Embed Size (px)
Citation preview

СОФИЙСКИ УНИВЕРСИТЕТ „СВ. КЛИМЕНТ
ОХРИДСКИ”
Билогически факултет
Катедра „Зоология и aнтропология”
ДИПЛОМНА РАБОТА
Фаунистични изследвания върху водните кончета (разред Odonata) от
района на Чокльово блато, Конявска планина
Ръководител катедра:
Доц. д-р Даниело Пешев
Научен ръководител:
Доц. д-р Пламен Митов
Дипломант:
Белчо Христов
Фак. номер: 20 094
София 2006 г.

- 2 -
СЪДЪРЖАНИЕ
I. Увод................................................................................................................................3 стр.
II. Преглед на досегашните проучвания за разред Odonata в района на
Югозападна България и Чокльово блато (Конявска планина) ...........................7 стр.
III. Цел и задачи …………………………….................................................................8 стр.
IV. Характеристика на Чокльово блато.....................................................................9 стр.
V. Материал и методи..................................................................................................21 стр.
VI. Резултати и обсъждания........................................................................................23 стр.
5.1. Фаунистични изследвания................................................................................27 стр.
5.2 Биологични, екологични и фенологични бележки за Odonata от района на
Чокльово блато................................................................................................................27 стр.
5.3. Зоогеографски анализ........................................................................................49 стр.
VII. Обобщени резултати и изводи……………………........................................…52 стр.
VIII. Приложения…………………..............................................................................53 стр.
IX. Използвана литература.........................................................................................77 стр.

- 3 -
I. Увод
Обект на настоящата дипломна работа са представителите на Разред Odonata
(Fabricius, 1793), известни у нас като ”Водни кончета”.
Водните кончета са едни от най-големите хищници в света на насекомите.
Повечето от тях са разпространени в тропиците, като броят на видовете им
закономерно намалява с приближаване на полярните райони.
Специфичният им начин на живот - тясно свързан с водоеми от различен тип
(езера, блата, торфища, реки, канали и др.), е причина те да се срещат почти навсякъде
по света. Водните кончета притежават изключително добри приспособления за
оцеляване в средата си на живот – аеродинамично тяло; големи фасетни очи,
обхващащи широк зрителен ъгъл; широки криле с отлично развити структури и
механизми за балансиране в полет и покой; яки и силно назъбени челюсти; предна
двойка крака, оформяща т. нар. ”летящ сак” и остри шипове по краката за захващане и
задържане на жертвата. Хищническият им начин на живот ги прави ненадминати
нападатели и унищожители на множество ларви и възрастни на комари, мухи и други
насекоми.
Водните кончета се използват от зоолозите при екологични анализи на редица
хабитати, т. к. привързаноста към водоемите ги прави надежден индикатор за
определяне състоянието на водите (степен на замърсеност, аерираност и други важни
показатели) и на влажните зони като цяло.
Досега от разреда в света са установени 5521 вида, групирани в 630 рода и 34
семейства. От тях в България се срещат 68 вида (10 семейства, 28 рода), като за нашата
фауна се очакват още 7 - 8 вида, които вероятно използват територията на нашата
страна като място за размножаване или по време на миграция (Маринов, 2003).
Най-големите бели петна в проучванията на одонатите у нас са планинските райони
по протежение на цялата ни западна граница, голяма част от Северна България, както и
Горно-Тракийската низина (Маринов, pers. com., Хубенов, 2005).
Всичко това, както и слабото познаване одонатната фауна на торфищата в
България ни накара да насочим нашето внимание към изследването на един слабо
проучен по отношение на безгръбначната фауна район. А именно Чокльово блато,
Конявска планина. Изборът му, беше определен и от обстоятелството, че то място с

- 4 -
висока орнитологична стойност и е обявено за „Защитена местност” (ДВ:
1022/30.12.1992 г.) поради две причини: 1) намира се на втория по големина
миграционен път в Европа за птиците – Via pontica; 2) в ливадите около блатото гнезди
глобално застрашеният от изчезване вид ливаден дърдавец - Crex crex.
В тази връзка натрупаните фаунистични, биологични и екологични данни за
водните кончета в района ще могат да се използват в бъдеще като база за сравнение и
анализ на антропогенните въздействия и за установяване на промени в статуса на
групата.
Влажните зони са сред най-важните природни територии на Планетата. Ето защо
въпросът за тяхното опазване и съхранение стои и в екологично отношение, и като
социално значим елемент за запазване на връзката между човека и естествената
природна среда (Lakatos, 1995).
.

- 5 -
Благодарности
За осъществяването на настоящата дипломна работа съм особено задължен на
дипломния ми ръководител доц. Пламен Митов. Благодаря му на първо място за
доверието, което ми гласува и за удоволствието, което изпитах при изследването на
тази интересна група безгръбначни животни. Благодаря му за напътствията, за
коректността и за неоценимата му помощ при цялостното разработване на настоящата
дипломна работа.
Особено признателен съм и на д-р Милен Маринов (гр. София). Без неговата подкрепа,
предоставена литература, съвети и насоки, тази работа не би била факт.
Сърдечна благодарност дължа на колегите доц. Спасимир Тонков, доц. Мая Стойнева и
доц. Димитринка Стоянова (катедра „Ботаника” БФ) за предоставената литература по
темата.
Благодаря и на колектива на катедра „Хидробиология” за предоставения ми достъпът
до непубликувани данни във дипломните работи на студентите защитили в тази
катедра.
Дълбока благодарност дължа и на Ръководството на кат. „Зоология и антропология” при
Биологическият факултет на СУ "Св. Климент Охридски", в лицето на неговият ръководител
доц. Даниело Пешев за предоставените ми пълни възможности и добри условия за работа.
Признателен съм на целият екип от Варненския Природонаучен музей, където
понастоящем работя, и особено на неговия директор ст. н. с. д-р Янаки Сивков за
тяхната подкрепа, разбиране, ценни съвети и творческа среда, при написването на
настоящата работа. Благодарен съм им и за ценните съвети, които ми дадоха при
написването на настоящата работа..
Благодарен съм и на всички мои колеги, които по време на работата ми по темата ми
помагаха, подкрепяха морално и се отнесоха с разбиране и търпение. Това са маг.

- 6 -
Атанас Грозданов, доц. Венцислав Делов, д-р Любомир Запрянов, Теодора Пеева и
Велин Раденков.
Накрая, но не на последно място, бих искал да благодаря на моите родители доц.
Симеон Христов и доц. Татяна Христова, както и на моя брат маг. Тодор Христов,
които, нито за миг не престанаха да ме подкрепят и силно вярваха в мен, в моите цели,
желание и успехи, които като студент и възпитаник на Биологически факултет на
Софийския Университет си поставих и следвах.

- 7 -
II. Досегашни изследвания върху водните кончета на България
Изследванията върху одонатите в България започват в края на ХІХ век.
Публикуваната до този момент информация за видовете у нас може да се открие в
около 140 научни и научнопопулярни статии, но като цяло развитието на
одонатологията у нас изостава от темповете и мащабите, с които се осъществява в
Европа. По-съществен пик в одонатологичните изследвания у нас се наблюдава през 70-
те години на миналия век. В този период по-важни са работите на БЕШОВСКИ (1965,
1967), BILEK (1966) и SCHEFFLER (1973). Най-активен по отношение на проучването на
българските водни кончета се приема периода от 80-те години на миналия век до наши
дни. В този период изпъкват работите на БЕШОВСКИ (1991), който съобщава 64 вида
одонати за България. По-късно той (БЕШОВСКИ, 1994) обобщава известните данни за
разреда у нас в том 23 на “Фауна на България”. В следващите години BESHOVSKI
& GASHTAROV (1997), MARINOV (1999, 2001, 2001a,b, f) публикуват нови за фауната
видове, като ревизират част от съобщените за България водни кончета. MARINOV
(2001b) описва нов за науката вид Somatochlora borisi от района на Източни Родопи.
Досегашни изследвания върху водните кончета от района на
Чокльово блато
Данните за одонатната фауна в изследвания район са фрагментарни и могат да се
намерят единствено в две работи. Първата е тази на БЕШОВСКИ (1967), в която за
Чокльовото блато той съобщава общо 7 вида (Lestes barbarus; L. virens; Enallagma
cyathigerum; Ishnura elegans; Libellula depressa; Orthetrum albistylum; Sympetrum
flaveolum). Втората е непубликуваната дипломна работа на ТОТЕВА (1995), в която се
съобщава за намирането в бентосните проби от блатото на одонатни ларви от родовете
Calopteryx, Lestes и Coenagrion.

- 8 -
III. Цели и задачи
Цел на изследването е да се установи видовият състав и да се проучат някои по-
важни биологични, екологични и зоогеографски особености на разред Odonata, с което
да се постави и началото на системни биологични и екологични изследвания на групата
в района.
За изпълнение на тази цел са набелязани и конкретно формулирани следните по-
важни задачи.
ЗАДАЧИ:
Да се установи видовият състав и разпространението на разред Odonata на
територията на изследвания район.
Да се определят фенологичните групи и сезонната активност на водните кончета.
Да се установи типа на обитаваните и предпочитаните от тях хабитати и да се
определи екологичният тип на изследваните водни кончета.
Да се категоризират зоогеографски водните кончета, влизащи в състава на
изследваната фауна и да се направи зоогеографски анализ на последната.

