Embed Size (px)
Citation preview

162
Rev. Cient. Prod. Anim., v.10, n.2, 2008
1 1Doutorando em Ciência Animal/UFPI, Bolsista da FAPEMA ([email protected])2DMV Reprodução Animal/UFMA ([email protected]). 3Doutoranda em Ciência Animal/UFPI ([email protected]). 4Laboratório de Reprodução Animal/CCA/UFPI ([email protected]).5Médico Veterinário ([email protected]). 6Mestranda em Medicina Veterinária/UEMA ([email protected]). 7DCCV/CCA/UFPI ([email protected]/[email protected]).
Efeito da Somatotropina Recombinante Bovina (rbST) na Resposta Ovulatória e na Qualidade dos Embriões de Vacas Nelore
Felipe de Jesus Moraes Junior1, José Ribamar de Souza Torres Junior2, Isolda Márcia do Nascimento3, Antônio de Sousa Júnior4, Hélcio Santos Correia5, Aline Brito da Silva6,
Rômulo José Vieira7, José Adalmir Torres de Souza7
RESUMO – Avaliou-se os efeitos da rbST na produção in vivo de embriões de oito vacas doadoras da raça Nelore. As superovulações foram realizadas em duas etapas, em Julho e Outubro de 2007. Na primeira etapa, quatro animais (G1) receberam apenas FSH e quatro (G2) receberam além do FSH, 250 mg de rbST, por via intramuscular, no dia denominado D0. Na segunda etapa, no sistema crossover, os animais que receberam apenas FSH, receberam 250 mg rbST. As superovulações foram iniciadas no D4, com 133 mg de FSH, aplicadas em quatro dias consecutivos, com doses decrescentes, a intervalo de 12 horas. Os implantes foram retirados em D7 após aplicação de duas doses de PGF2α em D6. No D8 foram aplicadas 200 μg de um análogo do GnRH e 12 a 24 horas após, as inseminações foram realizadas. Os embriões foram coletados no sétimo dia após a primeira inseminação. A resposta superovulátoria (presença de corpos lúteos), total de estruturas, estádios de desenvolvimento e qualidade dos embriões recuperados não foram afetados (P>0,05) pelo tratamento com a rbST. As médias obtidas nos tratamento G1 e G2 foram respectivamente: estruturas colhidas, 9,5±3,0 e 7,6±2,2; embriões viáveis, 8,1±2,6 e 5,0±1,6; estruturas não fertilizadas, 0,3±0,2 e 0,5±0,3; estruturas degenerados, 1,0±0,5 e 0,5±0,3; mórulas, 0,5±0,2 e 2,3±1,9; blastocistos iniciais, 2,2±2,1 e 1,3±0,9; blastocistos, 4,7±1,6 e 2,5±0,5; blastocistos expandidos, 0,6±0,3 e 0,5±0,5; embriões grau I 7,3±2,5 e 5,0±0,8; embriões grau II, 0,5±0,3 e 1,1±0,5 e embriões grau III, 0,2±0,1 e 0,5±0,5. A administração da rbST no D0, na dose de 250 mg, não constitui boa alternativa para melhorar a qualidade de embriões em vacas Nelore.
