
39
J. Japan. Soc. Hort. Sci. 82 (1): 39–50. 2013.
Available online at www.jstage.jst.go.jp/browse/jjshs1
JSHS © 2013
A Torenia (Torenia fournieri Lind. ex Fourn.) Novel Mutant ‘Flecked’ Produces
Variegated Flowers by Insertion of a DNA Transposon into an R2R3-MYB Gene
Takaaki Nishijima1,2*, Yasumasa Morita1**, Katsutomo Sasaki1, Masayoshi Nakayama1,
Hiroyasu Yamaguchi1, Norihiro Ohtsubo1, Tomoya Niki1,2 and Tomoko Niki1
1NARO Institute of Floricultural Science, Tsukuba 305-8519, Japan2Graduate School of Life and Environmental Sciences, University of Tsukuba, Tsukuba 305-8577, Japan
A novel torenia (Torenia fournieri Lind. ex Fourn.) mutant ‘flecked’, which bears variegated flowers, was obtained
from ethyl methanesulfonate-treated M2 plants. The lower lip of this mutant has small violet spots with a pale
violet background, while that of the normal type is solid violet. The mutant trait frequently reverted to a
semicircular violet sector or solid violet lower lip. Germinal revertant plants with a solid violet lower lip also
frequently occurred in S1 plants derived from self-pollinated mutant flowers. In the lower lip of the mutant type,
anthocyanin concentration was much lower than in the normal type. This was attributed to decreased expression
of the genes encoding anthocyanin biosynthesis enzymes, i.e. torenia chalcone synthase (TfCHS), flavanone 3-
hydroxylase (TfF3H), dihydroflavonol 4-reductase (TfDFR), anthocyanidin synthase (TfANS), and UDP-glucose
3-O-flavonoid glucosyltransferase (TfUFGT). In the lower lip of the mutant, expression of a gene encoding R2R3-
MYB transcription factor (TfMYB1, Torenia fournieri MYB1) was much lower than in the normal type and the
revertants; this was caused by insertion of a Enhancer/Suppressor-Mutator (En/Spm)-like transposon (Ttf1,
Transposon Torenia fournieri 1) in the 2nd intron of TfMYB1. Furthermore, it was found that the reversion of
anthocyanin accumulation in the lower lips correlated to excision of Ttf1 from the TfMYB1. Overexpression of
TfMYB1 in torenia caused anthocyanins to accumulate in the purple callus as a result of enhanced expression of
the five structural genes mentioned above, demonstrating that TfMYB1 regulates these genes. Therefore, we
concluded that a homozygous allele of Ttf1-inserted TfMYB1 caused the mutant phenotype. Ttf1 is a non-
autonomous element because Ttf1 does not have the DNA sequence encoding transposase. Based on these results,
potential uses of the flecked mutant for torenia breeding and transposon tagging are discussed.
Key Words: anthocyanin, En/Spm family, mutable allele, transposable element.
Introduction
Torenia (Torenia fournieri Lind. ex Fourn.) is mainly
used as a bedding or potted plant in the summer in Japan.
Torenia plants branch vigorously and bear many flowers,
forming a compact and neat appearance (Morise, 2001;
National Agriculture and Food Research Organization,
2006). Torenia grows well under a wide range of light
intensities, including direct sunlight to weak light in the
shade, and is also highly tolerant of high temperatures
during mid-summer that have been increased by recent
climate changes on account of global warming. Breeding
for flower color variation has produced torenia cultivars
with white, pink, red, and yellow flowers, in addition to
the original violet flower of the wild T. fournieri.;
however, variation in flower morphology is relatively
poor, lacking large-flowered and double-flowered
cultivars. If this defect could be overcome, and a highly
attractive appearance could be combined with the
excellent growth properties mentioned above, the
commercial value of torenia would increase significantly.
Several breeding techniques have been employed to
extend the genetic variation of torenia. Interspecific
crosses among Torenia species have increased the variety
of flower colors (Miyazaki et al., 2007); however, fertile
combinations of interspecific crosses have been limited
and variation in flower morphology is relatively poor
among Torenia species. Heavy ion-beam radiation and
gene recombination has extended the variation of flower
Received; May 12, 2012. Accepted; August 8, 2012.* Corresponding author (E-mail: [email protected]).
** Present address: Faculty of Agriculture, Kagawa University,Kagawa 761-0795, Japan.

T. Nishijima, Y. Morita, K. Sasaki, M. Nakayama, H. Yamaguchi, N. Ohtsubo, T. Niki and T. Niki40
color and morphology of T. fournieri (Aida et al., 2000;
Sasaki et al., 2008); however, strains with sufficiently
large flowers and double flowers have not yet been
reported. Considering the current status of torenia
breeding, it is important to exploit new tools to extend
the genetic variation.
Spontaneous mutations caused by transposable
elements have been important mutagens in breeding
floricultural plants (Inagaki et al., 1994; Matsubara et al.,
2005; Nakajima et al., 2005; Nitasaka, 2003, 2007).
Transposable elements with high transposition activity
often cause unstable traits (e.g. Goodrich et al., 1992;
Inagaki et al., 1994). If transposition activity is moderate,
without disrupting the practical uniformity of cultivars,
transposable elements will function as a useful mutagen
for breeding (Matsubara et al., 2005; Nakajima et al.,
2005; Nitasaka, 2003, 2007).
Transposable elements have been used to identify the
gene causing a mutation, if the mutation is caused by
the insertion of a transposable element (e.g. Coen et al.,
1990). In this technique, i.e. transposon tagging, the
causal gene is identified using the sequence of the
transposable element as a tag. Torenia has the following
excellent traits as a model floricultural plant for
transposon tagging: moderate flower size, making
observation of the flower structure easy; compact plant
size, enabling a large number of plants to be grown in
a limited area; easy and efficient propagation, both by
seed and cutting; and a high Agrobacterium-mediated
transformation rate (Aida et al., 2000). Torenia is also
an excellent model plant for the fertilization processes
as the ovary structure enables the live fertilization
process to be observed under a microscope (Okuda et al.,
2009). If an active transposable element is found in
torenia, an excellent transposon tagging system could
be established for both floricultural science and plant
molecular biology.
Transposable elements are classified into class I and
class II elements (Flavel et al., 1994). The DNA sequence
of a class I element (also called a retrotransposon) is
transcribed into RNA and then the RNA is reverse-
transcribed into cDNA. The resultant cDNA is integrated
into the genome by integrase. Thus, the class I element
propagates with the parent element remaining in the
original position of the genome. In contrast, a class II
element (also called a DNA transposon; simply called
‘transposon’ in the following text) is directly excised
and then integrated into another site in the genome by
transposase. Transposons in higher plants are classified
into three groups; the Ac/Ds (or hAT) superfamily, the
En/Spm (or CACTA) superfamily and the Mu family,
depending on their structure and length of target site
duplication (Flavel et al., 1994).
Variegated flowers are caused by excisions of active
transposons from the genes involved in anthocyanin
biosynthesis. In such unstable mutants, transposition
activity of the transposon can be evaluated visually
because the somatic excision frequency of the transposon
is reflected in the density of the colored spots or sectors
(Bonas et al., 1984; Goodrich et al., 1992; Inagaki et al.,
1994; Luo et al., 1991; Martin et al., 1985; Sommer et
al., 1985). The molecular basis for such a variegation
trait is a transposon-induced insertion mutation in the
genes encoding anthocyanin biosynthesis enzyme
(Bonas et al., 1984; Inagaki et al., 1994; Martin et al.,
1985; Sommer et al., 1985) and transcription factors
regulating the genes for anthocyanin biosynthesis have
been reported (Goodrich et al., 1992). An early process
of anthocyanin biosynthesis is the phenylpropanoid
pathway, which synthesizes 4-coumaloyl-CoA. 4-
Coumaloyl-CoA and malonyl-CoA are converted into
tetrahydroxy chalcone by chalcone synthase (CHS)
(Forkmann, 1994; Heller and Forkmann, 1994).
Tetrahydroxy chalcone is converted into flavanone by
chalcone isomerase (CHI), and then converted into
dihydroflavonol by flavanone-3-hydroxylase (F3H).
Dihydroflavonol is converted into leucoanthocyanidin
by dihydroflavonol 4-reductase (DFR), and then into
anthocyanidin by anthocyanidin synthase (ANS). The
anthocyanidin is then glycosylated to generate anthocy-
anin by UDP-glucose 3-O-glucosyltransferase (UFGT).