- 9 -
IV. Характеристика на Чокльово блато
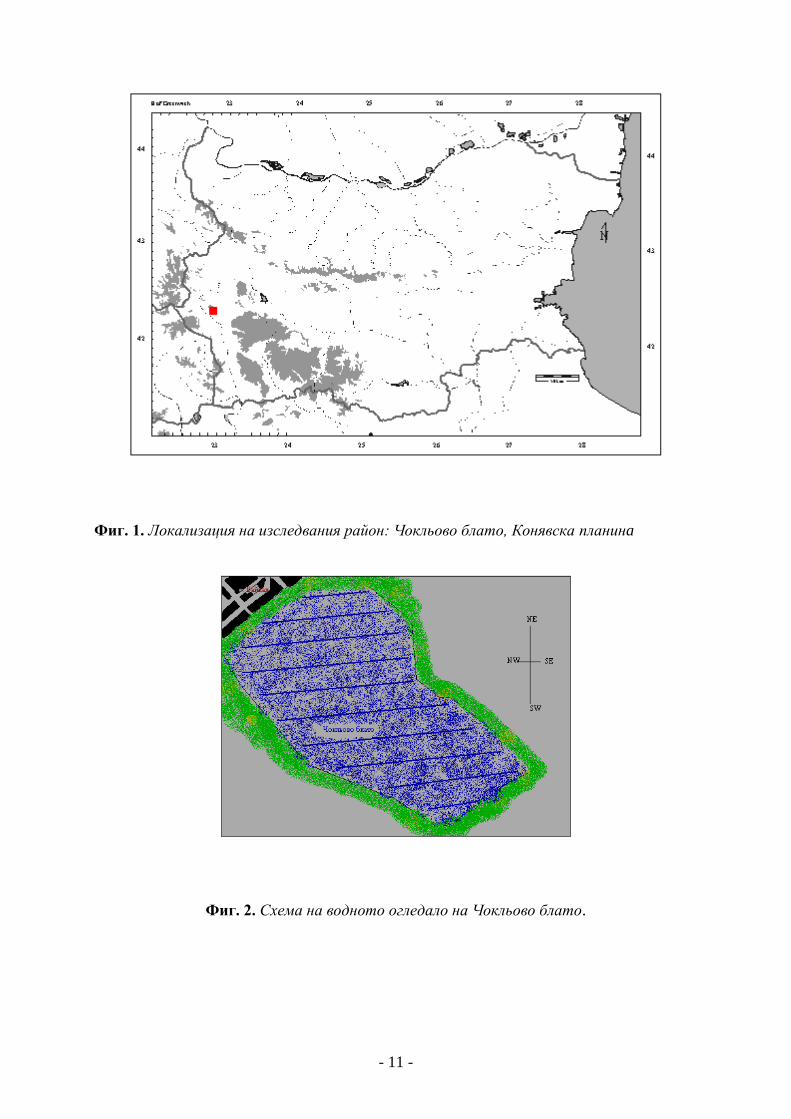
Географско положение, граници и климат
Конявска планина e част от Южната Милевско-Конявска планинска редица на
Краищенската област. Тя е разположена на границата между умерено-континенталния
и преходно-континентален климат; средната годишна температура достига 10-12 о С;
средната годишна сума на валежите е между 500 и 750 mm, с максимум през юли и
минимум през февруари; почвите са главно канелено-горски и хумусно-карбонатни, на
места ерозирали; горската растителност е представена от съобщества на Quercus cerris
L., Q. pubescens Willd., Carpinus orientalis Mill. (TONKOV & BOZHILOVA, 1992). Почти в
средата на Конявска планина тектоничната неогенно-кватернерна депресия е заета от
Чокльовото блато, което е със старохолоценска възраст. То е разположено в дясно от
пътя Радомир-Кюстендил, на 877 m надм. вис. (Г. БОНЧЕВ, 1929, КОНСТАНТИНОВ,
1973/74) (Фиг. 1). На север от Чокльово блато е разположен Димовгробският
(Кориловският) дял с най-висок връх – Димов гроб, 1056 м. Този дял се простира
между Радомирската котловина и долините на реките Струма, Блатешница и
Клисурска. (Николов, 1997).
Понастоящем повърхността на водното огледало на Чокльово блато е с площ 1,8 км² а
самата заблатена част е 4,5 км² (Фиг. 2). Крайбрежните части на блатото са обрасли с
папур, а във вътрешността на водоема се разполагат островчета от тръстика. Ъглите на
блатото са насочени към изворните части на реките Шегава и Драгомировска, а също
така и към течението на река Блатешица. Най-ниските му части се намират в
североизточния край и в тях водата се задържа по-дълго време. Водосборът на
отичащата се вода има много по-обширен обхват, включващ планинските склонове от
по-високата поясна ивица на Конявска планина. Площта на водосбора на Чокльовското
структурно понижение е 16,2 км².
По време на Втората световна война е започнала експлотация на блатото като източник
на торф за горене. В тази връзка по-късно (1950 година) чрез специална дренажна
система то е било напълно пресушено Едва през 1981 г. дренажната система е
премахната и блатото възстановено.

- 10 -
Според Г. Бончев (1929) Чокльовото блато лежи средноюрски глинесто-пясъчников
терен като то трябва да бъде поставено в групата на карстовите блата. По-късно обаче
Йорданов (1931) посочва, че Чокльово блато би трябвало да носи името планинско
тресавище, понеже и при максимално пълноводие, то няма присъщият вид на едно
типично блато. Понастоящем въз основа на получените за Чокльово блато резултати
(виж по-долу) за температурата, прозрачността, сапробността и хидрохимичните му
компоненти, като: 1) ниско съдържание на нитрати, фосфати, 2) минимално количество
нитрити, 3) ниски стойности на БПК (биологично потребление на кислород), 4) слабо
алкални стойности на pH, 5) нормално насищане с кислород, STOYNEVA & VALCHANOVA
(1997) го определят като фосфор-лимитиран, бета-мезосапробeн, еутрофeн,
полимиктичен , плитководен басейн със слабо алкални води.
- 11 -
Фиг. 1. Локализация на изследвания район: Чокльово блато, Конявска планинa
Фиг. 2. Схема на водното огледало на Чокльово блато.

- 12 -
Кратка хидрологична и хидрохимична характеристика на Чокльово блато (по
данни на ВУЧКОВА, 1995, STOYNEVA & VALCHANOVA, 1997)
Дълбочина
Тя се колебае в различните части на блатото, средно 100-120 см. Най-голяма тя е в
западната му част, където достига до 168-203 см, а най-малка е в източната - до около
124 см. Освен това дълбочината се променя през отделните сезони - през юни е най-
голяма (198-203 см), а през ноември достига 125 см.
Температура
Тя също търпи сезонни промени, (минимална от 7.5 о С през април до 26
о С през
септември) като на повърхността тя е в пряка зависимост от температурата на въздуха.
Средните стойности на водата през април е 7.85 о С, през юни е 18.12
о С, през юли -
21.25 о С, за септември - 23.75
о С, а през ноември спада на 9.97
о С). Поради
сравнително малката дълбочина на водата във водоема, температурни различия във
вертикално направление не са установени. В хоризонтално направление,
температурната разлика е минимална (около 16 о С).
Прозрачност
За нея значение имат прозрачността на планктона, разтворените вещества и газове,
както и количеството на наносите. Най-високата стойност на прозрачността е отчетна
през месец юни (средно 107.5 см), което може да се обясни с ниската численост на
фитопланктона (ВЪЛЧАНОВА, 1995). Най-ниска тя е през юли (60 см) и септември (67.5
см). Утайките на дъното се формират от автохтонни материали (главно гниеща водна
растителност и планктонен детрит) (ТОТЕВА, 1995).
Активна реакция на водата
Определя се от концентрацията на водородните йони. В хоризонтално направление рН
на водата на блатото се движи в границите 7,7 – 7,85. Най-високото рН на Чокльово
блато е измерено през месец ноември - 8,29, а най-ниско то е през април – 7,26. През
летните месеци рН в повърхностния слой се колебае между 7,6 и 7,7.

- 13 -
Кислородно съдържание и насищане
То зависи от различни фактори (абиотични: температура, вертикална и хоризонтална
циркулация; биотични: развитие на фотосинтезиращи организми, гнилостни процеси и
т. н.). Сезонните изменения на кислорода следват температурните (например: през
лятото при температура на водата 26 о С, кислородното съдържание е 96 %). В
хоризонтално направление в повърхностните слоеве не се наблюдават значителни
колебания. Кислородното насищане през летните месеци на повърхността на водата е
76%, през пролетта е по-ниско – 61%, а есента се повишава (септември достига 82%, а
ноември – 78%).
Кислородното съдържание е с най-високи средни стойности през месец ноември (8.96
mg/l), когато температурата на водата е ниска. С отмирането на основните продуценти
на кислород в периода ноември-април съдържанието на кислород в блатото силно
намалява и достига средни стойности 7.24 mg/l . През месеците юни-септември варира
от 6.24 до 7.03 mg/l. Този феномен може да се обясни с факта, че въпреки че юни-
септември е период на най-интензивно развитие на фитопланктона, то той е и период
на най-високо биохимично потребление на кислород и период на високи температури.
Количество на въглероден двуокис
В Чокльово блато то е средно между 5,18 mg/l и 10,8 mg/l. Намаленото количество на
въглероден двуокис във водата през месец юли корелира с „цъфтежа” на синьозелените
водорасли от род Microcystis.
Съдържанието на амоний и амоняк
В Чокльово блато то се колебае между 0,35 mg/l и 1,4 mg/l и е в пряка зависимост от
процесите на амонификация, денитрификация и сезонна динамика на фитопланктона.
Количеството на нитритите варира от 0 mg/l до 0,0005 mg/l, а на нитратите между
0,0009 mg/l и 0,015 mg/l. Тяхното колебание е резултат на протичащите процеси на
нитрификация и денитрификация.
Динамика на фосфатите
В Чокльово блато тя е тясно свързано с динамиката на развитие на фитопланктона.
Отчетените аналитични нули на съдържанието на фосфати в блатото през месеци като
юли, септември и ноември са свързани отново с интензивното развитие на синьо-

- 14 -
зелените водорасли. През пролетта (април) и началото на лятото (юни) съдържанието
им варира от 1.0 до 1.05 mg/l.
Кратка флористична характеристика на Чокльово блато
Бреговете на блатото (STOYNEVA & VALCHANOVA, 1997) са обрасли с Typha latifolia L.,
T. angustifolia L., Iris pseudacorus L. et al. Характерно за неговото водно огледало е
присъствието и развитието на хидатофитите Myriphyllum spicatum L., M. verticullatum
L., Ceratophyllum demersum L., Persicaria hydropiper (L.) Sparch, Potamogeton natans L.,
Urticularia vulgaris L. и U. minor L.
На неговата територия в миналото се е срещала и пет тичинковата върба Salix
pentandra L., която е включена в Червения списък на защитените от изчезване растения.
След пресушаване на блатото тя е изчезнала. Сега в специален разсадник в
югозападния край на блатото нейната популация се възстановява чрез изкуствено
засадени дръвчета.
Проучването на намерените макроостанки в Чокльово блато и спорово-поленовия
анализ дават голяма яснота за растителността, такава каквато е била преди хиляди
години и за една цялостна реконструкция на сукцесионните процеси в блатото
(ЧАКАЛОВА и кол. 1986, TONKOV & BOZHILOVA, 1992). Новите данни за холоценската
история на растителността (TONKOV & BOZHILOVA, 1992) показват, че в района от
късния бореал до късния атлантик (8000 до 6300 г. BP) е констатирано окупиране на
първичните дъбови гори от асоциции на Pinus-Abies; 2000 години по-късно се появяват
буковите съобщества, заедно с габъра Carpinus betulus с максимум 3000 г. BP; В
периода 8000-4700 г. BP тук вече се установяват следи от езерна растителност (Typha,
Sparganium). В периода 4700-3000 г. BP се наблюдава натрупване на торф от
Cyperaceae – доказателство за заблатяването на района; След 1200 г. BP растителността
е била вече сходна с тази на рецентната.

