solução e à sua temperatura absoluta. Considerando a constante de proporcionalidade o parâmetro , obtém-se:
TnVTV
nCT ,
que é uma expressão muito semelhante à equação dos gases ideais: nRTPV , onde P é a pressão a que o gás está sujeito; V o seu volume; n o número de moles; T a temperatura absoluta e R a constante dos gases ideais. Analogia que se torna tanto mais evidente quanto o facto de apresentar um valor muito próximo ao da constante dos gases ideais R.
É ainda de referir a notável influência que o fenómeno da osmose tem ao nível do equilíbrio dos tecidos. Repare-se que todas as células são revestidas por membranas semipermeáveis o que implica um enorme equilíbrio entre as pressões osmóticas no interior e no exterior das células para que estas não percam ou ganhem demasiada água com a consequente alteração do seu volume.
Mesmo ao nível das membranas, até aqui tem-se referido o transporte espontâneo, ou seja, o transporte de massa que ocorre no sentido das maiores concentrações para as menores concentrações. No entanto, várias são as situações em que o transporte ocorre em sentido contrário com o natural consumo energético. Este tipo de transporte, ao qual se dá o nome de transporte activo, é perpetrado por proteínas de membrana as bombas que são muitas vezes responsáveis pela manutenção das concentrações iónicas vitais para o funcionamento celular. Uma vez que este tipo de transporte envolve, geralmente, o transporte de iões, o seu funcionamento está relacionado com diferenças de potencial eléctrico existente entre o interior e o exterior das membranas e, por esse motivo, será abordado com maior detalhe no capítulo em que se abordará o comportamento eléctrico dos sistemas biológicos (EEG e ECG).
Como exemplo curioso de aplicação de alguns dos conceitos que se desenvolveram neste sub-capítulo, é interessante escrever algumas palavras sobre a importância da difusão na oxigenação dos tecidos. Como é do conhecimento geral, a

6
energia necessária para a manutenção do corpo humano é fornecida essencialmente pela oxidação dos alimentos que ingerimos, sendo, portanto, crucial o consumo de oxigénio. Verifica-se que a forma mais simples de oxigenar o corpo humano é através da difusão de moléculas de oxigénio através da pele. Porém, este mecanismo é muito pouco eficiente verificando-se que no homem, por exemplo, apenas 2% das suas necessidades em oxigénio poderão ser supridas através deste processo. Os pulmões são, pois, a forma encontrada para tornar o ritmo das trocas gasosas adequado às exigências. De facto, a superfície dos alvéolos é cerca de 50 vezes superior à superfície da pele e, além disso, a distância entre os alvéolos e os capilares é cerca de 4 x 10-5 cm, pelo que a troca de oxigénio e de dióxido de carbono através da difusão entre os pulmões e os capilares é bastante rápida. Quanto ao sentido do fluxo dos gases é fácil compreender que este se processa tendo em conta as diferenças de concentração dos mesmos: no caso do oxigénio será dos pulmões para os capilares, no caso do dióxido de carbono será dos capilares para os pulmões.
1.2 Estrutura de proteínas
A forma como as proteínas adquirem a sua conformação funcional tem sido um dos temas mais interessantes que tem ocupado a comunidade científica. Apesar de ser comummente aceite que essa conformação é alcançada por ser essa a estrutura energeticamente mais favorável, o facto de 1) existirem várias conformações com energias semelhantes; 2) ser usual as proteínas mudarem ligeiramente de conformação durante os processos relacionados com a sua função e 3) existirem conformações intermédias pelas quais as proteínas passam sempre durante o processo através do qual a sua conformação nativa é atingida, faz pensar que existam mecanismos bastante mais complexos que estabeleçam essa conformação.
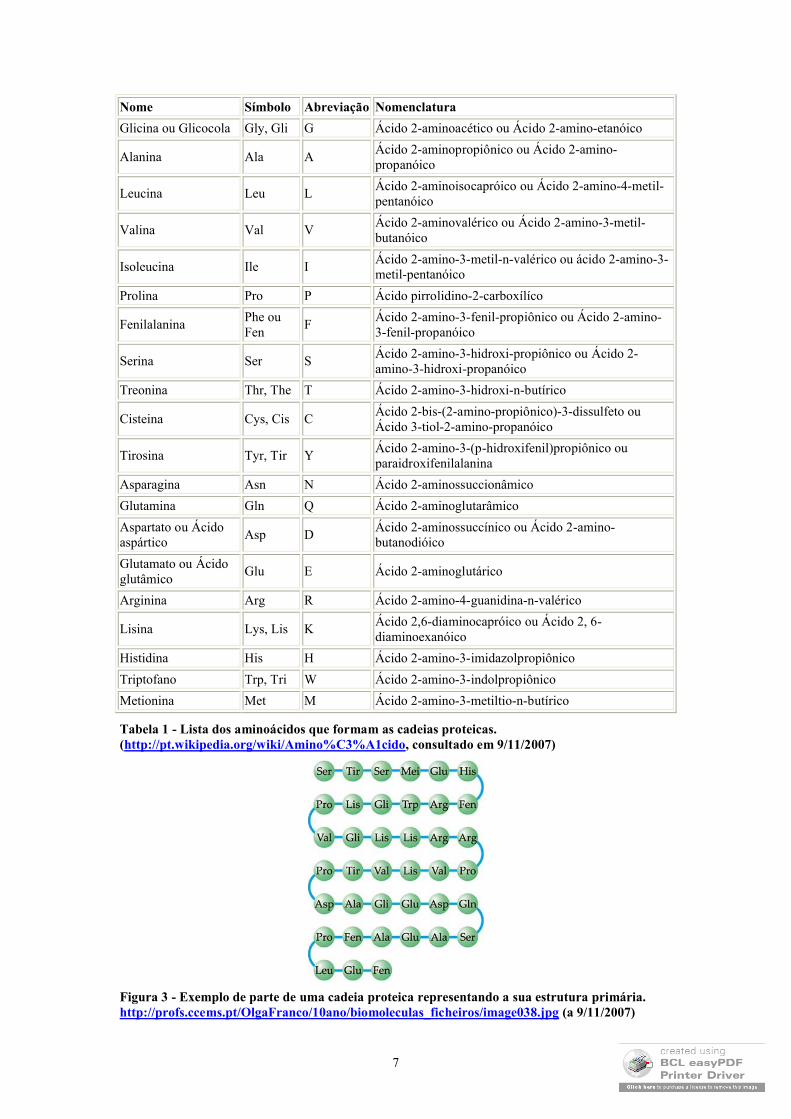
Como se sabe, as proteínas são formadas por aminoácidos (ver Tabela 1), que se ligam através de ligações covalentes: ligações peptídicas (ver Figura 2) que consistem na estrutura primária das proteínas (Figura 3).
Figura 2 - Exemplo de uma ligação peptídica entre dois aminoácidos. http://www.ciadaescola.com.br/zoom/materia.asp?materia=249 (consultado em 9/11/2007)
7
Nome Símbolo Abreviação Nomenclatura
Glicina ou Glicocola Gly, Gli G Ácido 2-aminoacético ou Ácido 2-amino-etanóico
Alanina Ala AÁcido 2-aminopropiônico ou Ácido 2-amino-propanóico
Leucina Leu LÁcido 2-aminoisocapróico ou Ácido 2-amino-4-metil-pentanóico
Valina Val VÁcido 2-aminovalérico ou Ácido 2-amino-3-metil-butanóico
Isoleucina Ile IÁcido 2-amino-3-metil-n-valérico ou ácido 2-amino-3-metil-pentanóico
Prolina Pro P Ácido pirrolidino-2-carboxílíco
FenilalaninaPhe ou Fen
FÁcido 2-amino-3-fenil-propiônico ou Ácido 2-amino-3-fenil-propanóico
Serina Ser SÁcido 2-amino-3-hidroxi-propiônico ou Ácido 2-amino-3-hidroxi-propanóico
Treonina Thr, The T Ácido 2-amino-3-hidroxi-n-butírico
Cisteina Cys, Cis CÁcido 2-bis-(2-amino-propiônico)-3-dissulfeto ou Ácido 3-tiol-2-amino-propanóico
Tirosina Tyr, Tir YÁcido 2-amino-3-(p-hidroxifenil)propiônico ou paraidroxifenilalanina
Asparagina Asn N Ácido 2-aminossuccionâmico
Glutamina Gln Q Ácido 2-aminoglutarâmico
Aspartato ou Ácido aspártico
Asp DÁcido 2-aminossuccínico ou Ácido 2-amino-butanodióico
Glutamato ou Ácido glutâmico
Glu E Ácido 2-aminoglutárico
Arginina Arg R Ácido 2-amino-4-guanidina-n-valérico
Lisina Lys, Lis KÁcido 2,6-diaminocapróico ou Ácido 2, 6-diaminoexanóico
Histidina His H Ácido 2-amino-3-imidazolpropiônico
Triptofano Trp, Tri W Ácido 2-amino-3-indolpropiônico
Metionina Met M Ácido 2-amino-3-metiltio-n-butírico
Tabela 1 - Lista dos aminoácidos que formam as cadeias proteicas. (http://pt.wikipedia.org/wiki/Amino%C3%A1cido, consultado em 9/11/2007)
Figura 3 - Exemplo de parte de uma cadeia proteica representando a sua estrutura primária. http://profs.ccems.pt/OlgaFranco/10ano/biomoleculas_ficheiros/image038.jpg (a 9/11/2007)
8
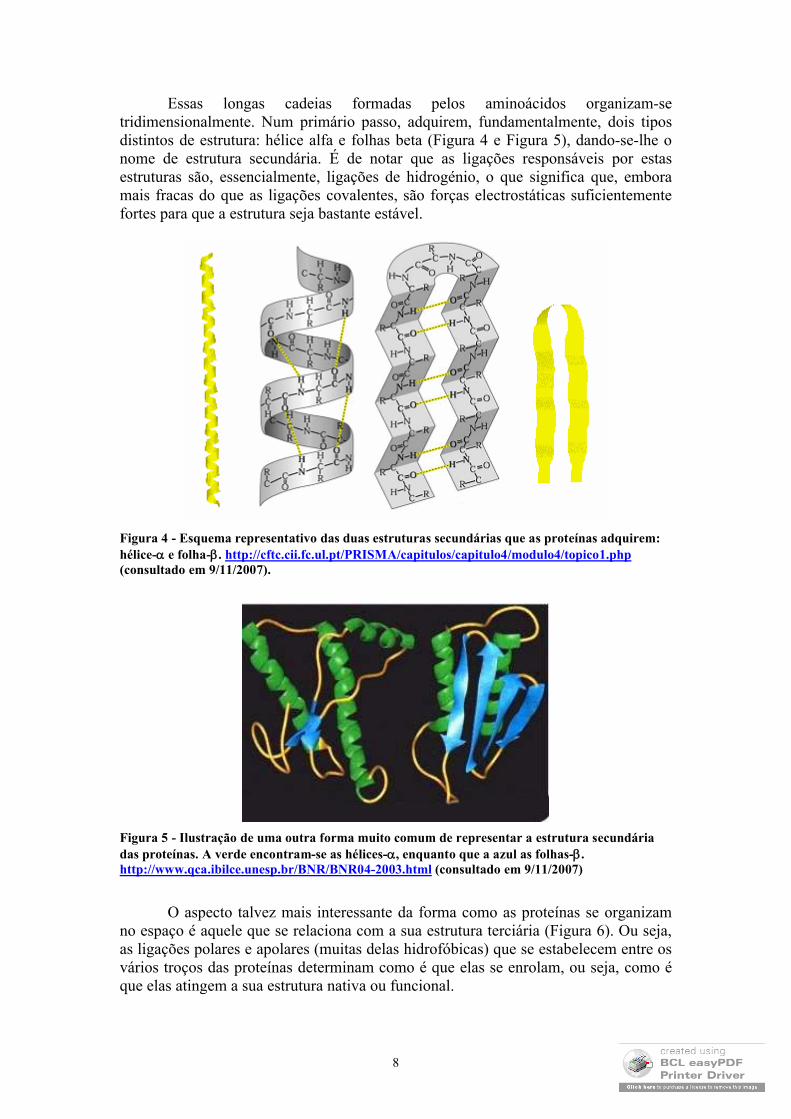
Essas longas cadeias formadas pelos aminoácidos organizam-se tridimensionalmente. Num primário passo, adquirem, fundamentalmente, dois tipos distintos de estrutura: hélice alfa e folhas beta (Figura 4 e Figura 5), dando-se-lhe o nome de estrutura secundária. É de notar que as ligações responsáveis por estas estruturas são, essencialmente, ligações de hidrogénio, o que significa que, embora mais fracas do que as ligações covalentes, são forças electrostáticas suficientemente fortes para que a estrutura seja bastante estável.
Figura 4 - Esquema representativo das duas estruturas secundárias que as proteínas adquirem: hélice- e folha-. http://cftc.cii.fc.ul.pt/PRISMA/capitulos/capitulo4/modulo4/topico1.php(consultado em 9/11/2007).
Figura 5 - Ilustração de uma outra forma muito comum de representar a estrutura secundária das proteínas. A verde encontram-se as hélices-, enquanto que a azul as folhas-.http://www.qca.ibilce.unesp.br/BNR/BNR04-2003.html (consultado em 9/11/2007)
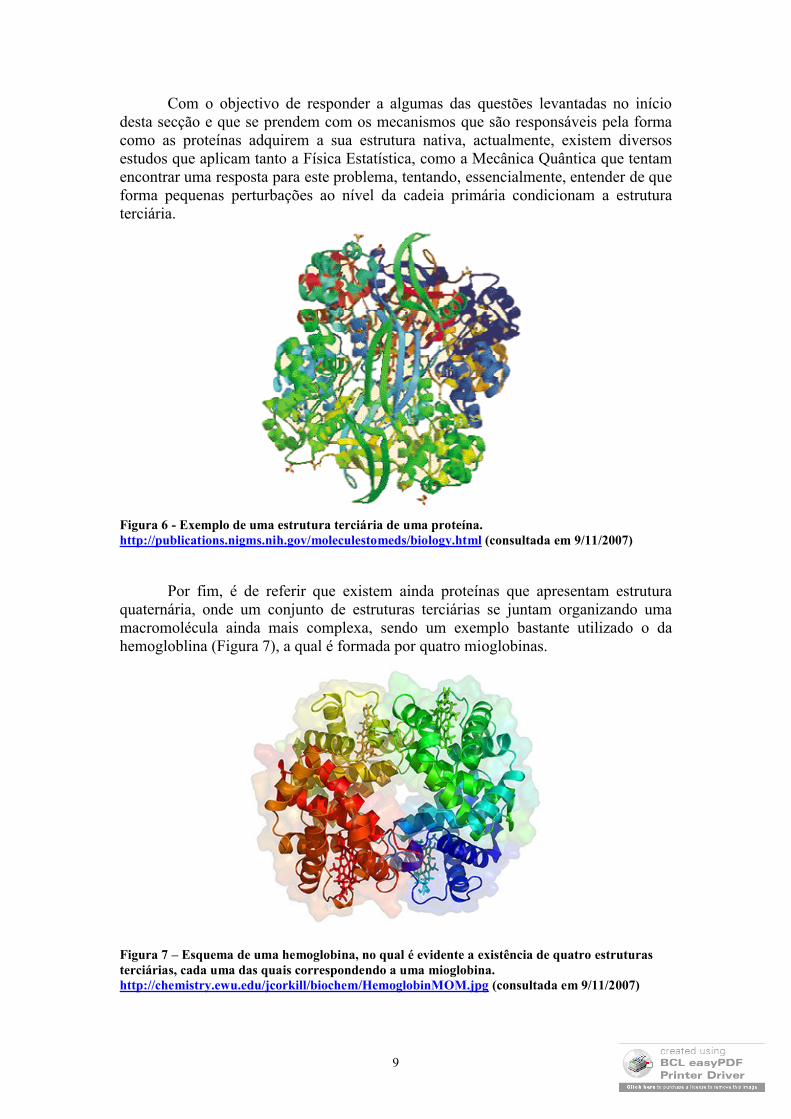
O aspecto talvez mais interessante da forma como as proteínas se organizam no espaço é aquele que se relaciona com a sua estrutura terciária (Figura 6). Ou seja, as ligações polares e apolares (muitas delas hidrofóbicas) que se estabelecem entre os vários troços das proteínas determinam como é que elas se enrolam, ou seja, como é que elas atingem a sua estrutura nativa ou funcional.
9
Com o objectivo de responder a algumas das questões levantadas no início desta secção e que se prendem com os mecanismos que são responsáveis pela forma como as proteínas adquirem a sua estrutura nativa, actualmente, existem diversos estudos que aplicam tanto a Física Estatística, como a Mecânica Quântica que tentam encontrar uma resposta para este problema, tentando, essencialmente, entender de que forma pequenas perturbações ao nível da cadeia primária condicionam a estrutura terciária.
Figura 6 - Exemplo de uma estrutura terciária de uma proteína. http://publications.nigms.nih.gov/moleculestomeds/biology.html (consultada em 9/11/2007)
Por fim, é de referir que existem ainda proteínas que apresentam estrutura quaternária, onde um conjunto de estruturas terciárias se juntam organizando uma macromolécula ainda mais complexa, sendo um exemplo bastante utilizado o da hemogloblina (Figura 7), a qual é formada por quatro mioglobinas.
Figura 7 – Esquema de uma hemoglobina, no qual é evidente a existência de quatro estruturas terciárias, cada uma das quais correspondendo a uma mioglobina. http://chemistry.ewu.edu/jcorkill/biochem/HemoglobinMOM.jpg (consultada em 9/11/2007)

10
1.3 Sensores médicos
Os instrumentos médicos podem ter funções de medida, monitorização, cirurgia ou terapia. E um dos elementos mais importantes destes aparelhos é, na maioria das vezes, os seus sensores. Um sensor é um elemento de medida que transforma ou amplifica uma determinada variável num parâmetro que é mais facilmente processado. Uma vez que tanto o processamento de sinal como o seu registo são feitos, cada vez mais, através da electrónica, os sensores tendem a ser transdutores que transformam a grandeza que se pretende medir em tensão.
Uma das dificuldades da construção de sensores biomédicos é a de, num grande número de casos, a gama na qual se pretende medir a grandeza ser muito larga, pelo que é exigido que a dependência entre a grandeza medida e a registada seja a mesma, em geral, linear, numa vasta gama de valores. Em seguida far-se-á uma resenha de alguns dos mais importantes transdutores utilizados em Medicina, dividindo-os segundo o tipo de variável que medem: sensores de grandezas mecânicas, de temperatura e eléctrodos. Nesta enumeração não são referidos os sensores químicos que detectam, por exemplo, as concentrações de pH e de gases, como o oxigénio e o dióxido de carbono, e os sensores bioquímicos constituídos por um sistema de reconhecimento específico, como um enzima ou um anticorpo, e uma estrutura que transforma uma reacção química numa grandeza mensurável, ou seja, num sinal óptico, eléctrico ou mecânico.1.3.1 Sensores de Grandezas Mecânicas
Os sensores de grandezas mecânicas são particularmente úteis no estudo dos músculos, do sistema circulatório e respiratório. Contam-se como os principais sensores desta categoria:
i) Sensores de tensão mecânica/pressão – Nestes sensores é utilizado o efeito piezoresistivo para medir tensões mecânicas, ou seja, o facto da resistência de um metal ou de um semicondutor ser dependente da pressão a que está sujeita. O principal problema destes sensores é o serem muito dependentes da temperatura, pelo que é necessário compensar este aspecto. Este tipo de sensores é utilizado, por exemplo, para medição directa da pressão sanguínea, ou seja, o detector de pressão é colocado no interior da via onde se pretende determinar a pressão. É de salientar, no entanto, que esta não é a forma mais usual de medir a pressão sanguínea, para o que se usa, habitualmente um método indirecto, utilizando um esfingmomanómetro que, associado a um estetoscópio, permite medir as pressões máxima e mínima (na mínima, o sangue começa a passar, mas com um fluxo turbulento e, na máxima, o sangue deixa simplesmente de passar).
ii) Potenciómetro – Transforma deslocamentos lineares ou rotacionais numa diferença de potencial, através do contacto com um elemento resistivo e usando a lei de Ohm. Em geral, a resolução destes sensores é limitada pelo tamanho das espiras de que a resistência é feita.
iii)Sensor de deslocamento indutivo – Considerando que a indutância mútua se calcula através da expressão L = N2r2/l, onde N é o número de espiras, r é o raio da espira, l o tamanho da espira e a permeabilidade do núcleo é possível determinar deslocamentos através do movimento no interior do núcleo de uma peça, em geral, ferrosa. Com este procedimento altera-se e, por conseguinte, Lpassa a ser uma medida do deslocamento.
iv)Condensador de placas – a capacidade de um condensador de placas é dada por: C = A/d, onde é a permitividade dieléctrica entre as placas; A é a área das

11
placas e d a distância entre elas. Deste modo, variando d, varia-se a capacidade do condensador.
v) Sensor de efeito piezoeléctrico – Estes sensores baseiam-se no efeito através do qual um cristal sujeito a uma força F desenvolve uma carga Q que cumpre a relação: Q = DF, onde D é o coeficiente piezoeléctrico do material. A força aplicada pode assim ser medida através de um amplificador de carga ou medindo a diferença de potencial, uma vez que esta é dada por V = Q/C, onde Cé a capacidade do cristal que pode ser calculada usando a expressão do condensador de placas.Estes sensores são, por exemplo, utilizados para determinar a velocidade sanguínea quer através de fluxómetros de tempo de trânsito como através do efeito de Doppler.
vi)Fluxómetros electromagnéticos – Estes instrumentos consideram o facto de um condutor em movimento com uma velocidade v, perpendicular a um campo magnético com densidade de fluxo B, gerar uma diferença de potencial que cumpre: V = 2rBv, onde r é o raio do condutor, considerado cilíndrico. No caso de se pretender medir a velocidade do sangue, este funciona como o condutor que se move e r, será, então, o raio da artéria.
1.3.2 Sensores de TemperaturaOs termómetros são aparelhos muito utilizados na clínica, uma vez que a
temperatura do corpo é uma grandeza muito constante e o seu desequilíbrio corresponde, geralmente, a uma situação de infecção. Para além desta utilização os termómetros têm ainda a função de monitorizar a temperatura dos tecidos irradiados por radiação laser. Para além dos populares termómetros que utilizam a variação de volume de um líquido com a temperatura, pode ainda considerar-se como aplicados à Medicina, os termómetros:
i) Termoresistivos – Estes sensores utilizam a variação da resistência dos metais e dos semicondutores com a temperatura, que, para variações pequenas de temperatura, segue a expressão: R = Ro[1 + (T - To)], onde Ro é a resistência a uma temperatura padrão, To, e o coeficiente de temperatura que pode ser positivo ou negativo. Estes sensores são parte integrante de um circuito electrónico que é, basicamente, um amplificador operacional.
ii) Termoeléctricos – Estes termómetros são os chamados termopares e utilizam o efeito de Seebeck. Este efeito indica que se forem considerados três troços de dois metais diferentes ligados através de duas junções, onde uma se encontra a uma temperatura padrão T1 e a outra à temperatura que se pretende medir T2, é gerada uma diferença de potencial que cumpre: V = (T1 - T2) + (T1
2 - T22). Se
a gama de temperaturas for pequena, o termo quadrático pode ser desprezável e, portanto, a diferença de potencial vem proporcional à diferença de temperaturas.
1.3.3 EléctrodosExistem inúmeras situações em que são necessárias medidas eléctricas na
clínica, especialmente aplicadas ao sistema nervoso e aos músculos, em particular ao músculo cardíaco. Os principais sensores nesta área são os eléctrodos, que se dividem em macroeléctrodos e microeléctrodos. Os primeiros medem a actividade de um tecido ou órgão, enquanto que os segundos medem a actividade ao nível celular. Refira-se que uma vez que os eléctrodos medem diferenças de potencial é necessário estabelecer-se um local para a colocação do eléctrodo de referência, o qual não pode ser excessivamente afastado da actividade que se pretende medir, pois corre-se o risco de ser sensível à actividade de outros tecidos, nem excessivamente perto para que não seja contaminado com a actividade em estudo.