Palavras-chave: bovino, FSH, GH
Rev. Cient. Prod. Anim., v.10, n.2, p.162-173, 2008

163
Rev. Cient. Prod. Anim., v.10, n.2, 2008
Effect of the Bovine Recombinant Somatotropin (rbST) in the Ovulatory Response and Embryos Quality of Nelore Cows
ABSTRACT - To evaluate the effect of rbST in the in vivo production of Nelore cows embryos a study was carried with eight givers. The superovulation was carried in 2007, in two stages, the first one in July and the second other in October. In the first stage, four animals (G1) received only FSH and four (G2) received besides the FSH, 250 mg of rbST, both by intramuscular application, in the day D0. In the second stage, in the crossover system, the animals that had received only FSH received 250 mg of rbST. The superovulation was initiated in the D4, with 133 mg of FSH, applied in four consecutive days, with decreasing doses in the 12 hours interval. The implantations were removed in the D7 after application of two doses of PGF2α in D6. In the D8, 200 μg of GnRH analogous were applied, and after 12 and 24 hours inseminations took place. The embryos were collected in the seventh day after the first insemination. The superovulated reply (presence of lutea corpora), total of structures, developmental stage and quality of the recouped embryos had not been affected (P>0.05) by the rbST treatment. The averages of G1 and G2 treatment were: recovered structures, 9.5±3.0 and 7.6±2.2; viable embryos, 8.1±2.6 and 5.0±1.6; not fertilized, 0.3±0.2 and 0.5±0.3; degenerated, 1.0±0.5 and 0.5±0.3; morulae, 0.5±0.2 and 2.3±1.9; early blastocysts, 2.2±2.1 and 1.3±0.9; blastocysts, 4.7±1.6 and 2.5±0.5; expanded blastocysts, 0.6±0.3 and 0.5±0.5; embryos degree I, 7.3±2.5 and 5.0±0.8; embryos degree II, 0.5±0.3 and 1.1±0.5 and embryos degree III, 0.2±0.1 and 0.5±0.5, respectively. The administration of 250 mg of rbST in the D0, do not improve the embryos quality in Nelore cows.
Key Words: bovine, FSH, GH
Introdução
O maior entrave dos programas de Transferência de Embriões (TE) continua sendo o elevado custo e a variabilidade da resposta ao tratamento superovulatório entre as doadoras. Apesar do intenso número de pesquisas desenvolvidas
em diversos países ainda não existe um protocolo capaz de contornar este problema. Alguns protocolos que sincronizam o desenvolvimento da onda folicular têm se revelado úteis por facilitarem o manejo das doadoras e produzirem o mesmo número de embriões dos tratamentos tradicionais utilizados (Nagano et al., 2004).

164
Rev. Cient. Prod. Anim., v.10, n.2, 2008
A utilização da somatotropina recombinante bovina (rbST) em doadoras no início do ciclo estral, tem sido pesquisada com o objetivo de aumentar a população de folículos recrutados e, conseqüentemente, melhorar a resposta superovulatória (Gong et al., 1996).
Os principais hormônios utilizados para superovular fêmeas são o hormônio folículo-estimulante (FSH) e a gonadotrofina coriônica eqüina (eCG) e quando associados à rbST têm a capacidade de aumentar o número de folículos recrutados e embriões viáveis (Gong et al., 1993; Gong et al., 1996).
A rbST foi um dos primeiros fatores de crescimento produzidos em escala comercial como proteína recombinante (Bauman, 1999). A maior quantidade de receptores para a rbST é encontrada no fígado, que é responsável pela produção do fator de crescimento semelhante à insulina (IGF-I). Entretanto a maioria dos outros tecidos também responde à rbST. Sobre o sistema reprodutor, esse hormônio atua sobre os receptores do útero e ovário induzindo-os à produção de IGF-I (Lucy, 2000).
Em vacas, arbST diminuiu o número de estruturas não fertilizadas, melhorou a taxa de sobrevivência embrionária e acelerou o desenvolvimento embrionário (Moreira et al., 2002). Por sua vez, Nagano et al. (2004) observaram que 500 mg de
rbST aplicados no dia zero do protocolo de FSH, em vacas, aumentaram o número total de embriões, de ovócito e de embriões viáveis.
Esta pesquisa foi realizada com o objetivo de avaliar a rbST na produção in vivo de embriões de vaca Nelore.
Material e Métodos
O experimento foi realizado na fazenda Canarana, município de Vitória do Mearim-MA, com coordenadas geográficas 03º27’44’’ latitude Sul e 44º52’14’’ longitude Oeste. O clima da região é quente e úmido, com o período chuvoso de dezembro a junho, temperatura de 22 a 35ºC e precipitação pluviométrica média anual 1.404 mm (NUGEO/UEMA, 2004). O experimento foi realizado nos meses de julho e outubro de 2007, correspondendo ao período de estiagem.