- 15 -
Кратка фаунистична характеристика на Чокльово блато*
Зообентос (по данни на ТОТЕВА, 1995)
Установената в Чокльово блато бентосна фауна включва 33 вида, спадащи към 3 типа.
Тип Mollusca с представители на Gastropoda и Bivalvia, тип Annelida с представители на
класове Hirudinea, Oligohaeta и тип Arthropoda - с класове Crustacea, Ostracoda, Araneae,
Insecta.
В дънните проби преобладават хирономидите, олигохетите и хаоборидите. Широко
разпространени са видовете Pisidum casertanum, Coretus corneus, Radix sp. и др. В
процентно отношение доминират Radix ovata, Pisidium casertanum, олигохетите и
хиронимидите. Други доминантни видове са Planorbis planorbis, Tubifex sp., Cyclops
vicinus, Lestes sp., както и представители на клас Hirudinea.
Видовото разнообразие на зообентоса в Чокльово блато според ТОТЕВА (1995) е ниско.
Индексът за видово разнообразие през различните сезони се колебае между 1-1,3.
следователно в структурата на бентоса не се наблюдават резки промени. Характерна
особеност на зообентоса е неговото мозаечно разпределение по дъното на блатото.
Зоопланктон (по данни на ТОТЕВА, 1995)
Зоопланктона е представен от 48 вида. Групите, към които спадат отделните
представители са следните: Rotatoria (24 вида), Cladocera (16), Copepoda (8); Семейство
Brachionidae e най-многобройно - представено с 9 вида. Кладоцерите в блатото
принадлежат на семействата Daphnidae – 9 вида, Bosminidae – 1 вид и един подвид,
Chydoridae – 5 вида; Най-многобройни видове от Copepoda са Cyclops vicinus,
Acanthocyslops robustus и Thermicyclops crassus. Изводите, които могат да се направят за
зоопланктона на Чокльово блато са следните – качествения и количествения състав са
константни и стабилни величини. Минимумът зоопланктон е наблюдаван през лятото, а
максимумът през есента (ноември) (Kozuharov, 2000).
Херпетофауна
Тук могат да се видят зелената водна жаба (Rana ridibunda), дървесната жаба (Hyla
arborea) (25.V.2005 г., Б. Христов leg.), зеленият гущер (Lacerta viridis), пепелянката
* Поради слабата проученост на фауната на Чокльовото блато, тук ще дадем както известните от
литературата данни, така и данните от нашите собствени наблюдения.

- 16 -
(Vipera ammodytes) (♀, 03.ІХ.2005, Б. Христов leg.), а във водоемите, честа беше и
жълтоухата водна змия (Natryx natrix).
Орнитофауна
Чокльово блато представлява орнитологично важно място, тъй като тук мигриращите
птици намират място за почивка и храна, а някои от тях, като белооката потапница и
малкият воден бик остават, за да отглеждат малките си. На територията на блатото
установихме гнездяща двойка и на редкия червеноврат гмурец (Podiceps grisegena). В
ливадите около блатото гнезди и ливадният дърдавец, вид застрашен от глобално
изчезване. В района на селото и около блатото се наблюдаваха традиционни за
гнездовата орнитофауна видове като: бял щъркел (Ciconia ciconia), обикновен мишелов
(Buteo buteo), сива овесарка (Emberiza calandra), черноглава стърчиопашка (Motacilla
flava feldegg) и други.
Хабитатни типове, установени на територията на Чокльово блато
Поради спецификата на своята биология водните кончета са надежден
индикатор за екологичната стабилност, качеството на екотонната зона вода-суша и за
хабитатната хетерогенност на водоемите (SCHMIDT, 1983; CHOVANEC, 1994, 2000). В
пряка връзка с индивидуалното значение на отделните видове одонати като екологични
индикатори са и различните класификации на хабитатите в зависимост от най-
характерните за тях одонатни ценози.* Поради това считаме за важно представянето в
настоящата разработка на характеристики на установените хабитатни типове на
територията на Чокльово блато, които са въз основа на хабитатната класификация на
Маринов (2001).
Торфище
Плитководни (до 0,50 м.) или по-дълбоки (до към 5,00 м.) водоеми. През летните
месеци по-плитките участъци могат и да пресъхнат.
* За територията на бившата ГДР JACOB (1969) използва синекологичния подход и описва 11
различни съобщества, като ключов пункт за един правилен биогеографски анализ. Основното в неговата
класификация е използването на така наречените от него ръководни (ценобионтни и ценофилни) и
съпътстващи (еврибионтни и ценоксенни) видове.

- 17 -
Според Маринов (2003) индикаторни (ръководни) са тези видове водни кончета, които
са тясно привързани към съответния хабитат, според екологичните изисквания на вида .
За този тип хабитат са ръководни видове са: Leucorrhinia dubia, Somatochlora arctica,
Coenagrion hastulatum, като освен тях като съпътстващи ги видове посочва: Aeshna
juncea, Libellula quadrimaculata, Somatochlora metallica, Cordulia aenea, Pyrhosomma
nymphula, Lestes sponsa, Lestes sponsa, Sympetum flaveolum, Aeshna cyanea, Erythromma
najas, като последните три вида са установени в района на Чокльово блато.
Сенчести участъци по дължина на басейни с течащи или слабо подвижни
води, с брегове или водна повърхност, обрасли с растителност
Характеристика: Ширината варира обикновено в границите 0,50-3,00 м., а
дълбочината – 0,01-0,50 м. Сенчестите участъци се получават от развитието на
дървесна растителност (в случая Salix sp.).
Според Маринов (2001) индикаторни (ръководни) за този тип хабитат са:
Caliaeschna microstigma, Somatochlora meridionalis, Somatochlora borisi, Cordulegaster
picta, Cordulegaster bidentata;
Като съпътстващи видове Маринов отняся следните видове водни кончета:
Cordulegaster heros, Cordulegaster insignis, Chalcolestes viridis, Calopteryx splendenновен
на тs; Последният вид бе и установен в блатото.
Тези видове могат да бъдат открити и сред растителните обраствания в съседни
на хабитата осветени участъци. Тези места се използват за снасяне на яйца (C.insignis),
копулация (C.viridis) или почивка (C.splendens) – за последният вид, беше многократно
установено. Почивката се изразява в продължителното състояние на покой върху
растителността – най – често тръстика.

- 18 -
Обрасли с растителност брегове до водоема с видимо течение,
преминаващи през осветени участъци
Характеристика: Размерите на тези басейни варират в по-широки граници от тези,
представляващи предния тип. Ширината може да надхвърли и 10,00 м., а дълбочината
до достигне повече от 1,00 м. Осветеността се дължи на факта, че околните дървета
(подобни по състав на тези, дадени за предния тип) са отдалечени от бреговете.
Ширините на отделните участъци допринася в значителна степен за повишаване на
осветеността. Растителността по бреговете е представена от групи с характерни
представители Carex sp., Lythrum sp.. Дълбочините и бързото течение обикновено не
позволяват развитието на растения във вътрешността на басейните. В по-плитките
участъци (до около 0,50 м.) понякога се развива Ranunnculus sp. (???).
Според Маринов (2001) индикаторни (ръководни) за този тип хабитат са Epallage
fatime и Calopteryx virgo – последния вид бе установен.
Като съпътстващи видове тук се отнасят два също така установени вида: Platycnemis
pennipes, Calopteryx splendens – и двата вида установени
Временни, обрасли с растителност участъци, обособени като
самостоятелни водоеми или формирани около други влажни зони.
Разположени са най-вече в слънчеви участъци
Характеристика: Варират изключително много по форма и големина. Най-
характерното за тях е, че са плитководни (до около 0,50 м.), като в по-топлите месеци
на годината отчасти или напълно пресъхват. Често се образуват при разливане на
водата от един основен басейн в следствие на пролетните пълноводия. Като друга
причина за тяхната поява може да се посочи липсата на постоянен водоток (изворна
или дъждовна вода) в единичните понижения в терена, където тези временни водоеми
се формират. Повърхността е обрасла изцяло с макрофитна растителност, като най-
характерните представители са: Phragmites australis, Typha sp.; Според Маринов (2001)

- 19 -
индикаторни (ръководни) за този тип хабитат са: Lestes sponsa, Sympetrum vulgatum,
Sympetrum flaveolum – последният е установен.
Открити слънчеви участъци всред макрофитните обраствания в или по
бреговете на постоянни водоеми
Характеристика: Вариациите в размерите на тези хабитати са изключително големи.
Определят се от степента на обраствания с макрофити. Последните са представени най-
вече от съобщества от Phragmites australis и Typha sp., които се развиват в басейни с
максимална дълбочина до към 1,00-1,50 м. В отделни случаи тези макрофити могат да
образуват собствен субстрат, където се вкореняват. Така се получават “островчета” от
растителност, които при по-високи стоежи на водоема могат да се откъснат и да се
движат в зависимост от посоката на вятъра. Затова определените тук като ръководни
видове могат да бъдат открити както по крайбрежията при добре развитите
обраствания, така и във вътрешността на водоема, следвайки “островчетата” от
макрофити.
Според Маринов (2001) индикаторни (ръководни) за този тип хабитат са Orthetrum
albistylum, Aeshna isosceles – и двата вида бяха установени на територията на Чокльово
блато.
От съпъдстващите видове бяха уловени следните представители: Libellula depressa,
Crocothemis erythraea, Anax imperator, Cordulia aeneа.
Плаваща/потопена растителност в стоящи или почти стоящи водоеми,
развита предимно в слънчеви участъци
Характеристика: Размери на тази хабитати няма да се посочват. Определящо при
избора на хабитата от страна на имагото е наличието на характерната растителност,

- 20 -
представена от Potamogeton sp. например, която бе и ръководната растителност в
балатото. Ръководни видове: Erythromma viridulum, Erythromma najas;
Хубаво е да се отбележи, че Чокльово блато е второто находище в България, където
тези два вида са установени заедно в един и същ биотоп заедно.
Съпътстващи видове: Anax imperator, Sympecma fusca; също установени видове.