12
i) Macroeléctrodos – Os macroeléctrodos que se mantêm em contacto com a pele podem ser de ligação metal/electrólito ou de ligação capacitiva. No primeiro caso tem-se uma peça metálica e um gel electrolítico que estabelece a ligação entre o tecido e o metal. Quanto aos eléctrodos capacitivos o contacto do metal com a pele é feito através de um isolador, por exemplo ar, e o que é medido é a corrente de deslocamento, em vez da corrente relativa ao movimento de cargas.
ii) Microeléctrodos – Existem basicamente três tipos de microeléctrodos: 1) aqueles que estão no interior de um capilar de vidro e mergulhados num electrólito que faz o contacto com a célula através de um orifício na ponta do capilar; 2) os totalmente metálicos revestidos por um isolador, que estabelecem o contacto directamente com a célula, através de um orifício no referido revestimento e 3) eléctrodos de estado sólido que são fabricados utilizando técnicas idênticas às de construção dos circuitos integrados.
1.4 Radioterapia usando radiação ionizante
O cancro, como é do conhecimento geral, é uma das principais causas de morte, pelo que os seus tratamentos tomam especial relevância. Deve chamar-se a atenção para o facto de, em muitos casos, se desenvolverem curas para o cancro, sem antes se ter conhecimento detalhado do seu mecanismo, dada a urgência de se ‘fazer algo’ pelo indivíduo afectado e não haver tempo para a compreensão pormenorizada das alterações que ocorrem ao nível das células malignas, uma vez que estas são extremamente complexas!
De uma forma simples, pode dizer-se que as células cancerosas são células cuja função se encontra alterada, não respondendo correctamente aos estímulos químicos que recebem das células vizinhas. Além disso, são, geralmente, células que se reproduzem muito rapidamente provocando tumores, sendo melhor ou pior diferenciadas dependendo do grau de parecença com as células das quais derivam.
Os métodos de tratamento de cancro são tanto melhores quanto melhorconseguirem eliminar as células cancerosas, mantendo intactas as normais.
As principais terapias do cancro são, basicamente, de três tipos: cirurgia, quimioterapia e terapias que usam radiação ionizante, podendo ser usadas separadamente ou em conjunto num regime de tratamento planeado pelo(s) médico(s).
No caso da radioterapia aceita-se como taxa de sucesso a percentagem de 65% de doentes que reagem favoravelmente ao tratamento (doentes que sobrevivem durante, pelo menos, 3 anos), sendo este relativo sucesso devido ao facto de as células cancerosas serem muito mais susceptíveis à radiação do que as células normais. Refira-se a este respeito que mesmo entre as células cancerosas existem as que são mais sensíveis à radiação do que outras, pelo que para a mesma dose, os efeitos podem ser muito diferentes. Há ainda uma questão digna de nota e que se prende com o facto de as células que se reproduzem mais rapidamente serem aquelas que mais sensíveis são à radiação, isto porque durante a mitose (forma de reprodução das células) estas se tornam mais vulneráveis, como se discutirá adiante.
Deve ainda atender-se a que a radioterapia pode ter duas acções: a radical, na qual se pretende irradicar completamente o cancro, e a paliativa, quando apenas se pretende diminuir a dor e outros sintomas desagradáveis em doentes em estados terminais da doença.
Atente-se ainda no facto de as doses poderem ser dadas de uma só vez (em geral quando o volume a eliminar é pequeno) ou em fracções (quando o volume é maior). Esta estratégia permite às células normais uma certa recuperação, sem a qual
13
não tolerariam toda a dose necessária para eliminar as células cancerosas. É de salientar, a este respeito, que também as células cancerosas exibem uma certa recuperação entre fracções do tratamento e, por isso, se deve ir aumentando ligeiramente a dose em cada sessão, para que se obter o mesmo efeito.
1.4.1 RadioterapiaConcentrando-nos na utilização das radiações no tratamento do cancro pode
considerar-se dois tipos de abordagem: a radioterapia ou teleterapia, quando a fonte de radiação se encontra no exterior do organismo, ou braquiterapia, quando a fonte é colocada em contacto com os tecidos, nalguns casos, no interior do organismo.
Existem diversos tipos de radiação que podem ser utilizados com estes propósitos, como raios-X e raios-, mas também electrões, neutrões, protões e mesões-pi. No entanto, aqueles que são mais extensivamente utilizados, são, indubitavelmente, os que envolvem radiação electromagnética (raios-X e ), pelo que serão esses que serão aqui discutidos.
Dado que a penetração da radiação electromagnética nos tecidos é tanto maior quanto maior for a sua energia, as fontes menos energéticas são, em geral, utilizadas para tratar lesões superficiais, enquanto que as mais energéticas são utilizadas para lesões profundas.
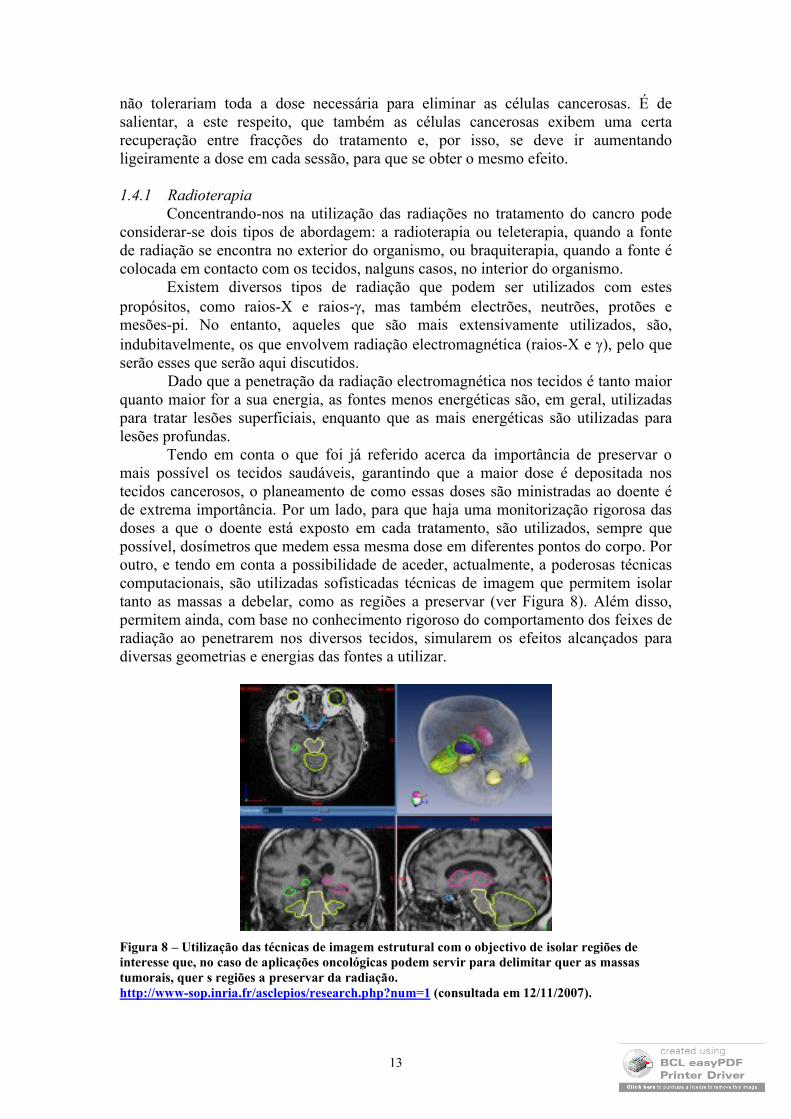
Tendo em conta o que foi já referido acerca da importância de preservar o mais possível os tecidos saudáveis, garantindo que a maior dose é depositada nos tecidos cancerosos, o planeamento de como essas doses são ministradas ao doente é de extrema importância. Por um lado, para que haja uma monitorização rigorosa das doses a que o doente está exposto em cada tratamento, são utilizados, sempre que possível, dosímetros que medem essa mesma dose em diferentes pontos do corpo. Por outro, e tendo em conta a possibilidade de aceder, actualmente, a poderosas técnicas computacionais, são utilizadas sofisticadas técnicas de imagem que permitem isolar tanto as massas a debelar, como as regiões a preservar (ver Figura 8). Além disso, permitem ainda, com base no conhecimento rigoroso do comportamento dos feixes de radiação ao penetrarem nos diversos tecidos, simularem os efeitos alcançados para diversas geometrias e energias das fontes a utilizar.
Figura 8 – Utilização das técnicas de imagem estrutural com o objectivo de isolar regiões de interesse que, no caso de aplicações oncológicas podem servir para delimitar quer as massas tumorais, quer s regiões a preservar da radiação.http://www-sop.inria.fr/asclepios/research.php?num=1 (consultada em 12/11/2007).

14
Figura 9 – Ilustração de um planeamento de radioterapia utilizando técnicas de imagem e simulações das doses, tendo em conta a geometria das fontes externas utilizadas. As linhas correspondem a regiões de igual dose. http://www.prism.co.uk/main.htm (consultada em 12/11/2007).
1.4.2 BraquiterapiaDá-se o nome de braquiterapia à terapia que usa radiação nuclear quando a
fonte é colocada sobre o tumor. Este tipo de tratamento pode ser classificado em superficial, intersticial ou em cavidades. A grande vantagem da braquiterapia relativamente à teleterapia é o facto de a dose ser mais concentrada na região do tumor, atendendo à lei do inverso do quadrado da distância.
No primeiro caso, contam-se, essencialmente, os cancros da pele, dos lábios ou dos olhos, sendo a fonte colocada a 0.5-1cm de distância da lesão. No caso de braquiterapia intersticial a fonte radioactiva é introduzida no órgão em tratamento por intermédio de agulhas. Enquanto que quando o tumor é localizado em cavidades naturais do corpo, as fontes são conduzidas através de tubos que são introduzidos nessas cavidades.
Neste tipo de terapia o Rádio tem vindo a ser substituído por Cobalto e Césio, uma vez que o primeiro não só tem uma energia muito elevada, como apresenta níveis elevados de toxicidade quer do ponto de vista químico, quer radioactivo, basta lembrar que o seu tempo de mais vida é de 1620 anos.
A principal desvantagem desta técnica é o risco acrescido para o pessoal médico, para-médico e auxiliar que manuseia as fontes radioactivas. Com o intuito de minorar estes efeitos, têm sido desenvolvidas várias técnicas onda as fontes só são introduzidas após a introdução dos tubos ou das agulhas nos locais apropriados, minimizando assim, o tempo durante o qual os sujeitos saudáveis são expostos à radiação.
No caso da braquiterapia intersticial a fonte a utilizar tem sido a de Irídio, uma vez que é um elemento bastante flexível o que permite a construção de agulhas suficientemente estreitas de modo a serem introduzidas nas agulhas um pouco mais largas que são previamente introduzidas no órgão com o recurso, caso seja necessário, a técnicas de imagem.
Nos tratamentos oncológicos internos, são os tubos que são primeiramente introduzidos, enquanto que as fontes, de forma esférica, ou oval, são colocadas no interior dos tubos através de pequenas pressões e retiradas recorrendo a vácuos fracos. Este mecanismo pode ser automatizado, evitando qualquer contacto do pessoal médico com as fontes radioactivas, uma vez que o comando do sistema pode ser for a da sala. Muitos destes sistemas são já construídos de forma a recolherem as fontes

15
caso a porta da sala se abra. O contacto com o doente é mantido quer visual, quer auditivamente.
Mais uma vez o equipamento informático é de extrema importância para o planeamento das doses a ministrar, podendo as fontes radioactivas estarem misturadas com elementos que não o são, apenas para obter a geometria pretendida.
Para terminar este capítulo deve-se referir a possibilidade de usar radiação como terapia de doenças não cancerosas. Existem dois exemplos paradigmáticos, que são o hipertiroidismo e uma doença de sangue caracterizada pelo aumento da viscosidade sanguínea, devido a um excesso de glóbulos vermelhos no sangue. Em ambos os casos a radiactividade é utilizada para diminuir a função de determinado órgão: no primeiro caso tem-se uma redução da função da tiroide, no segundo uma redução de produção de glóbulos vermelhos.
1.5 Lasers aplicados à Medicina
A radiação electromagnética proveniente dos lasers pode abranger a gama do espectro entre o infra-vermelho e o ultra-violeta, numa gama de frequências de, aproximadamente, 1014 Hz a 1016 Hz. Ora estas radiações, à excepção das de mais alta frequência, possuem energias que não são suficientes para quebrar as ligações químicas das moléculas. De forma que os seus principais efeitos são ao nível do aumento local da temperatura dos tecidos, verificando-se que estes efeitos são extremamente dependentes da forma como essa energia for depositada nos tecidos. Com o objectivo de dispor de grandezas que permitam avaliar estes dois factores: energia depositada e tempo durante o qual essa energia foi transferida, são definidas duas grandezas - a densidade de potência e a fluência. A primeira tem a expressão:
A
PI ,
onde P é a potência do laser (energia emitida por unidade de tempo) e A a área da secção recta do feixe. E a segunda:
EITF
onde TE e o tempo de exposição.
Analisando as duas expressões anteriores, pode concluir-se que, para a mesma densidade de potência, quanto menor a fluência, mais rápido será o processo de deposição de energia.
Existem, pois, dois efeitos distintos associados a diferentes formas de deposição de energia térmica nos tecidos através de irradiação com luz laser: a fotocoagulação (quando as densidades de potência dos lasers são menores e, portanto a deposição da mesma quantidade de energia é mais lenta) e a fotovaporização(quando as densidade de potência são maiores).
A fotocoagulação é, em certa medida, uma espécie de cozedura. Na verdade, os efeitos sobre os tecidos são muito semelhantes ao que acontece, por exemplo, ao cozermos um naco de carne. O aumento de temperatura irá conduzir à desnaturação das proteínas, ou seja, à perda da sua estrutura terciária, tornando-as disfuncionais. Em termos de aspecto físico, e apelando à nossa experiência quotidiana sabemos, por exemplo que a carne vermelha cozida, perde a sua cor tornando-se castanho

16
acinzentada. Este fenómeno deve-se, precisamente, à desnaturação da hemoglobina e da mioglobina (proteínas que são responsáveis pelo transporte do sangue nos vasos sanguíneos e nos músculos, respectivamente, e que conferem ao sangue a cor vermelha). Sabemos ainda que a carne depois de cozinhada se torna mais fácil de rasgar, devido à desnaturação do colagénio, proteína de estrutura de grande parte dos tecidos orgânicos. E, por fim, já todos tivemos a infeliz decepção de preparar uma refeição para várias pessoas e o naco de carne que cozinhámos, ter diminuído visivelmente, o seu volume. O que corresponde, precisamente, à evaporação (ainda que lenta) de uma parte significativa da água que o constituía. Tendo em conta estas considerações, é fácil compreender que a fotocoagulação pode ser utilizada para destruir tecidos, infligindo-lhes desnaturação das proteínas que os constituem. Embora se vá discutir adiante várias aplicações médicas, pode desde já adiantar-se que a fotocoagulação é utilizada, essencialmente, na destruição de tumores, em tratamentos da retina e como forma de evitar hemorragias (os vasos sanguíneos fotocoagulados são como que ‘selados’ e evitam a perda de sangue).
Apesar de a fotocoagulação envolver a deposição lenta de energia térmica nos tecidos, deve ter-se presente que esse processo não pode demorar demasiado tempo, caso contrário, corre-se o risco de o calor se transferir para os tecidos em redor e, por um lado, não ser eficiente relativamente aos efeitos pretendidos na região de acção, por outro, ir lesar tecidos adjacentes sobre os quais não se desejava agir. A este respeito, é pertinente introduzir-se um outro conceito que é o de tempo de relaxação térmico. É representado por TR e é o tempo necessário para que o calor depositado numa determinada região seja conduzido para fora dessa região, de forma a que a temperatura aumentada no tecido exposto diminua para metade. Com base nesta definição, compreende-se que para que a acção do laser seja eficiente deve cumprir-se a condição: Te<<TR.
Além disso, observou-se que as densidades de potência associadas ao efeito de fotocoagulação estão na gama entre 10 W/cm2 e 100 W/cm2, de modo que os lasers utilizados nestas aplicações possuem estas características.
Quando as densidades de potência dos lasers são superiores a 100 W/cm2 o efeito mais evidente é o de fotovaporização, ou seja, a temperatura de ebulição da água é rapidamente atingida e os tecidos são cortados. Por este motivo, estes lasers têm aplicações essencialmente ao nível cirúrgico, podendo funcionar como bisturi, ou como uma forma de remover tecidos extremamente precisa. Esta aplicação tem ainda a vantagem adicional de os tecidos adjacentes ao corte sofrerem fotocoagulação, evitando, desta forma, o surgimento de hemorragias.
Ainda no domínio dos lasers com densidades de potência elevadas, há a considerar aqueles cuja gama de frequências se situa no ultravioleta e cujas energia já interferem ao nível das ligações químicas. Neste caso, a transferência de energia não implica aumento da temperatura, sendo o corte dos tecidos devido à quebra das ligações químicas. A este fenómeno dá-se o nome de fotoablação.
A discussão sobre a forma de interacção da luz com os tecidos só fica completa referindo o facto de a absorção da luz pelos tecidos ser selectiva. Ou seja, como se sabe, algumas substâncias absorvem especificamente num dado c.d.o.,podendo essa especificidade ser utilizada em diferentes cenários.
Antes de mais, relembremo-nos que o espectro de absorção das moléculas é muito mais complexo do que o dos átomos, parecendo contínuos. O que significa que, embora absorvam preferencialmente em determinadas frequências o seu espectro não apresenta picos bem definidos como no caso dos átomos. No que respeita à constituição dos tecidos, pode dizer-se estes são formados por cerca de 70% de água e
17
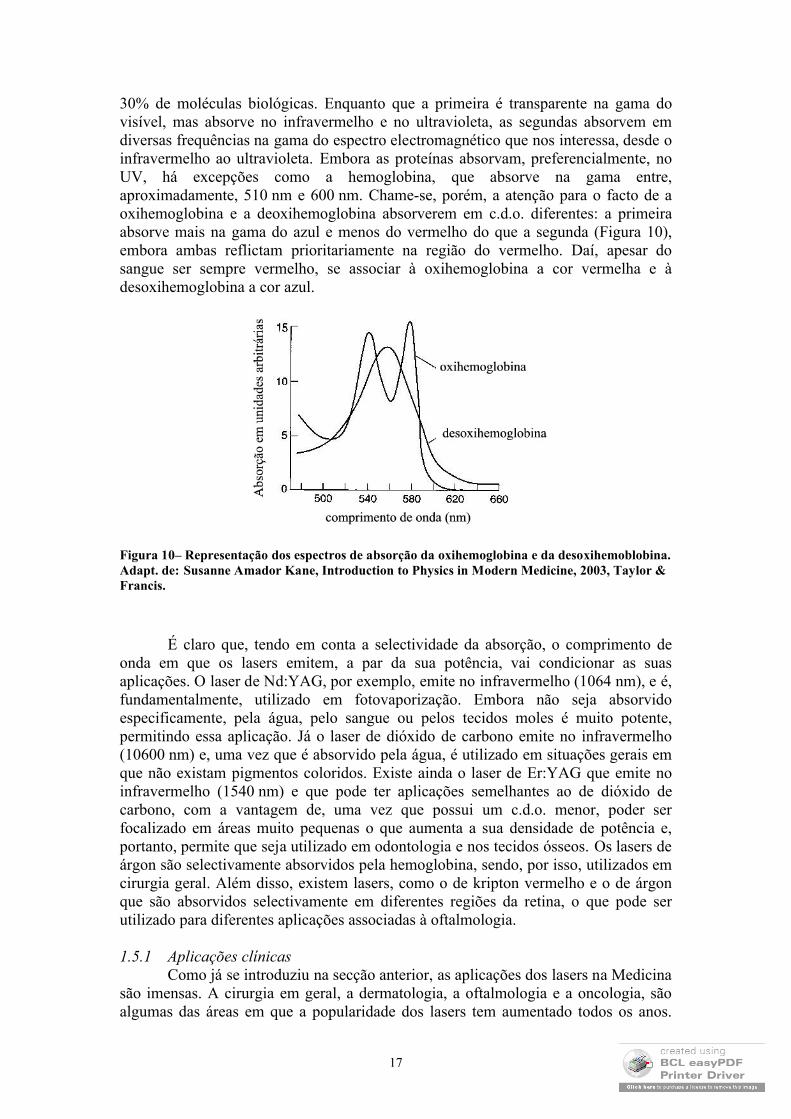
30% de moléculas biológicas. Enquanto que a primeira é transparente na gama do visível, mas absorve no infravermelho e no ultravioleta, as segundas absorvem em diversas frequências na gama do espectro electromagnético que nos interessa, desde o infravermelho ao ultravioleta. Embora as proteínas absorvam, preferencialmente, no UV, há excepções como a hemoglobina, que absorve na gama entre, aproximadamente, 510 nm e 600 nm. Chame-se, porém, a atenção para o facto de aoxihemoglobina e a deoxihemoglobina absorverem em c.d.o. diferentes: a primeira absorve mais na gama do azul e menos do vermelho do que a segunda (Figura 10), embora ambas reflictam prioritariamente na região do vermelho. Daí, apesar do sangue ser sempre vermelho, se associar à oxihemoglobina a cor vermelha e à desoxihemoglobina a cor azul.
Figura 10– Representação dos espectros de absorção da oxihemoglobina e da desoxihemoblobina. Adapt. de: Susanne Amador Kane, Introduction to Physics in Modern Medicine, 2003, Taylor & Francis.
É claro que, tendo em conta a selectividade da absorção, o comprimento de onda em que os lasers emitem, a par da sua potência, vai condicionar as suas aplicações. O laser de Nd:YAG, por exemplo, emite no infravermelho (1064 nm), e é, fundamentalmente, utilizado em fotovaporização. Embora não seja absorvido especificamente, pela água, pelo sangue ou pelos tecidos moles é muito potente, permitindo essa aplicação. Já o laser de dióxido de carbono emite no infravermelho (10600 nm) e, uma vez que é absorvido pela água, é utilizado em situações gerais em que não existam pigmentos coloridos. Existe ainda o laser de Er:YAG que emite no infravermelho (1540 nm) e que pode ter aplicações semelhantes ao de dióxido de carbono, com a vantagem de, uma vez que possui um c.d.o. menor, poder ser focalizado em áreas muito pequenas o que aumenta a sua densidade de potência e, portanto, permite que seja utilizado em odontologia e nos tecidos ósseos. Os lasers de árgon são selectivamente absorvidos pela hemoglobina, sendo, por isso, utilizados em cirurgia geral. Além disso, existem lasers, como o de kripton vermelho e o de árgon que são absorvidos selectivamente em diferentes regiões da retina, o que pode ser utilizado para diferentes aplicações associadas à oftalmologia.
1.5.1 Aplicações clínicasComo já se introduziu na secção anterior, as aplicações dos lasers na Medicina
são imensas. A cirurgia em geral, a dermatologia, a oftalmologia e a oncologia, são algumas das áreas em que a popularidade dos lasers tem aumentado todos os anos.
18
Em dermatologia, por exemplo, contam-se, por exemplo, as seguintes aplicações: 1) Remoção de cancros da pele. O laser de dióxido de carbono é muito utilizado para este efeito, actuando ao nível da remoção dos tecidos. 2) Na cirurgia estética. O mesmo tipo de laser pode ser utilizado para retirar camadas de pele muito finas, permitindo o rejuvenescimento de tecidos que tenham sido, por exemplo, queimados do sol. 3) Na cosmética. O laser de Er: YAG, por exemplo, é muito utilizado em depilação dita definitiva. 4) Ainda em cosmética, os lasers podem ser utilizados para remoção de manchas pigmentadas. Os lasers de corantes com c.d.o. no amarelo têm sido utilizados para destruir os vasos sanguíneos responsáveis pelas manchas tipo vinho do porto. E o laser de Nd:YAG e de rubi são utilizados na remoção de tatuagens. A este respeito é de referir que quando a cor das tatuagens coincide com a da hemoglobina e da melanina o tratamento é mais difícil, uma vez que implica também a destruição de tecidos saudáveis e sem tatuagem…
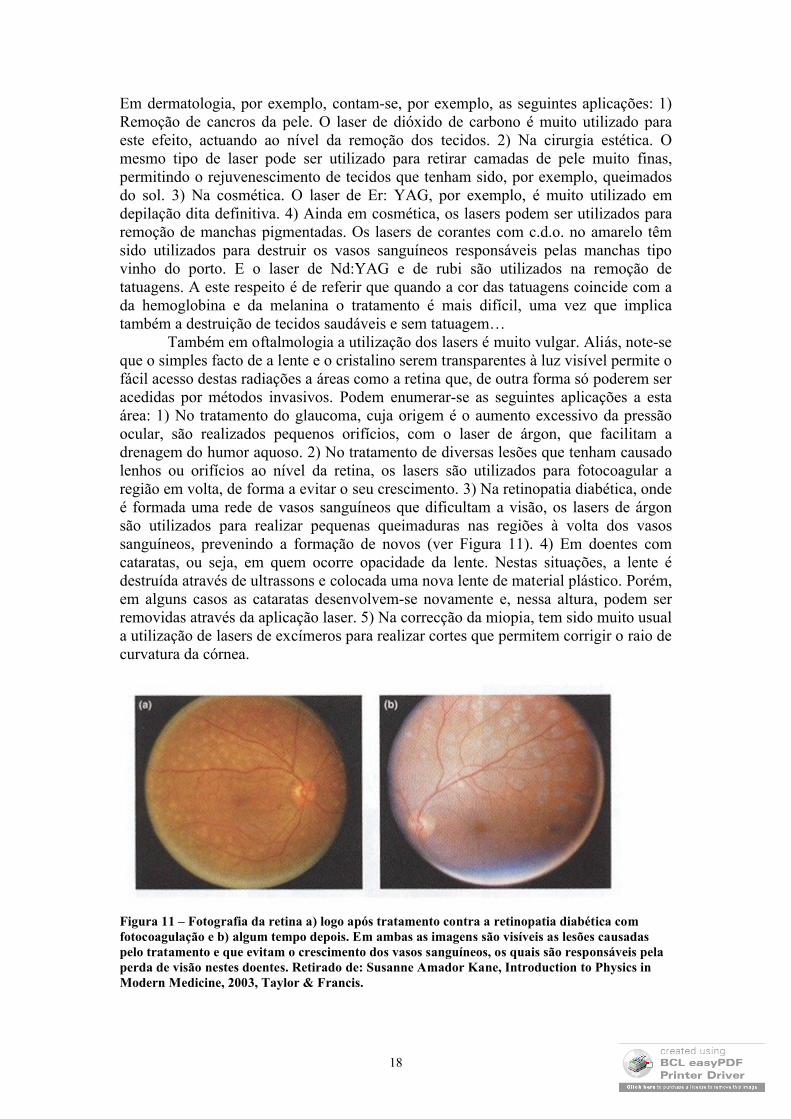
Também em oftalmologia a utilização dos lasers é muito vulgar. Aliás, note-se que o simples facto de a lente e o cristalino serem transparentes à luz visível permite o fácil acesso destas radiações a áreas como a retina que, de outra forma só poderem ser acedidas por métodos invasivos. Podem enumerar-se as seguintes aplicações a esta área: 1) No tratamento do glaucoma, cuja origem é o aumento excessivo da pressão ocular, são realizados pequenos orifícios, com o laser de árgon, que facilitam a drenagem do humor aquoso. 2) No tratamento de diversas lesões que tenham causado lenhos ou orifícios ao nível da retina, os lasers são utilizados para fotocoagular a região em volta, de forma a evitar o seu crescimento. 3) Na retinopatia diabética, onde é formada uma rede de vasos sanguíneos que dificultam a visão, os lasers de árgon são utilizados para realizar pequenas queimaduras nas regiões à volta dos vasos sanguíneos, prevenindo a formação de novos (ver Figura 11). 4) Em doentes com cataratas, ou seja, em quem ocorre opacidade da lente. Nestas situações, a lente é destruída através de ultrassons e colocada uma nova lente de material plástico. Porém, em alguns casos as cataratas desenvolvem-se novamente e, nessa altura, podem ser removidas através da aplicação laser. 5) Na correcção da miopia, tem sido muito usual a utilização de lasers de excímeros para realizar cortes que permitem corrigir o raio de curvatura da córnea.
Figura 11 – Fotografia da retina a) logo após tratamento contra a retinopatia diabética com fotocoagulação e b) algum tempo depois. Em ambas as imagens são visíveis as lesões causadas pelo tratamento e que evitam o crescimento dos vasos sanguíneos, os quais são responsáveis pela perda de visão nestes doentes. Retirado de: Susanne Amador Kane, Introduction to Physics in Modern Medicine, 2003, Taylor & Francis.

