Embed Size (px)
Citation preview

Cytochrome C OxidaseOctober 15
Angela Sanchez
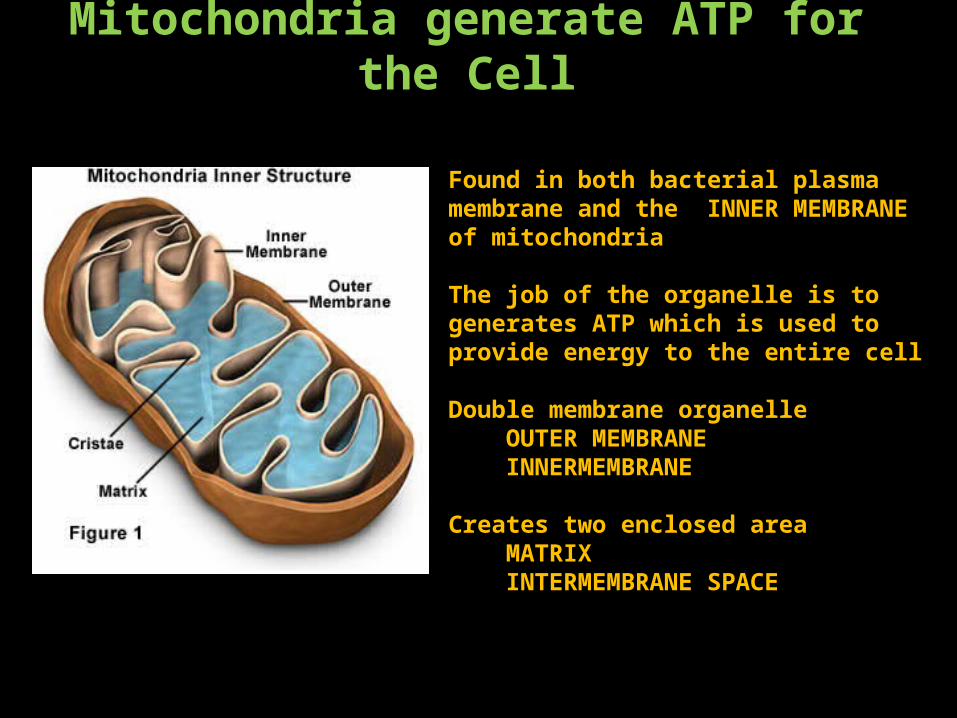
Mitochondria generate ATP for the Cell
Found in both bacterial plasma membrane and the INNER MEMBRANE of mitochondria
The job of the organelle is to generates ATP which is used to provide energy to the entire cell
Double membrane organelle OUTER MEMBRANE INNERMEMBRANE
Creates two enclosed area MATRIX INTERMEMBRANE SPACE
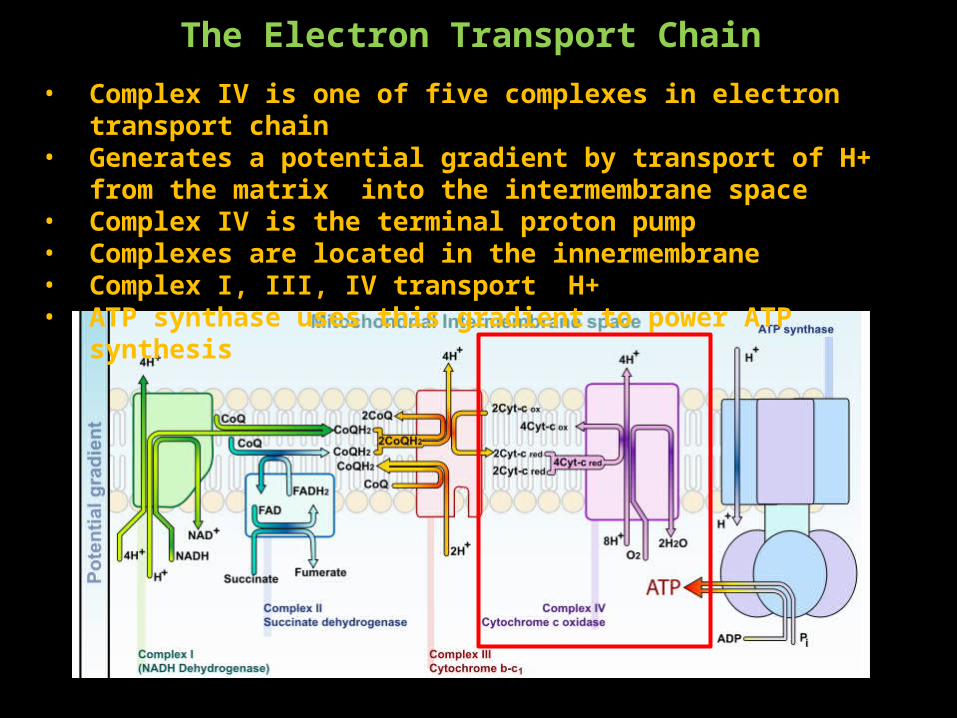
The Electron Transport Chain• Complex IV is one of five complexes in electron transport chain• Generates a potential gradient by transport of H+ from the matrix into the
intermembrane space• Complex IV is the terminal proton pump• Complexes are located in the innermembrane• Complex I, III, IV transport H+ • ATP synthase uses this gradient to power ATP synthesis
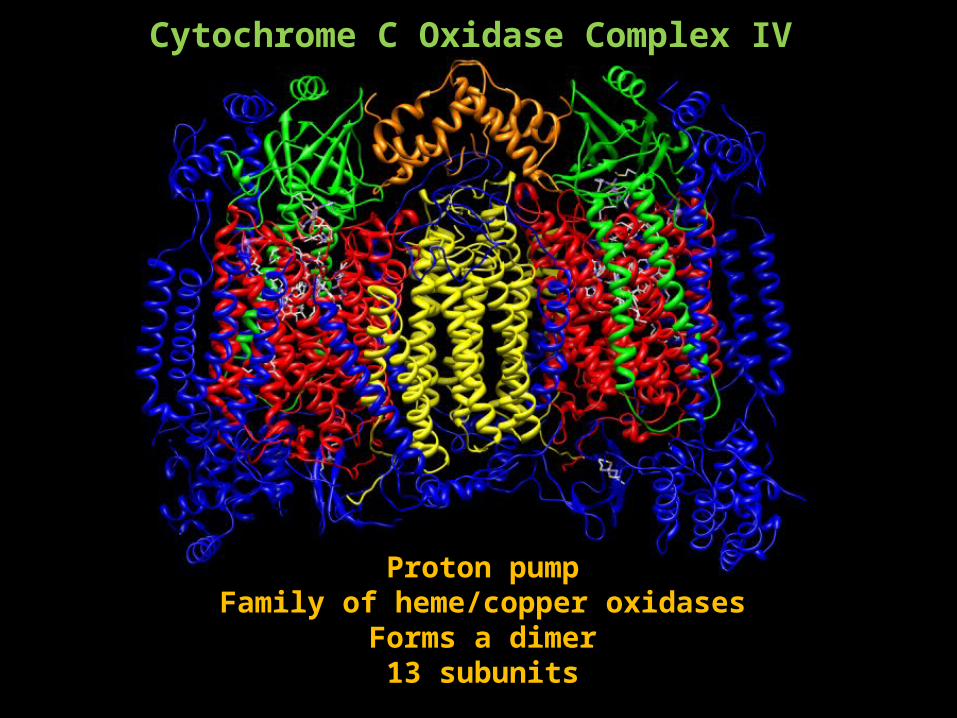
Cytochrome C Oxidase Complex IV
Proton pumpFamily of heme/copper oxidases
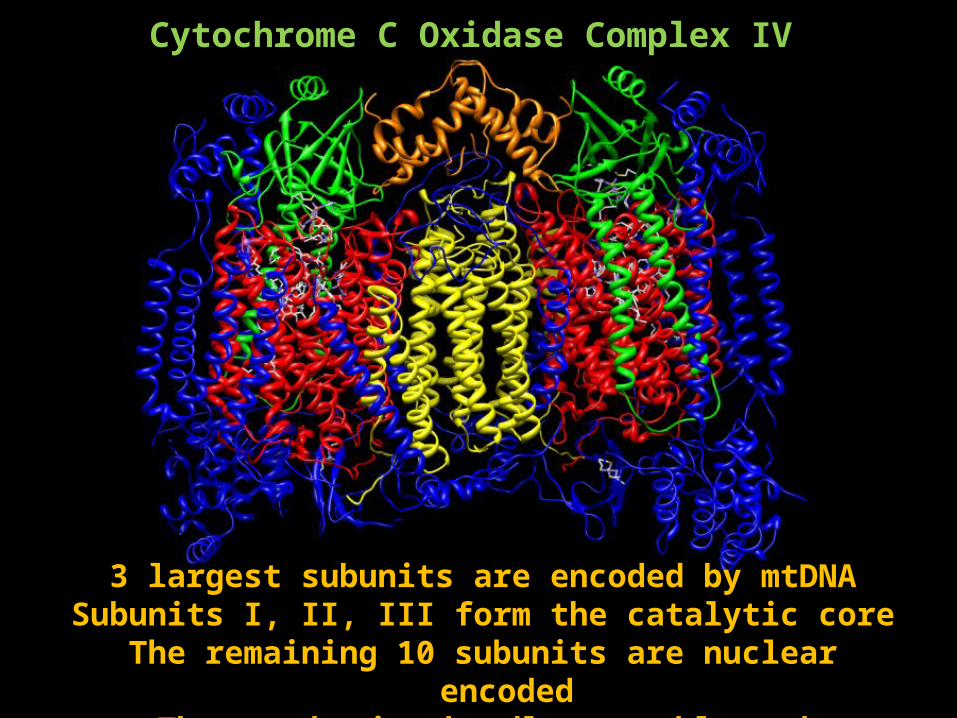
Forms a dimer13 subunits
Cytochrome C Oxidase Complex IV
3 largest subunits are encoded by mtDNASubunits I, II, III form the catalytic core
The remaining 10 subunits are nuclear encodedThese subunits handle assembly and regulation
Cytochrome C Oxidase Complex IV
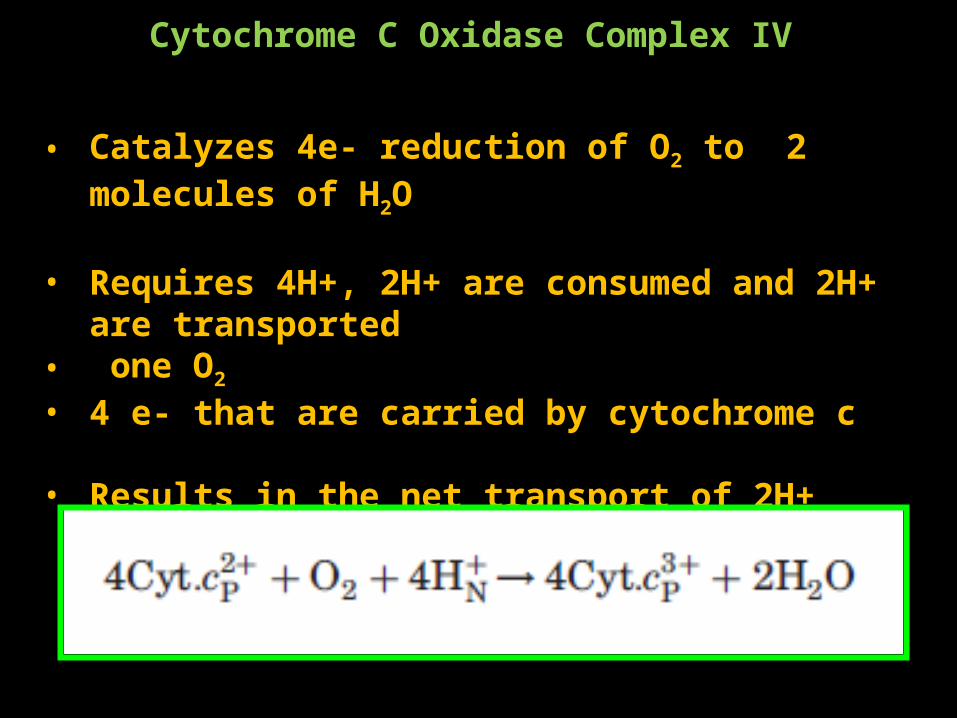
• Catalyzes 4e- reduction of O2 to 2 molecules of H2O
• Requires 4H+, 2H+ are consumed and 2H+ are transported• one O2
• 4 e- that are carried by cytochrome c
• Results in the net transport of 2H+
MAIN JOB IS TO MOVE PROTONS!!!