Foram utilizadas oito vacas puras de origem (PO) da raça Nelore, não lactantes, com peso vivo médio 525,37±26,84 kg e escore de condição corporal 3,0 a 3,5, em escala de 1 a 5, onde 1 corresponde ao animal muito magro e 5 ao muito gordo (Dirksen et al., 1993), mantidas em pastagem de Brachiaria brizantha, sem suplementação, com água e sal mineral à vontade. Os animais foram previamente examinados por palpação retal, no intuito de eliminar fêmeas com problemas clínico-
165
Rev. Cient. Prod. Anim., v.10, n.2, 2008
ginecológicos.Adotou-se o delineamento
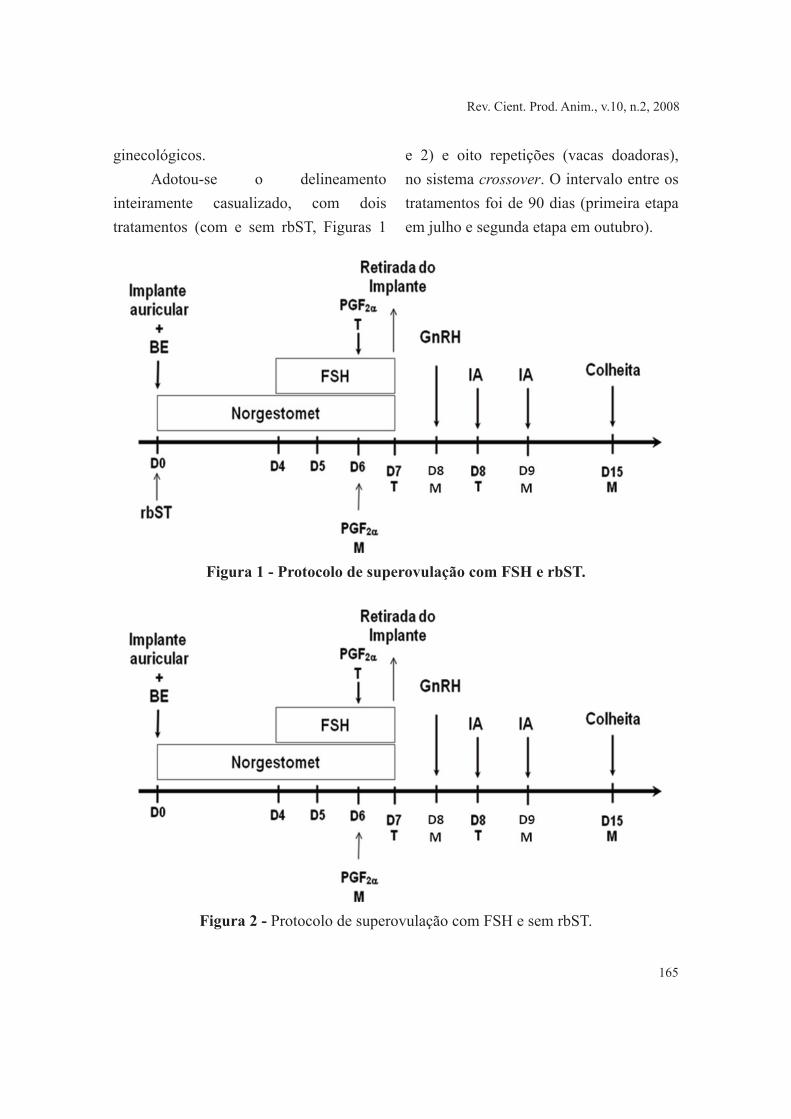
inteiramente casualizado, com dois tratamentos (com e sem rbST, Figuras 1
e 2) e oito repetições (vacas doadoras), no sistema crossover. O intervalo entre os tratamentos foi de 90 dias (primeira etapa em julho e segunda etapa em outubro).
Figura 1 - Protocolo de superovulação com FSH e rbST.
Figura 2 - Protocolo de superovulação com FSH e sem rbST.

166
Rev. Cient. Prod. Anim., v.10, n.2, 2008
As doadoras receberam dois implantes auriculares com 6 mg de Norgestomet (Crestar®, Intervet, Boxmeer, Holanda) e 2 mg de Benzoato de Estradiol (RIC-BE®, Syntex, Buenos Aires, Argentina) associados a 250 mg de rbST (Boostin®, Schering-Plough Saúde Animal Indústria e Comércio Ltda.) em um dia aleatório do ciclo estral, denominado dia zero (D0).