The expression of the genes encoding those anthocyanin
biosynthesis enzymes is directly regulated by the
following transcription factors, i.e. R2R3-MYB protein,
basic helix-loop-helix protein (bHLH) and WD40 repeats
protein (WDR) (Broun, 2005; Koes et al., 2005).
We isolated a novel torenia mutant with variegated
flowers from ethyl methanesulfonate-treated M2 plants.
The lower lip of the flowers of this mutant had small
violet spots with a pale violet background, while that of
the normal-type flower was solid violet. The mutant
frequently produced flowers with a semicircular violet
sector or solid violet lower lip. These phenotypes
suggested the involvement of a transposon. In this paper,
we identify the transposon responsible for this mutation.
Potential uses of the mutant in torenia breeding and
transposon tagging are also discussed.
Materials and Methods
Plant material and isolation of mutant
An open pollination cultivar ‘Common Violet’ (Takii
Seed Co., Kyoto, Japan) of torenia (Torenia fournieri
Lind. ex Fourn.) was used. The mutant was isolated as
follows. Approximately 1000 seeds were immersed in
100 mM aqueous ethyl methanesulfonate with gentle
shaking for 16 h at room temperature. The seeds were
then washed repeatedly with distilled water for 4 h and
then planted in plastic containers (44 cm length, 32 cm
width, and 7 cm height) filled with Metromix 350 (Sun
Gro Horticulture, BC, Canada). Seedlings were planted
in 72-cell plug trays (54 cm length, 28 cm width; 4 cm
length, 4 cm width, and 6 cm height for a cell) filled
with Kureha-Engei-Baido (Kureha Chemical Industry
Co. Ltd., Tokyo, Japan) and Metromix 350 (1 : 1 (v/v)).

J. Japan. Soc. Hort. Sci. 82 (1): 39–50. 2013. 41
The plants were grown in a glasshouse with natural
sunlight. The temperature was kept between 18–32°C.
Lateral stems were removed and a slow-releasing coated
fertilizer (Ecolong 100; Chisso Co., Tokyo, Japan) was
used for top-dressing. Two flowers per plant were self-
pollinated by hand. Approximately 4000 M2 plants were
grown as described above. A M2 plant bearing flowers
with variegated lower lips was selected and self-
pollinated. This mutant trait was unstable and somatic
and germinal reversions frequently occurred. In this
paper we refer to normal-type and the variegated mutant-
type flowers as NT and MT flowers, respectively.
Flowers with apparently complete somatic reversion and
flowers of apparently germinal revertant plants, both of
which have solid violet lower lips, are termed NT-SR
and NT-GR flowers, respectively. To maintain the mutant
strain, the MT flowers were selectively self-pollinated
and M5 plants were used for experiments. The plants
used for the cross-pollination experiment, analysis of
anthocyanins and gene expression were planted in pots
(9 cm in diameter and 12 cm in depth) filled with the
same medium as that used in the plug tray culture to
obtain highly stable and uniform growth.
Cross pollination experiment
The NT, MT, and NT-SR flowers were cross-pollinated
by hand in the combinations shown in Table 1. Six
flowers per plant of the seed and pollen parents were
used for crossing.
Analysis of anthocyanins and related compounds
For anthocyanin analysis, we collected 20 mg FW of
the lateral limbs of the lower lip of the opened flower;
the lower limb was excluded because it had a yellow
blotch. The samples were frozen in liquid nitrogen and
stored at −80°C until analysis. Anthocyanins were
extracted with 5% aqueous acetic acid. We analyzed
10 µL of the extract using an HP1100 high-performance
liquid chromatography system with a photodiode array
detector (Agilent Technologies/Yokogawa Analytical
Systems, Tokyo, Japan) and an Inertsil ODS-2 column
(4.6 mm × 250 mm; GL Science, Tokyo, Japan) at 40°C
with a flow rate of 0.8 mL·min−1. Absorption spectra
were monitored at 240–580 nm. A linear gradient of 10%
to 50% of solvent B (1.5% H3PO4, 40% acetonitrile,
50% acetic acid) in solvent A (1.5% H3PO4) was run
over 40 min.
Molecular cloning of the genes encoding transcription
factors regulating anthocyanin biosynthesis
The lateral limbs of the lower lips were collected at
the late corolla development stage (stage 7) when petal
pigmentation started (Nishijima and Shima, 2006). The
samples were frozen in liquid nitrogen and stored at
−80°C until use. Total RNA was obtained using an
RNeasy Plant Mini Kit (QIAGEN K. K., Tokyo, Japan).
cDNA was synthesized using a 1st Strand cDNA
Synthesis Kit for RT-PCR (AMV; Roche Applied
Science, IN, USA). Partial sequences of cDNAs
encoding R2R3-MYB protein (MYB) and basic helix-
loop-helix protein (bHLH) were obtained by nested PCR
using the following degenerate primers based on the
conserved amino acid sequences indicated in parenthe-
ses; forward 5'-AARWSITGYMGIYTIMGITGGYT-3'
(KSCRLRWL) and reverse 5'-YTTISWRCAICKIVWIG
CYTTIGT-3' (K(S/T)CR(I/S)AKT) for 1st PCR and
forward 5'-AAYMRITGGWSIYTIATHGCIGG-3' (N(K/
R)WSLIAG) and reverse 5'-YTTISWRCAICKIVWIGC
YTTIGT-3' (K(S/T)CR(I/S)AKT) for 2nd PCR for MYB;
forward 5'-TGGDSIGAYGGITWYTAYAAYGG-3' (W(G/
S)DG(F/Y)YNG) and reverse 5'-GTYTGIAYISIIGCIS
WYTTIGC-3' (TQ(I/V)(S/G/R)ASKA) for 1st PCR and
forward 5'-GGIGMIATHAARACIMGIAARAC-3' (G(A/
D)IKTRKT) and reverse 5'-CKISWRAAIACYTTISWI
TCIRC-3' (R(S/T)FVK(T/S)(D/E)(A/V)) for 2nd PCR
for bHLH. 5'- and 3'-RACE-PCR (Frohman et al., 1988)
was performed using the CapFishing Full-length cDNA
Premix Kit (Seegene, Seoul, South Korea) based on the
partial cDNA sequences of the genes. Full-length cDNAs
were obtained using the following primer sets: for-
ward 5'-ACGCACGTATAATAATTATTTCGCG-3' and
reverse 5'-GTGGTGATTATGATCATAATTAATTACCAC-
3' for torenia MYB1 (TfMYB1); forward 5'-ACGCACGT
ATAATAATTATTTCGCG-3' and reverse 5'-GTGGT
GATTATGATCATAATTAATTACCAC-3' for torenia
bHLH1 (TfbHLH1).
Expression analysis of the genes involved in anthocyanin
biosynthesis
To investigate the expression of the genes involved
in anthocyanin biosynthesis, cDNAs were obtained from
the lateral limbs of the lower lip in stage 7 as described
above. For total RNA extraction for cDNA synthesis,
we treated the samples with DNase to avoid potential
contamination with genomic DNA. For organ-specific
analysis of the genes, the following organs were
collected: sepal, limb, tube, stamen, and pistil of flowers
at stage 7, stems, young and adult leaves (about 1 cm
and 3–4 cm in length, respectively).
Primers for quantitative real-time PCR (qPCR) were
Table 1. Trait segregation of F1 and S1 plants derived from crossesamong the normal-type flower (NT), mutant-type flower(MT), and the flower apparently with complete somaticreversion (NT-SR).
z Plants with MT flowers include those bearing both MT and NT-SRflowers.
CrossPlant solely with NT
flowers(No. of plants (%))
Plant with MT flowersz (No. of plants (%))
NT × MT 70 (100) 0 (0)
MT × NT 69 (100) 0 (0)
MT × MT 115 (81.6) 26 (18.4)
NT-SR × NT-SR 133 (94.3) 8 (5.6)

T. Nishijima, Y. Morita, K. Sasaki, M. Nakayama, H. Yamaguchi, N. Ohtsubo, T. Niki and T. Niki42
designed to amplify 190–225 bp fragments of the 3'
regions of the cDNAs (listed in Table 2). Although two
DNA sequences for TfCHS are registered as TfCHS1
and TfCHS2, we designed a primer set for total
quantification of those genes without discriminating
between them. This is because the difference between
the two sequences is negligible for clear discrimination
as different genes. Primer sequences for torenia ACTIN3
(TfACT3) used as an internal standard were as described
previously (Niki et al., 2012). Gene expression was
quantified using a qPCR machine (LightCycler; Roche
Diagnostics, Mannheim, Germany) and SYBR Premix
Ex Taq (TakaraBio, Shiga, Japan). The reactions were
cycled with a preincubation step of 10 min at 95°C
followed by 50 cycles of 5 s at 95°C, 10 s at 60°C and
20 s at 72°C each. The raw data were analysed with
LightCycler software version 3.5 (Roche Diagnostics).
