Embed Size (px)
Citation preview

51
3-1.生物は細胞の分裂により生きている「細胞周期」
1個の受精卵から細胞は分裂を繰り返して増殖し、成人では 60 兆個もの
細胞になります。さらに、生きている間、私たちの体では、毎日多くの体細
胞が死んでいきます。生物が生きていくためには、たえず新しい細胞が作ら
れなくてはいけません。
私たちの体では、特定の細胞が細胞分裂を繰り返して新しい細胞を作って
います。このような細胞は、細胞分裂を行う分裂期(M 期)と分裂を行わな
い間期を繰り返しています。この周期性を細胞周期といいます。細胞周期に
おいて間期はさらに G1 期※ 1 ⇒ S 期⇒ G2 期※ 2 に分けられます。細胞が分裂
する際には、G1 期に DNA の合成の準備が行われ、S 期に DNA が複製されま
す。その後、細胞は G2 期にさらに成長して分裂の準備をします。G2 期の細
胞の核内には G1 期の 2 倍量の DNA が存在します。
細胞周期の長さは細胞によって異なりますが、ヒトの細胞が増殖する際の
細胞周期は 1 日程度で、そのうち S 期は 6 〜 8 時間、M 期は 1 時間程度です。
肝臓の細胞は通常は細胞分裂を停止しており(G0 期)、1 〜 2 年に 1 回しか
分裂しませんが、肝臓が切除されると G1 期に戻り、細胞分裂を繰り返します。
一方、腸の上皮細胞には毎日 2 回以上分裂するものがあります。多くの体細
胞は細胞周期を離れており、決まった役割を担うようになっています。
※ 1 G1 期:細胞外からの情報を感知して、DNA 複製期(S 期)に進むか、休止期(G0 期)に入るか、
分化するか、アポトーシス(参照:8-1.プログラムされた死「アポトーシス」p.87)に進むかを決
定します。
※ 2 DNA 複製が正常に終了した事を確認し、M 期に進行します。
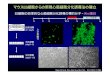
蛍光プローブ "Fucci" を用いた細胞周期のリアルタイム観察野沢-上阪 :供提ご真写 朝子 先生 , 宮脇 敦史 先生
(独立行政法人 科学技術振興機構 ERATO 宮脇生命時空間情報プロジェクト、独立行政法人 理化学研究所 脳科学総合研究センター 細胞機能探索技術開発チーム)
G1 G1 / S S G2 M
Fucci-S/G2/MGreen
Fucci-G1 Orange
merge+DIC
BrdU
10 μm
3. 細胞周期ー遺伝情報の再分配遺伝情報は細胞から細胞へと伝えられる
Pick Up from MBL
細胞周期のリアルタイム観察
Fucci(フーチ)
Fucci (Fluorescent Ubiqutination-
based Cell Cycle Indicator) は細胞
周期の進行を " リアルタイム " に
観察できる蛍光プローブです。S/
G2/M 期に緑色、G1 期にオレンジ
色の蛍光が核に観察されます。
Fucci は、S/G2/M 期に増加し G1
期には存在しない Geminin と、G1
期に増加し S/G2/M 期には存在し
ない Cdt1 という 2 つのタンパク
質に、それぞれ緑色 (monomeric
Azami-Green1) と オ レ ン ジ 色
(monomeric Kusabira-Orange2)
の蛍光タンパク質を融合して細胞
周期を可視化しています。
細胞の増殖や分化、がん細胞の挙
動などの生命現象における、細胞
周期の時間的、空間的なパターン
を解析することができます。
MG2
SG1

細胞周期調節に関わる種々のサイクリンと CDK
細胞周期エンジンの活性制御
52
3-2.細胞分裂のタイミングを決めるタンパク質「サイクリン」細胞周期で重要なことは正確に遺伝情報を複製し、2 つの細胞に分配するこ
とです。DNA に損傷がないか、DNA の複製が完了しているかなど、様々な段
階で細胞周期のチェックが行われています。