- 21 -
V. Материал и методи
Настоящето изследване се базира на обработката на 37 екземпляра одонати от 21
вида, събирани с помощта на стандартен ентомологичен сак в периода май – септември
2005 г. В този период Водните кончета бяха събирани ежемесечно, приблизително през
две седмици (25.05; 04.06; 11.06; 30.06; 09.07; 23.07; 12.08; 24.08; 03.09; 16.09), с цел да
се уловят видове с различна сезонна активност.
При събирането на материала бяха използвани предварително избрани
маршрути, които включваха и различни по тип хабитати.
Дънни проби за ларвен материал бяха събирани с помощта на стандартен
хидрологичен сак. Уловените живи ларви (Cordulia aenae и Coenagrion puella) бяха
отглеждани в лабораторни условия до тяхното имагиниране.
Определянето на уловените видове се извършваше след тяхното препариране и
етикетиране. Определянето на уловените видове се извършване с помоща на
бинокулярна лупа. Детерминирането на част от събраният материал (ларви, имаго и
екзувии) беше извършено от д-р Милен Маринов, за което му изказвам най-искрена
благодарност. Събраният материал се съхранява в личната колекция на автора, като
част от него ще бъде депонирана в ентомологичната колекция на катедра „Зоология и
антролопогия” при БФ на СУ.
При определяне на водните кончета беше използвана следната по важната
литература:
- В. Бешовски, „Фауна на България”, том 23, разред Odonata
- М. Маринов “Джобен полеви определител на водните кончета на България”
В настоящата дипломна работа се придържаме към класификацията на Бешовски
(1994) и Маринов (2000).
Картовият материал с данните за разпространението на всеки вид, както и
обобщените фенограми включени в настоящата работа ни бяха любезно предоставени
от д-р Милен Маринов, за което още веднъж му благодарим.
Предложената тук класификация на типовете хабитати e съобразенa с тaзи в
работите на Jacob (1969) и Маринов (2003). Използвайки понятията микрохабитат,
хабитат, биотоп и местообитание се придържаме към смисъла вложен в тях от Одум
(1975, 1986) Янков (1997), Devilliers-Terschuren (1996). Последните автори определят

- 22 -
хабитата по следния начин: „триизмерна част от пространството , обхващаща поне една
гранична зона между въздуха, водата и земята, която включва както неживата среда
така и обитаващите я съобщества от растения и животни. Отличава се с определена
хомогенност, характерен облик и относително еднообразни условия на цялата си
повърхност, като може да бъде изградена от по-ниски по ранг съставни единици или
самата тя да бъде част от по-големи такива”.
Представени са обобщените за България фенограми на ръководните видове водни
кончета. Използваме зоогеографския анализ на Геогриев (1978, 1982) в раздела
„Зоогеографски анализ”

- 23 -
VI. Резултати и обсъждане
В резултат на нашите изследвания и на обобщените данни от литературата (БЕШОВСКИ, 1994,
МАРИНОВ, 2003) се установи, че на територията на Чокльово блато се срещат общо 23 вида
одонати (Табл. 1), което прави 1/3 от видовия състав на българската одонатна фауна (ХУБЕНОВ,
2005). От тях 17 са нови за изследвания район, 4 вида се потвърдиха, а два (Lestes virens и
Enalagma cyathigerum), посочени в литературата за района не бяха установени
6.1. Фаунистичен анализ на установените видове
В изследвания район – Чокльово блато, Конявска планина, бяха установени
общо 23 вида водни кончета, представени в таксономичен ред в Таблица 1:

- 24 -
Таблица 1: видов списък на водните кончета от Чокльово блато
Вид
♂ ♀
Брой
екземпляри
1. Calopterix virgo ** 2
2. Calopterix splendens ** 1
3. Chalcolestes parvidens ** 1
4. Lestes virens * Бешовски, 1968
5. Lestes barbarus * 1
6. Sympecma fusca ** 2
7. Erythroma najas ** 3
8. Erythroma viridulum ** 1
9. Coenagrion puella ** 2
10. Coenagrion pulchelum ** 1 2
11. Enallagma cyathigerum * Бешовски, 1968
12. Platycnemis pennipes ** 1
13. Ishnura elegans * 2
14. Aeshna affinis ** 1
15. Aeshna cyanea ** 1
16. Aeshna isosceles ** 3
17. Anax imperator ** 1
18. Cordulia aenea ** 3
19. Libellula depressa * 1
20. Orthetrum albistylum* 1
21. Orthetrum brunneum ** 1
22. Crocothemis erythraea ** 1
23. Sympetrum flaveolum * 3 2
*- вид, известен и установяван преди извършените изследвания; ** - вид, установен по
време на изследванията и нов за региона;

- 25 -
Фиг.3. % разпределение на установените досега водни кончета, нови и
предполагаеми за региона
% разпределение на установени досега
видове, нови и предполагаеми
26%
23%
51%
Очаквани видове
Установени видове
видове
досега
Нови за района

- 26 -
С настоящата дипломна работа бяха установени 16 изцяло нови за региона вида, което
е 51 % от предполагаемият (заради липса на сигурни данни) видов състав за региона на
Чокльово блато, Конявска планина.
Установените видове водни кончета са широка разпространени в България. На
територията на Чокльово блато се срещат видове, които са със широка екологична
пластичност и предвид сукцесиите, които водоема търпи все още, не бяха открити по-
значими и редки за странат видове.
Според действащият в сила „Природозащитен списък” на Маринов (2003), има някои
видове водни кончета от района на Чокльово блато, които са със консервационна
значимост. Те се подразделят по следния начин:
Като „Застрашен вид” влиза Erythromma najas; Като „Уязвим” са видовете
Chalcolestes parvidens и Cordulia aenea;
„Почти застрашен” са видове като: Coenagrion pulchellum, Aeshna cyanea, Aeshna
isosceles , Libellula fulva, Sympetrum flaveolum
С „Най-малка консервационна значимост”са видовете: Calopteryx virgo, Caloptеryx
splendens, Lestes barbarus, Sympecma fusca, Platycnemis pennipes, Erythromma viridulum,
Coenagrion puella, Ischnura elegans, Aeshna affinis, Anax imperator, Libellula depressa,
Orthetrum albistylum, Orthetrum brunneum, Crocothemis erythraea.

- 27 -
6.2. Биологични и екологични бележки на установените
видове водни кончета на територията на Чокльово блато,
Конявска планина
Подразред Z Y G O P T E R A
Надсемейство CALOPTERYGOIDEA
Семейство CALOPTERYGIDAE Selys, 1853
Род Calopteryx Leach, 1815
Calopteryx virgo (Linnaeus, 1758)
Биология и екология: Видът е разпространен по крайбрежията на реките в
цялата страна. В България, както и в Чокльово блато се среща се заедно с други
типични реофилни представители като Calopteryx splendens, Platycnemis pennipes и др.,
като това бе установено и в Чокльово блато. C.virgo обитава по-често по-горните
течения на реките при надморска височина над 400-500 м. Тази тенденция се нарушава
в крайните югоизточни райони на страната. В някои течащи водоеми по
Черноморското крайбрежие и Странджа C.virgo е наблюдаван в по-големи количества
от C.splendens.
Според Маринов (2003) най-рано видът може да бъде видян през месец май, а
най-късно в началото на септември.

- 28 -
Calopteryx splendens (Harris, 1782)
Биология и екология: Ларвите на този вид са типични обитатели на реофилната
епифитна ценоза. Обитават хабитати подобни на тези, посочени за C.virgo.
У нас видът е характерен за средните и долните течения на реките в страната и
затова съобщенията от местата с надморска височина до към 200 метра значително
преобладават над останалите. За имагото е известно, че може да се отдалечава от
местата за репродуциране. Поради тази причина отделни екземпляри са откривани и на
по-големи надморски височини, обикновено без сигурни доказателства за
репродуциране.
Най-ранните наблюдения на вида в природата са от последите дни на април, а
най-късните са от началото на месец октомври.
Надсемейство LESTOIDEA
Семейство LESTIDAE Sеlys, 1840
Род Chalcolestes Kennedy, 1920
Chalcolestes parvidens (Artobolevski, 1929)
Биология и екология: Видът е намиран на много места в България главно в райони с
ниска надморска височина. Той обитава най-вече сенчести участъци сред близките до
водоемите дървесни видове. Сред подобни хабитати бе и откриван на територията на
Чокльово блато.
Характерно за имагото е голямата му привързаност към сенчести участъци, сред
крайбрежната дървесна растителност. Когато около водоема има само единични
дървета, а не обширен пояс, са наблюдавани струпвания на имагото високо в клоните.
Видът е типичен обитател на низинните райони- предимно до 100 м.н.в. Всички
съобщения от места с по-висока надморска височина са от Южна България.

- 29 -
Маринов (2003) съобщава за най-ранните, юли месец и най-късни, ноември
месец наблюдения на вида главно от Североизточна България.
Род Lestes Leach, 1815
Lestes barbarus (Fabricius, 1798)
Биология и екология: Той е един от най-масовите видове от род Lestes за
страната. Среща се около водоемите в цялата страна, разположени в местата с по-ниска
надморска височина. Съществуват и единични съобщения от някои високопланински
райони.
Ларвите на вида принадлежат към лимнофилната епифитна фитофилна биоценоза. Те
се срещат във временни водоеми. Могат да преживеят и при завишена соленост на
водата (до 13‰). Тази особеност обуславя разпространението на вида по цялата
Черноморска крайбрежна ивица..
Според Маринов (2003) най-ранните наблюдения на вида са от мачалото на май, а най-
късни от средата на октомври.
Lestes virens (Charpentier, 1825)
Биология и екология: Ларвите на вида принадлежат към лимнофилната епифитна
фитофилна биоценоза (БЕШОВСКИ 1968). За развитието си предпочитат временни, гъсто
обрасли водоеми. Плътно разположените растителни стъбла осигуряват добра защита и
на имагото в първоначалния момент след своето излюпване. В такива случаи са
ноблюдавани струпвания сред Typha над самата водна повърхност в осветени участъци.
В периода на полово съзряване имагото може да избере участъци, отдалачени на
значително разстояние от водоема. Копулация може да има и при самата водна
повърхност, като при това двойката каца по стъблата на потопените във водата

- 30 -
растения (Typha sp.). Яйцата се полагат отново в тъканта на стъблата на това растение.
В единично наблюдаваните случаи това се е осъществявало в участъци, непотопени във
водата.
Род Sympecma Burmeister, 1839
Sympecma fusca (Vander Linden, 1820)
Биология и екология:
Ларвите на вида са представители на лимнофилната епифитна фитофилна биоценоза.
Развиват се в постоянни водоеми. Обитават растителни обраствания в крайбрежните
плитки участъци. Същите се използват и от имагото след половото съзряване. Най-
вероятна причина при избора на хабитат от имагото е наличието на подходящи
субстрати за кацане, а не гъстотата на обрастване. При това те използват растителните
стъбла над водата или по бреговата ивица, както и техни остатъци, плуващи на
повърхността. Мъжките се разполагат обикновено на разстояние около 2 метра един от
друг. При по-големи концентрации те проявяват агресивност към съседите си. Такава
бе наблюдавана в част от наблюденията ни. От избраните за кацане места атакуват и
преминаващи екземпляри от други видове - Ischnura pumilio, например. При тях се
наблюдава голяма привързаност към избраните места и след всеки полет (проява на
агресивност или след хранене) кацат отново на същия субстрат.
S.fusca е известен като презимуващ вид.
Той се среща предимно в низинните части на страната. Съобщенията от
планинските райони с надморска височина около и над 1000 метра са малко.
S.fusca е най-рано появяващия се вид през годината

- 31 -
Семейство COENAGRIONIDAE Kirby, 1890
Род Erythromma Charpentier, 1840
Erythrommа najas (Charpentier, 1840)
Този вид е известен само от 5 находища в страната, едно от които се явява Чокльово
блато – ново за вида. Съобщенията за неговото откриване са върху единични
екземпляри. Най-добре представена популация, известна от територията на България, е
в района на Чаирските езера (Западни Родопи). E.najas е по-широко разпространен в
централните части на континента. Балканският полуостров е крайната южна граница на
ареала му. В тези географски ширини той е почти изцяло изместен от близкия до него
вид Erythromma viridulum. За сега тяхното съвместно съществуване е известно само от
резерват „Сребърна“, и сега чрез настоящето проучване - и в Чокльово блато.
Биология и екология: Ларвите на вида са представители на лимнофилната епифитна
фитофилна биоценоза. Обитатели са на постоянни водоеми. Като субстрат за кацане
имагото използва не само листа на повърхността на водата, но и връхните генеративни
части на растението и единичните сухи клонки над водата. Поради малкото
наблюдения допълнителни подробности във връзка с поведенческите особености на
имагото не могат да бъдат дадени.
Имагото е наблюдавано в период с широки граници: най-ранна дата е от май
месец, а най-късната от края на месец юли (Западни Родопи).

