Embed Size (px)
DESCRIPTION
第五章 植物体细胞胚胎发生. 第一节 体细胞胚的概念及其发生途径 第二节 人工种子. 第一节 体细胞胚的概念及其发生途径. 一、体细胞胚的概念. 体细胞胚( somatic embryo )又叫胚状体,是指离体培养条件下没有经过受精过程而形成的胚胎类似物。. 胚的发育图. 双子叶植物的胚胎发育过程. - PowerPoint PPT Presentation
Citation preview

第一节 体细胞胚的概念及其发生途径第一节 体细胞胚的概念及其发生途径 第二节 人工种子第二节 人工种子
第五章 植物体细胞胚胎发生第五章 植物体细胞胚胎发生

第一节 体细胞胚的概念及其发生途径第一节 体细胞胚的概念及其发生途径

体细胞胚( somatic embryo )又叫胚状体,是指离体培养条件下没有经过受精过程而形成的胚胎类似物。
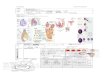
胚的发育图
一、体细胞胚的概念一、体细胞胚的概念

(A) 伸长期合子; (B) 合子经过第一次分裂 , 产生 1 个顶细胞 (绿色 )和一个基细胞 (粉红色 ); (C) 四分体胚 : 顶细胞经历 2 次纵向分裂,产生含有 4 个细胞的胚体和 2 个细胞的胚柄; (D) 16 细胞球形胚 : 8 细胞胚的所有细胞经过 1 次平周分裂 , 产生包含 8 个细胞的表皮原和 8 个细胞的内部组织; (E) 早期心形胚; (F) 心形胚 : 子叶和胚根原基已经出现; (G) 成熟胚胎。 1: 顶细胞 ; 2 : 基细胞 ; 3: 胚根原细胞 , 形成未来的 ROC 和根冠中央区 ; RM: 根尖生长点 ; SM: 茎尖生长点
双子叶植物的胚胎发育过程


花粉培养
单细胞悬浮培养
原生质体培养
组织器官培养
体细胞胚
花粉胚
非合子胚
非合子细胞
合子胚
无融合生殖胚
** 大量研究表明,大多数植物组织培养,单细胞悬浮培养、原生质体培养和花粉培养中都观察到体细胞胚胎发生或花粉胚胎发生( pollen embryogenesis ) ** 前者为二倍体的体细胞产生的胚状结构 ** 后者是由小孢子或其分裂产物等单倍体细胞产生的体细胞胚,通常称花粉胚( pollen embryos ),可发育成单倍体植株。
体细胞胚的产生体细胞胚的产生

** 体细胞胚或花粉胚都是指在植物组织培养中起源于一个非合子细胞,经过胚胎发生和胚胎发育过程形成的胚状结构,包括以下几点含义: 1、体细胞胚是组织培养的产物,区别于无融合生殖的胚,只限于在组织培养范围使用; 2 、体细胞胚起源于非合子细胞,区别于合子胚; 3 、体细胞胚的形成经历胚胎发育过程,区别于组织培养的器官发生中芽与根的分化。

植物体细胞胚胎发生和诱导器官发生相比具有明显的特点:
1、具有两极性 2、存在生理隔离 3、遗传性相对稳定 4、重演受精卵形态发生的特性

二、体细胞胚胎发生的方式 ** 植物组织培养过程中植物体细胞胚胎途径可归纳为两类:直接途径和间接途径。 ** 直接途径是直接从原外植体不经愈伤组织阶段发育而成 ** 间接方式是体胚从愈伤组织或悬浮细胞、有时也从已形成的体胚的一组细胞中发育而成,如香雪兰( Freesia refracta )花序外植体经直接体细胞胚胎发生途径形成再生植株。 ** 外植体形态学极性决定体细胞胚胎发生,所有体细胞胚都出现在花序轴切段的原形态学下端。

1 直接途径 直接发生体胚的来源细胞可以是外植体表皮、亚表皮、幼胚、悬浮培养的细胞和原生质体。 一般认为直接方式发生体胚是由原先就存在于外植体中的胚性细胞――预胚胎决定细胞( PEDCs )培养后直接进入胚胎发生而形成体胚,如柑桔属的珠心组织(在体内或离体情况)通过预胚胎决定细胞直接进行体胚发生的最好例子 .
2 间接途径
间接体细胞胚胎发生中,外植体已分化的细胞先脱分化,并对其发育命运重决定而诱导出胚性细胞――诱导胚胎决定细胞( IEDCs ),进而形成体细胞胚。

1 茎尖培养诱导形成胚性愈伤组织
MS 培养基附加 2.0mg/L 2,4-D
2 胚性愈伤组织悬浮培养增殖
MS 培养基附加 2.0mg/L 2,4-D
3 由细胞集合体形成体细胞胚
MS 培养基附加2.0mg/L 2,4-D
4 体细胞胚萌发并形成小植株 MS 培养基附加1.0mg/L ABA
图 甘薯品种栗子香胚性愈伤组织诱导体细胞胚和再生小植株

