Embed Size (px)
Citation preview

Aus der Ambulatorischen und Geburtshilflichen Tierklinik der Veterinärmedizinischen
Fakultät der Universität Leipzig
und der
Abteilung Veterinärmedizin des Landesbetriebes des Hessischen Landeslabors, Standort
Gießen
Untersuchungen zur Eutergesundheit in Milchviehbeständen des
Bundesstaates Jalisco, Mexiko
INAUGURAL-DISSERTATION
zur Erlangung des Grades eines Doctor medicinae veterinariae (Dr. med. vet.)
durch die Veterinärmedizinische Fakultät
der Universität Leipzig
eingereicht von
SYBILLE PETRA JÄGER
aus Frankfurt am Main
LEIPZIG, 2006

Mit Genehmigung der Veterinärmedizinischen Fakultät der Universität Leipzig
Dekan: Prof. Dr. Karsten Fehlhaber
Betreuer: Prof. Dr. Axel Sobiraj
Dr. Michael Zschöck
Gutachter: Prof. Dr. Axel Sobiraj, Ambulatorische und Geburtshilfliche Tierklinik,
Veterinärmedizinische Fakultät der Universität Leipzig
Dr. Michael Zschöck, Landesbetrieb des Hessischen Landeslabors,
Gießen
Prof. Dr. Martina Hoedemaker, Klinik für Rinder,
Tierärztliche Hochschule Hannover
Doz. Dr. Peggy Braun, Institut für Lebensmittelhygiene,
Veterinärmedizinische Fakultät der Universität Leipzig
Tag der Verteidigung: 13.06.2006

Inhaltsverzeichnis
1 Einleitung 1
2 Literaturübersicht 2.1 Mastitisformen 3 2.2 Mastitiserreger 6
2.2.1 Kontagiöse, euterassoziierte Mastitiserreger 8
2.2.1.1 Staphylococcus (S.) aureus
2.2.1.2 Äskulin-negative Streptokokken 14
2.2.1.2.1 Streptococcus (S.) agalactiae
2.2.1.2.2 Streptococcus (S.) dysgalactiae 18
2.2.1.3 Corynebacterium (C.) bovis 19
2.2.2 Umweltassozierte Mastitiserreger 21
2.2.2.1 Koagulase-negative Staphylokokken
2.2.2.2 Äskulin-positive Streptokokken 23
2.2.2.2.1 Streptococcus (S.) uberis
2.2.2.2.2 Enterococcus spp. 26
2.2.2.3 Enterobacteriaceae (Coliforme) 27
2.3 Pulsfeldgelelektrophorese (PFGE) 29
2.4 Wirtschaftliche Verluste durch Mastitiden 31
2.5 Klimatische Einflüsse auf das Mastitisgeschehen 35
2.6 Geschichte und heutiger Stand der Milchproduktion in Mexiko 36
2.7 Betriebsformen in Mexiko 41
2.7.1 Spezialisierte Betriebe
2.7.2 Halbspezialisierte Betriebe 42
2.7.3 Familiäre Hinterhofbetriebe 43
2.7.4 Doppelnutzungsbetriebe 44
2.8 Stand Jaliscos im mexikanischen Milchsystem 45
3 Tiere, Material und Methoden 3.1 Milcherzeugerbetriebe und Kuhbestand 49
3.2 Datenerhebung auf den Betrieben 51

3.3 Probenentnahme und Transport 51
3.4 California-Mastitis-Test 52
3.5 Euterpalpation 52
3.6 Zytobakteriologische Untersuchung der Milchproben 53
3.6.1 Anzüchtungsmedien 53
3.6.2 Subkultvierung der Kulturen 54
3.6.3 Identifizierung der Bakterienkulturen 54
3.7 PCR-vermittelter Nachweis speziesspezifischer Genabschnitte 58
3.7.1 Präparation der bakteriellen DNA 58
3.7.2 Verwendete Oligonucleotidprimer und Temperaturprogramme 58
3.7.2.1 Staphylococcus aureus 58
3.7.2.2 Streptococcus agalactiae 59
3.7.2.3 Streptococcus uberis 60
3.7.3 Agarosegelelektrophorese 60
3.7.4 Ethidiumbromidfärbung 60
3.8 Makrorestriktion der chromosomalen DNA und PFGE 61
3.8.1 Präparation der bakteriellen DNA 61
3.8.2 Restriktionsverdau der Gesamtzell-DNA 62 3.8.3 Laufbedingungen der PFGE 62
3.8.4 Auswertung der PFGE-Muster 63
3.9 Statistische Methoden. 63
4 Ergebnisse 4.1 Beschreibung der besuchten Betriebe 64
4.2 Beurteilung der Fütterung 65
4.3 Beurteilung der Melkhygiene und des Trockenstellens 66
4.4 Ergebnisse des CMT 68
4.5 Befunde der Euterpalpation und Sekretbeurteilung 69
4.6 Ergebnisse der bakteriologischsen Untersuchungen 70
4.7 Ergebnisse der molekularbiologischen Untersuchungen 71
4.8 Ergebnisse der Pulsfeldgelelektrophorese (PFGE) 72
4.9 Ergebnisse der statistischen Auswertung 73

5 Diskussion 5.1 Hygienische Verhältnisse von Ausläufen und Melkständen 80
5.2 Vorkommenshäufigkeit subklinischer und klinischer Mastitiden 81
5.3 Prävalenz der einzelnen Mastitiserreger 84
5.4 Melkhygiene und –technik 90
6 Zusammenfassung 95
7 Summary 97
8 Literaturverzeichnis 99
Anhang

Abkürzungsverzeichnis A. dest. Aqua destillata Abb. Abbildung Bp Basenpaare bzw. beziehungsweise C. Corynebacterium ca. circa cAMP Cycloadenomysinmonophosphat cfu colony forming unit cm Zentimeter CMT California-Mastitis-Test D diskriminatoscher Index DGDG Direcion general de ganadería DIN Deutsches Institut für Normung DNA Desoxyribonukleinsäure DVG Deutsche Veterinärmedizinische Gesellschaft E. Escherichia EDTA Ethylen-Diamin-Tetraessigsure et al. et alii g Gramm h Stunde I.E. Internationale Einheit IDF International Dairy Federation INEGI Instituto Nacional de Estadíistica, Geografía e Informática km Kilometer KNS Koagulase-negative Staphylokokken m Meter mg Milligramm ml Milliliter min Minute nm Nanometer NOM Norma oficial mexicana PCR Polymerase-Kettenreaktion pH pondus hydrogenii PFGE Pulsfeldgelelektrophorese RNA Ribnukleinsäure S. Staphylococcus, Streptococcus SAGAR Statistisches Zentrum für Ackerbau und Viehzucht in Mexiko SE Staphyokokken-Enterotoxine spp. Spezies ssp. Subspezies Tab. Tabelle TSST-1 Toxic Shock Syndrome Toxin-1 U Unit U/min Umdrehungen pro Minute UV Ultraviolett V Volt Vol. Volumen WHO Weltgesundheitsorganisation

z.B. zum Beispiel ºC Grad Celsius μg Mikrogramm μl Mikroliter 0.5x halbkonzentriert 10x zehnfach konzentriert


Meinen Eltern
und Freunden aus Mexiko

1
1 Einleitung
Im mexikanischen Staat Jalisco werden täglich 4,7 Millionen Liter Milch produziert. Dieser
ist mit einer Beteiligung an der nationalen Milchproduktion von 17,4% der größte
Milchproduzent Mexikos (SAGAR 2004). So werden zurzeit in diesem westlichen Staat
Mexikos über 800.000 Milchkühe gehalten (JOSÉ ZORRILLA et al. 1996). Insbesondere
durch eine zunehmende Industrialisierung der Betriebe wächst die Milchproduktion im
Einzugsbereich Guadalajaras, der Hauptstadt Jaliscos und zweitgrößten Stadt Mexikos, stetig.
Da Mexiko zur Deckung des nationalen Konsums an Trinkmilch und Frischmilchprodukten
sowie Käse trotz stetig wachsender Produktion immer noch auf enorme Importe insbesondere
aus den USA und Europa angewiesen ist, eröffnen sich für kleine und mittlere
landwirtschaftliche Betriebe im Milcherzeugergebiet Jaliscos gute Verdienstmöglichkeiten
durch die Haltung von Milchkühen. Jedoch kann nur mit gesunden Kühen wirtschaftlich
Milch produziert und den Verbrauchern ein hochwertiges Lebensmittel angeboten werden.
Weltweit die wirtschaftlich und lebensmittelhygienisch bedeutendste Erkrankung der
Milchkuh ist die Mastitis, die Entzündung der Milchdrüse.
Klinische und subklinische Mastitiden sind in den Milchviehherden weit verbreitet und führen
zu Veränderungen in der Zusammensetzung der Milch, zu Leistungsrückgang bis hin zum
vollständigen Funktionsverlust der betroffenen Milchdrüse, selten auch zum Tod des Tieres.
Die Milch erkrankter Kühe ist darüber hinaus mit bakteriellen Krankheitserregern
kontaminiert und daher für den menschlichen Verzehr nicht geeignet. Die Milch klinisch
erkrankter Tiere darf nicht zur Herstellung von Lebensmitteln genutzt werden. Aus den
genannten Gründen stellt die Mastitis ein erhebliches wirtschaftliches Problem dar. Trotz
dieser Bedeutung stehen kaum Informationen zur Mastitissituation in Jalisco und
insbesondere zu den Vorkommenshäufigkeiten der verschiedenen Mastitiserreger zur
Verfügung. Untersuchungen in anderen Regionen Mexikos haben gezeigt, dass mit einer
Prävalenz der subklinischen Mastitis zwischen 40 und 78% gerechnet werden muss (JUÁREZ
et al. 1980; AVILA et al. 1991). Bei an Mastitis erkrankten Schlachttieren in Jalisco konnten
überwiegend kontagiöse Mastitiserreger wie Streptococcus (S.) agalactiae und
Staphylococcus (S.) aureus isoliert werden (WOLTER et al. 2002).
Die primären Ziele der vorliegenden Arbeit lagen in der Ermittlung der aktuellen
Mastitissituation in Jalisco. Es wurden 33 Milchbetriebe besucht, deren Haltungs- und
Melkhygiene beurteilt, Viertelanfangsgemelkproben zur Melkzeit entnommen sowie darauf
folgend die zytobakteriologische Untersuchung dieser Milchproben im Labor durchgeführt,

2
um die Vorkommenshäufigkeiten der einzelnen Mastitiserreger nachzuweisen. Die isolierten
Erreger wurden für weitergehende Untersuchungen subkultiviert und asserviert. Zur
abschließenden Identifizierung der Feldisolate wurden neben den herkömmlichen
phänotypischen Techniken molekulare Untersuchungsverfahren wie etwa der Nachweis
speziesspezifischer Gene bzw. Genabschnitte mittels der Polymerasekettenreaktion (PCR)
durchgeführt. Zum Nachweis epidemiologischer Zusammenhänge wurde zusätzlich die
Makrorestriktionsanalyse der chromosomalen DNA mit anschließender
Pulsfeldgelelektrophorese eingesetzt. Desweiteren wurden von EL-SAYED et al. (im Druck)
durch PCR-vermittelte Analysen das Toxinbildungsvermögen bzw. das Toxin-Genmuster von
17 der asservierten S. aureus-Kulturen untersucht, wodurch die lebensmittelhygienische
Bedeutung dieses Erregers für die mexikanische Bevölkerung in Anbetracht der hohen
Rohmilchdirektvermarktung eruiert werden sollte. Mit Hilfe der statistischen
Datenauswertung wurde versucht, Zusammenhänge zwischen betriebswirtschaftlichen und
melkhygienischen Gegebenheiten und dem nachgewiesenen Erregerspektrum in den
Betrieben aufzuzeigen.
Nur durch genaue Kenntnis der Mastitissituation und Erregerhäufigkeiten wird es zukünftig
möglich sein, individuelle auf mexikanische Verhältnisse angepasste Sanierungsstrategien für
die Milchviehherden zu entwickeln und dauerhaft zu implementieren. Auf diese Weise könnte
die Milchproduktion gesteigert und der hohe wirtschaftliche Verlust der mexikanischen
Milchindustrie minimiert und gleichzeitig dem gesundheitlichen Verbraucherschutz genüge
getan werden.

3
2 Literaturübersicht
2.1 Mastitisformen Die Mastitis wird als eine Entzündung des Milchdrüsengewebes definiert (SMITS et al. 1998;
HERINDSTAD et al. 2000; RIFFON et al. 2001; ZADOKS et al. 2002). Sie gehört zu den
häufigsten Erkrankungen in Milchviehbetrieben und kommt weltweit vor. Wegen der
enormen wirtschaftlichen Verluste in der Milchproduktion und –verarbeitung ist sie der
wirtschaftlich bedeutendste Krankheitskomplex der Milchrinderhaltung. Diese Verluste
werden vor allem durch eine Verringerung der Milchleistung und ein frühzeitiges Merzen von
Milchkühen sowie ein Ansteigen der notwendigen tiermedizinischen Behandlungen
verursacht (CERON-MUÑOZ et al. 2002). Außerdem wird durch die Mastitis die hygienische
Wertigkeit der Milch beeinträchtigt. Aus diesem Grund wird diese Krankheit auch als eine der
teuersten Erkrankungen der Tierproduktion bezeichnet (KIELWEIN 1994; CORREA et al.
2002). Nach JAYARO et al. (1993) und FORSMAN et al. (1997) ist diese komplexe
Faktorenerkrankung ferner eine der am schwierigsten zu kontrollierenden Tierkrankheiten.
Nach WENDT (1998) lassen sich Differenzierungen der Erkrankung nach pathologischen,
klinischen und zeitlichen Aspekten sowie nach Art der auslösenden Ursache vornehmen.
Die Mastitis wird überwiegend durch pathogene Mikroorganismen verursacht, kann aber auch
durch Traumata sowie chemische, thermische oder andere nicht infektiöse Noxen ausgelöst
werden (DEGO u. TAREKE 2003). JAYARO et al. (1993) erwähnen, dass sich die Mastitis
vor allem dadurch von anderen Tierkrankheiten unterscheidet, dass viele verschiedene
Bakterienspezies eine Euterentzündung auslösen können. YANCEY (1999) gibt an, dass die
Mastitis durch ungefähr 135 verschiedene Agentien, zum Großteil Bakterien, hervorgerufen
werden kann.
Neben dem eigentlichen Drüsengewebe (Mastitis) können auch die Zitze (Thelitis) oder das
milchableitende Gangsystem (Galaktophoritis) von dem Entzündungsgeschehen betroffen
sein. Häufig treten Mischformen auf. In der Pathologie unterscheidet man je nach
Krankheitsverlauf, Sekretbeschaffenheit und Art der vorherrschenden Entzündungsmerkmale
katarrhalische, katarrhalisch-eitrige, hämorrhagisch-nekrotisierende, eitrig-abszedierende,
interstitielle und granulomatöse Mastitiden (WEISS 1999).
Anhand klinischer Gesichtspunkte lassen sich die klinische und die subklinische Mastitis
unterscheiden. Klinische Mastitiden sind hierbei durch sicht- und/oder tastbare Anzeichen
einer Entzündung erfassbar, während die subklinische Mastitis nur auf labordiagnostischem

4
Weg sowie die Änderung der kompositionellen Beschaffenheit der Milch zu ermitteln ist
(WENDT 1998; DVG, 2002).
Die klinischen Anzeichen reichen von Flocken und Fibrinklumpen im Vorgemelk mit leichter
Verringerung der Milchmenge bis hin zu deutlichen Entzündungszeichen des Euters wie
vermehrte Wärme, Rötung, Schwellung und Schmerzhaftigkeit. In solchen Fällen ist die
Milch makroskopisch meist deutlich in ihrem Charakter verändert, und es kann zum
vollständigen Versiegen des Milchflusses kommen. Das ermolkene Sekret kann hierbei
wässrig, blutig, serös, mukös, eitrig, jauchig oder breiig verändert sein. Gelegentlich tritt
begleitend eine starke Störung des Allgemeinbefindens mit Fieber auf. Mastitisbedingte
Todesfälle kommen vor.
Anhand ihres zeitlichen Verlaufs und der Ausprägung der Symptome können die akute,
subakute und chronische klinische Mastitis unterschieden werden. Während bei der akuten
Mastitis die oben erwähnten klinischen Symptome im Vordergrund stehen, fehlen bei der
subakuten Mastitis deutliche klinische Veränderungen des Euters. Meist zeigt sich lediglich
eine Flockenbildung im Anfangsgemelk. Als chronische Mastitis bezeichnet man ein
Stadium, in dem es nach einer klinischen Mastitis zu keiner Ausheilung des betroffenen
Euterviertels gekommen ist. Dieses neigt zur Atrophie oder zeigt dauerhaft anomale klinische
Befunde. Durch den vermehrten Ersatz des Drüsengewebes durch Bindegewebe kann es zu
einer fühlbaren Knotenbildung im Euterparenchym kommen. Die Milchbildung kann auf
einem Euterviertel auf Dauer versiegen, so dass es zur Ausbildung eines so genannten
Dreistrichs oder zum Abgang des Tieres wegen der Leistungsminderung kommt (WENDT
1998).
Bei der subklinischen Mastitis hingegen verhindern die Abwehrmechanismen der Kuh zwar
eine akute Entzündung, der Erreger bleibt aber im Euter nachweisbar und kann nicht
eliminiert werden. Es stellt sich ein krankhaftes Gleichgewicht zwischen Mastitiserreger und
Milchdrüse ein (ZSCHÖCK et al. 1998). Neben den drei Merkmalen Entzündung des Euters
ohne klinische Symptome, erhöhter Zellgehalt und Veränderung der chemischen
Zusammensetzung der Milch ist das vierte Charakteristikum der subklinischen Mastitis der
Nachweis von Mastitiserregern. Der Nachweis gilt als erbracht, wenn in zwei von drei
untersuchten Milchproben innerhalb eines Probeentnahmeintervalls von einer Woche
identische Erreger nachgewiesen werden können (DVG 2002).
Die Milch ist zwar grobsinnlich unauffällig, aber ihre chemische Zusammensetzung ist je
nach Grad der Erkrankung verändert. Es kommt zu einer Reduktion der Trockenmasse, und
die Konzentrationen an Gesamtkasein, Laktose, β-Laktoglobulin, α-Laktalbumin, Kalzium,

5
Phosphor, Kalium, Riboflavin und Ascorbinsäure sind unter anderem vermindert. Relativ
erhöht sind die Anteile an Immunglobulinen und an Serumalbumin. Ferner erhöhen sich die
Konzentrationen an Katalase, Natrium und Chlor (KIELWEIN 1994). Die subklinische
Infektion kann jedoch jederzeit zu einer klinisch apparenten Erkrankung werden, wenn der
Wirtsorganismus in seiner Abwehr z.B. durch Stressfaktoren geschwächt wird (ROLLE u.
MAYR 2002).
Die Deutsche Veterinärmedizinische Gesellschaft (DVG 2002) charakterisiert die Kategorien
der Eutergesundheit wie folgt:
1.) Normale Sekretion: Diese liegt vor, wenn die Euterviertel äußerlich keine krankhaften
Veränderungen erkennen lassen und die Milch weder einen erhöhten Zellgehalt
aufweist, noch Mastitiserreger im Sekret nachweisbar sind.
2.) Latente Infektion: Hier bewegt sich die Zellzahl im Normbereich, es können jedoch
pathogene Mikroorganismen im Sekret nachgewiesen werden. Eine direkte
Unterscheidung einer Infektion des Drüsengewebes oder einer alleinigen Besiedelung
des Strichkanals ist hierbei nicht möglich.
3.) Unspezifische Mastitis: Trotz subklinischer Befunde oder klinischer Symptome
können trotz erhöhter Zellzahl wiederholt keine Mastitiserreger im Sekret
nachgewiesen werden.
4.) Mastitis: Liegt eine Mastitis vor, lassen sich sowohl erhöhte Zellzahlen als auch
pathogene Mikroorganismen in Sekret nachweisen.
Ein wichtiges Kriterium für die Beurteilung des Gesundheitszustandes des Euters ist der
Zellgehalt bzw. die Zellzahl der Milch. Dieser gibt den Gehalt an somatischen Zellen in der
Milch an und wird pro Milliliter Milch angegeben. Die somatischen Zellen setzen sich aus
überalterten, verbrauchten Epithelzellen des Drüsengewebes sowie aus Abwehrzellen des
Blutes, wie Leukozyten, Makrophagen, Lymphozyten und Plasmazellen zusammen.
Während sich in der Milch gesunder Euterviertel nur wenige Zellen, vorwiegend
Epithelzellen, finden, steigt bei entzündlichen Veränderungen der Milchdrüse der Zellgehalt
an, wobei der Anteil der Abwehrzellen stark zunimmt (KIELWEIN 1994).
Eine strenge Grenze zwischen gesund und krank kann nicht gezogen werden. Allerdings kann
davon ausgegangen werden, dass ab einem bestimmten Zellzahlniveau mit hoher
Wahrscheinlichkeit eine entzündliche Erkrankung vorliegt, während in niedrigeren
Zellzahlbereichen die Wahrscheinlichkeit hierfür geringer ist.

6
Ein normaler Zellgehalt liegt auf Viertelgemelksebene bei 10.000 bis 100.000 Zellen/ml
Milch (KIELWEIN 1994; WENDT 1998). Bei Überschreitung dieses Wertes ist bereits eine
Änderung der kompositionellen Beschaffenheit der Milch zu erwarten (TOLLE et al. 1977).
Auf Herdensammelmilchebene liegt die Toleranzgrenze bei 150.000 Zellen/ml (ZSCHÖCK et
al. 1998).
Die Streubreite der Zellzahlwerte infizierter Euterviertel ist sehr hoch und bewegt sich
zwischen 100.000 und mehr als 10.000.000 Zellen/ml Milch (KIELWEIN 1994). Dabei
liegen teilweise erhebliche Schwankungen im Zellgehalt infizierter Euterviertel vor, die sich
durch das Wechselspiel zwischen den infektiösen Noxen und der Abwehrreaktion des
Organismus erklären lassen. Hierbei steht die Zellgehaltserhöhung nicht in linearer, sondern
in exponentieller Gesetzmäßigkeit zur Stärke der Noxe. Auch ist zu berücksichtigen, dass
Euterviertel in Bezug auf den Zellgehalt keine funktionelle Einheit darstellen, sondern dass
einzelne Bezirke der Milchdrüse in unterschiedlichem Grade zum Zellgehalt der Milch
beitragen. Die Höhe des Anstiegs des Milchzellgehaltes korreliert mit dem jeweiligen
Mastitiserreger (DJABRI et al. 2002).
Selbst in gut geführten Milchkuhherden macht ungefähr jede zweite Kuh pro Laktation eine
klinische Mastitis durch. Die Mastitisindex für klinisch euterkranke Tiere beträgt auch in
Herden mit geringeren Gesundheitsstörungen ständig 1,0 -1,5. Herden mit entsprechend
höheren Erkrankungsraten von über 2% können an den Rand der Rentabilität gelangen.
2.2 Mastitiserreger
Das Prinzip der ätiologischen Klassifizierung von Mastitiden ist insbesondere aus der Sicht
der mikrobiologischen Diagnostik zweckmäßig. Neben den Bakterien werden in weitaus
geringerem Umfang auch Hefen, Algen und Viren im infizierten Euter gefunden.
Letztgenannte treten jedoch meist nur bei Allgemeinerkrankungen mit Beteiligung der
Milchdrüse auf.
Die Mastitiserreger bestimmen in hohem Maße das Erscheinungsbild und den Verlauf der
Erkrankung. Ein eindeutiger Rückschluss vom klinischen Bild auf den ursächlichen Erreger
ist jedoch nicht möglich (KLEINSCHROTH et al. 1994; WENDT 1994; HEESCHEN 1998).
Die Mastitiserreger können entsprechend ihrer epidemiologischen Eigenschaften in
kontagiöse („klassische“) und konstitutionelle Erreger unterteilt werden (RIFFON et al.
2001).
Kontagiöse Mastitiserreger leben und vermehren sich ausschließlich in der Milchdrüse und
werden primär während des Melkvorgangs von Tier zu Tier übertragen. Dabei ist die

7
infizierte Milchdrüse das Reservoir für die Erreger, und sie können auf Dauer nur in diesem
überleben. Die Erreger sind stark an das Euter adaptiert und finden hier ideale Bedingungen
für ihre Vermehrung. Durch die Ausscheidung erregerhaltiger Milch kommt es zur
Kontamination von Euter- und Zitzenhaut, von wo die Erreger auf andere Körperstellen
übertragen oder in der Umwelt verbreitet werden können. Da die Melkerhände, die
Eutertücher und Melkbecher während des Melkvorgangs ebenfalls Kontakt mit der
kontaminierten Euterhaut und Milch haben, fungieren sie als Vektoren, mit deren Hilfe nicht
infizierte Euterviertel der selben oder anderer Kühe infiziert werden. Zu den kontagiösen
Mastitiserregern gehören Staphylococcus (S.) aureus, Streptococcus (S.) agalactiae, S.
dysgalactiae, S. canis, Mycoplasma spp. und Corynebacterium (C.) bovis (FOX u. GAY,
1993; SMITH et al. 1985; ZSCHÖCK 1999; RIFFON et al. 2001).
Die konstitutionellen Erreger hingegen sind ubiquitär in der Umgebung der Tiere verbreitet
und besiedeln von dort aus das Eutergewebe. Man findet sie auf der Euterhaut, im Haarkleid,
in der Einstreu, auf dem Stallboden und im Stallstaub. Auch die infizierte Milchdrüse stellt
eine Infektionsquelle dar, jedoch ist sie im Vergleich zu den kontagiösen Erregern von nur
untergeordneter Bedeutung (TOLLE et al. 1977; DVG 1994). Infektionen durch diese
Erreger entstehen insbesondere unter einem hohen Infektionsdruck infolge unhygienischer
Haltungsbedingungen und bei einem geschwächten Immunsystem. Das Infektionsrisiko ist in
der Zwischenmelkzeit am größten (DVG 2002), weswegen die durch umweltassoziierte
Erreger ausgelösten Mastitiden durch eine alleinige Verbesserung der Melkhygiene nicht
einzudämmen sind. Die Exposition des nicht infizierten Viertels mit den umweltassoziierten
Erregern tritt permanent auf (ZADOKS et al. 2001). Diese Form der Mastitis findet man
daher häufiger bei meist sehr gut geführten Milcherzeugerbetrieben, die ein Programm zur
Bekämpfung der kontagiösen Mastitiden durchführen und Durchschnittszellzahlen unter
200.000/ml Tankmilch aufweisen (PHUEKTES et al. 2001; ROSSITTO et al. 2002).
Zu der Gruppe der umweltassoziierten Mastitiserreger gehören verschiedene, meist Äskulin-
positive Streptococcus spp. außer den oben bereits genannten, verschiedenartige
gramnegative Bakterien, wie Escherichia (E.) coli, Klebsiellen und Enterobacter sowie die
Gruppe der Koagulase-negativen Staphylokokken (KNS). Unter den umweltassoziierten
Streptokokken ist insbesondere S. uberis zu erwähnen. Ebenfalls häufig nachweisbar sind S.
parauberis, S. bovis und die verschiedenen Enterokokkenspezies.
Die Mastitiserreger können auch entsprechend der Stärke des von ihnen im Kuheuter
ausgelösten Entzündungsgeschehens in majorpathogen und minorpathogen eingeteilt werden.
Als majorpathogen werden besonders häufig auftretende Keime, insbesondere mit hoher

8
Kontagiosität, Erreger klinischer Mastitiden, Erreger, die starke entzündliche Reaktionen
hervorrufen und Erreger, die besonders hohe finanzielle Verluste auslösen, angesprochen.
Hierzu gehören vor allem S. aureus, S. agalactiae, S. dysgalactiae, S. uberis sowie
Enterokokken, Mycoplasma spp. und die Gruppe der coliformen Keime (WILSON et al.
1997; ARIZNABARRETA et al. 2002; DJABRI et al. 2002).
Minorpathogen dagegen sind Erreger, die nur moderate Zellzahlerhöhungen hervorrufen und
selten klinische Symptome auslösen. Diese Infektionen treten häufig mit hohen Prävalenzen
auf. Zu diesen werden insbesondere verschiedene Corynebacterium spp., die KNS und
Mikrokokken gezählt.
Über 90% der Euterentzündungen des Rindes beruhen auf Infektionen mit S. aureus, S.
agalactiae, S. uberis und S. dysgalactiae (NEAVE 1975). Diese werden somit als die
dominierenden Mastitiserreger angesehen (BRAMLEY u. DODD 1984; JAYARAO et al.
1992; WILSON et al. 1997, FORSMANN et al. 1997, PHUEKTES et al. 2001). Die
Prävalenzen der verschiedenen Spezies variieren dabei geographisch, zeitlich und mit den im
Herdenmanagement verankerten Kontrollmaßnahmen (FORSMANN 1997).
2.2.1 Kontagiöse, euterassoziierte Mastitiserreger
2.2.1.1 Staphylococcus (S.) aureus Staphylococcus (S.) aureus wird zusammen mit den übrigen Vertretern der Gattung
Staphylococcaceae der Familie der Micrococcaceae zugeordnet. Der Gattungsname
Staphylococcus leitet sich aus dem griechischen Wort „staphyle“= Weintraube ab und wurde
vom schottischen Chirurgen OGSTON im Jahre 1882 geprägt, als dieser mikroskopisch
traubenförmig angeordnete Kugelmikroben in Eiter wahrnahm.
Staphylokokken sind Besiedler der Haut, der Hautdrüsen und der Schleimhäute des Menschen
und warmblütiger Tiere (KLOOS u. SCHLEIFER 1986).
Staphylokokken können aufgrund ihrer vorhandenen oder nicht vorhandenen Fähigkeit das
Enzym Koagulase zu bilden, welches zur Koagulierung von Blutplasma führt, in zwei große
Untergruppen, die Koagulase-positiven und –negativen Staphylokokken eingeteilt werden
(BAIRD-PARKER 1974). Die weitaus größere pathogene Bedeutung wird den Koagulase-
positiven Staphylokokken eingeräumt. Die Koagulase-positiven Staphylokokken lassen sich
durch Zuordnung zu unterschiedlichen Biovaren in S. aureus (Biovare A-D) und S.
intermedius (Biovare E und F) unterteilen. Von DEVRIESE (1977) wurde noch die Spezies S.
hyicus mit zwei Subspezies den Koagulase-positiven Staphylokokken zugeordnet.

9
Der weitaus wichtigste veterinärmedizinische Vertreter dieser Gruppe ist S. aureus.
S. aureus verursacht vielgestaltige Krankheitsbilder bei Mensch und Tier. Häufig sind die
Erreger an eitrigen Infektionsprozessen beteiligt, denen meist eine Verletzung oder andere
Prädispositionen des Wirtsorganismus vorausgegangen sind. Ferner werden beim Menschen
Septikämien, Endokarditiden, Osteomyelitiden, Lungenerkrankungen und das toxische
Schock-Syndrom ausgelöst (SUTRA u. POUTREL 1994; WESSON et al. 1998;
FITZGERALD et al. 2001). Als fakultativer Krankheitserreger tritt S. aureus beim Tier vor
allem in Zusammenhang mit Mastitis, Synovitis, Furunkelbildung, eitriger Dermatitis,
Abszessen, Phlegmonen, Pyämien und Septikämien auf. Die Krankheitsprozesse können
örtlich begrenzt sein, flächenhaften Charakter annehmen, aber auch in eine Septikämie mit
schwerem Krankheitsverlauf münden (ROLLE u. MAYR 2002).
Die Staphylokokken-Mastitis des Rindes ist die veterinärmedizinisch bedeutendste
Staphylokokken-Infektion. S. aureus ist weltweit der bedeutendste Erreger der subklinischen
Mastitis dieser Tierart. Aus 20-60% der Milchproben von subklinischen Mastitiden beim Rind
können Staphylokokken nachgewiesen werden (SEFFNER u. BERGMANN 1994). SUTRA
und POUTREL (1994) geben an, dass S. aureus mit 25-30% an intramammären Infektionen
des Rindes beteiligt ist. Aber auch wenn die chronische, subklinische Form am häufigsten
vorkommt, so kommen auch schwerwiegende klinische Euterentzündungen vor (ZSCHÖCK
et al. 1998). Diese treten meist akut bis perakut als hämorrhagisch-nekrotisierende oder
gangräneszierende Mastitis mit unter Umständen tödlichem Verlauf auf (SEFFNER u.
BERGMANN 1994) und häufen sich vor allem am Ende der Laktation und kurz vor bzw.
nach der Abkalbung (BRÜCKLER u. SCHWARZ 1994). EDWARDS und SMITH (1966)
führten detaillierte Studien an drei Herden durch, und zeigten, dass 22% der S. aureus-
Infektionen mit klinischen Erscheinungen einhergingen. Die S. aureus-Mastitiden verlaufen
als katarrhalisch-eitrige Mastitis und Galaktophoritis und verursachen der Milchindustrie
große wirtschaftliche Verluste durch eine Minderung der Milchproduktion sowohl in
Quantität als auch in Qualität (SCHALM et al. 1971; BRÜCKLER u. SCHWARZ 1994). Da
die Erkrankung häufig als subklinische Mastitis unerkannt vorkommt, stellt sie auch eine
mögliche Gefahr für die Gesundheit des Menschen dar (GILMOUR u. HARVEY 1990).
Einige der von diesen Infektionen isolierten Kulturen sind nämlich in der Lage extrazelluläre,
auf den Verdauungstrakt des Menschen wirkende Toxine zu bilden, die
Lebensmittelvergiftungen hervorrufen können (BRÜCKLER u. SCHWARZ 1994).
S. aureus kommt als Kommensale vor allem in den vorderen Nasenpartien und auf der Haut
von Rindern vor (LACEY 1981). MATTHEWS et al. (1986) zeigten außerdem, dass 7% der

10
Strichkanäle der Färsen einer Herde, die kurz vor der Geburt standen, mit dem Erreger
besiedelt waren. Das Hauptreservoir ist die infizierte Milchdrüse (NEAVE et al. 1969;
MATOS et al. 1991). Neuinfektionen erfolgen in der Regel über den Strichkanal. Eine
Ausbreitung findet dann meist über die natürlichen Gangsysteme der Drüsen statt.
Daneben ist ein Eindringen der Erreger über Zitzen- und Zitzenhautverletzungen möglich
(SCHALM et al. 1971). Diese entstehen häufig durch nicht korrektes Melken. MATOS et al.
(1991) zeigten mit ihren Untersuchungen, dass 40% der Zitzenläsionen einer Herde mit S.
aureus kolonisiert waren. NEAVE (1971) wies nach, dass es in einer Herde mit ungenügender
Melkhygiene zu einer Kontamination der Strichkanalöffnungen mit S. aureus bei 60% der
gesunden Viertel und bei 85% der Viertel mit Zitzenhautverletzungen kommen konnte. S.
aureus ist bei weitem nicht so stark an das Euter adaptiert wie z.B. S. agalactiae. Daher
vergehen meist längere Zeiträume, bis Staphylokokken-Neuinfektionen einer Herde zu
nachweisbaren Erkrankungen führen (SEFFNER u. BERGMANN 1994).
Übertragen werden die Keime durch Kontakt- und Schmierinfektionen, hauptsächlich beim
Melken (NEWBOULD 1968; MATOS et al. 1991). Als Vektoren dienen hier insbesondere
die Melkerhände, Euterreinigungstücher und Melkzeuge. Insbesondere aus Haarrissen älterer
Zitzengummis kann in S. aureus-Problembetrieben der Erreger häufig isoliert werden
(SEFFNER u. BERGMANN 1994). Auch das Zurückfließen von Milch innerhalb eines
Melkzeugs nach Lufteinbrüchen kann zur Erregerausbreitung beitragen. MATOS et al. (1991)
konnten in 50% der Luftproben aus den Melkständen in Problembetrieben während der
Melkzeit S. aureus nachweisen, sodass von der möglichen Rolle von Aerosolen bei der
Erregerübertragung ausgegangen werden muss. Andere Quellen, vor allem in der Umwelt der
Tiere, werden für die Transmission der Erkrankung ebenfalls diskutiert (BRAMLEY u.
DODD 1984; ROBERSON et al. 1994).
Meist sind mehrere Viertel eines Tieres betroffen, wobei anfangs meist zunächst die
Hinterviertel infiziert sind (SEFFNER u. BERGMANN 1994).
Nach der Infektion über den Strichkanal lagert sich S. aureus bevorzugt an den Epithelzellen
des Strichkanals und in noch stärkerem Maße in der Drüsenzisterne und in den großen
Milchgängen an (FROST et al. 1977; ROLLE u. MAYR 2002). Diese selektive Adhärenz, die
auf Oberflächenstrukturen der Bakterien, Rezeptoren der Wirtszellen und hydrophoben
Interaktionen beruht, ist ein wichtiger Faktor in der Pathogenese der Staphylokokken-
Mastitis. Dieser initialen Adhärenz folgt die Multiplikation der Erreger und ihre Verbreitung
über das Gangsystem des Euters. WESSON et al. (1998) beschreiben eine durch S. aureus
verursachte Apoptose der Epithelzellen des Rindereuters.

11
Die weitere Ausbreitung der Staphylokokken im Gewebe erfolgt mit Hilfe mutmaßlicher
Virulenzfaktoren wie der Hämolysine, der Leukozidine, der Staphylokinase und der
Hyaluronidase (ROLLE u. MAYR 2002).
S. aureus dringt tief in das Eutergewebe ein und kann sich dort abkapseln. Besonders nach
Invasion der Epithelzellen kann der Erreger intrazellulär über mehrere Monate persistieren
und wird meist nicht einmal in der Trockenstehphase eliminiert (BRAMLEY u. DODD
1984). Ein Grund hierfür ist, dass S. aureus in der Lage ist, eine Reihe von Substanzen zu
bilden, mit deren Hilfe die Erreger den Abwehrmechanismen des Wirts widerstehen und
diesen schädigen können (BRÜCKLER u. SCHWARZ 1994).
Im fortgeschrittenen Stadium der S. aureus-Mastitis kann es durch Gewebsnekrosen zu
fibrotischen Veränderungen bis hin zu einer vollständigen bindegewebigen Atrophie eines
Euterviertels kommen (PLATONOW u. BLOBEL 1963).
Die Behandlung ist trotz moderner Chemotherapeutika und unterstützender Maßnahmen nicht
immer erfolgreich. Meist können im Euter trotz in vitro-Empfindlichkeit des Erregers keine
ausreichenden Wirkstoffkonzentrationen erreicht werden, da die Koagulase-bedingte
Plasmagerinnung und die Keimverklumpung die Erreger vor dem Chemotherapeutikum
abschotten. Durch die bei der chronischen Mastitis auftretenden Gewebsnekrosen und
fibrotischen Veränderungen wird zudem die Diffusion der Substanzen behindert oder sogar
unmöglich gemacht. Auch können viele Antibiotika nicht zu den in den Phagolysosomen der
Makrophagen vorliegenden Keimen vordringen (SUTRA u. POUTREL 1994). Durch zu
niedrige Wirkspiegel und zu kurze bzw. zu lange Behandlungszeiten kann es außerdem zur
Förderung einer Resistenzentwicklung kommen. Bezüglich der antibiotischen
Empfindlichkeit des Erregers resultierte in den vergangenen Jahren eine zunehmende
Resistenz gegenüber Penicillin G durch die Bildung von β-Laktamase.
Ferner können sich Staphylokokken im Gewebe vorübergehend in eine Art
Stoffwechselruhelage begeben, sodass Chemotherapeutika nicht wirksam werden. So ist bei
chronischen Rindermastitiden bedingt durch S. aureus kein Heilungserfolg über 80% zu
erwarten (BRÜCKLER u. SCHWARZ 1994). PHILPOT und NICKERSON (1991) sowie
PYÖRÄLÄ und PYÖRÄLÄ (1994) geben an, dass 20-70% der antibiotischen Behandlungen
während der Trockenstehphase effektiv sind und 10-30% der S. aureus-Mastitisfälle während
der Laktation durch eine antibiotische Behandlung geheilt werden können. Auch SUTRA und
POUTREL (1994) geben für eine antibiotische Therapie in der Laktation Heilungschancen
von weniger als 50% an. Therapieresistente Tiere sollten abgeschafft werden, da sie aufgrund

12
der Erregereigenschaften eine ständige Infektionsgefahr für die übrigen Tiere des Bestandes
darstellen.
Die Bekämpfung der S. aureus-Mastitis liegt vor allem in der Prävention der
Erregerübertragung von Tier zu Tier. Hierdurch kann die Mastitisinzidenz langfristig
reduziert werden (NEAVE et al. 1969). Jedoch ist die Sanierung einer Herde selbst bei
strengster Einhaltung der Hygienevorschriften schwierig, und es kann insbesondere durch die
ubiquitäre Verbreitung des Erregers und sein Vorkommen außerhalb des Wirtsorganismus
durch kontaminierte Einstreu, Futter, Equipment oder Kontakt mit der Haut anderer Tiere und
des Menschen immer wieder zu Neuinfektionen kommen (ROBERSON et al. 1994). Neuere
epidemiologische Untersuchungen von SOMMERHÄUSER 2002 und SOMMERHÄUSER et
al. (2003) von S. aureus-Isolaten aus mehreren hessischen Milchviehbetrieben zeigten, dass
der Erreger neben seiner typischen kuh- bzw. euterassoziierten kontagiösen Form auch mit
den epidemiologischen Charakteristika von umweltassoziierten Keimen auftreten kann. Diese
Isolate treten vor allem in Herden mit niedriger S. aureus-Prävalenz und geringer
Ausbreitungstendenz auf und scheinen die Kontrollmaßnahmen für kontagiöse Mastitiserreger
zu umgehen. Daher führen sie trotz Durchführung von Sanierungsprogrammen zu hohen
Neuinfektionsraten innerhalb der betroffenen Herden.
Lebensmittelhygienisch von besonderer Bedeutung ist das Vermögen einiger
Staphylokokken, Enterotoxine zu bilden. Dieses ist insbesondere für S. aureus beschrieben, es
wurden aber auch bei S. intermedius und bei verschiedenen KNS wie z.B. S. epidermidis und
S. haemolyticus Enterotoxine nachgewiesen (UNTERMANN 1994).
Staphylokokken-Enterotoxine (SE) sind einfache Proteine, die aus einer einzigen
Polypeptidkette bestehen. Sie haben ein Molekulargewicht von 28 000 bis 35 000 Dalton. Die
SE sind resistent gegenüber proteolytischen Enzymen wie Trypsin, Chymotrypsin, Renin und
Papain (UNTERMANN 1994). Aufgrund ihrer einfachen Struktur sind sie außerdem äußerst
hitzestabil. So überstehen sie die Pasteurisierung und das Abkochen, sodass in einem
pasteurisierten Milchprodukt noch aktives SE vorhanden sein kann, obwohl die
Staphylokokken selbst abgetötet sind (BONE et al. 1989; OTTO 1991). Es gibt verschiedene
serologisch unterschiedliche Typen dieser Toxine. Neben den bekannten klassischen
Enterotoxinen SEA, SEB, SEC, SED und SEE wurden in den letzten Jahren vier weitere
Enterotoxine beschrieben: SEG und SEI (MUNSON et al. 1998), SEH (REN et al. 1994) und
SEJ (ZHANG et al. 1998). Desweiteren wurden in jüngster Zeit mit den SEK, SEL, SEM,

13
SEN und SEO fünf ganz neue Typen von Enterotoxinen entdeckt (JARRAUD et al. 2001;
ORWIN et al. 2001; ORWIN et al. 2002; LETERTRE et al. 2003).
Das Toxinbildungsvermögen boviner S. aureus-Stämme variiert stark und wird daher in der
Literatur unterschiedlich angegeben. HEESCHEN (1994) gibt an, dass etwa 5-10% der
Erreger der bovinen Mastitis ein oder mehrere verschiedene Enterotoxine bilden können.
MÜLLER (1992) schätzt die Rate auf 4-40%. LARSEN et al. (2000) überprüften 414 bovine
S. aureus-Stämme, und nur einer von ihnen beherbergte Gene zur Produktion von SEC und
TSST-1. Von 166 bovinen S. aureus-Isolaten aus Korea, die LIM et al. (2004) mit Hilfe der
PCR auf das Vorkommen von Enterotoxingenen untersuchten, beherbergten 32 Isolate das
SEA-Gen, drei Isolate das SEB-Gen und nur ein Isolat das SEC-Gen. ZSCHÖCK et al.
(2004) überprüften ebenfalls mittels PCR 533 Mastitisisolate auf das Vorkommen von SEC-
Gen und TSST-1-Gen und fanden eine Prävalenz von 9,3%. In einer weiteren Untersuchung
konnten ZSCHÖCK et al. (2005) bei 58,7% von 104 untersuchten Stämme aus deutschen
Milchviehbetrieben eines oder Kombinationen der neuen Enterotoxine SEG-SEJ nachweisen.
Von diesen positiven Stämmen produzierten wiederum 60,7% zusätzlich eines der klassischen
Enterotoxine SEA-SEE und TSST-1.
Interessanterweise fanden PUIG de CENTORBI et al. (1992) bei der Untersuchung von 27
Stämmen aus Argentinien kein einziges Enterotoxin- oder TSST-1 produzierendes Isolat.
Die zur Enterotoxikose führende Kontamination von Milch und Milcherzeugnissen ist nach
Meinung von KIELWEIN (1994) und UNTERMANN (1994) nicht bedingt durch eine
primäre Kontamination durch das erkrankte Tier, sondern durch die sekundäre Kontamination
der Milch durch infizierte Wunden und Hautausschläge an den Händen des Personals. Andere
Autoren vermuten aber, dass die Milch erkrankter Tiere die Hauptquelle tierischen Ursprungs
für enterotoxinbildende Staphylokokken darstellt (BRYAN 1983; BERGDOLL 1989;
GILMOUR u. HARVEY 1990).
Bereits die Menge von 0,1-0,2 µg Enterotoxin kann bei in der Abwehrkraft geschwächten
Menschen Erkrankungen hervorrufen (EVENSON et al. 1988). Im Durchschnitt werden
jedoch bei oraler Aufnahme von 10 µg Intoxikationen ausgelöst (BERGDOLL 1970;
KIELWEIN 1994). Die klinischen Symptome reichen von Übelkeit, Erbrechen und Durchfall,
Kopfschmerzen und Wadenkrämpfen bis hin zu schweren Kreislaufstörungen, die bei
vorgeschädigten Personen zu lebensbedrohlichen Schockzuständen führen können. In der
Regel ist die Erkrankung bereits nach 1-2 Tagen überwunden.
Die Kontamination eines Lebensmittels mit geringen Mengen enterotoxinbildender
Staphylokokken stellt meist keine akute Gesundheitsgefährdung dar. Erst wenn es nach einer

14
intensiven Vermehrung der toxinbildenden Erreger im Lebensmittel auf Keimzahlen bis zu
105 - 106 Erreger/g kommt, ist mit einer Krankheitsauslösung durch Bildung von
Enterotoxinen zu rechnen (TATINI et al. 1971; GILBERT 1974; KIELWEIN 1994;
HEESCHEN 1998). In Rohmilch wird die Toxinbildung durch die natürliche Begleitflora aus
überwiegend proteolytischen und saccharolytischen Bakterien mit kurzen Generationszeiten
gehemmt. Besondere Verhältnisse bestehen jedoch, wenn Rückstände von Antiinfektiva in
der Milch vorhanden sind, die eine natürliche Milchflora in ihrer Entwicklung hemmen und
enterotoxinbildenden Staphylokokken, wenn diese eine Resistenz aufweisen, eine starke
Vermehrung ermöglichen (ZEHREN u. ZEHREN 1968; TATINI et al. 1971).
Die Häufigkeit von Staphylokokken-Lebensmittelvergiftungen lässt sich schwer einschätzen.
Nach MEAD et al. (1999) liegt die geschätzte jährliche Zahl der Erkrankungsfälle in den
USA bei über 180.000.
2.2.1.2 Äskulin-negative Streptokokken
2.2.1.2.1 Streptococcus (S.) agalactiae S. agalactiae ist der Erreger des so genannten „Gelben Galtes“, einer hochkontagiösen
Euterentzündung des Rindes, die in allen Ländern mit intensiver Milchwirtschaft
nachgewiesen werden kann (LÄMMLER u. HAHN 1994). Der Begriff Galt stammt aus dem
Süddeutschen bzw. Schweizerischen und bedeutet Milchrückgang bzw. völliges Versiegen
der Milch. Der Erreger wird serologisch der Lancefield-Gruppe B zugeordnet. Eine weitere
Subtypisierung von S. agalactiae ist mit Hilfe seiner Typenantigene, wie den Polysaccharid-
Antigenen Ia, Ib, II, III, IV, V und den Protein-Antigenen c, R und X möglich. Diese sind
besonders bei epidemiologischen Untersuchungen von Bedeutung (HEJLICEK 1994). Der
Erreger reagiert Äskulin-negativ, sein positives CAMP-Phänomen erlaubt eine zuverlässige
Diagnose. Die seit dem Ende des 19. Jahrhunderts bekannte Eutererkrankung stellt ein
schwerwiegendes wirtschaftliches und milchhygienisches Problem dar, weil es durch den
Erreger zu einer starken Erhöhung des Milchzellgehaltes sowie der Gesamtkeimzahl durch
Primärkontamination und somit zu einer Verminderung der Milchqualität, vor allem auch der
–quantität kommt.
Durch S. agalactiae ausgelöste Mastitiden nehmen meist einen subklinischen und chronischen
Verlauf, seltener löst der Erreger klinisch-katharrhalische Mastitiden aus (ERSKINE et al.
1996). Meist tritt die Erkrankung bestandsweise gehäuft auf. In betroffenen Betrieben liegt
die Herdenprävalenz zwischen 10 und 50% (KEEFE 1997). Ferner wird die Infektion zu den

15
Erkrankungen gezählt, die bei Tieren mit einer hohen Milchleistung häufiger vorkommt als
bei Tieren mit einer geringeren Laktationsleistung.
Mittels Genotypisierung von S. agalactiae-Isolaten konnte die Kontagiosität dieses Erregers
bestätigt werden. So zeigten BASSEGIO et al. (1997) und WANG et al. (1999) mit Hilfe der
Makrorestriktionsanalyse, dass meist ein einziger Klon für das Mastitisgeschehen in einer
Herde verantwortlich ist. Mit der gleichen Methode untersuchten MERL et al. (2003) 79 S.
agalactiae-Isolate aus sieben hessischen Milchviehbetrieben und konnten zeigen, dass in
jedem einzelnen Betrieb identische oder zumindest eng miteinander verwandte Stämme für
die Mastitissituation verantwortlich waren. Die Stämme der einzelnen Betriebe unterschieden
sich jedoch deutlich voneinander und ließen keinerlei epidemiologischen Zusammenhang
zwischen den Einzelbetrieben erkennen.
Insgesamt ist in Ländern mit intensiver Milchwirtschaft die Verbreitung des Erregers als
direkte Folge seiner hohen Empfindlichkeit gegenüber Penicillin G deutlich zurückgegangen
(BRAMLEY u. DODD 1984). Vor der Entwicklung der Antibiotika galt S. agalactiae als
einer der Haupterreger von Mastitiden (KEEFE 1997). Von allen Streptokokken-Arten kam er
mit Abstand am häufigsten vor (TOLLE 1982; VECHT et al. 1989).
Zwischen 1942 und 1962 fiel der prozentuale Anteil nachgewiesener S. agalactiae-
Infektionen durch den Einsatz von Antibiotika von 44% auf 4% aller nachgewiesenen
Erreger-bedingten Mastitiden (WILSON 1963). Im Bundesland Hessen konnte nach der
Einführung von flächendeckenden Präventiv- und Kontrollmaßnahmen die Prävalenz noch
weiter gesenkt werden (WOLTER et al. 1999). Durch die Auswertung der von KLOPPERT et
al. (1999) publizierten Daten des Eutergesundheitsdienstes Hessen kamen die Autoren auf
eine geschätzte Infektionsrate in Hessen für das Jahr 1998 von 0,5% aller Betriebe bzw. 0,3%
aller Milchkühe. In schlecht geführten Betrieben neigt der Erreger zur schnellen Ausbreitung
(WILSON et al. 1997).
S. agalactiae ist ein obligater Keim der Milchgänge des Kuheuters, welches im infizierten
Zustand das Hauptreservoir darstellt (SEELEMANN 1932; KEEFE 1997). Im Euter findet S.
agalactiae optimale Verhältnisse zur Vermehrung vor und wird mit der Milch in hohen
Mengen ausgeschieden (ROLLE u. MAYR 2002). In infizierten Herden kann der Erreger in
der Luft, der Einstreu, sämtlichen mit kontaminierter Milch in Berührung kommenden
Gegenständen, sowie an den Händen des Melkpersonals nachgewiesen werden. Laut
HEJLICEK (1994) bleibt er auf den Händen und der Kleidung des Melkpersonals bis zu 10
Tage infektionsfähig. Diese Vektoren gehören somit neben den Eutertüchern und
Melkbechern der Melkmaschine zu den häufigsten Überträgern (SEELEMANN 1954;

16
PHILPOT 1975). Auch durch kontaminierte Zitzentauchlösungen können die Erreger
verbreitet werden. Junge Kühe sind widerstandsfähiger gegen den Gelben Galt als ältere
(HEJLICEK 1994).
Der Erreger dringt über den Strichkanal ein und zeigt eine selektive Adhärenz zu den
Epithelzellen des Strichkanals, besonders aber zur Milchzisterne und den großen
Milchgängen, wo er sich in großer Zahl anlagert (ROLLE u. MAYR 2002). Hier findet laut
SCHULZ (1994) die erste intramammäre Auseinandersetzung zwischen Wirtsorganismus und
Erreger statt. Nach FOX und GAY (1993) dringt S. agalactiae nicht in das Drüsengewebe ein,
sondern heftet sich den Epithelzellen an, die unter anderem durch die Milchsäureproduktion
und toxischen Substanzen, die aus den zerfallenden Streptokokken freiwerden, zerstört
werden. Von den distalen Abschnitten der Milchdrüse breitet sich die Infektion kanalikulär in
die höher gelegenen Milchgänge und die Alveolen aus (HEJLICEK 1994). Ein einmal
infiziertes Euter kann mehrere Laktationsperioden hindurch den Erreger beherbergen und
somit eine ständige Infektionsquelle für alle Tiere des Bestandes darstellen (ROLLE u.
MAYR 2002). Dies liegt vor allem an der niedrigen Selbstheilungsrate (FARNSWORTH
1987).
Die Erkrankung verläuft meist chronisch, da das vorangehende akute Stadium durch seine
kurze Dauer klinisch oft unbemerkt bleibt. Die ersten Anzeichen sind Veränderungen der
Milch, wie eine schmutzige Rahmschicht und grau-weißes durchsichtiges Aussehen. Zu
Beginn der Erkrankung liegt eine katarrhalische Entzündung der Milchgänge und Alveolen
vor. Später kommt es zur Atrophie des Alveolarepithels, verbunden mit einer Verkleinerung
der Alveolen und einer Reduktion bzw. einer kompletten Einstellung der Milchsekretion in
den betroffenen Drüsenläppchen. Gleichzeitig entwickelt sich häufig eine proliferative
Entzündung der Milchgänge mit knotigen Verdickungen der Zisternenwand (HEIDRICH u.
RENK 1963). Zu diesem Zeitpunkt treten dann auch Eiterflocken im Anfangsgemelk auf. Es
kommt zu einer Vermehrung des interstitiellen Bindegewebes, und die Drüsenläppchen
atrophieren. In den seltener vorkommenden akuten Fällen steht eine katarrhalisch-eitrige
Galaktophoritis mit Hyperämie und Ödem des Unterhautbindegewebes im Vordergrund.
WENDT (1998) gibt den Anteil klinischer Mastitisfälle mit etwa 5% an. Die Tagesleistung
kann sich nach HEJLICEK (1994) auf bis zu einem Zehntel der sonstigen Menge verringern,
wobei die Milch meist mit zahlreichen Kasein- und Eiterflocken durchsetzt ist. STADTFELD
und NITZSCHKE (1980) beschrieben ferner, dass sich die klinische Form der S. agalactiae-
Mastitis durch eine mehr oder weniger deutliche Gelbverfärbung der Milch äußere.

