Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE MATO GROSSO
CÂMPUS UNIVERSITARIO DE SINOP
INSTITUTO DE CIÊNCIAS AGRÁRIAS E AMBIENTAIS
EFEITO DA APLICAÇÃO DO BIOCARVÃO NOS ATRIBUTOS
MICROBIOLOGICOS DO SOLO, CULTIVADO COM TECA, NO BIOMA
AMAZÔNIA
Mirelly Mioranza
Engenheira Agrônoma
2016

UNIVERSIDADE FEDERAL DE MATO GROSSO
CÂMPUS UNIVERSITARIO DE SINOP
INSTITUTO DE CIÊNCIAS AGRÁRIAS E AMBIENTAIS
EFEITO DA APLICAÇÃO DO BIOCARVÃO NOS ATRIBUTOS
MICROBIOLOGICOS DO SOLO, CULTIVADO COM TECA, NO BIOMA
AMAZÔNIA
Mirelly Mioranza
Orientador: Dr. Anderson Ferreira
Dissertação apresentada ao Programa de Pós Graduação em Agronomia, como parte das exigências para a obtenção do título de Mestre em Agronomia. Área de concentração: Ciências do Solo.
Fevereiro de 2016



DADOS CURRICULARES DO AUTOR
Mirelly Mioranza - filha de Gilmar Mioranza e Aparecida de Fátima
Montagneri Mioranza nasceu em 23 de Agosto de 1990, na cidade de
Amambai, estado de Mato Grosso do Sul. Em 2008 ingressou no curso de
Agronomia pela Universidade Federal de Mato Grosso (UFMT), campus
universitário de Sinop-MT, obtendo o titulo de Engenheira Agrônoma em 2013.
Ingressou no Curso de Pós Graduação em Agronomia - Ciência do Solo da
UFMT em março de 2014. Em março de 2014 ingressou no curso de Mestrado
em Agronomia pelo Programa de Pós-Graduação em Agronomia pela mesma
instituição, obtendo o título de Mestre em Agronomia em 2016.

Odiei as palavras e as
amei, e espero tê-las usado
direito.
Markus Zusak- The Book Thief

OFEREÇO
Aquele ao qual devemos toda honra e toda glória.
”Omnia possum in eo qui me confortat”
(Fp, 4,13)
DEDICO
Aos meus pais, Fátima e Gilmar Mioranza, pelo apoio, incentivo e pela confiança depositada em
mim ao longo desses anos. Amo vocês!
Uma pequena Teoria
As pessoas só observam as cores do dia no começo e no fim, mas, para mim, está muito claro
que o dia se funde através de uma multidão de matizes e entonações, a cada momento que
passa. Uma só hora pode consistir em milhares de cores diferentes.
Amarelos céreos, azuis borrifados de nuvens. Escuridões enevoadas.
No meu ramo de atividade, faço questão de notá-los.
Markus Zusak- The Book Thief

AGRADECIMENTOS
Agradeço a Deus, por me dar saúde, sabedoria e fé para a
concretização dos meus objetivos.
Aos meus pais, Gilmar Mioranza e Fátima, pelo incentivo, pela educação
que me proporcionaram e pelos princípios que me ensinaram. Por entenderem
minha ausência constante e acreditar no meu caminho, sabendo que essa é
mais uma etapa vencida na busca de um sonho.
Aos meus irmãos, Francielly e Marcos Vinicius por fazerem parte da
minha vida, cada um do seu jeito, mas cada qual com sua importância e
contribuição.
A Universidade Federal de Mato Grosso- Campus de Sinop e ao
Programa de Pós Graduação em Agronomia pela oportunidade de estudo
desde a graduação ate a realização deste Mestrado.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior –
CAPES e a Fundação de Amparo à Pesquisa do Estado de Mato Grosso –
FAPEMAT, pela concessão da bolsa de estudo.
Aos professores do curso de Pós-Graduação em Agronomia, por todo o
conhecimento transmitido. Muito obrigada!
Ao meu orientador, Dr. Anderson Ferreira, pela orientação, pelo
incentivo, confiança e pela contribuição marcante na minha formação
profissional. Obrigada pela amizade também.
Obrigada a todos da Embrapa que de alguma forma tenham contribuído
com este projeto, em especial aos meninos do campo Enio e Tiago que sempre
nos ajudaram nos momentos ‘temidos’ de coleta. Sou muito grata a vocês.
Aos pesquisadores da Embrapa: Fabiana Rezende pela gerencia dos
recursos do projeto e Flavio Tardin pela paciência e as aulas de estatísticas.
Essas informações são valiosas.
Aos técnicos do laboratório de Biologia Molecular, Joyce e Bruno - sem
vocês essa pesquisa não seria possível.
Agradeço imensamente as minhas amigas agrônomas Kaynara
Kavasaki, Lana Bäumgartner e Tairiny Oliveira (não é agrônoma, mas te
considero pacas). Espero que nossa amizade seja eterna. Amo vocês!

Agradeço minha parceira de laboratório Jacqueline Jesus, pelo
companheirismo, pela amizade e confiança. Sem você esta jornada não teria a
mesma graça. E agora vai me aguentar por mais quatro anos, rumo ao
doutorado. Obrigada!
As minhas queridas Embrapianas, Andressa, Carol, Camila, Luana,
Maíra, Mari e Carolina (mamãe do João), pelos momentos de descontração,
pelos ‘coffees’, pelos ‘Happy Hour’ muito necessários. Enfim, obrigada por
fazerem desta jornada bem mais divertida! Arrasaram gatas!!
Aos estagiários da equipe “Pró- enzima”: Okada, Bruce, Vinicius, Junior
e Elisa. Sou muito grata a vocês, por me ajudarem sempre, mesmo tendo que
pesar solo toda vez que não dava certo. Vocês são demais. Obrigada,
obrigada!
Agradeço aos meus companheiros de viagem: Tati, Kaynara, Jacque,
Vini, Heitor, Lana e Andréia. Minhas “férias” foram muito mais divertidas com
vocês. Agradeço também aos novos amigos que fiz durante esses momentos.
Obrigada a todos do Grupo “Tudo Junto e Misturado”. Quem tem
amigos, nunca esta só. Vocês são demais!!
A todos que contribuíram, direta ou indiretamente, para a realização
deste trabalho.
E por fim, agradeço a banda Engenheiros do Hawaii pelas musicas
inspiradoras. Vocês permitiram que eu concluísse as analises de DGGE. Valeu!
Meus Sinceros Agradecimentos!!!

SUMÁRIO
RESUMO ......................................................................................................... 12
ABSTRACT ...................................................................................................... 13
CAPÍTULO 1 – CONSIDERAÇÕES GERAIS ................................................... 14
1. INTRODUÇÃO ............................................................................................. 14
2. REVISÃO DE LITERATURA ........................................................................ 16
2.1. Teca (Tectona grandis Linn f) ............................................................... 16
2.2. Origem do biocarvão ............................................................................. 17
2.2.1. Biocarvão e a sua interação com o solo ........................................ 19
2.3. Qualidade do solo ................................................................................. 23
2.4. Microbiota do Solo ................................................................................ 24
2.5. Indicadores Microbiológicos do solo ..................................................... 24
2.5.1. Atividade Enzimática ...................................................................... 25
2.5.2. Diversidade Microbiana através da técnica de DGGE ................... 26
3. REFERÊNCIAS BIBLIOGRÁFICAS ............................................................. 27
CAPÍTULO 2 – Efeito da aplicação do biocarvão nos atributos
microbiologicos do solo, cultivado com teca, no bioma amazônia ........... 37
RESUMO ......................................................................................................... 37
1.INTRODUÇÃO .............................................................................................. 38
2. MATERIAIS E MÉTODOS............................................................................ 40
2.1. Descrição da Área de Estudo ................................................................ 40
2.2. Plantio da Teca ..................................................................................... 41
2.3. Produção do Biocarvão ......................................................................... 41
2.4. Tratamentos .......................................................................................... 42
2.5. Área de Referência ............................................................................... 42
2.6. Amostragem de solo ............................................................................. 43
2.7. Umidade e pH do solo ........................................................................... 43

2.8. Atividade Enzimática ............................................................................. 43
2.9.Caracterização da comunidade de bactérias (gene 16S rDNA) ............. 44
2.9.1. Extração de DNA e Reação em Cadeia de Polimerase (PCR) ...... 44
2.9.2. Gel em Eletroforese com Gradiente Desnaturante (DGGE) .......... 45
2.10. Análise Estatística ............................................................................... 46
4. RESULTADOS ............................................................................................. 47
4.1. Atividade Enzimática, pH e umidade do solo ........................................ 47
4.2. Diversidade bacteriana (Índice de Shannon) ........................................ 49
4.3. Estrutura da comunidade de bactérias totais (gene 16S rDNA) ............ 50
4.4. Relação das alterações nas comunidades microbianas com atributos do
solo. .................................................................................................................. 52
5. DISCUSSÃO ................................................................................................ 55
6. CONCLUSÃO ............................................................................................... 61
7. AGRADECIMENTOS ................................................................................... 62
8. REFERÊNCIAS BIBLIOGRÁFICAS ............................................................ 62
9. ANEXOS ...................................................................................................... 68

EFEITO DA APLICAÇÃO DO BIOCARVÃO NOS ATRIBUTOS
MICROBIOLOGICOS DO SOLO, CULTIVADO COM TECA, NO BIOMA
AMAZÔNIA
RESUMO - O biocarvão é o produto formado a partir da decomposição térmica da biomassa, em ambiente fechado, através de suprimento limitado de oxigênio e de temperaturas relativamente baixas e utilizado como condicionador de solo. O objetivo desta pesquisa foi avaliar o uso do biocarvão como condicionador do solo e seu efeito na atividade da β-glicosidase, fosfatase ácida e diversidade bacteriana do solo em experimento de Teca com três e quinze meses de duração, em um latossolo vermelho-amarelo, localizados em Sinop - Mato Grosso, Brasil. Os tratamentos consistiram de: T1- Muda sem biocarvão; T2- Muda com biocarvão; T3- Muda sem biocarvão + biocarvão ativado aplicado no solo; T4- Muda com biocarvão + biocarvão ativado aplicado no solo; T5- Muda sem biocarvão + biocarvão não ativado aplicado no solo; T6- Muda com biocarvão + biocarvão não ativado aplicado no solo. Foi determinada a atividade de duas enzimas relacionadas ao ciclo do C e P e a biodiversidade bacteriana foi avaliada por meio da técnica de DGGE com iniciadores universais para a região 16S rDNA de bactérias. Não houve incremento na atividade da β-glicosidase quinze meses após aplicação do biocarvão, mas sua forma ativada proporcionou a maior atividade desta enzima, ao contrário a fosfatase ácida apresentou uma baixa atividade nos primeiros 3 meses e após quinze meses com aplicação do biocarvão no solo a atividade desta enzima foi maior. Em relação à umidade e pH do solo, não foi alterado pela presença do biocarvão. A estrutura e diversidade da comunidade bacteriana no solo sofreram influencia mais acentuada do biocarvão com quinze meses de aplicação no solo. Palavras-chave: DGGE, condicionador de solo, β-glicosidase, fosfatase ácida, ecologia microbiana

EFFECT OF APPLICATION OF BIOCHAR ON MICROBIOLOGICAL SOIL
PROPERTIES, CULTIVATED WITH TECA, IN THE AMAZON BIOME
ABSTRACT – The biochar product is formed from the thermal decomposition of
biomass in a closed environment by limited supply of oxygen and at relatively
low temperatures and used as a soil conditioner. The aim of this study was to
evaluate the use of biochar as a soil conditioner and its effect on the β-
glucosidase activity, acid phosphatase and bacterial diversity in soil Teak
experiment with fifteen and three months duration in a oxisol, located in Sinop -
Mato Grosso, Brazil.The treatments were: T1 seedling without biocarvão ; T2
seedling with biocarvão; T3 seedling without biocarvão + activated biocarvão
applied to the soil; T4 seedling with biocarvão + activated biocarvão applied to
the soil; T5- seedling without biocarvão + biocarvão not activated applied to the
soil; T6- seedling with biocarvão + biocarvão not activated applied to the soil. It
determined the activity of two enzymes related to the cycle C and P and
bacterial biodiversity was evaluated by DGGE technique using universal
primers for the bacterial 16S rDNA region. There was no increase in the activity
of β-glucosidase fifteen months after application of biochar, but its activated
form accompanied by increased activity of this enzyme, unlike the acid
phosphatase had a low activity in the first 3 months and after fifteen months on
application of biochar in the soil the activity of this enzyme was higher. With
regard to moisture and soil pH was not altered by the presence of biochar. The
structure and diversity of the bacterial community in the soil suffered more
pronounced influence of biochar fifteen months of soil application.
KEYWORDS: DGGE, soil conditioner, β-gluicosidase, acid phosphatase, microbial ecology

14
CAPÍTULO 1 – CONSIDERAÇÕES GERAIS
1. INTRODUÇÃO
Em ambientes tropicais, o processo de degradação dos solos encontra-
se intimamente relacionado à dinâmica da matéria orgânica (CAMARGO,
1999). Diversos autores têm ressaltado que a converção da vegetação nativa
em área de produção agrícola pode reduzir drasticamente os teores de matéria
orgânica e consequentemente alterando a qualidade dos solos, principalmente
no Cerrado/Amazônia, devido ao menor suprimento de resíduos orgânicos e ao
aumento na taxa de decomposição (FREIXO et al., 2002; BAYER et al., 2004;
CHEN et al., 2004; MACEDO et al., 2008).
Os microrganismos são a parte viva e mais ativa da matéria orgânica do
solo, e são muito importantes nos processo de decomposição de resíduos,
ciclagem de nutrientes, fixação biológica do nitrogênio, decomposição de
material mineral, solubilização de nutrientes, formação da matéria orgânica e
estrutura do solo (ZILLI et al., 2003; KARLEN et al., 2003). Em condições
naturais os solos tropicais, possuem baixa fertilidade natural, porém
características físicas e biológicas favoráveis. Uma vez incorporados ao
processo produtivo, para que proporcionem condições adequadas ao
crescimento das plantas cultivadas, é necessário o aumento da sua fertilidade
química, por meio de calagens, gessagens e adubações (CORAZZA et al.,
2004).
O cultivo agrícola, entretanto, promove alterações em atributos físicos,
químicos e biológicos, com o passar dos anos (BAYER et al., 1999). Assim,
mesmo que uma área cultivada apresente fertilidade química mais elevada do
que uma área sob vegetação nativa, e essa fertilidade seja adequada para
obtenção de produtividades elevadas, outros atributos do solo serão alterados,
o que pode comprometer a eficiência dos sistemas e a sustentabilidade dos
sistemas de produção implantados (SILVA et al., 2009).
Atualmente, a fronteira agrícola brasileira está inserida dentro da
Amazônia Legal, particularmente, as maiores áreas estão no estado de Mato

15 Grosso (DOMINGUES E BERMANN, 2012). Nesse sentido, novas alternativas
para manter o manejo adequado e conservação da qualidade de solos devem
ser exploradas como, por exemplo, uso de biocarvão como condicionadores de
solos. Para regiões como o Cerrado e áreas de transição com Amazônia, onde
as praticas de queimadas são comuns, a quantidade e a qualidade de MOS ao
passar do tempo podem ser influenciadas pela produção de carvão vegetal,
que representa uma fração de carbono orgânico total (COT), estável no solo
(FONTANA, 2006).
Biocarvão é o produto da combustão incompleta de restos de material
orgânico ou de culturas voltadas à sua produção. Possui alto teor de C e
grande área superficial devido a sua porosidade em consequência do processo
de queima na ausência do oxigênio (MADARI at al., 2009). Apresenta baixo
teor de nitrogênio, o que resulta em uma elevada relação de C:N. Segundo Bird
et al. (1999), o biocarvão pode ser significativamente degrado em uma escala
de décadas ou milhares de anos em solos tropicais. A biodegradação do
biocarvão no solo é um processo relativamente lento que resulta na
mobilização do carbono e na alteração das propriedades da superfície do
carvão, aumentando a concentração de sítios quimicamente reativos que
podem fazer ligações com os nutrientes presentes e contribuírem para
aumentar a CTC do solo. (GLASER et al., 2003).
Dentre os possíveis benefícios ambientais que podem ser alcançados
com uso de biocarvão podemos citar: (1) mitigação nas mudanças do clima; (2)
melhoria na fertilidade dos solos; e (3) redução da poluição ambiental
(LEHMANN et al., 2011). Devido a sua enorme importância na manutenção da
produtividade dos ecossistemas agrícolas e florestais, torna-se cada vez mais
frequentes as pesquisas que colaboram para um melhor entendimento sobre
as propriedades microbiológicas dos solos. Nesse ponto, conhecer a
quantidade, diversidade e atividade do componente microbiológico do solo,
pode auxiliar no entendimento das mudanças no balanço de carbono e fluxo de
energia, na ciclagem de nutrientes e na sua fertilidade, pois a atividade e
biomassa microbiana são consideradas indicadores importantes de degradação
do solo (RIBEIRO, 2012).
Dessa maneira, objetiva-se manter o carbono orgânico no solo,
utilizando o biocarvão, através da avaliação de alguns atributos microbiológicos

16 em cultivo com Teca no interior do Mato Grosso, como condicionador dos solos
tropicais.
2. REVISÃO DE LITERATURA
2.1. Teca (Tectona grandis Linn f)
A espécie Tectona grandis Linn f., popularmente conhecida como Teca
(Brasil) ou Teak (Índia), é uma espécie arbórea natural das florestas do
sudoeste asiático (SÁNCHEZ, 2002). Esta arvore pertence à família
Lamiaceae, é uma espécie arbórea decídua da floresta tropical, caducifólia que
perde suas folhas na estação seca, apresenta um fuste cilíndrico que pode
alcançar alturas maiores que 25 m quando cresce em condições
edafoclimáticas favoráveis (SHIMIZU et al., 2007).
O bom crescimento e alta qualidade da madeira estão associados a
alguns fatores ambientais, tais como, a profundidade do solo, a capacidade de
retenção de água, solos com boa drenagem, ricos em cálcio, relevo suave e
fertilidade mediana. A teca se desenvolve melhor com precipitações anuais
variando de 1250 a 3750 mm e temperaturas que oscilam entre os 13°C e
37°C, desenvolvendo-se melhor aos 25ºC (SETH & KAUL, 1978).
O plantio de teca, tem se mostrado nos últimos anos como uma das
grandes essências florestais de cultivo intensivo no Brasil, sobretudo no Estado
de Mato Grosso, com mais de 48.000 hectares de área cultivada, conforme
constatado por Shimizu et al. (2007). Neste Estado, o cultivo de Teca está em
franca expansão, tendo em vista o alto valor comercial de sua madeira,
associado às condições edafoclimáticas favoráveis encontradas em diversas
regiões (ÂNGELO et al.,2009).
A área de florestas plantadas com teca no Brasil atingiu, em 2009,
estimados 65.240 ha, enquanto em 2008 o total foi de 58.810 ha. Tais áreas
representam crescimento de 10,9% na área plantada deste grupo de espécies
no país indicando o interesse crescente por tal gênero florestal, principalmente
em função das expectativas de retorno financeiro em projetos com a mesma

