Embed Size (px)
DESCRIPTION
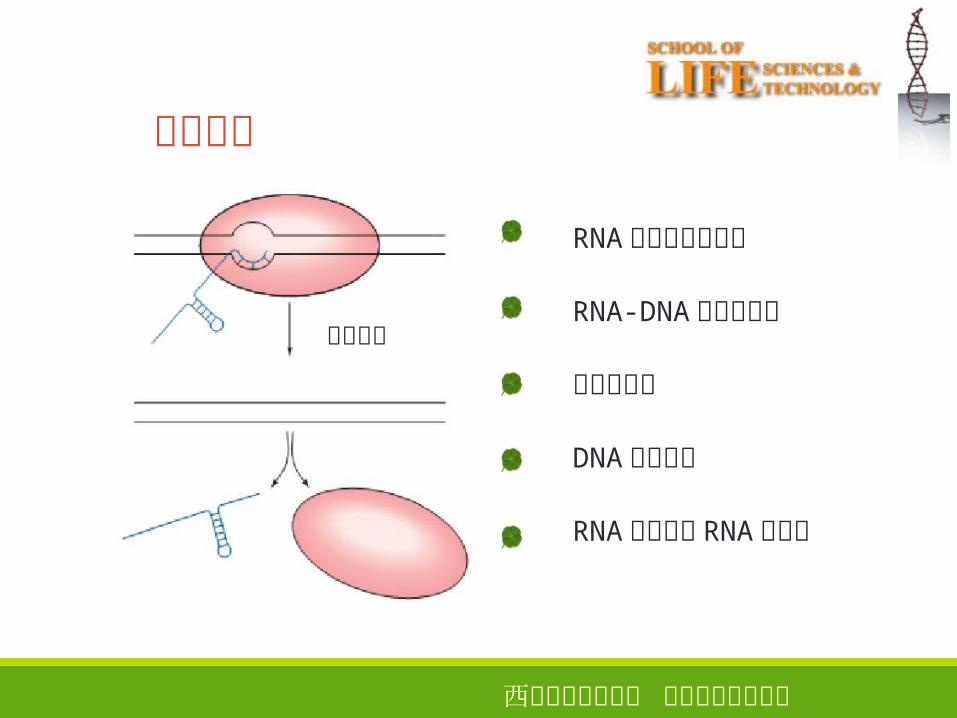
第三章 生物信息的传递(上) —— 从 DNA 到 RNA. 基本概念 RNA 的结构、分类和功能 转录的基本过程 RNA 聚合酶、启动子 原核生物与真核生物 mRNA 的特征比较 转录后加工 RNA 合成与 DNA 合成异同点. Contents. 生物体以 DNA 为模板合成 RNA 的过程 。. 转录. RNA. DNA. 一、 基本概念. 转录 (transcription) :. 原料 : NTP (ATP, UTP, GTP, CTP) 模板 :DNA 酶 : RNA 聚合酶 其他蛋白质因子. 参与转录的物质. - PowerPoint PPT Presentation
Citation preview
西安电子科技大学 生命科学技术学院
第三章 生物信息的传递(上) ——从 DNA到 RNA

西安电子科技大学 生命科学技术学院
• 基本概念• RNA 的结构、分类和功能• 转录的基本过程• RNA 聚合酶、启动子• 原核生物与真核生物 mRNA 的特征比较• 转录后加工• RNA 合成与 DNA 合成异同点
Contents
西安电子科技大学 生命科学技术学院
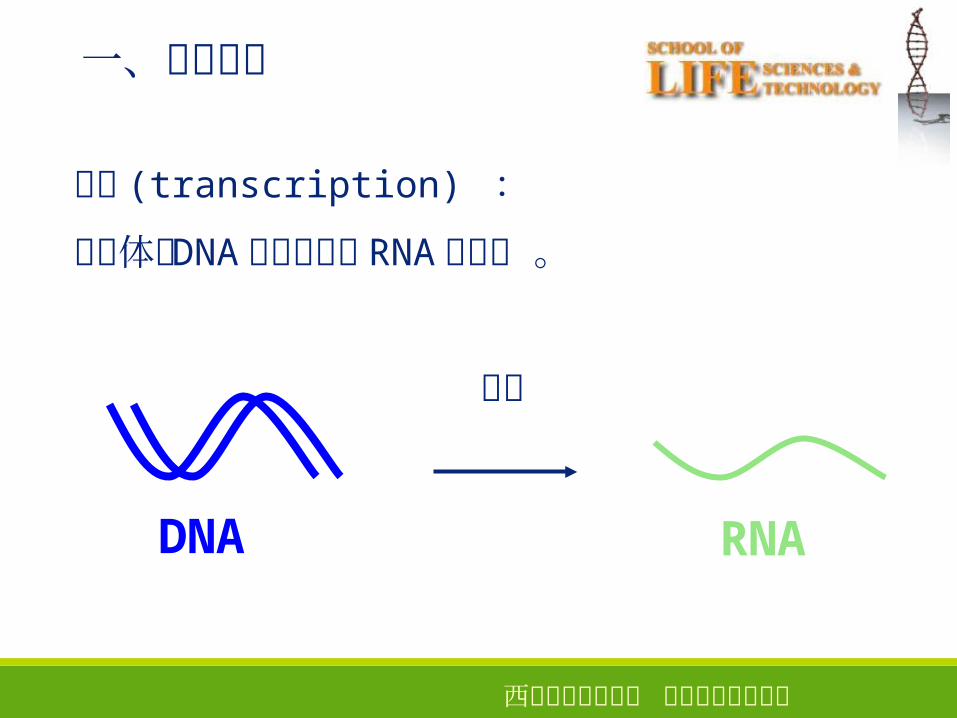
一、基本概念
