Embed Size (px)
Citation preview

CITOESQUELETODR. BRADY BELTRAN GARATE

CONCEPTOS Citoplasma: contenido de una célula que está situado dentro de su
membrana plasmática pero, en el caso de las células eucariontes,fuera del núcleo.
Citosol: contenido del principal compartimiento del citoplasma, conexclusión de los orgánulos limitados por membrana. Fracción celularque queda después de eliminación de las membranas, loscomponentes citoesqueléticos y otros orgánulos.
Citoesqueleto: Sistema de filamentos proteicos del citoplasma deuna célula eucarionte que le da su forma y la capacidad demovimiento dirigido.

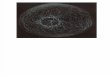
CITOESQUELETO Red compleja de filamentos proteicos que se extiende por todo el
citoplasma.
Estructura muy dinámica que se reorganiza de manera continua a medidaque la célula cambia de forma, se divide y responde a su ambiente
Responsable directo de diversos movimientos a gran escala (deslizamientosobre una superficie, contracción muscular, cambios de forma celulardurante el desarrollo embrionario, segregación de cromosomas, etc)
Controla la localización de los orgánulos, además de suministrar lamaquinaria para el transporte entre ellos
Tres filamentos proteicos: Filamentos Intermedios, Microtúbulos yFilamentos de Actina; cada uno con diferentes propiedades mecánicas yformados por una subunidad proteica distinta.


FILAMENTOS INTERMEDIOS Gran resistencia a la tensión.
Permiten que las células toleren las fuerzas mecánicas asociadas conel estiramiento
Son los más resistentes y estables
Forman una red por todo el citoplasma que rodea al núcleo y seextiende hacia la periferia celular. Suelen estar anclados a lamembrana en el sitio de uniones intercelulares como losdesmosomas; lo que establece una conexión mecánica continuaentre las células en todo el tejido
También forman la lámina nuclear

Red sólida y resistenteCuerdas formadas por hebras largasy retorcidas de proteínas fibrosascon una cabeza globular N-terminal,una cola globular C-terminal y undominio bastoniforme alargadocentral
Los dominios bastoniformes sonsimilares en tamaño y secuencia deaminoácidos, cuando se agrupansiempre forman filamentos de undiámetro y una estructura internasimilares.Las cabezas y las colas globularesvarían mucho y al encontrarseexpuestas en la superficie delfilamento interactúan con otroscomponentes del citoplasma.

FILAMENTOS INTERMEDIOS Son muy abundantes en los axones de las células nerviosas, células
musculares y epiteliales.
Se agrupan en 4 clases:
Filamentos de Queratina en las células epiteliales
Filamentos de Vimentina y relacionados (Células del tejido conectivo,muscular y glíal)
Neurofilamentos de las células nerviosas
Láminas nucleares

EPIDERMÓLISIS AMPOLLAR SIMPLE Mutaciones en los genes de la queratina interfieren con la formación
de filamentos de queratina en la epidermis piel muy vulnerable alas lesiones mecánicas

FILAMENTOS INTERMEDIOS Muchos de los filamentos intermedios son estabilizados y reforzados
aun más por proteínas accesorias (Plectina) que forman unionescruzadas entre los haces de filamentos formando una estructuraresistente.
Además de mantener juntos los haces de filamentos intermediosestas proteínas unen los filamentos intermedios a los Microtúbulos,a los filamentos de actina y a estructuras adherentes de losdesmosomas.

FILAMENTOS INTERMEDIOSLámina Nuclear:
Proteínas llamadas laminas, queforman una red bidimensional.
Se desensamblan y se vuelven aformar en cada división celular.
Esto es controlado porfosforilación y desfosforilaciónde las laminas porproteincinasas.