17 (ABRAF, 2010). Atualmente, os plantios totalizam 67.329 ha e representam
apenas 0,4% da produção de madeira plantada no Brasil (ABRAF, 2013).
A produtividade média obtida para teca está na ordem de 15 m³/ha.ano,
representando uma produção estimada de 0,7 milhões de m³ por ano dividida
entre a Região Norte (32,6%) e Centro-oeste (67,4%) (ABRAF, 2013). Segundo
Finger et al., (2001), a produção mundial de madeira de teca é estimada em 3
milhões de m³/ano, o que é extremamente baixa pela demanda atual dessa
espécie no mercado exterior. No momento, o reflorestamento com teca no
Brasil surge como uma ótima opção de investimento.
2.2. Origem do biocarvão
O biocarvão (biochar) é o produto da degradação térmica de materiais
orgânicos na ausência de ar (pirólise) e difere do carvão vegetal devido sua
aplicação como condicionador de solo (LEHMANN et al., 2011).
O biocarvão é o produto formado a partir da decomposição térmica da
biomassa, em ambiente fechado, através de suprimento limitado de oxigênio e
de temperaturas relativamente baixas (<700ºC) (LEHMANN E JOSEPH, 2009).
E de acordo com Sohi et al. (2010), o biocarvão apenas deve ser utilizado
quando existir potencial para: melhorar a qualidade do solo, a produtividade
agrícola e contribuir para a mitigação da emissão de gases do efeito estufa a
longo prazo.
Solos antropogênicos são formados através de milhares de anos, devido
à decomposição térmica do material vegetal e a continua reposição da matéria
orgânica no solo. O biocarvão esta normalmente relacionada com estes solos,
por sua semelhança no processo de formação, sugere-se também uma
conexão com a “Terra Preta de Índio” (TPI), um solo altamente fértil encontrado
na Amazônia, devido principalmente à alta capacidade de troca de cátions
(CTC), aos altos teores de matéria orgânica e nutrientes como fósforo,
potássio, nitrogênio e cálcio (NOVOTNY et al., 2009).
No Brasil, exemplo do efeito benéfico do biocarvão (ou carbono
pirogênico) para a fertilidade dos solos tropicais são os solos chamados de
terra preta de índio (ou terra preta arqueológica), que são solos antrópicos dos

18 povos pré-colombianos da Bacia Amazônica (KAMPF E KERN, 2005). Esses
solos contêm elevado teor de carbono pirogênico que chega a ser 2,5 vezes
maior que o teor desse material em solos adjacentes não antrópicos. Esse
carbono, que provavelmente se originou de queimas periódicas de lavouras e
resíduos da atividade diária previamente carbonizada, foi incorporado ao solo
pelo homem e pela atividade biológica no solo durante centenas ou milhares de
anos.
Evidencias indicam de que as TPI são depósitos culturais criados a partir
de adição de artefatos como peças de cerâmicas, ferramentas de pedras e
carvão ao redor das áreas habitadas e da manipulação de material orgânico
em áreas associadas ao cultivo. O fogo é um componente crucial na formação
desses solos por contribuir na forma de carvão e cinza, o que aumenta o pH do
solo (devido aos carbonatos presentes nas cinzas vegetais) e suprime a
toxidade do alumínio nas raízes vegetais e microbiota do solo (TEIXEIRA et al.,
2010; GLASER et al., 2001; GLASER et al., 2002).
O solo amazônico é geralmente arenoso ou argiloso, tem pouco
nutriente e apresenta uma fina camada superficial de húmus, proveniente das
florestas. Contudo em algumas regiões amazônicas são encontradas manchas
profundas de solos, escura e muito fértil, ao contrário do que é normalmente
encontrado em quase toda região. O aumento da atividade biótica e da
capacidade de retenção de nutrientes através da deposição de material
orgânico e carvão vegetal pode ser a principal razão pela persistência desses
solos, por um longo período de tempo mesmo após o término do seu manejo
cultural (TEIXEIRA et al., 2010).
Para regiões como o Cerrado e áreas de transição com Amazônia, onde
as praticas de queimadas são comuns, a quantidade e a qualidade de MOS ao
passar do tempo podem ser influenciadas pela produção de carvão vegetal,
que representa uma fração de carbono orgânico total (COT), estável no solo
(FONTANA, 2006). Biocarvão (BC) é o produto da combustão incompleta de
restos de material orgânico ou de culturas voltadas à sua produção, apresenta
baixo teor de nitrogênio, o que resulta em uma elevada relação C:N e possui
alto teor de C e grande área superficial devido a sua porosidade em
consequência do processo de queima na ausência do oxigênio (MADARI et al.,
2009).

19
Segundo Bird et al. (1999), o BC pode ser significativamente degradado
em uma escala de décadas ou milhares de anos em solos tropicais. A
biodegradação do BC no solo é um processo relativamente lento que resulta na
mobilização do carbono e na alteração das propriedades da superfície do
carvão, aumentando a concentração de sítios quimicamente reativos que
podem fazer ligações com os nutrientes presentes e contribuírem para
aumentar a CTC do solo (GLASER et al., 2003). Dentre os possíveis benefícios
ambientais que podem ser alcançados com uso de biocarvão podemos citar:
(1) mitigação nas mudanças do clima; (2) melhoria na fertilidade dos solos; e
(3) redução de poluição ambiental (LEHMANN et al., 2011).
Estudos tem demonstrado também a utilização do biocarvão como
alternativa para melhorar a fertilidade dos solos, devido a um rearranjo dos
átomos de carbono, aumentando a porosidade do material, isto proporciona
melhoria nas propriedades químicas, principalmente pela alta superfície
específica resultando em aumento da capacidade de troca de cátions e
adsorção de nutrientes (ATKINSON et al., 2010). Em solos onde o sistema de
manejo possibilita acúmulo de material orgânico, como a adição de biocarvão,
as condições são favoráveis à fertilidade do solo, ao desenvolvimento
microbiano, além da reciclagem de resíduos e de nutrientes (MADARI et al.,
2009).
2.2.1. Biocarvão e a sua interação com o solo
A melhoria na fertilidade do solo tem sido atribuída ao aumento da
capacidade de troca catiônica, as alterações no pH do solo e as contribuições
diretas de nutrientes fornecidos pelo biochar (MAJOR et al., 2010;XU et al.,
2013; WANG et al., 2015). Além disso, devido a sua estrutura de carbono
aromático, o biochar é mais resistente á decomposição microbiana e pode
persistir no solo de 1000 a 10.000 anos, mantendo assim, estoque de carbono
no solo (WARNOCK et al., 2007; STEINER et al., 2007; RUTIGLIANO et al.,
2013).
As interações entre matéria orgânica e a superfície coloidal das argilas
são dependentes da distribuição dos grupos funcionais, da matéria orgânica,

20 da polaridade dos componentes, da composição e concentração dos cátions e
ânions na solução do solo. A área de superfície específica do biochar é de 200-
400 m2 g-1, é comparável a da argila e superior a da areia (PETTER et al.,
2012). Tais parâmetros levam a hipótese de que a presença do biochar em
solos arenosos pode suprir de certa forma a limitada disponibilidade de água,
baixa capacidade de troca catiônica, disponibilidade de nutrientes e atividade
de microrganismos. Reações complexas similares estão propensas a ter lugar
nas superfícies dos biocarvãos, principalmente se estes biocarvões
apresentam alto conteúdo mineral (JOSEPH et al., 2010).
O biochar apresenta cinzas, que são fontes de P, K e outros elementos,
que podem ser mais solúveis e acessíveis nos biocarvões que na matéria
orgânica não pirolisada. O efeito do biocarvão na disponibilidade destes
elementos pode explicar alguns impactos na produção de algumas culturas a
curto prazo, especialmente pelo fato de que, alguns elementos não podem ser
disponibilizados simplesmente pelo aumento da quantidade de matéria
orgânica no solo (SOHI et al., 2010).
Em solos adubados com uma mistura de cinzas e carvão, proveniente
de um usina termoelétrica, Omil et al. (2013) observaram que não foi
necessária a aplicação suplementar de P com aplicações anuais da mistura,
durante três anos, em doses de 4,5 t ha-1, no crescimento de um povoamento
de Pinus radiata, com anos de idade. Os autores ainda ressaltaram que os
tratamentos com a mistura, proporcionaram menores perdas de nutrientes por
lixiviação e aumento no teor de matéria orgânica do solo, mesmo em solo
arenoso. Petter et al. (2012) concluíram que a aplicação de biocarvão de
eucalipto em aplicações de 8, 16 e 32 t ha-1 em campo, afetou positivamente a
fertilidade do solo, aumentando principalmente a disponibilidade de Ca, P, e
aumentando o pH e o carbono orgânico total dos solos.
Estudando a influência do biocarvão em duas doses, 8 ou 20 t ha-1,
aplicadas uma única vez, na qualidade do solo adubado, Major et al. (2010)
observaram, após quatro anos da aplicação em savana colombiana, que o
biocarvão, produzido a partir de resíduos de madeira, aumentou principalmente
a disponibilidade de Ca e Mg e o pH, e diminuiu a acidez trocável.
A capacidade de troca catiônica está relacionada á área de superfície
específica, mas é altamente dependente de sítios quimicamente reativos.

21 Esses sítios vão se formando ao longo dos anos ao passo que partículas são
atacadas por microrganismos no solo, alterando as características químicas e
físicas do biochar (COHEN-OFRI et al., 2006). Essas alterações segundo estes
autores, se dão pelo aumento de cargas elétricas oriundas da oxidação dos
compostos bioquímicos, levando a um aumento no número de compostos
fenólicos, hidroxílicos, carbonilicos e quinonas e, consequentemente, a um
aumento no número de cargas negativas resultante da substituição pelas
cargas positivas no processo de oxidação. Talvez esta seja à explicação do
aumento da CTC do biochar com o passar dos anos em solos que contem esse
material.
Um aumento no pH do solo após a aplicação do biocarvão é
frequentemente relatado em diversos tipo de solo (GLASER et al., 2002;
AMELOOT et al, 2013.; XU et al., 2014). Isto é devido ao pH alcalino do
biocarvão, que é positivamente relacionado com a sua temperatura de
produção e do tipo de matéria-prima (à base de madeira tende a ter um pH
mais elevado do que o biocarvão a partir de resíduos de culturas e
compostagem). Outra razão para o aumento do pH no solo é a presença de
grupos fenólicos, carboxilo e hidroxilo carregados negativamente sobre
superfícies de biocarvão que se ligam aos H + da solução do solo, reduzindo
desse modo a sua concentração aumentando assim, o pH. A influência positiva
do biocarvão no aumento pH do solo é mais acentuada em solos ácidos e com
baixo teor de matéria orgânica, provavelmente porque está ligada à capacidade
de tamponamento do pH do solo (GUL et al., 2015).
A presença de biochar pode contribuir para algumas alterações nas
propriedades físicas do solo, tais como a estrutura, porosidade, diâmetro dos
poros e densidade em função de sua maior área superficial especifica
(PETTER et al., 2012). Ainda segundo esses autores, a mudança nas
características físicas do solo devido à incorporação do biochar pode resultar
em maior produtividade das plantas, devido a maior disponibilidade de água na
zona próxima ao sistema radicular e também devido à porosidade do biochar
que pode aumentar a capacidade de retenção de água, especialmente em
solos de textura arenosa, reflexo das estruturas celulares da matéria prima que
é normalmente produzido. Contudo, a estrutura aromática, que tem
características hidrofóbicas, pode reduzir a penetração de água nos espaços

22 porosos dos agregados do solo, aumentando assim a estabilidade dos
agregados (GLASER et al., 2002).
Todas essas alterações nas propriedades físicas podem levar a
mudanças em outras propriedades do solo, principalmente químicas e
biológicas, em função do surgimento de sítios quimicamente reativos e habitats
de proteção para microrganismos do solo (BRADY E WEIL, 2008).
A estrutura porosa do biochar, sua elevada área superficial interna e sua
capacidade de absorver matéria orgânica, gases e nutriente inorgânicos
fornecem um habitat ideal para os microrganismos do solo colonizar, crescer e
reproduzir, principalmente bactérias, actinomicetos e fungos micorrizicos
arbusculares (THIES et al., 2009). Dependendo do tamanho do poro, diferentes
microrganismos terão acesso aos espaços internos, tais como bactérias (entre
0,3 a 3 mm), fungos (2-80 mm) e protozoários (7-30 mm). Os macroporos do
biochar (> 200 nm) representam, provavelmente, o habitat mais protegido uma
vez que são do tamanho certo para acomodar as bactérias (QUILLIAM et al.,
2013), embora o biochar também apresente microporos (<2 nm) e mesoporos
(2-50 nm) que podem armazenar água e substancias dissolvidas que são
necessárias para o metabolismo microbiano (BREWER E BROWN, 2012).
Muitos autores tem sugerido que os poros do biochar agem como
microhabitats para a colonização microbiana, onde eles são protegidos dos
seus predadores naturais ou por serem menos competitivos no solo, poderem
se estabelecer (WARNOCK et al., 2007; RUTIGLIANO et al., 2014; GUL et al.,
2015). O tamanho e abundancia dos poros é influenciado pela matéria-prima e
da temperatura de produção do biocarvão, onde maior a temperança, resulta
em mais água e volatilização de matéria orgânica, criando assim, poros
maiores (BREWER E BROWN, 2012).
O aumento da biomassa e diversidade microbiana no solo é vista como
um resultado proporcionado pela adição do biochar. SUN et al., (2013)
verificaram um aumento 3 vezes mais no Índice de Shannon-Weiner para o
perfil bacteriano de DGGE 16S rDNA no solo com biochar comparado com o
controle durante uma incubação de 96 dias. Hu et al. (2014) verificaram uma
diversidade bacteriana maior de 12%, 30% e 37% e uma diversidade de fungos
menor de 17%, 40% e 23% nos índices de Shannon-Weiner, Simpsons e Chao
respectivamente, em solo argiloso de floresta. Esta detecção de gêneros

23 /espécies devido a um aumento no tamanho e densidade das populações
microbianas, em função do biochar, baseada no DNA fornece aos
pesquisadores a capacidade de usar ferramentas moleculares para sondar
ainda mais os processos microbianos que são afetados em solos com a
aplicação deste material.
2.3. Qualidade do solo
A qualidade do solo, em um ecossistema, compreende o equilíbrio entre
os processos geológicos, hidrológicos, químicos, físicos e biológicos do solo
(BRUGGEN E SEMENOV, 2000; SPOSITO E ZABEL, 2003; ZILLI et al., 2003).
O conceito de qualidade do solo surgiu no final da década de 70 e
durante os 10 anos seguintes estiveram muito associados ao conceito de
fertilidade (KARLEN et al., 2003). Acreditava-se, por exemplo, que um solo
quimicamente rico era um solo com alta qualidade, isto porque tinha a
capacidade de prover a produção agrícola. Entretanto, a percepção de
qualidade do solo evoluiu e, num entendimento mais amplo, percebe-se que
não basta apenas o solo apresentar alta fertilidade, mas, também, possuir boa
estruturação e uma alta diversidade de organismos.
Essa evolução do conceito de qualidade do solo, fez com que sua
complexidade de avaliação fosse ampliada, uma vez que todos estes
parâmetros estão intimamente associados, podendo ser avaliados por vários
indicadores. Nos estudos de ZILLI et al. (2003) avaliou-se a diversidade
microbiana como indicador de qualidade do solo (IQS), estabeleceu as
seguintes características ideais de um indicador de qualidade do solo: ser
capaz de responder, de forma rápida e acurada, a um distúrbio no solo; refletir
os aspectos do funcionamento do ecossistema; possuir processo de avaliação;
ser economicamente viável. Ainda neste contexto, Visser e Parkinson (1992)
mencionam que precisa ter uma distribuição universal e independente de
sazonalidade.
O tipo de manejo do solo interfere diretamente nos atributos físicos,
químicos e biológicos do solo (CARTER, 1986). Essas interferências são
promovidas principalmente pelas modificações na temperatura e umidade do

24 solo, qualidade do resíduo vegetal depositado e atividade dos microrganismos
(CATTELAN E VIDOR, 1990).
2.4. Microbiota do Solo
Os microrganismos ocupam em torno de 0,5% do espaço poroso do
solo. O solo não rizosférico, é considerado um deserto nutricional, devido à
ausência de substratos necessários ao metabolismo microbiano e as condições
físico-químicas nem sempre favoráveis. Dessa forma, é muito importante a
natureza dos compostos que fornecem carbono, nutrientes e energia ao
metabolismo celular, pois a microbiota do solo utiliza os resíduos vegetais,
animais e de outros microrganismos para a sua reprodução (MOREIRA E
SIQUEIRA, 2006).
As bactérias, que tem geralmente diâmetro aproximado de 0,5 μm,
localizam-se na matéria orgânica, nos poros e no interior dos agregados. A
ocupação dos poros é devido ao seu tamanho e fornecimento de gases e água
(MOREIRA E SIQUEIRA, 2006).
O solo e, consequentemente, a microbiota são continuamente
influenciados por condições do ambiente, como a umidade, temperatura,
manejo do cultivo e precipitação (CARDOSO E FREITAS, 1992). Por essa
razão, e condições de estresses abióticos e bióticos, apenas parte desses
microrganismos encontram-se em estado ativo, cerca de 15 a 30 % das
bactérias e 10% dos fungos. Essa parte ativa dos microrganismos é
denominada de biomassa microbiana, respondendo por 90% da atividade
microbiana no solo (SIQUEIRA E FRANCO, 1988).
2.5. Indicadores Microbiológicos do solo
Os indicadores de qualidade do solo podem ser classificados como
físicos, químicos e biológicos (ZILLI et al., 2003). Dentre estes, o biológico
possui uma capacidade de responder rapidamente às alterações no solo,
fornecendo informações rápidas sobre mudanças nas propriedades do solo. Os
indicadores biológicos podem ser mensurados pela composição da microbiota
e sua biomassa, diversidade genética de microrganismo e da atividade