19
Os lasers têm também sido aplicados com sucesso em odontologia. Neste âmbito, contam-se 1) a remoção de tumores e de tecidos em excesso e 2) a remoção de placa bacteriana.
Uma última aplicação que nos parece digna de nota é a de remoção de tumores, não através de cirurgia, mas utilizando-se a técnica de terapia fotodinâmica. Nesta técnica, o indivíduo é injectado com uma substância com afinidade às células cancerosas. Essa substância é formada por moléculas fotosensíveis que, uma vez expostas a luz com determinado c.d.o. sofrem alterações tais que destroem as células a que estão ligadas. Este processo de destruição de células cancerosas é, quanto a nós, muito interessante, sendo necessário ter-se em atenção que o indivíduo deve permanecer às escuras até a substância injectada ter-se fixado na região do tumor, caso contrário, corre-se o risco de serem destruídas células sãs.
2 IMAGENS MÉDICAS
Como é sabido, as técnicas de imagem utilizam equipamentos extremamente sofisticados do ponto de vista técnico e informático, recorrendo a modelos físicos e matemáticos muito complexos. Por este motivo, este é um capítulo importante no âmbito da Física Médica tentando-se discutir uma parte das técnicas de imagem actualmente utilizadas na clínica.
As imagens médicas tornaram-se num meio de diagnóstico muito atraente para os médicos, uma vez que permitem, em geral com grande rigor, obter informação sobre os órgãos internos, sem recorrer à cirurgia. Por este motivo, são considerados pelo meio clínico como métodos não-invasivos, uma vez que dispensam a ablação dos tecidos. Deve, no entanto, estar-se alertado para o facto de algumas das técnicas de imagem envolverem a utilização de radiações que, embora não cortem os tecidos, têm sobre eles efeitos lesivos que devem ser considerados e, tanto quanto possível, minorados. São estes os argumentos utilizados pelos investigadores que evitam classificar algumas técnicas de imagem, nomeadamente as que utilizam radiação ionizante (raios-X e raios ), como técnicas não-invasivas.
Refira-se ainda que, actualmente, os diversos tipos de imagens médicas disponíveis em qualquer hospital permitem obter informação quantitativa tanto quanto à estrutura, como quanto à função dos diversos órgãos. Sendo importante atender aos aspectos clínicos apresentados pelo doente quando se pede ao radiologista um determinado tipo de imagem. É de notar, ainda, que esta área da Medicina é talvez aquela onde mais facilmente se reflecte a necessidade da interdisciplinaridade, sendo comum encontrar num serviço de radiologia, pessoal com diferentes tipos de formação como sejam, médicos, enfermeiros, técnicos, engenheiros electrotécnicos, físicos e informáticos.
2.1 Radiografia e Tomografia Axial Computorizada
Os raios-X não só foram as primeiras radiações a serem utilizadas em equipamento médico, como continuam a ter uma enorme importância ao nível clínico, basta pensar-se no elevado número de radiografias realizadas por ano num hospital!Além disso, uma vez que as primeiras imagens médicas foram obtidas com raios-X, uma parte dos conceitos e técnicas de imagem são directamente importados destes dispositivos.

20
Uma radiografia não é mais do que a imagem (projecção) dos fotões que atravessam o doente e que são gravados num detector. A informação que se obtém é respeitante à atenuação dos raios-X no organismo, ou seja, é esta característica que permite discriminar os diferentes tecidos biológicos. Os fotões detectados podem ser primários, quando não são desviados, ou secundários, quando existe interacção que implique desvio, sendo os primeiros, aqueles que contêm a informação relevante.
Quanto ao objectivo da Tomografia Axial Computorizada (TAC) é o de obter imagens bidimensionais em diferentes ângulos, de modo a conseguir, através de métodos de reconstrução, obter imagens tri-dimensionais. Este procedimento permite, por um lado, melhorar o contraste dos tecidos, por outro, obter informação em profundidade.
Como em qualquer outra substância, a atenuação sofrida por um feixe de raios-X em tecidos biológicos ocorre graças a diversas interacções entre a radiação e os tecidos. De facto, atendendo a que a fracção de energia perdida por um feixe, quando ultrapassa um material, é proporcional à sua espessura dx:
xI
Id
d
então, é válida a expressão:heII 0
onde, é o coeficiente de atenuação do material, h a sua espessura, I0 a intensidade inicial do feixe e I a intensidade do feixe após ter atravessado o material.
É, pois, com base nesta expressão que se obtêm as imagens de raios-X, uma vez que uma radiografia é a projecção dos fotões que atravessam o indivíduo e que são medidos por um detector (ver Figura 12). Ou seja, os tecidos são discriminados mediante o seu coeficiente de atenuação.
Figura 12 – Esquema da formação de uma radiografia (adapt. W.R. Hendee, E.R. Ritenour, 1992).
Quanto à Tomografia Axial Computorizada (TAC) o seu objectivo é construir imagens a três dimensões com base em projecções bi-dimensionais obtidas paradiferentes ângulos. Na Figura 13 é possível observar, a título ilustrativo, o esquema de uma das últimas gerações destes equipamentos.

21
Figura 13 – Esquema de uma TAC de 5ª geração (adapt. W.R. Hendee, E.R. Ritenour, 1992).
Os dispositivos de visualização de imagem são, em geral, monitores de TV que permitem mostrar as imagens em tons de cinzento e que, através de um processamento de imagem adequado, permitem aumentar o contraste das imagens.
2.2 Tomografias de Fotão Único e Tomografia de Emissão de PositrõesAs imagens de Medicina Nuclear são obtidas introduzindo no corpo
determinadas substâncias (radiofármacos), importantes na realização de uma função orgânica específica e marcadas por isótopos radioactivos (radionuclidos). A actividade destes radionuclidos é detectada e, através dela, obtém-se informação sobre a função em estudo. Existem várias diferenças entre estas imagens e as obtidas com raios-X, das quais se destaca o facto de a informação associada às imagens de Medicina Nuclear ser funcional e não estrutural ou anatómica, como as primeiras. Além disso, em termos de reconstrução da imagem, nas de Medicina Nuclear não se tem informação sobre a localização da fonte, conhecendo-se apenas a localização dos detectores.
Relativamente aos tipos de imagem que se obtêm, é possível construir imagens planares (projecção bi-dimensional), imagens dinâmicas (que contêm informação sobre a função do órgão longo do tempo) e tomografia (informação tri-dimensional).
Tal como nas imagens de raios-X, também as imagens de Medicina Nuclear podem ser planares ou tomográficas. Relativamente às primeiras, estas podem ser estáticas ou dinâmicas. Nas imagens estáticas, os órgãos aparecem imóveis e sem alterações da actividade ao longo do tempo (ver Figura 14), nas dinâmicas, constrói-se
22
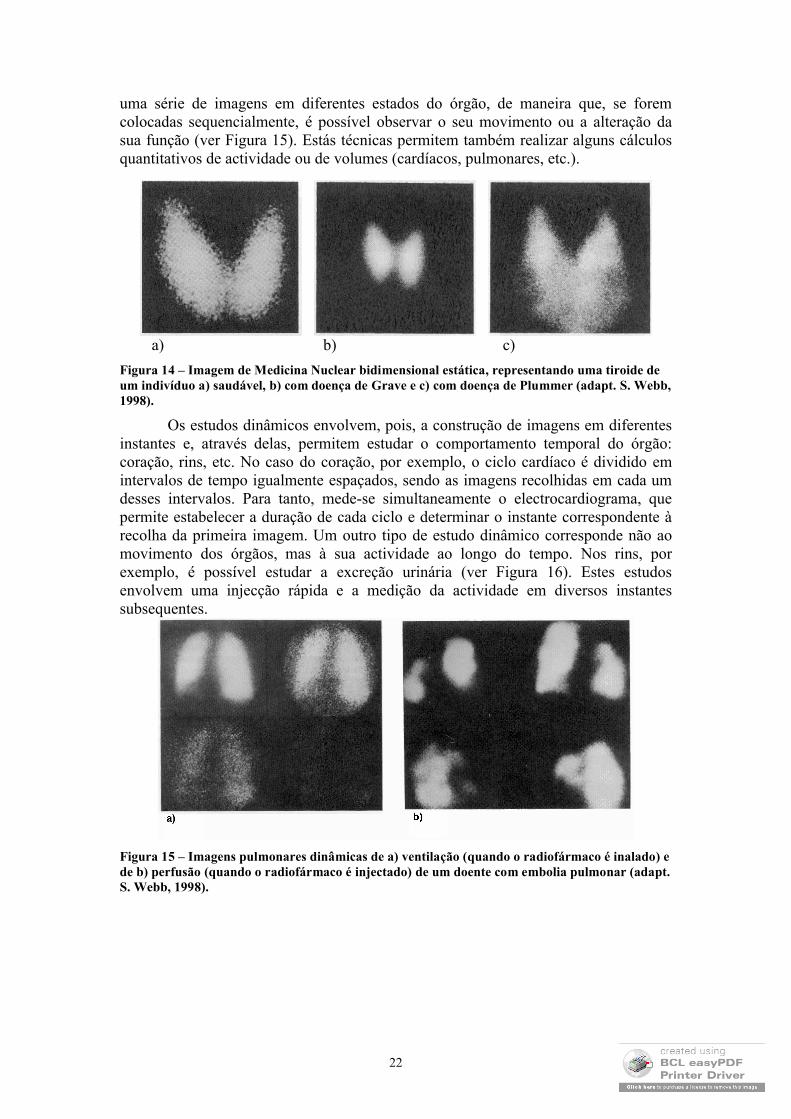
uma série de imagens em diferentes estados do órgão, de maneira que, se forem colocadas sequencialmente, é possível observar o seu movimento ou a alteração da sua função (ver Figura 15). Estás técnicas permitem também realizar alguns cálculos quantitativos de actividade ou de volumes (cardíacos, pulmonares, etc.).
a) b) c)
Figura 14 – Imagem de Medicina Nuclear bidimensional estática, representando uma tiroide de um indivíduo a) saudável, b) com doença de Grave e c) com doença de Plummer (adapt. S. Webb, 1998).
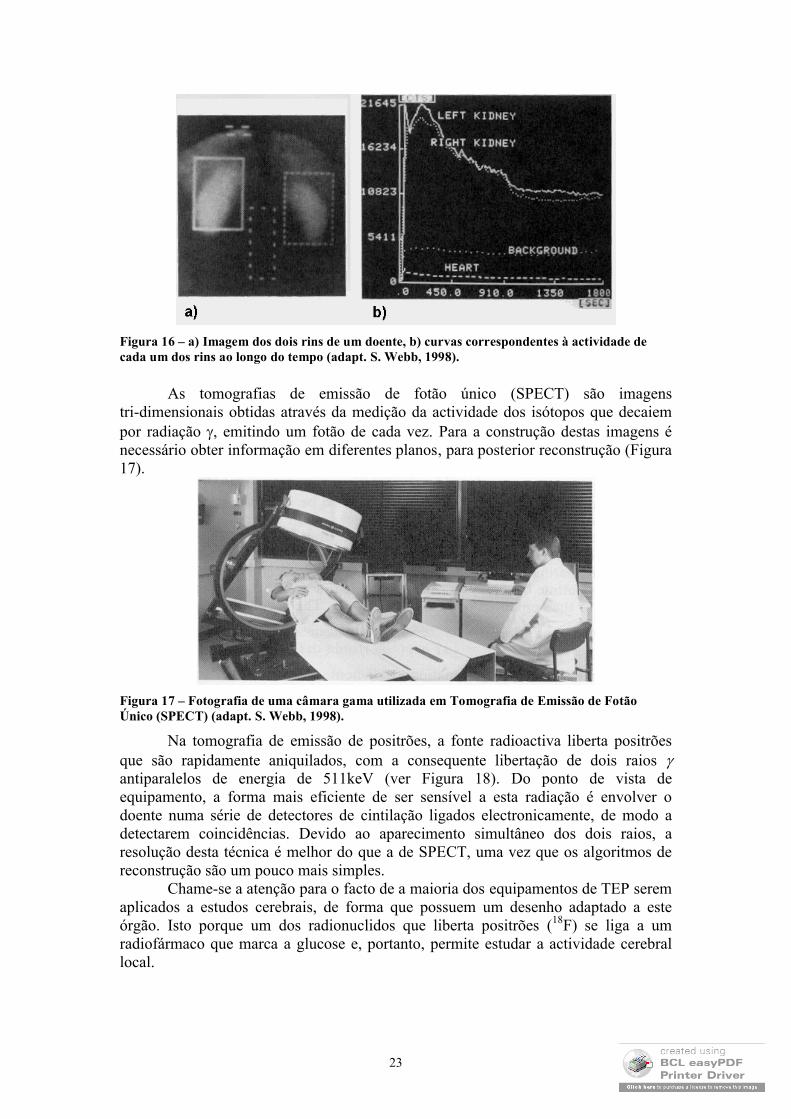
Os estudos dinâmicos envolvem, pois, a construção de imagens em diferentes instantes e, através delas, permitem estudar o comportamento temporal do órgão: coração, rins, etc. No caso do coração, por exemplo, o ciclo cardíaco é dividido em intervalos de tempo igualmente espaçados, sendo as imagens recolhidas em cada um desses intervalos. Para tanto, mede-se simultaneamente o electrocardiograma, que permite estabelecer a duração de cada ciclo e determinar o instante correspondente à recolha da primeira imagem. Um outro tipo de estudo dinâmico corresponde não ao movimento dos órgãos, mas à sua actividade ao longo do tempo. Nos rins, por exemplo, é possível estudar a excreção urinária (ver Figura 16). Estes estudos envolvem uma injecção rápida e a medição da actividade em diversos instantes subsequentes.
Figura 15 – Imagens pulmonares dinâmicas de a) ventilação (quando o radiofármaco é inalado) e de b) perfusão (quando o radiofármaco é injectado) de um doente com embolia pulmonar (adapt. S. Webb, 1998).
23
Figura 16 – a) Imagem dos dois rins de um doente, b) curvas correspondentes à actividade de cada um dos rins ao longo do tempo (adapt. S. Webb, 1998).
As tomografias de emissão de fotão único (SPECT) são imagens tri-dimensionais obtidas através da medição da actividade dos isótopos que decaiem por radiação , emitindo um fotão de cada vez. Para a construção destas imagens é necessário obter informação em diferentes planos, para posterior reconstrução (Figura 17).
Figura 17 – Fotografia de uma câmara gama utilizada em Tomografia de Emissão de Fotão Único (SPECT) (adapt. S. Webb, 1998).
Na tomografia de emissão de positrões, a fonte radioactiva liberta positrões que são rapidamente aniquilados, com a consequente libertação de dois raios antiparalelos de energia de 511keV (ver Figura 18). Do ponto de vista de equipamento, a forma mais eficiente de ser sensível a esta radiação é envolver o doente numa série de detectores de cintilação ligados electronicamente, de modo a detectarem coincidências. Devido ao aparecimento simultâneo dos dois raios, a resolução desta técnica é melhor do que a de SPECT, uma vez que os algoritmos de reconstrução são um pouco mais simples.
Chame-se a atenção para o facto de a maioria dos equipamentos de TEP serem aplicados a estudos cerebrais, de forma que possuem um desenho adaptado a este órgão. Isto porque um dos radionuclidos que liberta positrões (18F) se liga a um radiofármaco que marca a glucose e, portanto, permite estudar a actividade cerebral local.

24
Figura 18 – Esquema da radiação libertada aquando da aniquilação de um positrão com um electrão no ponto P. Os detectores D1 e D2 acusam a chegada simultânea de radiação com energia aproximada de 511 keV, fornecendo essa informação ao equipamento computacional que irá processá-la e reconstruir a imagem (adapt. W.R. Hendee, E.R. Ritenour, 1992).
2.3 Ecografias
Uma vez que os ultra-sons, tal como os raios-X, são radiações que penetram e interagem com os tecidos, a radiação que é transmitida, bem como a radiação que é dispersa, contêm informação acerca das estruturas internas do corpo.
No caso dos ultra-sons, o comportamento a estudar é relativo às reflexões que ocorrem quando os ultra-sons atravessam meios com diferentes impedâncias acústicas. Ou seja, envia-se um pulso de ultra-som para o interior do organismo e mede-se a chegada dos ecos correspondentes à reflexão do mesmo nas paredes dos órgãos.
2.3.1 Ecografia tradicional – efeito de eco, noção de impedância acústicaHistoricamente, a ecografia terá tido início quando, em 1880, foi descoberto o
efeito piezoeléctrico por Pierre e Jacques Curie, que observaram que certos cristais, quando sujeitos a uma pressão, são responsáveis pelo aparecimento de uma diferença de potencial nas suas extremidades, do mesmo modo que, quando sujeitos a uma diferença de potencial, se deformam. Estas propriedades foram desenvolvidas, tendo sido a sua primeira aplicação ao nível das comunicações em alto mar como emissores e receptores de ondas mecânicas de alta frequência, ou seja, de ondas de ultra-som. Nos anos 30 aplicou-se esta tecnologia para fins terapêuticos do tipo oncológico e apenas nos anos 40 se iniciou a aplicação dos ultra-sons ao diagnóstico clínico.
Para compreender o movimento das ondas mecânicas nos fluidos atente-se no seguinte: considera-se que as moléculas que constituem um fluido estão em movimento aleatório contínuo, quando as moléculas são perturbadas por uma força que age sobre elas (por exemplo um pistão em movimento oscilatório), o que se verifica é que são criadas zonas de compressão (aumento da pressão) e zonas de rarefacção (diminuição da pressão), conforme o pistão se movimenta contra as moléculas ou se afasta das mesmas. Estas zonas vão-se propagando ao longo do meio, de modo que se estabelece uma onda longitudinal (a direcção de propagação é paralela ao movimento das moléculas). Quando estas ondas possuem frequências entre 20 e 20 000 Hz são ondas sonoras audíveis pelo ouvido humano, abaixo dessa

25
gama são chamadas infra-sons, enquanto que acima dessa gama são chamados ultra-sons. Os ultra-sons utilizados no diagnóstico têm, habitualmente, frequências entre 1 e 20 MHz.
Nos sólidos a direcção do movimento das moléculas é ligeiramente diferente da direcção de propagação das ondas. No entanto, no caso dos tecidos biológicos, este efeito só é notório nos ossos.
Os ultra-sons, tal como as restantes ondas, são caracterizados por um comprimento de onda (), uma frequência () e uma amplitude. A velocidade de propagação dos ultra-sons é, em praticamente todos os tecidos orgânicos, cerca de 1540 m/s, sendo a sua expressão dada por:
c
Uma vez que os ultra-sons transportam energia através de um meio, é possível caracterizar um feixe de ultra-sons pela energia que transporta por unidade de tempo, ou seja, pela sua potência, ou pela sua intensidade, que não é mais do que a potênciapor unidade de área.
Verifica-se que, nas situações clínicas, a onda reflectida pode ser dez vezes mais pequena do que a onda transmitida e que, para estruturas que se encontram a 10 cm da superfície, a diferença pode ser de um factor de 10 biliões. Por este motivo, as escalas mais apropriadas para lidar com os ultra-sons são logarítmicas, nomeadamente, em acústica, utiliza-se o decibel, definido através da relação:
0
log10I
IdB
Geralmente, esta expressão é utilizada considerando a onda transmitida como a onda de referência, embora não haja muita consonância a este respeito.
A intensidade dos ultra-sons é dependente da pressão máxima no meio (Pmáx) através da relação:
c
PI
2
2máx
Onde é a densidade do meio e c é a velocidade das ondas nesse mesmo meio. De tal forma que a expressão de dB poderá também ser dada por:
0máx
máxlog20P
PdB
onde 0máxP é a pressão máxima do meio correspondente à onda de referência.
No que respeita à velocidade dos ultra-sons em diferentes meios, facilmente se verifica que, enquanto nos gases, a velocidade é relativamente baixa, devido ao facto de uma molécula no meio gasoso poder mover-se em distâncias razoáveis sem interferir com moléculas vizinhas; nos sólidos a velocidade dos ultra-sons é bastante mais elevada, pelo motivo inverso. Nos líquidos, os ultra-sons possuem velocidades intermédias. Ao nível dos tecidos biológicos, exceptuando os pulmões e os ossos, os restantes tecidos comportam-se, a este respeito, como líquidos.

