Embed Size (px)
DESCRIPTION
CHAPTER 5. 細胞分裂與 DNA 複製. 細胞分裂和繁殖不一定相同. 在許多的植物與真菌中,一大群聚集的細胞也許會終止活動,或者親代生物體會釋放單細胞的孢子,成長為個別新的多細胞生物體。新的個體與親代遺傳具有相同的特性,稱為 無性 (asexual) 或 營養繁殖 (vegetative reproduction) 。相較於有性 (sexual) 繁殖,每個細胞從兩個分別的親代得到大致相等的遺傳訊息,因此基因有新的組合。. - PowerPoint PPT Presentation
Citation preview

細胞分裂與 DNA 複製細胞分裂與 DNA 複製

2 CHAPTER 5 DNA細胞分裂與 複製
細胞分裂和繁殖不一定相同細胞分裂和繁殖不一定相同在許多的植物與真菌中,一大群聚集的細胞也許會終止活動,或者親代生物體會釋放單細胞的孢子,成長為個別新的多細胞生物體。新的個體與親代遺傳具有相同的特性,稱為無性 (asexual) 或營養繁殖 (vegetative reproduction) 。相較於有性 (sexual) 繁殖,每個細胞從兩個分別的親代得到大致相等的遺傳訊息,因此基因有新的組合。

3 CHAPTER 5 DNA細胞分裂與 複製
細菌在沒有細胞分裂時,且牽涉到相似的 DNA 小片段從一個細胞 ( 供應者; the donor) 往另一個細胞 ( 接受者; the recipient) 的輸送。這種 DNA的側向輸送 (sideways transfer) ,發生在同一世代的細胞間,被認為是水平的基因傳播 (horizontal gene transmission) 。相反的,垂直基因傳播 (vertical gene transmission) 為基因從之前的世代傳到下個新的世代。因此垂直傳播包含了細胞分裂的所有形式與繁殖,可創造一個新的複製基因體,不論是有性或無性。
4 CHAPTER 5 DNA細胞分裂與 複製
DNA 複製發生在複製叉上,是一種兩階段的過程DNA 複製發生在複製叉上,是一種兩階段的過程
複製 (replicaion) 為一種始祖細胞的 DNA 加倍的過程,發生於細胞分裂之前。細胞分裂時,每一個子代會得到一份完整的複製 DNA ,與原本親代的遺傳物質相同。

5 CHAPTER 5 DNA細胞分裂與 複製
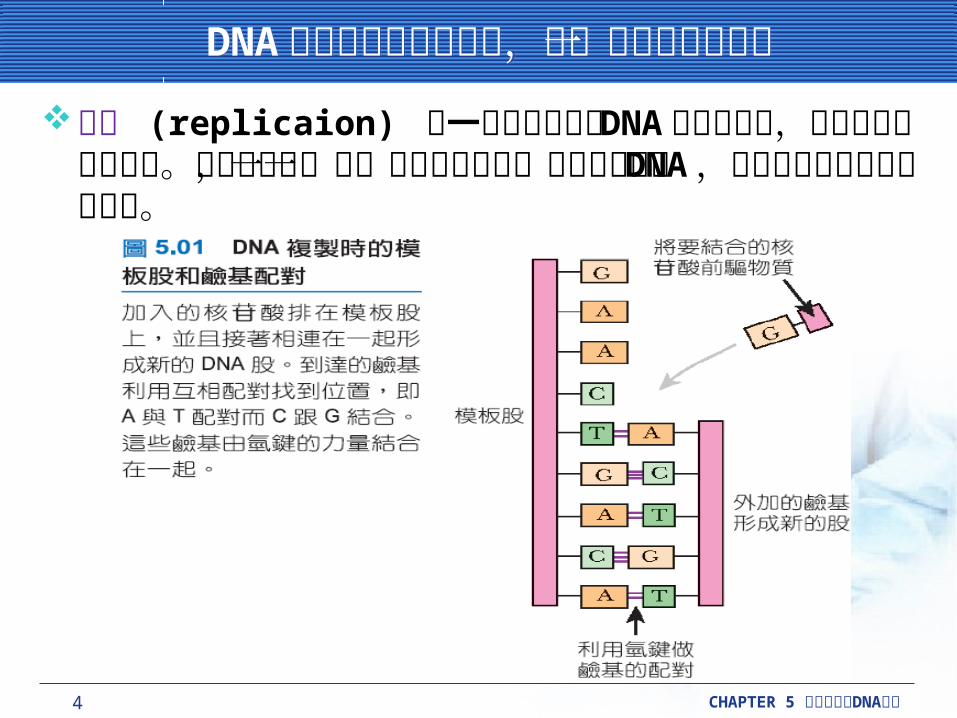
複製叉是 DNA 複製時,酵素解開 DNA 結構時,結合在已解開的單股 DNA 的區域。兩條新的 DNA 股合成都在複製叉 (replication fork) 上,此交叉點會沿著親代 DNA 移動。
複製叉包含 DNA 分開的區域,再加上負責合成的蛋白質集合,有時稱為複製體 (replisome) 。複製的結果為兩個雙股 DNA ,而且序列都與原來的相同。子代細胞其中一個有原本細胞左邊的一股,另一個具有原本細胞右邊的一股。複製是採半保留的 (semi-conservative) 方式,因為每個子代都保留一半原來的 DNA 物質。
6 CHAPTER 5 DNA細胞分裂與 複製

7 CHAPTER 5 DNA細胞分裂與 複製
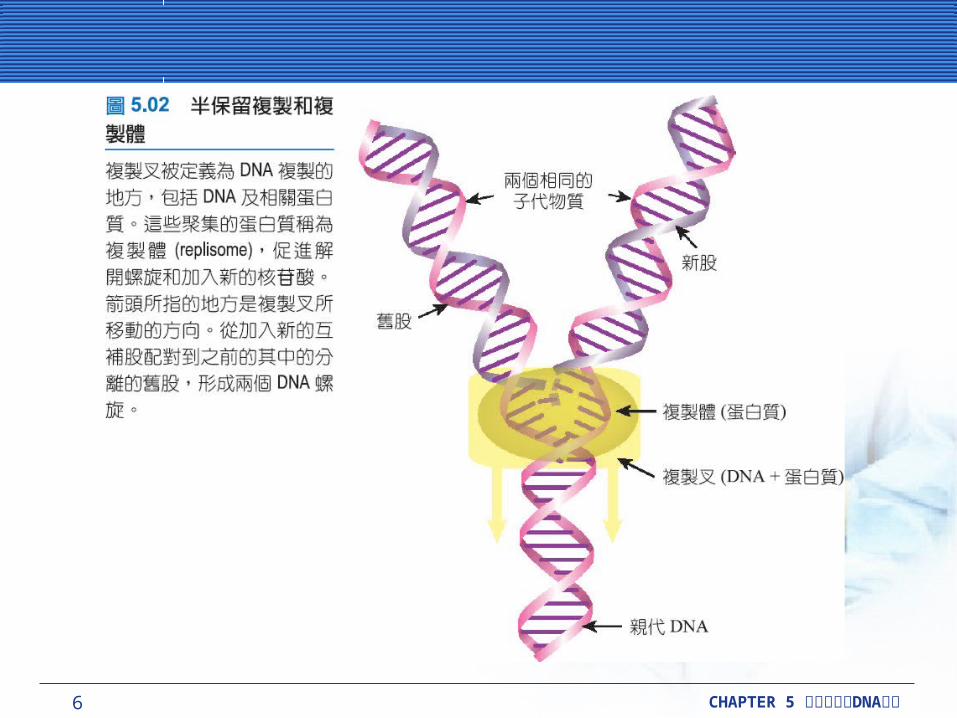
超螺旋引起複製的問題超螺旋引起複製的問題超螺旋的細菌染色體為環狀的,而且兩個複製叉以相對
的方向繞著環狀進行作用 。這過程就是雙向複製 (bi-directional replication) 。環狀的 DNA 分裂時,中間會出現像希臘字母 θ(theta) 的構造,所以這種模式也稱作θ 複製 (theta-replication) 。