17
Meist ist zunächst ein Euterviertel von der Infektion betroffen, nachfolgend erkranken dann
auch die übrigen Viertel der Kuh.
Im Verlauf der chronischen Form lassen sich eine Asymmetrie des Euters, strangförmige
Verdickungen der proliferierten Milchkanäle und knotige Bezirke im Drüsenteil feststellen
(ROLLE u. MAYR 2002). Der „Gelbe Galt“ führt meistens zu einem Verlust großer Teile des
milchbildenden Gewebes, das auch nach erfolgreicher Behandlung nicht mehr regeneriert
(HEJLICEK 1994). WENDT (1998) gibt die Milchverluste durch Gewebsuntergang mit 15-
35% an.
Da S. agalactiae längere Zeit nur im Euter überleben kann und gegenüber einer Penicillin G-
Therapie empfindlich ist, ist eine Eradikation des Keimes durch Verbesserung der
Melkhygiene und eine lokale Behandlung der infizierten Tiere mit Antibiotika aus
geschlossenen Herden möglich (KEEFE 1997).
Die antibiotische Therapie einer mit Galt infizierten Herde erreicht während der Laktation
Heilungsraten von über 90% und ist damit signifikant erfolgreicher als Spontanheilungen
(WILSON et al. 1999; WOLTER et al. 1999). Die Spontanheilungsrate während des
Trockenstehens wird von NATZKE et al. (1975) mit 36-42% angegeben. Bei der Behandlung
wird die hohe Empfindlichkeit des Erregers gegenüber Penicillin G genutzt. HEJLICEK
(1994) gibt diese mit 95,6-100% an.
ERSKINE und EBERHART (1990) konnten in ihrer Studie zeigen, dass in 12 Herden der
durchschnittliche Zellgehalt der Tankmilch mittels der Laktationstherapie innerhalb von 30
Tagen von 918.000 auf 439.000 Zellen/ml Milch gesenkt werden konnte. Durch eine weitere
Verbesserung der Herdenmanagements wie z.B. das Zitzendippen nach dem Melken und das
konsequente Trockenstellen aller Tiere mit Langzeitantibiotika gelang eine weitere Senkung
innerhalb eines Jahres auf durchschnittlich 268.000 Zellen. Die Autoren zeigten ferner, dass
durch die Durchführung dieses Programms ein Überschuss von 512 kg Milch und 14 kg Fett
pro Kuh innerhalb des ersten Jahres erwirtschaftet werden konnte, wodurch die
Kosteneffizienz der so genannten Blitztherapie eindeutig belegt werden konnte.
Die Infektionsmöglichkeit für den Menschen durch den Verzehr von kontaminierter Rohmilch
ist noch nicht vollkommen geklärt. Es können jedoch beim Melkpersonal verstärkt
Racheninfektionen auftreten. HAHN et al. (1972) konnten bei Patienten einen statistischen
Zusammenhang zwischen Rohmilchverzehr und dem Nachweis von B-Streptokokken
feststellen.

18
S. agalactiae ist ein Keim, der bei vielen Tierarten vorkommen kann. Neben dem Rind konnte
er unter anderem von Hund, Katze, Schwein und Nutria isoliert werden (SMOLA 1993,
YILDIRIM et al. 2002). Auch in der Humanmedizin besitzen Streptokokken der Lancefield-
Gruppe B eine herausragende Bedeutung. So sind sie ein bedeutender Krankheitserreger der
so genannten Neugeborenenseptikämien, die häufig perakut verlaufen und mit letal
verlaufenden Meningitiden einhergehen (BAKER, 1980; ROSS, 1984). Hier wird der Erreger
nach seinem antigenen Verhalten als Group B-Streptococcus (GBS) bezeichnet. Neben der
Infektion von Neugeborenen und Schwangeren sind B-Streptokokken als ursächliche Erreger
von Pneumonien, Pyelonephritiden, Arthritiden, Endokarditiden, abszedierenden Mastitiden
und Septikämien bekannt (ROSS 1984). Diese verlaufen häufig mit einer hohen
Mortalitätsrate und treten gehäuft bei älteren Patienten mit chronischen Erkrankungen bzw.
bei immunsupprimierten Personen auf (OPAL et al. 1988).
Das Vorkommen von B-Streptokokken bei Rind und Mensch wirft die Frage auf, inwieweit
zwischen den beiden Populationen ein epidemiologischer Zusammenhang besteht. Obwohl
die Bedeutung als Zooanthroponose beschrieben wurde (NIELSEN 1987), konnten von
zahlreichen Autoren deutliche Unterschiede in kulturellen, biochemischen, serologischen und
molekularbiologischen Eigenschaften zwischen vom Mensch und Rind isolierten Kulturen
aufgezeigt werden (FINCH u. MARTIN, 1984; DEVRIESE 1991; LÄMMLER et al. 1993;
BECKER 1994; KIELWEIN 1994; LÄMMLER u. HAHN 1994; DMITRIEV et al. 1999;
YILDIRIM et al. 2002). Danach scheinen Infektionen zwischen Mensch und Rind nur in
Ausnahmefällen möglich zu sein. Auch LÄMMLER und HAHN (1994) halten eine
Ansteckung über erkrankte Tiere oder rohe Milch für eher unwahrscheinlich. Es ist allerdings
experimentell gelungen, bei Rindern eine Mastitis mit Laktose-negativen B-Streptokokken
des humanen Biotyps auszulösen (JENSEN 1982).
2.2.1.2.2 Streptococcus dysgalactiae S. dysgalactiae wird der Lancefield-Gruppe C zugeordnet und gilt als bedeutende Ursache
von Infektionen bei Mensch und Tier. Auch dieser Erreger zählt zu den Äskulin-negativen
Streptokokken, lässt sich jedoch durch seine negative CAMP-Reaktion eindeutig von S.
agalactiae unterscheiden. Der Keim kommt hauptsächlich beim Rind und Schwein vor.
Außer in der Milchdrüse sind die Erreger vor allem in den Tonsillen (DALEEL u. FROST
1967), in Hautverletzungen, der gesunden sowie entzündeten Gebärmutter (HEJLICEK 1994)
und im Urogenitaltrakt zu finden, selten jedoch auf der gesunden Zitzenhaut (PHILPOT

19
1975). Diese Quellen spielen eine wesentliche Rolle bei der Entstehung einzelner
Mastitisfälle innerhalb einer Herde.
Der Anteil dieses Erregers am Mastitisgeschehen wird zwischen 5,1 und 20% angegeben
(HEJLICEK 1994). Sowohl die akute klinische, als auch die subklinische Form verlaufen als
katarrhalische Galaktophoritis und Mastitis. EDWARDS und SMITH (1966) fanden in einer
detaillierten Studie unter Einbeziehung von drei Herden, dass S. dysgalactiae für 7,4% aller
klinischen Mastitisfälle verantwortlich war. Sie zeigten außerdem, dass 90% der S.
dysgalactiae-Infektionen des Euters ein klinisches Erkrankungsbild hatten und nur 10% der
Infektionen subklinisch verliefen. Meist beobachtet man bei einer S. dysgalactiae-Infektion
eine starke Erhöhung der Zahl somatischer Zellen. Da nur vereinzelt Mastitisfälle im Bestand
auftreten, wird dem Erreger im Gegensatz zu S. agalactiae keine hohe Kontagiosität
zugesprochen. Die kontaminierte Milch scheint dennoch auch hier der häufigste
Übertragungsvektor zu sein. Der Erreger besitzt die Fähigkeit, für 5-20 Monate in der einmal
infizierten Milchdrüse zu überleben. Diese persistierenden Infektionen konnten von WANG
et al. (1999) mit Hilfe der Makrorestriktionsanalyse und der Pulsfeldgelelektrophorese
nachgewiesen werden. Die meisten Infektionen entstehen galaktogen in der Trockenstehphase
und der frühen Laktation (SMITH et al. 1985; SMITH u. HOGAN 1993).
WANG et al. (1999) fanden bei ihren Untersuchungen von 52 Isolaten aus fünf Betrieben
mittels Pulsfeldgelelektrophorese sechs verschiedene Typen, wobei meist ein oder zwei pro
Bestand dominierten. Ein Typ konnte in drei der fünf Betriebe isoliert werden. Dies zeigt,
dass trotz der weit verbreiteten Annahme, dass es sich bei diesem Erreger um einen
Umweltkeim handelt, die Übertragung von Kuh zu Kuh während der Melkzeit durchaus auch
wahrscheinlich ist. Übereinstimmende Aussagen findet man bei SMITH und HOGAN (1993)
und BASSEGIO et al. (1997). Zusammenfassend ist S. dysgalactiae sowohl als kuhassoziiert
gleichermaßen auch als umweltassoziiert einzuordnen (DVG 2002).
2.2.1.3 Corynebacterium (C.) bovis Corynebakterien sind in der Natur weit verbreitet. Man findet sie im Erdboden, im Wasser
und auf der Haut von Tieren.
Corynebacterium (C). bovis kommt ausschließlich bei Rindern vor und wird sehr häufig im
Zitzenkanal gesunder Tiere nachgewiesen. Da der Erreger die Eigenschaft hat, Fett zu
hydrolysieren, wurde er zunächst Bacterium lipolyticum genannt. Corynebacterium spp.
zählen aufgrund ihres epidemiologischen Verhaltens zu den wenig kontagiösen
Mastitiserregern. Trotz ihrer häufigen Isolierung aus Milchproben ist ihre Bedeutung als

20
Mastitiserreger umstritten. Seit Jahrzehnten wurde C. bovis als Saprophyt angesehen. So
isolierte BOURLAND (1967) den Keim aus 90% der antiseptisch entnommenen Milchproben
einer Kuhherde, ohne dass Anzeichen einer Mastitis vorlagen. Bei den von WILSON und
RICHARDS (1980) durchgeführten Untersuchungen konnte in 70% der Milchproben ein
positiver Befund für C. bovis erbracht werden. BLACK et al. (1972) konnten durch Punktion
der Zitzenzisterne zeigen, dass gewisse C. bovis-Besiedlungen auf den Strichkanal beschränkt
sind, und die Erreger während der Probenentnahme aus dem Strichkanal in die Milchprobe
gespült werden.
COBB und WALLEY (1962) widersprachen erstmals der These, dass C. bovis nur als
Saprophyt im Milchsekret vorkommt. So nahmen sie intramammäre Inokulationen von 2ml
einer C. bovis-Serumbouillonkultur mit und ohne Inaktivierung durch Erhitzung vor. Die
Kühe zeigten nach Inokulation der nicht erhitzten Kultur teilweise geringe
Temperaturerhöhungen oder Flockenbeimengungen in der Milch. Dies wurde als Zeichen
einer Bedeutung des Erregers als Mastitis-Verursacher gewertet.
Auch RIZK und JANETSCHKE (1972) erwähnten, dass C. bovis unter bestimmten
begünstigenden Bedingungen, wie z.B. schlechter Melkhygiene eine Mastitis beim Rind
auslösen kann. Der Keim breitet sich vor allem dann in Beständen aus, wenn keine
Zitzendesinfektion nach dem Melken durchgeführt wird (BLOBEL 1980).
SELBITZ (1992) und ROLLE und MAYR (1993) bringen den Erreger mit milden klinischen
Mastitiden in Zusammenhang, und WATTS und ROSBACH (2000) beobachteten eine
verminderte Milchleistung beim Vorliegen von C. bovis-Infektionen. Auch BRAMLEY
(1975) beschreibt, dass der Erreger selten eine augenfällige Leukozytose auslöst, und
lediglich eine geringfügige Erhöhung des Zellgehaltes im Vergleich mit nicht C. bovis-
infizierten Nachbarvierteln nachweisbar ist.
Generell wird C. bovis heute zu den minorpathogenen Mastitiserregern gezählt, da er zwar zur
Infektion der Milchdrüse befähigt ist und auch Entzündungen auslösen kann, die aber meist
subklinisch verlaufen und im Bezug auf den Anstieg des somatischen Zellgehaltes eher milde
verlaufen (RÖDER 1985). SOBIRAJ et al. (1997) stufen den Erreger als nicht pathogen ein.
Weitere Diskussionen werden über einen schützenden Effekt von C. bovis vor einer Infektion
mit majorpathogenen Keimen geführt. So konnte gezeigt werden, dass eine Besiedelung mit
C. bovis vor Infektionen mit S. aureus und anderen kontagiösen Mastitiserregern schützt
(MATTHEWS et al. 1991). Die Inzidenz von C. bovis kann nach BRAMLEY (1975) durch
Zitzentauchen und gezieltes antibiotisches Trockenstellen aller Tiere deutlich gesenkt werden.

21
2.2.2 Umweltassoziierte Mastitiserreger
2.2.2.1 Koagulase-negative Staphylokokken (KNS) KNS sind Erreger, die in der Umgebung der Tiere häufig vorkommen. Derzeit werden 28
Spezies des Genus Staphylococcus unterschieden, von denen 25 zu den KNS gezählt werden.
Im Euter des Rindes lassen sich nach den Ergebnissen verschiedener Untersuchungen vor
allem S. xylosus, S. chromogenes, S. epidermidis, S. sciuri, S. haemolyticus, S. hyicus, S.
simulans, und S. cohnii nachweisen (DEVRIESE 1977; HARMON u. LANGLOIS 1989;
TODHUNTER et al. 1993). Nach KUDINHA und SIMANGO (2002) sind die KNS normale
Besiedler der gesunden Euterhaut und stellen somit eine ständige Quelle für die Kolonisation
des Zitzenendes und des Strickkanals dar, weswegen sie gleichzeitig auch zu den
kuhassoziierten Bakterien gerechnet werden. Die Autoren konnten in ihrer Studie zeigen, dass
die KNS heute zu den Hauptursachen boviner Mastitiden gehören.
MATOS et al. (1991) konnten S. xylosus neben den übrigen Staphylokokkenspezies als
dominierende Staphylokokkenart in 90% der von ihnen untersuchten Einstreuproben
nachweisen. KUDINHA und SIMANGO (2002) konnten S. chromogenes, S. epidermidis und
S. hominis am häufigsten aus Milchproben isolieren und stellen die Hypothese auf, dass es
sich bei diesen drei Erregern um die wichtigsten Mastitiserreger aus der Gruppe der KNS
handelt. S. epidermidis und S. chromogenes werden häufig auf der menschlichen Haut
gefunden, so dass eine Übertragung vom Menschen auf das Tier während des Melkens
möglich scheint.
Wegen ihrer geringen Pathogenität werden die KNS zu den minorpathogenen
Mastitiserregern gerechnet. So werden sie häufig mit einer milden Entzündungsform des
Euters und einer leichten Leukozytose ohne Auftreten klinischer Erscheinungen
nachgewiesen (TODHUNTER et al. 1993; SEFFNER u. BERGMANN 1994, KUDINHA u.
SIMANGO 2002). DAVIDSON et al. (1992) vertreten die Auffassung, dass der Nachweis
von KNS auf Einzeltierebene einen relativ unwichtigen Befund darstellt. Liegt jedoch eine
hohe Herdenprävalenz vor, kann ein signifikanter Anstieg der Tankmilchzellzahlen ausgelöst
werden. SOBIRAJ et al. (1997) sehen die Erreger eher als nicht pathogen an.
Der Anteil der KNS an Mastitisfällen, vor allem bei Färsen, hat im letzten Jahrzehnt jedoch
stark zugenommen (MYLLYS et al. 1998). Studien von JONES et al. (1982), TIMMS und
SCHULTZ (1987) sowie BENITES et al. (2002) zeigen außerdem, dass die KNS in
bestimmten Herden die am häufigsten isolierte Erregergruppe darstellten. KUDINHA und
SIMANGO (2002) zeigen, dass KNS häufig eine entzündliche Antwort des Euters
hervorrufen, die sich bei 95% der von ihnen isolierten KNS-Kulturen in erhöhten Zellgehalten

22
äußerte. Die Autoren vermuten, dass die Erreger wahrscheinlich eine höhere Pathogenität
besitzen, als bisher angenommen wurde. So beschrieben STABENFELDT und SPENCER
(1965) die histologischen Veränderungen von 13 mit KNS infizierten Eutern. Sie konnten
überwiegend chronisch entzündliche Veränderungen mit starker Infiltration mit neutrophilen
Granulozyten nachweisen.
Andererseits macht eine Infektion mit S. epidermidis das Euter weniger empfindlich für
nachfolgende Infektionen mit S. agalactiae oder E. coli (BRAMLEY 1978). Dieses Ergebnis
bestätigt die Hypothese, dass gering euterpathogene Infektionserreger, die den Zitzenkanal
und die Oberfläche der Zitzen besiedeln, die Gefahr einer Infektion durch majorpathogene
Mastitiserreger mindern können. Nach BRAMLEY (1975) ist diese günstige Wirkung jedoch
eher durch eine verstärkte Leukozytose bedingt und nicht einer direkten Hemmung durch
diese Keimart zuzuschreiben. Ferner wäre die Inhibition schon durch die Anwesenheit der
KNS im Strichkanal denkbar. DAVIDSON et al. (1992) konnten im Gegensatz dazu nicht
bestätigen, dass eine Infektion des Euters mit KNS das Viertel vor einer Superinfektion mit S.
aureus schützt. Die Untersuchungsergebnisse ergaben keinen statistisch signifikanten
Unterschied zwischen einer Untersuchungsgruppe mit KNS infizierten Eutervierteln und einer
Gruppe, deren Viertel nicht infiziert waren.
In neuer Zeit werden KNS immer öfter als wichtige Mastitiserreger, die Ursache von
ökonomischen Verlusten sind, angesprochen (TIMMS u. SCHULTZ 1987; DAVIDSON et al.
1992). Obwohl geographische Unterschiede auftreten, scheinen weltweit S. hyicus ssp.
chromogenes und S. simulans sowie S. epidermidis und S. xylosus die am häufigsten aus
bovinen Milchproben zu isolierenden Spezies zu sein (BIRGERSSON et al. 1992;
AARESTRUP et al. 1995).
S. hyicus ssp. chromogenes scheint hierbei der am stärksten pathogene Vertreter der Gruppe
der KNS zu sein und kann ernste Mastitiden hervorrufen (DEVRIESE et al. 2002). ZHANG
und MADDOX (2000) fanden bei ihren Studien keine statistisch signifikanten Unterschiede
in den Entzündungsparametern zwischen Euterinfektionen durch S. aureus bzw. S. hyicus ssp.
chromogenes. KUDINHA und SIMANGO (2002) zeigten jedoch, dass S. hyicus ssp.
chromogenes und S. epidermidis wesentlich häufiger bei subklinischen als bei klinischen
Mastitiden isoliert werden konnten. S. hominis und die übrigen KNS-Spezies konnten sie im
Gegensatz dazu fast ausschließlich in Fällen mit subklinischer Mastitis isolieren. Nach
AARESTRUP et al. (1995) werden vor allem bei S. xylosus-Infektionen hohe Zellgehalte
nachgewiesen. Nach DAVIDSON et al. (1992) steigt das Infektionsrisiko im Laufe der
Laktation an. So lag die Prävalenz für Euterinfektionen durch KNS in den ersten fünf

23
Laktationsmonaten zwischen 4,8 und 6,4%, während sie in den letzten vier Monaten der
Laktation 14,2-16,6% betrug.
MOELLER et al. (1999) konnten einige Pathogenitätsunterschiede zwischen den
verschiedenen KNS-Spezies feststellen. So konnte S. simulans weit häufiger aus klinischen
Mastitiden isoliert werden als die übrigen Spezies.
KNS-Infektionen neigen zur Chronizität. So ermittelten TODHUNTER et al. (1993) eine
durchschnittliche Infektionsdauer von 222 Tagen. Die Autoren zeigten, dass die Erkrankung
besonders häufig bei Erstkalbinnen sowie bei älteren Kühen nach der Trockenstehphase
auftrat. Die Prävalenz dieser Erreger kann durch die Verwendung eines Dippmittels nach dem
Melken gesenkt werden (HARMON et al. 1986).
2.2.2.2 Äskulin-positive Streptokokken
2.2.2.2.1 Streptococcus uberis S. uberis ist ein klassischer Mastitiserreger bei Rind, Schaf, Büffel, Schwein und Katze.
Etwa 15-50% der Stämme reagieren mit dem Streptokokken-E-Serum. Diese Spezies besitzt
gemeinsam mit den übrigen umweltassoziierten Streptokokken und Enterokokken die
Fähigkeit, Äskulin abzubauen. Daher zählen sie zur Gruppe der Äskulin-positiven
Streptokokken. S. uberis löst weltweit in einem hohen Prozentsatz katharralisch-klinische
Mastitiden aus (PHUEKTES et al. 2001). Als Krankheitserreger wird S. uberis außer bei
Mastitiden auch bei Endometritiden einschließlich Aborten sowie bei Endokarditiden
nachgewiesen. Nach SMITS et al. (1998) ist S. uberis für 12-14% der klinischen Mastitiden
verantwortlich, wird aber auch häufig bei subklinischen Mastitiden isoliert. KING (1981) gibt
an, dass es insbesondere bei fehlender Behandlung durch eine lange Erkrankungsperiode zu
großen Milchmengenverlusten kommen kann. EDWARDS und SMITH (1966) berichteten,
dass S. uberis für 7% der klinischen Mastitisfälle verantwortlich war, und dass in ihrem
Untersuchungsgut 50% der S. uberis-Infektionen mit klinischen Erscheinungen einhergingen.
BLOBEL (1980) hingegen beschreibt den Erreger als wenig Milchdrüsen-pathogen. Auch
KING (1981) beschreibt eine überwiegend milde Verlaufsform, die nur mit leichten
Sekretveränderungen einhergeht. Aber auch eine Verlaufsform, die bis zur Zerstörung des
betroffenen Euterviertels führen könne, sei möglich. Nach HEJLICEK (1994) sind für die
klinische Ausprägung prädisponierende Einflüsse notwendig. SOBIRAJ et al. (1988) fanden
neben mild verlaufenden Galaktophoritiden bei einem Drittel der mit S. uberis infizierten
Euterviertel akute Mastitiden der katharrhalischen und phlegmonösen Form bei

24
Erstkalbinnen. MILNE et al. (2002) untersuchten 2257 Milchproben klinischer Mastitiden
und konnten S. uberis als häufigsten Erreger in 37% der Proben nachweisen.
Die Verbreitung von S. uberis-Infektionen des Euters variiert stark in den verschiedenen
Regionen. KING (1981) gibt die durchschnittliche S. uberis-Inzidenz mit 4% der Kühe bzw.
1,5% der Viertel an, erwähnt jedoch, dass unter besonderen Umständen in bestimmten Herden
S. uberis der Leitkeim für Mastitiden werden könne. Hierfür kämen insbesondere häufige
antibiotische Euterbehandlungen und verschiedene umweltassoziierte Faktoren als Auslöser
in Betracht.
WANG et al. (1999) geben an, dass S. uberis mit 60-80% heute die am häufigsten
nachgewiesene Streptokokkenart in Kuhmilch ist. Von besonderem epidemiologischem
Interesse ist das Vorkommen dieses Erregers im Gegensatz zu den anderen Mastitis
auslösenden Streptokokken auch außerhalb der Milchdrüse des Rindes (KING 1981;
LÄMMLER u. HAHN 1994). Der Erreger kann von der Bauchwand, den Lippen, Tonsillen,
der Euterhaut, dem Urogenitaltrakt, aus dem Pansen sowie dem Kot und der Einstreu isoliert
werden (BEERWERTH u. KÖSER 1965; CULLEN 1966; WINKENWERDER 1966;
DALEEL u. FROST 1967; SCHALM et al. 1971; WENDT 1994). Eine wesentliche
Bedeutung für die Verbreitung scheint hierbei dem Rinderkot zuzukommen. So konnte aus
Bodenproben einer Weide, die von einer mit S. uberis infizierten Herde begangen worden
war, der Erreger mehrfach isoliert werden (CULLEN 1969). HUGHES (1960) und CULLEN
(1966) hingegen sahen als Hauptreservoir vor allem die Lippenhaut der Tiere an. Der Erreger
kann durch Belecken andere Körperstellen kontaminieren.
HEJLICEK (1994) gibt an, dass in Herden mit S. uberis-Euterinfektionen der Erreger aus
51,6% der Haut- und 85,8% der Umweltproben isoliert werden konnte. BRAMLEY et al.
(1978) konnten den Erreger in hohem Maße aus Einstreumaterial isolieren. Hier scheint sogar
eine Erregervermehrung möglich. Die Vorkommenshäufigkeit von S. uberis in der nächsten
Umgebung der Milchdrüse steht allerdings in keinem direkten Verhältnis zum
Infektionsrisiko. So folgt einer Besiedelung der Zitzenspitze nicht automatisch eine Infektion
des Drüsengewebes. KING (1981) vermutet, dass bei einem Nachweis von S. uberis
außerhalb des Euters eher von einer Kontamination als von einer Kolonisierung oder gar
pathogenen Aktivität auszugehen ist.
Ähnlich wie bei S. dysgalactiae haben auch die S. uberis-Mastitiden keinen seuchenhaften
Charakter. Mittels der Pulsfeldgelelektrophorese konnten WANG et al. (1999) nachweisen,
dass die Isolate innerhalb einer Herde genetisch sehr heterogen sind, da nur sehr selten
verschiedene Kühe Erreger mit identischen Makrorestriktionsmustern im Euter beherbergten.

25
Das zeigt, dass die Transmission von S. uberis von Kuh zu Kuh während des Melkaktes
unwahrscheinlich ist. Die Autoren konnten durch den Nachweis von bis zu fünf Monate lang
persistierenden S. uberis-Infektionen bei 18 von 20 Tieren ferner Hinweise für den
chronischen Charakter der Infektionen erbringen. Bei acht dieser 20 Kühe wurden die
Infektionen durch mehrere Makrorestriktionstypen hervorgerufen. HEJLICEK (1994)
beschreibt chronische S. uberis-Infektionen von 5-20, in seltenen Fällen mit über 20 Monaten
Dauer.
Aufgrund dieser Befunde erscheint eine Verbreitung des Erregers während der Melkzeit
unwahrscheinlich. Vielmehr wird angenommen, dass aufgrund seines ubiquitären
Vorkommens als Umweltkontaminante S. uberis in der Zwischenmelkzeit die Zitzenkuppen
kontaminiert und dann über den Strichkanal in die Milchdrüse eindringt. Er wird bei älteren
Kühen häufig in der Trockenstehperiode isoliert und scheint auch an den klinischen
Mastitiden in der Frühlaktation beteiligt zu sein (COUSINS et al. 1980; SMITH u. HOGAN
1993). OLIVER et al. (1956) konnten zeigen, dass der Erreger bei Kühen im ersten
Laktationsmonat zu 11% nachgewiesen wurde, diese Nachweisrate fällt bis zum neunten
Laktationsmonat auf 1,8%. Nach PHILPOT (1975) ist S. uberis der am häufigsten
nachgewiesene Mastitiserreger in der frühen Trockenstehperiode. NEAVE et al. (1950)
fanden, dass es insbesondere in den ersten drei Wochen des Trockenstehens zu
Neuinfektionen kommt. Auch HEJLICEK (1994) gibt die Neuinfektionsrate in der Zeit des
Trockenstehens mit 40% an und begründet dies damit, dass in diesem Zeitraum der
ausspülende Milchfluss fehle. LACY-HULBERT und HILLERTON (1994) beschreiben eine
signifikant höhere S. uberis-Infektionsrate bei Kühen mit kürzerer Länge des Strichkanals
sowie mit einem aus dem Strichkanal entfernbaren Keratingewicht von weniger als 1,8 mg.
Ferner tritt die S. uberis-Infektion vermehrt bei Erstkalbinnen auf (SOBIRAJ et al. 1988).
Bei der Behandlung werden gute Ergebnisse (87-95% iger Erfolg) mit Penicillin G und
halbsynthetischen Penicillin-Präparaten in der Trockenstehphase erzielt (HEJLICEK 1994).
Um das Überleben und die Vermehrung von S. uberis an Orten außerhalb des Euters
einzuschränken, wird den Milchviehhaltern verstärkte Sauberkeit im Fress-, Lauf- und vor
allem im Liegebereich empfohlen (WANG et al. 1999).
Neben S. uberis werden häufiger auch S. parauberis, S. bovis, S. mitis, S. salivarius, S.
saccarolyticus und Aerococcus viridans aus Milch Mastitis-kranker Kühe isoliert. Diese
werden zusammen hinsichtlich ihres biochemischen Verhaltens bei der Spaltung von Äskulin
als Äskulin-positive Streptokokken angesprochen. Aerococcus viridans wird beim Menschen

26
vor allem aus der Mundhöhle, dem Rachen, aus Zahnplaques, dem Blut, dem
Respirationstrakt, dem Intestinum und aus dem weiblichen Genitale isoliert. Beim Rind wird
er häufig aus der Milch, aus den Tonsillen und aus dem Genitaltrakt ohne Vorliegen von
Krankheitserscheinungen nachgewiesen.
2.2.2.2.2 Enterococcus spp. Enterokokken bilden innerhalb der Gattung Streptococcus eine Gruppe, die sich durch ihr
ubiquitäres Vorkommen, ihre auffallenden biologischen Eigenschaften und ihren
abweichenden Zellaufbau von den übrigen Streptokokken unterscheiden. Sie sind fakultativ
pathogen und werden als Verursacher von Lebensmittelinfektionen beim Menschen diskutiert.
Durch ihre weite Verbreitung und ihr massives Auftreten im Kot existieren breite
Infektionsmöglichkeiten, und Interpretationen bezüglich der ätiologischen Bedeutung bei
Infektionen werden erschwert. Obwohl Enterokokken sehr häufig aus Milch isoliert werden
konnten, lässt dies keine zuverlässige Aussage über ihre tatsächliche originäre Bedeutung in
der Milchdrüse zu, da oftmals eine postsekretorische Kontamination vorzuliegen scheint
(BLOBEL 1980). Enterokokken sind Kommensalen und Saprophyten des Magendarmtraktes
von Mensch und Tier und treten als fäkal kontaminierende Mikroflora in der Stallumwelt auf.
Aus der direkten Umgebung der Tiere können sie dennoch in die Milchdrüse des Rindes
eindringen und subklinische, gelegentlich auch klinische Mastitiden auslösen. In der
Milchdrüse können sie für längere Zeit persistieren.
Am häufigsten werden E. faecium, E. faecalis und E. avium aus Milchproben isoliert.
Enterokokken mit dem Lancefield-Gruppenantigen D verursachen beim Rind nur vereinzelt
Mastitiden. So beträgt nach HEJLICEK (1994) ihr Anteil an den Mastitisstreptokokken in
Frankreich 10%, in Österreich jedoch 56,9% (hiervon 5% S. bovis).
Von besonderer Bedeutung ist ihre natürliche Resistenz gegen die allgemein bei der
Mastitistherapie verwendeten Chemotherapeutika.
Umstritten ist bis heute die Rolle der Enterokokken bei den so genannten „unspezifischen“
Lebensmittelintoxikationen. Vergiftungen, bei denen aus den inkriminierten Lebensmitteln
hohe Keimzahlen an Enterokokken isoliert werden konnten, wurden häufig beschrieben.
Überwiegend wurden E. faecalis und E. faecium nachgewiesen. Die Inkubationszeit betrug 2-
36 Stunden, die Hauptsymptome waren Übelkeit, Erbrechen, Leibschmerzen und Durchfall.
In fast allen Fällen konnten aus den Proben aber auch andere Bakterien isoliert werden.
Tierexperimente und Versuche an menschlichen Freiwilligen haben unterschiedliche
Resultate und kaum Hinweise auf eine gesundheitsschädliche Eigenschaft der Enterokokken

27
ergeben. Man nimmt an, dass sowohl die Erregeranzahl, als auch die Beschaffenheit des
Substrates sowie das Wachstumsstadium des Erregers eine Bedeutung haben. Erwähnenswert
ist auch die Eigenschaft der Enterokokken zur Bildung von Aminosäure-Decarboxylase und
damit die Förderung der Entstehung von biogenen Aminen (FEHLHABER 2004).
In Rohmilch liegt der prozentuale Anteil von Enterokokken meist unter 5% (HAHN 1984).
Der Autor äußert die Meinung, dass die Bedeutung der Enterokokken als
Lebensmittelvergifter im Vergleich zu anderen mikrobiellen Ursachen sehr gering ist und
insgesamt keine nennenswerte Gefahr darstellt.
2.2.2.3 Enterobacteriaceae (Coliforme) Die Enterobacteriaceae sind als Bestandteil der natürlichen Darmflora von Mensch und
Säugetieren klassische Umwelterreger. Die im klinischen Verlauf etwa gleichartigen durch
Enterobacteriaceae verursachten Mastitiden werden überwiegend durch Escherichia (E.) coli
hervorgerufen, weswegen sie häufig auch als Coli-Mastitis bezeichnet werden. Mit größerem
Abstand folgen Klebsiella pneumoniae, Enterobacter agglomerans, Enterobacter cloacae,
Citrobacter diversus, Serratia marcescens, Serratia odorifera, Proteus spp. und Salmonella
spp., die in der Routinediagnostik oftmals nicht weiter differenziert und als coliforme Keime
zusammengefasst werden. E. coli besitzt eine Reihe von Virulenzfaktoren wie z.B.
Endotoxine, Adhäsionsfaktoren, Enterotoxine und Zytokine und ist an Krankheitsprozessen
wie Enteritiden, Septikämien und Arthritiden unterschiedlicher Tierspezies ursächlich
beteiligt.
Der Anteil von E. coli an der gesamten Darmflora beträgt im Zustand der Eubiose nicht mehr
als 1% (ROLLE u. MAYR 2002). Neben ihrer Beteiligung an den Verdauungsprozessen
haben die coliformen Keime mit der Produktion von Thiamin, Folsäure sowie Vitamin C und
K wichtige physiologische Funktionen. Die weite Verbreitung der Bakterienart in der Umwelt
steht in engem Zusammenhang mit der ständigen Ausscheidung über den Kot (SELBITZ
1992). Sowohl die Verteilung der Serovare, als auch das Resistenzverhalten berechtigen zu
der Annahme, dass der Darmkanal des Rindes Reservoir für Mastitis auslösende Stämme ist
(BERGMANN 1994). So dürfte für die Coli-Mastitis kontaminierte Einstreu die Quelle für
eine Euterbesiedelung und Infektion sein. Klebsiella und Enterobacter ssp. sind unabhängig
von einer fäkalen Kontamination in vielen Umweltmaterialien vorhanden. Nach
BERGMANN (1994) sind außerdem prädisponiernde Faktoren wie z.B. resistenzmindernde
Stresseinwirkungen und hohe Erregerkonzentration für die Infektion erforderlich. Der Grund
für die wachsende Bedeutung der Coli-Mastitis sind ihre zunehmende Inzidenz und die

28
ernsten Symptome der Erkrankung. Sie tritt überwiegend als perakut bis akut und bösartig
verlaufende, seltener chronische oder subklinische Euterentzündung vor allem peripartal bei
Einzeltieren auf. Die Erkrankung mehrer Kühe einer Herde zum gleichen Zeitpunkt kommt so
gut wie nicht vor. Eine Übertragung von Kuh zu Kuh wird daher nahezu ausgeschlossen. Im
Stall gehaltene Tiere erkranken weitaus häufiger als auf der Weide gehaltene. Überwiegend
ist nur ein Viertel, häufig ein Hinterviertel, betroffen. Eine Zunahme der Erkrankungen in den
letzten Jahren wird mit intensivierter Milcherzeugung unter den Bedingungen der Stallhaltung
in Verbindung gebracht. So kann ihr Anteil in Problembeständen 25% des
Mastitiserregerspektrums betragen (BERGMANN 1994).
Der Anteil der durch coliforme Keime ausgelösten Mastitisfälle variiert sehr zwischen
verschiedenen Regionen. So liegt dieser in Finnland unter 20%, während in Israel mehr als
60% der Mastitisfälle durch coliforme Keime bedingt sind (KAIPAINEN et al. 2002). Nach
HOGAN et al. (1989) und SCHUKKEN et al. (1990) ist E. coli der am meisten isolierte
Erreger bei klinischen Mastitisfällen in Herden mit einem niedrigen Tankmilchzellgehalt.
Beim perakuten Verlauf treten durch die Bakterien- und deren Endotoxinwirkung neben den
lokalen entzündlichen Veränderungen auch starke Allgemeinsymptome auf. Es kommt zu
hohem Fieber mit stark erhöhter Pulsfrequenz, Anorexie, Muskelzittern und Festliegen. Das
betroffene Viertel ist hochgradig geschwollen, vermehrt warm, schmerzhaft sowie von harter
Konsistenz. Das mengenmäßig verminderte Sekret ist klar, enthält kleine Fibrin- oder
Eiterflocken. Gehäuft tritt diese Form der Mastitis kurz nach dem Abkalben auf. Die
Krankheit endet ohne Therapie häufig letal oder mit völligem Funktionsverlust des
betroffenen Viertels. Hierfür verantwortlich ist insbesondere das schnelle Auftreten von
massiven Gewebsveränderungen in der Milchdrüse. So lassen sich bereits eine Stunde nach
einer experimentellen Infektion erste Nekrosen der Epithelzellen der Zitzen und Milchgänge
nachweisen. Bereits zwei Stunden post infectionem weisen entnommene Milchproben die
typischen Mastitis-bedingten Sekretveränderungen auf, und erste Allgemeinsymptome treten
in Erscheinung. Die Symptomatik einer akuten Mastitis ist etwa fünf Stunden post
infectionem voll ausgebildet (BERGMANN 1994).
Der galaktogene Infektionsweg gilt als der häufigste, jedoch wird auch der hämatogene aus
Darminfektionen und puerperalen Störungen diskutiert (ROLLE u. MAYR 2002). WEIGT
und DREIST (1988) hingegen sehen in jeweils 30% der Coli-Mastitiden Geburtsstörungen,
andere Organ- und Allgemeinerkrankungen sowie Euter- und Zitzenverletzungen als
Auslöser, während sie den galaktogenen Infektionsweg nur in 10% der Fälle und meist nach
melkhygienischen Mängeln als Ursache ansehen.

