
Biología Sensorial Animal
Teórica #22
Otros estímulos:
Gravitacional
Campo magnético

ERICH VON HOLST • Invaluables contribuciones al desarrollo de la neuroetología.
• Extraordinaria habilidad experimental, trabajo de doctorado,
eliminando segmentos del cuerpo de lombriz, pero dejando el cordón
nervioso intacto, demostró el patrón de movimiento estaba controlado
por un ritmo endógeno producido por el sistema nervioso central (hoy
esto se conoce como Central Pattern Generator).
• Además trabajó en biofísica del vuelo de aves, mecanismo del control
neuronal del comportamiento (estimulación eléctrica cerebro de
pollos). Con Mittelstaed formuló el principio de reaferencia, modelo
que explica cómo animales y humanos distinguir movimientos de las
imágenes en la retina causados por el movimiento de objetos de
aquellos producidos por el movimiento del propio cuerpo.
• Además, era un eximio músico y un extraordinario luthier.
• Konrad Lorenz: “Estoy convencido de que si él (Von Holst) estuviese
todavía vivo, estaría ahora aquí en Estocolmo”.

Sin otolito
S = shearing force (cte)
P = pressure force
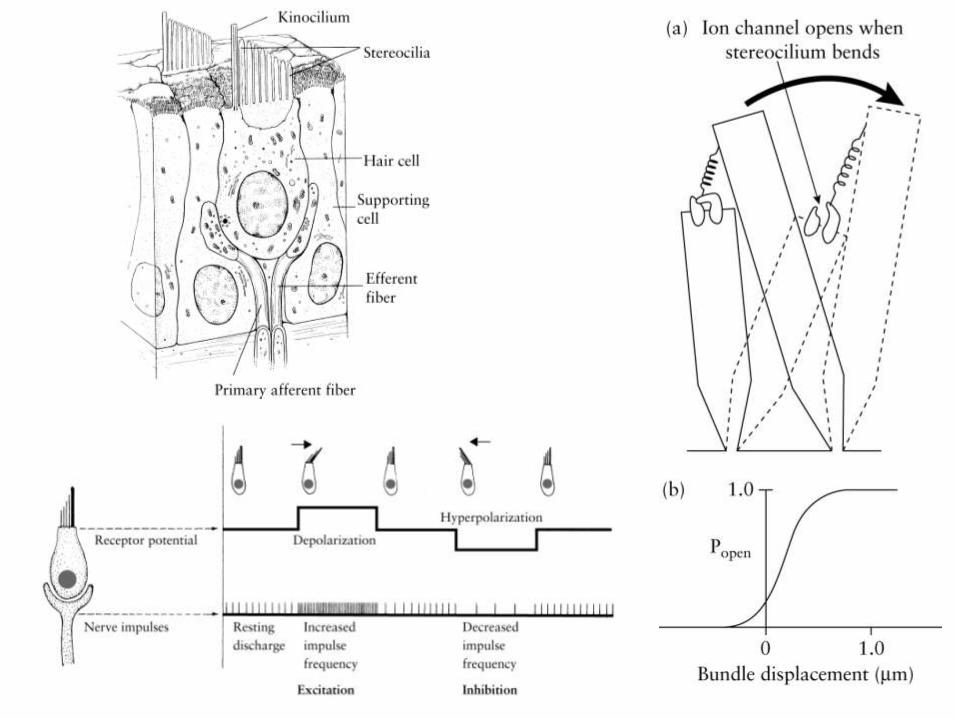