8 CHAPTER 5 DNA細胞分裂與 複製
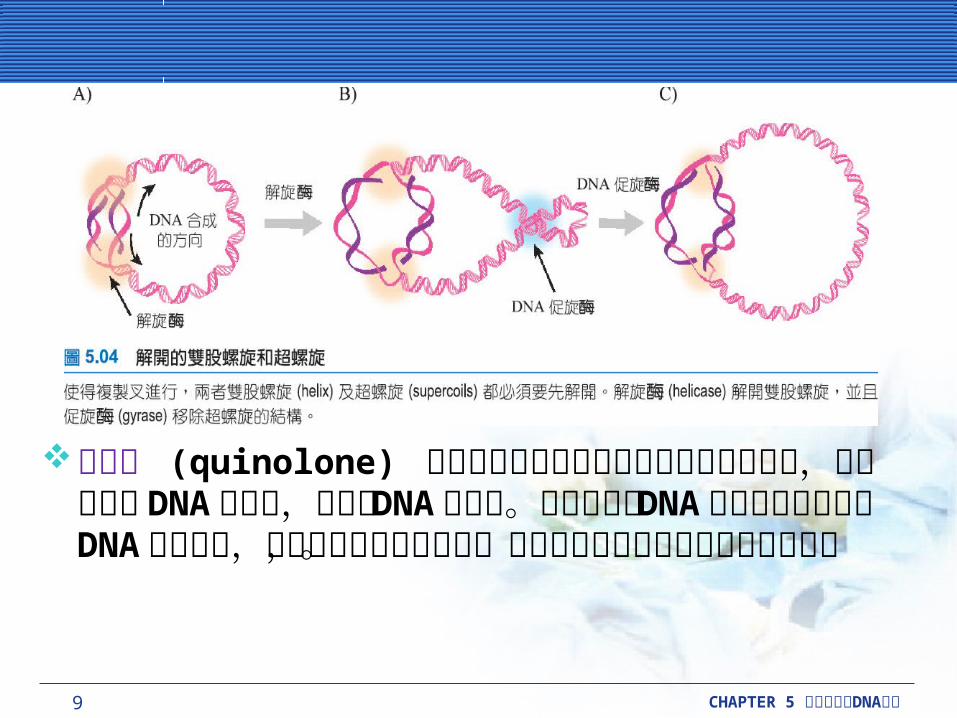
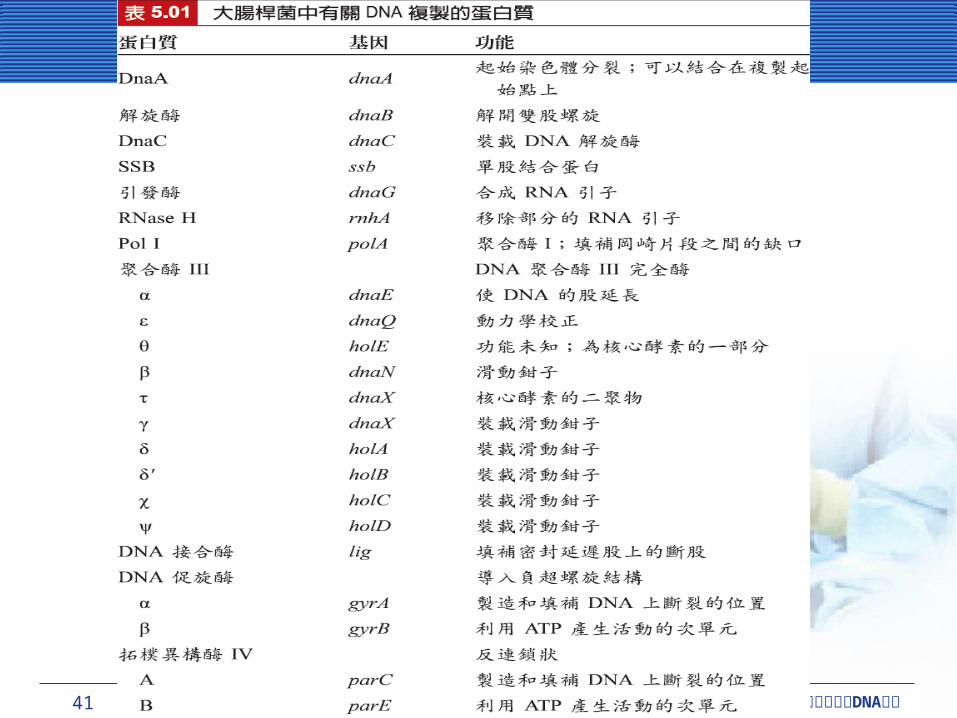
在大腸桿菌中有兩種第二型拓樸異構酶,即 DNA促旋酶 (DNA gyrase) 與拓樸異構酶 IV (topoisomerase IV) ,影響了 DNA 的複製。
DNA 促旋酶會結合在複製叉的前端 DNA 上,引起可以取消正螺旋的負螺旋。淨結果 (net result) 為DNA 促旋酶移除複製叉前面的超螺旋結構。拓樸異構酶 IV 在過程中或其他方面有協助的作用,但主要的功能,是解開複製完成後的子代 DNA 物質。
9 CHAPTER 5 DNA細胞分裂與 複製
喹啉酮 (quinolone) 抗生素利用在第二型拓樸異構酶上的作用,抑制了細菌 DNA 的複製,特別是 DNA促旋酶。受到抑制的 DNA 促旋酶仍然結合在 DNA的位置上,並且阻止複製叉的移動,而整個反應的結果為導致細胞死亡。
10 CHAPTER 5 DNA細胞分裂與 複製
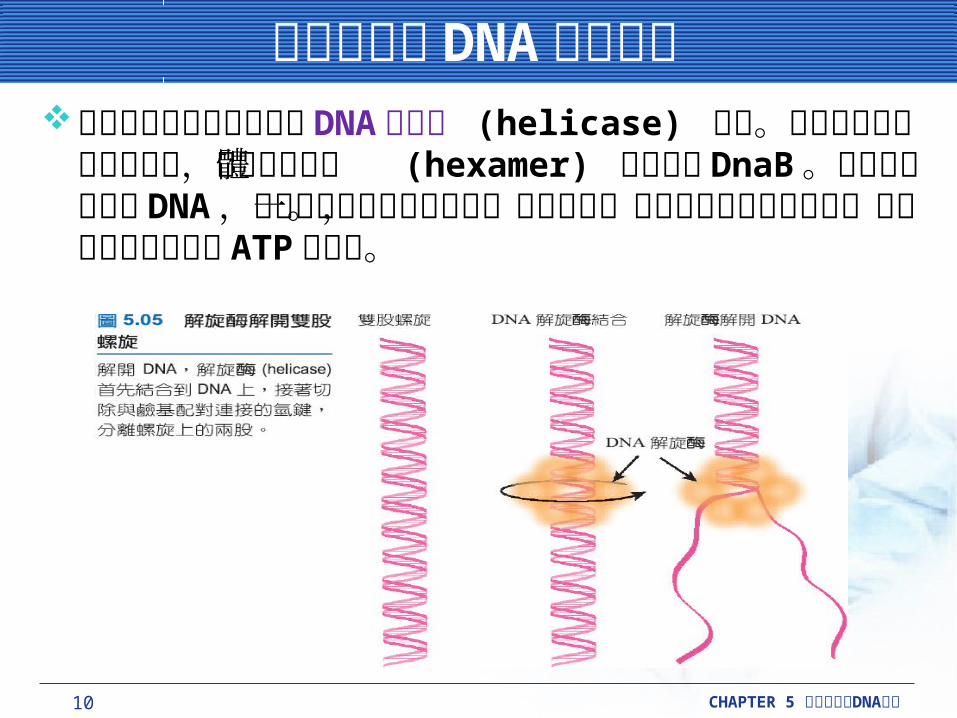
雙股分離在 DNA 合成之前雙股分離在 DNA 合成之前雙股螺旋構造會自動地被 DNA 解旋酶 (helicase) 解開。
大腸桿菌最主要的解旋酶,是形成六合體 (hexamer) 的蛋白質 DnaB 。解旋酶不會損毀 DNA ,只是干擾維持鹼基聚合在一起的氫鍵。此過程需要能量的供應,而能量由解旋酶降解 ATP 所提供。
11 CHAPTER 5 DNA細胞分裂與 複製
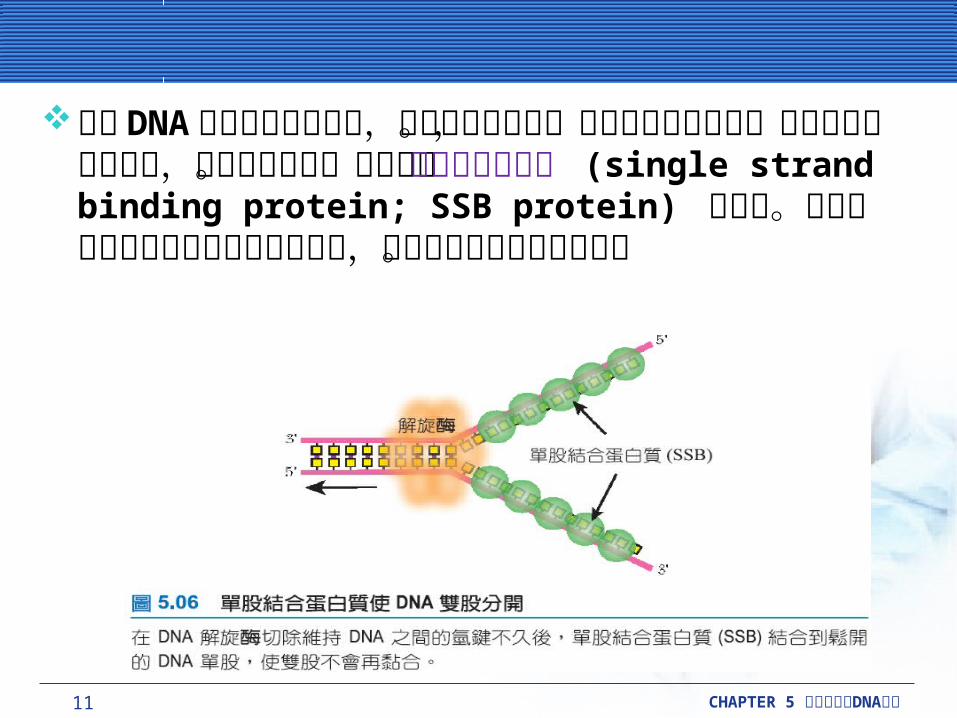
親代 DNA 分離的兩股為互補,所以可互相配對。為了製造出新的股,原來的兩股必須分離,當作模板使用。這工作由單股結合蛋白質 (single strand binding protein; SSB protein) 來完成。此蛋白質可以結合在未配對的單股上,避免兩段原來股重新黏合。
12 CHAPTER 5 DNA細胞分裂與 複製
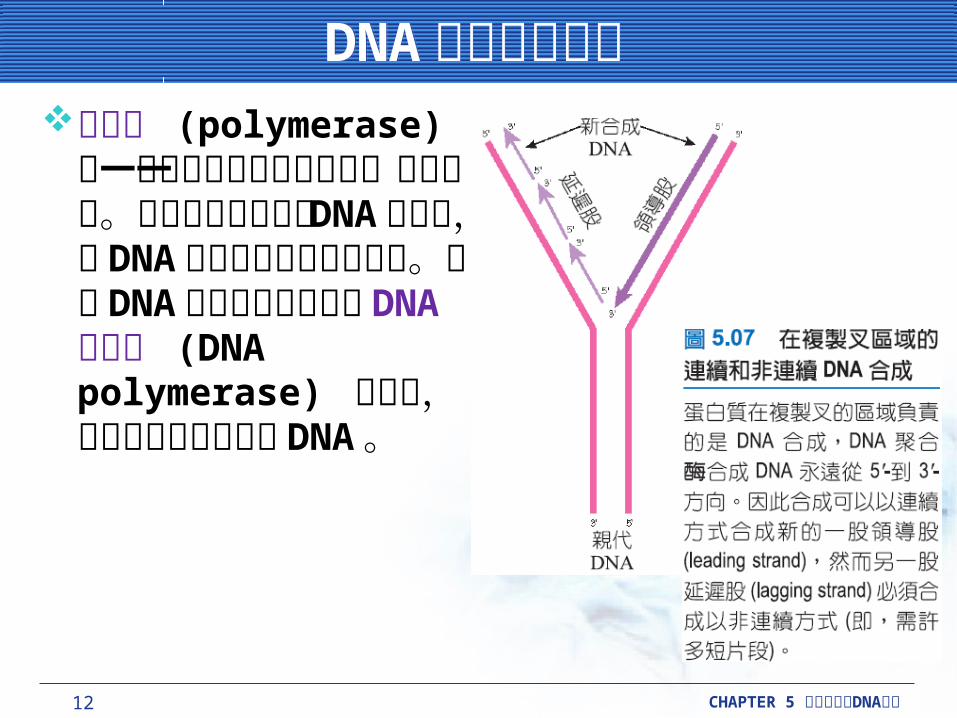
DNA 聚合酶的特性DNA 聚合酶的特性聚合酶 (polymerase) 為一種可以將核苷酸聚集在一起的酵素。細菌細胞包含幾種 DNA 聚合酶,對 DNA 複製與修復有不同功能。有關DNA 複製的問題根源於DNA 聚合酶 (DNA polymerase) 的特性,此酵素負責製造新的DNA 。