生物体以 DNA 为模板合成 RNA 的过程 。
转录
RNADNA
转录 (transcription) :

西安电子科技大学 生命科学技术学院
参与转录的物质
原料 : NTP (ATP, UTP, GTP, CTP)
模板 :DNA
酶 : RNA 聚合酶其他蛋白质因子
RNA 合成方向: 5' 3'
西安电子科技大学 生命科学技术学院
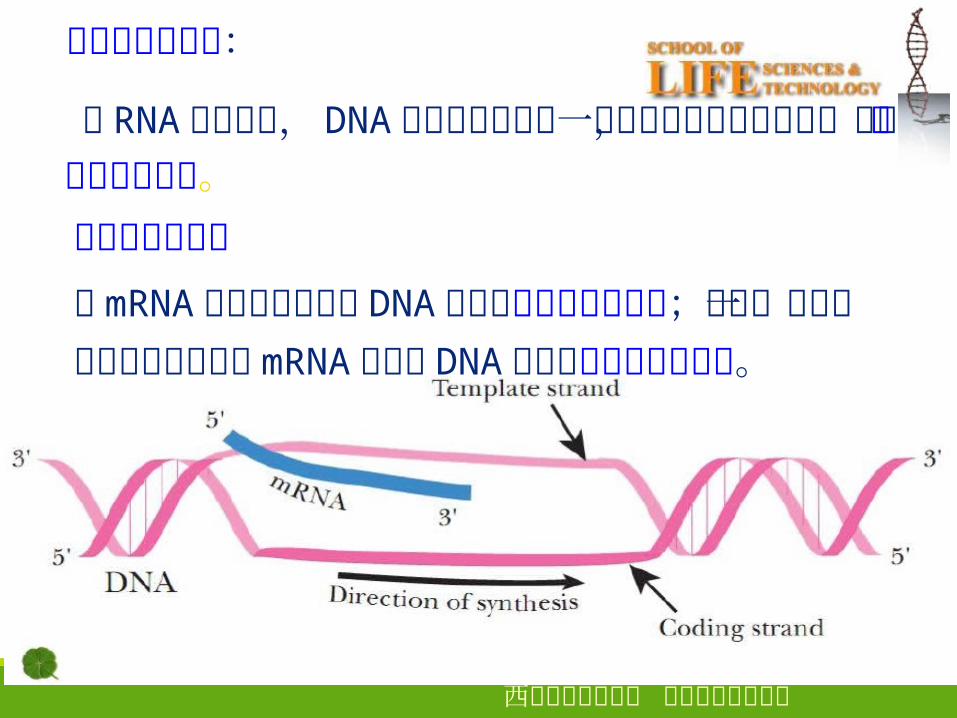
转录的不对称性:
在 RNA 的合成中, DNA 的二条链中仅有一条链可作为转录的模板,称为转录的不对称性。编码链与模板链与mRNA 序列相同的那条 DNA 链称为编码链或有义链;将另一条根据碱基互补原则指导mRNA 合成的 DNA 链称为模板链或反义链。

西安电子科技大学 生命科学技术学院
• 基本概念• RNA 的结构、分类和功能• 转录的基本过程• RNA 聚合酶、启动子• 原核生物与真核生物 mRNA 的特征比较• 转录后加工• RNA 合成与 DNA 合成异同点
Contents
西安电子科技大学 生命科学技术学院
RNA的结构特点
RNA含有核糖和嘧啶,通常是单链线性分子
RNA链自身折叠形成局部双螺旋发夹结构、凸结构、环结构A-U,C-G,� �G-U双螺旋 RNA小沟宽而浅,没有序列特异性信息;大沟狭且深,不适合与蛋白质作用
RNA可折叠形成复杂的三级结构
4