Cytochrome C Oxidase Reaction Cofactors
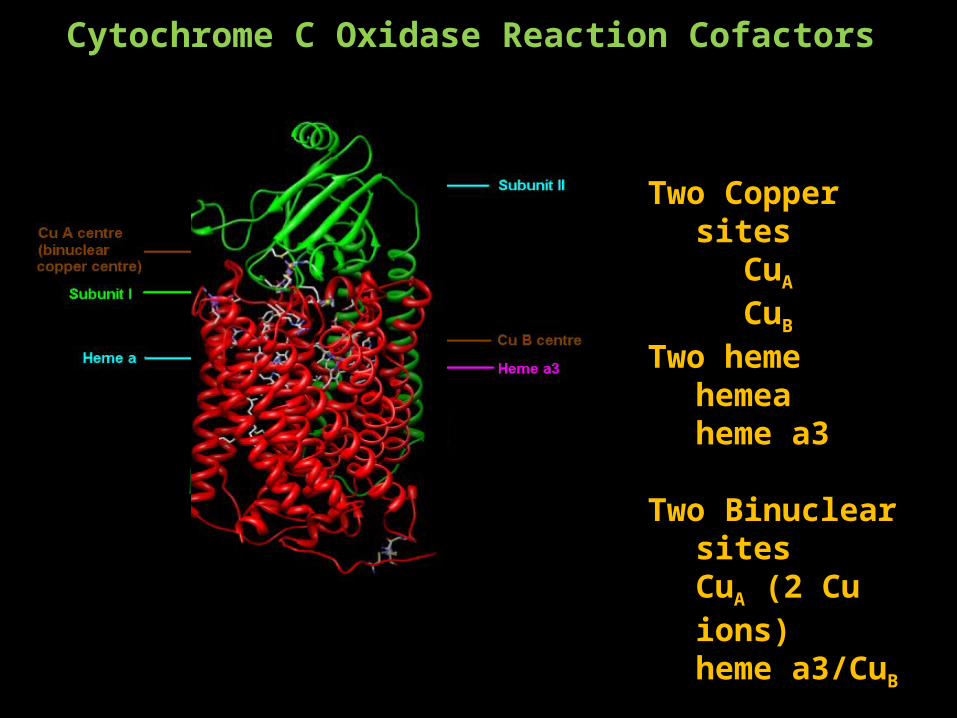
Two Copper sitesCuA
CuB
Two hemehemeaheme a3
Two Binuclear sitesCuA (2 Cu ions)heme a3/CuB
Cytochrome C Oxidase Complex IV Subunit II
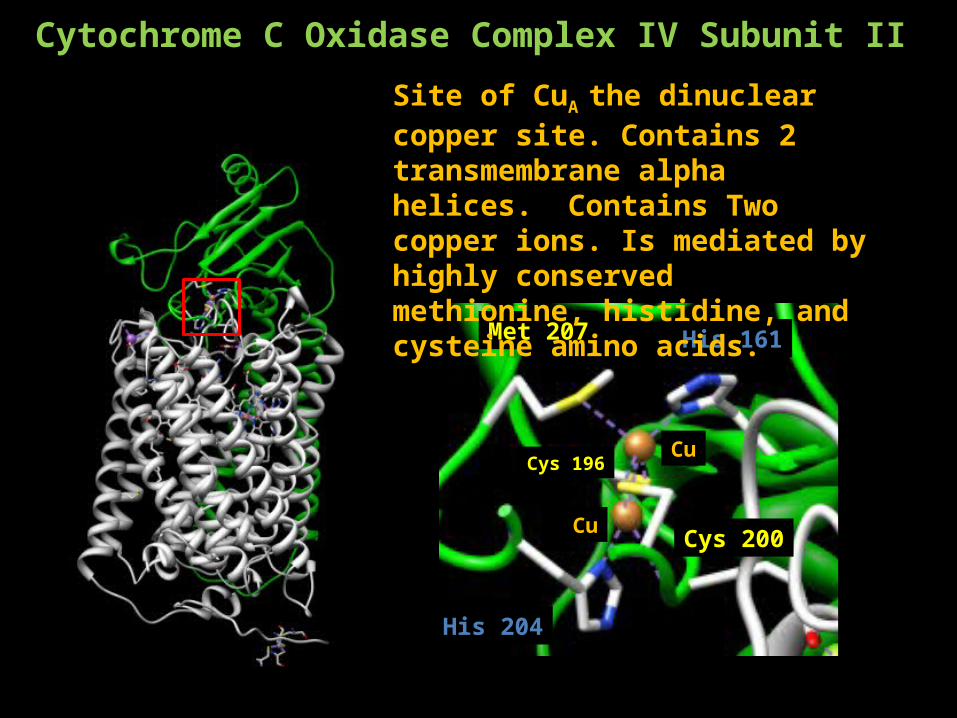
Met 207
Cys 200
Cys 196Cu
Cu
His 161
His 204
Site of CuA the dinuclear copper site. Contains 2 transmembrane alpha helices. Contains Two copper ions. Is mediated by highly conserved methionine, histidine, and cysteine amino acids.
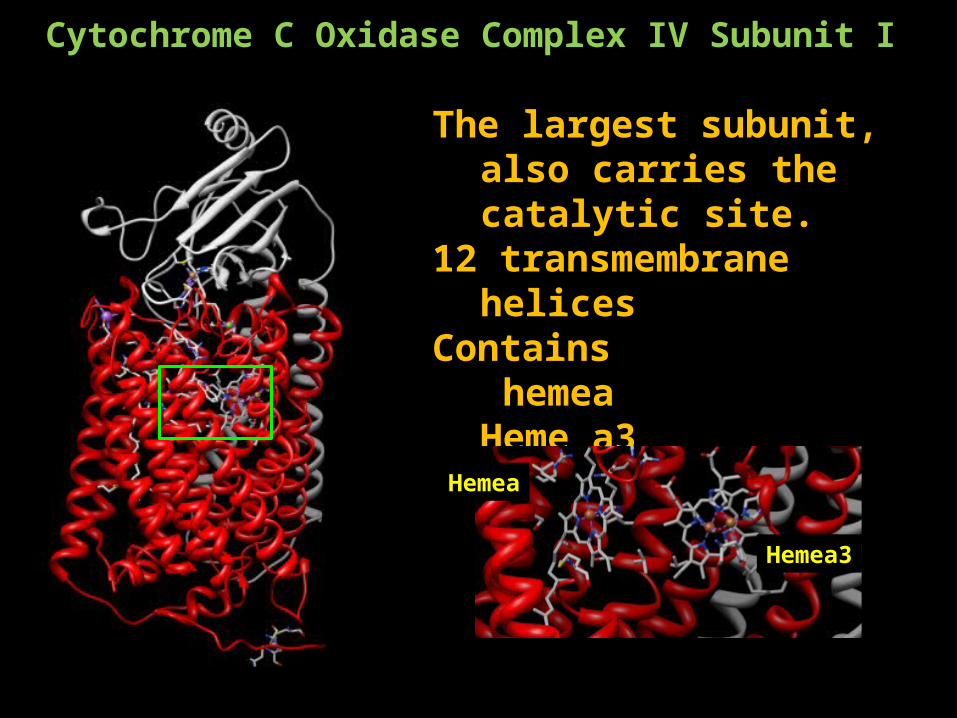
Cytochrome C Oxidase Complex IV Subunit I
The largest subunit, also carries the catalytic site.