13 CHAPTER 5 DNA細胞分裂與 複製
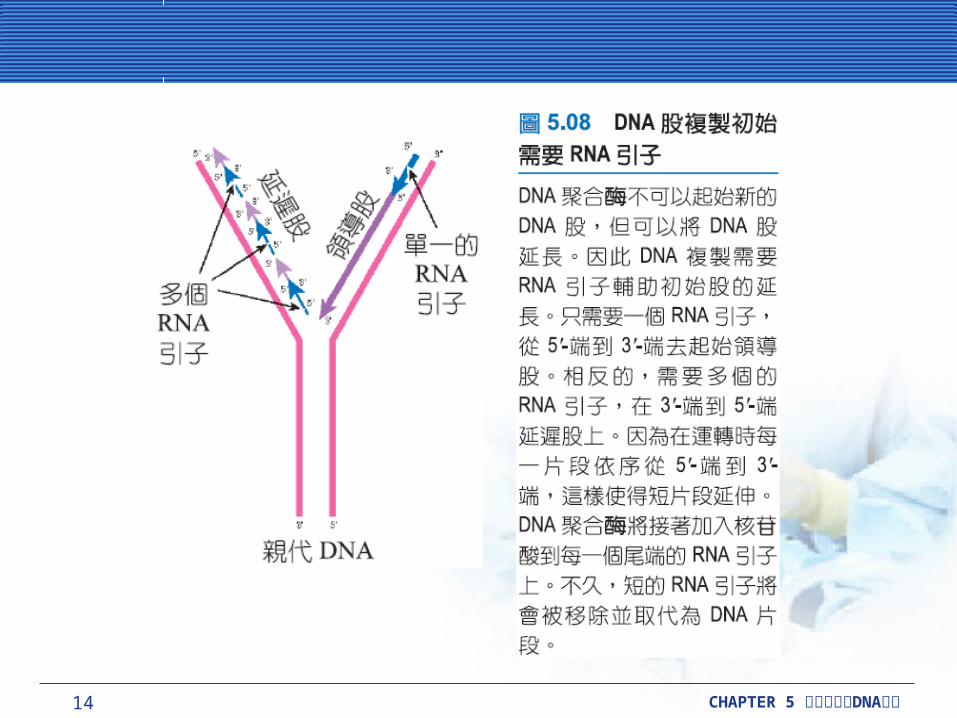
DNA 聚合酶利用其他酵素製造的短片段的 RNA 引子,去起始新股的合成。與 DNA 聚合酶不同的RNA 聚合酶 (RNA polymerase) 可以起始新股的合成。
一個特殊的 RNA 聚合酶,如引發酶 (primase) ;蛋白質 DnaG ,在細菌 DNA 合成製造期間,主要負責起始 DNA 股合成的 RNA 引子。
14 CHAPTER 5 DNA細胞分裂與 複製

15 CHAPTER 5 DNA細胞分裂與 複製
核苷酸的聚合核苷酸的聚合不論是 DNA 或 RNA ,所有核苷酸的合成方向皆為 5’-
至 3’- 。外加進來的核苷酸會附在 3’- 的羥基 (hydroxyl group) 上。
DNA 合成的前驅物質為去氧核糖核苷三磷酸或稱為去氧核糖核苷酸 (deoxyribonucleoside 5’-triphosphate; deoxy-NTP) ,包含 dATP 、 dGTP 、 dCTP 和 dTTP 。從去氧核糖的外部與功能區分,可以分為 α 、 β 與 γ磷酸根。當聚合時, α 與 β磷酸根之間的高能鍵結會被分開,釋出能量驅使聚合作用的進行。最外邊的兩個鄰酸根 (β 與 γ磷酸根 ) 會以焦磷酸根 (pyrophosphate) 的形態釋出。新的鍵結會在外加核苷酸最裡面的磷酸根 (α磷酸根 ) 形成,且增長在 DNA尾端,先前核苷酸的 3’-OH 也會有鍵結的形成
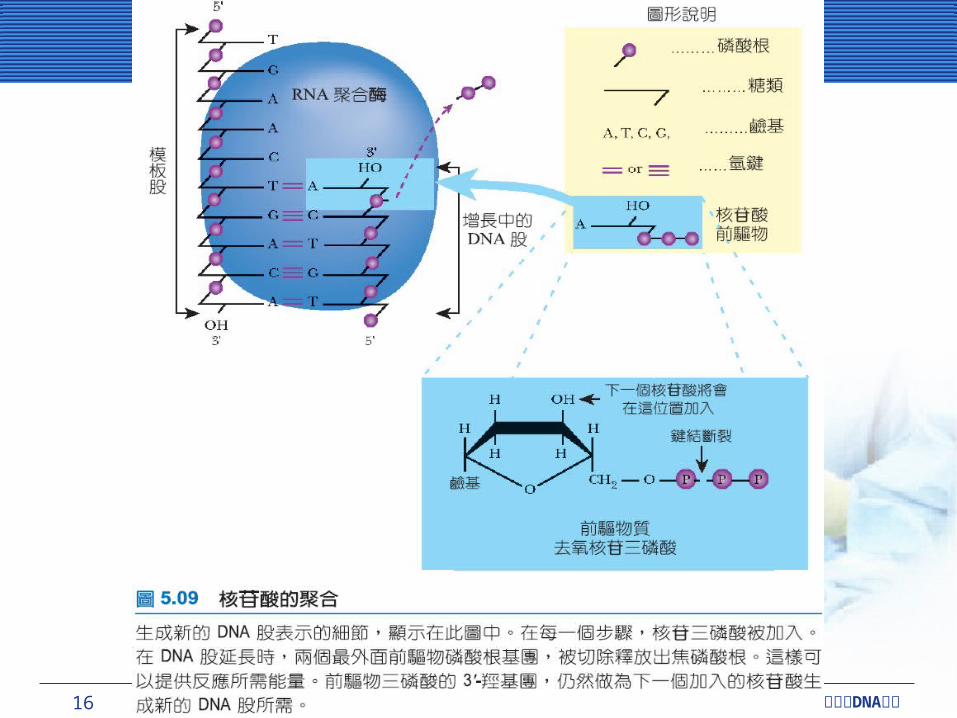
16 CHAPTER 5 DNA細胞分裂與 複製

17 CHAPTER 5 DNA細胞分裂與 複製
供應 DNA 合成的前驅物質供應 DNA 合成的前驅物質DNA 前驅物質包含去氧核糖,是由相對應的含核糖的核苷酸所合成的。
核糖核苷酸還原酶 (ribonucleotide reductase) 催化,使核糖 (ribose) 還原成去氧核糖 (deoxyribose) 。這方面也作用在雙磷酸根的衍生物 (ADP 、 GDP 、 CDP 和UDP) 。激酶 (kinase) 稍後會在dADP 、 dGDP 、 dCDP 上加入第三個磷酸根,形成dATP 、 dGTP 、 dCTP 。 dUDP則有不同的安排, DNA 沒有包括尿嘧啶 (uracil) ,但包含另一含有甲基的衍生物胸腺嘧啶 (thymine) 。在甲基化 (methylation) 進行之前, dUDP 會先移除一個磷酸根,轉變成 dUMP 。然後胸苷酸合成酶 (thymidylate synthetase) 加入一個甲基,所以 dUMP 會轉變成dTMP 。最後兩個磷酸根加入,形成 dTTP 。
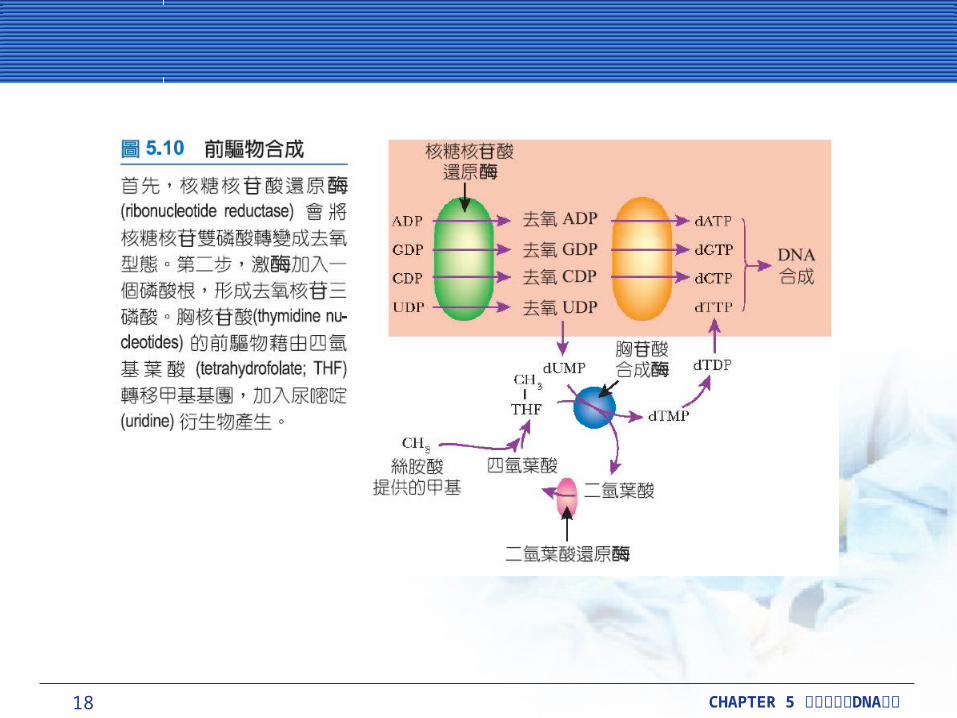
18 CHAPTER 5 DNA細胞分裂與 複製

19 CHAPTER 5 DNA細胞分裂與 複製
胸腺嘧啶的甲基由輔助因子 (cofactor) 四氫葉酸 (tetrahydrofolate; THF) 攜帶,而四氫葉酸會在反應中氧化成二氫葉酸 (dihydrofolate; DHF) 。 DHF必須還原成 THF ,當作 DNA 合成用。參與作用的酵素為二氫葉酸還原酶 (dihydrofolate reductase) ,在細菌中會被抗生素三甲氧二胺嘧啶 (trimethoprim) 所抑制。除此之外,合成嘌呤類物質,如腺嘌呤 (adenine) 和鳥糞嘌呤 (guanine) ,也需要 THF攜帶單碳的物質。

20 CHAPTER 5 DNA細胞分裂與 複製
真核的二氫葉酸還原酶會被甲氨喋呤 (methotrexate; amethopterin) 所抑制。因為腫瘤的生長會影響癌症細胞的快速分裂與 DNA 複製,因此甲氨喋呤常被用來做抗癌藥劑。
磺胺 (sulfonamide) 類的抗生素會主動抑制葉酸 (folate) 輔助因子的合成。動物體不能合成葉酸,但在飲食中必須攝取,所以在合理的劑量下,磺胺為無害的。