As superovulações foram realizadas com 133 mg do FSH (Folltropin®; Bioniche Canadá Inc., Ontário, Canadá) divididas em oito doses decrescentes, aplicadas duas vezes ao dia, por quatro dias consecutivos. Quando da 5ª e 6ª administração de FSH as doadoras receberam 150 μg de D-Cloprostenol (Prolise®, Syntex, Buenos Aires, Argentina) e à 8ª aplicação foi retirado o implante auricular. No D8 foram administrados 200 μg de um análogo do GnRH (Fertagyl®, Intervet, Boxmeer, Holanda). A inseminação artificial, realizada com sêmen de uma única partida de um touro, foi efetivada 12 e 24 h após aplicação do GnRH. Dois tratamentos, um com rbST e outro sem rbST foram definidos e estão apresentados nas Figuras 1 e 2.
No D15 do programa, os corpos lúteos foram quantificados através de ultra-sonografia, utilizando um aparelho Pie Medical, munido de um transdutor linear de 6/8 MHz. A seguir, realizou-se anestesia epidural e higienização da região perineal,
procedendo-se então a colheita das estruturas. Utilizou-se um cateter de duas vias com um balão em sua extremidade (sonda de Foley, N. 20), sendo este inflado após passagem pela cérvix e posicionado no corno uterino, vedando a passagem para o corpo uterino, impedindo o refluxo de líquidos. O meio utilizado para lavagem foi o PBS (Phosphate Buffer Solution) aquecido a 37ºC, utilizando cerca de 500 ml para lavagem de cada corno uterino. O líquido recolhido foi passado em filtro apropriado, onde ficaram retidas as estruturas em cerca de 50 ml do meio. Repetido o processo no outro corno uterino, o conteúdo foi colocado em uma placa de Petri, para identificação e captura de estruturas, com auxílio de uma lupa estereoscópia sob aumento de 40X. As estruturas capturadas foram colocadas em outra placa de Petri com meio de manipulação (HOLDING Embriocare) e foram classificados quanto aos estádios de desenvolvimento (Lindner & Wright,1983) e avaliados quanto à qualidade (Kennedy et al., 1983).
Os estádios de desenvolvimento foram classificados: estádio 1 (mórula: aglomerado celular em cuja superfície blastômeros individuais podem ser distinguidos); estádio 2 (mórula compacta: blastômeros individuais não podem ser distinguidos na superfície do embrião); estádio 3 (blastocisto inicial: uma pequena cavidade, a blastocele, está visível e a massa

167
Rev. Cient. Prod. Anim., v.10, n.2, 2008
celular interna começa a se formar); estádio 4 (blastocisto: o embrião ocupa a maior parte dentro da zona pelúcida, a massa celular interna começa a tornar-se mais distinta, mas o diâmetro global do embrião, incluindo a zona pelúcida, permanece inalterado); e estádio 5 (blastocisto expandido: o diâmetro embrionário está aumentado e a espessura da zona pelúcida pode ser reduzida para aproximadamente 1/3 da espessura original). Quanto à qualidade, os embriões foram classificados em quatro graus: grau I (excelente: estádio de desenvolvimento com zona pelúcida intacta e esférica, massa celular homogênea com células de tamanho uniforme, nenhum ou poucos fragmentos celulares no espaço perivitelino); grau II (bom: alterações mínimas na forma e coloração com relação ao grau I, alguns fragmentos ou debris celulares no espaço perivitelino e/ou pequenas formações vesiculares nos blastômeros); grau III (regular: claras alterações comparadas com o grau II, embora com a maior parte da massa celular intacta); e grau IV (ruim: muitos fragmentos ou debris celulares no espaço perivitelino, vesículas maiores e em maior número e claras mudanças degenerativas nos blastômeros, com menos da metade da massa celular intacta).
Após a avaliação, os embriões (mórulas e blastocistos) de graus I e II foram colocados em uma placa de Petri contendo
meio de congelação (etilenoglicol), onde permaneceram por 10 min. Em seguida, foram envasados em palhetas de 0,25 mL contendo cinco colunas, duas de PBS nas extremidades e três mais internas de etilenoglicol, sendo que a coluna central continha o embrião. As palhetas, depois de vedadas com lacradores plásticos e identificadas, foram congeladas utilizando-se uma máquina modelo TK 3000®.