胚性愈伤组织胚性愈伤组织 非胚性愈伤组织非胚性愈伤组织
颜色颜色 淡黄淡黄 白色白色表面表面 光滑光滑 粗糙粗糙质地质地 坚硬但易碎坚硬但易碎 松散,水渍状松散,水渍状
胚性细胞的多少胚性细胞的多少 大量大量 极少或无极少或无细胞形态细胞形态 小,圆球形小,圆球形 管状或不规则形管状或不规则形细胞质细胞质 浓厚,染色深浓厚,染色深 稀薄,染色浅稀薄,染色浅内含物内含物 多多 少少
细胞核及核仁细胞核及核仁 核明显,大,核仁核明显,大,核仁多多
核小,核仁少,或核小,核仁少,或无核无核
胚性细胞团特征:成簇、成团,体积小,细胞质致密

三、体细胞胚的起源
** 体细胞胚胎发生途径有直接途径和间接途径,但无论哪一种发生途径,绝大多数研究报道认为体细胞胚胎起源于单细胞 ,从多种植物中都观察到单个胚性细胞,有不均等分裂的二细胞原胚和均等分裂的二细胞原胚、多细胞原胚、球形胚直到成熟胚。也有研究认为起源于多细胞。

** 在同一块愈伤组织或一个胚性细胞复合体上可以观察到多个不同发育时期的体细胞胚,从单个原始细胞到多细胞的原胚、球形胚、鱼雷胚直至具子叶的胚状体,这是由体细胞胚发生和分裂的不同步所致。
香蕉心形体细胞胚 可可心形胚 A和球形胚 B
A
B
1 体细胞胚的单细胞起源

野胡萝卜的胚状体 A. 心形期胚状体 B. 成熟胚状体
C. 早心形期胚状体 D. 两个成熟胚状体共一个胚柄

a) 胚性细胞第一次分裂; b-d) 原胚第一次细胞分裂,鳄梨树形胚柄细胞仍保持单个细胞;
e) 胚柄细胞开始增殖;f-k) 原胚和胚柄发育的不同时期; l)球形体细胞胚。
(ep)= 原胚; (s)=胚柄
图 仙人掌科体细胞胚胎早期发育(由单细胞形成球形胚时期)

a 胚性叶片愈伤组织; b 球形细胞; c 伸长细胞; d 两个细胞时期; e 四个细胞时期; f 原胚; g 球形胚; h 心形胚; i 早期鱼雷胚; j 成熟鱼雷胚; k 不同时期的体细胞胚; l 体细胞胚的萌发; m 再生小植株。
图 木豆幼苗叶片愈伤组织悬浮培养体细胞胚发育过程

** 有的学者观察到体细胞胚来源于多细胞组成的胚性细胞复合体,王亚馥等人也观察到这种胚性细胞团,但用同位素脉冲标记实验即可证明这些细胞团也是由一个单细胞连续分裂而形成,经进一步分化和分裂形成不同发育时期的体细胞胚,因而这些体细胞胚实质也可能是起源于单细胞。当然也不排除有些植物的体细胞胚的确是起源于多细胞,仍有待深入研究。
**总之,体细胞胚胎的发生是一个非常复杂的生理过程,仍有许多问题尚需进一步研究。
2 体细胞胚的多细胞起源

四、体细胞转变为胚性细胞的机制
** 合子胚来源于受精卵;体细胞胚是非合子胚,未经过雌雄配子受精这一过程, Somatic Embryos 又是如何形成?
机制是什么?
** 体细胞胚无论起源于哪一种细胞,都存在着一个共同的问题,即体细胞是如何转变为胚性细胞而形成体细胞胚的。大量研究事实表明,植物细胞在一定培养条件下,具胚胎发生能力可能是一个普遍现象。

1 体细胞胚发生理论—渐变理论
( 1)体细胞胚胎发生实质
** 体细胞胚胎发生实质是细胞分化的问题,而细胞分化的分子基础则是基因差别表达与调控的结果。有关细胞分化的分子机制有许多理论,其中最具普遍性的是渐变理论,认为一个细胞系的细胞质因子间的互相作用是一个原发稳定基因组中基因活化的结果,从而使该细胞系与其它细胞系不同。
外植体 脱分化 愈伤组织细胞分化 胚性细
胞
非胚性细胞
基因调控

( 2)渐变理论认为细胞分化可分为几个阶段? 分为两个阶段: 第一阶段为预定阶段,细胞接受了或预定了特殊发育命运,但并未表现出可见的特化标志,这种特性一旦建立就可以在细胞继代培养中得以稳定保持。 第二阶段是表达阶段,在适宜条件诱导下,预定的细胞( pro-embroygenic determined cells, PEDC )发生一系列生理生化和形态上的变化,表现出分化的特性,但由于这种表达与培养过程中诱导因素关系密切,而表现为一种生理反应,是不稳定的。
( 3)渐变理论的不足 (一)未揭示体细胞胚胎发生的实质,以及细胞的预定性是如何实现的问题。 (二)不能解释经多次继代培养的细胞往往会失去胚胎发生能力,因其理论认为预定性细胞在以后的培养过程中处于稳定状态。 (三)不能解释体细胞胚胎直接发生途径和间接发生途径可因条件不同而相互转化的问题。