21 CHAPTER 5 DNA細胞分裂與 複製
DNA 聚合酶延長 DNA 單股DNA 聚合酶延長 DNA 單股DNA 聚合酶 III (polymerase III, Pol III) 是重要的酵素,主要是在染色體複製期間牽涉到 DNA 的延長。
滑動鉗子 (sliding clamp) 由兩個半圓形的 DnaN蛋白質次單元組成。它像一個門簾包圍著 DNA 模板股到處滑動,而且需要其他不同的蛋白質 ( 如HolA 、 B 、 C 和 D ,再加上 γ 次單元 ) ,這就是所謂的鉗子裝載複合物 (clamp-loading complex) 裝載這個滑動鉗子 (sliding clamp) 到 DNA 上。
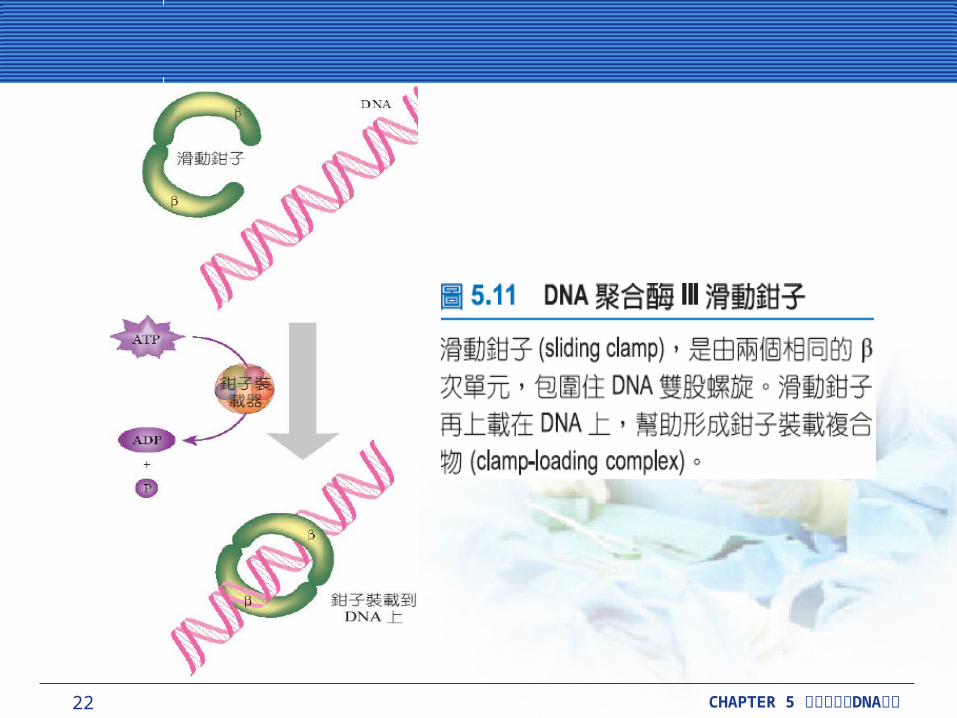
22 CHAPTER 5 DNA細胞分裂與 複製

23 CHAPTER 5 DNA細胞分裂與 複製
「滑動鉗子」會形成巨大複雜的核心酵素 (core enzyme) 到 DNA 上。核心酵素可合成 DNA ,包含三個小次單元, DnaE (α-次單元;聚合化 ) , DnaQ (ε-次單元;校正 ) 和 HolE (θ-次單元;功能未知 ) 。兩個核心結合後,其中一個主要負責製造新的 DNA 股,是利用一對 (tau) 次單位連接在一起。單一鉗子裝載複合物被兩個核心結合後共享。
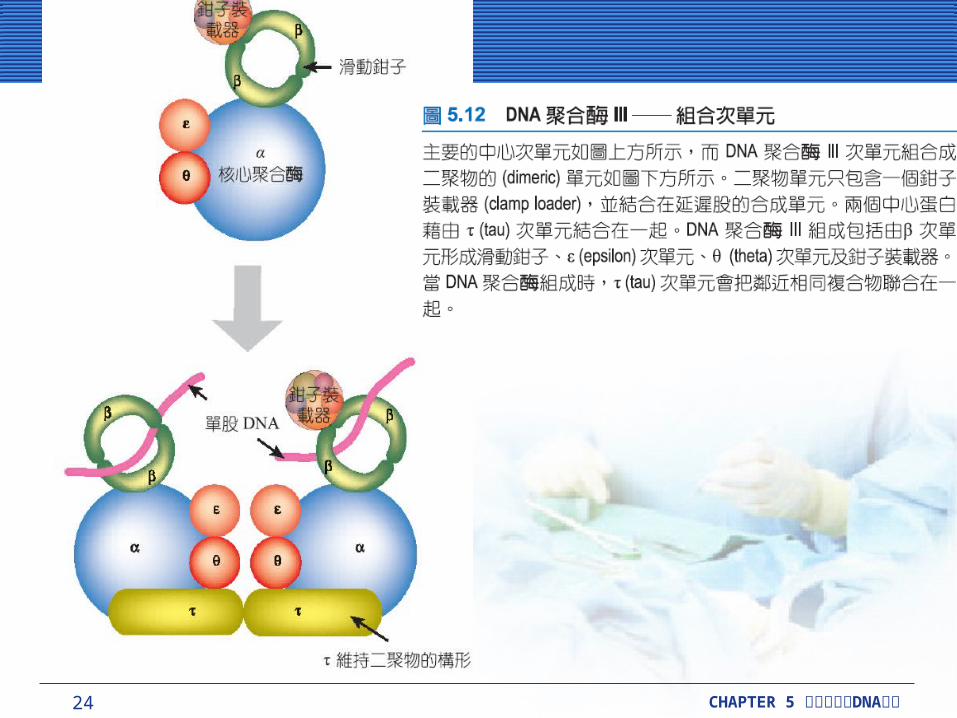
24 CHAPTER 5 DNA細胞分裂與 複製
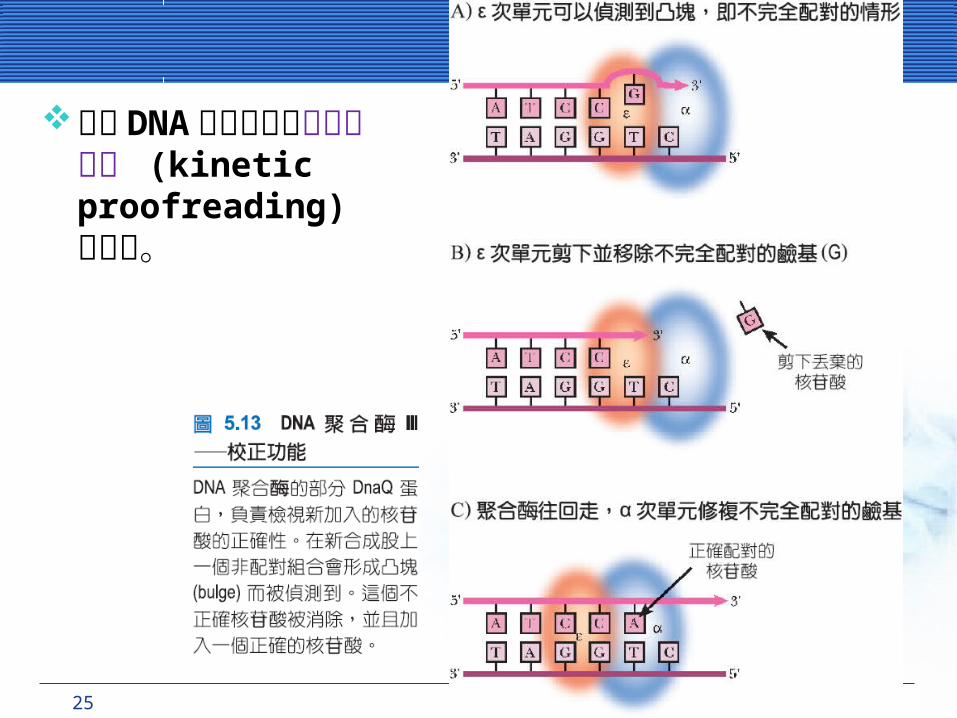
25 CHAPTER 5 DNA細胞分裂與 複製
許多 DNA 聚合酶具有動力學校正 (kinetic proofreading) 的功能。