Otros estímulos
Campo magnético

Especie de ave paseriforme de la familia
Turdidae. El ave adulto mide 12,5-14,0 cm de
largo, pesa 16-22 g, y tiene una envergadura de
20-22 cm. Es un ave diurna, aunque
ocasionalmente caza insectos en las noches de
luna, o cerca de luz artificial. Se alimenta
principalmente de invertebrados terrestres,
como arañas, lombrices e insectos. En otoño e
invierno, complementa su dieta habitual con
bayas y frutas.
Su área de distribución abarca Europa, el
noroeste de Asia y el norte de África, y se
extiende hasta el oeste de Siberia en el oriente,
en el sureste hasta la cordillera del Cáucaso, y
en el sur hasta Argelia. Incluye también las Islas
Británicas y la Macaronesia. No se encuentra en
Islandia. Los petirrojos de Escandinavia y Rusia
migran hacia Europa occidental para escapar de
los inviernos más fríos en su territorio de
anidación.
El petirrojo europeo (Erithacus rubecula)

Magnetorrecepción
• La brújula magnética aviar del Petirrojo Europeo
(Erithacus rubecula) ha sido ampliamente estudiada y
utiliza magnetorrecepción basada en la visión, en el cual la
capacidad de detectar el campo magnético de la tierra para
la navegación se ve afectado por la luz que entra en el ojo
del ave. Aunque todavía no se entiende por completo el
mecanismo físico de su sentido magnético, se considera
que podría incluir entrelazamiento cuántico de espines de
electrones.

Metodo para cuantificar (Emlen)
a) y b) con estrellas “falsas”, c) sin
Raw data vector

Bobina (“coil”) de Helmholtz
Aparato para generar una región uniforme
de campo magnético. Nombrado en honor
al físico Alemán Hermann von Helmholtz.
Consiste en dos bobinas magnéticas
circulares idénticas colocadas
simétricamente a cada lado del área
experimental y separadas por una distancia
igual al radio de la bobina. Cada bobina
lleva la misma corriente eléctrica.
Se puede utilizar para cancelar el campo
magnético terrestre.
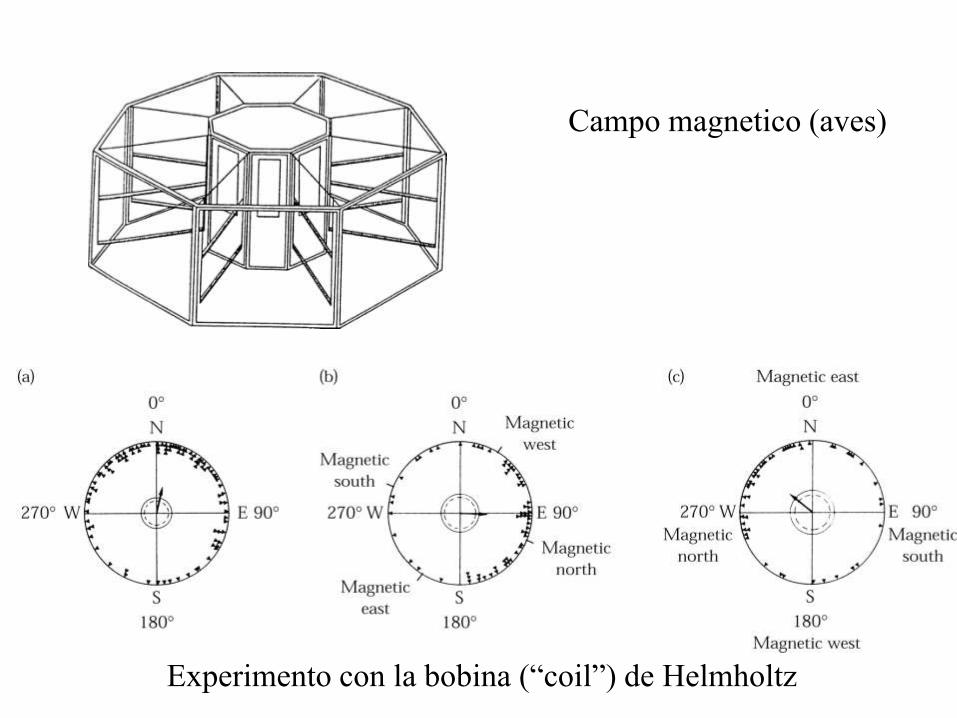
Campo magnetico (aves)
Experimento con la bobina (“coil”) de Helmholtz

Campo magnetico (aves)
Efecto VW

Modo funcional del compás magnético
• La forma en la cual la información
direccional se deriva de las líneas
magnéticas permite distinguir dos tipos de
mecanismos:
– compás de inclinación
– compás de polaridad.