- 32 -
Erythromma viridulum (Charpentier, 1840)
Видът е широко разпространен у нас.
Биология и екология: Ларвите на вида са представители на лимнофилната епифитна
фитофилна биоценоза. Те също обитават постоянни водоеми. Определящото за
репродуцирането и успешното развитие на ларвите е развитието на плаваща по
повърхността водна растителност. Възрастните показват голяма привързаност към
субстратите за кацане - листа или растителни остатъци на повърхността на водата. Те
слабо реагират при дразнения и сред преминаването на опасността отново се завръщат
към растителния субстрат. Съюите места се използват за оплождане и снасяне на
яйцата.
Имагото на Erythroma viridulum nай-ранното може да се наблюдава през средата на май
месец, а най-късно началото на септември.
Род Coenagrion Kirby, 1890
Coenagrion puella (Linnaeus, 1758)
Широко разпространен. Въпреки големия брой съобщения за вида не е установена
някаква типична за C. puella закономерност в разпространението му. Повечето данни от
районите с ниска надморска височина (до 100 м) са във връзка с преобладаващия брой
изследвания, провеждани в тези места.
Най-ранно наблюдение на вида е от месец май, (Южна България), а най-късните са от
средата на август (планински райони).

- 33 -
Coenagrion pulchellum (Vander Linden, 1825)
Този вид е със северен тип на разпространение и поради това у нас е масов в Северна
България, планините до 1500 м надм. вис (торфища в Западни Родопи) и по
Черноморското ни крайбрежие. Този феномен най-вероятно може да бъде обяснен с
относително еднаквите климатични условия, които предлагат крайбрежията и високите
планини (виж. JOSIFOV 1988).
Биология и екология: Ларвите на вида са представители на лимнофилната
епифитна фитофилна биоценоза. Обитават постоянни водоеми. Възможна е
копулацията да се осъществява също така далеч от водоема (до 20-30 м). При снасяне
партньорите остават двамата, като мъжкия е в изправено положение. Женските снасят в
тъкант на растения в близост до брега. Когато двойките избират листа на повърхността
на водата, те могат да се разположат непосредствено една до друга без проява на
агресивност към съседите.
Най – рано вида се наблюдава през първите дни на май, а най-късно е откриван към
средата на август (Западна България).
Род Enallagma Charpentier, 1840
Enallagma cyathigerum (Charpentier, 1840)
Биология и екология: За пръв път е открита за България по време на изследване в
Чокльовото блато, а след това и във водоем от Лозенската планина (Бешовски 1968).
Обитатели са на лимнофилната фитофилна ценоза. Екземплярите от Лозенската
планина са открити всред нападали от околните дървета листа и растителни части от
хидрофити. Копулация им се осъществява в кацнало положение, като при това двойката
избира растителни обраствания, разположени в слънчени участъци по бреговата ивица.

- 34 -
Надсемейство COENAGRIONOIDEA
Семейство PLATYCNEMIDAE Sélys, 1863
Род Platycnemis Burmeister, 1839
Platycnemis pennipes (Pallas, 1771)
Един масов вид, откриван с много екземпляри из цялата страна. Той е сред най-често
споменаваните видове при различен вид изследвания - фаунистични одонатологични,
хидробиологични и др.
Биология и екология: в различна форма от своето развитие Platicnemis pennipes сесреща
главно по плаващи по повърхността листа на Potamogeton sp. и др. растения, върху
крайбрежната растителност или сред околните обраствания от храсти и дървета.
Последните най-често се използват като места за полово съзряване, но големи
струпвания от полово зрели представители и на двата пола също са откривани в тези
места.
Род Ischnura Charpentier, 1890
Ischnura elegans (Vander Linden, 1820)
У нас I. elegans е широко разпространен в низинните части на страната, като над
700 м надм. Височина числеността му рязко спада.
Биология и екология: Ларвите му са представители на лимнофилната епифитна
фитофилна биоценоза. Обитатели са на постоянни водоеми. Това е обща
характеристика, обобщаваща най-честите наблюдения. Видът може да бъде установен
допълнително и във водоеми с бавно течение, както и в разливи в околните на
басейните площи. Не са установени някакви предпочитания на имагото към територия
около, до или във вътрешността на водоемите. Прави впечатление, че числеността на
имагото не намалява осезателно дори и при значителни промени във времето –
увеличаване на облачността, дъжд и др.

- 35 -
Видът е широко разпространен в низинните райони. Броят на находищата, разположени
над 700 м.н.в. значително намалява.
Най-ранните наблюдения са от средата на май месец от Североизточна България, а най-
късните са от средата на октомври от Южното Черноморие.
Подразред A N I S O P T E R A
Надсемейство AESHNOIDEA
Семейство AESHNIDAE Leach, 1815
Род Aeshna Fabricius, 1775
Aeshna affinis Vander Linden, 1823
У нас видът е разпространен почти изцяло в низинните части на страната. Данните от
планинските райони са откъслечни.
Биология и екология: Ларвите на вида са представители на лимнофилната епифитна
фитофилна биоценоза. Обитатели са на постоянни водоеми. Могат да се развиват и в
условия на леко завишена соленост на водата (до 4 ‰). Имагинират ниско по
растителните стъбла. Откривани са и във вътрешността на гъсто обрасли тръстикови
масиви.
Двете крайни месеца, очертаващи периода на летене на A.affinis в България са май и
октомври и са от наблюдения, проведени в Южната част на страната. В края на май
видът се появява и по черноморското крайбрежие.

- 36 -
Aeshna cyanea (Műller, 1764)
В България видът е обитава главно планинските райони с надморска височина 700
– 2100 метра, където се среща сред торфища или глациални водоеми.
Биология и екология: Ларвите на вида са представители на лимнофилната
епифитна фитофилна биоценоза. Обитатели са на постоянни водоеми.
За имагиниране Aeshna cyanea използва нискостъблената крайбрежна растителност.
Полово зрелите мъжки имат много характерен нисък полет в близост до водоемите.
Често се задържат продължително във въздуха под сянката на дървета или крайбрежна
растителност.
Най-рано е наблюдаван в началото на юли. Като представител на планинската фауна
Aeshna cyanea има пик на активност на имагото през летните месеци. Най-късно е
наблюдаван в края на септември.
Aeshna isosceles (Műller, 1767)
Aeshna isosceles у нас е ограничен с райони в малка надморска височина (100-200
м).
Биологияи екология: Ларвите на вида са представители на лимнофилната епифитна
фитофилна биоценоза. Обитатели са на постоянни водоеми. След имагинирането
индивидите се отдалечават от водата до 500 м. В тези места водят скрит начин на живот
като винаги избират участъци, разположени ниско до земната повърхност. Независимо
дали се намират сред тревна или храстова растителност около водоемите мъжките имат
характерно поведение – патрулират малки ”заливчета” сред тръстиковите обраствания.
Както обикновено са разположени във вътрешността на водоема. Поради вдадеността
им в растителните обраствания те са защитени от вятъра. Използват се много често
като имагото продължително време застава на едно място във въздуха преди да се
премести в търсене на друг такъв участък. Рядко избират позиции за кацане при самата
вода. Ако там има мъжки от същия или други видове (Anax imperator, например)
атакуват. Могат да стоят на едно и също стъбло с Orthetrum albistylum без да са

- 37 -
агресивни един към друг. Такова агресивно поведение беше наблюдавано по време на
едно от посещенията на Чокльово блато.
Копулацията се осъществява в кацнало положение в близост до брега като
двойката каца ниско сред растенията.
Видът е разпространен изключително в низинните райони. Почти всички
съобщения от местата с надморска височина над 300 метра са върху единични
екземпляри.
Най-ранното за вида наблюдения са през месец май, а най-късните средата на юли, до
към края на юли, за Югозападна България.
Род Anax Leach, 1815
Anax imperator Leach, 1815
Anax imperator е широко разпространен в цялата страна. Той е сред един от най-
често срещаните анизоптери през почти цялия активен период на летене на водните
кончета в България.
Биология и екология: Ларвите на вида са представители на лимнофилната
епифитна фитофилна биоценоза. Обитатели са на постоянни водоеми. Откривани са
сред стъблата и листата на крайбрежната и плаващата по повърхността растителност.
Екзувии са откривани и в участъци от реки със слабо течение на водата предимно
долните течения и разливи в приустиевите части. Около водата полово зрелите мъжки
облитат участъци обикновено 3 – 4 метра покрай крайбрежната растителност и
дължина 1-2 метра. Често застават на едно място във въздуха в микрохабитати,
подобни на тези, описани при A.isosceles. Атаките между мъжките от същия вид са
много яростни, при което често се сблъскат с челата си. Атаките им към мъжки от
други родове (Orthetrum, Libellula) са редки. За снасяне женските използват растителни
части по повърхността на водата. Мъжкият може да остане в тандем с женската, но
последната може да снася и сама. Хранещи се полово зрели индивиди са откривани в
територии, отдалечени от водата и над 50 метра. При това те облитат с много бърз
полет връхните части на ниската тревиста растителност.

- 38 -
Ограниченото му разпространение във височина вероятно се дължи на
намаляване на подходящите места за развитие на ларвите му.
Най-ранното наблюдения са от месец май, а най-късните от средата на септември.
Надсемейство LIBELLULOIDEA
Семейство CORDULIIDAE McLachlan, 1870
Род Cordulia Leach, 1815
Cordulia aenea (Linnaeus, 1758)
Cordulia aenae е вид със северен тип на разпространение и поради това у нас се
среща главно в планинските райони на Южна България и в низинните на Северна
България.
Биологияи екология: Ларвите на вида са представители на лимнофилната бентосна
пелофилна биоценоза. Обитават постоянни водоеми, където се срещат в по-осветените
им части. При имагиниране се използват стъблата на крайбрежната растителност.
Тенерални са откривани сред околните на водоема храсти и дървета (до 35-40 метра).
Тези места вероятно се използват за полово съзряване. При водата полово зрелите
мъжки облитат крайбрежната ивица с бавен полет ниско до водната повърхност. Това
поведение е забелязано при водоеми, чиито брегове са обрасли с крайбрежна
растителност. В по-открити басейни имагото почти не се доближава до брега и
обикновено се придържа във вътрешността. Копулацията се осъществява в кацнало
положение, като двойката избира ниски участъци всред обрастванията в близост до
брега на водоема (около 5-6 метра). Женските снасят неохранявани. При това те
избират участъци до брега или във вътрешността на водоема. Вероятно от значение за
тях е развитието на повърхностни обраствания (от водорасли или висши растения). При
първото ни посещение за събиране на материал на 25.05.2005, на около 5-7 метра
извън водната площ открихме екзувии на Cordulia aenae.