26 CHAPTER 5 DNA細胞分裂與 複製
複製叉是複雜的複製叉是複雜的複製叉被定義為,在 DNA 分子被複製的這個範圍中所有結構的組成。它包括 DNA 被 DNA 促旋酶 (gyrase) 和解旋酶 (helicase) 解開的區帶,也包括單股 DNA延伸中,去幫助維持單股的單股結合蛋白質 (SSB) 。它也包括兩個 DNA 聚合酶 III 分子,這也製造出兩個新的 DNA 雙股。領導股持續地被製造出,而延遲股是製造成一段段 1000 ~ 2000鹼基長的小片段,這就是所謂的岡崎片段 (Okazaki fragments) 。
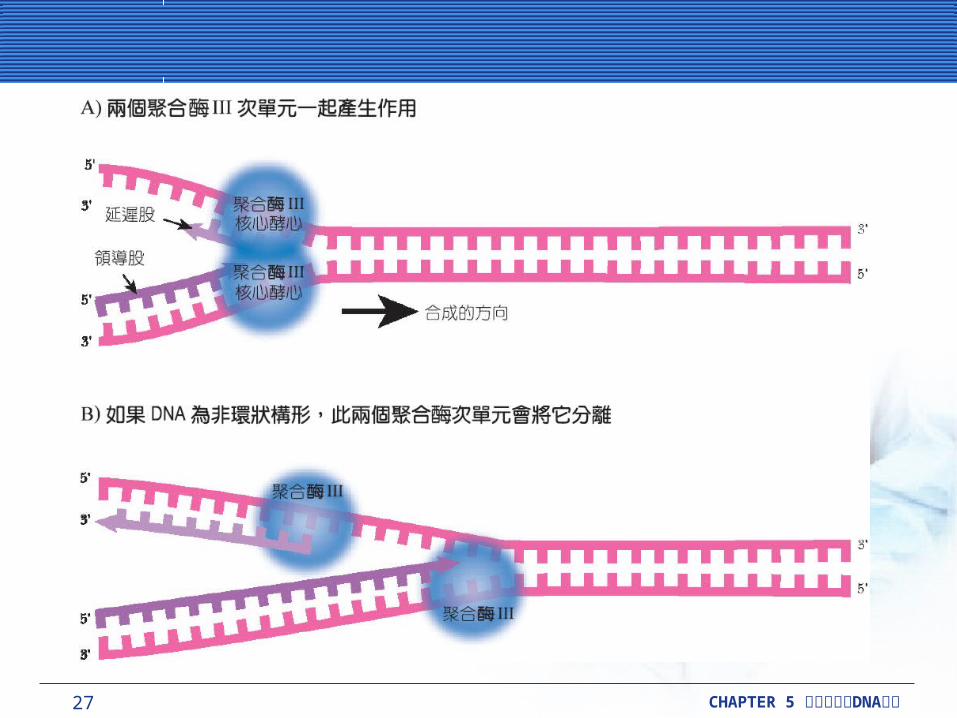
27 CHAPTER 5 DNA細胞分裂與 複製
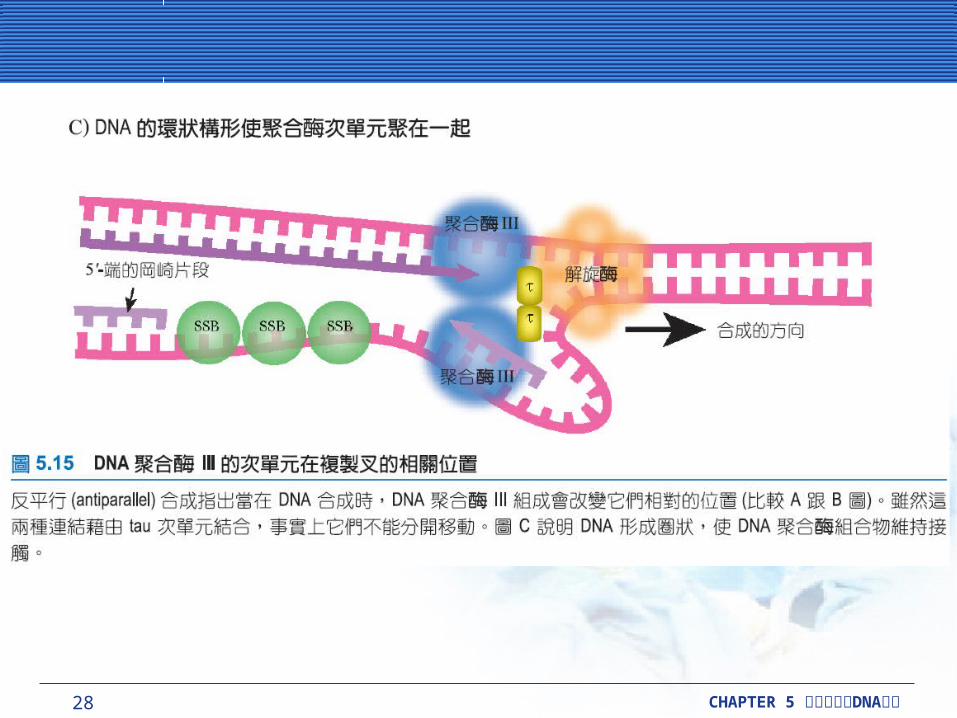
28 CHAPTER 5 DNA細胞分裂與 複製
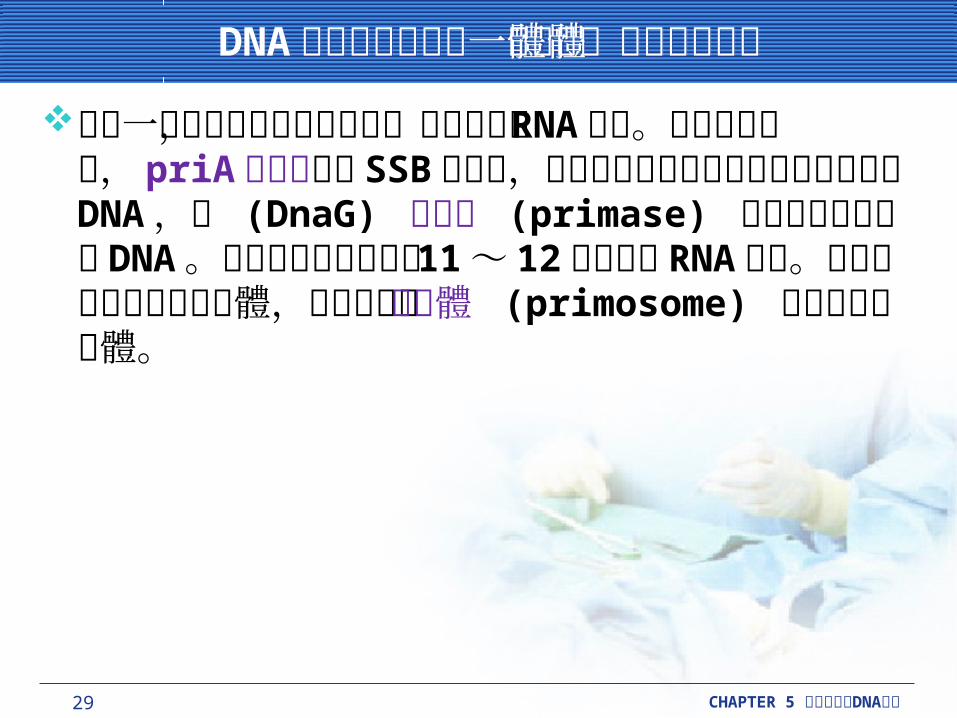
29 CHAPTER 5 DNA細胞分裂與 複製
DNA 間斷的合成需要一個引發體的蛋白複合體DNA 間斷的合成需要一個引發體的蛋白複合體
當每一個岡崎片段開始進行時,就必須要有 RNA引子。在開始進行時, priA 蛋白質取代 SSB 蛋白質,這個蛋白質有義務的去結合未配對的 DNA ,使 (DnaG) 引發酶 (primase) 能夠捆住延長中的DNA 。引發酶能夠製造出約 11 ~ 12鹼基長的RNA 引子。這個初期所形成的複合體,有時被稱作引發體 (primosome) 的蛋白質複合體。
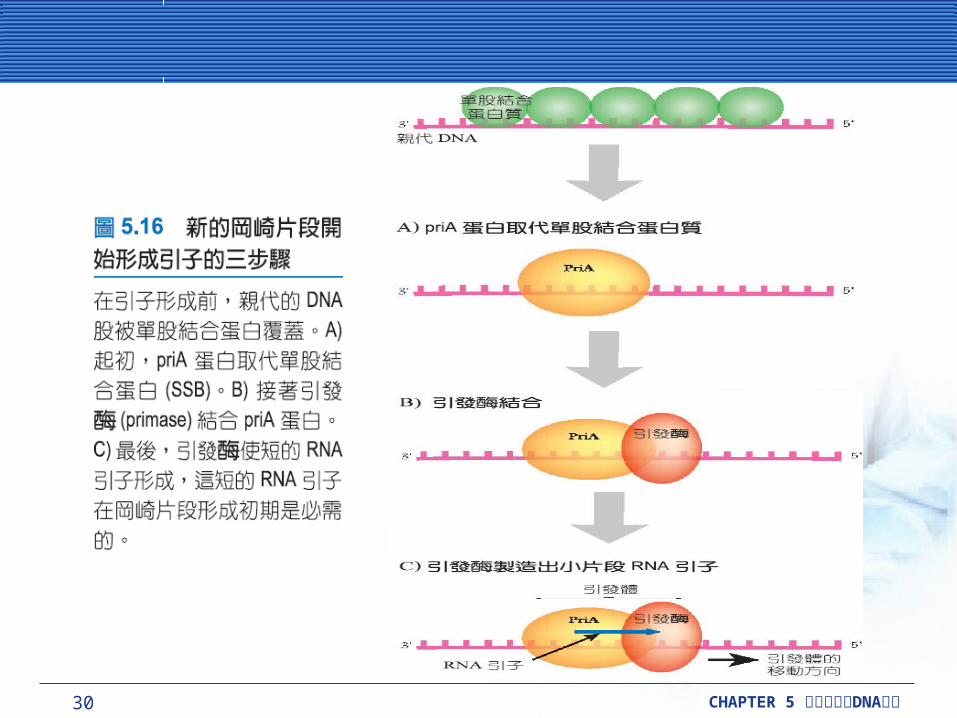
30 CHAPTER 5 DNA細胞分裂與 複製

31 CHAPTER 5 DNA細胞分裂與 複製
延遲股的完成延遲股的完成連接岡崎片段形成完整的 DNA需要 2 或者 3 酵素,接替順序為:核糖核酸酶 H (ribonuclease H, RNase H) 、 DNA 聚合酶 I (DNA polymerase I, Pol I) 與DNA 接合酶 (DNA ligase) 。
DNA 聚合酶 I 會分解 RNA 引子,且會利用分解的 RNA 來填滿缺口,最後 DNA 接合酶連結尾端。RNA 雙股螺旋中的 RNA 股,會移除每個 RNA 引子,而且 DNA 聚合酶 I只會移除 RNA 引子的最後幾個鹼基。
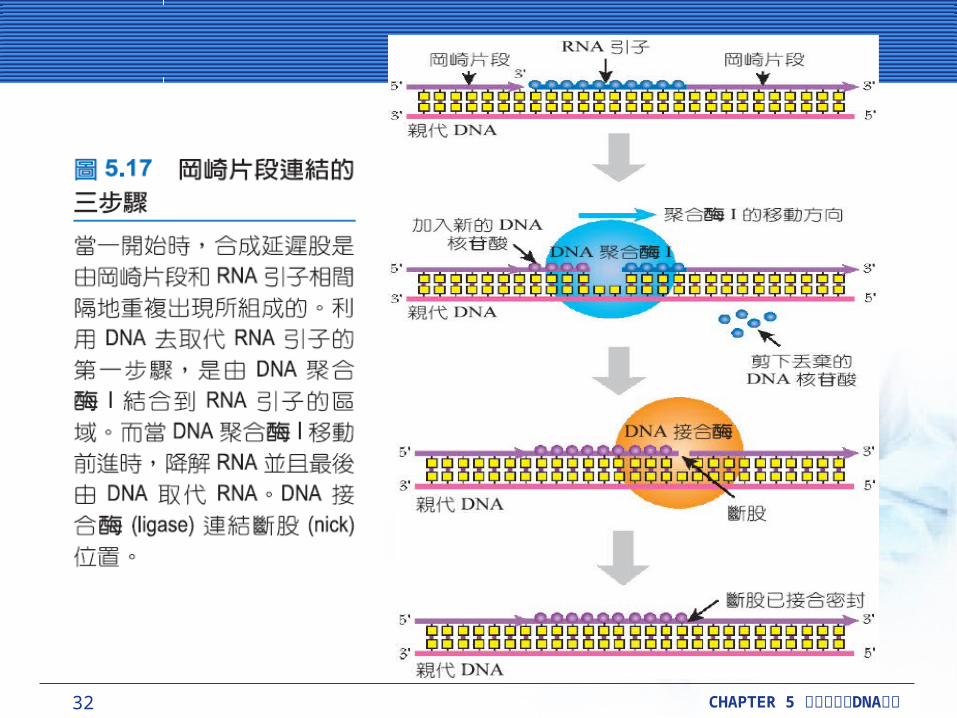
32 CHAPTER 5 DNA細胞分裂與 複製

33 CHAPTER 5 DNA細胞分裂與 複製
DNA 聚合酶 I 具有像 Pol III 的動力學校正 (kinetic proofreading) 能力。
DNA 聚合酶 I 起始於斷股缺口,然後沿著 DNA移位,並切開核苷酸的 5’- 端方向,再用放射性核苷酸來取代。而 DNA 聚合酶 I 和 DNA 接合酶,在遺傳工程扮演很重要的角色。