**按预决定观点,从外植体组织或细胞直接形成体细胞胚的途径,需要一种诱导物的合成或抑制物的消除,来恢复有丝分裂活动和促进胚胎发育。这时生长素对胚胎发生常起抑制作用,去掉生长素并给以必要的条件就使细胞进入分裂周期,并进行胚胎发生的表达。
** 从外植体经过脱分化形成愈伤组织,间接产生体细胞胚的途径,则需要分化细胞的重新预决定,称为诱导的胚胎预决定的细胞( induced embroygenically determined cells,IEDC ),这时则需要生长素,诱导细胞进入细胞周期。生长素不仅作为使细胞发生分裂所必需的物质,而且也是使细胞进入胚胎状态所必需的因素。
( 4)按预决定观点,生长素对由直接途径和间接途径体细胞胚发生的作用

五、体细胞胚发育成植株的能力
1 体细胞胚发育成植株的能力
** 一般来说胚胎一旦被分化出来或甚至完成诱导之后转移到基本培养基上即可发育成熟,并可形成小植株。
** 如陈东方等( 1986 )对棒头草幼穗培养的研究表明,体细胞胚形成之后,可以在原来培养基上或 MS无任何附加成分的培养基上以及 1/2MS 加 IAA 或 NAA 0.5mg/L 和 6-BA 0.5mg/L 培养基上萌发形成小植株。

1 茎尖培养诱导形成胚性愈伤组织
MS 培养基附加 2.0mg/L 2,4-D
2 胚性愈伤组织悬浮培养增殖
MS 培养基附加 2.0mg/L 2,4-D
3 由细胞集合体形成体细胞胚
MS 培养基附加2.0mg/L 2,4-D
4 体细胞胚萌发并形成小植株 MS 培养基附加1.0mg/L ABA
图 甘薯品种栗子香胚性愈伤组织诱导体细胞胚和再生小植株

2 体细胞胚萌发的两种不正常现象
体细胞胚萌发时观察到两种不正常萌发现象:
** 一种是不完全萌发,即只有芽而无根或只有根而无芽 .
**另一种是早熟萌发,即体胚还未表现出完整的胚结构之前就已萌发,萌发时盾片膨大形成一叶状体,在叶状体基部长出芽。
注意:体细胞胚胎途径的形态发生能力要比器官分化途径保持更长久。

六、影响体细胞胚发生的因素
11 、基因型与体细胞胚发生 、基因型与体细胞胚发生 基因型是影响体细胞胚发生的重要因素,体基因型是影响体细胞胚发生的重要因素,体
细胞胚的诱导率和每个胚性外植体上体细胞胚的细胞胚的诱导率和每个胚性外植体上体细胞胚的发生率因基因型而异。 发生率因基因型而异。

( a )不同基因型体细胞胚诱导率; ( b )不同基因型每个胚性外植体上体细胞胚的数目
图 6.7 在相同培养基上 Feijoa sellowiana Berg 不同基因型体细胞胚诱导 (引自 Miguel 等, 2001 )

22 、植物激素与体细胞胚发生 、植物激素与体细胞胚发生 ** 离体植物细胞在开始往往缺乏合成生长素和细胞分裂素的能力,但是在大多数情况下,这些细胞的分裂和分化以及形态建成过程中又必需生长素和细胞分裂素两种激素的共同作用。
** 在培养介质中添加不同种类或不同浓度的外源激素诱导形态发生已受到广泛的重视。
**但最关键的是组织内部和体细胞胚发生部位的内源激素代谢动态和平衡。

(( 11 )内源激素与体细胞胚发生 )内源激素与体细胞胚发生 早在胡萝卜细胞悬浮培养体细胞胚发生中发现, 2, 4-D 在诱导胚性细胞
早期是必需的,而且 2 , 4-D 是通过影响 IAA 结合蛋白起作用,其实质是促进 IAA 结合蛋白的形成,提高细胞对 IAA 的敏感性从而诱导胚性细胞的形成。
水稻细胞培养中也发现,由于 2,4-D促进细胞内源 IAA 含量提高进而诱导胚性细胞的形成,并认为内源 IAA 含量上升或维持在较高水平,是胚性细胞出现的一个共同标志。
一些研究表明, ABA 对植物体细胞胚的发生与发育具有重要作用。 外源 ABA 与内源 ABA 对体细胞胚发生起到相互调节和促进的作用,而且通
过补充提高外源 ABA ,可以明显提高体细胞胚发生的频率与质量。 荔枝体细胞胚胎发生早期的内源激素 ABA 与 IAA 和 ABA 与 CTK(细胞分裂素)的质量比增大,将有利于荔枝体细胞球形胚的形成。