PROGERIA Laminopatía asociada a mutaciones
en el gen LMNA.
Disrupción del ensamblaje normal dela envoltura nuclear, la funciónnuclear y la lámina nuclear. Efectoprofundo en la integridad del tejidoconectivo.
Crítico para el soporte nuclear y parala organización de la cromatina.
Núcleos con alteracionesestructurales como herniaciones ylóbulos. Molecularmente presentanun defecto en el mecanismo dereparación del ADN

MICROTÚBULOS Tubos proteicos huecos, largos y relativamente rígidos,
que tienen la capacidad de desensamblarse con rapidezen un sitio y ensamblarse de nuevo en otro. Seoriginan en el centrosoma
Se extienden hacia la periferia celular formando unsistema de guías intracelulares a lo largo de las cualesse desplazan vesículas, orgánulos y otros componentescelulares.
En la mitosis los Microtúbulos se desensamblan y luegose reensamblan en el huso mitótico, que aporta lamaquinaria que permitirá la segregación equitativa delos cromosomas
También pueden formar estructuras permanentes:cilios y flagelos
La función organizativa de los Microtúbulos dependede su asociación con proteínas accesorias sobre todomotoras que propulsan orgánulos a lo largo de suscarriles citoesqueléticos

MICROTÚBULOS Subunidades: moléculas de tubulina. Dímero α y β tubulina estrechamente unidas por
enlaces no covalentes.
Los dímeros de tubulina se apilan para formar la pared del microtúbulo cilíndricohueco: 13 protofilamentos paralelos
Polaridad estructural (β tubulina extremo + y el opuesto (α) negativo)
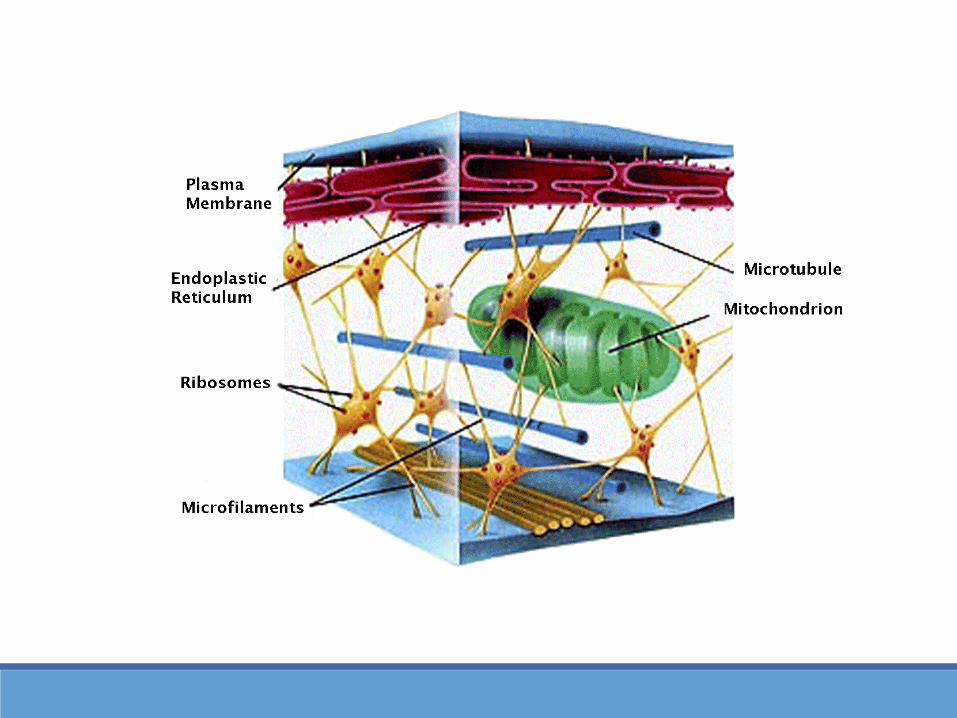
MICROTÚBULOS: CENTROSOMA Centrosoma principal centro
organizador de Microtúbulos en lascélulas animales; controla número,localización y su orientación en elcitoplasma.
Contienen estructuras anularesformadas por tubulina y cada anilloes el punto de partida o sitio denucleación para el crecimiento delmicrotúbulo.
Contiene un par de centriolos(constituidos por Microtúbulos cortosde disposición cilíndrica) funciónexacta no conocida
Cuerpos basales: centrosorganizadores de Microtúbulos en loscilios y flagelos