- 39 -
Семейство LIBELLULIDAE Latreille, 1802
Род Libellula Linnaeus, 1758
Libellula depressa (Linnaeus, 1758)
Среща се край водоемите из цялата страна.
Биология и екология: Ларвите на вида са представители на лимнофилната бентосна
пелофилна биоценоза. Обитават постоянни водоеми. Близката до водоема тревиста
растителност се използва като място за полово съзряване или почивка през неактивния
за тях период през деня. Това са обикновено моменти когато териториите при брега
временно се засенчват от облаци. Представители на този вид могат да бъдат видяни
кацнали по единични сухи клонки до брега на водоема. При ограничено пространство
до водоема и висока концентрация на мъжките те почти не кацат, а са в непрекъснато
движение за охрана на територията си. Непрекъснат полет около водата е наблюдаван и
в случай на липса на подходящи за тях места за кацане. Оплождането се осъществява
във въздуха, като двойката лети над водата или се измества над тревистата
растителност в случай, че територията е активно облитана от други мъжки. Женските
снасят охранявани. Не е установена строга закономерност по отношение на местата за
снасяне, но най-често се придържат в близост до крайбрежната растителност и полагат
яйцата си в почти изцяло свободната от хидрофитни обраствания водна повърхност.
Най-рано видът може да се наблюдава през към края на май, а най-късно до към
средата на август. В низинните части на страната L.depressa спира да лети обикновено
до към началото (рядко средата) на юли.

- 40 -
Род Orthetrum Newmann, 1833
Orthetrum albistylum (Sélys, 1848)
Широко разпространен вид. У нас в райони с малка надморска височина (до 300 м)
видът е откриван из цялата страна. Ограничения в неговото разпространение на
територията на България поставят известната екологична специализация.
Екология и биология: Ларвите на вида са представители на лимнофилната бентосна
биоценоза. Обитават постоянни водоеми. Имаго е откривано по дължина на течащи
водоеми без да е доказано репродуцирането му в тези места. Най-вероятно те използват
прилежащите до реките площи, чието водно ниво се поддържа от постоянно заливане
от страна на речните води. От значение за тях са крайбрежните макрофитни
обраствания, както и тези развиващи се във вътрешността на водоемите. В
прилежащите до водните басейни площи имагото е откривано в полет над тревистата
растителност. Характерно е, че при тези движения те се преместват без чести
задържания във въздуха или резки смени на посоката. Полетът на имагото при водата
се различава съществено. Копулацията се осъществява във въздуха или двойката може
да кацне. Не е забелязана строга зависимост в това отношение. Като места за кацане се
избират растителни части от обрастванията или двойката може да престои върху
хидрофити над самата водна повърхност.
Видът е характерен обитател на районите с ниска надморска височина до около 300
метра. Сигурни доказателства за репродуциране са откривани и в места, разположени
на около 600-700 м.н.в.
Най-ранните наблюдения са от края на април и най-късното средата на септември
наблюдения върху имаго са от Северна България.

- 41 -
Orthetrum brunneum (Fonscolombe, 1848)
Видът е широко разпространен за територията на България.
Биология и екология: Ларвите на вида са представители на лимнофилната бентосна
биоценоза. Обитават постоянни водоеми. От значение за тяхното развитие е наличието
макар слабо течение на водата. Откриват се често в повърхностни отводнителни
канали, потоци, образувани от изворна вода, приустиевата част на притоци към по-
големи реки и др. Екзувии са откривани по растителни обраствания над самата вода.
При водоемите мъжките установяват участъци, които активно бранят. Привързаността
към точно определено място за кацане не е строга и след приключване на полета за
защита на територия те могат да кацнат в участък в непосредствена близост до преди
това избрания. След първоначалното захващане за копулация двойката лети във
въздуха до избора на място за кацане. Най-често тези места са в близост до
територията, избрана преди това от мъжкия индивид. След приключване на
копулацията женската остава кацнала за известен период (до около 2 минути) преди да
започне снасяне. През това време, а и при самото последвало снасяне, мъжкият активно
я охранява.
Род Crocothemis Brauer, 1868
Crocothemis erythraea (Brullé, 1832)
Видът е широко разпространен из цялата страна
Биология и екология: Ларвите на вида са представители на литореофилната бентосна
екологична подгрупа. Обитават постоянни водоеми. Установено е, че издържат и на
завишена соленост на водоемите дори и когато промените в този параметър са
значителни през сезона. Копулацията се осъществява последователно във въздуха и в
кацнало положение – по растителни части в близост до предварително избраните места
за кацане от мъжкия. Снасянето не следва веднага след разделянето, а партньорите
остават кацнали върху растенията. Женската снася охранявана. При това се избират
открити участъци от водната повърхност или такива, обрасли с хидрофити. В
територии извън водоема имагото е откривано кацнало главно сред гъсто обрасли с

- 42 -
тревна растителност места. Вероятно те се използват за полово съзряване и при
нощуване. Кацането направо върху пръстта не е характерно за вида. Ако това се
осъществи, то индивидите не остават дълго време върху този субстрат.
Видът е широко разпространен в низинните части на страната, като във височина е
откриван най-често до към 300 м. Допълнителните наблюдения от някои по-
високопланински райони показват, че C.erythraea е възможно да се репродуцира и в
тези места.
Имаго на вида е наблюдавано в период с много широки граници: най-рано през средата
на май (S Черноморие), най-къснна дата е от средата на октомври (NE България).
Род Sympetrum Newmann, 1833
Sympetrum flaveolum (Linnaeus, 1758)
Разпространението на Sympetrum flaveolum на територията на България е типично
за видовете със северен произход. В преобладаващата си част се среща в планинските
райони на Южна България, както и в единични места в Стара планина. Съобщенията от
низинните части са за резерват ”Сребърна”, Североизточна България, Аладжа манастир
и в Конявска планина.
Биология и екология: Ларвите на вида са представители на лимнофилната бентосна
пелофилна биоценоза. Обитават временни водоеми. Откривани са и в крайбрежните
заливни части на постоянни басейни, което е в съответствие с известната за рода
тенденция. Наблюденията върху имагото са оскъдни. Най-често са наблюдавани
кацнали сред гъсто обрасли с макрофити осветени микрохабитати по крайбрежните
зони. Прави впечатление, че имагото рядко каца по изолирани от останалата
растителност стъбла. Вероятната причина за това може да е свързана с характерните им
морфологични белези – широки жълти ивици по крилата, които ги правят по-
забележими от страна на насекомоядните птици
Планински вид. Броят на съобщенията рязко се увеличава след границата от 600 м.н.в.
До момента наблюденията от най-високите части (2100-2200 м.н.в.) са единични и в
някои случаи без доказателства за репродуциране, но се предполага, че S.flaveolum
може да се размножава и в тези надморски височини.

- 43 -
Очаквани видове
Като имаме предвид 1) физикогеографските и климатични особености на Конявска
планина и района, 2) наличието на определени хабитатни типове както и 3) видовия
състав на водните кончета, населяващи територията на Югозападна България, тук може
да се очаква намирането на още 10 екологично пластични вида одонати. Това са:
Chalcolestes viridis, Lestes virens, Lestes sponsa, Lestes dryas, Pyrosoma nymphula,
Enallagma cyathigerum, Ishnura pumillio, Aeshna mixta, Sympetrum vulgatum, Sympetrum
striolatum. За района на Чокльовото блато може да се очаква и представители на род
Еpitheca, известен за сега от територията на Сърбия.
Фенологична характеристика на одонатите от района на Чокльовото
блато
Според МАРИНОВ (2003) водните кончета у нас могат да бъдат разпределени в седем
фенологични групи. По отношение на фенологичните си особености водните кончета
от територията на Чокльово блато попадат в пет от тях.
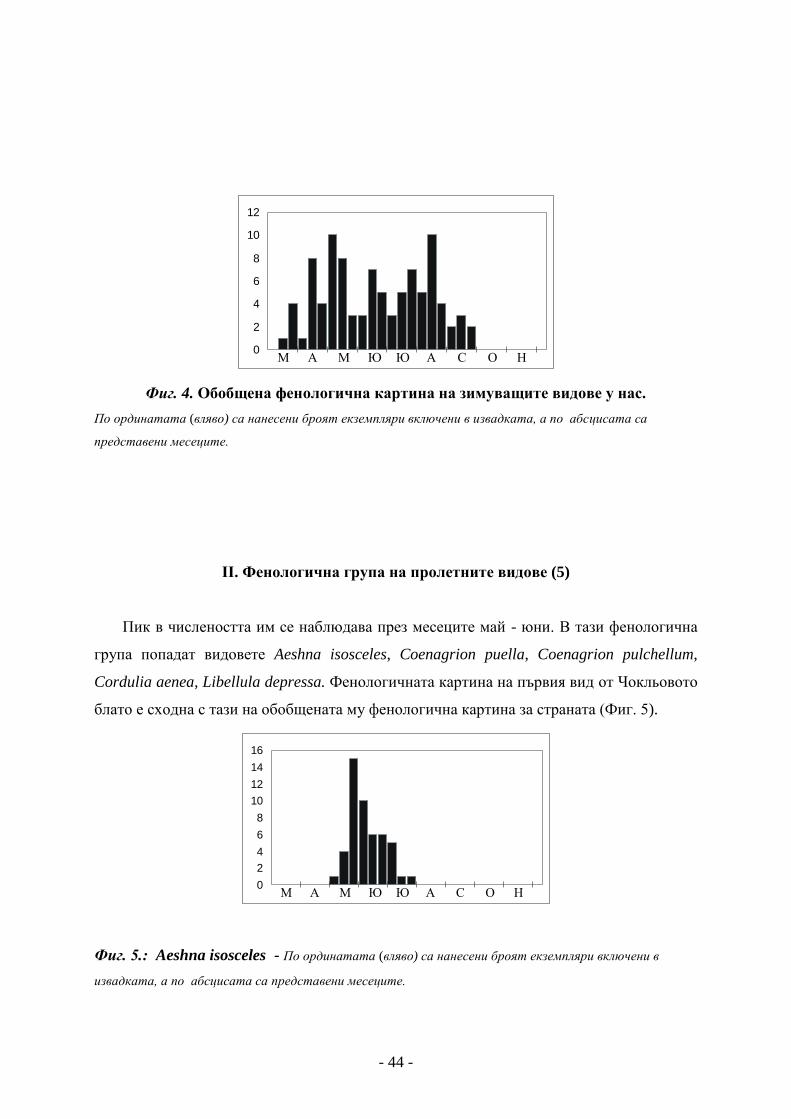
І. Фенологична група на зимуващите видове (1)
При тях имагото презимува и с повишаването на температурата се появява през
следващата година. Малко от тях остават да летят до юни. Наблюдавания при тях пик
на числеността в края на юни е във връзка главно с появата на новото поколение. Ако
сравним обобщената фенологична картина на зимуващите водни кончета у нас (Фиг.4.)
с тази от изследвания район, ще установим тяхното сходство
Вид, който попада в тази фенологична група е Sympecma fusca, определен от МАРИНОВ
(2003) като ръководен вид. Този вид беше установен от нас още при първото ни
посещение на Чокльовото блато (25.05.2005).
- 44 -
0
2
4
6
8
10
12
0
2
4
6
8
10
12
М А М Ю Ю А С О Н
Фиг. 4. Обобщена фенологична картина на зимуващите видове у нас.
По ординатата (вляво) са нанесени броят екземпляри включени в извадката, а по абсцисата са
представени месеците.
ІІ. Фенологична група на пролетните видове (5)
Пик в числеността им се наблюдава през месеците май - юни. В тази фенологична
група попадат видовете Aeshna isosceles, Coenagrion puella, Coenagrion pulchellum,
Cordulia aenea, Libellula depressa. Фенологичната картина на първия вид от Чокльовото
блато е сходна с тази на обобщената му фенологична картина за страната (Фиг. 5).
0
2
4
6
8
10
12
14
16
0
2
4
6
8
10
12
14
16
М А М Ю Ю А С О Н
Фиг. 5.: Aeshna isosceles - По ординатата (вляво) са нанесени броят екземпляри включени в
извадката, а по абсцисата са представени месеците.

