Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DO CEARÁ
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
FACULDADE DE VETERINÁRIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
FRANCISCO RENAN ARAGÃO LINHARES
OTIMIZAÇÃO DA CRIOPRESERVAÇÃO SEMINAL E FERTILIZAÇÃO
ASSISTIDA EM Cyprinus carpio (ACTINOPTERYGII) NO SEMIÁRIDO
BRASILEIRO
FORTALEZA - CEARÁ
2016

FRANCISCO RENAN ARAGÃO LINHARES
OTIMIZAÇÃO DA CRIOPRESERVAÇÃO SEMINAL E FERTILIZAÇÃO ASSISTIDA
EM Cyprinus carpio (ACTINOPTERYGII) NO SEMIÁRIDO BRASILEIRO
Tese apresentada ao Curso de Doutorado em
Reprodução e Sanidade Animal do Programa
de Pós-Graduação em Ciências Veterinárias da
Faculdade de Veterinária da Universidade
Estadual do Ceará, como requisito parcial à
obtenção do título de doutor em Ciências
Veterinárias. Área de Concentração:
Reprodução e Sanidade Animal.
Orientador: Prof.a Dr.a Carminda Sandra Brito
Salmito Vanderley
FORTALEZA - CEARÁ
2016



Dedico à minha avó materna, Francisca
Aragão, por ter contribuído diretamente nesta
conquista.

AGRADECIMENTOS
A Deus por toda a oportunidade que me foi concedida durante minha vida.
Ao Programa de Pós-Graduação em Ciências Veterinárias - PPGCV, da Faculdade de
Veterinária, da Universidade Estadual do Ceará, pela oportunidade de fazer parte do corpo
discente.
Ao Núcleo Integrado de Biotecnologia (NIB) e Laboratório de Biotecnologia da Reprodução
de Peixes (LBRP) da Universidade Estadual do Ceará (UECE) que através do uso de suas
instalações e equipamentos, tornou possível a conclusão do experimento.
Ao Departamento Nacional de Obras Contra as Secas (DNOCS), pelo apoio de laboratórios e
estruturas de pesquisa para realização do experimento.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pela bolsa de
estudo durante o período de doutorado.
À minha avó materna Francisca Aragão que apesar das condições difíceis naquela época,
sempre priorizou a educação de todos os filhos e netos e por fazer parte de minha educação
em todos os sentidos lá na minha infância.
À família Aragão Linhares pelos exemplos de determinação para cumprir com eficiência
minhas tarefas diárias.
À minha esposa Liliane Chagas Pascoal pela força de vontade para atingir os objetivos
traçados e aguentar meus estresses de cada dia.
À família Chagas Pascoal por todo o acolhimento e apoio desde o momento em que adentrei
na família.
Ao Professor Dr. José Ferreira Nunes, por toda a orientação que me foi passada e confiança
de fazer parte de sua equipe do Núcleo Integrado de Biotecnologia desde o início de minha
vida acadêmica.
À Dra. Cristiane Clemente de Mello Salgueiro, pelo meu acompanhamento durante o início de
minha iniciação científica e pela sua disponibilidade em sempre estar me ajudando nas horas
que mais preciso.

À Dra. Carminda Sandra Brito Salmito Vanderley por ter acreditado no meu trabalho desde o
início da graduação e por sua orientação.
À Dra. Maria Audália Marques de Carvalho pelo apoio como coorientadora durante o período
de desenvolvimento da pesquisa, contribuição intelectual e membro avaliador da tese.
À Dra. Caroline Vieira Feitosa pela contribuição intelectual como membro avaliador da
referida Tese.
À Dra. Mônica Aline pela contribuição intelectual como membro avaliador da tese.
À Dra. Liliane Veras pelo apoio intelectual durante o período de desenvolvimento da pesquisa
e apoio de suas experiências em campo.
Ao Dr. Francisco José Lopes Cajado pela contribuição intelectual como membro avaliador da
referida tese.
Ao Dr. Cláudio Cabral e ao Me. Assis Montenegro pela realização da estatística.
À José Agenor Galvão pelos ensinamentos práticos durante a realização dos experimentos e
pelo total apoio a minha pesquisa científica no DNOCS.
Aos pós-graduandos Júlia Trugílio Lopes, Larissa Teixeira Nunes, Mayara Setúbal Oliveira,
Jordana Leite Sampaio, Priscila Silva de Almeida Monteiro, João Paulo Silva Pinheiro,
Renata Vieira do Nascimento e Vanessa Alves Pereira por todo o apoio fornecido aos meus
experimentos em Fortaleza e Pentecoste.
Aos alunos de iniciação científica, Rômulo Roberto Ribeiro Pinheiro, Thais Maia Torres,
Yasmim Maia Ferreira, Edna Raquel Paiva Teodozia, Cibele Castro Monteiro, Eric Silva
Antério, Filipe Oliveira Ferreira, Karen Emanuelly Pinheiro Gomes, Jéssica Uchôa Pinheiro,
Francisco Yan Tavares Reis, José Pedro Vieira Arruda Júnior, Maria Mayara Aguiar Lima,
Sayansk Queiroz da Silva, Vanessa Sayonara de Farias Monteiro e José de Souza Junior pela
disponibilidade e disposição para cumprir com eficiência suas tarefas.
Aos integrantes do Núcleo Integrado de Biotecnologia, em especial a Iraci Clemente de Melo
e Henna Roberta Quito.

RESUMO
A carpa comum, introduzida no nordeste do Brasil pelo Departamento Nacional de Obras
Contras às Secas (DNOCS) no ano de 1977, é uma das espécies mais cultivadas em todo o
mundo. Por representar uma espécie com grande valor comercial, os objetivos desse trabalho
foram: (1) avaliar o efeito da suplementação das vitaminas C (ácido ascórbico) ou E (α-
tocoferol) ao meio ACP-104 sobre a qualidade do sêmen pós-descongelado da carpa comum;
(2) definir a melhor proporção do número de espermatozoides por ovócito (dose inseminante)
utilizando sêmen in natura da espécie. Para o experimento 1, foram formados oito pools a
partir da coleta de sêmen de 15 machos. As amostras seminais coletadas foram avaliadas
quanto à motilidade total, à velocidade, ao percentual de espermatozoides normais, à
vitalidade espermática e a duração da motilidade antes e depois da criopreservação seminal.
Esta foi realizada em meio ACP-104 acrescido de dimetilsulfóxido (DMSO) ou etilenoglicol
(EG), suplementados ou não com vitamina C ou E. As amostras foram então congeladas em
vapor de nitrogênio líquido em dry shipper e estocadas em nitrogênio líquido (-196 °C) até a
descongelação e análises. O experimento 2, utilizou-se de um pool de sêmen de 5 machos e
um pool de ova de 2 fêmeas. Os gametas coletados foram então fertilizados nas seguintes
proporções de espermatozoides/ovócito: D1-15.152, D2-60.491, D3-179.584, D4-238.072 e
D5-298.563. Além disso, foi registrado em imagens o desenvolvimento embrionário da carpa
comum. No que diz respeito aos resultados do experimento 1, foi observado que o EG
suplementado com vitamina E produziu resultado significativamente superior de motilidade
total, normalidade espermática e duração da motilidade em relação ao DMSO, concluindo-se
que o EG deve ser, portanto, o crioprotetor de escolha a ser utilizado com o ACP-104
suplementado ou não com vitamina E. Para o experimento 2, verificou-se que a porcentagem
de fertilização aumentou de forma linear, atingindo um platô em 45,5% na proporção de
208.295 espermatozoides/ovócito. Assim, recomenda-se o uso da dose inseminante de
aproximadamente 200.000 espermatozoides/ovócito na rotina de fertilização assistida dessa
espécie no nordeste brasileiro. Além disso, o desenvolvimento embrionário da carpa em
latitude equatorial segue a cronologia semelhante ao relatado para a espécie em clima
temperado.
Palavras-chave: Criopreservação. Antioxidantes. Espermatozoides/ovócito. Embriologia.

ABSTRACT
The common carp, introduced in northeastern Brazil by the National Department of Works
Cons Drought (DNOCS) in 1977, is one of the most cultivated species in the world. To
represent a species with high commercial value, the objectives of this study were: (1) evaluate
the effect of supplementation of vitamins C (ascorbic acid) or vitamin E (α-tocoferol) with
ACP-104 medium on the quality of post-thawed of common carp; (2) determine the best ratio
of the number of sperm per oocyte using semen in natura. For the experiment 1, semen pools
from 15 selected males were formed. Seminal samples were evaluated for total motility,
velocity, percentage of normal sperm, sperm vitality and duration of total motility before and
after the seminal cryopreservation. This was performed in extenders ACP-104 plus DMSO or
EG supplemented or not with vitamin C or E. The samples were frozen in vapors of nitrogen
into dry shipper and stored in liquid nitrogen (-196 °C) until thawing and analysis. The
experiment 2, semen pool from 5 selected males and eggs pool from 2 selected female were
formed. Gametes collected were then fertilized in the following proportions of sperm/oocyte
D1-15.152, D2-60.491, D3-179.584, D4-238.072 and D5-298.563. Moreover, monitoring of
embryonic development was done. For the results of experiment 1, EG supplemented with
vitamin E, produced significantly better results overall motility, sperm normality and duration
of motility relative to DMSO, it concluded that the EG should be the cryoprotectant of choice
for use with the ACP-104 supplemented or not with vitamin E. For the experiment 2, it was
found that the fertilization rate increased linearly up to the proportion of 208.295
spermatozoa/oocyte, and, from this point, the fertilization rate was maintained at 45,5%.
Therefore, we recommend the use of the insemination dose of approximately 200.000
sperm/oocyte in the artificial fertilization routine of common carp in brazilian northeast.
Furthermore, embryonic development of carp in equatorial latitude follows the chronology
similar to that reported for the species in temperate zones.
Key-words: Cryopreservation. Antioxidants. Epermatozoa/oocyte. Embryology.

LISTA DE ILUSTRAÇÕES
CAPÍTULO 1
Figura 1- Estágios do desenvolvimento embrionário de carpa comum (Cyprinus carpio).
(A) formação de mórula, (B) formação de blástula e (C) gástrula....................32
CAPÍTULO 4
Figura 1- Taxas de fertilização assistida de ovócitos de carpa comum (Cyprinus carpio)
fertilizados artificialmente com diferentes relações espermatozoides/ovócito.
Para x<208.295, y=1,6*10-4+11,7559. Para x>208.295, y=45,49
(R2=93,30).........................................................................................................73
Figura 2- Estágios do desenvolvimento embrionário de carpa comum (Cyprinus carpio).
Primeiras segmentações (A e B), formação de mórula (C), formação de blástula
(D), gástrula inicial (E), 50% de gastrulação (F), 80% de gastrulação (G), 90%
de gastrulação (H), fechamento do blastóporo (I), diferenciação do embrião, da
vesícula ótica (vo) e formação dos somitos (so) (J), região caudal destacada do
vitelo (K), embrião com movimentação dentro da membrana coriônica (L),
eclosão (M). Aumento de 40x...........................................................................75

LISTA DE TABELAS
REVISÃO DE LITERATURA
Tabela 1 - Taxas de fertilização e eclosão (%) após a realização de fertilização artificial
utilizando diferentes doses inseminantes com sêmen descongelado de algumas
espécies de ciprinídeos................................................................................. .....21
CAPÍTULO 1
Tabela 1 - Médias dos valores físicos, químicos e microbiológicos da água do açude
Pereira de Miranda nas unidades experimentais durante o experimento.
Pentecoste, Ceará...............................................................................................32
CAPÍTULO 2
Tabela 1 - Taxas de fertilização e eclosão (%) após a realização de fertilização artificial
utilizando diferentes doses inseminantes com sêmen descongelado de algumas
espécies de ciprinídeos......................................................................................46
CAPÍTULO 3
Tabela 1 - Efeito de diferentes tipos de crioprotetores sobre os parâmetros cinéticos de
espermatozoides frescos (grupo controle) e descongelados de Cyprinus carpio
criopreservados em ACP-104 sem vitaminas....................................................58
Tabela 2 - Efeito de diferentes tipos de crioprotetores sobre o percentual de
espermatozoides normais e vitalidade de espermatozoides frescos (grupo
controle) e descongelados de Cyprinus carpio criopreservados em ACP-104
sem vitaminas....................................................................................................59
Tabela 3- Efeito de diferentes tipos de crioprotetores acrescidos de vitaminas sobre os
parâmetros de motilidade total (%), espermatozoides morfologicamente
normais (%) e vitalidade espermática (%) de espermatozoides frescos (grupo

controle) e descongelados de Cyprinus carpio criopreservados em ACP-
104.....................................................................................................................60
Tabela 4 - Efeito de diferentes tipos de crioprotetores acrescidos de vitaminas sobre a
duração da motilidade (segundos) de espermatozoides frescos (grupo controle)
e descongelados de Cyprinus carpio criopreservados em ACP-
104.....................................................................................................................60
CAPÍTULO 4
Tabela 1 - Taxa de fertilização e eclosão (%) de ovos de carpa comum (Cyprinus carpio)
fertilizados com diferentes proporções de espermatozoides (sptzs) por
ovócito...............................................................................................................72

LISTA DE ABREVIATURAS
ALH Deslocamento lateral da cabeça
ACP Água de coco em pó
BCF Frequência de batimento cruzado
CASA Análise espermática auxiliada por computador
CEUA Comitê de Ética para o Uso de Animais
CPAq Centro de Pesquisas em Aquicultura
DMSO Dimetilsulfóxido
DNOCS Departamento Nacional de Obras Contra às Secas
EG Etilenoglicol
EHC Extrato hipofisário de carpa
LBRP Laboratório de Biotecnologia da Reprodução de Peixes
mOsm Miliosmole
NIB Núcleo Integrado de Biotecnologia
SCA Sperm Class Analyser
sptz Espermatozoide
STR Retilinearidade
LIN Linearidade
UECE Universidade Estadual do Ceará VAP – Velocidade média do percurso
VCL Velocidade do percurso curvilinear
VSL Velocidade em linha reta
WOB Oscilação

SUMÁRIO
1 INTRODUÇÃO...........................................................................................................13
2 REVISÃO DE LITERATURA..................................................................................13
2.1 ESPÉCIE.......................................................................................................................13
2.2 TAXONOMIA..............................................................................................................14
2.3 REPRODUÇÃO............................................................................................................14
2.4 CRIOPRESERVAÇÃO................................................................................................15
2.4.1 Diluição seminal e armazenamento do sêmen..........................................................15
2.4.2 Adição de vitaminas C ou E aos meios de congelação.............................................17
2.5 AVALIAÇÃO DO SÊMEN IN NATURA E PÓS-DESCONGELADO.......................19
2.6 FERTILIZAÇÃO ASSISTIDA ARTIFICIAL..............................................................20
3 JUSTIFICATIVA........................................................................................................23
4 HIPÓTESE CIENTÍFICA.........................................................................................24
5 OBJETIVOS................................................................................................................25
5.1 OBJETIVO GERAL.....................................................................................................25
5.2 OBJETIVOS ESPECÍFICOS........................................................................................25
6 CAPÍTULO 1...............................................................................................................26
7 CAPÍTULO 2...............................................................................................................36
8 CAPÍTULO 3...............................................................................................................51
9 CAPÍTULO 4...............................................................................................................67
10 CONCLUSÕES...........................................................................................................79
11 PERSPECTIVAS........................................................................................................80
12 REFERÊNCIA BIBLIOGRÁFICA...........................................................................81

13
1 INTRODUÇÃO
O brasileiro Rodolfo von Ihering (1883-1939), diretor da Comissão Técnica de
Piscicultura do Nordeste e pioneiro em fecundação artificial de peixes por meio da descoberta
da inseminação artificial de peixes, colocou o Brasil em nível mundial em piscicultura.
Porém, constatando a pobreza qualitativa e quantitativa da ictiofauna do semiárido nordestino,
representada por apenas dez espécies de valor comercial, o brasileiro e a sua equipe passaram
a estudar inúmeras espécies de outras bacias hidrográficas, a partir de 1935, com o objetivo de
serem introduzidas e aclimatadas na região (DNOCS, 2009). Dentre as espécies, destaca-se a
carpa comum (Cyprinus carpio vr hungaricus), trazida da Hungria em 1986. Este peixe é
considerado de alto valor comercial, forte apelo culinário e esportivo, sendo uma das espécies
mais cultivadas em todo o mundo, representando 6,14% do total da produção mundial
aquícola (FAO, 2008) e 76,7% da produção de pescado do Brasil (ECHEVENGUÁ, 2006).
Nesse contexto, o desenvolvimento de biotécnicas como a criopreservação gamética
vem contribuindo para o crescimento da piscicultura mundialmente. A criopreservação
espermática destaca-se como uma ferramenta essencial para a reprodução artificial,
possibilitando a preservação de espécies de peixes ameaçadas de extinção (VIVEIROS et al.,
2012a), além da renovação e do aumento da variabilidade genética de peixes comerciais.
Estudos recentes realizados pela equipe do Laboratório de Biotecnologia da
Reprodução de Peixes pertencente à Universidade Estadual do Ceará obtiveram resultados
próximos e satisfatórios aos parâmetros seminais encontrados na literatura para carpa comum
(LINHARES et al., 2015). No entanto, o protocolo de criopreservação seminal desenvolvido
para a espécie citada acima utilizando o diluente ACP-104 acrescido de diferentes
crioprotetores e suplementado com vitaminas C ou E ainda não foi empregado. Além disso, a
literatura também não relata a caracterização do desenvolvimento embrionário e a
determinação da dose inseminante da espécie no nordeste brasileiro.
Dessa forma, a realização destes procedimentos contribuirá para a introdução de novas
linhagens de carpa comum nos açudes públicos e privados utilizando um protocolo
padronizado de criopreservação seminal e fertilização artificial, fortalecendo a piscicultura
comercial no estado do Ceará.
2 REVISÃO DE LITERATURA
2.1 ESPÉCIE

14
A carpa comum, nativa do litoral do mar Cáspio e Aral, é considerada uma das
espécies de peixes de água doce domesticadas mais antigas, cultivadas há mais de 2000 anos
na China (BALON, 1995). Devido a sua tolerância e resistência a uma ampla variedade de
habitats aquáticos, estes animais se espalharam por todo o mundo (BRITTON et al., 2007).
São animais que vivem em grupo, apresentando hábito alimentar onívoro, com tendência à
iliofagia (FROESE; PAULY, 2007). É uma espécie típica de ambientes lênticos, capaz de
suportar baixos níveis de oxigênio dissolvido (3,2 mg/L) (MACÊDO et al., 2007). A
temperatura ideal para seu bom desenvolvimento varia de 24 a 28 °C, porém, apresenta uma
faixa tolerável de 8 a 30 °C, sendo assim, um peixe rústico e de fácil adaptação (GRAEFF;
PRUNER, 1999). As carpas geralmente apresentam um comprimento variando entre 30 e 60
cm e pesam entre 0,5 e 4 kg (TOMELLERI; EBERLE, 1990).
2.2 TAXONOMIA
Reino: Animalia
Filo: Chordata
Classe: Actinopterygii
Ordem: Cypriniformes
Família: Cyprinidae
Gênero: Cyprinus
Espécie: Cyprinus carpio
2.3 REPRODUÇÃO
O ciclo reprodutivo dos peixes teleósteos depende de vários fatores dentre eles o
fotoperíodo, temperatura, salinidade, pH, tubidez e precipitação pluviométrica (BROMAGE
et al., 2001; GLASSER et al., 2004). Em regiões tropicais e subtropicais as carpas geralmente
atingem a maturação sexual durante o primeiro ano de vida e pode desovar várias vezes
dentro de um determinado ano (SIVAKUMARAN et al., 2003). Kucharczyk et al. (2008)
reforçam que é possível produzir alevinos de carpa comum durante todo o ano, sem a
diminuição da qualidade dos gametas e alevinos. Segundo Moreira et al. (2001), a
temperatura ótima da água para se realizar a reprodução induzida das carpas comuns, situa-se
entre 22 a 28ºC.

15
No ambiente natural, as fêmeas desovam em águas paradas com a presença de
macrófitas, liberando cerca de 100 a 230 g de ovócitos por quilograma de peso corporal, uma
média de 100.000 a 300.000 ovócitos.Kg-1 (LINHART et al., 1995). Sem o cuidado parental,
os ovócitos são colocados sobre plantas aquáticas que, ao entrarem em contato com a água,
adquirem adesividade e aumentam de três a quatro vezes seu volume. A fertilização ocorre
externamente e ao final do desenvolvimento embrionário, os ovos eclodem. Os alevinos
recém-eclodidos tornam-se capazes de se alimentar de minúsculos animais aquáticos após o
desaparecimento do saco vitelínico (WOYNAROVICH, 1993).
2.4 CRIOPRESERVAÇÃO
Desde que Blaxter (1953) realizou os primeiros estudos de criopreservação de gametas
de peixes, muitas pesquisas têm sido voltadas para a criopreservação do sêmen de várias
espécies de pescado, incluindo a carpa comum (Cyprinus carpio). A introdução desta
biotécnica na reprodução animal tem sido de grande importância para a aquicultura,
contribuindo para aumentar a propagação de material genético de alto valor em menor espaço
de tempo.
A execução de um protocolo de criopreservação de sêmen incluem ajustes de taxas de
diluição, escolhas de diluentes e diluidores, tempo de equilíbrio entre o crioprotetor e os
espermatozoides, método de envase adequado, taxas de resfriamento e descongelamento das
amostras e relação entre espermatozoides:ovócitos durante a realização de testes de
fertilização. A presença de várias etapas dificulta a padronização de protocolos de congelação
seminal devido às diferentes interações espécie-específicas do volume, composição seminal e
suas características bioquímicas em cada etapa do processo (YANG; TIERSCH, 2009). Neste
contexto, abordaremos as variações das etapas que abrange o processo de crioprervação
seminal de carpa comum.
2.4.1 Diluição seminal e armazenamento do sêmen
Os espermatozoides, quando submetidos às alterações rigorosas de temperaturas,
estão sujeitos aos danos ocasionados pelo frio, denominados crionjúrias (SILVA; GUERRA,
2011). Estes danos podem levar a um decréscimo nas características espermáticas, como a
velocidade, motilidade, capacidade de fertilização e subsequente redução das taxas de eclosão
(LI et al., 2010). Dessa forma, após a coleta, o sêmen é submetido à etapa de diluição

16
resultante da adição de soluções protetoras capazes de minimizar os danos causados pelas
crioinjúrias. Estas soluções são chamadas de crioprotetores, atuando na diminuição do ponto
de congelação da água, reduzindo o estresse osmótico e os efeitos da cristalização sobre a
estrutura celular (SALMITO-VANDERLEY et al., 2012).
Devido à especificidade própria de cada grupo animal, torna-se fundamental a
adequação da composição do diluidor as características seminais de cada espécie. Na
literatura são relatadas soluções contendo apenas sais, apenas açucares ou uma combinação
dos mesmos para a criopreservação seminal de peixes. A solução de Alsever, cuja composição
é formada por NaCl e citrato de sódio, é considerado um dos diluidores mais conhecidos
utilizado em ciprinídeos (NAHIDUZZAMAN et al., 2012). Além disso, muitos autores
também utilizam um diluidor descrito por Kurokura et al. (1984) resultantes da combinação
dos sais NaCl, KCl, CaCl2 e NaHCO3 (AKÇAY et al., 2004; RANI; MUNUSWAMY, 2014;
YAVAS et al. 2014). Outro tipo de diluidor utilizado em ciprinídeos é a glicose (HORVÁTH
et al., 2007). Esta age como substrato de energia, componente osmótico e agente crioprotetor
em função do seu alto peso molecular, contribuindo para o equilíbrio osmótico e atuando
como substituto de eletrólitos (HOLT, 2000). Em geral, os diluidores acrescidos de açucares
são capazes de melhorar a motilidade espermática durante a criopreservação seminal de
algumas espécies de ciprinídeos (URBÁNYI et al., 2006). A combinação de sais (NaCl, KCl)
e açucares (frutose, glicose e sacarose) podem ser utilizados da mesma forma como diluidores
de sêmen de carpa comum (IRAWAN et al., 2010).
Outra alternativa, é a utilização do diluidor água de coco em pó (ACP), considerada
uma das inovações mais recentes da biotecnologia do sêmen de peixes desenvolvido por
pesquisadores da Universidade Estadual do Ceará-Brasil. O ACP é composto por sais,
proteínas, açúcares, vitaminas, gorduras neutras, além de indutores da divisão celular e
diversos eletrólitos, proporcionando meio nutritivo completo para os espermatozoides.
Estudos com a água de coco já foram realizados em várias espécies como caprinos (NUNES,
1995), ovinos (ARAÚJO, 1990) e suínos (TONIOLLI; MESQUITA, 1990). Estudos iniciais
em peixes mostraram resultados satisfatórios na combinação do ACP com os crioprotetores
DMSO (VELASQUEZ-MEDINA, 2008; VIEIRA, 2011) e metilglicol (VIVEIROS et al.,
2008), podendo ser utilizado também como diluidor para a criopreservação seminal de carpa
comum (LINHARES et al., 2015).
Os diluentes seminais também atuam como carreadores de crioprotetores, agindo em
conjunto para proteger os espermatozoides contra os efeitos deletérios do congelamento. Os
crioprotetores podem ser classificados como internos ou externos. Os crioprotetores internos