12 transmembrane helices Contains
hemea Heme a3CuB
Hemea
Hemea3
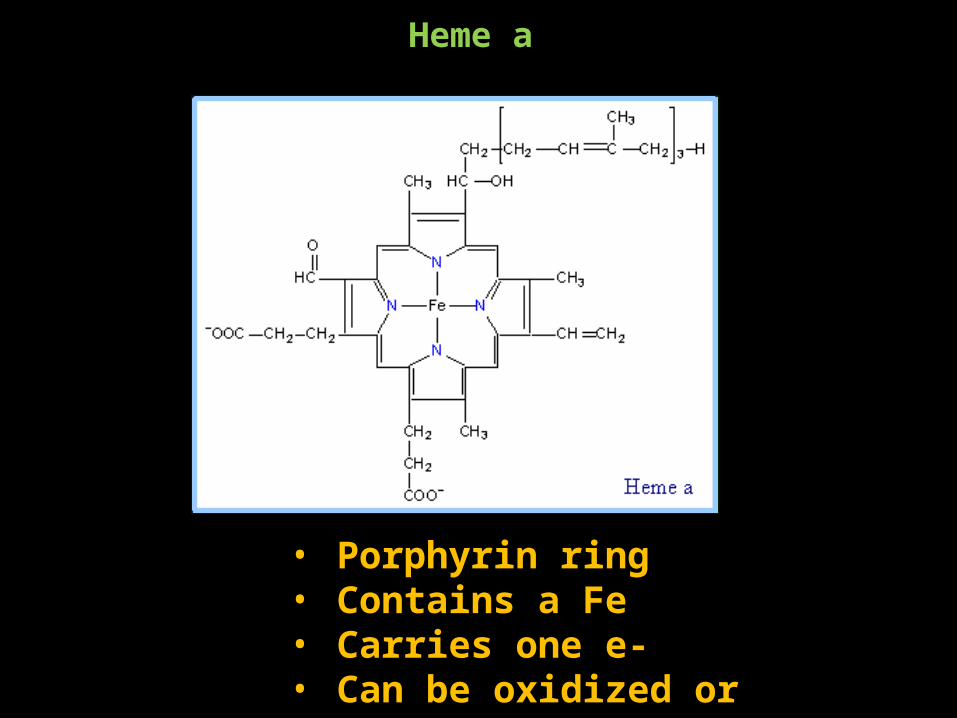
Heme a
• Porphyrin ring• Contains a Fe• Carries one e-• Can be oxidized or reduced
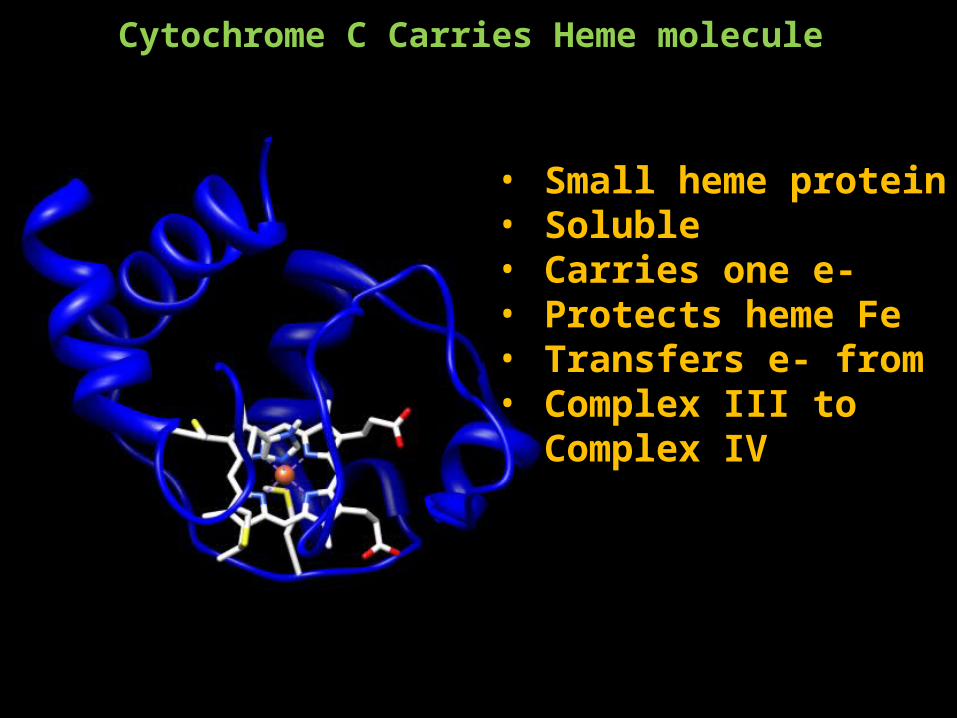
Cytochrome C Carries Heme molecule
• Small heme protein• Soluble• Carries one e-• Protects heme Fe• Transfers e- from • Complex III to Complex IV
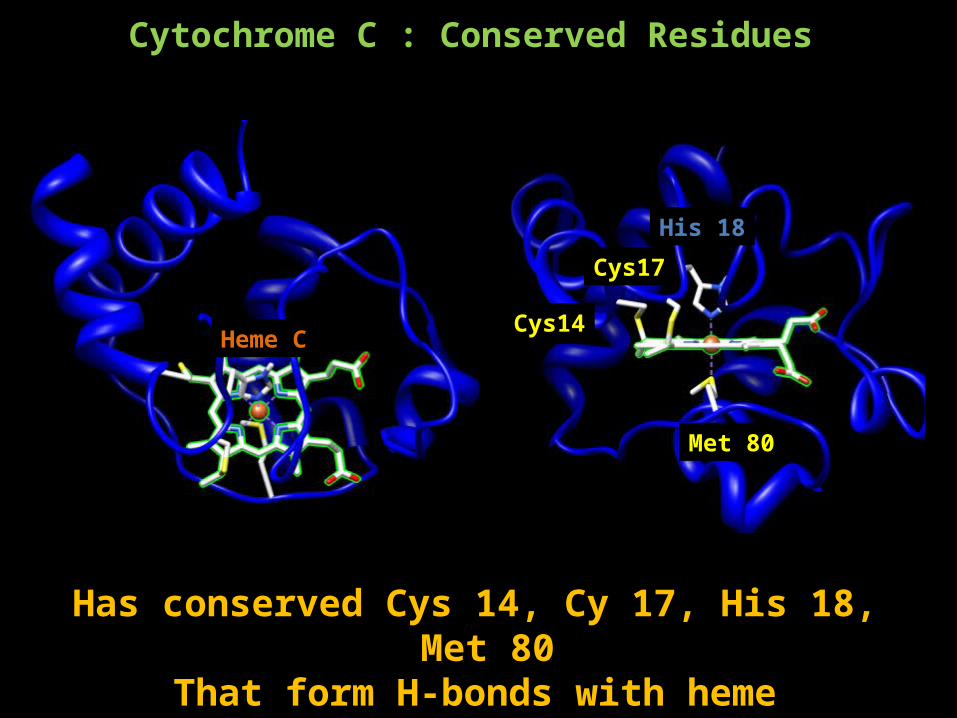
Cytochrome C : Conserved Residues
Heme C
His 18
Cys14
Cys17
Met 80
Has conserved Cys 14, Cy 17, His 18, Met 80 That form H-bonds with heme
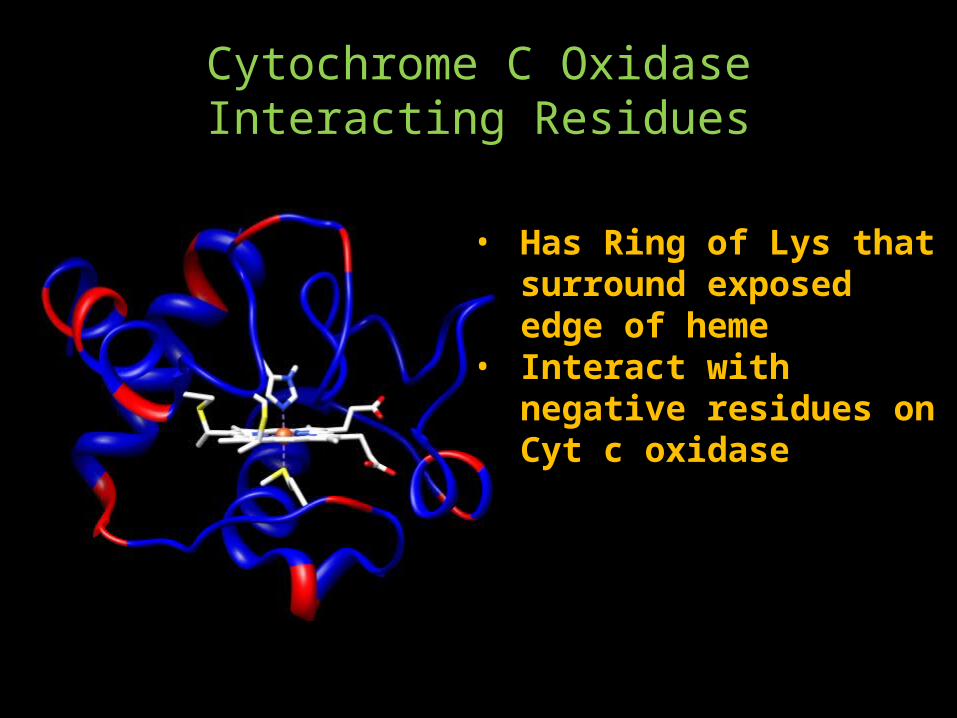
Cytochrome C Oxidase Interacting Residues
• Has Ring of Lys that surround exposed edge of heme
• Interact with negative residues on Cyt c oxidase

Two Mechanisms of Proton TransferDirect coupling Indirect coupling
• Reduction and protonation occur on the same molecule (cyt bc1)
• Neutral species diffuses to other side and releases e- and H+
OR• e- and H+ are transferred to an
acceptor• Each from a distinct side of the
membrane
• Requires energy generated from an exergenic reaction
• Energy can come from ATPlightRedox Reaction
Cytochrome C Oxidase uses the energy generated from the reduction of O2 to pump protons
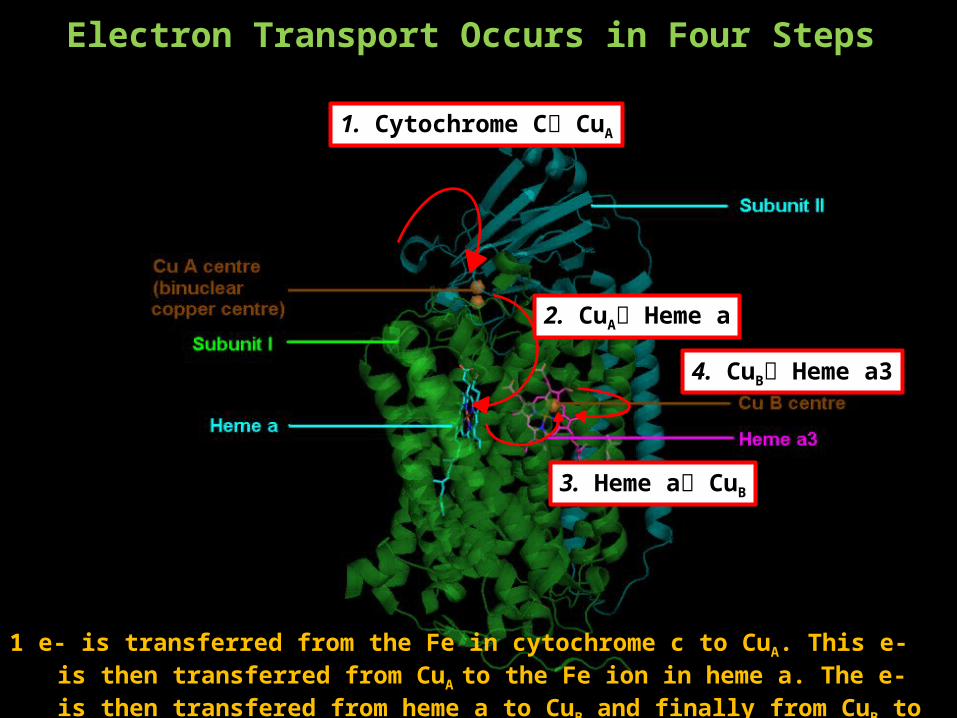
Electron Transport Occurs in Four Steps
1. Cytochrome C CuA
2. CuA Heme a
3. Heme a CuB
4. CuB Heme a3
1 e- is transferred from the Fe in cytochrome c to CuA. This e- is then transferred from CuA to the Fe ion in heme a. The e- is then transfered from heme a to CuB and finally from CuB to Fe in heme a3 where oxygen reduction occurs.