西安电子科技大学 生命科学技术学院
5
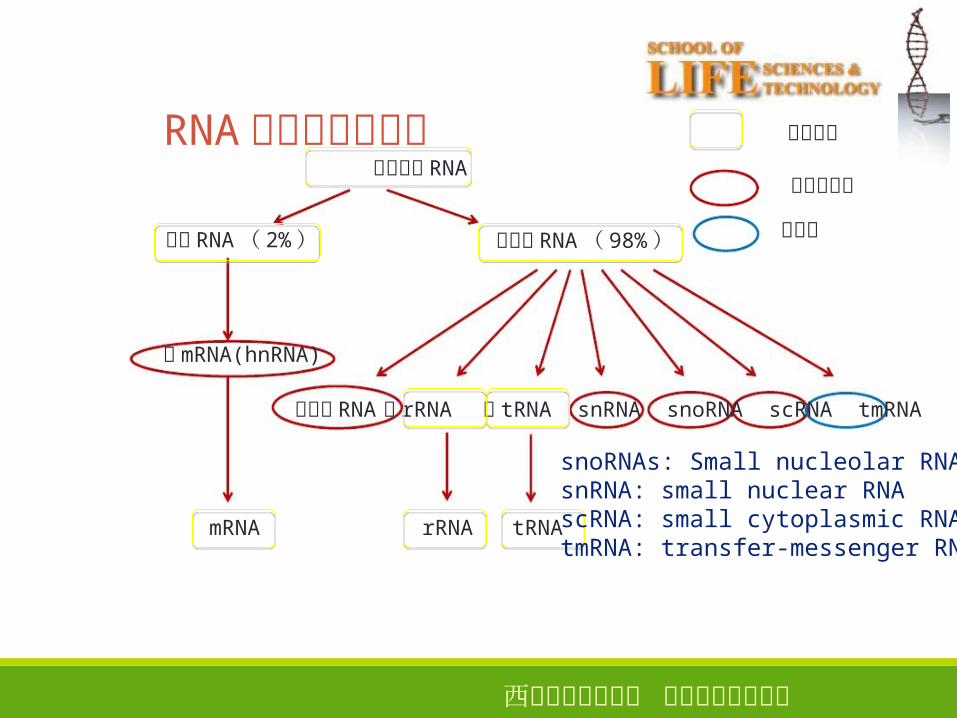
RNA在细胞中的分布细胞内总 RNA
编码 RNA( 2%)
前mRNA(hnRNA)
mRNA
非编码 RNA( 98%)
非编码 RNA前 rRNA 前 tRNA snRNA snoRNA scRNA tmRNA
rRNA tRNA
仅细菌
所有生物
仅真核生物
snoRNAs: Small nucleolar RNAssnRNA: small nuclear RNAscRNA: small cytoplasmic RNAtmRNA: transfer-messenger RNA
西安电子科技大学 生命科学技术学院
RNA的功能
信息分子贮藏和转移遗传信息
功能分子作为蛋白质合成的主要参与者
部分 RNA作为核酶在细胞中催化反应
参与基因的表达调控
某些病毒中,是遗传物质

西安电子科技大学 生命科学技术学院
• 基本概念• RNA 的结构、分类和功能• 转录的基本过程• RNA 聚合酶、启动子• 原核生物与真核生物 mRNA 的特征比较• 转录后加工• RNA 合成与 DNA 合成异同点
Contents
西安电子科技大学 生命科学技术学院
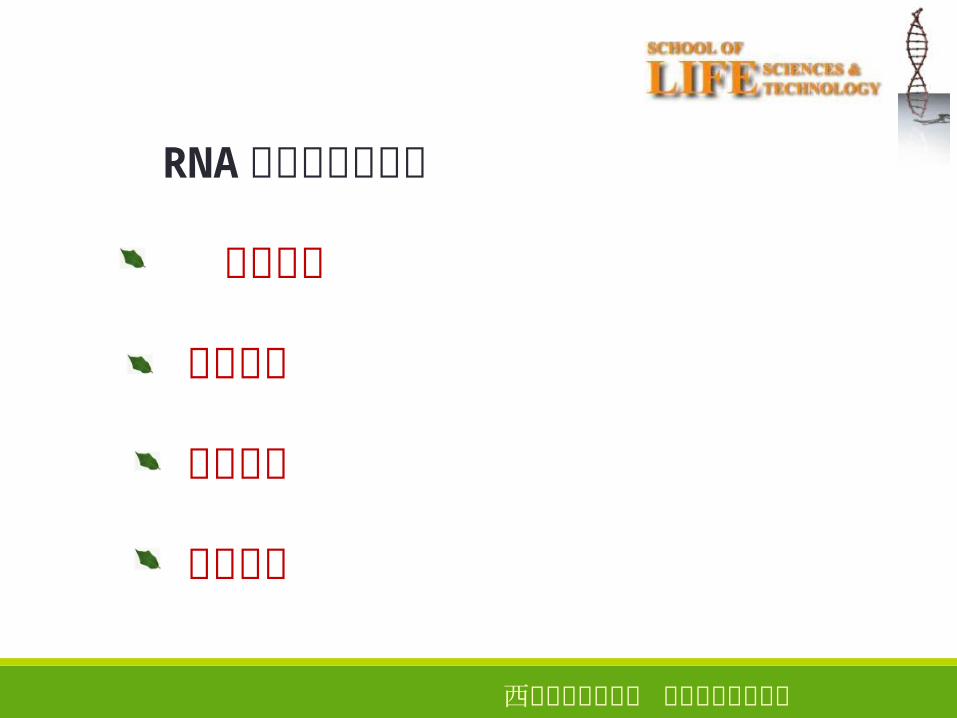
RNA转录的基本过程
模板识别
转录起始
转录延伸
转录终止