17
atuam na redução do ponto crioscópio da água intracelular evitando a formação de cristais de
gelo e o comprometimento da integridade da membrana espermática interna durante as
mudanças de temperatura.
Os crioprotetores internos mais comumente utilizados em ciprinídeos são
dimetilsulfóxido (DMSO), metanol, propilenoglicol, dimetilacetamida (DMA), glicerol e
etanol (SULTANA et al., 2010; IRAWAN et al., 2010; AKÇAY et al., 2004). Na maioria dos
estudos, o DMSO fornece os melhores resultados, provavelmente devido a sua rápida
penetração em espermatozoides e sua interação com os fosfolipídios da membrana
espermática (SUQUET et al., 2000). Os crioprotetores externos atuam na estabilização da
membrana plasmática e restauração dos fosfolipídeos perdidos pelos espermatozoides durante
o congelamento. No entanto, estes não são utilizados frequentemente em ciprinídeos, sendo
relatadas na literatura à adição de gema de ovo-citrato ou somente gema de ovo as soluções
diluidoras de sêmen de carpa comum (SULTANA et al., 2010; ÖĞRETMEN et al., 2014;
YAVAS et al. 2014).
Outro ponto a ser levado em consideração é o ajuste das concentrações dos
crioprotetores antes de serem misturados aos diluentes, pois os mesmos são tóxicos as células
em temperatura ambiente. Comumente as concentrações dos crioprotetores são ajustados a 5
ou 10%, pois as pesquisas apontam que em altas concentrações alguns crioprotetores pode ser
prejudicial à motilidade espermática (RANI; MUNUSWAMY, 2014).
Após a diluição o sêmen estará pronto para o armazenamento. Os equipamentos mais
utilizados para congelar as amostras são as caixas térmicas de poliestireno, refrigeradores
programáveis, dry shippers e botijões criogênicos (TAITSON et al., 2007; IRAWAN, et al.,
2010; NAHIDUZZAMAN et al., 2012). Após o equilíbrio térmico, o sêmen congelado é
transferido para um botijão de nitrogênio líquido, onde pode ser conservado por longos
períodos à -196 °C sem efeitos deletérios (ASHWOOD-SMITH, 1980).
2.4.2 Adição de vitaminas C ou E aos meios de congelação
O processo de congelamento e descongelamento pode ocasionar danos a membrana
plasmática e outras organelas celulares devido ao estresse oxidativo, choque térmico e
formação de cristais de gelo intracelulares (GUTHRIE; WELCH, 2005). O estresse oxidativo
é ocasionado pela geração excessiva de espécies reativas do metabolismo do oxigênio (ROS)
(MAIA; BICUDO, 2009). As ROS (O2-, OH•, HO2
• e H2O2) são intermediários químicos

18
reativos continuamente produzidas pelo metabolismo aeróbio celular que podem comprometer
a viabilidade e a função espermática (SŁOWIŃSKA et al., 2013).
Para combater a produção excessiva de ROS e o estresse oxidativo, o sêmen apresenta
um sistema de defesa antioxidante intra e extracelular formado por componentes enzimáticos
e não enzimáticos (SANOCKA; KURPISZ, 2004). Os antioxidantes enzimáticos são
representados pelo superóxido dismutase (SOD), catalase (CAT) e sistema glutatião
peroxidase/glutatião redutase (GSH-Px/GSH-Red), enquanto os não-enzimáticos são α-
tocoferol, ascorbato, urato, piruvato, glutatião, taurina e hipotaurina (SILVA; GUERRA,
2012).
No entanto, o sistema intracelular de defesa antioxidante do espermatozoide é mais
susceptível a ação das ROS, devido à escassez de citoplasma que é perdido durante a
maturação celular (CARVALHO et al., 2002). Além disso, Stradaioli et al. (2007) observaram
que a crioproservação reduz os níveis dos antioxidantes presentes tanto na célula quanto no
plasma seminal, fortalecendo a evidência de que, entre as causas da deterioração da qualidade
do sêmen após um ciclo de congelação e descongelação, estão aquelas ligadas ao estresse
oxidativo.
Dessa forma, pesquisas relacionadas à adição de antioxidantes aos diluidores seminais
de peixes têm sido realizadas a fim de melhorar a qualidade do sêmen criopreservado por
meio da prevenção ou redução do processo peroxidativo. Neste contexto, podem ser
destacados entre os antioxidantes já estudados com esta finalidade a vitamina E e a vitamina
C (NAVARRO et al., 2014; MARTINEZ-PÁRAMO et al., 2012).
A vitamina E (α-tocoferol e derivados) é um antioxidante não enzimático
(MANEESH; JAYALEKSHMI, 2006), encontrada em quantidades baixas em membranas
celulares de mamíferos e no plasma seminal que pode proteger os espermatozoides contra
danos oxidativos do DNA e da membrana (SIKKA, 2004). Este antioxidante é capaz prevenir
a peroxidação lipídica e suprimir, por conseguinte, a produção de malonaldeido (AITKEN;
CLARKSON, 1988), o que é possível pelo fato de atuar como quelante das ROS produzidas
durante a lipoperoxidação (FERREIRA; MATSUBARA, 1997).
O ácido ascórbico ou vitamina C atua neutralizando as ROS e RNS por meio de
reações de redução (BARREIROS et al., 2006). Dessa forma, a inibição da peroxidação
lipídica promovida por este antioxidante ocorre sobre as membranas celulares, atuando
eficientemente sobre os radicais O2˙˙, H2O2, ClO˙, OH˙, OOH˙ (VASCONCELOS et al.,
2007). A vitamina C também atua sinergicamente com a vitamina E, através da geração de

19
tocoferol, reduzindo a produção de ROS e protegendo o espermatozoide contra danos no
DNA (GUERRA et al., 2004).
2.5 AVALIAÇÃO DO SÊMEN IN NATURA E PÓS-DESCONGELADO
O método ideal para predizer a fertilidade dos machos reprodutores é pelo exame das
características seminais à campo. Portanto, a qualidade espermática torna-se um pré-requisito
para a seleção dos animais em programas de reprodução artificial. A avaliação do ejaculado
deve ser rápida e eficiente, preservando a qualidade inicial e a capacidade fertilizante.
Amostras seminais são avaliadas tendo em vista várias características físicas como o aspecto,
a cor, o volume, o pH, a osmolaridade, a concentração, a morfologia e a motilidade
espermática. A partir desses resultados qualquer desvio para os padrões de uma determinada
espécie pode ser correlacionado com a fertilidade (HAFEZ, 1995).
O aspecto seminal pode ser facilmente classificado de forma subjetiva, podendo ser
classificado em viscoso, líquido e indeterminado (SUQUET et al., 1992), enquanto o pH do
sêmen de peixes de água doce varia de 6,5 a 8,5 e a osmolaridade em torno de 300 mOsm/Kg
(TABARES et al., 2005; MORISAWA; SUSUKI, 1980).
A concentração espermática é uma das medidas quantitativas mais importantes
utilizadas na pesquisa e rotina de avaliação do sêmen de peixes (SILVEIRA et al., 1987). A
determinação da concentração espermática contribui para a padronização da taxa de diluição
sêmen:diluidor apropriada para a criopreservação seminal e posterior utilização em programas
de fertilização assistida.
A qualidade espermática está diretamente relacionada com a motilidade dos gametas
masculinos, uma vez que esta característica constitui um pré-requisito para a fertilização dos
ovócitos (RURANGWA et al., 2004). A técnica convencional, baseada na observação através
de microscópio óptico, é utilizada pela maioria dos laboratórios para realizar análise seminal.
Embora sujeita a distintos graus de imprecisão, devido à subjetividade da avaliação a partir de
escalas arbitrárias (CHAMBEYRON; ZOHAR, 1990), este método é tão confiável quanto à
análise objetiva desde que realizada por um técnico treinado.
O sistema de análise seminal auxiliado por computador consiste basicamente de um
microscópio de contraste de fase acoplado a uma câmera de vídeo. As imagens geradas são
reproduzidas e analisadas de forma eficiente e objetiva por meio do software Sperm Class
Analyser – SCA instalado no sistema operacional de um computador adaptado a estes
equipamentos. A automatização desta análise permite maior objetividade e rapidez, uma vez

20
que é realizada numa menor fração do tempo requerido pela avaliação subjetiva. A utilização
deste equipamento contribuiu para a produção de trabalhos mais detalhados sobre a atividade
dos espermatozoides, avaliando os parâmetros de rápidos, médios, lentos, estáticos,
velocidade do percurso curvilinear (VCL), velocidade do percurso médio (VAP), velocidade
em linha reta (VSL), retilinearidade (STR), linearidade (LIN), oscilação (WOB), frequência
de batimento cruzado (BCF), e deslocamento lateral da cabeça (ALH). Os parâmetros de
velocidade, VCL, VAP e VSL, descrevem o movimento do espermatozoide, enquanto, os
parâmetros LIN, STR e WOB representam as relações entre estas velocidades VSL e VCL;
VSL e VAP; VAP e VCL, respectivamente.
2.6 FERTILIZAÇÃO ASSISTIDA ARTIFICIAL
O sucesso do protocolo de criopreservação é verificado através da descongelação do
sêmen em banho de imersão térmica e avaliação da qualidade espermática. A fertilização
artificial é a etapa final para a análise da qualidade dos gametas, sendo considerado um dos
indicadores mais seguros e eficientes para se comprovar a condição do material
criopreservado. A biotécnica pode ser realizada por dois métodos diferentes: o método úmido
que se baseia na mistura de gametas masculinos e femininos coletados simultaneamente com
água (SULTANA et al., 2010) e o método a seco que mistura primeiramente os gametas
acrescentando-se água em seguida (AKÇAY et al., 2004).
A reprodução artificial em carpa é mais complexa porque o procedimento de
inseminação requer uma solução específica para evitar a aderência entre os ovócitos que
ocorre normalmente ao entrarem ao contato com a água. A aderência é oriunda do envelope
vitelino que reveste os ovócitos e é formado por carboidratos desidratados (LINHART et al.,
1995). Essas camadas acelulares envoltórias servem para proteção, nutrição e barreira pela
qual os espermatozoides devem atravessar (MONTANARI, 2013). Entretanto, essa mesma
aderência é responsável pela formação de uma massa de ova que dificulta o encontro da
micrópila pelos espermatozoides, prejudicando o processo de fertilização.
A solução comumente utilizada pode ser formulada de duas formas conforme a
quantidade de ureia adicionada. Uma delas é composta por 3 g de ureia e a outra por 20 g de
ureia juntamente com 4 g de NaCl para cada litro de água. Essas soluções atuam na ativação
dos espermatozoides e remoção da adesividade dos ovócitos durante agitação contínua por 1
hora a 1 hora e 30 minutos. Após esse período adiciona-se uma solução de ácido tânico
(0,5g/L) por 20 segundos, removendo-a em seguida (BILLARD et al., 1995). Vale ressaltar

21
que muitos autores relatam a utilização da solução contendo 3% de ureia (IRAWAN et al.,
2010; HORVÁTH et al., 2007). No entanto, Billard et al. (1995) afirmam que a solução
contendo 20% de ureia é uma solução melhorada, necessitando de menos tempo de agitação
para a remoção da adesividade dos ovócitos. Além disso, pode-se utilizar também o leite em
pó composto por 27,2% de gordura, 26,6g de albumina, 37,2g de açúcar do leite, 5,8g de íons,
0,2g de lecitina e 3g de água em 100g do mesmo para a preparação de uma solução de 40g de
leite em pó por litro de água declorada ou limpa. A solução de leite em pó deve ser adicionada
lentamente misturando-se um volume de 1L da solução por Kg de ovócitos. Após 60 minutos,
adiciona-se lentamente mais água durante 10 minutos para substituição da solução de leite e
em seguida ovócitos podem ser transferidos para as incubadoras (LINHART et al., 2003).
Durante os testes de fertilização necessita-se definir a dose inseminante ideal, ou seja,
a proporção mínima de espermatozoides por ovócito capaz de produzir a máxima
sobrevivência em termos de larvas eclodidas. Esta técnica conserva e economiza gametas,
melhora a utilização do macho e contribui para um aumento da variabilidade genética. O
sucesso de uma fertilização artificial utilizando uma dose inseminante é determinado por meio
da taxa de fertilização, ou seja, a partir de uma coleta aproximada de 150 embriões contam-se
aqueles em estágio de gástrula e divide-se pelo número de ovócitos fertilizados (HORVÁTH
et al., 2003). Da mesma forma, também é calculada a taxa de eclosão por meio da contagem
do número de larvas eclodidas dividida pelo número total de ovócitos fertilizados. A definição
da dose inseminante utilizando sêmen criopreservado de ciprinídeos já foi estabelecida por
alguns autores obtendo diferentes resultados de taxa de fertilização e eclosão (Quadro 1).
Tabela 1. Taxas de fertilização e eclosão (%) após a realização de fertilização artificial utilizando
diferentes doses inseminantes com sêmen descongelado de algumas espécies de ciprinídeos. Espécies Meios de congelação Dose
inseminante
(sptzs:ovócito)
Taxa de
fertilização
(%)
Taxa de
eclosão
(%)
Referências
Diluidor Diluente
Carpa
comum
(Cyprinus
carpio)
Glicose Metanol 10% 2,6-3,4x104:1 74 ± 15 67 ± 17 Horváth et al.
(2003)
Carpa
comum
(Cyprinus
carpio)
Kurokura +
própolis
Dimetilsulfóxido
10% + gema de
ovo 10%
1,0 x105:1 95,0 ± 0,6 60,3 ± 2,0 Öğretmen et
al. (2014)
Carpa
comum
(Cyprinus
carpio)
Kurokura +
trealose
Dimetilacetamida
20%
1,0 x105:1 98 80±2 Warnecke &
Pluta (2003)
Carpa
comum
(Cyprinus
NaCl +
Sacarose
DMSO 10% 2,0x105:1 73,6±6,5 62,8±5,9 Irawan et al.
(2010)

22
carpio)
Carpa
espelho
(Cyprinus
carpio)
NaCl + KCl
+ Tris
Dimetilacetamida
15%
2,5x105:1 25,9 - Akçay et al.
(2004)
Carpa
comum
(Cyprinus
carpio)
Kurokura DMSO 10% 1,8-2,4x105:1 56±10 52±9 Linhart et al.
(2000)
Carpa
indiana
(Labeo
calbasu)
Alsever DMSO 10% ou
Metanol 10%
4,1x105:1 65±3 ou
63±4
45±2 ou
45±3
Nahiduzzama
n et al. (2012)
Posteriormente, os ovócitos fertilizados são transferidos para um sistema de
incubação, desenvolvido de acordo com as especificidades do animal. As incubadoras podem
apresentar dimensões de 0,6m de altura e 0,7m de diâmetro com capacidade para 90L
(NAHIDUZZAMAN et al., 2012). É importante ressaltar que o suprimento de oxigênio,
temperatura adequada e fluxo contínuo de água são fundamentais para o desenvolvimento
embrionário regular. Este é comumente descrito pelos autores em fases divididas em zigoto,
primeiras clivagens, mórula, blástula, gástrula, fechamento do blastóporo, diferenciação do
embrião e embrião eclodido, sendo o tipo de clivagem meroblástica discoidal. Durante o
surgimento de cada fase é registrado o tempo (horas ou minutos) após o início da fertilização.
Sividanes et al. (2012) observou as primeiras segmentações com 22 minutos, estágio
de mórula com 1 hora e 43 minutos, blástula com 2 horas e 27 minutos, gástrula com 5 horas
e 50 minutos, fechamento do blastóporo com 9 horas e 49 minutos, diferenciação do embrião
com 12 horas e 56 minutos e a eclosão das larvas com 20 horas e 3 minutos após a fertilização
a uma temperatura de 24 °C. Enquanto Nica et al. (2012) verificou as primeiras segmentações
com 50 minutos, estágio de mórula e blástula com 50 minutos a 1 hora e 35 minutos, gástrula
com 6 horas e 15 minutos, diferenciação do embrião com 15 horas e eclosão das larvas com
50 horas após a fertilização a uma temperatura de 22 °C. Portanto percebe-se que os tempos
dos registros embrionários e eclosão dependem principalmente da temperatura da água de
incubação dos embriões (SORENSEN et al., 1966). Do exposto, torna-se fundamental o
conhecimento da embriologia detalhada da carpa comum para uma melhor compreensão da
biologia da espécie.

23
3 JUSTIFICATIVA
O desenvolvimento de biotécnicas como a indução hormonal, a reprodução artificial
em cativeiro e a criopreservação de gametas vem contribuído para aumentar a produção de
pescado, permitindo o crescimento deste setor mundialmente. No entanto, são inúmeros os
protocolos que visam à produção de carpa comum devido às diferenças espécie-específicas
das características gaméticas (YANG; TIERSCH, 2009). Em todo o mundo, já foram
desenolvidos protocolos de criopreservação de sêmen para mais de 200 espécies de pescado
(TIERSCH et al., 2007). No entanto, ainda não existe na literatura um protocolo definido de
criopreservação seminal de carpa comum utilizando o diluente ACP-104 suplementado com
vitaminas C ou E. A execução deste protocolo investigará a capacidade de aumentar a
qualidade seminal e proteger os espermatozoides de carpa comum do efeito tóxico dos
radicais livres durante o processo de criopreservação seminal. Além disso, o estabelecimento
da dose inseminante usando sêmen in natura e a caracterização do desenvolvimento
embrionário desta espécie no nordeste do Brasil auxiliará a definição do melhor protocolo a
ser utilizado com sêmen criopreservado em água de coco em pó, possibilitando um aumento
na variabilidade genética dos plantéis nordestinos e diminuição do grau de endocruzamentos
dos estoques reprodutores.

24
4 HIPÓTESE CIENTÍFICA
O diluidor ACP-104 suplementado com vitamina C e combinado com 10% do
crioprotetor DMSO apresentou melhor desempenho na qualidade seminal após a
criopreservação de carpa comum quando comparado ao mesmo meio sem
suplementação.
A dose inseminante de 200.000 espermatozoides/ovócito utilizando sêmen in natura
produziu uma taxa aceitável de alevinos de carpa comum pós-fertilização no Nordeste
do Brasil.
A cronologia do desenvolvimento embrionário da carpa comum no Nordeste do Brasil
não é semelhante ao relatado para a espécie em clima temperado, porém as imagens
registradas estão de acordo com o clima temperado.

25
5 OBJETIVOS
5.1 OBJETIVO GERAL
Avaliar o efeito de diferentes antioxidantes e crioprotetores sobre a qualidade
do sêmen criopreservado de carpa comum (Cyprinus carpio) e definir a melhor
proporção do número de espermatozoides por ovócito (dose inseminante)
utilizando sêmen in natura;
5.2 OBJETIVOS ESPECÍFICOS
Otimizar o protocolo existente definindo inicialmente o crioprotetor que deve
ser usado em associação com o meio ACP-104 para a criopreservação seminal
de carpa comum;
Registrar e caracterizar em imagens o desenvolvimento embrionário de carpa
comum no nordeste do Brasil (3°45'00" de latitude sul e 39°10'24" de
longitude oeste);

26
6 CAPÍTULO 1
Influência da qualidade da água sobre o desenvolvimento embrionário de carpa comum
(Cyprinus carpio)
Water quality influence on the embryonic development of common carp
Periódico: Ciência Animal (ISSN: 0104-3773)
Qualis: B4

27
RESUMO
O estado do Ceará se destaca pela presença de açudes que auxiliam a piscicultura. Dentre
estes reservatórios de água, destaca-se o açude Pereira de Miranda, que se encontra
atualmente em estado de eutrofização. Dessa forma, a presente pesquisa objetivou avaliar a
influência da qualidade da água do açude Pereira de Miranda sobre o desenvolvimento
embrionário de carpa comum. Para tanto, os gametas obtidos foram fertilizados e o
acompanhamento do desenvolvimento embrionário foi realizado até a eclosão dos ovos para
se estimar as taxas de fertilização e eclosão. O tipo de clivagem foi meroblástica discoidal,
típica de ovos telolécitos. A taxa de fertilização média encontrada foi de 17,25 ± 1,19%.
Porém, a taxa de eclosão foi nula, observando-se embriões degenerados na sua totalidade. A
baixa taxa de fertilização e não eclosão pode estar relacionada as alterações da maioria dos
parâmetros de qualidade de água analisados tais como: temperatura (28,63 ± 0,67 ºC),
oxigênio dissolvido (4,38 ± 0,41 mg/L O2), dureza (216,6 mg/L de CaCO3), alcalinidade (156
mg/L de CaCO3), pH (7,5 a 10,9), amônia (1,09 mg/L) e nitrito (0,104 mg/L) que
permaneceram fora da faixa ideal para o desenvolvimento inicial desta espécie. Assim,
conclui-se que a eutrofização do açude Pereira de Miranda pode prejudicar a reprodução de
carpa comum nas estações de piscicultura abastecidas pelo açude, o que torna urgente a
definição de uma série de ações para melhoria de suas águas.
Palavras-chave: fertilização assistida, piscicultura, reprodução

28
Influência da qualidade da água sobre o desenvolvimento embrionário de carpa comum
(cyprinus carpio)
Francisco Renan Aragão LINHARES1*, Larissa Teixeira NUNES1, Júlia Trugilio LOPES1,
José Agenor Soares GALVÃO2, José Ferreira NUNES1, Carminda Sandra Brito SALMITO-
VANDERLEY1
1Universidade Estadual do Ceará. 4Departamento Nacional de Obras Contra as Secas -
DNOCS
Abstract
The state of Ceará is highlighted by the presence of dams that help fish farming. Among these
water reservoirs, there is the dam Pereira de Miranda, which is currently suffer from
eutrophication. Thus, the present study aimed to evaluate the influence of the reservoir water
quality Pereira de Miranda on the embryonic development of common carp. Therefore, the
obtained gametes were fertilized and monitoring of embryonic development was carried out
until the eggs hatch to estimate the fertilization and hatching rates. The type of cleavage was
meroblastic discoidal typical of telolecithal eggs. The average fertilization rate was found to
be 17.25 ± 1.19%. However, the hatching rate was zero, observing degenerate embryos in its
entirety. The low rate of fertilization and not hatch may be related changes in most water
quality parameters analyzed such as temperature (28.63 ± 0.67 ºC), dissolved oxygen (4.38 ±
0.41 mg/L O2), hardness (216.6 mg/L CaCO3), alkalinity (156 mg/L CaCO3), pH (7.5 to
10.9), ammonia (1.09 mg/L) and nitrite (0.104 mg/L) which have remained outside the
optimal range for the initial development of this species. Thus, it is concluded that the
eutrophication of the dam Pereira de Miranda can impair reproduction of common carp in fish
culture stations supplied by the dam, which makes it urgent to design a series of actions to
improve their waters.
Keywords: assisted fertilization, fish farming, reproduction
Introdução
Atualmente, a carpa comum (Cyprinus carpio) representa o quarto pescado da
aquicultura continental mais produzido no Brasil (Boletim estatístico da pesca e aquicultura,
2011). De acordo com o mesmo boletim, o Nordeste representa a segunda maior produção
aquícola continental de pescado, estando o estado do Ceará na liderança entre as regiões

29
nacionais. A espécie foi introduzida no final do século XIX com a chegada dos primeiros
imigrantes (Silva, 2005) e no Nordeste, através do Departamento Nacional de Obras Contras
às Secas (DNOCS) no ano de 1977 (DNOCS, 2009).
Neste contexto, os açudes construídos no Ceará apresentam grande importância para a
piscicultura do estado. Dentre estes reservatórios de água, destaca-se o açude Pereira de
Miranda, com barragem localizada no município de Pentecoste, Ceará e distante 85 km de
Fortaleza. O mesmo, foi construído pelo DNOCS entre os anos de 1950 e 1957, visando o
abastecimento de água, a irrigação, a geração de energia elétrica e a piscicultura. No entanto,
o açude citado encontra-se em estado de eutrofização, podendo prejudicar entre outras
utilidades a piscicultura da região (COGERH, 2015). Diante deste quadro, a avaliação dos
parâmetros físicos, químicos e microbiológicos da água do açude em questão, será
fundamental para se monitorar a sobrevivência e reprodução dos peixes no ambiente em que
vivem.
Dessa forma, a presente pesquisa objetivou avaliar a influência da qualidade da água,
por meio de análises de parâmetros fiscos, químicos e microbiológicos do açude em estado de
eutrofização Pereira de Miranda sobre o desenvolvimento embrionário de carpa comum.
Material e métodos
A pesquisa foi aprovada pelo Comitê de Ética para o Uso de Animais da Universidade
Estadual do Ceará (11516665-3/63). O trabalho foi realizado no Centro de Pesquisas em
Aquicultura (CPAq) do Departamento Nacional de Obras Contra as Secas (DNOCS) em
Pentecoste - Ceará, situando-se a 3° 45' 00" de latitude sul e 39° 10' 24" de longitude oeste,
local onde o abastecimento de água é realizada pelo açude Pereira de Miranda.
Para a realização da fertilização assistida, um pool de sêmen foi formado a partir da
coleta de sêmen de três machos sexualmente maduros de comprimento médio de 43,2 ± 2,8
cm e peso médio de 1,51 ± 0,2 kg, que receberam dose única de extrato hipofisário de carpa
(EHC) de 1 mg/kg de peso vivo. Doze horas após a indução hormonal da reprodução, os
animais foram mergulhados individualmente num tanque contendo solução anestésica de
Eugenol (Cequímica Ltda., Brasil) diluído em álcool absoluto e água (1:10:1000) por
aproximadamente dois minutos. Durante a coleta do sêmen, os animais foram contidos em
decúbito lateral sobre uma esponja (D33) e envolvidos com um pano úmido sobre os olhos
para minimizar o estresse. O orifício urogenital foi enxuto com papel toalha e o sêmen foi
coletado em tubos graduados de polietileno realizando-se uma pressão abdominal no sentido
ântero-posterior.