29
Coli-Mastitiden häufen sich zum Zeitpunkt der Geburt bis zu drei Monate post partum
(BERGMANN 1994). Durch die rasche Vermehrung der coliformen Keime und die sofort
einsetzende Endotoxinproduktion kommt es insbesondere durch die toxinbedingte Freisetzung
von Histamin und Prostaglandinen zu einer Schädigung der Gefäßmembranen und damit der
Mikrozirkulation. Es bilden sich Gefäßthromben und in der Folge Mangeldurchblutung mit
Nekrose größerer Euterabschnitte mit dem Ergebnis einer hämorrhagisch-nekrotisierenden
Mastitis.
LAM et al. (1996) konnten in ihren Untersuchungen 3-18 verschiedene E. coli-Genotypen in
unterschiedlichen Herden isolieren. Verschiedene Viertel einer Kuh waren dabei meist durch
verschiedene Genotypen infiziert worden. Dies spricht für die umweltassoziierte Übertragung
der Erreger. Eine Übertragung von Viertel zu Viertel wird als unwahrscheinlich angesehen.
Wenn allerdings mehr als eine klinische E. coli-Mastitis pro Kuh/Viertel und Laktation
auftrat, konnte meist derselbe Genotyp isoliert werden. Die Autoren äußern daher die
Vermutung, dass auch eine chronische subklinische Coli-Mastitis vorkommen kann.
2.3 Pulsfeldgelelektrophorese (PFGE)
Im Laufe der vergangenen Jahre wurden neue molekularbiologische Typisierungssysteme zur
Klärung epidemiologischer Fragestellungen herangezogen. Theoretisch wird für jedes System
vorausgesetzt, dass epidemiologisch verwandte Bakterienisolate, die im Rahmen der klonalen
Expansion aus einem gemeinsamen Vorläuferstamm hervorgegangen sind, charakteristische
Eigenschaften teilen, welche sie von anderen, epidemiologisch nicht verwandten Isolaten
unterscheidet. Die Verwendungsmöglichkeit dieser speziellen Eigenschaften hängt dabei von
der Stabilität der Eigenschaften innerhalb eines Stammes und der Diversität innerhalb einer
Spezies ab. Die Diversität reflektiert die evolutionäre genetische Divergenz, die durch
zufällige, nicht letale Mutationen entsteht, welche im Laufe der Zeit akkumulieren. Solche
Mutationen sind dann nachweisbar, wenn sie an Stellen auftreten, für die mit den heutigen
Methoden Detektionsmöglichkeiten bestehen (MASLOW et al. 1993, SCHMITZ et al. 1995).
Durch Digestion der chromosomalen DNA mit selten schneidenden Restriktionsenzymen und
anschließender PFGE können z.B. Mutationen an den spezifischen Schnittstellen
nachgewiesen werden. Diese Schnittstellen dienen hierbei als selektionsneutrale Marker, die
im Verlauf der Evolution des Bakterienchromosoms über Rearrangements oder Mutationen an
einer Stelle verloren gehen und an anderer Stelle neu entstehen können.
Laut GOERING (1998) eignen sich für GC-reiche Sequenzen insbesondere Enzyme, die
Hexanukleotide aus A und T erkennen, und analog für AT-reiche Genome Enzyme, die eine

30
hexamere Sequenz aus G oder C erkennen. So schneidet die in vorliegender Arbeit
verwendete Endonuclease SmaI z.B. CCC-GGG. Idealerweise sollte nach Schneidung aus der
Gesamt-DNA eine überschaubare Anzahl von 7-20 Fragmente entstehen.
Bei der konventionellen Gelelektrophorese mit einem konstanten elektrischen Feld wandern
die durch ihr Phosphatgruppengerüst negativ geladenen DNA-Moleküle in Richtung auf die
Anode (SCHMITZ et al. 1995). Mit dieser Methode können Moleküle bis zu einer Größe von
50 Kilobasenpaaren (Kb) aufgetrennt werden. Die Moleküle wandern hierbei umso
langsamer, je größer sie sind (GARDINER 1991). Größere Moleküle können nicht horizontal
durch die Poren des Gels wandern, da die Porengröße des Gels hierfür zu klein ist. Deswegen
richten die Moleküle ihre Längsachse parallel zu den elektrischen Feldlinien aus und wandern
somit in Schlangenlinien durch die Poren des Gels. Direkte Folge ist, dass alle diese Moleküle
in einer einzigen Bande wandern, und eine Auftrennung und Größenbestimmung nicht mehr
möglich ist (WINTER u. DRIESEL 1988).
SCHWARTZ und CANTOR (1984) gelang die Entwicklung einer neuen Technologie zur
Überwindung dieses physikalischen Phänomens. Bei der PFGE werden mehrere Elektroden in
einem bestimmten Winkel zueinander angeordnet, welche abwechselnd ein elektrisches Feld
erzeugen, wodurch sich die Richtung der Feldlinien in bestimmten Zeitintervallen regelmäßig
verändert. Die DNA-Moleküle müssen daher ihre Ausrichtung und Konformation ständig
erneut ändern, um ihre Längsachse parallel zu den elektrischen Feldlinien ausrichten zu
können (SCHMITZ et al. 1995). Diese Umorientierung nimmt bei größeren Molekülen eine
längere Zeit in Anspruch als bei kleinen Molekülen, so dass die Nettowandergeschwindigkeit
der kleineren Moleküle größer ist (WINTER u. DRIESEL 1988). Folglich verbleibt den
Molekülen mit zunehmender Größe immer weniger Zeit für die Wanderung entlang der
Feldrichtung, wodurch es zur Auftrennung der linearen DNA-Moleküle nach deren
Molekulargewicht kommt. Mit Hilfe dieser Technologie können somit auch Moleküle mit
einer Größenordnung von 50 bis 6000 Kb aufgetrennt werden. Die ständig wechselnde
Richtung des Stromflusses wird über die gesamte Elektrophoresezeit aufrechterhalten, so dass
die DNA einem Zickzackpfad folgt, der in seiner Nettobewegung jedoch dem unteren Ende
des Gels entgegenstrebt.
Um alternierende elektrische Felder erzeugen zu können, wurden zahlreiche Apparaturen
entwickelt, die sich in der Geometrie ihrer Elektrophoresekammer sowie Form, Anzahl und
Stellung der Elektroden unterscheiden. Am häufigsten wird das "contour clamped
homogeneous electric field" (CHEF)-System eingesetzt, welches auch in der vorliegenden
Arbeit verwendet wurde. 24 Elektroden sind hier in einer Elektrophoresekammer hexagonal

31
im Winkel von 120° zueinander angeordnet, die beiden sich gegenüberliegenden Seiten des
Sechsecks werden abwechselnd aktiviert.
Laut GROTHEUS und TÜMMLER (1991) ist die PFGE ein geeignetes Verfahren, um
Infektionsquellen und Übertragungswege von bakteriellen Erregern zu ermitteln und die
Diversität und klonale Struktur von Bakterienkulturen aufzuklären. Nach Meinung von
RÖMLING et al. (1994 u. 1995) stellt die PFGE den Goldstandard für die epidemiologische
Typisierung fast aller infektiöser Mikroorganismen dar. Mit Hilfe des visuellen Vergleichs
der Fragmentlängenmuster kann der Verwandtschaftsgrad der Stämme bestimmt werden.
Nach TENOVER et al. (1995) definiert das Fragmentlängenmuster den Genotyp eines
Bakterienstammes. So zeichnen sich nicht miteinander verwandte Klone durch
unterschiedliche Fragmentlängenmuster aus, während Varianten eines Klons ähnliche oder
identische Muster besitzen. Isolate, die in bis zu drei Bandenpositionen differieren, können
aus einem einzigen genetischen Event entstanden sein und deshalb epidemiologisch eng
verwandte Untertypen eines Stammes darstellen. Liegen mehr als drei Abweichungen im
Bandenmuster vor, so können diese nach TENOVER et al. (1995) miteinander verwandt sein,
wobei der Verwandtschaftsgrad nicht ganz so eng liege.
Bei der PFGE werden die Bakterien in Agarosegelblöckchen eingebettet, die Organismen in
situ lysiert, wodurch das Bakterienchromosom gegen Scherbruch geschützt wird. Es folgt die
Digestion der chromosomalen DNA mit spezifischen Restriktionsenzymen, welche seltene
Schnittstellen haben, wodurch die DNA in eine überschaubare Zahl von Fragmenten
gespalten wird. Die Agaroseblöckchen mit den DNA-Fragmenten werden in die
Aussparungen eines 1%igen Agarosegels eingebettet, und die Fragmente werden in einer
Apparatur, welche die Richtung der elektrischen Strömung ändern kann, ihrer Größe nach in
ein bestimmtes Restriktionsmuster aufgetrennt. Durch den Vergleich dieser spezifischen
Muster kann der Verwandtschaftsgrad der Isolate bestimmt werden (TENOVER et al. 1995).
2.4 Wirtschaftliche Verluste durch Mastitiden Die geschätzten Kosten für die klinische Mastitis variieren von Land zu Land und beinhalten
die Ausgaben für die Behandlungs- und Tierarztkosten, die verminderte Milchleistung
während der verbleibenden Laktation, die wegen antibiotischer Rückstände zu verwerfende
Milchmenge, die frühzeitige Merzung von Tieren mit einem hohen genetischen Wert und
deren Remontierung, die Extraarbeit, die Verminderung der Milchqualität und das erhöhte
Risiko einer rezidivierenden Erkrankung zu einem späteren Zeitpunkt (HERINGSTAD et al.
2000). Die Mastitis ist von größter ökonomischer Bedeutung für die Milchindustrie, da sie

32
eine verminderte Milchqualität und einen großen Produktionsverlust sowie einen erhöhten
Bedarf an Medikamenten und tiermedizinischer Betreuung verursacht (WANG et al. 1999).
Alle Mastitiden haben Einfluss auf die substantielle Zusammensetzung und die
technologische Wertigkeit der Milch (TOLLE et al. 1977). Subklinische Mastitiden treten in
der Regel als Bestandsproblem auf. Der Milchverlust durch subklinische Mastitiden ist in der
Gesamtsumme um ein Vielfaches höher als durch klinische Mastitiden, da diese auf
Herdenbasis sehr viel verbreiteter sind als klinische Fälle (WENDT 1998). Nach ZSCHÖCK
et al. (1998) kommt die subklinische Mastitis 20-50mal häufiger vor als die klinische
Mastitis.
Ihre Bedeutung liegt im wirtschaftlichen Schaden, in erster Linie verursacht durch
Milchminderleistung, durch die eingeschränkte Verwertungsmöglichkeit infolge der
veränderten Milchzusammensetzung, durch erhöhte Arzneimittel- und Tierarztkosten, durch
das Verkehrsverbot für die Milch von mit Antibiotika behandelten Kühen und schließlich
durch frühzeitiges Merzen erkrankter Tiere (KIELWEIN 1994). So sind Eutererkrankungen
neben Fruchtbarkeitsstörungen die häufigste Abgangsursache für Kühe aus einem Bestand.
Nach Angaben von KIELWEIN (1994) ist für 70% der durch subklinische Mastitiden
verursachten Schäden die verminderte Milchproduktion und für 14% die frühzeitige Merzung
betroffener Kühe verantwortlich. Pro erkrankter Kuh entstehen in der BRD pro Jahr
finanzielle Einbußen von 100 Euro (KIELWEIN 1994; ZSCHÖCK et al. 1998).
Weitere Berechnungen ergaben, dass der Milchverlust durch subklinische Mastitiden im
Durchschnitt bei 17,1% pro Viertel und 9,7% pro Tier liegt (TOLLE et al. 1977; BLOBEL
1980). Nach JANZEN (1970) liegt die tägliche Milcheinbuße pro betroffenem Viertel
zwischen 0,76-5,86 Litern.
WENDT (1998) gibt den Verlust bei Milchzellgehalten von über 400.000 Zellen/ml Milch
zwischen 7,5 und 12% an. Die Verluste der Milchleistung nehmen mit steigendem
Milchzellgehalt linear zu. Die angegebenen Verluste entstehen nicht nur während der
Laktation, in der der Zellgehalt erhöht war, sondern ebenfalls abhängig vom Milchzellgehalt
auch in den Folgelaktationen. Nach KIELWEIN (1994) setzt die durch Mastitis bedingte
Leistungsminderung im Bereich um 100.000 Zellen/ml ein und kann bis zu 20% des
erkrankten Viertels betragen. Die technologische Wertigkeit der Milch mastitiskranker Kühe
ist beeinträchtigt durch den vermehrten Gehalt an Chlor- und Natriumionen, eine Reduktion
der fettfreien Trockenmasse, ein vermindertes Koagulierungsvermögen der Kaseine, eine
Erhöhung des Gehaltes an originären Enzymen, eine verminderte Hitzestabilität, eine
unspezifische Hemmung der Starterkulturen und durch die Gefahr des Auftretens von

33
Hemmstoffen in der Milch (KIELWEIN 1994). Nach JANZEN (1970) sinkt der Fett- bzw.
Laktosegehalt der Milch um 0,1-0,45% bzw. 0,1-0,77%. Weiterhin verändert sich der
prozentuale Anteil des Calciums, Magnesiums und Kaliums, wodurch es zu einer
Geschmacksbeeinflussung und zu Verarbeitungsschwierigkeiten in der Molkerei kommt, die
zu Qualitätsmängeln des Endproduktes führen können. Die Veränderungen der
Milchzusammensetzung bedingen auch eine verminderte ernährungsphysiologische
Wertigkeit der Milch. Erhöhte Milchzellgehalte korrelieren mit einer schlechten
Milchqualität, einer reduzierten Qualität der hergestellten Milchprodukte und einer
verringerten Haltbarkeit.
GRAY und SCHALM (1962) zeigten durch einen Vergleich mit CMT-negativen Vierteln,
dass es bei Vierteln mit einem CMT-Ergebnis von 1+, 2+ bzw. 3+ zu einer Verminderung der
Milchmenge um 10, 16 bzw. 24,5% kommt. Auch PHILPOT (1967) untersuchte den Einfluss
der subklinischen Mastitis auf die Milchmenge und kam zu dem Ergebnis, dass die
Milchleistung bei einem CMT-Ergebnis von 1+, 2+ bzw. 3+ um 11,4, 25,6 bzw. 45,5%
reduziert ist.
Nach WILSON et al. (1997) liegen die Verluste pro Fall einer intramammären Infektion
zwischen 130 und 320 US$. HERINGSTAD et al. (2000) geben die Kosten pro klinischem
Mastitisfall in den USA mit 108 bis 122 US$ an. WELLENBERG et al. (2002) schätzen den
weltweiten Verlust durch die Mastitis jährlich auf 35 Milliarden US$. Der finanzielle Verlust
pro Laktation war am höchsten für Pasteurella spp. mit 500,12 US$, Mycoplasma spp. mit
451,63 US$ und S. agalactiae mit 388,19 US$. Die Werte für S. aureus und Streptococcus
spp. lagen mit 185,51 US$ bzw. 143,65 US$ ebenfalls hoch. Für die KNS konnten WILSON
et al. (1997) einen finanziellen Verlust von ungefähr 100 US$ pro Fall ermitteln. Für die
Schätzungen dieser Werte verwendeten die Autoren die ermittelten Unterschiede der 305-
Tage Milchleistung von mit dem jeweiligen Erreger infizierten zu gesunden Tieren.
Die Kontrolle dieser kostenverursachenden Erkrankung muss auf ein kontinuierliches
Präventionsprogramm und die Eliminierung bestehender Infektionen aufbauen. Durch die
Einführung und konsequente Beibehaltung eines solchen Programms kann laut JANZEN
(1970) der durch die Mastitis verursachte finanzielle Verlust um 50% reduziert werden. Ein
spezielles Kontrollprogramm kann den durch Mastitiden verursachten Milchverlust, die
Anzahl der Euterbehandlungen und den ökonomischen jährlichen Verlust pro Kuh signifikant
reduzieren.
Da Gewinnspannen der Milchindustrie direkt von der Eutergesundheit abhängen, haben in
vielen Ländern wie z.B. auch in Deutschland die milchverarbeitenden Organisationen

34
begonnen, schlechte Milchqualität mit Milchgeldabzügen zu bestrafen und für gute
Milchqualität Prämien zu zahlen. Es hat sich gezeigt, dass die Milchviehhalter sehr gut auf
diese Systeme reagieren, und dass die Milchqualität in Ländern mit oben genannten
Vorgehensweisen stetig gesteigert werden konnte. In Finnland konnte der Zellgehalt der
Anlieferungsmilch durch Prämienzahlungen für besonders zellarme Milch auf den niedrigsten
Wert der gesamten EU mit 132.000 Zellen/ml gesenkt werden (HALLEN-SANDGREN
2000).
Auch in Mexiko haben einzelne Molkereien ähnliche Programme eingeführt, jedoch gibt es
bisher noch keinen im offiziellen Normenprogramm Mexikos (Norma Oficial de México =
NOM) verankerten oberen Zellzahlgrenzwert für Anlieferungsmilch. Auch ohne stützende
Gesetzgebung zeichnet sich bei den milchverarbeitenden Betrieben Mexikos der Trend ab,
Anlieferungsmilch mit geringen Zell- und Keimzahlen zu fordern. Das Ziel der Unternehmen
ist es, langfristig Milch mit einem Zellgehalt von weniger als 200.000 Zellen/ml mit einem
Keimgehalt von weniger als 10.000 cfu/ml aufzukaufen (ARREGUÍN 2003).
Momentan regeln in Mexiko die NOMs den Austausch von Produkten innerhalb eines Landes
und zwischen verschiedenen Ländern. Durch die Normen werden Regeln für die Qualität der
Produkte aufgestellt und so der Handel und der Wettbewerb stabilisiert.
Die NOMs liefern obligatorische Regeln, Attribute und Charakteristiken für ein bestimmtes
Produkt, einen Prozess, eine Aktivität, einen Service, eine Produktionsmethode, sowie
Symbole, Marken und Etikettierungen für die fertigen Produkte.
Da Milch und Milchprodukte eine Lebensmittelgruppe großer Wichtigkeit für die
menschliche Ernährung darstellen, wird in den 19 derzeit gültigen und zwischen 1980 und
2003 publizierten mexikanischen Normen festgesetzt, dass die Produkte vor ihrer
Kommerzialisierung und Konsumierung bestimmten hygienischen Behandlungsprozessen
unterzogen werden müssen, damit von den Produkten keinerlei Risiko für den Konsumenten
ausgehen kann. Basierend auf diesen Normen haben die Produzenten die Möglichkeit eine
gerechte Vergütung für das von ihnen erzeugte Produkt zu erhalten (ARREGUÍN 2003).
Die momentan für die mexikanische Milchwirtschaft gültigen Normen sind folgende:
- NOM-155-SCFI-2003
Diese Norm setzt die kommerziellen Namen der verschiedenen Milchtypen, deren
physiochemische Eigenschaften sowie die kommerziellen Informationen der Etiketten und
bestimmte Probemethoden fest. Sie beinhaltet eine Beschreibung für die Homogenisierung,
Pasteurisierung und thermische Verarbeitung von Milch.

35
- NOM-184-SSA1-2002
Diese Norm setzt sanitäre und hygienische Anforderungen fest, welche Milch für den
menschlichen Konsum erfüllen muss, wenn sie in Mexiko hergestellt und in Umlauf gebracht
oder aus dem Ausland importiert werden soll.
- NMX-F-026-1997-SCFI
In dieser Norm erfolgt die Namensgebung für die verschiedenen Milchtypen im Einklang mit
den Verarbeitungsprozessen sowie für die Mindestanforderungen an die Qualität. Sie besagt,
dass Milch von gesunden Kühen stammen muss und 15 Tage vor bis 5 Tage nach der Geburt
keine Milch für den menschlichen Konsum gewonnen werden darf. Sie muss für die
Unbedenklichkeit für den Verbraucher einer thermischen Behandlung unterzogen werden.
Weiter darf Milch einem Filtrationsprozess, der Homogenisierung oder anderen
Behandlungen ausgesetzt werden, wenn sie hierbei nicht kontaminiert und in ihren
ernährungsphysiologischen Werten verändert wird.
Neben den genannten Normen plant das nationale Komitee für Normierung die Aufnahme der
Rohmilch mit ihren physiochemischen und biologischen Eigenschaften sowie geeigneten
Probemethoden. Nach Vollendung dieses Projektes PROY-NMX-F-700-COFOCALEC-2003
soll die Norm NMX-F-026-1997-SCFI außer Kraft treten. Mit Hilfe dieser neuen Norm und
der gründlichen Revision und Aktualisierung der momentan gültigen Normen aus den 80er
Jahren könnte ein wichtiger Schritt in Richtung Mastitisprävention und Verbraucherschutz
erreicht werden.
2.5 Klimatische Einflüsse auf das Mastitisgeschehen Das Klima hat einen großen Einfluss auf das Wohlbefinden der Tiere und somit auch auf die
Mastitisinzidenz. Die Behaglichkeitszone im Bezug auf die Temperatur für Milchkühe liegt
zwischen 5 und 17°C. Innerhalb dieses Bereiches tritt für die Tiere kein Stress auf. Fallen die
Temperaturen unter 5°C, steigt lediglich der Erhaltungsbedarf der Tiere an. Bei Temperaturen
über 24°C sind die Tiere einer Stress-Situation ausgesetzt, so dass die Milchmengenleistung
bei gleichzeitigem Anstieg der Zellzahl sinkt (KIELWEIN 1994). Guadalajara liegt 20 Grad
05 `nördlicher Breite und 103 Grad 05 `östlicher Länge auf 1540 Metern Höhe. Die
durchschnittlichen Tagestemperaturen im Sommer liegen in Jalisco bei 28°C. In der Nacht
kühlt die Luft auf 23°C ab. Die Wintertemperaturen liegen bei 23°C am Tag und 10°C in der
Nacht.

36
Zu extreme Umgebungstemperaturen, die über dem biologischen Optimum liegen, führen bei
Milchkühen schnell zu hitzebedingtem Stress. Hierbei sinken die Abwehrkräfte, und die Tiere
werden anfälliger für Infektionen. Dabei erfolgt die Beeinflussung teils über die Erhöhung des
Energieumsatzes, teils über Veränderungen des intermediären Stoffwechsels sowie über
Veränderungen der Hormonkonstellationen. Der Hitzestress führt zu einer starken
Reduzierung der Raufutteraufnahme und des Wiederkauens, wodurch es zu einer verringerten
Produktion von Fettsäuren und zu Veränderungen im Acetat/Propionat-Verhältnis kommt.
Der pH-Wert sowie die Elektrolyt-Konzentration im Pansen sinken, was mit einem erhöhten
Natrium-Verlust über die Harnausscheidung und mit einem erhöhten Kalium-Verlust über die
Haut verbunden ist (MIELKE 1994).
Bei höheren Außentemperaturen, die oft mit einer erhöhten Luftfeuchte verbunden sind,
finden Keime außerdem ideale Wachstumsbedingungen, und durch ihre stärkere Vermehrung
steigt der Infektionsdruck auf die Kühe an. Durch die gesunkenen Abwehrkräfte infolge des
temperaturbedingten Stresses kann dann eine Invasion des Euters mit Mastitiserregern leichter
erfolgen (ZSCHÖCK et al. 1998). Bei Wärmestress steigt zudem die Wasseraufnahme,
wodurch die Tiere wiederum weniger Futter aufnehmen. Neben der erhöhten
Krankheitsanfälligkeit ist der Hitzestress der entscheidende limitierende Faktor für die
Milchproduktion in tropischen Regionen (KIELWEIN 1994).
Einen weiteren Einfluss auf die Mastitisinzidenz vor allem mit umweltassoziierten Erregern
haben nach SMITH und HOGAN (1993) die vermehrten Niederschläge in der Regenzeit, da
die Ausläufe und die Triebwege zum Melkstand zu diesem Zeitpunkt sehr matschig sind und
es zu einem erhöhten Kontaminationsrisiko für die Euter kommt. Insbesondere die teilweise
schlechten hygienischen Verhältnisse in Mexiko und die feuchten klimatischen Bedingungen
der Regenzeit begünstigen die Übersiedelung ubiquitärer Erreger vom Boden auf die
Milchdrüse der Tiere.
2.6 Geschichte und heutiger Stand der Milchproduktion in Mexiko Die Milchviehhaltung hat in Mexiko eine lange Tradition und begann etwa vor 400 Jahren,
als die Spanier die ersten Milchkühe einführten. Die Ureinwohner hielten bis dahin kaum
domestizierte Tiere, wenn man von einigen alten Rinderrassen, die aus Kuba eingeführt
worden waren, absieht. Es folgte eine Vergrößerung der Rinderpopulation, die mit der
extensiven Nutzung großer Landstriche einherging. Nachdem es zu einer staatlichen
Limitierung der Bodennutzung kam, folgte die Viehhaltung auf Haziendas, wo die Rinder zur
Produktion von Fleisch und Milch für den Binnenmarkt Mexikos gehalten wurden. Aus

37
diesen Haziendas entwickelten sich im 19. Jahrhundert produktive Einheiten mit
Bodenrechten und festangestellten Arbeitern für den internen Milch- und Fleischmarkt. Die
Milchwirtschaft entwickelte sich ohne jegliche Technisierung vor allem auf den Haziendas
nahe Mexiko Stadt und Guadalajara, den Hauptzentren des Milchkonsums. Die von den
Spaniern eingeführten Tiere waren jedoch keine reinen Milchviehrinder.
Die sozialen Umstrukturierungen im 19. Jahrhundert sowie die Revolution von 1910 setzten
der Entwicklung der Milchviehhaltung stark zu, da es durch Hungersnöte der mexikanischen
Bevölkerung zu einer extremen Dezimierung der Viehherden kam.
Zwischen 1910 und 1928 verstärkte man die Einfuhr von speziellen Milchviehrassen wie
Holstein Friesian, Jersey, Guernsey, Ayrshire und Brown Swiss, um die vorhandene
Tierpopulation genetisch zu verbessern und dadurch die nationale Milchproduktion zu
steigern. Ab 1940 folgte die Stabilisierung des mexikanischen Marktes durch einen
fortschreitenden Ausbau der Transportwege und eine industrielle Transformation. Diese
Entwicklung wurde durch die stetig steigende Nachfrage nach Kuhmilch gefördert. Etwa zur
gleichen Zeit traten erstmals Forderungen nach Lösung der lebensmittelhygienischen
Probleme, die mit der Milchproduktion verbunden waren, auf. Es kam zu ersten gesetzlichen
Festlegungen für die Produktion und Kommerzialisierung von Milchprodukten, da
insbesondere die Konservierung und die schlechte Hygiene im Umgang mit dem Produkt zu
Problemen führten.
So entstanden zwischen 1950 und 1970 die ersten Pasteurisierungsanlagen, um Milch mit den
nötigen hygienischen Erfordernissen anbieten zu können. Der Großteil der Bevölkerung
bevorzugte zu diesem Zeitpunkt noch Rohmilch als Nahrungsmittel.
In den 50er und 60er Jahren begann die Spezialisierung der Milchproduktion. Der Staat bot an
Milchprojekten interessierte Produzenten Kredite zur Unterstützung an. Hierdurch haben sich
allmählich große Milchviehbetriebe entwickelt, die sich in die Produktionskette der
milchverarbeitenden Industrie integrierten. Jedoch hielt trotz dieses Trends der Großteil der
mexikanischen Milchviehhalter an ihren traditionellen Produktions- und Haltungsformen fest.
Zwischen 1972 und 1994 zeigte die Milchproduktion ein relativ konstantes Wachstum. Der
Anteil der importierten Milchvolumina am nationalen Konsum verringerte sich demzufolge in
den 70er Jahren kontinuierlich, um schließlich unter die 10%-Marke zu fallen. Von den
importierten Volumina nimmt das Milchpulver aus Voll- bzw. entrahmter Milch die
wichtigste Stellung ein. In den 80er Jahren setzte eine erhöhte Nachfrage der Bevölkerung
nach dem Lebensmittel Milch ein, der die nationale Produktion nicht gewachsen war. Durch
eine drastische Erhöhung der Importmenge wurde versucht, die nationale Produktion zu

38
komplettieren. Diese Entscheidung hatte jedoch dramatische Auswirkungen: Durch das
internationale Angebot und die niedrigen Preise für importiertes Milchpulver fiel der den
mexikanischen Produzenten pro Liter ausbezahlte Preis unter die realen Produktionskosten, so
dass viele Betriebe an den Rand der Rentabilität gerieten. Das importierte Milchpulver und
andere Milchprodukte wie Laktose, Molke und Fette, welche im Einkauf noch günstiger als
Milchpulver waren, wurden von der mexikanischen Industrie aufgekauft und zur Herstellung
von Milchersatzprodukten verwendet, welche mit nationalen, aus Frischmilch hergestellten
Milchprodukten konkurrierten (JOSÉ ZORRILLA et al. 1996). Ferner überschwemmten im
Ausland hergestellte Milchprodukte den mexikanischen Markt. So wurden ausländische
Milchmarken und -produkte im Gegensatz zu den nationalen Marken zu einem bis zu 50%
niedrigeren Preis, der teilweise unter den nationalen Produktionskosten liegt,
kommerzialisiert (VELAZCO, 2002). Ein weiterer Punkt, der die Entwicklung der
mexikanischen Milchwirtschaft hemmte, war der seit 1974 zum Schutze des Konsumenten
durch den Staat fixierte Preis für pasteurisierte Milch. Als Grund für die Festsetzung des
Preises nannte man die Steigerung des Milchkonsums und die Ermöglichung des Erwerbs
dieses Grundnahrungsmittels für Jeden trotz des permanent sinkenden Einkommens. So lag
1987 trotz der hohen Importe die täglich pro Person verfügbare Milchmenge bei 267 ml. Seit
1989 steigerte sich die Verfügbarkeit durch das Wachstum der Produktion und der
Importmenge auf ein seit 25 Jahren nicht mehr erreichtes Niveau. Pro Person waren täglich
über 371 ml Milch verfügbar, diese Menge ist aber immer noch weit von der täglich
empfohlenen Aufnahmemenge der Food and Agriculture Organization (FAO) von 500 ml
entfernt.
Der dem Erzeuger gezahlte Milchpreis wird durch eine Reihe von Gegebenheiten festgesetzt,
die der industrielle Milchkäufer bestimmt. Hierbei spielen vor allem Qualitätserfordernisse,
die mit der Saisonalität schwankenden benötigten Milchvolumina und der durch die seit 1991
durchgeführten offiziellen Milchpulverversteigerungen aus den Importen festgesetzte
Referenzwert eine bedeutende Rolle.
Durch die extremen jahreszeitlichen Schwankungen der mexikanischen Produktion beträgt
der Milchpreis in der Trockenzeit 3 Pesos/kg Milch (ca. 0,23 €) und in der Regensaison, in
der die größten Volumina produziert werden, nur 2,53 Pesos/kg (ca. 0,19 €) Milch.
Durch die Abschaffung des staatlich kontrollierten Milchpreises seit Ende 1996, die
Förderung der Technisierung der Milchviehbetriebe in den 90er Jahren, den Rückgang der
Inflation und der Zinshöhe und den Zuwachs bei den Arbeitsstellen wurde von 1990-1998 ein
durchschnittliches jährliches Wachstum der Produktion von 3,9% ausgelöst. Durch die

39
erstgenannte Maßnahme wurde das Ende der Regulation von Seiten des Staates eingeleitet
und der Beginn für einen neoliberalen Markt gesetzt. DEL VALLE (2002) bemerkt, dass
durch die verstärkte Technisierung der Betriebe eine Reduzierung der Produktionskosten und
eine Verbesserung der Milchqualität erreicht werden konnte, die in einer verbesserten
Wettbewerbsfähigkeit Mexikos resultiert. Mit dem Wachstum der landeseigenen Produktion
kam es zu einer stetigen Verringerung der Importmenge. So betrug der prozentuale Anteil der
Importe am nationalen Konsum 1990 noch 46% und 1998 nur noch 15,8%. Seit 1998 werden
die importierten Milchmengen überwiegend für die sozialen Programme des Staates und zur
industriellen Weiterverarbeitung verwendet. Obwohl die mexikanische Milchproduktion in
den letzten Jahren ein ununterbrochenes Wachstum registriert, ist das Land immer noch der
größte Milchpulverimporteur weltweit, da es für Mexiko unmöglich ist, die Selbstversorgung
dieses Grundnahrungsmittels zu erlangen. So erwirbt Mexiko 33% des auf dem Weltmarkt
kommerzialisierten Milchpulvers (FIRA, 1996). Die kommerzielle Balance auf dem Gebiet
des Milchsektors ist stark defizitär, und 1993 wurde ein absoluter Tiefpunkt mit einem Defizit
von 433,9 Millionen US$ erreicht (DGDG, 1999).
Unter den beschriebenen Bedingungen verbesserten sich die Perspektiven und die
Produktionsvoraussetzungen für die mexikanischen Milchproduzenten wieder erheblich.
Gleichzeitig konnte die Rentabilität gesteigert werden.
Da es sich bei Kuhmilch um ein Grundnahrungsmittel handelt, besitzt der Milchsektor
Mexikos unter sämtlichen landwirtschaftlichen Aktivitäten eine hohe Priorität.
Die Wichtigkeit des Milchsektors gemeinsam mit der Milch verarbeitenden Industrie zeigt
sich durch seine Schaffung von 1,5 Millionen Arbeitsplätzen und seinen Beitrag zum
nationalen Bruttosozialprodukt mit 1,2%. Die Finanzierungsgruppe Serfin führte 1995 eine
Studie durch und ermittelte einen ungefähren jährlichen Verkaufswert für Milchprodukte von
12.000 Millionen Pesos. Dies entspricht ungefähr einer Milliarde US$ (DEL VALLE et al.
1997).
Die Situation des mexikanischen Milchsektors war von Anfang an stark mit den Bewegungen
der Milchproduktion in der Welt verbunden. Die Weltproduktion wird vor allem von den am
weitesten entwickelten Ländern wie den USA, Kanada, Neuseeland, Australien und einigen
europäischen Ländern bestimmt. Diese produzieren Überschüsse, und ihre transnationalen
Unternehmen dirigieren und kontrollieren den Weltmarkt, der bis zu den 90er Jahren ein
stetiges Wachstum zeigte. 1990 wurde der bisherige Höhepunkt der Weltproduktion mit 441,3
Millionen Tonnen Milch erreicht. Danach kam es jedoch zu einem Produktionsrückgang, so
dass 1999 noch 387 Millionen Tonnen produziert wurden.
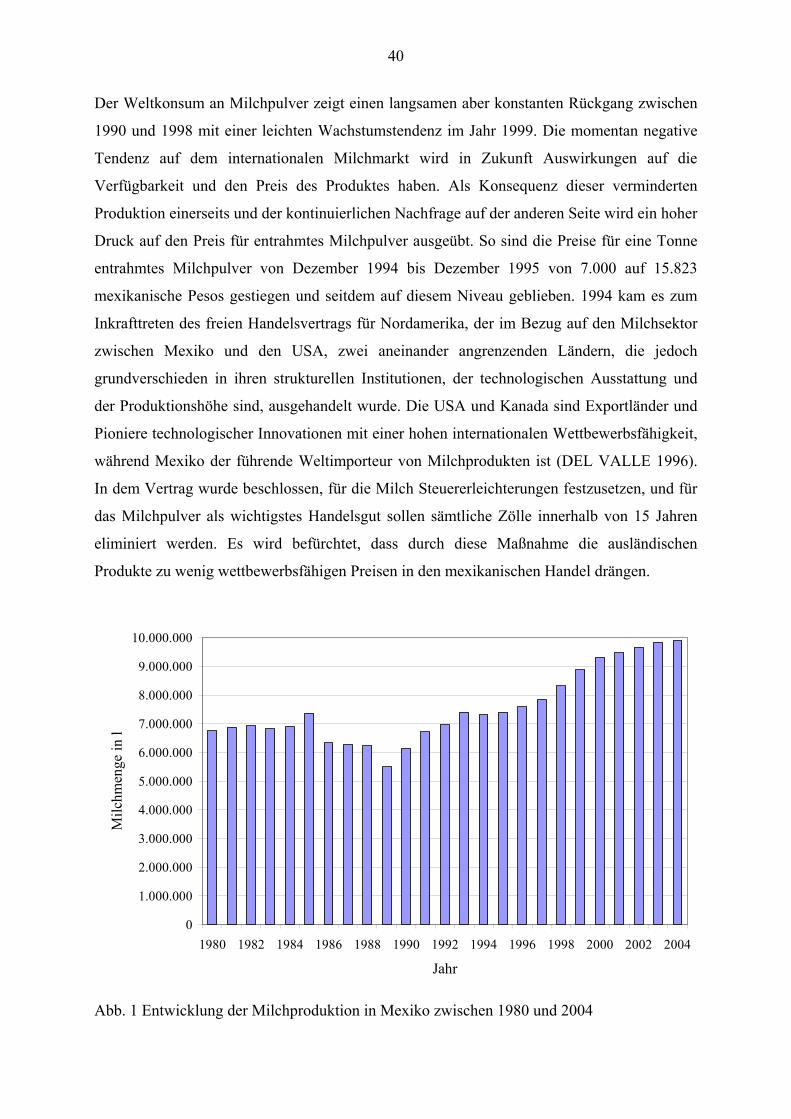
40
Der Weltkonsum an Milchpulver zeigt einen langsamen aber konstanten Rückgang zwischen
1990 und 1998 mit einer leichten Wachstumstendenz im Jahr 1999. Die momentan negative
Tendenz auf dem internationalen Milchmarkt wird in Zukunft Auswirkungen auf die
Verfügbarkeit und den Preis des Produktes haben. Als Konsequenz dieser verminderten
Produktion einerseits und der kontinuierlichen Nachfrage auf der anderen Seite wird ein hoher
Druck auf den Preis für entrahmtes Milchpulver ausgeübt. So sind die Preise für eine Tonne
entrahmtes Milchpulver von Dezember 1994 bis Dezember 1995 von 7.000 auf 15.823
mexikanische Pesos gestiegen und seitdem auf diesem Niveau geblieben. 1994 kam es zum
Inkrafttreten des freien Handelsvertrags für Nordamerika, der im Bezug auf den Milchsektor
zwischen Mexiko und den USA, zwei aneinander angrenzenden Ländern, die jedoch
grundverschieden in ihren strukturellen Institutionen, der technologischen Ausstattung und
der Produktionshöhe sind, ausgehandelt wurde. Die USA und Kanada sind Exportländer und
Pioniere technologischer Innovationen mit einer hohen internationalen Wettbewerbsfähigkeit,
während Mexiko der führende Weltimporteur von Milchprodukten ist (DEL VALLE 1996).
In dem Vertrag wurde beschlossen, für die Milch Steuererleichterungen festzusetzen, und für
das Milchpulver als wichtigstes Handelsgut sollen sämtliche Zölle innerhalb von 15 Jahren
eliminiert werden. Es wird befürchtet, dass durch diese Maßnahme die ausländischen
Produkte zu wenig wettbewerbsfähigen Preisen in den mexikanischen Handel drängen.
0
1.000.000
2.000.000
3.000.000
4.000.000
5.000.000
6.000.000
7.000.000
8.000.000
9.000.000
10.000.000
1980 1982 1984 1986 1988 1990 1992 1994 1996 1998 2000 2002 2004
Jahr
Milc
hmen
ge in
l
Abb. 1 Entwicklung der Milchproduktion in Mexiko zwischen 1980 und 2004

41
2.7 Betriebsformen in Mexiko
Die aktuelle Milchproduktion in Mexiko setzt sich laut DEL VALLE (2002) sehr heterogen
zusammen. Dies ist unter anderem die Folge der unterschiedlichen Bodenbeschaffenheit,
Klimaverhältnisse und der Infrastruktur der einzelnen Regionen. Die verschiedenen
Betriebsformen lassen sich aufgrund der technischen Ausstattung, der Herdengröße und dem
erzielten Ertrag unterscheiden. Die DIRECION GENERAL DE GANDERÍA (1999) teilt die
mexikanischen Betriebe in vier verschiedene Gruppen ein: Auf Milchproduktion spezialisierte
Betriebe, halbspezialisierte Betriebe, familiäre Hinterhofbetriebe und
Doppelnutzungssysteme.
2.7.1 Spezialisierte Betriebe Die spezialisierten Betriebe in Mexiko arbeiten hauptsächlich mit einseitig auf Milchleistung
gezüchteten Rassen, vorwiegend mit Holstein Friesian und in weit geringerem Grade mit
Brown Swiss und Jersey. Die durchschnittliche Tierzahl liegt bei 300-400 Tieren (DGDG
1999) bzw. 100-300 Tieren (DEL VALLE 1997). In diesen Betrieben werden mit
durchschnittlichen Leistungen von 5.000-6.500 Litern pro Kuh und Laktation bei einer
Laktationsdauer von 305 Tagen die größten Renditen erzielt (FIRA 1996).
Laut DGDG (1999) sind die spezialisierten Betriebe im hohen Maße technisiert, so dass
sowohl auf den Feldern als auch bei der Melkarbeit moderne Maschinen eingesetzt werden
und die Möglichkeit zur Kühlhaltung und zum Abtransport der Milch besteht. Die Tiere
werden überwiegend aufgestallt gehalten. Große Bemühungen zur genetischen Verbesserung
des Tierbestandes und für eine optimale tiermedizinische Versorgung werden durchgeführt.
Eine Berechnung der optimalen Fütterungsmenge wird für die einzelnen Tiere bzw.
Tiergruppen vorgenommen. Es wird Silage und Kraftfutterkonzentrat gefüttert. Die
produzierte Milch wird ausschließlich an die großen Molkereien wie Lala oder Sello Rojo
abgegeben, Rohmilchabgabe an Privatpersonen findet keine statt (FIRA 1996). 1998 wurden
in den spezialisierten Betrieben 4.196 Millionen Liter Milch produziert, was einen Anteil von
50,5% der gesamten Produktion entspricht. Diese Menge wurde von 17% der
Milchviehpopulation ganz Mexikos produziert (DGDG 1999). Die durchschnittliche jährliche
Milchleistung pro Tier liegt bei 5.667 Litern Milch. Die spezialisierten Betriebssysteme sind
sehr empfindlich für Variationen des Wechselkurses und Veränderungen der
Importkonditionen für die benötigte technische Ausstattung und für den Zukauf von
Zuchttieren zum Zweck der genetischen Verbesserung (DEL VALLE 1997). Man findet diese
Betriebsform zumeist in den Hochflächen sowie den trockenen oder halbtrockenen Zonen des

42
Nordens wie in Durango, Coahuila, Guanajuato und Jalisco, wo sich auch die großen
milchverarbeitenden Unternehmen befinden (DGDG 1999).
2.7.2 Halbspezialisierte Betriebe Die halbspezialisierte Betriebsform charakterisiert sich mit einer Herdengröße von etwa 180-
200 Tieren, deren genetisches Potential jedoch nicht so hoch ist wie bei den Tieren der
spezialisierten Betriebe (DGDG 1999). Es dominieren auch hier die Milchviehrassen Holstein
Friesian und Brown Swiss, es kommen jedoch auch Criollos und Zebus vor (DEL VALLE
1997). Wegen des geringeren Grades der genetischen Reinheit werden nur Leistungen von
18-20 Litern täglich bei einer Laktationsdauer von 280-305 Tagen erreicht. Die Tiere werden
häufig halbaufgestallt auf fest eingezäunten Flächen gehalten, die teilüberdacht sein können.
Den in den halbspezialisierten Betrieben gehaltenen Tieren stehen pro Großvieheinheit etwa
10-50 Hektar Land zur Verfügung. Die Installationen sind an die Nutzung des Milchviehs
adaptiert. Es wird von Hand, mit Einzelmelkzeugen oder Melkanlagen mit wenigen
Melkzeugen gemolken. Oft mangelt es an Kühl- und Aufbewahrungsmöglichkeiten für die
Milch, wodurch nur ein mittleres technologisches Niveau in Infrastruktur und Ausstattung
erreicht wird. Sind keine Kühlmöglichkeiten vorhanden, wird die Milch nach der Melkzeit
behelfsmäßig in Kannen im Wasserbad gekühlt und direkt von der Molkerei abgeholt. Einige
Erzeuger transportieren ihre Milch selbst zum nächstgelegenen Sammelkühltank oder zu
Zwischenhändlern.
Die selbst erzeugten Agrarprodukte sowie täglicher Weidegang stellen die Basis der
Milchviehfütterung dar. In geringerem Maße, meist nur zur Melkzeit, wird Kraftfutter
zugefüttert (DEL VALLE 2002).
Die meisten halbspezialisierten Betriebe führen eine Kontrolle der Produktion und
Reproduktion durch. Die künstliche Besamung ist weit verbreitet. Jedoch erhält meist nur ein
kleiner Prozentteil der Tierpopulation Sperma eines Bullen mit hoher genetischer Qualität,
weswegen die genetische Entwicklung nur langsam vorangeht. Auf der anderen Seite kommt
es durch das kurze reproduktive Leben der Tiere häufig zu einem Verlust von genetischem
Potential.
1998 wurde in den halbspezialisierten Betrieben 15% der Milchviehpopulation Mexikos
gehalten, welche 1.717 Millionen Liter Milch produzierte. Dies entspricht 20,63% der
nationalen Produktion (DGDG 1999). Pro Kuh wurden durchschnittlich 2.786 Liter Milch pro
Jahr erwirtschaftet. Diese Betriebsform findet sich vor allem in Baja California, Colima,
Chihuahua (DGDG 1999) und häufig auch in Jalisco (DEL VALLE 1997).

43
2.7.3 Familiäre Hinterhofbetriebe
Die Betriebsform der familiären Hinterhofbetriebe stellt das traditionelle Haltungssystem
Mexikos dar (DGDG 1999). Die Tiere werden auf kleinen Flächen, meist in Hinterhofställen,
gehalten (LOSADA et al. 1998). Auch hier überwiegen die Rassen Holstein Friesian und
Brown Swiss (DVDG 1999), jeweils mit niedriger genetischer Qualität (DEL VALLE 1997).
Im Durchschnitt findet man 2-10 Tiere mit einer täglichen Leistung von 6-12 Litern bei einer
Laktationsdauer von 210-260 Tagen (LOSADA et al.1998; DGDG 1999). Das technologische
Niveau ist niedrig. Es dominiert das Handmelken, nur selten stehen einfache Melkzeuge zur
Verfügung. Nach LOSADA et al. (1998) wird in 66% der Betriebe die Arbeitskraft nur einer
Familie eingesetzt. Nur 20% der Familien leben allein von der Milchproduktion. Die übrigen
Familien supplementieren diesen Verdienst mit anderen Einkommensquellen. DDie Tierhalter
führen keine vorbeugenden tiermedizinischen Maßnahmen oder Bestrebungen zur
genetischen Verbesserung durch. Es überwiegt der Natursprung, nur vereinzelt wird die
künstliche Besamung angewendet. Die Futtergrundlage stellt Feldfutter, und falls vorhanden,
der Weidegang dar. Kraftfutter wird während des Melkvorgangs ohne Rationsberechnung
zugefüttert. In diesem System wurden 1998 etwa 780 Millionen Liter Milch von 8% der
Milchviehpopulation produziert. Diese Menge entspricht 9,4% der nationalen Produktion
(DGDG 1999). Jede Kuh gab durchschnittlich 2.230 Liter Milch im Jahr. DEL VALLE
(2002) gibt die durchschnittliche erzielte Jahresleistung der Tiere sogar nur 630 Litern an.
Die meiste Milch wird selbst konsumiert oder als Rohmilch an Zwischenhändler und die
Nachbarschaft verkauft. Dieses System findet sich vor allem in Jaslico, gefolgt von México,
Michoacán, Hidalgo und anderen Staaten.
Abb. 2 Prozentuale Verteilung der einzelnen Betriebsformen in Mexiko (DGDG 1999)
Doppelnutzung 60%
Familienbetriebe 8%
Spezialisiert 17%
Halbspezialisiert 15%

44
2.7.4 Doppelnutzungsbetriebe
Die Betriebsform der Doppelnutzung für Milch- und Fleischproduktion kommt vor allem in
den tropischen Regionen des Landes vor. Es werden meist 30-40 Tiere gehalten, die bei einer
durchschnittlichen Laktationsdauer von 120-180 Tagen täglich 3-9 Liter Milch produzieren
(DGDG 1999). DEL VALLE (2002) gibt die durchschnittliche jährliche Milchleistung der
Doppelnutzungsrinder mit etwa 600 Litern an.
Es werden überwiegend Zebus und Zebu-Kreuzungen und nur in geringem Maße Brown
Swiss, Holstein Friesian und Simmentaler extensiv auf der Weide gehalten (DOMÍNGUEZ
1996). Falls eine Zufütterung stattfindet, werden Nebenprodukte der Landwirtschaft
verwendet. Der große Vorteil dieser Haltungsform ist in seiner Flexibilität zu sehen. So kann
entsprechend der aktuellen Nachfrage des Marktes entweder verstärkt Milch oder Fleisch
produziert werden.
Es wird fast ausschließlich von Hand gemolken. Die Milch stellt die erste Einkommensquelle
für die Erhaltung und Weiterführung des Betriebes dar, bis die Tiere für die
Fleischgewinnung verkauft werden. Mittlerweile gibt es aber schon Betriebe, die den
kommerziellen Charakter spezialisierter Milchproduktion besitzen. Die Milch wird meist
direkt als Rohmilch an Konsumenten, an Zwischenhändler zur Produktion von Käse oder vor
allem in der Hauptsaison zu Billigpreisen an die Industrie verkauft. Generell ist das
Management stark verbesserungswürdig. Es werden keine tiermedizinischen Vorsorgungen
oder genetischen Verbesserungen angestrebt. Die Probleme dieser Systeme liegen vor allem
in der Tiergesundheit, der Milchkonservierung und dem Produkttransport (FIRA 1996). 1998
wurden in diesem System 60% der nationalen Milchviehpopulation gehalten, die 1.622
Millionen Liter Milch produzierten. Diese Menge entspricht 19,5% der nationalen Produktion
(DGDG 1999). Dies entspricht einer durchschnittlichen jährlichen Milchleistung pro Kuh von
662 Litern.
1991 wurden in ganz Mexiko 24.611.862 Rinder gezählt. 11.006.737 dieser Tiere sind über
dreijährige Kühe (INEGI 1991). Im Jahr 1997 wurden in den Betriebsformen Spezialisiert,
Halbspezialisiert und in den Familienbetrieben 3.480.199 Tiere gehalten, von denen
1.879.352 Kühe im reproduktiven Alter waren. Im System der Doppelnutzung befinden sich
6.090.752 Rinder mit 2.369.912 Kühen. Die übrigen Tiere werden zur reinen
Fleischproduktion gehalten (DGDG 1999). Im Staat Jalisco wurden 1999 2.263.698 Rinder
gehalten, mit denen 1.563.605 Liter Milch produziert wurden (INEGI 2000).
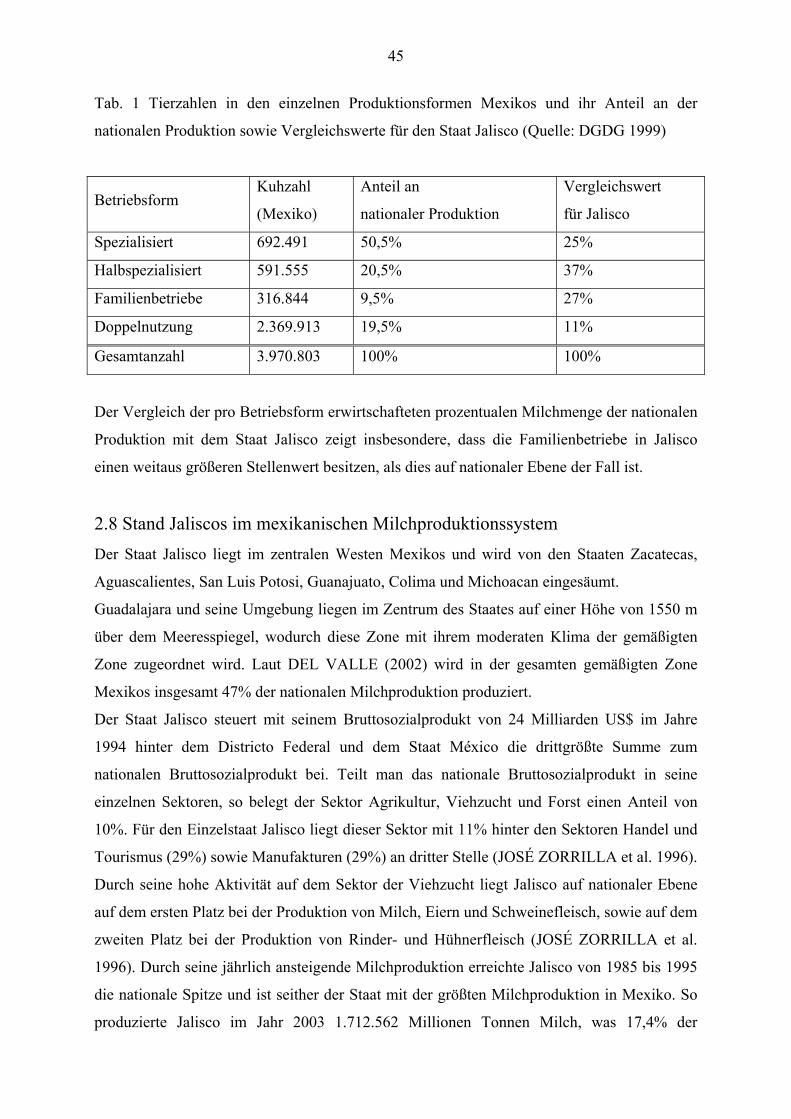
45
Tab. 1 Tierzahlen in den einzelnen Produktionsformen Mexikos und ihr Anteil an der
nationalen Produktion sowie Vergleichswerte für den Staat Jalisco (Quelle: DGDG 1999)
Betriebsform Kuhzahl
(Mexiko)
Anteil an
nationaler Produktion
Vergleichswert
für Jalisco
Spezialisiert 692.491 50,5% 25%
Halbspezialisiert 591.555 20,5% 37%
Familienbetriebe 316.844 9,5% 27%
Doppelnutzung 2.369.913 19,5% 11%
Gesamtanzahl 3.970.803 100% 100%
Der Vergleich der pro Betriebsform erwirtschafteten prozentualen Milchmenge der nationalen
Produktion mit dem Staat Jalisco zeigt insbesondere, dass die Familienbetriebe in Jalisco
einen weitaus größeren Stellenwert besitzen, als dies auf nationaler Ebene der Fall ist.
2.8 Stand Jaliscos im mexikanischen Milchproduktionssystem Der Staat Jalisco liegt im zentralen Westen Mexikos und wird von den Staaten Zacatecas,
Aguascalientes, San Luis Potosi, Guanajuato, Colima und Michoacan eingesäumt.
Guadalajara und seine Umgebung liegen im Zentrum des Staates auf einer Höhe von 1550 m
über dem Meeresspiegel, wodurch diese Zone mit ihrem moderaten Klima der gemäßigten
Zone zugeordnet wird. Laut DEL VALLE (2002) wird in der gesamten gemäßigten Zone
Mexikos insgesamt 47% der nationalen Milchproduktion produziert.
Der Staat Jalisco steuert mit seinem Bruttosozialprodukt von 24 Milliarden US$ im Jahre
1994 hinter dem Districto Federal und dem Staat México die drittgrößte Summe zum
nationalen Bruttosozialprodukt bei. Teilt man das nationale Bruttosozialprodukt in seine
einzelnen Sektoren, so belegt der Sektor Agrikultur, Viehzucht und Forst einen Anteil von
10%. Für den Einzelstaat Jalisco liegt dieser Sektor mit 11% hinter den Sektoren Handel und
Tourismus (29%) sowie Manufakturen (29%) an dritter Stelle (JOSÉ ZORRILLA et al. 1996).
Durch seine hohe Aktivität auf dem Sektor der Viehzucht liegt Jalisco auf nationaler Ebene
auf dem ersten Platz bei der Produktion von Milch, Eiern und Schweinefleisch, sowie auf dem
zweiten Platz bei der Produktion von Rinder- und Hühnerfleisch (JOSÉ ZORRILLA et al.
1996). Durch seine jährlich ansteigende Milchproduktion erreichte Jalisco von 1985 bis 1995
die nationale Spitze und ist seither der Staat mit der größten Milchproduktion in Mexiko. So
produzierte Jalisco im Jahr 2003 1.712.562 Millionen Tonnen Milch, was 17,4% der

46
nationalen Produktion entsprach, welche im Jahr 2003 insgesamt 9.842.422 Millionen Tonnen
betrug (SAGAR 2004).
Ein Grund für diese führende Stellung Jaliscos mag die traditionelle Milchviehhaltung des
Staates sein. So lebt in Jalisco eine große Anzahl milchviehhaltender Familien, die diese
Aktivität von Generation zu Generation weitergeben. Durch diese starke kulturelle
Verankerung kam es bereits in den 40er Jahren des letzten Jahrhunderts zu einer
Spezialisierung der Landwirtschaft dieses Staates auf die Milchproduktion (DEL VALLE et
al. 1997).
So beträgt der Milchkuhbestand Jaliscos mit 828.000 Tieren etwa 20% des nationalen
Viehbestandes (JOSÉ ZORRILLA et al. 1996).
Wie in der gesamten mexikanischen Milchindustrie zeigt sich jedoch auch im Staat Jalisco
eine große Heterogenität der milchproduzierenden Einheiten. So wirtschaften 10% der
Betriebe Jaliscos mit großen Herden von 100-500 Tieren. Diese hochspezialisierten
Produktionssysteme befinden sich überwiegend in den Regionen Lagos de Moreno,
Jalostotitlan und Tepatitlan (JOSÉ ZORRILLA et al. 1996). Weitere 20% halten Herden
zwischen 25-100 Tieren, während die übrigen 70% Familienbetriebe sind, die meist in
Hinterhöfen 6-20 Tiere melken (DEL VALLE et al. 1997). Diese Kleinbetriebe befinden sich
überwiegend in der Region Los Altos de Jalisco, der Region mit der größten Milchproduktion
Jaliscos, während sich an der tropischen Westküste des Staates überwiegend
Doppelnutzungsbetriebe befinden, die Milch und Fleisch gleichermaßen produzieren.
Die Kleinbetriebe führen nur 25% ihrer Produktion der industriellen Verarbeitung zu,
während die mittleren und großen Betriebe 100% des gewonnenen Produktes an die
Milchindustrie liefern.
Diese Kleinbetriebe zeigen eine flexible Spezialisierung. Es überwiegen die Rassen Holstein
Friesian, Criollo und Kreuzungen. Die durchschnittliche Leistung pro Kuh liegt bei 10 (8-24)
Litern pro Tag (DEL VALLE et al. 1997). Die Betriebe kommen mit einer minimalen
Infrastruktur zurecht. So wird meist von Hand gemolken, oder es kommen überwiegend
veraltete, dagegen nur selten moderne Melksysteme zum Einsatz. Oftmals werden die Tiere
ganztags auf Weiden gehalten und nur für den Melkvorgang in Ausläufe getrieben. Folglich
wird die Grundfutterversorgung durch den Weidegang sichergestellt, der durch Rationen von
Maissilage und Kraftfutter aufgebessert wird (DEL VALLE et al. 1997). Die genetische
Verbesserung erfolgt überwiegend durch die Selektion der Bullen für den Natursprung und
nur in etwa 15% durch die künstliche Besamung.