細胞周期のそれぞれのステージへの移行(チェックポイントの通過)に関与
するのが、サイクリン -Cdc2 キナーゼ※ 1 複合体で、細胞周期のエンジンとも
呼ばれています。サイクリンは細胞周期のある時期でのみ発現されて、Cdc2
キナーゼを活性化し、その後すぐに分解されます。サイクリンは数種類(ヒト
では 15 種類)存在し、それぞれ働く時期が異なり、細胞周期に応じて濃度が
変化します。Cdc2 キナーゼにも類似のタンパク質が複数存在し、まとめてサ
イクリン依存リン酸化酵素 (Cyclin-dependent protein kinases: CDKs) と名
づけられています(図.細胞周期調節に関わる種々のサイクリンと CDKs)。
ヒトでは、4 種類の細胞周期エンジンが働いています。例えば、G2 期から
M 期への移行では、まずサイクリン B が CDK1 に結合して複合体を形成しま
す。CDK1 は、2 つの自己リン酸化されたアミノ酸残基(14 番目のトレオニ
ンと 15 番目のチロシン)が、Cdc25 によって両方とも脱リン酸化されるこ
とにより活性化し、M 期へと移行します。また、M 期の終了にはサイクリン
B の分解が必須です(図.細胞周期エンジンの活性制御)。
一方、細胞周期エンジンのブレーキとして備わっているのが CKI というタン
パク質です。CKI は CDK やサイクリン -Cdc2 キナーゼ複合体に結合すること
により、サイクリン -Cdc2 キナーゼ複合体の活性を阻害します。CKI が正常
に働かなくなると、サイクリン -Cdc2 キナーゼ複合体の活性が暴走して、細
胞は際限なく分裂し始め、がん細胞となります。
※ 1 キナーゼ:アミノ酸をリン酸化する酵素。
タンパク質の機能調節とリン酸化・脱リ
ン酸化については p.66 をご覧下さい。
Pick Up from MBL
細胞周期関連キナーゼ活性測定キット
細胞周期、細胞分裂等の様々な細
胞内シグナル伝達に関与するキ
ナーゼの異常は、多くのがんで見
られます。これらに特異的なアク
チベーター、インヒビターは疾患
の治療薬として注目されており、
キナーゼ活性測定は、新規治療薬
開発の優れた指標になると考えら
れています。
CycLex 社ではこれらの分子やキ
ナーゼ活性を簡単、迅速に測定で
きる ELISA キットを開発・販売し
ています。
G1 期
S 期
G2 期
サイクリン D-CDK4/6
サイクリン B-CDK1
サイクリンのユビキチン化P P
UUUU
サイクリン分解サイクリン分解
CDK不活性型
サイクリンの分解によりCDK は不活化します。
Cdc25
オーロラ A
サイクリン D1-CDK4サイクリン E-CDK2
活性化されたサイクリン -CDK 複合体
P
チェックポイントの通過
P
PP
CDK
サイクリンリン酸化
阻害タンパク質
脱リン酸化
不活性型CDK 活性型
Cdc7-ASK
染色体分離
適応 腫瘍形成
M期破壊
DNA 複製開始
Chk1/Chk2
ATM/ATR/DNA-PK
サイクリン A-CDK2
PLK1
細胞周期の進行とサイクリン-CDKの活性制御
M 期
G1 期
S 期
G2 期
サイクリン D-CDK4/6
サイクリン B-CDK1
サイクリンのユビキチン化P P
UUUU
サイクリン分解
CDK不活性型
サイクリンの分解によりCDK は不活化します。
Cdc25
オーロラ A
サイクリン D1-CDK4サイクリン E-CDK2
活性化されたサイクリン -CDK 複合体
P
チェックポイントの通過
P
PP
CDK
サイクリンリン酸化
阻害タンパク質
脱リン酸化
不活性型CDK 活性型
Cdc7-ASK
染色体分離
適応 腫瘍形成
M期破壊
DNA 複製開始
Chk1/Chk2
ATM/ATR/DNA-PK
サイクリン A-CDK2
PLK1
細胞周期の進行とサイクリン-CDKの活性制御
M 期

DNA と染色体
53
3-3.膨大な遺伝情報をコンパクトに収納「染色体の構造」