30
As amostras foram conservadas sobre gelo em escamas em caixas térmicas a uma
temperatura de aproximadamente 5 °C até a realização da fertilização artificial assistida. Para
a seleção das amostras, foi estimada a análise subjetiva da motilidade utilizando um
microscópio óptico (Nikon Eclipse E200 – ampliação de x400) após a mistura de uma
alíquota de 2 µL de sêmen e 100 µL de água do tanque, considerando-se como 0% (zero)
nenhum espermatozoide móvel e 100% (cem) todos os espermatozoides móveis. Somente as
amostras com o valor superior a 80% de motilidade foram utilizadas, enquanto aquelas
contaminadas com água, fezes, urina ou sangue foram descartadas.
Para a coleta de ovócitos, foram utilizadas duas fêmeas, de comprimento médio de 46
± 5,7 cm e peso médio de 2,38 ± 0,6 kg, submetidas à indução da desova pela aplicação de
duas doses de 0,5 e 5,0 mg/kg de peso vivo de EHC, num intervalo de doze horas entre elas.
A partir do material coletado, formou-se um pool de ova do qual foram retiradas cinco
amostras de 25 g (aproximadamente 21.800 ovócitos) de ovócitos para fertilização a seco de
cada amostra com 157,5 µL de sêmen fresco. Essa dose de sêmen correspondia à proporção
aproximada de 200.000 espermatozoides por ovócito (Irawan et al., 2010). Os gametas foram
então misturados suavemente durante dois minutos a fim de evitar injúrias nos ovócitos.
Para a remoção da adesividade dos ovócitos, cada amostra foi imersa durante uma
hora em 100 mL de solução de ureia, sob homogeneização constante. Completado os 60
minutos, as amostras foram lavadas com 100 mL de solução de ácido tânico puríssimo (0,5
g/L) (Vetec Ltda., Brasil) durante 20 segundos (Billard et al., 1995). Para a remoção desta
última solução, todas as amostras (n=5) foram então lavadas três vezes com 100 mL de água
do tanque. Posteriormente, cada uma das amostras foram transferidas para incubadoras
confeccionadas a partir de canos de PVC (policloreto) teladas no fundo de volume
aproximado de 1 L.
Inicialmente, o acompanhamento do desenvolvimento embrionário foi realizado a cada
meia hora e posteriormente a cada hora até a eclosão dos ovos. Estes foram removidos das
incubadoras com auxílio de uma peneira e depositados em placas de Petri para observações
em microscópio óptico (Nikon Eclipse E200 – ampliação de x400), equipado com uma
câmera fotográfica digital, a fim de documentar e melhor visualizar os ovos. As taxas de
fertilização foram calculadas por meio da contagem do número de embriões em estágio de
gástrula pelo total de ovócitos. A taxa de eclosão foi estimada contando-se o número de larvas
e dividindo pelo número total de ovócitos fertilizados. Foram observados aleatoriamente 200
embriões de cada incubadora para ambos os parâmetros avaliados, sendo realizada uma média
das cinco replicatas.

31
Paralelamente ao experimento, foram analisados parâmetros físicos, químicos e
microbiológicos da água utilizada na incubação dos embriões. A temperatura, o oxigênio
dissolvido (OD) e o pH foram medidos in loco de três em três horas desde o início da
fertilização assistida até as fases finais do desenvolvimento embrionário, no qual utilizou-se
um medidor de oxigênio dissolvido e temperatura (Phtek, modelo DO-100) e pHmetro
(KASVI, modelo K39-0014P). Para os parâmetros alcalinidade, dureza, amônia e nitrito
foram coletadas amostras de água das incubadoras de embriões do CPAq – DNOCS e
refrigeradas a 4°C conforme recomendações do Guia da CETESB (Companhia de Tecnologia
de Saneamento Ambiental) até serem processadas em Fortaleza pela Superintendência
Estadual do Meio Ambiente do Ceará (SEMACE).
Para os parâmetros microbiológicos, foi analisada o número mais provável de
coliformes termotolerantes e o número mais provável de Pseudomonas. Para tanto, as
amostras também foram processadas no mesmo dia pelo LABOMAR: Instituto de Ciências do
Mar – UFC. No local, as amostras foram serialmente diluídas em solução salina estéril até 10-
5. O Número Mais Provável (NMP) de coliformes termotolerantes foi determinado segundo a
técnica dos tubos múltiplos usando meio de cultura Caldo Lauryl MUG (Difco). Para se obter
o NMP de coliformes termotolerantes, consultou-se a tabela de Mc Crady (APHA, 2005). O
NMP de Pseudomonas spp. foi obtido através da técnica de tubos múltiplos usando caldo
asparagina, como citado por Cabrini & Gallo (2001). Dos tubos positivos (com turvação)
foram repicadas alçadas para placas de ágar cetrimide e incubadas a 35 °C por 48 h. O
surgimento de coloração esverdeada em toda a placa ou de colônias com luminescência verde
indicou resultado positivo para a presença de Pseudomonas spp. Para a contagem de bactérias
heterotróficas cultiváveis foi utilizada a técnica de plaqueamento pour plate onde uma
alíquota de 1 mL de cada diluição sucessiva foi colocado em placa de Petri e coberto com o
meio de cultivo Plate Count Agar (PCA), misturado e incubadas em estufa a 35–37 ºC por 48
h de acordo com o Standard Methods (APHA, 2005).
Resultados e discussão
As imagens registradas para o desenvolvimento embrionário se enquadram no padrão
relatado pela literatura para a espécie carpa comum (Cyprinus carpio). O tipo de clivagem foi
meroblástica discoidal típica de ovos telolétcitos e o desenvolvimento embrionário,
apresentou três períodos bem definidos, divididos em mórula, blástula e gástrula (Leite et al.,
2013) (Fig. 1).

32
Figura 1. Estágios do desenvolvimento embrionário de carpa comum (Cyprinus carpio). (A)
formação de mórula, (B) formação de blástula e (C) gástrula.
A taxa de fertilização média encontrada foi de 17,25 ± 1,19%, sendo considerada
baixa quando comparada a 75,6 ± 7,5% observada por Irawan et al. (2010) para sêmen fresco.
A taxa de eclosão foi nula, observando-se embriões degenerados na sua totalidade.
A temperatura é considerada um fator importante para se monitorar a má formação nas
larvas, pois segundo Moreira et al. (2001), a temperatura ótima da água para se realizar a
reprodução induzida das carpas comuns, situa-se entre 22 a 28 ºC. Assim, a temperatura
média de 28,63 ± 0,67 ºC (Tab. 1) encontrada no presente trabalho está no limite para os
padrões de outros estudos na área de desenvolvimento inicial de carpas, podendo ter
ocasionado má formação dos embriões e não eclosão.
Tabela 1. Médias dos valores físicos, químicos e microbiológicos da água do açude Pereira de Miranda nas
unidades experimentais durante o experimento. Pentecoste, Ceará.
Parâmetros Média ± D. P.
Temperatura (°C) 28,63 ± 0,67
Oxigênio dissolvido (mg/L O2) 4,38 ± 0,41
Dureza (mg/L CaCO3) 216,6
Alcalinidade (mg/L CaCO3) 156
pH (0-14) 7,5 a 10,9
Amônia (mg/L) 1,09
Nitrito (mg/L) 0,104
Coliformes termotolerantes (100 mL-1) 17 x 102
Pseudomonas sp. (100 mL-1) 4,5 x 102
A concentração de oxigênio dissolvido para um desenvolvimento embrionário ideal
em carpa comum encontra-se entre 6 a 9 mg L-1 O2, sendo considerada uma das espécies mais
exigentes (Boryshpolets et al., 2009). No entanto, o valor médio encontrado para o oxigênio
dissolvido nas incubadoras dos embriões foi de 4,38 ± 0,41 mg/L O2 (Tab. 1), sendo inferior
ao relatado na literatura para a espécie.
A dureza e a alcalinidade apresentaram níveis de 216,6 e 156 mg/L de CaCO3 (Tab. 1)
podendo ser classificadas como águas brandamente duras e, portanto, sem aplicação biológica

33
(GRAEFF; MONDARDO, 2006). Além disso, esses dois parâmetros podem ter ocasionando
uma grande oscilação do pH de 7,5 a 10,9 (Tab. 1).
A concentração de amônia encontrada de 1,09 mg/L (Tab. 1) foi considerada alta,
apesar de permanecer dentro da tolerância de 0,6 e 2 mg/L usualmente reportados. O nível de
nitrito foi de 0,104 mg/L, estando fora das concentrações letais para a espécie estudada
(Golombieski et al., 2005).
No que diz respeito às análises bacteriológicas, verificou-se a ocorrência de coliformes
termotolerantes (17 x 102 100 mL-1) (Tab.1) indicando presença de efluentes de esgotos
domésticos, assim como também a ocorrência de Pseudomonas sp. (4,5 x 102 100 mL-1)
(Tab.1), consideradas patógenos oportunistas na infecção de ovos de peixes. De fato,
Sorensen et al. (2014) verificou que a interferência microbriana em ovos fertilizados de
enguia (Anguilla anguilla) afetou significativamente a taxa de eclosão e a longevidade larval.
Além disso, observou-se na água a presença de uma grande quantidade de zooplâncton,
podendo ser responsável também pela degeneração de embriões em desenvolvimento.
Podemos concluir que a eutrofização do açude Pereira de Miranda pode prejudicar a
reprodução de carpa comum nas estações de piscicultura abastecidas pelo açude, o que torna
urgente a definição de uma série de ações para melhoria de suas águas.
Agradecimentos
Agradecemos ao Departamento Nacional de Obras Contra Secas (DNOCS) e
Laboratório de Biotecnologia da Reprodução de Peixes (LBRP), por fornecer as instalações e
as amostras utilizadas nos experimentos. A Financiadora de Estudos e Projetos (FINEP) pelo
apoio financeiro e ao Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq) pela concessão de bolsa de estudos.
Referências bibliográficas
Companhia de Gestão dos Recursos Hídricos – COGERH. Qualidade da água do açude
pereira de miranda, ceará: avaliação e recomendações. Fortaleza, 2015. Acessado em 15
outubro de 2012. Disponível em: http://www.srh.ce.gov.br/publicacoes/artigos/Qualidade-
Agua-%20Acude-%20Pereira-de-Miranda-Ceara-%20Avaliacao-
%20Recomendacoes.pdf/view?searchterm=None.

34
APHA - American Public Health Association. American Water Works Association, Water
Environment Federation. Standard Methods for The Examination of Water and Wastewater
Analysis. 21st ed. Washington, D.C.: American Public Health Association, 2005.
BILLARD, R.; COSSON, J.; PERCHEC, G.; LINHART, O. Biology of sperm and artificial
reproduction in carp. Aquaculture, v. 129, p. 95-l 12, 1995.
BORYSHPOLETS, S.; DZYUBA, B.; RODINA, M.; LI, P.; HULAK, M.; GELA, D.;
LINHART, O. Freeze-thawing as the factor of spontaneous activation of spermatozoa motility
in common carp (Cyprinus carpio L.). Cryobiology, v. 59, p. 291-296, 2009.
CABRINI, K. T.; GALLO, C. R. Avaliação da qualidade microbiológica de águas minerais
envasadas. Revista Higiene Alimentar, Mirandópolis, v. 15, n. 90-91, p. 83-92, 2001.
Departamento Nacional de Obras Contra as Secas. Relatório 2008. Fortaleza, 2009. Acessado
em: 8 outubro de 2012. Disponível em:
http://www.dnocs.gov.br/php/CGU/dnocs_relatorio_anual_2008.pdf.
GRAEFF, A.; MONDARDO, M. Influência do probiótico no crescimento das carpas comum
(Cyprinus carpio l., 1758) na fase de recria. Revista Eletrônica de Veterinária, v.7, n.11,
2006.
GOLOMBIESKI, J. I.; MARCHEZAN, E.; MONTI, M. B.; STORCK, L.; CAMARGO, E.
R.; SANTOS, F. M. Qualidade da água no consórcio de peixes com arroz irrigado. Ciência
Rural, v. 35, n. 6, p. 1263-1268, 2005.
IRAWAN, H.; VUTHIPHANDCHAI, V.; NIMRAT, S. The effect of extenders,
cryoprotectants and cryopreservation methods on common carp (Cyprinus carpio) sperm.
Animal reproduction science, v. 122, p. 236-243, 2010.
LEITE, L. V.; MELO, M. A. P.; OLIVEIRA, F. C. E.; PINHEIRO, J. P. S.; CAMPELLO, C.
C.; NUNES, J. F.; SALMITO-VANDERLEY, C. S. B. Determinação da dose inseminante e
embriogênese na fertilização artificial de tambaqui (Colossoma macropomum). Arquivo
Brasileiro de Medicina Veterinária e Zootecnia, v. 65, n. 2, p. 421-429, 2013.

35
MINISTÉRIO DA PESCA E AQUICULTURA. Boletim Estatístico da Pesca e Aquicultura
2011. Acessado em: 9 fevereiro 2013. Disponível em:
http://www.mpa.gov.br/files/docs/Boletim_MPA_2011_pub.pdf.
MOREIRA, H. L. M.; VARGAS, L.; RIBEIRO, R. P.; ZIMMERMANN, S. Fundamentos da
moderna aquicultura. Canoas: Editora ULBRA, 2001, 200 p.
SILVA, N. J. R. Dinâmicas de desenvolvimento da piscicultura e políticas públicas no Vale
do Ribeira/SP e Alto Vale do Itajaí/SC – Brasil. Tese (Doutorado em aqüicultura e à école
nationale supérieure agronomique de rennes) - Universidade Estadual Paulista. Curso de Pós
Graduação em Aqüicultura do Centro de Aqüicultura, Universidade Estadual Paulista,
Jaboticabal, Brasil. 2005, 544p.
SORENSEN, S. R.; SKOV, P. V.; LAUESEN, P.; TOMKIEWICZ, J.; BOSSIER, P.;
SCHRYVER, P. D. Microbial interference and potential control in culture of European eel
(Anguilla anguilla) embryos and larvae. Aquaculture, v. 426-427, p. 1-8, 2014.

36
7 CAPÍTULO 2
Biotécnicas aplicadas a reprodução de ciprinídeos
Biotechs applied the reproduction of cyprinids
Periódico: Acta Veterinaria Brasilica (ISSN: 1981-5484)
Qualis: Qualis: B3

37
RESUMO
Esta revisão relata as variações de cada etapa que abrange o processo de crioprervação
seminal e fertilização assistida artificial em ciprinídeos. Os diluidores mais utilizados para a
criopreservação do sêmen de carpa são uma combinação de sais e açucares associados aos
crioprotetores DMSO ou metanol. Para o armazenamento em nitrogênio líquido as amostras
podem ser envasadas em palhetas de diferentes volumes e congeladas em caixas térmicas de
poliestireno, refrigeradores programáveis, dry shippers ou botijões criogênicos. Após o
descongelamento, uma alíquota do sêmen criopreservado é retirada para o cálculo da dose
inseminante e realização da fertilização assistida artificial. Em seguida, os ovos fertilizados
são tratados com uréia, NaCl e ácido tânico e transferidos para um sistema de incubação.
Diante de uma variedade de protocolos relatados pela literatura, ainda é fundamental o
aperfeiçoamento continuado destas biotécnicas para permitir o aprimoramento dos programas
de reprodução de ciprinídeos.
Palavras-chave: Cyprinus sp; criopreservação seminal; fertilização assistida.

38
Biotécnicas aplicadas a reprodução de ciprinídeos
Carminda Sandra Brito Salmito-Vanderley1*, Francisco Renan Aragão Linhares 2, Maria
Audália Marques de Carvalho 3, Mayara Setúbal Oliveira 4, José Ferreira Nunes5
1 Profa. Doutora Adjunta da Universidade Estadual do Ceará (UECE) e Pesquisadora do
Programa de Pós-Graduação em Ciências Veterinárias (PPGCV). * Autor para
correspondência: E-mail: [email protected].
2 Doutorando em Reprodução e Sanidade Animal pelo PPGCV – UECE.
3 Doutora em Biotecnologia pela Rede Nordeste de Biotecnologia – UECE.
4 Mestranda em Reprodução e Sanidade Animal pelo PPGCV – UECE.
5 Prof. Doutor Titular da Universidade Estadual do Ceará e Pesquisador do PPGCV.
Abstract
This review reports the variations of each stage which covers the process of sperm
cryopreservation and artificial fertilization in cyprinids. The most commonly used extenders
for sperm cryopreservation of carp are a combination of salts and sugars associated with
DMSO or methanol. The samples can be packaged in straws of different volumes for storage
in liquid nitrogen and frozen in styrofoam boxes, computer-controlled freezer, dry shippers or
cryogenic vessel. After thawing an aliquot of cryopreserved semen is taken for the calculation
of insemination dose and realization of artificial assisted fertilization. Then fertilized eggs are
treated with urea, NaCl and tannic acid and transferred to incubation system. Faced with a
variety of protocols reported in the literature, it is still essential to continued improvement of
these biotechnologies to enable the improvement of breeding programs for cyprinids.
Keywords: Cyprinus sp; sperm cryopreservation; artificial fertilization.
Introdução
Constatando a pobreza da ictiofauna do semiárido nordestino, o brasileiro Rodolfo von
Ihering, diretor da Comissão Técnica de Piscicultura do Nordeste, passou a estudar inúmeras
espécies de outras bacias hidrográficas, a partir de 1935, com o objetivo de serem

39
introduzidas na região. Dentre as espécies de alto valor comercial, destaca-se a carpa comum
(Cyprinus carpio vr hungaricus), trazida da Hungria em 1986 (DNOCS, 2009).
Nesse contexto, o desenvolvimento de biotécnicas reprodutivas de base como a
indução hormonal e a reprodução artificial em cativeiro vem contribuindo para aumentar a
produção de pescado, permitindo o crescimento deste setor mundialmente. Outra biotécnica
reprodutiva bastante difundida em peixes é a criopreservação seminal. Em todo o mundo, já
foram desenvolvidos protocolos de criopreservação de sêmen para mais de 200 espécies de
pescado (Tiersch et al., 2007). Na carpa comum, são inúmeros os protocolos de
criopreservação e fertilização assistida que visam a sua produção.
Assim, diante da importância da aplicação destas biotécnicas, este artigo apresentará
uma revisão sobre o conhecimento atual das técnicas de criopreservação seminal e fertilização
assistida utilizadas na produção de ciprinídeos.
Criopreservação
Desde que Blaxter (1953) realizou os primeiros estudos de criopreservação de gametas
de peixes, muitas pesquisas têm sido voltadas para a criopreservação do sêmen de várias
espécies de pescado, incluindo a carpa comum (Cyprinus carpio). A introdução desta
biotécnica na reprodução animal contribui para aumentar a propagação de material genético
de alto valor em menor espaço de tempo.
A execução de um protocolo de criopreservação de sêmen inclue ajustes de taxas de
diluição, escolhas de diluentes e crioprotetores, tempo de equilíbrio entre o crioprotetor e os
espermatozoides, método de envase adequado, taxas de resfriamento e taxas de descongelação
das amostras. Diante da presença de várias etapas e suas interações surgem dificuldades para a
padronização de protocolos de congelação seminal (Yang & Tiersch, 2009). Nesse contexto,
abordaremos as variações de cada etapa que abrange o processo de crioprervação seminal de
carpa comum encontradas na literatura nos últimos anos.
Diluição seminal

40
Os espermatozoides, quando submetidos às alterações rigorosas de temperaturas, estão
sujeitos à danos ocasionados pelo frio, denominados crionjúrias (Silva & Guerra, 2011). Estes
danos podem levar a um decréscimo nas características espermáticas, como a velocidade,
motilidade, capacidade de fertilização e subsequente redução das taxas de eclosão (Li et al.,
2010). Dessa forma, após a coleta, o sêmen é submetido à etapa de diluição resultante da
adição de soluções protetoras capazes de minimizar os danos causados pelas crioinjúrias.
Estas soluções são chamadas de crioprotetores, atuando na diminuição do ponto de
congelação da água, reduzindo o estresse osmótico e os efeitos da cristalização sobre a
estrutura celular (Salmito-Vanderley et al., 2012).
Devido à especificidade própria de cada grupo animal, torna-se fundamental a
adequação da composição do diluidor as características seminais de cada espécie. Na
literatura são relatadas soluções contendo apenas sais, apenas açucares ou uma combinação
dos mesmos para a criopreservação seminal de peixes. A solução de Alsever, cuja composição
é formada por NaCl e citrato de sódio, é considerado um dos diluidores mais conhecidos
utilizados em ciprinídeos (Nahiduzzaman et al., 2012). Além disso, muitos autores também
utilizam um diluidor descrito por Kurokura et al. (1984) resultantes da combinação dos sais
NaCl, KCl, CaCl2 e NaHCO3 (Akçay et al., 2004; Rani & Munuswamy, 2014). Outro tipo de
diluidor utilizado em ciprinídeos é a glicose (Horváth et al., 2007). Esta age como substrato
de energia, componente osmótico e agente crioprotetor em função do seu alto peso molecular,
contribuindo para o equilíbrio osmótico e atuando como substituto de eletrólitos (Holt, 2000).
Em geral, os diluidores acrescidos de açucares são capazes de melhorar a motilidade
espermática durante a criopreservação seminal de algumas espécies de ciprinídeos (Urbányi et
al., 2006). A combinação de sais (NaCl, KCl) e açucares (frutose, glicose e sacarose) podem
ser utilizados da mesma forma como diluidores de sêmen de carpa comum (Irawan et al.,
2010).
Outra alternativa, é a utilização do diluidor água de coco em pó (ACP®), composto
por sais, proteínas, açúcares, vitaminas, gorduras neutras, além de indutores da divisão celular
e diversos eletrólitos. O ACP® é considerado uma das inovações mais recentes da
biotecnologia do sêmen de peixe e já foi utilizado com sucesso por Linhares (2012) para a
criopreservação seminal de carpa comum. Recentemente, Öğretmen et al. (2014) obtiveram
resultados satisfatórios de motilidade espermática, taxas de fertilização e eclosão utilizando
extrato bruto de própolis como diluidor para a criopreservação de sêmen de carpa comum,

41
sendo identificado compostos fenólicos, terpenóides, flavonoides, ácidos graxos, açucares e
outros componentes.
Os diluentes seminais também atuam como carreadores de crioprotetores, agindo em
conjunto para proteger os espermatozoides contra os efeitos deletérios da congelação. Os
crioprotetores podem ser classificados como internos ou externos. Os crioprotetores internos
atuam na redução do ponto crioscópio da água intracelular evitando a formação de cristais de
gelo e o comprometimento da integridade da membrana espermática interna durante as
mudanças de temperatura. Os crioprotetores internos comumente utilizados em ciprinídeos
são dimetilsulfóxido (DMSO), metanol, propilenoglicol, dimetilacetamida (DMA), glicerol e
etanol (Akçay et al., 2004; Irawan et al., 2010; Sultana et al., 2010). Na maioria dos estudos, o
DMSO fornece os melhores resultados, provavelmente devido a sua rápida penetração em
espermatozoides e sua interação com os fosfolipídios da membrana espermática (Suquet et al.,
2000). Os crioprotetores externos atuam na estabilização da membrana plasmática e
restauração dos fosfolipídeos perdidos pelos espermatozoides durante a congelação. No
entanto, estes não são utilizados frequentemente em ciprinídeos, sendo relatadas na literatura
à adição de gema de ovo-citrato ou somente gema de ovo as soluções diluidoras de sêmen de
carpa comum (Sultana et al., 2010; Öğretmen et al., 2014).
Após a diluição, os crioprotetores requerem um tempo de equilíbrio para penetrar nas
células. Este é um processo dinâmico e depende da permeabilidade dos espermatozoides,
crioprotetores e as suas concentrações (Yang & Tiersch, 2009). Irawan et al. (2010) obtiveram
uma taxa de motilidade espermática de 94,5±3,3% utilizando como diluidor NaCl e sacarose,
diluente DMSO 10% e tempo de equilíbrio de 10 min a 25 °C para a criopreservação seminal
de carpa comum. Porém, utilizando as mesmas condições de criopreservação seminal da
espécie, Linhares (2012) iniciou o processo de congelamento das amostras sem a utilização de
tempo de equilíbrio obtendo 61,8±10,7% de motilidade espermática. Por outro lado,
Nahiduzzaman et al. (2012) observaram que quando as amostras de sêmen de carpa indiana
(Labeo calbasu) foram criopreservadas com Alsever e DMSO 5%, maiores taxas de
motilidade foram obtidos, aproximadamente 90±0%, em todos os tempos de equilíbrio (5, 10,
15, 20 e 25 min) testados a 4 °C. No entanto, Akçay et al. (2004) obteve uma taxa de
motilidade espermática de 55% utilizando um tempo de equilíbrio de 45 min a 4 °C para
criopreservar sêmen de carpa comum diluído em NaCl, KCl, CaCl2, NaHCO3 e DMSO 15%.