El compás de inclinación
• Descripto primero en aves migratorias. Aves cambian la
preferencia de dirección cuando el componente horizontal del
campo magnético ambiental cambia.
• Sin embargo, ese cambio también sucede cuando el componente
vertical se invierte. Cuando ambos componentes cambian, las aves
testeadas responden igual que el campo geomagnético.
• Las aves obviamente ignoran la polaridad del campo magnético y
derivan de información direccional de la inclinación del eje de las
líneas del campo. Fundamentalmente diferente de una brújula.
• Compás de inclinación se encontró en todas las especies de aves
testeadas hasta el momento. Característica común dentro de la
clase aves. Sin embargo, también hallado en salamandras
acuáticas y jóvenes tortugas de mar.

Compás
de
inclinación
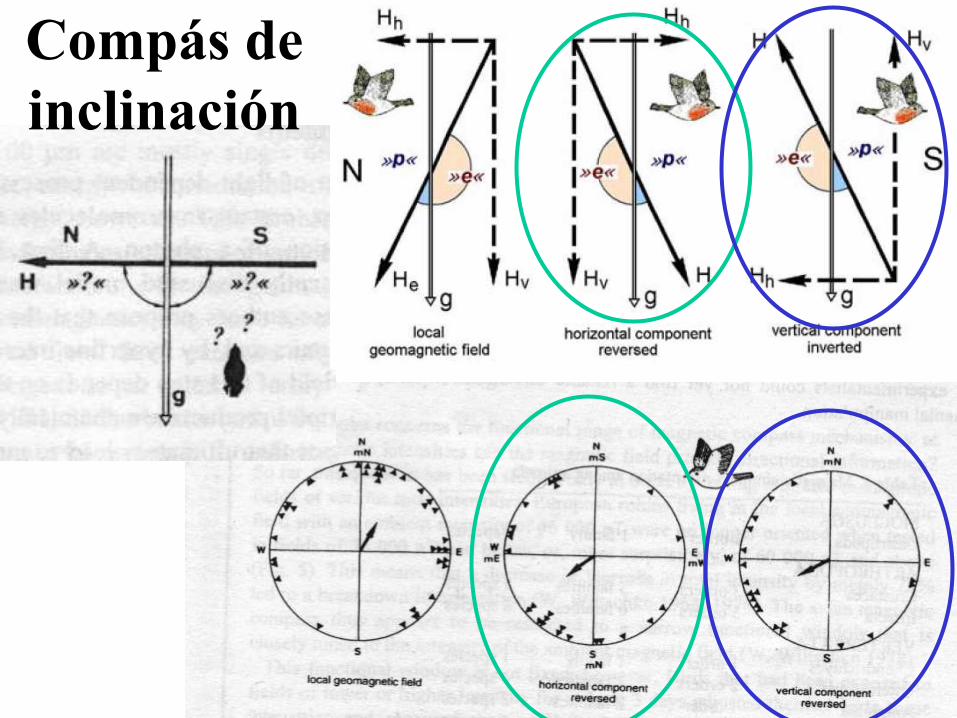
Compás de
inclinación

El compás de polaridad
• Evidencia experimental sugiere un segundo tipo de compás
magnético en peces salmónidos, anfibios y roedores. Estos animales
no cambian su orientación cuando el componente vertical del
campo magnético se invierte. Compás basado en la polaridad de las
líneas del campo magnético (como brújula).
• El modo funcional del compás magnético en invertebrados ha sido
analizado sólo en dos especies. Langostas marinas y escarabajos de
la línea en Tenebrio molitor, ambos casos sugiere compás basado en
la polaridad de las líneas del campo magnético. No se conoce cuán
ampliamente distribuido puede estar el compás de polaridad dentro
de los artrópodos.