25 enzimática, para mitigação de possíveis estratégias de manejo
conservacionistas do solo (KARLEN et al., 2003).
2.5.1. Atividade Enzimática
O estudo da atividade enzimática tem sido reportado como indicador
efetivo da qualidade do solo, da decomposiçã da matéria orgânica e da
disponibilidade de nutrientes. As enzimas presentes no solo, em geral, são
produzidas por microrganismos, via intra e extracelulares. A atividade
enzimática do solo resulta principalmente da ação de enzimas extracelulares
que podem estar livres na solução do solo, adsorvidas nos coloides ou
imobilizadas em complexos húmicos, estas enzimas são mais importantes na
biodegradação de macromoléculas orgânicas no solo, devido às interações das
enzimas com os coloides do solo (MOREIRA E SIQUEIRA, 2006).
Lisboa et al. (2012) explicam que devido a baixa concentração de
enzimas no solo, as determinações são mensuradas indiretamente, através da
medida da sua atividade e não de quantidade. Geralmente, a atividade é
avaliada por meio da adição de um substrato específico para cada enzima, em
uma concentração conhecida, onde ocorrerá a ligação e, posteriormente, a
quebra desse substrato, e com a incubação sob condições ótimas de pH e
temperatura quantifica-se o produto (TABATABAI, 1994).
A escolha das enzimas a serem analisadas para avaliar a qualidade do
solo baseia-se na sua sensibilidade ao manejo do solo, na decomposição da
matéria orgânica e operacionalidade da analise. As enzimas mais comumente
avaliadas são as hidrolases ligadas aos ciclos do C, N, P e S.
As glicosidases podem ser encontradas em plantas, animais e
microrganismos, catalisam reações de hidrólise de celulose em glicose, cujos
produtos são importantes fontes de carbono para os microrganismos do solo.
As fosfatases são fundamentais na mineralização do fósforo e,
consequentemente, na ciclagem deste nutriente no ambiente. Elas estão
amplamente distribuídas no solo e tem sido estudadas porque catalisam a
hidrolise de fósforo orgânico e inorgânico, disponibilizando-o assim para as

26 plantas. De acordo com seu pH ótimo de ação, podem ser classificadas como
ácidas (pH 6,5) ou alcalinas (pH 11) (TABATABAI, 1994).
No Brasil, maioria dos estudos realizados envolvendo a atividade
enzimática do solo foi em regiões de clima tropical, principalmente no Cerrado
brasileiro. Nessas regiões, geralmente observa-se que a atividade da β-
glicosidase varia de 24 a 35 μg p-nitrofenol g-1 solo h-1 na profundidade de 0-5
cm, sendo que nem sempre as áreas de mata nativa apresentam os maiores
valores quando comparados aos solos cultivados (MENDES, 2006). Em
relação à fosfatase ácida, ela geralmente é superior em áreas de vegetação
nativa, e os valores normalmente encontrados variam de 260 a 1500 μg p-
nitrofenol g-1 solo h-1 (MATSUOKA et al., 2006)
Apesar de serem indicadores sensíveis para avaliar a qualidade do solo,
faz-se necessário o estabelecimento de cuidadosas relações destas com o
clima e solo, uma vez que, esta análise é muito influenciada por variações
ambientais, como o manejo do solo, pH, temperatura, umidade, precipitação,
composição da microbiota e outros.
2.5.2. Diversidade Microbiana através da técnica de DGGE
Aproximadamente 9.500 espécies de procariotos já foram descritas ao
longo dos anos. No entanto, por meio de técnicas moleculares, sabe-se que
isso representa apenas cerca de 1 a 10% da biodiversidade existente no
planeta (THOMPSON et al., 2005). Atualmente, existem diversos métodos
moleculares disponíveis para o estudo de comunidades microbianas
independentes de cultivo. Eles se baseiam principalmente em técnicas que
utilizam a manipulação dos ácidos nucleicos, ou seja, o DNA ou o RNA
extraído diretamente do solo, entre os quais se podem citar o DGGE,
Sequenciamento, PCR em tempo real entre outros (MUYZER, 1993;
CARDENAS E TIEDJE, 2008).
A técnica de eletroforese em gel com gradiente desnaturante (DGGE) se
baseia na mobilidade eletroforética de fragmentos de DNA parcialmente
desnaturados de mesmos tamanhos, mas de diferentes sequências de pares
de base amplificados por PCR de uma mesma amostra ambiental em um gel

27 de poliacrilamida que contém gradiente linear desnaturante crescente
composto por ureia e formamida (MUYZER, 1993). Quando sequencias de
mesmos pares de bases atingem a temperatura de desnaturação a uma
determinada posição no gel, a migração praticamente cessa. Diferentes teores
de G+C na dupla fita, irão parar de migrar em diferentes posições devido à
variação de gradiente de desnaturação e, assim, podem ser separados pelo
DGGE (RACHID, 2010). O DNA amplificado é separado de acordo com a
quantidade de grampos G+C dos diferentes fragmentos, construindo assim um
perfil de bandas que representam a impressão digital da estrutura da
comunidade microbiana em diferentes ambientes.
Uma sequencia de DNA rica em guanina e citosina (grampo de GC),
variando entre 30 e 40 bases é anexada no final 5’ do oligonucleotideo iniciador
forward, modificando dessa forma o padrão de desnaturação em que
praticamente 100 % das sequencias podem ser identificadas (MUYZER E
SMALLA, 1998). Segundo Øvreås et al. (1997) a região V3 do rDNA 16S é a
região mais estudada para avaliar a comunidade microbiana através da técnica
de DGGE, por apresentar a maior quantidade de bandas no gel e produzir um
perfil mais significativo.
O DGGE é uma técnica de fingerprinting relativamente rápida, pouco
onerosa e possibilita a análise simultânea de varias amostras. No entanto, o
DGGE apresenta algumas limitações, onde somente fragmentos de até 500 pb
podem ser separados no gel de acrilamida e apenas espécies predominantes
presentes na comunidade podem ser detectadas, em que, a população
detectada no gel pode corresponder a menos de 1% dos organismos existentes
na amostra (FROMIN et al., 2002).
3. REFERÊNCIAS BIBLIOGRÁFICAS
ABRAF – Associação Brasileira de Produtores de Florestas Plantadas. Anuário
estatístico da ABRAF 2010 ano base 2009. Brasília. 140 p., 2010.
ABRAF – Associação Brasileira de Produtores de Florestas Plantadas. Anuário
estatístico da ABRAF 2013 ano base 2012. Brasília. 142 p., 2013.

28 AMELOOT, N., NEVE, S.D., JEGAJEEVAGAN, K., YILDIZ, G., BUCHAN, D.,
FUNKUIN, Y.N., PRINS, W., BOUCKAERT, L., SLEUTEL, S. Short-term CO2
and N2O emissions and microbial properties of biochar amended sandy loam
soils. S. Soil Biology & Biochemistry, v.57, p.401–410, 2013
AMELOOT, N.; SLEUTEL, S.; CASE, S.D.C.; ALBERTI, G.; McNAMARRA,
N.P.; ZAVALLONI, C.; VERVISCH, B.; VEDOVE, G.D.; DE NEVE, S. C
mineralization and microbial activity in four biochar field experiments several
years after incorporation. Soil Biology & Biochemistry, v. 78, p. 195-203,
2014.
ÂNGELO, H.; SILVA, V. S. M.; A. N. SOUZA.; GATTO, A. C. Aspectos
financeiros da produção de Teca no Estado de Mato Grosso. FLORESTA,
Curitiba, PR, v. 39, n. 1, p. 23-32, jan./mar. 2009.
ATKINSON, C. J.; FITZGERALD, J. D.; HIPPS, N. A. Potential mechanisms for
achieving agricultural benefits from biochar application to temperate soils: a
review. Plant Soil, v. 337, p. 1–18, 2010.
BAYER, C.; MARTIN-NETO, L.; MIELNICZUK, J.; PAVINATO, A.
Armazenamento de carbono em frações lábeis da matéria orgânica de um
Latossolo Vermelho sob plantio direto. Pesquisa Agropecuária Brasileira,
v.39, p.677-683, 2004.
BAYER, C.; MIELNICZUK, J. Dinâmica e função da matéria orgânica. In:
SANTOS, G.A.; CAMARGO, F.A.O. Fundamentos da matéria orgânica:
Ecossistemas tropicais e subtropicias. Genesis Press, Porto Alegre, RS,
Brasil, p.9-26, 1999.
BIRD, M. I. et al. Stability of elemental carbon in savanna soil. Global
Biogeochemical Cycles, Washington, DC, v. 13, n. 4, p. 923-932, 1999.
BREWER, C.E., BROWN, R.C. Biochar. In: Sayigh, A. (Ed.), Comprehensive
Renewable Energy. Elsevier, Oxford, p. 357–384, 2012.

29 BRUGGEN, A. H. C.; SEMENOV, A. M. In search of biological indicators for soil
health and disease suppression. Applied Soil Ecology, v. 15, p.13-24, 2000.
CAMARGO, F.A.O.; SANTOS, G.A.; GUERRA, J.G.M. Macromoléculas e
substancias húmicas. In: SANTOS, G.A.; CAMARGO, F.A.O. Fundamentos da
matéria orgânica do solo: ecossistemas tropicias e subtropicias. Porto Alegre:
Genesis, p.183-196, 1998.
CARDENAS, E.; TIEDJE, J. M. New tools for discovering and characterizing
microbial diversity. Current Opnion in Biotechnology, Amsterdam, v. 19 p.
544-549, 2008.
CARDOSO, E. J. B. N.; FREITAS, S. S. A rizosfera. In: CARDOSO, E. J. B. N.;
TSAI, S. M.; NEVES, M. C. P. Microbiologia do solo. Campinas: Sociedade
Brasileira de Ciência do Solo, 1992. p.41-57.
CARTER, M. R. Microbial biomass as na index for tillage-induced changes in
soil biological properties. Soil and Tillage Research, v. 7, p. 29-40, 1986.
CATTELAN, A. J.; VIDOR, C. Flutuações na biomassa, atividade e população
microbiana do solo, em função de variações ambientais. Revista Brasileira de
Ciência do Solo, v.14, p.133-142, 1990.
CHAVEZ.L.F.; ESCCOBAR.L.F.; ANGHINONI.I.; CARVALHO.P.C.F.;
MEURER.E.J. Diversidade metabólica e atividade microbiana no solo em
sistema de integração lavoura-pecuaria sob intensidade de pastejo. Pesquisa
Agropecuaria Brasileira. Brasilia. V.46 n.10 p.1254-1261, 2011.
CHEN, C.R.; XU, Z.H.; MATHERS, N.J. Soil carbon pools in adjacent natural
and plantation forests of subtropical Australia. Soil Science Society of
America Journal, v.68, p.282-291, 2004.
COHEN-OFRI, I.; WEINER, L.; BOARETTO, E.; MINTZ, G.; WEINER, S.
Modern and fossil charcoal: aspects of structure and diagenesis. Journal of
Archaeological Science, Boston, v. 33, n. 3, p. 428-439, 2006.

30 CORAZZA, E.J.; SILVA, J.E.; RESK, D.V.S.; GOMES, A.C. Comportamento de
diferentes sistemas de manejo como fonte ou depósito de carbono em relação
a vegetação do cerrado. Revista Brasileira de Ciência do Solo, v.23, p.425-
432, 1999.
DOMINGUES, M.S.; BERMANN, C. O arco de desflorestamento na Amazônia:
da pecuária á soja. Ambiente e Sociedade, v.15, 2012.
http://dx.doi.org/10.1590/S1414-753X2012000200002.
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - Embrapa. Centro
Nacional de Pesquisa de Solos. Manual de métodos de análises de solo. 2.
ed. Rio de Janeiro: Embrapa Solos, 1997. 212p.
FINGER, Z; FINGER, F. A.; DRESCHER, R. Teca (Tectona grandis L.f.): plante
esta idéia. In: Simpósio Brasileiro de Pós-graduação em Engenharia, 1., 2001,
Santa Maria-RS. Anais... Santa Maria: UFSM, 2001. CD-Rom.
FONTANA, A. Caracterização química e espectroscópica da matéria
orgânica em solos do Brasil. Dissertação (Mestrado em Agronomia: Ciência
do solo) – Instituto de Agronomia, Departamento de solos, Universidade
Federal do Rio de Janeiro, Seropédica, 2006.
FREIXO, A.A.; DE MACHADO, P.L.O.A.; SANTOS, H.P.; SILVA, C.A.;
FADIGAS, F.S. Soil organic carbon and fraction of a Rhodic Ferrasol under the
influence of tillage and crop rotation systems in southern Brazil. Soil & Tillage
Research, v.64, p.221-230, 2002.
FROMIN, N.; HAMELIN, J.; TARNAWSKI, S.; ROESTI, D.; JOURDAIN-
MISEREZ, K.; FORESTIER, N.; TEYSSIER-CUVELLE, S.; GILLET, F.;
ARAGNO, M.; ROSSI, P. Statistical analysis of denaturing gel
electrophoresis (DGE) fingerprinting patterns. Environmental
Microbiology; v. 4, n. 11, p. 634-643, 2002.

31 GLASER, B.; HAUMEIER, L.; GUGGENBERGER, G.; ZECH, W. The “Terra
Preta” Phenomenon: A model for sustainable agriculture in the humid tropics’,
Naturwissenschaften, vol 88, p. 37–41, 2001.
GLASER, B.; LEHMANN, J.; ZECH, W. Ameliorating physical and chemical
properties of highly weathered soil in the tropic with charcoal – a review.
Biology and Fertility of Soils, Berlin, v. 35, n. 4, p. 219-230. 2002.
GLASER, B; GUGGENBERGER, G.; ZECH, W; RUIVO, M. L.. Soil organic
matter stability in Amazonian Dark Earths. In: LEHMANN, J. et al. (Eds.).
Amazonian dark earths: origin, properties, management. Dordrecht : Kluwer
Academic, p. 141-158, 2003.
HEUER, H.; KRSEK, M.; BAKER, P.; SMALLA, K.; WELLINGTON, E.M.H.
Analysis of the actinomycete communities by specific amplification of genes
encoding 16S rRNA and gel-electrophoresis separation in denaturing gradients.
Applied & Environmental Microbiology, v. 63, p. 3233-3241, 1997.
HU, L., CAO, L., ZHANG, R. Bacterial and fungal taxon changes in soil
microbial community composition induced by short-term biochar amendment in
red oxidized loam soil. World J. Microb. Biotechnol, v.30, p.1085–1092, 2014.
JOSEPH, S.; CAMPS-ARBESTAIN, M.; LIN, Y.; MUNROE, P.; CHIA, C. H.;
HOOK, J.; VAN ZWIETEN, L.; KIMBER, S.; COWIE, A.; SINGH, B. P.;
LEHMANN, J.; FOIDL, N.; SMERNIK, R. J.; AMONETTE, J. E. An investigation
into the reactions of biochar in soil. Australian Journal of Soil Research, v.
48, p. 501–515, 2010.
KARLEN, D. L.; DITZLER, C. A.; ANDREWS, S. S. Soil quality: why and how?
Geoderma, Amsterdam, v. 114, p. 145-156, 2003.
KARLEN, D. L.; DITZLER, C. A.; ANDREWS, S. S. Soil quality: why and how?
Geoderma, Amsterdam, v. 114, p. 145-156, 2003.

32 LEHMANN, J., RILLIG, M. C.; THIES, J.; MASIELLO, C. A.; HOCKADAY, W.
C.; CROWLEY, D. Biochar effects on soil biota – A review. Soil Biology &
Biochemistry, v. 43, p. 1812-1836, 2011.
LEHMANN, J.; JOSEPH, S. Biochar for Enviromental Management: An
Introduction. In: LEHMANN, J.; SOSEPH, S. (ed). Biochar for Enviromental
Management: Science and Technology. 1. Ed. earthscan, Londres, 2009. p.
4-18.
LISBOA, B. B.; VARGAS, L. K.; SILVEIRA, A. O. da; MARTINS, A. F.;
SELBACH, P. A. Indicadores Microbianos de Qualidade do Solo em Diferentes
Sistemas de Manejo. Revista Brasileira de Ciência do Solo, v. 36, p. 45-55,
2012.
MACEDO, M.O.; RESENDE, A.S.; GARCIA, P.C.; BODDEY, R.M.; JANTALIA,
C.P.; URQUIAGA, S.; CAMPELLO, E.F.C.; FRANCO, A.A. Changes in soil C
and N stocks and nutrient dynamics 13 years after recovery of degraded land
using leguminous nitrogen-fixing trees. Forest Ecology and Management,
v.255, p.1516-1524, 2008.
MADARI, B. E. et al. Matéria orgânica dos solos antrópicos da Amazônia (Terra
Preta de Índio): suas características e papel na sustentabilidade da fertilidade
do solo. In: TEIXEIRA, W. G. et al. As terras pretas de índio da Amazônia:
sua caracterização e uso deste conhecimento na criação de novas áreas.
Manaus: EMBRAPA Amazônia Ocidental, p. 174-187, 2009.
MAJOR, J.; RONDON, M.; MOLINA, D.; RIHA, S.J.; LEHMANN, J. Maize yield
and nutrition during 4 years after biochar application to a Colombian savanna
Oxisol. Plant and Soil, v. 333, p. 117–128, 2010.
MATSUOKA, M. Atributos biológicos de solo cultivados com videira na
região da Serra Gaúcha. 2006. 152f. Tese (Doutorado em Ciência do Solo) –
Universidade Federaldo Rio Grande do Sul, Porto Alegre - RS, 2006.

33 MOREIRA, F. M. S.; SIQUEIRA, J. O. Microbiologia e Bioquímica do Solo.
Lavras: UFLA, 2006. 729p.
MUYZER, G.; SMALLA, K. Application of denaturing gradient gel
electrophoresis (DGGE) and temperature gradient gel electrophoresis (TGGE)
in microbial ecology. Antonie Van Leewenhoek, v. 73, p. 127-141, 1998.
MUYZER, G.; WALL, E. C. DE; UITTERLINDEN, A. G. Profiling of complex
microbial populations by denaturing gradient gel electrophoresis analysis of
polymerase chain reaction-amplified genes coding for 16S rRNA. Applied and
Environmental Microbiology, Baltimore, v. 59, p. 695-700, 1993.
NOVOTNY, E.; HAYES, M. H. B.; MADARI, B. E.; BONAGAMBA, T. J.;
AZEVEDO, E. R. de; SOUZA, A. A. de; SONG, G.; NOGUEIRA, C. M.;
MANGRICH, A. S. Lessons from the Terra Preta de Índios of the Amazon
Region for the utilization of charcoal for soil amendment. Journal of the
Brazilian Chemical Society, v. 20, n. 6, p. 1003-1010, 2009.
OMIL, B.; PIÑEIRO, V.; MERINO, A. Soil and tree responses to the application
of wood ash containing charcoal in two soils with contrasting properties. Forest
Ecology and Management, v. 295, p.199–212, 2013.
ØVEREÅS, L.; FORNEY, L.; DAAE, F. L.; TORSVIK, V. Distribution of
bacterioplankton in meromictic lake Sælenvannet, as determined by Denaturing
Gradient Gel Electrophoresis of PCR-amplified gene fragments coding for 16s
rRNA. Applied and Environmental Microbiology, Baltimore, v. 63, p. 3367-
3373, 1997.
PETTER, F. A.; MADARI, B. E.; SILVA, M. A. S.; CARNEIRO, M. A. C.;
CARVALHO, M. T. ,.; MARIMON JR, B. H.; PACHECO, L. P. Soil fertility and
upland rice yield after biochar application in the Cerrado. Pesquisa
Agropecuaria Brasileira, Brasília, v.47, n.5, p.699-706, 2012.