MICROTÚBULOS: INESTABILIDAD DINÁMICA
Los Microtúbulos en crecimiento presentan inestabilidad dinámica:crecen y se retraen por adición o pérdida de subunidades αβ, debidoa la capacidad intrínseca de las moléculas de tubulina de hidrolizarGTP.
Cada dímero de tubulina libre contiene una molécula de GTPestrechamente unida que es hidrolizada a GDP poco después de laadición de la subunidad a un microtúbulo en crecimiento.
Las moléculas de tubulina asociadas a GTP se agrupan de maneraeficiente, mientras que las moléculas de tubulina portadoras de GDPtienen una conformación distinta y se unen más laxamente entre sí


MICROTÚBULOS: INESTABILIDAD DINÁMICA
La inestabilidad relativa de los Microtúbulos les permite presentarun remodelado rápido, característica crucial para su función.
Es posible evitar su desensamblaje si su extremo + es estabilizado demanera permanente por unión a otra molécula o estructura celular.
Fármacos que impiden la polimerización o la despolimerización de latubulina: Colchicina, taxol, vincristina, vinblastina
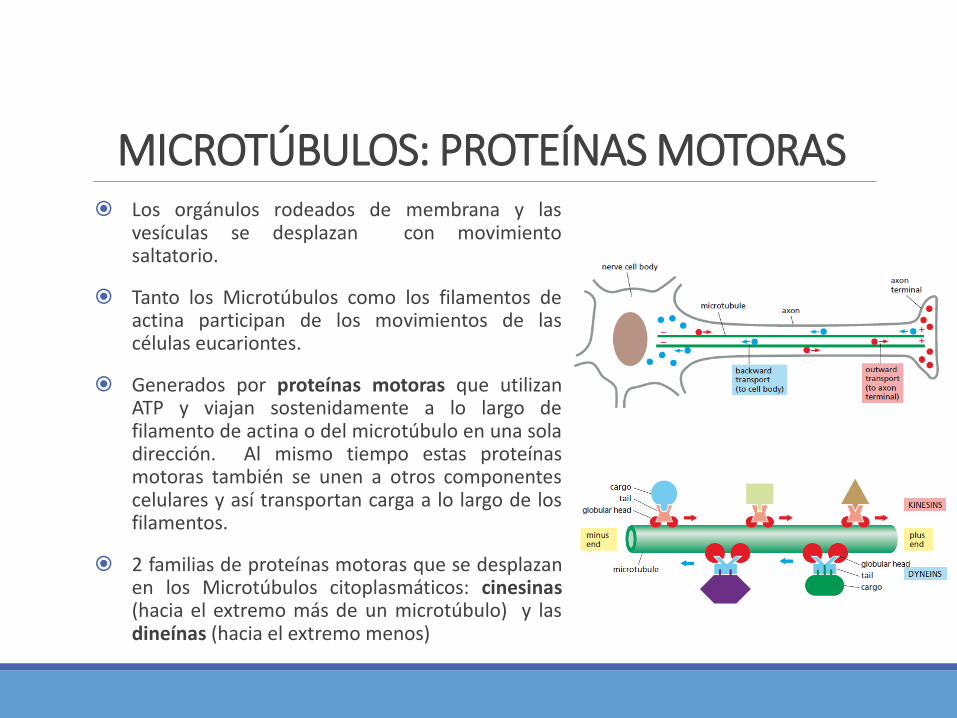
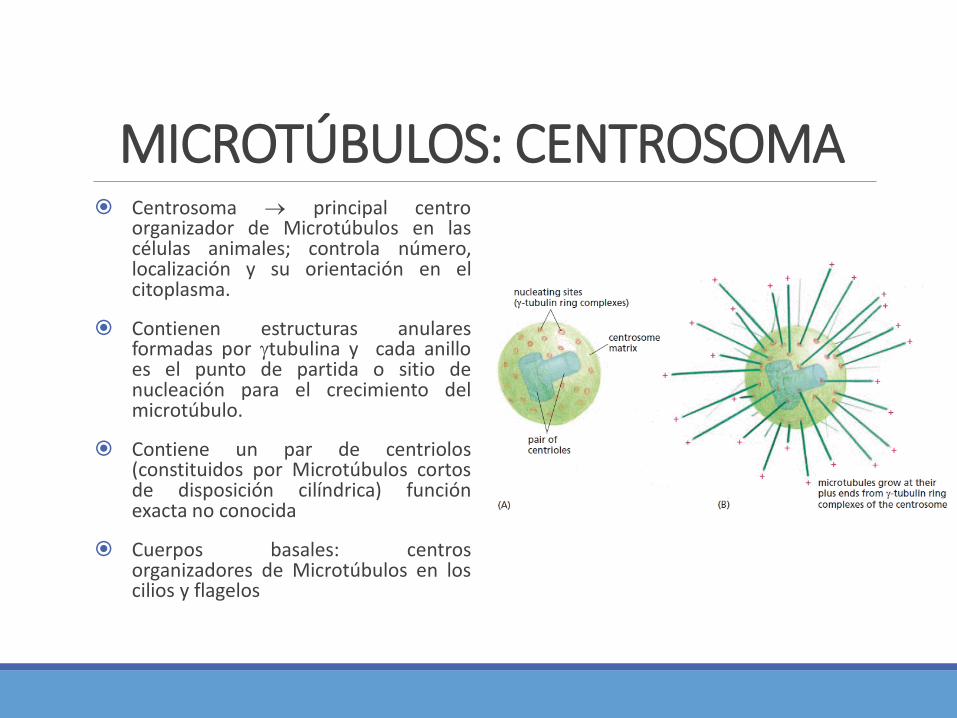
MICROTÚBULOS: PROTEÍNAS MOTORAS Los orgánulos rodeados de membrana y las
vesículas se desplazan con movimientosaltatorio.
Tanto los Microtúbulos como los filamentos deactina participan de los movimientos de lascélulas eucariontes.
Generados por proteínas motoras que utilizanATP y viajan sostenidamente a lo largo defilamento de actina o del microtúbulo en una soladirección. Al mismo tiempo estas proteínasmotoras también se unen a otros componentescelulares y así transportan carga a lo largo de losfilamentos.
2 familias de proteínas motoras que se desplazanen los Microtúbulos citoplasmáticos: cinesinas(hacia el extremo más de un microtúbulo) y lasdineínas (hacia el extremo menos)