Plasmids (pGEM-T Easy vector) harbouring cDNAs of
the corresponding genes were used to obtain the standard
curves. The ratio of the expression of each gene with
that of TfACT3 was calculated. The expression analysis
was conducted independently in triplicate.
Construction of transgenic callus overexpressing
TfMYB1
The entire coding sequence of TfMYB1 cDNA was
amplified by PCR with the following primer set that
creates SalI and NotI sites: forward 5'-CACGGTCGAC
AATTATTTCGCGTTCAAATGAGTAGA-3' and re-
verse 5'-ATGAGCGGCCGCAATTACCACCATAAC
ATGGTCTCCTC-3' (single underline, restriction site;
double underline, start and stop codons). The amplified
fragment was inserted into pGreen binary Ti vector
(Hellens et al., 2000) at the SalI/NotI site downstream
of the CaMV 35S promoter. The construct was
transformed into Agrobacterium tumefaciens strain
EHA105, and then torenia ‘Common Violet’ was
transformed according to Aida et al. (2000). Plants
transformed with empty vector were used as the control.
T0 transgenic calli were used for analysis of anthocyanin
and gene expression as described above.
Molecular cloning of torenia transposon (Ttf1) and the
correspondence of Ttf1 insertion with mutant trait
The genomic sequence including the full-length open
reading frame of TfMYB1 was amplified by PCR using
the same primer set used for amplification of the full-
length cDNA as described above. Genomic DNA was
extracted from lateral limbs of lower lips using
MagExtractor Plant Genome Kit (TOYOBO, Osaka,
Japan) and used as templates. Comparing the amplified
sequences among NT, MT, and NT-SR flowers, we iden-
tified the DNA sequence of the torenia transposon (Ttf1,
Transposon Torenia fournieri 1) inserted in TfMYB1.
To confirm that insertion of Ttf1 into TfMYB1 actually
caused the mutant phenotype, the correlation between
the TfMYB1 genotype and the mutant trait in the S1segregants derived from self-pollination of the variegated
mutant flowers was investigated. We have indicated the
genotype of Ttf1-inserted TfMYB1 as myb1Ttf1, which is
a recessive allele, as proved below. The genotype of
TfMYB1 without Ttf1 insertion is indicated as MYB1.
We used 5 S1 mutant plants bearing solely MT flowers,
22 mutant plants bearing both MT and NT-SR flowers
within a plant and 18 apparent germinal revertant plants
bearing solely NT-GR flowers for the analysis. Lateral
limbs of lower lips of a MT and a NT-SR flowers were
collected from a mutant plant, while that of a NT-SR
flower was collected from a germinal revertant plant.
Sample collection was conducted at flower opening and
genomic DNA was extracted as described above. qPCR
analysis was performed as described above to confirm
the TfMYB1 genotype. Quantitative analysis was needed
because the lower lip of the MT flower has small violet
spots possibly caused by somatic reversion, although we
excluded MT flowers with large violet sectors from the
analysis. We used the following 2 primer sets (Fig. 11A).
Primer set A, forward 5'-CATTTGGCGACAGACCG
TTTC-3' and reverse 5'-TTTAATCGGCGGTTAATAAA
TGTTTC-3', amplifies the 153 bp sequence between the
right subterminal region of Ttf1 and the 2nd exon of
TfMYB1 in the myb1Ttf1 genotype, but does not amplify
any sequence of the MYB1 genotype. Primer set B,
forward 5'-TGAAGCTCACTTTGGTGCATTAAAC-3'
and reverse 5'-TTTAATCGGCGGTTAATAAATGTTTC-
3', amplifies a 104 bp sequence of MYB1 genotype, but
Table 2. Primers used for gene expression analysis using quantitative real-time PCR.
Target gene (Accession No.) Forward primer (5'-3') Reverse primer (5'-3')
TfCHS (AB548585 and AB548586) GAGGAACGTGCTGAGCGAGT ATTAATTGGGCGTGCTGTGC
TfCHI (AB548584) TCTTTTCACTCAATCACCTTCCG CACTATTTCAGGAACAACTCAGAGAGTC
TfF3H (AB548588) AGCCCATCACTTATGGAGAAATGTAC ACCTTAAGACGAATACGACATAGAGAATAT
TfDFR (AB548587) AATTTGAGGGAATTGACAAGGACA TTCGATGCATGTTCTATTCTATCTTATG
TfANS (AB548583) CCAAGGAGAAGATCGTGCTCA CAATAAACTCCACCCATCACTCAAC
TfUFGT (AB548591) GCGGCGTGTTCACTGAGAAT TTCCCAATCTTTGCACCGAT
TfMYB1 (AB719455) AGCAGATGATTCAAGATGAACGC CACCATAACATGGTCTCCTCCTC
TfbHLH1 (AB719456) AAGTTTCAGACAAGGAAGTGTTGATC TGAAGAGCCTGCCTTATCACACTAG
TfACTIN3 (AB330989) TTCACAGAAAATGCTTCTAAACAAAGAC AACGACACATAATGAAAGAACACAAGA

J. Japan. Soc. Hort. Sci. 82 (1): 39–50. 2013. 43
does not amplify any sequence of the myb1Ttf1 genotype
because the sequence to be amplified is too long
(3073 bp). qPCR analysis was performed as described
above using genomic DNA as a template. Plasmids
(pGEM-T Easy vector) harbouring the myb1Ttf1 and
MYB1 genotypes of TfMYB1 were used to obtain standard
curves. TfACT3 was employed as an internal standard
using the primer set described in Table 2. The genomic
sequence amplified by this primer set was the same size
as that of the cDNA sequence.
Estimation of copy number of Ttf1 and related elements
Genomic DNA extracted from normal type plants was
digested by EcoRI and XbaI, and subjected to Southern
blot analysis as described previously (Niki et al., 2001).
Probe construction and signal detection were performed
using a DIG DNA Labelling Kit and DIG Luminescent
Detection Kit, respectively, following the manufacturer’s
instructions (Roche Applied Science, Penzberg,
Germany). DNA sequence spanning 1712 bp from 520
to 2231 bp (from the left terminal) of Ttf1, which
excludes the subterminal and terminal regions, was used
as probe 1, while that spanning 377 bp from 2437 to
2813 bp within the right subterminal region was used as
probe 2.
Detection of Ttf1 in normal-type plant
To investigate whether the DNA sequence of Ttf1 is
mutated by EMS treatment, we performed molecular
cloning of Ttf1 in the normal-type plant. The full-length
Ttf1 sequence in the normal-type plant was obtained by
PCR using a primer set based on the terminal sequence
of Ttf1, forward 5'-CACAGCAAAAAAACATGTTTT
TAGCGACAGG-3' and reverse 5'-TCACACTAC
AAAATATAAGGGTTTTAGCGACGGA-3'. We further
conducted TAIL-PCR (Liu and Whittier, 1995) to
confirm the DNA sequence at those primer-binding sites.