(( 22 )外源激素与体细胞胚发生 )外源激素与体细胞胚发生 ① 2 , 4-D 与体细胞胚发生 2 , 4-D 是诱导多种植物离体培养体细胞转变为胚性细胞的重要激素。
单子叶植物和双子叶植物诱导体细胞胚胎发生时,所要求的 2,4-D 的浓度不同,对单子叶植物而言,一般要求较高浓度的 2,4-D ,其浓度范围为 0.5-5mg/L ,通常使用浓度为 2mg/L ;
而对双子叶植物而言,一般要求较低浓度的 2,4-D ,其浓度范围为 0.02-1mg/L ,通常使用的浓度为 0.1mg/L 。
在体细胞胚胎发生的诱导阶段,培养基中必需添加 2,4-D ,而在体细胞胚胎形成阶段,要降低培养基中 2,4-D 的浓度或去除 2,4-D ,以保证体细胞胚的正常生长。
在胡萝卜细胞培养中,加入 2, 4-D 后诱导一些特异性多肽或蛋白质形成,当去除生长素后,这些多肽和蛋白质也随之消失。但在诱导出胚性细胞后,及时除去培养基中的 2, 4-D ,胚性细胞在进一步分化和发育的同时释放糖蛋白 GP65( 65kDa )

表 在 MS固体培养基上不同浓度 2,4-D 对木豆幼苗叶片外植体诱导和原胚细胞形成的影响
2,4-D(μM) Callus induction(%)
Proembryonic cell*(%)
0.00 0.0 0.0
2.26 3.2d 3.4d
4.52 7.4c 10.2c
6.78 38.6a 37.4a
9.04 18.2b 12.8b
11.30 3.6c 2.2c

表 在具有不同浓度 2,4-D 的 MS液体培养基上木豆不同时期体细胞胚诱导率
2,4-D(μM)
收集细胞的体积 (10th day)
体细胞胚发育时期
球形胚 (%) 心形胚 (%) 鱼雷胚 (%)
0.00 0 0 0 0
1.12 2.8e 2.4e 1.8e 1.2e
2.26 3.2d 7.6d 2.4d 1.6d
3.38 4.6c 20.2b 8.2b 5.2b
4.52 5.2b 36.2a 20.4a 16.4a
5.64 5.6b 18.6c 6.6c 3.6c
6.78 6.2a 7.4d 1.6e 1.2e

Origin of embryoids in embryo-derived callus culture. (a) Single cells (arrow) and two-celled stage (double arrows). x137. (b)Four-celled stage (arrowhead). x137. (c) Multicelled proembryo containing eight to ten cells. x137 (d) Epidermized lobular
stage. x34. (e) Heart shape embryoid that remain in the callus. X34. (f) Several compact embryoids in different stages of formation. x14.

A,C,E 为离体培养 6 周后的体细胞胚; B,D,F 为离体培养 5个月后形成的体细胞胚
A:品种 GK7 ,在添加 83.0 mM centrophenoxine培养基上 ; B: 品种 AT120 ,在添加 124.4 mM centrophenoxine 培养基上 ; C: 品种 59-4144 ,在添加 12.4 mM dicamba 培养基上 ; D:品种 AT120 ,在添加 83.0 mM dicamba培养基上 ; E: 品种 VC1 ,在添加 83.0 mM picloram 培养基上 ; and F:品种 VC1 ,在添加 124.4 mM picloram 培养基上 .
图 含不同种类和不同浓度生长素固体培养基上成熟花生胚轴体细胞胚诱导

② 其它生长激素与体细胞胚发生 除 2,4-D 外, ABA 对植物体细胞胚的发生也起到重要的调
控作用。 研究表明, ABA 具有促进苈蒿体细胞胚结构正常化的显著
效应。 在浓度适宜时, ABA 能抑制多种异常体细胞胚的发生,而
且 ABA 加入培养基的时间愈早其效应也越明显。 较低浓度的 ABA 对保持籼稻愈伤组织的胚性结构具有重要
意义,可明显提高这种胚性愈伤组织分化出苗率。 在拐芹中 ABA 与 NAA 共同作用可诱导一些游离氨基酸含
量的变化和特异蛋白的形成。 ABA 和 PEG 对胡萝卜 (Daucus carota L.) 体细胞胚发生具
有调控作用 大量的研究表明,大量的研究表明, ABAABA 对某些植物体细胞胚胎发生特异性对某些植物体细胞胚胎发生特异性
基因的表达起调控作用,激活相关基因的表达,合成贮藏蛋基因的表达起调控作用,激活相关基因的表达,合成贮藏蛋白、晚期胚胎发生丰富蛋白和胚胎发生的特异性蛋白。白、晚期胚胎发生丰富蛋白和胚胎发生的特异性蛋白。

3 多胺(腐胺、尸胺、亚精胺、高亚精胺、精胺等)和乙烯与体细胞胚发生
** 外源多胺对体胚发生的作用除了取决于植物种类及其内源多胺的状况之外,还关系到它们如何被吸收、运输及降解等问题。多胺与体细胞胚发生的关系在胡萝卜的研究中较多。

举例说明: ** 已发现胡萝卜的胚性细胞中腐胺、精胺含量比非胚性的高,在胡萝卜体细胞胚发生的前胚时期多胺含量一般较低,从球形胚、心形胚到鱼雷胚时期,精胺和亚精胺逐渐升高,心形胚时期以腐胺为主,鱼雷胚时期则富含亚精胺。 ** 在胡萝卜愈伤组织的体细胞胚发生中发现乙烯抑制体细胞胚的发生,而乙烯形成酶抑制剂 Ni2+、 Co2+可以促进其体细胞胚发生。 ** 有研究表明,只在胚细胞分化早期才与多胺生物合成关系较密切。