34 QUILLIAM, R.S., GLANVILLE, H.C., WADE, S.C., JONES, D.L. Life in the
‘charosphere’ – does biochar in agricultural soil provide a significant habitat for
microorganisms?. Soil Biology & Biochemistry, v.65, p.287–293, 2013.
RACHID, C. T. C. C. Comunidade bacteriana, atributos do solo e fluxo de
gases em solo sob Cerrado e cana-de-açucar. 2010, 126f. Dissertação
(Mestrado em Quimica na Agricultura e no Ambiente). Centro de Energia
Nuclear na Agricultura- Universidade de São Paulo – USP, Piracicaba- SP,
2010.
RIBEIRO, G.H.S. Qualidade biológica de solos de cerrado em áreas
degradadas e sob diferentes manejos de cana-de-açucar. Dissertação
(Mestrado). Universidade Católica de Brasília- UCB, Brasília, 2012.
RUTIGLIANO, F.A.; ROMANO, M.; MARZAOLLI, R.; BAGLIVO, L.; BARONTI,
S.; MIGLIETTA, F.; CASTALDI, S. Effect of biochar addition on soil microbial
community in a wheat crop. European Journal of soil Biology. V.60, p. 9-15,
2014.
SETH, S.K., KAUL, O.N. Tropical forest ecosystems of India: the teak forests
(as a case study of silviculture and management). Tropical Forest
Ecosystems: A State-of-knowledge Report. UNESCO, Paris, pp. 628–640,
1978.
SHIMIZU, J.Y.; KLEIN, H; OLIVEIRA, J.R.V. de. Diagnóstico das plantações
florestais em Mato Grosso. Cuiabá, MT: Central de Texto, 2007.
SIQUEIRA, J. O.; FRANCO, A. A. Biotecnologia do solo: Fundamentos e
perspectivas. Brasília: MEC Ministério da Educação, ABEAS; Lavras: ESAL,
FAEPE, 1988. 236p.
SOHI, S. P.; KRULL, E.; LOPEZ-CAPEL, R.; BOL, A. Review of biochar and its
use and function in soil. In: SPARKS, D. L. (Ed.). Advances in Agronomy.
Burlington: Academic Press, 2010, p.47-82.

35 SPERA, S. T.; SANTOS, H. P. dos; FONTANELI, R. S.; TOMMM, G. O.
Integração lavoura e pecuária e os atributos físicos de solo manejado sob
sistema plantio direto. Revista Brasileira de Ciência do Solo, v. 33, p.130,
2009.
SPOSITO, G.; ZABEL, A.The assessment of soil quality. Geoderma, v. 114, p.
143-144, 2003.
STEINER, C.; TEIXEIRA, W. G.; LEHMANN, J.; NEHLS, T.; de MACÊDO, J. J.
V.; BLUM, W. E. H.; ZECH, W. Long term efects of manure, charcoal and
mineral fertilization on crop production and fertility on a highly weathered
Central Amazonian upland soil. Plant and Soil, The Hague, v. 291, n. 1-2, p.
275-290, 2007.
SUN, D., MENG, J., CHEN, W. Effects of abiotic components induced by
biochar on microbial communities. Acta Agric. Scand, v.63, p.633–641, 2013.
TABATABAI, M. A. Soil enzymes. In: WEAVER, R. W.; SCOTT, A.;
BOTTOMELEY, P. J. Methods of soil analysis: microbiological and
biochemical properties. Madison: Soil Science Society of America, Part 2.
1994. p. 778-835. (Special Publication, 5).
THIES, J. E.; RILLIG, M. C. Characteristics of Biochar: biological Properties. In:
LEHMANN, J.; JOSEPH, S. Biochar for environmental management:
Science and Technology. London: Earthscan. 2009. p. 85-106.
THOMPSON, F. L.; OIVEIRA V. M.; AZEVEDO, J. L.; ARAUJO, W. L.; INAVIO,
C. A.; SELEGHIM, M. H. R.; KITAJIMA, E. W. Taxonomia microbiana, de
procariontes, de fungos, de protozoários e de vírus. Brasília: Centro de Gestão
e Estudos Estratégicos, CGEE, 2005. 53 p.
VISSER, S.; PARKINSON, D. Soil Biological criteria as indicators of soil quality:
Soil microorganisms. American Journal of Alternative Agriculture, v. 7, p.
33-37, 1992.

36 WANG, X.; SONG, D.; LIANG, G.; ZHANG, Q.; AI, C.; ZHOU, W. Maize biochar
addition rate influences soil enzyme activity and microbial community
composition in a fluvo-aquic soil. Applied Soil Ecology, v.96, p.265-272, 2015.
WARNOCK, D. D.; LEHMANN, J.; KUYPER, T. W.; RILLIG, M. C. Mycorrhizal
responses to biochar in soil – concepts and mechanisms. Plant Soil, v. 300, p.
9–20, 2007.
XU, G.; SUN, J.; SHAO, H.; CHANG, S. Biochar had effects on phosphorus
sorption and desorption in three soils with differing acidity. Ecological
Engineering, v.62, p.54-60, 2014.
ZILLI, J. E.; RUMJANEK, N. G.; XAVIER, G. R.; COUTINHO, H. L. C.; NEVES,
M.C. P. Diversidade microbiana como indicador de qualidade do solo.
Cadernos de Ciência e Tecnologia, v. 20, p. 391-411, 2003.

37 CAPÍTULO 2 – EFEITO DA APLICAÇÃO DO BIOCARVÃO NOS ATRIBUTOS
MICROBIOLOGICOS DO SOLO, CULTIVADO COM TECA, NO BIOMA
AMAZÔNIA
RESUMO - Biocarvão é um material orgânico que contém um elevado teor de
carbono, produzido por pirólise, na ausência de oxigênio. Possui uma estrutura
aromática, tornando-o estável e altamente resistente a degradação química e
biológica no solo. O objetivo desta pesquisa foi avaliar o uso do biocarvão
como condicionador do solo e seu efeito na atividade da β-glicosidase,
fosfatase ácida e diversidade bacteriana do solo em experimento de Teca com
três e quinze meses de duração, em um latossolo vermelho-amarelo,
localizados em Sinop - Mato Grosso, Brasil. Os tratamentos consistiram de: T1-
Muda sem biocarvão; T2- Muda com biocarvão; T3- Muda sem biocarvão +
biocarvão ativado aplicado no solo; T4- Muda com biocarvão + biocarvão
ativado aplicado no solo; T5- Muda sem biocarvão + biocarvão não ativado
aplicado no solo; T6- Muda com biocarvão + biocarvão não ativado aplicado no
solo. Foi determinada a atividade de duas enzimas relacionadas ao ciclo do C e
P e a biodiversidade bacteriana foi avaliada por meio da técnica de DGGE com
iniciadores universais para a região 16S rDNA de bactérias. Não houve
incremento na atividade da β-glicosidase quinze meses após aplicação do
biocarvão, mas sua forma ativada proporcionou a maior atividade desta
enzima, ao contrário a fosfatase ácida apresentou uma baixa atividade nos
primeiros 3 meses e após quinze meses com aplicação do biocarvão no solo a
atividade desta enzima foi maior. Em relação à umidade e pH do solo, não foi
alterado pela presença do biocarvão. A estrutura e diversidade da comunidade
bacteriana no solo sofreram influencia mais acentuada do biocarvão com
quinze meses de aplicação no solo.
Palavras-chave: DGGE, condicionador de solo, β-glicosidase, fosfatase ácida, microbial ecology

38
1. INTRODUÇÃO
A Amazônia brasileira, uma das principais regiões produtoras de
madeira tropical do mundo, tem sofrido com a falta de oferta de madeira natural
legalizada. A Tectona grandis Linn f., ou simplesmente Teca, é uma espécie
que vem ganhando grande destaque dentro do cenário florestal brasileiro nos
últimos anos, sobremaneira no Estado de Mato Grosso, devido às condições
edafoclimáticas favoráveis (ZIECH, 2011).
A madeira da espécie é utilizada principalmente na indústria naval,
construção civil e indústria moveleira na fabricação de assoalhos e de decks.
Em Mato grosso a área plantada é de 48.526,2 há, e se encontra distribuída
em 42 municípios, indicando o interesse crescente por tal gênero florestal,
principalmente em função das expectativas de retorno financeiro em projetos
com a mesma (SHIMIZU et al., 2007). Atualmente, os plantios totalizam 67.329
ha e representam apenas 0,4% da produção de madeira plantada no Brasil
(ABRAF, 2013).
Embora a Teca seja encontrada em diversas regiões dentro da faixa
tropical e subtropical, a produtividade observada difere para cada uma destas
regiões (ZIECH, 2011). No estado de Mato Grosso os povoamentos de Teca
apresentam produtividades extremamente variáveis em função da diversidade
de condições física e química do solo (SHIMIZU et al., 2007).
O processo de transformação física e química do solo agrícola é
condicionado, em parte pela calagem, que apresenta ação dispersante de
agregados e, em parte, pela atividade biológica, que promove a mineralização
da matéria orgânica estável do solo (SPERA et al., 2009). Uma alternativa para
manter a matéria orgânica e melhorar a fertilidade do solo é o uso de resíduos
orgânicos carbonizados ou biocarvão (THIES et al., 2015). Esse biocarvão é
produzido por meio de um processo chamado pirólise o qual permite a
decomposição térmica do material orgânico em condições limitadas de
suprimento de oxigênio e em temperaturas em torno de 500ºC (LEHMANN E
JOSEPH, 2009).
Usado como condicionador de solo, o biocarvão pode aumentar a
disponibilidade de nutrientes para as plantas e melhorar as propriedades físicas

39 e biológicas do solo, promovendo um rearranjo dos átomos de carbono
aromáticos, aumentando a porosidade do material (AMELOOT et al.,2014). Isto
pode proporcionar melhoria nas propriedades químicas, principalmente pela
alta superfície específica resultando em aumento da capacidade de troca de
cátions e adsorção de nutrientes e tornando-o muito mais resistente à
decomposição microbiana (LEHMANN, 2007; ATKINSON et al., 2010).
A estrutura porosa do biocarvão serve como habitat para os
microrganismos do solo, devido à área superficial e a sua capacidade de
adsorver material orgânico solúvel, gases e nutrientes inorgânicos, podendo
providenciar um habitat para os microrganismos colonizar (LEHMANN E
JOSEPH, 2015). Além disso, a adição do biocarvão ao solo pode alterar as
propriedades físico-químicas do solo (estrutura e pH), os quais influenciam
diretamente na atividade e biomassa microbiana, bem como, na composição da
comunidade de microrganismos (LEHMANN et al., 2011).
Para a avaliação da qualidade de um solo, tem sido difundida também a
necessidade de identificação de parâmetros indicativos do seu estado de
conservação e/ou degradação.
Os estudos sobre bioindicadores mostram que os microrganismos do
solo, devido às características como abundância e atividades bioquímicas,
apresentam alto potencial de uso na avaliação da degradação do solo
(MOREIRA E SIQUEIRA, 2006). Segundo Araujo e Monteiro (2007), a atividade
enzimática do solo esta inter-relacionada com a matéria orgânica, com as
propriedades físicas e com a atividade e biomasssa microbiana podendo
funcionar como um bom indicador de mudanças na qualidade do solo.
Como resultado da sua sensibilidade as mudanças ambientais, a
atividade enzimática e a estrutura e abundancia da microbiota do solo tem sido
amplamente utilizados como indicadores das mudanças na qualidade do solo
(WANG et al., 2015). Atualmente, existem diversos métodos moleculares
disponíveis para o estudo de comunidades microbianas independentes de
cultivo. Eles se baseiam principalmente em técnicas que utilizam a
manipulação dos ácidos nucleicos, ou seja, o DNA ou o RNA extraído
diretamente do solo (RACHID, 2010).
Nesse sentido, diante do potencial do uso de biocarvão como alternativa
na melhoria e conservação dos solos e a possibilidade de usar a biologia

40 molecular para estudar o a comunidade microbiana no solo os objetivos deste
trabalho foram: i) avaliar as possíveis influencia dessa pratica na atividade
enzimática relacionada ao ciclo do carbono e fósforo; ii) verificar a influencia da
aplicação do biocarvão na diversidade bacteriana e na interação com o solo em
Sinop, Mato Grosso.
2. MATERIAIS E MÉTODOS
2.1. Descrição da Área de Estudo
O experimento foi implantado na área experimental da Embrapa
Agrossilvipastoril (Anexo A), no bioma Amazônia, município de Sinop, MT (11°
52′ 23″ Sul, 55° 29′ 54″ Oeste, a 384 m de altitude).
O clima da região segundo a classificação de Köppen é do tipo Am,
apresenta característica de clima de monção, com moderada estação seca e
ocorrência de precipitação média mensal inferior a 60 mm (ALVAREZ et al.,
2013).
O solo é classificado como Latossolo Vermelho Amarelo distrófico típico,
textura argilosa, horizonte A moderado e com relevo plano (DIEL et al., 2014).
Antes da implantação do experimento foram determinados os atributos
químicos e granulométricos na camada de 0–20 cm: pH em CaCL2 de 5,1; 13
mg dm-3 de P (Mehlich-1); 62 mg dm-3 de K (Mehlich-1); 2,6 cmolc dm-3 de Ca;
0,7 cmolc dm-3 de Mg; 0,0 cmolc dm-3 de Al; CTC de 7,4 cmolc dm-3 e 30,4 g
dm-3 de matéria orgânica (MO). A textura do solo é argilosa com 283 g kg-1 de
areia, 150 g kg-1 de silte e 567 g kg-1 de argila. Para correção da acidez,
aplicou-se 1,5 ton.ha-1 de calcário em área total de acordo com as indicações
agronômicas.
Foram analisados os dados climáticos como: precipitação, umidade e
temperatura médias mensais, de outubro de 2013 a abril de 2015, para
observações das alterações climáticas cumulativas de 6 meses anteriormente
as coletas. A precipitação total acumulada durante o período da chuva
(outubro-abril) em 2014 foi de 2200,55 mm com a temperatura média de 25º C
e umidade relativa do ar de 82,06%. No período da chuva em 2015 a
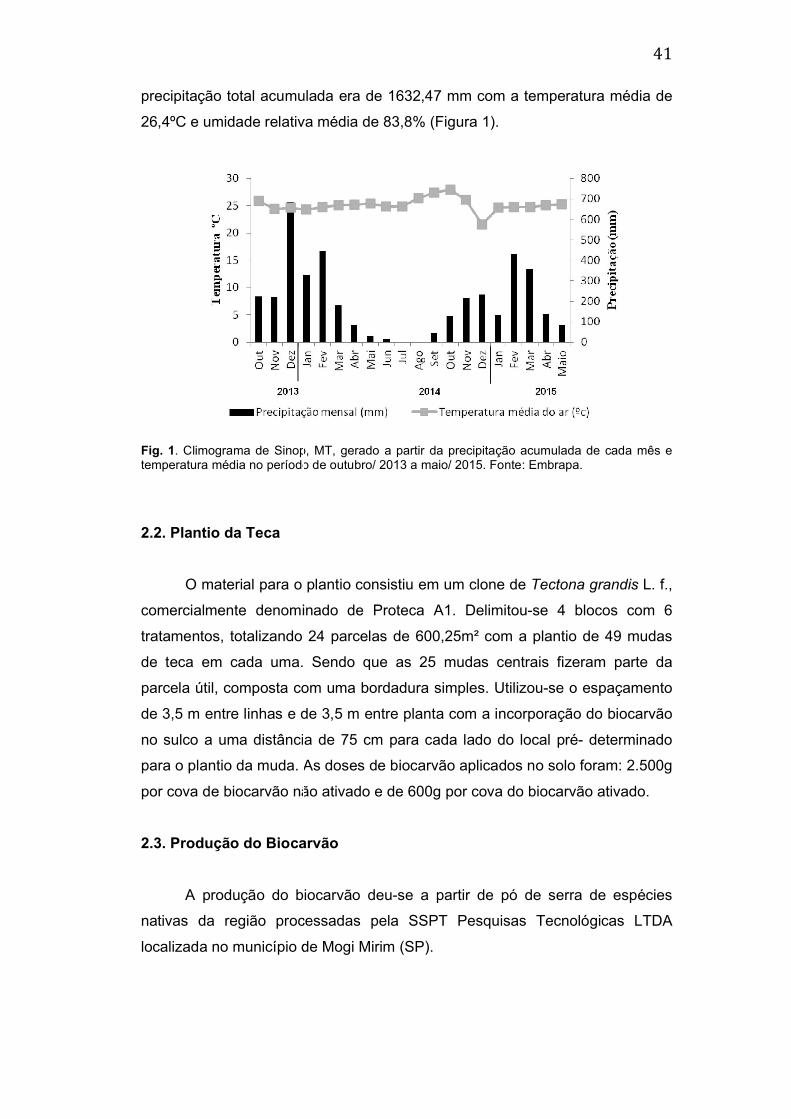
precipitação total acumulada era de 1632,47 mm com a temperatura média de
26,4ºC e umidade relativa média de 83,8% (Fig
Fig. 1. Climograma de Sinop, MT, gerado a partir da precipitação acumulada de cada mês e temperatura média no período de outubro/ 2013 a maio/ 2015. Fonte: Embrapa.
2.2. Plantio da Teca
O material para o plantio consistiu em um clone de
comercialmente denominado de Proteca A1. Delimitou
tratamentos, totalizando 24 parcelas de 600,25m² com a plantio de 49 mudas
de teca em cada uma. Sendo que as 25 mudas centrais fizeram parte da
parcela útil, composta com uma bordadura simples. Utilizou
de 3,5 m entre linhas e de 3,5 m entre planta com a
no sulco a uma distância de 75 cm para cada lado do local pré
para o plantio da muda. As doses de biocarvão aplicados no solo foram: 2.500g
por cova de biocarvão não ativado e de 600g por cova do biocarvão ativado.
2.3. Produção do Biocarvão
A produção do biocarvão deu
nativas da região processadas pela SSPT Pesquisas Tecnológicas LTDA
localizada no município de Mogi Mirim (SP).
precipitação total acumulada era de 1632,47 mm com a temperatura média de
26,4ºC e umidade relativa média de 83,8% (Figura 1).
. Climograma de Sinop, MT, gerado a partir da precipitação acumulada de cada mês e mperatura média no período de outubro/ 2013 a maio/ 2015. Fonte: Embrapa.
O material para o plantio consistiu em um clone de Tectona grandis
comercialmente denominado de Proteca A1. Delimitou-se 4 blocos com 6
talizando 24 parcelas de 600,25m² com a plantio de 49 mudas
de teca em cada uma. Sendo que as 25 mudas centrais fizeram parte da
parcela útil, composta com uma bordadura simples. Utilizou-se o espaçamento
de 3,5 m entre linhas e de 3,5 m entre planta com a incorporação do biocarvão
no sulco a uma distância de 75 cm para cada lado do local pré
para o plantio da muda. As doses de biocarvão aplicados no solo foram: 2.500g
por cova de biocarvão não ativado e de 600g por cova do biocarvão ativado.
2.3. Produção do Biocarvão
A produção do biocarvão deu-se a partir de pó de serra de espécies
nativas da região processadas pela SSPT Pesquisas Tecnológicas LTDA
localizada no município de Mogi Mirim (SP).
41
precipitação total acumulada era de 1632,47 mm com a temperatura média de
. Climograma de Sinop, MT, gerado a partir da precipitação acumulada de cada mês e mperatura média no período de outubro/ 2013 a maio/ 2015. Fonte: Embrapa.
Tectona grandis L. f.,
se 4 blocos com 6
talizando 24 parcelas de 600,25m² com a plantio de 49 mudas
de teca em cada uma. Sendo que as 25 mudas centrais fizeram parte da
se o espaçamento
incorporação do biocarvão
no sulco a uma distância de 75 cm para cada lado do local pré- determinado
para o plantio da muda. As doses de biocarvão aplicados no solo foram: 2.500g
por cova de biocarvão não ativado e de 600g por cova do biocarvão ativado.
se a partir de pó de serra de espécies
nativas da região processadas pela SSPT Pesquisas Tecnológicas LTDA