42
Portanto, a variedade dos tempos de equilíbrio relatadas na literatura é esperada
devido a grande quantidade de diluidores utilizados para a família dos ciprinídeos. Além
disso, a taxa de diluição é um fator a ser considerado.
Taxas de diluição e armazenamento do sêmen
As concentrações dos diluentes precisam ser ajustadas antes de serem misturados aos
diluidores, pois os mesmos são tóxicos as células em temperatura ambiente. Comumente as
concentrações dos crioprotetores são ajustados a 5 ou 10%, pois as pesquisas apontam que em
altas concentrações alguns crioprotetores pode ser prejudicial à motilidade espermática. Rani
& Munuswamy (2014) observaram menores taxas de viabilidade espermática (35%)
utilizando 20% de DMSO, viabilidade mediana (61,5%, 62,3%, 60,8%) utilizando glicerol
(10% e 20%) e DMSO 15% e viabilidade máxima (75,8%) utilizando 10% de DMSO em
amostras de sêmen de carpa comum conservadas a 4 °C. Após a verificação da melhor
concentração do crioprotetor e formação do diluente, o sêmen é submetido a diferentes taxas
de diluição para se verificar a melhor proporção entre as quantidades de diluente e sêmen. A
literatura relata a utilização de diferentes taxas de diluição para a criopreservação do sêmen de
carpa comum tais como 1:1, 1:3, 1:5, 1:6, 1:7 e 1:9 (Linhart et al., 2000; Warnecke & Pluta,
2003; Sultana et al., 2010; Linhares, 2012).
Para o armazenamento em nitrogênio líquido a mistura (sêmen:diluente) deve ser
envasada em recipientes padronizados para uniformizar o processo de resfrigeração e evitar
variações de transferência de calor. Para tal finalidade, podem ser utilizados palhetas de 0,25
mL, 0,5 mL, 1,2 mL, 2 mL e 5 mL (Linhart et al., 2000; Akçay et al., 2004; Horváth et al.,
2007; Irawan et al., 2010). Em seguida, o sêmen é submetido a uma queda gradual de
temperatura que influenciará o equilíbrio osmótico e o pH de soluções intra e extracelulares
durante a congelação. O ideal é que a taxa de resfrigeração seja rápida o suficiente para
minimizar o tempo de exposição às soluções concentradas e lenta o suficiente para minimizar
a quantidade de formação de gelo intracelular (Yang & Tiersch, 2009). Essa taxa de
resfriamento varia de acordo com o equipamento utilizado para fazê-lo.
Equipamentos de congelação

43
Os equipamentos utilizados para congelar as amostras de sêmen são as caixas térmicas
de poliestireno, refrigeradores programáveis, dry shippers e botijões criogênicos. A caixa
térmica de isopor é formada por poliestireno, dentro da qual é adaptada uma bandeja metálica
distante do fundo alguns centímetros acima da superfície do nitrogênio líquido. As dimensões
(comprimento x largura x altura) das caixas utilizadas podem variar de 27-40 cm x 18-24 cm
x 28-33 cm (Lahnsteiner et al., 2000; Linhares, 2012). A altura ideal para a congelação de
sêmen acima da superfície do nitrogênio líquido varia entre as espécies de ciprinídeos, sendo
utilizadas 2, 3, 4 e 6 cm (Irawan et al., 2010; Linhares, 2012). A temperatura na caixa pode
variar de -100 a -130 °C dependendo da quantidade de nitrogênio líquido despejado. As
amostras são suspensas sob a bandeja e congeladas por um período específico em vapor de
nitrogênio líquido podendo ser deixadas no interior da caixa por 3, 10, 20 ou 25 min (Alvarez
et al., 2003; Urbányi et al., 2006; Boryshpolets et al., 2009; Irawan et al., 2010). Dependendo
da espécie de peixe e da velocidade de resfriamento, o método de congelação de sêmen em
caixa térmica de poliestireno pode ser tão eficaz quanto à utilização de um refrigerador
programável.
Este último equipamento citado anteriormente é programado para controlar o
resfriamento gradual de amostras biológicas antes de serem imersas em nitrogênio líquido. A
redução da temperatura é realizada durante um período determinado pelo programador
estabelecendo-se um intervalo de temperatura inicial e final para o término do resfriamento.
Este processo é chamado de curva de resfriamento e pode variar entre as taxas de resfriamento
e intervalos de temperatura inicial e final. Nahiduzzaman et al. (2012), em apenas uma etapa,
resfriou as amostras de 5°C a -80°C numa taxa de resfriamento de 10°C/min enquanto Irawan
et al. (2010) utilizou a mesma taxa de resfriamento porém um intervalo de temperatura
diferente de 25°C a -40°C. Sultana et al. (2010), utilizando duas etapas, primeiramente
resfriou as amostras de 5°C a -4°C numa taxa de resfriamento de 4°C/min e em seguida
prosseguiu o resfriamento de -4°C a -80°C sob uma taxa de 10°C/min. No entanto, Linhart et
al. (2000) utilizou um protocolo diferente resfriando primeiramente as amostras de 4°C a -9°C
numa taxa de resfriamento de 4°C/min e em seguida continuou de -11°C a -80°C sob uma
taxa de 11°C/min.
O dry shipper é um contêiner para transportar com segurança amostras em
temperaturas criogênicas. Trata-se de um botijão em alumínio, cujo interior está recoberto por
um material de absorção de nitrogênio líquido responsável pela congelação a vapor de
amostras de sêmen a uma taxa de resfriamento de aproximadamente 25–40 °C min-1 com

44
estabilização gradual de -181 °C em 3 min (Taitson et al., 2007). O equipamento foi utilizado
com sucesso para carpa comum, obtendo boas taxas de motilidade espermática quando
comparado à caixa de poliestireno comumente utilizada para a espécie (Linhares, 2012). No
entanto, são escassas as pesquisas que relatam a utilização do dry shipper para criopreservar
sêmen de carpa comum. O dry shipper é utilizado com frequência para as espécies nativas
brasileiras tais como Brycon insignis e Brycon opalinus obtendo resultados positivos de
motilidade, fertilização e eclosão (Viveiros et al., 2012a,b).
Após o equilíbrio térmico, o sêmen congelado é transferido para um botijão de
nitrogênio líquido, onde pode ser conservado por longos períodos (Li et al., 2010) até ser
utilizado para fertilização.
Fertilização
O sucesso do protocolo de criopreservação é verificado através da descongelação do
sêmen em banho de imersão térmica e avaliação da qualidade espermática, sendo a
fertilização artificial a etapa final para a análise da qualidade dos gametas. A biotécnica pode
ser realizada por dois métodos diferentes: o método úmido que se baseia na mistura de
gametas masculinos e femininos coletados simultaneamente com água (Sultana et al., 2010) e
o método a seco que mistura primeiramente os gametas acrescentando-se água em seguida
(Akçay et al., 2004).
A reprodução artificial em carpa é mais complexa porque o procedimento de
inseminação requer uma solução específica para evitar a aderência entre os ovócitos que
ocorre normalmente na água doce. A aderência é oriunda do envelope vitelino que reveste os
ovócitos e é formado por carboidratos desidratados (Linhart et al., 1995). Essas camadas
acelulares envoltórias servem para proteção, nutrição e barreira pela qual os espermatozoides
devem atravessar (Montanari, 2013). Entretanto, essa mesma aderência é responsável pela
formação de uma massa de ova que dificulta o encontro da micrópila pelos espermatozoides,
prejudicando o processo de fertilização.
A solução comumente utilizada pode ser formulada de duas formas conforme a
quantidade de ureia adicionada. Uma delas é composta por 3 g de ureia e a outra por 20 g de
ureia juntamente com 4 g de NaCl para cada litro de água. Essas soluções atuam na ativação
dos espermatozoides e remoção da adesividade dos ovócitos durante agitação contínua por 1

45
hora a 1 hora e 30 minutos. Após esse período adiciona-se uma solução de ácido tânico
(0,5g/L) por 20 segundos, removendo-a em seguida (Billard et al., 1995). Vale ressaltar que
muitos autores relatam a utilização da solução contendo 3% de ureia (Horváth et al., 2007;
Irawan et al., 2010). No entanto, Billard et al. (1995) afirmam que a solução contendo 20% de
ureia é uma solução melhorada, necessitando de menos tempo de agitação para a remoção da
adesividade dos ovócitos. Além disso, pode-se utilizar também o leite em pó composto por
27,2% de gordura, 26,6g de albumina, 37,2g de açúcar do leite, 5,8g de íons, 0,2g de lecitina e
3g de água em 100g do mesmo para a preparação de uma solução de 40g de leite em pó por
litro de água declorada ou limpa. A solução de leite em pó deve ser adicionada lentamente
misturando-se um volume de 1 L da solução por Kg de ovócitos. Após 60 minutos, adiciona-
se lentamente mais água durante 10 minutos para substituição da solução de leite e em
seguida ovócitos podem ser transferidos para as incubadoras (Linhart et al., 2003).
Durante os testes de fertilização necessita-se definir a dose inseminante ideal, ou seja,
a proporção mínima de espermatozoides por ovócito capaz de produzir a máxima
sobrevivência em termos de larvas eclodidas. Esta técnica conserva e economiza gametas,
melhora a utilização do macho e contribui para um aumento da variabilidade genética. O
sucesso de uma fertilização artificial utilizando uma dose inseminante é determinado por meio
da taxa de fertilização, ou seja, a partir de uma coleta aproximada de 150 embriões contam-se
aqueles em estágio de gástrula e divide-se pelo número de ovócitos fertilizados (Horváth et
al., 2003). Da mesma forma, também é calculada a taxa de eclosão por meio da contagem do
número de larvas eclodidas dividida pelo número total de ovócitos fertilizados. A definição da
dose inseminante utilizando sêmen criopreservado de ciprinídeos já foi estabelecida por
alguns autores obtendo diferentes resultados de taxa de fertilização e eclosão (Quadro 1).
Posteriormente, os ovócitos fertilizados são transferidos para um sistema de
incubação, desenvolvido de acordo com as especificidades do animal. As incubadoras podem
apresentar dimensões de 0,6m de altura e 0,7m de diâmetro com capacidade para 90L
(Nahiduzzaman et al., 2012). É importante ressaltar que o suprimento de oxigênio,
temperatura adequada e fluxo contínuo de água são fundamentais para o desenvolvimento
embrionário regular. Este é comumente descrito pelos autores em fases divididas em zigoto,
primeiras clivagens, mórula, blástula, gástrula, fechamento do blastóporo, diferenciação do
embrião e embrião eclodido, sendo o tipo de clivagem meroblástica discoidal. Durante o
surgimento de cada fase é registrado o tempo (horas ou minutos) após o início da fertilização.
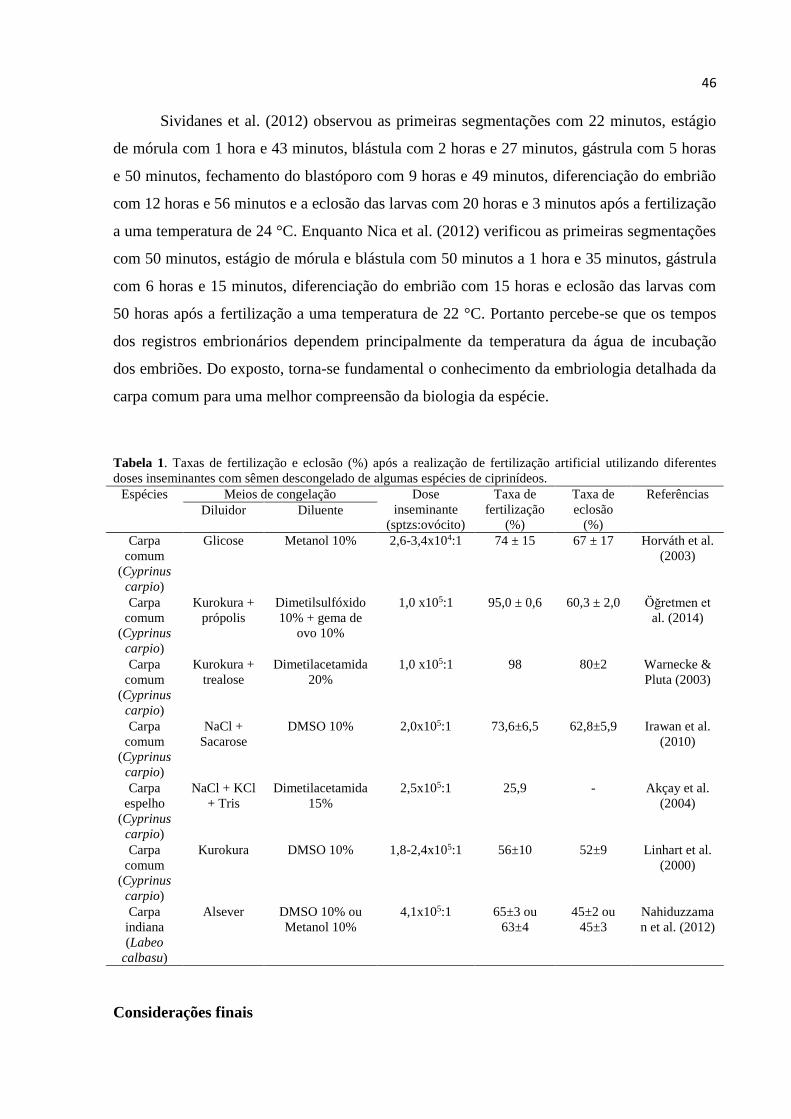
46
Sividanes et al. (2012) observou as primeiras segmentações com 22 minutos, estágio
de mórula com 1 hora e 43 minutos, blástula com 2 horas e 27 minutos, gástrula com 5 horas
e 50 minutos, fechamento do blastóporo com 9 horas e 49 minutos, diferenciação do embrião
com 12 horas e 56 minutos e a eclosão das larvas com 20 horas e 3 minutos após a fertilização
a uma temperatura de 24 °C. Enquanto Nica et al. (2012) verificou as primeiras segmentações
com 50 minutos, estágio de mórula e blástula com 50 minutos a 1 hora e 35 minutos, gástrula
com 6 horas e 15 minutos, diferenciação do embrião com 15 horas e eclosão das larvas com
50 horas após a fertilização a uma temperatura de 22 °C. Portanto percebe-se que os tempos
dos registros embrionários dependem principalmente da temperatura da água de incubação
dos embriões. Do exposto, torna-se fundamental o conhecimento da embriologia detalhada da
carpa comum para uma melhor compreensão da biologia da espécie.
Tabela 1. Taxas de fertilização e eclosão (%) após a realização de fertilização artificial utilizando diferentes
doses inseminantes com sêmen descongelado de algumas espécies de ciprinídeos.
Espécies Meios de congelação Dose
inseminante
(sptzs:ovócito)
Taxa de
fertilização
(%)
Taxa de
eclosão
(%)
Referências
Diluidor Diluente
Carpa
comum
(Cyprinus
carpio)
Glicose Metanol 10% 2,6-3,4x104:1 74 ± 15 67 ± 17 Horváth et al.
(2003)
Carpa
comum
(Cyprinus
carpio)
Kurokura +
própolis
Dimetilsulfóxido
10% + gema de
ovo 10%
1,0 x105:1 95,0 ± 0,6 60,3 ± 2,0 Öğretmen et
al. (2014)
Carpa
comum
(Cyprinus
carpio)
Kurokura +
trealose
Dimetilacetamida
20%
1,0 x105:1 98 80±2 Warnecke &
Pluta (2003)
Carpa
comum
(Cyprinus
carpio)
NaCl +
Sacarose
DMSO 10% 2,0x105:1 73,6±6,5 62,8±5,9 Irawan et al.
(2010)
Carpa
espelho
(Cyprinus
carpio)
NaCl + KCl
+ Tris
Dimetilacetamida
15%
2,5x105:1 25,9 - Akçay et al.
(2004)
Carpa
comum
(Cyprinus
carpio)
Kurokura DMSO 10% 1,8-2,4x105:1 56±10 52±9 Linhart et al.
(2000)
Carpa
indiana
(Labeo
calbasu)
Alsever DMSO 10% ou
Metanol 10%
4,1x105:1 65±3 ou
63±4
45±2 ou
45±3
Nahiduzzama
n et al. (2012)
Considerações finais

47
As biotécnicas utilizadas na produção de carpa comum vêm avançando e contribuindo
para aprimorar programas de reprodução de ciprinídeos. No entanto, diante de uma variedade
de ferramentas empregadas na produção de alevinos não há uma padronização nos protocolos
de criopreservação e reprodução assistida. A literatura relata diferenças entre diluidores, taxas
de diluição, armazenamento do sêmen, equipamentos de congelação e técnicas de fertilização
assistida em um mesmo grupo de animais, tornando esse conjunto de biotécnicas
inconsistentes e de difícil difusão. Portanto, percebe-se que os protocolos são desenvolvidos
para alcançar determinados objetivos conforme as necessidades e condições de pesquisa de
cada grupo.
Neste contexto, é fundamental o aperfeiçoamento continuado destas biotécnicas,
permitindo futuramente a preservação de espécies em via de extinção bem como aquelas de
potencial produtivo capazes de favorecer a geração de proteínas de origem animal adequadas
para aniquilar a fome proteica que assola parte da população mundial.
Referências bibliográficas
Alvarez B., Fuentes R., Pimentel R., Abad Z., Cabrera E., Pimentel E. & Arenal A. 2003.
High fry production rates using post-thaw silver carp (Hypophthalmichthys molitrix)
spermatozoa under farming conditions. Aquaculture. 220:195–201.
Akçay E., Bozkurt Y., Seçer S. & Tekün N. 2004. Cryopreservation of Mirror Carp Semen.
Turkish journal of veterinary and animal sciences. 28:837-843.
Billard R., Cosson J., Perchec G. & Linhart O. 1995. Biology of sperm and artificial
reproduction in carp. Aquaculture. 129:95-l 12.
Blaxter J.H.S. 1953. Sperm storage and cross-fertilization of spring and autumn spawning
herring. Nature. 172:1189-90.
Boryshpolets S., Dzyuba B., Rodina M., Li P., Hulak M., Gela D. & Linhart O. 2009. Freeze-
thawing as the factor of spontaneous activation of spermatozoa motility in common carp
(Cyprinus carpio L.). Cryobiology. 59:291–296.

48
Departamento Nacional de Obras Contra as Secas. Relatório 2008. Capturado em 8 de out.
2012. Online. Disponível na Internet
http://www.dnocs.gov.br/php/CGU/dnocs_relatorio_anual_2008.pdf
Holt W.V. 2000. Fundamental aspects of sperm cryobiology: The importance of species and
individual differences. Theriogenology. 53:47-58.
Horváth A., Miskolczi E. & Urbányi B. 2003. Cryopreservation of common carp sperm.
Aquatic Living Resources. 16:457–460.
Horváth A., Miskolczi E., Mihalffy S., Osz K., Szabo K. & Urbanyi B. 2007.
Cryopreservation of common carp (Cyprinus carpio) sperm in 1.2 and 5 ml straws and
occurrence of haploids among larvae produced with cryopreserved sperm. Cryobiology.
54:251–257.
Irawan H., Vuthiphandchai V. & Nimrat S. 2010. The effect of extenders, cryoprotectants and
cryopreservation methods on common carp (Cyprinus carpio) sperm. Animal reproduction
science. 122:236-243.
Kurokura H., Hirano R., Tomita M. & Iwahashi M. 1984. Cryopreservation of carp sperm.
Aquaculture. 37:267-273.
Lahnsteiner F., Berger B., Horváth A., Urbanyi B. & Weismann T. 2000. Cryopreservation of
spermatozoa in cyprinid fishes. Theriogenology. 54:1477-1496.
Linhares F.R.A. 2012. Efeito de diferentes taxas de diluição e protocolos de congelação sobre
a cinética e morfologia de espermatozoides de carpa comum (Cyprinus carpio)
criopreservados em água de coco em pó (ACP-104). Dissertação de mestrado, Universidade
Estadual do Ceará, Fortaleza. 80p.
Linhart O., Kudob S., Billard R., Slechtad V. & Mikodina E.V. 1995. Morphology,
composition and fertilization of carp eggs: a review. Aquaculture. 129:75-93.
Linhart O., Rodina M. & Cosson J. 2000. Cryopreservation of Sperm in Common Carp
Cyprinus carpio: Sperm Motility and Hatching Success of Embryos. Cryobiology. 41:241-
250.
Linhart O., Rodina M., Gela D., Kocour M. & Rodriguez M. 2003. Improvement of common
carp artificial reproduction using enzyme for elimination of egg stickiness. Aquatic Living
Resources. 16:450–456.

49
Li P., Hulak M., Koubek P., Sulc M., Dzyuba B., Boryshpolets S., Rodina M., Gela D.,
Manaskova-Postlerova P., Peknicova J. & Linhart O. 2010. Ice-age endurance: the effects of
cryopreservation on proteins of sperm of common carp, Cyprinus carpio L. Theriogenology.
74(3):413-23.
Montanari T. 2013. Embriologia: texto, atlas e roteiro de aulas práticas [recurso eletrônico]. 1ª
ed. Ed. do autor, Porto Alegre. Capturado em 10 de out. 2013. Online. Disponível na Internet
http://www.ufrgs.br/livrodeembrio/
Nahiduzzaman M., Hassanb M., Roya P.K., Hossainc A., Hossaina M.A.R. & Tiersch T.R.
2012. Sperm cryopreservation of the Indian major carp, Labeo calbasu: Effects of
cryoprotectants, cooling rates and thawing rates on egg fertilization. Animal Reproduction
Science. 136:133–138.
Nica A., Cristea V., Gheorghe D., Hoha G.V. & Enache I.B. 2012. Embryonic and larval
development of japanese ornamental carp Cyprinus carpio (Linnaeus, 1758). Lucrări
Ştiinţifice - Seria Zootehnie. 58:116-120.
Öğretmen F., İnanan B.E. & Öztürk M. 2014. Protective effects of propolis on
cryopreservation of common carp (Cyprinus carpio) sperm. Cryobiology. 68:107–112.
Rani K.U. & Munuswamy N. 2014. Preliminary studies on the cryopreservation of
spermatozoa in the fresh water fish common carp (Cyprinus carpio L.). Journal of Coastal
Life Medicine. 2(3):181-186.
Salmito-Vanderley C.S.B., Vieira M.J.A.F., Leite L.V., Oliveira F.C.E., Linhares F.R.A.,
Salgueiro C.C.M. & Nunes J.F. 2012. Meios de congelação para conservação de sêmen de
peixes da família Characidae. Ciência Animal. 22(1):255-268.
Silva S.V. & Guerra M.M.P. 2011. Efeitos da criopreservação sobre as células espermáticas e
alternativas para redução das crioinjúrias. Revista Brasileira de Reprodução Animal.
35(4):370-384.
Sividanes V.P., Dutra F.M. & Mendonça P.P. 2012. Desenvolvimento embrionário da carpa
prateada, Hypophthalmichthys molitrix (Valenciennes, 1844). Revista Brasileira de Ciência
Veterinária. 19(1):21-25.
Sultana M., Nahiduzzaman M., Hassan M.M., Khanam M.U.H. & Hossain M.A.R. 2010.
Fertility of cryopreserved common carp (Cyprinus carpio) spermatozoa. University journal of
zoology. 28:51-55.