7
西安电子科技大学 生命科学技术学院
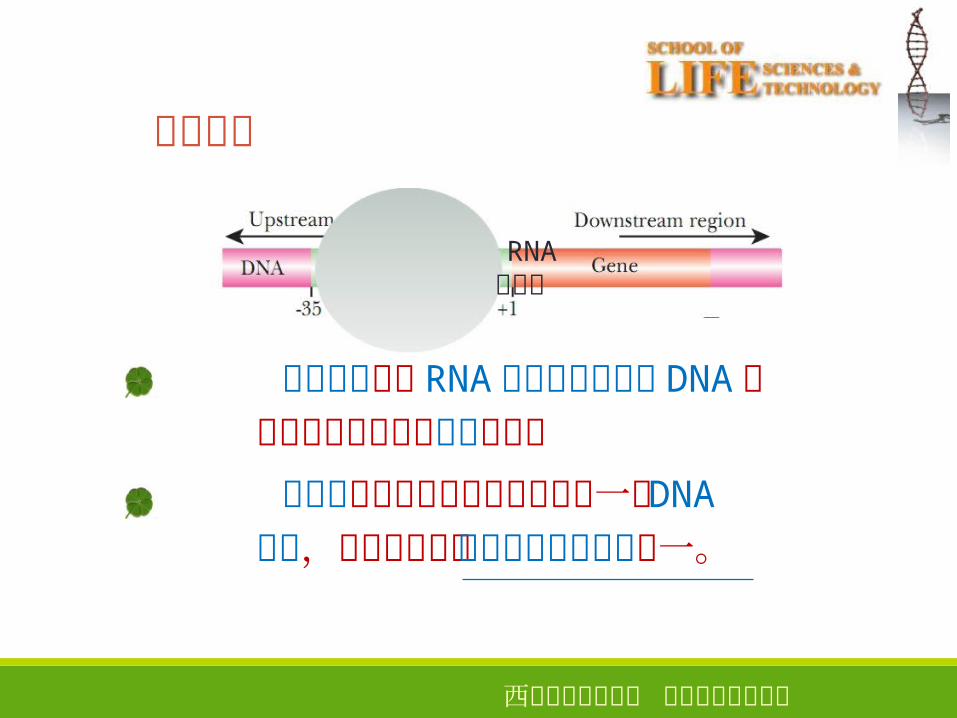
模板识别
8
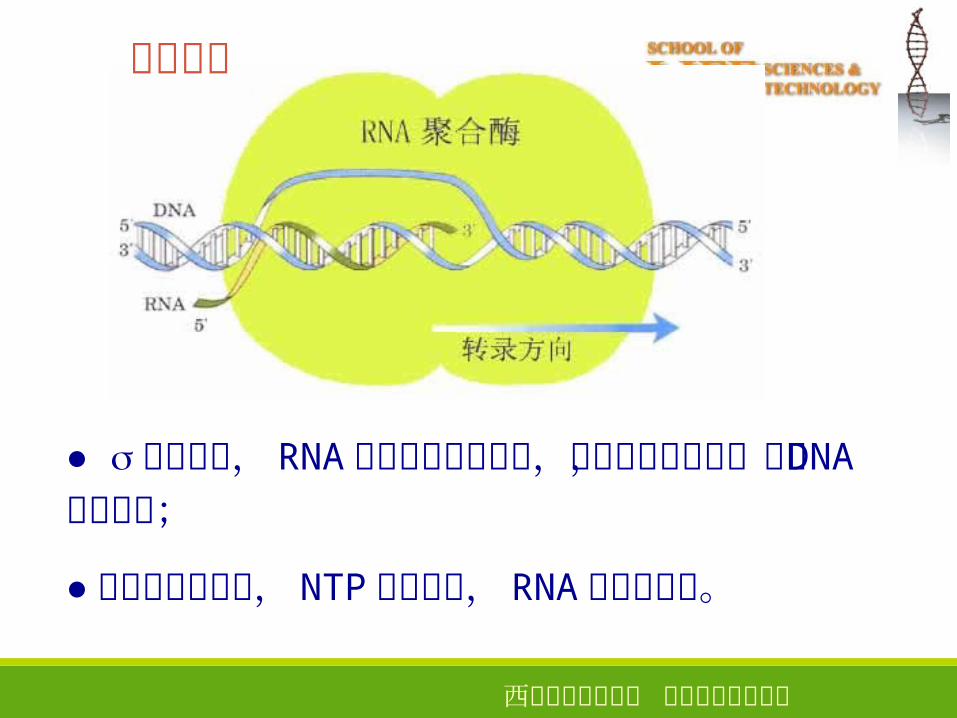
RNA聚合酶
模板识别是指 RNA聚合酶与启动子 DNA双链相互作用并与之结合的过程
启动子是基因转录起始所必需的一段 DNA序列,是基因表达的上游顺式作用元件之一。
西安电子科技大学 生命科学技术学院
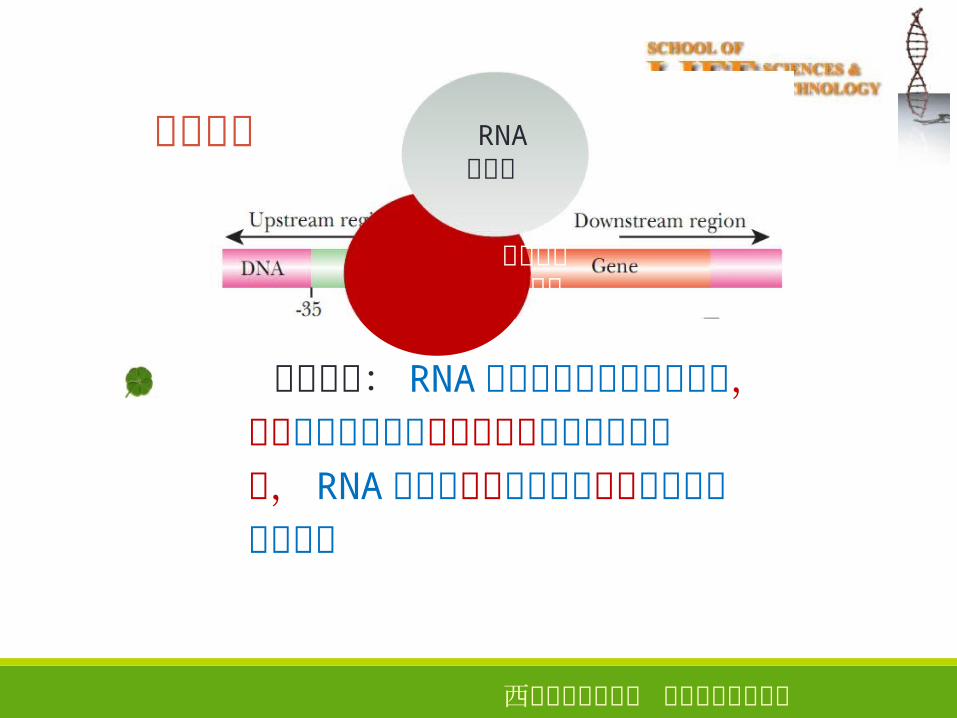
模板识别
9
转录调控因子
真核生物: RNA聚合酶不能识别启动子区,需要转录调控因子按特定顺序结合于启动子上, RNA聚合酶才能与之结合形成转录起始前复合物
RNA聚合酶
西安电子科技大学 生命科学技术学院
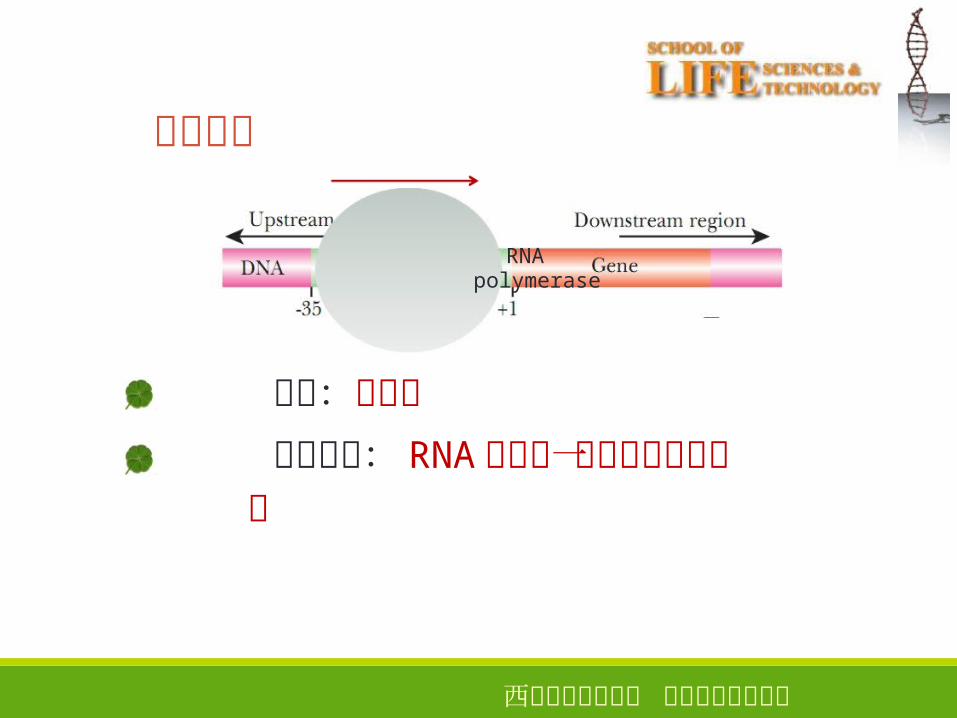
转录起始
10
RNApolymerase
引物:不需要
转录起始: RNA链上第一个核苷酸键的产生
西安电子科技大学 生命科学技术学院
1.� 聚合酶和启动子结合成封闭 复合物
2.� 封闭复合物变成开放复合物
3.� 开放复合物结合新生 RNA , 形成三元复合物
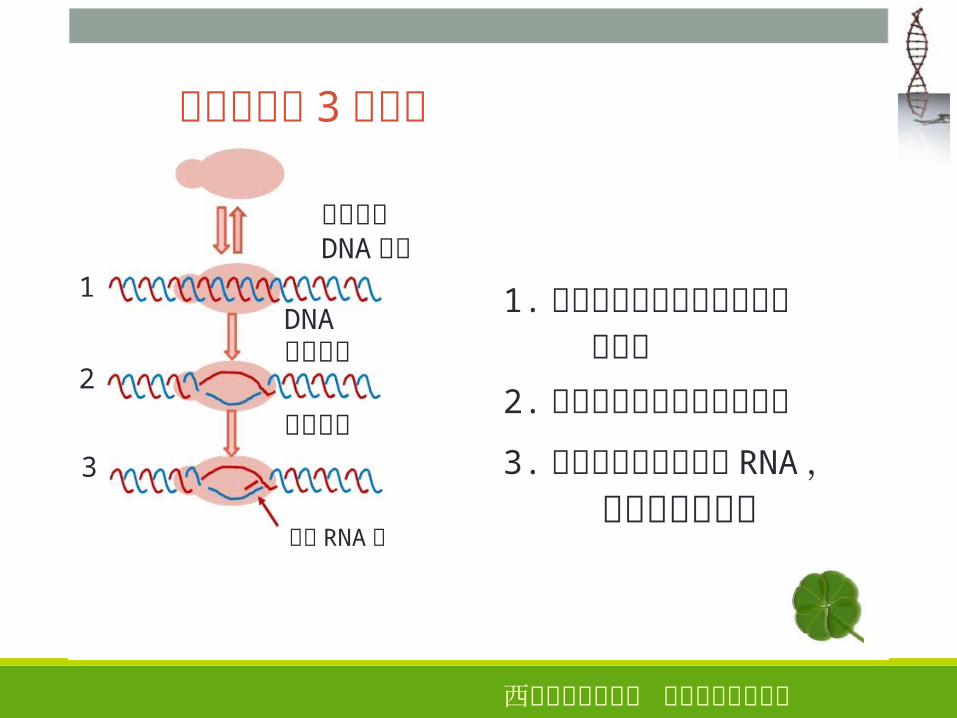
转录起始的 3个阶段
聚合酶与DNA结合
1
2
3
DNA开始解链