42
Foram utilizados dois tipos de biocarvão: não ativado e ativado. Para
obtenção do biocarvão não ativado processou-se o pó de serra em reator de
pirólise lenta (Forno Vertical), com 25 minutos de residência no forno, a 450°C.
Para obtenção do biocarvão ativado foi necessário aumentar a temperatura do
reator de pirólise (Forno Tubular Horizontal) para 650°C e injetar vapor de água
durante o processo. O tratamento de ativação realizou-se com intuito de
eliminar resíduos do processo de pirólise, desobstruir os poros do biocarvão e
disponibilizar radicais orgânicos na superfície do carvão.
2.4. Tratamentos
Os tratamentos consistiram de: T1- Muda sem biocarvão; T2- Muda com
biocarvão; T3- Muda sem biocarvão + biocarvão ativado aplicado no solo; T4-
Muda com biocarvão + biocarvão ativado aplicado no solo; T5- Muda sem
biocarvão + biocarvão não ativado aplicado no solo; T6- Muda com biocarvão +
biocarvão não ativado aplicado no solo. Aplicou-se em todos os tratamentos a
mesma quantidade de adubo: 60 g por cova de NPK (20-00-20), 120g por cova
de P2O5 e 12 g por cova de FTE BR (3,9% enxofre, 1,8% de boro, 2,0% de
manganês e 9,0% de zinco).
Foram utilizados dois tipos de muda: muda com substrato composto de
100% substrato comercial (Mac Plant) e muda com substrato composto de 25%
de biocarvão ativado + 75% de substrato comercial.
2.5. Área de Referência
A 1ª área denominada de Pousio está localizada próximo aos blocos do
experimento da fazenda experimental da Embrapa Agrossilvipastoril e desde
2010 deixou de ser área agrícola sendo naturalmente revegetada por
gramíneas invasoras.
A 2ª área está coberta por Mata Nativa composta por Floresta Estacional
Perenifolia (HAIDAR et al., 2013). O fragmento avaliado está localizado
próximo à área experimental da fazenda da Embrapa Agrossilvipastoril,
formando corredor ecológico a partir da mata ciliar do rio Teles Pires com
extensão total de 7,3 km e largura média de 200 m.

43 2.6. Amostragem de solo
A coleta das amostras de solo foi em abril dos anos de 2014
(experimento com três meses no campo) e 2015 (experimento com quinze
meses no campo), caracterizado pelo final do período chuvoso. Com auxilio de
um trado tipo holandês, as amostras de solo deformadas foram coletadas na
profundidade de 0-10 cm. Em cada tratamento foram coletadas 20 amostras
simples para formar uma amostra composta por parcela. As coletas foram
realizadas a 40 cm de distancia entre as plantas e sempre na área útil da
mesma.
O solo coletado foi homogeneizado, acondicionado em caixas térmicas
contendo gelo e transportados para ao Laboratório de Microbiologia do Solo e
Biologia Molecular da Embrapa Agrossilvipastoril. As amostras foram
armazenadas em câmara fria a ± 4 °C até a execução das análises de
atividade enzimática. Para as analises moleculares, o solo foi armazenado a -
80ºC.
2.7. Umidade e pH do solo
O teor de umidade do solo foi realizado por meio do método da umidade
atual, onde as amostras de campo foram secas em estufa a 105 °C, por 24
horas, ao final o peso da água foi obtido pela diferença entre o peso da
amostra úmida e seca (EMBRAPA, 1997). O pH em água foi determinado para
cada tratamento nos dois períodos avaliados (EMBRAPA, 1997).
2.8. Atividade Enzimática
Foi determinada a atividade de duas enzimas: β-glicosidase e fosfatase
ácida conforme o método proposto por (TABATABAI, 1994). O principio do
método é baseado na determinação colorimétrica do p-nitrofenol liberado pela
ação da enzima, quando o solo é incubado com substrato especifico. Para as
análises de β-glicosidase o substrato utilizado na reação foi o p-nitrofenil- β-D-
glucopiranosídeo 0,05M (PNG 0,05M) e para a Fosfatase Ácida o substrato
utilizado na reação foi o p-nitrophenyl phosphate 0,05M (PNF 0,05M).

44
Para cada amostra de solo coletada no campo, necessitou duas
repetições analíticas no laboratório, mais o controle. A concentração de p-
nitrofenol produzido foi estimada a partir de uma curva padrão preparada com
concentrações conhecidas de p- nitrofenol (0; 10; 20; 30; 40 e 50 μg p-
nitrofenol mL-1) com R2 > 0,99, na regressão linear, após, os resultados foram
expressos em μg p-nitrofenol g-1 solo seco h-1 (TABATABAI E BREMNER,
1969).
2.9. Caracterização da comunidade de bactérias (gene 16S rDNA)
2.9.1. Extração de DNA e Reação em Cadeia de Polimerase (PCR)
O DNA genômico do solo foi extraído com PowerSoil™ DNA isolation kit
(MoBio Laboratories, Carlsbad, CA., USA), conforme protocolo descrito pelo
fabricante. A qualidade do material extraído foi verificada em gel de agarose a
0,8% e a quantificação do mesmo foi realizada em NanoDrop (Thermo
Scientific).
Na amplificação do gene 16S rDNA, utilizou-se o seguinte conjunto de
primers: iniciador 799F (5’- AACMGGATTAGATACCKG-3’) e finalizador 1492R
(5’- TACGGYTACCTTGTTACG-ACT-3’). Foram realizadas duas reações de
PCR, para evitar a amplificação de possíveis DNAs de cloroplastos presentes
no material extraído do solo (CHELIUS E TRIPLETT, 2001). A solução da
primeira reação continha: 1 μL do DNA total extraído (5-20 ng), 2,5 μL de
tampão 10x com MgCl2 (Sigma), 0,2 mM de cada desoxirribonucleotídeos
trifosfato, 0,4 μM de cada primer, 2,5 U de Taq DNA polimerase (Sigma), 0,2
mg de BSA (albumina de soro bovino) e água ultra pura estéril (Milli-Q) para
volume final de 25 μL.
A primeira PCR foi conduzida em termociclador (Bio-Rad T100TM
Thermal Cycler) programado para realizar desnaturação inicial a 95ºC por 3
min, seguido de 35 ciclos a 94°C por 20 seg, 53ºC por 40 seg, 72ºC por 40 seg
e com extensão final de 72ºC por 7 min. O produto da PCR foi corado com
GelRedTM 20x aplicado em gel de agarose 1,5 % (p/v), submetido a
eletroforese de 70 V por 55 min e observado sobre luz ultravioleta. Para cada
amostra, dois fragmentos de DNA foram observados, correspondendo a 1090

45 pb e 735 pb referentes ao DNA mitocondrial e bacteriano respectivamente,
posteriormente o gel foi fotodocumentado (Loccus Biotecnologia).
Os amplicons resultantes dessa amplificação foram utilizados como
moldes para uma nova PCR-2 especifica que abrangem a região V3 do DNA
ribossômico 16S (OVREAS et al., 1997), realizados com o conjunto de primers:
U968GC (5’-grampo CG+ AACGCGAAGAACCTTAC-3’) e 378R (5’-
CGGTGTGTACAAGGCCCGGGAACG-3’). A amplificação foi feita em solução
contendo: 0,5 μL do produto da PCR1, 0,4 μM de ambos os primers, 2,5 μL de
tampão 10x com MgCl2 (Sigma), 0,2 mM de cada desoxirribonucleotídeos
trifosfato, 2,5 U de Taq DNA polimerase (Sigma), 1 % de formamida e água
ultrapura estéril (Milli-Q) para volume final de 25 μL.
A reação foi programada para realizar desnaturação inicial a 94ºC por 4
min, seguido de 35 ciclos a 94ºC por 1 min, 56ºC por 1 min, 72ºC por 1 min e
extensão final de 72ºC por 10 min. O resultado das amplificações foi verificado
em gel de agarose 1,5 % (p/v), coradas com GelRedTM 20x, submetidas a
corrente de 70 Volts por 55 min e observadas sobre luz ultravioleta. A presença
e o tamanho do gene bacteriano amplificado, 410pb foi certificado e
fotodocumentado (Loccus Biotecnologia). A quantificação do mesmo foi
realizada em NanoDrop (Thermo Scientific).
2.9.2. Gel em Eletroforese com Gradiente Desnaturante (DGGE)
Os produtos amplificados na PCR-2 foram analisados através da técnica
de DGGE utilizando-se um equipamento DcodeTM Universal Mutation
Detection System (Bio-Rad Inc., Hercules, USA). Os géis de acrilamida:bis-
acrilamida (37:1 m:m) 6% (p/v), preparados com gradiente desnaturante linear
de 35% - 65%, os gradientes formados a partir de soluções estoque de
poliacrilamida, contendo 0 a 100% de desnaturante (100 % corresponde a 7 M
de ureia e 30% de formamida). Aproximadamente 200 ng foram aplicados em
cada slot do gel. O tempo de eletroforese foi de 17 horas a 60ºC e 60v em TAE
1,0 X (10 Mm de Tris-Acetato e 0,5 Mm de EDTA, pH 8,0). Após a eletroforese,
os géis foram corados por 30 minutos com GelRedTM, fotografados em luz UV,
utilizando-se um sistema de captura de imagem (Loccus Biotecnologia) e

46 analisados através do programa BioNumerics versão 7.1 (Applied Maths, Sint
Martens Latem, Bélgica).
A determinação da diversidade de bactérias totais no solo foi por meio
do Índice de Shannon (H’) baseada na intensidade das bandas presentes no
gel do DGGE em que foi considerado o número total de bandas da canaleta e a
intensidade individual de cada banda.
2.10. Análise Estatística
Todos os dados de atividade enzimática, índice de shannon, pH e
umidade do solo foram analisados em relação à normalidade (Kolmogorov-
Sminov & Liliefors e Shapiro-Wilk’s) e homocedasticidade (Cochran C, Hartley,
Bartlett). Quando essas condições foram respeitadas, foi executada a análise
de variância (ANOVA) seguida do teste de Tukey para diferenciação de média
(ao nível de 5% de probabilidade) nos casos de diferenças significativas.
O Índice de Shannon (H’), que indica a diversidade de espécies, foi
determinado pelo número de bandas presentes em cada amostra do gel e o
total do número de indivíduos pela intensidade das bandas utilizando o
programa Bionumerics versão 7.1 (Applied Maths, Sint Martens Latem,
Bélgica). O resultado desse índice foi submetido á análise de variância e as
comparações de médias foram feitas através do teste de Tukey (p,<0,05). Para
a análise da estrutura das comunidades microbianas, foram geradas matrizes
de presença e ausência das bandas detectadas nos perfis do gel do DGGE.
Essa matriz foi utilizada na analise de componentes principais (PCA) para cada
ano de experimento.
Com os dados de presença e ausência de bandas, foi primeiramente
realisada a analise de correlação (DCA) entre as amostras e variáveis
(bandas), que forneceu informação sobre a distribuição dos dados de espécies
em cada grupo microbiano avaliado. Em casos onde a distribuição foi linear
(valor do primeiro eixo <4,00), a análise multivariada foi realizada por análise
de redundância (RDA), enquanto que valores >4,00 (distribuição normal)
indicam a melhor análise por correspondência canônica (CCA) (ANDREOTE et
al., 2009).

47
A correlação dos dados ambientais com a ocorrência das bandas foi
determinada pela Análise de Redundância (RDA), realizada no software
Canoco (Canoco 4.5, Biometris, Wageningen, Holanda). As bandas observadas
nos géis de DGGE foram consideradas como espécies e o teste estatístico de
permutação de Monte Carlo foi aplicado considerando 999 permutações
aleatórias, permitindo avaliar a significância (valor de p) dos fatores ambientais
na distribuição de espécies das amostras. Nestas análises foram também
obtidos os valores de λA (Lambda-1), que mostram as porcentagens das
variâncias explicadas pelos fatores ambientais independentemente. Como
variáveis ambientais foram usadas a atividade das enzimas β-glicosidase e
fosfatase ácida, Índice de Shannon (H’), pH e Umidade Gravimétrica do solo.
4. RESULTADOS
4.1. Atividade Enzimática, pH e umidade do solo
A atividade da enzima β-glicosidase apresentou um comportamento
distinto quanto a aplicação do biocarvão no solo nos primeiros 3 meses de
implantação em 2014 (Figura 2A). Na enzima β-glicosidase, os maiores valores
de atividade foram obtidos nos tratamentos com a aplicação do biocarvão em
sua forma ativada no solo, T3 e T4 (51,91 e 58,77 μg p-nitrofenol h-1 g solo
seco-1, respectivamente) e a menor atividade obtida foi no tratamento T5 com
aplicação do biocarvão não ativado (39,58 μg p-nitrofenol h-1 g solo seco-1).
A atividade enzimática após 15 meses de experimento com biocarvão
também apresentou a maior atividade da β-glicosidase no tratamento T3 com o
uso do biocarvão ativado (65,59 μg p-nitrofenol h-1 g solo seco-1) e a menor
atividade nos tratamentos T5 e T6 (37,07 e 35,22 μg p-nitrofenol h-1 g solo
seco-1, respectivamente) utilizando o biocarvão não ativado (Figura 2B). As
áreas de referência nos dois anos avaliados apresentaram valores de β-
glicosidase menores quando comparados com os tratamentos com o uso do
biocarvão.
Após três meses de aplicação do biocarvão no solo, a maior atividade da
fosfatase ácida foi encontrada foi no tratamento T1 (controle). Os tratamentos
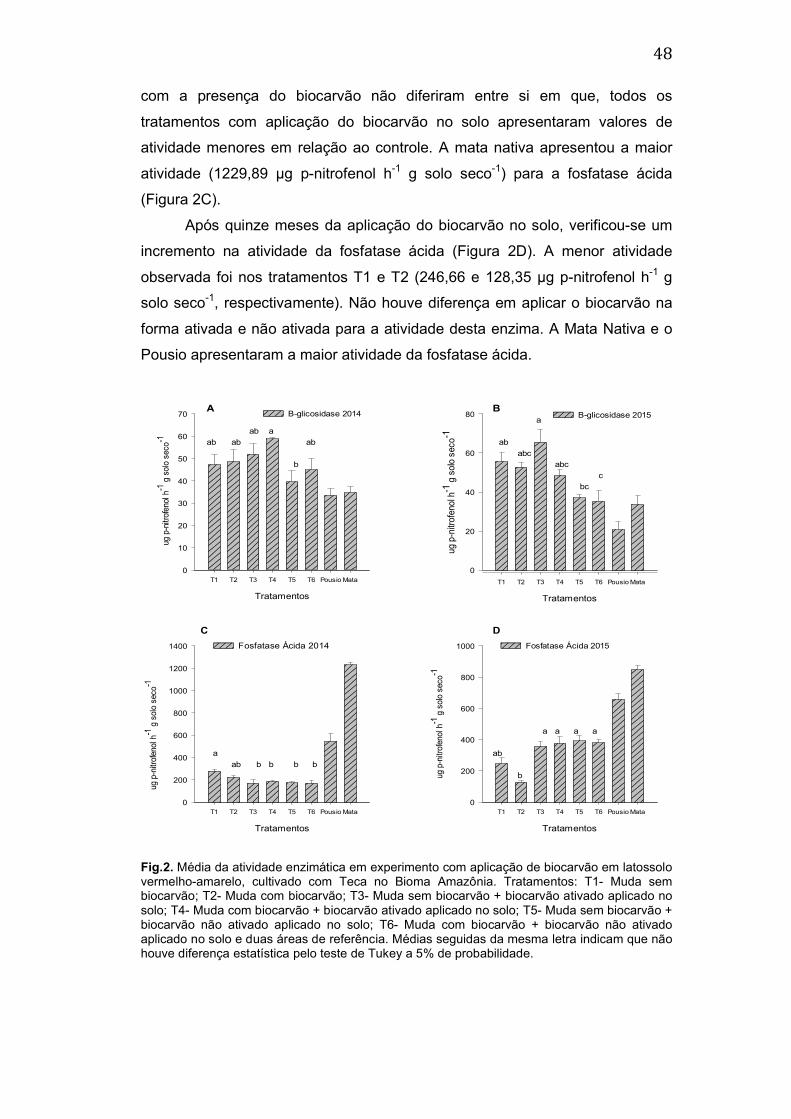
48 com a presença do biocarvão não diferiram entre si em que, todos os
tratamentos com aplicação do biocarvão no solo apresentaram valores de
atividade menores em relação ao controle. A mata nativa apresentou a maior
atividade (1229,89 μg p-nitrofenol h-1 g solo seco-1) para a fosfatase ácida
(Figura 2C).
Após quinze meses da aplicação do biocarvão no solo, verificou-se um
incremento na atividade da fosfatase ácida (Figura 2D). A menor atividade
observada foi nos tratamentos T1 e T2 (246,66 e 128,35 μg p-nitrofenol h-1 g
solo seco-1, respectivamente). Não houve diferença em aplicar o biocarvão na
forma ativada e não ativada para a atividade desta enzima. A Mata Nativa e o
Pousio apresentaram a maior atividade da fosfatase ácida.
Tratamentos
T1 T2 T3 T4 T5 T6 Pousio Mata
0
200
400
600
800
1000
1200
1400 Fosfatase Ácida 2014
Tratamentos
T1 T2 T3 T4 T5 T6 Pousio Mata
0
200
400
600
800
1000 Fosfatase Ácida 2015
a
Tratamentos
T1 T2 T3 T4 T5 T6 Pousio Mata
ug p
-nitr
ofen
ol h
-1 g
sol
o se
co-1
0
20
40
60
80 B-glicosidase 2015
Tratamentos
T1 T2 T3 T4 T5 T6 Pousio Mata
0
10
20
30
40
50
60
70 B-glicosidase 2014
ug p
-nitr
ofen
ol h
-1 g
sol
o se
co-1
ug p
-nitr
ofen
ol h
-1 g
sol
o se
co-1
ug p
-nitr
ofen
ol h
-1 g
sol
o se
co-1
ab ab
ab a
b
ab ab
abc
a
abc
bc
c
a
ab b b b b
ab
b
a a a a
A B
C D
Fig.2. Média da atividade enzimática em experimento com aplicação de biocarvão em latossolo vermelho-amarelo, cultivado com Teca no Bioma Amazônia. Tratamentos: T1- Muda sem biocarvão; T2- Muda com biocarvão; T3- Muda sem biocarvão + biocarvão ativado aplicado no solo; T4- Muda com biocarvão + biocarvão ativado aplicado no solo; T5- Muda sem biocarvão + biocarvão não ativado aplicado no solo; T6- Muda com biocarvão + biocarvão não ativado aplicado no solo e duas áreas de referência. Médias seguidas da mesma letra indicam que não houve diferença estatística pelo teste de Tukey a 5% de probabilidade.