Compás de
polaridad

Interacción entre información del
compás celeste y magnético

Rango de Intensidad
¿A qué intensidades magnéticas
puede el campo magnético
proveer información direccional?

Rango de Intensidad • Hasta el momento sólo se probó aves migrando a distintas intensidades
totales de campos magnéticos.
• Petirrojos europeos viviendo en campos geomagnéticos locales
(intensidad ambiental 46000 nT) no se orientaban en campo de 34000 nT
o menos y sorprendentemente a 60000 nT o más.
• Compás restringido a ventana funcional muy angosta. Sintonizada a
intensidad campo magnético ambiente (solo 25% flexible).
• Ventana no es fija. Aves expuestas ≠ intensidades de campos magnéticos
(3 o más días) vuelven a poseer la habilidad de orientarse. Incluso a
intensidades de 16000 o 150000 nT.
• Proceso adaptación no es simple corrimiento. Aves adaptadas no
perdieron habilidad de orientarse en el campo geomagnético local pero si
a intensidades intermedias.
• No se conoce en otros animales. ¿Ventana funcional limitada
característica del compás de inclinación o también del de polaridad?

Rango de Intensidad

Mecanismos hipotéticos de
receptores magnéticos • Inducción electromagnética
– Kalmjim (Scripps). Rayas y tiburones. Mar muy conductor, pez se mueve (excepto paralela a líneas magnéticas) induce corriente. Órgano de Lorenzini puede detectar pequeños cambios en voltaje asociado con corriente inducida.
• Magneto recepción química
– Schulten (U. Illinois) influencia del campo magnético externo en el spin de electrón cuando pasa de molécula dador a aceptor (fotoreceptores).
• Receptores basados en magnetita (Fe3O4)
– Se encontró en bacteria, trucha, etc

Posibles procesos de transducción
• Una amplia variedad de mecanismos han sido
propuestos como procesos de transducción. Hoy
la discusión se focaliza fundamentalmente en dos
hipótesis: (1) magneto recepción mediada por
estado excitado de macromoléculas, en particular
foto pigmentos, y (2) magneto recepción basada
en materia ferro magnética como cristales de
magnetita. Otros, híbrido de los dos mencionados.
• Magneto recepción por fotopigmentos
• Mecano recepción basada en magnetita

Mecanismo dependiente de luz.
Proponen que la exitación de
fotopigmentos de la retina pobla estados
tripletes cuya estabilidad depende del
campo magnético. Sería necesario un
receptor capaz de discriminar entre
pigmentos en estado triplete de singlete.
Magneto recepción por fotopigmentos
A radical-pair compass seems
implausible: the energetic
interaction of the geomagnetic
field (25–65 μT) with a single
molecule is more than a million
times smaller than the molecule’s
thermal energy, kBT, under
physiological conditions

Magneto recepción por fotopigmentos
• Para mediar información magnética por este mecanismo un arreglo
ordenado de macromoléculas orientadas en diferentes direcciones
espaciales es requerido, ej. fotopigmentos ojo de vertebrados.
• Diseñaron modelos de magnetorecepción con características
funcionales del compás magnético. Cambios en intensidad alterarían
el patrón de la retina sin afectar su simetría.
• Habilidad para explicar características importantes de respuestas de
aves bajo diferentes condiciones magnéticas hicieron a este modelo
atractivo. El compás de polaridad (peces, roedores y dos especies
artrópodos), parece incompatible con la respuesta axial del mecanismo
basado en el fotoreceptor .




El petirrojo europeo (Erithacus rubecula)
Magnetorecepción:
Efecto de longitud de onda

Proponen que la magnetita actúa como receptor, siguiendo las líneas de
campo magnético. Esta rotación en la magnetita podría ser detectada por
distintos mecanismos (cambio de conductancia de membrana, canales
iónicos acoplados a la partícula, partículas acopladas a células pilosas
con mecanorreceptores, etc).
Magneto recepción basada en magnetita
• Material biológico permanentemente
magnético recién 1962 descripto en diente de
rádula de quitones. Mediciones de remanencia
y espectrometría, identificaron cristales
magnéticos en tejidos de amplia variedad de
animales (especulaciones acerca del posible
rol en magneto recepción).