转录起始
新生 RNA链
西安电子科技大学 生命科学技术学院
转录起始
12
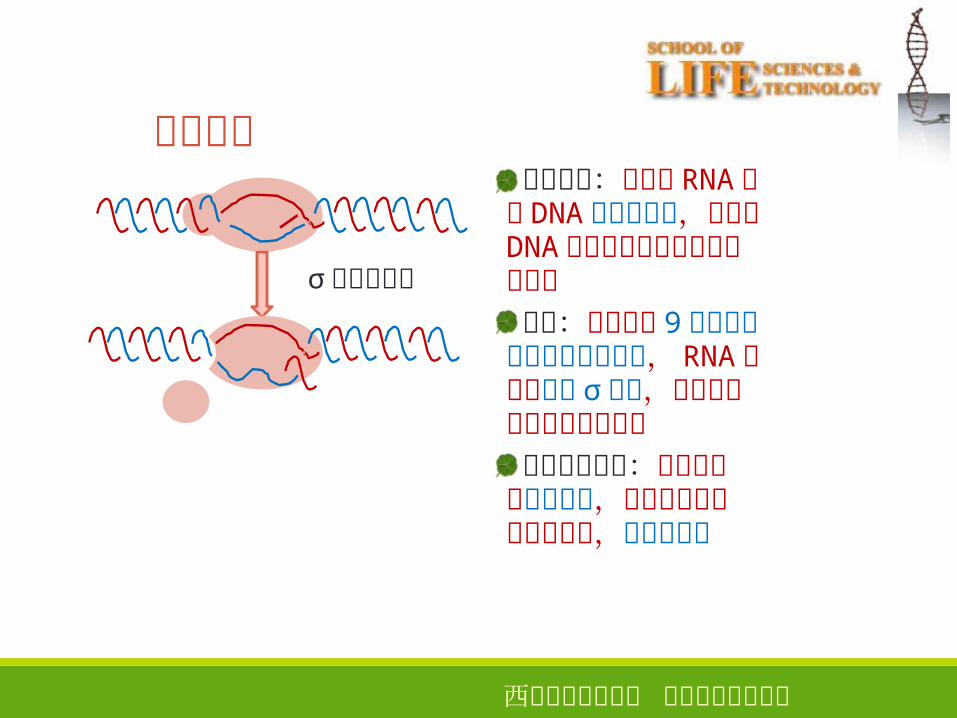
σ因子被释放
重新开始:新生的 RNA链与 DNA结合不牢固,容易从DNA链上掉下来导致转录重新开始成功:成功合成 9个以上核苷酸离开启动子区, RNA聚合酶释放 σ因子,转录就进入正常的延伸阶段启动子的强弱:通过启动子时间越短,基因转录起始的频率越高,启动子越强
西安电子科技大学 生命科学技术学院
转录起始中的概念
聚合酶全酶负责启动子的选择和转录的开始
聚合酶全酶中的核心酶负责 RNA的延伸
真核生物RNA聚合酶
转录因子 TF (七种)
西安电子科技大学 生命科学技术学院
转录延伸 14
● 亚基脱落, RNA 聚合酶核心酶变构,与模板结合松弛,沿着 DNA 模板前移;
● 在核心酶作用下, NTP 不断聚合, RNA 链不断延长。
西安电子科技大学 生命科学技术学院
转录终止
17
转录终止
RNA聚合酶不再聚合
RNA-DNA杂合物分离
转录泡瓦解
DNA恢复双链
RNA聚合酶和 RNA链释放

西安电子科技大学 生命科学技术学院
终止子( terminator)的分类
根据体外 RNA聚合酶是否需要辅助因子参与才能终止 RNA的延伸,大肠杆菌的终止子分为:不依赖于 ρ因子的终止
依赖于 ρ 因子的终止 ( Rho-dependent terminator )