26
Os mecanismos de atenuação dos ultra-sons são, como no caso das radiações: a absorção, a reflexão e a dispersão.
O comportamento dos ultra-sons quando encontram um obstáculo depende do tamanho desse obstáculo quando comparado com o comprimento de onda. Se o obstáculo é maior do que o comprimento de onda, sendo o obstáculo relativamente suave, o feixe mantém as mesmas propriedades, mesmo que mude de direcção. Neste caso, uma parte do feixe é reflectido, outra parte é transmitida através de um feixe de menor intensidade. Esta reflexão tem o nome de reflexão especular. Quando os obstáculos são de tamanho semelhante ou menor do que o comprimento de onda dos ultra-sons, o feixe é disperso. É claro que neste mecanismo também há uma parte do feixe que é desviado de 180º, mas geralmente depois de ter sido desviado por diversas vezes, neste caso será um desvio não especular.
Repare-se que a reflexão especular permite obter informação sobre as interfaces, enquanto que a não especular dá informação sobre a estrutura dos diferentes órgãos.
Também no caso dos ultra-sons (tal como acontecia com os raios-X de baixa energia) a atenuação referente aos ossos é bastante acentuada, pelo que se torna difícil visualizar estruturas (ou tecidos) que se encontrem por detrás dos ossos. Verifica-se que uma boa aproximação do coeficiente de atenuação dos tecidos moles é 0.9, onde é a frequência dos ultra-sons. Pelo que, nestes tecidos, a atenuação dos ultra-sons é tanto mais importante, quanto maior for a frequência dos mesmos.
Para compreender o que se passa ao nível de uma interface, é importante definir a grandeza impedância acústica:
cZ
onde é a densidade do meio e c a velocidade dos ultra-sons nesse meio. Ao considerar um feixe de ultra-sons que incida perpendicularmente numa superfície, a fracção de energia incidente que é reflectida (ou coeficiente de reflexão) é dada por:
2
12
12
ZZ
ZZR
em que Z1 é a impedância acústica do primeiro meio e Z2 é a impedância acústica do segundo. Do mesmo modo, o coeficiente de transmissão (fracção da energia incidente que é transmitida) é definido como:
221
214
ZZ
ZZT
Quanto maior for a diferença entre as impedâncias acústicas, maior será a fracção de energia reflectida e menor a fracção de energia transmitida. Por exemplo, na interface ar/tecido ou ar/água a maior parte da energia é reflectida. Na interface músculo/fígado praticamente toda a energia é transmitida. A construção das imagens médicas baseia-se, então, nos ecos recebidos após a reflexão nas paredes dos órgãos. Para tanto, é necessário garantir que o ângulo de incidência é praticamente perpendicular à interface que se pretende visualizar, pelo menos nos casos em que o emissor é o mesmo que o receptor.

27
Lembre-se, a este respeito, que as leis que governam a refracção são semelhantes tanto nos ultra-sons como na óptica, ou seja:
2
1
2
1
sen
sen
c
c
Donde se retira que a refracção é uma das principais causas de artefactos nas imagens de ultra-sons, uma vez que pode ser responsável pela chegada de ecos ao sensor que não provenham directamente das interfaces.
O principal mecanismo de absorção de ultra-sons é a transformação da energia do feixe em energia térmica. A absorção de ultra-sons nos tecidos é um fenómeno bastante complicado que varia de um modo complexo com a frequência. De facto, o que se verifica é que existem frequências características dos tecidos para as quais a atenuação é muito grande (frequências de relaxação), sendo menores quando nos afastamos dessas frequências.
Tendo em conta o que foi esclarecido anteriormente, as interfaces de meios com impedâncias acústicas muito diferentes, são responsáveis por grandes reflexões nos feixes de ultra-sons. Por este motivo, é imprescindível que se utilize um gel entre o cristal e a pele cuja impedância seja intermédia entre estes dois meios, para, assim, maximizar a energia do feixe que atravessa a pele. Além disso, o gel deve ter uma espessura tão pequena quanto possível para que a atenuação seja mínima.
Os transdutores das imagens ecográficas são cristais piezoeléctricos que transformam energia cinética em energia eléctrica e vice-versa. Em geral, estes transdutores são formados por mais do que um cristal, embora a maior parte das suas características possam ser ilustradas considerando apenas um. Existem muitos cristais que exibem este efeito a temperaturas baixas, no entanto, poucos são os que podem ser aproveitados para diagnóstico por não operarem a temperaturas ambientais.
2.3.2 Formação da imagemExistem, basicamente, três modos de visualização da informação obtida
através dos ecos de ultra-som: o modo A (de amplitude), o modo B (de brilho) e o modo M (de movimento). No modo A informação é visualizada através de sinais num osciloscópio (ver Figura 19) (no eixo dos xx está representado o tempo, ou, admitindo que se conhece a velocidade dos ultra-sons, a distância e, no eixo dos yy a amplitude do eco. Este modo permite conhecer a distância das estruturas que se encontram perpendiculares ao feixe.
Figura 19 – Representação da formação de imagens em modo A (adapt. W.R. Hendee, E.R. Ritenour, 1992).

28
No modo B obtém-se uma imagem propriamente dita, o que envolve um modo de varrimento (ver Figura 20). A informação é visualizada num plano xy, em que as zonas mais brilhantes são aquelas que correspondem a maiores amplitudes de eco (visualizam-se os contornos). Estas imagens podem ser estáticas ou dinâmicas. Nas segundas o movimento pode ser representado em tempo real, sendo particularmente úteis para seguir o funcionamento de órgãos como o coração ou as respectivas válvulas.
Figura 20 – Representação da formação de imagens em modo B (adapt. W.R. Hendee, E.R. Ritenour, 1992).
Por fim, o modo M é utilizado, fundamentalmente, em ecocardiografia e a posição do eco em cada instante é representada do seguinte modo: a profundidade das estruturas que se pretende estudar são representadas verticalmente, enquanto que na horizontal é representado o tempo (ver Figura 21).
Figura 21 – Imagem de ecocardiografia em modo M. No eixo dos xx encontra-se o tempo, enquanto que no eixo dos yy a distância ao detector (adapt. W.R. Hendee, E.R. Ritenour, 1992).
No que respeita ao processamento do sinal, existem, essencialmente, três factores que determinam a amplitude dos ecos: as diferenças de impedâncias acústicas nas interfaces em estudo, a atenuação dos ultra-sons nos tecidos intervenientes e a amplitude do sinal transmitido. O parâmetro ao qual pretendemos ser sensíveis é a diferença de impedâncias acústicas, de maneira que a atenuação nos tecidos funciona como factor indesejável. Para obviar este efeito, utiliza-se um mecanismo de

29
compensação em que os ecos mais tardios e, consequentemente, provenientes de profundidades maiores, são multiplicados por factores de correcção que compensam a atenuação à qual estes foram sujeitos. Refira-se que esta compensação se faz, em geral, on-line, ou seja, vão se fazendo sucessivos ajustes para melhorar a imagem, consoante o órgão que se estiver a observar.
Existem ainda outros tipos de processamento que envolvem a rejeição de sinais demasiado elevados ou demasiado pequenos para serem considerados, filtragens e todo o tipo de análise que permite reforçar as características da imagem como o contraste, a nitidez, etc.
2.3.3 Ecografia de efeito de DopplerNa ecografia de efeito de Doppler são obtidas imagens dinâmicas do fluxo
sanguíneo. Estas imagens têm diferentes aplicações como a monitorização dos batimentos cardíacos fetais, detecção de embolias, monitorização da pressão sanguínea, caracterização do fluxo sanguíneo e localização de oclusões em vasos sanguíneos.
O efeito de Doppler consiste na alteração da frequência de ondas sonoras quando existe uma velocidade relativa entre a fonte e o receptor das ondas. Considere-se que a fonte de ultra-sons está a mover-se na direcção do receptor com uma velocidade vs. Após um intervalo de tempo t depois da criação de uma determinada frente de onda, a distância entre a frente de onda e a fonte é de (c-vs)t, o que significa que o comprimento de onda do ultra-som na direcção do movimento é diminuído para (ver Figura 22):
0v
vc s
onde 0 é a frequência do ultra-som ao sair da fonte. É fácil verificar que a variação na frequência é, então, dada por:
S
S
vc
v0
que toma a forma:
c
vS0
quando se considera a velocidade da fonte muito menor que a velocidade do ultra-som no meio.
Figura 22 – Esquema explicativo do efeito de Doppler (adapt. W.R. Hendee, E.R. Ritenour, 1992).

30
O mesmo efeito se verifica quando é o detector que se encontra em movimento.
Quando a direcção é contrária (em vez de ser no sentido da fonte e do detector se aproximarem, é no sentido de se afastarem) a variação da frequência vem negativa o que significa que esta diminui.
Um último caso a considerar (que é aliás o que mais nos importa neste contexto), é a situação em que os ultra-sons são reflectidos por um objecto em movimento. Nesse caso o objecto funciona como detector quando o feixe o atinge e funciona como fonte quando o feixe é reflectido, de modo que provoca um desvio na frequência que é o dobro dos desvios atrás considerados.
No caso mais geral, em que o feixe não tem a direcção do movimento o desvio causado pelo movimento do objecto reflector móvel é dado por:
cos2 0
c
v
sendo o ângulo formado pela direcção do movimento com a propagação do feixe.Nestas imagens o objecto reflector é a estrutura da qual se pretende conhecer a
velocidade, de modo que é necessário possuir-se equipamento sensível ao desvio de frequência.
No caso de dispositivos de ultra-sons contínuos é necessário existir um sensor com dois cristais, um dos quais emite e o outro recebe, estando ambos a operar ininterruptamente. A informação recebida é proveniente da interferência dos sinais reflectidos por estruturas imóveis e por superfícies em movimento. Deste modo, é necessário retirar o sinal emitido do recebido, de modo que subtracção seja o desvio em frequência que se procura.
A diferença de frequências é amplificada, filtrada e, finalmente, ouvida, uma vez que essa variação abrange a gama de frequências audíveis.
Este tipo de equipamento apresenta uma grande limitação no que respeita à localização da fonte responsável pelo desvio na frequência. Para colmatar este problema, existem os equipamentos de Doppler pulsados, onde um trem de ondas é emitido, passado algum tempo é gravado o eco, e só posteriormente, emitido novo trem de ondas. Deste modo, uma vez que se conhece a velocidade dos ultra-sons nos tecidos biológicos, sabe-se, automaticamente, a distância da fonte responsável por aquele sinal. Por seu lado, este mecanismo envolve um tempo de espera maior e limita a gama de frequências a que se é sensível, uma vez que discretiza o sinal.
Repare-se, finalmente, que nestes últimos equipamentos é possível registar em simultâneo imagens estáticas e dinâmicas. Na maioria das vezes, opta-se por construir uma imagem em tons de cinzento das estruturas imóveis, sobre as quais se projecta, segundo um código de cores, a imagem dinâmica dos reflectores móveis.
2.3.4 Aplicações clínicasSendo a ecografia uma das técnicas de imagem médica mais seguras, ela é
utilizada num grande número de situações, apesar da sua fraca resolução e falta de nitidez.
A ecografia cerebral é apenas utilizada nos primeiros três meses de idade, quando o crânio não está ainda completamente formado e é possível aos ultra-sons penetrarem até ao encéfalo. Nessa fase, é possível detectar algumas massas tumorais, casos de hidroencefalia e hemorragias.
31
São ainda realizadas ecografias à tiróide; à mama, onde se tornou um popular método de rastreio de lesões (em particular quando a mamografia está contra-indicada) e ao útero (com ou sem sonda).
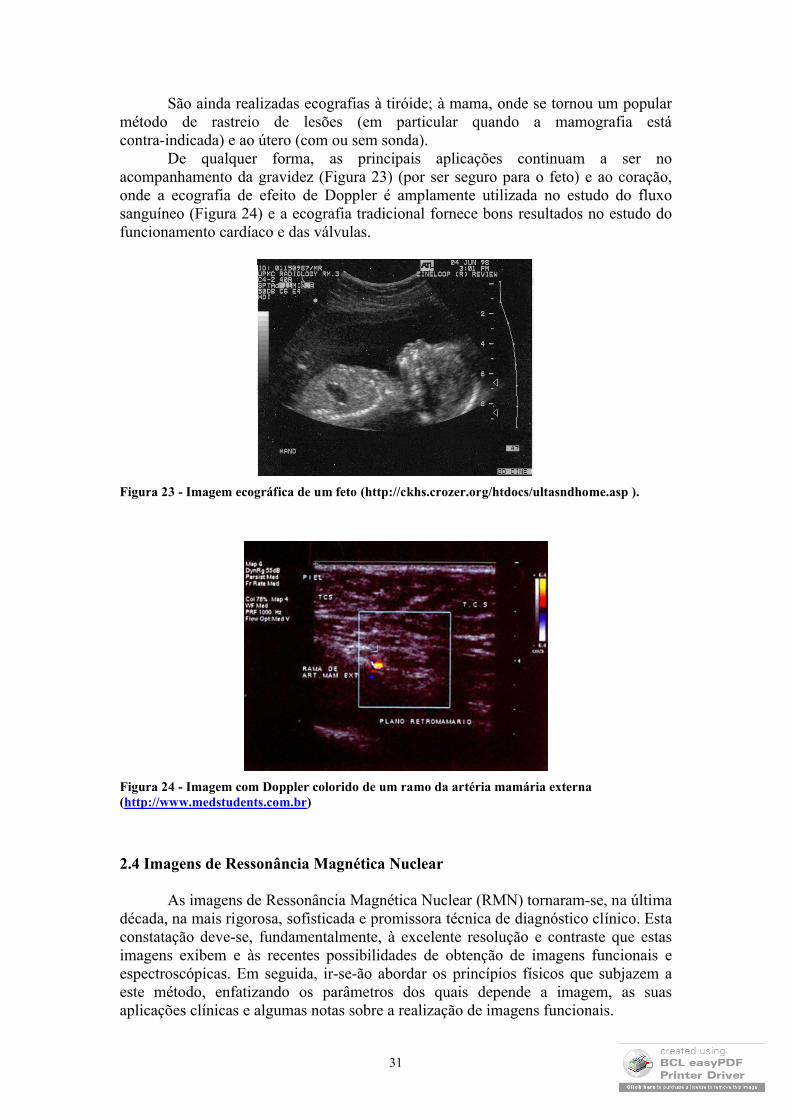
De qualquer forma, as principais aplicações continuam a ser no acompanhamento da gravidez (Figura 23) (por ser seguro para o feto) e ao coração, onde a ecografia de efeito de Doppler é amplamente utilizada no estudo do fluxo sanguíneo (Figura 24) e a ecografia tradicional fornece bons resultados no estudo do funcionamento cardíaco e das válvulas.
Figura 23 - Imagem ecográfica de um feto (http://ckhs.crozer.org/htdocs/ultasndhome.asp ).
Figura 24 - Imagem com Doppler colorido de um ramo da artéria mamária externa (http://www.medstudents.com.br)
2.4 Imagens de Ressonância Magnética Nuclear
As imagens de Ressonância Magnética Nuclear (RMN) tornaram-se, na última década, na mais rigorosa, sofisticada e promissora técnica de diagnóstico clínico. Esta constatação deve-se, fundamentalmente, à excelente resolução e contraste que estas imagens exibem e às recentes possibilidades de obtenção de imagens funcionais e espectroscópicas. Em seguida, ir-se-ão abordar os princípios físicos que subjazem a este método, enfatizando os parâmetros dos quais depende a imagem, as suas aplicações clínicas e algumas notas sobre a realização de imagens funcionais.

32
2.4.1 Princípios físicosOs princípios físicos que permitem a formação de imagens de RMN são
importados da Física Nuclear, nomeadamente, das teorias quânticas que explicam o comportamento dos spins nucleares. No entanto, a abordagem clássica deste tema é, a este nível, suficiente para compreender o mecanismo de obtenção das imagens. De modo que será esta a abordagem aqui explanada.
Magnetização dos tecidos na presença de um campo magnético - De um modo análogo ao efeito de Zeeman nos electrões4, também os núcleos com spin diferente de zero só adquirem valores distintos de energia, se estiverem na presença de um campo magnético. Classicamente, aceita-se que um spin pode ser compreendido como um momento magnético que precessa em torno de um eixo (ver Figura 25).
Figura 25 – Aproximação de um spin a um íman (momento magnético que gira em torno de um eixo). (adapt. R.B. Lufkin, 1990)
Concentremo-nos nos núcleos com spin ±1/2. Quando estes não estão sujeitos a qualquer campo magnético, o eixo ao redor do qual o spin precessa é completamente aleatório, de modo que a magnetização total é nula (ver Figura 26).
Figura 26 – Spins na ausência de campo magnético externo (adapt. R.B. Lufkin, 1990).
No entanto, quando um campo magnético é aplicado, os spins passam a rodar em torno do eixo do campo paralela (estado de energia mais baixa – spin +1/2) ou antiparalelamente (estado de energia mais alta – spin –1/2) (ver Figura 27).
4 Os dois electrões que podem povoar uma orbital atómica possuem a mesma energia quer estejam no estado de spin +1/2, quer estejam no estado de spin –1/2. A sua energia só é diferenciada quando se sujeita o átomo a um campo magnético exterior.

33
Figura 27 – Spins num meio onde se estabeleceu um campo magnético B0. M0 tem o significado de magnetização total do meio (adapt. R.B. Lufkin, 1990).
Devido à diferença de energias dos dois estados, a população no estado de energia mais baixa é mais povoado do que o estado de energia mais alta5. Por este motivo, a magnetização total deixa de ser nula e passa a ter a direcção do campo z, (ver Figura 28), pois os spins, embora façam com o eixo do campo magnético externo um determinado ângulo, não estão em fase, encontrando-se aleatoriamente distribuídos sobre um cone, pelo que a sua componente xy se anula, sobrevivendo, apenas, a componente segundo z.
Figura 28 – Representação de spins a precessarem em torno de um campo magnético externo (Bo) e magnetização total do meio (Mo) (adapt. R.B. Lufkin, 1990).
Tendo em conta o que foi dito, uma das exigências para a utilização desta técnica é que os núcleos em estudo apresentem spin diferente de zero e, de preferência, com spin ±1/2, visto que são os mais simples de interpretar. Na verdade, em imagens médicas, os núcleos utilizados são os de hidrogénio (protões), uma vez que cumprem estas condições e são muito abundantes, o que permite obter um sinal de grande amplitude.
Aplicação de um campo de rádio-frequência: noção de ressonância -Mantendo presente o modelo clássico, é possível definir a frequência à qual os núcleos giram (também chamada frequência de Larmor) como sendo proporcional a um parâmetro característico do núcleo (constante giromagnética - ) e à amplitude do campo magnético externo aplicado aos núcleos (B0):
0Bf
5 Note-se, no entanto, que a diferença é muito pequena (aproximadamente dois spins em cada milhão).

34
Tendo em conta a ordem de grandeza da constante giromagnética dos núcleos e as amplitudes dos campos aplicados (cerca de 1T6) a frequência de Larmor corresponde à gama das rádio-frequências. Deste modo, como facilmente se prevê, se sujeitarmos os núcleos a um campo de rádio-frequências interferiremos com estes, através de um fenómeno de ressonância. Na verdade, ao aplicar-se estes campos actua sobre o comportamento dos spins de duas formas:
Aumenta-se a população de spins anti-paralelos. Coloca-se os spins em fase.Nestas condições, a magnetização total muda de direcção, passando a exibir
componente xy. A amplitude e a duração dos pulsos de rádio-frequência a que os spins são sujeitos, determinam os seus efeitos. São de referir os chamados pulsos de 90º, que são responsáveis por a magnetização passar da direcção z para o plano xy(ver Figura 29).
Figura 29 – Consequências da aplicação de um campo de rádio-frequência na magnetização total. Exemplo de um pulso de 90º (adapt. R.B. Lufkin, 1990).
O sinal medido, nesta técnica, não é mais do que a magnetização transversal (xy), à qual se acede através de uma bobina colocada segundo um dos eixos (x ou y) (ver Figura 30).
Figura 30 – O sinal medido corresponde à componente transversal da magnetização (adapt. R.B. Lufkin, 1990).
6 Actualmente, há já equipamentos a utilizarem campos de 5 T, ou até com valores mais elevados, mas campos magnéticos de 1T continuam a ser os mais utilizados em rotina clínica.
As populações igualam-se.
Os spins ficam em fase.

35
Com base no que até aqui foi exposto, facilmente se compreende que o sinal medido seja proporcional à densidade protónica do meio (é de relembrar que os núcleos utilizados são os de hidrogénio). Concentrando-nos no instante em que o pulso de rádio-frequência é aplicado, o sinal medido será tanto maior quanto maior for o número de spins presentes no meio em estudo (ver Figura 31). Assim, este será, logo à partida, um parâmetro de diferenciação dos tecidos: o sinal será mais ou menos intenso, consoante a maior ou menor concentração de hidrogénio (voltar-se-á a este assunto mais adiante). A este respeito deve ter-se em conta que os tecidos biológicos não diferem significativamente quanto à concentração de átomos de hidrogénio, pelo que, embora este seja um factor a considerar, não é o principal responsável pelo excelente contraste revelado por esta técnica.
Figura 31 – Dependência da magnetização total com o número de protões. Quanto maior for o número de protões, maior será a magnetização total (adapt. R.B. Lufkin, 1990).
Noção de tempo de relaxação spin/spin (T2) e spin/rede (T1) - Retome-se a sequência de acontecimentos: após a aplicação de um campo magnético estático, responsável pelo aparecimento de uma magnetização segundo a sua direcção; aplica-se um campo magnético oscilatório (na gama das rádio-frequências), que roda a magnetização de um determinado ângulo (por exemplo, de 90º), devido ao facto de alterar (igualado, no caso do pulso de 90º) as populações dos spins de +1/2 e de –1/2 e os pôr a rodar em fase. Logo após o pulso de rádio-frequência ter deixado de actuar, a tendência natural do sistema é regressar ao estado inicial. Ou seja, haver reorganização do povoamento dos spins e a desfasagem dos mesmos. Na realidade estes dois processos são independentes e correspondem a diferentes fenómenos de relaxação.
Concentremo-nos no mecanismo de desfasagem dos spins. Repare-se que se a frequência de cada spin fosse exactamente a mesma, estes manter-se-iam em fase. Mas o que se verifica é que as frequências de precessão de cada spin vão ser ligeiramente diferentes. Esta circunstância deve-se, por um lado, ao facto de o campo magnético estático imposto não ser perfeitamente uniforme, apresentando heterogeneidades no espaço; por outro, o próprio meio onde os spins estão inseridos apresenta campos locais que são gerados pela presença de outros spins. Por este motivo, os spins vão-se desfasar, a magnetização no plano xy vai tornando-se menor, o que corresponde a um decaimento no sinal medido (FID – Free Induction Decay) (ver Figura 32 e Figura 33).