- 45 -
ІІІ. Фенологична група на пролетно-летните видове (8)
При тях се наблюдават два изразени пика в активността им. Първият е в края на
май и началото на юни, а вторият - в средата/края на юли и началото на август.
Към тази група се отнасят видовете Calopteryx virgo, Calopterix splendens, Platycnemis
pennipes, Ischnura elegans, Anax imperator, Orthetrum albistylum, Orthetrum brunneum,
Croccothemis erythraea.
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
30
35
40
М А М Ю Ю А С О Н
Фиг. 6.: Ischnura elegans - По ординатата (вляво) са нанесени броят екземпляри включени в
извадката, а по абсцисата са представени месеците.
ІV. Фенологична група на летните видове (6)
При тях пикът в тяхната численост е изместен в периода юли-август. В тази група са
включени одонатите: Lestes barbarus, Aeshna affinis, Aeshna cyanea, Erythroma viridulum,
Erithroma najas, Sympetrum flaveolum;

- 46 -
0
2
4
6
8
10
0
2
4
6
8
10
М А М Ю Ю А С О Н
Фиг. 7.: Sympetrum flaveolum - По ординатата (вляво) са нанесени броят екземпляри включени в
извадката, а по абсцисата са представени месеците.
V. Фенологична група на лятно-есенните видове (1)
При тях се наблюдава ясно изразен фенологичен пик през август, последван от
пикове в края на септември и средата на октомври. Към нея се отнасят Chalcolestes
parvidens.
0
1
2
3
4
5
6
М А М Ю Ю А С О Н
Фиг. 8.: Chalcolestes parvidens - По ординатата (вляво) са нанесени броят екземпляри включени в
извадката, а по абсцисата са представени месеците

- 47 -
Групата на зимуващите и на лятно-есенните видове включва най-малък брой видове,
съответно по един; следвани от групата на пролетните (5) и летните видове (7). Най-
многобройна е тази на пролетно-летните видове, включваща 9 вида водни кончета.
Може да се направи извода, че в резултат на фенологичните си особености водните
кончета по-лесно и ефективно разпределят ресурсите на средата и избягват силната
териториална и трофична конкуренция.
На фиг.9 става ясно, че доминира групата на „Пролетно летните видове”, следвана от
„Летните видове” и „Пролетните видове”. Най-слабо са изразени групите на
„Зимуващите видове‟ и тези на „Лятно – есенните видове”, тъй като в извадката
участват само с по 1 представител.
І. Фенологична група на зимуващите видове (1)
Sympecma fusca
ІІ. Фенологична група на пролетните видове (5)
Aeshna isosceles, Coenagrion puella, Coenagrion pulchellum, Cordulia aenea, Libellula
depressa.
ІІІ. Фенологична група на пролетно-летните видове (9)
Calopteryx virgo, Calopterix splendens, Ishnura elegans, Enallagma cyathigerum,
Platycnemis pennipes, Anax imperator, Orthetrum albistylum, Orthetrum brunneum,
Crocothemis erythraea
ІV. Фенологична група на летните видове (7)
Lestes barbarus, Lestes virens, Aeshna affinis, Aeshna cyanea, Erythroma viridulum,
Erithroma najas Sympetrum flaveolum
V. Фенологична група на лятно-есенните видове (1)
Chalcolestes parvidens

- 48 -
4%
22%
40%
30%4%
зимуващите
видове
пролетните видове
пролетно-летните
видове
летните видове
лятно-есенните
видове
Фиг. 9.
% разпределение на установените видове водни кончета по фенологични групи

- 49 -
6.3. Зоогеографска характеристика
Според зоогеографското райониране на територията на България (по ГЕОРГИЕВ, 1994)
(вж. фиг. 11), Конявска планина (респ. Чокльово блато) се отнася към Рило–Родопската
зоогеографска област. Въз основа на събрания материал от водни кончета в района на
Чокльово блато, може да се направи следния зоогеографски анализ: установените 21
вида водни кончета се подразделят в следните съществуващи по ГЕОРГИЕВ (1994)
зоогеографски групи: най-силно представени, с равен процент участие са „Европейско-
медитеранската” и „Холомедитеранската” групи, следвани от „Евро-сибирската”,
„Холопалеарктическата” групи . С по-малък и еднакъв процент се нареждат след тях
„Холарктичната” и „Източномедитеранска” групи. И като цяло, „Евро-сибирският тип”
доминира с 15 вида, над „Медитеранския”, предтсавен от 8 вида. (вж. фиг. 10)
БОРЕАЛЕН КОМПЛЕКС
І. Евро-сибирски тип
1. Холарктична група, представена от 1 вид: Enallagma cyathigerum
2. Холопалеарктична група, представена от 3 вида: Calopteryx virgo, Ischnura
elegans, Sympetrum flaveolum.
3. Евро-сибирска група, представена от 4 вида: Erythromma najas, Coenagrion
pulchellum, Cordulia aenea, Orthetrum albistylum,
4. Европейска група с 2 представени вида: Libellula depressa, Aeshna cyanea.
5. Европейско-медитеранска група с 5 представени вида: Calopteryx splendens,
Platycnemis pennipes, Coenagrion puella, Aeshna affinis, Aeshna isosceles;
ІІ. Медитерански тип
1. Холомедитеранска група с 5 вида: Lestes barbarus, Lestes virens, Erythromma
viridulum, Sympecma fusca, Orthetrum brunneum
2. Източномедитеранска група с 1 вид: Chalcolestes parvidens.
3. Медитерано-афротропична група с 2 вида: Anax imperator, Croccothemis
erythraea

- 50 -
% разпределение на зоогеографските групи водни
кончета на територията на Чокльово блато
4%
13%
17%
9%22%
22%
4% 9%
Холарктичнагрупа
Холопалеарктична група
Евро-сибирскагрупа
Европейска група
Европейско-медитеранскагрупа
Холомедитеранска група
Източномедитеранска група
Медитерано-афротропичнагрупа
Фиг. 10. % разпределение на зоогеографските групи водни кончета от района на
Чокльово блато, Конявска планина

- 51 -
Фиг. 11. Зоогеографско райониране на територията на България според ГЕОРГИЕВ
(1994) и точното положение (отбелязано с точка) на Чокльово блато, попадащо в
Рило – Родопския район на Северна зоогеографска област. Останалите области, са
дадени за общо сведение:
І. Северна зоогеографска подобласт: 1. Дунавски район, 2. Старопланински район, 3.
Рило-Родопски район;
ІІ. Южна зоогеографска подобласт: 4. Струмско-Местенски район, 5. Тракийски
район, 6. Черноморски район, 7. Странджански район.

- 52 -
VII. Обобщени резултати и изводи
Въз основа на извършените и изложените в настоящата дипломна работа изследвания
можем да обобщим следните по-важни резултати и изводи:
1) Съставен е пълен списък на установените видове водни кончета от разред
Odonata от района на Чокльово блато. Този списък включва 23 вида, отнасящи
се към 15 рода и 7 семейства. От тези видове, 17 са нови за изследвания район.
2) В „Природозащитен списък” на Маринов, 2003, влизат 4 вида от установените,
в следните категории: „Застрашен” - Erythroma najas, „Уязвим” - Cordulia
aenae, Chalcolestes parvidens; „Почти застрашен” - Sympetrum flaveolum;
3) Чокльово блато е се явява второто място на територията на България, (за първи
път се наблюдава в резерват „Сребърна”, Маринов, 2003) където видове от род
Erythroma – Е. najas и E. viridulum се срещат в един и същ хабитат. Считаме
този факт, като едно рядко и интересно явление.
4) Установените данни, натрупаният опит и анализите ни показват, че Чокльово
блато разполага с много добър фаунистичен ресурс водни кончета.
Създадени са естествени условия за развитие на многообразна одонатна фауна.
За една по-добра база данни, ще са нужни още по-задълбочени изследвания в
екологичен, фенологичен аспект, както и фаунистичен разбира се.