18

西安电子科技大学 生命科学技术学院
不依赖因子的终止
1 、终止位点上游一般存在一个富含 GC 碱基的二重对称区,转录产生的 RNA 形成发夹结构;
2 、在终止位点前面有一段由 4-8个 A 组成的序列,转录产生的 RNA的 3’ 端为寡聚 U
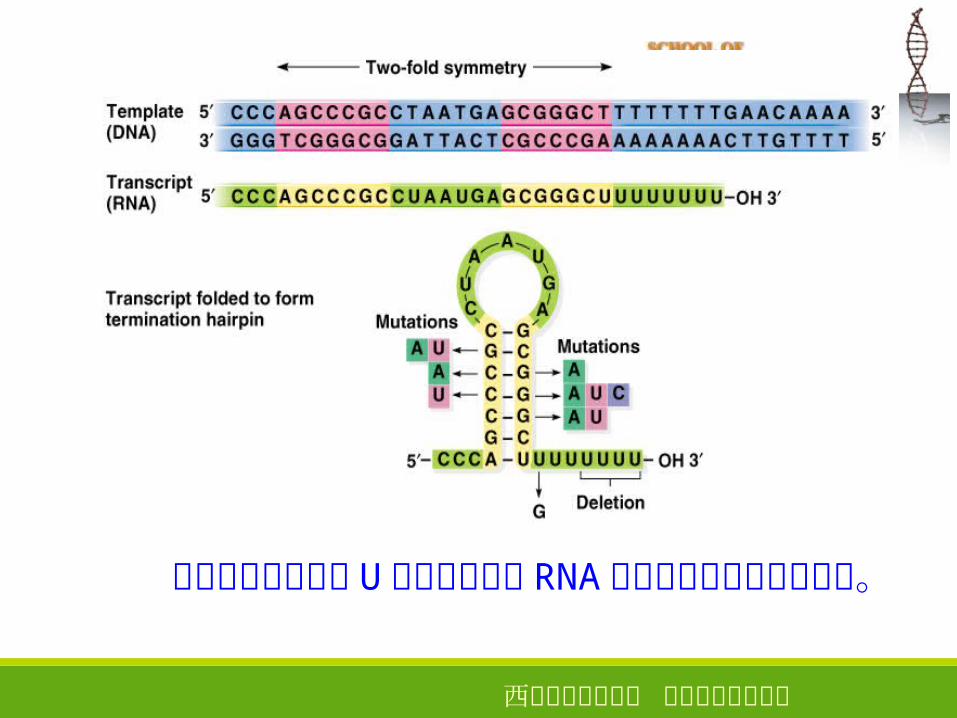
西安电子科技大学 生命科学技术学院
发夹式结构和寡聚 U 的共同作用使 RNA从三元复合物中解离出来。
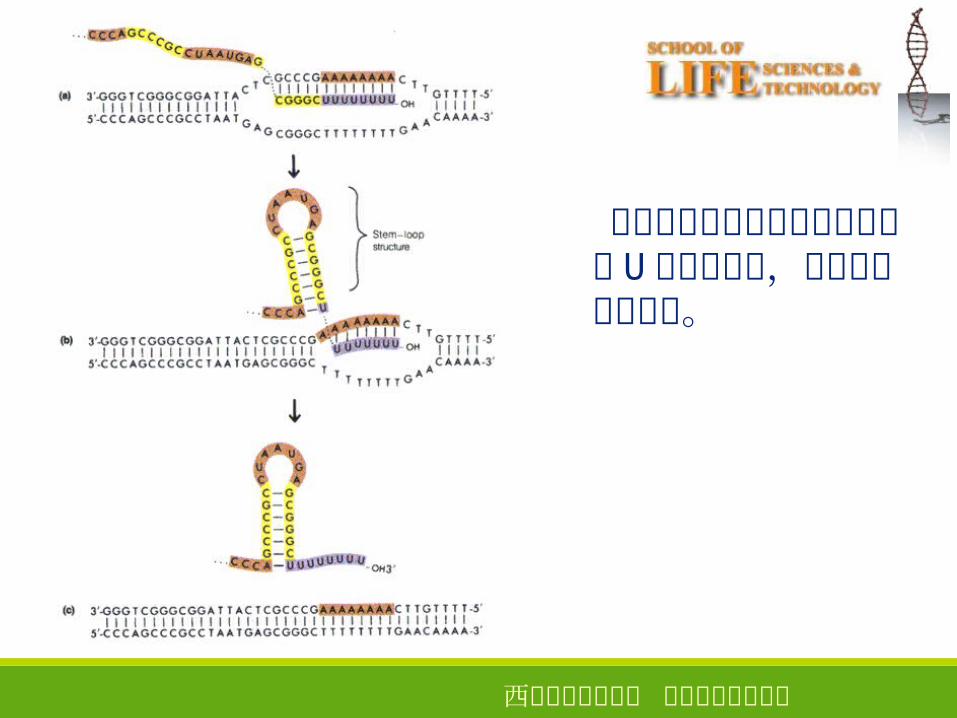
西安电子科技大学 生命科学技术学院
终止效率与二重对称序列和寡聚 U 的长短有关,长度增加效率提高。
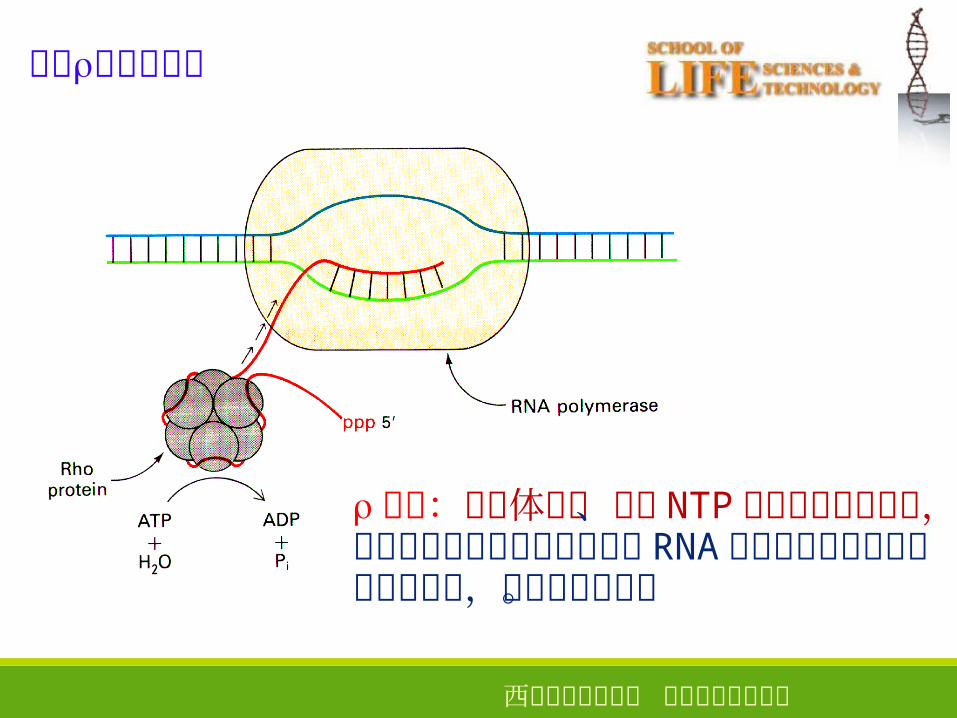
西安电子科技大学 生命科学技术学院
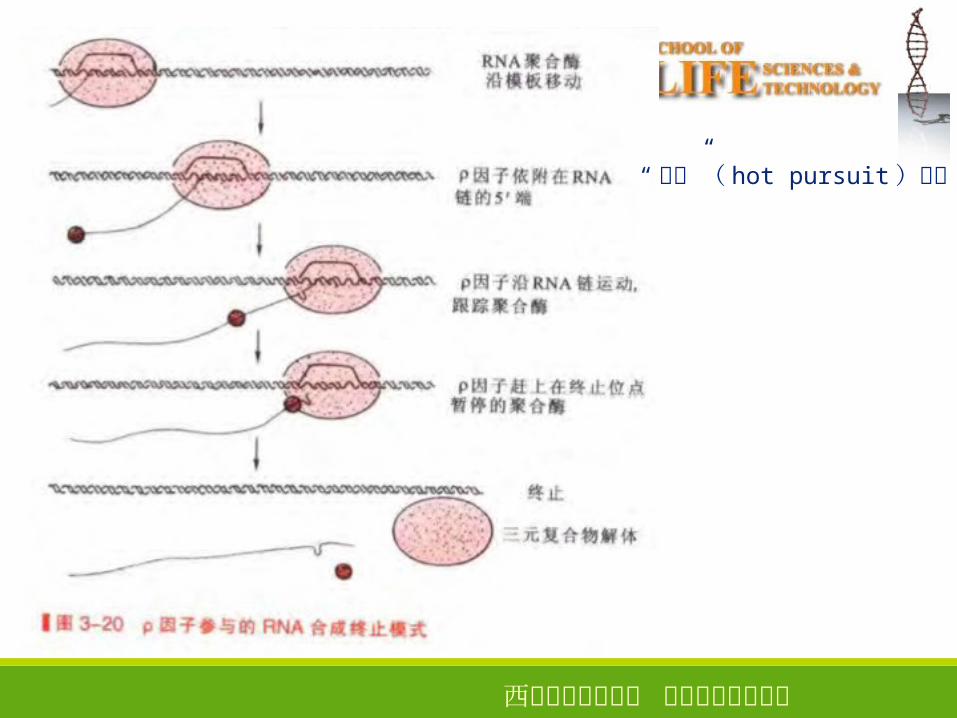
依赖因子的终止
因子:六聚体蛋白、具有 NTP酶和解螺旋酶活性,水解各种核甘三磷酸促使新生 RNA 链从三元转录复合物中解离出来,从而终止转录。
西安电子科技大学 生命科学技术学院
“ 穷追”( hot pursuit )模型

西安电子科技大学 生命科学技术学院
转录的基本过程
西安电子科技大学 生命科学技术学院
抗终止
破坏终止位点 RNA的茎环结构转录和翻译偶联