49
Não houve diferença estatística para o pH do solo com 3 e 15 meses de
experimento (Anexo B). A média dos tratamentos para o pH do solo no primeiro
ano foi de 4,71 e no segundo de 4,64. O pH das áreas de referencia (Pousio e
Mata Nativa) observados foram 3,82 e 3,18, respectivamente. Quanto à
Umidade Gravimétrica (UG%) do solo, esta também não apresentou diferença
significativa, sendo a média dos tratamentos de 2014 de 16,65% e de 2015 de
24,92%, refletindo a maior precipitação acumulada no mês de abril neste ano,
período em que foram coletadas as amostras de solo.
4.2. Diversidade bacteriana (Índice de Shannon)
O uso do biocarvão ativado aplicado no solo proporcionou a maior
diversidade bacteriana (2,72) em experimento com 3 meses de aplicação do
biocarvão, no ano de 2014, sendo o valor mais próximo da Mata Nativa (2,79) e
foram superiores aos tratamentos com biocarvão não ativado (T5 e T6), que
apresentaram os menores valores de diversidade bacteriana (2,31 e 2,21,
respectivamente) (Tabela 1). Os tratamentos T5 e T6, caracterizados pela
aplicação do biocarvão não ativado aplicado no solo apresentaram os menores
valores de diversidade bacteriana (2,31 e 2,21 respectivamente), valores estes,
próximos ao observado na área de Pousio.
Tabela 1 Índice de Shannon (H’) dos perfis de DGGE de comunidades bacterianas totais do solo, cultivado com Teca no Bioma Amazônia.
Tratamento Índice de Shannon (H’) 2014
Índice de Shannon (H’) 2015
T1 2,5734ab 2,0990b
T2 2,4536bc 2,3852a
T3 2,7121a 2,4554a
T4 2,3985bc 2,4839a
T5 2,3103c 2,5080a
T6 2,2171c 2,5859a
Pousio 2,0422 2,2117
Mata Nativa 2,7957 2,5902
CV (%) 4,68 4,60
T1- Muda sem biocarvão; T2- Muda com biocarvão; T3- Muda sem biocarvão + biocarvão ativado aplicado no solo; T4- Muda com biocarvão + biocarvão ativado aplicado no solo; T5- Muda sem biocarvão + biocarvão não ativado aplicado no solo; T6- Muda com biocarvão +
/////
///

50 biocarvão não ativado aplicado no solo e duas áreas de referência. Médias seguidas da mesma letra indicam que não houve diferença estatística pelo teste de Tukey a 5% de probabilidade.
No ano de 2015 em experimento com 15 meses de aplicação do
biocarvão, a maior diversidade também foi encontrada na área de Mata Nativa
(2,59). Entre os tratamentos, somente o T1 (sem aplicação do biocarvão)
diferiu estatisticamente dos demais. Os valores de diversidade entre os
tratamentos T2 ao T6 foram bastante semelhantes e muito próximos da área de
referencia de Mata Nativa, sendo novamente superiores ao Pousio, que junto
com o tratamento controle, apresentaram a menor diversidade bacteriana do
solo no ano de 2015.
4.3. Estrutura da comunidade de bactérias totais (gene 16S rDNA)
Os géis obtidos a partir de amostras de solo coletadas em experimento com
três e quinze meses de aplicação do biocarvão permitiram a visualização de
diferentes perfis das comunidades bacterianas (ANEXO C). A estrutura da
comunidade bacteriana total do solo foi caracterizada através da análise do
perfil de bandas pela amplificação dos genes 16S rDNA com a reação da
polimerase em cadeia combinada com DGGE.
A técnica de PCR-DGGE possibilitou identificar diferenças na
composição das comunidades bacterianas do solo no ano de 2014 em função
dos tratamentos, em que, o uso do solo (implantação de uma área manejada
com Teca) distinguiu-se da área de Pousio e de Mata Nativa (Figura 3). A
ordenação gerada por meio da PCA (Analise de Componentes Principais)
separou os tratamentos em três grupos distintos e praticamente equidistantes,
sugerindo que a comunidade bacteriana total do solo se estruturou de três
maneiras diferentes e não há maior ou menor relação de similaridade entre
duas comunidades em relação á outra. A relativa proximidade entre as
repetições mostra um padrão de estrutura dentro de cada tratamento.
O eixo 2 explicou 12.4 % da variância e conseguiu discriminar os três
grupos, com maior força a área manejada com Teca (com e sem biocarvão),
enquanto o eixo 1 explicou 20.8% da variância e agrupou a área manejada com
a Mata Nativa e as colocou distante da área de Pousio.

51
Fig.3. Analise de Componentes Principais (PCA) do perfil estrutural das comunidades bacterianas do solo por PCR-DGGE em experimento com três meses de aplicação do biocarvão (2014). T1- Muda sem biocarvão; T2- Muda com biocarvão; T3- Muda sem biocarvão + biocarvão ativado aplicado no solo; T4- Muda com biocarvão + biocarvão ativado aplicado no solo; T5- Muda sem biocarvão + biocarvão não ativado aplicado no solo; T6- Muda com biocarvão + biocarvão não ativado aplicado no solo.
A análise da estrutura da comunidade bacteriana total do solo no ano de
2015 (experimento com quinze meses de implantação) revelou padrão de
resposta semelhante á análise do ano anterior (Figura 4). Contudo, a
comunidade de bactérias dentre os tratamentos cultivados com Teca,
apresentaram uma variação maior na estrutura da comunidade quanto à
aplicação ou não do biocarvão e em relação a sua forma ativada no solo.
O eixo 1 explicou 18.1 % da variância e discrimina de forma mais tênue
os tratamentos T1 (controle) e T3 (aplicação de biocarvão ativado no solo)
sem, contudo, distingui-las dos demais tratamentos da área cultivada com
Teca. Já, o eixo 2 explicou 11.2% da variância e discrimina de forma marcante
a área de Mata Nativa das demais áreas estudadas.
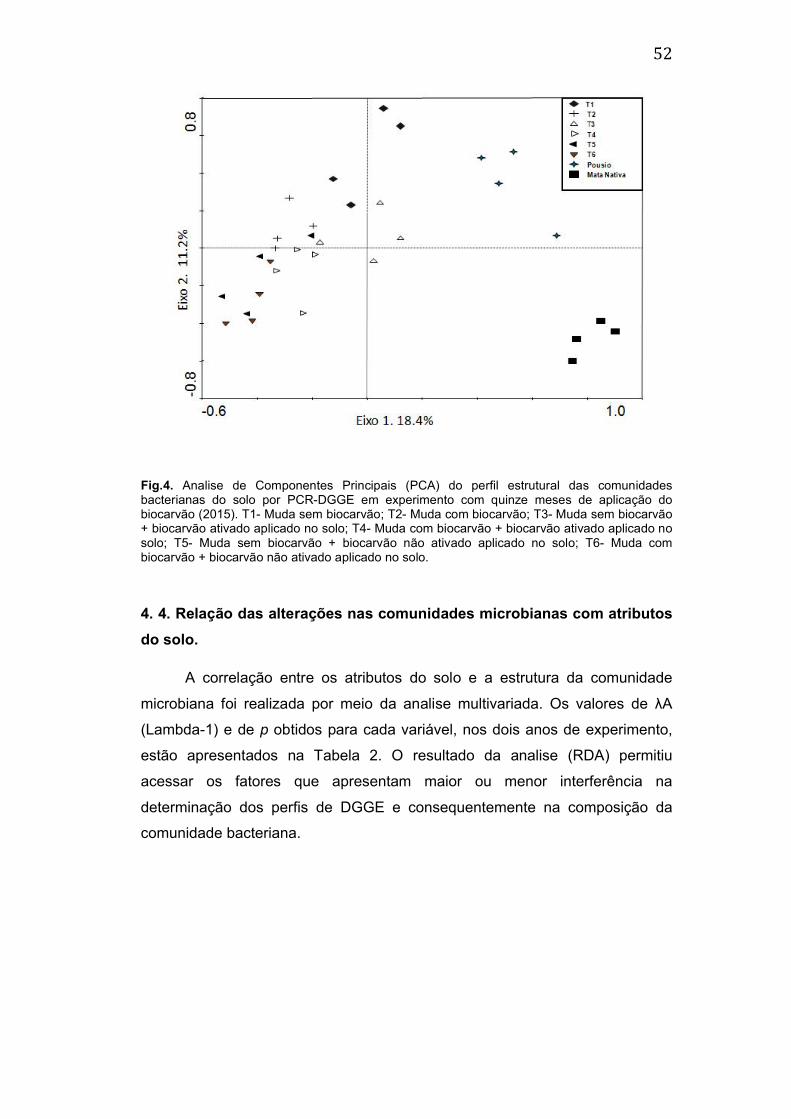
52
Fig.4. Analise de Componentes Principais (PCA) do perfil estrutural das comunidades bacterianas do solo por PCR-DGGE em experimento com quinze meses de aplicação do biocarvão (2015). T1- Muda sem biocarvão; T2- Muda com biocarvão; T3- Muda sem biocarvão + biocarvão ativado aplicado no solo; T4- Muda com biocarvão + biocarvão ativado aplicado no solo; T5- Muda sem biocarvão + biocarvão não ativado aplicado no solo; T6- Muda com biocarvão + biocarvão não ativado aplicado no solo.
4. 4. Relação das alterações nas comunidades microbianas com atributos
do solo.
A correlação entre os atributos do solo e a estrutura da comunidade
microbiana foi realizada por meio da analise multivariada. Os valores de λA
(Lambda-1) e de p obtidos para cada variável, nos dois anos de experimento,
estão apresentados na Tabela 2. O resultado da analise (RDA) permitiu
acessar os fatores que apresentam maior ou menor interferência na
determinação dos perfis de DGGE e consequentemente na composição da
comunidade bacteriana.
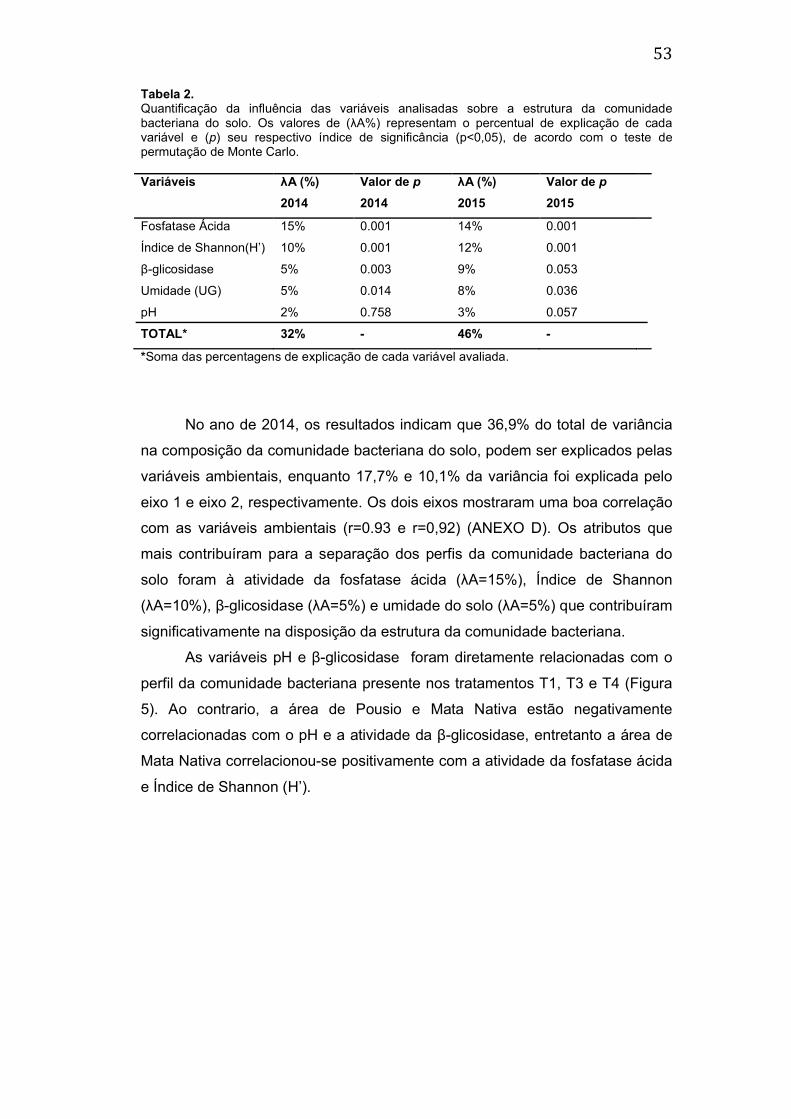
53 Tabela 2. Quantificação da influência das variáveis analisadas sobre a estrutura da comunidade bacteriana do solo. Os valores de (λA%) representam o percentual de explicação de cada variável e (p) seu respectivo índice de significância (p<0,05), de acordo com o teste de permutação de Monte Carlo.
Variáveis λA (%)
2014
Valor de p
2014
λA (%)
2015
Valor de p
2015
Fosfatase Ácida 15% 0.001 14% 0.001
Índice de Shannon(H’) 10% 0.001 12% 0.001
β-glicosidase 5% 0.003 9% 0.053
Umidade (UG) 5% 0.014 8% 0.036
pH 2% 0.758 3% 0.057
TOTAL* 32% - 46% -
*Soma das percentagens de explicação de cada variável avaliada.
No ano de 2014, os resultados indicam que 36,9% do total de variância
na composição da comunidade bacteriana do solo, podem ser explicados pelas
variáveis ambientais, enquanto 17,7% e 10,1% da variância foi explicada pelo
eixo 1 e eixo 2, respectivamente. Os dois eixos mostraram uma boa correlação
com as variáveis ambientais (r=0.93 e r=0,92) (ANEXO D). Os atributos que
mais contribuíram para a separação dos perfis da comunidade bacteriana do
solo foram à atividade da fosfatase ácida (λA=15%), Índice de Shannon
(λA=10%), β-glicosidase (λA=5%) e umidade do solo (λA=5%) que contribuíram
significativamente na disposição da estrutura da comunidade bacteriana.
As variáveis pH e β-glicosidase foram diretamente relacionadas com o
perfil da comunidade bacteriana presente nos tratamentos T1, T3 e T4 (Figura
5). Ao contrario, a área de Pousio e Mata Nativa estão negativamente
correlacionadas com o pH e a atividade da β-glicosidase, entretanto a área de
Mata Nativa correlacionou-se positivamente com a atividade da fosfatase ácida
e Índice de Shannon (H’).
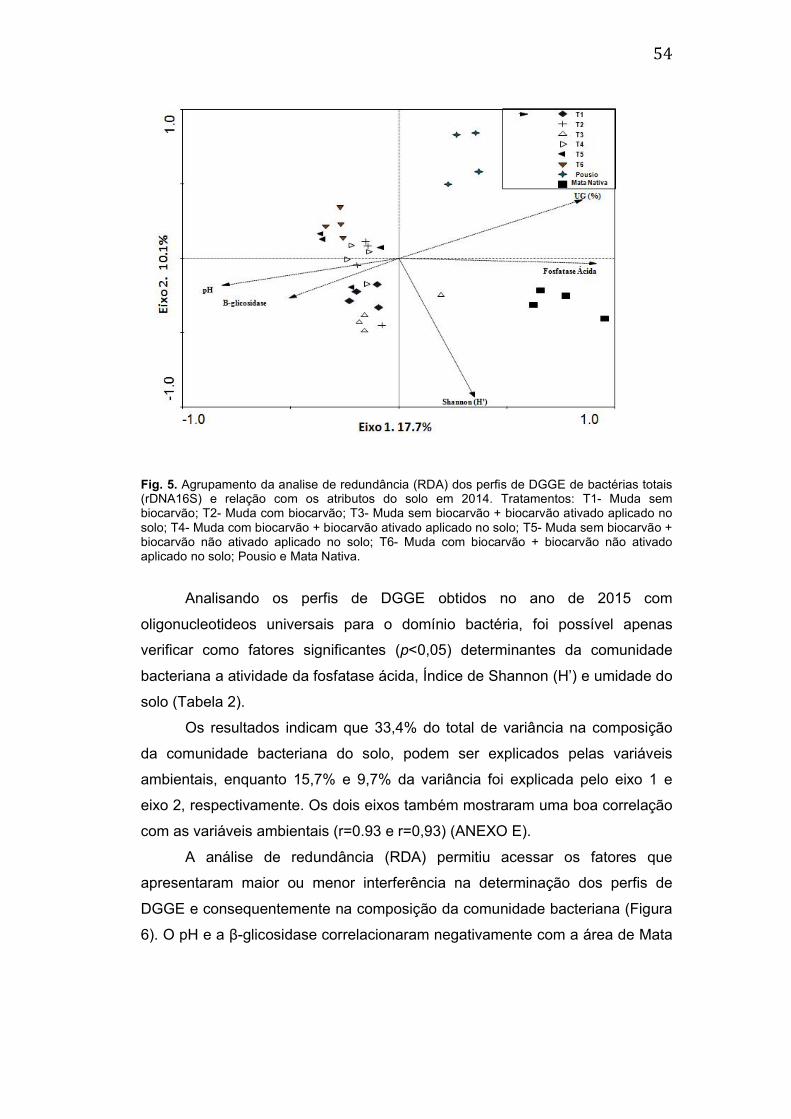
54
Fig. 5. Agrupamento da analise de redundância (RDA) dos perfis de DGGE de bactérias totais (rDNA16S) e relação com os atributos do solo em 2014. Tratamentos: T1- Muda sem biocarvão; T2- Muda com biocarvão; T3- Muda sem biocarvão + biocarvão ativado aplicado no solo; T4- Muda com biocarvão + biocarvão ativado aplicado no solo; T5- Muda sem biocarvão + biocarvão não ativado aplicado no solo; T6- Muda com biocarvão + biocarvão não ativado aplicado no solo; Pousio e Mata Nativa.
Analisando os perfis de DGGE obtidos no ano de 2015 com
oligonucleotideos universais para o domínio bactéria, foi possível apenas
verificar como fatores significantes (p<0,05) determinantes da comunidade
bacteriana a atividade da fosfatase ácida, Índice de Shannon (H’) e umidade do
solo (Tabela 2).
Os resultados indicam que 33,4% do total de variância na composição
da comunidade bacteriana do solo, podem ser explicados pelas variáveis
ambientais, enquanto 15,7% e 9,7% da variância foi explicada pelo eixo 1 e
eixo 2, respectivamente. Os dois eixos também mostraram uma boa correlação
com as variáveis ambientais (r=0.93 e r=0,93) (ANEXO E).
A análise de redundância (RDA) permitiu acessar os fatores que
apresentaram maior ou menor interferência na determinação dos perfis de
DGGE e consequentemente na composição da comunidade bacteriana (Figura
6). O pH e a β-glicosidase correlacionaram negativamente com a área de Mata
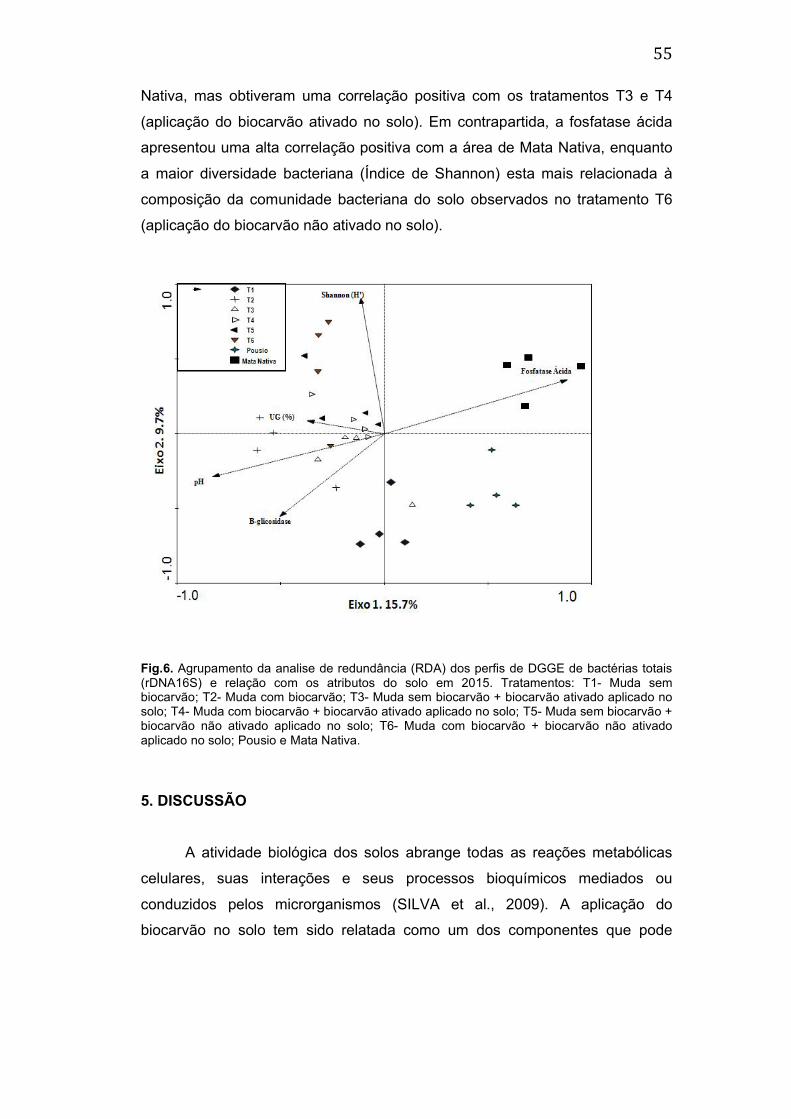
55 Nativa, mas obtiveram uma correlação positiva com os tratamentos T3 e T4
(aplicação do biocarvão ativado no solo). Em contrapartida, a fosfatase ácida
apresentou uma alta correlação positiva com a área de Mata Nativa, enquanto
a maior diversidade bacteriana (Índice de Shannon) esta mais relacionada à
composição da comunidade bacteriana do solo observados no tratamento T6
(aplicação do biocarvão não ativado no solo).
Fig.6. Agrupamento da analise de redundância (RDA) dos perfis de DGGE de bactérias totais (rDNA16S) e relação com os atributos do solo em 2015. Tratamentos: T1- Muda sem biocarvão; T2- Muda com biocarvão; T3- Muda sem biocarvão + biocarvão ativado aplicado no solo; T4- Muda com biocarvão + biocarvão ativado aplicado no solo; T5- Muda sem biocarvão + biocarvão não ativado aplicado no solo; T6- Muda com biocarvão + biocarvão não ativado aplicado no solo; Pousio e Mata Nativa.
5. DISCUSSÃO
A atividade biológica dos solos abrange todas as reações metabólicas
celulares, suas interações e seus processos bioquímicos mediados ou
conduzidos pelos microrganismos (SILVA et al., 2009). A aplicação do
biocarvão no solo tem sido relatada como um dos componentes que pode