Results and Discussion
Mutant traits and its heredity
In the normal-type plant, the limb of the upper lip of
the flower was solid pale violet, while that of the lower
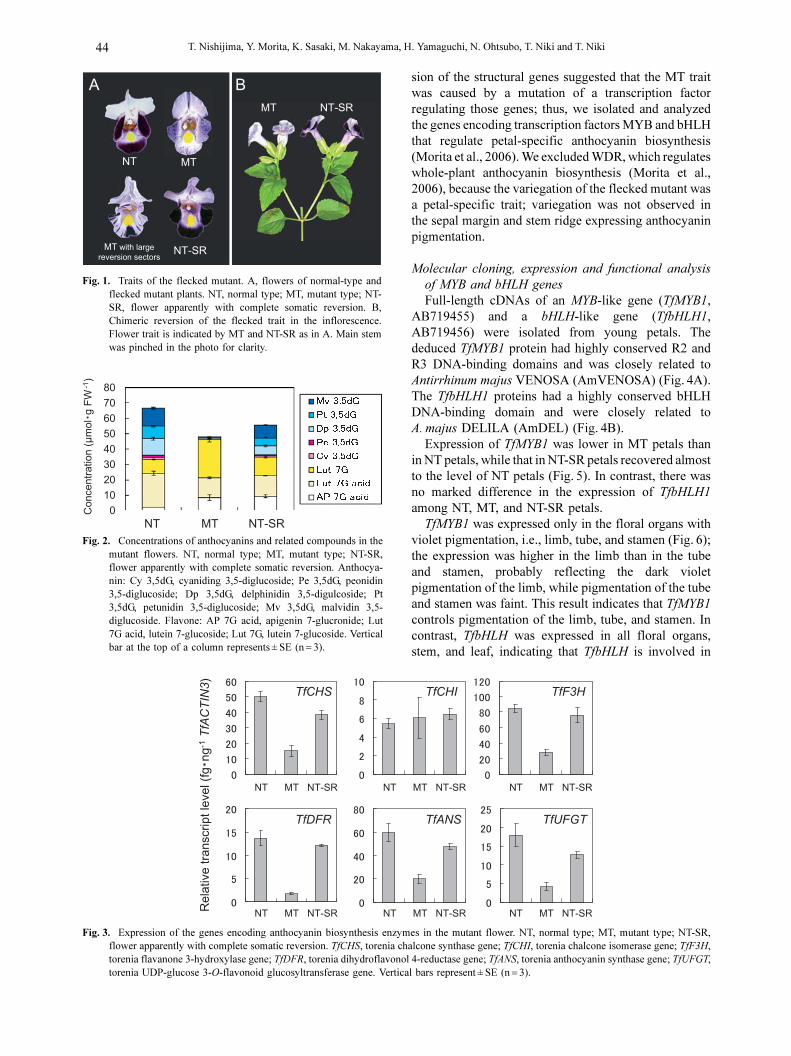
lip was solid violet (Fig. 1A, NT, normal type); however,
the lower lip of the mutant had small violet spots with
a pale violet background (Fig. 1A, MT, mutant type).
This mutant frequently displayed a semicircular violet
sector on its pale violet lower lips (Fig. 1A, MT with
large reversion sectors) or solid violet lower lip (Fig. 1A,
NT-SR, normal type apparently with complete somatic
reversion). The stamen and tube of NT flower had pale
violet pigmentation. The stamen pigmentation of MT
flower was paler than that of NT flower (photo not shown
because the color difference could not be clearly
photographed due to pale pigmentation), although
variegation was not clearly observed in MT flower. No
difference in color depth between NT and MT flowers
was observed in the tube. MT and NT-SR flowers
occurred chimerically within a plant (Fig. 1B). Once a
NT-SR flower occurred, the more apical flowers
seemingly generated from the same cell lineage were
always NT-SR. The pale violet background of the MT
flower indicated that anthocyanin biosynthesis leaked
slightly. Since those mutable traits resembled those of
the ‘flecked’ mutant of Ipomoea nil described by Hiraga
(1763), we named the torenia mutant and its trait (i.e.
variegated lower lip) as ‘flecked’. We refer to the limb
of the lower lip as simply ‘petal’ or ‘flower’ in the
following text unless otherwise stated.
All F1 plants derived from the cross between MT and
NT flowers showed the normal-type trait, suggesting
that MT is recessive (Table 1). If the MT or NT-SR
flower was self-pollinated, some S1 plants had MT
flowers. The ratio of S1 plants to MT flowers was higher
in the progeny of MT flowers than NT-SR flowers. If it
is hypothesized that the flecked mutation is caused by
a single recessive allele, the low rate of S1 plants with
the mutant trait (i.e. 18.4% as shown in Table 1) suggests
a high reversion rate. Of course, this is not direct evidence
of a high germinal reversion rate but is an indirect
indication. This is because apparent discrimination of
MT and NT-SR flowers depends on the anthocyanin
pigmentation pattern expressed in the L1 layer, while
reproductive cells are generated from the L2 layer which
is scarcely competent for anthocyanin biosynthesis.
Therefore, it is possible that the genotype of the causal
allele in the L2 layer is different from that estimated
from the petal pigmentation pattern appearing in the L1
layer.
Analysis of anthocyanins and expression of the genes
encoding anthocyanin biosynthesis enzymes
Anthocyanins (cyaniding 3,5-diglucoside, Cy 3,5dG;
peonidin 3,5-diglucoside, Pe 3,5dG; delphinidin 3,5-
digulcoside, Dp 3,5dG; petunidin 3,5-diglucoside, Pt
3,5dG; malvidin 3,5-diglucoside, Mv 3,5dG) and
flavones (apigenin 7-glucronide, AP 7G acid; lutein 7-
glucronide, Lut 7G acid; lutein 7-glucoside, Lut 7G)
listed in Figure 2 were identified from the petals. MT
petals had much lower anthocyanin and slightly higher
total flavone concentrations than NT petals (Fig. 2).
Anthocyanin and flavone concentrations of NT-SR petals
recovered to levels similar to those of NT petals. These
results suggest that inhibition of anthocyanin biosynthe-
sis mainly occurs in biosynthesis steps after that
catalyzed by F3H.
Expressions of TfCHS, TfF3H, TfDFR, TfANS, and
TfUFGT were lower in MT petals than in NT petals
(Fig. 3). In NT-SR petals, these genes were expressed at
almost the same levels as in NT petals. Expression of
TfCHI did not change markedly among the samples.
Estimation from the endogenous concentrations of
anthocyanins and related compounds suggested that CHS
had substantial activity even in MT flowers in which
TfCHS was down-regulated. Synchronous low expres-
T. Nishijima, Y. Morita, K. Sasaki, M. Nakayama, H. Yamaguchi, N. Ohtsubo, T. Niki and T. Niki44
sion of the structural genes suggested that the MT trait
was caused by a mutation of a transcription factor
regulating those genes; thus, we isolated and analyzed
the genes encoding transcription factors MYB and bHLH
that regulate petal-specific anthocyanin biosynthesis
(Morita et al., 2006). We excluded WDR, which regulates
whole-plant anthocyanin biosynthesis (Morita et al.,
2006), because the variegation of the flecked mutant was
a petal-specific trait; variegation was not observed in
the sepal margin and stem ridge expressing anthocyanin
pigmentation.
Molecular cloning, expression and functional analysis
of MYB and bHLH genes
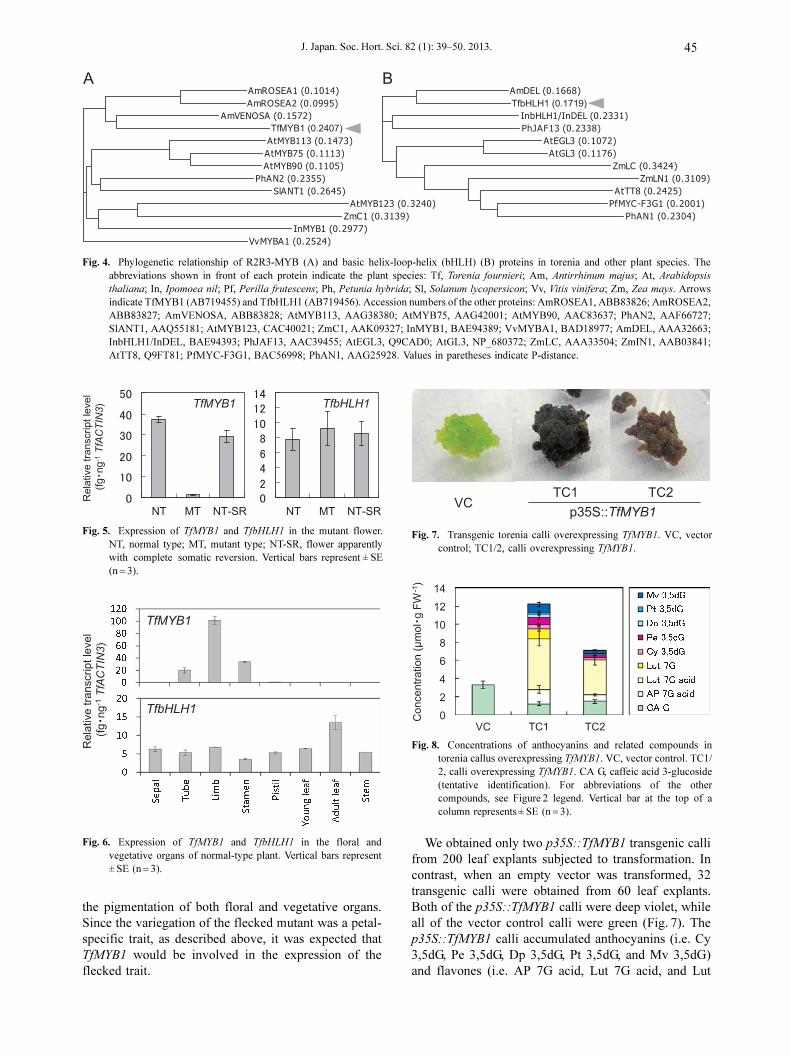
Full-length cDNAs of an MYB-like gene (TfMYB1,
AB719455) and a bHLH-like gene (TfbHLH1,
AB719456) were isolated from young petals. The
deduced TfMYB1 protein had highly conserved R2 and
R3 DNA-binding domains and was closely related to
Antirrhinum majus VENOSA (AmVENOSA) (Fig. 4A).