4 糖类和金属离子与体细胞胚发生
① 糖类
**糖类对诱导植物体细胞胚发生也是不可缺少的重要成分。
**研究表明,小麦的愈伤组织一旦分化为胚性细胞后就有淀粉粒的积累,在胚性细胞分化与发育的整个过程中,淀粉的两次合成高峰均在发育的重要转折期,为体细胞胚的进一步发育和分化提供必要物质和能量基础。淀粉的积累与胚性细胞分化能力和体细胞胚发育时期的转折密切有关。
** 在茴香的胚性细胞中也富含淀粉粒,在体细胞胚发育中同样出现两次峰值,同时认为淀粉出现的峰值与核酸和蛋白质合成密切相关,因此,表明淀粉为蛋白质和核酸的合成奠定了物质基础。

碳水化合物 浓度( mM) 胚胎发育时期 球形胚(%)
心形胚(%)
鱼雷胚(%)
葡萄糖 Glucose
55.49 2.4f 1.2h 1.2g 110.99 6.8f 2.6h 2.4f 166.48 10.2e 5.0f 4.2e 221.98 13.6c 7.2d 6.4d
果糖
Fructose
55.49 6.2f 2.8g 2.4f 110.99 12.4c 5.2f 4.6c 166.48 6.4f 5.0f 4.2e 221.98 2.4h 1.2h 1.2g
蔗糖
Sucrose
29.21 12.4c 6.8c 6.2d 58.43 20.2b 16.2b 14.6b 87.64 36.2a 20.4a 16.4a 116.86 12.6d 10.2c 8.4c
麦芽糖 Maltose
27.75 2.4h 1.2h n.d 55.51 5.4g 2.6g 1.2g 83.26 1.2i n.d n.d 111.02 n.d n.d n.d
表 在 4.52μM/L 2,4-D 培养基上不同碳水化合物
对木豆愈伤组织悬浮培养体细胞胚发生的影响

② 金属离子 **金属离子对体细胞胚的发生也有影响。 ** 在小麦愈伤组织诱导体细胞胚发生中也发现 Zn2+、Cu2+、 Co2+、 Mn2+等金属离子具有重要的促进作用,不仅可提高诱导频率,而且还可以加速胚性愈伤组织的形成,因此,在培养基中加入适合浓度金属离子可提高体细胞胚发生的频率。 ** 有研究表明,金属离子可能是通过促进多胺合成而提高体细胞胚发生频率。

5 外植体与体细胞胚发生
不同基因型、外植体体细胞胚发生能力不同。如
胡萝卜子叶不同部位培养诱导的发育过程不同。只有
表皮下细胞具有真正细胞全能性和有能力直接产生体
细胞胚(胚胎发生区),而不用通过愈伤组织阶段的
诱导。

表皮下细胞 根发生区细胞 芽发生区细胞
胡萝卜子叶不同部位外植体示意图

6 光与体细胞胚发生
品种 Jack(光照强度50-60 µE m2 s-1 )
品种 Fayette(光照强度 50-60 µE m2 s-1 )
品种 Jack(光照强度 5-10 µE m2 s-1 )
品种 Fayette(光照强度 5-10 µE m2 s-1 )
(图 光对大豆不同品种子叶体细胞胚发生的影响 )

第二节 人工种子
一、人工种子的概念
人工种子:又称合成种子或体细胞种子,是指将植物离体培养的胚状体或芽包裹在含有养分和保护功能的人工胚乳和人工种皮中的类似种子的颗粒。
** 人工种子可在一定条件下萌发生长,形成完整的植株。

人工种子
依靠植物细胞“全能性”,美国首先发明“人工种子”

人工种子 www.agr.hokudai.ac.jp 不定胚形成人工种子 www.brs.nihon-u.ac.jp
人工种子大量生产 kankyo08.en.a.u-tokyo.ac.jp
人工种子结构示意图

二、人工种子研究历史
**1978年 Murashige提出人工种子概念
**1986年 Redenbaugh 等成功地利用藻酸钠包埋单个体细胞胚,生产人工种子。
**胡萝卜、棉花、玉米、甘蓝、莴苣、苜蓿等人工种子制作获得成功。
** 在我国人工种子的研究已列入重点攻关项目,并已取得一些重要的研究成果。

三、人工种子各部分功能 1 体细胞胚 为了使人工种子在一定条件下萌发生长,并形成完整植株,
人工种子首先应该具备一个能够很好地发育成完整植株能力的胚,它具有胚根和胚芽的双极性,经原胚、球形胚、心形胚、鱼雷胚和子叶形胚等不同阶段发育而成。
2 、人工胚乳 人工胚乳为体细胞胚萌发提供营养物质。根据使用者的目的,
可向人工胚乳内自由添加有利于胚状体正常萌发所需的各种营养成分、防病虫物质、植物激素等,也是人工种子优越于天然种子之处。
33 、人工种皮 、人工种皮 包裹在人工种子的最外层,对人工种子起到保护作用,保护包裹在人工种子的最外层,对人工种子起到保护作用,保护
人工种子内部的水分和营养不致丧失和防止外部物理冲击,人工种子内部的水分和营养不致丧失和防止外部物理冲击,还要保证通气。 还要保证通气。