36
Figura 32 – Mecanismo de desfasagem dos spins, com consequente decaimento do sinal (adapt. R.B. Lufkin, 1990).
Figura 33 – Esquema do decaimento provocado pela desfasagem dos spins (adapt. R.B. Lufkin, 1990).
Verifica-se que este decaimento do sinal medido é exponencial. E, por conseguinte, é caracterizado por uma constante de tempo. A esta constante de tempo dá-se o nome de T2
* e é definida como o tempo necessário para que o sinal (magnetização transversal – perpendicular ao campo magnético estático) decaia para 37% do seu valor máximo (atingido aquando da aplicação do pulso de rádio-frequência).
Observe-se, uma vez mais, que a grandeza T2* contém informação sobre as
interacções spin/spin (que é a que nos interessa, uma vez que está relacionada com a estrutura do tecido), mas está contaminada com as heterogeneidades do campo magnético estático, cujos efeitos são muito maiores do que os referentes aos campos locais, devido à presença dos spins vizinhos. Numa abordagem mais aprofundada sobre esta técnica referir-se-ia o procedimento que nos permite separar estas duas componentes.
Como já foi anteriormente mencionado, existe ainda um outro mecanismo de relaxação que envolve troca de energia com o exterior, no sentido de repor as populações iniciais respeitantes aos níveis energéticos de spin. Enquanto o pulso de rádio-frequência actua existe excitação de spins que se encontravam no nível de energia mais baixo (paralelo com o campo magnético estático) para o estado de energia mais alto (anti-paralelo). A partir do momento que o pulso cessa as populações tendem a reassumir a situação inicial, ou seja, a magnetização longitudinal (segundo o campo magnético estático) retoma o valor inicial (ver Figura 34). Este mecanismo ocorre através de libertação de energia para o meio e é caracterizado por uma constante de tempo T1, à qual se dá o nome de tempo de relaxação spin/rede. T1
é, analogamente a T2* (ou a T2, quando apenas se considera as interacções spin/spin),
o tempo que demora a magnetização longitudinal a recuperar 63% do seu valor

37
máximo. Como facilmente se compreende, também este parâmetro contém informação sobre os tecidos, uma vez que a maior ou menor facilidade com que os spins libertam energia para o meio, terá necessariamente que estar relacionada com a estrutura do meio onde estes se encontram. É desta forma que T1 é utilizado para obter contraste entre os tecidos.
Figura 34 – Mecanismo de recuperação da magnetização longitudinal, devido à reorganização das populações de spin entre os estados energéticos, com consequente libertação de energia para o meio (adapt. R.B. Lufkin, 1990).
Intensidade do sinal - Nos subcapítulos anteriores foram introduzidos os três parâmetros através dos quais as imagens são construídas. Vejamos, neste ponto, de que maneira é que cada um deles influencia a intensidade do sinal.
Tendo em conta que o sinal medido em imagens de RMN é a magnetização transversal total dos tecidos, seja, então, I a intensidade desse sinal e observe-se que:
1. A intensidade do sinal é tanto mais intensa quanto maior for a densidade protónica (n).
nI α
2. A intensidade do sinal depende de T2 segundo a expressão (ver Figura 35):) f(eI /T-TE 2
3. A intensidade do sinal depende de T1 segundo a expressão (ver Figura 35):)-e f(I /T-TR 11
Figura 35 – Esquema explicativo do comportamento da magnetização devido a T2 e devido a T1
(adapt. R.B. Lufkin, 1990).

38
Uma forma simples de compreender o comportamento da magnetização devido a T1 e a T2 é imaginando os casos limite em que T1 >> T2 e T1 << T2. No primeiro caso o vector magnetização começaria por rodar do plano xy para o eixo z direcção do campo magnético estático e, seguidamente, aumentaria a sua amplitude, enquanto que no segundo, o módulo da magnetização aumentaria e só depois sofreria rotação.
Refira-se, a este propósito, que a gama de valores de T1 e T2 em tecidos biológicos são, respectivamente, [200 ms,2000 ms] e [50 ms, 500 ms].
Imagens com contraste em densidade protónica, T1 e T2. - Retomando o que ficou expresso no subcapítulo anterior, pode dizer-se que o sinal medido em RMN depende da densidade protónica, do tempo de relaxação spin/spin e do tempo de relaxação spin/rede da seguinte forma:
)e (I α α /TT/T-T RE 12 1
Esta expressão permite compreender de que forma é que é possível manipular o contraste das imagens, enfatizando os parâmetros que se pretende. Quem já teve algum contacto com imagens de RMN sabe que existem imagens em densidade protónica, em T1 e em T2.
Imagine-se que existem dois tecidos que possuem tempos de relaxação muito próximos, embora sejam caracterizados por densidades protónicas muito distintas. Nesse caso, o bom senso sugere-nos que se utilizem intervalos de tempo entre dois pulsos de 180º (TE) muito curtos e intervalos entre dois pulsos de 90º (TR) muito longos. Nesta situação, a primeira exponencial tende a saturar e a expressão entre parêntesis também. Pelo que, a forma de separar os tecidos é fundamentalmente através da densidade protónica n.
De igual forma, quando se pretende separar dois tecidos a partir da diferença entre os seus tempos de relaxação T1, deve-se utilizar TE e TR curtos7. Na Figura 36 é possível compreender este procedimento através da análise da intensidade do sinal em função do tempo, quando o tecido é caracterizado por um T1 curto ou longo. Repare-se que para obter um maior contraste entre os tecidos deve-se deixar passar pouco tempo.
Figura 36 – Gráfico da intensidade do sinal em função do tempo para dois tecidos caracterizados por tempos de relaxação spin/rede diferentes (adapt. R.B. Lufkin, 1990).
7 Note-se que o parâmetro n não é susceptível de ser manipulado.

39
Por fim, quando se pretende enfatizar as diferenças em T2, a sequência a utilizar deverá ser caracterizada por um TE e um TR longos. Na Figura 37 estão ilustradas duas curvas de intensidade do sinal em função do tempo, no caso de um tecido com um T2 curto e outro com um T2 longo. Da observação da figura, facilmente se compreende que a maior distinção é feita ao fim de algum tempo.
Figura 37 – Gráfico da intensidade do sinal em função do tempo para dois tecidos caracterizados por tempos de relaxação spin/spin diferentes (adapt. R.B. Lufkin, 1990).
Falta referir o que significam os tons de cinzento nas imagens de RMN. Perante o que ficou expresso nos parágrafos anteriores, é fácil admitir que regiões com uma densidade protónica elevada aparecem mais brancas (intensidade de sinal elevada), como é o caso de tecidos gordos e fluidos. No outro extremo, encontram-se a escuro as áreas com densidade protónica baixa, sendo exemplos calcificações, ar, tecidos fibrosos e osso cortical.
O valor de T1 depende, como já foi descrito, da maior ou menor facilidade que o tecido tem de receber energia na gama das rádio-frequências adequada. Verifica-se que, enquanto a água apresenta um T1 longo, o colestrol, por exemplo, apresenta um T1 curto. Esta observação deve-se, fundamentalmente, ao facto de os movimentos no colestrol serem mais lento e, por isso, mais próximos da frequência de Larmor dos átomos de Hidrogénio. É interessante observar que, em muitas situações, a água que se encontra livre nos tecidos se liga (ainda que por ligações fracas) às fronteiras de muitas moléculas. Em tecidos em que este mecanismo ocorre, o tempo T1 da água tende a diminuir. Na Tabela 2estão apresentados os valores de T1 e de T2 para alguns tecidos orgânicos, considerando uma frequência de 20 MHz. Repare-se que, por exemplo, no sangue, cujo principal constituinte é a água, o tempo de relaxação spin/rede é muito mais baixo do que o da água pura, porque se estabelecem as tais ligações que se referiram anteriormente, entre a água e os restantes constituintes sanguíneos.
Deste modo, para imagens com contraste em T1 aparecem a branco tecidos como a gordura, fluidos com proteínas, moléculas lipídicas, hemorragias subagudas e a melanina. A escuro apresentam-se regiões com neoplasmas, edemas, inflamações, fluidos puros e o líquido céfalo-raquidiano.
T1 (ms) T2 (ms)Sangue 900 200Músculo 500 35Gordura 200 60Água 3000 3000
Tabela 2 – Valores de T1 e de T2 para alguns tecidos biológicos para pulsos de 20 MHz.
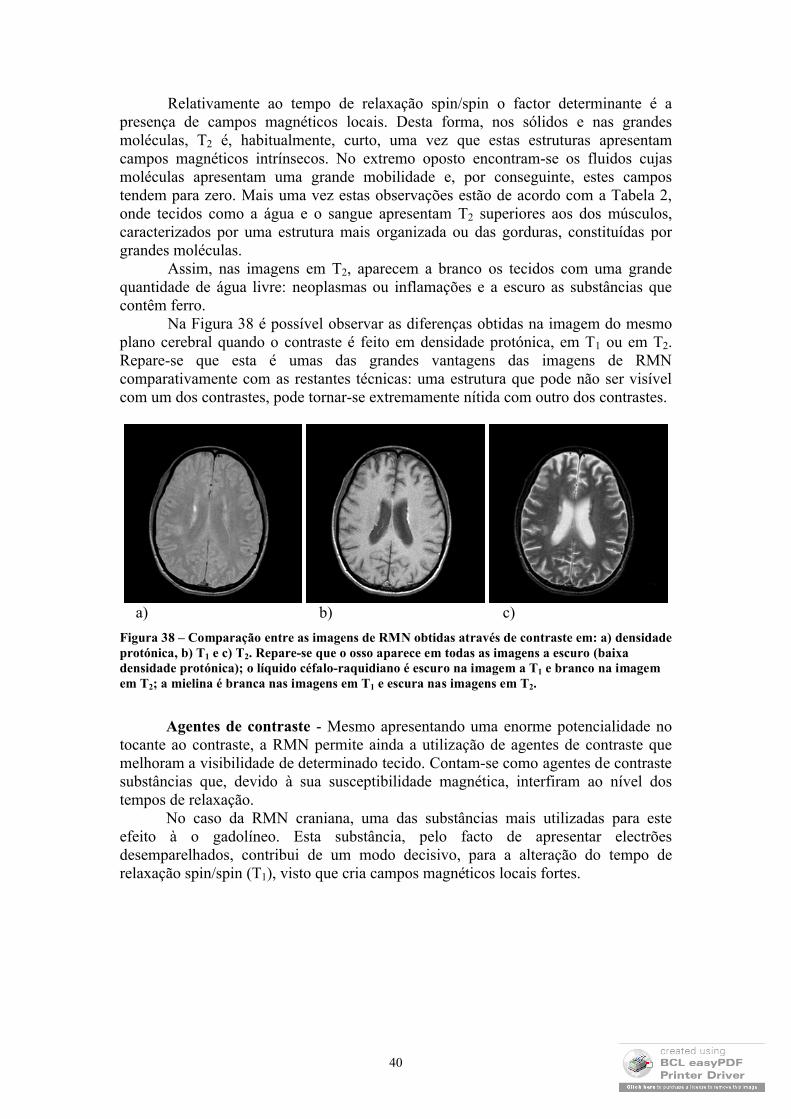
40
Relativamente ao tempo de relaxação spin/spin o factor determinante é a presença de campos magnéticos locais. Desta forma, nos sólidos e nas grandes moléculas, T2 é, habitualmente, curto, uma vez que estas estruturas apresentam campos magnéticos intrínsecos. No extremo oposto encontram-se os fluidos cujas moléculas apresentam uma grande mobilidade e, por conseguinte, estes campos tendem para zero. Mais uma vez estas observações estão de acordo com a Tabela 2, onde tecidos como a água e o sangue apresentam T2 superiores aos dos músculos, caracterizados por uma estrutura mais organizada ou das gorduras, constituídas por grandes moléculas.
Assim, nas imagens em T2, aparecem a branco os tecidos com uma grande quantidade de água livre: neoplasmas ou inflamações e a escuro as substâncias que contêm ferro.
Na Figura 38 é possível observar as diferenças obtidas na imagem do mesmo plano cerebral quando o contraste é feito em densidade protónica, em T1 ou em T2. Repare-se que esta é umas das grandes vantagens das imagens de RMN comparativamente com as restantes técnicas: uma estrutura que pode não ser visível com um dos contrastes, pode tornar-se extremamente nítida com outro dos contrastes.
a) b) c)
Figura 38 – Comparação entre as imagens de RMN obtidas através de contraste em: a) densidade protónica, b) T1 e c) T2. Repare-se que o osso aparece em todas as imagens a escuro (baixa densidade protónica); o líquido céfalo-raquidiano é escuro na imagem a T1 e branco na imagem em T2; a mielina é branca nas imagens em T1 e escura nas imagens em T2.
Agentes de contraste - Mesmo apresentando uma enorme potencialidade no tocante ao contraste, a RMN permite ainda a utilização de agentes de contraste que melhoram a visibilidade de determinado tecido. Contam-se como agentes de contraste substâncias que, devido à sua susceptibilidade magnética, interfiram ao nível dos tempos de relaxação.
No caso da RMN craniana, uma das substâncias mais utilizadas para este efeito à o gadolíneo. Esta substância, pelo facto de apresentar electrões desemparelhados, contribui de um modo decisivo, para a alteração do tempo de relaxação spin/spin (T1), visto que cria campos magnéticos locais fortes.
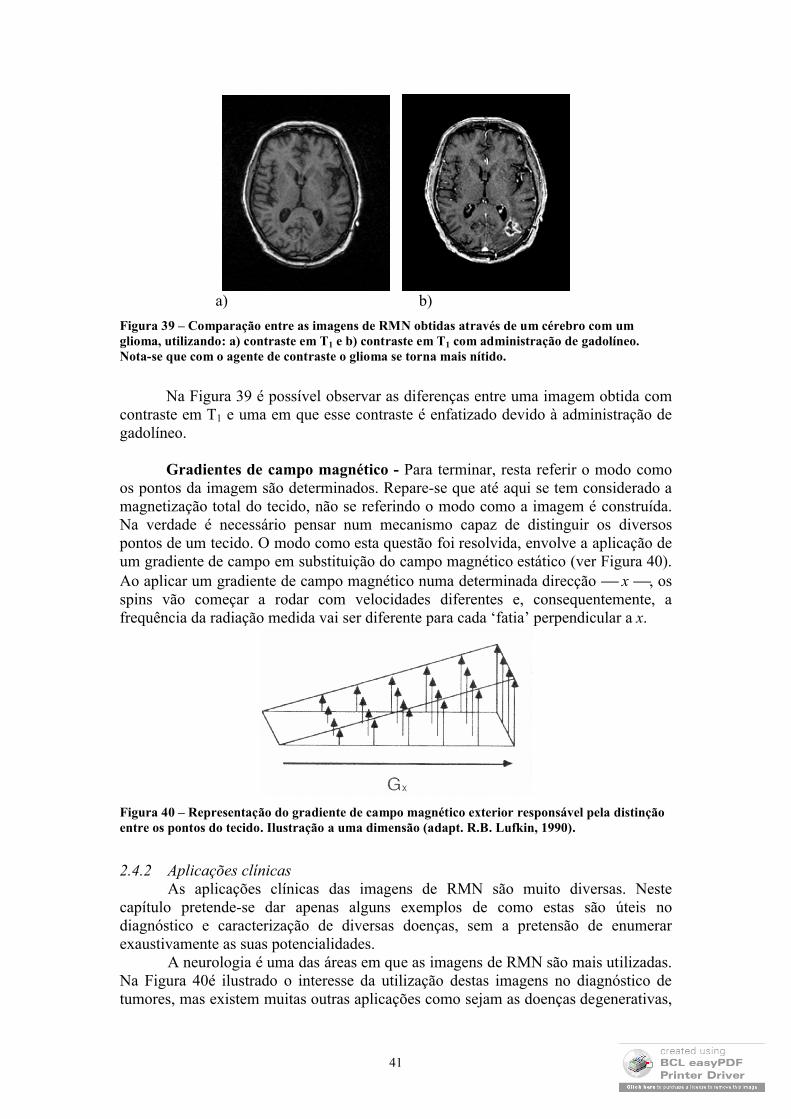
41
a) b)
Figura 39 – Comparação entre as imagens de RMN obtidas através de um cérebro com um glioma, utilizando: a) contraste em T1 e b) contraste em T1 com administração de gadolíneo. Nota-se que com o agente de contraste o glioma se torna mais nítido.
Na Figura 39 é possível observar as diferenças entre uma imagem obtida com contraste em T1 e uma em que esse contraste é enfatizado devido à administração de gadolíneo.
Gradientes de campo magnético - Para terminar, resta referir o modo como os pontos da imagem são determinados. Repare-se que até aqui se tem considerado a magnetização total do tecido, não se referindo o modo como a imagem é construída. Na verdade é necessário pensar num mecanismo capaz de distinguir os diversos pontos de um tecido. O modo como esta questão foi resolvida, envolve a aplicação de um gradiente de campo em substituição do campo magnético estático (ver Figura 40). Ao aplicar um gradiente de campo magnético numa determinada direcção x , os spins vão começar a rodar com velocidades diferentes e, consequentemente, a frequência da radiação medida vai ser diferente para cada ‘fatia’ perpendicular a x.
Figura 40 – Representação do gradiente de campo magnético exterior responsável pela distinção entre os pontos do tecido. Ilustração a uma dimensão (adapt. R.B. Lufkin, 1990).
2.4.2 Aplicações clínicasAs aplicações clínicas das imagens de RMN são muito diversas. Neste
capítulo pretende-se dar apenas alguns exemplos de como estas são úteis no diagnóstico e caracterização de diversas doenças, sem a pretensão de enumerar exaustivamente as suas potencialidades.
A neurologia é uma das áreas em que as imagens de RMN são mais utilizadas. Na Figura 40é ilustrado o interesse da utilização destas imagens no diagnóstico de tumores, mas existem muitas outras aplicações como sejam as doenças degenerativas,
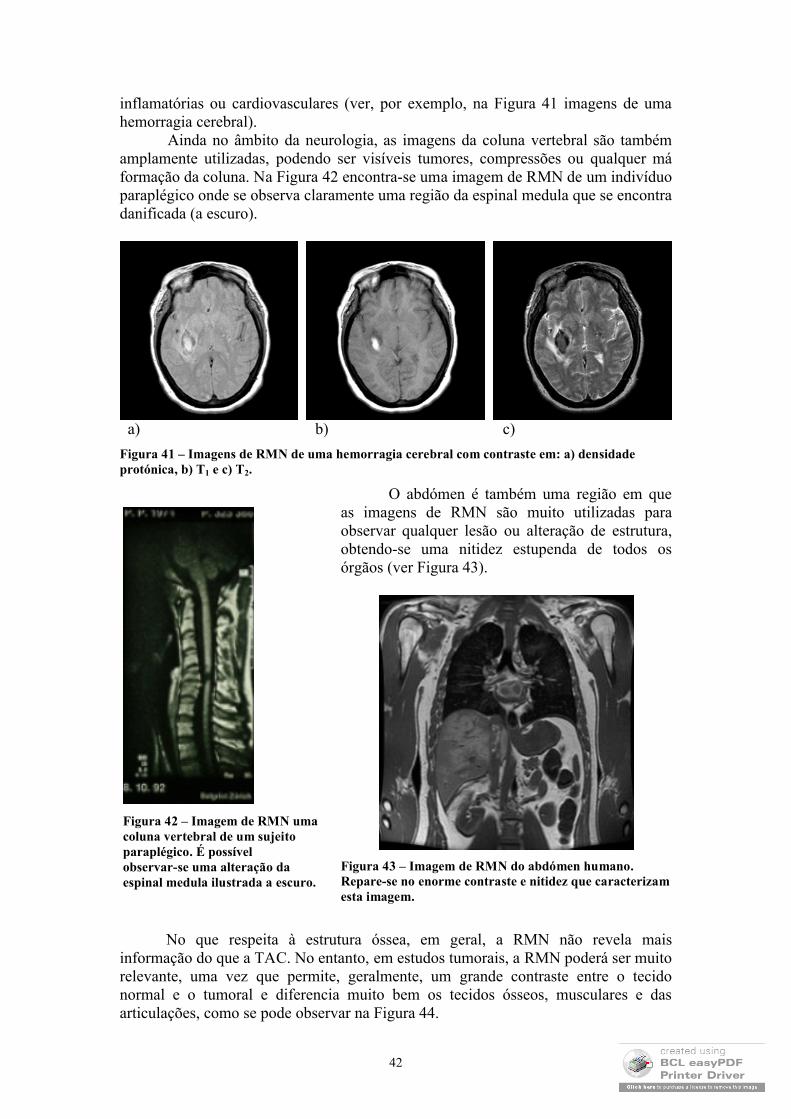
42
Figura 42 – Imagem de RMN uma coluna vertebral de um sujeito paraplégico. É possível observar-se uma alteração da espinal medula ilustrada a escuro.
inflamatórias ou cardiovasculares (ver, por exemplo, na Figura 41 imagens de uma hemorragia cerebral).
Ainda no âmbito da neurologia, as imagens da coluna vertebral são também amplamente utilizadas, podendo ser visíveis tumores, compressões ou qualquer má formação da coluna. Na Figura 42 encontra-se uma imagem de RMN de um indivíduo paraplégico onde se observa claramente uma região da espinal medula que se encontra danificada (a escuro).
a) b) c)
Figura 41 – Imagens de RMN de uma hemorragia cerebral com contraste em: a) densidade protónica, b) T1 e c) T2.
O abdómen é também uma região em que as imagens de RMN são muito utilizadas para observar qualquer lesão ou alteração de estrutura, obtendo-se uma nitidez estupenda de todos os órgãos (ver Figura 43).
Figura 43 – Imagem de RMN do abdómen humano. Repare-se no enorme contraste e nitidez que caracterizam esta imagem.
No que respeita à estrutura óssea, em geral, a RMN não revela mais informação do que a TAC. No entanto, em estudos tumorais, a RMN poderá ser muito relevante, uma vez que permite, geralmente, um grande contraste entre o tecido normal e o tumoral e diferencia muito bem os tecidos ósseos, musculares e das articulações, como se pode observar na Figura 44.

43
Figura 44 – Imagem de RMN dos joelhos de um indivíduo. É, uma vez mais notório um grande contraste entre os diferentes tecidos.
Em resumo, poder-se-á concluir que a RMN permite a obtenção de imagens muito nítidas de praticamente todos os tecidos humanos, sendo de extrema utilidade quando se exige detalhe e nitidez de qualquer órgão. É particularmente utilizada para a detecção de lesões ou alterações das estruturas dos tecidos.
2.4.3 Imagens funcionaisComo já ficou expresso no capítulo anterior a RMN é uma poderosa técnica de
imagem, permitindo uma enorme resolução espacial e um detalhe sobre as diversas estruturas cerebrais não atingido por qualquer outra técnica de imagem médica. No entanto, até há poucos anos atrás, esta técnica permitia apenas aceder a informação anatómica, não sendo capaz de fornecer dados acerca da função do órgão em estudo.
No entanto, o melhoramento das capacidades do equipamento de RMN, tem vindo a permitir estender esta técnica a outros domínios, nomeadamente, a conseguir informação relevante associada ao fluxo sanguíneo, revelando-se uma poderosa ferramenta, principalmente, ao nível do estudo do funcionamento cerebral. Até então, estes estudos funcionais limitavam-se ao processamento de dados electroencefalográficos que, embora tendo uma enorme resolução temporal apresentam uma fraca resolução espacial, e a estudos de imagens de medicina nuclear (como a tomografia de emissão de positrões) que, além de exigir equipamento muito caro, devido aos radionuclidos, e envolver radiação ionizante, atinge resoluções apenas de 0.5 a 1.0 cm. Esta moderna técnica de imagem que em seguida se descreve é caracterizada por ser não invasiva, possuir uma resolução espacial de poucos milímetros e uma resolução temporal que pode ser menor do que um milisegundo, permitindo aceder aos níveis superiores do funcionamento cerebral.
Princípios físicos - Se recordarmos que desde 1936 se sabe que a susceptibilidade magnética da oxihemoglobina e da desoxihemoglobina é diferente, e que o parâmetro T2 da ressonância magnética nuclear é sensível às heterogeneidades do meio, facilmente se compreende que, através da medição de T2, seja possível obter alguma informação acerca dos diferentes estados da hemoglobina. Na verdade, começou-se por observar que a taxa de decaimento, T2, era menor quando estávamos na presença de desoxihemoglobina, quando comparada com a taxa associada à oxihemoglobina. Ou seja, observou-se, em 1990, que os vasos sanguíneos apareciam mais brilhantes (o que pode corresponder a um aumento de T2) quando o sangue era mais oxigenado. Este efeito foi interpretado como correspondendo a um aumento das

44
heterogeneidades locais do campo correspondente ao estado desoxigenado da hemoglobina e ficou conhecido pelas iniciais BOLD, tendo estas o significado de Blood Oxygenation Level Dependent. Um ano mais tarde, era possível seguir este sinal no tempo, observando-se as mudanças operadas quando um animal respirava uma atmosfera pobre em oxigénio.
Esta observação tem permitido obter informação sobre o funcionamento cerebral uma vez que, quando uma determinada área cerebral é activada, verifica-se um aumento de fluxo sanguíneo nesse zona, esse aumento é maior do que o aumento do consumo de oxigénio, de tal forma que, no sangue venoso, passa a existir mais sangue oxigenado do que anteriormente, o que aumenta o sinal de RMN. Ou seja, este é um mecanismo em que o próprio sangue funciona como agente de contraste endógeno.
Características do Sinal - A percentagem de sinal de RMN que corresponde ao sangue é de aproximadamente 6% na matéria cinzenta e ainda menos na matéria branca. Se pensarmos que as diferenças ocorridas no fluxo sanguíneo durante a activação cerebral são entre 2-5% para campos de cerca de 1,5 T e até cerca de 15% em campos de 4 T, verifica-se que as diferenças no sinal às quais pretendemos ser sensíveis, são, de facto, muito pequenas. De modo que a forma de obter imagens passa por estabelecer diferenças adequadas entre duas situações: uma que funciona como controlo e a outra como paradigma, que é, em geral, uma situação de estímulo que visa determinada função8.
Observa-se que entre o estímulo e a resposta máxima em termos de imagem existe um certo tempo de atraso de cerca de 4 s, o que limita a possibilidade de seguir o processamento de informação ao nível neuronal ou de resposta psico-física, como se consegue em EEG ou com técnicas de microeléctrodos. Observe-se que este tempo de atraso é dependente da área do encéfalo que está a ser estimulada.
Aplicações - Os primeiros resultados obtidos com esta técnica terão sido, obviamente, os referentes a respostas a estímulos visuais, motores e auditivos. Numa primeira abordagem a resposta ao estímulo parece variar linearmente com a intensidade do estímulo, embora ainda possa haver algum trabalho para estipular a gama de comportamento linear e o limiar de sensibilidade.
As aplicações mais sofisticadas desta técnica são as que envolvem funções mais complexas como as que estão relacionadas com a linguagem ou a idealização de imagens e de movimentos
A título ilustrativo apresenta-se na Figura 45 um estudo realizado na Universidade de Cambridge, que pretende compreender alguns mecanismos subjacentes à memória semântica dos indivíduos.
8 Estratagema também utilizado nas imagens de Medicina Nuclear.