当氨基酸浓度较低时,核糖体滞留在串联密码子上,导致mRNA不能形成茎环结构
依赖于蛋白质因子的转录抗终止λ-噬菌体中的 N蛋白具有转录抗终止作用
21
西安电子科技大学 生命科学技术学院
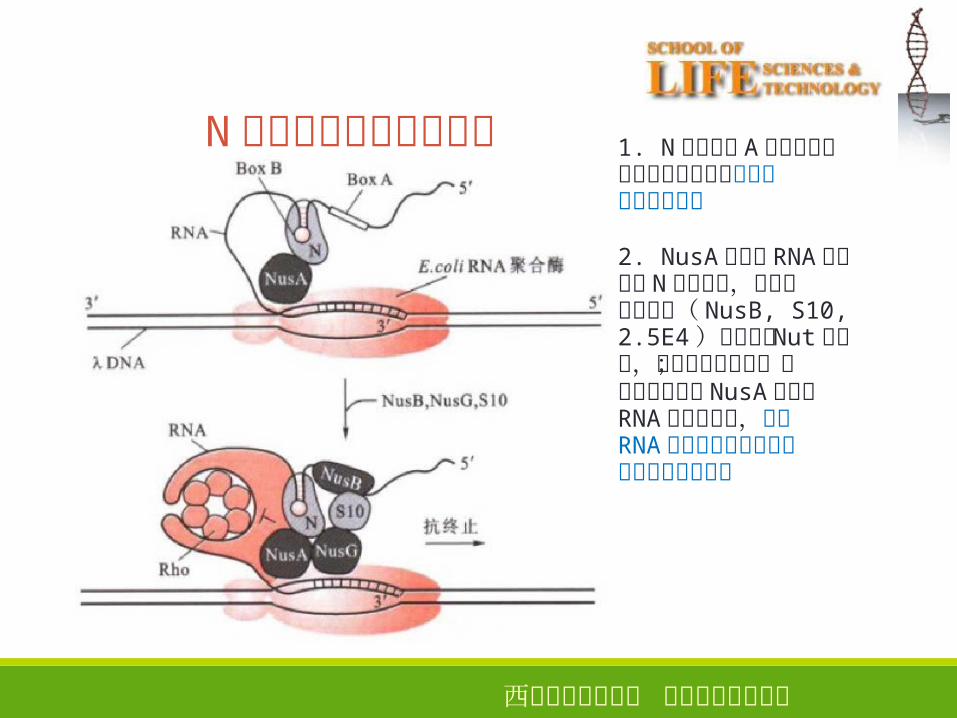
N蛋白引起的转录抗终止
22
1. N蛋白识别 A区和对称二重序列转录形成的茎环结构并与之结合
2. NusA蛋白与 RNA聚合酶和 N蛋白结合,当其它三种蛋白( NusB, S10,2.5E4)都结合到 Nut位点时,形成蛋白复合物;该复合物通过与 NusA蛋白和RNA聚合酶结合,改变RNA聚合酶的构象,使其对终止信号不敏感

西安电子科技大学 生命科学技术学院
• 基本概念• RNA 的结构、分类和功能• 转录的基本过程• RNA 聚合酶、启动子• 原核生物与真核生物 mRNA 的特征比较• 转录后加工• RNA 合成与 DNA 合成异同点
Contents
西安电子科技大学 生命科学技术学院
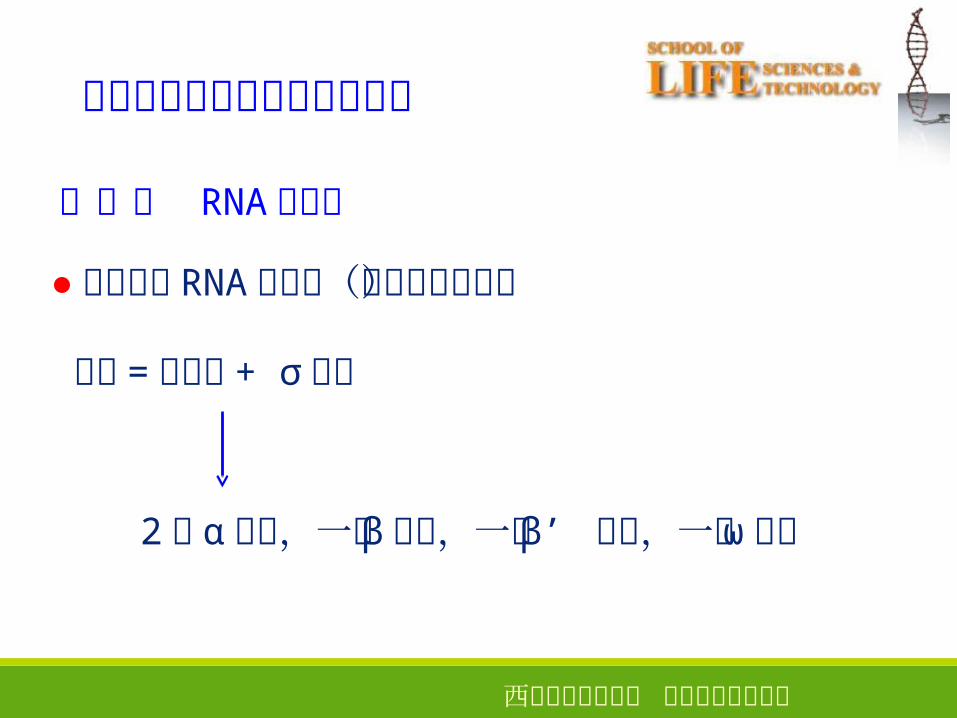
参与转录起始的关键酶与元件
(一) RNA 聚合酶
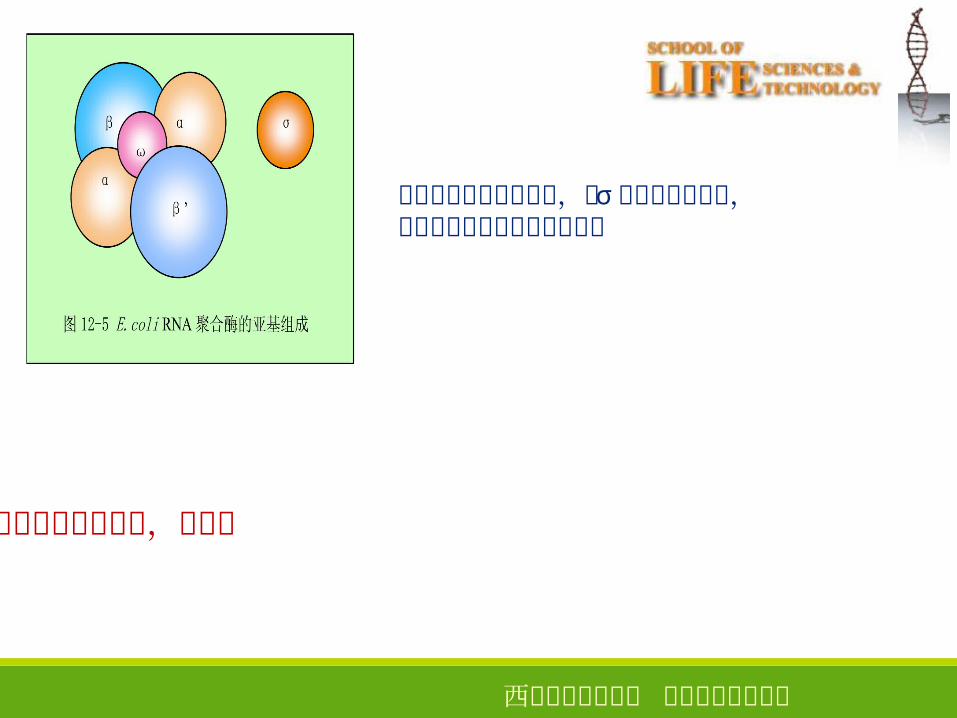
●原核生物 RNA 聚合酶(大肠杆菌为例)
全酶 = 核心酶 + σ 因子
2 个 α 亚基,一个 β 亚基,一个 β’ 亚基,一个 ω 亚基