50
Suquet M., Dreanno C., Fauvel C., Cosson J. & Billard R. 2000. Cryopreservation of sperm in
marine fish. Aquaculture Research. 31(3):231-243.
Taitson P.F., Chami E. & Godinho H.P. 2007. Gene banking of the neotropical fish Leporinus
obtusidens (Valenciennes, 1836): a protocol to freeze its sperm in the field. Animal
reproduction science. 105(3-4):283-291.
Tiersch T.R., Yang H., Jenkins J.A. & Dong Q. 2007. Sperm cryopreservation in fish and
shellfish. Society of Reproduction and Fertility Supplement. 65:93–508.
Urbányi B., Szabó T., Miskolczi E., Mihálffy S., Vranovics K. & Horváth Á. 2006.
Successful fertilization and hatching of four European cyprinid species using cryopreserved
sperm. Journal of Applied Ichthyology. 22:201–204.
Viveiros A.T.M., Isaú Z.A., Caneppele D. & Leal M.C. 2012a. Sperm cryopreservation
affects postthaw motility, but not embryogenesis or larval growth in the Brazilian fish Brycon
insignis (Characiformes). Theriogenology. 78:803–810.
Viveiros A.T.M., Orfão L.H., Nascimento A.F., Corrêa F.M. & Caneppele D. 2012b. Effects
of extenders, cryoprotectants and freezing methods on sperm quality of the threatened
Brazilian freshwater fish pirapitinga-do-sul Brycon opalinus (Characiformes).
Theriogenology. 78:361–368.
Warnecke D. & Pluta H. 2003. Motility and fertilizing capacity of frozen/thawed common
carp (Cyprinus carpio L.) sperm using dimethyl-acetamide as the main cryoprotectant.
Aquaculture. 215:167–185.
Yang H. & Tiersch T. R. 2009. Current status of sperm cryopreservation in biomedical
research fish models: zebrafish, medaka and xiphophorus. Comparative Biochemistry and
Physiology - Part C: Toxicology & Pharmacology. 149(2):224–232.

51
8 CAPÍTULO 3
Efeito de vitaminas adicionadas ao diluente ACP-104 sobre a qualidade do sêmen
criopreservado de carpa comum (Cyprinus carpio)
Effect of vitamins added to the ACP-104 extender on the quality of cryopreserved semen
of common carp (Cyprinus carpio)
Periódico: Arquivo Brasileiro de Medicina Veterinária e Zootecnia (ISSN: 1678-4162)
Fator de impacto: 0,269 (Qualis A2)

52
RESUMO
O objetivo foi alcançado a partir da divisão da pesquisa em dois experimentos: (1) aperfeiçoar
o protocolo de congelação utilizando água de coco em pó (ACP-104) como diluente para a
criopreservação seminal de carpa comum; (2) avaliar o efeito da suplementação das vitaminas
C (ácido ascórbico) ou E (α-tocoferol) sobre os melhores diluidores testados no experimento 1
na qualidade do sêmen pós-descongelado da espécie. Para o experimento 1, foram formados
oito pools de sêmen provenientes de 14 machos selecionados. As amostras seminais coletadas
foram avaliadas quanto à motilidade total, à velocidade, ao percentual de espermatozoides
normais e à vitalidade espermática antes e depois da criopreservação seminal. Esta foi
realizada em meio ACP-104 acrescido de dimetilsulfóxido (DMSO), ou etilenoglicol (EG), ou
glicerol, ou metanol, todos à concentração de 10% diluídos em 1:3 (sêmen:diluidor). As
amostras foram então congeladas em vapor de nitrogênio líquido em dry shipper e estocadas
em nitrogênio líquido (-196 °C). Para o experimento 2, foram formados oito pools a partir da
coleta de sêmen de 15 machos. As amostras seminais foram avaliadas seguindo as mesmas
análises do experimento 1, acrescentando-se a duração da motilidade total. A criopreservação
seminal utilizou-se dos meios ACP-104 acrescido de DMSO ou EG, suplementados ou não
com vitamina C ou E. Os melhores resultados encontrados no experimento 1 foram obtidos
com o DMSO e o EG. Estes não diferiram significativamente entre si para a motilidade total
(24% e 28%; p>0,05) e normalidade espermática (32% e 26%; p>0,05) respectivamente. Para
o experimento 2, o EG suplementado com vitamina E produziu significativamente resultados
superiores de motilidade total, normalidade espermática e duração da motilidade em relação
ao DMSO, concluindo-se que o EG deve ser, portanto, o crioprotetor de escolha a ser
utilizado com o ACP-104 suplementado ou não com vitamina E.
Palavras-chave: Cyprinus carpio, sêmen, criopreservação, antioxidantes, ACP-104

53
Efeito de vitaminas adicionadas ao diluente ACP-104 sobre a qualidade do sêmen
criopreservado de carpa comum (Cyprinus carpio)
F. R. A. Linhares1*, M. S. Oliveira1, J. F. Nunes1, M. A. M. Carvalho1, C. C. Campello1, C. S.
B. Salmito-Vanderley1
1Universidade Estadual do Ceará – UECE – Fortaleza, CE
Abstract
The objective was achieved from the research division in two experiments: (1) improving the
freezing protocol using powdered coconut water (ACP-104) as a diluent for the
cryopreservation seminal of common carp; (2) evaluate the effect of supplementation of
vitamins C (ascorbic acid) or vitamin E (α-tocoferol) with the best extenders tested in
experiment 1 on the quality of post-thawed. For the experiment 1, semen pools from 14
selected males were formed. Seminal samples were evaluated for total motility, velocity,
percentage of normal sperm and sperm vitality before and after the seminal cryopreservation.
This was done in ACP-104 extender plus dimethyl sulfoxide (DMSO), or ethylene glycol
(EG), or glycerol or methanol all at concentration 10% diluted in 1:3 (semen:extender). The
samples were frozen in vapors of nitrogen into dry shipper and stored in liquid nitrogen (-196
°C). For the experiment 2, eight pools were formed from the 15 male. The semen samples
were evaluated following the same analysis of the experiment 1 adding duration of total
motility. The sperm cryopreservation was performed in extenders ACP-104 plus DMSO or
EG supplemented or not with vitamin C or E. The best results found in Experiment 1 were
obtained with DMSO and EG. They do not differ significantly for total motility (24% and
28%; p>0.05) and normal sperm (32% and 26%; p>0.05) respectively. For experiment 2, EG
supplemented with vitamin E, produced significantly better results overall motility, sperm
normality and duration of motility relative to DMSO. In conclusion the EG should be the
cryoprotectant of choice for use with the ACP-104 supplemented or not with vitamin E.
Keywords: Cyprinus carpio, sperm, cryopreservation, antioxidants, ACP-104

54
Introdução
O diluente ACP-104 preparado à base de água de coco vem sendo utilizado com
relativo sucesso na criopreservação do sêmen de peixes (Carvalho et al., 2014). No entanto
sua eficiência ainda é baixa, como diluente para a criopreservação, em termos de conservação
da motilidade e de velocidades espermáticas quando os resultados são comparados ao sêmen
in natura (Linhares et al., 2015). Esse baixo desempenho tem sido observado em outros
diluidores de criopreservação e foram correlacionados com o estresse térmico, osmótico e
oxidativo (Li et al., 2010). Desse modo são necessários novos estudos que possibilitem
aumentar a eficiência do meio ACP-104.
O estresse oxidativo é ocasionado pela geração excessiva de espécies reativas do
metabolismo do oxigênio conhecidas como ROS (O2-, OH•, HO2
• e H2O2) (Martinez-Páramo
et al., 2012). As ROS podem prejudicar a função espermática; reduzir viabilidade, motilidade
e integridade de membrana; aumentar a peroxidação lipídica e comprometer a atividade de
algumas enzimas após criopreservação (Słowińska et al., 2013). Para combater a produção
excessiva de ROS e o estresse oxidativo, o sêmen apresenta um sistema de defesa
antioxidante intra e extracelular formado por componentes enzimáticos e não enzimáticos
(LAHNSTEINER; MANSOUR, 2010). No entanto o sistema intracelular de defesa
antioxidante do espermatozoide é mais susceptível à ação das ROS, devido à escassez de
citoplasma, o qual é perdido durante a maturação celular (Carvalho et al., 2002). Alguns
estudos têm demonstrado que a adição de antioxidantes aos meios de congelação de sêmen de
peixes ajuda a proteger os espermatozoides, reduzindo os danos provocados pelo estresse
oxidativo (Kutluyer et al., 2014; Navarro et al., 2014).
As vitaminas C (ácido ascórbico) e E (α-tocoferol e derivados) são antioxidantes não
enzimáticos que protegem os espermatozoides de danos oxidativos do DNA e previnem a
deterioração oxidativa de lipídios poli-insaturados presentes na membrana que rodeia as
células e organelas celulares (Martinez-Páramo et al., 2012). Contudo não existem
publicações sobre a suplementação das vitaminas C ou E ao diluente ACP-104 quanto a sua
capacidade de aumentar a qualidade seminal e proteger os espermatozoides de carpa comum
do efeito tóxico dos radicais livres. Portanto o objetivo da presente pesquisa foi otimizar o
protocolo existente definindo inicialmente qual crioprotetor deve ser usado em associação
com o meio ACP-104 para a criopreservação seminal de carpa comum e posteriormente, após

55
seleção do crioprotetor, avaliar o efeito da suplementação da vitamina C ou E ao diluidor
escolhido sobre a qualidade do sêmen após a descongelação.
Material e métodos
A pesquisa foi aprovada pelo Comitê de Ética para o Uso de Animais da Universidade
Estadual do Ceará (CEUA/UECE; 11516665-3/63). Os animais utilizados pertenciam ao
Centro de Pesquisas em Aquicultura (CPAq) do Departamento Nacional de Obras Contra as
Secas (DNOCS) (Pentecoste - Ceará).
A pesquisa foi dividida em dois experimentos: (1) aperfeiçoar o protocolo de
congelação utilizando água de coco em pó (ACP-104) como diluente para a criopreservação
seminal de carpa comum; (2) avaliar o efeito da suplementação das vitaminas C e E sobre os
melhores crioprotetores selecionados no experimento 1 com base na qualidade do sêmen pós-
descongelação. A coleta de sêmen foi realizada em janeiro de 2015. No primeiro experimento,
o sêmen foi obtido a partir de 14 machos sexualmente maduros (46,36 ± 3,63 cm e 1,68 ±
0,41 Kg), que foram estimulados à espermiação após dose única de extrato hipofisário de
carpa (EHC; Danúbio Aquicultura Ltda., Brasil) de 1 mg/Kg de peso vivo. Os animais foram
previamente sedados em solução de eugenol (Cequímica Ltda., Brasil), diluída em álcool
absoluto e água (1:10:10000). Após 12 horas da indução hormonal, o sêmen foi colhido por
meio de massagem abdominal, evitando-se contaminação com água, fezes, urina ou sangue. O
sêmen foi coletado em tubos graduados e as amostras conservadas em caixas térmicas (~ 5
°C). O volume seminal (mL) foi aferido diretamente nos tubos; o pH, por meio de fitas de pH
(Merck-Alemanha) e a concentração espermática, por contagem em câmara de Neubauer. A
análise subjetiva da motilidade foi estimada com o auxílio de um microscópio ótico (Nikon
Eclipse E200 - ampliação de 100x) após a mistura de 2 µL de sêmen e 100 µL de água do
tanque. Considerando-se como 0% nenhum espermatozoide móvel e 100% todos os
espermatozoides móveis, somente as amostras com motilidade superior a 80% foram
utilizadas.
A partir das amostras selecionadas, foram formados oito pools, cada um constituído
pela combinação das amostras de três machos de forma aleatória com algumas outras carpas
contribuindo mais de uma vez para a formação dos pools. Cada pool foi dividido em quatro
alíquotas, diluído na proporção de 1:3 (sêmen:diluidor) em diluente ACP-104 (ACP
Biotecnologia, Fortaleza, Ceará, Brasil) e acrescido de dimetilsulfóxido (DMSO),

56
etilenoglicol (EG), glicerol ou metanol, todos na concentração de 10% (Cequímica Ltda.,
Brasil). Dessa forma, os quatro criodiluidores para cada pool foram: ACP-104 + DMSO;
ACP-104 + EG; ACP-104 + glicerol e ACP-104 + metanol. O grupo controle correspondeu ao
sêmen in natura. A solução crioprotetora foi preparada de acordo com Linhares et al. (2015).
No segundo experimento, verificou-se o efeito da suplementação com vitamina C ou E
nos melhores criodiluentes do experimento 1 sobre a qualidade seminal. Para o sêmen obtido
de 15 machos sexualmente maduros (52,27 ± 5,56 cm; 2,37 ± 0,80 Kg), seguiu-se o mesmo
procedimento: indução hormonal, coleta e preparação das amostras, congelação e avaliação
para ambos os experimentos. Cada pool (n=8) foi dividido em seis alíquotas e diluído na
proporção de 1:3 (sêmen:diluidor) em diluente ACP-104 acrescido de 10% de DMSO ou EG
suplementados ou não com vitamina C (0,0001 mg; ácido ascórbico) ou E (0,0001 mg; α-
tocoferol) conforme Navarro et al. (2014). Assim foram formados seis tratamentos: ACP-104
+ DMSO; ACP-104 + DMSO + vitamina C; ACP-104 + DMSO + vitamina E; ACP-104 +
EG; ACP-104 + EG + vitamina C e ACP-104 + EG + vitamina E. O grupo controle
correspondeu ao sêmen in natura.
A criopreservação seminal realizada nos dois experimentos seguiu a seguinte
metodologia: os criodiluidores foram envasados em palhetas de 0,25 mL, distribuídas em
hastes de alumínio e congelados em dry shipper (Cryofarm) por dez minutos numa taxa de
resfriamento de aproximadamente 25-40 °C/min com estabilização gradual de -181 °C em 3
min. Em seguida as hastes de alumínio foram transferidas para um botijão com nitrogênio
líquido (-196 °C) e transportadas até o Laboratório de Biotecnologia da Reprodução de Peixes
do Núcleo Integrado de Biotecnologia (LBRP-NIB) da UECE (Fortaleza - Ceará). Após uma
semana de estocagem, as amostras seminais foram descongeladas em banho-maria a 25 °C
por 30 segundos e analisadas quanto aos parâmetros cinéticos (motilidade total e velocidades
espermáticas), à vitalidade e à morfologia espermática. No experimento 2, a análise da
duração da motilidade espermática foi acrescentada.
Por meio da análise objetiva computadorizada (CASA), o sêmen foi avaliado quanto
aos parâmetros de porcentagem de motilidade total (velocidade espermática >10 μm/s), de
velocidade curvilinear (VCL), de velocidade em linha reta (VSL) e de velocidade média do
percurso (VAP). Cada imagem (n=25) foi analisada utilizando-se a configuração padrão para
peixes estabelecida pelo Sperm Class Analyser software (SCATM 2005, Microptics, S.L.
versão 3.2.0, Barcelona, Espanha). Para ativação, 2 μL de sêmen foram misturados com 100

57
μL de NaCl (0,29%) em câmara de Makler® e as amostras foram analisadas em microscópio
ótico com contraste de fase (Nikon® H550S, ECLIPSE 50i, Japan, 400x) conectado a uma
vídeo câmera. As imagens foram gravadas de três a quatro segundos após a ativação.
A vitalidade espermática foi avaliada a partir de esfregaços corados (10 µL eosina:10
µL nigrosina:5 µL sêmen), contando-se 200 espermatozoides por lâmina em microscópio
ótico (400x). Considerou-se espermatozoides vivos com membrana intacta (cabeça não
corada) e espermatozoides mortos com ruptura na membrana plasmática (forte coloração
avermelhada).
Para a análise morfológica espermática, esfregaços realizados com sêmen fixado em
citrato formol 4% foram corados com rosa bengala conforme Miliorini et al. (2011),
registrando-se os espermatozoides normais (%) em microscópio ótico (400x). Foram
observados 200 espermatozoides (100 por lâmina).
A duração da motilidade (segundos) consistiu na observação dos espermatozoides em
microscópio ótico (400x; CASA), acionando-se o cronômetro desde o momento da ativação
do sêmen até que restassem somente 10% de células móveis.
No experimento 1, um total de 256 palhetas foram criopreservadas, resultantes da
combinação de 8 palhetas x 4 tratamentos x 8 pools. Para o experimento 2, foram
criopreservadas 384 palhetas, resultantes da combinação de 8 palhetas x 6 tratamentos x 8
pools.
Os dados foram inicialmente submetidos aos testes de Shapiro-Wilk e Bartlett para
confirmação dos requerimentos com o objetivo de realização da análise de variância
(distribuição normal dos resíduos e homocedasticidade, respectivamente). Nos casos em que
não houve homogeneidade de variâncias entre os tratamentos, considerando os dados brutos,
foram feitas tentativas de transformação (logarítmica, radicial e angular). Para algumas
variáveis, a transformação logarítmica resultou em enquadramento nos requerimentos da
ANOVA. A análise de variância foi então executada considerando um delineamento
inteiramente casualizado em arranjo fatorial 2 x 3, sendo dois tratamentos e três
possibilidades de inclusão de vitaminas (vitaminas C, E ou nenhuma vitamina). Nos casos em
que houve significância dos efeitos principais ou da interação, foi aplicado o teste de Student-
Newman-Keuls (SNK) para comparação das médias. Comparações com o grupo controle
foram feitas usando-se o teste de Dunnett. Em alguns casos, persistiu a homocedasticidade,

58
mesmo após as tentativas de transformação, e foi aplicado o teste não paramétrico de Kruskal-
Wallis. Em todas as situações, diferenças foram consideradas significativas quando p<0,05, e
os resultados foram apresentados como média ± desvio padrão.
Resultados
Para o experimento 1, as amostras do sêmen in natura apresentaram valor médio de
motilidade subjetiva igual a 96,50 ± 6,27%, concentração espermática média igual a 19,59 ±
5,85x109 espermatozoides/mL, volume médio de 7,54 ± 6,20 mL e pH médio de 8,25 ± 0,26.
Em todos os tratamentos, independente do crioprotetor utilizado e do acréscimo ou
não de vitaminas, o sêmen descongelado mostrou uma redução significativa (p<0,05) em
todos os parâmetros seminais avaliados (motilidade total, velocidades, percentual de
espermatozoides normais, vitalidade e duração da motilidade) em comparação ao sêmen in
natura (Tab. 1, 2, 3 e 4).
Quando o sêmen criopreservado com DMSO foi comparado ao EG, os resultados não
diferiram entre si (p>0,05). Contudo, ao serem comparados com o glicerol e metanol,
mostraram motilidade total significativamente superior (p<0,05; Tab. 1). Efeitos deletérios
sobre a qualidade seminal foram observados com o glicerol e metanol com apenas 2% e 11%
de motilidade total respectivamente. As velocidades espermáticas do sêmen criopreservado
em glicerol e metanol foram abaixo (p<0,05) dos demais crioprotetores (Tab. 1), quando
observou-se aglutinação dos espermatozoides logo após a descongelação das amostras com
estes dois crioprotetores.
O percentual de espermatozoides normais diminuiu em média 21% após a
criopreservação seminal ao comparar-se o sêmen in natura (Tab. 2). Apesar do alto percentual
de espermatozoides vivos (~ 92%) no grupo controle, apenas cerca de 47% de
espermatozoides foram normais, influenciando negativamente os resultados do sêmen
descongelado. Considerando-se como critério principal a motilidade total, seguido da
velocidade e da normalidade espermática, os melhores tratamentos em ordem decrescente
foram EG, DMSO, metanol e glicerol (Tab. 1 e 2).
Tabela 1. Efeito de diferentes tipos de crioprotetores sobre os parâmetros cinéticos de espermatozoides frescos

59
(grupo controle) e descongelados de Cyprinus carpio criopreservados em ACP-104 sem vitaminas.
Crioprotetor
Parâmetros cinéticos
Motilidade total (%) VCL (μm/s) VSL (μm/s) VAP (μm/s)
Controle 95,06±4,34 86,77±21,27 37,51±15,09 68,54±19,70
DMSO 24,38±15,37*a 37,68±3,95*a 22,08±3,76*a 31,30±4,81*a
EG 28,30±15,21*a 41,70±5,46*a 25,95±5,19*a 36,10±5,11*a
Glicerol 2,65±3,72*c 15,39±16,65*c 6,55±7,22*c 11,14±12,11*c
Metanol 11,05±9,48*b 34,50±4,56*b 17,05±4,30*b 26,99±4,73*b
*Representa diferença significativa em relação ao controle. Letras minúsculas distintas representam diferenças
significativas entre linhas. O controle corresponde ao sêmen in natura. ACP- 104: água de coco em pó para
sêmen de teleósteos de água doce de fertilização externa; DMSO: dimetilsulfóxido; EG: etilenoglicol; VCL:
velocidade curvilinear; VSL: velocidade retilinear; VAP: velocidade média do percurso.
Tabela 2. Efeito de diferentes tipos de crioprotetores sobre o percentual de espermatozoides normais e
vitalidade de espermatozoides frescos (grupo controle) e descongelados de Cyprinus carpio
criopreservados em ACP-104 sem vitaminas.
Crioprotetor Espermatozoides normais (%) Vitalidade (%)
Controle 46,62 ± 13,44 91,75 ± 3,23
DMSO 31,94 ± 6,97*a 37,16 ± 10,76*a
EG 26,44 ± 6,92*ab 25,71 ± 7,39*b
Glicerol 20,50 ± 8,36*b 28,05 ± 14,78*ab
Metanol 25,06 ± 6,79*ab 28,09 ± 8,99*ab
*Representa diferença significativa em relação ao controle. Letras minúsculas distintas representam
diferenças significativas entre linhas. O controle corresponde ao sêmen in natura. ACP-104: água de
coco em pó para sêmen de teleósteos de água doce de fertilização externa; DMSO: dimetilsulfóxido; EG:
etilenoglicol.
Para o segundo experimento, as amostras seminais coletadas apresentaram taxa média
de motilidade subjetiva de 90,13 ± 22,92%, concentração espermática média de 27,36 ±
9,83x109 espermatozoides/mL, volume médio de 6,65 ± 6,97 mL e pH médio de 7,23 ± 1,61.
Os crioprotetores DMSO e EG foram, então, selecionados para o segundo experimento
para se avaliar o efeito da suplementação de vitamina C ou E no meio ACP-104 contendo
DMSO ou EG sobre a qualidade espermática.
O acréscimo de vitamina C ou E ao EG não teve nenhum efeito quando comparado ao
tratamento sem vitamina (p>0,05; Tab. 3). Contudo a adição de vitamina C ao DMSO
aumentou a taxa de motilidade total em 11% quando comparado ao tratamento sem vitamina e
em 17% quando comparado ao tratamento com vitamina E (Tab. 3). Independente do
acréscimo ou não de vitaminas, o EG foi o crioprotetor que obteve maiores índices de
normalidades espermáticas. Dentro dos tratamentos envolvendo o DMSO, o acréscimo de
vitamina C proporcionou maiores índices de normalidade espermática, enquanto o acréscimo
de vitamina E não produziu um efeito benéfico na concentração testada (Tab. 3).