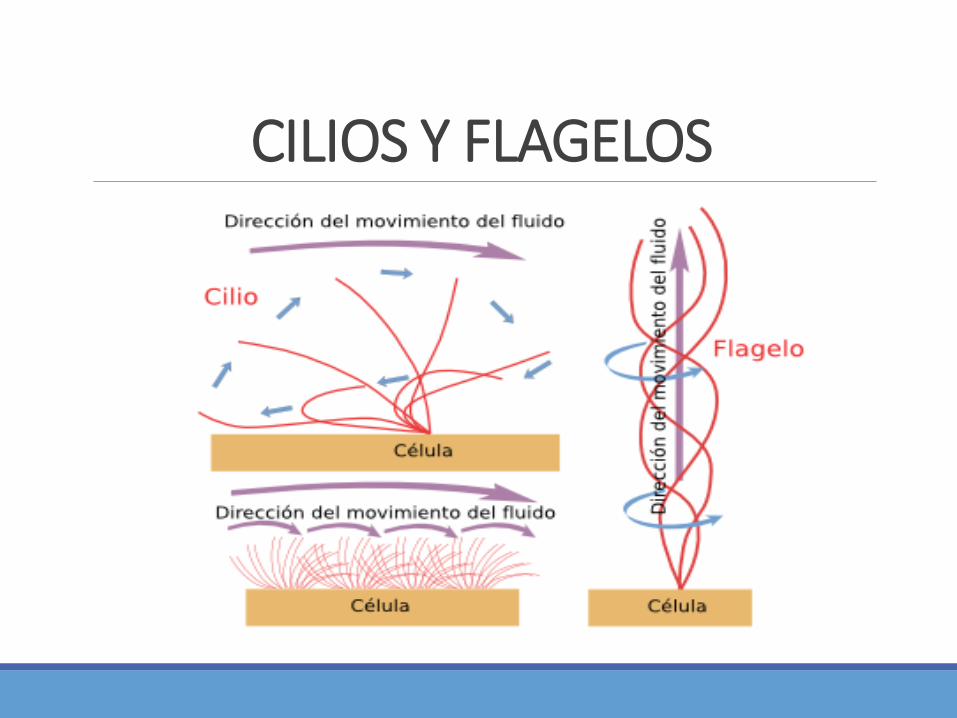
MICROTÚBULOS: CILIOS Y FLAGELOS Los cilios y flagelos están formados por microtúbulos estables movidos por la dineína ciliar. Disposición
“9+2”
Cilios: estructuras piliformes 0.25 um de diámetro, cubiertas de membrana plasmática, que parten de lasuperficie de varios tipos de células eucariontes, contienen una porción central formada por un haz demicrotúbulos estables que crecen a partir de un cuerpo basal localizado en el citoplasma, el cuerpo basalactúa como centro organizador del cilio.
Los flagelos que impulsan a los espermatozoides y a muchos protozoos presentan una estructura internasimilar a la de los cilios pero son mucho más largos. Los flagelos desplazan a toda la célula a través de unmedio líquido