The TfbHLH1 proteins had a highly conserved bHLH
DNA-binding domain and were closely related to
A. majus DELILA (AmDEL) (Fig. 4B).
Expression of TfMYB1 was lower in MT petals than
in NT petals, while that in NT-SR petals recovered almost
to the level of NT petals (Fig. 5). In contrast, there was
no marked difference in the expression of TfbHLH1
among NT, MT, and NT-SR petals.
TfMYB1 was expressed only in the floral organs with
violet pigmentation, i.e., limb, tube, and stamen (Fig. 6);
the expression was higher in the limb than in the tube
and stamen, probably reflecting the dark violet
pigmentation of the limb, while pigmentation of the tube
and stamen was faint. This result indicates that TfMYB1
controls pigmentation of the limb, tube, and stamen. In
contrast, TfbHLH was expressed in all floral organs,
stem, and leaf, indicating that TfbHLH is involved in
Fig. 1. Traits of the flecked mutant. A, flowers of normal-type andflecked mutant plants. NT, normal type; MT, mutant type; NT-SR, flower apparently with complete somatic reversion. B,Chimeric reversion of the flecked trait in the inflorescence.Flower trait is indicated by MT and NT-SR as in A. Main stemwas pinched in the photo for clarity.
Fig. 2. Concentrations of anthocyanins and related compounds in themutant flowers. NT, normal type; MT, mutant type; NT-SR,flower apparently with complete somatic reversion. Anthocya-nin: Cy 3,5dG, cyaniding 3,5-diglucoside; Pe 3,5dG, peonidin3,5-diglucoside; Dp 3,5dG, delphinidin 3,5-digulcoside; Pt3,5dG, petunidin 3,5-diglucoside; Mv 3,5dG, malvidin 3,5-diglucoside. Flavone: AP 7G acid, apigenin 7-glucronide; Lut7G acid, lutein 7-glucoside; Lut 7G, lutein 7-glucoside. Verticalbar at the top of a column represents ± SE (n = 3).
Fig. 3. Expression of the genes encoding anthocyanin biosynthesis enzymes in the mutant flower. NT, normal type; MT, mutant type; NT-SR,flower apparently with complete somatic reversion. TfCHS, torenia chalcone synthase gene; TfCHI, torenia chalcone isomerase gene; TfF3H,torenia flavanone 3-hydroxylase gene; TfDFR, torenia dihydroflavonol 4-reductase gene; TfANS, torenia anthocyanin synthase gene; TfUFGT,torenia UDP-glucose 3-O-flavonoid glucosyltransferase gene. Vertical bars represent ± SE (n = 3).
J. Japan. Soc. Hort. Sci. 82 (1): 39–50. 2013. 45
the pigmentation of both floral and vegetative organs.
Since the variegation of the flecked mutant was a petal-
specific trait, as described above, it was expected that
TfMYB1 would be involved in the expression of the
flecked trait.
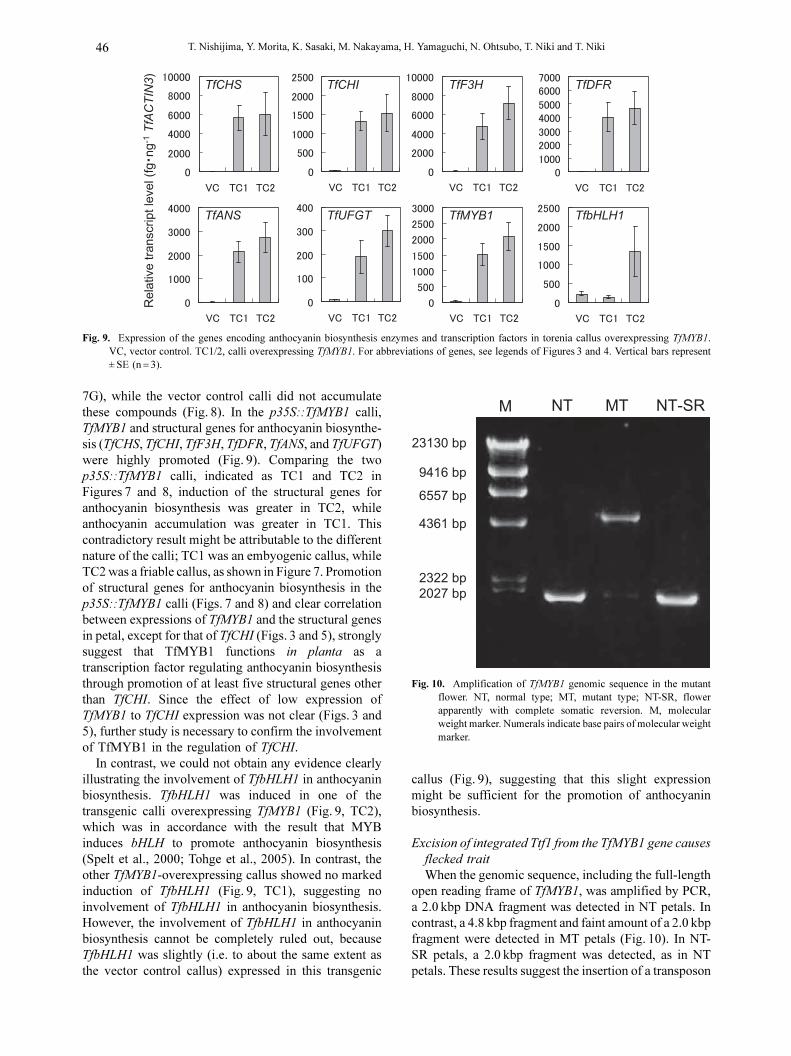
We obtained only two p35S::TfMYB1 transgenic calli
from 200 leaf explants subjected to transformation. In
contrast, when an empty vector was transformed, 32
transgenic calli were obtained from 60 leaf explants.
Both of the p35S::TfMYB1 calli were deep violet, while
all of the vector control calli were green (Fig. 7). The
p35S::TfMYB1 calli accumulated anthocyanins (i.e. Cy
3,5dG, Pe 3,5dG, Dp 3,5dG, Pt 3,5dG, and Mv 3,5dG)
and flavones (i.e. AP 7G acid, Lut 7G acid, and Lut
Fig. 4. Phylogenetic relationship of R2R3-MYB (A) and basic helix-loop-helix (bHLH) (B) proteins in torenia and other plant species. Theabbreviations shown in front of each protein indicate the plant species: Tf, Torenia fournieri; Am, Antirrhinum majus; At, Arabidopsis
thaliana; In, Ipomoea nil; Pf, Perilla frutescens; Ph, Petunia hybrida; Sl, Solanum lycopersicon; Vv, Vitis vinifera; Zm, Zea mays. Arrowsindicate TfMYB1 (AB719455) and TfbHLH1 (AB719456). Accession numbers of the other proteins: AmROSEA1, ABB83826; AmROSEA2,ABB83827; AmVENOSA, ABB83828; AtMYB113, AAG38380; AtMYB75, AAG42001; AtMYB90, AAC83637; PhAN2, AAF66727;SlANT1, AAQ55181; AtMYB123, CAC40021; ZmC1, AAK09327; InMYB1, BAE94389; VvMYBA1, BAD18977; AmDEL, AAA32663;InbHLH1/InDEL, BAE94393; PhJAF13, AAC39455; AtEGL3, Q9CAD0; AtGL3, NP_680372; ZmLC, AAA33504; ZmIN1, AAB03841;AtTT8, Q9FT81; PfMYC-F3G1, BAC56998; PhAN1, AAG25928. Values in paretheses indicate P-distance.
Fig. 5. Expression of TfMYB1 and TfbHLH1 in the mutant flower.NT, normal type; MT, mutant type; NT-SR, flower apparentlywith complete somatic reversion. Vertical bars represent ± SE(n = 3).
Fig. 6. Expression of TfMYB1 and TfbHLH1 in the floral andvegetative organs of normal-type plant. Vertical bars represent± SE (n = 3).