45
Figura 45 – As áreas assinaladas a roxo são as activas quando se estimula o indivíduo da seguinte forma: pede-se ao sujeito para observar uma sequência de três objectos (insectos, por exemplo) e, por fim, mostra-se um quarto objecto e o sujeito deve decidir se este pertence ou não à mesma família de objectos (cortesia de P. Tavares, P. Fletcher e L.K. Tyler).
3 ELECTROENCEFALOGRAFIA
Este capítulo tem um duplo objectivo: por um lado introduzir algumas notas sobre a fisiologia do sistema nervoso central, por outro, discutir algumas questões relacionadas com os equipamentos de electroencefalografia, os quais permitem registar, em tempo real, a expressão da actividade cerebral, medida ao nível do escalpe.
3.1 Aspectos da electricidade do sistema nervoso
Sendo a electricidade um fenómeno extremamente eficiente quer no transporte de informação, quer na sua sincronização, não é de admirar que os sistemas biológicos a utilizem nas mais sofisticadas e delicadas funções do corpo humano. Na verdade, qualquer que seja a situação em que exista transporte iónico, os fenómenos eléctricos marcam uma indelével presença, revelando-se de particular interesse no processamento dos sinais nervosos e na actividade muscular. Neste sub-capítulo serão, pois, abordados os mecanismos associados à electricidade do sistema nervoso nas suas diversas vertentes: criação, manutenção e transporte de informação quer ao nível celular, quer num âmbito mais geral, relacionado com a organização cerebral.
3.1.1 As células gliaisAlgumas das funções mais interessantes das células (nomeadamente das
células cerebrais) estão associadas às propriedades das suas membranas e à forma como elas determinam a diferença de potencial que se estabelece entre o interior e o exterior das células. Quando as concentrações iónicas são diferentes no interior das células relativamente ao exterior, há tendência, como se sabe, para os iões fluírem no sentido das mais altas concentrações para as mais baixas. Porém, ao saírem da célula, os iões, uma vez que são partículas carregadas, provocam diferenças de potencial que se opõem à saída e/ou entrada de mais iões. Há, pois, uma diferença de potencial, a

46
partir da qual deixa de haver fluxo iónico9, uma vez que a tendência provocada pelo gradiente de concentrações é, nessa circunstância, totalmente contrabalançada pelo gradiente de potencial que se estabelece. Ora a equação que governa a dependência do potencial eléctrico com as concentrações iónicas no interior e no exterior de uma célula, no estado de equilíbrio, é a equação de Goldman que, para iões monovalentes, toma a forma:
oi
io
+
+ln
F
RT=
JPKP
JPKPV
JJ
KK
JJ
KK
,
onde: R - constante dos gases raros (8.3144 J mol-1 K-1);T - temperatura (em kelvin);F - constante de Faraday (9.6487 x 104 C mol-1);K - percorre todos os iões positivos envolvidos no processo;J - percorre todos os iões negativos envolvidos no processo;Pn - permeabilidade da membrana ao ião n;no - concentração do ião n no exterior da célula, no equilíbrio;ni - concentração do ião n no interior da célula, no equilíbrio.
Em relação às células neurogliais10, por exemplo, verifica-se que a permeabilidade da membrana ao potássio é muito superior à de outro qualquer ião e, portanto, a equação anterior reduz-se à conhecida equação de Nernst, aplicada ao potássio (K+):
i+
o+
K
Kln
F
RT=V .
Substituindo as variáveis pelos seus valores aproximados, ou seja: R = 8.3143 J.K-1.mol-1
T = 310.15 KF = 9.6487 x 104 C.mol-1
K+o = 3 x 10-3 MK+i = 0.09 M
obtém-se: V = -90.9 mV.
Este valor é totalmente comprovado pelas medidas experimentais, que apontam para a existência de um potencial de cerca de -90mV no interior das células gliais. Refira-se, ainda a este respeito, que a dependência do potencial com as concentrações de potássio segue de perto o comportamento sugerido pela equação
9 Na verdade, o fluxo iónico continua a existir, mas as partículas que entram são totalmente contrabalançadas com as que saem, gerando-se, desta forma, um equilíbrio dinâmico.10 As células neurogliais são, habitualmente, de pequena dimensão e circundam os corpos celulares e os axónios das células nervosas. A elas se atribuem diversas funções tais como: 1) conferir firmeza aos tecidos cerebrais, analogamente ao tecido conjuntivo de outras regiões do corpo, isolando, por vezes, grupos neuronais; 2) remover os detritos resultantes da morte celular; 3) formar a mielina que envolve alguns axónios; 4) remover os neurotransmissores químicos, após estes terem sido libertados pelos neurónios; 5) permitir armazenar K+, de modo a manter estável a concentração extracelular deste ião; 6) conduzir os neurónios para as regiões correctas durante o seu desenvolvimento e guiar o crescimento dos axónios; 7) participar nas funções correspondentes à barreira hemato-encefálica; 8) ter funções nutrientes.

47
Nernst, de modo que se admite que o factor determinante para o aparecimento do potencial das células gliais é o transporte passivo de iões K+ através de canais selectivos a este ião e presentes na membrana celular.
Estas considerações conduzem-nos ao facto de as células neurogliais se comportarem como reguladoras da concentração de K+ no exterior da célula. Como se observará adiante, a alteração da concentração de K+ no espaço extracelular é um dos factores modeladores do funcionamento neuronal, de modo que um desequilíbrio na função das células neurogliais ao nível da sua função homeostática relativamente às concentrações de K+, pode desencadear um anormal processamento de informação por parte dos neurónios.
3.1.2 As células nervosas ou neuróniosOs neurónios são as células
responsáveis por todo o tratamento da informação envolvida nos processos cerebrais. Tipicamente, um neurónio é constituído por quatro regiões diferenciadas: as dendrites, o corpo celular ou soma, o axónio e os seus terminais pré-sinápticos, correspondendo cada um deles, respectivamente, à entrada, integração, condução e transmissão da informação (ver Figura 46). O corpo celular é considerado o centro metabólico e integrador da célula, nele se encontra o núcleo, o retículo endoplasmático e o sistema de Golgi. Às dendrites ou árvore dendrítica fluem numerosos terminais de outros neurónios, sendo este elemento considerado como a região através da qual, tipicamente, entra a informação. O axónio encontra-se, de uma forma geral, do lado oposto à maioria das dendrites e é responsável pela condução da informação até outros neurónios ou até aos músculos. Refira-se que alguns axónios são revestidos por uma camada de mielina(formada por determinadas células gliais as células de Schwann) que é diversas vezes interrompida em regiões a que se dá o nome de nós de Ranvier. Esta disposição do envolvimento isolante, permite que a informação seja mais rapidamente conduzida. Quanto aos terminais do axónio, são estes que estabelecem a comunicação entre dois neurónios, através do contacto directo entre as membranas de ambos sinapse eléctrica ou, mais comummente, mediada por neurotransmissores sinapse química.
Figura 46 - Esquema das diferentes estruturas de um neurónio: as dendrites; o corpo celular com o núcleo e o citoplasma; o axónio com a blindagem de mielina e os nós de Ranvier e os terminais sinápticos. (Adapt. Kandel et al, 1995).

48
3.1.3 O potencial de repousoA característica mais determinante das células nervosas é a sua excitabilidade,
a qual está intimamente relacionada com as propriedades do seu estado de repouso. Os neurónios encontram-se, no estado de equilíbrio, a cerca de -70mV relativamente ao exterior (note-se que este é um valor médio que depende fortemente do tipo de neurónios que se considere), e quando este valor sofre uma alteração de duas ou três dezenas de mV no sentido positivo, este desequilíbrio acentua-se e o neurónio passa a um estado excitado.
O estado de repouso neuronal resulta da interacção de diversos factores: a permeabilidade da membrana aos iões presentes no espaço intra e extracelular, as concentrações desses iões, o transporte activo através da membrana e a diferença de potencial entre o interior e o exterior da mesma.
Tal como já se referiu em relação às células neurogliais, também o potencial de repouso dos neurónios é regido, em primeira aproximação, pela equação de Goldman. Como exemplo ilustrativo pode tomar-se para as diversas concentrações dos iões envolvidos as encontradas no axónio gigante da lula, as quais, apesar de serem tipicamente 3 ou 4 vezes superiores às encontradas nos neurónios dos mamíferos são, em termos relativos, idênticas às destes. Assim, as concentrações iónicas tomam os valores: K+o=20mM; K+i=400mM; Na+o=440mM; Na+i=50mM; Cl-o=560mM; Cl-i=52mM, sendo as permeabilidades relativas: PK
+=1; PNa+=0.04; PCl-=0.45. Donde resulta, para o potencial de repouso, à
temperatura de 25oC: V= -60.9mV. Este potencial negativo está relacionado com o facto de a permeabilidade de membrana para o potássio ser muito maior do que para o sódio (numa relação de 25 para 1). Assim, e uma vez que o gradiente de concentrações do ião K+ é no sentido da saída deste do interior para o exterior, cria-se um potencial negativo que não é compensado com o fluxo de iões Na+ para o interior, visto que a permeabilidade da membrana é, para este ião, muito pequena.
É de referir que o potencial de repouso, que corresponde ao fluxo passivo de iões através de canais de membrana selectivos, sendo, portanto, determinado pelos gradientes de concentração e pela permeabilidade relativa da membrana aos diferentes iões, não é o de equilíbrio para o K+ ou para o Na+ isoladamente, o que implica que cada um destes iões tenda a fluir continuamente por transporte passivo (o K+ de dentro para fora o seu potencial de equilíbrio é de aproximadamente -77mV; o Na+
de fora para dentro o seu potencial de equilíbrio é de aproximadamente 56mV). Quanto ao Cl-, tendo um potencial de equilíbrio de cerca de -61mV, não tende a exibir, nestas condições, fluxo efectivo. Quando, no entanto, o potencial de repouso da membrana apresenta valores inferiores/superiores, o ião Cl- tende a sair/entrar na célula levando a um reajuste das suas concentrações. Na realidade, verifica-se que existem fluxos iónicos correspondentes aos iões K+ e Na+ segundo as direcções esperadas, mas que são compensados, como se mencionará adiante, através de transporte activo isto é, com gasto energético. Quanto ao ião Cl- apenas algumas células nervosas apresentam transporte activo deste ião e, neste caso, será também, completamente contrabalançado pelo transporte passivo através de canais membranares.

49
Figura 47 - Esquema de alguns canais iónicos existentes numa membrana neuronal: os canais de Na+ e de K+ responsáveis pelo transporte passivo destes iões (refira-se que a permeabilidade relativa da membrana a estes dois iões é de 0.04 para 1); os canais de Na+ e de K+ dependentes do potencial e que são responsáveis pela excitabilidade da membrana e a bomba de Na+/K+ que mantém as concentrações destes dois iões. (Adapt. Le Cerveau, 1984).
Um transporte activo extremamente importante ao nível neuronal é o associado à bomba de sódio/potássio. Esta bomba é uma proteína que, através da hidrólise de uma molécula de adenosina trifosfato (ATP), transporta três iões Na+ para o exterior do neurónio e dois de K+ para o seu interior, contrariando o seu fluxo passivo (ver Figura 47). Esta bomba, para além de manter as concentrações iónicas do Na+ e do K+ nos níveis necessários para manter o potencial de membrana nos valores anteriormente calculados, aumenta-o em cerca de 10%, devido ao facto de ser electrogénica (isto é, por cada dois iões positivos que entram na célula, saem três iões do mesmo sinal, gerando-se, assim, uma diferença de potencial negativa no interior relativamente ao exterior).
3.1.3 O potencial de acçãoQuando a célula, habitualmente polarizada com valores de cerca de -70mV,
sofre uma despolarização de duas ou três dezenas de mV, esta é acentuada, através de mecanismos de realimentação positiva, atingindo aproximadamente 40mV. Em seguida, tem lugar uma repolarização e, após uma breve hiperpolarização (que pode atingir cerca de -90mV), o seu valor de equilíbrio é retomado (Figura 48). A esta descrição corresponde o potencial de acção que dura, tipicamente, cerca de 2ms, podendo variar entre 1 e 10ms e é o responsável pela transmissão de informação aolongo do axónio.
Figura 48 - Esquema do potencial de acção. Quando o potencial da membrana atinge cerca de -55mV abrem-se os canais de Na+ dependentes da tensão, responsáveis pela despolarização. Ao fim de algumas décimas de ms são abertos os canais de K+ e fechados os de Na+ de modo a repolarizar a membrana. O fecho tardio dos canais de K+ implica um período final de hiperpolarização. (Adap. Tortora e Grabowski, 1996).

50
A criação deste potencial deve-se à existência, na membrana neuronal, de canais de Na+ e de K+ dependentes do potencial. Ou seja, os canais abrem quando se verifica uma despolarização na membrana. Estes canais aumentam a permeabilidade da membrana aos referidos iões de modo que, em conformidade com o que foi exposto anteriormente, passa a existir maior fluxo de iões Na+ para dentro da célula e de iões K+ para fora. Quanto à morfologia do sinal, esta é determinada pelas respostas temporais de ambos os tipos de canais. Assim, como os canais de sódio abrem mais rapidamente de que os de K+, o potencial de membrana aumenta abruptamente, devido à entrada de iões Na+ para o interior do neurónio. Durante essa subida, abrem os canais de K+ que fluem em sentido contrário e, portanto, se opõem à subida do potencial. Este facto, conjuntamente com a circunstância de os canais de Na+ também se fecharem rapidamente, é o responsável pela repolarização da membrana. Como os canais de K+ são lentos a fechar, verifica-se a hiperpolarização já mencionada.
O funcionamento dos canais explica ainda, a necessidade de atingir um determinado patamar de despolarização para o aparecimento do potencial de acção e a existência de um período refractário, durante o qual não é possível o surgimento de novo potencial de acção na mesma porção de membrana. Observa-se, pois, que, quando a despolarização não atinge um determinado valor, o potencial de acção não é desencadeado. Esta circunstância verifica-se porque o aumento de permeabilidade ao Na+ suscitado por uma pequena despolarização é totalmente compensado pelos iões K+ que, mesmo no estado de repouso, tendem a fluir para o exterior do neurónio. Quanto ao período refractário durante o qual não é possível a criação de um novo potencial de acção, existem essencialmente dois factores que o determinam: a manutenção de canais de K+ abertos para além da reposição do potencial de repouso e a existência de um estado de inactivação dos canais de Na+. O primeiro está relacionado com o facto de, durante o período de hiperpolarização ser necessária uma maior despolarização para alcançar o patamar correspondente ao despoletar do potencial de acção. O segundo, com a impossibilidade de reactivação dos canais de Na+ nos instantes posteriores ao seu fecho.
Refira-se também que os potenciais de acção se propagam ao longo do axónio, transmitindo a informação de um dos seus extremos para o outro. Esta condução é feita do seguinte modo: a criação de um potencial de acção numa determinada região do axónio aumenta o potencial de membrana em redor dessa região, ora quando essepotencial atinge o patamar anteriormente referido, novo potencial de acção é criado e assim sucessivamente em relação às regiões adjacentes. Há, porém, uma questão que se deve ressalvar: a unidireccionalidade dessa propagação. De facto, se fosse possível gerar um potencial de acção a meio de um axónio, este propagar-se-ia em ambos os sentidos. No entanto, os potenciais de acção surgem habitualmente no início do axónio, uma vez que é ao nível do soma que ocorre a integração da informação que aflui ao neurónio e é aí que se determina o aparecimento ou não do potencial de acção. Assim, verifica-se que este se propaga apenas num sentido, graças ao período refractário a que se aludiu anteriormente.
3.1.4 O papel da mielina na propagação dos potenciais de acçãoResta enfatizar o papel da mielina na condução do sinal. As células neurogliais
que envolvem alguns neurónios aumentam drasticamente a resistência eléctrica destes, de modo que, praticamente, só é possível a criação de potenciais de acção nas regiões onde esta blindagem se interrompe nos nós de Ranvier, distanciados entre si cerca de 1 ou 2 mm. Supondo que num desses nós se gerou um potencial de acção, o aumento do potencial de membrana propaga-se através do citoplasma até ao nó de

51
Ranvier mais próximo, onde se irá formar novo potencial de acção. Este sistema de condução tem como principal objectivo o aumento da velocidade de propagação dos sinais que pode ser, nos casos mais eficazes, cerca de 100 vezes maior. Além da rapidez de propagação, este sistema tem como vantagem adicional a possibilidade de aumentar a frequência dos potenciais de acção por períodos de tempo mais prolongados, sem saturar os tecidos, uma vez que as trocas iónicas são muito menores.
3.1.5 As sinapsesAs sinapses são, como já se referiu, as regiões de contacto entre dois
neurónios. Dividem-se em sinapses eléctricas e químicas. As primeiras são pouco frequentes e nelas a célula pré-sináptica está fisicamente ligada à pós-sináptica. O seu funcionamento limita-se ao contacto entre os citoplasmas das duas células através de canais de pequena resistência, de modo que o potencial de acção, ao chegar ao terminal da célula pré-sináptica, se replica na célula pós-sináptica (ver Figura 49). Estas sinapses não apresentam características modeladoras tão versáteis como as sinapses químicas, no entanto, outras vantagens lhes são inerentes, tais como a ausência de atraso na transmissão do sinal de uma célula para a outra e a facilidade no aparecimento de sincronia num grupo de células onde esta possa, eventualmente, ser desejável.
Figura 49 - Esquema de uma sinapse eléctrica. O contacto entre a célula pré-sináptica e a pós-sináptica é feito através de canais que permitem uma transmissão rápida do sinal e facilitam uma possível sincronia entre as células. (Adapt. Kandel et al, 1995).
No caso das sinapses químicas, a transmissão de informação é modelada por substâncias libertadas pela célula pré-sináptica os neurotransmissores. O mecanismo é o seguinte: os sinais atingem o terminal do axónio, abrem canais de iões cálcio, cuja entrada para o interior da célula desencadeia a libertação de neurotransmissores para o espaço entre os dois neurónios (fenda sináptica); na célula pós-sináptica encontram-se receptores sensíveis a estes neurotransmissores químicos, de modo que, quando detectam a presença destas substâncias, induzem fluxos iónicos que alteram a polarização da membrana (ver Figura 50). Este processo permite uma versatilidade muito grande, uma vez que a modificação da polarização tanto pode ser no sentido da despolarização como no da hiperpolarização. Ou seja, dependendo dos canais iónicos que são abertos, assim a chegada de sinais ao neurónio pré-sináptico pode suscitar o aparecimento de potenciais de acção no neurónio pós-sináptico ou inibi-lo. Deste modo, relativamente às sinapses químicas, consideram-se sinapses excitatórias ou inibitórias consoante o sentido da polarização que provocam na célula pós-sináptica.

52
Figura 50 - Esquema de uma sinapse química. Quando o potencial de acção chega ao terminal nervoso da célula pré-sináptica desencadeia a entrada de iões Ca++ que vão, por sua vez, motivar a libertação de neurotransmissores contidos em vesículas que se fundem à membrana pré-sináptica. Os neurotransmissores irão ser reconhecidos por receptores existentes na célula pós-sináptica que irão desencadear processos responsáveis pela alteração do estado de polarização do neurónio pós-sináptico. (Adapt. Beatty, 1995).
Um conceito que emerge da discussão anterior é o de potencial pós-sináptico: dá-se o nome de potencial pós-sináptico à alteração do potencial de membrana, provocada pela actividade da sinapse, a qual pode, como já se referiu, ser excitatória ou inibitória. A cada potencial de acção que atinge o terminal de um neurónio pré-sináptico pode estar associado um potencial pós-sináptico de, aproximadamente, 1 mV o que significa, tendo em consideração o valor do patamar a partir do qual surge um potencial de acção, que é necessária a soma de vários potenciais para o desencadeamento destes sinais no neurónio pós-sináptico. Na prática verifica-se que, estabelecendo cada neurónio centenas de sinapses (pode chegar a estabelecer, nos casos em que o número de ligações é maior, cerca de 150 000 sinapses, o seu comportamento é determinado pela integração de todas as fontes de informação que a ele afluem. Essa integração espacial e temporal é responsável pela duração dos potenciais pós-sinápticos que pode ser na ordem do segundo, até vários minutos. Deste modo, as sinapses excitatórias e inibitórias, ao coexistirem no mesmo neurónio, são responsáveis pelo facto da resposta desse neurónio a diferentes estímulos, ser uma integração complexa dos diversos pulsos que a ele afluem. Ou seja, um neurónio estimulado por diversas sinapses, pode ou não criar potenciais de acção, dependendo das sinapses inibitórias que, em simultâneo com as excitatórias, se tornem activas. Assim, o aparecimento de potenciais pós-sinápticos excitatórios num neurónio é tão determinante na criação de potenciais de acção como o surgimento de potenciais pós-sinápticos inibitórios, uma vez que o efeito destes últimos pode reduzir ou cancelar o efeito dos primeiros.
3.2 Organização cerebral e actividade eléctricaNas secções anteriores foram abordadas diversas vertentes da actividade
eléctrica das células cerebrais. Falta, porém, realçar alguns aspectos relacionados com fenómenos eléctricos que envolvem populações de neurónios. A electroencefalografia, técnica a que nos referiremos com maior detalhe nos sub-capítulos posteriores e que consiste na medição de potenciais eléctricos ao nível

53
do escalpe, revela-nos que existe actividade eléctrica cerebral síncrona. Isto é, verifica-se que existem grupos de neurónios cuja actividade ocorre em simultâneo, de modo que geram ritmos susceptíveis de ser medidos no exterior do crânio. Por este motivo, cedo se especulou no sentido de o cérebro se organizar segundo circuitos neuronais cada um dos quais responsável pelo processamento de um determinado tipo de informação. Esta ideia tem sido corroborada por numerosos estudos que apontam para a especificidade de determinadas regiões do cérebro. Aceita-se que no lobo occipital se encontram os córtices visuais, associados ao processamento da visão; nos lobos temporais os córtices auditivos, no lobo parietal os córtices somato-sensoriais e no lobo frontal os córtices motor e pré-motor. Refira-se, no entanto, que, se a comunidade científica assume unanimemente que as tarefas mais simples associadas à percepção dos sentidos estão razoavelmente localizadas no cérebro, não é menos verdade que, ao nível das actividades com carácter cognitivo mais evidente, muito se tem especulado. De facto, as evidências experimentais apontam para que as tarefas mais complexas relacionadas com a memória, a aprendizagem ou as emoções, abranjam áreas cerebrais muito amplas e deslocalizadas. Estas áreas ou circuitos parecem trabalhar separadamente, numa espécie de processamento em paralelo, onde cada uma se encontra envolvida num aspecto particular da tarefa comum. O que se mantém sem resposta é o modo como finalmente toda essa informação é coligida, guardada e recuperada em novas situações.
3.3 Equipamento
O electroencefalograma (EEG) tem um princípio muito semelhante ao do ECG, mas mede a actividade eléctrica cerebral. Em termos gerais estabelecem-se dois tipos de exame: os registos espontâneos, onde as diferenças de potencial são medidas continuamente e sem a presença de estímulos exteriores, e os registos evocados que são os potenciais associados à resposta cerebral a um estímulo que pode ser visual, auditivo ou sensorial.
3.3.1 Registos electroencefalográficosOs primeiros registos de EEG são datados de 1928 e, logo após as primeiras
medições da actividade cerebral não-invasivamente, tornou-se claro que as características do traçado eram fortemente dependentes do estado de repouso do indivíduo. De facto, uma parte significativa do registo de EEG espontâneo é extremamente irregular, tornando-se difícil a sua caracterização. Porém, quer por simples inspecção visual, quer através do recurso a técnicas de Transformadas de Fourier que permitem o cálculo dos espectros de potência dos registos, facilmente se verifica que existem frequências dominantes correspondentes a diferentes estados de vigília (naFigura 51 encontram-se esquematizados exemplos de diversos traçados espontâneos). Além disso, é possível detectar diversas patologias através da análise do EEG, uma vez que se podem observar quer notórias alterações nos padrões de frequências considerados normais, quer a existência de grafo-elementos específicos da doença.