真核細胞では、ミトコンドリアや葉緑体の DNA を除き、大部分の DNA は
核に詰め込まれています。核内の DNA は、様々なタンパク質と結合して「ク
ロマチン」という太い紐状の構造を形成しています。真核細胞の分裂期には、
クロマチンは凝縮して光学顕微鏡で観察できる「染色体」になり、分裂前の染
色体は染色分体がセントロメアという領域でくっついた X 字状の構造となって
います。クロマチンの基本構造が、ヒストンというタンパク質と結合したヌク
レオソームです。4 種類のヒストン(H2A、H2B、H3、H4)が、2 個ずつ集
まった 8 量体のヒストンコアに DNA が巻き付いた構造をヌクレオソームと呼
びます。ヌクレオソームに 5 種類目のヒストン H1 が結合し、さらに大きな
螺旋を巻いて線維状の構造になります。そしてさらに他のタンパク質も加わっ
てコイル状の太いクロマチン線維を作ります。
ヒトの染色体には、22 種類の常染色体と X と Y と呼ばれる性染色体があり
ます。女性は常染色体を 2 セットと X 染色体を 2 本、男性は常染色体を 2 セッ
トと X 染色体と Y 染色体を 1 本ずつ持っており、母親と父親からそれぞれ 23
本の染色体を受け継いでいます。
ヒトの体細胞 1 個の DNA の長さは、約 2 m、ヒト 1 人の細胞数は 60 兆個
あるといわれていますから、体内にある DNA を全てつなぎあわせると 1,200
億 km にもなります。ちなみに、太陽と地球間の距離は約 1.5 億 km で、太陽
系の直径はその約 100 倍の約 150 億 km です。この約 10 倍の長さの DNA
が私たち一人一人の細胞の中に入っているのです。
DNA の二重らせん
ヒストンタンパク
2 nm
10 nm
300 nm
1,400 nm
セントロメア
ヌクレオソーム
染色体(分裂期)(光学顕微鏡で観察)
クロマチン繊維(電子顕微鏡で 観察)
30 nm
8 量体のヒストンコア
H2AH3H4 H2B
DNAと染色体
Pick Up from MBL
ヒト人工染色体(HAC)
MBL ネットワーク会社クロモリ
サーチがヒト培養細胞内で人工的
に構築した染色体で、ベクターと
して用いられます。
染色体には、
(1) 染色体 DNA の複製を開始さ
せる「複製起点」
(2) 細胞分裂期における染色体の
均等分配に重要である
「セントロメア」
(3) 染色体の末端構造の維持に必
要な「テロメア」
の三領域が必要とされます。
HAC ベクターは、これら必要最小
限の機能領域と希望の遺伝子で構
成されています。
目的の細胞株に取り込まれた HAC
ベクターは、宿主細胞とは独立に
存在し、宿主細胞染色体と同調し
て倍化・分裂し、次世代の細胞に
安定に受け継がれてゆきます。
HAC はトランスジェニックマウス
の作製にも利用されています。
セントロメア
テロメア
複製起点
遺伝子
染色体
人工染色体
◎ ヒト培養細胞内で人工的に構築した染色体
◎ 必要最小限の機能領域で構成
◎ 希望の遺伝子を挿入可能

54
細胞分裂期(M期)
間期
2)前中期(Prometaphase) 3)中期(Metaphase)1)前期(Prophase)
中心体は分離して紡錘体極となる。
中心体
核膜
核小体
細胞質細胞膜
5)終期(Telophase)
4)後期(Anaphase)
細胞骨格の一部をなしていた細胞質微小管が崩壊し、紡錘体の形成が始まる。培養細胞では細胞の形態が変化し、M 期の間、細胞は丸くなって接着性が減少する。また、間期に分散していたクロマチンは凝縮して染色体を形成し始める。G2 期に小さくなった核小体は前期~前中期にかけて消失する。
HEp-2 細胞のような接着性の細胞は、細胞質を延ばし、容器やスライドグラスの表面に張り付いている。G1 期は DNA 合成の準備時期で、ヒストンの合成も始まる。S期に入ると DNA は DNA ポリメラーゼによって複製される。G2 期で、分裂の前にDNA 複製が完全に行われたかチェックされる。間期にはクロマチンは解きほぐされて核全体に広がっている。