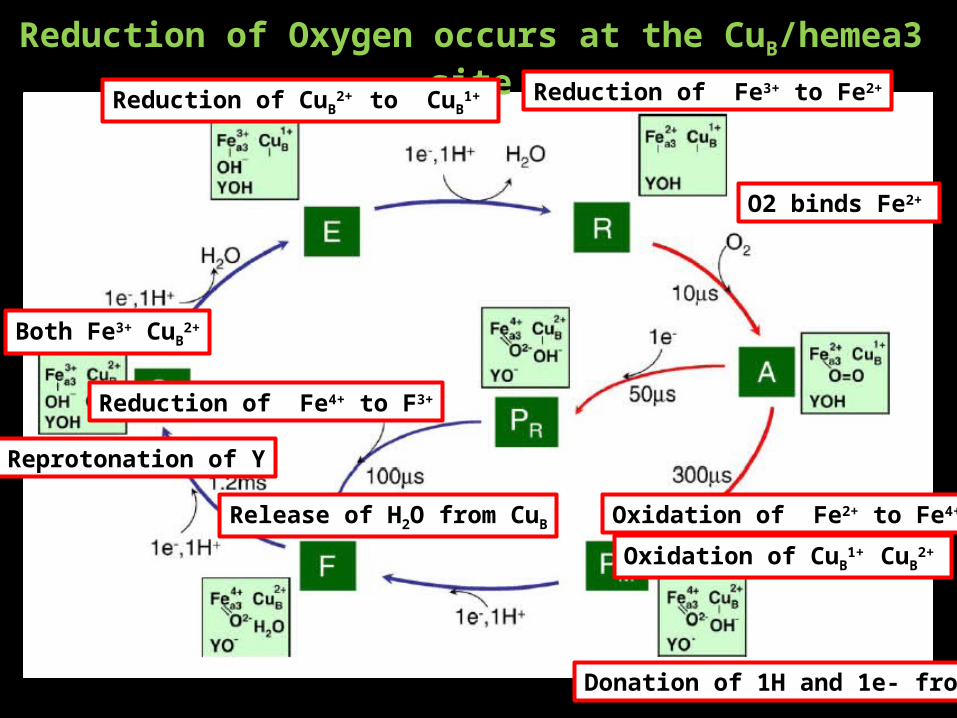
Reduction of Oxygen occurs at the CuB/hemea3 site
Both Fe3+ CuB2+
Reduction of CuB2+ to CuB
1+ Reduction of Fe3+ to Fe2+
O2 binds Fe2+
Oxidation of Fe2+ to Fe4+
Oxidation of CuB1+ CuB
2+
Release of H2O from CuB
Donation of 1H and 1e- from Y
Reprotonation of Y
Reduction of Fe4+ to F3+
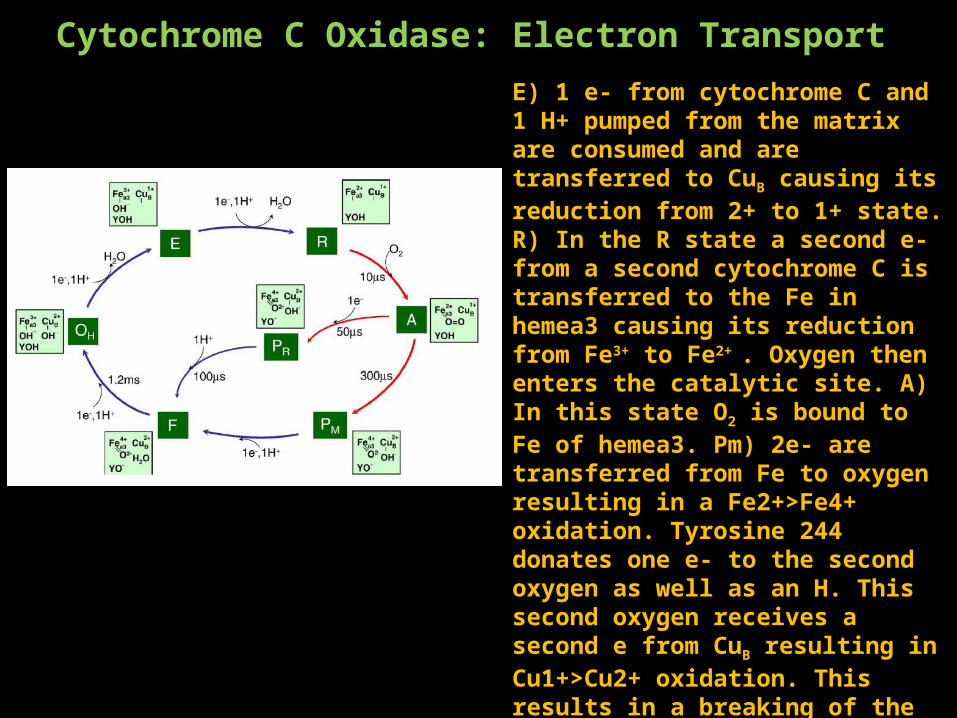
Cytochrome C Oxidase: Electron TransportE) 1 e- from cytochrome C and 1 H+ pumped from the matrix are consumed and are transferred to CuB causing its reduction from 2+ to 1+ state. R) In the R state a second e- from a second cytochrome C is transferred to the Fe in hemea3 causing its reduction from Fe3+ to Fe2+ . Oxygen then enters the catalytic site. A) In this state O2 is bound to Fe of hemea3. Pm) 2e- are transferred from Fe to oxygen resulting in a Fe2+>Fe4+ oxidation. Tyrosine 244 donates one e- to the second oxygen as well as an H. This second oxygen receives a second e from CuB resulting in Cu1+>Cu2+ oxidation. This results in a breaking of the O-O bond. F) The addition of a third e- from cytochrome c and a H+ from the matrix results in the release of H2O from CuB. O) A fourth and final e- from cytochrome c and a fourth H+ from the matrix results in reprotonation of Tyr244 and reduction of Fe from Fe4+>Fe3+ completing the cycle.
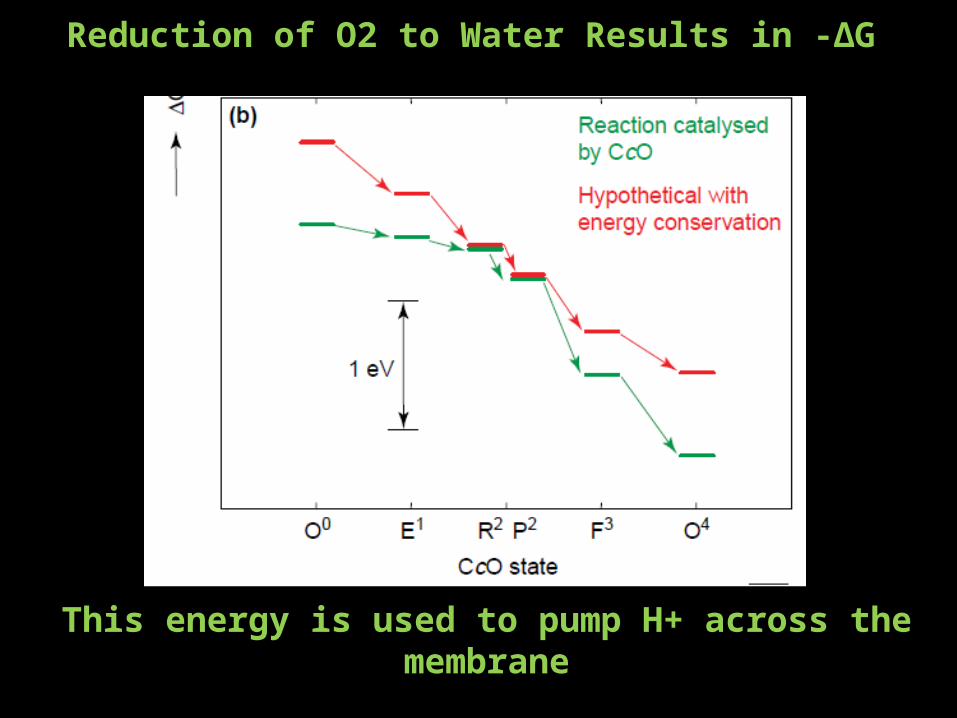
Reduction of O2 to Water Results in -∆G
This energy is used to pump H+ across the membrane
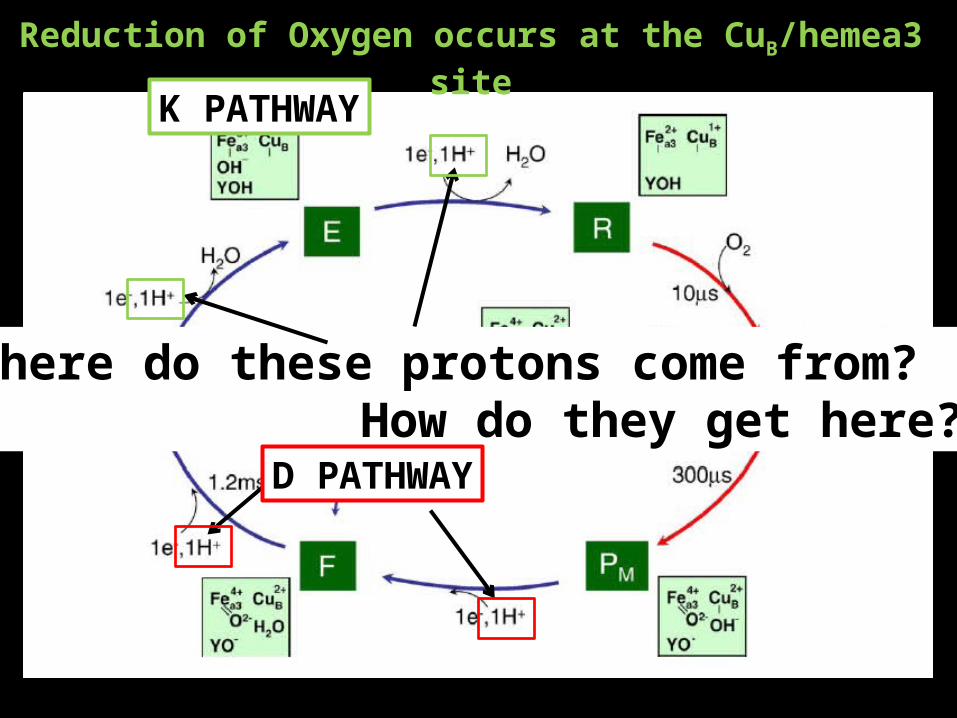
Reduction of Oxygen occurs at the CuB/hemea3 site
K PATHWAY
Where do these protons come from? How do they get here?