四、人工种子种类 根据包裹体细胞胚材料的不同,可以将人工种子分为以
下 4 种类型( Redenbaush等, 1991 )。 ( 1 )裸露的或休眠的繁殖体,可以直接播种,如休眠的微鳞茎和微型薯等。
( 2 )人工种皮包裹的繁殖体。 ( 3 )水凝胶包埋的胚状体、不定芽以及休眠的小鳞茎等繁殖体,水凝胶中可含多种养分和激素,促进繁殖体的生长。
( 4 )液胶包埋的繁殖体。

五、人工种子与天然种子比较有其自身的特点: ( 1)可对一些自然条件下不结实的或种子很昂贵的植物进行繁殖。 ( 2)固定杂种优势,使 Fl 杂交种可多代利用,使优良的单株能快速繁殖成无性系品种,从而大大缩短育种年限。 ( 3)节约粮食。 ( 4)在人工种子的包裹材料里加入各种生长调节物质、菌 肥、农药等,可人为地影响控制作物生长发育和抗性。 ( 5 ) 可以保存及快速繁殖脱病毒苗,克服某些植物由于长期营养繁殖所积累的病毒病等。 ( 6 ) 与试管苗相比成本低,运输方便 ( 体积小 ) ,可直接播种和机械化操作。

六、目前人工种子涉及的问题 1 、高质量和数量体细胞胚的诱导。 2、体细胞胚发生的同步控制。 3、人工种皮的制造与生产。 4、人工胚乳的研制。 5、工艺流程。 6、贮藏。
**获得体细胞胚是生产人工种子的基础,其发生频率
的高低、胚的质量和体细胞胚发生的同步控制是生产
人工种子的关键。

*** 人工种子制备包括胚状体的诱导与同步化、人工种子的包埋、人工胚乳的制作、人工种皮的制作、人工种子贮藏等步骤内容。
1 、高质量和高产量的体细胞胚诱导与同步化控制
体细胞胚的诱导与同步化控制方法:
七 人工种子的研制

手工选择:无菌条件下逐个筛选。 过筛选择:胚性细胞悬浮培养液分别通过不同
孔径滤网过筛、培养、再过滤。 不连续密度梯度离心:通过 Fi col l 不同浓度
产生不同密度梯度溶液,对不同比重细胞进行筛选。
渗透压分选:基于体细胞胚不同发育阶段渗透压明显变化,选择比较一致的同一发育时期的体细胞胚。
植物胚性细胞分级仪筛选:根据不同发育阶段体细胞胚比重不同,通过筛选液(一般为2%蔗糖,进样速度 15mL/mi n,分选液流速 20mL/mi n)经数分钟分级筛选,以获得一定纯化的成熟胚。
低温处理同步化:冷处理或与营养饥饿相结合,以增加细胞同步化。
饥饿法:除去悬浮细胞生长所需基本成分,导
致进入静止生长期,然后恢复营养,以促进细胞同步化。
阻断和解除法:适当阻断细胞循环过程,使培养细胞同步化,然后解除阻断,以控制体细胞胚发育同步化。
有丝分裂阻止:利用秋水仙碱抑制细胞有丝分裂,以达同步化,但处理时间不宜过长,避免引起不正常有丝分裂。
化学方法
物理方法
体细胞胚诱导与同步化控制
体细胞胚诱导与同步化控制

a 成熟的体细胞胚; b 用无菌吸管吸取与包埋剂混合的体细胞胚 c 带体细胞胚的液滴滴入 2% CaCl2溶液后形成的白色半透明的包埋丸; d 人工种皮包埋制成的人工种子
人工种子的制
备

苜蓿体细胞胚的生产与植株的获得www.plant.uoguelph.ca/research/embryo/embryo.htm

2 、人工种子包埋 获得发育正常、形态上较为一致的体细胞胚后,就要用适合的材料进行包埋。进行包埋时应做到以下几点: ( 1)包埋材料必须对体细胞胚无损害和无毒性,并能保护胚和胚发芽的能力; ( 2)同时在制造、贮藏、运输和种植过程中必须具有耐久性; ( 3)胶囊必须含有养分和植物激素以及植物生长和发育所需的成分和水分,并要求制作的人工种子适合于农业机械化播种。