47
Generell hat das Milchgeld in Jalisco an Wichtigkeit für das familiäre Einkommen eingebüßt.
Die großen Betriebe produzieren vielerorts Luzerne, Hafer, Mais und Mastfutter für Schweine
und Hühner, um ein Zusatzeinkommen zu haben. Die kleinen Produzenten suchen eine zweite
Einnahmequelle außerhalb ihres Betriebs.
In Gegensatz zu vielen anderen Staaten Mexikos, in denen die milchverarbeitenden Industrien
eine Monopolstellung innehaben, existieren in Jalisco mehrere hundert verschiedene
Unternehmen, welche die Milch aufkaufen. Durch diese Marktlage erhalten die Produzenten
eine größere Beweglichkeit bei der Suche nach optimalen Konditionen für den Verkauf ihres
Produktes (DEL VALLE et al. 1997). Von den die Milch aufkaufenden Unternehmen
verlangen 67% keinerlei Qualitätsprüfung der Anlieferungsmilch. Die restlichen 33% fordern
eine Hygieneprüfung. In 65% der Fälle bestehen für die Milchabnahme nur mündliche
Vereinbarungen zwischen Abnehmer und Produzent.
Den Produzenten wird hierbei die Abnahme ihres Produktes zugesichert. Ferner fördern die
Unternehmen den Erwerb von Kühltanks durch Kommissionen für die individuellen
Produzenten und Produktionsgesellschaften und bieten technische Beratung sowie Bürgung
bei Krediten an.
Trotz des Wettbewerbs der vielen Unternehmen üben diese die Kontrolle über die Milchpreise
und die Qualität des Produktes aus. So fordern immer mehr Unternehmen die Lieferung von
gekühlter Milch, wodurch es 1990 in Jalisco zur Einführung des für das Land Mexiko
einmaligen Programms für den Erwerb von Kühltanks kam. Dieses Programm zählt zu einer
der erfolgreichsten Aktionen auf dem Milchsektor. Die Umsetzung erfolgte durch die
gemeinsame Beteiligung der privaten Industrie, der Regierung und der Produzenten. Der
Erfolg des Programms zeigt sich in der ständigen Zunahme der Anzahl der Kühltanks von 20
in Jahr 1992 auf 270 in Jahr 1995 (JOSE ZORRILLA et al. 1996).
Jedoch wurden durch die Einführung der Kühltanks die Kosten für die Einsammlung und
Kühlung der Milch von den Unternehmen auf die Produzenten übertragen. Und obwohl die
Produzenten nun bessere Preise für ihr Produkt erhalten, argumentieren viele, dass die
Kosten, die durch die Auflagen der Unternehmen ausgelöst wurden, über dem Mehrverdienst
durch die Lieferung einer qualitativ hochwertigeren Milch liegen. Obwohl dies in einigen
Fällen zutrifft, muss gleichzeitig darauf hingewiesen werden, dass die Unternehmen an der
Abnahme von ungekühlter Milch kein Interesse mehr zeigen, so dass außerhalb dieser neuen
Organisation kaum noch Möglichkeiten für den Produzenten bestehen, sein Produkt zu
verkaufen.
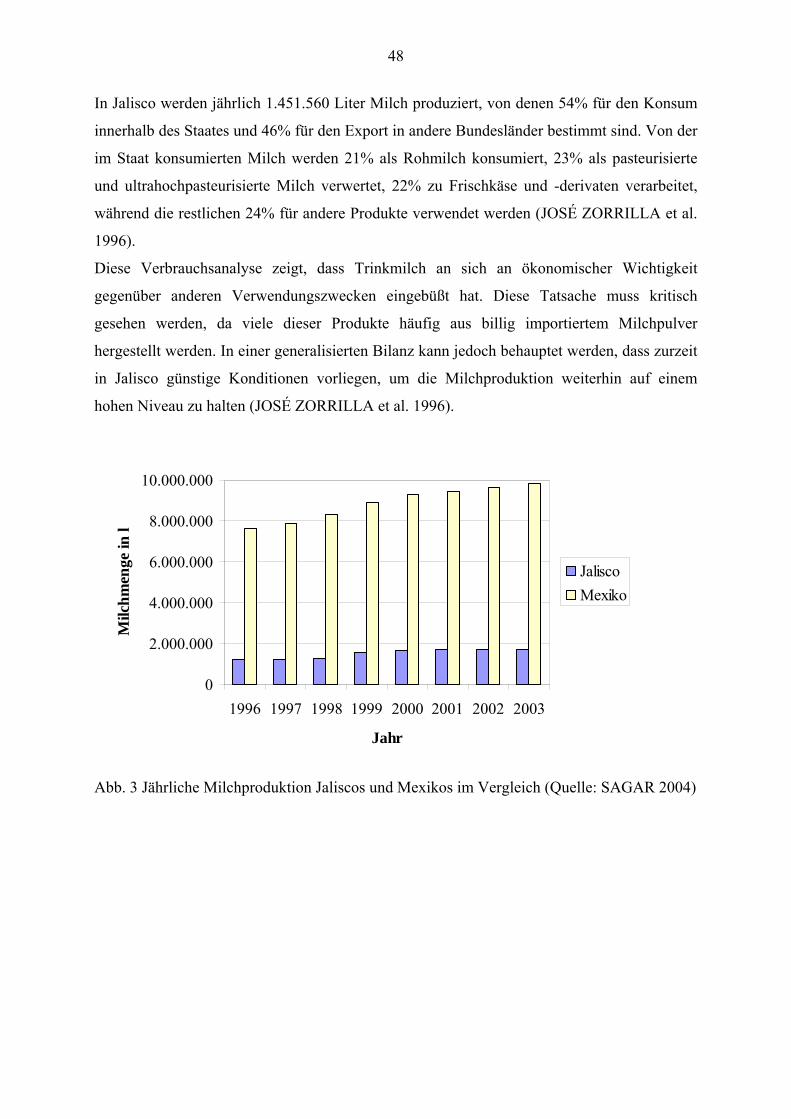
48
In Jalisco werden jährlich 1.451.560 Liter Milch produziert, von denen 54% für den Konsum
innerhalb des Staates und 46% für den Export in andere Bundesländer bestimmt sind. Von der
im Staat konsumierten Milch werden 21% als Rohmilch konsumiert, 23% als pasteurisierte
und ultrahochpasteurisierte Milch verwertet, 22% zu Frischkäse und -derivaten verarbeitet,
während die restlichen 24% für andere Produkte verwendet werden (JOSÉ ZORRILLA et al.
1996).
Diese Verbrauchsanalyse zeigt, dass Trinkmilch an sich an ökonomischer Wichtigkeit
gegenüber anderen Verwendungszwecken eingebüßt hat. Diese Tatsache muss kritisch
gesehen werden, da viele dieser Produkte häufig aus billig importiertem Milchpulver
hergestellt werden. In einer generalisierten Bilanz kann jedoch behauptet werden, dass zurzeit
in Jalisco günstige Konditionen vorliegen, um die Milchproduktion weiterhin auf einem
hohen Niveau zu halten (JOSÉ ZORRILLA et al. 1996).
0
2.000.000
4.000.000
6.000.000
8.000.000
10.000.000
1996 1997 1998 1999 2000 2001 2002 2003
Jahr
Milc
hmen
ge in
l
JaliscoMexiko
Abb. 3 Jährliche Milchproduktion Jaliscos und Mexikos im Vergleich (Quelle: SAGAR 2004)

49
3 Tiere, Material und Methoden
3.1 Milcherzeugerbetriebe und Tierbestände Im Zeitraum von Oktober 2002 bis Juni 2003 wurden 33 milchproduzierende Betriebe aus
dem näheren Umfeld von Guadalajara, Jalisco, in die Studie aufgenommen.
Um eine möglichst große räumliche Streuung der Milcherzeugerbetriebe zu erreichen, war
neben einer Beprobung sämtlicher in Jalisco vorkommender unterschiedlicher
Betriebsstrukturen die geographische Lage Kriterium für die Aufnahme in die Studie.
Die Anzahl der ausgewählten Betriebe, nach Betriebsformen geordnet, lässt sich der
folgenden Tabelle entnehmen:
Tab. 2 Aufteilung der besuchten 33 Betriebe nach Betriebsformen
Betriebsform Anzahl Betriebe Prozentualer Anteil
Spezialisierte Betriebe 5 15,2%
Halbspezialisierte Betriebe 17 51,5%
Familiäre Hinterhofbetriebe 11 33,3%
Die räumliche Verteilung der Betriebe kann aus der Abb. 19 auf Seite XV des Anhangs
entnommen werden. Die Betriebe lagen zwischen 0,5 und ca. 120 km auseinander.
Die durchschnittliche Herdengröße der beprobten Betriebe lag bei 66 Tieren. Die folgende
Tabelle zeigt eine Aufteilung der Betriebe nach Anzahl der gehaltenen Tiere:
Tab. 3 Aufteilung der 33 Betriebe nach Anzahl der Milchkühe
Anzahl Kühe Anzahl Betriebe Prozentualer Anteil
<25 11 33,4%
25-75 7 21,2%
75-100 8 24,2%
>100 7 21,2%

50
Überwiegend setzten sich die Bestände aus Tieren der Rasse Holstein Friesian zusammen.
Nur vereinzelt wurden daneben Kühe der Rasse Brown Swiss, Criollo oder Jersey gehalten.
Die Kühe befanden sich in unterschiedlichen Laktationsstadien. Die durchschnittliche
Milchleistung der Betriebe lag bei 19,75 Litern pro Kuh und Tag. Die Angaben zur
Milchleistung wurden durch die Betriebsleiter im Rahmen einer Befragung gemacht, da in
Mexiko die Milchleistungsprüfung mit Erfassung der Milchmenge nicht üblich ist. Folgende
Tabelle zeigt die Aufteilung der Betriebe nach Milchleistung in Litern/Tag:
Tab. 4 Aufteilung der 33 Betriebe nach der Milchproduktion in Litern pro Kuh und Tag
Tägliche Milchleistung pro Kuh in Litern Anzahl Betriebe Prozentualer Anteil
<15 4 12,1%
15-20 10 30,3%
21-25 17 51,5%
26-30 2 6,1%
Die größeren Betriebe sind modern ausgestattet und Melkmaschinen, während in den
Kleinbetrieben überwiegend von Hand gemolken wird.
In der folgenden Tabelle wurden die Betriebe den verschiedenen Melksystemen zugeordnet:
Tab. 5 Aufteilung der 33 Betriebe nach Melksystemen
Melksystem Anzahl Betriebe Prozentualer Anteil
Handmelken 7 21,2%
Eimermelkanlage stationär 5 15,2%
Eimermelkanlage mobil 3 9,1%
Rohrmelkanlage 18 54,5%
16 Betriebe (48,5%) geben Rohmilch an Privatpersonen des näheren Umkreises ab. 17
Betriebe (51,5%) liefern die Milch ausschließlich an Molkereien oder an Zwischenhändler,
wo sie vor der weiteren Verarbeitung zum größten Teil einer Pasteurisierung unterzogen wird.

51
Da nicht alle mexikanischen Molkereien die Anlieferungsmilch auf die Zellzahl hin
untersuchen und deshalb nicht zu allen Betrieben genaue Daten zur Verfügung standen,
können nur die Durchschnittszellzahlen der Tankmilch von 15 der 33 Betriebe angegeben
werden. Die Tankmilchzellzahlen bewegten sich zwischen 234.000 und 1.500.000 Zellen/ml
mit einem geometrischen Mittel von 623.000 Zellen/ml Milch.
Die zu beprobenden Kühe wurden vor Ort nach dem Zufallsprinzip ausgewählt. Teilweise
wurden auf Wunsch des Betriebsleiters verdächtige Kühe oder laut Vorbericht bereits
mehrfach an Mastitis erkrankte Tiere zusätzlich beprobt.
Es wurden möglichst viele Proben pro Betrieb genommen. In Betrieben mit weniger als 35
Milchkühen wurden alle Tiere in die Untersuchung aufgenommen. Ansonsten wurden
Teilbestandsuntersuchungen von jeweils 25-35 Tieren durchgeführt.
3.2 Datenerhebung auf den Betrieben mit Beurteilung der Melk- und
Stallhygiene sowie weiterer betriebswirtschaftlicher Faktoren Zu Beginn eines Besuches wurde der Betrieb besichtigt. Hierbei wurden die Ausläufe und
Weideflächen, der Melkplatz mit den Melkzeugen und, falls vorhanden, die Milchkammer mit
dem zur Kühlung bestimmten Tank in Augenschein genommen und hinsichtlich ihrer
hygienischen Verhältnisse bewertet. Während des Melkens wurde eine Beurteilung der
körperlichen Verfassung der Tiere sowie des Melkregimes einschließlich der Euterreinigung,
des Vormelkens, der Einhaltung einer Melkreihenfolge, der Durchführung des Zitzendippens
und der allgemeinen Melkhygiene vorgenommen. Nach der Probenentnahme wurde der
Betriebsleiter zu allen milchhygienisch relevanten Daten, zur Fütterung und
Rationsberechung, dem Trockenstellen und zu weiteren gesundheitlichen Problemen der Tiere
befragt. Der Fragebogen sollte einen Einblick in die verschiedenen Betriebsformen geben und
Faktoren und Managementdetails aufdecken, die das Auftreten von Mastitiden begünstigen.
Soweit vorhanden erfolgte Einsicht in die Milchgeldabrechnungen.
3.3 Milchprobenentnahme und Transport Die Probenentnahme erfolgte gemäß den Leitlinien zur Entnahme von Milchproben unter
antiseptischen Bedingungen der Deutschen Veterinärmedizinischen Gesellschaft (2000). Die
Zitzen wurden von den Melkern wie zum normalen Melkvorgang gereinigt und das
Vorgemelk ermolken. Anschließend wurde ein Milchstrahl verworfen und je Zitze 2-3
Milchstrahlen zur Beurteilung der Sekretbeschaffenheit und zur darauf folgenden
Durchführung des CMT in eine Testschale gemolken. Die Zitzenkuppen wurden anschließend

52
gründlich mit einem separaten alkoholgetränkten Zellstofftupfer gereinigt und desinfiziert.
Dann wurden etwa 10ml Milch pro Viertel in sterile Plastikröhrchen gemolken, welche sofort
verschlossen und gekennzeichnet wurden. Die Röhrchen wurden für den Transport in
geeignete Behältnisse verbracht und umgehend bei 4-5°C gekühlt.
3.4 California-Mastitis-Test (CMT) Der CMT wurde nach SCHALM und NOORLANDER (1957) durchgeführt. In die vier
Schalen einer Prüfplatte wurden aus jedem Euterviertel 2-3 Milchstrahlen gemolken und unter
langsam kreisenden Bewegungen mit der gleichen Menge des Standard-Schalm-Reagenz
vermischt. Gleichzeitig wurde die Milch auf visuelle Abnormalitäten wie abnormale Farbe
oder Flocken überprüft.
Die Beurteilung erfolgte in Anlehnung an ROSENBERGER (1990):
0 Negativ: Probe bleibt flüssig und zeigt keinerlei Anzeichen von Schlierenbildung oder
Konsistenzveränderung (< 200.000 Zellen/ml)
1 Leicht positiv: Beim Kippen der Testschale von vorne nach hinten tritt eine leichte
Schlierenbildung auf, es zeigt sich jedoch keine Tendenz zur Gelbildung (200.000-
800.000 Zellen/ml)
2 Deutlich positiv: Mischung verdickt sich augenblicklich, geringgradige Gelbildung,
bei kreisenden Bewegungen der Testschale strebt die Mischung zur Mitte, bei
Beendigung der Bewegung verteilt sich die Mischung wieder (800.000-1.500.000
Zellen/ml)
3 Stark positiv: Gelformung mit konvexer Oberfläche, keine Verteilung nach
Beendigung der Bewegung (> 1.500.000 Zellen/ml)
Für die Auswertung der Daten wurde in Anlehnung an die Mastitis-Definition der DVG
(2002) bereits ein einfach positives CMT-Ergebnis als eine Sekretionsstörung bzw. bei
gleichzeitigem Nachweis eines Mastitiserregers als subklinische Mastitis bewertet.
3.5 Euterpalpation Nach dem vollständigen Ausmelken der Kühe wurde jedes Euter auf Anzeichen einer
klinischen Mastitis hin untersucht. Hierbei erfolgte eine Palpation der vier Drüsenkomplexe
nach ROSENBERGER (1990). Hierbei wurde auf die Körnung des Drüsenparenchyms,

53
Knotenbildung, diffuse Verhärtungen und Schwellungen sowie Schmerzhaftigkeit geachtet.
Die Dokumentation des Euterpalpationsbefundes erfolgte nach folgendem nach
ROSENBERGER (1990) modifizierten Schema:
0 Eutergewebe insgesamt feinkörnig und weich
1 Grobkörniges bzw. -knotiges Drüsengewebe
2 Prall-derbes Eutergewebe
3 Einzelne oder mehrere Knoten
4 Eutergewebe diffus verhärtet oder akut geschwollen (vermehrt warm, schmerzhaft)
3.6 Zytobakteriologische Untersuchung der Milchproben
3.6.1 Anzüchtungsmedien Die Anzüchtung und weitere Differenzierung der Erreger fand im Labor des Departamento de
Salud Publica der Universidad de Guadalajara statt. Der Ausstrich der Milchproben erfolgte
nach spätestens 12h auf Schafblutagarplatten, zu deren Herstellung Blutagar-Basis (Merck,
Darmstadt, Deutschland) verwendet wurde (s. Anhang).
Zur besseren Differenzierbarkeit von Streptokokken wurde dem Medium 0,01% Äskulin
zugesetzt. Das Glycosid Äskulin zeigt im UV-Licht Fluoreszenz. Durch Abbau des Äskulins
in Glucose und Äskuletin geht die Fluoreszenz in der Umgebung einer Kolonie verloren.
Somit lassen sich Äskulin abbauende von Äskulin nicht verwertenden Stämmen
unterscheiden. Das Medium wurde nach den Angaben des Herstellers angefertigt, 15min bei
120˚C autoklaviert und auf ca. 50˚C abgekühlt. Nach Zugabe von 5% aseptisch
entnommenem, defibriniertem Schafblut erfolgte das Gießen des Flüssigagars in sterile
Petrischalen. Vor dem Ausstreichen auf die Nährböden wurden die Proben auf
Zimmertemperatur erwärmt und sorgfältig durchmischt. Mit einem sterilen Glasstab wurde
anschließend pro Probe etwa 0,01 ml auf ein Viertel einer Blutagarplatte ausgestrichen. Die
Platten wurden bei 37˚C unter aeroben Bedingungen inkubiert und jeweils nach 24 und 48h
abgelesen.
Zur Differenzierung der Enterobactericae wurde der McConkey-Agar (Oxoid GmbH, Wesel,
Deutschland) eingesetzt (s. Anhang). Der Nährboden wurde nach Angaben des Herstellers
angefertigt, 15min bei 120˚C autoklaviert und auf 50˚C abgekühlt. Danach erfolgte das
Gießen des Flüssigagars in sterile Petrischalen.

54
Zur Identifizierung von Escherichia coli wurde das SIM (Schwefel-Reduktion, Indol-
Produktion, Motilität)-Medium von Merck (Darmstadt, Deutschland) eingesetzt (s. Anhang).
Das Medium wurde nach Herstellerangaben angefertigt und jeweils 4 ml in verschließbare
Reagenzgläser gefüllt. Diese wurden für 20min bei 121˚C autoklaviert.
Zum Nachweis von Hefen in Proben mit über einer Millionen Zellen/ml wurden diese auf
Sabouraud-2%-Glucose-Agar (Merck, Darmstadt, Deutschland) als Selektivnährboden
ausgestrichen (s. Anhang).
Das Medium wurde nach den Angaben des Herstellers angefertigt, 15min bei 120˚C
autoklaviert und auf 50˚C abgekühlt. Danach erfolgte das Gießen des Flüssigagars in sterile
Petrischalen. Die Inkubation erfolgte für 48h bei 20˚C.
Für die Kultivierung der Bakterien in flüssigem Medium, wie dies für die
Pulsfeldgelelektrophorese erforderlich war, kam Todd-Hewitt-Bouillon (THB; Oxoid GmbH,
Wesel, Deutschland) zum Einsatz (s. Anhang).
Die Bouillon wurde nach Angaben des Herstellers angefertigt, mit 0,2% Agar (Merck,
Darmstadt, Deutschland) versetzt, zu je 40 ml in Reagenzgläser abgefüllt und für 1h bei 95˚C
im Dampftopf erhitzt. Die Bebrütung erfolgte für 18-24h bei 37˚C.
3.6.2 Subkultivierung der Bakterienkulturen Die Bakterienkulturen wurden nach Anzüchtung bei 4˚C aufbewahrt und innerhalb eines
Zeitraums von zwei Wochen in Rinderserum mit 6% Glucose überführt und anschließend bei
-20˚C bis zur weiteren Untersuchung aufbewahrt. Die Wiederanzüchtung der Isolate erfolgte
nach Auftauen auf Raumtemperatur durch Ausstreichen der Suspensionen auf Schafblut-
Äskulin-Platten nach 3.6.1 und Bebrütung für 24h bei 37˚C.
3.6.3 Identifizierung der Bakterienkulturen Die nach 3.6.1 ausgestrichenen und bebrüteten Proben wurden nach 24 und 48h auf
bakterielles Wachstum überprüft. Die Koloniemorphologie, hämolytische Charakteristika, das
Gramfärbeverhalten und das Verhalten beim Katalase-Test wurden überprüft.
Es wurden drei Kategorien für die Anzahl der gewachsenen Kolonien verwendet:
1= < 6 gleichartige Kolonien (geringgradiges Wachstum)
2= 6-15 gleichartige Kolonien (mittelgradiges Wachstum)
3= > 15 gleichartige Kolonien (hochgradiges Wachstum)

55
Als S. aureus wurden solche Kulturen angesprochen, die 1-3 mm große, glatte, unregelmäßige
cremefarbene bis goldgelbe Kolonien auf Blutagar bildeten und von typischen
Hämolysezonen umgeben wurden. Das α-Hämolysin produziert eine klare Zone vollständiger
Hämolyse, während das β-Hämolysin eine deutlich abgegrenzte Zone einer unvollständigen
Hämolyse verursacht. Selten kann auch eine abgegrenzte schmale Zone einer vollständigen
Hämolyse auftreten, die durch das δ-Hämolysin ausgelöst wird.
Die Beurteilung der Bakterienmorphologie erfolgte nach Gram-Färbung im Lichtmikroskop
bei 1000facher Vergrößerung. Staphylokokken sind grampositive Bakterien mit einem
Durchmesser von ca. 1 µm. Sie sind kugelförmig und meist in Form von traubenartigen
Haufen unregelmäßig angeordnet. Zur Abgrenzung der Katalase-positiven Staphylokokken
von den Katalase-negativen Streptokokken kam der Katalase-Test nach BRÜCKLER und
SCHWARZ (1994) zur Anwendung. Bei diesem Test wurden einige Kolonien der zu
prüfenden Kultur mit einer sterilen Öse auf einem Objektträger mit einer 3%-igen H2O2-
Lösung verrieben. Eine positive Reaktion zeigte sich in einer deutlichen Bläschenbildung. Zur
Differenzierung zwischen S. aureus und der Gruppe der KNS wurde der Röhrchen-
Koagulase-Test nach KLOOS und LAMBE (1991) durchgeführt. Zu 300µl mit Citrat
ungerinnbar gemachten Kaninchenplasmas, gewonnen aus sterilem Citratblut (1 Vol. 4%
Natriumzitrat zu 9 Vol. Blut) in einer Verdünnung von 1:5 mit einer 0,14 mol/l
Natriumchlorid-Lösung, wurden je drei Kolonien der zu untersuchenden Bakterienkulturen
gegeben. Die Beurteilung erfolgte nach einer Bebrütung von 24h bei 37˚C.
Ein Isolat wurde als S. aureus angesprochen, wenn ein vollständiges oder überwiegend
zusammenhängendes Koagulum zu erkennen war. Blieb der Inhalt flüssig, wurden die
Kolonien als Koagulase-negative Staphylokokken (KNS) angesprochen.
Als Streptokokken wurden zarte, durchscheinende Kolonien mit bis zu 1 mm Größe
angesprochen, die von keiner (γ), einer unvollständigen vergrünenden (α) oder einer
vollständigen Hämolyse (β) umgeben sind. Nach Gramfärbung zeigen sich im
Lichtmikroskop grampositive Kokken, die einen Durchmesser unter 2 µm besitzen und in
Paaren oder unterschiedlich langen Ketten angeordnet sind. Der oben beschriebene Katalase-
Test ist bei Streptokokken negativ. Unter der UV-Lampe können Äskulin-abbauende und
Äskulin nicht verwertende Kulturen anhand der Fluoreszenz um die gewachsenen Kolonien
unterschieden werden und wurden je nach Verhalten als Äskulin-positive bzw. Äskulin-
negative Streptokokken angesprochen.

56
Zur Differenzierung der Äskulin-negativen Streptokokken wurde der CAMP-Test nach
CHRISTIE et al. (1944) herangezogen. Ein S. aureus- Stamm, der eine breite unvollständige
Hämolyse (β-Hämolysin) bildet, wird quer über eine Blutagarplatte nach 3.6.1 ausgestrichen.
Die zu untersuchenden Streptokokken-Stämme werden senkrecht zum S. aureus- Stamm
ausgestrichen, wobei ein Zwischenraum von 2-3 mm bestehen bleiben soll. Eine positive
Kontrolle wird mitgeführt. Nach 24-stündiger Inkubation bei 37˚C erfolgt die Auswertung.
Ein CAMP-Phänomen liegt vor, wenn eine halbmondförmige Zone einer vollständigen
Hämolyse im Bereich der unvollständigen Hämolyse des S. aureus- Stammes vorliegt. Die
CAMP-Reaktion wird auf die synergistische Wirkung des Staphylokokken β-Toxins, einer
Sphingomyelinase, und dem B-Streptokokken CAMP-Faktor zurückgeführt. Der CAMP-
Faktor wird als thermostabiles Protein mit einem Molekulargewicht von 23.500 Dalton und
einem isoelektrischen Punkt bei pH 8,3 beschrieben (BERNHEIMER et al. 1979). Das
CAMP-Protein reagiert nicht enzymatisch mit Ceramid-Phosphat, welches nach vorheriger
Einwirkung der Staphylokokken-Sphingomyelinase C aus dem Sphingomyelin der
Erythrozytenmembran entsteht. Die Bindung des CAMP-Faktors führt zur Desorganisation
der Lipiddoppelschicht und schließlich zur Hämolyse (BERNHEIMER et al. 1979). S.
agalactiae reagiert positiv im CAMP-Test, während S. dysgalactiae keinen CAMP-Faktor
besitzt. S. agalactiae-Kolonien zeigen zumeist eine β-Hämolyse, während S. dysgalactiae
eine α-Hämolyse oder keine Hämolyse zeigt.
Zur weiteren Differenzierung der Äskulin-positiven Streptokokken in die verschiedenen
Streptokokken- und Enterokokkenspezies wurde bei einigen Stämmen das kommerziell
erhältliche api20 Strep-System (Biomérieux, Nürtingen, Deutschland) eingesetzt. Die meisten
Äskulin-positiven Streptokokken treten ohne Hämolyse auf. Nur Aerococcus viridans zeigt
eine α-Hämolyse mit deutlicher Vergrünung des Agars.
Als Corynebacterium spp. wurden Kolonien angesprochen, die erst nach 48-stündiger
Inkubation der beimpften Platten erkennbar wurden mit einem Durchmesser von maximal 1
mm. Die Kolonien befinden sich meist in dem Bereich des Ausstrichs, in dem die Milchfette
vorliegen. Es ist keine Hämolyse erkennbar. Die Kolonien besitzen eine raue Oberfläche und
wölben sich häufig nach oben. Im Gramausstrich zeigen sich grampositive pleomorphe
Stäbchen mit einer Größe von 0,5 x 2 µm. Auffällig ist häufig eine V-förmige oder parallele
Lagerung.

57
Enterobacteriaceae sind gramnegative, peritrich begeißelte Stäbchen, die keine Sporen bilden.
Sie sind fakultativ anaerob und verfügen über Katalase. In der milchhygienischen
Routinediagnostik wird zwischen E. coli und sonstigen coliformen Keimen unterschieden.
Als coliforme Keime werden 2-4 mm große gräuliche, feucht-glänzende runde Kolonien mit
scharfer Begrenzung angesprochen. Teilweise konnte ein stark schleimiges, mukoides
Wachstum oder eine deutliche Hämolyse beobachtet werden. Ein Wachstum auf Mc Conkey-
Agar nach 3.6.1 war immer vorhanden. Anhand des Farbumschlages von hellrosa zu pink
können Laktose-positive Kulturen sofort angesprochen werden. Zur Abgrenzung von E. coli
von den übrigen coliformen Keimen wurde dann die Überimpfung einiger Kolonien in ein
SIM-Medium nach 3.6.1 durchgeführt. Eine positive Indol-Reaktion wurde als Nachweis von
E. coli gewertet.
Traten große, flache, kompakte oder auch große raue Kolonien mit zahlreichen Ausläufern
auf, so wurden diese als Bacillus (B.) spp. beurteilt, wobei Bazillen in einer großen Variation
von Kulturmorphologien auftreten. Lag eine Hämolyse um die Kolonie herum vor, wurde die
Kultur als B. cereus angesprochen. Im Grampräparat zeigen sich grampositive Stäbchen mit
Endosporen. Sie sind etwa 1 x 4 µm groß.
Als Hefen wurden Kulturen angesprochen, die meist erst nach 48h ihre volle Ausbildung mit
einem Durchmesser von etwa 2 mm zeigten. Sie treten in weißen und cremefarbenen
Varianten auf, und ihre Oberfläche ist meistens rau. Nach Überimpfung auf Sabouraud-2%-
Glucose-Agar nach 3.6.1 tritt im Gegensatz zu anderen Kulturen nach Bebrütung bei 20˚C
Wachstum auf. Im Lichtmikroskop zeigen sich runde bis ovale Zellen mit einem Durchmesser
von 5-9 µm. Häufig sind Sprossung oder Hyphen erkennbar.
Die Isolation von mehr als drei unterschiedlichen Bakterienarten aus einer Milchprobe wurde
als Kontamination angesprochen. Kontaminierte Proben wurden von der Auswertung
ausgenommen. Nachweise von zwei unterschiedlichen Spezies in einer Probe wurden als ein
Mastitisfall für jedes einzelne Pathogen gewertet.

58
3.7 PCR-vermittelter Nachweis speziesspezifischer Genabschnitte
3.7.1 Präparation der bakteriellen DNA Alle Schritte der PCR wurden im PCR-Labor des Landesbetriebs Hessisches Landeslabor
Giessen durchgeführt. Zur Gewinnung der als „template“ in der PCR eingesetzten DNA kam
eine ursprünglich von BENTLEY et al. (1993) beschriebene, modifizierte Methode zum
Einsatz. Die zu untersuchenden Kulturen wurden zunächst auf Blutagar nach 3.6.1
subkultiviert und über Nacht bei 37˚C unter aeroben Bedingungen bebrütet. Nach erneuter
Reinheitskontrolle wurden dann etwa 5-10 Kolonien in 100µl TE-Puffer (s. Anhang) in 1,5 ml
Reaktionsgefäße überführt.
Bei Vorliegen von Staphylokokken-Kulturen wurde nun 5µl Lysostaphin (1,8 U/µl,Sigma,
Deisenhofen, Deutschland), bei Streptokokken-Kulturen hingegen 5µl Mutanolysin (10 U/µl;
Sigma, Deisenhofen, Deutschland) zugesetzt. Es folgte eine Inkubation im Wasserbad bei
37˚C für 1h, wodurch die Bakterienzellwände lysiert wurden. Zur Deproteinisierung erfolgte
die Zugabe von 2µl bzw. 10µl Proteinase K (14,8mg/ml, Roche, Mannheim, Deutschland)
und eine zweistündige Inkubation im Wasserbad bei 56˚C. Durch anschließendes Kochen der
Proben für 10min erfolgte die Inaktivierung der Proteinase K. Anschließend wurde die Probe
zentrifugiert (1min, 10.000 x g) und der Überstand entweder direkt in die PCR eingesetzt oder
bei -20˚C eingefroren.
3.7.2 Verwendete Oligonucleotidprimer und Thermocycler-
Temperaturprogramme
3.7.2.1 Staphylococcus aureus Zur Amplifizierung eines speziesspezifischen Abschnitts des 23 S rRNA-Gens von S. aureus
wurden die von STRAUB et al. (1999) beschriebenen Primer Staur4 mit der Sequenz 5'-ACG
GAG TTA CAA AGG ACG AC-3' und Staur6 mit der Sequenz 5'-AGC TCA GCC TTA
ACG AGT AC-3' in der PCR eingesetzt. Alle verwendeten Primer wurden von MWG-
Biotech (Ebersberg, Deutschland) bezogen.
Der Mastermix wurde nach folgendem Protokoll pipettiert:
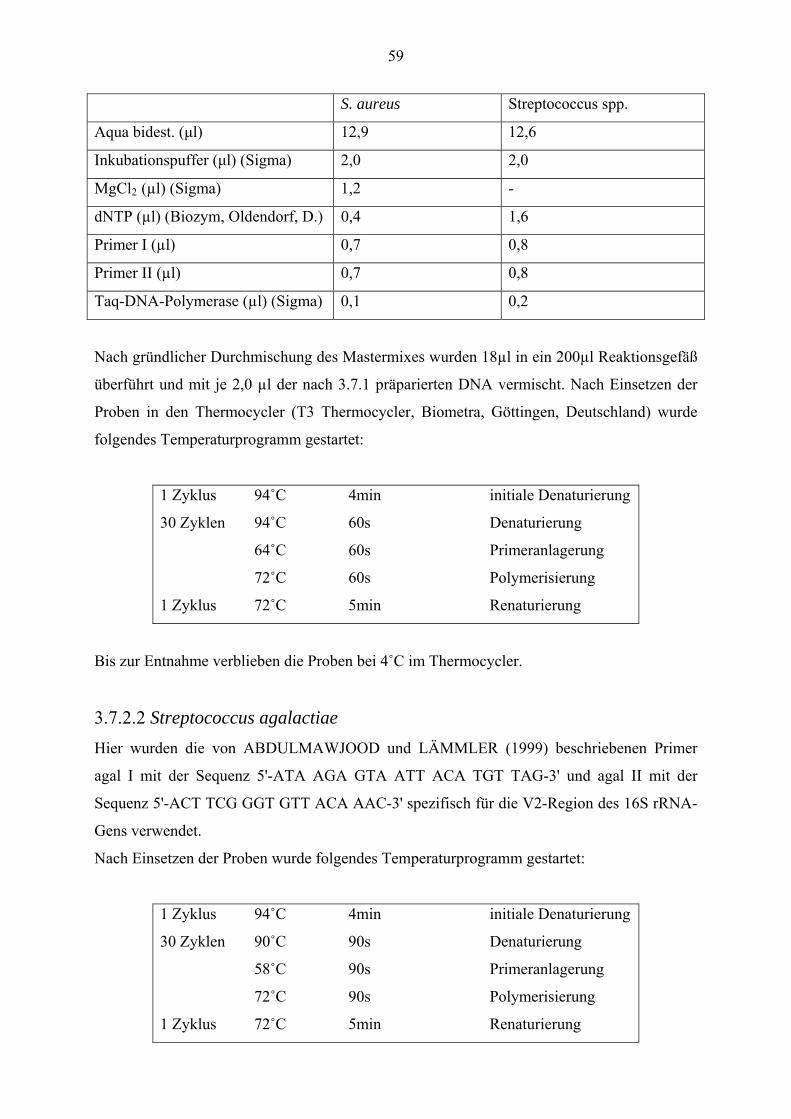
59
S. aureus Streptococcus spp.
Aqua bidest. (μl) 12,9 12,6
Inkubationspuffer (μl) (Sigma) 2,0 2,0
MgCl2 (µl) (Sigma) 1,2 -
dNTP (µl) (Biozym, Oldendorf, D.) 0,4 1,6
Primer І (µl) 0,7 0,8
Primer ІІ (µl) 0,7 0,8
Taq-DNA-Polymerase (µl) (Sigma) 0,1 0,2
Nach gründlicher Durchmischung des Mastermixes wurden 18µl in ein 200µl Reaktionsgefäß
überführt und mit je 2,0 µl der nach 3.7.1 präparierten DNA vermischt. Nach Einsetzen der
Proben in den Thermocycler (T3 Thermocycler, Biometra, Göttingen, Deutschland) wurde
folgendes Temperaturprogramm gestartet:
1 Zyklus 94˚C 4min initiale Denaturierung
30 Zyklen 94˚C 60s Denaturierung
64˚C 60s Primeranlagerung
72˚C 60s Polymerisierung
1 Zyklus 72˚C 5min Renaturierung
Bis zur Entnahme verblieben die Proben bei 4˚C im Thermocycler.
3.7.2.2 Streptococcus agalactiae Hier wurden die von ABDULMAWJOOD und LÄMMLER (1999) beschriebenen Primer
agal І mit der Sequenz 5'-ATA AGA GTA ATT ACA TGT TAG-3' und agal ІІ mit der
Sequenz 5'-ACT TCG GGT GTT ACA AAC-3' spezifisch für die V2-Region des 16S rRNA-
Gens verwendet.
Nach Einsetzen der Proben wurde folgendes Temperaturprogramm gestartet:
1 Zyklus 94˚C 4min initiale Denaturierung
30 Zyklen 90˚C 90s Denaturierung
58˚C 90s Primeranlagerung
72˚C 90s Polymerisierung
1 Zyklus 72˚C 5min Renaturierung

60
3.7.2.3 Streptococcus uberis Die Amplifizierung eines speziesspezifischen Abschnitts des 16S rRNA-Gens von S. uberis
wurde mit den von BROWN und BROWN (1995) und OKA et al. (1999) beschriebenen
Primern ub І mit der Sequenz 5'-CGCATGACTTATCA-3' und ub ІІ mit der Sequenz 5'-
GCCTTTAACTTCAGACTTATCA-3' sowie dem unter 3.7.2.2 beschriebenen
Temperaturprogramm durchgeführt.
Desweiteren wurden 17 S. aureus-Stämme von EL-SAYED et al. (im Druck) mit Hilfe der
PCR auf das Vorkommen der Gene für die S. aureus-Enterotoxine A-J (entA- entJ) und das
Toxic-shock-syndrome-toxin 1 (tst) untersucht. Der Nachweis beschränkte sich auf die
Untersuchung von jeweils einem Isolat stellvertretend für jeden ermittelten S. aureus-Betrieb.
3.7.3 Agarosegelelektrophorese Der Nachweis der Amplifikate erfolgte mittels Elektrophorese in einem 2%-igen Agarosegel.
Hierfür wurden zunächst 0,7g Agarose NEEO (Roth, Karlsruhe, Deutschland) in 35ml TBE-
Puffer (40mmol/l Tris, 1% [v/v] Essigsäure, 1mmol/l EDTA) durch mehrminütiges Kochen in
der Mikrowelle in Lösung gebracht und nach Abkühlung im Wasserbad auf etwa 56˚C in eine
Flachbettform mit eingesetzten Kämmen gegossen. Nach dem Abkühlen wurden die Kämme
entfernt, und in die entstandenen Mulden konnte jeweils 8µl des PCR-Produkts nach
Vermischung mit jeweils 3µl Probenpuffer (50% Glycerin, 50mmol/l EDTA, 0,25%
Bromphenolblau) pipettiert werden. Die elektrophoretische Auftrennung erfolgte im
Horizontalgel für 1h bei 100V, wobei als Laufpuffer ebenfalls TBE-Puffer diente. Eine
Bestimmung der Größe der PCR-Produkte ermöglichte ein ebenfalls in das Gel aufgetragener
Marker (GeneRulerTM DNA Ladder Mix, MBI Fermentas, St. Leon-Rot; 100Bp DNA Ladder,
Gibco/BRL, Eggenstein; DNA-Längenstandard VIII, Roche, Mannheim).
3.7.4 Ethidiumbromidfärbung Im Anschluss an die Elektrophorese erfolgte die Dokumentation durch Färbung des Gels für
20min in einer wässrigen Ethidiumbromid-Lösung (5μg/ml, Sigma, Deisenhofen,
Deutschland), wobei sich das fluoreszierende Ethidiumbromid als interkalierende Substanz in
die Doppelstränge der DNA einlagerte. Überschüssiges Ethidiumbromid wurde durch kurzes
Schwenken in Aqua dest. entfernt. Die Auswertung und photographische Dokumentation
erfolgte anschließend unter UV-Licht (Gel-Videodokumentationssystem Image Master, VDS,

61
Pharmacia Biotech, Freiburg, Deutschland), wobei sich die DNA-Fragmente als helle Banden
vor dunklem Hintergrund darstellten.
3.8 Makrorestriktionsanalyse der chromosomalen DNA und
Pulsfeldgelelektrophorese (DNA-Fingerprinting)
3.8.1 Präparation der bakteriellen DNA Zur Erstellung eines DNA-Fingerprints erfolgte zunächst die Präparation der bakteriellen
Gesamtzell-DNA von 27 ausgewählten Isolaten nach einer modifizierten Methode von
MASLOW et al. (1993) und THIELE et al. (1993). Aus jedem Betrieb mit positivem
Erregernachweis wurden 1-4 Isolate ausgewählt. Die auf Blutagar nach 3.6.1 angezüchteten
Reinkulturen der zu untersuchenden Bakterienisolate wurden in 40ml THB nach 3.6.1
überführt und für 24h bei 37˚C unter aeroben Bedingungen bebrütet. Anschließend wurden
die Bakteriensuspensionen für 10min bei 4.000 x g abzentrifugiert und zweimal in TE-Puffer
(10mmol/l Tris-HCl (pH 8,0), 1mmol/l EDTA) gewaschen. Das Sediment wurde daraufhin in
0,25ml TE-Puffer resuspendiert und bei einer Wellenlänge von 620nm (Spectronic 20,
Bausch und Lomb, New York) photometrisch auf 5% Transmission eingestellt. Aus der
eingestellten Suspension wurden je Kultur vier Agaroseblöckchen hergestellt. Hierzu wurde
200µl der auf 55˚C vorgewärmten Bakteriensuspension mit 205µl einer 1%-igen „low
melting, low gelling“ InCert-Agarose (Biozym Diagnostik, Hess. Oldendorf, Deutschland)
sorgfältig vermischt und in eine Gießkammer (100µl, BioRad, München, Deutschland)
pipettiert. Bis zum Erstarren der Blöckchen wurden diese auf Eis inkubiert. Der Aufschluss
der Bakterienzellwand erfolgte in 200µl Lysispuffer (6mmol/l Tris [pH 7,6], 1 mol/l NaCl,
10mmol/l EDTA [pH 7,6], 0,5% Brij-58 (Polyoxyethylene-20-Cetyl-Ether (Sigma,
Deisendorf, Deutschland), 0,2% Natriumdodecylsulfat (Sigma), 0,5% NatriumIauroylsarcosin
(Sigma)) unter Zusatz von 5mg/ml Lysozym (Sigma) und 5 U/ml Lysostaphin (Sigma) über
Nacht bei 37˚C. Eine anschließende Deproteinisierung erfolgte durch Zusatz von Proteinase K
(Endkonzentration 0,5 μg/ml; Boehringer, Mannheim, Deutschland) und eine Inkubation im
Wasserbad bei 56˚C über Nacht. Nach zweimaligem Waschen der Blöckchen mit 200µl TE-
Puffer und jeweils 30- minütiger Inkubation bei Raumtemperatur sowie zweimaligem
Waschen mit 200µl TE-Puffer unter Zusatz von Phenylmethylsulfonylfluorid
(Endkonzentration 1,0mmol/l; Sigma) und einer Inkubation für 1h bei 56˚C zur Inaktivierung
der Proteinase K schlossen sich nochmals zwei Waschschritte mit jeweils 200µl TE-Puffer
und eine Inkubation bei Raumtemperatur für 30min an. Die Blöckchen konnten nun mit den

62
Restriktionsenzymen verdaut werden oder verblieben bis zur weiteren Verwendung bei 4˚C in
TE-Puffer im Kühlschrank.
3.8.2 Restriktionsverdau der Gesamtzell-DNA
Der Restriktionsverdau der nach 3.8.1 präparierten chromosomalen DNA erfolgte mit Hilfe
der Restriktionsendonucelease SmaI (Promega, Mannheim, Deutschland). Der
Restriktionsansatz setzte sich wie folgt zusammen (Ansatz pro Blöckchen):
Aqua dest. 158µl
Puffer J (Promega) 20µl
Bovines Serumalbumin (10 x BSA, Promega) 20µl
SmaI 2 µl
Die Agaroseblöckchen wurden für 5h bei 25˚C inkubiert.
3.8.3 Laufbedingungen der Pulsfeldgelelektrophorese (PFGE) Zur Auftrennung der Fragmente der nach 3.8.1 präparierten und nach 3.8.2 geschnittenen
DNA mittels PFGE wurden die Agaroseblöckchen in ein 1%-iges Agarosegel (1,1g Agarose
in 110ml 0,5x TBE-Puffer (45mmol/l Tris, 45mmol/l Borat, 1,0mmol/l EDTA (pH 8,0))
eingebettet. Nach Verbringen der Blöckchen in die Vertiefungen des Gels und Verschließen
der Vertiefungen mit flüssigem 1%-igem Agarosegel erfolgte die Elektrophorese in 2 Litern
0,5xTBE-Puffer in einem Chef DR ІІ-Elekrophoresegerät (Bio-Rad, München, Deutschland)
bei 14˚C unter folgenden Bedingungen:
1. Lauf 2. Lauf
Spannung 5 V/cm 6 V/cm
Anfangsumschaltzeit 0,1s 9,0s
Endumschaltzeit 11,0s 40,0s
Dauer 8h 17h
Als DNA-Längenstandard diente λ-ladder PFGE Marker (Bio-Rad).
Anschließend folgte das Färben und die Dokumentation des Gels wie in unter 3.7.4
beschrieben.