60
Tabela 3. Efeito de diferentes tipos de crioprotetores acrescidos de vitaminas sobre os parâmetros de motilidade
total (%), espermatozoides morfologicamente normais (%) e vitalidade espermática (%) de espermatozoides
frescos (grupo controle) e descongelados de Cyprinus carpio criopreservados em ACP-104.
Parâmetros
avaliados
Controle Crioprotetor Vitaminas
Sem vitamina C E
Motilidade total 96,14 ± 5,89 DMSO 34,63 ± 6,29*Bb 46,04 ± 9,94*Aa 28,87 ± 7,44*Bb
EG 48,22 ± 9,46*Aa 53,93 ± 12,83*Aa 52,50 ± 15,27*Aa
Espermatozoides
normais
83,71 ± 13,29 DMSO 32,00 ± 6,90*Ab 40,17 ± 5,67*Aa 21,83 ± 5,08*Bc
EG 57,17 ± 14,85*Aa 49,50 ± 7,48*Aa 53,67 ± 18,29*Aa
Vitalidade 97,44 ± 2,24 DMSO 47,12 ± 7,12*Ba 67,33 ± 12,69*Aa 74,50 ± 11,29*Aa
EG 36,75 ± 7,74*Bb 48,58 ±
8,12*ABb
61,08 ±
22,50*Aab
*Representa diferença significativa em relação ao controle. Letras maiúsculas distintas representam diferenças
significativas entre colunas dentro de cada parâmetro analisado (motilidade total, espermatozoides normais e
vitalidade). Letras minúsculas distintas representam diferenças significativas entre linhas dentro de cada
parâmetro analisado (motilidade total, espermatozoides normais e vitalidade). O controle corresponde ao sêmen
in natura. ACP-104: água de coco em pó para sêmen de teleósteos de água doce de fertilização externa; DMSO:
dimetilsulfóxido; EG: etilenoglicol.
Quanto à vitalidade espermática, o número de células viáveis, após o acréscimo de
vitamina C ou E, foi superior ao tratamento sem vitamina usando-se o DMSO como o
crioprotetor (Tab. 3; p<0,05). O uso do EG acrescido de vitamina E foi superior quando
comparado ao tratamento sem vitamina ou acrescido de vitamina C (p<0,05).
O crioprotetor EG foi capaz de manter a duração da motilidade espermática constante,
independente do acréscimo ou não de vitamina C ou E (Tab. 4), não havendo diferença
significativa (p>0,05).
Tabela 4. Efeito de diferentes tipos de crioprotetores acrescidos de vitaminas sobre a duração da
motilidade (segundos) de espermatozoides frescos (grupo controle) e descongelados de Cyprinus
carpio criopreservados em ACP-104.
Controle 168,14 ± 55,34
Crioprotetor Vitaminas
Sem vitamina C E
DMSO 65,57 ± 10,77*ABa 82,37 ± 25,66*Aa 50,67 ± 20,22*Ba
EG 81,67 ± 30,60*Aa 90,83 ± 9,64*Aa 70,54 ± 11,50*Aa
*Representa diferença significativa em relação ao controle. Letras maiúsculas distintas representam
diferenças significativas entre colunas. Letras minúsculas distintas representam diferenças
significativas entre linhas. O controle corresponde ao sêmen in natura. ACP- 104: água de coco em pó
para sêmen de teleósteos de água doce de fertilização externa; DMSO: dimetilsulfóxido; EG:
etilenoglicol.
Foi observado que a duração da motilidade reduziu em 32 segundos após o acréscimo
de vitamina E ao DMSO (50,67 ± 20,22%) quando comparada ao tratamento acrescido de
vitamina C (82,37 ± 25,66), havendo diferença significativa (p<0,05) entre os dois

61
tratamentos (Tab. 4). No presente estudo, a dose de 0,0001 mg utilizada para a vitamina E
combinada com DMSO 10% não foi benéfica.
Discussão
Este é o primeiro trabalho que avalia a interação do diluente ACP-104 com quatro
diferentes crioprotetores na criopreservação do sêmen de carpa comum. Além disso avalia de
forma original o efeito da suplementação de vitamina C ou E ao meio ACP-104 contendo
DMSO ou EG.
A redução da qualidade seminal após criopreservação, já observada no sêmen
descongelado de várias espécies, vem sendo atribuída principalmente a fatores como estresses
osmótico, térmico e oxidativo (Martinez-Páramo et al., 2012). Durante o processo de
criopreservação, os espermatozoides são submetidos à diluição do plasma seminal e a
temperaturas negativas em curto espaço de tempo. A diluição tem, assim, uma ação
multifatorial sobre a resistência dos espermatozoides contra lesões do congelamento (Akçay
et al., 2004), que dependem da taxa de diluição e da interação entre a composição química do
diluente com o tipo e a concentração de crioprotetor utilizada (Viveiros et al., 2012),
produzindo resultados diferenciados em relação à espécie. Além disso a diluição seminal
reduz a concentração de antioxidantes presente no plasma seminal, dificultando o combate aos
radicais livres referencias. Isso favorece a perda da qualidade seminal devido a danos
celulares, tais como alterações na estrutura da bicamada lipídica da membrana espermática,
baixa regulação de proteínas de membrana e enzimas citoplasmáticas, perda da integridade de
membrana e fragmentação de DNA (Li et al., 2010).
Os crioprotetores DMSO e metanol foram utilizados na presente pesquisa por serem
comumente utilizados para o sêmen de carpa (Salmito-Vanderley et al., 2014). O EG, ainda
pouco utilizado na criopreservação de sêmen da espécie estudada, foi escolhido com base nos
estudos de Li et al. (2013) por produzir menos danos osmóticos e oxidativos quando
comparado ao DMSO. Embora ambos alterem o estado de fosforilação das proteínas sobre os
resíduos de tirosina e treonina (Li et al., 2013), o EG tem sido menos prejudicial a este efeito
(Li et al., 2010; Li et al., 2013). O glicerol é pouco empregado na criopreservação do sêmen
de peixes, principalmente na concentração de 10%, pois tem elevada pressão osmótica e
viscosidade, o que promove uma aglutinação espermática e injúrias celulares irreversíveis

62
(Irawan et al., 2010). Entretanto foi escolhido para que fosse investigada a interação entre ele
e o diluente ACP-104.
A adição de antioxidantes aos diluidores de criopreservação com o intuito de melhorar
a qualidade espermática após a criopreservação vem sendo objeto de estudos por diferentes
grupos de pesquisas no mundo (Kutluyer et al., 2014; Navarro et al., 2014). Alguns destes
trabalhos têm revelado o efeito positivo de certos antioxidantes sobre a redução dos níveis de
ROS e sobre a motilidade total, a qual melhora; na duração da motilidade e na redução das
anormalidades espermáticas. Contudo este efeito positivo depende do tipo e da concentração
do antioxidante (Cao e Cutler, 1997).
As vitaminas C e E são conhecidas como potentes antioxidantes capazes de eliminar
os radicais livres e seus efeitos negativos. A vitamina C interage com a vitamina E e com o
selênio na manutenção da atividade de algumas enzimas como a superóxido dismutase (SOD)
e a glutationa peroxidase que são importantes para a eliminação de radicais oxidantes
produzidos pelo metabolismo espermático (Navarro et al., 2009). Entretanto a interação da
vitamina C com E juntamente com o ACP-104 não foi investigada no presente estudo.
A água de coco verde, que é base para o diluente ACP-104, é rica em aminoácidos,
vitaminas, sais minerais, carboidratos, além de possuir baixo teor de lipídios saturados e
insaturados (Carvalho et al., 2006). A vitamina C, presente no coco verde, é capaz de
proporcionar uma ação anti-inflamatória e antioxidante ao eliminar diretamente os radicais
livres e inibir a produção in vitro de substâncias reativas ao ácido tiobarbitúrico (TBARS)
(Carvalho et al., 2006; Santos et al., 2013), destacando assim a importância da água de coco
como uma alternativa no combate as ROS.
O processo de liofilização da água de coco reduz drasticamente os níveis de seus
compostos, muito embora a concentração de ácido ascórbico se mantenha constante segundo
Santos et al. (2013). No produto ACP, após a liofilização, detectou-se para cada 100 g do pó,
16,51 mg de vitamina C (Carvalho, 2013). Todavia o volume de diluente ACP, utilizado nesse
estudo, continha apenas 0,034 mg de vitamina C para cada mL de sêmen criopreservado,
sugerindo-se que a concentração de ácido ascórbico tanto do diluente quanto da
suplementação pode ter sido insuficiente para atuar na eliminação dos ROS. Deste modo é
necessário investigar concentrações mais elevadas de vitaminas C e E e suas interações com
outros antioxidantes na suplementação do meio ACP-104 capazes de favorecer um aumento
significativo na qualidade do sêmen pós-descongelado que seja comparável ao sêmen fresco.

63
Outro ponto a ser levado em consideração é o estresse que os peixes podem ter sofrido
durante o período de estiagem que assola a região onde foram realizados os experimentos
(CPAq, DNOCS – Pentecoste - CE). A crise hídrica vem diminuindo a níveis críticos a água
dos reservatórios (FUNCEME, 2015), o que pode ser um dos fatores que provocam alterações
na qualidade de água (Linhares et al., 2013) e na maturação dos gametas, justificando as
baixas taxas de velocidades dos espermatozoides e o alto percentual de anormalidades
espermáticas encontrados na pesquisa. Apesar disso a morfologia dos espermatozoides está
dentro dos padrões de normalidade exigidos para outros animais pelo Colégio Brasileiro de
Reprodução Animal (2013).
A duração da motilidade espermática é considerada um parâmetro importante a ser
avaliado, pois os espermatozoides necessitam de um tempo para encontrar a micrópila do
ovócito e fertilizá-la. Na presente pesquisa, o EG livre ou não de vitaminas e o DMSO
adicionado à vitamina C foram os tratamentos que apresentaram duração de motilidade
suficiente para os espermatozoides encontrarem a micrópila.
Conclusão
Conclui-se que o EG suplementado ou não com vitamina E seja o crioprotetor de
escolha a ser utilizado com o meio ACP-104 para criopreservar espermatozoides de carpa, por
apresentar melhor desempenho em termos de motilidade, de normalidade, de vitalidade e de
duração da motilidade espermática.
Referências
AKÇAY, E.; BOZKURT, Y.; SEÇER, S.; TEKÜN, N. Cryopreservation of mirror carp
semen. Turk. J. Vet. Anim. Sci., v.28, p.837-843, 2004.
CAO, G.; CUTLER, R. G. High concentration of antioxidants may not improve defense
against oxidative stress. Arch. Gerontol. Geriatr., v.17, p.189-201, 1997.
CARVALHO, J.M.; MAIA, G.A.; SOUSA, P.H.M.; MAIA JR., G.A. Água-de-coco:
Propriedades nutricionais, funcionais e processamento. Semin., Ciênc. Agrár., v.27, n.3,
p.437-452, 2006.

64
CARVALHO, M.A.M. Água de coco em pó na criopreservação do sêmen de carpa comum,
Cyprinus carpio L. 2013. 123f. Tese (Doutorado em Biotecnologia) – Rede Nordeste de
Biotecnologia, Universidade Estadual do Ceará, Fortaleza, CE.
CARVALHO, M.A.M.; LINHARES, F.R.A.; NUNES, J.F. et al. Água de coco como diluente
para o sêmen de peixes de água doce de fertilização externa. Rev. Bras. Hig. San. Anim., v.08,
n.4, p.203-222, 2014.
CARVALHO, O.F.; FERREIRA, J.D.J.; SILVEIRA, N.A.; FRENEAU, G.E. Efeito oxidativo
do óxido nítrico e infertilidade no macho. J. Bras. Patol. Med. Lab., v.38, n.1, p.33-38, 2002.
FUNCEME (Fundação Cearense de Meteorologia e Recursos Hídricos). Calendário das
chuvas no estado do Ceará. Disponível em:
<http://www.funceme.br/app/calendario/produto/municipios/maxima/diario>. Acessado em: 1
jun. 2015.
IRAWAN, H.; VUTHIPHANDCHAI, V.; NIMRAT, S. The effect of extenders,
cryoprotectants and cryopreservation methods on common carp (Cyprinus carpio) sperm.
Anim. Reprod. Sci., v.122, p.236-243, 2010.
KUTLUYER, F.; KAYIM, M.; ÖĞRETMEN, F. et al. Cryopreservation of rainbow trout
Oncorhynchus mykiss spermatozoa: Effects of extender supplemented with different
antioxidants on sperm motility, velocity and fertility. Cryobiology, v.69, p.462-466, 2014.
LAHNSTEINER, F.; MANSOUR, N. A comparative study on antioxidant systems in semen
of species of the Percidae, Salmonidae, Cyprinidae, and Lotidae for improving semen storage
techniques. Aquaculture, v.307, p.130-140, 2010.
LI, P.; HULAK, M.; LI, Z.H. et al. Cryopreservation of common carp (Cyprinus carpio L.)
sperm induces protein phosphorylation in tyrosine and threonine residues. Theriogenology,
v.80, p.84-89, 2013.
LI, P.; LI, Z.H.; DZYUBA, B. et al. Evaluating the impacts of osmotic and oxidative stress on
common carp (Cyprinus carpio, L.) sperm caused by cryopreservation techniques. Biol.
Reprod., v.83, p.852-858, 2010.

65
LINHARES, F.R.A.; NUNES, L.T.; LOPES, J.T. et al. Influência da qualidade da água sobre
o desenvolvimento embrionário de carpa comum (Cyprinus carpio). Ciênc. Anim., v.23, n.2,
p.29-35, 2013.
LINHARES, F.R.A.; SALMITO-VANDERLEY, C.S.B.; CARVALHO, M.A.M. et al.
Cinética e morfologia de espermatozoides de carpa comum criopreservados em água de coco
em pó ACP-104. Arq. Bras. Med. Vet. Zootec., v.67, n.5, p.1313-1320, 2015.
MANUAL para exames andrológicos e avaliação de sêmen animal. 3ed. Belo Horizonte:
CBRA, 2013.
MARTINEZ-PÁRAMO, S.; DIOGO, P.; DINIS, M.T. et al. Incorporation of ascorbic acid
and α-tocopherol to the extender media to enhance antioxidant system of cryopreserved sea
bass sperm. Theriogenology, v.77, n.1-6, p.1129-1136, 2012.
MILIORINI, A.B.; MURGAS, L.D.S.; ROSA, P.V. et al. A morphological classification
proposal for curimba (Prochilodus lineatus) sperm damages after cryopreservation. Aquac.
Res., v.42, p.177-187, 2011.
NAVARRO, R.D.; NAVARRO, F.K.S.P.; FELIZARDO, V.O. et al. Semen quality of
curimba (Prochilodus lineatus) cryopreserved with vitamins. Acta Sci., Technol., v.36, n.1,
p.55-60, 2014.
NAVARRO, R.D.; RIBEIRO FILHO, O.P.; FERREIRA, W.M.; PEREIRA, F.K.S. A
importância da vitamina E, C e A na reprodução de peixes. Rev. Bras. Reprod. Anim., v.33,
n.1, p.20-25, 2009.
SALMITO-VANDERLEY, C.S.B.; LINHARES, F.R.A.; CARVALHO, M.A.M. et al.
Biotécnicas aplicadas a reprodução de ciprinídeos. Acta Vet. Bras., v.8, supl.2, p.292-298,
2014.
SANTOS, J.L.A.; BISPO, V.S.; FILHO, A.B.C. et al. Evaluation of chemical constituents and
antioxidant activity of coconut water (Cocus nucifera L.) and caffeic acid in cell culture. An.
Acad. Bras. Cienc., v.85, n.4, p.1235-1246, 2013.
SŁOWIŃSKA, M.; NYNCA, J.; CEJKO, B.I. et al. Total antioxidant capacity of fish seminal
plasma. Aquaculture, v.400-401, p.101-104, 2013.

66
VIVEIROS, A.T.M.; ORFÃO, L.H.; NASCIMENTO, A.F. et al. Effects of extenders,
cryoprotectants and freezing methods on sperm quality of the threatened Brazilian freshwater
fish pirapitinga-do-sul Brycon opalinus (Characiformes). Theriogenology, v.78, n.2, p.361-
368, 2012.

67
9 CAPÍTULO 4
Determinação da dose inseminante e embriogênese na fertilização assistida
de carpa comum no nordeste brasileiro
Determination of insemination dose and embryonic development in the artificial
fertilization of common carp in brazilian northeast
Periódico: Ciência Rural (ISSN: 1678-4596)
Fator de impacto: 0,383 (Qualis B1)

68
RESUMO
A presente pesquisa dividiu-se em duas etapas: a primeira visou determinar a dose
inseminante para fertilização assistida da carpa comum (Cyprinus carpio), e a segunda
objetivou o registro em imagens e tempo de desenvolvimento de cada fase embrionária desta
espécie em latitude equatorial. Para execução da primeira etapa, os gametas foram coletados
de reprodutores induzidos hormonalmente e fertilizados nas seguintes proporções de
espermatozoides/ovócito: D1-15.152, D2-60.491, D3-179.584, D4-238.072 e D5-298.563.
Para a segunda etapa, o acompanhamento do desenvolvimento embrionário foi realizado por
meio de observações periódicas em estereoscópio até a eclosão das larvas. A porcentagem de
fertilização aumentou de forma linear, atingindo um platô em 45,5% na proporção de 208.295
espermatozoides/ovócito. Observou-se que os parâmetros de qualidade de água
permaneceram constantes e dentro dos padrões para o desenvolvimento inicial desta espécie.
O desenvolvimento embrionário apresentou três períodos bem definidos, divididos em mórula
(2 horas), blástula (3 horas) e gástrula (5 horas). O início da eclosão ocorreu entre 42 horas e
30 minutos e 45 horas após a fertilização, o que equivale a um intervalo de 1165,14 e 1235,7
horas-grau. Assim, recomenda-se o uso da dose inseminante de aproximadamente 200.000
espermatozoides/ovócito na rotina de fertilização assistida dessa espécie no nordeste
brasileiro. Além disso, o desenvolvimento embrionário da carpa em latitude equatorial segue
a cronologia semelhante ao relatado para a espécie em clima temperado.
Palavras-chave: Cyprinus carpio, embriologia, espermatozoides/ovócito, reprodução.

69
Determinação da dose inseminante e embriogênese na fertilização assistida
de carpa comum no nordeste brasileiro
Francisco Renan Aragão LinharesI Jordana Sampaio LeiteI Liliane Veras LeiteI Priscila Silva
de AlmeidaI Rômulo Roberto Ribeiro PinheiroI Assis Rubens MontenegroII Carminda Sandra
Brito Salmito-VanderleyI*
I*Laboratório de Biotecnologia da Reprodução de Peixes, Programa de Pós-Graduação em
Ciências Veterinárias, Faculdade de Veterinária, Universidade Estadual do Ceará (UECE),
Fortaleza, CE, Brasil. E-mail: [email protected]. Autor para correspondência.
IIPrograma de Doutorado Integrado, Departamento de Zootecnia, Universidade Federal do
Ceará (UFC), Fortaleza, CE, Brasil.
Abstract
This research divided into two stages: the first aimed to determine the insemination
dose for artificial fertilization of common carp (Cyprinus carpio), and the second aimed to
record images and time in the development of each embryonic stage of the species in
equatorial latitude. For the implementation of the first stage, the gametes were collected from
induced breeding hormonally and fertilized in the following proportions of sperm/oocyte D1-
15152, D2-60491, D3-179584, D4-238072 and D5-298.563. For the second step, monitoring
of embryonic development was done through periodic observations in stereoscope until
hatching. The fertilization rate increased linearly up to the proportion of 208.295
spermatozoa/oocyte, and, from this point, the fertilization rate was maintained at 45,5%. It
was observed that the water quality parameters and remained constant within the standards for
the initial development of this species. The embryonic development showed three well
defined periods, divided into morula (2 hours), blastocyst (3 hours) and gastrula (5 hours).
The start of the hatching occurred between 42 hours and 30 minutes and 45 hours after
fertilization, which is equivalent to range of 1165.14 and 1235.7 hours-degree. Therefore, we
recommend the use of the insemination dose of approximately 200.000 sperm/oocyte in the
artificial fertilization routine of common carp in brazilian northeast. Moreover, embryonic
development of carp in equatorial latitude follows the chronology similar to that reported for
the species in temperate zones.
Key-words: Cyprinus carpio, embryology, spermatozoa/oocyte, reproduction.

70
Introdução
A carpa comum foi introduzida no Nordeste do Brasil por meio do Departamento
Nacional de Obras Contra as Secas (DNOCS) no ano de 1977 (DNOCS, 2009), e nos últimos
anos representa o quarto pescado da aquicultura continental mais produzido no país
(BOLETIM ESTATÍSTICO DA PESCA E AQUICULTURA, 2011). De acordo com o
mesmo boletim, o Nordeste representa a segunda maior produção aquícola continental de
pescado entre as regiões nacionais.
Nesse contexto, biotecnias aplicadas à fertilização assistida em peixes surgem como
ferramentas essenciais para auxiliar a crescente demanda de pescado no Brasil. A biotecnia de
fertilização assistida consiste em fertilizar o maior número de ovócitos com a menor
quantidade de espermatozoides, garantindo a máxima fertilidade com maior economia de
sêmen (LEITE et al., 2013). Porém, apesar da fertilização assistida de carpa comum ser
dominada (SALMITO-VANDERLEY et al., 2014), ainda são utilizados diferentes volumes
de sêmen por ovócito (dose inseminante), surgindo uma dúvida sobre a escolha da dose mais
eficiente para a realização da técnica.
Para avaliar o sucesso da fertilização assistida, torna-se necessário o acompanhamento
do desenvolvimento embrionário, visando a obtenção de embriões viáveis. Apesar de existir
relatos sobre o desenvolvimento embrionário da espécie em regiões de clima temperado
(NICA et al., 2012), o registro de imagens e a cronologia deste desenvolvimento para carpa
comum em latitude equatorial no Nordeste do Brasil não são relatados na literatura. A
compreensão destes eventos embrionários auxiliará na produção de novas linhagens de carpa
comum no Nordeste do Brasil.
Desta forma, os objetivos da presente pesquisa foram a determinação da dose
inseminante, ou a proporção de espermatozoide/ovócito, adequada para uso na fertilização
assistida de carpa comum (Cyprinus carpio) e o registro de imagens e do tempo de
desenvolvimento de cada fase embrionária para esta espécie em condições naturais de latitude
equatorial do nordeste do Brasil.
Material e métodos
A pesquisa foi aprovada pelo Comitê de Ética para o Uso de Animais da Universidade
Estadual do Ceará (CEUA/UECE; 11516665-3/63). O experimento foi realizado nos meses de
dezembro de 2014 e janeiro de 2015 no Centro de Pesquisas em Aquicultura (CPAq) do
DNOCS em Pentecoste - Ceará, situado a 3° 45' 00" de latitude sul e 39° 10' 24" de longitude
oeste.

71
O experimento foi dividido em duas etapas: (1) determinar a dose inseminante, ou
seja, a melhor proporção do número de espermatozoides por ovócitos; (2) registrar imagens e
acompanhar a cronologia do desenvolvimento embrionário. Para a etapa 1, o sêmen foi obtido
de 5 machos sexualmente maduros (43,5±2,13cm; 1,9±0,6kg) que receberam dose única de
extrato hipofisário de carpa (EHC; Danúbio Aquicultura Ltda., Brasil) de 1mg/kg de peso
vivo. Doze horas após a indução hormonal da reprodução, foi formado um pool do sêmen
coletado por meio de massagem abdominal.
O volume seminal foi aferido por meio de tubos graduados, o pH por fitas de pH
(MERCK-Alemanha) e a concentração espermática por contagem em câmara de Neubauer.
A análise subjetiva da motilidade (0 - 100%) foi estimada com o auxílio de um
microscópio ótico (Nikon Eclipse E200 - ampliação de 100x) após a adição de 100µL de água
do tanque a 2µL de sêmen. Os ovócitos foram obtidos de duas fêmeas sexualmente maduras
(53±8,49cm; 4,15±0,92kg) submetidas à indução da desova pela aplicação de duas doses de
EHC (0,5 e 5,0mg/kg, com intervalo de doze horas entre elas). Após a aplicação da segunda
dose hormonal, a temperatura da água do tanque foi verificada a cada hora para determinar o
momento da desova (tempo de latência) em horas-grau.
Foi formado um pool de ova do qual foram retiradas três amostras de 1g para estimar
a concentração de ovócitos. Do mesmo pool de ova, também foram retiradas 5 amostras de
25g, que foram fertilizadas com 20μL, 80μL, 237,5μL, 315μL ou 395μL do pool de sêmen em
quintuplicata. Essas quantidades de sêmen corresponderam às seguintes doses inseminanes:
15.152 (D1), 60.491 (D2), 179.584 (D3), 238.072 (D4) e 298.563 (D5) espermatozoides/
ovócito. Em seguida, utilizou-se o protocolo preconizado por BILLARD et al. (1995) para a
desagregação dos ovos. Estes logo foram transferidos para incubadoras cônicas de fluxo
constante com capacidade para 20L.
Para a segunda etapa experimental, exemplares de embriões foram observados em
microscópio ótico (ampliação de 40x) equipado com uma câmera fotográfica digital. A taxa
de fertilização foi calculada por meio da contagem de embriões em estágio de gástrula
dividido pelo total de ovócitos. A taxa de eclosão foi estimada quando pelo menos 50% das
larvas eclodiram, contando-se o número de larvas e dividindo pelo número total de ovos
fertilizados. Foram observados aleatoriamente 200 embriões de cada incubadora para ambos
os parâmetros avaliados.
Simultaneamente ao experimento, foram analisados temperatura, oxigênio dissolvido
(Phtek, modelo DO-100) e pH (KASVI, modelo K39-0014P) da água das incubadoras de três
em três horas, desde o início da fertilização assistida até as fases finais do desenvolvimento