D PATHWAY
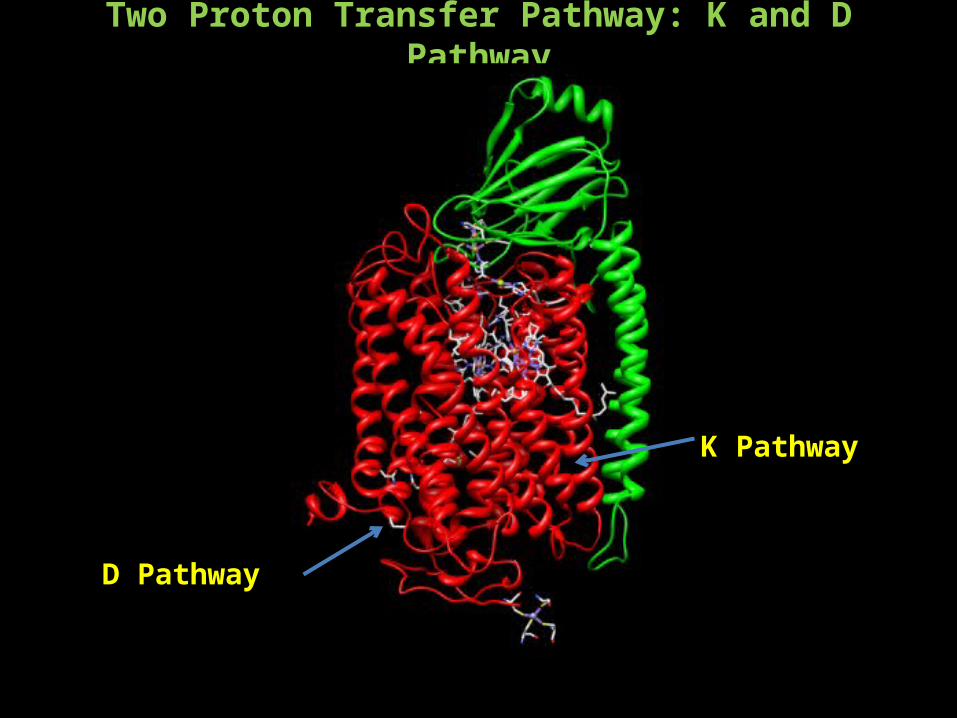
Two Proton Transfer Pathway: K and D Pathway
D Pathway
K Pathway
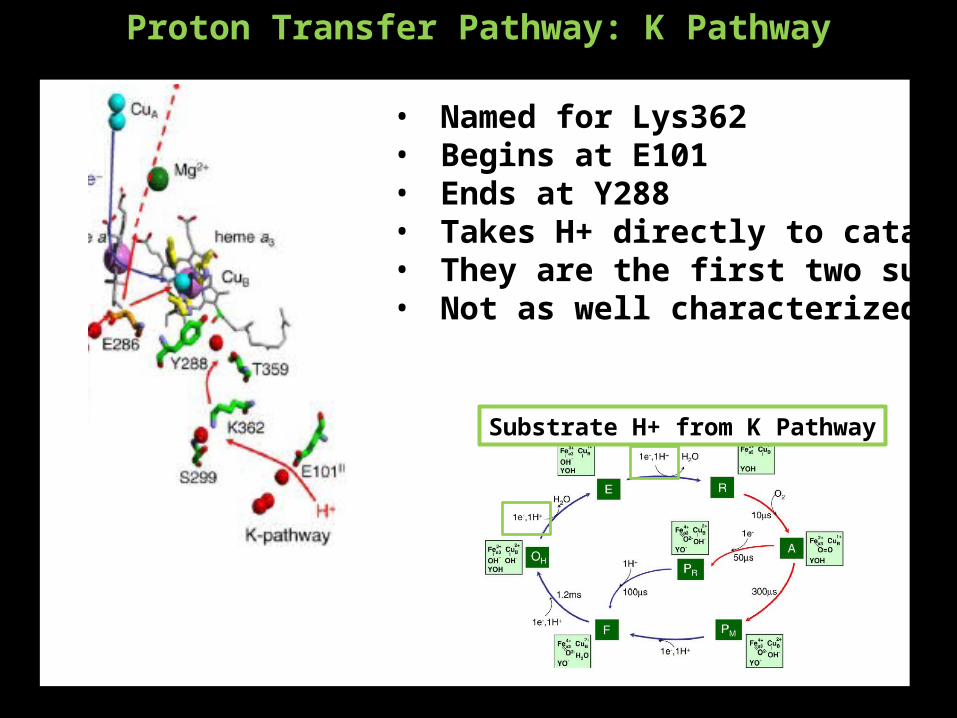
Proton Transfer Pathway: K Pathway
• Named for Lys362• Begins at E101• Ends at Y288• Takes H+ directly to catalytic site• They are the first two substrate H+• Not as well characterized
Substrate H+ from K Pathway
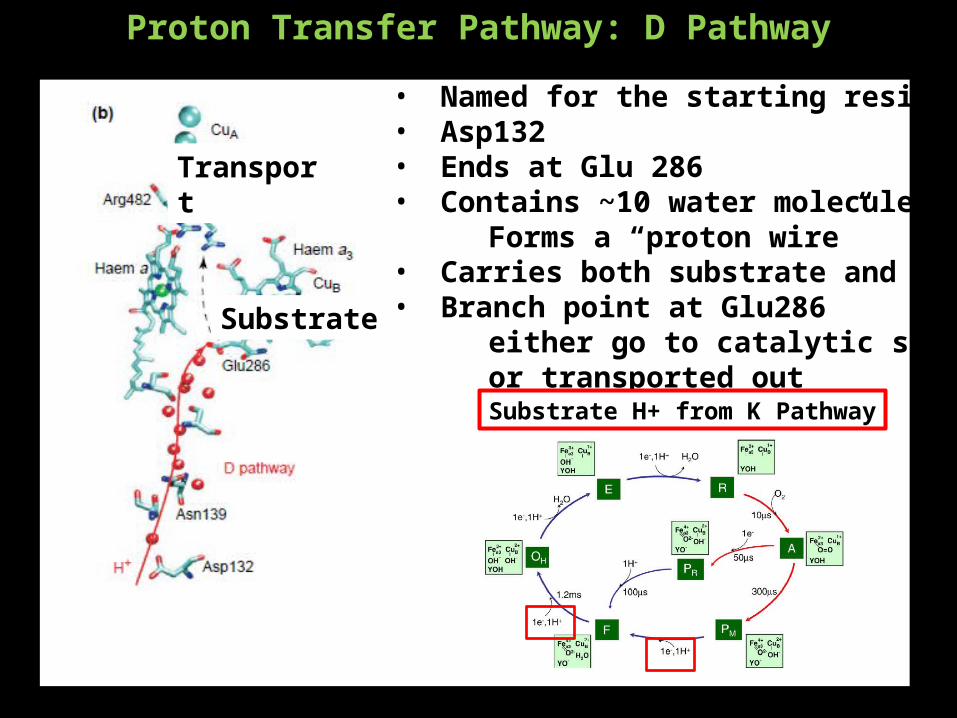
Proton Transfer Pathway: D Pathway
• Named for the starting residue• Asp132• Ends at Glu 286• Contains ~10 water molecules
Forms a “proton wire”• Carries both substrate and transport H+• Branch point at Glu286
either go to catalytic site as substrateor transported out
Substrate
Transport
Substrate H+ from K Pathway
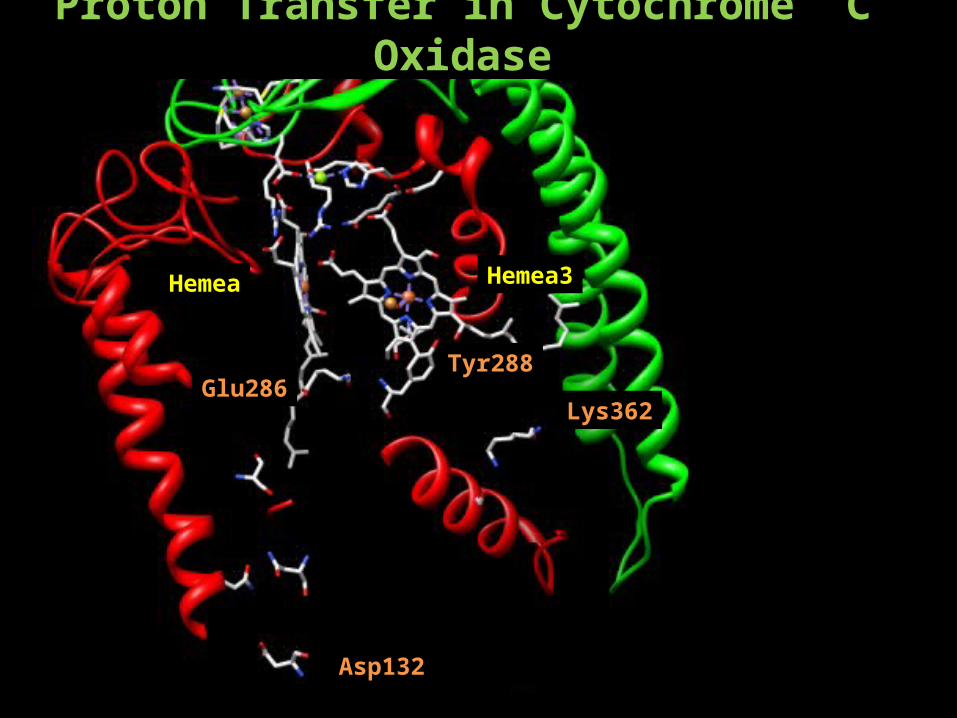
Proton Transfer in Cytochrome C Oxidase
Hemea Hemea3
Tyr288
Lys362
Asp132
Glu286
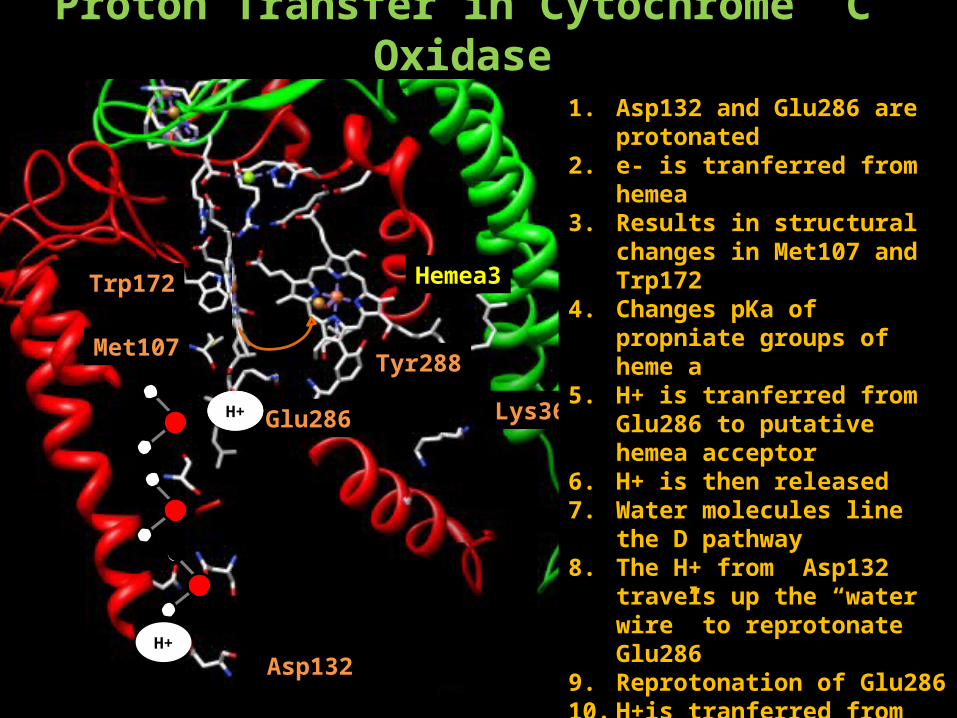
Proton Transfer in Cytochrome C Oxidase
Hemea3
Tyr288
Lys362
Asp132H+
Hemea
Glu286H+H+
Trp172
Met107
1. Asp132 and Glu286 are protonated
2. e- is tranferred from hemea3. Results in structural changes in
Met107 and Trp1724. Changes pKa of propniate groups
of heme a5. H+ is tranferred from Glu286 to
putative hemea acceptor6. H+ is then released7. Water molecules line the D
pathway8. The H+ from Asp132 travels up
the “water wire” to reprotonate Glu286
9. Reprotonation of Glu28610. H+is tranferred from Glu286 to
CuB.