Os dados foram analisados utilizando-se o pacote estatístico SAS (1997), para obtenção de médias e erros padrões dos parâmetros avaliados (total de estruturas, embriões viáveis, estruturas não fertilizadas, estruturas degeneradas, estádios de desenvolvimento e qualidade embrionária). Foi utilizado o teste não-paramétrico Wilcoxon, com probabilidade de 5%.
Resultados e Discussão
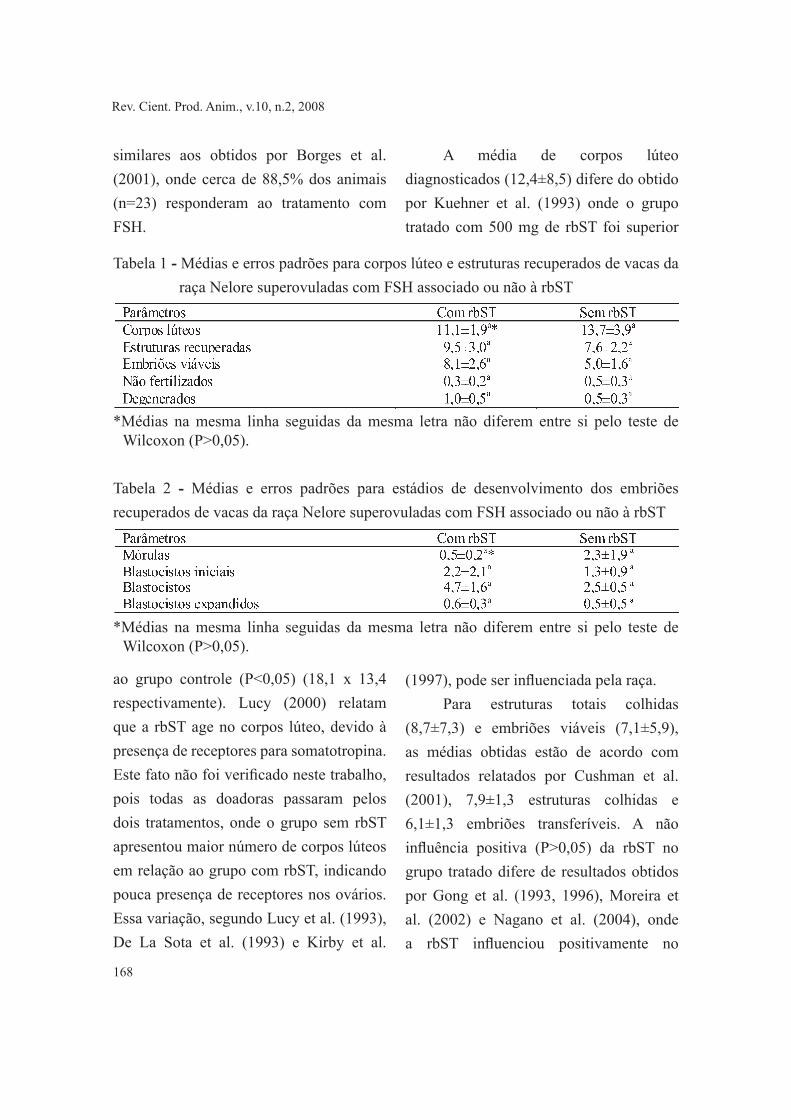
Para corpos lúteos, estruturas recuperadas (embriões viáveis, não fertilizados e degenerados) e estádios de embriões (mórulas, blastocistos iniciais, blastocistos e blastocistos expandidos) não houve efeito (P>0,05) dos tratamentos, onde as médias gerais foram 12,4±8,5; 9,9±8,7 e 7,5±6,1, respectivamente, (Tabelas 1 e 2). Das 16 superovulações, 14 (87,5%) responderam ao tratamento com FSH com três ou mais ovulações. Estes dados foram
168
Rev. Cient. Prod. Anim., v.10, n.2, 2008
similares aos obtidos por Borges et al. (2001), onde cerca de 88,5% dos animais (n=23) responderam ao tratamento com FSH.