72
embrionário. Também foram analisados os parâmetros de alcalinidade, de dureza, de amônia e
de nitrito conforme recomendações do Guia da Companhia de Tecnologia de Saneamento
Ambiental e analisadas pela Superintendência Estadual do Meio Ambiente do Ceará. A etapa
2 da pesquisa também foi realizada em quintuplicata, obedecendo as mesmas condições
experimentais.
Para as análises estatísticas, os dados foram incialmente submetidos aos testes de
Shapiro-Wilk e Bartlett para confirmação dos requerimentos para realização da análise de
variância (distribuição normal dos resíduos e homocedasticidade, respectivamente). A análise
de variância foi então executada, considerando um delineamento inteiramente casualizado, no
qual o efeito da quantidade de espermatozoides por ovócito foi avaliado. Nos casos em que
houve significância do efeito, foi aplicado o teste de Student-Newman-Keuls (SNK) para
comparação das médias. Aplicou-se o modelo de regressão segmentada ou descontínua
(Modelo “Broken Line” ou “Linear Response Plateau”) por meio do procedimento NLIN do
programa SAS (2002).
Resultados e discussão
Os valores médios de pH e volume seminais e de concentração e motilidade
espermáticas registrados foram 7,33±0,29, 9,19±2,84mL, 26,6±2,55x109 espermatozoides mL-
1 e 95%, respectivamente. A quantidade mínima e máxima de ova liberada foi de 111 e 638g,
respectivamente, com uma concentração média de 1408±146,5 ovócitos por grama de ova. As
fêmeas desovaram após 196,75±8,13 horas-grau do final do tratamento hormonal,
correspondendo a um tempo de latência de 7 horas. Todos estes valores foram semelhantes
aos parâmetros observados na literatura para carpa comum (IRAWAN et al., 2010).
No que diz respeito à etapa 1, foi verificado que as taxas de fertilização aumentaram
gradativamente até a proporção de 208.295 espermatozoides/ovócito (Tabela 1). Quando a
proporção foi aumentada para 238.072 e 298.563 espermatozoides/ovócito, a porcentagem de
ovócitos fertilizados permaneceu constante.
Tabela 1 - Taxa de fertilização e eclosão (%) de ovos de carpa comum (Cyprinus carpio) fertilizados com
diferentes proporções de espermatozoides (sptzs) por ovócito.
Dose inseminante Proporção sptzs/
ovócito
Taxa de
fertilização±DP (%)
Taxa de eclosão±DP
(%)
D1 15.152 13,50±2,09a 83,80±13,34
D2 60.491 22,50±4,91b 65,00±15,05
D3 179.584 40,50±2,12c 83,25±10,68
D4 238.072 45,80±7,12c 70,80±12,75
D5 298.563 46,60±3,49c 71,40±9,13
Letras minúsculas sobrescritas distintas representam diferenças significativas entre linhas.
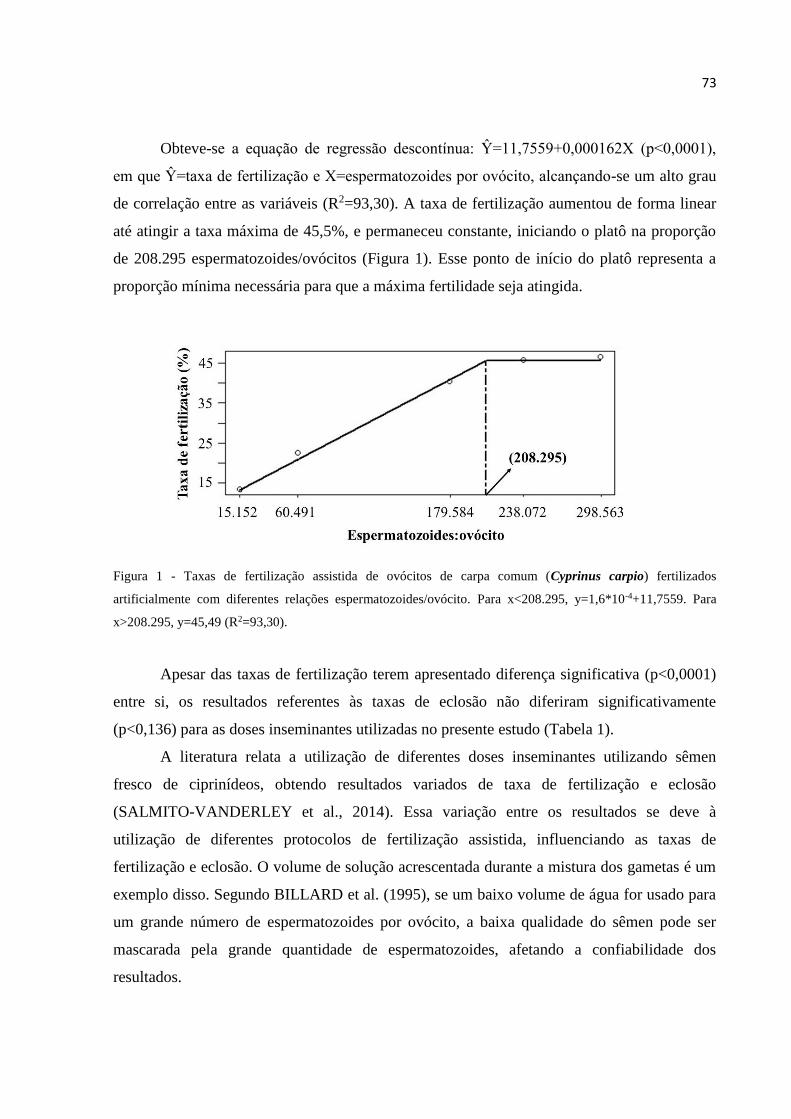
73
Obteve-se a equação de regressão descontínua: Ŷ=11,7559+0,000162X (p<0,0001),
em que Ŷ=taxa de fertilização e X=espermatozoides por ovócito, alcançando-se um alto grau
de correlação entre as variáveis (R2=93,30). A taxa de fertilização aumentou de forma linear
até atingir a taxa máxima de 45,5%, e permaneceu constante, iniciando o platô na proporção
de 208.295 espermatozoides/ovócitos (Figura 1). Esse ponto de início do platô representa a
proporção mínima necessária para que a máxima fertilidade seja atingida.
Figura 1 - Taxas de fertilização assistida de ovócitos de carpa comum (Cyprinus carpio) fertilizados
artificialmente com diferentes relações espermatozoides/ovócito. Para x<208.295, y=1,6*10-4+11,7559. Para
x>208.295, y=45,49 (R2=93,30).
Apesar das taxas de fertilização terem apresentado diferença significativa (p<0,0001)
entre si, os resultados referentes às taxas de eclosão não diferiram significativamente
(p<0,136) para as doses inseminantes utilizadas no presente estudo (Tabela 1).
A literatura relata a utilização de diferentes doses inseminantes utilizando sêmen
fresco de ciprinídeos, obtendo resultados variados de taxa de fertilização e eclosão
(SALMITO-VANDERLEY et al., 2014). Essa variação entre os resultados se deve à
utilização de diferentes protocolos de fertilização assistida, influenciando as taxas de
fertilização e eclosão. O volume de solução acrescentada durante a mistura dos gametas é um
exemplo disso. Segundo BILLARD et al. (1995), se um baixo volume de água for usado para
um grande número de espermatozoides por ovócito, a baixa qualidade do sêmen pode ser
mascarada pela grande quantidade de espermatozoides, afetando a confiabilidade dos
resultados.

74
Outro ponto ligado a variação do protocolo de fertilização assistida é a quantidade
excessiva de espermatozoides durante o ajuste da proporção espermatozoides/ovócito. O
aumento desta proporção pode ocasionar acúmulo de espermatozoides na micrópila, gerando
poliespermia e/ou competição entre os espermatozoides (KWANTONG & BART, 2009). A
razão ótima de espermatozoides/ovócito verificadas no presente estudo foi de 208.295. No
entanto, o aumento da proporção de espermatozoides/ovócito para 298.563 manteve-se no
platô de fertilização (45,5%), não sendo observado efeito do excesso de espermatozoides de
carpa comum sobre as taxas de fertilização.
Além dos fatores nutricionais, sociais, idade, fisiologia, manejo, estresse e etc., a
qualidade da água também pode influenciar na qualidade dos gametas, interferindo nos
resultados de dose inseminante (VALDEBENITO et al., 2013). Apesar disso, a variação da
temperatura da água (26 a 29,4°C) durante o experimento não prejudicou as taxas de eclosão
encontradas no presente estudo, não atingindo temperaturas iguais ou acima de 30°C. De fato,
segundo SAPKALE et al. (2011), só há diminuição da taxa de eclosão quando há um aumento
da temperatura de 26 para 30°C. Além disso, os teores de oxigênio dissolvido na água (6,10 a
8,61mg L-1 O2), alcalinidade (160 a 166mg L-1 de CaCO3), concentração de amônia (0,19 a
0,82mg L-1) e nitrito (0,018 e 0,107mg L-1) se mantiveram dentro da faixa recomendada por
BORYSHPOLETS et al. (2009), TAVARES (1995), LUKOWICZ (1982) e ARANA (1997)
respectivamente. Apesar dos valores encontrados para a dureza (273,6 a 284mg L-1 de
CaCO3) se apresentarem acima dos parâmetros admissíveis (30 a 100mg L-1 de CaCO3), o pH
médio (7,5) não oscilou, mantendo-se dentro do aceitável (SAPKALE et al., 2011).
No que diz respeito à segunda etapa experimental, observou-se que as imagens
geradas durante o acompanhamento do desenvolvimento embrionário se enquadraram no
padrão de clivagem meroblástica discoidal, típica de ovos telolécitos (LEITE et al., 2013).
Pôde-se observar que a formação da blástula foi evidenciada com uma média de 3
horas após a fertilização (~82,61 horas-grau) (Figura 2D). Nessa fase observou-se um
abaulamento do polo animal. A fase de gástrula foi verificada entre 5 horas e 5 horas e 30
minutos após a fertilização (137,3 e 151,86 horas-grau) (Figura 2E). Essa fase foi
caracterizada pela movimentação de células do polo animal para o polo vegetal, recobrindo
gradativamente a região vitelínica. Conforme as células se movimentavam, a fase de gástrula
avançou para as fases de 50%, 80% e 90% de recobrimento do vitelo, culminando com o
fechamento do blastóporo. Esta última fase ocorreu em até 8 horas após a fertilização
(~220,28 horas-grau) (Figura 2F, G, H e I).

75
Figura 2 - Estágios do desenvolvimento embrionário de carpa comum (Cyprinus carpio). Primeiras
segmentações (A e B), formação de mórula (C), formação de blástula (D), gástrula inicial (E), 50% de
gastrulação (F), 80% de gastrulação (G), 90% de gastrulação (H), fechamento do blastóporo (I), diferenciação do
embrião, da vesícula ótica (vo) e formação dos somitos (so) (J), região caudal destacada do vitelo (K), embrião
com movimentação dentro da membrana coriônica (L), eclosão (M). Aumento de 40x.
A diferenciação do embrião para um formato larval se deu entre 14 e 16 horas após a
fertilização (384,44 e 441,76 horas-grau), tornando-se notório o aparecimento dos somitos e o
início da encefalização, com o surgimento da vesícula ótica (Figura 2J). O início do
desprendimento da cauda do vitelo ocorreu entre 22 e 25 horas após a fertilização (604,12 e
690,25 horas-grau) (Figura 2K). Nessa fase, foram observados os movimentos de contração
muscular, tornando-se mais intensos com o avanço do desenvolvimento embrionário. O
embrião apresentou movimento de natação dentro da membrana de fertilização entre 40 e 43

76
horas após a fertilização (1104,4 e 1180,78 horas-grau) (Figura 2L), até o momento em que a
larva eclodiu e tornou-se livre. Mesmo antes da eclosão, já era possível visualizar o
aparecimento de pigmentação ao longo do embrião e o início da circulação sanguínea,
evidenciada pelos batimentos do coração. A eclosão das larvas ocorreu entre 42 horas e 30
minutos e 45 horas após a fertilização (1165,14 e 1235,7 horas-grau) (Figura 2M).
Os tempos dos registros embrionários e eclosão dependem principalmente da
temperatura da água de incubação dos embriões (SAPKALE et al., 2011). Porém, observou-se
que o tempo para se atingir o estágio de fechamento do blastóporo (220,28 horas-grau após a
fertilização) e de eclosão (1165,14 e 1235,7 horas-grau após a fertilização) a uma temperatura
média de 27,5°C no presente estudo foi próximo ao relatado por NICA et al. (2012) para
ciprinídeos em regiões temperadas (231 horas-grau; 1100 horas-grau, respectivamente, a
22ºC).
Conclusões
Diante do exposto, recomenda-se o uso da dose inseminante de aproximadamente
208.295 espermatozoides/ovócito para a fertilização assistida dessa espécie no nordeste
brasileiro, garantindo boa fertilidade e economia de gametas. Além disso, o desenvolvimento
embrionário da carpa, em latitude equatorial no Nordeste do Brasil, segue o padrão para ovos
telolécitos, tendo cronologia semelhante ao relatado para a espécie em clima temperado.
Salienta-se que as informações obtidas contribuirão para a realização de programas de
fertilização assistida da espécie no Nordeste do Brasil.
Referências
ARANA, L.V. Princípios químicos de qualidade da água em aquicultura: uma revisão
para peixes e camarões. Florianópolis: UFSC, 1997. 166p.
BILLARD, R. et al. Biology of sperm and artificial reproduction in carp. Aquaculture, v.129,
p.95-112, 1995. Disponível em:
<http://www.sciencedirect.com/science/article/pii/004484869400231C>. Acesso em: 13 fev.
2015. doi:10.1016/0044-8486(94)00231-C.

77
BORYSHPOLETS, S. et al. Freeze-thawing as the factor of spontaneous activation of
spermatozoa motility in common carp (Cyprinus carpio L.). Cryobiology, v.59, p.291-296,
2009. Disponível em:
<http://www.sciencedirect.com/science/article/pii/S0011224009001229>. Acesso em: 15 fev.
2015. doi:10.1016/j.cryobiol.2009.08.005.
DEPARTAMENTO NACIONAL DE OBRAS CONTRA AS SECAS. Relatório 2008.
Fortaleza, 2009. Acesso em: 09 fev. 2015. Online. Disponível em:
http://www.dnocs.gov.br/php/CGU/dnocs_relatorio_anual_2008.pdf
IRAWAN, H. et al. The effect of extenders, cryoprotectants and cryopreservation methods on
common carp (Cyprinus carpio) sperm. Animal Reproduction Science, v.122, p.236-243,
2010. Disponível em: <http://www.animalreproductionscience.com/article/S0378-
4320(10)00401-X/abstract>. Acesso em: 25 fev. 2015. doi:
10.1016/j.anireprosci.2010.08.017.
KWANTONG, S.; BART, A.N. Fertilization efficiency of cryopreserved sperm from striped
catfish, Pangasius hypophthalmus (Sauvage). Aquaculture Research, v.40, p.292-297,
2009. Disponível em: <http://onlinelibrary.wiley.com/doi/10.1111/j.1365-
2109.2008.02088.x/abstract;jsessionid=2E9EF1ACD1397E854634617C04FE6662.f04t03>.
Acesso em: 23 fev. 2015. doi: 10.1111/j.1365-2109.2008.02088.x.
LEITE, L.V. et al. Determinação da dose inseminante e embriogênese na fertilização artificial
de tambaqui (Colossoma macropomum). Arquivo Brasileiro de Medicina Veterinária e
Zootecnia, v.65, n.2, p.421-429, 2013. Disponível em:
<http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0102-
09352013000200018&lng=pt&nrm=iso&tlng=en>. Acesso em: 03 mar. 2015. doi:
10.1590/S0102-09352013000200018.
LINHART, O. et al. Common carp (Cyprinus carpio Linneaus, 1758) male fertilization
potency with secure number of spermatozoa per ova. Journal of Applied Ichthyology, v.31,
p.169-173, 2015. Disponível em:
<http://onlinelibrary.wiley.com/doi/10.1111/jai.12736/abstract>. Acesso em: 05 mar. 2015.
doi: 10.1111/jai.12736.
LUKOWICZ, M.V. Intensive carp Cyprinus carpio (L.) is rearing in a farm pond in southern
Germany and its effects on water quality. Aquaculture Engineers, v.1, n.2, p.121-137, 1982.
Disponível em: <http://www.sciencedirect.com/science/article/pii/0144860982900048>.
Acesso em: 10 mar. 2015. doi: 10.1016/0144-8609(82)90004-8.

78
MINISTÉRIO DA PESCA E AQUICULTURA. Boletim Estatístico da Pesca e
Aquicultura. 2011. Acesso: em 09 fev. 2015. Online. Disponível em:
http://www.icmbio.gov.br/cepsul/images/stories/biblioteca/download/estatistica/est_2011_bol
__bra.pdf.
NICA, A. et al. Embryonic and larval development of japanese ornamental carp Cyprinus
carpio (Linnaeus, 1758). Lucrări Ştiinţifice - Seria Zootehnie, v.58, p.116-120, 2012.
Disponível em: <http://www.univagro-
iasi.ro/revista_zoo/ro/documente/Pdf_Vol_58/Aurelia_Nica.pdf>. Acesso em: 10 mar. 2015.
SALMITO-VANDERLEY, C.S.B. et al. Biotécnicas aplicadas a reprodução de ciprinídeos.
Acta Veterinaria Brasilica, v.8, supl.2, p.292-298, 2014. Disponível em:
<http://periodicos.ufersa.edu.br/revistas/index.php/acta/article/view/3938>. Acesso em: 23
mar. 2015. doi: dx.doi.org/10.21708/avb.2014.8.0.3938
SAPKALE, P.H. et al. Optimal water temperature and pH for development of eggs and
growth of spawn of common carp (Cyprinus carpio). Journal of Applied Animal Research,
v.39, n.4, p.339-345, 2011. Disponível em:
<http://www.tandfonline.com/doi/abs/10.1080/09712119.2011.620269>. Acesso em: 18 mar.
2015. doi: 10.1080/09712119.2011.620269.
TAVARES, L.H.S. Limnologia aplicada a aquicultura. Jaboticabal: FUNEP, 1995. 70p.
VALDEBENITO, I.I. et al. Gamete quality in fish: evaluation parameters and determining
factors. Zygote, v.23, p.177-197, 2013. Disponível em:
<http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=9635466&fileI
d=S0967199413000506>. Acesso em: 22 mar. 2015. doi: 10.1017/S0967199413000506.

79
10 CONCLUSÕES
Recomenda-se o uso da dose inseminante de aproximadamente 208.295
espermatozoides/ovócito para a fertilização assistida dessa espécie no nordeste brasileiro,
garantindo boa fertilidade e economia de gametas. Além disso, o desenvolvimento
embrionário da carpa, em latitude equatorial no Nordeste do Brasil, segue o padrão para ovos
telolécitos, tendo cronologia semelhante ao relatado para a espécie em clima temperado.

80
11 PERSPECTIVAS
Na presente pesquisa, observou-se que o efeito da suplementação das vitaminas C ou
E ao meio ACP-104 sobre a qualidade do sêmen pós-descongelado de carpa comum não foi
comparável ao sêmen fresco. Deste modo, é necessário investigar concentrações mais
elevadas desses antioxidantes e suas interações entre si e com outros antioxidantes na
suplementação do meio ACP-104 capazes de favorecer um aumento significativo na
qualidade do sêmen pós-descongelado que seja comparável ao sêmen fresco.
Durante os experimentos para determinação da dose inseminante de carpa comum no
Centro de Pesquisas em Aquicultura (CPAq, DNOCS – Pentecoste - CE) foi constatada
problemas sérios na qualidade da água onde se encontravam os embriões em
desenvolvimento. Esse fator influenciou de algum modo alguns resultados referentes a
determinação da dose inseminante na presente pesquisa, como baixas taxas de velocidades
dos espermatozoides e o alto percentual de anormalidades espermáticas. Diante do exposto,
torna-se necessário a realização de futuras pesquisas com condições mais adequadas, visto
que a eutrofização do açude Pereira de Miranda pode prejudicar a reprodução de carpa
comum nas estações de piscicultura abastecidas pelo açude. Apesar disso, a pesquisa poderá
auxiliar na produção e introdução de novas linhagens de carpa comum via fertilização
assistida, contribuindo efetivamente para a consolidação da piscicultura comercial no
Nordeste do Brasil.

81
12 REFERÊNCIAS BIBLIOGRÁFICAS
AITKEN, R. J.; CLARKSON, J. S. Significance of reactive oxygen species and antioxidants
in defining the efficacy of sperm preparation techniques. Journal of Andrology, v. 6, n. 6, p.
367-376, 1988.
AKÇAY, E.; BOZKURT, Y.; SEÇER, S. et al. Cryopreservation of Mirror Carp Semen.
Turkish journal of veterinary and animal sciences, v. 28, p. 837-843, 2004.
ALVAREZ, B.; FUENTES, R.; PIMENTEL, R.; ABAD, Z.; CABRERA, E.; PIMENTEL, E.;
ARENAL, A. High fry production rates using post-thaw silver carp (Hypophthalmichthys
molitrix) spermatozoa under farming conditions. Aquaculture, v. 220, p. 195–201, 2003.
APHA - American Public Health Association. American Water Works Association, Water
Environment Federation. Standard Methods for The Examination of Water and Wastewater
Analysis. 21st ed. Washington, D.C.: American Public Health Association, 2005.
ARANA, L. V. Princípios químicos de qualidade da água em aquicultura: uma revisão para
peixes e camarões. Florianópolis: UFSC, 1997. 166p.
ARAÚJO, I. S. 1990. Utilização da água de coco e do leite glicosado como diluidor do sêmen
ovino. Fortaleza, 35p. Monografia (Especialização em Produção e Reprodução de Pequenos
Ruminantes da Universidade Estadual do Ceará - UECE, 1990).
ASHWOOD-SMITH, M. J. Low temperature preservation of cells, tissues and organs. In:
ASHWOOD-SMITH, M. J.; FARRANT, J. (eds.), Low temperature preservation in medicine
and biology, Turnbridge Wells: Pitman Medical Ltd, 1980. p. 19-44.
BALON, E. K. Origin and domestication of the wild carp, Cyprinus carpio: from Roman
gourmets to the swimming flowers. Aquaculture, v. 129, p. 3-48, 1995.
BARREIROS, A. L. B. S.; DAVID, J. M.; DAVID, J. P. Estresse oxidativo: relação entre
geração de espécies reativas e defesa do organismo. Química Nova, v. 29, n. 1, p. 113-123,
2006.

82
BILLARD, R.; COSSON, J.; PERCHEC, G.; LINHART, O. Biology of sperm and artificial
reproduction in carp. Aquaculture, v. 129, p. 95-l 12, 1995.
BLAXTER, J. H. S. Sperm storage and cross-fertilization of spring and autumn spawning
herring. Nature, v. 172, p. 1189-90, 1953.
BORYSHPOLETS, S.; DZYUBA, B.; RODINA, M.; LI, P.; HULAK, M.; GELA, D.;
LINHART O. Freeze-thawing as the factor of spontaneous activation of spermatozoa
motility in common carp (Cyprinus carpio L.). Cryobiology, v. 59, p. 291–296, 2009.
BRITTON, J. R.; BOAR, R. R.; GREY, J.; FOSTER, J.; LUGONZO, J.; HARPER, D. M.
From introduction to fishery dominance: the initial impacts of the invasive carp Cyprinus
carpio in Lake Naivasha, Kenya, 1999 to 2006. Journal of Fish Biology, v. 71, p.239-257,
2007.
BROMAGE, N.; PORTER, M.; RANDALL, C. The environmental regulation of maturation
in farmed finfish with special reference to the role of photoperiod and melatonin.
Aquaculture, v. 197, p. 63-98, 2001.
CABRINI, K. T.; GALLO, C. R. Avaliação da qualidade microbiológica de águas minerais
envasadas. Revista Higiene Alimentar, Mirandópolis, v. 15, n. 90-91, p. 83-92, 2001.
CARVALHO, J. M.; MAIA, G. A.; SOUSA, P. H. M.; MAIA JR., G. A. Água-de-coco:
Propriedades nutricionais, funcionais e processamento. Semina, Ciências Agrárias, v.27, n.3,
p.437-452, 2006.
CARVALHO, M. A. M. Água de coco em pó na criopreservação do sêmen de carpa comum,
Cyprinus carpio L. 2013. 123f. Tese (Doutorado em Biotecnologia) – Rede Nordeste de
Biotecnologia, Universidade Estadual do Ceará, Fortaleza, CE.
CARVALHO, M. A. M.; LINHARES, F. R. A.; NUNES, J. F.; SALGUEIRO, C. C. M.;
COSTA, R. B.; SALES, R. O. Água de coco como diluente para o sêmen de peixes de água

83
doce de fertilização externa. Revista Brasileira de Higiene e Sanidade Animal, v.08, n.4,
p.203-222, 2014.
CARVALHO, O. F.; FERREIRA, J. D. J.; SILVEIRA, N. A.; FRENEAU, G. E. Efeito
oxidativo do óxido nítrico e infertilidade no macho. Jornal Brasileiro de Patologia e
Medicina Laboratorial, v. 38, n. 1, p. 33-38, 2002.
CHAMBEYRON, F.; ZOHAR, Y. A diluent for sperm cryopreservation of gilthead seabream,
Sparus aurata. Aquaculture, v. 90, p. 345-352, 1990.
Companhia de Gestão dos Recursos Hídricos – COGERH. Qualidade da água do açude
pereira de miranda, ceará: avaliação e recomendações. Fortaleza, 2015. Acessado em 15
outubro de 2012. Disponível em: http://www.srh.ce.gov.br/publicacoes/artigos/Qualidade-
Agua-%20Acude-%20Pereira-de-Miranda-Ceara-%20Avaliacao-
%20Recomendacoes.pdf/view?searchterm=None.
CAO, G.; CUTLER, R. G. High concentration of antioxidants may not improve defense
against oxidative stress. Arch. Gerontol. Geriatr., v.17, p.189-201, 1997.
Departamento Nacional de Obras Contra as Secas. Relatório 2008. Fortaleza, 2009.
Disponível em: <http://www.dnocs.gov.br/php/CGU/dnocs_relatorio_anual_2008.pdf>.
Acessado em: 8 out. 2012.
ECHEVENGUÁ, M. M. Avaliação da qualidade da carne da carpa húngara (Cyprinus carpio)
de diferentes sistemas de cultivo da região Sul do Brasil. 2006. Dissertação (Mestrado) -
Universidade Federal do Rio Grande, Rio Grande do Sul.
FAO Yearbook, 2008. Fishery and Aquaculture Statistics, 2006. Food and agriculture
organization of the united nations, Rome, 57p.
FERREIRA, A. L. A.; MATSUBARA, J. S. Radicais livres: Conceitos, doenças relacionadas,
sistema de defesa e estresse oxidativo. Revista da Associação Médica Brasileira, v. 43, n. 1,
p. 61-68, 1997.