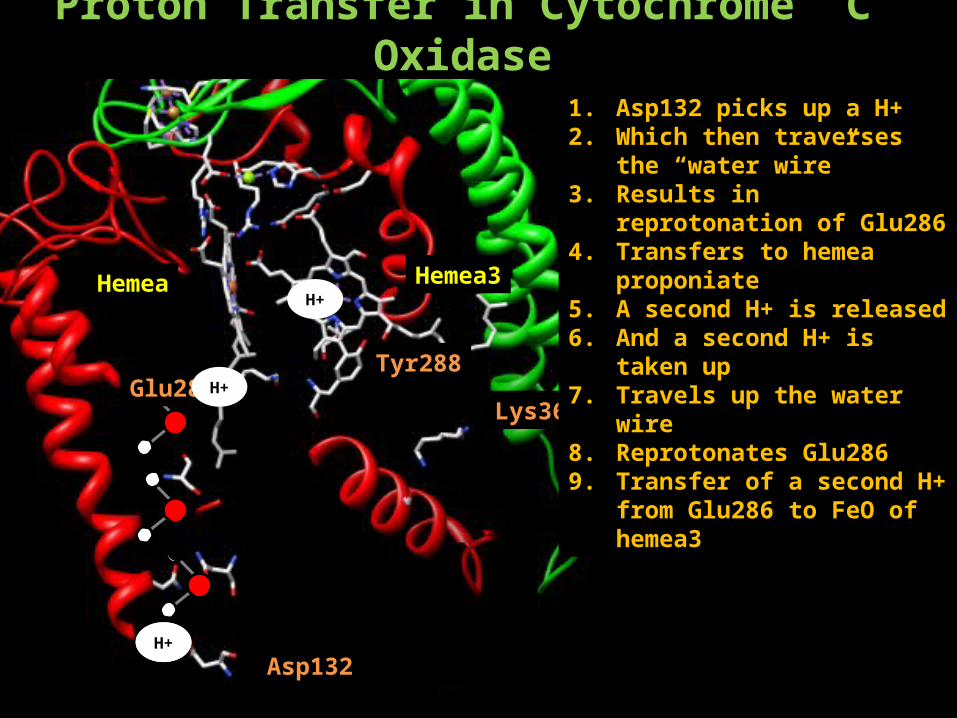
Proton Transfer in Cytochrome C Oxidase
Hemea Hemea3
Tyr288
Lys362
Asp132H+
Glu286
H+
1. Asp132 picks up a H+2. Which then traverses the “water
wire”3. Results in reprotonation of
Glu2864. Transfers to hemea proponiate 5. A second H+ is released6. And a second H+ is taken up7. Travels up the water wire 8. Reprotonates Glu2869. Transfer of a second H+ from
Glu286 to FeO of hemea3
H+
H+
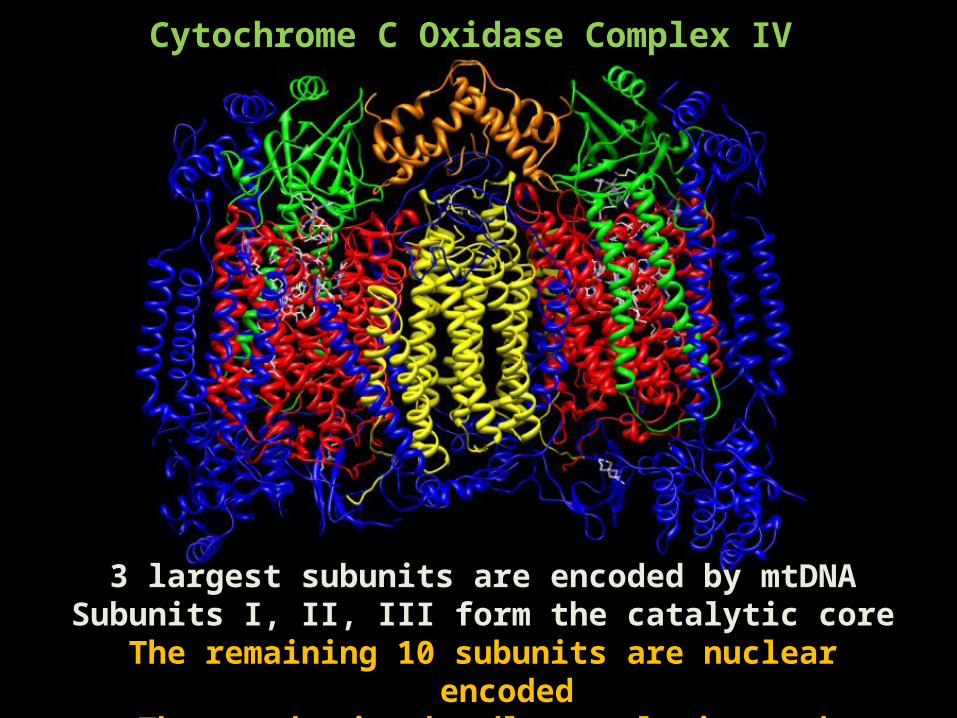
Cytochrome C Oxidase Complex IV
3 largest subunits are encoded by mtDNASubunits I, II, III form the catalytic core
The remaining 10 subunits are nuclear encodedThese subunits handle regulation and assembly
Important Inputs for Respiration
O2
ADP
ATP

Reasons for Regulation
• Would want increased respiration when demands are high• Might want to limit O2 consumption if oxygen is limited• Might want to increase respiration in response to low ATP
• Can be accomplished by Transcriptional regulation
Altered subunit expressionDirect inhibition or activation of the protein itself
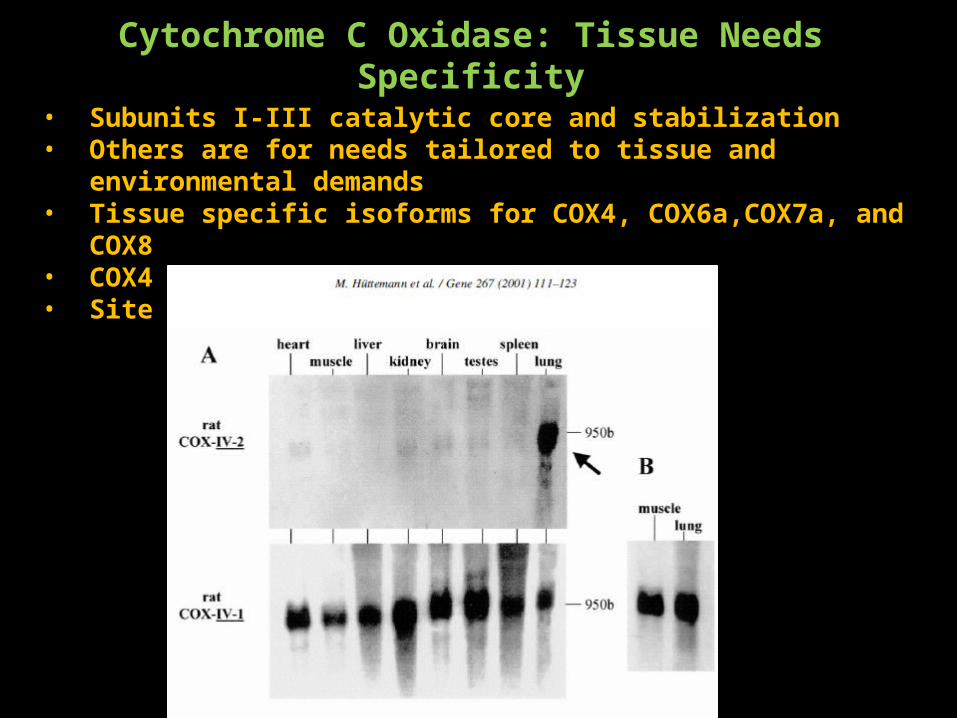
Cytochrome C Oxidase: Tissue Needs Specificity
• Subunits I-III catalytic core and stabilization• Others are for needs tailored to tissue and environmental demands• Tissue specific isoforms for COX4, COX6a,COX7a, and COX8• COX4 has a lung specific isoform• Site of ATP binding?
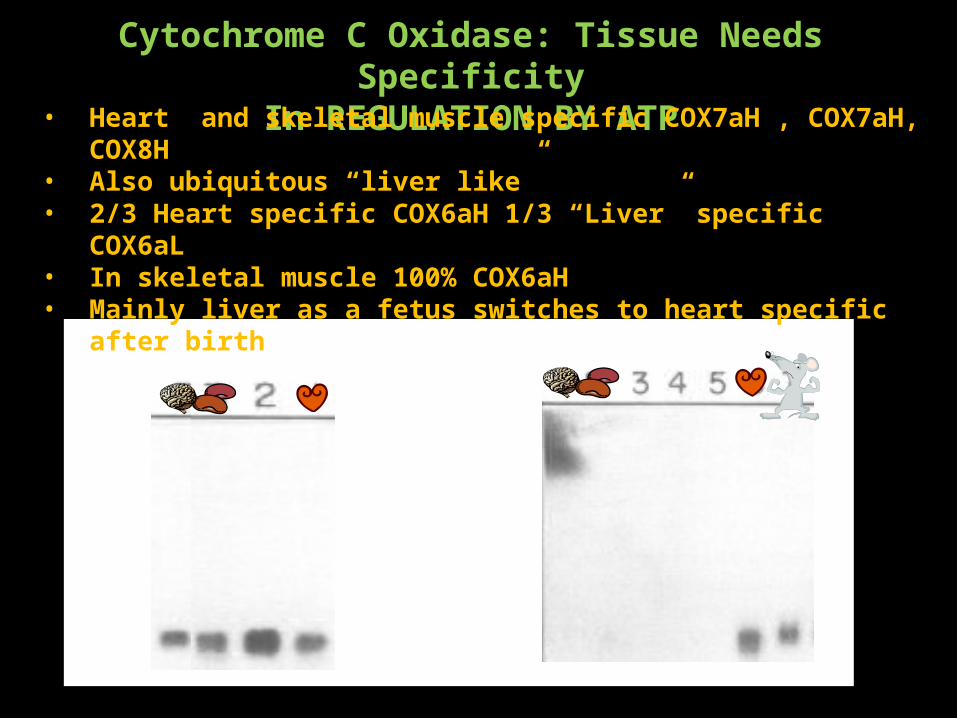
Cytochrome C Oxidase: Tissue Needs SpecificityIn REGULATION BY ATP
• Heart and skeletal muscle specific COX7aH , COX7aH, COX8H• Also ubiquitous “liver like” • 2/3 Heart specific COX6aH 1/3 “Liver” specific COX6aL• In skeletal muscle 100% COX6aH• Mainly liver as a fetus switches to heart specific after birth
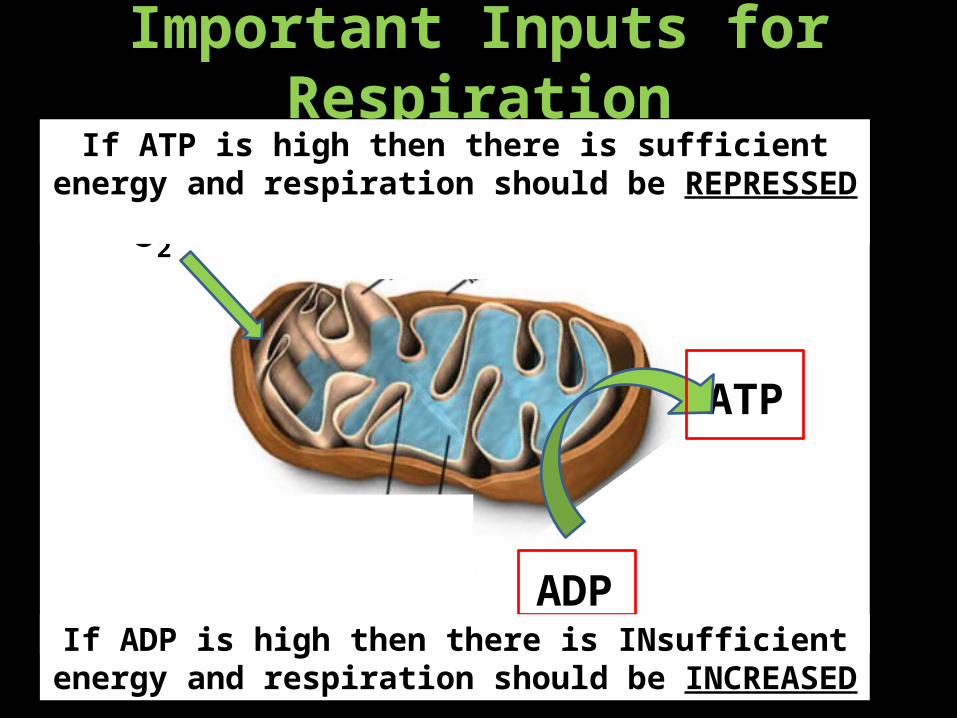
Important Inputs for Respiration
O2
ADP
ATP
If ATP is high then there is sufficient energy and respiration should be REPRESSED
If ADP is high then there is INsufficient energy and respiration should be INCREASED

ATP/ADP Ratio Regulates Cytochrome C Oxidase Activity
High ATP decreases H+/e-
High ADP stimulates COX activity
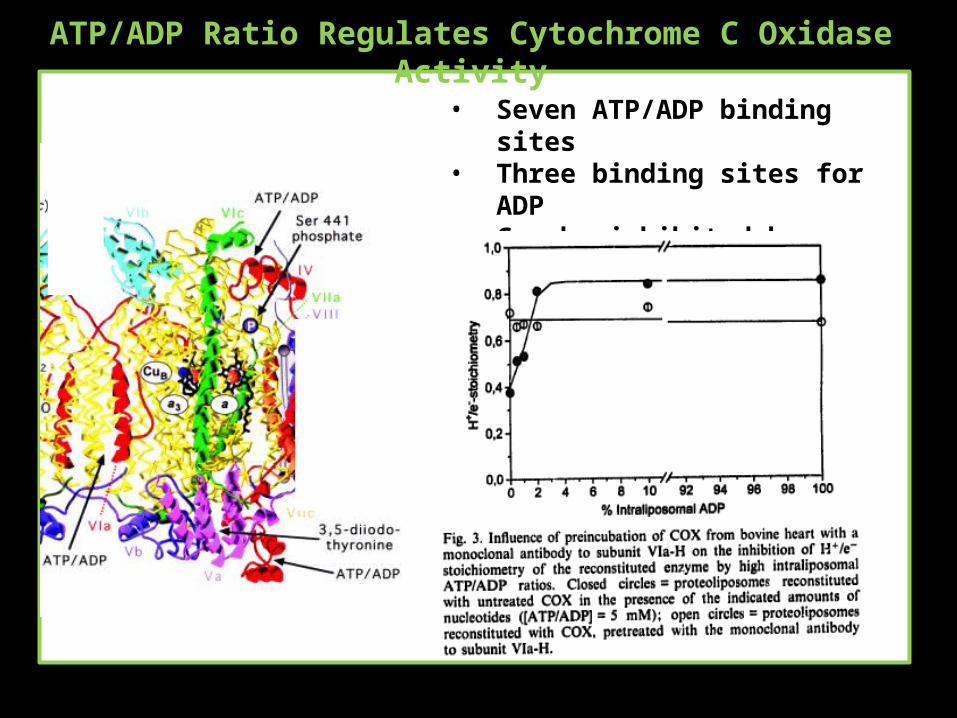
ATP/ADP Ratio Regulates Cytochrome C Oxidase Activity