63
3.8.4 Auswertung der PFGE-Muster Die durch die PFGE erhaltenen Bandenmuster wurden von zwei unabhängigen Personen
visuell ausgewertet. In Anlehnung an TENOVER et al. (1995) wurden Isolate als nicht
unterscheidbar und zum identischen Makrorestriktionstyp gehörend angesprochen, wenn ihre
Bandenmuster vollständig übereinstimmten. Hatten zwei Muster die gleiche Anzahl von
Banden und unterschieden sich nur in einer Bande wurde dies als ein Verlust bzw. eine
Aufnahme von DNA gewertet. Isolate mit solchen Bandenmustern wurden als Subklone des
entsprechenden PFGE-Genotyps eingestuft. Gleiches galt für Isolate, die infolge der
Aufnahme bzw. des Verlustes einer Schnittstelle für das Restriktionsenzym in ihrem
Bandenmuster einen Unterschied von zwei bis drei Banden aufwiesen. Sofern Isolate, die als
Subklone eines PFGE-Genotyps eingestuft waren, in epidemiologischem Zusammenhang
standen, d.h. aus einer Herde stammten, wurden die Muster als identisch gewertet. Bei
größerem Unterschied der Bandenmuster wurden die Isolate als unterschiedlich klassifiziert
und in verschiedene PFGE-Genotypen eingeordnet.
3.9 Statistische Methoden Die Datenauswertung erfolgte auf den Rechnern im lokalen Rechnernetzwerk (LAN) der
Arbeitsgruppe Biomathematik und Datenverarbeitung des Fachbereichs Veterinärmedizin der
Justus-Liebig-Universität Giessen. Die statistischen Auswertungen wurden unter Verwendung
des Statistikprogrammpakets BMDP/Dynamic, Release 7.0, (DIXON 1993) durchgeführt.
Bei der Bewertung der statistischen Signifikanzen wurde das Signifikanzniveau α = 0,05
zugrunde gelegt, d.h. Ergebnisse mit p≤ 0,05 wurden als statistisch signifikant angesehen. Die
Untersuchung der Zusammenhänge erfolgte bei quantitiativen Merkmalen mit Hilfe von
Korrelations- bzw. Regressionsanalysen mit dem Programm BMDP6D unter Angabe des
Korrelationskoeffizienten (r) und der Regressionsgeraden (y = m·x + b) bzw. mit dem
Rangkorrelationskoeffizienten nach Spearman. Die statistischen Zusammenhänge wurden mit
dem Fisher-Test/Chi-Quadrat-Test überprüft.

64
4 Ergebnisse
4.1 Beschreibung der besuchten Betriebe Auf den 33 besuchten Betrieben mit insgesamt 2205 Milchkühen wurden zu 95% Tiere der
Rasse Holstein Friesian gehalten. Die übrigen 5% setzten sich aus den Rassen Jersey, Brown
Swiss, Criollo und Simmental zusammen. In Übereinstimmung mit den Witterungsverhältnissen muss man in Mexiko das Jahr in die
Regen- und Trockenzeit einteilen. Die Trockenzeit reicht von November bis Mai, während die
Regenzeit von Juni bis Oktober andauert. Von den 33 Betrieben wurden 23 (69,7%) zum
Zeitpunkt der Trockenzeit und 10 (30,3%) in der Regenzeit besucht.
25 (75,8%) der 33 Betriebe halten ihre Tiere halbaufgestallt in eingezäunten Ausläufen mit
provisorischen Überdachungen zum Sonnenschutz. Zusätzlich zu dieser Haltung gewähren
fünf von diesen 25 Betrieben (20%) ihren Tieren tagsüber Weidegang, fünf (20%) Betriebe
lassen ihre Tiere nur während der Regenzeit weiden und 15 (60%) halten ihre Tiere ohne
Weidegang.
Acht (24,2%) der 33 Betriebe halten ihre Tiere das ganze Jahr auf der Weide und treiben diese
nur zur Melkzeit in die provisorischen Melkstände.
Zum Reinigungsintervall ihrer Ausläufe machten die 25 betroffenen Betriebsleiter folgende
Angaben:
Keinerlei Reinigung (2; 8%), einmal im Jahr (1; 4%), monatlich (1; 4%), alle 14 Tage (3;
12%), einmal pro Woche (8; 32%), zweimal pro Woche (3, 12%) oder alle 2 Tage (4; 16%).
Keine Angaben machten drei Betriebsleiter (12%).
Die Sauberkeit der Ausläufe wurde bei Besichtigung der Betriebe wie folgt bewertet:
Tab. 6 Einteilung der 25 Betriebe mit Auslaufhaltung nach der Sauberkeit ihrer Auläufe
Hygienischer Zustand der Ausläufe Anzahl Betriebe Prozentualer Anteil
Sehr gut 0 0%
Gut 10 40%
Mäßig 13 52%
Schlecht 2 8%
In der Regenzeit sind insbesondere die Haupttriebwege der Tiere extrem matschig, so dass die
Kühe teilweise bis zu den Karpalgelenken im Matsch stehen, und damit die Zitzen einer

65
möglichen Kontamination ausgesetzt sind. Häufig verfügen die Ausläufe über keine
geeigneten Vorrichtungen zum Ablaufen des Regenwassers. In der Sommerzeit besteht der
Bodenbelag vieler Ausläufe überwiegend aus eingetrocknetem Kot. Auf einigen Ausläufen
stehen den Tieren keine Dächer zum Schutz vor der Sonnenbestrahlung zur Verfügung.
Beispiele zur Auslaufsauberkeit zeigen die Abb. 7 und 8 auf S. III des Anhangs.
Auf die Frage nach gehäuft auftretenden gesundheitlichen Problemen nannten 15 Betrieben
(45,5%) Aborte. Fünf (15,2%) der Betriebsleiter gaben bei der Befragung an Probleme mit
verlängerten Zwischenkalbezeiten zu haben. Ferner wurden folgende Erkrankungen
angegeben, die hier entsprechend der Häufigkeit ihres Auftretens genannt werden:
Piroplasmose, Anaplasmose, Leptospirose, Retentio secundinarum, Pneumonien, Tuberkulose
und Brucellose. Das Auftreten von klinischen Mastitisfällen beurteilten 17 (51,5%) der
Betriebsleiter als selten, 13 (39,4%) als häufig und drei (9,1%) als sehr häufig. In 20 der
Betriebe (60,6%) kommt die künstliche Besamung zum Einsatz. Die übrigen Betriebe halten
meist ihren eigenen Bullen, der die Kühe im Natursprung deckt.
4.2 Beurteilung der Fütterung Der Ernährungszustand der Tiere wurde in sieben (21,2%) Herden mit sehr gut, bei 16
(48,5%) Betrieben als gut und bei vier (12,1%) Betrieben als mäßig beurteilt. Der
Ernährungszustand der Tiere der übrigen sechs (18,2%) Betriebe schwankte stark innerhalb
der einzelnen Herden, auch kümmernde Tiere kamen vor.
Tab. 7 Einteilung der Betriebe nach Art der Rationsberechnung der einzelnen Milchkühe
Art der Rationsberechnung Anzahl der Betriebe Prozentualer Anteil
Keinerlei Berechnung 18 54,5
Grob geschätzt 12 36,4
Exakte Berechnung 3 9,1
Bei den auf der Weide gehaltenen Tieren haben die Kühe generell nur während der Melkzeit
die Möglichkeit zur Kraftfutteraufnahme. Die Tiere werden zum Melken in den
Fangfressgittern eingesperrt und während des Melkvorgangs gefüttert.

66
Auch bei den in Ausläufen gehaltenen Tieren erfolgt die Kraftfuttergabe häufig während des
Melkens in den Fangfressgittern. Nur in den spezialisierten Betrieben haben die Tiere
melkzeitunabhängig Zugang zu Kraftfutter.
Zur Fütterung der Tiere kommen neben den Spezialkraftfuttermischungen für Milchkühe auch
Soja, Mais, Baumwollsamen, Weizenspelzen, Zuckerrohr, Brauereiabfälle, Luzerne und Hafer
zum Einsatz. Als Grundfutter werden häufig Maissilage oder –stroh und Leguminosen
vorgelegt. Die Kraftfuttermenge liegt zwischen 5-12 kg/Kuh/Tag. Gelegentlich wird den
Tieren Mineralfutter zugefüttert.
4.3 Beurteilung der Melkhygiene und des Trockenstellens Zum Zeitpunkt des Melkvorgangs sind nur bei drei (9,1%) Betrieben die Melkplätze als
sauber einzustufen. Die übrigen melken ihre Tiere in provisorischen Melkständen, die meist
keinen befestigten und leicht zu reinigenden Boden besitzen. Meist findet man einen feuchten
Boden mit einer Schicht aus Futterresten, Einstreu und Dung vor. Der während des Melkens
anfallende Kot wird meist nur grob entfernt. Eine den Melkvorgang abschließende Reinigung
bzw. Desinfektion des Melkbereiches findet gar nicht bzw. nur sporadisch statt. Eine extreme
Belästigung mit Fliegen war in vielen Melkständen eine direkte Folge dieser
Bodenverschmutzung. Die meisten Betriebe melken ihre Tiere in den Fangfressgittern
während der Kraftfuttergabe.
Tab. 8 Zustand der Melkzeuge in den 26 maschinenmelkenden Betrieben
Zustand der Melkzeuge Anzahl der Betriebe Prozentualer Anteil
Sehr gut 6 23,1
Gut 13 50,0
Mäßig 3 11,5
Schlecht 4 15,4
Die Betriebe melken mit 2-12 Melkzeugen. Auf jedes Melkzeug kommen zwischen acht und
43 Kühe/Melkzeit, durchschnittlich sind es 15 Kühe pro Melkzeug/Melkzeit. Die
Zitzengummis werden nach Angaben der Besitzer in neun (34,6%) Betrieben alle 3-4 Monate,
in sechs (23,1%) Betrieben alle 6 Monate, in fünf (19,2%) Betrieben alle 10-12 Monate und in
sechs (23,1%) Betrieben erst, wenn die Gummis defekt und brüchig sind, gewechselt.
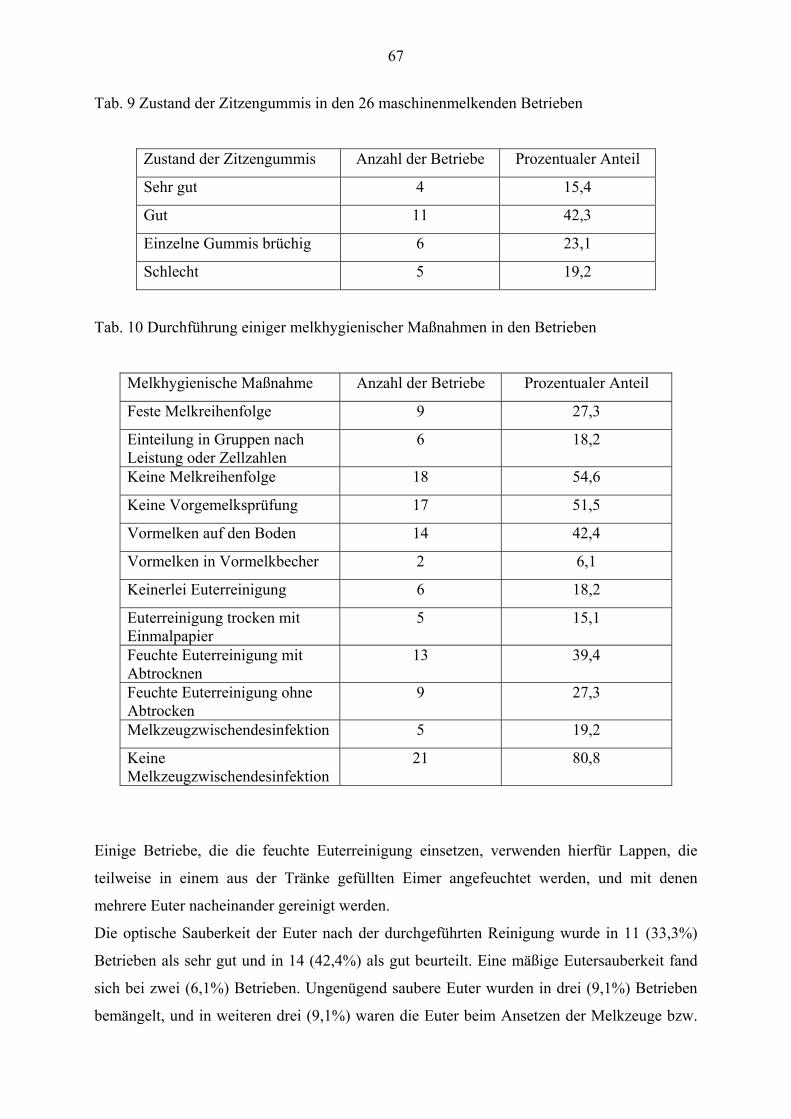
67
Tab. 9 Zustand der Zitzengummis in den 26 maschinenmelkenden Betrieben
Zustand der Zitzengummis Anzahl der Betriebe Prozentualer Anteil
Sehr gut 4 15,4
Gut 11 42,3
Einzelne Gummis brüchig 6 23,1
Schlecht 5 19,2
Tab. 10 Durchführung einiger melkhygienischer Maßnahmen in den Betrieben
Melkhygienische Maßnahme Anzahl der Betriebe Prozentualer Anteil
Feste Melkreihenfolge 9 27,3
Einteilung in Gruppen nach Leistung oder Zellzahlen
6 18,2
Keine Melkreihenfolge 18 54,6
Keine Vorgemelksprüfung 17 51,5
Vormelken auf den Boden 14 42,4
Vormelken in Vormelkbecher 2 6,1
Keinerlei Euterreinigung 6 18,2
Euterreinigung trocken mit Einmalpapier
5 15,1
Feuchte Euterreinigung mit Abtrocknen
13 39,4
Feuchte Euterreinigung ohne Abtrocken
9 27,3
Melkzeugzwischendesinfektion 5 19,2
Keine Melkzeugzwischendesinfektion
21 80,8
Einige Betriebe, die die feuchte Euterreinigung einsetzen, verwenden hierfür Lappen, die
teilweise in einem aus der Tränke gefüllten Eimer angefeuchtet werden, und mit denen
mehrere Euter nacheinander gereinigt werden.
Die optische Sauberkeit der Euter nach der durchgeführten Reinigung wurde in 11 (33,3%)
Betrieben als sehr gut und in 14 (42,4%) als gut beurteilt. Eine mäßige Eutersauberkeit fand
sich bei zwei (6,1%) Betrieben. Ungenügend saubere Euter wurden in drei (9,1%) Betrieben
bemängelt, und in weiteren drei (9,1%) waren die Euter beim Ansetzen der Melkzeuge bzw.

68
vor dem Handmelken noch nass. In einigen Betrieben wurden die Euter nach dem
Maschinenmelken mit der Hand nachgemolken.
In den sieben handmelkenden Betrieben erfolgte das Melken meist mit Vollhandgriff. Hierfür
wurden die Zitzen mit der Milch anderer Kühe aus dem Melkeimer schlüpfrig gemacht.
Andere Melker verwendeten hierfür Melkfett, wobei sie ihre Hände vor Entnahme des Fettes
nicht wuschen. Das Ansetzen der Melkzeuge wurde in allen Betrieben ohne Fehler
durchgeführt. Ein Blindmelken konnte in fünf (19,2%) der Fällen festgestellt werden.
Das Zitzentauchen mit jodhaltigen Lösungen nach dem Melken gehörte bei 21 (63,6%) der
Betriebe zum festen Bestandteil des Melkens, von diesen tauchen acht (38,1%) Betriebe die
Zitzen zusätzlich noch vor dem Melken mit einem nicht jodhaltigen Desinfektionsmittel. Die
übrigen 12 (36,4%) Betriebe dippen die Zitzen weder vor noch nach dem Melken.
Ein Milchtank für die Kühlung und Aufbewahrung der Milch war in 20 (60,6%) Betrieben
vorhanden. Die übrigen 13 (39,4%) Betriebe hatten keinen eigenen Milchtank. Hier wurde die
Milch entweder von den Melkern sofort zu Sammelkühltanks oder direkt zu
Zwischenhändlern gefahren oder bis zur Abholung durch die Molkerei provisorisch im
Wasserbad gekühlt.
Der durchschnittlich pro Liter erzielte Milchpreis lag bei 3,12 mexikanische Pesos mit
Schwankungen zwischen 2,2 und 5,5 Pesos. Die höheren Preise werden beim Direktverkauf
von Rohmilch erzielt. Ein Liter pasteurisierte Milch kostet im Supermarkt etwa 8 Pesos.
Sechzehn (48,5%) der Betriebe stellen ihre Kühe trocken, indem sie Melkzeiten überspringen
und die Futtermenge radikal reduzieren. Das derartige Trockenstellen nimmt 1-2 Wochen in
Anspruch. Von diesen 16 Betrieben verwenden sieben (43,8%) einen antibiotischen
Trockensteller. Von einer Melkzeit auf die folgende stellen 17 (51,5%) der Betriebe ihre Kühe
trocken. In 11 (64,7%) dieser 17 Betriebe kommen antibiotische Trockensteller zum Einsatz.
4.4 Ergebnisse des California-Mastitis-Test (CMT) Von den 752 untersuchten Kühen wurden 2937 Euterviertel mit Hilfe des CMT untersucht.
Insgesamt zeigten 1996 (66,9%) eine positive Reaktion, wobei 1087 (37%) deutlich bis stark
positiv reagierten, was einem Zellgehalt von > 500.000 Zellen/ml entspricht. Tab. 11 zeigt die
Resultate der 2937 durchgeführten Tests. Der geometrische Mittelwert der durchgeführten
Tests lag bei 1,19.

69
Tab. 11 Resultate des CMT von 2937 untersuchten Viertelgemelksproben
Stufe Anzahl Prozentualer Anteil
0 972 33,1%
1 878 29,9%
2 706 24,0%
3 381 13,0%
Bei 105 (3,5%) der Viertel konnte anhand makroskopischer Veränderungen der Milch bzw.
akut entzündlicher Veränderungen am Euter die Diagnose klinische Mastitis gestellt werden.
Neunundzwanzig (1%) Viertel waren verödet, so dass weder der CMT durchgeführt, noch
eine Milchprobe für die bakteriologische Untersuchung entnommen werden konnte. Der
Verdacht auf eine subklinische Mastitis wurde bei einem mindestens einfach positiven CMT-
Ergebnis bei gleichzeitig unverändertem Viertelgemelk und unauffälligem Palpationsbefund
ausgesprochen.
4.5 Befunde der Euterpalpation und Sekretbeurteilung Von den 3008 untersuchten Eutervierteln konnten 448 (14,9%) wegen zu erheblicher Störung
des normalen Melkablaufes oder aus Sicherheitsgründen nicht palpiert werden. Ein
physiologischer Palpationsbefund konnte bei 1929 (75,4%) der untersuchten 2560 Viertel
erhoben werden. Bei 154 (6,0%) der Viertel erschien das Drüsenparenchym grobkörnig.
Neunundneunzig (3,9%) Viertel waren von prall-elastischer Konsistenz, so dass die
durchgängige Palpation des kompletten Drüsenparenchyms nicht möglich war. In 289
(11,3%) der Viertel konnten kleinere bis große derbe Knoten ermittelt werden, während 89
(3,5%) Viertel akut entzündlich geschwollen oder komplett verhärtet erschienen.
Bei 2925 (98,2 %) Milchproben konnten keinerlei makroskopisch erkennbare
Sekretionsstörungen festgestellt werden. Deutliche Flockenbildung zeigten 21 (0,7%) der
Proben, eine deutliche Gelbfärbung 33 (1,1%) Proben.
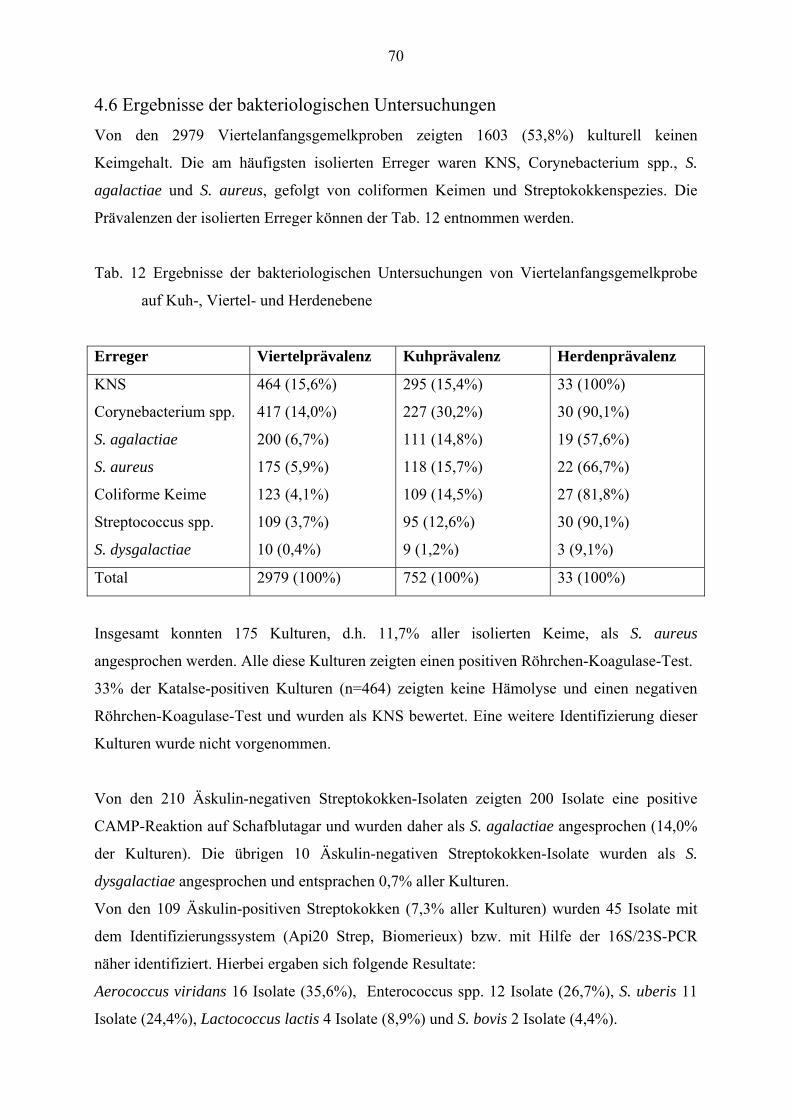
70
4.6 Ergebnisse der bakteriologischen Untersuchungen
Von den 2979 Viertelanfangsgemelkproben zeigten 1603 (53,8%) kulturell keinen
Keimgehalt. Die am häufigsten isolierten Erreger waren KNS, Corynebacterium spp., S.
agalactiae und S. aureus, gefolgt von coliformen Keimen und Streptokokkenspezies. Die
Prävalenzen der isolierten Erreger können der Tab. 12 entnommen werden.
Tab. 12 Ergebnisse der bakteriologischen Untersuchungen von Viertelanfangsgemelkprobe
auf Kuh-, Viertel- und Herdenebene
Erreger Viertelprävalenz Kuhprävalenz Herdenprävalenz
KNS
Corynebacterium spp.
S. agalactiae
S. aureus
Coliforme Keime
Streptococcus spp.
S. dysgalactiae
464 (15,6%)
417 (14,0%)
200 (6,7%)
175 (5,9%)
123 (4,1%)
109 (3,7%)
10 (0,4%)
295 (15,4%)
227 (30,2%)
111 (14,8%)
118 (15,7%)
109 (14,5%)
95 (12,6%)
9 (1,2%)
33 (100%)
30 (90,1%)
19 (57,6%)
22 (66,7%)
27 (81,8%)
30 (90,1%)
3 (9,1%)
Total 2979 (100%) 752 (100%) 33 (100%)
Insgesamt konnten 175 Kulturen, d.h. 11,7% aller isolierten Keime, als S. aureus
angesprochen werden. Alle diese Kulturen zeigten einen positiven Röhrchen-Koagulase-Test.
33% der Katalse-positiven Kulturen (n=464) zeigten keine Hämolyse und einen negativen
Röhrchen-Koagulase-Test und wurden als KNS bewertet. Eine weitere Identifizierung dieser
Kulturen wurde nicht vorgenommen.
Von den 210 Äskulin-negativen Streptokokken-Isolaten zeigten 200 Isolate eine positive
CAMP-Reaktion auf Schafblutagar und wurden daher als S. agalactiae angesprochen (14,0%
der Kulturen). Die übrigen 10 Äskulin-negativen Streptokokken-Isolate wurden als S.
dysgalactiae angesprochen und entsprachen 0,7% aller Kulturen.
Von den 109 Äskulin-positiven Streptokokken (7,3% aller Kulturen) wurden 45 Isolate mit
dem Identifizierungssystem (Api20 Strep, Biomerieux) bzw. mit Hilfe der 16S/23S-PCR
näher identifiziert. Hierbei ergaben sich folgende Resultate:
Aerococcus viridans 16 Isolate (35,6%), Enterococcus spp. 12 Isolate (26,7%), S. uberis 11
Isolate (24,4%), Lactococcus lactis 4 Isolate (8,9%) und S. bovis 2 Isolate (4,4%).

71
4.7 Ergebnisse der molekularbiologischen Untersuchungen
Die Amplifizierung des speziesspezifischen Bereichs des 23S rRNA-Gens (siehe 3.7.2.1) mit
den Oligonukleotidprimern Staur4 und Staur6 ergab bei allen 35 untersuchten koagulase-
positiven Staphylokokken-Kulturen ein einheitliches Amplifikat mit der Größe von 1270
Basenpaaren (Bp). Bei dem als Negativkontrolle mitgeführten S. epidermidis-Stamm konnte
unter Verwendung dieser Primer kein Amplifikat erzeugt werden. Es konnte somit mittels der
molekularbiologischen Technik der Nachweis erbracht werden, dass es sich bei den
untersuchten koagulase-positiven Staphylokokken um S. aureus-Isolate handelt. Die
zugehörigeAbbildung (Abb.13) befindet sich auf S. X des Anhangs.
Das Amplifikat der nach 3.7.2.2 durchgeführten Untersuchung des 16 S rRNA-Gens von 40
CAMP-Faktor positiven, Äskulin-negativen Streptokokken-Kulturen wies eine einheitliche
Größe von 1250 Bp auf. Der als Negativ-Kontrolle mitgeführte S. canis- Stamm ergab kein
Amplifikat. Hierdurch konnte der molekularbiologische Nachweis erbracht werden, dass es
sich bei den CAMP-Faktor positiven, Äskulin-negativen Streptokokken um S. agalactiae-
Isolate handelt. Die zugehörige Abbildung (Abb.14) befindet sich auf S. XI des Anhangs.
Die Amplifizierung des speziesspezifischen Bereichs des 16S rRNA-Gens von 25 Äskulin-
positiven Streptokokken-Kulturen mit den Oligonukleotidprimern ubІ und ubІІ (siehe 3.7.2.3)
ergab bei 11 der untersuchten Kulturen ein Amplifikat mit der einheitlichen Größe von 445
Bp. Hierdurch konnte molekularbiologisch nachgewiesen werden, dass es sich bei diesen
positiven Kulturen um Isolate von S. uberis handelt. Die übrigen Kulturen ohne Amplifikat
gehörten anderen Streptokokkenspezies wie z.B. Enterococcus faecalis oder S. bovis an.
Unter Verwendung der Primer ubІ und ubІІ ließ sich keine Amplifizierung der
entsprechenden Größe darstellen. Die entsprechende Abbildung (Abb. 15) befindet sich auf S.
XI des Anhangs.
EL-SAYED et al. (im Druck) konnten unter den 17 mittels Multiplex-PCR auf SE-Toxingene
untersuchten S. aureus-Stämmen nur einen einzigen Stamm (5,9%) mit einem 580 Bp
Amplifikat für das SEI Gen nachweisen. Bei keinem der untersuchten Stämme konnten Gene
für die SEA, SEB, SEC, SED, SEE, SEG, SEJ und das TSST-1 gefunden werden.
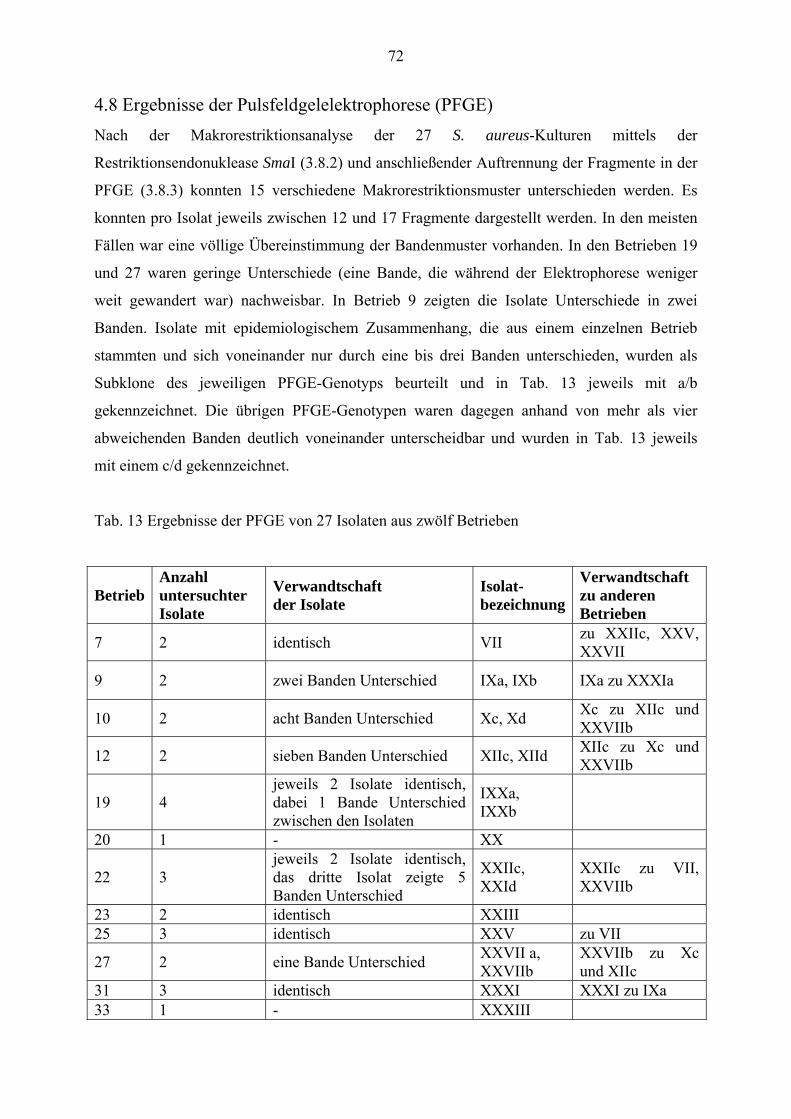
72
4.8 Ergebnisse der Pulsfeldgelelektrophorese (PFGE)
Nach der Makrorestriktionsanalyse der 27 S. aureus-Kulturen mittels der
Restriktionsendonuklease SmaІ (3.8.2) und anschließender Auftrennung der Fragmente in der
PFGE (3.8.3) konnten 15 verschiedene Makrorestriktionsmuster unterschieden werden. Es
konnten pro Isolat jeweils zwischen 12 und 17 Fragmente dargestellt werden. In den meisten
Fällen war eine völlige Übereinstimmung der Bandenmuster vorhanden. In den Betrieben 19
und 27 waren geringe Unterschiede (eine Bande, die während der Elektrophorese weniger
weit gewandert war) nachweisbar. In Betrieb 9 zeigten die Isolate Unterschiede in zwei
Banden. Isolate mit epidemiologischem Zusammenhang, die aus einem einzelnen Betrieb
stammten und sich voneinander nur durch eine bis drei Banden unterschieden, wurden als
Subklone des jeweiligen PFGE-Genotyps beurteilt und in Tab. 13 jeweils mit a/b
gekennzeichnet. Die übrigen PFGE-Genotypen waren dagegen anhand von mehr als vier
abweichenden Banden deutlich voneinander unterscheidbar und wurden in Tab. 13 jeweils
mit einem c/d gekennzeichnet.
Tab. 13 Ergebnisse der PFGE von 27 Isolaten aus zwölf Betrieben
Betrieb Anzahl untersuchter Isolate
Verwandtschaft der Isolate
Isolat- bezeichnung
Verwandtschaft zu anderen Betrieben
7 2 identisch VII zu XXIIc, XXV, XXVII
9 2 zwei Banden Unterschied IXa, IXb IXa zu XXXIa
10 2 acht Banden Unterschied Xc, Xd Xc zu XIIc und XXVIIb
12 2 sieben Banden Unterschied XIIc, XIId XIIc zu Xc und XXVIIb
19 4 jeweils 2 Isolate identisch, dabei 1 Bande Unterschied zwischen den Isolaten
IXXa, IXXb
20 1 - XX
22 3 jeweils 2 Isolate identisch, das dritte Isolat zeigte 5 Banden Unterschied
XXIIc, XXId
XXIIc zu VII, XXVIIb
23 2 identisch XXIII 25 3 identisch XXV zu VII
27 2 eine Bande Unterschied XXVII a, XXVIIb
XXVIIb zu Xc und XIIc
31 3 identisch XXXI XXXI zu IXa 33 1 - XXXIII

73
Interessanterweise unterschieden sich viele der Makrorestriktionstypen zwischen den
untersuchten Betrieben nur durch 1-3 Banden, wodurch auf eine starke Verwandtschaft dieser
Isolate untereinander geschlossen werden kann. So unterscheidet sich Isolat VII von Isolat
XXV und XXVIIc nur durch jeweils eine einzige Bande und von Isolat XXIIa durch zwei
Banden. Isolat Xc und XIIc scheinen identisch und unterscheiden sich von Isolat XXVIIb nur
durch eine einzige Bande. Auch die Isolate IXa und XXXI zeigten bei Vergleich ihrer
Bandenmuster nur in einer Bande einen Unterschied.
4.9 Ergebnisse der statistischen Auswertung Die einfaktorielle Auswertung der erhobenen Daten zeigte statistisch signifikante
Unterschiede bei der Vorkommenshäufigkeit der einzelnen Mastitiserreger in Bezug auf
Betriebsformen oder einzelne melkhygienische Maßnahmen.
So konnte S. aureus in den spezialisierten Betrieben mit einer Prävalenz von 1,3%
nachgewiesen werden, während S. agalactiae in gar keinem spezialisierten Betrieb isoliert
werden konnte. In den halbspezialisierten Betrieben stiegen die Nachweisraten auf jeweils
6,7%. Die höchste Prävalenz für beide Erreger konnte in den Familienbetrieben mit 7,4% für
S. aureus bzw. 12,7% für S. agalactiae gefunden werden. Diese Unterschiede waren für beide
Erreger hoch signifikant (p<0,0001). Da überwiegend die Familienbetriebe auch Rohmilch an
Privatpersonen verkaufen, kam S. aureus in Betrieben mit Rohmilchabgabe mit 10,4% fast
dreimal häufiger vor als in Betrieben, die ihre Milch direkt an die Molkereien liefern (3,1%).
Für S. agalactiae liegen die Werte bei 9,7% gegenüber 4,9% (p<0,0001).
In Bezug zu den verschiedenen Melksystemen, konnten S. aureus und S. agalactiae bei
handmelkenden Betrieben mit jeweils 4,8%, bei festen Eimermelkanlagen mit 8,2% bzw.
22,4%, bei mobilen Eimermelkanlagen zu 2,1% bzw. 5,8% und bei Rohrmelkanlagen mit
6,2% bzw. 4% nachgewiesen werden (p=0,0022 und p<0,0001).
Auch das Vormelken hatte einen signifikanten (p=0,0093 bzw. p<0,0001) Einfluss auf die
Vorkommenshäufigkeit der kontagiösen Mastitiserreger. Nicht vormelkende Betriebe zeigten
mit 7,0 bzw. 9,5% eine höhere Prävalenz für S. aureus bzw. S. agalactiae als Betriebe, die auf
den Boden vormelken (4,9% bzw. 4,0%). Die niedrigste Prävalenz zeigte sich beim
Vormelken mit Verwendung eines Vormelkbechers (2,9% bzw. 1,3%).
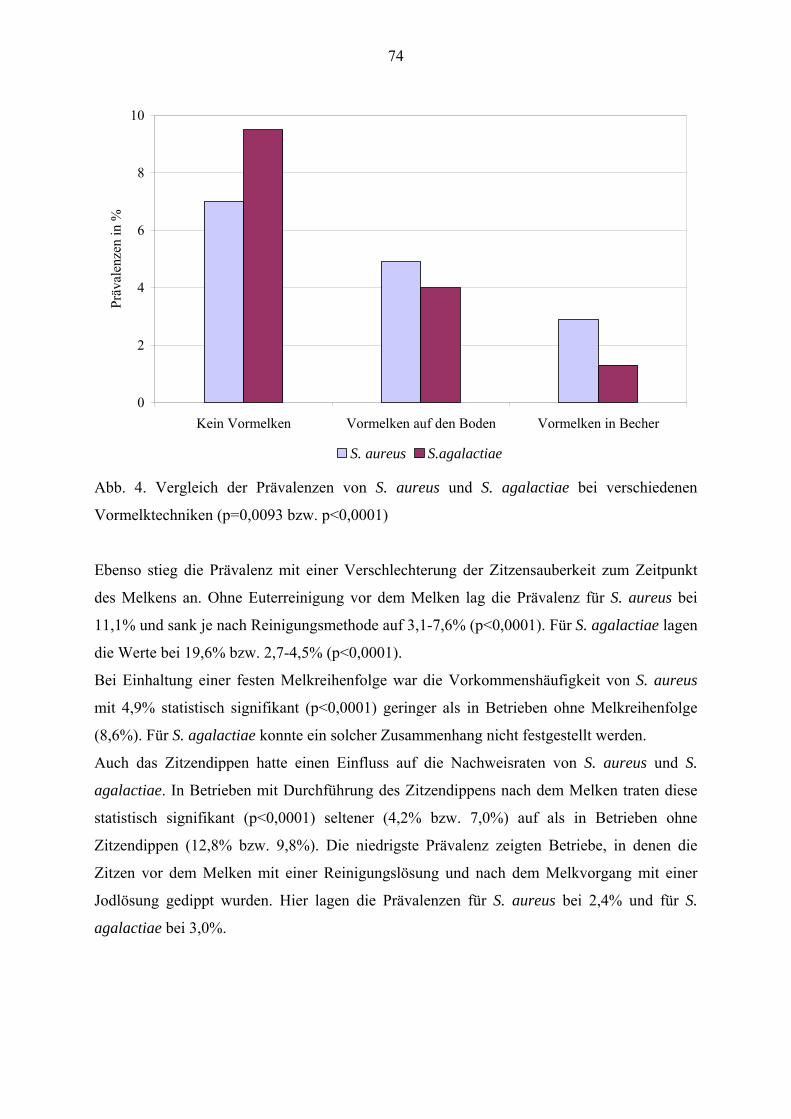
74
0
2
4
6
8
10
Kein Vormelken Vormelken auf den Boden Vormelken in Becher
Präv
alen
zen
in %
S. aureus S.agalactiae Abb. 4. Vergleich der Prävalenzen von S. aureus und S. agalactiae bei verschiedenen
Vormelktechniken (p=0,0093 bzw. p<0,0001)
Ebenso stieg die Prävalenz mit einer Verschlechterung der Zitzensauberkeit zum Zeitpunkt
des Melkens an. Ohne Euterreinigung vor dem Melken lag die Prävalenz für S. aureus bei
11,1% und sank je nach Reinigungsmethode auf 3,1-7,6% (p<0,0001). Für S. agalactiae lagen
die Werte bei 19,6% bzw. 2,7-4,5% (p<0,0001).
Bei Einhaltung einer festen Melkreihenfolge war die Vorkommenshäufigkeit von S. aureus
mit 4,9% statistisch signifikant (p<0,0001) geringer als in Betrieben ohne Melkreihenfolge
(8,6%). Für S. agalactiae konnte ein solcher Zusammenhang nicht festgestellt werden.
Auch das Zitzendippen hatte einen Einfluss auf die Nachweisraten von S. aureus und S.
agalactiae. In Betrieben mit Durchführung des Zitzendippens nach dem Melken traten diese
statistisch signifikant (p<0,0001) seltener (4,2% bzw. 7,0%) auf als in Betrieben ohne
Zitzendippen (12,8% bzw. 9,8%). Die niedrigste Prävalenz zeigten Betriebe, in denen die
Zitzen vor dem Melken mit einer Reinigungslösung und nach dem Melkvorgang mit einer
Jodlösung gedippt wurden. Hier lagen die Prävalenzen für S. aureus bei 2,4% und für S.
agalactiae bei 3,0%.
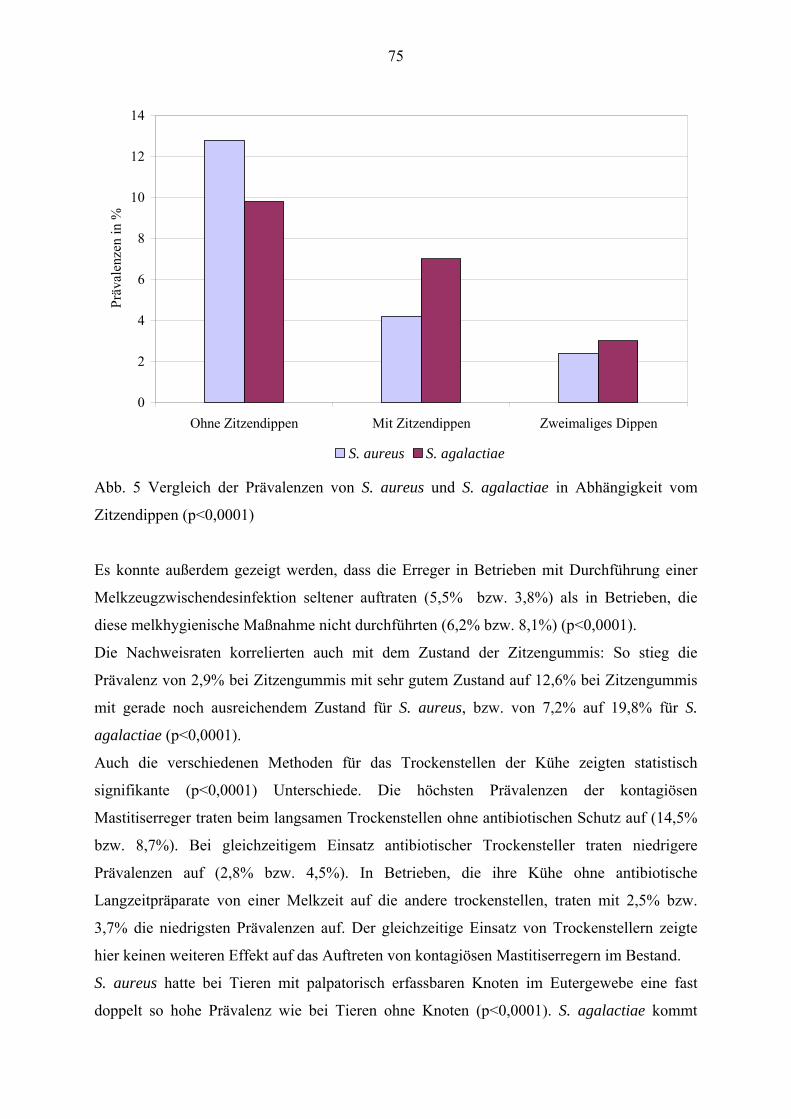
75
0
2
4
6
8
10
12
14
Ohne Zitzendippen Mit Zitzendippen Zweimaliges Dippen
Präv
alen
zen
in %
S. aureus S. agalactiae Abb. 5 Vergleich der Prävalenzen von S. aureus und S. agalactiae in Abhängigkeit vom
Zitzendippen (p<0,0001)
Es konnte außerdem gezeigt werden, dass die Erreger in Betrieben mit Durchführung einer
Melkzeugzwischendesinfektion seltener auftraten (5,5% bzw. 3,8%) als in Betrieben, die
diese melkhygienische Maßnahme nicht durchführten (6,2% bzw. 8,1%) (p<0,0001).
Die Nachweisraten korrelierten auch mit dem Zustand der Zitzengummis: So stieg die
Prävalenz von 2,9% bei Zitzengummis mit sehr gutem Zustand auf 12,6% bei Zitzengummis
mit gerade noch ausreichendem Zustand für S. aureus, bzw. von 7,2% auf 19,8% für S.
agalactiae (p<0,0001).
Auch die verschiedenen Methoden für das Trockenstellen der Kühe zeigten statistisch
signifikante (p<0,0001) Unterschiede. Die höchsten Prävalenzen der kontagiösen
Mastitiserreger traten beim langsamen Trockenstellen ohne antibiotischen Schutz auf (14,5%
bzw. 8,7%). Bei gleichzeitigem Einsatz antibiotischer Trockensteller traten niedrigere
Prävalenzen auf (2,8% bzw. 4,5%). In Betrieben, die ihre Kühe ohne antibiotische
Langzeitpräparate von einer Melkzeit auf die andere trockenstellen, traten mit 2,5% bzw.
3,7% die niedrigsten Prävalenzen auf. Der gleichzeitige Einsatz von Trockenstellern zeigte
hier keinen weiteren Effekt auf das Auftreten von kontagiösen Mastitiserregern im Bestand.
S. aureus hatte bei Tieren mit palpatorisch erfassbaren Knoten im Eutergewebe eine fast
doppelt so hohe Prävalenz wie bei Tieren ohne Knoten (p<0,0001). S. agalactiae kommt
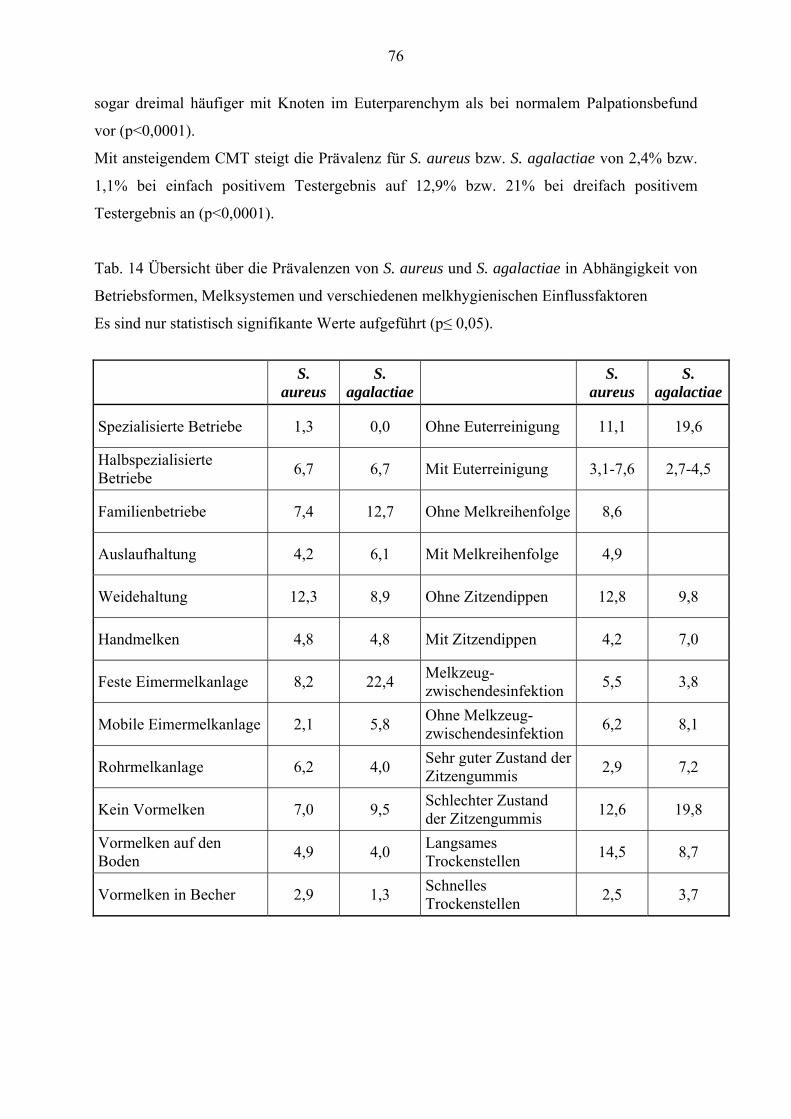
76
sogar dreimal häufiger mit Knoten im Euterparenchym als bei normalem Palpationsbefund
vor (p<0,0001).
Mit ansteigendem CMT steigt die Prävalenz für S. aureus bzw. S. agalactiae von 2,4% bzw.
1,1% bei einfach positivem Testergebnis auf 12,9% bzw. 21% bei dreifach positivem
Testergebnis an (p<0,0001).
Tab. 14 Übersicht über die Prävalenzen von S. aureus und S. agalactiae in Abhängigkeit von
Betriebsformen, Melksystemen und verschiedenen melkhygienischen Einflussfaktoren
Es sind nur statistisch signifikante Werte aufgeführt (p≤ 0,05).
S. aureus
S. agalactiae S.
aureus S.
agalactiae
Spezialisierte Betriebe 1,3 0,0 Ohne Euterreinigung 11,1 19,6
Halbspezialisierte Betriebe 6,7 6,7 Mit Euterreinigung 3,1-7,6 2,7-4,5
Familienbetriebe 7,4 12,7 Ohne Melkreihenfolge 8,6
Auslaufhaltung 4,2 6,1 Mit Melkreihenfolge 4,9
Weidehaltung 12,3 8,9 Ohne Zitzendippen 12,8 9,8
Handmelken 4,8 4,8 Mit Zitzendippen 4,2 7,0
Feste Eimermelkanlage 8,2 22,4 Melkzeug-zwischendesinfektion 5,5 3,8
Mobile Eimermelkanlage 2,1 5,8 Ohne Melkzeug-zwischendesinfektion 6,2 8,1
Rohrmelkanlage 6,2 4,0 Sehr guter Zustand der Zitzengummis 2,9 7,2
Kein Vormelken 7,0 9,5 Schlechter Zustand der Zitzengummis 12,6 19,8
Vormelken auf den Boden 4,9 4,0 Langsames
Trockenstellen 14,5 8,7
Vormelken in Becher 2,9 1,3 Schnelles Trockenstellen 2,5 3,7
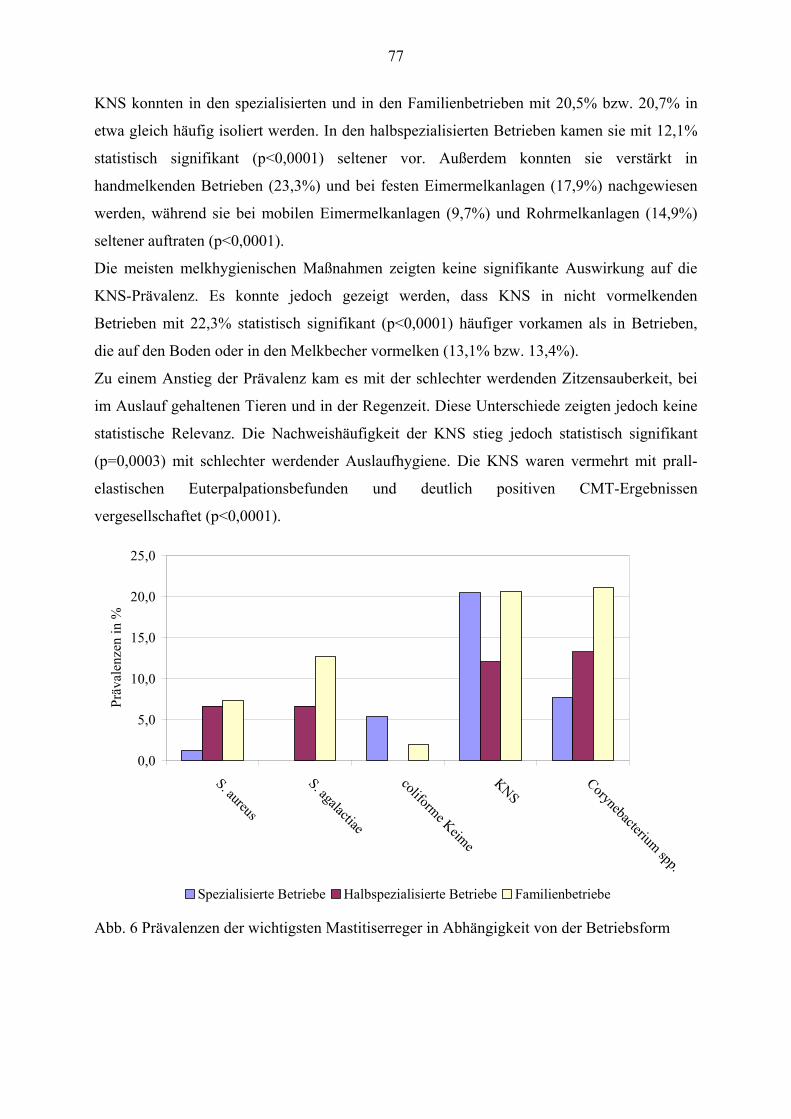
77
KNS konnten in den spezialisierten und in den Familienbetrieben mit 20,5% bzw. 20,7% in
etwa gleich häufig isoliert werden. In den halbspezialisierten Betrieben kamen sie mit 12,1%
statistisch signifikant (p<0,0001) seltener vor. Außerdem konnten sie verstärkt in
handmelkenden Betrieben (23,3%) und bei festen Eimermelkanlagen (17,9%) nachgewiesen
werden, während sie bei mobilen Eimermelkanlagen (9,7%) und Rohrmelkanlagen (14,9%)
seltener auftraten (p<0,0001).
Die meisten melkhygienischen Maßnahmen zeigten keine signifikante Auswirkung auf die
KNS-Prävalenz. Es konnte jedoch gezeigt werden, dass KNS in nicht vormelkenden
Betrieben mit 22,3% statistisch signifikant (p<0,0001) häufiger vorkamen als in Betrieben,
die auf den Boden oder in den Melkbecher vormelken (13,1% bzw. 13,4%).
Zu einem Anstieg der Prävalenz kam es mit der schlechter werdenden Zitzensauberkeit, bei
im Auslauf gehaltenen Tieren und in der Regenzeit. Diese Unterschiede zeigten jedoch keine
statistische Relevanz. Die Nachweishäufigkeit der KNS stieg jedoch statistisch signifikant
(p=0,0003) mit schlechter werdender Auslaufhygiene. Die KNS waren vermehrt mit prall-
elastischen Euterpalpationsbefunden und deutlich positiven CMT-Ergebnissen
vergesellschaftet (p<0,0001).
0,0
5,0
10,0
15,0
20,0
25,0
S. aureus
S. agalactiae
coliforme Keime
KNSCorynebacterium spp.
Präv
alen
zen
in %
Spezialisierte Betriebe Halbspezialisierte Betriebe Familienbetriebe Abb. 6 Prävalenzen der wichtigsten Mastitiserreger in Abhängigkeit von der Betriebsform