56 modular os atributos químicos e físicos do solo (LEHMANN et al., 2011), em
que, a atividade enzimática e a diversidade microbiana do solo são importantes
e sensíveis indicadores microbiológicos de qualidade de solos (BASTIDA et al.,
2008; LU et al., 2015). Nesse sentido, foram avaliados no presente estudo o
efeito do biocarvão na atividade enzimática e na biodiversidade bacteriana do
solo cultivado com Teca.
Devido à capacidade de adsorver compostos orgânicos e inorgânicos,
alguns autores tem demonstrado que o biocarvão pode inibir a atividade de
algumas enzimas no solo ou os seus substratos que ficam adsorvidos nos
sítios de reação (JIN, 2010; BAILEY et al., 2011; LEHMANN et al., 2011). Para
os estudos de curto prazo, no entanto, é ainda necessário determinar se as
propriedades físicas do biocarvão (por exemplo, porosidade e grande área de
superfície) vão beneficiar a atividade dos microrganismos do solo (Warnock et
al, 2007) ou não (QUILLIAM et al., 2013), podendo contribuir também com a
capacidade de adsorção melhorada de compostos polares e apolares (ZHAO et
al., 2015a).
Entretanto, no presente estudo, foi verificado que a atividade enzimática
da β-glicosidade e fosfatase ácida é afetada positivamente por meio da
aplicação de biocarvão ativado, um importante condicionador de solo. O
biocarvão ativado apresenta maior área de superfície, volume e diâmetro dos
poros criados durante a pirolise (HONGJIE, 2014).
O tamanho dos poros favorece o movimento de raízes através do solo, o
que pode ser um dos fatores responsáveis pela maior produção de enzimas na
presença do biocarvão ativado (JAAFAR et al., 2015). Esses mesmos autores,
verificaram a presença de pequenas partículas de solo no interior desses poros
e concluíram que o aumento da porosidade do biocarvão fornece um habitat
seguro, água e nutrientes para a reprodução dos microrganismos, favorecendo
a atividade microbiana no solo (JOSEPH et al., 2010).
Além das características físicas do biocarvão, algumas propriedades
químicas do solo correlacionam-se com a atividade da β-glicosidade. O grande
volume de material orgânico em decomposição no solo (30,4 g dm-3 teor de
matéria orgânica no solo) podem ter contribuído também para atividade desta
enzima, uma vez que, a matéria orgânica fornece substrato para sua ação e
também protege as enzimas do solo em suas formas ativas (SOUZA E

57 LOBATO, 2004). Essa proteção ocorre devido à formação de complexos
enzimas-compostos húmicos (DENG E TABATABAI, 1997), deixando a β-
glicosidade indisponível para a degradação e desnaturação pelas enzimas
proteolíticas naturalmente presentes no solo das áreas cultivadas.
β-glicosidade e celobio-hidrolase são as principais controladoras da
dinâmica do carbono no solo. Estas enzimas refletem fortemente a
mineralização do C e são frequentemente utilizadas para indicar a taxa de
incorporação de carbono lábil no solo (SINSABAUGH et al., 2008). Liao et al.
(2016) observaram a maior atividade destas enzimas a medida que aumentava
a dose de biocarvão aplicado no solo, em comparação com o tratamento
controle (0 t ha-1). Os autores concluíram que a adição do biocarvão no solo
promove a mineralização do carbono, proporcionando substratos para estas
enzimas, além de que, os compostos voláteis liberados durante a produção do
biocarvão em baixas temperaturas de pirólise (350ºC a 500ºC) podem estimular
a atividade da β-glicosidade.
Wu et al. (2013) observaram que a aplicação do biocarvão não
proporcionou efeito sobre a atividade da β-glicosidade. Os autores sugeriram
que as formas de C do biocarvão produzidos a partir da palha de trigo eram
muito recalcitrantes e não estimularam a atividade enzimática do solo.
Os maiores valores de atividade enzimática observados em solos sob
Mata Nativa ocorrem devido ao maior aporte de resíduos nestes locais e
também pela diversidade deste material vegetal (LISBOA et al., 2012).
Resultados esses não observados na nossa pesquisa, em que a área de Mata
Nativa apresentou a menor atividade enzimática da β-glicosidade nos dois anos
de experimento. Nestes casos, outros fatores também deve estar influenciando,
como se verificou em nossa pesquisa em que a β-glicosidade correlacionou-se
positivamente com o pH nos dois anos de experimento.
Os solos da Mata Nativa apresentam um menor valor de pH quando
comparados com os valores da área cultivada, além disso, a composição dos
resíduos adicionado ao solo podem influenciar na atividade enzimática. Os
resíduos mais lignificados tendem a ser mais difíceis de decompor, reduzindo
assim a quantidade de resíduos carbonados, que são substrato da β-
glicosidade (BAILEY et al., 2011; PAZ FERREIRO et al., 2014).

58
Matsuoka et al. (2003), relataram que a atividade da β-glicosidase foi
maior no sistema de cultivo perene de uva do que na mata e na área de soja
convencional. Os autores atribuem esse fato aos restos vegetais de folhas,
frutos, galhos e flores na entrelinha da videira, pois a β-glicosidase atua na
etapa final da decomposição da celulose, sendo responsável pela hidrólise dos
resíduos de celobiose e formando o açúcar simples β-D-glucose.
Assim como a matéria orgânica, as argilas do solo também podem
estabilizar as enzimas e manter sua atividade por longos períodos de tempo
(ALLISON, 2006). Segundo Turner et al. (2002), esse mecanismo pode ser
direto, ou indireto, seja adsorvendo em sua superfície as próprias enzimas, ou
realizando uma proteção física a matéria orgânica, nutrientes e microrganismos
do solo.
Trazi (2014) relata que o biocarvão melhora as propriedades químicas
do solo em campo, após 30 meses de sua aplicação, principalmente pela
redução da acidez do solo e pelo aumento dos valores de saturação por bases,
K e P.
Em relação à atividade da fosfatase ácida, nos primeiros três meses, a
adubação com fósforo pode ter influenciado a baixa atividade desta enzima,
provavelmente devido a resíduos de fósforo disponíveis no solo, resultantes da
aplicação de fertilizante fosfatado (120 g de P2O5 por cova). Quanto maior a
disponibilidade de P solúvel no solo, menor é a atividade da enzima fosfatase
ácida (GATIBONE et al., 2008), uma vez que ela é liberada pelas plantas e
microrganismos quando há baixa disponibilidade de P solúvel no solo.
A fosfatase ácida pode estar presente nos exsudados liberados pelas
raízes das plantas, ao passo que, a fosfatase alcalina origina-se dos fungos e
bactérias do solo. Nos estudos de Jin et al. (2016), eles verificaram maior
atividade da fosfatase ácida nas amostras controles (sem aplicação do
biocarvão) em relação a fosfatase alcalina, em que a atividade da fosfatase
ácida diminuiu com adição do biocarvão. Os autores concluíram que esta
variação na atividade da fosfatase ácida e alcalina podem estar atribuídas a
aplicação do biocarvão, ao pH do solo e a disponibilidade de P.
De modo geral, as áreas sob vegetação nativa apresentam uma maior
atividade da fosfatase ácida, e as áreas cultivadas, os menores valores,
corroborando com os resultados obtidos por Evangelista et al. (2012) que

59 observaram a maior atividade da fosfatase ácida em área nativa de Cerrado no
período chuvoso e de estiagem. Outra razão para a maior atividade da
fosfatase ácida em áreas de vegetação nativa é a presença de grande
quantidade de carbono e fósforo orgânico contido na biomassa microbiana do
solo e baixa disponibilidade de fósforo inorgânico (GATIBONI et al., 2008).
Elzobair et al. (2015) observaram um efeito neutro do biocarvão no
potencial enzimático mas, observaram que a atividade da fosfatase ácida foi
maior após três anos de aplicação do biocarvão no solo. Entretanto, Carneiro et
al. (2008) verificaram que a fosfatase ácida não foi capaz de diferenciar os
manejos e usos do solo estudado, não encontrando diferenças significativas
entre si e também não encontraram correlação dela com a fertilidade de um
latossolo vermelho.
Segundo Dick et al. (1988), a matéria orgânica é outro fator que pode
influenciar na atividade desta enzima, uma vez que apresenta efeitos benéficos
sobre a comunidade microbiana, além de proteger fisicamente e manter as
enzimas na sua forma ativa no solo. Bailey et al. (2011) relataram que a
aplicação do biocarvão no solo geralmente aumenta a atividade enzimática
relacionados ao ciclo do N e P e reduz a atividade enzimática envolvidos no
ciclo do C.
A análise redundância revelou uma alta correlação da β-glicosidade com
o pH do solo, enquanto para a fosfatase ácida a correlação foi negativa, sendo
mais influenciada pelo uso do solo. Deng e Tabatabai (1997) afirmaram que a
maior atividade dessa enzima se expressa com pH do solo em torno de 5,0 e
que a atividade da fosfatase ácida tende a diminuir com o aumento do pH. As
alterações na atividade enzimática dos solos pode ser uma resposta das
alterações químicas após aplicação do biocarvão (ABUJABHA et al., 2015).
Enquanto outros estudos tem relatado um aumento no pH do solo após
a aplicação do biocarvão (KIMETU et., 2008; RUTIGLIANO et al., 2014;
AMELOOT et al., 2014; XU et al., 2014), nossos resultados mostraram que não
houve diferença no pH com aplicação do biocarvão em relação ao controle,
mas verificamos uma diminuição do pH após quinze meses da aplicação no
solo. O efeito do biocarvão no pH do solo depende do valor do pH do próprio
biocarvão, das condições de pirólise e do material usado para a sua produção
(LEHMANN et al., 2011). Um aumento da matéria orgânica após a adição do

60 biocarvão também pode diminuir o pH do solo, devido á atividade microbiana e
ácidos orgânicos liberados durante a decomposição (ABUJABHA et al., 2015).
Em nosso estudo, após 3 meses de aplicação do biocarvão no solo,
verificamos que o uso do solo foi o responsável pela mudança na estrutura da
comunidade bacteriana, onde houve a separação bem distinta de 3 grupos
(solo cultivado com biocarvão, solo de Mata Nativa e uma área não manejada
de solo com plantas espontâneas), em que a microbiota do solo foi influenciada
pelo uso do solo. As comunidades vegetais podem modificar as condições dos
micro-ambientes ocupados pelos microrganismos, devido ao aporte de material
vegetal e exsudados liberados, alterando assim as diversas populações na
comunidade microbiana do solo (NDAW et al., 2009), refletindo na maior
diversidade bacteriana observada nesse trabalho.
Evidenciou-se uma alteração na estrutura da comunidade bacteriana no
solo cultivado com teca influenciado pela adição do biocarvão no solo após
quinze meses. As alterações nas propriedades físico-químicas do solo após a
aplicação do biocarvão, tais como pH, teor de água, atividade enzimática,
diversidade microbiana e principalmente teor de carbono orgânico podem ser a
principal razão para alterações nas populações bacterianas do solo (LEHMANN
et al., 2011; KELLY et al., 2014; ABUJABHA et al., 2015).
A diversidade bacteriana, em nossos estudos, representada pelo Índice
de Shannon (H’) foi maior na Mata Nativa e no tratamento com aplicação do
biocarvão ativado. Nos estudos realizados por Liao et al. (2016), trabalhando
com aplicação de três doses de biocarvão (0 t ha-1; 2,5 t ha-1 e 4,5 t ha-1)
observaram aumento na diversidade bacteriana do solo representada pelo
Índice de Shannon (H’) a medida que incrementava as doses de biocarvão no
solo. Em nossa pesquisa, após quinze meses, a diversidade da Mata Nativa
ainda foi maior, e os tratamentos com o biocarvão apresentaram um valor de
Índice de Shannon (H’) semelhantes, em que as superfícies e poros do
biocarvão fornecem um habitat para os microrganismos, além de melhorar a
densidade, pH e a circulação de ar, água e nutrientes no interior da matriz do
solo (GUL et al., 2015).
Há uma ampla evidencia na literatura que a adição do biocarvão melhora
as propriedades físico-químicas do solo, refletindo na atividade dos
microrganismos do solo. A influência que o biocarvão tem nas comunidades

61 microbianas é fortemente afetada pelo tempo em que ele é adicionado e pelas
características do solo (THIES et al., 2015). As reações que ocorrem quando o
biocarvão recém-preparado é adicionado ao solo, é bastante diferente
daquelas que ocorrem depois que as superfícies do biocarvão foram afetadas
por processos bióticos e abióticos (ZHAO et al., 2015a ; ZHAO et al., 2015b).
Joseph et al. (2010) atribuem alguns fatores que podem influenciar as
características do biocarvão e suas reações no solo, como, a composição e
disponibilidade de nutrientes, matéria orgânica, textura do solo, condições de
pH, biota do solo, tipo de material vegetal, a proximidade do biocarvão da
rizosfera e as variações temporais da umidade do solo. Todos os fatores, em
conjunto, irão modular as alterações na atividade, abundancia e diversidade de
comunidades microbianas no solo.
6. CONCLUSÃO
A atividade da β-glicosidade é favorecida pela aplicação do biocarvão
ativado no solo. Contudo, a fosfatase ácida é mais influenciada pelo pH e uso
do solo, em relação a aplicação do biocarvão.
A área de Mata Nativa, apresentou a maior atividade da fosfatase ácida
e diversidade bacteriana e a menor atividade da β-glicosidade, que pode estar
relacionada ao pH mais ácido e a composição da matéria orgânica nestas
áreas.
As mudanças na estrutura das comunidades podem estar associadas
com o aumento da macroporosidade do biocarvão ativado, proporcionando um
nicho seguro para os microrganismos do solo, embora seja desconhecido em
que medida os efeitos benéficos do biocarvão mantém-se constante ao longo
do tempo.
A aplicação do biocarvão e o seu uso como condicionador do solo,
apresentam resultados variáveis nas características do solo, influenciando na
diversidade da comunidade bacteriana pelo tempo em que ele é adicionado.