54
Figura 51 - Exemplos de diversos traçados de EEG espontâneo. Os fusos são sinais que aparecem no início do sono, as ondas delta são típicas dos instantes de sono mais profundo, o ritmo beta aparece tanto durante a fase de sono REM (Rapid Eyes Movements), como durante a vigília quando o indivíduo se encontra com a sua atenção focalizada e o ritmo beta aparece em repouso. (Adapt. Guyton e Hall, 1996).
Quanto aos potenciais evocados há a referir que quando um sujeito é estimulado visual, auditiva ou sensorialmente desencadeia-se, nos hemisférios cerebrais, um processo complexo de recolha, transmissão e processamento de informação que corresponde às diversas etapas da percepção. Após o estímulo, existem, pois, neurónios, ou grupos de neurónios, que se encontram activos em simultâneo ou sequencialmente dando origem a potenciais eléctricos passíveis de ser medidos ao nível do escalpe. Refira-se, porém, que as amplitudes dos potenciais correspondentes ao estímulo (potenciais ou respostas evocadas), quando comparadas com as dos potenciais espontâneos constantemente presentes no registo electroencefalográfico, são várias vezes menores (os registos espontâneos têm tipicamente, uma amplitude de 10-30V, enquanto os potenciais evocados têm uma amplitude mínima de 0.5V). Por este motivo, com o intuito de tornar visível os potenciais evocados (PE), é usual repetir os estímulos (habitualmente entre 50 e 200 vezes, dependendo do tipo de estímulo e da relação sinal/ruído que se pretender) e ir somando os registos dos potenciais referentes aos instantes subsequentes. Partindo do princípio que os potenciais espontâneos são independentes do estímulo, este procedimento permite fazer emergir os PE do ruído provocado pelos potenciais espontâneos (ver Figura 52).
Os PE referentes a um determinado estímulo podem apresentar diversos picos de amplitude aos quais se dá o nome de componentes (ver Figura 53). Cada uma destas componentes está relacionada com uma ou mais etapas de processamento de informação e é caracterizada pela sua latência (intervalo de tempo entre o estímulo e a componente). Actualmente, existem diversos estudos onde são comparadas as latências das diferentes componentes dos PE relativos a indivíduos saudáveis e a doentes, verificando-se, em muitos casos, desvios significativos. Assim, a medição de PE em doentes com distúrbios neurológicos é cada vez mais uma prática corrente na clínica, tendo-se revelado como uma importante ferramenta ao nível do diagnóstico.
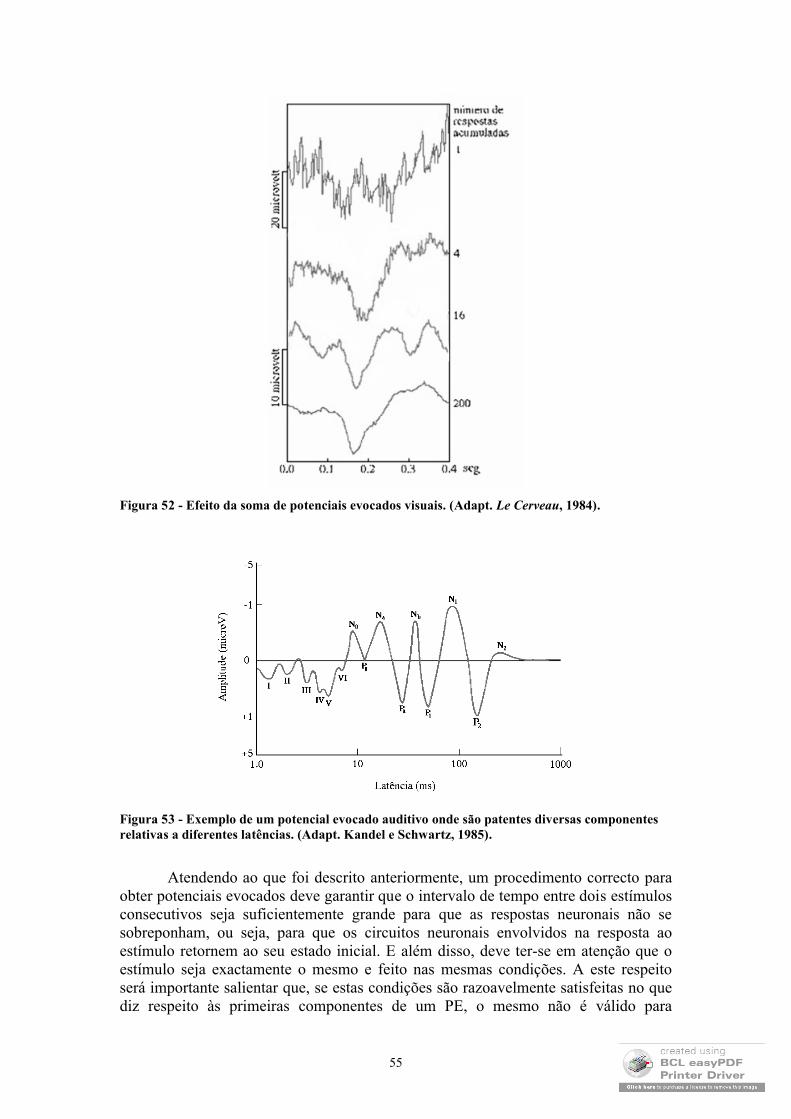
55
Figura 52 - Efeito da soma de potenciais evocados visuais. (Adapt. Le Cerveau, 1984).
Figura 53 - Exemplo de um potencial evocado auditivo onde são patentes diversas componentes relativas a diferentes latências. (Adapt. Kandel e Schwartz, 1985).
Atendendo ao que foi descrito anteriormente, um procedimento correcto para obter potenciais evocados deve garantir que o intervalo de tempo entre dois estímulos consecutivos seja suficientemente grande para que as respostas neuronais não se sobreponham, ou seja, para que os circuitos neuronais envolvidos na resposta ao estímulo retornem ao seu estado inicial. E além disso, deve ter-se em atenção que o estímulo seja exactamente o mesmo e feito nas mesmas condições. A este respeito será importante salientar que, se estas condições são razoavelmente satisfeitas no que diz respeito às primeiras componentes de um PE, o mesmo não é válido para

56
componentes de maior latência. Na realidade, e como seria de esperar, conforme a latência vai aumentando, os potenciais medidos vão sendo progressivamente mais complexos, estando relacionados com processamentos cada vez mais elaborados. Assim, se as primeiras componentes dependem maioritariamente do tipo de estímulo esão respeitantes à simples percepção do mesmo, as de latência mais elevada são fortemente dependentes de mecanismos cognitivos como a atenção ou a expectativa, pelo que se torna difícil garantir que o indivíduo mantenha a mesma atitude cognitiva perante o estímulo desde o início do exame até ao seu final.
3.3.2 HardwareEm traços gerais, os modernos equipamentos de EEG resumem-se a dois
grandes blocos: o sistema de medida (eléctrodos, amplificadores e restante material de registo) e todo um conjunto logicial necessário para o processamento dos dados (técnicas de imagem, transformadas de Fourier de processamento rápido (FFT11), estatística de comparação de populações, localização de fontes eléctricas neuronais, etc.).
Tendo em conta que a posição dos eléctrodos no escalpe é uma questão que deve obedecer a determinados critérios, em 1958 um comité da Federação Internacional das Sociedades de Electroencefalografia e Neurofisiologia Clínica12
emitiu uma recomendação onde descrevia um modo de padronizar a distribuição dos eléctrodos sobre o escalpe: o sistema internacional 10/20. O principal objectivo desta recomendação foi permitir uma fácil comparação entre os resultados obtidos em diferentes laboratórios.
Na Figura 54 encontra-se esquematizada a distribuição sugerida, com a nomenclatura dos diversos eléctrodos. Esta distribuição pressupõe o posicionamento de 21 eléctrodos.
Figura 54 - Posição esquemática dos eléctrodos segundo o sistema internacional 10/20. (Adapt. Lewine e Orrison Jr., 1995).
A colocação dos eléctrodos reveste-se de grande importância, visto que a qualidade das medidas electroencefalográficas é extremamente dependente do modo como o contacto entre os eléctrodos e o escalpe é feito. Para tanto, é necessário esfregar os locais onde os eléctrodos serão posicionados com um gel electrolítico. Este procedimento permite limpar essa região, favorecendo o contacto. Em seguida, colocam-se os eléctrodos nas posições padronizadas, usando para o efeito um capacete ou uma touca que de adapta à medida das diferentes cabeças. Existem diversos tipos de eléctrodos, que variam na forma e no modo de fixação: podem ser 11 Do inglês - Fast Fourier Transform.12 International Federation of Societies for Electroencephalography and Clinical Neurophysiology.

57
aderentes à pele, não precisando de fixação adicional; podem usar um sistema de mola para se segurarem à pele (estes eléctrodos não podem ser usados no escalpe e são unicamente usados nas orelhas, como eléctrodos de referência) ou podem ser fixos através de uma pasta que facilita a aderência do eléctrodo. O uso de toucas onde os eléctrodos são fixos, torna mais fácil manter os eléctrodos na mesma posição durante todo o exame. Por fim, deve garantir-se que a impedância de todos os eléctrodos é a adequado ao registo.
A escolha dos eléctrodos de referência tem sido amplamente discutida, devendo ser criteriosa, visto que se o(s) eléctrodo(s) de referência estiver(em) contaminado(s) com a actividade que se pretender observar, os dados ficam mascarados e pode mesmo não se conseguir discernir os sinais em causa. As estratégias de registo dos potenciais dividem-se em: montagens monopolares montagens em que as medidas de todos os eléctrodos são feitas com referência a um ou mais eléctrodos e montagens bipolares a medida de cada canal é referente à diferença entre dois eléctrodos, geralmente adjacentes. As montagens monopolares, apesar de serem aquelas que permitem uma mais fácil comparação entre os registos, têm a dificuldade de nenhum eléctrodo ser suficientemente isento de actividade eléctrica. De um modo geral, a média de dois eléctrodos colocados sobre as orelhas ou sobre os mentos é considerada como uma boa opção, ainda que mesmo assim haja sempre contaminação por actividade eléctrica. Uma outra referência que se usa é a média dos valores medidos em todos os eléctrodos. Esta é uma forma de contornar situações em que a referência não seja muito boa, ainda que apresente a desvantagem de alisar as amplitudes medidas, em particular para actividades elevadas.
Apesar de as questões relacionadas com os eléctrodos possuírem uma enorme relevância num exame de EEG, existem ainda outros factores que devem ser enfatizados: a calibração do equipamento, a sua sensibilidade e resolução, a sua resposta temporal e os filtros utilizados. Durante o protocolo inicial para o registo dos potenciais, após a colocação dos eléctrodos e a medida das suas impedâncias, procede-se à calibração dos amplificadores. Nessa operação sujeita-se os amplificadores a uma tensão nula, de modo a fazer o ajuste do zero e, em seguida, a uma determinada tensão, de modo a calibrá-los.
Uma questão importante que se coloca é o da resolução do equipamento de medida. Nos aparelhos mais antigos, em que o registo era apenas gráfico, a resolução do mesmo estava associado à diferença entre duas tensões próximas capazes de fazer mexer a caneta. Actualmente, as medidas são, normalmente, arquivadas digitalmente (disco rígido, disco óptico, diskette, etc.), após serem submetidas a um conversor analógico-digital (CAD), cujos níveis de amplitude determinam a sua resolução. Em termos de valor absoluto de tensão, esta resolução pode variar de exame para exame, uma vez que é sempre possível definir a gama de valores que se pretendem medir através de adequadas amplificações (valores mais baixos para potenciais evocados, por exemplo). Associada à resolução existe ainda uma outra grandeza, importante em qualquer sistema de medida, que é a sensibilidade (valor mínimo capaz de ser registado). Tendo em conta a sua definição, numa primeira abordagem, a sensibilidade poderá ser confundida com resolução, no entanto, note-se que a sensibilidade poderá ser menor do que a resolução, devido à presença de ruído no equipamento. Assim, a sensibilidade está dependente da qualidade do equipamento e o seu valor deve ser tipo em conta em medições que envolvam pequenas amplitudes do sinal.

58
Uma outra característica de um exame de EEG é a sua taxa de aquisição. Para além de uma quantização em amplitude (imposta pelo CAD), um sinal de EEG é também quantizado em tempo, de modo que um sinal de EEG não é mais do que uma série temporal, sendo o tempo entre amostras determinado pela rapidez com que o equipamento electrónico consegue recolher e guardar informação. A escolha da taxa de aquisição ou amostragem do sinal, para além das limitações técnicas, deve ser escolhida atendendo a um compromisso entre a resolução temporal que se pretender e a memória ocupada pelos dados. De facto, alguns dos potenciais evocados sensitivos, por exemplo, de maior interesse, têm latências na ordem das dezenas de milisegundo, pelo que é comum, nestes casos, usarem-se taxas de aquisição superiores a 500Hz. No entanto, para exames de actividade espontânea cujo registo corresponde, em geral, a cerca de 20 minutos, usar frequências na ordem dos 500Hz para esses registos na rotina clínica, colocaria alguns problemas de memória na manipulação desses dados.
O tipo de filtros utilizados num registo de EEG é uma outra questão muito pertinente. De facto, a escolha de um filtro é sempre um compromisso entre o que se pretende eliminar de um sinal exterior e o que se pretende manter do sinal medido. Mais uma vez, a escolha dos filtros a aplicar deve ser feita em consonância com o tipo de medidas que se está a efectuar. Ou seja, as frequências características do sinal a analisar devem ser mantidas, na medida do possível, inalteradas. Refira-se, a este propósito, que actualmente, com a construção de filtros digitais bastante fiáveis, tende-se a abandonar o uso de filtros analógicos muito limitativos, os quais não permitem a recuperação do sinal. Em vez destes tende-se a usar filtros digitais que podem ser aplicados após a recolha dos dados.
3.3.3 SoftwareUma grande parte dos procedimentos abordados nas últimas secções:
determinação da impedância dos eléctrodos, calibração dos amplificadores, escolha da amplificação dos sinais, da sua taxa de aquisição e dos filtros aplicados, são, nos equipamentos modernos, controlados logicialmente. Para além destas especificidades técnicas, o operador pode ainda controlar o tipo de registo que pretende efectuar: registo espontâneo ou potenciais evocados e, neste último caso, explicitar os parâmetros dos estímulos tipo de estímulo, frequência e intensidade do mesmo, tempo de análise, etc. Esta forma de fixar os parâmetros é muito eficiente no que respeita a padronizar os resultados, uma vez que para cada exame existe um menu por defeito que só não será o usado em casos especiais.
Os equipamentos mais modernos de EEG são acompanhados por um módulo logicial completo que permite a análise detalhada dos sinais recolhidos. Nesta secção pretende-se referir algumas das capacidades desse equipamento.
Mapas - Antes de mais, é possível visualizar os sinais medidos quer de um modo tradicional registo de uma curva que contém a amplitude do sinal ao longo do tempo13 quer em forma de mapa a actividade medida é interpolada para diferentes pontos da cabeça e visualizada num mapa cerebral bi-dimensional (Figura55).
13 A título de curiosidade, mencione-se que, por convenção, os sinais de EEG são apresentados com os potenciais negativos no sentido positivo do eixo dos yy e os potenciais positivos no sentido negativo do eixo dos yy.

59
Figura 55 - Exemplo de um mapa cerebral onde estão representados os potenciais medidos e o resultado da interpolação dos mesmos.
Médias de pontas – É possível seleccionar vários sinais e somá-los, fazendo-os coincidir no tempo (escolhe-se o instante de amplitude máxima e sobrepõem-se os sinais). Este procedimento permite obter uma melhor relação sinal/ruído, no entanto, pode revelar-se falacioso, uma vez que é possível realizar somas de sinais que não provenham da mesma origem. Para evitar este inconveniente, o operador deve escolher apenas sinais que sejam caracterizados por topologias inequivocamente similares.
Estatística - Com o intuito de proceder a uma objectiva avaliação do carácter dos traçados de EEG, cedo se tentou determinar parâmetros ou grandezas que os caracterizassem. Desta forma, surgiu a ideia de aplicar aos registos electroencefalográficos técnicas estatísticas desenvolvidas para sistemas cujo comportamento é muito complexo e imprevisível. Neste contexto, é possível, calcular a média do sinal, os seus momentos, etc. e comparar diferentes sinais ou diferentes troços de um mesmo sinal, através destas grandezas. Este tipo de análise permite ainda a comparação entre populações, sendo, deste modo, possível, avaliar a influência de determinado factor no EEG de um indivíduo normal ou doente.
Transformadas de Fourier - Uma área que tem sido extremamente explorada no processamento de dados electroencefalográficos é o das técnicas de transformada de Fourier rápida. Aliás, dada a importância de que os ritmos cerebrais se revestem esta ferramenta tem sido amplamente desenvolvida, sendo possível, por exemplo: 1) a construção de mapas correspondentes a determinadas bandas de frequência, o que permite estabelecer relações entre estas e as regiões cerebrais e 2) a comparação dos espectros correspondentes a diferentes populações ou à mesma população, mas em diferentes condições.
Uso de diferentes montagens montagem laplaciana - Uma outra possibilidade que se coloca ao operador que dispõe dos dados guardados digitalmente é a de alterar a montagem dos eléctrodos e pesquisar se existe alguma informação que se torne mais visível após essa modificação. Assim, como foi anteriormente discutido, de um modo geral, os dados são recolhidos através de uma montagem monopolar (habitualmente, usando um ou dois eléctrodos de referência) e é depois possível visualizá-los em montagens monopolares correspondentes a outras referências (média de todos os eléctrodos, por exemplo) ou em montagens bipolares (basta calcular as diferenças entre eléctrodos).

60
4 ELECTROCARDIOGRAFIA (Facultativo)
Este capítulo terá uma organização análoga à do anterior. Ou seja, começar-se-á por referir alguns aspectos da fisiologia dos músculos, com especial ênfase para o músculo cardíaco, e terminar-se-á com a explicação de como se regista a actividade eléctrica do coração, utilizando electrocardiografia.
4.1 Aspectos da electricidade dos músculos
Tal como o sistema nervoso central, também a actividade muscular depende de uma forma crucial das propriedades eléctricas das células constituintes dos músculos. Os músculos dividem-se em três categorias: os músculos esqueléticos, os lisos e o cardíaco. Os primeiros encontram-se essencialmente ligados aos ossos e a sua contracção, geralmente voluntária, é responsável pelo suporte e movimento do esqueleto. Os lisos são músculos que envolvem diversos órgãos ou estruturas como o estômago, os intestinos ou os vasos sanguíneos e as suas contracções, involuntárias e coordenadas pelo sistema nervoso autónomo e pelo sistema endócrino, implicam o movimento desses órgãos e estruturas. Quanto ao coração, apresenta características de ambos os tipos de músculo (esquelético e liso) e tem a particularidade de contrair espontaneamente.
4.1.1 Os músculos esqueléticosOs músculos esqueléticos são constituídos por células denominadas fibras
musculares que, por sua vez, provêm, em termos de desenvolvimento fetal, de um conjunto de células indiferenciadas e mononucleadas às quais se dá o nome de mioblastos (ver Figura 56). Estas fibras têm diâmetros entre 10 a 100 m e comprimentos que podem ir até 20 cm, não apresentando capacidade de reprodução ao longo da vida.
Quando vistas ao microscópio a principal característica das fibras musculares é apresentarem um padrão de estrias que podem ser observadas, em esquema, na Figura 5714. Estas estrias correspondem à existência de pequenos filamentos com diferentes espessuras organizados em finos cilindros no interior do citoplasma das fibras musculares. O arranjo destes filamentos obedece a um padrão que se repete, cuja unidade fundamental é o sarcómero. Cada uma destas unidades contém, no centro, um conjunto de filamentos grossos que constituem a chamada banda A e em cada um dos extremos um conjunto de filamentos finos que constituem a banda I. Além disso, os filamentos finos sobrepõem-se, numa certa extensão, com os filamentos grossos e pertencem simultaneamente a dois sarcómeros (ver Figura 57). À divisão entre dois sarcómeros dá-se o nome de linha Z.
14 Esta propriedade é comum ao músculo cardíaco e responsável por estes dois tipos de músculos serem também conhecidos por estriados.
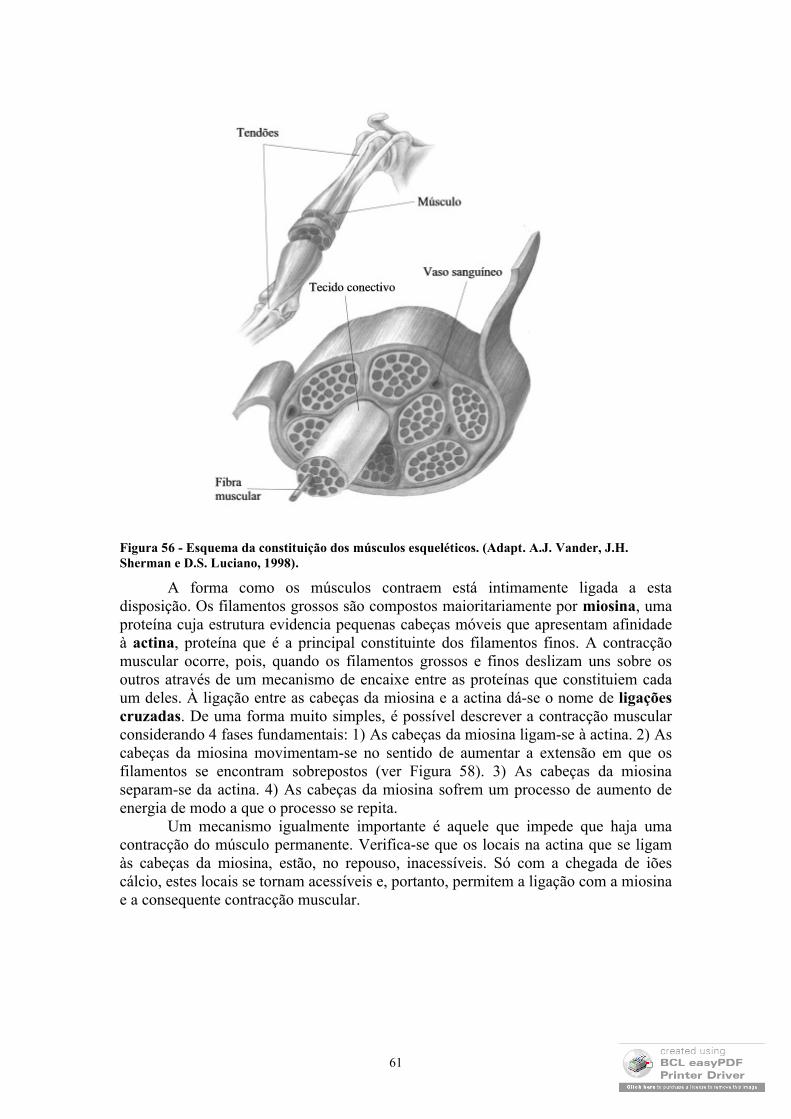
61
Figura 56 - Esquema da constituição dos músculos esqueléticos. (Adapt. A.J. Vander, J.H. Sherman e D.S. Luciano, 1998).
A forma como os músculos contraem está intimamente ligada a esta disposição. Os filamentos grossos são compostos maioritariamente por miosina, uma proteína cuja estrutura evidencia pequenas cabeças móveis que apresentam afinidade à actina, proteína que é a principal constituinte dos filamentos finos. A contracção muscular ocorre, pois, quando os filamentos grossos e finos deslizam uns sobre os outros através de um mecanismo de encaixe entre as proteínas que constituiem cada um deles. À ligação entre as cabeças da miosina e a actina dá-se o nome de ligações cruzadas. De uma forma muito simples, é possível descrever a contracção muscular considerando 4 fases fundamentais: 1) As cabeças da miosina ligam-se à actina. 2) As cabeças da miosina movimentam-se no sentido de aumentar a extensão em que os filamentos se encontram sobrepostos (ver Figura 58). 3) As cabeças da miosina separam-se da actina. 4) As cabeças da miosina sofrem um processo de aumento de energia de modo a que o processo se repita.
Um mecanismo igualmente importante é aquele que impede que haja uma contracção do músculo permanente. Verifica-se que os locais na actina que se ligam às cabeças da miosina, estão, no repouso, inacessíveis. Só com a chegada de iões cálcio, estes locais se tornam acessíveis e, portanto, permitem a ligação com a miosina e a consequente contracção muscular.