78
Die Corynebacterium spp. kamen mit 21,2% signifikant (p<0,0001) häufiger in
Familienbetrieben vor. In spezialisierten bzw. halbspezialisierten Betrieben lag ihre
Nachweishäufigkeit bei 7,7% bzw. 13,3%. Des Weiteren traten sie bei festen
Eimermelkanlagen (28,4%) häufiger als bei Rohrmelkanlagen (12,8%) und in handmelkenden
Betrieben (9,2%) auf. Die geringste Prävalenz ergab sich bei mobilen Eimermelkanlagen mit
7,9% (p<0,0001).
In Betrieben mit Einhaltung einer festen Melkreihenfolge oder Durchführung des
Zitzendippens traten Corynebacterium spp. seltener auf als ohne Durchführung dieser
Maßnahmen (p<0,0001). Bezüglich der Zitzensauberkeit war ein statistisch signifikanter
(p<0,0001) Unterschied zwischen Kühen mit und ohne Erregernachweis vorhanden. So stieg
die Nachweishäufigkeit mit schlechter werdender Zitzensauberkeit an. Die Erreger waren am
häufigsten mit einem leichten Anstieg des Zellgehaltes und einem grobkörnigen
Euterpalpationsbefund vergesellschaftet.
Coliforme Keime traten mit 5,4% am häufigsten in den spezialisierten Betrieben auf. Die
geringste Prävalenz zeigten sie mit 2,0% in den Familienbetrieben (p=0,008).
Ein Einfluss der Haltungsform auf das Auftreten konnte nicht ermittelt werden. Jedoch stieg
die Prävalenz signifikant mit sinkender Auslaufhygiene an. (p=0,002).
Von allen melkhygienischen Maßnahmen zeigte nur die Verwendung von antibiotischen
Trockenstellern einen Einfluss auf die Vorkommenshäufigkeit dieser Erreger. Bei sofortigem
Trockenstellen ohne Trockensteller konnten sie zu 6,1% und bei gleichzeitiger Verwendung
eines antibiotischen Trockenstellers zu 2,2% isoliert werden (p<0,0001). Bei dem langsamen
Trockenstellen war dieser Unterschied statistisch nicht signifikant. Coliforme Keime waren
meist mit hohen bis sehr hohen Zellzahlen vergesellschaftet und traten vermehrt in Eutern mit
Knotenbildung auf (p<0,0001).
Die Äskulin-positiven Streptokokken ließen sich häufiger bei Auslaufhaltung (4,0%) als bei
reiner Weidehaltung (2,2%) nachweisen (p=0,03). Auch eine mangelhafte Auslaufhygiene
war statistisch signifikant mit einem vermehrten Auftreten dieser umweltassoziierten Erreger
verbunden (p=0,006). Außerdem waren sie vermehrt bei Kühen mit schlechter
Zitzensauberkeit nachweisbar (p=0,0311). Die Äskulin-positiven Streptokokken waren
häufiger mit hohen Zellzahlen und einem prall-elastischen Euterpalpationsbefund
vergesellschaftet als mit niedrigeren Zellzahlen und anderen Palpationsbefunden (p<0,0001).
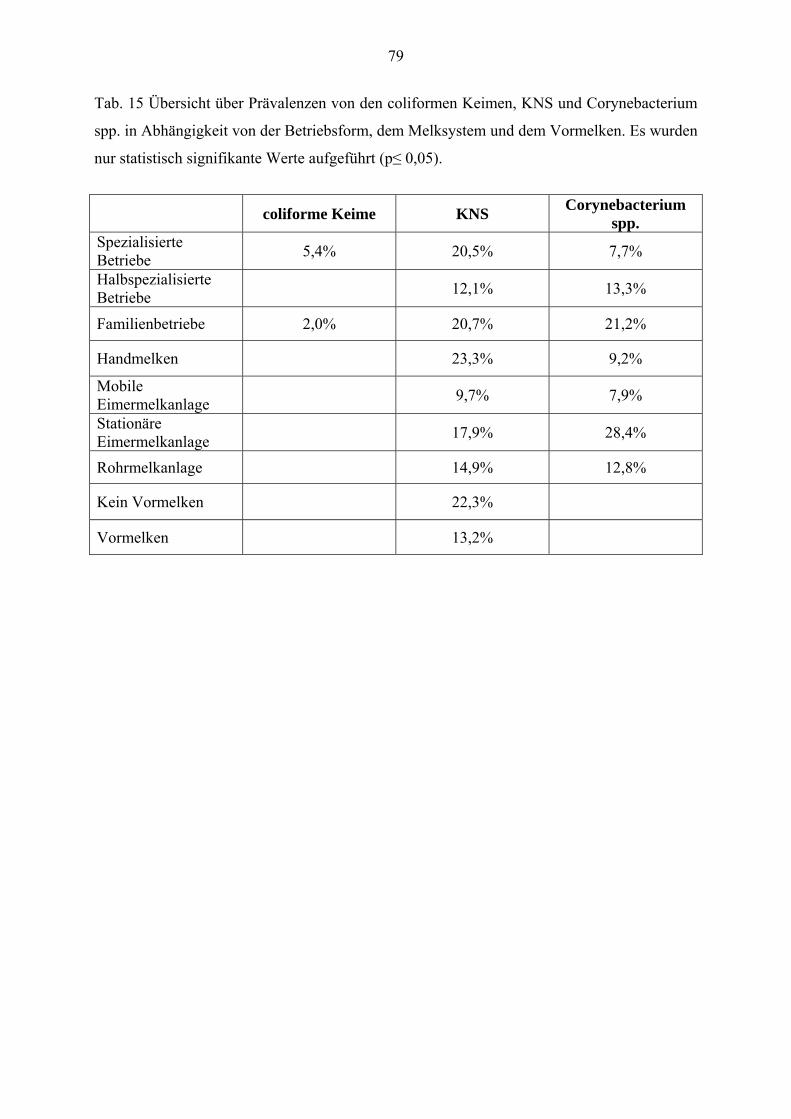
79
Tab. 15 Übersicht über Prävalenzen von den coliformen Keimen, KNS und Corynebacterium
spp. in Abhängigkeit von der Betriebsform, dem Melksystem und dem Vormelken. Es wurden
nur statistisch signifikante Werte aufgeführt (p≤ 0,05).
coliforme Keime KNS Corynebacterium spp.
Spezialisierte Betriebe 5,4% 20,5% 7,7%
Halbspezialisierte Betriebe 12,1% 13,3%
Familienbetriebe 2,0% 20,7% 21,2%
Handmelken 23,3% 9,2%
Mobile Eimermelkanlage 9,7% 7,9%
Stationäre Eimermelkanlage 17,9% 28,4%
Rohrmelkanlage 14,9% 12,8%
Kein Vormelken 22,3%
Vormelken 13,2%

80
5 Diskussion
5.1 Hygienische Verhältnisse von Ausläufen und Melkständen Die Beurteilung der Sauberkeit der Ausläufe gibt einen Hinweis auf den auf die Kuh
wirkenden Erregerdruck. Als hohen Keimdruck bezeichnet man eine starke Ansammlung von
Bakterien in der Umwelt der Tiere. Dazu gehört auch die Sauberkeit des Melkstandes. Wird
dieser nicht nach jedem Melken gereinigt, kommt es zu einer Ansammlung und eventuell zu
einer Vermehrung von Bakterien, durch welche der auf die Tiere einwirkende Keimdruck
ansteigt. Stresssituationen, unausgeglichene Fütterung, schlechte Stall- und Melkhygiene wie
auch geschwächte Abwehrreaktionen begünstigen Manifestationen von Euterinfektionen
(KIELWEIN 1994). Die Beurteilung der Betriebe zeigte, dass in 60,6% die Sauberkeit der
Ausläufe als mäßig bis schlecht anzusehen ist. Die von den Betriebsleitern bei der Befragung
angegebenen Reinigungsintervalle waren in den meisten Fällen zu lang. Durch die fehlenden
Ablaufmöglichkeiten für Regenwasser kommt es in der Regenzeit häufig zu extrem
matschigen Böden, wodurch eine Kontamination der Zitzen häufig unvermeidbar wird.
Zusammen mit der häufig ungenügenden Reinigung der Zitzen vor dem Melken besteht
hierbei eine lebensmittelhygienische Problematik. In der Sommerzeit besteht der Bodenbelag
vieler Ausläufe überwiegend aus eingetrocknetem Kot. Ähnlich sieht die Situation auf den
Melkplätzen aus: Zum Zeitpunkt des Melkvorgangs sind nur bei drei (9,1%) Betrieben die
Melkplätze als sauber einzustufen. In vielen Betrieben besitzen die Melkplätze keinen
befestigten, leicht zu reinigenden Boden. Häufig ist der feuchte Boden mit einem Gemisch
aus Futterresten, Einstreu und Dung bedeckt. Eine den Melkvorgang abschließende Reinigung
bzw. Desinfektion des Melkbereiches findet gar nicht bzw. nur oberflächlich statt. Durch
diese Gegebenheiten muss von einer starken Keimansammlung und -vermehrung auf dem
Boden ausgegangen werden. Diese ist insbesondere bei gleichzeitigem Vormelken auf den
Boden, wobei Milch vom Boden an die Zitzen zurückspritzen kann, als kritisch für den
Eutergesundheitszustand einer Herde anzusehen. Es muss daher davon ausgegangen werden,
dass die Mastitissituation in Jalisco mit der ungenügenden Haltungs- und Melkstandhygiene
im direkten Zusammenhang steht und durch eine Behebungung dieses und weiterer Mängel
verbessert werden könnte. Diese Ergebnisse sind typisch für Länder ohne
Masitiskontrollprogramme. So stufte FRESE (2003) die allgemeine Farmhygiene in
äthiopischen Betrieben in 43% als schlecht ein. Nur 19% der Farmen wurde die generelle
Hygiene als gut eingestuft.

81
5.2 Vorkommenshäufigkeit subklinischer und klinischer Mastitiden
Von den 2937 vorliegenden CMT-Ergebnissen zeigten 62,9% ein positives und von diesen
37% ein deutlich bis stark positives Ergebnis. Nach GRAY und SCHALM (1962) liegt der
Produktionsverlust bei einem einfach, mittel oder stark positivem CMT-Ergebnis in etwa bei
10%, 16% bzw. 24%. Dies zeigt, dass die Mehrzahl der mexikanischen Betriebe weit unter
ihrem möglichen Potential produzieren.
Ein Vergleich mit Ergebnissen aus anderen Regionen Mexikos zeigt, dass dieser Wert
durchaus typisch für die Milchwirtschaft in Mexiko ist. So zeigte JUÁREZ et al. (1980), dass
im Gebiet Valle de México eine Inzidenz der subklinischen Mastitis von 78% herrschte.
AVILA et al. (1991) konnten bei Zweinutzungsrindern in den feucht-tropischen Zonen
Mexikos eine Inzidenz von 39,7% nachweisen. Ferner untersuchten AVILA et al. (2001)
1092 Viertel in Xochimilco und fanden eine Inzidenz der subklinischen Eutererkrankungen
von 34%. DOMÍNGUEZ (1996) sammelte über 5 Jahre Daten aus ganz Mexiko und schätzt
die Inzidenz der subklinischen Mastitis im gesamten Land auf etwa 50%.
Auch beim Vergleich dieses Wertes mit in anderen Ländern gefundenen Daten zeigt sich,
dass die hohe Mastitisinzidenz keineswegs ungewöhnlich ist. PUIG DE CENTORBI et al.
(1992) untersuchten 652 Viertel in Argentinien, von denen 131 (80,3%) ein positives und
hiervon immerhin 46,6% ein deutlich bis stark positives CMT-Ergebnis zeigten. FRESE
(2003) konnte subklinische Mastitiden in äthiopischen Milchviehherden mit einer Prävalenz
von 52,7% nachweisen. LAFI et al. (1994) fanden bei ihrer Untersuchung von 63 Herden in
Jordanien, dass 60% der Viertel einen Milchzellgehalt von über 283.000 Zellen/ml aufwiesen.
THOMPSON und HOUSTON (1967) geben an, dass in Australien 50% aller untersuchten
Kuhviertel ein positives und hiervon 34% ein deutlich bis stark positives CMT-Resultat
zeigten. Diese Ergebnisse liegen ungefähr in der gleichen Größenordnung, wie der in Jalisco
gefundene Wert. Sie liegen jedoch weitaus höher als vorliegende Untersuchungsergebnisse
aus Finnland und Schweden, welche auf Kuhebene eine Inzidenz von 37 bzw. 30% angeben
(MYLLYS et. al. 1998; Swedish Dairy Association 2000). In Hessen betrug die
durchschnittliche Zellzahl der Anlieferungsmilch im Jahr 2004 176.000 Zellen pro ml Milch
(ADR 2004). Diese Differenzen können durch das vollständige Fehlen von
Eutergesundheitsprogrammen zum Untersuchungszeitpunkt in Mexiko, Argentinien,
Jordanien und in Australien sowie das mangelhafte Management bei der Viehhaltung sowie
die hygienischen Konditionen der Stalleinrichtungen erklärt werden. In Schweden, Finnland
und Hessen wurden zum Untersuchungszeitpunkt schon Eutergesundheitsprogramme
durchgeführt, wodurch die Mastitisinzidenz offensichtlich gesenkt werden konnte.

82
Bei der Beurteilung von nur auf CMT-Ergebnissen beruhenden Studien muss jedoch immer
berücksichtigt werden, dass der CMT zwar eine verlässliche indirekte Methode zur Schätzung
des Milchzellgehaltes von individuellen Vierteln darstellt, seine Probleme vor allem aber
darin liegen, dass seine Sensitivität unter anderem von der Subjektivität der anwendenden
Person abhängig ist. In der vorliegenden Studie wurde der CMT meist von einem häufiger
wechselnden Team mexikanischer Studenten der Tiermedizin oder Landwirtschaft
durchgeführt, weswegen von gewissen Schwankungen bei der Interpretation ausgegangen
werden muss.
Als indirekte Methode zur Bestimmung der Leukozytenzahl kann der CMT lediglich ein
Indikator für eine Zellzahlerhöhung durch eine Euterentzündung sein.
Bei der Interpretation der Ergebnisse muss ebenfalls in Betracht gezogen werden, dass der
von der DVG (2002) festgesetzte Grenzwert für Milchzellgehalte für mexikanische
Verhältnisse zu niedrig angesetzt sein könnte. So fand GIRAUDO et al. (1995) z. B. bei
seinen Untersuchungen in argentinischen Milchviehherden, dass der arithmetrische Mittelwert
bei bakteriologisch-negativen Vierteln bei 494.000 Zellen/ml lag. Auch GIANNEECHINI et
al. (2002) setzten für die Auswertung ihrer Ergebnisse in Uruguay den Schwellenwert für die
subklinische Mastitis auf 300.000 Zellen/ml, um den aktuellen Verhältnissen in Uruguay und
Südamerika besser gerecht zu werden. Laut FRESE (2003) ist ein Schwellenwert von 300.000
Zellen pro ml/Milch zu niedrig angesetzt, da die Zellzahlen auch offensichtlich gesunder
Euterviertel erhöht sind. PUIG DE CENTORBI et al. (1992) werteten bei ihren
Untersuchungen in Argentinien nur deutlich bis stark positive CMT-Ergebnisse als
subklinische Mastitiden. DOHOO und MEEK (1982) sowie BROLUND (1985) setzen die
Grenze, welche die eutergesunden von euterkranken Vierteln trennt, zwischen 200.000 und
500.000 Zellen/ml Milch. Für die vorliegende Arbeit wurde bereits ein einfach positiver CMT
als eine Sekretionsstörung gewertet. Dies war bei 62,9% der untersuchten Viertel vorhanden.
Wäre erst ein zweifach positives CMT-Ergebnis als Eutergesundheitsstörung gewertet
worden, wäre der Prozentsatz der betroffenen Viertel auf 37,7% gesunken.
Die klinische Mastitis wurde im Gegensatz zur subklinischen Mastitis nicht durch
Überschreitung eines bestimmten Zellzahlgrenzwertes, sondern durch visuell abnormales
Eutersekret und/oder ein vermehrt warmes, akut geschwollenes oder schmerzhaftes Euter mit
oder ohne Beeinträchtigung des Allgemeinbefindens definiert. Die Untersuchungen zeigten,
dass 76 (2,5%) aller untersuchten Viertel zum Untersuchungszeitpunkt nach dieser Definition
an einer klinischen Mastitis erkrankt waren. AVILA et al. (2001) konnten 3,5% klinisch
erkrankte Euterviertel bei der Untersuchung von 273 Kühen in Xochimilco, Mexiko

83
nachweisen. ALARCǑN (1997) untersuchte die jahreszeitlichen Schwankungen des
Auftretens von klinischen Mastitiden bei in tropischen Regionen gehaltenen Milchviehherden
und konnte bei seinen Untersuchungen je nach Jahreszeit in 2,1 bis 6,5% der Viertel eine
klinische Erkrankung nachweisen.
Die eigenen Ergebnisse stimmen gut mit den Untersuchungen von DANIEL et al. (1982) in
Australien überein, die in 26 Herden in Queensland eine Viertelprävalenz von 2,6% fanden.
BISHI (1998) untersuchte Milchviehherden in Äthiopien und zeigte, dass 1,6% der
untersuchten Euter klinische Erkrankungsanzeichen zeigten. Mc CLURE et al. (1966) fanden
in Australien eine Erkrankungsrate an klinischer Mastitis von 3,4%. FRESE (2003) konnte in
äthiopischen Betrieben eine Prävalenz von nahezu 5% nachweisen. GIANNEECHINI et al.
(2002) stellten bei ihren Untersuchungen in Uruguay eine Inzidenz von 14,4 Fällen/100
Kuhjahren in Risiko fest. In Schweden, Norwegen und Dänemark wird die Inzidenz von
PLYM-FORSHELL et al. (1995) mit 21, 30 bzw. 56 Fällen/100 Kuhjahren in Risiko
angegeben, und liegt somit über den eigenen ermittelten Werten. Nach BARLTETT et al.
(1992) können Mastitisinzidenzen verschiedener geographischer Lokalisationen bzw. Länder
nur unter Vorbehalt miteinander verglichen werden, da die Unterschiede der Inzidenz der
klinischen Mastitiden in Milchviehherden mit vielen verschiedenen Faktoren wie z.B. dem
Klima, der Rasse, dem Produktionslevel und dem Herdenmanagement in enger Verbindung
stehen. Ferner müssen auch jahreszeitliche und ökologische Einflüsse berücksichtigt werden.
Wichtig ist auch, dass es unmöglich ist, die tatsächliche Inzidenz klinischer Mastitiden durch
eine einmalige Herdenuntersuchung festzustellen, da diese meist nur von kurzer Dauer sind
und deshalb nach der Gesundung und Elimination des Erregers schnell durch eine einzelne
Beprobung nicht entdeckt werden.
In Mexiko überwiegen mittlere bis hohe Zellgehalte in der Herdensammelmilch, und beim
direkten Vergleich mit den skandinavischen Ländern fällt eine weitaus niedrigeres Auftreten
klinischer Mastitiden auf. Diese negative Korrelation zwischen Zellgehalt der Tankmilch und
der Inzidenz klinischer Mastitiden zeigten bereits ERSKINE et al. (1998), die in Herden mit
niedrigem Tankmilchzellgehalt eine weitaus höhere Inzidenz klinischer Mastitiden
nachweisen konnten als in Herden mit hohen Tankmilchzellzahlen. Auch SCHUKKEN et al.
(1990) beschrieben, dass Farmen mit einem sehr geringen Tankmilchzellgehalt einen Anstieg
der klinischen Mastitisfälle mit einer hohen Prävalenz von E. coli-Infektionen zeigen.

84
5.3 Prävalenz der einzelnen Mastitiserreger Nach dem Ausstreichen auf Blutagar zeigten 53,8% der untersuchten
Viertelanfangsgemelksproben kulturell keinen Keimgehalt. Diese Rate liegt über den meisten
in der Literatur gemachten Angaben, lässt sich aber durch die Tatsache erklären, dass
keinerlei Vorselektion der zur bakteriologischen Untersuchung verwendeten Proben durch
z.B. den CMT stattfand, sondern sämtliche entnommene Proben bakteriologisch untersucht
wurden. So konnten THOMPSON und HOUSTON (1967) in zwei verschiedenen
Untersuchungsgruppen bei 41 bzw. 37% der Milchproben kein bakterielles Wachstum
nachweisen. Die Autoren untersuchten jedoch nur Proben, die mindestens eine einfach
positive Reaktion beim CMT zeigten, wodurch kein direkter Vergleich dieser Ergebnisse mit
den Resultaten der vorliegenden Untersuchung möglich ist. Vergleichbar ist hingegen das
Ergebnis von WILSON et al. (1997), die Milchproben von 108.312 Milchkühen in den
Staaten New York und Pennsylvania ebenfalls ohne Vorselektion untersuchten, und hierbei in
51,5% der Fälle kulturell-bakteriologisch keine Erreger isolieren konnten.
Die aus den Viertelgemelksproben Mastitis-kranker Kühe isolierten Erreger sind KNS 15,6%,
Corynebacterium spp. 14,0%, S. agalactiae 6,7%, S. aureus 5,9%, coliforme Keime 4,1%,
Streptococcus spp. 3,7% und sonstige Keime (Bazillus spp. Nocardia spp. Candida spp.)
1,7%. Diese Resultate unterscheiden sich substantiell von in anderen Ländern durchgeführten
Studien.
Einen ähnlichen prozentualen Anteil an KNS konnte in einer Studie von HARMON und
LANGLOIS (1989) aus Milchproben einer Milchviehherde in Kentucky mit 14,7% isoliert
werden. Auch FRESE (2003) konnten in seinen Studien in Äthiopien deutlich mehr fakultativ
pathogene Erreger als strikt euterpathogene Erreger nachweisen. Seine Nachweisraten der
KNS lagen je nach Betriebsform bei 12,1-39,4%. Auch BRAMLEY (1975) fand in einer
britischen Herde, in der kein Zitzendippen nach dem Melken praktiziert wurde, eine
Prävalenz von 13,4%, während er in einer anderen Herde, in der regelmäßig gedippt wurde,
eine Prävalenz von 6,3% nachweisen konnte. KUDINHA und SIMANGO (2002) konnten bei
ihrer Untersuchung von 406 Milchproben mit 22,9% bzw. 33,2% einen sehr hohen Anteil der
KNS an klinischen bzw. subklinischen Mastitiden feststellen. Gleichzeitig zeigten sie, dass
95% dieser Isolate aus Milchproben mit erhöhtem Zellgehalt isoliert wurden. Dies beweist,
dass die Isolate Auslöser von Euterparenchyminfekionen sind und nicht allein von
Strichkanalbesiedelungen stammen. Die hohe Isolierungsrate der KNS in der vorliegenden

85
Arbeit könnte durch den protektiven Effekt, welchen KNS gegenüber einer Superinfektion
mit E. coli, S. agalactiae und S. aureus ausüben sollen, erklärt werden. Interessanterweise
konnten nämlich in Herden mit hohen (bis 100%) Prozentsätzen an KNS kaum
majorpathogene Erreger nachgewiesen werden. In Herden, in denen überwiegend S. aureus
oder S. agalactiae nachgewiesen wurden, lag die Herdenprävalenz für KNS sehr viel
niedriger.
Zu ähnlichen Ergebnissen kam ABDELLA (1996) bei ihren Untersuchungen in mehreren
Milchviehherden in Äthiopien. Sie zeigte, dass die Präsenz von KNS das Wachstum potentiell
pathogener Keime hemmte und deswegen Viertel mit KNS-Nachweis weniger empfindlich
für eine Infektion mit majorpathogene Keimen waren. Die hohe Nachweisrate der KNS in
subklinisch erkrankten Vierteln zeigt jedoch auch, dass die KNS eine große Rolle als
Mastitiserreger spielen können.
Die eigenen Untersuchungen zeigten außerdem eine hohe Prävalenz von Corynebacterium
spp.. Die Rolle der Corynebakterien als Mastitiserreger ist nicht eindeutig geklärt.
ROBINSON und HARWOOD (1998) vertreten die Auffassung, dass Corynebacterium spp.
imstande sind Mastitiden auszulösen, während sie von anderen Autoren als opportunistische
Erreger und Bewohner des Strichkanals angesehen werden (RAINARD 1987). So sind die
Zellzahlen meist nur moderat erhöht und die Milchmenge, wenn überhaupt, geringgradig
reduziert. Corynebacterium spp. haben auch dadurch das Interesse geweckt, dass verschiedene
Autoren ihre protektive Wirkung in Bezug auf majorpathogene Mastitiserreger beschrieben.
So sollen mit C. bovis infizierte Viertel weniger anfällig gegen Infektionen mit
majorpathogenen Keimen sein als nicht infizierte (RAINARD 1987, LAM et al. 1997).
Hierdurch könnte sich die verglichen mit anderen Studien geringere Prävalenz von S. aureus
in der vorliegenden Studie erklären lassen.
Die hohe Prävalenz von S. agalactiae in der vorliegenden Kuhpopulation ist eine direkte
Reflexion des mangelhaften Herdenmanagements. So konnten KLOPPERT et al. (1999) in
Hessen zeigen, dass durch die Einführung geeigneter Mastitiskontrollprogramme die
Prävalenz von S. agalactiae in der kontrollierten Kuhpopulation auf einen Wert von 0,2%
gesenkt werden konnte. Auch in der Schweiz und in Finnland, zwei Ländern, in denen die
Milchhygiene weit fortgeschritten ist, konnten BUSATO (1997) bzw. MYLLYS et al. (1998)
nur eine Prävalenz von 0,8% bzw. 0,12% nachweisen. Deutlich höher ist die Nachweisrate in
Ländern, in denen keine gezielten Mastitiskontrollprogramme zur Anwendung kommen. So

86
lag die Prävalenz in Äthiopien nach WORKINEH et al. (2002) bei 7,4%, und
GIANNEECHINI et al. (2002) konnten für Uruguay eine Prävalenz von 11,3% feststellen.
Interessanterweise konnte in spezialisierten Betrieben Jaliscos, in denen eine konsequente
Melkhygiene betrieben wird, kein S. agalactiae nachgewiesen werden. Diese Ergebnisse
unterscheiden sich deutlich von den Untersuchungen von FRESE (2003) in Äthiopien, der S.
agalactiae mehr als doppelt so häufig in großen periurbanen Milchbetrieben nachweisen
konnte als in kleinen urbanen Betrieben. Besonders hoch hingegen war die Prävalenz in den
Familienbetrieben, in denen Zitzendippen und antibiotisches Trockenstellen der Tiere kaum
bis keine Anwendung finden. S. agalactiae kann nach KEEFE (1997) mit Hilfe der
antibiotischen Blitztherapie und einer konsequenten Verbesserung der Melkhygiene schnell
und erfolgreich aus einem geschlossenen Bestand eliminiert werden.
Das in Jalisco nachgewiesene Vorkommen von S. aureus mit einem Anteil von 5,9% an den
gesamten Milchproben lag höher als Nachweisquoten in Herden mit einer niedrigen Zellzahl
der Herdensammelmilch und niedriger als Nachweisquoten in Herden mit sehr hohem
Tankmilchzellgehalt. WILSON et al. (1997) fanden bei ihren mehrjährigen Untersuchungen
in den USA eine S. aureus-Prävalenz von 9,1%. Eine ähnliche Nachweisrate fanden BISHI et
al. (1998) mit 9,5% der untersuchten Proben in Äthiopien. Wieder zeigt sich, dass in Ländern
mit guter Milchhygiene wie Finnland, dem Vereinigten Königreich und Hessen/Deutschland
relativ geringe Prävalenzen von 3,5 bzw. 3,0% (MYLLYS et al. 1998; KLOPPERT et al.
1999; MILNE et al. 2002) nachgewiesen werden konnten, während die Prävalenzen in
Ländern mit fehlenden Mastitiskontrollprogrammen deutlich höher liegen. FRESE (2003)
konnte in äthiopischen Betrieben eine Prävalenz von 15,8% nachweisen. Teilweise werden
sogar Prävalenzen von bis zu 34,2% in Zimbabwe oder sogar 38,5% in Australien
beschrieben (KUDINHA u. SIMANGO 2002; DANIEL et al. 1982). Eine mit Jalisco
vergleichbare Rate fand sich mit einer Prävalenz von 7,6% in Brasilien (BENITES et al.
2002). Ferner konnte eine statistische Abhängigkeit der Vorkommenshäufigkeit des Erregers
von der Zwischendesinfektion der Melkzeuge, der Anwendung von antibiotikahaltigen
Trockenstellern, dem Vormelken, der Einhaltung einer Melkreihenfolge, dem Dippen, der Art
der Euterreinigung und dem Zustand der Zitzengummis festgestellt werden. Durch
Optimierung all dieser Faktoren könnte die Prävalenz von S. aureus auch in Jalisco mit
Sicherheit erheblich gesenkt werden.
Die Makrorestriktionsanalyse der Gesamt-DNA von 27 S. aureus Stämmen aus 12 Betrieben
wurde zum Nachweis epidemiologischer Zusammenhänge innerhalb der einzelnen Betriebe

87
sowie zwischen den Betrieben durchgeführt. Nach KAPUR et al. (1995) ist ein besseres
Wissen über die Verteilung der infektiösen Stämme in den Milchviehherden für die
Entwicklung von Strategien zur Reduzierung der Infektionsverbreitung unerlässlich.
Die Untersuchungen zeigten, dass die Isolate von vier Betrieben (Betrieb 7, 23, 25, 31)
jeweils identisch waren und keine Unterschiede im Restriktionsmuster zeigten. Durch die
ebenfalls fehlende Variation der Bandenmuster innerhalb der Betriebe ist anzunehmen, dass
in jedem Bestand ein einzelner Bakterienklon für die Mastitissituation verantwortlich ist.
Diese Beobachtungen bestätigen Untersuchungen von ANNEMÜLLER et al. (1999) und
ZSCHÖCK et al. (1999), die die Kontagiosität der S. aureus-Mastitis ebenfalls mittels
Makrorestriktionsanalyse nachwiesen. So breitet sich ein einzelner Klon innerhalb der Herde
aus. Von diesen Klonen wird vermutet, dass sie aufgund eines erhöhten
Übertragungspotentials, einer gesteigerten Resistenz gegenüber den Abwehrmechanismen des
Wirtes und ihrer erhöhten Neigung im Wirt zu persistieren, eine erhöhte Virulenz besitzen. So
ist z.B. bekannt, dass Isolate des dominierenden Genotyps eine erhöhte Resistenz gegenüber
der bakteriziden Aktivität der neutrophilen Granulozyten aufweisen (SCHLEGELOVÁ et al.
2003).
Innerhalb des Betriebs 19 unterschieden sich die Muster durch ein Fragment. Dieses Ergebnis
ist laut GOERING (1998) zwar möglich, aber äußerst selten. So könnte die Deletion eines
DNA-Fragmentes mit beiden flankierenden Schnittstellen zum Verlust eines einzelnen
Fragmentes im PFGE-Muster führen. Die relative Sicherheit solcher genetischen „events“
lässt vermuten, dass die meisten beschriebenen Einzelfragmentdifferenzen in Wirklichkeit
Zweifragmentunterschiede sind, bei denen eines der Fragmente hinter einem gleich großen
versteckt ist. In Betrieb 27 zeigten die untersuchten Isolate Abweichungen in Form von zwei
Banden. Diese Differenz könnte nach GOERING (1998) durch Insertion bzw. Deletion eines
DNA-Sequenzabschnittes außerhalb einer SmaI-Schnittstelle hervorgerufen worden sein.
Diese Kulturen sind durch ihre geringen Differenzen vom Ursprungsklon innerhalb des
jeweiligen Betriebes als Subklone eines Makrorestriktionstyps zu werten. Die subklonale
Heterogenität ist nach ZADOKS et al. (2000) durch die zeitliche Evolution bedingt. Dieses
Phänomen wurde für humane S. aureus Stämme beschrieben (O´BRIEN et al. 1999). Die
Variation des genetischen Inhalts der Chromosomen von S. aureus kann mit dem Verlust von
nicht essentiellen, aber klinisch und epidemiologisch relevanten Genen wie z.B. Virulenz-
oder Resistenzgenen vergesellschaftet sein, wodurch sich das seltenere Vorkommen
bestimmter Klone erklären lässt (PREVOST et al. 1992; MATSUNAGA et al. 1993).

88
In den Betrieben 10, 12 und 22 zeigten die Isolate in der PFGE einen Unterschied von mehr
als fünf Banden. Nach BANNERMANN et al. (1995) werden Isolate mit mehr als drei
Unterschieden im Bandenmuster als unterschiedliche, nicht miteinander verwandte Stämme
angesprochen und daher unterschiedlichen Makrorestriktionstypen zugeordnet.
Die Mehrzahl der in den 12 Betrieben nachwiesenen Genotype konnte in mehreren Herden
gefunden werden. Dieses Phänomen zeigte sich auch in anderen Studien (AARESTRUP et al.
1995; FITZGERALD et al. 1997; ANNEMÜLLER et al. 1999; SU et al. 1999; ZADOKS
2000). Die genannten Autoren konnten jeweils eine limitierte Anzahl von dominierenden S.
aureus-Klonen sowohl innerhalb einer Herde als auch zwischen unterschiedlichen
Milchviehherden nachweisen. Dies lässt annehmen, dass bestimmte in der Umwelt
vorkommende Varianten einen Vorteil für die Auslösung einer intramammären Infektion
besitzen. Nach AARESTRUP et al. (1995) und MULLARKY et al. (2001) haben sie
außerdem eine höhere Resistenz gegenüber dem Immunsystem des Wirtsorganismus. Solche
Klone haben nach FITZGERALD et al. (1997) eine weite geographische Verbreitung, die
ihnen durch besondere Faktoren für eine effektive Verbreitung in der bovinen Umwelt
ermöglicht wird. So konnte eine limitierte Anzahl dominierender Typen in verschiedenen
Herden, Regionen, Ländern und sogar Kontinenten gefunden werden.
Meist lässt sich jedoch, wie auch bei den hier überprüften Betrieben, eine gewisse genetische
Heterogenität der Stämme innerhalb geschlossener Populationen nachweisen (MATTHEWS
et al. 1994; KAPUR et al. 1995; ANNEMÜLLER et al. 1999; FITZGERALD et al. 1997;
MYLLYS et al. 1997, SOMMERHÄUSER 2002). Selbst bei einzelnen Tieren oder innerhalb
eines Euterviertels können gelegentlich verschiedene S. aureus-Klone gefunden werden.
EL-SAYED et al. (im Druck) konnten unter den 17 auf SE-Toxingene untersuchten S. aureus-
Stämmen lediglich einen einzigen Stamm (5,9%) mit einem 580 bp Amplifikat für das SEI
Gen nachweisen. Keiner der untersuchten Stämme beherbergte Gene für die SEA, SEB, SEC,
SED, SEE, SEG, SEJ und das TSST-1.
Dieses Ergebnis überrascht nicht, wenn man es mit den Untersuchungsresultaten von PUIG
de CENTORBI et al. (1992) vergleicht. Diese fanden bei 27 Stämmen aus Argentinien kein
Enterotoxin- oder TSST-1 produzierendes Isolat. Die geographische Verteilung von
enterotoxinbildenden S. aureus-Stämmen ist weltweit sehr unterschiedlich. So zeigten
Untersuchungen in Dänemark, dass hier keine Enterotoxingene bei den S. aureus-Stämmen
nachgewiesen werden konnten, während sie in den übrigen skandinavischen Ländern recht
häufig vertreten waren (TOLLERSRUD et al. 2000). Da von den in Jalisco isolierten

89
Stämmen nur 17 Isolate auf ihr Enterotoxinbildungsvermögen untersucht worden sind, ist die
Untersuchung einer größeren Anzahl von Stämmen notwendig, um mit größerer Sicherheit
Aussagen über das Vorkommen von Enterotoxinbildnern in Mexiko und ihre
lebensmittelhygienische Bedeutung für die mexikanische Bevölkerung machen zu können.
Dies ist insbesondere in Anbetracht der hohen Rohmilchdirektvermarktung in Jalisco von
großer Bedeutung, da gezeigt werden konnte, dass die Prävalenz von S. aureus in
Familienbetrieben, bei denen der Verkauf von Rohmilch sehr verbreitet ist, besonders hoch
ist. Vor diesem Hintergrund muss ein potentielles gesundheitliches Risiko zumindest
vermutet werden.
Bei der Interpretation der Prävalenz der coliformen Keime muss nach GOLDBERG et al.
(1992) das Haltungsmanagement miteinbezogen werden. So konnten die Autoren
umweltassoziierte Bakterien bei Weidehaltung der Tiere seltener auf der Zitzenhaut
nachweisen als bei aufgestallten Tieren. Dieses Phänomen lässt sich durch das höhere
Expositionsrisiko der aufgestallten Tiere erklären, während bei weidenden Tieren die
bakterielle Kontamination der Zitzen geringer ist. Ein Risikofaktor liegt allerdings in den
häufig matschigen Weidearealen, an denen sich die Tiere vermehrt zusammenfinden. So
können morastige Areale an Wasserstellen, Eingängen oder Triebwegen zum Melkstand
durch vermehrte Zitzenkontamination zu einer erhöhten Inzidenz von Mastitiden mit
umweltassoziierten Erregern in Milchviehherden führen (SMITH u. HOGAN 1995). Eine
derartige Korrelation konnte anhand der eigenen Ergebnisse jedoch nicht gefunden werden. In
Jalisco konnten die coliformen Keime in der Trockenzeit doppelt so häufig nachgewiesen
werden wie in der Regenzeit. Ferner konnten sie in Familienbetrieben weitaus seltener isoliert
werden als in den halbspezialisierten und spezialisierten Betrieben. Dies lässt sich dadurch
erklären, dass coliforme Keime in Betrieben mit einem hohen Zellgehalt der
Herdensammelmilch infolge einer S. aureus bzw. S. agalactiae-Infektion, wie dies in den
Familienbetrieben häufig der Fall ist, keine so große Bedeutung haben wie in hygienisch
einwandfrei geführten Betrieben mit einem geringen Zellgehalt der Herdensammelmilch. Die
in Jalisco nachgewiesene Prävalenz von 4,1% korreliert mit Ergebnissen aus Äthiopien und
Zimbabwe, zwei Ländern mit einem ähnlich hohen Zellgehalt in der Anlieferungsmilch. Hier
konnten BISHI et al. (1992) bzw. KUDINAH et al. (2002) einen Anteil von 5,2% bzw. 5,4%
nachweisen. FRESE (2003) konnten bei seinen Untersuchungen in äthiopischen Betrieben gar
keine coliformen Keime nachweisen. In Schweden und Finnland, zwei Ländern mit hohem
Milchhygienestandard, konnten HALLEN-SANDGREN (2000) und MYLLYS et al. (1998)

90
coliforme Keime zu 1,5% bzw. 0,29% aus Milchproben von an subklinischer Mastitis
erkrankten Kühen isolieren. Erwähnt werden muss hierbei, dass die coliformen Keime zu
einem wesentlich höheren Anteil aus Milchproben von klinischen Mastitiden isoliert werden
können. So wiesen MILNE et al. (2002) aus Milchproben von klinischen Erkrankungen im
Vereinigten Königreich eine Prävalenz von 23% nach.
Äskulin-positive Streptokokken wurden in Jalisco nur zu 3,7% isoliert. Eine weitergehende
Identifizierung wurde an 45 Isolaten durchgeführt, wobei zu 35,6% Aerokokkenspezies, zu
26,7% Enterokokkenspezies und zu 24,4% S. uberis differenziert wurden. Aerokokken und
Enterokokken werden häufig als Kontamimaten angesehen. S. uberis hingegen gilt eindeutig
als umweltassoziierter Mastitiserreger. Eine mit Jalisco vergleichbare Nachweisrate wurde
von BISHI et al. (1998) mit einer Prävalenz von 3,8% in Äthiopien und von DANIEL et al.
(1982) mit 3,4% in Australien beschrieben. FRESE (2003) konnte in Äthiopien ebenfalls eine
S. uberis-Prävalenz von unter 5% nachweisen. In den Ländern mit konsequent durchgeführter
Milchhygiene haben die Äskulin-positiven Streptokokken in den letzten Jahren durch den
gravierenden Rückgang von S. agalactiae zahlenmäßig bzw. relativ an Bedeutung gewonnen.
So konnte S. uberis in Schweden und Hessen/Deutschland, zwei Ländern mit niedriger S.
agalactiae-Prävalenz, von HALLEN-SANDGREN (2000) und KLOPPERT et al. (1999) aus
6,4% bzw. 7,3% der Proben isoliert werden. In der Schweiz liegen die Prävalenzen für die
erste bzw. zweite Laktationshälfte nach BUSATO et al. (1997) bei 19,4 bzw. 15,6%.
S. dysgalactiae, Bazillus spp., Nocardia spp. und Candida spp. konnten nur ganz vereinzelt
(0,4%, 1,0%, 0,6% und 0,1%) aus den untersuchten Proben isoliert werden. Eine ähnlich
niedrige Prävalenz für S. dysgalactiae beschreibt lediglich MYLLYS et al. (1998) mit 0,08%
für Finnland. In den übrigen Ländern schwanken die Prävalenzen zwischen 1,6% in Brasilien
(BENITES et al. 2002) und 6,6% in Australien (DANIEL et al. 1982).
5.4 Melkhygiene und -technik Die vorliegende Arbeit konnte zeigen, dass kontagiöse und umweltassoziierte Mastitiserreger
wie auch die so genannten minorpathogenen Bakterien in Mexiko weit verbreitet und am
Mastitisgeschehen beteiligt sind. Bei der Überprüfung der Betriebe zeigten sich
erwartungsgemäß viele Defizite insbesondere bei der Melkhygiene und der Melktechnik.
Nach THOMPSON und HOUSTON (1967) wirkt sich vor allem das Fehlen einer adäquaten
Melkhygiene auf eine schnelle Ausbreitung von Euterinfektionen aus. Die Milchviehhalter