Tabela 1 - Médias e erros padrões para corpos lúteo e estruturas recuperados de vacas da raça Nelore superovuladas com FSH associado ou não à rbST
*Médias na mesma linha seguidas da mesma letra não diferem entre si pelo teste de Wilcoxon (P>0,05).
Tabela 2 - Médias e erros padrões para estádios de desenvolvimento dos embriões recuperados de vacas da raça Nelore superovuladas com FSH associado ou não à rbST
*Médias na mesma linha seguidas da mesma letra não diferem entre si pelo teste de Wilcoxon (P>0,05).
ao grupo controle (P<0,05) (18,1 x 13,4 respectivamente). Lucy (2000) relatam que a rbST age no corpos lúteo, devido à presença de receptores para somatotropina. Este fato não foi verificado neste trabalho, pois todas as doadoras passaram pelos dois tratamentos, onde o grupo sem rbST apresentou maior número de corpos lúteos em relação ao grupo com rbST, indicando pouca presença de receptores nos ovários. Essa variação, segundo Lucy et al. (1993), De La Sota et al. (1993) e Kirby et al.
(1997), pode ser influenciada pela raça.Para estruturas totais colhidas
(8,7±7,3) e embriões viáveis (7,1±5,9), as médias obtidas estão de acordo com resultados relatados por Cushman et al. (2001), 7,9±1,3 estruturas colhidas e 6,1±1,3 embriões transferíveis. A não influência positiva (P>0,05) da rbST no grupo tratado difere de resultados obtidos por Gong et al. (1993, 1996), Moreira et al. (2002) e Nagano et al. (2004), onde a rbST influenciou positivamente no
A média de corpos lúteo diagnosticados (12,4±8,5) difere do obtido por Kuehner et al. (1993) onde o grupo tratado com 500 mg de rbST foi superior

169
Rev. Cient. Prod. Anim., v.10, n.2, 2008
recrutamento dos folículos, no aumento do número de folículos ovulatórios e, conseqüentemente, no de embriões viáveis. Resultados semelhantes também foram relatados por Neves et al. (2005) em três grupos experimentais: controle 0 mg, 250 mg e 500 mg de rbST. O número de estruturas recuperadas pelos autores foi 8,47±5,60; 9,36±8,37 e 8,86±6,88 e de embriões viáveis 3,33±2,61; 5,91±9,31 e 4,79±5,79, respectivamente, e por Borges et al. (2001), onde as médias das estruturas totais 15,3±9,5 e 17,4±10,9 e embriões viáveis 8,5±8,3 e 11,0±10,0 foram menores em animais tratados que no grupo controle. Segundo esses autores, a utilização do rbST antes do início da superovulação não aumenta as médias de estruturas colhidas e embriões viáveis.
Um fator que pode ter contribuído para a não influência da rbST sobre as estruturas colhidas e embriões viáveis, foi a dose utilizada, pois, segundo Rieger et al. (1991), em vacas da raça holandesa, para estímulo dos ovários são necessárias doses maiores que 500 mg de rbST. De modo geral, fêmeas zebuínas necessitam de menor dose hormonal para estimulação ovariana quando comparada com fêmeas de raças européias. Na presente pesquisa a raça foi Nelore e a dose utilizada 250mg. Outro fator que também pode ter influenciado os resultados é a raça, já observado em vacas da raça Holandesa (Hasler et al., 1983),
Angus, Brahman (Segerson et al., 1984), Charolês, Simental e Hereford (Breuel et al., 1991).
A ausência do efeito do tratamento sobre embriões degenerados e não fertilizados, verificada nesta pesquisa, corrobora com os dados de Nagano et al. (2004) e contrapõe-se aos resultados de Moreira et al. (2002), que atribuem à rbST o aumento da taxa de fertilização e melhoria na qualidade dos embriões. Lucy et al. (1995), atribuem os efeitos diretos e indiretos a este hormônio, pois a rbST aumenta as concentrações IGF-I, podendo melhorar a qualidade embrionária, o que não foi observado nesta pesquisa.