Fig. 7. Transgenic torenia calli overexpressing TfMYB1. VC, vectorcontrol; TC1/2, calli overexpressing TfMYB1.
Fig. 8. Concentrations of anthocyanins and related compounds intorenia callus overexpressing TfMYB1. VC, vector control. TC1/2, calli overexpressing TfMYB1. CA G, caffeic acid 3-glucoside(tentative identification). For abbreviations of the othercompounds, see Figure 2 legend. Vertical bar at the top of acolumn represents ± SE (n = 3).
T. Nishijima, Y. Morita, K. Sasaki, M. Nakayama, H. Yamaguchi, N. Ohtsubo, T. Niki and T. Niki46
7G), while the vector control calli did not accumulate
these compounds (Fig. 8). In the p35S::TfMYB1 calli,
TfMYB1 and structural genes for anthocyanin biosynthe-
sis (TfCHS, TfCHI, TfF3H, TfDFR, TfANS, and TfUFGT)
were highly promoted (Fig. 9). Comparing the two
p35S::TfMYB1 calli, indicated as TC1 and TC2 in
Figures 7 and 8, induction of the structural genes for
anthocyanin biosynthesis was greater in TC2, while
anthocyanin accumulation was greater in TC1. This
contradictory result might be attributable to the different
nature of the calli; TC1 was an embyogenic callus, while
TC2 was a friable callus, as shown in Figure 7. Promotion
of structural genes for anthocyanin biosynthesis in the
p35S::TfMYB1 calli (Figs. 7 and 8) and clear correlation
between expressions of TfMYB1 and the structural genes
in petal, except for that of TfCHI (Figs. 3 and 5), strongly
suggest that TfMYB1 functions in planta as a
transcription factor regulating anthocyanin biosynthesis
through promotion of at least five structural genes other
than TfCHI. Since the effect of low expression of
TfMYB1 to TfCHI expression was not clear (Figs. 3 and
5), further study is necessary to confirm the involvement
of TfMYB1 in the regulation of TfCHI.
In contrast, we could not obtain any evidence clearly
illustrating the involvement of TfbHLH1 in anthocyanin
biosynthesis. TfbHLH1 was induced in one of the
transgenic calli overexpressing TfMYB1 (Fig. 9, TC2),
which was in accordance with the result that MYB
induces bHLH to promote anthocyanin biosynthesis
(Spelt et al., 2000; Tohge et al., 2005). In contrast, the
other TfMYB1-overexpressing callus showed no marked
induction of TfbHLH1 (Fig. 9, TC1), suggesting no
involvement of TfbHLH1 in anthocyanin biosynthesis.
However, the involvement of TfbHLH1 in anthocyanin
biosynthesis cannot be completely ruled out, because
TfbHLH1 was slightly (i.e. to about the same extent as
the vector control callus) expressed in this transgenic
callus (Fig. 9), suggesting that this slight expression
might be sufficient for the promotion of anthocyanin
biosynthesis.
Excision of integrated Ttf1 from the TfMYB1 gene causes
flecked trait
When the genomic sequence, including the full-length
open reading frame of TfMYB1, was amplified by PCR,
a 2.0 kbp DNA fragment was detected in NT petals. In
contrast, a 4.8 kbp fragment and faint amount of a 2.0 kbp
fragment were detected in MT petals (Fig. 10). In NT-
SR petals, a 2.0 kbp fragment was detected, as in NT
petals. These results suggest the insertion of a transposon
Fig. 9. Expression of the genes encoding anthocyanin biosynthesis enzymes and transcription factors in torenia callus overexpressing TfMYB1.VC, vector control. TC1/2, calli overexpressing TfMYB1. For abbreviations of genes, see legends of Figures 3 and 4. Vertical bars represent± SE (n = 3).
Fig. 10. Amplification of TfMYB1 genomic sequence in the mutantflower. NT, normal type; MT, mutant type; NT-SR, flowerapparently with complete somatic reversion. M, molecularweight marker. Numerals indicate base pairs of molecular weightmarker.

J. Japan. Soc. Hort. Sci. 82 (1): 39–50. 2013. 47
in TfMYB1. In contrast, amplification of the TfbHLH1
genomic sequence, including the full-length open
reading frame, gave an approximately 2.4 kbp fragment
in all of the NT, MT, and NT-SR petals, suggesting no
insertion of transposon (data not shown). Molecular
cloning of the 2.0 kbp DNA fragment of TfMYB1 showed
that TfMYB1 consisted of 3 exons and 2 introns
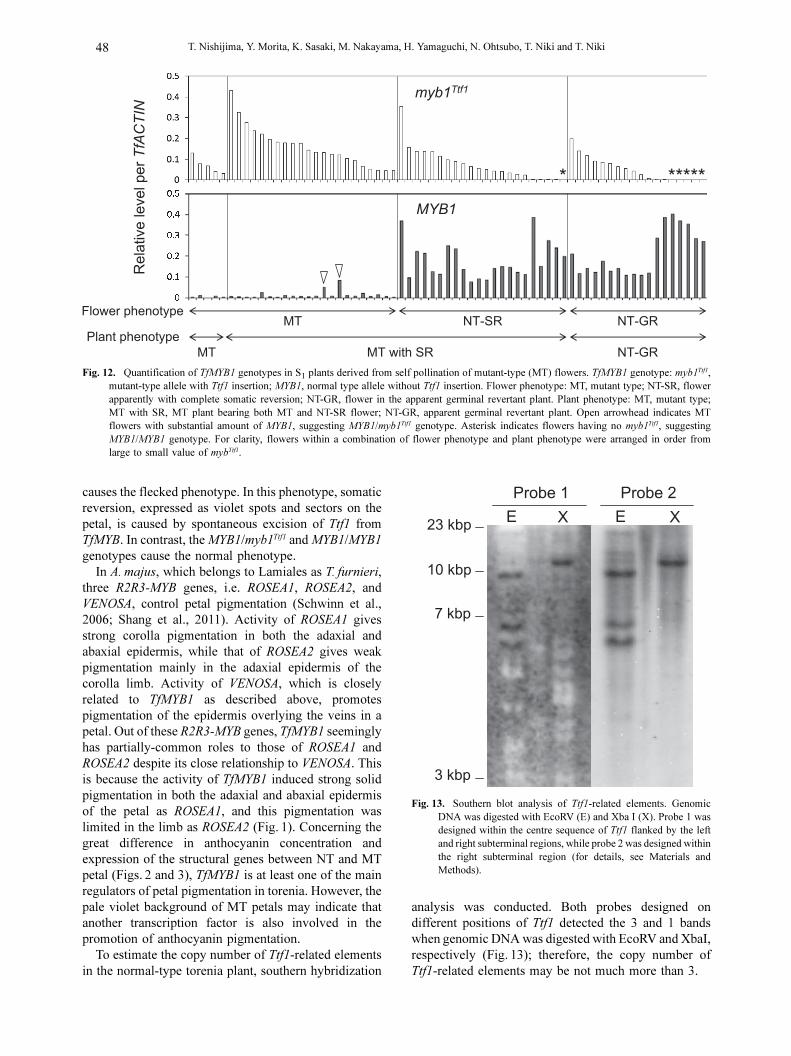
(Fig. 11A). Further, sequence analysis of the 4.8 kbp
fragment showed insertion of a 2893 bp DNA sequence
in the 2nd intron of TfMYB1 (Fig. 11A). This inserted
sequence (named Ttf1 as described in Materials and
Methods, AB719454) had the typical structure of a
transposon (Fig. 11B, C). The left subterminal 264 bp
and right subterminal 456 bp regions had repeats of the
6 bp CGACAG motif. The right subterminal region had
102 bp and 37 bp tandem repeats. Both left and right
terminals had terminal inverted repeats. The DNA
sequence of the terminal inverted repeat suggested that
Ttf1 belongs to the family of En/Spm transposon
(Fig. 11C). Ttf1 was flanked by 3-bp direct repeat TGA
that was identical to the integration site (Fig. 11B). This
result also supports the idea that Ttf1 belongs to the
family of En/Spm transposon, because the Spm/En
transposon and its relatives generate 3-bp target
duplications when integrated (Gierl and Saedler, 1992).