91
der besuchten Betriebe waren im Allgemeinen unwissend über den Gesundheitsstatus ihrer
Herden, selbst wenn sehr hohe Infektionsraten vorlagen. Bei der Eingangsbefragung gaben
54% der Landwirte an, nur selten Probleme mit Mastitiden zu haben. 37,5% schätzen ihre
Mastitisproblematik als häufig ein, und immerhin 9,5% gaben an, dass ihre Kühe sehr häufig
an Euterentzündungen erkranken. Diese Selbsteinschätzung liegt fern der Realität von 62,9%
subklinischen und 2,5% klinischen Mastitiden zum Untersuchungszeitpunkt. Auch über den
bestehenden Zusammenhang von Hygiene, Zellzahlen der Einzeltiere und der Tankmilch
sowie der Milchleistung waren sich viele Tierhalter nicht bewusst. Die meisten Mastitiden in
Mexiko verlaufen subklinisch und werden daher keiner Behandlung zugeführt, weswegen die
sie verursachenden kontagiösen Erreger als Ansteckungsquelle für eutergesunde Kühe einer
Herde dienen. Die hohe Präsenz von subklinischen Reservoiren, das Fehlen eines
Gesundheitsmonitorings und das empirische Behandeln der klinischen Fälle zeigt große
Defizite im Management auf. Auch FRESE (2003) beschreibt in seinen Untersuchungen in
Milchviehbetrieben Äthiopiens das totale Fehlen von Kenntnissen über den Einfluss der
Melk-, Haltungs- und Stallhygiene sowie des Managements auf die Eutergesundheit, und
vermutet deswegen so hohe Mastitisprävalenzen in Äthiopien.
Da die Mehrzahl der mexikanischen Milchbetriebe mit halbextensiven und halbaufgestallten
Systemen geführt werden, und der Melkvorgang mit wenig bis keiner technischen Assistenz
durchgeführt wird, ist es sehr schwierig neue Praktiken vorzuschlagen und einzuführen.
Jedoch wurden die ungenügende Hygiene und das Herdenmanagement auf den meisten
Betrieben angesprochen. Eine Verbesserung der Mastitissituation in Jalisco wird nur durch
intensive Aufklärung der Betriebsleiter und Einführung von speziell auf die einzelnen
Betriebe angepassten Programmen zur Bekämpfung und Prävention der Mastitis zu erreichen
sein. Zur Erlangung und Sicherung einer stabilen Eutergesundheit müssen neben der
Melktechnik und Melkhygiene weitere Faktoren optimiert werden. Da die Haltung einen
entscheidenden Einfluss auf das Wohlbefinden der Tiere hat, sollte sie innerhalb der
gegebenen Voraussetzungen verbessert werden, einschließlich der Hygiene auf den Ausläufen
und in den Melkständen. Nur durch regelmäßige Beseitigung des Tierkotes kann der
Keimdruck nachhaltig gesenkt werden. Als weitere Maßnahme müssten die Tiere bedarfs-
und wiederkäuergerecht gefüttert werden, um ein Leistungsoptimum für jede einzelne Kuh zu
erreichen. Hierfür müssen Futteranalysen durchgeführt und anschließend der Bedarf jedes
einzelnen Tieres im jeweiligen Laktationsstadium berechnet werden. Insbesondere ein
Energie- oder Eiweißmangel beeinträchtigen die Abwehrkräfte des Gesamtorganismus und
des Euters. Wichtig ist der ständige Zugang zu Grundfutter guter Qualität. Die alleinige

92
Zufütterung von Kraftfutter während des Melkvorgangs stellt keine tiergerechte Lösung dar.
Besser wäre es, Kraftfutter häufig in kleinen Portionen auch außerhalb der Melkzeiten
anzubieten.
Eine fehlerhafte Melkarbeit und Melktechnik wirken sich ebenfalls negativ auf die
Eutergesundheit aus. Insbesondere die Melkzeuge müssen hygienische und technische
Mindestanforderungen erfüllen. In Jalisco waren jedoch viele Melkmaschinen veraltert und
wurden über Jahre nicht auf ihre Funktionstüchtigkeit hin überprüft. Als weiterer wichtiger
Bestandteil sollten die Zitzengummis mindestens zweimal im Jahr gewechselt werden, da
diese nach spätestens diesem Zeitraum ihre Elastizität verlieren. Sind sie überaltert bilden sich
Risse auf der Gummioberfläche, die keine ausreichende Reinigung ermöglichen, wodurch es
zu einer Keimansammlung kommt. Überalterte Zitzengummis beeinträchtigen direkt den
Zustand der Zitzen. Die Auswertung der Daten zeigte, dass die Prävalenz für S. aureus in
Herden mit Zitzengummis in sehr guten Zustand bei 2,9%, in Herden mit Zitzengummis in
nur ausreichendem Zustand jedoch bei 12,6% lag. Der Boden des Melkplatzes sollte befestigt
sein, so dass er mit Wasser gereinigt anschließend abtrocknen kann. Eine ausreichende
Wasserversorgung hierfür ist auch in der Trockenzeit sicherzustellen.
Nur durch eine optimale Melkhygiene können Ausbreitung und Vermehrung von
Mastitiserregern innerhalb einer Herde verringert werden. Für jeden Betrieb müssen die
einzelnen Maßnahmen auf das vorliegende Erregerspektrum, die örtlichen Begebenheiten und
Möglichkeiten angepasst werden. Die statistische Auswertung der Daten konnte zeigen, dass
die Prävalenz von S. aureus in Betrieben mit einer festen Melkreihenfolge bzw. mehreren
Melkgruppen signifikant geringer war als in Herden ohne Melkreihenfolge. Für die
Durchführung dieser Maßnahme ist die Herde in mindestens zwei Gruppen mit eutergesunden
bzw. mit kontagiösen Mastitiserregern infizierten Tieren aufzuteilen, und die eutergesunden
Tiere sind stets vor den kranken Tieren zu melken.
Des Weiteren konnten statistisch signifikant geringere Prävalenzen kontagiöser
Mastitiserreger in Herden in denen regelmäßig vorgemolken wird, gefunden werden.
Besonders niedrige Prävalenzen waren vorhanden, wenn dafür ein Vormelkbecher verwendet
wurde. Der große Nutzen des Vormelkbechers ist bei konsequenter Anwendung die
Verhinderung der Kontamination des Melkplatzbodens mit Mastitiserregern. Ferner können
durch diese zweimal tägliche Kontrolle klinische Entzündungen schnell erkannt und adäquat
behandelt werden. Auf diese Weise werden die Tiere schnell als Erregerreservoire
identifiziert. Auch die Euterreinigung zeigte sich als wichtige Einflussgröße auf die Prävalenz
mit kontgiösen Mastitiserregern. Diese wurde in vielen Betrieben nicht optimal durchgeführt.

93
Auf keinen Fall darf dasselbe Tuch für mehrere oder gar alle Kühe einer Herde benutzt
werden. Selbst wenn das Tuch in Desinfektionslösung eingelegt wird, ist mit einer
erheblichen bakteriellen Kontamination zu rechnen. Stattdessen muss für jede Kuh ein
frisches Reinigungstuch verwendet werden. Bei geringgradiger Verschmutzung reicht meist
eine trockene Reinigung der Zitzen. Angetrocknete Kotreste müssen sowohl zum Schutz der
Kuh als auch aus lebensmittelhygienischer Sicht restlos entfernt werden. Wird zum Reinigen
Wasser verwendet, so müssen die Euter und Zitzen anschließend auf jeden Fall getrocknet
werden, bevor die Melkzeuge angesetzt werden. Die Daten zeigten, dass die höchste S.
aureus-Prävalenz in den Herden nachgewiesen werden konnte, in denen die Zitzen zum
Melkzeitpunkt noch nass waren. In Herden ohne jegliche Euterreinigung ließ sich S. aureus
mit einer Prävalenz von 11,1% nachweisen, während je nach angewendeter
Reinigungsmethode Prävalenzen zwischen 3,1 und 7,6% auftraten. In Beständen mit
kontagiösen Mastitiserregern empfiehlt sich außerdem die Zwischendesinfektion der
Melkzeuge nach jeder gemolkenen Kuh (DVG 2002). Dies konnte durch den statistisch
signifikanten Unterschied der Nachweishäufigkeit von S. aureus bzw. S. agalactiae von 6,2%
bzw. 8,1% ohne Desinfektion zu 5,5% bzw. 3,8% mit Desinfektion belegt werden.
Einen weiteren wichtigen Faktor der Mastitiskontrolle stellt das Zitzendippen dar (HOGAN et
al. 1987). Hierbei werden direkt nach der Abnahme des Melkzeuges Mastitiserreger auf den
Zitzen sämtlicher Tiere durch Zitzentauchen in eine geeignete Desinfektionslösung abgetötet.
Das Dippmittel ist hierbei mindestens einmal täglich zu erneuern (ZSCHÖCK et al.1998).
Alle Kühe eines Betriebes müssen 6-8 Wochen vor dem Abkalben trockengestellt werden.
Das schlagartige Trockenstellen von einer Melkzeit auf die andere ist hierfür die Methode der
Wahl (ZSCHÖCK et al 1998). Das in vielen mexikanischen Betrieben bevorzugte langsame
Trockenstellen durch das Überspringen von Melkzeiten bei gleichzeitiger Reduzierung der
Futtermenge ist nicht zu empfehlen. Sämtliche Tiere sollten außerdem unter antibiotischem
Schutz trockengestellt werden, wobei es zu einer Ausheilung subklinischer Euterinfektionen
und zur Senkung von Neuinfektionen kommt (ZSCHÖCK et al. 1998). Das Merzen
therapieresistenter Tiere ist für die Kontrolle und Prävention der Mastitis anzuraten (DVG
2002).
In Mexiko werden all diese Maßnahmen bisher nur sporadisch bzw. in vielen Betrieben
überhaupt nicht durchgeführt.
Zur Kontrolle der umweltassoziierten Mastitiserreger belegten die Daten, dass sich eine
vermehrte Auslaufhygiene und eine verbesserte Euterreinigung vor dem Melken als wirksam
erweisen. Die Prävalenz von S. uberis war interessanterweise in Betrieben mit Durchführung

94
der Melkzeugzwischendesinfektion geringer als in den übrigen Betrieben. Bei den coliformen
Keimen konnte ein positiver Einfluss durch Vormelken und Zitzentauchen gezeigt werden.
Die Auswertung der Daten in Bezug auf die Corynebacterium spp. zeigte, dass sie bei
Einhaltung einer Melkreihenfolge, bei Durchführung einer feuchten Euterreinigung mit
anschließendem Abtrocknen der Zitzen sowie bei gezieltem antibiotischem Trockenstellen
aller Tiere statistisch signifikant seltener nachweisbar waren.
Anhand der vorgeführten Verbesserungsvorschläge können für jeden einzelnen Betrieb
umsetzbare vorbeugende, kontrollierende und korrigierende Maßnahmen ausgearbeitet und
den Betriebsleitern unterbreitet werden, mit dem Ziel, Milch mit einem besseren hygienischen
und gesundheitlichen Status zu produzieren.
Der von uns in Mexiko bestimmte durchschnittliche Zellzahlwert von 623.000 Zellen/ml
Milch ist die direkte Folge der schlechten Mastitisprävention und –kontrolle. Auch wenn in
Mexiko einige Molkereien Zellzahlbestimmungen der Anlieferungsmilch durchführen und
langfristig den Aufkauf von Milch mit weniger als 200.000 Zellen/ml anstreben, gibt es
bislang noch keine mit der Milchgüte-Verordnung vergleichbare gesetzliche Verankerung der
Mindestanforderungen von Anlieferungsmilch in den Normas Oficial de México. Eine
gesetzliche Zellzahlobergrenze wie diese in Deutschland in der Milchverordnung vom 1. Juli
2000 festgesetzt ist, wäre vonnöten. Nur so können die Milchviehhalter zu einer verstärkten
Mastitiskontrolle, die letztendlich sowohl aus wirtschaftlicher, tierschützerischer als auch aus
lebensmittelhygienischer Sicht erforderlich ist, bewegt werden.
Die momentan gültigen offiziellen Normen bieten keine Grundlage für eine derartige
Forderung, jedoch könnte eine Aus- bzw. Überarbeitung der gängigen Normen den
Problemen Abhilfe schaffen. Es bleibt zu hoffen, dass das nationale Komitee für Normierung
mit der Umsetzung des Projektes PROY-NMX-F-700-COFOCALEC-2003, das die
physiochemischen und biologischen Eigenschaften der Rohmilch regeln soll, diesen
Anforderungen nahe kommen wird. Mit Hilfe einer neuen Norm könnte ein wichtiger Schritt
in Richtung Mastitisprävention und Verbraucherschutz in Mexiko erreicht werden.

95
6 Zusammenfassung
Sybille Petra Jäger
Untersuchungen zur Eutergesundheit in Milchviehbeständen des Bundesstaates Jalisco,
Mexiko
Ambulatorische und Geburtshilfliche Tierklinik der Veterinärmedizinischen Fakultät der
Universität Leipzig
und
Abteilung Veterinärmedizin des Landesbetriebes Hessisches Landeslabor, Gießen
Eingereicht im März 2006
(116 S., 6 Abb., 15 Tab., 242 Lit., Anhang mit 13 Abb., 2 Tab.)
Schlüsselworte: Eutergesundheit, Mastitisinzidenz, Jalisco, Mexiko, Erregerprävalenzen,
Zytobakteriologie, PCR, Pulsfeldgelelektrophorese, Toxine, Melkhygiene,
Sanierungsmaßnahmen.
Die vorliegende Arbeit hatte zum Ziel, das Vorkommen subklinischer und klinischer
Eutergesundheitsstörungen in 33 Milchviehherden in Jalisco, Mexiko, aufzuzeigen. Von 2937
mittels CMT untersuchten Eutervierteln zeigten 1996 (66,9%) eine positive und hiervon 1087
(37%) eine deutlich bis stark positive Reaktion. Im Abgleich mit den bakteriologischen
Untersuchungen ergab sich eine Prävalenz an subklinischen Mastitiden in Höhe von 43,7%.
Klinische Mastitiden ließen sich zu 2,5% nachweisen.
53,8% der untersuchten Milchproben zeigten bakteriologisch keinen Keimgehalt. Aus den
übrigen Proben konnten zu 15,6% KNS, 14,0% Corynebacterium spp. 6,7% S. agalactiae,
5,9% S. aureus, 4,1% coliforme Keime, 3,7% Streptococcus spp. und 1,7% sonstige Keime
(Bacillus spp., Nocardia spp., Candida spp.) isoliert werden. Diese Ergebnisse zeigen, dass
neben den kontagösen Mastitiserregern wie S. aureus und S. agalactiae minorpathogene
Erreger zu einem hohen Anteil am Mastitisgeschehen in Jalisco beteiligt sind.
Mit Hilfe der Pulsfeldgelelektrophorese konnte gezeigt werden, dass in jedem der Betriebe, in
denen S. aureus isoliert werden konnte, überwiegend ein einzelner Genotyp für das
Mastitisgeschehen verantwortlich war. Die betriebsspezifischen Genotypen zeigten
überwiegend eine enge Verwandtschaft zu den Genotypen aus anderen Betrieben. Hierdurch
konnte der kontagiöse Charakter dieses Mastitiserregers und ein dominierendes Vorkommen
bestimmter S. aureus-Klone belegt werden.

96
Durch die Besichtigung der Betriebe und Befragung der Betriebsleiter konnten Defizite in der
Haltungs- und Melkhygiene aufgezeigt werden. Sie korrelierten statistisch signifikant mit
erhöhter Mastitisprävalenz und mit erhöhten Nachweisraten von kontagiösen
Mastitiserregern. Vorbeugende, kontrollierende und korrigierende Maßnahmen wurden
vorgeschlagen und erörtert. Durch die genannten Programme zur Vorbeuge und Kontrolle der
Mastitis könnte die Milchproduktion in Jalisco und Mexiko um bis zu 20% gesteigert und
somit das starke Defizit des mexikanischen Milchsektors sowie die hohen
Milchpulverimportmengen minimiert werden. Da die verminderte bakteriologische Qualität
der Milch von an subklinischer Mastitis erkrankten Kühen außerdem, insbesondere vor dem
Hintergrund des hohen Rohmilchkonsums, ein mögliches gesundheitliches Risiko für den
Verbraucher darstellt, wurden 17 der aus den Betrieben isolierten S. aureus-Isolate auf ihr
Toxinbildungsvermögen und ihre Toxin-Genmuster hin untersucht. Nur bei einem Stamm
konnte ein Amplifikat für das SEI Gen nachgewiesen werden. Keiner der untersuchten
Stämme beherbergte Gene für die SEA, SEB, SEC, SED, SEE, SEG, SEJ und das TSST-1.
Trotz dieser geringen Nachweisrate sollte in Mexiko durch Einführung eines konsequent
durchgeführten Mastitiskontrollprogramms Milch mit einer höheren hygienischen Wertigkeit
produziert und somit das vom Lebensmittel Milch für den Verbraucher ausgehende
gesundheitliche Risiko reduziert werden.

97
7 Summary
Sybille Petra Jäger
Investigations on the udder health in dairy herds of the federal state Jalisco, Mexico.
Large Animal Clinic for Theriogenology and Ambulatory Services, Faculty of Veterinary
Medicine, Universitiy of Leipzig
and
Abteilung Veterinärmedizin des Landesbetriebes Hessisches Landeslabor, Gießen 2006
Submitted in March 2006
(116 pages, 6 figures, 15 tables, 242 lit; appendix with 13 figures, 2 tables)
Keywords: udder health, mastitis incidence, Jalisco, Mexico, pathogen prevalences,
cytobacteriology, PCR, pulsed-field gel electrophoresis, toxins, milking hygiene, control
programs.
Aim of the present work was to prove the occurrence of subclinic and clinic disturbances of
udder health in 33 herds of dairy cattle in Jalisco, Mexico. 1996 (66.9 %) out of 2937 udder
quarters examined by means of CMT showed a positive reaction, 1087 (37%) out of these
reactions were from clearly up to significantly positive reactions. Compared to the
bacteriological examinations the prevalence for subclinic mastitides came up to 43.7%. On
the other hand clinical mastitides could be proved in 2.5%.
In 53.8% of the examined quarter milk samples there was no bacteriological pathogen
content. From the rest of the samples we could isolate CNS (15.4%), Corynebacterium spp.
(13.9%) S. agalactiae (6.6%), S. aureus (5.8%), coliform pathogens (3.6%) and others
(Bacillus spp., Nocardia spp., Candida spp.) (1.7%). These results demonstrate a significant
share of minor pathogens beside contagious mastitis pathogens as S. aureus and S. agalactiae
in masititis incidents in Jalisco.
By means of the pulsed-field-gel-electrophoresis we proved that in each of those farms where
S. aureus had been isolated, only one genotype was responsible for mastitis incidents. The
farm specific genotypes mostly showed a close relationship to the genotypes of other farms.
Therefore the contagious character of mastitis pathogens and the dominating occurrence of
certain S. aureus clones could be proved.
By inspecting the farms and questioning work managers deficits in hygienic keeping and
milking could be demonstrated. Statistically they correlate significantly with an increased
mastitis prevalence and increased proving rates of contagious mastitis pathogens.
Prophylactic, controlling and correcting measurements were supposed and discussed. Those

98
prophylactic and controlling programmes for mastitis could elevate milk production in Jalisco
and Mexico by up to 20% and therefore reduce the large deficit in the Mexican milk sector as
well as the large amount of import of powdered milk. The decreased bacteriological quality of
milk of cows with subclinical mastitis means a possible health risk for the consumer,
especially considering the consumption of raw milk. Therefore 17 S. aureus isolates of those
farms were examined according to their toxigenic ability and their toxin gene pattern. Only in
one strain an amplification for the SEI gene could be proved. None of the examined strains
contained genes for SEA, SEB, SEC, SED SEE, SEG, SEJ and TSST-1.
In spite of this low proving rate milk of a higher hygienic value should be produced in
Mexico by introducing a mastitis controlling programme carried out consequently and thereby
reducing the health risk of milk as a comestible for the consumer.

99
8 Literaturverzeichnis
Aarestrup FM, Wegener HC, Rosdahl VT, Jensen NE. Staphylococcal and other bacterial species associated with intramammary infections in danish dairy herds. Acta Vet Scand. 1995;36:475-87. Abdella M. Bacterial causes of bovine mastitis in Wondogenet, Ethiopia. J Vet Med B.1996; 43:379-84. Abdulmawjood A, Lämmler C. Amplification of 16S ribosomal RNA gene sequences for the identification of streptococci of Lancefield group B. Res Vet Sci. 1999; 67:159-62. Arbeitsgemeinschaft Deutscher Rinderzüchter e.V. Jahresbericht der Milchproduktion in Deutschland. 2004. Alarcón RF. Frecuencia de la mastitis en vacas Holstein-Cebu y susceptibilidad quimiotherapeutica de las bacterias asociadas. Estudio retrospectivo. (Tesis de Licenciatura). México. D.F : Facultad de Medicina Veterinaria y Zootecnica. UNAM. México. D.F.; 1997. Annemüller C, Lämmler C, Zschöck M. Genotyping of Staphylococcus aureus from bovine mastitis.Vet Microbiol. 1999;69:217-24.
Ariznabarreta A, Gonzalo C, San Primitivo F. Microbiological quality and somatic cell count of ewe milk with special reference to staphylococci. J Dairy Sci. 2002; 85:1370-5. Arreguín Reyes BR. Normalizacacíon de la leche en México, avances y perspectivas. Proceedings del III Congreso Nacional de Control de Mastitis y Calidad de la leche 21 al 23 de junio del 2003-05-24 en Léon, Gto. México; 2003. Avila TS, Blanco OMA, Romero AT. Mastitis y producción de leche en el trópico húmedo. México: SUA-FMVZ Facultad de Medicina Veterinaria y Zootecnia, UNAM, México D.F.; 1991. Avila TS, Gutiérrez, CAJ, Sanchez GJI, Canizal JE, Torres VS. Prevalencia de mastitis y glandulas improductivas en hatos pequeños en la cuenca lechera de Xochimilco, México, D.F. Proceedings del III Congreso Nacional de Control de Mastitis y Calidad de la leche 21 al 23 de junio del 2001-05-24 en Léon, Gto. México; 2001. Baird-Parker AC. Family Micrococcacae. In: Gibbons NE. Bergey´s manual of determinative bacteriology, 8th Ed. The Williams & Wilkins Comp., Baltimore; 1974:478. Baker CJ. Group B streptococcal infections. In: Stollermann GH. (ed) Advances in internal medicin, Yearbook medical publishers, Chicago; 1980. Bannermann TL, Hancock GA, Tenover FC, Miller JM. Pulsed-field gel electrophoresis as a replacement for bacteriophage typing of Staphylococcus aureus. J Clin Microbiol. 1995;33: 551-5.

100
Bassegio N, Mansell PD, Browning JW, Browning GF. Strain differentiation of isolates of streptococci from bovine mastitis by pulsed-field gel electrophoresis. Mol Cell Prob. 1997; 11:349-54. Becker H. Streptococcus agalactiae (Group B streptococci). In: The significance of pathogenic microorganisms in raw milk. International Dairy Federation. Bruessels, Belgium. 1994:43-54.
Berrwerth W, Köser A. Streptokokken verschiedener serologischer Gruppen als Mastitiserreger und ihr Vorkommen außerhalb des Rindereuters. Milchwissenschaft 1965;20: 590-3. Benites NR, Guerra JL, Melville PA, Costa EO. Aetiology and histopathology of bovine mastitis of espontaneous occurrence. J Vet Med B Infect Dis Vet Public Health. 2002;49:366-70.
Bergdoll MS. Enterotoxins. In: Montie TC, Kadis S, Ajl SJ (eds). Microbial toxins. Academic press, New York and London. 1970;3:265-26. Bergdoll MS. Staphylococcus aureus. In: Doyle MP (ed). Foodborne bacterial pathogens. New York and Basel. Marcel Dekker. 1989:463-23. Bergmann A. Escherichia coli-, Klebsiella- und Enterobacter-Mastitiden. In: Wendt K, Bostedt H, Mielke H, Fuchs HW (eds). Euter- und Gesäugekrankheiten. Gustav Fischer Verlag Jena, Stuttgart; 1994. Bernheimer AW, Linder R, Avigad LS. Nature and mechanism of action of the CAMP protein of group B streptococci. Infect Immun. 1979;23:838-44. Bishi AS. Cross-sectional and longitudinal prospective study of bovine clinical and sub-clinical mastitis in peri-urban and urban dairy production systems in the Addis Ababa region, Ethiopia. (Msc thesis). Berlin: Addis Ababa University and Free Universitiy of Berlin, Germany; 1998. Black RT, Marshall RT, Bourland CT. California mastitis test reactivity and bacterial invasions in quarters infected with Corynebacterium bovis. J Dairy Sci. 1972;55:1016-7. Bramley AJ. Infection of the udder with coagulase-negative micrococci and Corynebacterium bovis. In: Dood FH, Griffin TK, Kingwill RG (eds). Proceedings, IDF Seminar Mastitis Control, Bruessels, Belgium. 1975;85:377-81. Bramley AJ, Shean MFH, Kingwill RG. Proceedings of XX International Dairy Congress Paris. 1978:142. Bramley AJ. The effect of subclinical Staphylococcus epidermidis infection of the lactating bovine udder on its susceptibility to infection with Streptococcus agalactiae or Escherichia coli. Br Vet J. 1978;134:146-51. Bramley AJ, Dodd FH. Reviews of the progress in dairy science: Mastitis control-progress and prospects. J Dairy Res. 198;51:481-512.

101
Brückler J, Schwarz S. Staphylokokken-Infektionen und –Enterotoxine. In: Handbuch der bakteriellen Infektionen bei Tieren. Bd.2/ Teil1. Gustav Fischer Verlag Jena, Stuttgart. 1994. Bryan FL. Epidemiology of milk-borne diseases. J Food Prot. 1983;46:637-49. Bone FJ, Bogie D, Morgan-Jones SC. Staphylococcal food poisoning from sheep milk cheese. Epidemiol Inf. 1989;103:449-58. Bourland CT. Mastitis due to Corynebacterium bovis and estrogen interaction. J Dairy Sci. 1967;50:978. Brolund L. Cell counts in bovine milk. Causes of variation for diagnosis of subclinical mastitis. Review. Acta Vet Scand Suppl. 1985;80:1-123. Busato A, Trachsel P, Schallibaum M, Blum JW. Udder health and risk factors for subclinical mastitis in organic dairy farms in Switzerland. Prev Vet Med. 1997;44:205-20. Ceron-Muñoz M, Tonhati H, Duarte J, Oliveira J, Muñoz-Berrcal M, Juardo-Gamez H. Factors affecting somatic cell counts and their relations with milk and milk constituent yield in buffalos. J Dairy Sci. 2002;85:2885-9. Cobb RW, Walley T. Corynebacterium bovis as a probable cause of bovine mastitis. Ebenda. 1962;74:101. Correa MGP, Marin JM. O-serogroups, eae gene and EAF plasmid in Escherichia coli isolates from cases of bovine mastitis in Brazil. Vet Microbiol. 2002;85:125-32. Christie R, Atkins NE, Munch-Petersen D. A note on a lytic phenomenon shown by group B streptococci. Aust J Exp Biol Med Sci. 1944;22:197-200.
Cullen GA. The ecology of Streptococcus uberis. Br Vet J. 1966;122:333-9. Cullen GA. Isolation of Streptococcus uberis from the rumen of cows and from soil. Vet Rec. 1969;85:114-18. Daleel EE, Frost AJ. Some observations on the bacterial flora of the bovine tonsil. Br Vet J. 1967;123:232-36. Daniel RC, O`Boyle D, Marek MS, Frost AJ. A survey of clinical mastitis in South-East Queensland dairy herds. Aust Vet J. 1982;58:143-7. Davidson TJ, Dohoo IR, Donald AW, Hariharan H, Collins K. A cohort study of coagulase negative staphylococcal mastitis in selected dairy herds in Prince Edward Island. Can J Vet Res. 1992;56:275-80. Dego OK, Tareke F. Bovine mastitis in selected areas of southern Ethiopia. Trop Anim Health Prod. 2003;35:197-205. Del Valle MR, Aguilar Cruz C, Hernández Tinajero A. Efectos económico-sociales de la política neoliberal en el sistema lácteo mexicano. La reestructuración productivo en dos microregiones; 1997.

102
Del Valle MR. El sistema lacteo mexicano, Investigadora Titular del Instituto de Investigaciones Económicas de la UNAM México; 1997. Del Valle MR. Producción de leche de gandado bovino, Investigadora Titular del Instituto de Investigaciones Económicas de la UNAM México; 2002. Deutsche Veterinärmedizinische Gesellschaft. Leitlinien zur Entnahme von Milchproben unter antiseptischen Bedingungen und Leitlinien zur Isolierung und Identifizierung von Mastitiserregern. Sachverständigenausschuss „Subklinische Mastitis“ des Arbeitskreises Eutergesundheit der Fachgruppe Milchhygiene des Arbeitsgebietes Lebensmittelhygiene, 2000, DVG Giessen. Deutsche Veterinärmedizinische Gesellschaft. Leitlinien zur Bekämpfung der Mastitis des Rindes als Bestandsproblem. Fachgruppe „Milchhygiene“, Sachverständigenausschuss „Subklinische Mastitis“. 2002, DVG Giessen. Devriese LA. Identification of clumping factor negative staphylococci isolated from cow´s udders. Res Vet Sci. 1977;27:313-20. Devriese LA. Streptococcal ecovars associated with different animal species: epidemiological significance of serogroups and biotyps. J Appl Bacteriol. 1991;71:478-83. Devriese LA, Baelea M, Vaneechoutteb A, Haeseboucka MF. Identifiaction and antimicrobial susceptibility of Staphylococcus chromogenes isolates from intramammary infections of dairy cows. Vet Microbiol. 2002;87:175-82. Direcion general de ganadería. Situacion actual y perspectiva de la produccion de leche de ganado bovino en México, 1990-2000. Centro de Estadistica Agropecuaria, Direccion General de Estudios del Sector Acropecuaria-SAGAR y del Departamento de Agricultura de los Estados Unidos Mexicanos; 1999. Dixon WJ. BMDP Statistical software manual, Volume 1 and 2. University of California Press, Berkeley, Los Angeles, London. 1993. Djabri B, Barielle N, Beaudeau F, Seegers H. Quarter milk somatic cell count in infected diary cows: a meta analysis. Vet Res. 2002;33:335-57. Dmitriev A, Tkaciková L, Suvorov A, Kantikova M, Mikula I, Totolyan A. Comparative genetic study of group B streptococcal strains of human and bovine origin. Folia Microbiol. 1999;44:449-53. Dohoo IR, Meek AH. Somatic cell counts in bovine milk. Can Vet J. 1982;23:119-25. Domínguez MP. Milk quality and mastitis control in México. National Mastitis Council Annual Meeting Proceedings. México D.F.. 1996:30-31. Edwards SJ, Smith GS. Epidemiology of mastitis in three dairy herds. J Comp Pathol Therap. 1966;46:211.

103
El-Sayed A, Alber J, Jäger S, Zschöck M, Wolter W, Castañeda VH. Comparative studies on genotypic properties of Staphylococcus aureus isolated from clinical and subclinical mastitic cows in México. Revista Veterinaria México. 2006;37. Erskine RJ, Eberhart RJ. Herd benefit-to-cost ratio and effects of a bovine mastitis control program the includes blitz treatment of Streptococcus agalactiae. J Am Vet Med Ass. 1990;196:1230-5. Erskine RJ, Bartlett PC, Johnson GL, Halbert LW. Intramuscular administration of ceftiofur sodium versus intramammary infusion of penicillin/novobiocin for treatment of Streptococcus agalactiae in dairy cows. J Am Vet Med Ass. 1996;208:258-60. Erskine RJ, Eberhart RJ, Hutchinson LJ, Spencer SB, Campbell MA. Incidence and types of clinical mastitis in dairy herds with high and low somatic cell counts. J Am Vet Med Ass. 1996;192:761-5.
Evenson ML, Hinds MW, Bernstein RS, Bergdoll MS. Estimation of human dose of staphylococcal enterotoxin A from large outbreak of staphylococcal food poisoning involving choclate milk. Int J Food Microbiol. 1988;7:311-316. Farnsworth RJ. Indications of contagious and environmental mastitis pathogens in a diary herd. Proc. 26 th Ann Meet Natl Mastitis Council. 1987;26:151-5. Fehlhaber K. Streptokokken- und unspezifische Lebensmittelvergiftungen. In: Einführung in die Lebensmittelhygiene. Sinell HJ (ed). Parey Verlag, Stuttgart; 2004. Fira (Banco de México). Oportunidades de desarollo de la lechería en México. Boletín Informativo v. XXIX. 1996;294:1-36. Fitzgerald RJ, Meaney WJ, Hartigan PJ, Smyth CJ, Kapur V. Fine-structure molecular epidemiological analysis of Staphylococcus aureus recoverd from cows. Epidemiol Infect. 1997;119:261-9. Fitzgerald RJ, Su C, Aarestrup FM, Dangler CA, Sordillo LM. Prevalence of coagulase gen polymorphism in Staphylococcus aureus isolates causing bovine mastitis. Can J Vet Res. 1997;59:124-8. Fitzgerald RJ, Sturdevant DE, Mackie SM, Gill SR, Musser JM. Evolutionary genomics of Staphylococcus aureus: Insight into the origin of methicillin-resistant strains and the toxic shock syndrome epidemic. Proceedings of the National Academy of Sciences of the United States of America. 2001;98:8821-6. Finch LA, Martin DR. Human and bovine group B streptococci: two distinct populations. J Appl Bacteriol. 1984;57:273-8. Frese ML. Eine Studie zur Prävalenz und Inzidenz boviner Mastitis in großen und kleinen Milchproduktionsbetrieben urbanen und periurbanen Regionen von Addis Ababa und Debre Zeit, Äthiopien. [Dissertation vet. med.]. Berlin: Univ. Berlin; 2003.

104
Frost AJ, Wanasinghe DD, Woolcovk JB. Some factors affecting selective adherence of microorganisms in the bovine mammary gland. Infect Immun. 1988;15:245-53. Fox LK, Gay JM. Contagious mastitis.Veterinary clinics of North America. Food Anim Prac. 1993;9:475-87. Gianneechini R, Concha C, Rivero R, Delucci I, Moreno Lopez J. Occurrence of clinical and sub-clinical mastitis in dairy herds in the west littoral region in Uruguay. Acta Vet Scand. 2002;43:221-30. Gilbert RJ. Staphylococcal food poisoning and botulism. Postgraduate Medical J. 1974;50: 603-11. Gilmour A, Harvey J. Staphylococci in milk and milk products. Soc Appl Bacteriol Symp Ser. 1990;19:147-66. Giraudo J, Rampone H, Martinez L, Calzolari A. Recuento de células somáticas en leche bovina de cuartos mamarios con y sin aislamiento microbiano (Counting of somatic cells in bovine milk from mammary quarters with and without isolation of micro-organisms).Revista de Medicina Veterinaria. 1995;76:6-10. Goering RV. The molecular epidemiology of nosocomial infection. An overview of principles, applications and interpretations. In: Spencer et al. (eds). Rapid detection of infectious agents. Plenum Press, New York, 1998:131-157. Goldberg JJ, Wildmann EE, Pankey JW, Kunkel JR, Howard DB, Murphy BM. The influence of intensively managed rotational grazing, traditional continuous grazing and confinement housing on bulk tank milk quality and udder health. J Dairy Sci. 1992;75:96-104. Gordillo ME, Singh KV, Baker CJ, Murray BE. Typing of group B streptococci: Comparison of pulsed-field gel electrophoresis and conventional electrophoresis. J Clin Microbiol. 1993;31:1430-4. Gray DM, Schalm OW. The mastitis variable in milk yield as estimated by the California Mastitis test. Am J Vet Res.1962; 23:541. Grothues D, Tümmler B. New approaches in genome analysis by pulsed-field gel electrophoresis application to the analysis of Pseudomonas species. Mol Microbiol. 1991;5: 2763-76.
Hahn G, Heeschen W, Tolle A. Wechselbeziehungen zwischen Infektionen mit Infektionen der serologischen Gruppe B bei Mensch und Rind. Fortschritte der Veterinärmedizin. 1972;17:189-197. Hallen-Sandgren CH. Mjölk kor. Natur och Kultur/LTs förlag, Helsingborg, Sweden. 2000:179-200. Harmon RJ, Crist WL, Hemken RW, Langlois BE. Prevalence of minor udder pathogens after intramammary dry treatment. J Dairy Sci. 1986;69:843-9.

105
Harmon RJ, Langlois BE. Mastitis due to coagulase-negative Staphylococcus species. Agricult Pract. 1989;10:29-34. Heeschen W. In: Wendt K, Bostedt H, Mielke H, Fuchs HW. (eds). Euter- und Gesäugekrankheiten. Gustav Fischer Verlag Jena, Stuttgart. 1998:145-6. Heeschen WH. Milk hygiene and milk safety in the european and international markets. Kieler Milchwirtschaftliche Forschungsberichte 1998;50:53-77. Heidrich HJ, Renk W. Krankheiten der Milchdrüse bei Haustieren.Verlag Paul Parey, Berlin-Hamburg; 1963. Hejlicek K. Mastitis durch Streptococcus agalactiae (Gelber Galt). In: Wendt K, Bostedt H, Mielke H, Fuchs HW. (eds). Euter- und Gesäugekrankheiten. Gustav Fischer Verlag Jena, Stuttgart. 1998:332-46. Heringstad B, Klemetsdal G, Ruane J. Selection of mastitis resistance in dairy cattle: a review with focus on the situation in the nordic countries. Livestock Prod Sci. 2000;64:95-106. Hogan JS, White DG, Pankey JW. Effects of teat dipping on intramammary infections by staphylococci other than Staphylococcus aureus. J Dairy Sci. 1987;70:873-9. Hogan JS, Smith KL, Hoblet KH. Field survey of clinical mastitis in low somatic cell count herds. J Dairy Sci. 1989;72:1547-56. Hughes DL. Some epidemiological patterns in bovine mastitis. Vet Rec. 1960;72:485-98. Inegi. Resulados Defintivos del VII Censo Agrícola-Ganadero de los Estados Unidos Méxicanos 1991 del Instituto Nacional de Estadíistica, Geografía e Informática; 1991. Inegi. Anuário 2000. Censo Agrícola-Ganadero del Gobierno del Estado Jalisco. 2000:401-17.
Janzen, JJ. Economic losses resulting from mastitis. A review. J Dairy Sci. 1970;53:1151-61.
Jarraud S, Peyrat MA, Lim A, Tristan A, Bes M, Mougel C, Etienne J, Vandenesch F, Bonneville M, Lina G. Egc, a highly prevalent operon of enterotoxin gene, forms a putative nursery of superantigens in Staphylococcus aureus. J Immunol. 2001;166:669-77. Jayarao BM, Dore JJ, Oliver SP. Restriction fragment length polymorphism analysis of 16S ribosomal DNA of Streptococcus and Enterococcus species of bovine origin. J Clin Microbiol. 1992;33:2235-40. Jensen NE. Experimental bovine group-B streptococcal mastitis induced by strains of human and bovine origin. Nord Veterinaermed. 1982;34:441-50. Jones GM, Pearson RE, Heald CW, Vinson WE. Milk loss, somatic cell counts and udder infections in Virgina herds. Proc. 21st Annu. Mtg. Natl. Mastitis Council, Louisville, KY. Natl. Mastitis Council, Inc. Arlington, VA. 1982:31.

106
José Zorrila R, Francisco Ramirez JV, Raul Martinez. Jalisco en el sistema lechero nacional, situación actual y perspectivas. Publicación Especial Numero 1, Diciembre de 1996, Instituto nacional de investigaciones forestales, agricolas y pecuarias (INIFAP), Centro de investigacion regional del pacifico, Centro campo experimental, Centro de Jalisco, Av. Morelos 1514 Col. Americana, Guadalajara, Jalisco, Mexico. 1996;1. Juarez E, Ruiz SH, Avila TS. Relación entre la prueba de California y los tipos de bacterias, aisladas de vacas Holstein-friesian del Valle de México. Memorias del VI Congreso Nacional de Buiatría; 1980 septiembre 12-14. Mérida (Yucatán) México. México, (D.F.) Associación Mexicana Veterinarios Especialistas en Bovinos, A.C., 1980. Kaipainen T, Pohjanvirta S, Shpigel NY, Shwimmer A, Pyörälä S, Pelkonen S. Virulence factors of Escherichia coli isolated from bovine clinical mastitis.Vet Microbiol. 2002;85:37-46. Kapur V, Schischo WM, Greer RS, Whittam TS, Musser JM. Molecular population genetic analysis of Staphylococcus aureus recovered from cows. J Clin Microbiol. 1995;33:376-80. Keefe GP. Streptococcus agalactiae mastitis: A review. Can Vet J. 1997;38:429-37. Kielwein G. Leitfaden der Milchkunde und Milchhygiene. 3. Aufl. Blackwell-Verlag, Berlin; 1994. King JS. Streptococcus uberis: A review of its role as a causative organism of bovine mastitis. Characteristics of the organism. Br Vet J. 1981;137:36-52. Kleinschroth E, Rabold K, Deneke J. Mastitis. Euterkrankheiten erkennen, vorbeugen und behandeln. top agrar. 1994:8-16. Kloos WE, Schleifer KH. Genus IV. Staphylococcus Rosenbach 1884. In: Sneath PHA, Nair NS, Sharpe ME, Holt JG (eds). Bergey´s manual of systematic bacteriology. Vol. 2. Williams and Willkins, Baltimore. 1986;2:1013-35. Kloppert B, Wolter W, Risse K, Zschöck M. Erregerspektrum bei boviner subklinischer Mastitis in hessischen Milcherzeugerbetrieben. In: Tagungsbericht des 23. Kongresses der Deutschen Veterinärmedizinischen Gesellschaft e.V. in Bad Nauheim 13.04-16.04.1999:350-5. Kudinha T, Simango C. Prevalence of coagulase-negative staphylococci in bovine mastitis in Zimbabwe. Tydsk S Afr Vet. 2002;73:62-5. Lacey RW. Are resistant bacteria from animal and poultry an important threat to the treatment of human infections? In: Jolly DW, Miller DJ, Ross DB. 5th AVI-Symposium, “Ten years on the swann” 1981:127-44. Lacy-Hulbert SJ, Hillerton JE. Physical characteristics of the bovine teat canal and their influence on suceptibility to streptococcal infection. J Dairy Res. 1994;43:395-404. Larsen HD, Huda A, Eriksen HNR, Jensen NE. Difference between Danish bovine and human Staphylococcus aureus isolates in posession of superantigens. Vet Microbiol. 2000;76:153-62.

107
Lämmler C, Wibawan IWT, Pasaribu FH, Warsa UC. Vergleichende Untersuchungen von Streptokokken der serologischen Gruppe B, isoliert aus Untersuchungsmaterial von Rind und Mensch in Deutschland und Indonesien. Tierärztl Umsch. 1993;48:171-75. Lämmler C, Hahn G. Streptokokken-Infektionen. In: Handbuch der bakteriellen Infektionen bei Tieren Band 2/Teil 2, Gustav Fischer Verlag Jena, Stuttgart; 1994. Lafi S, Al-Rawashdeh O, Na`was T, Hailat N. National cross-sectional study of mastitis in dairy cattle in Jordan. Trop Anim Health Prod. 1994;26:168-74. Lam TJGM, Lipman LJA, Schukken YH, Gaastra W, Brand A. Epidemiological characteristics of bovine clinical mastitis caused by Staphylococcus aureus and Escherichia coli studied by DNA fingerprinting. Am J Vet Res. 1996;57:9-42. Lam TJGM, Schukken YH, van Vliet JH, Grommers FJ, Tielen JM, Brand A. Effect of natural infection with minor pathogens on susceptibility to natural infection with major pathogens in the bovine mammary gland. Am J Vet Res. 1997;58:17-22. Letertre C, Perelle S, Dilasser F, Fach P. A strategy based on 5' nuclease multiplex PCR to detect enterotoxin genes sea to sej of Staphylococcus aureus. Mol Cell Prob. 2003;17:227-35. Lim SK, Joo YS, Moon JS, Lee AR, Nam HW, Wee SH, Koh HB. Molecular typing of enterotoxigenic Staphylococcus aureus isolated from bovine mastitis in Korea. J Vet Med Sci. 2004;66:581-4. Losada H, Bennett R, Cortes J, Viejra J, Arias L, Soriano R, Avaleta Z. The production of milk from dairy herds in the suburban conditions of Mexico City: II. The case of the region of Xochimilco in the south-east of the city. Livestock Res Rural Dev. 1998;10. Maslow JN, Slutsky AM, Arbeit AD. Application of pulsed-field gel electrophoresis to molecular epidemiology. In: Persing DH, Smith TF, Tenover FC, White TJ (eds). Diagnostic molecular microbiology. American Society of Microbiology, Washington D.C. 1993:563-72. Matos JS, White DG, Harmon RJ, Langlois BE. Isolation of Staphylococcus aureus from sites other than the lactating mammary gland. J Dairy Sci. 1991;74:1544-9. Matsunaga T, Kamata S, Kakinichi N, Uchida A. Characteristics of Staphylococcus aureus ioslated of peracute, acute and chronic bovine mastitis. J Vet Med Sci. 1993;55:297-300. Matthews KM, Harmon RJ, Langlois BE, Christ WL, Hemken RW. Use of a latex teat dip with germicide during the prepartum period. J Dairy Sci. 1986;71:1940. Matthews KR, Harmon RJ, Langlois BE. Effect of naturally occurring coagulase-negative staphylococci infections on new infections by mastitis pathogens in the bovine udder. J Dairy Sci. 1991;74:1855-9. Matthews KR, Kumar SJ, O´Connor SA, Harmon RJ, Pankey J, Fox LK, Oliver SP. Genomic fingerprints of Staphylococcus aureus of bovine origin by polymerase chain reaction-based DNA fingerprinting. Epidemiol Infect. 1994;112:177-86.

108
Mc Clure TJ, Hughes KL, Dowell AE, Murphy SM, Joyce E. Mastitis in dairy cattle in the Moss Vale District of New South Wales. Aust Vet J. 1966;42:194-8. Mead PS, Slutsker L, Dietz V, Mc Caig LF, Bresee JS, Shapiso C. Foodrelated illness and death in the United States. Emerg Inf Dis. 1994;5:607-25. Merl K, Abdulmawjood A, Lämmler C, Zschöck M. Determination of epidemiological relationships of Streptococcus agalactiae isolated from bovine mastitis. FEMS Microbiol Letters. 2002;26:87-92.
Mielke H. Physiologie der Laktation. In: Wendt K, Bostedt H, Mielke H, Fuchs HW. Euter- und Gesäugekrankheiten. Gustav Fischer Verlag Jena, Stuttgart; 1994. Milne MH, Barrett DC, Fitzpatrick JL, Biggs AM. Prevalence and aetiology of clinical mastitis on dairy farms in Devon. Vet Rec. 2002;24:241-3. Moeller FA, Daugaard HL, Jensen NE. Characterization of Staphylococcus simulans strains isolated from cases of bovine mastitis. Vet Microbiol. 1999;66:165-70. Müller C. Charakterisierung von Staphylokokken aus Mastitismilchproben der Region Nord-Ost-Schweiz. (Vet.-med. Diss.) Zürich: Universität von Zürich; 1992. Mullarky IK, Su C, Frieze N, Park YH, Sordillo LM. Staphylococcus aureus arg genotypes with enterotoxin production capabilities can resist neutrophil bactericidal activity. Infect Immun. 2001;69:45-51. Munson SH, Tremaine MT, Bentley MJ, Rodney AW. Identification and characterisation of staphylococcal enterotoxin types G and I from Staphylococcus aureus. Infect Immun. 1998;66:3337-48. Myllys V, Ridll J, Björkroth J, Biese I, Pyörälä S. Persistence of bovine mastitis of Staphylococcus aureus clones as assessed by random amplified polymorphic DNA analysis, ribotyping and biotyping. Vet Microbiol. 1997;51:245-51. Myllys V, Asplund K, Brofeldt E, Hrvelä-Koski V, Honkanen-Buzalske T, Junttila J, Kulkas L, Mymmkangas O, Niskanen M, Saloniemi H, Sandholm M, Saranpäa T. Bovine mastitis in Finland in 1988 and 1995 – Changes in prevalences and antimicrobial resistance. Acta Vet Scand. 1998;39:119-26.
Natzke RP, Everett RW, Bray DR. Effect of drying off practices on mastitis infection. J Dairy Sci. 1975;58:1828-32. Neave, FH. Diagnosis of mastitis by bacteriological methods alone. Proceedings of seminar mastitis control. International Dairy Federation, Brüssel. 1975:19-36. Neave FH, Dodd FH, Kingwell RG, Westgarth DR. Control of mastitis in the dairy herd by hygiene and managemant. J Dairy Sci. 1975;52:696-707. Neave FK, Dodd FH, Henriques E. J Dairy Res. 1950;17:37.