核膜は崩壊して断片となり、核質と細胞質は混ざり合う。セントロメアの両側に動原体と呼ばれるタンパク質複合体が形成され、そこに紡錘体微小管の一部(動原体微小管)が付着する。染色体は動原体微小管に引っ張られ活発に動く。
M 期の中では安定した時期であり、染色体は両極の中間で一つの平面状に並ぶ。
各染色体で対をなしていた動原体が分離し、娘染色体は紡錘体極に向かって引かれて行く。
娘染色体が両極に到達し、動原体微小管は消失する。染色体の周囲に再び核膜が形成され、細胞質と核質が分けられる。また、染色体は脱凝縮し、クロマチンは分散する。核小体は複数の点として観察されるようになり、G1 期には融合して1つとなる。
アクチンフィラメントを主成分とする収縮環の収縮によって細胞は 2 つに分かれる。分裂の最後には分裂溝の細胞質がくびれて中央体 (midbody) となり、2 つの娘細胞のつなぎとしてしばらく残っている。
6)細胞質分裂(Cytokinesis)細胞周期
M
G1
G0
G2
S
間期
分裂期
核膜核小体
細胞質細胞膜
紡錘体極
紡錘体極
動原体
動原体微小管
動原体微小管が短くなる。
中央体(微小管が重なりあった部分)
中心体を核として間期の微小管配列が形成される。
核膜の断片
分裂、増殖している細胞を観察すると、まず核内の
成分が凝縮して染色体となり、この染色体が核の両
側に移動して核が 2 つに分裂した後、細胞質が分裂
して 2 つの細胞となる。分裂したそれぞれの細胞は
しばらく時間をおき、再び分裂を繰り返す。この核
の分裂と細胞質の分裂の時期を細胞分裂期 (M 期 ;
Mitosis phase) といい、それ以外の時期を間期
(interphase) という。間期はさらに、G1 期 (Gap 1
phase)、S 期 (Synthesis phase)、G2 期 (Gap 2
phase) に分けられている。また G1 期の途中から休
止期 G0 期に入る細胞もある。
この図は細胞周期の模式図と、実際の細胞の写真(MBL「フルオロHEPANAテスト」を
抗セントロメア抗体で染色した写真)をあわせたものです。

55
細胞分裂期(M期)
間期
2)前中期(Prometaphase) 3)中期(Metaphase)1)前期(Prophase)
中心体は分離して紡錘体極となる。
中心体
核膜
核小体
細胞質細胞膜
5)終期(Telophase)
4)後期(Anaphase)
細胞骨格の一部をなしていた細胞質微小管が崩壊し、紡錘体の形成が始まる。培養細胞では細胞の形態が変化し、M 期の間、細胞は丸くなって接着性が減少する。また、間期に分散していたクロマチンは凝縮して染色体を形成し始める。G2 期に小さくなった核小体は前期~前中期にかけて消失する。
HEp-2 細胞のような接着性の細胞は、細胞質を延ばし、容器やスライドグラスの表面に張り付いている。G1 期は DNA 合成の準備時期で、ヒストンの合成も始まる。S期に入ると DNA は DNA ポリメラーゼによって複製される。G2 期で、分裂の前にDNA 複製が完全に行われたかチェックされる。間期にはクロマチンは解きほぐされて核全体に広がっている。
核膜は崩壊して断片となり、核質と細胞質は混ざり合う。セントロメアの両側に動原体と呼ばれるタンパク質複合体が形成され、そこに紡錘体微小管の一部(動原体微小管)が付着する。染色体は動原体微小管に引っ張られ活発に動く。
M 期の中では安定した時期であり、染色体は両極の中間で一つの平面状に並ぶ。
各染色体で対をなしていた動原体が分離し、娘染色体は紡錘体極に向かって引かれて行く。
娘染色体が両極に到達し、動原体微小管は消失する。染色体の周囲に再び核膜が形成され、細胞質と核質が分けられる。また、染色体は脱凝縮し、クロマチンは分散する。核小体は複数の点として観察されるようになり、G1 期には融合して1つとなる。