A rbST não afetou (P>0,05) os estádios de desenvolvimento embrionários quando colhidos sete dias após a primeira inseminação. As maiores médias, tanto no tratamento com rbST como sem rbST referem-se aos estádios de blatocisto inicial (1,8±4,6) e blastocisto (3,7±3,6). Coelho (1986), ao obter embriões no sétimo dia de superovulação, também não observou diferença nas médias de mórulas compactas e blastocistos jovens em taurinos (Bos taurus), como também no número de mórulas e mórulas compactas em zebuínos (Bos indicus), demonstrando a variabilidade individual no desenvolvimento e na qualidade embrionária. Variações de 24 a 48 h na idade dos embriões podem ocorrer no desenvolvimento embrionário (Lindner

170
Rev. Cient. Prod. Anim., v.10, n.2, 2008
e Wright, 1983). Deste modo, é importante considerar o dia das colheitas dos embriões em função da raça do animal. As médias
e erros padrões da qualidade dos embriões recuperados estão apresentados na Tabela 3.
Tabela 3 - Médias e erros padrões para qualidade dos embriões recuperados em vacas da raça Nelore superovuladas com FSH associado ou não à rbST
*Médias na mesma linha seguidas da mesma letra não diferem entre si pelo teste de Wilcoxon (P>0,05).
A rbST também não antecipou a
emergência da onda folicular, melhorando a
qualidade do folículo, e, conseqüentemente,
a qualidade embrionária. Neste sentido,
Maffili et al. (2001) relataram que o
uso de rbST, em associação com a
superovulação não aumentou a quantidade
de embriões produzidos, mas melhorou
o desenvolvimento e a qualidade dos
mesmos.
Conclusões
Em vacas da raça Nelore
superovuladas com FSH não se justifica
o tratamento prévio com rbST visando-se
melhoria da resposta ovariana quanto ao
número e qualidade dos embriões.
Referências Bibliográficas
BAUMAN, D.E. Bovine somatotropin and lactation: from basic science to commercial application. Domestic Animal Endocrinology, v.17, n.2-3, p.101-116, 1999.
BORGES, A.M.; TORRES, C.A.A.; RUAS, J.R.M. et al. Resposta superovulatória de novilhas mestiças holandês-zebu tratadas com somatotropina bovina recombinante (rbST). Revista Brasileira de Zootecnia, v.30, n.5, p.1439-1444, 2001.
BREUEL, K.F.; BAKER, R.D.; BUTCHER, R.L. et al. Effects of breed, age of donor and dosage of follicle stimulating hormone on the superovulatory response of beef cows. Theriogenology, v.36, n.2, p.241-255, 1991.

171
Rev. Cient. Prod. Anim., v.10, n.2, 2008
COELHO, E.N. Alguns aspectos da transferência de embriões em bovinos. Belo Horizonte: Escola de Veterinária da UFMG, 1986. 83p. Dissertação (Mestrado em Medicina Veterinária) - Universidade Federal de Minas Gerais, 1986.
CUSHMAN, R.A.; SOUZA, J.C.; HELGPETH, V.S. et al. Effect of longterm treatment with recombinant bovine somatotropin and estradiol on hormone concentrations and ovulatory response of superovulated cattle. Theriogenology, v.55, n.7, p.1533-1547, 2001.
DE LA SOTA, R.L.; LUCY, M.C.; STAPLES, C.R. et al. Effect of recombinant bovine somatotropin (sometribove) on ovarian function in lactating and nonlactating cows. Journal of Dairy Science, v.76, n.4, p.1002-1013, 1993.
DIRKSEN, G.; GRUNDER, H.; STOBER, M. et al. Exame clínico dos bovinos. 3.ed. Rio de Janeiro: Guanabara Koogan, 1993. p.72.
GONG, J.G.; BRAMLEY, T.A.; WILMUT, I. et al. Effect of recombinant bovine somatotropin on the superovulatory response to pregnant mare serum gonadotropin in heifers. Biology of Reproduction, v.48, p.1141-1149, 1993.
GONG, J.G.; BRAMLEY, T.A.; WILMUT, I. et al. Pretreatment with recombinant bovine somatotropin enhances the superovulatory response to FSH in heifers. Theriogenology, v.45, n.3, p.611-622, 1996.