- 53 -
VIII. ПРИЛОЖЕНИЯ
ИЗПОЛЗВАНИ ОЗНАЧЕНИЯ ПРИ РАЗПРОСТРАНЕНИЕТО НА ВИДОВЕТЕ
ВОДНИ КОНЧЕТА В БЪЛГАРИЯ
Тези означения са нанесени върху всяко едно находище на водни кончета на
територията на Република България. Освен че показват неговото общо
разпространение, показват и допълнителни база данни. А именно:
Представените карти на общо разпространение, вътрешно разпространение, и
фенология на всеки един вид са по Маринов (2003)
.
0
5
10
15
20
25
0
5
10
15
20
25
М А М Ю Ю А С О Н фиг. 12: Фенограми: По ординатата (вляво) са нанесени броят
екземпляри включени в извадката, а по абсцисата са представени месеците. В дясно по ординатата са
метрите надморска височина.
Данни, нуждаещи се от потвърждение
Данни преди 1984 г. без сигурни доказателства за репродуциране
Данни преди 1984 г. със сигурни/вероятни доказателства за репродуциране
Данни след 1984 г. без сигурни доказателства за репродуциране
Данни след 1984 г. със сигурни/вероятни доказателства за репродуциране

- 54 -
Табло 1
Calopteryx virgo (Linnaeus, 1758)
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
1
Calopteryx virgo
(Linnaeus, 1758)
Общо разпространение на вида в Европа и България
0
5
10
15
20
25
0
5
10
15
20
25
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 55 -
Табло 2
Calopteryx splendens
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
2
Calopteryx splendens
(Harris, 1782)
Общо разпространение на вида в Европа и България
0
5
10
15
20
25
30
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 56 -
Табло 3
Chalcolestes parvidens
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
5
Chalcolestes parvidens
(Artobolevski, 1929)
Общо разпространение на вида в Европа и България
0
1
2
3
4
5
6
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 57 -
Табло 4
Lestes barbarus
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
6
Lestes barbarus
(Fabricius, 1798)
Общо разпространение на вида в Европа и България
0
5
10
15
20
0
5
10
15
20
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 58 -
Табло 5
Lestes virens
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
7
Lestes virens
(Charpentier, 1825)
Общо разпространение на вида в Европа и България
0
1
2
3
4
5
6
7
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 59 -
Табло 6
Sympecma fusca
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
11
Sympecma fusca
(Vander Linden, 1820)
Общо разпространение на вида в Европа и България
0
2
4
6
8
10
12
0
2
4
6
8
10
12
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 60 -
Табло 7
Erythrommа najas
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
14
Erythromma najas
(Hansemann, 1823)
Общо разпространение на вида в Европа и България
Липсват достатъчни данни за фенологична характеристика на ви

- 61 -
Табло 8
Erythromma viridulum
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
15
Erythroma viridulum
(Charpentier, 1840)
Общо разпространение на вида в Европа и България
0
2
4
6
8
10
12
14
16
0
2
4
6
8
10
12
14
16
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 62 -
Табло 9
Coenagrion puella
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
19
Coenagrion puella
(Linnaeus, 1758)
Общо разпространение на вида в Европа и България
0
5
10
15
20
25
30
0
5
10
15
20
25
30
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 63 -
Табло 10
Coenagrion pulchellum
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
20
Coenagrion pulchellum
(Vander Linden, 1825)
Общо разпространение на вида в Европа и България
0
2
4
6
8
10
12
0
2
4
6
8
10
12
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 64 -
Табло 11
Enallagma cyathigerum
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
22
Enallagma cyathigerum
(Charpentier, 1840)
Общо разпространение на вида в Европа и България
0
2
4
6
8
10
12
0
2
4
6
8
10
12
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 65 -
Табло 12
Platycnemis pennipes
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
12
Platycnemis pennipes
(Pallas, 1771)
Общо разпространение на вида в Европа и България
0
5
10
15
20
25
30
0
5
10
15
20
25
30
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 66 -
Табло 13
Ischnura elegans
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
50 km
1200 m
UTM GridZones 34, 35
N
24
Ischnura elegans
(Vander Linden, 1820)
Общо разпространение на вида в Европа и България
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
30
35
40
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 67 -
Табло 14
Aeshna affinis
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
28
Aeshna affinis
Vander Linden, 1823
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
Общо разпространение на вида в Европа и България
0
1
2
3
4
5
6
7
0
1
2
3
4
5
6
7
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 68 -
Табло 15
Aeshna cyanea
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
29
Aeshna cyanea
(Müller, 1764)
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
Общо разпространение на вида в Европа и България
0
1
2
3
4
5
0
1
2
3
4
5
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месе

- 69 -
Табло 16
Aeshna isosceles
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
30
Aeshna isosceles
(Müller, 1767)
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
Общо разпространение на вида в Европа и България
0
2
4
6
8
10
12
14
16
0
2
4
6
8
10
12
14
16
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 70 -
Табло 17
Anax imperator
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
31
Anax imperator
Leach, 1815
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
Общо разпространение на вида в Европа и България
0
2
4
6
8
10
12
14
16
0
2
4
6
8
10
12
14
16
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 71 -
Табло 18
Cordulia aenea
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
44
Cordulia aenea
(Linnaeus, 1758)
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
Общо разпространение на вида в Европа и България
0
1
2
3
4
5
6
7
0
1
2
3
4
5
6
7
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 72 -
Табло 19
Libellula depressa
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
52
Libellula depressa
(Linnaeus, 1758)
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
Общо разпространение на вида в Европа и България
0
5
10
15
20
25
0
5
10
15
20
25
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 73 -
Табло 20
Orthetrum albistylum
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
54
Orthetrum albistylum
(Sélys, 1848)
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
Общо разпространение на вида в Европа и България
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 74 -
Табло 21
Orthetrum brunneum
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
55
Orthetrum brunneum
(Fonscolombe, 1848)
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
Общо разпространение на вида в Европа и България
0
5
10
15
20
25
0
5
10
15
20
25
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци
- 75 -
Табло 22
Crocothemis erythraea
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
57
Crocothemis erythraea
(Brullé, 1832)
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
Общо разпространение на вида в Европа и България
0
5
10
15
20
25
0
5
10
15
20
25
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 76 -
Табло 23
Sympetrum flaveolum
42
44
43
23 24 27 282625
24 272625 2823E of Greenwich
44
43
42
62
Sympetrum flaveolum
(Linnaeus, 1758)
1
2
3
4
5
6
7
8
9
0 1 2 3 4 5 6 7 8 9
Общо разпространение на вида в Европа и България
0
2
4
6
8
10
0
2
4
6
8
10
М А М Ю Ю А С О Н Фенограма, илюстрираща пиковете на летене по месеци

- 77 -
IX. ИЗПОЛЗВАНА ЛИТЕРАТУРА
Бешовски, В., 1967. – Екологичен преглед на ларвите на Odonata от българските реки.
– Изв. на Зоол. и-т с музей, БАН, ХХІV: 5-20.
Бешовски, В., 1968. – Ларви на Odonata от басейните със стоящи води в България. –
Изв. на Зоол. и-т с музей, БАН, ХХVІ: 5-27.
Бешовски, В., 1991. – Преглед на видовете Odonata, съобщени за България. – Първа
нац. конф. по ентомол.: 34-39.
Бешовски, В., 1994. – Фауна на България. – Изд. на БАН, т.23: 372 с.
Бешовски, В., М. Маринов, 1993. – Водните кончета (Odonata) от биосферния
резерват Сребърна. - Втора нац. конф. по ентомол.: 9-13.
Вучкова, Ц., (1995): Динамика на основните хидрохимични показатели в защитена
местност Чокльово блато, във връзка с биологичния режим – СУ, Кат. Хидробиология и
Ихтиология, Дипл. раб., 52 с.
Вълчанова, М. 1995: Фитопланкон на Чокльово блато, дипломна работа
Георгиев, В., 1978. – Проблеми на зоогеографското райониране на България. – Acta
zool. Bulg., 11: 41-48.
Георгиев, В., 1982. – Зоогеографски райони въз основа на сухоземната фауна.– В:
“География на България. Т. І. Физическа география. Природни условия и
ресурси”, БАН: 472-477.
Държавен вестник, брой 7, 1993

- 78 -
Йорданов, Д. 1931. Фитогеографски изучавания на блатата в България във връзка с
висшата им растителност. – Год. СУ, Физ.-мат. Фак., 75 – 156
Маринов, М., 2000. – Джобен полеви определител на водните кончета на България. –
„Eventus“ ООД, София: 104 с.
Маринов, М., 2003. – дисертационен труд на тема. – Хорология, биотопна и хабитатна
обвързаност на насекомите от разред Odonata в България, София:
Николов, В., 1997: - Планините, академично издателство „Проф. Марин Дринов”, 219
с.
ОДУМ, Ю. (1975): Основы экологии. Мир, М., 740 с.
ОДУМ, Ю. (1986): Экология., т. I и II, Мир, М., 704 с.
Тотева, И. (1995): Динамика на качествения, количествения и структурния състав на
зообентоса в защитена местност Чокльово блато. , дипломна работа, 40 с.
ХУБЕНОВ, З. (2005): Ентомофаунистично разнообразие на България. В: Петрова А.
(ред.). Current state of Bulgarian biodiversity – problems and perspectives. Bulgarian
Bioplatform, Dragon, Sofia, с. 173-198.
Чакалова, Е., Стоянова, Д., Тонков, С. (1986): Растителни макроостанки от Чокльово
блато (Конявска планина) 41-46 с.
Червена книга на Р България. Изчезнали, застрашени от изчезване и редки растения
и животни в два тома. 1984. София, БАН
ЯНКОВ, П. (1997): Местообитания на птиците в България. - В: Орнитологично влажни
места в България. БДЗП. Природозащитна поредица. Книга 1. Костадинова, И. (съст.).
БДЗП, София, стр. 18-23.

- 79 -
Askew, R., 1988. – The dragonflies of Europe. – Harley Books, Martins, Great Horkesley:
294 pp.
Chovanec, A., 1994. – Libellen als bioindikatoren. – ANAX 1(1): 1-9.
Chovanec, A., 2000. – Dragonflies (Insecta: Odonata) as indicators of the ecological integrity
of aquatic systems – a new assessment approach. – Verh. Internat. Verein.
Limnol., 27: 1-4.
DEVILLIERS, P. & DEVILLIERS-TERSCHUREN, J. (1996): A classification of Palearctic
habitats. Nature and Environment, No 78, Council of Europe, Strasburg, 9-53.
JOSIFOV, M. (1988): Über den zoogeographischen Charakter der Südeuropäischen Insektenfauna
unter besonderer Berücksichtigung der Heteropteren. - Ber. nat.-med. Verein Innsbruck, 75: 177-
184.
Kozuharov, D. (2000): Investigation of the zooplankton composition in Chokliovo swamp,
Annual of Sofia University “St Kliment Ohridski”, Faculty of Biology, Book 1 – Zoology,
Volume 91, 2000; 131-141 pp
Lakatos, G., 1995 – State of wetlands in Hungary. IUCN Wetland Programme Newsletter (in
press)
Marinov, M., 1999. – Chalcolestes parvidens (Artobolevski) and Somatochlora meridionalis
Nielsen in Bulgaria (Zygoptera: Lestidae, Anisoptera: Corduliidae). – Notulae
Odonatologicae, vol. 5, No 3: 31-33.
Marinov, M., 2000a. – An atypical Caliaeschna microstigma (Schneider) habitat in Bulgaria
(Anisoptera: Aeshnidae). – Notulae Odonatologice, vol. 5, No 6: 83.
Marinov, M., 2001b. – Somatochlora borisi spec. nov., a new European dragonfly species
from Bulgaria (Anisoptera: Corduliidae). – IDF-report 3 (1/2): 9-16.

- 80 -
Marinov, M., 2001f. – Does Coenagrion mercuriale (Charpentier, 1840) occur in Bulgaria? -
Exuviae 8(1): 13-19.
Schmidt, E., 1983. – Odonaten als Bioindikatoren fur mitteleuropdische Feuchgebiete. – Veh.
Dtsch. Zool. Ges., 1983: 131-136.
Tonkov & Bozilova, 1992 – Paleoecological investigation of Tschokljovo marsh (Konjavska
mountain), Annuair de L`universite de Sofia “St. Kliment Ohridksi”, Faculte
de biologie, Livre 2 – Botanique, Tome 83

- 81 -

- 82 -





