アクチンフィラメントを主成分とする収縮環の収縮によって細胞は 2 つに分かれる。分裂の最後には分裂溝の細胞質がくびれて中央体 (midbody) となり、2 つの娘細胞のつなぎとしてしばらく残っている。
6)細胞質分裂(Cytokinesis)細胞周期
M
G1
G0
G2
S
間期
分裂期
核膜核小体
細胞質細胞膜
紡錘体極
紡錘体極
動原体
動原体微小管
動原体微小管が短くなる。
中央体(微小管が重なりあった部分)
中心体を核として間期の微小管配列が形成される。
核膜の断片
分裂、増殖している細胞を観察すると、まず核内の
成分が凝縮して染色体となり、この染色体が核の両
側に移動して核が 2 つに分裂した後、細胞質が分裂
して 2 つの細胞となる。分裂したそれぞれの細胞は
しばらく時間をおき、再び分裂を繰り返す。この核
の分裂と細胞質の分裂の時期を細胞分裂期 (M 期 ;
Mitosis phase) といい、それ以外の時期を間期
(interphase) という。間期はさらに、G1 期 (Gap 1
phase)、S 期 (Synthesis phase)、G2 期 (Gap 2
phase) に分けられている。また G1 期の途中から休
止期 G0 期に入る細胞もある。
この図は細胞周期の模式図と、実際の細胞の写真(MBL「フルオロHEPANAテスト」を
抗セントロメア抗体で染色した写真)をあわせたものです。

56
3-4.設計図は正確にコピーされる「DNA 複製の仕組み」DNA は、A が塩基対を形成する相手は必ず T、G が塩基対を形成する相手
は必ず C、と決まっています。そのため、DNA が複製される(2 倍に増える)
際には片方の鎖を鋳型として、裏返しの塩基配列をもつ、もう一方の鎖を合成
します。こうして、設計図は次々と正確にコピーされていきます。
DNA の複製には、DNA 鎖を合成する DNA ポリメラーゼをはじめとして、
多くの酵素が関与しています。細菌からヒトまで、複製の機構は基本的には同
じですが、真核生物は原核生物よりも複雑な仕組みをもちます。
複製は複製起点から始まります。複製起点からヘリカーゼという酵素が
DNA 二本鎖を一本鎖にほどきます。そこにプライマー(DNA ポリメラーゼ
が合成を開始する際に必要な短い核酸の断片)が結合し、一本鎖の DNA を鋳
型にして DNA ポリメラーゼが新しい DNA 鎖を合成します。DNA には方向性
があり、一方の末端を 5' 末端、もう一方を 3' 末端と呼びます(参照:2-5.
DNA の美しい構造「二重らせん」p.50)。DNA ポリメラーゼは 5' から 3' の
方向に DNA を合成するため、3' から 5' 方向の DNA(リーディング鎖)を鋳
型にした場合の複製は 5' から 3' の方向に連続的に進みます。一方、5' から
3' 方向の DNA(ラギング鎖)を鋳型にした場合は 5' から 3' の方向に 100
- 200 のヌクレオチド分の DNA(岡崎フラグメント、参照:研究者コラム「岡
崎フラグメントの発見」)の複製を繰り返し、酵素の働きによってプライマー
が取り除かれ、それが DNA 鎖に置き換えられ、DNA リガーゼによって結合さ
れます(図.DNA の複製)。
2 本の親 DNA 鎖がほどけて娘鎖が合成される部分は、3 本の二本鎖 DNA
が観察されるので、複製フォークと呼びます。複製フォークが進行するさらに
先の方ではトポイソメラーゼが DNA 鎖を切断してねじれを解消したあと、再
結合しています。
研究者コラム
岡崎令治
「岡崎フラグメントの発見」
DNA は、互いに逆向きのポリヌク
レオチド鎖が対合した二重らせん構
造なので、DNA 複製の際は、全体
として、一方の鎖は 5' → 3' 方向
に、他方の鎖は 3' → 5' 方向に伸長
します。しかし、DNA ポリメラー
ゼはポリヌクレオチド鎖を 5' → 3'
方向にしか伸長しません。3' → 5'
方向の伸長はどうなっているのか?