HASLER, J.F.; McCAULEY, A.D.; SCHERMERHOM, E.C. et al. Superovulatory responses of Holstein cows. Theriogenology, v.19, n.1, p.83-99, 1983.
KENNEDY, L.G., BOLAND, M.P., GORDON, I. The effect of embryo quality at freezing on subsequent development of thawed cow embryos. Theriogenology, v.19, n.6, p.823-832, 1983.
KIRBY, C.J.; SMITH, M.F.; KEISLER, D.H. et al. Follicular function in lactating dairy cows treated with sustained-release bovine somatotropin. Journal of Dairy Science, v.80, n.2, p.273-285, 1997.
KUEHNER, L.F.; RIEGER, D.; WALTON, J.S. et al. The effect of a depot injection of recombinant bovine somatotropin on follicular development and embryo yield in superovulation Holstein heifers. Theriogenology, v.40, n.5, p.1003-1013, 1993.

172
Rev. Cient. Prod. Anim., v.10, n.2, 2008
LINDNER, G.M.; WRIGTH, R.W. Bovine
embryo morphology and evaluation.
Theriogenology, v.20, n.4, p.407-416,
1983.
LUCY, M.C. Regulation of follicular
growth by somatotropin and insulin-like
growth factors in cattle. Journal of Dairy
Science, v.83, n.7, p.1635-1647, 2000.
LUCY, M.C.; COLLIER, R.J.;
KITCHNELL, M.L. et al.
Immunohistochemical and nucleic
acid analysis of somatotropin receptor
populations in the bovine ovary. Biology of
Reproduction, v.48, p.1219-1227, 1993.
LUCY, M.C.; TATCHER, W.W.; COLLIER,
R.J. et al. Effects of somatotropin the
conceptus, uterus, and ovary during
maternal recognition of pregnancy on
cattle. Domestic Animal Endocrinology,
v.12, n.1, p.73-82, 1995.
MAFFILI, V.V.; TORRES, C.A.A.;
BORGES, A.M. et al. Efeitos da
somatotropina bovina na resposta
superovulatória e na taxa de clivagem de
zigotos de camundongos (Mus musculus).
Revista Brasileira de Reprodução
Animal, v.25, n.3, p.377-379, 2001.
MOREIRA, F.; BADINGA, L.; BURNLEY, C. et al. Bovine somatotropin increases embryonic development in superovulated cows and improves post-transfer pregnancy rates when given to lactating recipient cows. Theriogenology, v.57, n.4, p.1371-1387, 2002.
NAGANO, A.Y.; WEISS, R.R.; BÜCHELE, J.M. et al. A somatotropina bovina recombinamento (rbST) na superovulação de fêmeas bovinas. Archives of Veterinary Science, v.9, n.2, p.101-106, 2004.
NEVES, E.F.; RAMOS, A.F.; MARQUES JÚNIOR, A.P. Pré-tratamento com somatotropina bovina (rbST) na superovulação de doadoras da raça Holandesa. Arquivos Brasileiro de Medicina Veterinária e Zootecnia, v.57, n.2, p.205-209, 2005
NÚCLEO GEOAMBIENTAL DA UNIVERSIDADE ESTADUAL DO MARANHÃO (NUGEO/UEMA). O Clima da Região. 2004. Disponível em: <http://www.labgeo.uema.br/>. Acessado em: 19 Janeiro. 2008.
RIEGER, D.; WALTON, J.S.; GOODWIN, M.L. et al. The effect of co-treatment with recombinant bovine somatotrophin on plasma progesterone concentration and number of embryos collected from superovulated Holstein heifers. Theriogenology, v.35, n.5, p.863-868, 1991.

173
Rev. Cient. Prod. Anim., v.10, n.2, 2008
SAS INSTITUTE. SAS/STAT software: changes and enhancements trhough release 6.12. Cary: Satatistical Analysis System Institute, 1997.
SEGERSON, E.C; HANSEN, T.R.; LIBBY, D.W. et al. Ovarian and uterine morphology and function in Angus and Brahman cows. Journal of Animal Science, v.59, p.1026-1046, 1984.





