34 CHAPTER 5 DNA細胞分裂與 複製
染色體複製起始於 oriC染色體複製起始於 oriC原核染色體 DNA 複製時,起始在染色體一個特別的地方,我們稱作複製起始點 (origin of replication) ,在環狀的兩個方向進行。
這個起始複合物 (initiation complex) 包括 5 個蛋白質: DnaA 、 DnaB 、 DnaC、促旋酶和 SSB 。這些只有 DnaA 有一個特別的染色體起始點,其餘的只牽涉到新合成的岡崎片段。這個起始點即 oriC ,有三次 13-鹼基重複 (13-base repeat) ,這些鹼基包括 GATCTNTTNTTTT ,接著是四次 9-鹼基重複,為 TTATNCANA 。這些序列都有 AT-rich ,是這些序列的特色,有助於分辨這些序列。
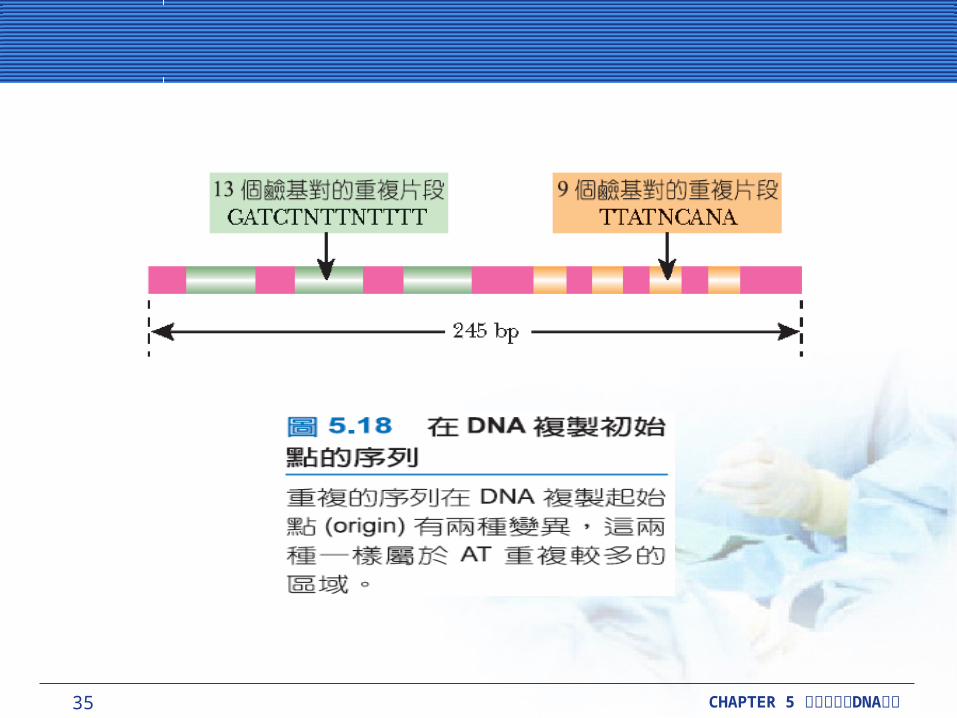
35 CHAPTER 5 DNA細胞分裂與 複製

36 CHAPTER 5 DNA細胞分裂與 複製
首先, DnaA 蛋白質 (DnaA protein) 和四次的 9-鹼基重複結合。約有 30 個叢聚的 DnaA 蛋白質結合在 oriC ,並且纏繞它們。
接下來, DnaA 蛋白質打開 DNA 所有的三次 13-鹼基重複區域。
六次方體的 DnaB 解旋酶,加入伴隨著 6 個 DnaC蛋白質,需要裝載 DnaB 蛋白質到 DNA 上。然後DnaB 蛋白質從三次 13-鹼基重複區域中取代了DnaA 蛋白質,並開始解開 DNA 而產生了複製叉。第二個 DnaB 蛋白質六次單元,也產生了一個複製叉,往相反的方向移動。

37 CHAPTER 5 DNA細胞分裂與 複製

38 CHAPTER 5 DNA細胞分裂與 複製
DNA甲基化和附著到細胞膜可以控制複製初始DNA甲基化和附著到細胞膜可以控制複製初始
控制複製初始, DNA 的甲基化在初始位置和附著到細胞膜。這個 oriC 區域包含全部 11 個可以被Dam甲基酶辨識的 GATC 序列。這些迴文的 (palindromic) 區域被 Dam甲基酶甲基化在 DNA雙股上的腺嘌呤。在複製前,每一個 GATC 在大腸桿菌染色體上,包括這些初始的複製區域,將被完全甲基化。複製之後立即將先前的 DNA 股甲基化,但新合成的 DNA 股還未甲基化。因此 DNA 是半甲基化的 (hemi-methylated) 。
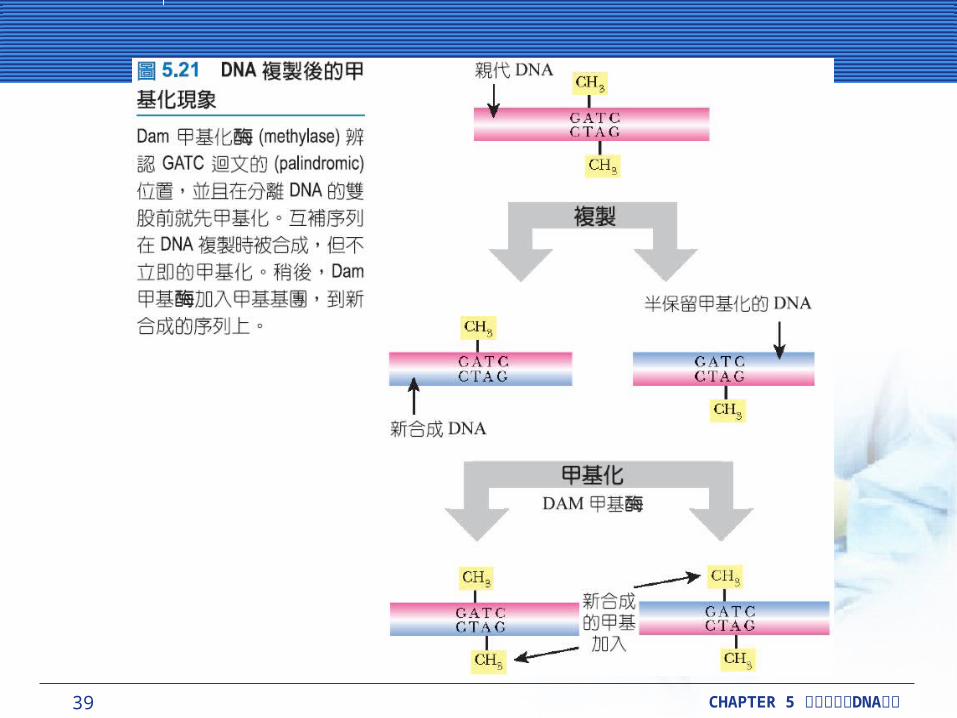
39 CHAPTER 5 DNA細胞分裂與 複製

40 CHAPTER 5 DNA細胞分裂與 複製
當在半保留甲基化的時候基因的轉錄被抑制,初始複製反應時整體的 DnaA 蛋白量減少。半甲基化位置不可以使用在初始複製反應。半保留甲基化DNA 會藉由隔離蛋白 A (SeqA, sequestration protein) 的幫助,結合到細胞膜上,而完全甲基化的 DNA則不會。
41 CHAPTER 5 DNA細胞分裂與 複製

42 CHAPTER 5 DNA細胞分裂與 複製
染色體複製結束在 terC染色體複製結束在 terC當兩個複製叉遇到染色體的終點 (terminus) ,複製結束。這區域附近有許多終點位置 (Ter site) ,防止複製叉前進移動 。 TerC 、 TerB 、 TerF防止複製叉以順時針方式移動,而 TerA 、 TerD 和 TerE防止複製以逆時針針方式移動。
43 CHAPTER 5 DNA細胞分裂與 複製
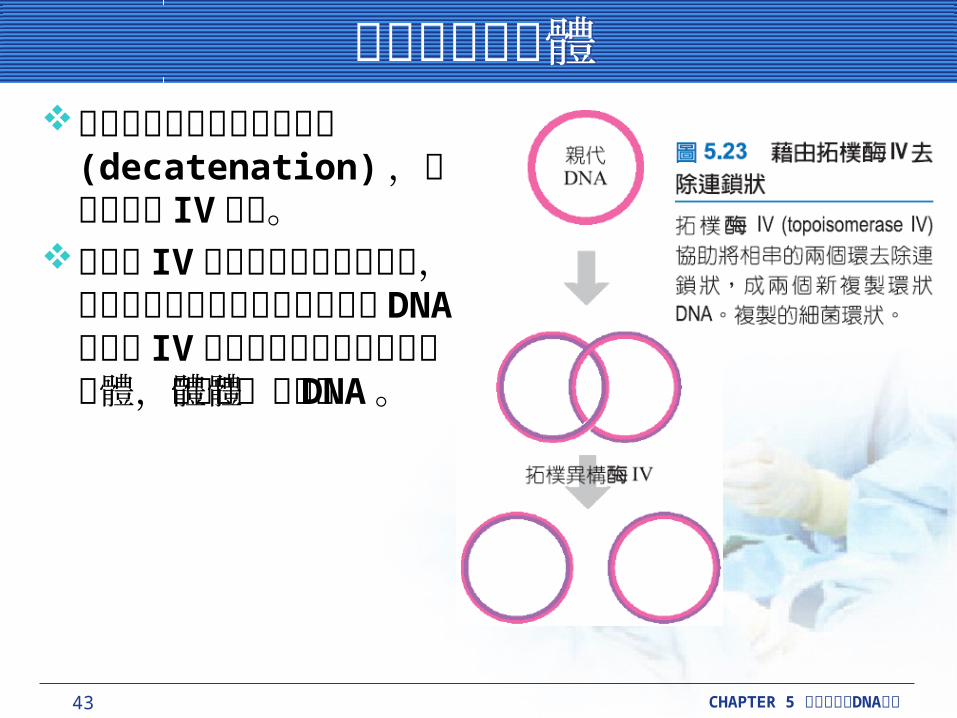
鬆開子代染色體鬆開子代染色體將連成鎖狀的環進行反連鎖 (decatenation) ,需要拓樸酶 IV 作用。
拓樸酶 IV 在複製叉的後方被發現,當複製進行時可以鬆開新合成的DNA 。拓樸酶 IV 也可以解開完成複製的環狀體,如染色體或質體DNA 。