3 包埋方法: 经研究认为藻酸盐所形成的胶囊是目前最好的凝胶包埋材料,所采用的主要方法是滴注和装膜两种。
( 1 )滴注法 制备人工种子是将选择好的体细胞胚悬浮于含 2%~ 3%的藻酸钠溶液中,然后用一个塑料吸管吸注含体细胞胚的藻酸钠悬滴加入到 0.1mol/LCaCl2溶液中,这时离子间发生交换而形成胶囊,胶囊的直径依赖于吸管的内径和吸注的速度,胶囊的大小,视体细胞胚的大小和发芽能力而定,一般采用吸管口径为 3- 4mm ,可得直径为 4- 5mm 的胶囊。体细胞胚包在胶囊中的位置不宜处在中央,宜偏在一边以利萌发。至于聚合时间长短则依赖于所用试剂浓度。一般藻酸钠的浓度控制在 2%- 3% ,在 CaCl2 中停留实践不要超过 10min ,以免胶囊太硬而影响发芽。形成胶囊后转入无菌水中冲洗,最后在 1/2MS培养基上作发芽和转换试验或进一步包裹人工种皮。

滴注法

人工种子包埋过程示意图

( 2)装膜法 制备人工种子是把体细胞胚混合到一个温度较高的胶液中,然后滴注到一个有小坑的微滴板上,随着温度降低变为凝胶而形成胶丸。

4 、人工种皮的研制 美国杜邦公司生产的 Elvax4260 是一种较好的“人工种皮”材料,是由乙烯、乙酸和丙烯酸三种物质共聚的产物,用它作为人工种子最外层的包裹剂,即人工种皮取得较好效果 。

2 、用 Elvax作为人工种皮的具体操作程序为: ( 1)、将 4g藻酸钙胶囊置于 20mL 含 0.1g葡萄糖和含 0.2mL甘油的氢氧化钠溶液中搅拌 1min; ( 2)、在 50mL环乙烷中加入 10%的 Elvax4260; ( 3)、在 40℃条件下溶解 5g硬脂酸, 10g鲸醋醇和 25g鲸醋取代物,另加 295mL石油醚和 155mL 二氯甲烷; (4) 、 将藻酸钙胶囊放在上述的混合液中浸泡 10s ,取出后用热风吹干。如此重复 4- 5次,最后用石油醚冲洗干净,经尼龙布过虑后在空气中风干即可。

体细胞胚与人工种子

1 、人工种子转换定义
**转换( Transformation)指人工种子在一定条件下,萌发、生长、形成完整植株的过程。
2、转换方法
**转换的方法可分为无菌条件下的转换和土壤条件下的转换。
八、人工种子的转换试验

( 1 )、无菌条件的转换:其也称离体条件的转换。一般是将新制成的人工种子播种在 1/4MS 培养基,附加 1.5% 麦芽糖、8g/L 的琼脂中,培养后统计人工种子形成完整植株的数目,即人工种子的转化率。
( 2)、无菌条件转换率的高低主要取决于:
( a)、对提高体细胞胚质量的培养基成分的研究;
( b)、改进转换条件的研究。如麦芽糖代替蔗糖,有利于体细胞胚的萌发和转换。

体细胞胚发育成植株,大致经历几个阶段:
发芽 根系的发育 芽分生组织的生长和发育
真叶的生长 芽和根的连接 正常植株生长
兰花人工种子的生产及其获得的小植株

( 3 )、土壤条件的转换:也称活体条件下的转换。人工种子真正目的是直接播种于土壤,即在活体条件下,使转换成功。 ( 4 )土壤条件下转换采用方法: a 、无土培养试验:目前以蛭石或珍珠岩试验较多,附加低浓度无机盐, 1/6 的 MS 培养基, 0.75%麦芽糖有利于转换。 b 、土壤试验:人工种子的土壤转化试验报道较少。苜蓿人工种子的直接土壤转化试验已达 20% ,水稻不定芽研制的人工种子经适当地过渡性培养,在土壤中的转化率为 10% 。 ( 5 )、土壤条件的转换人工种子转化率低的主要限制因子 a 、研究无机盐在转换中的重要性时表明,没有硝酸钾、硫酸镁、氯化钙时就不会发生转换,缺乏磷酸铵,转化率从 30%降至 5% 。显然这些无机盐成分作为人工种子营养肥料是不可缺少的。 b 、 0.75%( W/V )的麦芽糖有利于提高转化率。因此推测,碳源在人工种子转换中也是限制因子。 因此,必须在人工种子中贮藏必需的养分或供给外源营养物质。

九、人工种子的储藏与萌发 人工种子储藏时间的长短和萌发率的高低对人工种子的应
用至关重要。人工种子的寿命,其最理想的状态是能够与真正的合子胚种子的寿命一样长,只有这样,才有利于人工种子的储藏和萌发。
此外,海藻酸盐胶囊本身又抑制呼吸,更加缩短了体细胞胚的寿命。因此,储藏和萌发是人工种子研制的主要难点之一。
延长人工种子储藏时间和提高其萌发率的主要方法包括低温法、干燥法、抑制法、液体石蜡法等以及这些方法的组合。
干燥法和低温法是目前应用最多的方法。