CILIOS Y FLAGELOS
ESQUEMA DE UN CILIO (CORTE TRANSVERSAL)
CILIOS Y FLAGELOS

CILIOS Y FLAGELOS
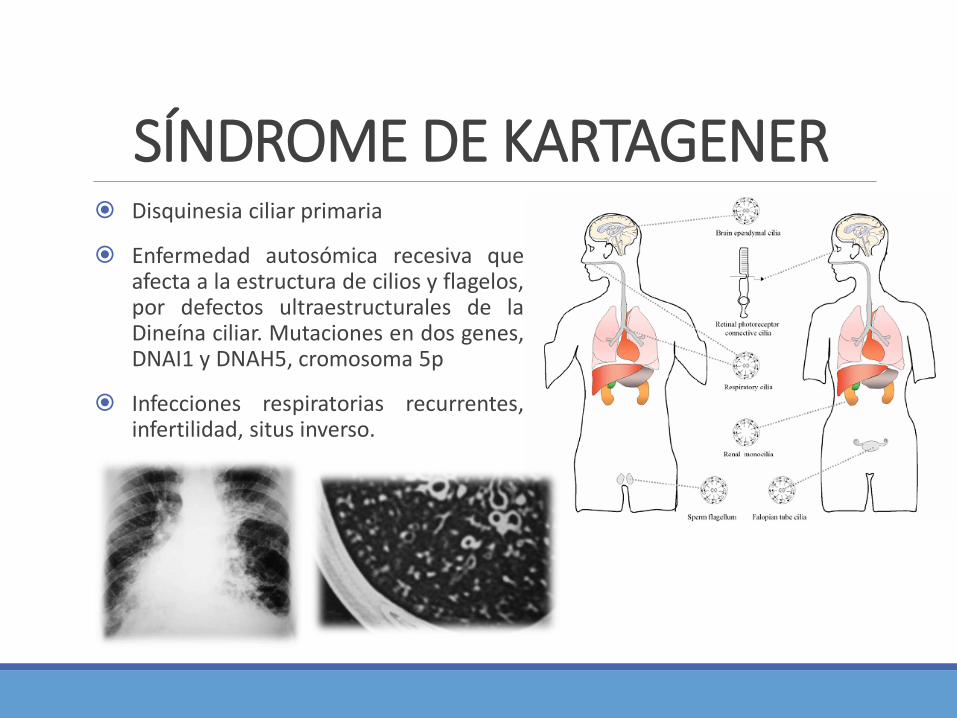
Disquinesia ciliar primaria
Enfermedad autosómica recesiva queafecta a la estructura de cilios y flagelos,por defectos ultraestructurales de laDineína ciliar. Mutaciones en dos genes,DNAI1 y DNAH5, cromosoma 5p
Infecciones respiratorias recurrentes,infertilidad, situs inverso.
SÍNDROME DE KARTAGENER

FILAMENTOS DE ACTINA Esenciales para movimientos, sobre todo los relacionados con la
superficie celular.
Al igual que los microtúbulos muchos filamentos de actina soninestables pero también pueden formar estructuras celularesestables, como el aparato contráctil del músculo.
Se asocian con proteínas fijadoras de actina, que posibilitan que losfilamentos cumplan diversas funciones celulares.