No DNA sequences encoding known proteins, including
transposase, were detected in Ttf1, suggesting that Ttf1
is a non-autonomous element. Moreover, in the 2.0 kbp
fragment of NT-SR petals, 6 bp of the target duplication
sequence was converted to 3- and 4-bp new sequences
(Fig. 11B), which were generated as footprints of Ttf1
excision.
qPCR analysis of genomic DNA in S1 segregants
showed clear correspondence between the quantities of
TfMYB1 genotypes (i.e. MYB1 and myb1Ttf1) and the
flower traits (Fig. 12). Most MT flowers exhibited a high
quantity of mybTtf1, while slight MYB1 was detected,
probably because of somatic reversion, observed as small
violet spots on the petal (Fig. 1A MT). This result
indicates that the genotype of MT flowers is basically
myb1Ttf1/myb1Ttf1; however, 2 out of 27 MT flowers
exhibited a substantial amount of MYB1 (indicated by
open arrowhead in Fig. 12). Since these two flowers
exhibited a low expression of TfMYB1, as expected from
their MT phenotype (data not shown), the MYB1/myb1Ttf1
or MYB1/MYB1 genotypes very probably occurred in
the L2 and/or L3 layers of the petals, which scarcely
induce anthocyanin biosynthesis.
In contrast, all NT-SR and NT-GR flowers exhibited
a high quantity of MYB1. myb1Ttf1 was detected in 21
out of 22 NT-SR flowers, suggesting that these flowers
have MYB1/myb1Ttf1 genotype, while the other NT-SR
flower had no detectable myb1Ttf (indicated by an asterisk
in Fig. 12), suggesting that this flower has MYB1/MYB1
genotype. In NT-GR flowers, MYB1 was detected in 13
out of 18 flowers, suggesting that these flowers have
MYB1/myb1Ttf1 genotype; however, the rate of flowers
without detectable myb1Ttf, which probably have MYB1/
MYB1 genotype, increased to 5 out of 18 flowers. These
results correspond with the natural prediction that
germinal revertant plants have a higher rate of MYB1/
MYB1 flowers than mutant plants with somatic reversion.
Overall, we concluded that the homozygous allele of
Ttf1-inserted TfMYB1 (i.e. genotype myb1Ttf1/myb1Ttf1)
Fig. 11. Characterization of torenia transposon Ttf1 (Transposon Torenia fournieli 1). A: Structure of TfMYB1 carrying Ttf1. Obliquely-stripedsquare, 5' UTR; grey square, coding region; dashed line, intron. Oblique double lines on the square indicating Ttf1 represent abbreviationbecause Ttf1 is too long to be illustrated to the right scale. Primer binding sites for qPCR analysis of TfMYB1 genotype are indicated byarrows; black arrow, forward primer for specific detection of TfMYB1 without Ttf1 insertion (MYB1 genotype); open arrow; forward primerfor specific detection of TfMYB1 with Ttf1 insertion (myb1Ttf1 genotype); grey arrow, common reverse primer. B: Structure of Ttf1 (uppercolumn), DNA sequence around Ttf1 insertion site of TfMYB1 (lower column), and footprint sequences formed by excision of Ttf1 (TCGAand TCA). Open square, subterminal region; large black triangle, terminal inverted repeat; small black triangle, 6 bp CGACAG motif; largegrey pentagon, 102 bp tandem repeat; small grey pentagon, 37 bp tandem repeat. Sequences of TGA underlined in TfMYB1 sequence are3 bp target duplications. C: DNA sequence of terminal inverted repeats (TIR) of Ttf1.
T. Nishijima, Y. Morita, K. Sasaki, M. Nakayama, H. Yamaguchi, N. Ohtsubo, T. Niki and T. Niki48
causes the flecked phenotype. In this phenotype, somatic
reversion, expressed as violet spots and sectors on the
petal, is caused by spontaneous excision of Ttf1 from
TfMYB. In contrast, the MYB1/myb1Ttf1 and MYB1/MYB1
genotypes cause the normal phenotype.
In A. majus, which belongs to Lamiales as T. furnieri,
three R2R3-MYB genes, i.e. ROSEA1, ROSEA2, and
VENOSA, control petal pigmentation (Schwinn et al.,
2006; Shang et al., 2011). Activity of ROSEA1 gives
strong corolla pigmentation in both the adaxial and
abaxial epidermis, while that of ROSEA2 gives weak
pigmentation mainly in the adaxial epidermis of the
corolla limb. Activity of VENOSA, which is closely
related to TfMYB1 as described above, promotes
pigmentation of the epidermis overlying the veins in a
petal. Out of these R2R3-MYB genes, TfMYB1 seemingly
has partially-common roles to those of ROSEA1 and
ROSEA2 despite its close relationship to VENOSA. This
is because the activity of TfMYB1 induced strong solid
pigmentation in both the adaxial and abaxial epidermis
of the petal as ROSEA1, and this pigmentation was
limited in the limb as ROSEA2 (Fig. 1). Concerning the
great difference in anthocyanin concentration and
expression of the structural genes between NT and MT
petal (Figs. 2 and 3), TfMYB1 is at least one of the main
regulators of petal pigmentation in torenia. However, the
pale violet background of MT petals may indicate that
another transcription factor is also involved in the
promotion of anthocyanin pigmentation.
To estimate the copy number of Ttf1-related elements
in the normal-type torenia plant, southern hybridization
analysis was conducted. Both probes designed on
different positions of Ttf1 detected the 3 and 1 bands
when genomic DNA was digested with EcoRV and XbaI,
respectively (Fig. 13); therefore, the copy number of
Ttf1-related elements may be not much more than 3.
Fig. 12. Quantification of TfMYB1 genotypes in S1 plants derived from self pollination of mutant-type (MT) flowers. TfMYB1 genotype: myb1Ttf1,mutant-type allele with Ttf1 insertion; MYB1, normal type allele without Ttf1 insertion. Flower phenotype: MT, mutant type; NT-SR, flowerapparently with complete somatic reversion; NT-GR, flower in the apparent germinal revertant plant. Plant phenotype: MT, mutant type;MT with SR, MT plant bearing both MT and NT-SR flower; NT-GR, apparent germinal revertant plant. Open arrowhead indicates MTflowers with substantial amount of MYB1, suggesting MYB1/myb1Ttf1 genotype. Asterisk indicates flowers having no myb1Ttf1, suggestingMYB1/MYB1 genotype. For clarity, flowers within a combination of flower phenotype and plant phenotype were arranged in order fromlarge to small value of mybTtf1.
Fig. 13. Southern blot analysis of Ttf1-related elements. GenomicDNA was digested with EcoRV (E) and Xba I (X). Probe 1 wasdesigned within the centre sequence of Ttf1 flanked by the leftand right subterminal regions, while probe 2 was designed withinthe right subterminal region (for details, see Materials andMethods).

J. Japan. Soc. Hort. Sci. 82 (1): 39–50. 2013. 49
In normal-type plants, we detected a DNA sequence
that was identical to that of Ttf1 in the flecked mutant;
therefore, the high transposition activity of Ttf1 in the
mutant cannot be attributed to a mutation in the base
sequence of Ttf1.
Potential use of Ttf1 in breeding and transposon tagging
of torenia
The high germinal reversion rate of the flecked mutant,
as shown in Table 1, indicates high transposition activity
of Ttf1 in the reproductive cells and/or L2 layer, which
is highly likely to cause novel mutations. When Ttf1 is
used as a mutagen for torenia breeding, the high
transposition activity may also make the mutant
phenotype unstable; however, the mutation might be
stabilized if Ttf1 and the allele promoting transposition
of Ttf1 (possibly the allele encoding the transposase for
Ttf1) are genetically separated. The allele promoting Ttf1
transposition may be homozygous in the flecked mutant,
because a mutant plant with no reversion was not
separated by M6 generation. We are currently
constructing an F2 population derived from cross
pollination of NT and MT flowers, which may enable
genetic separation of Ttf1 and the allele promoting Ttf1
transposition.
Since the copy number of the Ttf1-related elements
in the torenia genome may be small, as discussed above,
Ttf1 could potentially be useful for transposon tagging.
If Ttf1 is the sole transposon with high transposition
activity out of the related elements, an efficient
transposon tagging system could be established using
the flecked mutant. To confirm this, complete analysis
of the Ttf1-related elements is necessary.
Acknowledgements
We thank Mrs. Tomoko Kurobe for her technical
assistance.
Literature Cited
Aida, R., S. Kishimoto, Y. Tanaka and M. Shibata. 2000.
Modification of flower color in torenia (Torenia fournieri
Lind.) by genetic transformation. Plant Sci. 153: 33–42.
Bonas, U., H. Sommer and H. Saedler. 1984. The 17-kb Tam1
element of Antirrhinum majus induces a 3-bp duplication
upon integration into the chalcone synthase gene. EMBO J.
3: 1015–1019.
Broun, P. 2005. Transcriptional control of flavonoid biosynthesis:
a complex network of conserved regulators involved in
multiple aspects of differentiation in Arabidopsis. Curr. Opin.