62 7. AGRADECIMENTOS
Os autores agradecem à Fundação de Amparo à Pesquisa do Estado de
Mato Grosso (FAPEMAT), à Coordenação de Aperfeiçoamento de Pessoal de
Nível Superior (CAPES), a pesquisadora Dr. Fabiana de Abreu Rezende pelo
apoio e incentivo à pesquisa, a Universidade Federal de Mato Grosso e ao
programa de pós-graduação em Agronomia da UFMT pelo apoio no
desenvolvimento deste trabalho e a Embrapa Agrossilvipastoril pelo incentivo e
amparo na execução desta pesquisa.
8. REFERÊNCIAS BIBLIOGRÁFICAS
ABRAF – Associação Brasileira de Produtores de Florestas Plantadas. Anuário ABRAF – Associação Brasileira de Produtores de Florestas Plantadas. Anuário estatístico da ABRAF 2013 ano base 2012. Brasília. 142 p., 2013.
ABUJABHAH, I.; BOUND, S.A.; DOYLE, R.; BOWMAN, J.P. Effects of biochar and compost amendments on soil physico-chemical properties and the total community within a temperate agricultural soil. Applied Soil Ecology, 2015. Hppt://dx.doi.org/10.1016/j.apsoil.2015.10.021
ALLISON, S.D. Soil minerals and humic acids alter enzyme stability: implications for ecosystem processes. Biogeochemistry, v.81, p.361-373, 2006.
ALVAREZ, C.A.; STAPE, J.L.; SENTELHAS, P.C.; GONÇALVES, J.L.M.; SPAROVEK, G. Köppen’s climate classification map for Brazil. Gebrüder Bomtraeger, 2013. Doi: 10.1127/0941-2948/2013/0507.
AMELOOT, N.; SLEUTEL, S.; CASE, S.D.C.; ALBERTI, G.; MCNAMARRA, N.P.; ZAVALLONI, C.; VERVISCH, B.; VEDOVE, G.D.; DE NEVE, S. C mineralization and microbial activity in four biochar field experiments several years after incorporation. Soil Biology & Biochemistry, v. 78, p. 195-203, 2014.
ANDREOTE, F.D.; AZEVEDO, J.L.; ARAUJO, W. L. Assessing the diversity of bacterial communities associated with plants. Brazilian Journal of Microbiology, v.40, p.417-432, 2009.
ARAUJO, S.D.F.; MONTEIRO, R.T.R. Indicadores biológicos de qualidade do solo. Bioscience Journal, v.23, p.66-75, 2007.

63 ATKINSON, C. J.; FITZGERALD, J. D.; HIPPS, N. A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: a review. Plant Soil, v. 337, p. 1–18, 2010.
BAILEY, V.L., FANSLER, S.J., SMITH, J.L., BOLTON, H. Reconciling apparent variability in effects of biochar amendment on soil enzyme activities by assay optimization. Soil Biology & Biochemistry, v.43, p.296–301, 2011.
BASTIDA, F.; ZSOLNAY.; HERNÁNDEZ, T.; GARCÍA, C. Past, presente and future of soil quality índices: A biological perspective. Geoderma, v.147, p.159-171, 2008.
CARNEIRO, A.A.C.; ASSIS, P.C.R.; MELO, L.B.C.; PEREIRA, H.S.; PAULINO, H.B.; NETO, A.N.S. Atributos bioquímicos em dois solos de cerrado sob diferentes sistemas de manejo e uso. Pesquisa Agropecuária Tropical, v.38, p. 276-283, 2008.
CHELIUS, M. K.; TRIPLETT, E. W. The diversity of archaea and bacteria in association with the roots of Zea mays L. Microbial Ecology, v.41, p.252-263, 2001.
DENG, S.P.; TABATABAI, M.A. Effect of tillage and residue management on enzyme activities in soils: II. Phosphatases and arysulfatase. Biology and Fertility Soils, v.24, p. 141-146, 1997.
DICK, R.P.; RASMUNSSEN, P.E.; KERLE, E.A. Influence of long-term residue management on soil enzyme activities in relation to soil chemical properties of a wheat-fallow system. Biology and Fertility of Soils, v.24, p. 159-164, 1988.
DIEL, D.; BEHLING, M.; NETO, A.L.F. Distribuição horizontal e vertical de fósforo em sistemas de cultivos exclusivos de soja e de integração lavoura-pecuária-floresta. Pesquisa Agropecuária Brasileira, v.49, p.639-647, 2014.
ELZOBAIR, K.A.; STROMBERGER, M.E.; IPPOLITO, J. Stabilizing effect of biochar on soil extracellular enzymes aftera denaturing stress. Chemosphere, 2015. http://dx.doi. org/10.1016/j.chemosphere.
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Centro Nacional de Pesquisa de Solos. Manual de métodos de análises de solo. 2. ed. Rio de Janeiro: Embrapa Solos, 1997. 212p.
EVANGELISTA, C.R.; PARTELLI, F.L.; FERREIRA, E.P.B.; CORRECHEL, V. Atividade enzimática do solo sob sistema de produção orgânica e convencional na cultura da cana-de-açúcar em Goiás. Ciências Agrárias, v.33, p.1251-1262, 2012.

64 GATIBONI, L. C.; KAMINSKI, J.; RHEINHEIMER, D. S.; BRUNETTO, G. Fósforo da biomassa microbiana e atividade de fosfatases ácidas durante a diminuição do fósforo disponível no solo. Pesquisa Agropecuária Brasileira, Brasília, v. 43, n. 8, p. 1085-1091, 2008.
GUL, S.; WHALEN, J.K.; THOMAS, B.W.; SACHDEVA, V.; DENG, H. Phyco-chemical properties and microbial responses in biochar -amended soils: Mechanisms and future directions. Agriculture, Ecosystems and Enviroment, v. 206, p.46-59, 2015.
HAIDAR, R.F.; FAGG, J.M.F.; PINTO, J.R.R. Florestas estacionais e áreas de ecótono no estado do Tocantins, Brasil: parâmetros estruturais, classificação das fitofisionomias florestais e subsídios para conservação. Acta Amazonica, v.43, p.261-290, 2013.
HONGJIE, Z. 2014. Biocarvão Effects on Soil Microbial Biomass and Activity. Thesis, The University of Guelph, Ontario, Canadá.
JAAFAR, N.M.; CLODE, P.L.; ABBOTT, L.K. Soil microbial responses to biochar varying in particle size, surface and pore properties. Pedosphere, v.25, p.770-780, 2015.
JIN, H., 2010. Characterization of Microbial Life Colorizing biochar and amended Soils. PhD Dissertation, Cornell University, Ithaca, NY.
JIN, Y.; LIANG, X.; HE, M.; LIU, Y.; TIAN, G.; SHI, J. Manure biochar influence upon soil properties, phosphorus distribuition and phosphatase activities: A microcosm incubation study. Chemosphere, v.142, p.128-135, 2016.
JOSEPH, S.; CAMPS-ARBESTAIN, M.; LIN, Y.; MUNROE, P.; CHIA, C. H.; HOOK, J.; VAN ZWIETEN, L.; KIMBER, S.; COWIE, A.; SINGH, B. P.; LEHMANN, J.; FOIDL, N.; SMERNIK, R. J.; AMONETTE, J. E. An investigation into the reactions of biocarvão in soil. Australian Journal of Soil Research, v. 48, p. 501–515, 2010.
KELLY, C.N.; PELTZ, C.D.; STANTON, M.; RUTHERFORD, D.W.; ROSTAD, C.E. Biochar application to hardrock mine tailings: soil quality, microbial activity, and toxic element sorption. Applied Geochemistry, v.43, p.35–48, 2014.
KIMETU, J. M.; LEHMANN, J.; NGOZE, S. O.; MUGENDI, D. N.; KINYANGI, J. M.; RIHA, S.; VERCHOT, L.; RECHA, J. W.; PELL, A. N. Reversibility of soil productivity decline with organic matter of differing quality along a degradation gradient. Ecosystems, v.11, p.726–739, 2008.
LEHMANN, J.; JOSEPH, S. Biochar for Enviromental Management: An Introduction. In: LEHMANN, J.; JOSEPH, S. (ed). Biochar for Enviromental

65 Management: Science and Technology. 1. Ed. earthscan, Londres, 2009. p. 4-18.
LEHMANN, J.; JOSEPH, S. Biochar for Enviromental Management: An Introduction. In: LEHMANN, J.; JOSEPH, S. (ed). Biochar for Enviromental Management: Science, Technology and Implementation. 2. Ed. earthscan, Londres, 2015. p. 327-352.
LEHMANN, J.; RILLIG, M. C.; THIES, J.; MASIELLO, C. A.; HOCKADAY, W. C.; CROWLEY, D. Biochar effects on soil biota – A review. Soil Biology & Biochemistry, v. 43, p. 1812-1836, 2011.
LEHMANN, J.A. Handful of carbon. Nature, v.447, p.143-144, 2007.
LIAO, N.; LI, Q.; ZHANG, W.; ZHOW, G.; MA, L.; MIN, W.; YE, J.; HOU, Z. Effects of biochar on soil microbial community composition and activity in drip-irrigated desert soil. European Journal os Soil Biology, v.72, p.27-34, 2016.
LISBOA, B. B.; VARGAS, L. K.; SILVEIRA, A. O.; MARTINS, A. F.; SELBACH, P. A. Indicadores microbianos de qualidade do solo em diferentes sistemas de manejo. Revista Brasileira de Ciência do Solo, v.36, p.45-55, 2012.
LU. H.; LASHARI. M.S.; LIU. X.; JI. H.; LI. L.; ZHENG. J.; KIBUE. G. W.; JOSEPH. S.; PAN. G. Changes in soil microbial community structure and enzyme activity with amendment of biochar -manure compost and pyroligneous solution in a saline soil from Central China. European Journal of Soil Biology, 2015. http://dx.doi.org/10.1016/j.ejsobi.2015.07.005
MATSUOKA, M.; MENDES, I.C.; LOUREIRO, M.F. Biomassa microbiana e atividade enzimática em solos sob vegetação nativa e sistemas agrícolas anuais e perenes na região de Primavera do Leste (MT). Revista Brasileira de Ciência do Solo, v.27, p.425-433, 2003.
MOREIRA, F. M. S.; SIQUEIRA, J. O. Microbiologia e Bioquímica do Solo. Lavras: UFLA, 2006. 729p.
NDAW, S. M.; GAMA-RODRIGUES, A. C.; GAMA-RODRIGUES, E. F.; SALES, K. R.; ROSADO, A. S. Relationships between bacterial diversity, microbial biomass, and litter quality in soils under different plant covers in northern Rio de Janeiro State, Brazil. Canadian Journal of Microbiology, 2009. Doi: 10.1139/w09-066.
OVREAS, L., FORNEY, L., DAAE, F.L., TORVISK, V. Distribution of bacterioplankton in meromictic Lake Saelenvanet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S r RNA. Applied and Environmental Microbiology, v.63, p.3367-3373, 1997.

66 PAZ-FERREIRO, J.; FU, S.; MÉNDEZ, A.; GASCO, G. Interactive effects of biochar and the earthworm Pontoscolex corethrurus on plant productivity and soil enzymes activities. Journal of Soils Sediments, v.14, p.483–494, 2014.
QUILLIAM, R.S.; GLANVILLE, H.C.; WADE, S.C.; JONES, D.L. Lie in the ‘charosphere’ — does biochar in agricultural soil provide a significant habitat for microorganisms?. Soil Biology & Biochemistry, v.65, p.287–293, 2013.
RACHID, C.T.C.C. 2010. Comunidade bacteriana, atributos do solo e fluxo de gases em solo sob Cerrado e cana-de-açúcar. Dissertação, Universidade de São Paulo- Centro de Energia Nuclear na Agricultura, Piracicaba, SP, Brasil.
RUTIGLIANO, F.A.; ROMANO, M.; MARZAOLLI, R.; BAGLIVO, L.; BARONTI, S.; MIGLIETTA, F.; CASTALDI, S. Effect of biochar addition on soil microbial community in a wheat crop. European Journal of soil Biology, v.60, p. 9-15, 2014.
SHIMIZU, J.Y.; KLEIN, H; OLIVEIRA, J.R.V. DE. Diagnóstico das plantações florestais em Mato Grosso. Cuiabá, MT: Central de Texto, 2007.
SILVA, L.G.; MENDES, I.C.; REIS JUNIOR, F.B.; FERNANDES, M.F.; MELO, J.T.; KATO, E. Atributos físicos, químicos e biológicos de um latossolo de cerrado em plantio de espécies florestais. Pesquisa Agropecuária Brasileira, v.44, p.613-620, 2009. SINSABAUGH, R.L.; LAUBER, C.L.; WEINTRAUB, M.N.; AHMED, B.; CRENSHAW, A.C.; CONTOSTA, A.R.; FREY, C.S.; GALLO, M.E.; GARTNER, T.B.; HOBBIE, S.E.; HOLLAND, K.; KEELER, J.S.; POWERSM, M.; STURSOVA, C.; TAKACSVESBACH, M.P.; WALDROP, M.D.; WALLENSTEIN, D.R.; ZACK, L.H.; ZEGLIN. Stoichiometry of soil enzyme activity at global scale, Ecology Letters, v.11, p.1252-1264, 2008. SOUZA, D.M.G.; LOBATO, E. Cerrado: Correção do solo e adubação. 2º edição, 2004.
SPERA, S.T.; SANTOS, H.P. DOS; FONTANELI, R.S.; TOMMM, G.O. Integração lavoura e pecuária e os atributos físicos de solo manejado sob sistema plantio direto. Revista Brasileira de Ciência do Solo, v. 33, p.130, 2009.
TABATABAI, M. A. Soil enzymes. In: Weaver, R. W.; Scott, A.; Bottomeley, P. J. Methods of soil analysis: microbiological and biochemical properties. Madison: Soil Science Society of America, Part 2, p.778-835, 1994.
TABATABAI, M.A.; BREMNER, J. M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biology and Biochemistry, v. 1, p. 301-307, 1969.

67 THIES, J.E.; RILLIG, M.C.; GRABER, E.R, 2015. Biochar effects on the abundance, activity and diversity of the soil biota. In: Lehmann, J., Joseph, S. (Eds.), Biochar for Environmental Management: Science, Technology and Implementation. Earthscan, London, pp. 327–389.
TRAZZI, P.A, 2014. Uso do Biocarvão na produção de mudas e no crescimento inicial de Pinus taeda L. Tese de doutorado, Universidade Federal do Paraná, Curitiba, Brasil.
TURNER, B.L.; HOPKINS, D.W.; HAYGARTH, P.M.; OSTLE, N. B-Glucosidase activity in pasture soils. Applied Soil Ecology, v.20, p.157-162, 2002.
WANG, X.; SONG, D.; LIANG, G.; ZHANG, Q.; AI, C.; ZHOU, W. Maize biochar addition rate influences soil enzyme activity and microbial community composition in a fluvo-aquic soil. Applied Soil Ecology, v.96, p.265-272, 2015.
WARNOCK, D.; LEHMANN, J.; KUYPER, T.; RILLIG, M. Mycorrhizal responses to biochar in soil — concepts and mechanisms. Plant Soil, v.300, p.9–20, 2007.
WU, F.; WANG, J. S.; CHANG, S.X.; STARTSEV, A. Contrasting effects of wheat straw and its biochar on greenhouse gas emissions and enzyme activities in a Chernozemic soil. Biology Fertily Soils, v.49, p.555-565, 2013.
XU, G.; SUN, J.; SHAO, H.; CHANG, S. Biocarvão had effects on phosphorus sorption and desorption in three soils with differing acidity. Ecological Engineering, v.62, p.54-60, 2014.
ZHAO, R.; COLES, N.; WU, J. Carbon mineralization following additions of fresh and aged biochar to an infertile soil. Catena, v.125, p.183-189, 2015a.
ZHAO, R.; COLES, N.; KONG, Z.; WU. Effects of aged and fresh biochars on soil acidity under different incubation conditions. Soil & Tillage Research, v.146, p.133-138, 2015b.
ZIECH, B.G. 2011. Determinação da capacidade produtiva e do índice de espaçamento relativo de Tectona grandis Linn f. em glória d’oeste/MT. Dissertação, Universidade Federal de Mato Grosso, Cuiaba, MT.

68 9. ANEXOS
EFEITO DA APLICAÇÃO DO BIOCARVÃO NOS ATRIBUTOS
MICROBIOLOGICOS DO SOLO, CULTIVADO COM TECA, NO BIOMA
AMAZÔNIA
Mirelly Mioranzaa, Jacqueline Jesus Nogueira da Silvab, Fabiana Abreu de Rezendec, Onã da
Silva Freddid, Anderson Ferreirac
a Programa de Pós-Graduação em Agronomia (UFMT, Sinop-MT) bPrograma de Pós-Graduação em Ciências Ambientais (UFMT, Sinop-MT) c Embrapa Agrossilvipastoril (Sinop-MT)
d Universidade Federal de Mato Grosso (UFMT, Sinop-MT)
ANEXO A
Fig. 1. Fotografia aérea da unidade experimental do cultivo de Teca (Tectona grandi f. L) - Embrapa Agrossilvipastoril, 2015.

69 ANEXO B Média do pH e Umidade Gravimétrica do solo avaliado em experimento com 3 meses (2014) e 15 meses (2015) de aplicação do biocarvão em latossolo vermelho-amarelo, cultivado com Teca no Bioma Amazônia.
Tratamento pH em água
2014
pH em água
2015
UG (%)
2014
UG (%)
2015
T1 4,53a 4,40a 16,09a 25,05a
T2 4,73a 4,83a 16,96a 24,95a
T3 4,87a 4,72a 17,41a 25,43a
T4 4,80a 4,67a 16,27a 26,64a
T5 4,91a 4,69a 16,94ª 24,56a
T6 4,39a 4,64a 16,28a 24,92a
Pousio 3,82 4,31 22,11 23,12
Mata Nativa 3,18 3,33 21,04 24,10
CV (%) 5,49 6,24 6,12 4,66
T1- Muda sem biocarvão; T2- Muda com biocarvão; T3- Muda sem biocarvão + biocarvão ativado aplicado no solo; T4- Muda com biocarvão + biocarvão ativado aplicado no solo; T5- Muda sem biocarvão + biocarvão não ativado aplicado no solo; T6- Muda com biocarvão + biocarvão não ativado aplicado no solo e duas áreas de referência. Médias seguidas da mesma letra indicam que não houve diferença estatística pelo teste de Tukey a 5% de probabilidade. ANEXO C
Gel de DGGE, para os tratamentos T1, T2 e T3 em 2014 (A) e tratamentos T4, T5 e T6 em 2015 (B) de bactérias (rDNA 16S) obtidos com gradiente de desnaturante de 35-65% com quatro repetições para cada tratamento.
A B
70 ANEXO D Resumo da analise de redundância (RDA). Seleção automática realizada com o teste de permutação de Monte Carlo em 2014.
Axes 1 2 3 4
Eigenvalues 0.177 0.101 0.058 0.019
Species-environment correlations 0.936 0.929 0.81 0.652
Cumulative percentage variance
of species data 17.7 27.8 33.6 35.5
of species-environment relation 48.1 75.6 91.2 96.4
Sum of all canonical eigenvalues 0.369
ANEXO E Resumo da analise de redundância (RDA). Seleção automática realizada com o teste de permutação de Monte Carlo em 2015.
Axes 1 2 3 4
Eigenvalues 0.157 0.097 0.051 0.022
Species-environment correlations 0.936 0.938 0.778 0.678
Cumulative percentage variance
of species data 15.7 25.4 30.5 32.6
of species-environment relation 46.9 78.8 91.1 97.6
Sum of all canonical eigenvalues 0.334





