62
Figura 57 - Esquema da constituição das miofibrilhas constituintes dos músculos esqueléticos. (Adapt. A.J. Vander, J.H. Sherman e D.S. Luciano, 1998).
Figura 58 - Ilustração de como o movimento das cabeças da miosina é responsável pela alteração das dimensões dos músculos esqueléticos, determinando a sua contracção ou distensão. (Adapt. A.J. Vander, J.H. Sherman e D.S. Luciano, 1998).
É neste ponto que os aspectos eléctricos se tornam importantes na compreensão da contracção muscular. A membrana das fibras musculares é, tal como a dos neurónios, capaz de gerar e propagar potenciais de acção. No caso dos músculos o efeito crucial do aparecimento de um potencial de acção é a libertação de iões cálcio15, que vão, por sua vez, induzir a contracção muscular segundo o mecanismo anteriormente descrito. A este respeito, observa-se que, um único potencial de acção de 1 a 2 ms provoca um estado de contracção muscular que se prolonga tipicamente por cerca de 100 ms. Este facto deve-se a que enquanto a libertação de iões cálcio é 15 Neste mecanismo de libertação de cálcio existe uma estrutura constituinte do músculo esquelético o retículo sarcoplasmático que se reveste de particular interesse no armazenamento e libertação dos iões cálcio. No entanto, uma discussão aprofundada sobre o seu papel neste mecanismo encontra-se fora dos objectivos desta disciplina.

63
bastante rápida, a sua remoção é um processo mais lento e, portanto, o repouso, após a contracção, é mais demorado de atingir.
Falta referir ainda a origem dos potenciais de acção: Os músculos esqueléticos encontram-se ligados a terminais nervosos de neurónios motores. Estes neurónios estabelecem ligações com as fibras musculares, as junções neuromusculares16, e são eles que controlam a actividade dos músculos (ver Figura 59).
Figura 59 - Esquema das junções neuromusculares que governam o movimento dos músculos. Na alínea a) encontra-se representado um único neurónio motor, enquanto que na alínea b) encontram-se representados dois neurónios, deixando claro que cada neurónio não controla necessariamente fibras musculares adjacentes (Adapt. A.J. Vander, J.H. Sherman e D.S. Luciano, 1998).
Os mecanismos de troca de informação ao nível das junções neuromusculares são muito semelhantes aos que ocorrem nas sinapses químicas: o potencial de acção ao atingir o terminal do axónio, liberta neurotransmissores que são reconhecidos pela membrana da fibra muscular que, ao detectá-los, desencadeia a abertura de canais de cálcio, iniciando o processo de contracção.
4.1.2 Os músculos lisosConforme se referiu anteriormente, os músculos lisos recobrem a maior parte
dos órgãos e uma parte dos vasos sanguíneos, conferindo-lhes movimentos de contracção e distensão. Distingue-os dos músculos esqueléticos o facto de não apresentarem uma estrutura estriada e dos nervos que os controlam provirem do sistema nervoso autónomo, o que torna os seus movimentos involuntários. As fibras constituintes do tecido do músculo liso, ao contrário das fibras do músculo esquelético, são células em forma de fuso, com apenas um núcleo e que se reproduzem. No interior destas células encontram-se também filamentos de miosina (filamentos grossos) e de actina (filamentos finos), no entanto, a sua organização não 16 Cada neurónio motor estabelece ligação com mais do que uma fibra muscular, mas cada fibra muscular só recebe informação de um único neurónio. Além disso, refira-se que fibras musculares associadas ao mesmo neurónio não têm necessariamente que ser adjacentes (ver Figura 59).

64
é semelhante à encontrada nos músculos esqueléticos. Os filamentos encontram-se igualmente sobrepostos, sendo o mecanismo de contracção semelhante ao anteriormente explicado a propósito dos músculos esqueléticos, mas em vez dos filamentos se encontrarem paralelos uns aos outros, encontram-se ancorados à membrana da célula e a certos pontos do citoplasma denominados corpos densos (ver Figura 60). Esta geometria faz com que quando os filamentos deslizam uns sobre os outros as dimensões da célula diminuam e, portanto, o músculo se contraia.
Figura 60 - Esquema das ligações entre os filamentos finos e grossos num músculo liso em duas situações distintas: quando o músculo se encontra relaxado e quando se encontra contraído. (Adapt. A.J. Vander, J.H. Sherman e D.S. Luciano, 1998).
Embora, em detalhe, o mecanismo bioquímico através do qual as cabeças da miosina se ligam à actina seja diferente, a presença do cálcio continua a ser o factor que despoleta a ligação das duas proteínas. É interessante observar que a contracção dos músculos lisos é dependente da quantidade de cálcio libertada, uma vez que um potencial de acção induz contracção apenas numa porção das fibras dos músculos lisos17. Além disso, a contracção dos músculos lisos tem uma duração que pode chegar a vários segundos, visto que os mecanismos responsáveis pela sua remoção são muito lentos.
Para além do que foi já referido como diferenças entre os músculos esqueléticos e os músculos lisos, há ainda a acrescentar o facto de alguns músculos lisos apresentarem, tal como o coração, a possibilidade de gerarem, espontaneamente, potenciais de acção. Nestes músculos o potencial de membrana, ao invés de se manter constante, vai continuamente despolarizando. Deste modo, a dado momento, é atingido o limiar de excitação e gerado um potencial de acção. Após a repolarização o ciclo repete-se, gerando-se sinais ritmadamente (ver Figura 61).
17 Recorde-se que nos músculos esqueléticos apenas um potencial de acção é capaz de induzir contracção em todo o músculo.
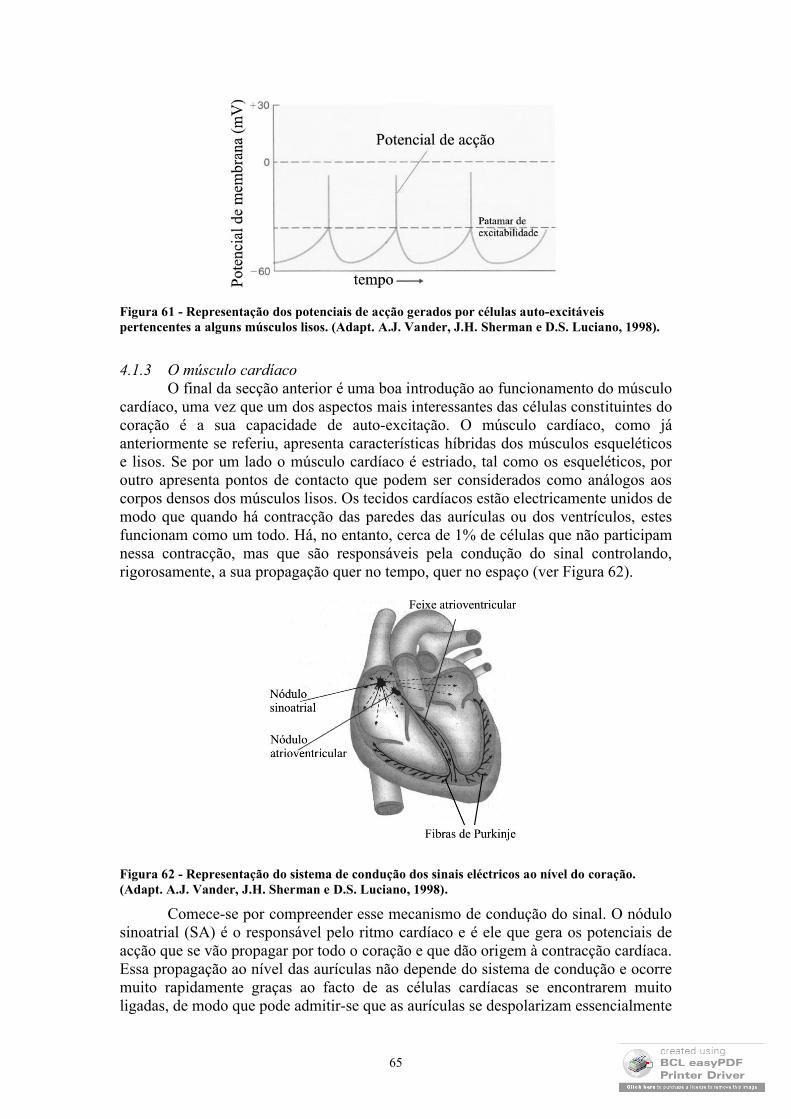
65
Figura 61 - Representação dos potenciais de acção gerados por células auto-excitáveis pertencentes a alguns músculos lisos. (Adapt. A.J. Vander, J.H. Sherman e D.S. Luciano, 1998).
4.1.3 O músculo cardíacoO final da secção anterior é uma boa introdução ao funcionamento do músculo
cardíaco, uma vez que um dos aspectos mais interessantes das células constituintes do coração é a sua capacidade de auto-excitação. O músculo cardíaco, como já anteriormente se referiu, apresenta características híbridas dos músculos esqueléticos e lisos. Se por um lado o músculo cardíaco é estriado, tal como os esqueléticos, por outro apresenta pontos de contacto que podem ser considerados como análogos aos corpos densos dos músculos lisos. Os tecidos cardíacos estão electricamente unidos de modo que quando há contracção das paredes das aurículas ou dos ventrículos, estes funcionam como um todo. Há, no entanto, cerca de 1% de células que não participam nessa contracção, mas que são responsáveis pela condução do sinal controlando, rigorosamente, a sua propagação quer no tempo, quer no espaço (ver Figura 62).
Figura 62 - Representação do sistema de condução dos sinais eléctricos ao nível do coração. (Adapt. A.J. Vander, J.H. Sherman e D.S. Luciano, 1998).
Comece-se por compreender esse mecanismo de condução do sinal. O nódulo sinoatrial (SA) é o responsável pelo ritmo cardíaco e é ele que gera os potenciais de acção que se vão propagar por todo o coração e que dão origem à contracção cardíaca. Essa propagação ao nível das aurículas não depende do sistema de condução e ocorre muito rapidamente graças ao facto de as células cardíacas se encontrarem muito ligadas, de modo que pode admitir-se que as aurículas se despolarizam essencialmente
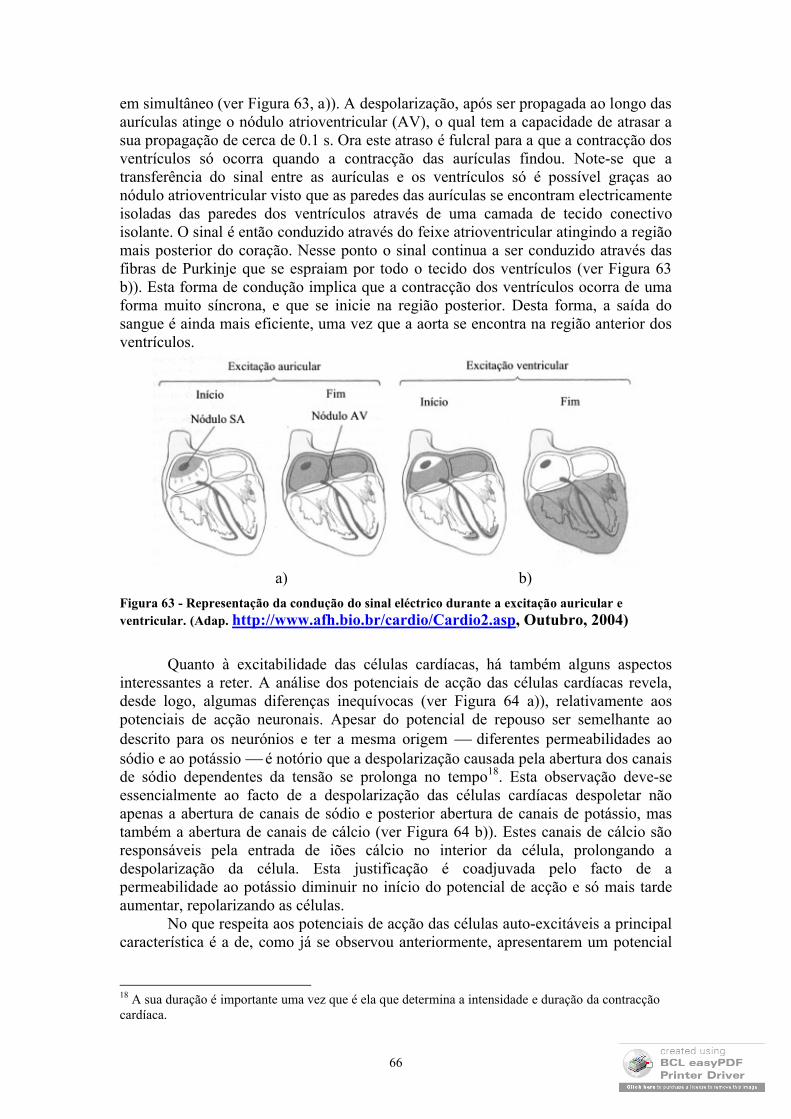
66
em simultâneo (ver Figura 63, a)). A despolarização, após ser propagada ao longo das aurículas atinge o nódulo atrioventricular (AV), o qual tem a capacidade de atrasar a sua propagação de cerca de 0.1 s. Ora este atraso é fulcral para a que a contracção dos ventrículos só ocorra quando a contracção das aurículas findou. Note-se que a transferência do sinal entre as aurículas e os ventrículos só é possível graças ao nódulo atrioventricular visto que as paredes das aurículas se encontram electricamente isoladas das paredes dos ventrículos através de uma camada de tecido conectivo isolante. O sinal é então conduzido através do feixe atrioventricular atingindo a região mais posterior do coração. Nesse ponto o sinal continua a ser conduzido através das fibras de Purkinje que se espraiam por todo o tecido dos ventrículos (ver Figura 63b)). Esta forma de condução implica que a contracção dos ventrículos ocorra de uma forma muito síncrona, e que se inicie na região posterior. Desta forma, a saída do sangue é ainda mais eficiente, uma vez que a aorta se encontra na região anterior dos ventrículos.
a) b)
Figura 63 - Representação da condução do sinal eléctrico durante a excitação auricular e ventricular. (Adap. http://www.afh.bio.br/cardio/Cardio2.asp, Outubro, 2004)
Quanto à excitabilidade das células cardíacas, há também alguns aspectos interessantes a reter. A análise dos potenciais de acção das células cardíacas revela, desde logo, algumas diferenças inequívocas (ver Figura 64 a)), relativamente aos potenciais de acção neuronais. Apesar do potencial de repouso ser semelhante ao descrito para os neurónios e ter a mesma origem diferentes permeabilidades ao sódio e ao potássio é notório que a despolarização causada pela abertura dos canais de sódio dependentes da tensão se prolonga no tempo18. Esta observação deve-se essencialmente ao facto de a despolarização das células cardíacas despoletar não apenas a abertura de canais de sódio e posterior abertura de canais de potássio, mas também a abertura de canais de cálcio (ver Figura 64 b)). Estes canais de cálcio são responsáveis pela entrada de iões cálcio no interior da célula, prolongando a despolarização da célula. Esta justificação é coadjuvada pelo facto de a permeabilidade ao potássio diminuir no início do potencial de acção e só mais tarde aumentar, repolarizando as células.
No que respeita aos potenciais de acção das células auto-excitáveis a principal característica é a de, como já se observou anteriormente, apresentarem um potencial
18 A sua duração é importante uma vez que é ela que determina a intensidade e duração da contracção cardíaca.
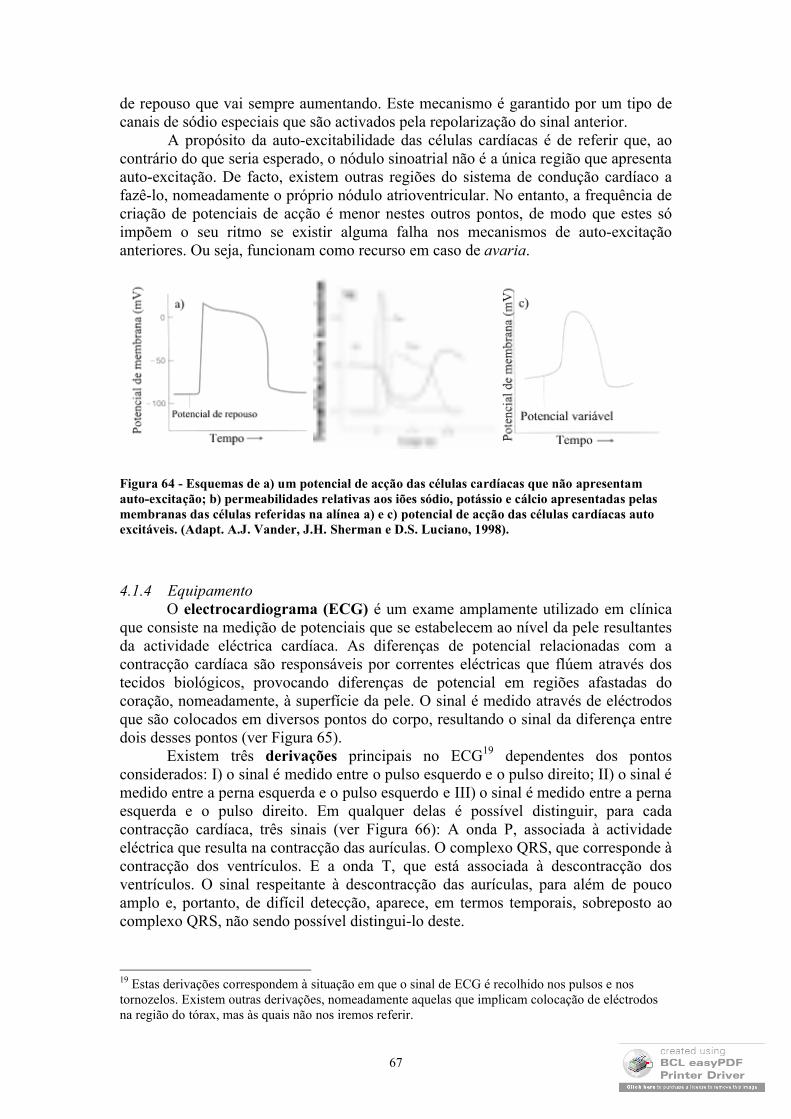
67
de repouso que vai sempre aumentando. Este mecanismo é garantido por um tipo de canais de sódio especiais que são activados pela repolarização do sinal anterior.
A propósito da auto-excitabilidade das células cardíacas é de referir que, ao contrário do que seria esperado, o nódulo sinoatrial não é a única região que apresenta auto-excitação. De facto, existem outras regiões do sistema de condução cardíaco a fazê-lo, nomeadamente o próprio nódulo atrioventricular. No entanto, a frequência de criação de potenciais de acção é menor nestes outros pontos, de modo que estes só impõem o seu ritmo se existir alguma falha nos mecanismos de auto-excitação anteriores. Ou seja, funcionam como recurso em caso de avaria.
Figura 64 - Esquemas de a) um potencial de acção das células cardíacas que não apresentam auto-excitação; b) permeabilidades relativas aos iões sódio, potássio e cálcio apresentadas pelas membranas das células referidas na alínea a) e c) potencial de acção das células cardíacas auto excitáveis. (Adapt. A.J. Vander, J.H. Sherman e D.S. Luciano, 1998).
4.1.4 EquipamentoO electrocardiograma (ECG) é um exame amplamente utilizado em clínica
que consiste na medição de potenciais que se estabelecem ao nível da pele resultantes da actividade eléctrica cardíaca. As diferenças de potencial relacionadas com a contracção cardíaca são responsáveis por correntes eléctricas que flúem através dos tecidos biológicos, provocando diferenças de potencial em regiões afastadas do coração, nomeadamente, à superfície da pele. O sinal é medido através de eléctrodos que são colocados em diversos pontos do corpo, resultando o sinal da diferença entre dois desses pontos (ver Figura 65).
Existem três derivações principais no ECG19 dependentes dos pontos considerados: I) o sinal é medido entre o pulso esquerdo e o pulso direito; II) o sinal é medido entre a perna esquerda e o pulso esquerdo e III) o sinal é medido entre a perna esquerda e o pulso direito. Em qualquer delas é possível distinguir, para cada contracção cardíaca, três sinais (ver Figura 66): A onda P, associada à actividade eléctrica que resulta na contracção das aurículas. O complexo QRS, que corresponde à contracção dos ventrículos. E a onda T, que está associada à descontracção dos ventrículos. O sinal respeitante à descontracção das aurículas, para além de pouco amplo e, portanto, de difícil detecção, aparece, em termos temporais, sobreposto ao complexo QRS, não sendo possível distingui-lo deste.
19 Estas derivações correspondem à situação em que o sinal de ECG é recolhido nos pulsos e nos tornozelos. Existem outras derivações, nomeadamente aquelas que implicam colocação de eléctrodos na região do tórax, mas às quais não nos iremos referir.

68
Figura 65 - Esquema de um electrocardiógrafo. (Adapt. de P. Davidovits, 2001).
Figura 66 - Sinal típico de electrocardiografia. (Adapt. de P. Davidovits, 2001).
Os sinais medidos são na ordem dos milivolt, de modo que, embora sejam filtrados com o objectivo de os libertar de frequências indesejáveis, são, geralmente, muito fáceis de medir. De qualquer forma, é necessário utilizar um gel electrolítico que garanta um bom contacto eléctrico entre os eléctrodos e a pele e o sinal será medido com melhor qualidade quando a pele se encontra limpa.
Embora actualmente sejam já correntemente usados equipamentos de ECG digitais20, na prática clínica continuam a ser amplamente utilizados os aparelhos com registo em papel. Apesar destes últimos não possibilitarem uma avaliação quantitativa do sinal rigorosa, geralmente, para um clínico experiente, a sua análise visual estudo das amplitudes relativas dos picos, a sua relação temporal e morfologia das ondas é suficiente para detectar anomalias ao nível do funcionamento cardíaco.
20 No capítulo referente ao EEG foram já feitos alguns comentários sobre as características dos sinais registados com equipamentos digitais que são igualmente válidos para os equipamentos de ECG.