• Seven ATP/ADP binding sites• Three binding sites for ADP• Can be inhibited by antibodies for
the binding site
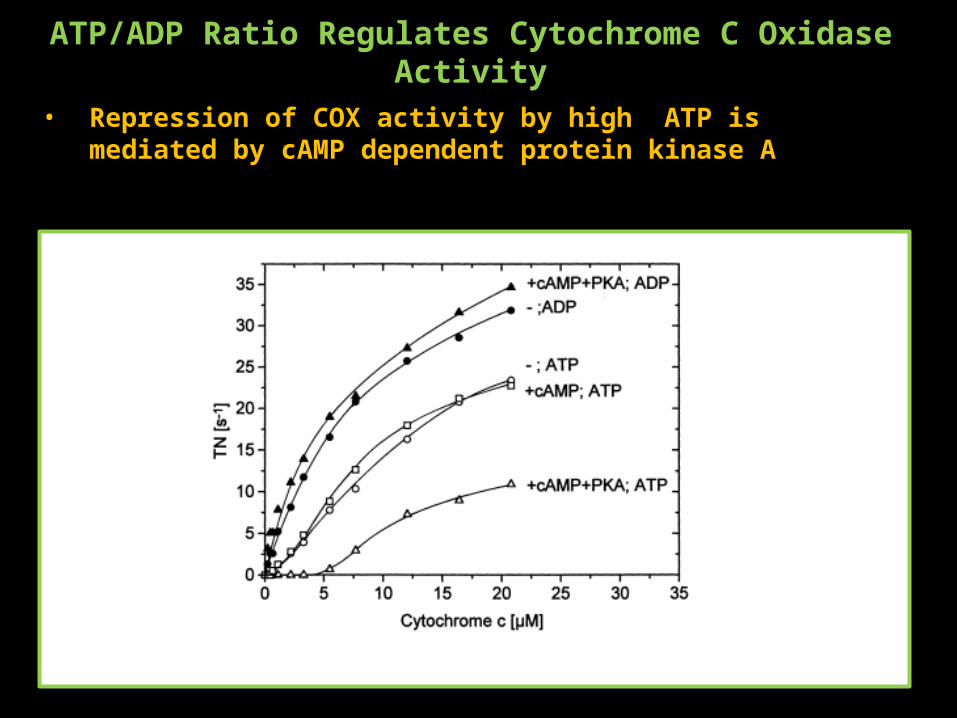
ATP/ADP Ratio Regulates Cytochrome C Oxidase Activity
• Repression of COX activity by high ATP is mediated by cAMP dependent protein kinase A

Important Inputs for Respiration
O2
ADP
ATP
When O2 is limited it must be conserved for use in other reactions so respiration should be REPRESSED under hypoxic
conditions
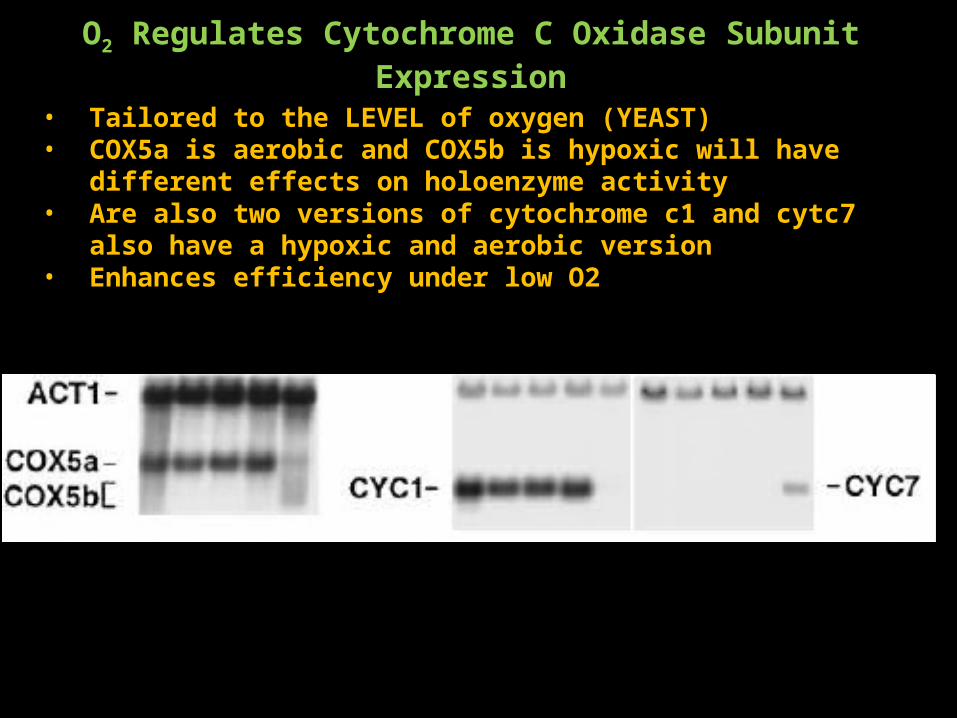
O2 Regulates Cytochrome C Oxidase Subunit Expression
• Tailored to the LEVEL of oxygen (YEAST)• COX5a is aerobic and COX5b is hypoxic will have different effects on
holoenzyme activity• Are also two versions of cytochrome c1 and cytc7 also have a hypoxic and
aerobic version• Enhances efficiency under low O2

Hypoxia Inducible Factor 1
• Transcription factor that responds to low O2 (hypoxia)• Two subunits: α and β• Hydroxylated by HIF-prolyl-hydroxylases on conserved prolines• Results in ubiquitination and degradation by the proteasome• HIF-prolyl-hydroxylases use O2 to hydroxylate• When O2 is low hydroxylation is inhibited
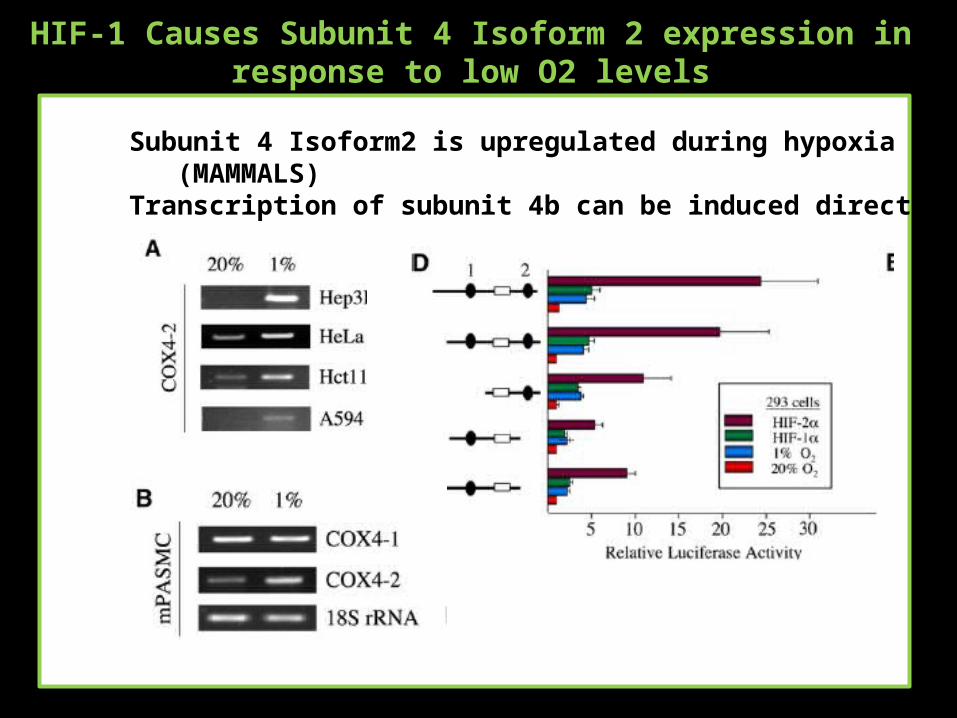
HIF-1 Causes Subunit 4 Isoform 2 expression in response to low O2 levels
Subunit 4 Isoform2 is upregulated during hypoxia (MAMMALS)Transcription of subunit 4b can be induced directly by HIF-1
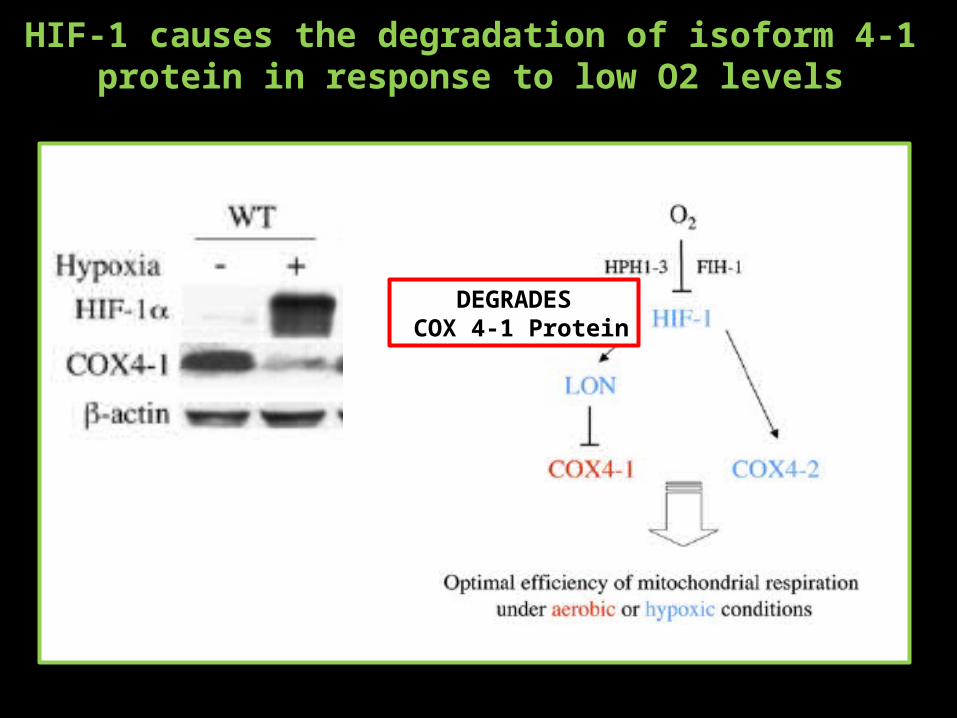
HIF-1 causes the degradation of isoform 4-1 protein in response to low O2 levels
DEGRADES COX 4-1 Protein
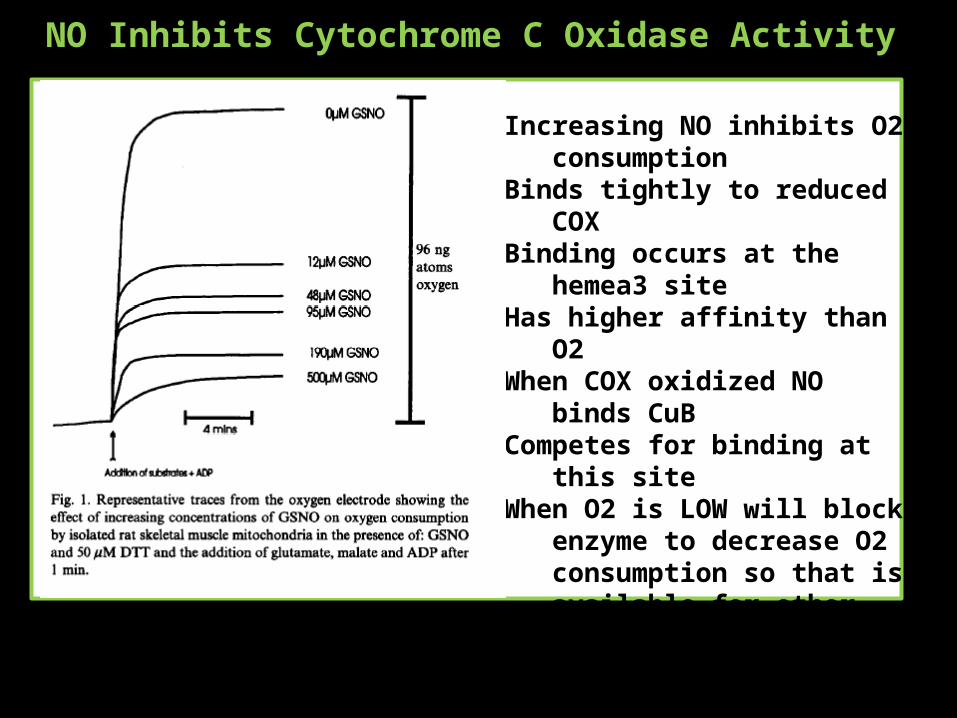
NO Inhibits Cytochrome C Oxidase Activity
Increasing NO inhibits O2 consumption
Binds tightly to reduced COXBinding occurs at the hemea3 siteHas higher affinity than O2When COX oxidized NO binds CuBCompetes for binding at this siteWhen O2 is LOW will block enzyme
to decrease O2 consumption so that is available for other processes

The production of ATP is necessary for life
O2
ATP
ADP
Making ATP is essential for life, even for bacteria. If an organism lacks the ability to pump protons, ability to generate ATP will be compromised and the organism is not viable

Cytochrome C Oxidase and Disease