Magneto recepción basada en magnetita
• Propiedades magnéticas partículas de magnetita dependen de su
tamaño. La energía magnética de orientación a menudo justo
sobrepasa de la energía térmica del fondo. Este problema puede
solucionarse por un arreglo ordenado de partículas en cadenas o
en agrupamientos donde sus momentos son sumados lo cual
aumenta su interacción con el campo geomagnético.
• Material magnético permanente responde a polaridad, por lo
tanto debieran indicar la dirección polar del campo magnético.
• Consideraciones teóricas también muestran que mecanismos
basados en magnética no sólo serían capaces de mediar de
indicar la dirección del campo magnético sino también serían
suficientemente sensibles para detectar pequeños cambios en
intensidad siempre cuando un número suficiente de partículas de
magnéticas estén involucradas.

Mecanismos de magnetorecepción:
Evidencia experimental • Procesos dependiente de luz del modelo fotopigmento, fácil testear
hipótesis: magnetorecepción imposible en ausencia de luz.
• Palomas volviendo a casa y salamandras, información del compás
magnético interrumpida en oscuridad total. Mecanismo de compás
de aves y anfibios basado en proceso dependiente de luz.
• Salmones, tortugas marinas y roedores capaces dirección en
oscuridad total; compás magnético basado en mecanismo
independiente de la luz.
• Llamativo que no hay correlación entre dependencia de luz y la
forma funcional del compás (polaridad o inclinación). Todas las
posibles combinaciones se encuentran dentro los vertebrados.
• Dentro de los artrópodos, las langostas de mar, abejas y escarabajos
no necesitan luz para orientación. Sugiere mecanismo de
magnetorecepción independiente de luz (moscas de la fruta no).

Mecanismos de magnetorecepción:
Evidencia experimental

El petirrojo europeo (Erithacus rubecula)

S Engels et al. (2014) Nature
Magnetic compass orientation of migratory European robins
tested at the University of Oldenburg requires aluminium screening.


Connecting and disconnecting the grounding of the screens turns
on and off the birds’ magnetic compass orientation capabilities.
S Engels et al. (2014) Nature

Artificially produced broadband electromagnetic noise disrupts
the magnetic compass orientation of birds tested inside the
grounded aluminium-screened huts.
S Engels et al. (2014) Nature

The disruptive effect of broadband electromagnetic noise on
magnetic compass orientation is not limited to a single narrow
frequency range.
S Engels et al. (2014) Nature
d, e: very low amplitud
f, g: green trace ca. 20–450 kHZ

In a rural location, European robins show magnetic compass
orientation without screening.
S Engels et al. (2014) Nature
University campus Rural area