FILAMENTOS DE ACTINA Más delgados, más flexibles y más
cortos que los microtúbulos. 7nmde diámetro. Mucho másnumerosos.
Haces con uniones cruzadas oredes cuya resistencia es muchomayor que la de los filamentosaislados.
Cada filamento es una cadenaretorcida de moléculas globularesde actina idénticas, que apuntanen la misma dirección a lo largodel eje de la cadena. Polaridadestructural con un extremo+ y unextremo -

FILAMENTOS DE ACTINA

FILAMENTOS DE ACTINA La actina y la tubulina se
polimerizan por mecanismossimilares, en este caso usa ATP

FILAMENTOS DE ACTINA La mitad de la actina está ensamblada en filamentos y la otra mitad
se halla en forma de monómeros en el citosol.
Timosina y profilina se unen a estos monómeros lo que les impideagregarse a los extremos de los filamentos de actina
Forminas y las proteínas relacionada con actina promueven suensamblaje
Asimismo existen proteínas fijadoras unidas a los filamentosensamblados, controlando su comportamiento.
Mantener haces paralelos en las microvellosidades, formarenlaces cruzados, propiedad de gel, fragmentadoras (gelsolina),proteínas motoras
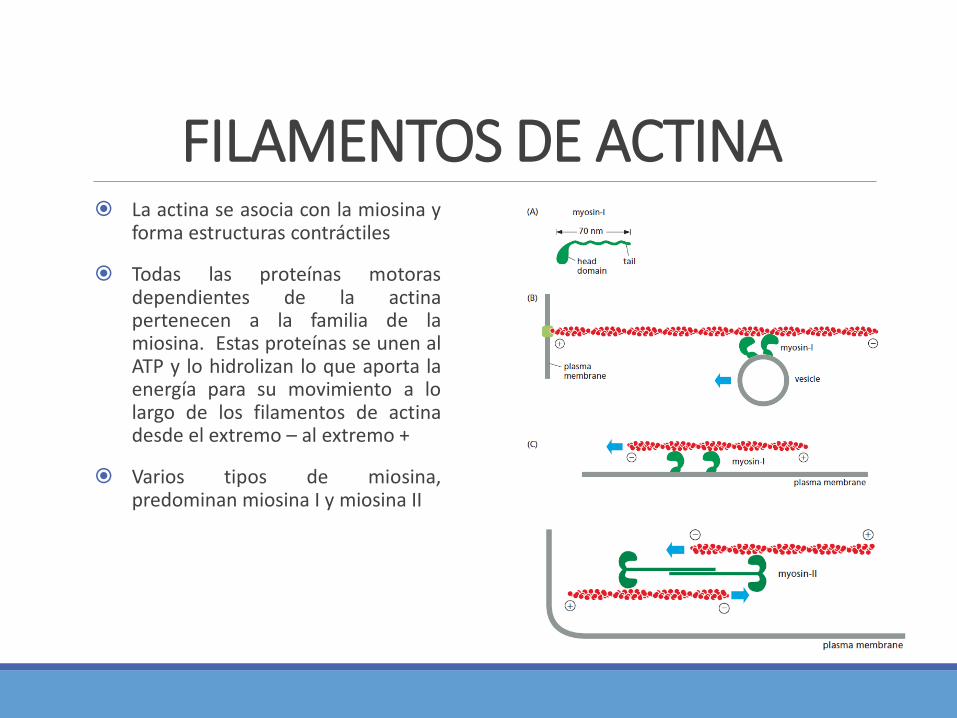
FILAMENTOS DE ACTINA La actina se asocia con la miosina y
forma estructuras contráctiles
Todas las proteínas motorasdependientes de la actinapertenecen a la familia de lamiosina. Estas proteínas se unen alATP y lo hidrolizan lo que aporta laenergía para su movimiento a lolargo de los filamentos de actinadesde el extremo – al extremo +
Varios tipos de miosina,predominan miosina I y miosina II

FILAMENTOS DE ACTINA Señales extracelulares controlan la disposición de los filamentos de
actina, lo que permite que la célula reorganice su citoesqueleto enrespuesta al ambiente.
En el caso del citoesqueleto de actina estos reordenamientosestructurales son desencadenados por la activación de una variedadde proteínas receptoras incluidas en la membrana plasmática,después estas señales convergen en el interior de la célula sobre ungrupo de proteínas de unión a GTP: familia de proteínas Rho.