• Mitochondrial diseases typically manifest in early adulthood• Mutations in the core subunits (SU1, SU2, SU3) are rare • Affect brain, muscle, heart (tissues with high energy demand)• Only ONE mutation (COX6) has been isolated in nuclear encoded subunits• Suggests that these are not viable
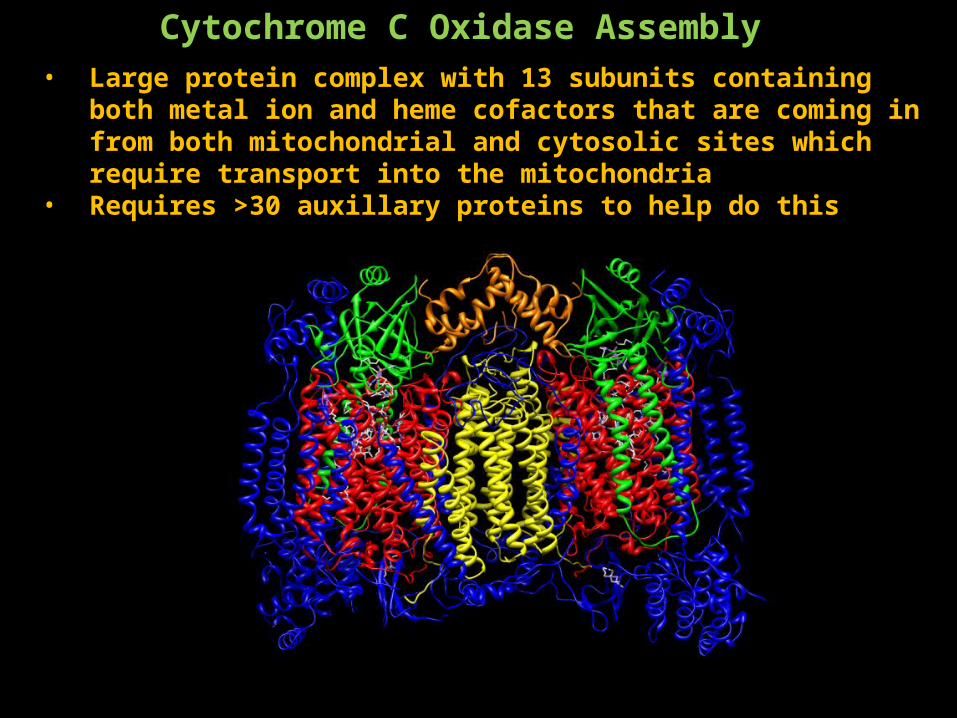
Cytochrome C Oxidase Assembly • Large protein complex with 13 subunits containing both metal ion and
heme cofactors that are coming in from both mitochondrial and cytosolic sites which require transport into the mitochondria
• Requires >30 auxillary proteins to help do this

Cytochrome C Oxidase Assembly • After translation, heme a is added to subunit I by COX10 and COX15• Cu atoms aare added to Subunit I by Sco1, Sco2,• Subunit 1 then binds Subunit 4/Subunit 5 complex• Phosphorylated SURF helps Subunit II bind the complex

Cytochrome C Oxidase Assembly Most disease causing mutations are in ASSEMBLY PROTEINS
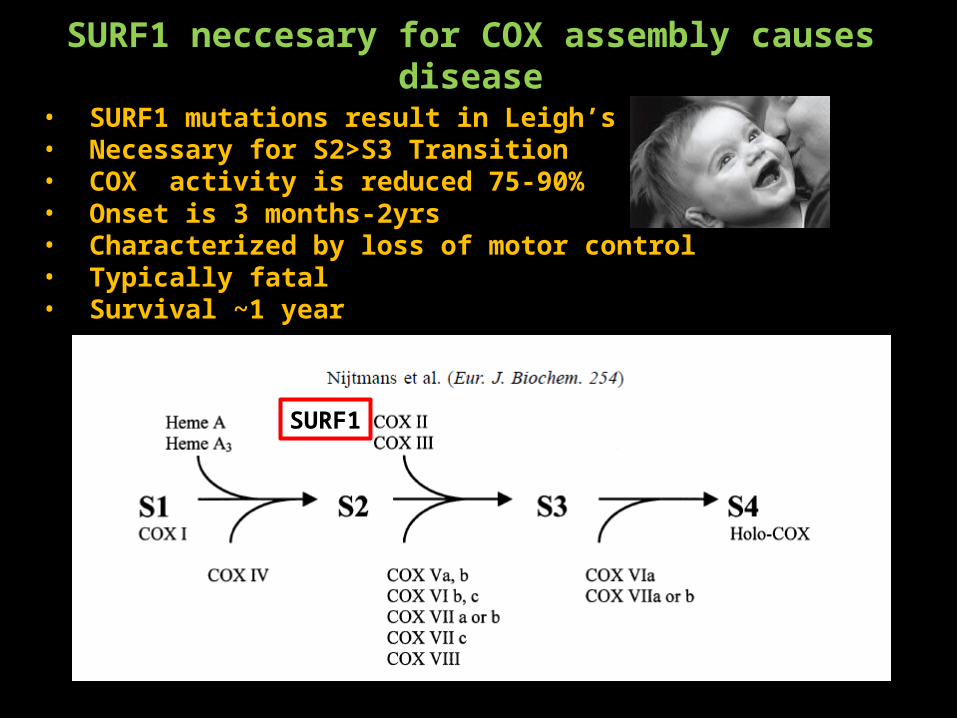
SURF1 neccesary for COX assembly causes disease
• SURF1 mutations result in Leigh’s Disesase• Necessary for S2>S3 Transition• COX activity is reduced 75-90%• Onset is 3 months-2yrs• Characterized by loss of motor control• Typically fatal• Survival ~1 year
SURF1

Cytochrome C Oxidase Summary• Catalyzes the reduction of O2 to H20• Uses this energy to generate a H+ gradient that is used to synthesize ATP• Electron transfer is a four step process that moves through two Cu and
heme molecules• The main reactions occur at the hemea3/CuB site• H+ are transferred through the K and D pathway• H+ traverse the D pathway via a “water wire” • Regulation of COX activity occurs due to
• Different energy demands by tissue or enviornment and is accomplished by• Subunit isoform switching• Direct interaction with feedback molecules
• Mutations in genes which regulate assembly of COX can result in disease

References
ADP/ATP ratios regulate CytCOx
Disease and cytochrome C
HIF-1 Regulates CytCOx
Oxygen Regulates CytCOx
NO Regulates Activity of CytCOxidase
Proton Pumping Mechanism D/K Pathway
Proton Pumpking (lined up with electron transfer steps)
CytC Electron Transfer Steps
Heart Specific Isoform
Lung Specific Isoform









![2018 - CyT - EG2 - R2[6215]](https://img.pdfslide.tips/doc/110x75/62bdaca1a4b7c7188a0d3f6b/2018-cyt-eg2-r26215.jpg)