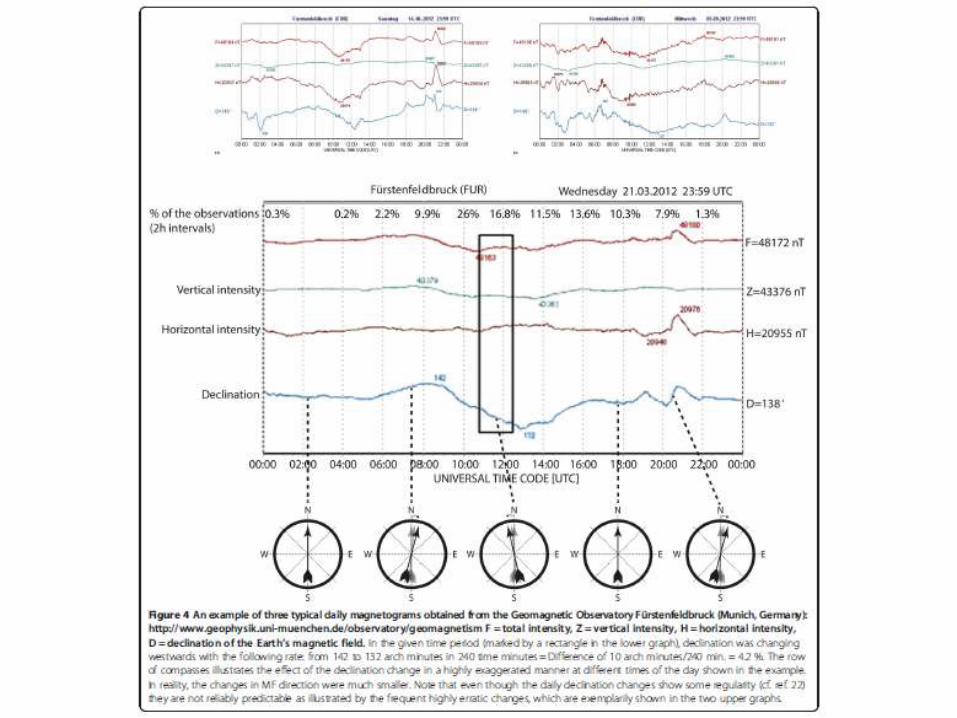
Interrupción de orientación magnética
• Con ruido electromagnético entre 20 y 5000 kHz
(≈ banda usada por radio AM).
• Con claves celestes presentes (sol o estrellas)
• Cuando la intensidad del background cambia
entre 20-30% con respecto a valores normales
• Tormentas geomagnéticas
• Algunos animales con presencia sólo de luz roja
(amanecer y anochecer) cuando el compas solar
más confiable

Salmónidos
• Famoso por habilidad de regresar a sus arroyos
natales a desovar luego de años. Imprinting de
claves olfatorias (Arthur Davis Hasler).
• Mecanismos para navegación en mar abierto.
Uso de compas solar e información magnética
• Apoyado por datos comportamentales
• Correlato estructural. Células con magnetita en
epitelio olfatorio conectadas vía nervio trigémino
• Registros electrofisiológicos nervio a cambio
campo magnético

Prof. Michael Walker
Univ. Auckland
Trucha arco iris (Oncorhynchus mykiss)
Entrenadas a discriminar una anomalía magnetica (Helmholtz coil)

Células en el epitelio olfatorio con cristales ricos en hierro (similar
a cadenas de magnetita de bacterias) inervadas por rama oftálmica
superficial del nervio trigémino

Navegación por campo magnético
en la langosta Panulirus argus

Tortugas marinas
• Al eclosionar utilizan 3 claves para orientarse y
adquirir preferencia de orientación magnética:
• Caminar hacia zona más baja y clara (luz; en
general no pueden ver el mar directamente)
• Nadar hacia la luz
• Mantener el curso con respecto a la dirección
de las olas

Tortugas marinas
• Orientación en tierra:
• Experimento: Eclosión en camino 4.1m con
luz tenue al final. Luego transfieren arena
experimental con agua en oscuridad, nadan
misma dirección que luz tenue.
• Cambio de campo magnético causa cambio en
dirección de nado.

Tortugas marinas

¿Cómo determinan la
dirección de
las olas?
Tortugas marinas
Pico de aceleración orbital ola 1m =
72cm/seg2 y detectada por oído interno
de vertebrado 5 cm/seg2

Tortugas marinas





Sensibilidad de los perros a pequeñas
variaciones del campo magnético
• Midiendo alineación eje cuerpo de 70 perros de 37 razas
durante defecación (n=1893) y orina (n=5582)
• Perros prefieren defecar con el eje del cuerpo alineado al
eje Norte-Sur bajo condiciones de campo magnético (CM)
estable (sólo 20% día)
• Primer demostración sensibilidad perros al CM
• Primer reacción comportamental predecible a las
fluctuaciones naturales del CM en un mamífero
• Se demostró una alta sensibilidad a pequeños cambios en
la polaridad en lugar de la intensidad como
biológicamente relevantes

Recommended

