44 CHAPTER 5 DNA細胞分裂與 複製

45 CHAPTER 5 DNA細胞分裂與 複製
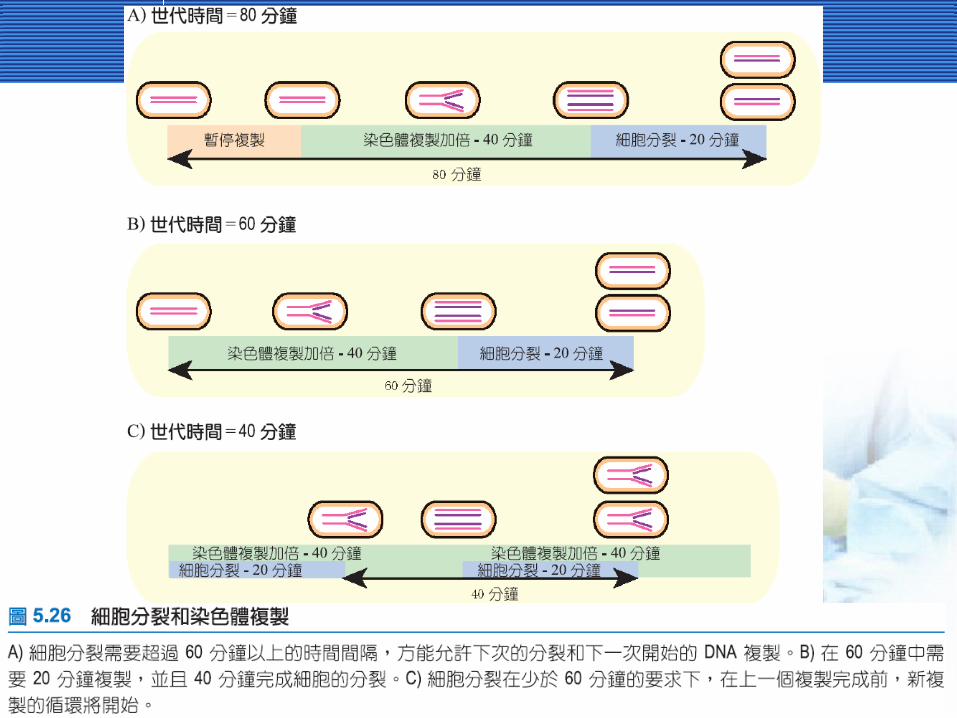
在染色體複製後才發生細菌細胞分裂在染色體複製後才發生細菌細胞分裂 細菌的細胞分裂相對來的簡單,並且簡易分成四
個時期。1. 複製染色體。2. 分開子代染色體。3. 細胞延長。4. 形成細胞壁,分離兩個子代細胞。

46 CHAPTER 5 DNA細胞分裂與 複製
47 CHAPTER 5 DNA細胞分裂與 複製
細菌花了多少時間進行複製細菌花了多少時間進行複製

48 CHAPTER 5 DNA細胞分裂與 複製
複製子的概念複製子的概念複製子 (replicon) 是在細胞中任何的 DNA ( 或
RNA) 分子,能夠存活和自我複製。伯氏疏螺旋菌 (Borrelia burgdorferi) ,造成萊姆症
(Lyme disease) ,它有直線狀染色體的末端髮夾狀序列。變鉛青鏈黴菌 (Streptomyces lividans) ,一種土壤中的生物體,有蛋白質以共價鍵的結合至染色體末端。
質體 (plasmid) 是另一群的複製子 (replicons) 。它們是屬於多餘的自我複製分子,對於宿主細胞生存是不需要的 。

49 CHAPTER 5 DNA細胞分裂與 複製
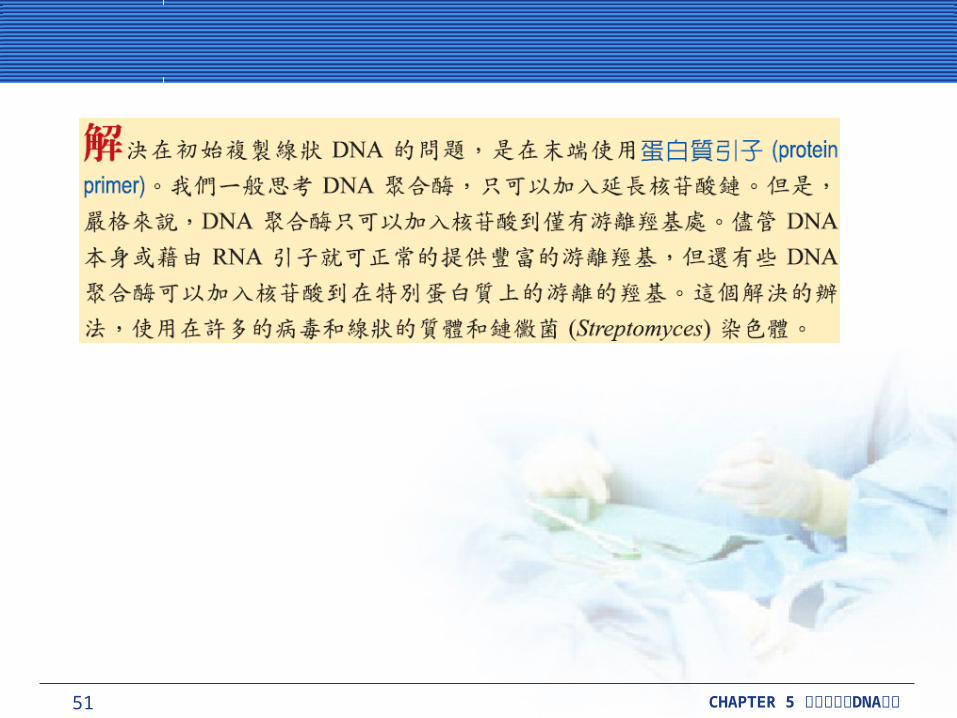
在真核細胞中複製直線狀 DNA在真核細胞中複製直線狀 DNA雖然 DNA 聚合酶只可以延長,而非初始,但新合成的 DNA 複製初始必須有 RNA 引子輔助。因為合成 DNA 的方向是從 5’- 到 3’- ,當線狀的 DNA 複製時,每個 RNA 引子必須正確位於 5’-末端的每個新合成 DNA 股。當末端 RNA 引子被移除時, RNA 引子結合的位置不會有 DNA 取代, DNA 聚合酶不會結合在此位置並做延長。如果沒有辦法去解決這樣的問題,每回合複製時, DNA 分子將縮短一個 RNA 引子長度。環狀原核細胞 DNA 分子,沒有末端所以沒有這樣的問題。
真核細胞為適應這樣複製線狀 DNA 問題,發展出一套位於染色體尾端稱為端粒 (telomere) 的構造。

50 CHAPTER 5 DNA細胞分裂與 複製
51 CHAPTER 5 DNA細胞分裂與 複製
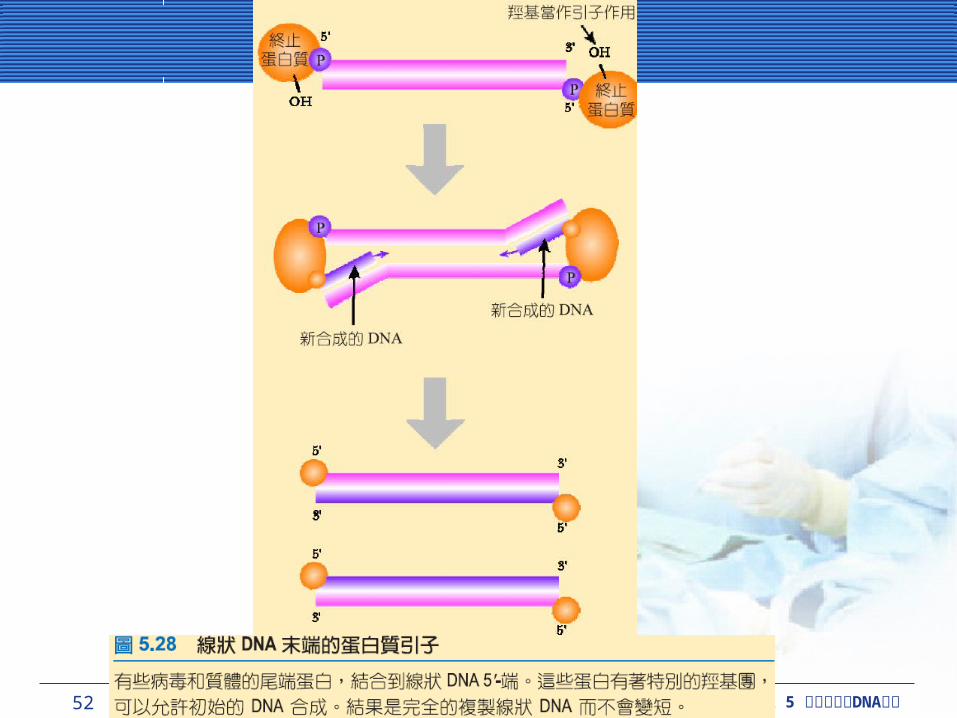
52 CHAPTER 5 DNA細胞分裂與 複製

53 CHAPTER 5 DNA細胞分裂與 複製
端粒包含多種連續重複片段的短序列,通常是 6-鹼基 (TTAGGG ,在脊椎動物身上包括人類 ) 。在每一個複製週期,染色體縮短並且許多端粒重複序列會遺失。此外,在每個週期之後細胞在端粒酶(telomerase) 存在時,遺失的 DNA稍候被加入許多 6-鹼基對重複序列到 3’-末端。端粒酶帶著小片段的 RNA ,配對到 6-鹼基對的端粒重複序列,讓端粒酶可以是辨識端粒序列,並且提醒端粒酶哪一段序列應該加入。
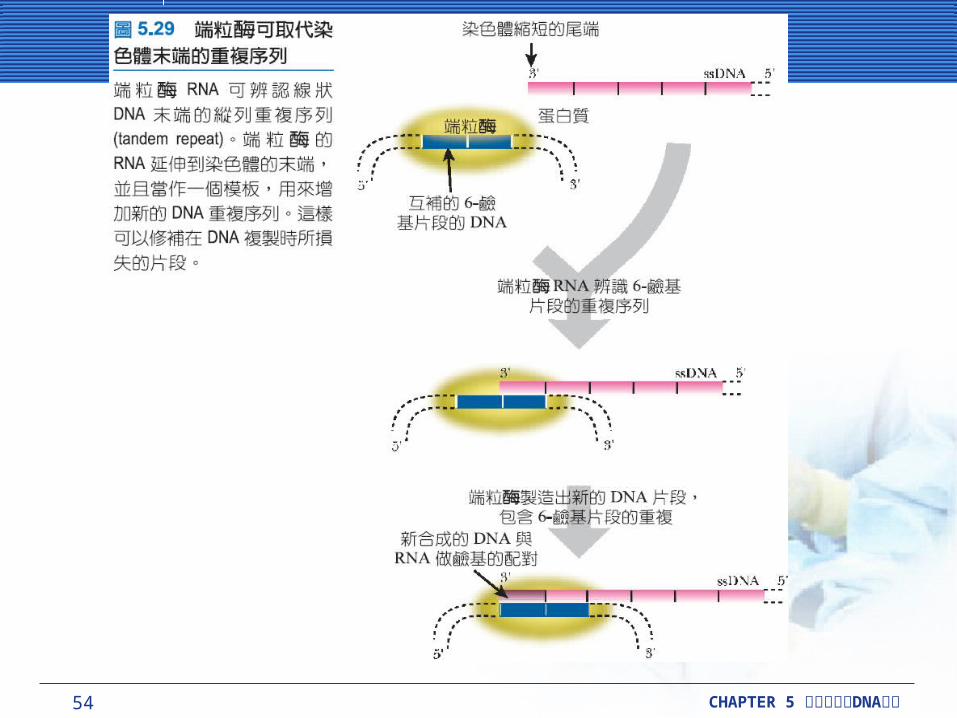
54 CHAPTER 5 DNA細胞分裂與 複製