岡崎らはこの矛盾を解消しうる仕組
みとして、3' → 5' 方向の伸長では、
5' → 3' 方向への短い鎖を合成し、
それを連結することで、全体として
は 3' → 5' 方向に伸長するという不
連続複製機構を考え、放射性同位体
で標識したヌクレオシドを用いた実
験で短い DNA 鎖が合成されること
を証明しました。さらに、1967 年
に DNA を連結する DNA リガーゼ
が発見され、岡崎の説は世界に認め
られました。ラギング鎖の DNA 断
片は「岡崎フラグメント」と呼ばれ
るようになりました。
1975 年、岡崎は中学生のときに広
島で被爆したことが原因の病で 44
歳の若さでこの世を去ります。その
後、妻の岡崎恒子ら(現・藤田保健
衛生大学名誉教授)が岡崎フラグメ
ントの合成開始から連結に至る全過
程の解明に成功しています。
3'
5'
3'
3'
3'
5'
5'
5'
5'
複製の進行方向
ラギング鎖の鋳型
リーディング鎖の鋳型
プライマー
プライマー
複製起点
親 DNAらせん
DNA ポリメラーゼ
DNA ポリメラーゼ
岡崎フラグメント
5'ヘリカーゼ
プライマーゼ
次に合成される岡崎フラグメント
新しく合成されたDNA
DNAの複製

いきものコラム 「テロメアが短かったクローン羊ドリー」
1996 年、イギリスで核移植により誕生したクロー
ン羊のドリーは、6 歳で肺の疾患のための安楽死に
よって、早すぎる死を迎えました(通常、羊の寿命
は 11 〜 12 歳)。ドリーは 6 歳のメスのヒツジの乳
腺細胞の核を、別の種類のヒツジの核を取り除いた
未受精卵に移植して、誕生しました。ドリーのテロ
メアは生まれつき短かったと報告されています。
しかし、ドリーの寿命に本当にテ
ロメアの短さが関係していたの
か、その因果関係ははっきりして
いません。 テロメラーゼによる DNA の伸長
DNA は複製を繰り返すと短くなる
57
3-5.老化を支配する時計「テロメア」
ラギング鎖の複製は、RNA プライマーが岡崎フラグメントに付加されるこ
とによっておこるので、3' 末端で RNA プライマーが取り除かれると、その
DNA 領域は複製されないままになります。こうして DNA 複製によって形成さ
れた新しい染色体は、それぞれ一本鎖の短い DNA を持つことになります。こ
の状況になると一本鎖の部分は、二本鎖の一部と一緒に切断され、複製のた
びに染色体が短くなります(図.DNA は複製を繰り返すと短くなる)。多くの
真核生物には、この染色体の末端に DNA の反復配列と末端安定性を維持する
タンパク質からなるテロメアという構造があります。ヒトのテロメアの場合、
TTAGGG という配列が 2,000 回以上繰り返されています。
ヒトの染色体は細胞分裂繰り返すたびに、テロメアの DNA 塩基対を失い、
約 60 〜 70 回細胞分裂を繰り返すとテロメアが短くなった細胞は分裂不能と
なって細胞の老化やアポトーシスが誘発されます。
しかし、骨髄幹細胞や生殖細胞のように絶えず分裂している細胞は、テロメ
アの短縮を「テロメラーゼ」という酵素の発現により克服しています(図.テ
ロメラーゼによる DNA の伸長)。テロメラーゼはテロメアを伸長させる鋳型と
なる RNA(TERC)と逆転写酵素活性を持つ触媒サブユニット(TERT)、その
他の制御サブユニットから構成されています。また、テロメラーゼは、90%
以上のヒトのがんで発現していて、ほとんどの正常細胞には発現しないことか
ら、テロメラーゼは抗がん剤の標的となっています。
Pick Up from MBL
テロメア伸長に関わるTERT 関連製品
近年、テロメラーゼ、とりわけ
その酵素複合体の律速因子であ
る TERT が創薬のターゲットとし
て注目されています。TERT はテ
ロメア維持機構以外の機能を介し
て、幹細胞の分化能維持に関わる
可能性が示唆されており、がん幹
細胞における再生能の維持に関す
る研究が注目されています。
弊社の TERT に対するモノクロー
ナル抗体は論文でも使用されてお
ります。また、ELISA で細胞可溶
化物や血清中の TERT を定量でき
る Assembly Human TERT Kit も
販売しています。
テロメラーゼがテロメラーゼRNAの相補的なテロメアの3’末端に結合する
塩基がRNAをテンプレートにして追加される
テロメラーゼが移動する
第2ステップが繰り返される
DNAポリメラーゼがラギング鎖を補完する
3’ 5’
3’
T T
G5’CT ACT GG CTCT AA GGG CT CT G
AC C C AAA
AGGG CT CT
C C3’ 5’
3’
T T
G5’CT ACT GG CTCT AA GGG CT CT G
AC C C AA A
AGGG CT CT