Plant Biol. 8: 272–279.
Coen, E. S., J. M. Romero, S. Doyle, R. Elliott, G. Murphy and
R. Carpenter. 1990. floricaula: A homeotic gene required for
flower development in Antirrhinum majus. Cell 63: 1311–
1322.
Flavell, A. J., S. R. Pearce and A. Kumar. 1994. Plant transposable
elements and the genome. Curr. Opin. Genet. Dev. 4: 838–844.
Forkmann, G. 1994. Biosynthesis of flavonoids. p. 499–535. In:
J. B. Harbone (ed.). The flavonoids. Chapman & Hall,
London.
Frohman, M. A., M. K. Dush and G. R. Martin. 1988. Rapid
production of full-length cDNAs from rare transcripts:
amplification using a single gene-specific oligonucleotide
primer. Proc. Natl. Acad. Sci. USA 85: 8998–9002.
Gierl, A. and H. Saedler. 1992. Plant-transposable elements and
gene tagging. Plant Mol. Biol. 19: 39–49.
Goodrich, J., R. Carpenter and E. S. Coen. 1992. A common gene
regulates pigmentation pattern in diverse plant species. Cell
68: 955–964.
Hellens, R. P., E. A. Edwards, N. R. Leyland, S. Bean and P. M.
Mullineaux. 2000. pGreen: a versatile flexible binary Ti vector
for Agrobacterium-mediated plant transformation. Plant Mol.
Biol. 42: 819–832.
Heller, W. and G. Forkmann. 1994. Genetics of flavonoids. p. 537–
564. In: J. B. Harbone (ed.). The flavonoids. Chapman &
Hall, London.
Hiraga, G. 1763 (reprinted in 1972). Butsurui-hinshitsu (In
Japanese). Yasaka-Shobo, Tokyo.
Inagaki, Y., Y. Hisatomi, T. Suzuki, K. Kasahara and S. Iida. 1994.
Isolation of a Suppressor-Mutator/Enhancer-like transpos-
able element, Tpn1, from Japanese morning glory bearing
variegated flowers. Plant Cell 6: 375–383.
Koes, R., W. Verweij and F. Quattrocchio. 2005. Flavonoids: a
colorful model for the regulation and evolution of biochemical
pathways. Trends Plant Sci. 10: 236–242.
Liu, Y. G. and R. F. Whittier. 1995. Thermal asymmetric interlaced
PCR: Automatable amplification and sequencing of insert
end fragments from P1 and YAC clones for chromosome
walking. Genomics 25: 674–681.
Luo, D., E. S. Coen, S. Doyle and R. Carpenter. 1991. Pigmentation
mutants produced by transposon mutagenesis in Antirrhinum
majus. Plant J. 1: 59–69.
Martin, C., R. Carpenter, H. Sommer, H. Saedler and E. S. Coen.
1985. Molecular analysis of instability in flower pigmentation
of Antirrhinum majus, following isolation of the pallida locus
by transposon tagging. EMBO J. 4: 1625–1630.
Matsubara, K., H. Kodama, H. Kokubun, H. Watanabe and T.
Ando. 2005. Two novel transposable elements in a
cytochrome P450 gene govern anthocyanin biosynthesis of
commercial petunias. Gene 358: 121–126.
Miyazaki, K., C. Kuwayama, K. Kageyama, S. Matsumoto and
H. Fukui. 2007. Relation between interspecific compatibility
and phylogenic analysis in Torenia. Shokubutsu Kankyo
Kogaku 19: 66–73 (In Japanese with English abstract).
Morise, N. 2001. Torenia. p. 567–568. In: Nosangyosonbunka-
kyokai (ed.). Nogyogijutsutaikei Kakihen 8 (In Japanese).
Nosangyosonbunkakyokai, Tokyo.
Morita, Y., M. Saitoh, A. Hoshino, E. Nitasaka and S. Iida. 2006.
Isolation of cDNAs for R2R3-MYB, bHLH and WDR
transcriptional regulators and identification of c and ca
mutations conferring white flowers in the Japanese morning
glory. Plant Cell Physiol. 47: 457–470.
Nakajima, T., K. Matsubara, H. Kodama, H. Kokubun, H.
Watanabe and T. Ando. 2005. Insertion and excision of a
transposable element governs the red floral phenotype in
commercial petunias. Theor. Appl. Genet. 110: 1038–1043.
National Agriculture and Food Research Organaization. 2006.
Torenia. p. 1112. In: National Agriculture and Food Research
Organization (ed.). NAROPEDIA (In Japanese). Nosan-
gyosonbunkakyokai, Tokyo.
Niki, T., M. Hirai, T. Niki, A. Kanno and T. Nishijima. 2012. Role
of homeotic genes in the morphology of forchlorfenuron-
induced paracorollas in Torenia fournieri Lind. J. Japan. Soc.
Hort. Sci. 81: 204–212.
Niki, T., T. Nishijima, M. Nakayama, T. Hisamatsu, N. Oyama-

T. Nishijima, Y. Morita, K. Sasaki, M. Nakayama, H. Yamaguchi, N. Ohtsubo, T. Niki and T. Niki50
Okubo, H. Yamazaki, P. Hedden, T. Lange, L. N. Mander
and M. Koshioka. 2001. Production of dwarf lettuce by
overexpressing a pumpkin gibberellin 20-oxidase gene. Plant
Physiol. 126: 965–972.
Nishijima, T. and K. Shima. 2006. Change in flower morphology
of Torenia fournieri Lind. induced by forchlorfenuron
application. Sci. Hortic. 109: 254–261.
Nitasaka, E. 2003. Insertion of an En/Spm-related transposable
element into a floral homeotic gene DUPLICATED causes a
double flower phenotype in the Japanese morning glory. Plant
J. 36: 522–531.
Nitasaka, E. 2007. Morphological mutants induced by transposable
elements in the Japanese morning glory. Gamma Field Symp.
46: 73–79.
Okuda, S., H. Tsutsui, K. Shiina, S. Sprunck, H. Takeuchi, R. Yui,
D. Kasahara, Y. Hamamura, A. Mizukami, D. Susaki, N.
Kawano, T. Sakakibara, S. Namiki, K. Itoh, K. Otsuka, M.
Matsuzaki, H. Nozaki, T. Kuroiwa, A. Nakano, M. Kanaoka,
T. Dresselhaus, N. Sasaki and T. Higashiyama. 2009.
Defensin-like polypeptide LUREs are pollen tube attractants
secreted from synergid cells. Nature 458: 357–361.
Sasaki, K., R. Aida, T. Niki, H. Yamaguchi, T. Narumi, T.
Nishijima, Y. Hayashi, H. Ryuto, N. Fukunishi, T. Abe and
N. Otsubo. 2008. High-efficiency improvement of transgenic
torenia flowers by ion beam irradiation. Plant Biotechnol. 25:
81–90.
Schwinn, K., J. Venail, Y. Shang, S. Mackay, V. Alm, E. Butelli,
R. Oyama, P. Bailey, K. Davies and C. Martin. 2006. A small
family of MYB-regulatory genes controls floral pigmentation
intensity and patterning in the genus Antirrhinum. Plant Cell
18: 831–851.
Shang, Y., J. Venail, S. Mackay, P. C. Bailey, K. E. Schwinn, P. E.
Jameson, C. R. Martin and K. M. Davies. 2011. The molecular
basis for venation patterning of pigmentation and its effect
on pollinator attraction in flowers of Antirrhinum. New
Phytologist 189: 602–615.
Sommer, H., R. Carpenter, B. J. Harrison and H. Saedler. 1985.
The transposable element of Tam3 of Antirrhinum majus
generates a novel type of sequence alterations upon excision.
Mol. Gen. Genet. 199: 225–231.
Spelt, C., F. Quattrocchio, J. N. M. Mol and R. Koes. 2000.
anthocyanin1 of petunia encodes a basic helix-loop-helix
protein that directly activates transcription of structural
anthocyanin genes. Plant Cell 12: 1619–1631.
Tohge, T., Y. Nishiyama, M. Yokota-Hirai, M. Yano, J. Nakajima,
M. Awazuhara, E. Inoue, H. Takahashi, D. B. Goodenowe,
M. Kitayama, M. Noji, M. Yamazaki and K. Saito. 2005.
Functional genomics by integrated analysis of metabolome
and transcriptome of Arabidopsis plants over-expressing an
MYB transcription factor. Plant J. 42: 218–235.
Recommended

