FILAMENTOS DE ACTINA: CONTRACCIÓN MUSCULAR
La miosina del músculo pertenece a la subfamilia miosina II
Deslizamiento de grupos de filamentos de actina de orientaciónopuesta entre sí.
Al agruparse los filamentos de actina y de miosina forman unfascículo, el haz genera una fuerza contráctil

FILAMENTOS DE ACTINA: CONTRACCIÓN MUSCULAR
Unidades contráctiles: sarcómeros (ensamblajes altamente organizados de filamentos de actina yde miosina II)
Contracción: acortamiento simultáneo de todos los sarcómeros provocado por el deslizamiento delos filamentos de actina sobre los de miosina sin modificar la longitud de ninguno de ellos
Extensión completa 3um // Contracción completa 2um

FILAMENTOS DE ACTINA: CONTRACCIÓN MUSCULAR
Luego del estímulo contráctil las cabezasde miosina comienzan a desplazarse a lolargo del filamento de actina en ciclosrepetidos de unión y separación
Durante cada ciclo una cabeza demiosina hidroliza un ATP. Estodetermina una serie de cambiosconformacionales de la molécula demiosina que desplazan el extremo de lacabeza alrededor de 5 nm a lo largo delfilamento de actina en la dirección delextremo más.
Este movimiento que se repite con cadaciclo de hidrólisis del ATP, propulsaunidireccionalmente la molécula demiosina a lo largo del filamento deactina
Al hacerlo la cabeza de miosinatracciona al filamento de actina y lohace deslizar contra el filamento demiosina. La acción coordinada demuchas cabezas de miosina provoca lacontracción del sarcómero
Completada la contracción el contactoentre las cabezas de miosina y losfilamentos de actina cesa por completoy el músculo se relaja
Cada cabeza de miosina se puede unir ala actina y separarse de ésta alrededorde 5 veces por segundo, lo que permiteque los filamentos de actina y demiosina se deslicen entre sí a unavelocidad de hasta 15um por segundo.

FILAMENTOS DE ACTINA: CONTRACCIÓN
MUSCULAR
1. Unión
2. Liberación
3. Enderezamiento
4. Generación de fuerza
5. Unión

FILAMENTOS DE ACTINA: CONTRACCIÓN MUSCULAR
La interacción molecular generadora de fuerza entre los filamentos de actina y miosina sólo tienelugar cuando el músculo esquelético recibe una señal proveniente del sistema nervioso.
Esta excitación eléctrica se propaga en milisegundos a lo largo de una serie de tubosmembranosos: túbulos transversos (o túbulos T).
Después la señal eléctrica se transmite al retículo sarcoplasmático: que rodea a cada miofibrillacomo una funda reticular y contiene una concentración elevada de Ca2+ y en respuesta a laexcitación eléctrica aferente gran parte de este calcio es liberado al citosol a través de canalesiónicos que se abren en la membrana del retículo sarcoplasmático como consecuencia del cambiodel voltaje a través de la membrana plasmática

FILAMENTOS DE ACTINA: CONTRACCIÓN MUSCULAR

FILAMENTOS DE ACTINA: CONTRACCIÓN MUSCULAR
En el músculo el Ca2+ interactúa con un interruptor molecular compuestopor proteínas accesorias especializadas estrechamente asociadas con losfilamentos de actina.
Tropomiosina: rígida y bastoniforme que se une al surco de la hélice deactina superponiéndose a siete monómeros, que impide que las cabezas demiosina se asocien con el filamento de actina
Troponina es un complejo que contiene una proteína sensible al Ca2+ quese asocia con el extremo de una molécula de tropomiosina. Cuandoaumenta el nivel citosólico de calcio este se une a la troponina e induce uncambio en su conformación, que determina a su vez que las moléculas detropomiosina modifiquen ligeramente su posición, lo que permite q lascabezas de miosina se unan al filamento de actina y desencadenen lacontracción