C C C A A UC C C A A UC
AGG
CTCT
GAGG CT CT
T T
G5’CT ACT GG CTCT AA GGG CT CT G
AC C C AA A
AGGG CT CT
C C C A A UC C C A A UC3’ 5’
3’
…
GAGG CT CT
T T
G5’CT ACT GG CTCT AA GGG CT CT G
AC C C AA A
AGGG CT CT
C C C A A UC C C A A UC3’ 5’
3’
GAGG CT CT
T T
G5’CT ACT GG CTCT AA GGG CT CT G
AC C C AA A
AGGG CT CT
C C T C C C A A T3’ 5’
GAGG CT CTGAGG CT CT
C A A T C C
…C A A
DNAポリメラーゼ
テロメラーゼ
3’
RNAプライマーラギング鎖
3’
3’
短縮
3’
RNAプライマーがはずれ、ラギンク鎖の鋳型DNAが切断される
3’
3’
T
G
5’
5’
CT ACT GG CTCT AA GGG CT CT G
AC C C AA A C A A T C C C A A T C
テロメア
TERT
TERC
TEP1 Dyskerin

様々な DNA 損傷と DNA 修復
58
3-6.傷ついた DNA をなおす仕組み「DNA 修復」遺伝情報の担い手である DNA は、安定に保持されなければいけません。し
かし、細胞内の DNA は外界からの紫外線や放射線、代謝の過程で生じる活性
酸素などによって、つねに化学的・物理的に傷ついています。DNA の損傷は、
細胞あるいは生物個体に様々な影響をおよぼします。例えば、多くの損傷は
DNA 複製時に DNA ポリメラーゼの進行を妨げ、DNA 複製、ひいては細胞増
殖を停止させ、その結果、細胞は死に至ります。さらに、DNA 損傷は突然変
異の原因となります。突然変異は個体間の多様性や生物進化につながる一方で、
細胞のがん化や老化を促進することもあります。
生物はこれらの有害な DNA 損傷を速やかに取り除くために、種々の DNA
修復機構を進化の過程で獲得し、遺伝情報である DNA の維持に努めています。
修復機構の一つに光回復酵素による修復があります。光回復酵素は紫外線に
よって損傷した塩基に直接結合し、可視光のエネルギーを利用して正常な塩基
に復帰させます。ヒトをはじめとするほ乳類では光回復酵素に類似のタンパク
質は存在しますが、DNA 修復活性は持たず、生物時計を司っていることが明
らかになっています(参照:6-7. 体内時計は遺伝子レベルで存在する「概日
リズム」p.76)。DNA 二本鎖の片方に生じた損傷を修復する機構として代表
的な機構は除去修復です。除去修復では損傷部位をいったん取り除いた後、あ
らためて正常な DNA 鎖を作り直します。X 線や γ 線などによって DNA 二本
鎖が切断された場合は切断端が再結合して、もとの DNA が復元します。これ
を組換え修復とよびます(図.様々な DNA 損傷と DNA 修復)。
DNA 複製時に DNA ポリメラーゼが引き起こすエラーもありますが、その頻
度は約1億回に1回と低い頻度です。DNA ポリメラーゼ自身に誤ったヌクレ
オチドを取り込んでもそれを検知し取り除く校正機能があるためです。
Pick Up from MBL
遺伝子・DNA の変異・多型検出キット
Surveyor® Mutation Detection Kits
MBL ネ ッ ト ワ ー ク 会 社 IDT-MBL
KK では、遺伝子や DNA の変異や
多型を検出するキットを販売してい
ます。セロリ由来の、変異箇所を特
異的に切断するヌクレアーゼ(CEL
ファミリータンパク質)、Surveyor
Nuclease の特徴的な活性を利用す
るキットです。代表的な変異解析法
の一つとして、多くの研究者に広く
使用されています。
Surveyor Nuclease は 2 本 鎖 DNA
内のミスマッチ箇所特異的なエンド
ヌクレアーゼです。ミスマッチ箇所
の 3' 側を切断します。
5’
3’
5’
3’
Surveyor Nuclease
Resulting fragments
5’
3’
5’
3’
5’
3’
5’
3’
Location of mismatch
5’
3’
5’
3’
DNAポリメラーゼの進行阻害
細胞増殖の停止
細胞死
個体の老化など
突然変異の蓄積
正常な細胞
DNA複製時におけるエラー DNA損傷
DNAポリメラーゼによる損傷の乗り越え
低頻度で誤ったヌクレオチドの取込みが起こる細胞のがん化、老化など
突然変異
損傷の修復(光回復酵素など)
除去修復
組換え修復などミスマッチ修復
DNAポリメラーゼによる校正