84
FROESE, R.; PAULY, D. (2007). FishBase: World Wide Web electronic publication.
Disponível em: <http://www.fishbase.org>. Acessado em: 23 ago. 2012.
FUNCEME (Fundação Cearense de Meteorologia e Recursos Hídricos). Calendário das
chuvas no estado do Ceará. Disponível em:
<http://www.funceme.br/app/calendario/produto/municipios/maxima/diario>. Acessado em: 1
jun. 2015.
GLASSER, F.; MIKOLAJCZYK, T.; JALABERT, B.; BAROILLER, J-F.; BRETON B.
Temperature effects along the reproductive axis during spawning induction of grass carp
(Ctenopharyngodon idella). General and Comparative Endocrinology, v. 136, p. 171-179,
2004.
GOLOMBIESKI, J. I.; MARCHEZAN, E.; MONTI, M. B.; STORCK, L.; CAMARGO, E.
R.; SANTOS, F. M. Qualidade da água no consórcio de peixes com arroz irrigado. Ciência
Rural, v. 35, n. 6, p. 1263-1268, 2005.
GRAEFF, A.; MONDARDO, M. Influência do probiótico no crescimento das carpas comum
(Cyprinus carpio l., 1758) na fase de recria. Revista Eletrônica de Veterinária, v.7, n.11,
2006.
GRAEFF, A.; PRUNER, E. N. Efeito da densidade de estocagem na produtividade final de
carpas, Cyprinus carpio Linnaeus, 1758 (var. specularis) na fase de engorda período -
inverno. Ciência e agrotecnologia, v. 23 (4), p. 958-967, 1999.
GUERRA, M. M. P.; EVANS, G.; MAXWELL, W. M. C. Papel de oxidantes e anti-oxidantes
na andrologia: Revisão de literatura. Revista Brasileira de Reprodução
Animal, v. 4, n. 4, p. 187-195, 2004.
GUTHRIE, H. D.; WELCH, G. R. Impact of storage prior to cyopreservation on plasma
menbrane function and fertility of boar sperm. Theriogenology, v. 63, p. 316-410, 2005.
HAFEZ, E. S. E. Reprodução animal. 6 ed. Barueri - SP: Manole, 1995, 582p.

85
HOLT, W. V. Fundamental aspects of sperm cryobiology: The importance of species and
individual differences. Theriogenology, v.53, p.47-58, 2000.
HORVÁTH, A.; MISKOLCZI, E.; URBÁNYI, B. Cryopreservation of common carp sperm.
Aquatic living resources, v. 16, p. 457-460, 2003.
HORVÁTH, A.; MISKOLCZI, E.; MIHALFFY, S.; OSZ, K.; SZABO, K.; URBANYI, B.
Cryopreservation of common carp (Cyprinus carpio) sperm in 1.2 and 5 ml straws and
occurrence of haploids among larvae produced with cryopreserved sperm. Cryobiology, v. 54,
p. 251–257, 2007.
IRAWAN, H.; VUTHIPHANDCHAI, V.; NIMRAT, S. The effect of extenders,
cryoprotectants and cryopreservation methods on common carp (Cyprinus carpio) sperm.
Animal reproduction science, v. 122, p. 236-243, 2010.
KUCHARCZYK, D.; TARGOŃSKA, K.; HLIWA, P.; GOMUŁKA, P.; KWIATKOWSKI,
M.; KREJSZEFF, S.; PERKOWSKI, J. Reproductive parameters of common carp (Cyprinus
carpio L) spawners during natural season and out-of-season spawning. Reproductive biology,
v. 8, n. 3, p. 285-289, 2008.
KUROKURA, H.; HIRANO, R.; TOMITA, M.; IWAHASHI, M. Cryopreservation of carp
sperm. Aquaculture, v. 37, p. 267-273, 1984.
KUTLUYER, F.; KAYIM, M.; ÖĞRETMEN, F. BÜYÜKLEBLEBICI, S.; TUNCER, P. B.
Cryopreservation of rainbow trout Oncorhynchus mykiss spermatozoa: Effects of extender
supplemented with different antioxidants on sperm motility, velocity and fertility.
Cryobiology, v.69, p.462-466, 2014.
KWANTONG, S.; BART, A. N. Fertilization efficiency of cryopreserved sperm from striped
catfish, Pangasius hypophthalmus (Sauvage). Aquaculture Research, v.40, p.292-297, 2009.
LAHNSTEINER, F.; BERGER, B.; HORVATH, A.; URBANYI, B.; WEISMANN, T.
Cryopreservation of spermatozoa in cyprinid fishes. Theriogenology, v. 54, p. 1477-1496,
2000.

86
LAHNSTEINER, F.; MANSOUR, N. A comparative study on antioxidant systems in semen
of species of the Percidae, Salmonidae, Cyprinidae, and Lotidae for improving semen storage
techniques. Aquaculture, v. 307, p. 130-140, 2010.
LEITE, L. V.; MELO, M. A. P.; OLIVEIRA, F. C. E.; PINHEIRO, J. P. S.; CAMPELLO, C.
C.; NUNES, J. F.; SALMITO-VANDERLEY, C. S. B. Determinação da dose inseminante e
embriogênese na fertilização artificial de tambaqui (Colossoma macropomum). Arquivo
Brasileiro de Medicina Veterinária e Zootecnia, v. 65, n. 2, p. 421-429, 2013.
LINHARES, F. R. A. Efeito das taxas de diluição e equipamentos de congelamento sobre a
cinética e morfologia de espermatozoides de carpa comum (Cyprinus carpio). 2012. 75f.
Dissertação (Mestrado) - Universidade Estadual do Ceará. Programa de Pós-Graduação em
Ciências Veterinárias, Universidade Estadual do Ceará, Fortaleza, Brasil.
LINHARES, F. R.A.; NUNES, L.T.; LOPES, J. T.; GALVÃO, J. A. S.; NUNES, J. F.;
SALMITOVANDERLEY, C. S. B. Influência da qualidade da água sobre o desenvolvimento
embrionário de carpa comum (Cyprinus carpio). Ciênc. Anim., v.23, n.2, p.29-35, 2013.
LINHARES, F. R. A.; SALMITO-VANDERLEY, C. S. B.; CARVALHO, M. A. M.;
PINHEIRO, R. R. R.; OLIVEIRA, F. C. E.; NUNES, J. F. Cinética e morfologia de
espermatozoides de carpa comum criopreservados em água de coco em pó ACP-104.
Arquivos Brasileiro de Medicina Veterinária e Zootecnia, v. 67, n.5, p. 1313-1320, 2015.
LI, P.; HULAK, M.; KOUBEK, P.; SULC, M.; DZYUBA, B.; BORYSHPOLETS, S.;
RODINA, M.; GELA, D.; MANASKOVA-POSTLEROVA, P.; PEKNICOVA, J.;
LINHART, O. Ice-age endurance: the effects of cryopreservation on proteins of sperm of
common carp, Cyprinus carpio L. Theriogenology, Aug; v. 74, n. 3, p. 413-23, 2010.
LI, P.; HULAK, M.; LI, Z. H.; SULC, M.; PSENICKA, M.; RODINA, M.; GELA, D.;
LINHART, O. Cryopreservation of common carp (Cyprinus carpio L.) sperm induces protein
phosphorylation in tyrosine and threonine residues. Theriogenology, v.80, p.84-89, 2013.

87
LINHART, O.; KUDOB, S.; BILLARD, R.; SLECHTAD, V.; MIKODINA, E. V.
Morphology, composition and fertilization of carp eggs: a review. Aquaculture, v. 129, p. 75-
93, 1995.
LINHART, O.; RODINA, M.; COSSON, J. Cryopreservation of Sperm in Common Carp
Cyprinus carpio: Sperm Motility and Hatching Success of Embryos. Cryobiology, v. 41, p.
241-250, 2000.
LINHART, O.; RODINA, M.; GELA, D.; KOCOUR, M.; RODRIGUEZ, M. Improvement of
common carp artificial reproduction using enzyme for elimination of egg stickiness. Aquatic
Living Resources, v. 16, p. 450–456, 2003.
LINHART, O.; RODINA, M.; KAŠPAR, V. Common carp (Cyprinus carpio Linneaus, 1758)
male fertilization potency with secure number of spermatozoa per ova. Journal of Applied
Ichthyology, v.31, p.169-173, 2015.
LUKOWICZ, M. V. Intensive carp Cyprinus carpio (L.) is rearing in a farm pond in southern
Germany and its effects on water quality. Aquaculture Engineers, v.1, n.2, p.121-137, 1982.
MACÊDO, H. R.; SILVA, A. J. N.; FERREIRA, D. M.; MARCELINO, J. F.; ARAÚJO, D.
M. (2007). Estudo de parâmetros físico-químicos para a criação de camarão marinho
Litopenaeus vannamei em água doce. In: II Congresso de Pesquisa e Inovação da Rede Norte
Nordeste de Educação Tecnológica. João Pessoa. Anais II CONNEPI, 4.
MAIA, M. S.; BICUDO, S. D. Radicais livres, antioxidantes e função espermática em
mamíferos: uma revisão. Revista Brasileira de Reprodução Animal, v. 33, n. 4, p. 183-193,
2009.
MANEESH, M.; JAYALEKSHMI, H. Role of reactive oxygenspecies and antioxidants on
pathophysiology of male reproduction. Indian Journal of Clinical Biochemistry, v. 21, n. 2, p.
80-89, 2006.
MANUAL para exames andrológicos e avaliação de sêmen animal. 3ed. Belo Horizonte:
CBRA, 2013.

88
MARTINEZ-PÁRAMO, S.; DIOGO, P.; DINIS, M.T.; HERRÁEZ, M. P.; SARASQUETE,
C.; CABRITA, E. Incorporation of ascorbic acid and α-tocopherol to the extender media to
enhance antioxidant system of cryopreserved sea bass sperm. Theriogenology, v.77, n.1-6,
p.1129-1136, 2012.
MILIORINI, A. B.; MURGAS, L. D. S.; ROSA, P. V.; OBERLENDER, G.; PEREIRA, G. J.
M; COSTA, D. V. A morphological classification proposal for curimba (Prochilodus
lineatus) sperm damages after cryopreservation. Aquac. Res., v.42, p.177-187, 2011.
MINISTÉRIO DA PESCA E AQUICULTURA. Boletim Estatístico da Pesca e Aquicultura
2011. Acessado em: 9 fevereiro 2013. Disponível em:
http://www.mpa.gov.br/files/docs/Boletim_MPA_2011_pub.pdf.
MONTANARI, T. 2013. Embriologia: texto, atlas e roteiro de aulas práticas [recurso
eletrônico]. 1ª ed. Ed. do autor, Porto Alegre. Capturado em 10 de out. 2013. Online.
Disponível na Internet http://www.ufrgs.br/livrodeembrio/
MOREIRA, H. L. M.; VARGAS, L.; RIBEIRO, R. P.; ZIMMERMANN, S. Fundamentos da
moderna aquicultura. Canoas: ULBRA, 2001, 200 p.
MORISAWA, M.; SUSUKI, K. Osmolality and potassium ion: their roles in initiation of
sperm motility in teleosts. Science, v. 210, p. 1145-1147, 1980.
NAHIDUZZAMAN; HASSANB, M.; ROYA, P. K.; HOSSAINC, A.; HOSSAINA, M. A.
R.; TIERSCH, T. R. Sperm cryopreservation of the Indian major carp, Labeo calbasu: Effects
of cryoprotectants, cooling rates and thawing rates on egg fertilization. Animal Reproduction
Science, v. 136, p. 133–138, 2012.
NAVARRO, R. D.; NAVARRO, F. K. S. P.; FELIZARDO, V. O.; MURGAS, L. D. S.;
ANDRADE, E. S. Semen quality of curimba (Prochilodus lineatus) cryopreserved with
vitamins. Acta Scientiarum. Technology, v. 36, n. 1, p. 55-60, 2014.

89
NAVARRO, R. D.; RIBEIRO FILHO, O. P.; FERREIRA, W. M.; PEREIRA, F. K. S. A
importância da vitamina E, C e A na reprodução de peixes. Rev. Bras. Reprod. Anim., v.33,
n.1, p.20-25, 2009.
NICA, A.; CRISTEA, V.; GHEORGHE, D.; HOHA, G. V.; ENACHE, I. B. Embryonic and
larval development of japanese ornamental carp Cyprinus carpio (Linnaeus, 1758). Lucrări
Ştiinţifice - Seria Zootehnie, v. 58, p. 116-120, 2012.
NUNES, J. F.; COMBARNOUS, Y. Utilização da água de coco e suas frações ativas como
diluente do sêmen de mamíferos domésticos. In: SIMPOSIO NACIONAL DE
BIOTECNOLOGIA DA REPRODUÇÃO DE MAMÍFEROS DOMÉSTICOS, 1995,
Fortaleza. Anais. Fortaleza.
ÖĞRETMEN F.; İNANAN B. E.; ÖZTÜRK M. Protective effects of propolis on
cryopreservation of common carp (Cyprinus carpio) sperm. Cryobiology, v. 68, p.107–112,
2014.
RANI, K. U.; MUNUSWAMY, N. Preliminary studies on the cryopreservation of
spermatozoa in the fresh water fish common carp (Cyprinus carpio L.). Journal of Coastal
Life Medicine, v. 2, n. 3, p. 181-186, 2014.
RURANGWA, E.; KIME, D. E.; OLLEVIER, F.; NASH, J. P. The measurement of sperm
motility and factors affecting sperm quality in cultured fish. Aquaculture, v. 234, p. 1-28,
2004.
SALMITO-VANDERLEY, C. S. B.; LINHARES, F. R. A.; CARVALHO, M. A. M.;
OLIVEIRA, M. S.; NUNES, J. F. Biotécnicas aplicadas a reprodução de ciprinídeos. Acta
Vet. Bras., v.8, supl.2, p.292-298, 2014.
SALMITO-VANDERLEY, C. S. B.; VIEIRA, M. J. A. F.; LEITE, L. V.; OLIVEIRA, F. C.
E.; LINHARES, F. R. A.; SALGUEIRO, C. C. M.; NUNES J. F. Meios de congelação para
conservação de sêmen de peixes da família Characidae. Ciência Animal, v. 22, n. 1, p. 255-
268, 2012.

90
SANOCKA, D.; KURPISZ, M. Reactive oxygen species and sperm cells. Reproductive
Biology and Endocrinology, v. 2, p. 12-18, 2004.
SANTOS, J. L. A.; BISPO, V. S.; FILHO, A. B. C.; PINTO, I. F. D.; DANTAS, L. S.;
VASCONCELOS, D. F.; ABREU, F. F.; MELO, D. A.; MATOS, I. A.; FREITAS, F. P.;
GOMES, O. F.; MEDEIROS, M. H. G.; MATOS, H. R. Evaluation of chemical constituents
and antioxidant activity of coconut water (Cocus nucifera L.) and caffeic acid in cell culture.
An. Acad. Bras. Cienc., v.85, n.4, p.1235-1246, 2013.
SAPKALE, P. H.; SINGH, R. K.; DESAI, A. S. Optimal water temperature and pH for
development of eggs and growth of spawn of common carp (Cyprinus carpio). Journal of
Applied Animal Research, v.39, n.4, p.339-345, 2011.
SIKKA, S. C. Role of oxidative stress and antioxidants in andrology and assisted reproductive
technology. Journal of Andrology, v. 25, n. 1, p. 5-18, 2004.
SILVA, N. J. R. Dinâmicas de desenvolvimento da piscicultura e políticas públicas no Vale
do Ribeira/SP e Alto Vale do Itajaí/SC – Brasil. 2005. 544f. Tese (Doutorado em aqüicultura
e à école nationale supérieure agronomique de rennes) - Universidade Estadual Paulista.
Curso de Pós-Graduação em Aqüicultura do Centro de Aqüicultura, Universidade Estadual
Paulista, Jaboticabal, Brasil.
SILVA, S. V.; GUERRA, M. M. P. Efeitos da criopreservação sobre as células espermáticas e
alternativas para redução das crioinjúrias. Revista Brasileira de Reprodução Animal, v. 35, n.
4, p. 370-384, 2011.
SILVEIRA, F. W.; KAVAMOTO, E. T.; RIGOLINO, M. G.; TABATA, Y. A. O método
espectrofotométrico na avaliação da concentração de espermatozoides da truta arco-íris,
Salmo irideus Gibbons. Boletim do Instituto Pesca, v.14, p. 69-73, 1987.
SIVAKUMARAN, K.; BROWN, P.; STOESSEL, D.; GILES, A. Maturation and
reproductive biology of female wild carp, Cyprinus carpio, in Victoria, Australia.
Environmental Biology of Fishes, v. 68, p. 321-332, 2003.

91
SIVIDANES, V. P.; DUTRA, F. M.; MENDONÇA, P. P. Desenvolvimento embrionário da
carpa prateada, Hypophthalmichthys molitrix (Valenciennes, 1844). Revista Brasileira de
Ciência Veterinária, v. 19, n. 1, p. 21-25, jan./abr. 2012.
SŁOWIŃSKA, M.; NYNCA, J.; CEJKO, B. I.; Dietrich, M. A.; HORVÁTH, Á.; URBÁNYI,
B.; KOTRIK, L.; CIERESZKO, A. Total antioxidant capacity of fish seminal plasma.
Aquaculture, v. 400-401, p. 101-104, 2013.
STRADAIOLI, G.; NORO, T.; SYLLA, L.; MONACI, M. Decrease in glutathione (GSH)
content in bovine sperm after cryopreservation: comparison between two extenders.
Theriogenology, v. 67, p. 1249-1255, 2007.
SULTANA, M.; NAHIDUZZAMAN, M.; HASSAN, M. M.; KHANAM, M. U. H.;
HOSSAIN, M. A. R. Fertility of cryopreserved common carp (Cyprinus carpio) spermatozoa.
University journal of zoology, v.28, p.51-55, 2010.
SUQUET, M.; DREANNO, C.; FAUVEL, C.; COSSON, J.; BILLARD, R. Cryopreservation
of sperm in marine fish. Aquaculture Research, v. 31, n. 3, p. 231-243, 2000.
SUQUET, M.; OMNES, M. H.; NORMANT, Y.; FAUVEL, C. Assessment of sperm
concentration and motility in turbot (Scophthulmus muximus). Aquaculture, v. 101, p. 177-
185, 1992.
SORENSEN, L.; BUSS, K.; BRADFORT, A. D. The artificial propagation of Escoid fishes in
Pennsylvania. Prog Fish Cult., v.28, p. 133-141, 1966.
SORENSEN, S. R.; SKOV, P. V.; LAUESEN, P.; TOMKIEWICZ, J.; BOSSIER, P.;
SCHRYVER, P. D. Microbial interference and potential control in culture of European eel
(Anguilla anguilla) embryos and larvae. Aquaculture, v. 426-427, p. 1-8, 2014.
TABARES, J.; TARAZONA, A.; OLIVEIRA, M. Fisiología de la activación del
espermatozoide en peces de agua dulce. Revista colombiana de Ciencias Pecuarias, v. 18, p.
149-16, 2005.

92
TAITSON, P. F.; CHAMI, E.; GODINHO, H. P. Gene banking of the neotropical fish
Leporinus obtusidens (Valenciennes, 1836): a protocol to freeze its sperm in the field. Animal
Reproduction Science, n. 105(3-4), p. 283-291, 2007.
TAVARES, L.H.S. Limnologia aplicada a aquicultura. Jaboticabal: FUNEP, 1995. 70p.
TOMELLERI, J.; EBERLE, M. Fishes of the Central United States. University Press of
Kansas, Lawrence, 1990.
TIERSCH, T. R.; YANG, H.; JENKINS, J. A.; DONG, Q. (2007). Sperm cryopreservation in
fish and shellfish. In: Roldan, E. R. S., Gomendio, M. (Eds.), Spermatology, Society of
Reproduction and Fertility Supplement 65. Nottingham University Press, Nottingham, pp. 93–
508.
TONIOLLI, R.; MESQUITA, S. M. Fertilidade de porcas inseminadas com sêmen diluído em
água de coco estabilizada e com BTS. Revista Brasileira de Reprodução Animal, v.14, p.249-
254, 1990.
URBÁNYI, B.; SZABÓ, T.; MISKOLCZI, E.; MIHÁLFFY, S.; VRANOVICS, K.;
HORVÁTH, Á. Successful fertilization and hatching of four European cyprinid species using
cryopreserved sperm. Journal of Applied Ichthyology, v. 22, p. 201–204, 2006.
VALDEBENITO, I. I.; GALLEGOS, P. C.; EFFER, B. R. Gamete quality in fish: evaluation
parameters and determining factors. Zygote, v.23, p.177-197, 2013.
VASCONCELOS, S. M. L.; GOULART, M. O. F.; MOURA, J. B. F.; MANFREDINI, V.;
BENFATO, M. S.; KUBOTA, L. T. Espécies reativas de oxigênio e de nitrogênio,
antioxidantes e marcadores de dano oxidativo em sangue humano: principais métodos
analíticos para sua determinação. Química Nova, v. 30, n. 5, p. 1323-1338, 2007.
VELASQUEZ-MEDINA, S. P. Criopreservação do sêmen de Pirapitinga Piaractus
brachypomus (Pisces, Characidae). 2008. 96f. Dissertação (Curso de Pós-Graduação Stricto
Sensu em Ciências Marinhas Tropicais), Universidade Federal do Ceará, Fortaleza.

93
VIEIRA, M. J. A. F.; CARVALHO, M. A. M.; SALMITO-VANDERLEY, C. S. B.;
SALGUEIRO, C. C. M.; VIVEIROS, A. T. M.; MOURA, A. A. A. N.; NUNES, J. F.
Características do sêmen de tambaqui (Colossoma macropomum) em latitude equatorial.
Archivos de zootecnia, v. 60, n. 232, p. 1263-1270, 2011.
VIVEIROS, A. T. M.; MARIA, A. M.; ÓRFÃO, L. H.; CARVALHO, M. A. M.; NUNES, J.
F. Powder coconut water (ACP) as extender for semen cryopreservation of Brazilian
migratory fish species. Cybium. v.32, n. 2, suppl 215, p.137, 2008.
VIVEIROS, A. T. M.; ISAÚ, Z. A.; CANEPPELE, D.; LEAL M. C. Sperm cryopreservation
affects postthaw motility, but not embryogenesis or larval growth in the Brazilian fish Brycon
insignis (Characiformes). Theriogenology, v. 78, p. 803–810, 2012a.
VIVEIROS, A. T. M.; ÓRFÃO, L. H.; NASCIMENTO, A. F.; CORRÊA, F. M.;
CANEPPELE, D. Effects of extenders, cryoprotectants and freezing methods on sperm
quality of the threatened Brazilian freshwater fish pirapitinga-do-sul Brycon opalinus
(Characiformes). Theriogenology, v. 78, p. 361–368, 2012b.
WARNECKE, D., PLUTA, H. Motility and fertilizing capacity of frozen/thawed common
carp (Cyprinus carpio L.) sperm using dimethyl-acetamide as the main cryoprotectant.
Aquaculture, v. 215, p. 167–185, 2003.
WOYNAROVICH, E. Manual de piscicultura. Ministério da Integração regional. Companhia
de desenvolvimento do vale do São Francisco (CODEVASF), Brasília 1993. Acesso em 25
outubro de 2012. Disponível em: http://www.abrapoa.org.br/livros%20download.html
YANG, H.; TIERSCH, T. R. Current status of sperm cryopreservation in biomedical research
fish models: zebrafish, medaka and xiphophorus. Comparative Biochemistry and Physiology -
Part C: Toxicology & Pharmacology, v. 149, n. 2, p. 224–232, 2009.
YAVAS, I.; BOZKURT, Y.; YILDIZ, C. Cryopreservation of scaly carp (Cyprinus carpio)
sperm: effect of different cryoprotectant concentrations on post-thaw motility, fertilization
and hatching success of embryos. Aquaculture International, v. 22, p. 141-148, 2014.





