55 CHAPTER 5 DNA細胞分裂與 複製
在端粒酶延長 3’- 端後,可以藉由正常的 RNA 引子啟動 (priming)、利用 DNA 聚合酶延伸和接合酶 (ligase) 相連,補入配對股。端粒重複片段也可以保護染色體末端,以防被外切酶降解。

56 CHAPTER 5 DNA細胞分裂與 複製
真核細胞染色體有多個的複製起始點真核細胞染色體有多個的複製起始點真核染色體通常很長並有許多複製起始點
(replication origins) ,沿著每一條染色體分散。在細菌中,複製是雙向的。一對複製叉開始於每個複製初始點,並且兩個複製叉接著移動相反方向。這些凸塊 (bulges) 在 DNA 進行分裂過程,通常被稱為複製泡 (replication bubble) 。
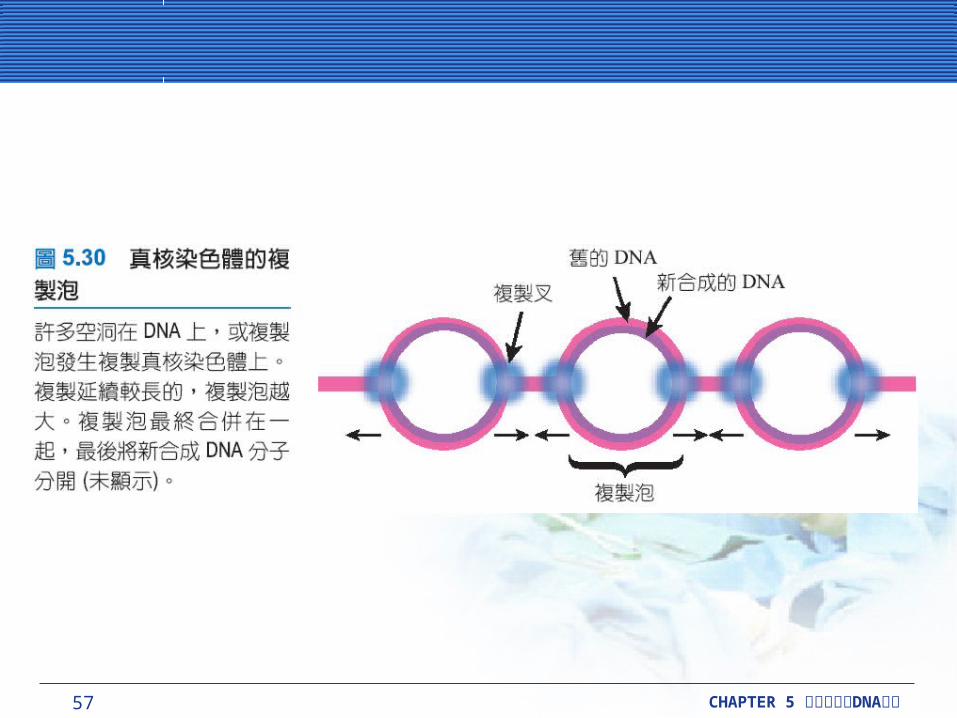
57 CHAPTER 5 DNA細胞分裂與 複製

58 CHAPTER 5 DNA細胞分裂與 複製
為了避免已經複製的 DNA 片段加倍的情況,每一個起始複製點,只能在每一個複製的循環中複製一次。這樣的情況藉由一個蛋白質複合物稱為複製許可因子 (replication licensing factor, RLF) 來達成,使得在下一個複製開始前 RLF 結合到每一個複製起始點 DNA 上,並在複製時被取代。只有當 RLF蛋白存在時, DNA 複製才可以進行。

59 CHAPTER 5 DNA細胞分裂與 複製
真核細胞 DNA 複製真核細胞 DNA 複製在真核細胞,有半保留複製 (semi-conservative
replication) 的發生。在動物細胞中,有兩個 DNA 聚合酶 (α 和 δ) 參與染色體的複製。 DNA 聚合酶 α(polymerase α) 是負責初始複製新的 DNA 股。它會伴隨著兩個小的蛋白質用來生成 RNA 引子。在 RNA 引子生成後, DNA 聚合酶 δ (polymerase δ) 會藉由三到四鹼基長度的 DNA ( 起始 DNA ; initiator DNA 或iDNA) 從 RNA 引子上做延長。
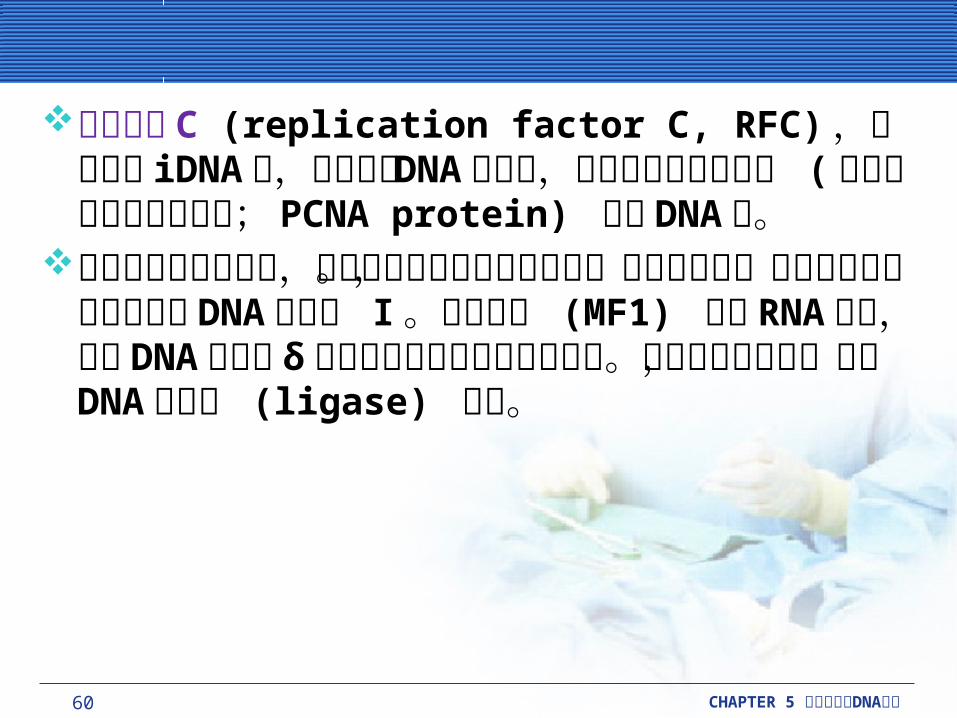
60 CHAPTER 5 DNA細胞分裂與 複製
複製因子 C (replication factor C, RFC) ,會結合到iDNA 上,並且搭載 DNA 聚合酶,加上本身的滑動鉗子 (增生細胞核抗原蛋白質; PCNA protein) 附在 DNA 上。
在動物和細菌細胞中,連結岡崎片段有顯著的不同。在動物細胞,沒有和細菌相等的雙功能 DNA 聚合酶 I 。藉外切酶 (MF1) 移除 RNA 引子,並由DNA 聚合酶 δ 行使功能在延遲股上填補間隔。而在細菌中間隔,是以 DNA 接合酶 (ligase) 填補。

61 CHAPTER 5 DNA細胞分裂與 複製
高等生物體的細胞分裂高等生物體的細胞分裂 個過程稱為有絲分裂,包含了許多運轉過程:
1.拆解母細胞的核膜2.染色體分裂3.染色體分開4.再建構兩個圍繞染色體的核膜5.母細胞的最後分裂,或細胞質分裂 (cytokinesis) 。

62 CHAPTER 5 DNA細胞分裂與 複製
有絲分裂的本身,只是真核細胞多時期細胞週期 (cell cycle) 中的某一個時期。 DNA 進行複製發生於細胞週期中的複製期或稱 S-期 (S-phase) 。 S期被分隔在真實自然的步驟即細胞分裂 ( 有絲分裂,mitosis) ,有兩個間隔時期或稱為 G-期 (G-phases) ,此時沒有任何較多的事件發生 ( 除了正常程序下的細胞活動和代謝外 ) 。 G1 、 S 和 G2一起構成細胞週期的間期 (interphase) 。
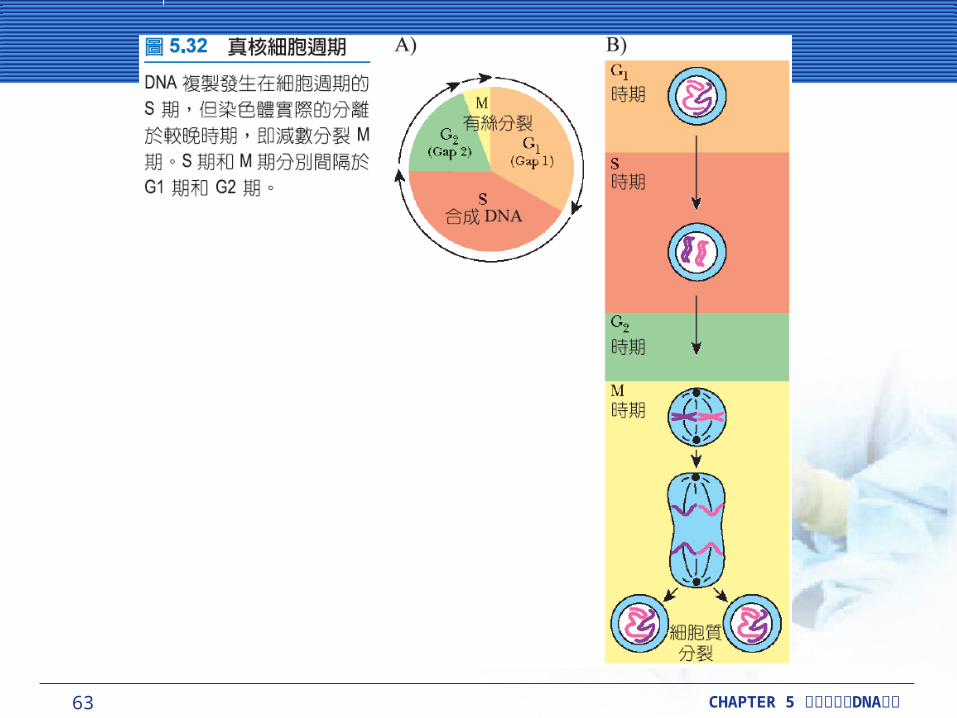
63 CHAPTER 5 DNA細胞分裂與 複製