( 1 )低温储藏技术 低温储藏是指在不伤害植物繁殖体的前提下 , 通过降低温度来降低繁殖
体的呼吸作用 , 使之进入休眠状态。 常用的温度一般是 4℃ 。在此温度下体细胞胚人工种子可以储存 1-2
个月。 如茶枝柑 ( C itrusreticu la ta 的人工种子 ,储存 1 个月仍具很高转化率。泡桐 ( Pau low n ia elonga ta 的人工种子在储藏 30 d 或 60 d 后体细胞胚的存活率分别是 67. 8% 和 53. 5% ,萌发率分别是 43. 2% 和 32. 4% 。
非体细胞胚人工种子可以在 4℃ 下储藏更长的时间 ,Mandal等 ( 2000 对4 种罗勒属 ( Ocim um ) 的植物腋芽进行包被 , 可保存 60 d, 并使植物耐寒性提高。
Datta 等 ( 1999 包被濒危植物地宝兰 ( Geodorum densif lorum ) 类原球体 ,储藏 120 d 后 , 生存能力与储藏前相比无明显变化。
马铃薯芽尖在 MS 培养基上培养 2 d 后 , 用海藻酸钠包被 ,储藏 270 d 、360 d 和 390 d 后在 MS 培养基上的萌发率分别是 100% 、 70. 8% 和 25% 。
由于人工种子没有像自然种子一样在储藏前进入休眠状态 ,随着低温储存时间的加长 , 包被体系内的含氧量降低 , 人工种子萌发率会下降

( 2 )液体石蜡储藏技术 液体石蜡作为经济、无毒稳定的液体物质 ,常被用来储藏细菌、真菌
和植物愈伤组织。美国已有报道 ,把人工种子放在液体石蜡中 ,保存时间可达 6 个月以上。
李修庆等 (1990) 研究胡萝卜人工种子的结果表明 : 人工种子在液体石蜡中短时间保存 ( 1 个月 )能较正常的生长 ,但时间一长 (79 d) , 人工种子苗的生长则明显比对照组差 ; 并发现液体石蜡对幼苗的呼吸和光合作用有一定的阻碍作用。
在常温下液体石蜡不能通过抑制发芽来储藏人工种子。干燥后的人工种子 , 在 2℃ 的液体石蜡中 , 2 个月后只有 2%萌发。
陈德富等 ( 1990 ) 对根芹 ( A piumg raveolens ) 体细胞胚人工种子的研究也得到同样结果。

( 3 ) 超低温储藏技术 超低温一般是指 - 80℃ 以下的低温 , 如超低温冰箱 ( - 80~ - 150
) ℃ 、液氮 ( - 196 )℃ 等。 在此温度下 , 植物活细胞内的物质代谢和生命活动几乎完全停止。
所以 , 植物繁殖体在超低温过程中不会引起遗传性状的改变 , 也不会丢失形态发生的潜能。
目前应用于人工种子超低温保存的方法主要是预培养 2干燥法 ,即人工种子经一定的预处理 , 并进行干燥 ,然后浸入液氮保存。
脱落酸 (ABA) 是一种植物逆境激素 , 可以诱导产生蛋白质、多胺、脯氨酸和淀粉等物质的增加 , 使体细胞胚在干燥处理中受到保护。
Nieves等 ( 2001) 对甘蔗晚盾片期胚用 3. 8μmol/L 的 ABA 或 4. 7μmol/L 的茉莉酸 ( jasmonic acid, JA)处理后进行包被 , 并在干燥器中用无水硅酸进行干燥处理 , 使其水含量降至 60% 或 30% 。结果发现存活的体细胞胚是由 ABA处理的胚 , 而用 JA处理不能达到此效果。
对于非体细胞胚人工种子 , Khor 等 ( 1998 ) 对龙舌兰原球茎用 10- 5M 的 ABA 处理 , 经干燥脱水与包埋 , 也可获得极高的存活率。

( 4)干燥法 干化的人工种子更像天然种子,具有更长的储藏寿命。 Gray等( 1987)在 23℃条件下干化鸭茅体细胞胚 21天,萌发率达到 12%。 Hammatt等( 1987)把大豆体细胞胚干化到原体积的 40%-50%后再吸水,萌发率仍达到 31%。干化增强人工种子幼苗的活力,可能与其超氧化物酶和过氧化物酶的活性显著提高,从而减轻低温贮藏对体细胞胚的伤害有关。

1 、人工种子的应用目前存在的问题
( 1 )许多有价值的基因作物,特别是那些不易获得种子和自然增殖率极低的植物,目前尚未显示出较好的体细胞胚发生能力,或胚的生活力差;
( 2 )有些作物虽然可以诱导出体细胞胚,但胚胎发生机制研究不够,而且同步化等技术尚未解决,因而难以达到大量控制同步化的目的;
( 3)人工种子的成本远高于自然种子;
( 4 )人工种子贮藏、运输和加工以及机械化播种,尚未完全解决,许多问题尚待进一步研究和探索。
十、 人工种子应用前景

2 、人工种子应用应该考虑哪些方面 要想人工种子技术得到应用: ( 1 )首先应考虑该作物的价值; ( 2 )要考虑那些很难获得种子,而且自然增殖率极低的植物。 例如:多年生名贵药材,高质量的木材和橡胶树或优质果树以及珍惜濒危植物等。3 、人工种子发展前景 ( 1 )人工种子的发展前景是客观的 ( 2 )它将是农业中一种革命性的繁殖系统 ( 3 )相信该技术会得到逐步完善,建立多种模式来研制人工种子,并逐步扩大应用范围。