109
Neave FK. The control of mastitis by hygiene. In: Dodd FH, Jackson ER (eds). Control of bovine mastitis. British Cattle Veterinary Association. 1971:55. Newbould FHS. Epizootiology of mastitis due to Staphylococcus aureus. J Am Vet Med Ass. 1969;153:1683-7. Nielsen AL. Group B-streptococci in humans as a cause of bovine mastitis. Dansk Veterinärsskr. 1987;70:154-7.
O´Brien FG, Pearman JW, Gracey M, Riley TV, Grubb WB. Community strain of methicillin resistant Staphylococcus aureus involved in a hospital outbreak. J Clin Microbiol. 1999;37:2858-63. Opal SM, Cross A, Palmer M, Almazan R. Group B streptococcal sepsis in adults and infants. Arch Intern Med. 1988;148:641-5. Orwin PM, Leung DY, Donahue HL, Novick RP, Schievert PM. Biochemical and biological properties of staphylococcal enterotoxin K. Infect Immun. 2001;69:360-6. Orwin PM, Leung DY, Tripp TJ, Bohach GA, Earhardt CA, Ohlendorf DH, Schlievert PM. Characterization of a novel staphylococcal enterotoxin-like superantigen, a member of the group V subfamily of pyrogenic toxins. Biochem. 2002;41:14033-40. Otto D. Untersuchungen über die Bedeutung Koagulase-positiver und Koagulase-negativer Staphylokokken für die Mastitis des Schafes. (Vet. Med. Diss.) Gießen: Justus-Liebig-Universität Gießen; 1991. Philpot WN. Influence of subclinical mastitis on milk production and milk composition. J Dairy Sci. 1967;41:1141. Philpot WN. Prevention of infection-hygiene. Proceedings of seminar of mastitis control 1975, International Dairy Federation, Brüssel; 1975:155-64. Philpot WN, Nickerson SC. Mastitis: Counter attack. Babson Bros. Co. Naperville, USA; 1991. Phuektes P, Mansell PD, Dyson RS, Hooper ND, Dick JS, Browning GF. Molecular epidemiology of Streptococcus uberis isolates from dairy cows with mastitis. J Clin Microbiol. 2001;39:1460-6. Platonow I, Blobel H. Therapeutic failures in chronic staphylococcal mastitis. J Am Vet Med Ass. 1963;142:1097-1101. Plym-Forshell K, Osteras O, Aagaard K, Kulkas L. Disease recording and cellcount data in 1993 in Sweden, Norway, Denmark and Finland. Proceedings of the third International Mastitis Seminar, Tel Aviv Israel. 1993;4:50-4. Prevost G, Jaulhac B, Piemont Y. DNA fingerprinting by pulsed-field gel electrophoresis is more effective than ribotyping in distinguishing among methicillin-resistant Staphylococcus aureus isolates. J Clin Microbiol. 1992;30:967-73.

110
Puig de Centorbi ON, de Cuadrado AMA, Alcaraz LE, Laciar AL, de Milan MC. Prevalence of Staphylococcus aureus isolated from subclinical bovine mastitis in dairies of the city of San Luis. Rev Argent Microbiol. 1992;24:73-80. Spanish. Pyörälä SHK, Pyörälä EO. Efficacy of bovine clinical mastitis therapy during lactation. Proceeding of XVII Nordic Veterinary Congress, 26-29 July 1994. Reykjavik, Iceland. Rainard P. Should mammary infections caused by Corynebacterium bovis and coagulase-negative staphylococci be eliminated? Ann Rech Vet. 1987;18:355-64. Ren K, Bannan JD, Panchli V, Cheung AL, Robbins JC, Fischetti VA, Zabriskie JB. Characterisation and biological properties of a new staphylococcal exotoxin. Exp Med. 1994;180:1675-83. Riffon RK, Sayasith H, Khalil P, Dubreuil M, Drolet S, Lagace J. Development of a rapid and sensitive test for identification of major pathogens in bovine mastitis by PCR. J Clin Microbiol. 2001;39:2584-9. Rizk G, Janetsckke P. Über die Rolle von Corynebacterium bovis im Mastitisgeschehen und Untersuchungen zu seiner biochemischen Aktivität. Mh Vet Med. 1972;27:470.
Roberson JR, Fox LK, Hancock DD, Gay JM, Besser TE. Ecology of Staphylococcus aureus isolated from varios sites on dairy farms. J Dairy Sci. 1994;77:3354-64. Robinson TC, Harwood DG. Mastitis in a dairy herd associated with Corynebacterium bovis. Vet Rec. 1998;143:147. Röder R. Beziehung zwischen Zellgehalt und bakteriologischem Befund von Viertelgemelksproben beim Deutschen Fleckvieh. (Vet.-med. Diss.) München: Ludwig-Maximilians-Universität München; 1985. Römling U, Heuer T, Tümmler B. Bacterial genome analysis by pulsed-field gel electrophoresis techniques. In: Chrambach A, Dunn MJ (eds). Advances in electrophoresis. Vol. VHC, Weinheim. 1994:353-40. Römling U, Fislage R, Tümmler B. Theorie und Anwendung der Makrorestriktionsanalyse für die klonale Analyse von Erregern. Immun Infekt. 1995;23:4-8. Rolle M, Mayr A. Bakterielle Krankheiten der Tiere. In: Medizinische Mikrobiologie, Infektions- und Seuchenlehre. 7. Auflage, Enke Verlag, Stuttgart; 2002. Rosenberger G. Die klinische Untersuchung des Rindes. Paul Parey Verlag; 1990. Ross PW. Group B streptococcus- profile of organism. J Med Microbiol. 1984;18:139-66. Rossitto PV, Ruiz L, Kikuchi Y, Glenn K, Liuz K, Watts JL. Antibiotic susceptibility patterns for environmental streptococci isolated from bovine mastitis in central California dairies. J Dairy Sci. 2002;85:132-8. Sagar. Programa de producción de leche y de sustitución de las importaciones. 19 Seiten und Anhänge; 1996.

111
Sagar. Boletín de leche, Februar 2004, Centro de Estadística Agropecuaria, 43 Seiten. Spanish. http://www.siap.sagarpa.gob.mx/modelos/boletlech/Leche-Feb04.zip. Saulnier P, Bourneix C, Prevot G. Andremont B. Random amplified polymorphic DNA assay is less discriminant than pulsed-field gel electrophoresis or typing strains of methicillin- resistant Staphylococcus aureus. J Clin Microbiol. 1993;31:982-5. Schalm OW, Carroll EJ, Jain NC. Bovine mastitis. Lea and Febiger, Philadelphia; 1971. Schalm OW, Noorlander DO. Experiments and observations leading to the development of the California Mastitis Test. J Am Vet Med Assoc. 1957;130:199-204. Schlegelová J, Dendis M, Benedík J, Babak V, Rysanek D. Staphylococcus aureus isolates from dairy cows and humans differ in coagulase genotype. Vet Microbiol. 2003;92:327-34. Schukken YH, Grommers FJ, van de Geer D. Risk factors for clinical mastitis in herds with a low bulk milk somatic cell count. 1. The data and the risk factors for all cases. J Dairy Sci. 1990;73:3463-71. Schulz J. Erkrankungen der Milchdrüse des Rindes: Grundsätze. In: Wendt K, Bostedt H, Mielke H, Fuchs HW (eds). Euter- und Gesäugekrankheiten. Gustav Fischer Verlag Jena, Stuttgart; 1994. Seelemann M. Die Streptokokkeninfektionen des Euters. Verlag Schaper, Hannover; 1932.
Seelemann M. Biologie der Streptokokken, 2.Auflage, Verlag Hans Carl, Nürnberg; 1954.
Seffner W, Bergmann A. Staphylokokkeninfektionen. In: Wendt K, Bostedt H, Mielke H, Fuchs HW (eds). Euter- und Gesäugekrankheiten. Gustav Fischer Verlag Jena, Stuttgart; 1994. Selbitz HJ. Lehrbuch der veterinärmedizinischen Bakteriologie. Gustav Fischer Verlag Jena, Stuttgart; 1992. Smith KL, Todhunter DA, Schoenberger PS. Environmental mastitis: cause, prevalence, prevention. J Dairy Sci.1985;68:1531-53. Smits E, Burvenich C, Guidry AJ, Roets E. In vitro expression of adhesion receptors and diapedesis by polymorphonuclear neutrophils during experimentally induced Streptococcus uberis mastitis. Infect Immun. 1998;6:2529-34. Smola J. Pig and nutria as hosts of Streptococcus agalactiae. Abstr. 12th Lancefield Internat. Symp. Streptococci and Streptococcal Diseases, Saint Petersburg (Russia). 1998:121. Sobiraj A, Ostertag HU, Peip D, Bostedt H, Kielwein G. Klinische und bakteriologische Untersuchungsbefunde zur Mastitishäufigkeit erstmalig laktierender Rinder intra und post partum. Tierärztl Prax. 1988;16:243-9.

112
Sobiraj A, Kron A, Schollmeyer U, Failing K. Bundesweite Untersuchungen zur Erregerverteilung und in vitro-Resistenz euterpathogener Bakterien in der Milch von Kühen mit subklinischer Mastitis. Tierärztl Prax. 1997;25(2):108-15. Sommerhäuser J. Untersuchungen mittels Geno- und Phänotypisierung zur Epidemiologie von Staphylococcus aureus als Erreger subklinischer Mastitiden in hessischen Milcherzeugerbetrieben im Zuge von Bestandssanierungsmaßnahmen. [Dissertation vet. med.]. Leipzig: Univ. Leipzig; 2002. Sommerhäuser J, Kloppert B, Wolter W, Zschöck M, Sobiraj A Failing K. The epidemiology of Staphylococcus aureus infections from subclinical mastitis in dairy cows during a control programme. Vet Microbiol. 2003;96:91-102. Stabenfeldt GH, Spencer GR. The lesions in bovine udders shedding hemolytic coagulase-positive staphylococci. Pat Vet. 1965;2:585-600. Stadtfeld H, Nitzschke E. Comparative studies of cultural demonstration of mastitis streptococci in udder health service. Berl Munch Tierärztl Wschr. 1980;93:441-4. Sutra L, Poutrel B. Virulence factors involved in the pathogenesis of bovine intramammary infections due to Stayphylococcus aureus. J Med Microbiol. 194;40:79-89. Swedish Dairy Association. Djurhälsovard 1999/2000 (Animal health 1999/2000). Svenkmjölk, Eskilstuna, Sweden; 2000. Tatini SR, Jezeski JJ, Olson JC, Casman EP. Factors influencing the production of staphylococcal enterotoxin A in milk. J Dairy Sci. 1971;54:312-20. Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, Swanminathan A. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33:2233-9. Thiele D, Willems H, Köpf G, Krauss H. Polymorphism in DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. Eur J Epidemiol. 1993;29:419-25. Thompson WH, Houston BA. A survey of the incidence of bovine mastitis in dairy herds in two districts in Victoria. Austr Vet J. 1967;43:558-63. Timms LL, Schultz LH. Dynamics and significance of coagulase-negative staphylococcal intramammary infections. J Dairy Sci. 1987;70:2648-57. Tolle A, Heeschen W, Hamann J. Grundlagen einer systematischen Bekämpfung der subklinischen Mastitis des Rindes. Kieler Milchw Forschungsber. 1987;29:3-103. Tolle A. Die subklinische Kokkenmastitis des Rindes. Zentralbl Vet Med B. 1982;29:329-58. Tollersrud T, Kenny K, Caugant DA, Lund A. Characterisation of isolates of Staphylococcus aureus from acute, chronic and subclinical mastitis in cows in Norway. APMIS. 2000;108:565-72.

113
Vecht U, Wisselink HJ, Defize PR. Dutch national mastitis survey. The effect of herd and animal factors on somatic cell count. Neth Milk Dairy J. 1989;43:425-35.
Velazco J. Es México un pais con litros de oportunidad. Guia lactea. 2002;7:17-9. Wang SM, Deighton MA, Caapstick JA, Gerraty N. Epidemiological typing of bovine streptococci by pulsed-field gel electrophoresis. Epidem Infect. 1999;123:317-24. Watts JL, Rossbach S. Susceptibilities of Corynebacterium bovis and Corynebacterium amylocolatum isolates from bovine mammary glands to 15 antimicrobial agents. Antimicrob Agents Chemother. 2000;44:3476-7. Weigt U, Dreist B. Gastrointestinale Störungen des Rindes als Wegbereiter der hämatogen bedingten Kolimastitis. In: Deutsche Veterinärmedizinische Gesellschaft: Tagung der Fachgruppe Bakteriologie, Rauischholzhausen, 8.-10.6.1988, DVG Gießen. 1988:177-85. Weiss E. Weibliche Geschlechtsorgane. In: Dahme E, Weiss E (eds). Grundriss der pathologischen Anatomie der Haustiere. Enke Verlag Stuttgart. 1999:313-17. Wellenberg GJ, van der Poel WHM, van Oirschot JT. Viral infections and bovine mastitis: a review. Vet Microbiol. 2002;2361:2-21. Wendt K. Handbuch Mastitis. Kamlage Verlag, Osnabrück; 1998. Wesson CA, Liou LE, Todd KM, Bohach GA, Rumble WR, Bayles KW. Staphylococcus aureus agr and sar global regulators influence internationalization and induction of apoptosis. Infection Immun.1998;66:5238-43. Wilson CD. The microbiology of bovine mastitis in Great Britain. Bull Off Int Epizoot. 1963;60:533-51. Wilson CD, Richards MS. A survey of mastitis in the British dairy herd. Vet Rec. 1980;106:431. Wilson DJ, Gonzalez RN, Das HH. Bovine mastitis pathogens in New York and Pennsylvania: Prevalence and effects on somatic cell count and milk production. J Dairy Sci. 1997;80:2592-8. Wilson DJ, Gonzalez RN, Case KL, Garrisn LL, Gröhn YT. Comparison of seven antibiotic treatments with no treatment for bacteriological efficiacy against bovine mastitis pathogens. J Dairy Sci. 1999;82:1664-70. Winkenwerder W. Vorkommen und Βedeutung der sogenannten nicht hämolysierenden Streptokokken im Genitaltrakt von Rindern. Zuchthygiene. 1966;1:170. Workineh S, Bayleyegn M, Mekonnen H, Potgieter LN. Prevalence and aetiology of mastitis in cows from two major Ethiopian diaries. Trop Anim Health Prod. 2002;34:19-25. Wolter W, Kloppert B, Zschöck M. Verbreitung und Bekämpfung von S. agalactiae in Hessen. In: Deutsche Veterinärmedizinische Gesellschaft: 40. Arbeitstagung des

114
Arbeitsgebietes Lebensmittelhygiene, Garmisch-Partenkirchen, 29.9.-1.10.1999, DVG Gießen; 1999:640-8. Wolter W, Castañeda HV, Jung HP, Zschöck M. Occurence and prevalence of bacterial pathogens in bovine mastitis in Jalisco, México. Milchwissenschaften. 2002;3. Yancey RJ. Vaccines and diagnostic methods for bovine mastitis: fact and fiction. Adv Vet Med. 1999;41:257-73. Yildirim AÖ, Lämmler C, Weiss R, Kopp P. Pheno- and genotypic properties of streptococci of serological group B of canine and feline origin. FEMS Microbiol Letters. 2002;212:187-92. Zadoks R, Van Leeuwen W, Berkema H. Application of pulsed-field gel electrophoresis and binary typing as tools in veterinary clinical microbiology and molecular analysis of bovine and human Staphylococcus aureus isolates. Infect Immun. 2000;38:1931-9. Zadoks RN, Allore HG, Barkema HW, Sampimo OC, Gröhn YT, Schukken YH. Analysis of an outbreak of Streptococcus uberis mastitis. J Dairy Sci. 2001;84:590-9. Zadoks RN, van Leeuwen WB, Kreft D, Fox LK, Berkema HW, Schukken YH, van Belkum A. Comparison of Staphylococcus aureus isolates from bovine and human skin, milking equipment, and bovine milk by phage typing, pulsed-field gel electrophoresis, and binary typing. J Clin Microbiol. 2002;40:3894-902. Zehren VL, Zehren VF. Examination of large quantities of cheese for staphylococcal enterotoxin. J Dairy Sci. 1968;51:635-44. Zhang S, Iandolo JJ, Stewart GC. The enterotoxin D plasmid of Staphylococcus aureus encodes a second enterotoxin determinant (sej). FEMS Microbiol Lett. 1998;168:227-33. Zhang S, Maddox CW. Cytotoxic activity of coagulase-negative staphylococci in bovine mastitis. Infection and Immunity, 2000;68:1102-8. Zschöck M, Kloppert B, Wolter W, Seufert H, Schwarz HP, Kötting C. Zellzahlen der Milch in großen Beständen. In: Rationalisierungs-Kuratorium für Landwirtschaft (RKL), Rendsburg-Osterrönfeld, 1998:487-536. Zschöck M. Aktuelle Aspekte zur Diagnostik von Mastitiserregern-Stand und Entwicklungstendenzen, Arbeitskreis „Eutergesundheit“ der Deutschen Veterinärmedizinischen Gesellschaft Thema: Trends in der Milchproduktion- Wandel in der Mastitisbekämpfung. Hannover, vom 27. bis 28. Mai. 1999:37-45. Zschöck M, Sommerhäuser J, Castañeda H. Relatedness of Staphylococcus aureus isolates from bovine mammary gland suffering from mastitis in a single herd. J Dairy Res. 2000;67:429-35. Zschöck M, Risse K, Sommerhäuser J. Occurrence and clonal relatedness of sec/tst-gene positive Staphylococcus aureus isolates of quartermilk samples of cows suffering from mastitis. Letters in Applied Microbiol. 2004;38:493-8.

115
Zschöck M, Kloppert B, Wolter W, Hamann HP, Lämmler C. Pattern of enterotoxin genes seg, seh, sei and sej positive Staphylococcus aureus isolated from bovine mastitis. Vet Microbiol. 2005;108:243-9.
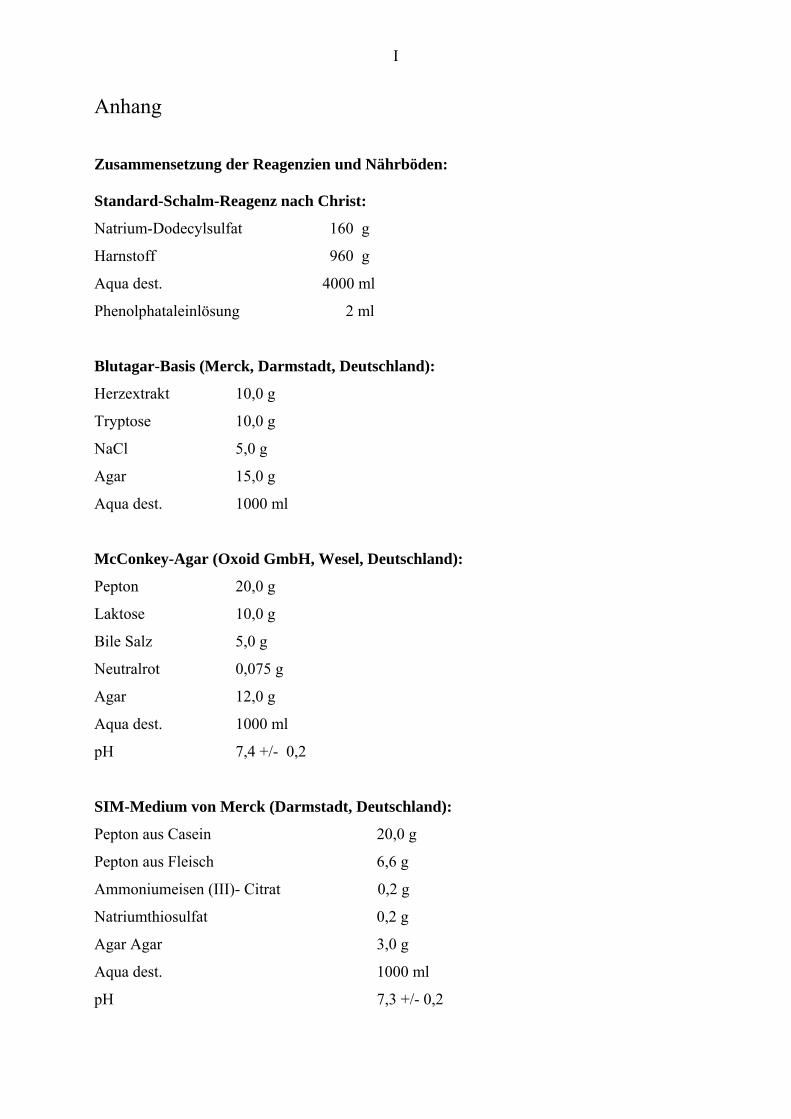
I
Anhang Zusammensetzung der Reagenzien und Nährböden: Standard-Schalm-Reagenz nach Christ:
Natrium-Dodecylsulfat 160 g
Harnstoff 960 g
Aqua dest. 4000 ml
Phenolphataleinlösung 2 ml
Blutagar-Basis (Merck, Darmstadt, Deutschland):
Herzextrakt 10,0 g
Tryptose 10,0 g
NaCl 5,0 g
Agar 15,0 g
Aqua dest. 1000 ml
McConkey-Agar (Oxoid GmbH, Wesel, Deutschland):
Pepton 20,0 g
Laktose 10,0 g
Bile Salz 5,0 g
Neutralrot 0,075 g
Agar 12,0 g
Aqua dest. 1000 ml
pH 7,4 +/- 0,2
SIM-Medium von Merck (Darmstadt, Deutschland):
Pepton aus Casein 20,0 g
Pepton aus Fleisch 6,6 g
Ammoniumeisen (III)- Citrat 0,2 g
Natriumthiosulfat 0,2 g
Agar Agar 3,0 g
Aqua dest. 1000 ml
pH 7,3 +/- 0,2

II
Sabouraud-2%-Glucose-Agar (Merck, Darmstadt, Deutschland):
Pepton 10,0 g
D (+)-Glucose 20,0 g
Agar 17,0 g
Aqua dest. 1000 ml
pH 5,6 +/- 0,2
Todd-Hewitt-Bouillon (THB; Oxoid GmbH, Wesel, Deutschland):
Hirn-, Herzextrakt und Peptone 27,5 g
Glucose 2,0 g
Natriumchlorid 5,0 g
Dinatriumhydrogenphosphat 2,5 g
Aqua dest. 1000 ml
pH 7,4 +/- 0,2
TE-Puffer:
Tris-HCl (10mmol/l) 1,21 g
EDTA (1mmol/l) 372,5mg
Aqua dest. 1000 ml
pH 7,6

III
Abb. 7 Beispiel für einen sauberen Auslauf bei reiner Auslaufhaltung (Note 2)
Abb. 8 Beispiel für einen ungenügenden sauberen Auslauf (Note 4)

IV
Abb. 9 Melkstand mit Rohrmelkanlage eines spezialisierten Betriebs
Abb. 10 Typischer Melkstand mit Fangfressgittern und Rohrmelkanlage der halbspezialisierten Betriebsformen

V
Abb. 11 Mobile Eimermelkanlage mit Motor in einem Familienbetrieb
Abb. 12 Typische familiäre Hinterhofhaltung
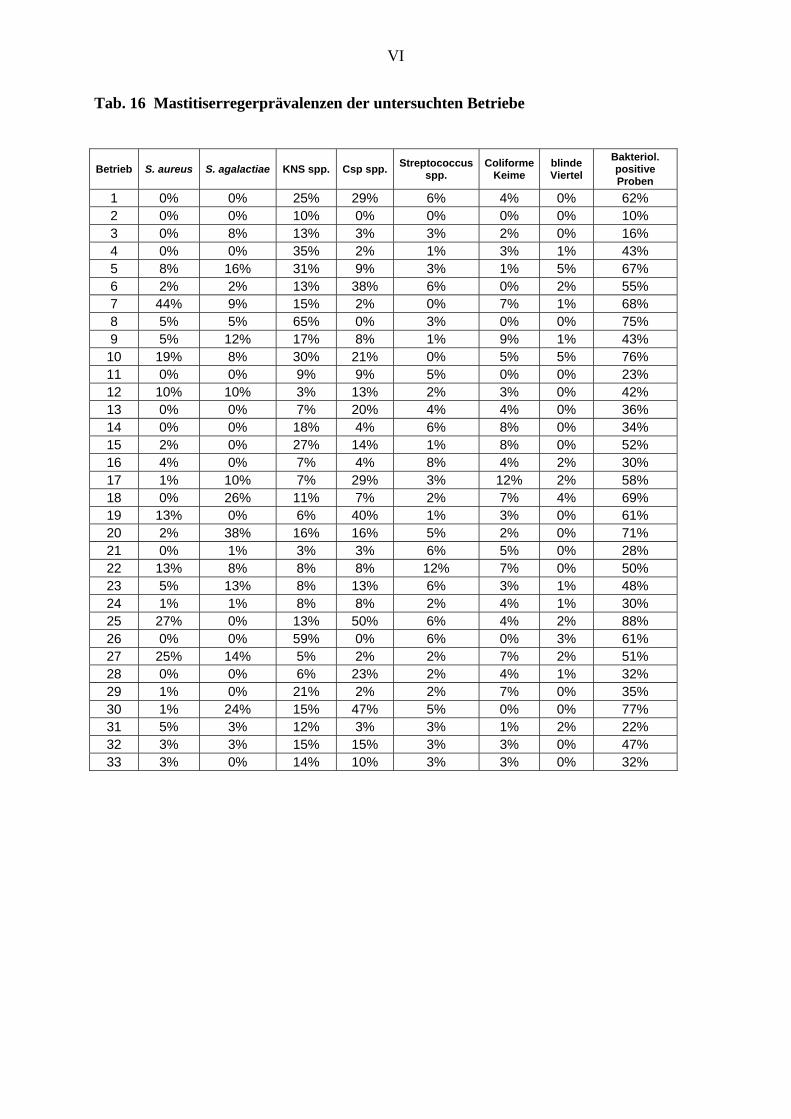
VI
Tab. 16 Mastitiserregerprävalenzen der untersuchten Betriebe
Betrieb S. aureus S. agalactiae KNS spp. Csp spp. Streptococcus spp.
ColiformeKeime
blinde Viertel
Bakteriol. positive Proben
1 0% 0% 25% 29% 6% 4% 0% 62% 2 0% 0% 10% 0% 0% 0% 0% 10% 3 0% 8% 13% 3% 3% 2% 0% 16% 4 0% 0% 35% 2% 1% 3% 1% 43% 5 8% 16% 31% 9% 3% 1% 5% 67% 6 2% 2% 13% 38% 6% 0% 2% 55% 7 44% 9% 15% 2% 0% 7% 1% 68% 8 5% 5% 65% 0% 3% 0% 0% 75% 9 5% 12% 17% 8% 1% 9% 1% 43%
10 19% 8% 30% 21% 0% 5% 5% 76% 11 0% 0% 9% 9% 5% 0% 0% 23% 12 10% 10% 3% 13% 2% 3% 0% 42% 13 0% 0% 7% 20% 4% 4% 0% 36% 14 0% 0% 18% 4% 6% 8% 0% 34% 15 2% 0% 27% 14% 1% 8% 0% 52% 16 4% 0% 7% 4% 8% 4% 2% 30% 17 1% 10% 7% 29% 3% 12% 2% 58% 18 0% 26% 11% 7% 2% 7% 4% 69% 19 13% 0% 6% 40% 1% 3% 0% 61% 20 2% 38% 16% 16% 5% 2% 0% 71% 21 0% 1% 3% 3% 6% 5% 0% 28% 22 13% 8% 8% 8% 12% 7% 0% 50% 23 5% 13% 8% 13% 6% 3% 1% 48% 24 1% 1% 8% 8% 2% 4% 1% 30% 25 27% 0% 13% 50% 6% 4% 2% 88% 26 0% 0% 59% 0% 6% 0% 3% 61% 27 25% 14% 5% 2% 2% 7% 2% 51% 28 0% 0% 6% 23% 2% 4% 1% 32% 29 1% 0% 21% 2% 2% 7% 0% 35% 30 1% 24% 15% 47% 5% 0% 0% 77% 31 5% 3% 12% 3% 3% 1% 2% 22% 32 3% 3% 15% 15% 3% 3% 0% 47% 33 3% 0% 14% 10% 3% 3% 0% 32%

VII
Tab. 17 Übersicht über einige auf den 33 Betrieben erfassten Daten
In der Tabelle verwendete Abkürzungen: Betriebsform: 1 spezialisiert 2 halbspezialisiert 4 Familienbetrieb Haltungsform: 1 Auslauf 2 Weide Hygiene des Auslaufs bzw. der Melkzeuge:
1 sehr gut 2 gut 3 befriedigend 4 ausreichend
Melksystem:
1 Handmelken 2 Eimermelkanlage mobil 3 Eimermelkanlage stationär 4 Rohrmelkanlage
Vormelken:
0 Kein Vormelken 1 auf den Boden 2 mit Melkbecher
Euterreinigung:
0 keine Reinigung 1 trockene Reinigung mit Papier 2 feuchte Reinigung mit nachfolgendem Abtrocknen mit Papier 3 feuchte Reinigung ohne Abtrocknen
Melkreihenfolge:
0 keine 1 feste Melkreihenfolge 2 Kühe werden in verschiedenen Gruppen gemolken
Zwischendesinfektion:
0 nein 1 ja
Dippen: 0 kein Dippen 1 Dippen nach dem Melken mit Jodlösung 2 Dippen vor und nach dem Melken mit einer Reinigungs- bzw. mit einer Jodlösung

VIII
Trockenstellen: 0 langsam mit übersprungenen Melkzeiten und Futterreduzierung 1 wie oben jedoch mit Verwendung eines antibiotischen Trockenstellers 2 von einer Melkzeit auf die andere ohne Trockensteller 3 von einer Melkzeit auf die andere mit Trockensteller 4 nur nach vorheriger Euterkontrolle mittels CMT

IX
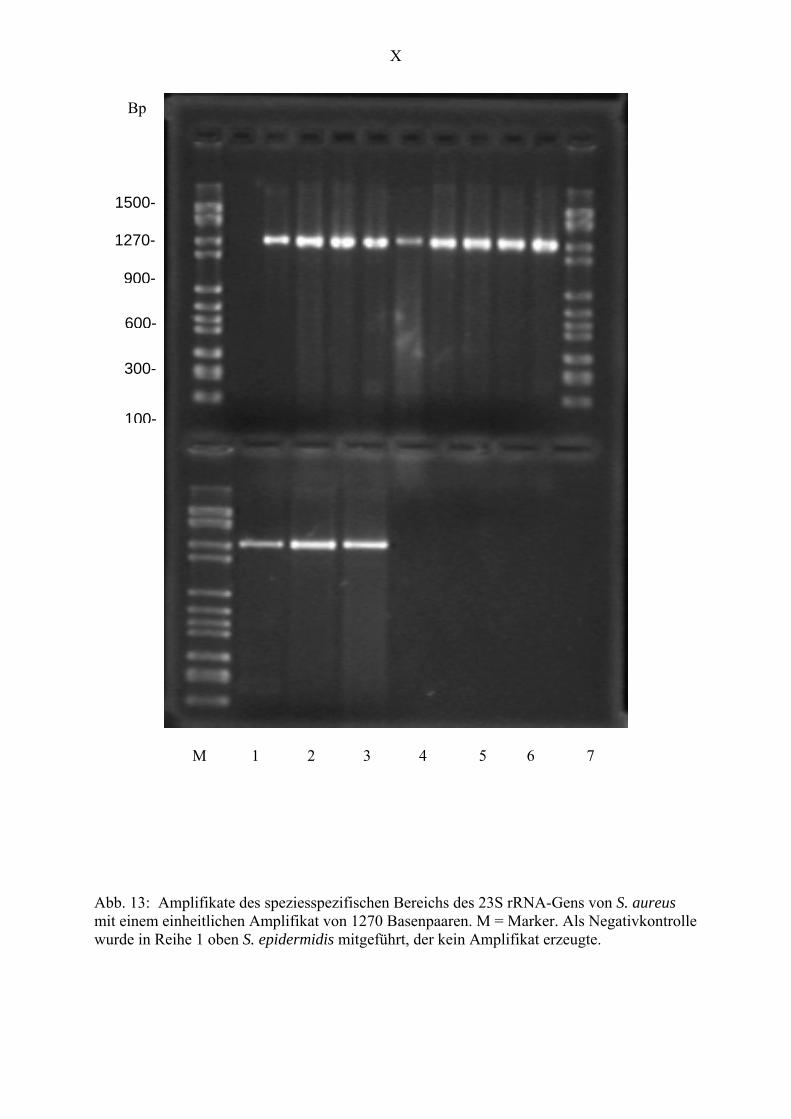
X
M 1 2 3 4 5 6 7 Abb. 13: Amplifikate des speziesspezifischen Bereichs des 23S rRNA-Gens von S. aureus mit einem einheitlichen Amplifikat von 1270 Basenpaaren. M = Marker. Als Negativkontrolle wurde in Reihe 1 oben S. epidermidis mitgeführt, der kein Amplifikat erzeugte.
1270-
900-
600-
100-
300-
Bp
1500-

XI
M 1 2 3 4 5 6 7 8 9 10 11
Abb. 14: Amplifikate des 16 S rRNA-Gens von 10 S. agalactiae-Isolaten mit einer
einheitlichen Größe von 1250 Bp. Als Negativ-Kontrolle wurde auf Postion 1 ein S. canis-
Stamm mitgeführt, der kein Amplifikat erzeugte.
M 1 2 3 4 5 6 7
Abb. 15: Amplifikate des speziesspezifischen Bereichs des 16S rRNA-Gens von S. uberis mit
der einheitlichen Größe von 445 Bp (Reihe 1, 4, 5, 7). Hiermit konnte nachgewiesen werden,
dass es sich bei den positiven Kulturen um Isolate von S. uberis handelt. Die übrigen Kulturen
(Reihe 2, 3, 6) ohne Amplifikat gehörten anderen Streptokokkenspezies wie z.B.
Enterococcus faecalis oder S. bovis an, weswegen unter Verwendung der Primer ub І und ub
ІІ keine Amplifizierung der entsprechenden Region möglich war.
1250-
1500-
Bp
900-
600-
300-
100-
900-
1500-
600- 445-
300-
100-
Bp
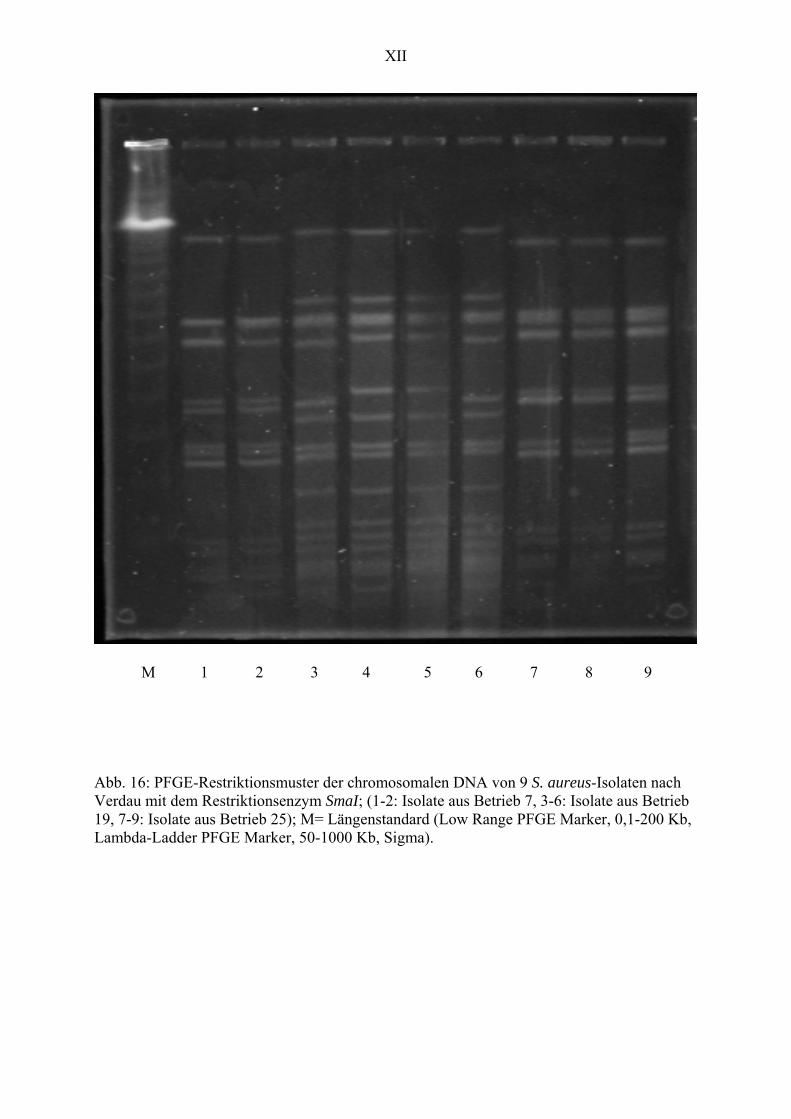
XII
M 1 2 3 4 5 6 7 8 9 Abb. 16: PFGE-Restriktionsmuster der chromosomalen DNA von 9 S. aureus-Isolaten nach Verdau mit dem Restriktionsenzym SmaI; (1-2: Isolate aus Betrieb 7, 3-6: Isolate aus Betrieb 19, 7-9: Isolate aus Betrieb 25); M= Längenstandard (Low Range PFGE Marker, 0,1-200 Kb, Lambda-Ladder PFGE Marker, 50-1000 Kb, Sigma).
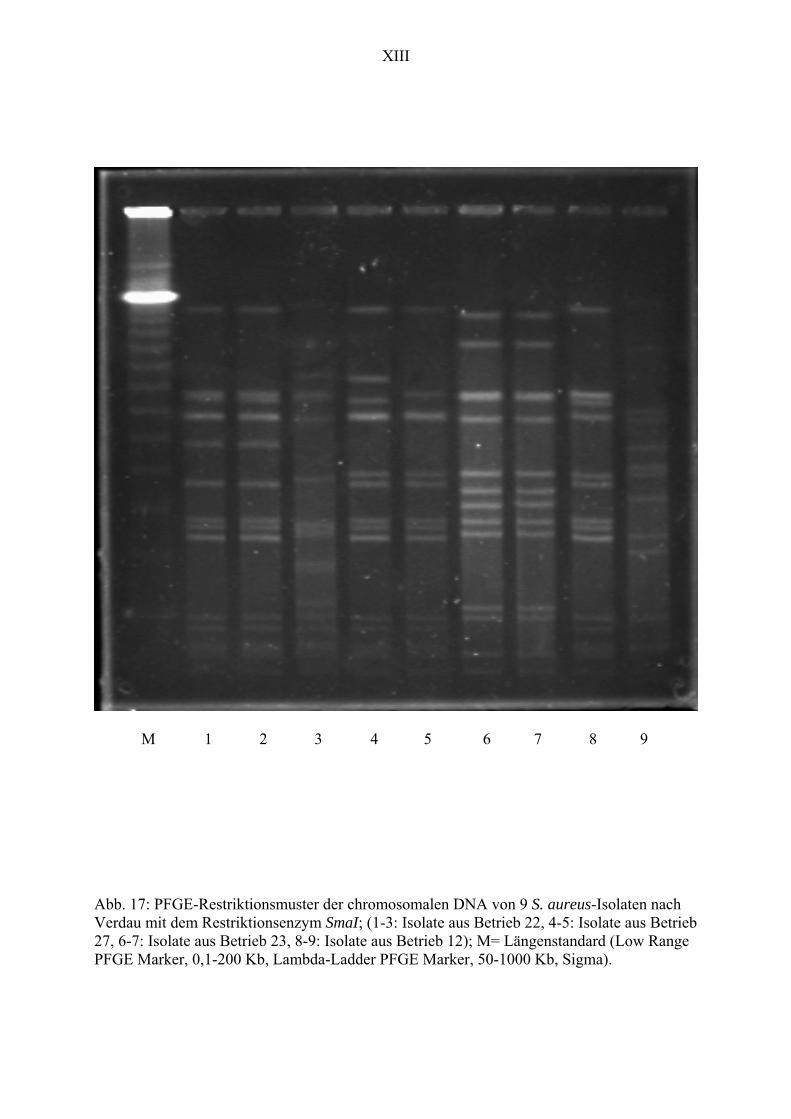
XIII
M 1 2 3 4 5 6 7 8 9 Abb. 17: PFGE-Restriktionsmuster der chromosomalen DNA von 9 S. aureus-Isolaten nach Verdau mit dem Restriktionsenzym SmaI; (1-3: Isolate aus Betrieb 22, 4-5: Isolate aus Betrieb 27, 6-7: Isolate aus Betrieb 23, 8-9: Isolate aus Betrieb 12); M= Längenstandard (Low Range PFGE Marker, 0,1-200 Kb, Lambda-Ladder PFGE Marker, 50-1000 Kb, Sigma).
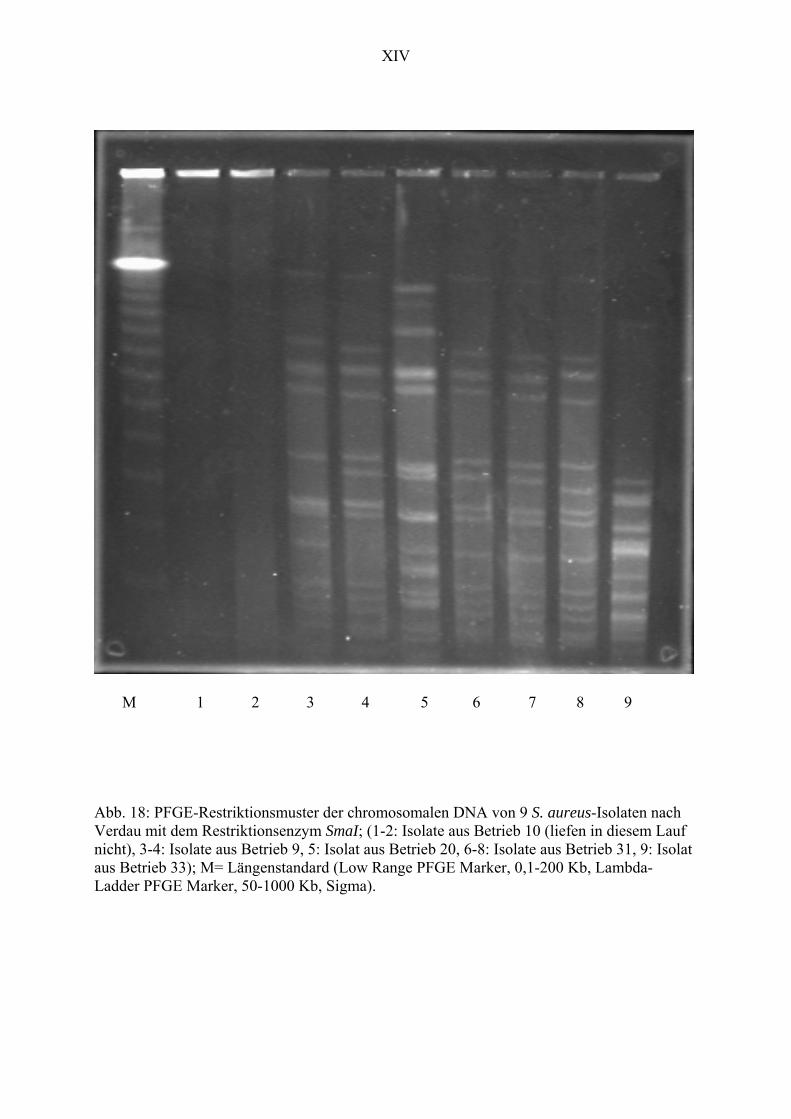
XIV
M 1 2 3 4 5 6 7 8 9 Abb. 18: PFGE-Restriktionsmuster der chromosomalen DNA von 9 S. aureus-Isolaten nach Verdau mit dem Restriktionsenzym SmaI; (1-2: Isolate aus Betrieb 10 (liefen in diesem Lauf nicht), 3-4: Isolate aus Betrieb 9, 5: Isolat aus Betrieb 20, 6-8: Isolate aus Betrieb 31, 9: Isolat aus Betrieb 33); M= Längenstandard (Low Range PFGE Marker, 0,1-200 Kb, Lambda-Ladder PFGE Marker, 50-1000 Kb, Sigma).
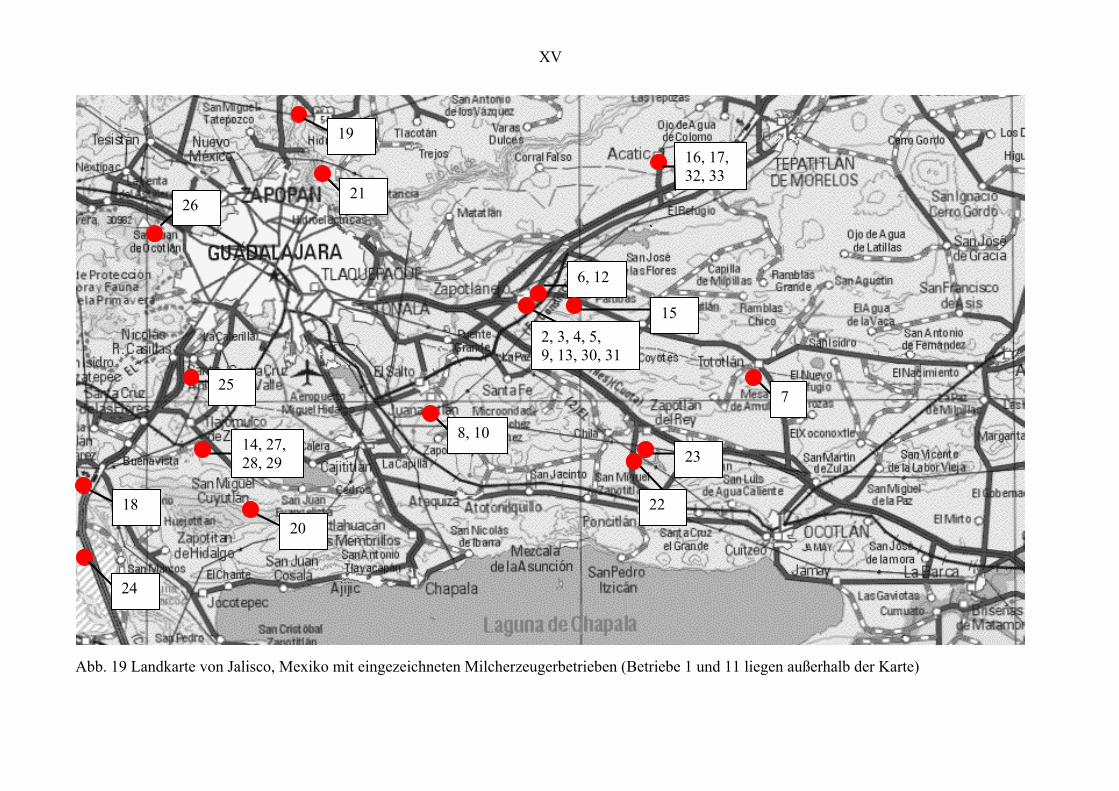
XV
Abb. 19 Landkarte von Jalisco, Mexiko mit eingezeichneten Milcherzeugerbetrieben (Betriebe 1 und 11 liegen außerhalb der Karte)
16, 17, 32, 33
23
22 20
8, 10
18
14, 27, 28, 29
25
26 21
19
2, 3, 4, 5, 9, 13, 30, 31
7
6, 12
15
24

Zum Gelingen dieser Arbeit möchte ich mich bei folgenden Menschen bedanken: Dr. Wilfried Wolter und Dr. Bärbel Kloppert für die freundliche Überlassung des Themas, die Korrekturen und aufmunternden Worte. Prof. Dr. Hugo Castañeda und meinen mexikanischen Studenten für die nette Zusammenarbeit und tatkräftige Unterstützung bei der Probenentnahme in Mexiko. Hans-Peter Jung, Silke Zimmermann, Tanja Puchtinger und Jens Wiessner für die Einarbeitung in die verschiedenen Untersuchungsmethoden am Hessischen Landeslabor Gießen. Dr. Michael Zschöck und Prof. Dr. Axel Sobiraj für die Betreuung der Arbeit. Dr. Klaus Failing für die statistische Auswertung meiner Daten. Meinen Eltern und Freunden, ohne deren Unterstützung eine Fertigstellung dieser Arbeit nicht möglich gewesen wäre. Mein letzter Dank gilt dem DAAD für die finanzielle Unterstützung meines Aufenthaltes in Mexiko.





