西安电子科技大学 生命科学技术学院
转录起始过程需要全酶,由 σ 因子辨认起始点,延伸过程仅需要核心酶的催化
全酶:Mw 465 000
α亚基:核心酶组装,启动子识别β和 β’亚基:催化中心σ因子:辨认起始点,增加聚合酶对启动子的亲和力,降低其
对非专一位点的亲和力

西安电子科技大学 生命科学技术学院
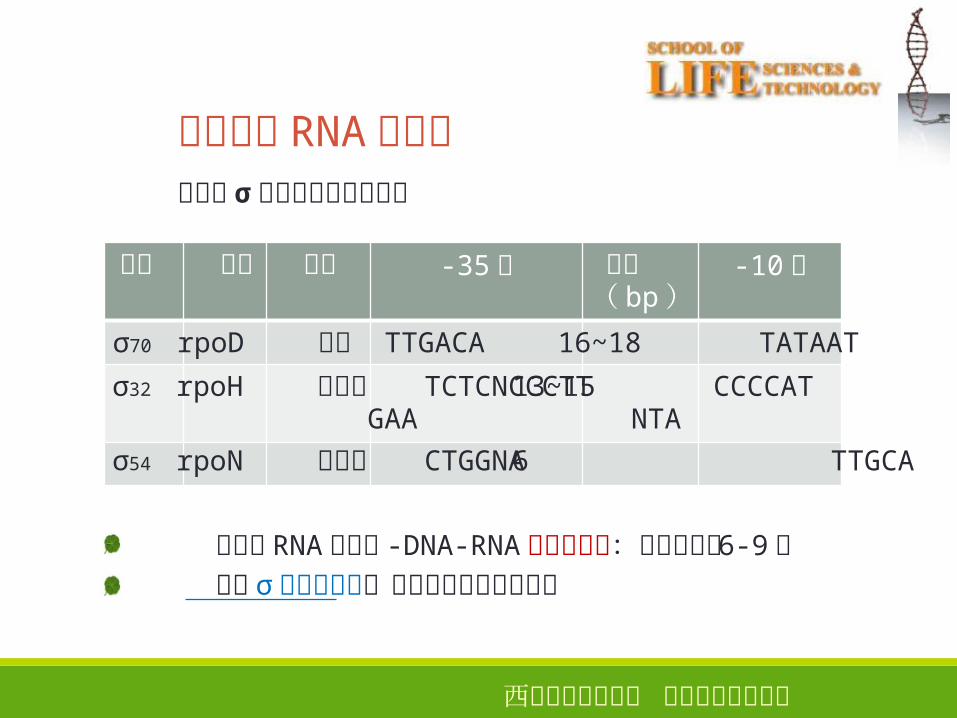
大肠杆菌 RNA 聚合酶的组成分析
亚基
基因 相对分子量
亚基数
组分 功能
α rpoA 36500 2 核心酶 核心酶组装,启动子识别
β rpoB 151000
1 核心酶 β和 β' 共同形成 RNA合成的活性中心
β' rpoC 155000
1 核心酶
ω ? 11000 1 核心酶 ?
σrpoD 70000 1 σ 因子 存在多种 σ 因子,用于
识别不同的启动子
西安电子科技大学 生命科学技术学院
因子 基因 功能 -35区 间隔( bp)
-10区
σ70 rpoD 广泛 TTGACA 16~18 TATAAT
σ32
σ54
rpoH 热休克 TCTCNCCCTTGAA
rpoN 氮代谢 CTGGNA
13~15 CCCCATNTA
6 TTGCA
25
稳定的 RNA聚合酶 -DNA-RNA三元复合物:核苷酸合成 6-9个随后 σ因子的释放,转录从起始到延伸阶段
原核生物 RNA聚合酶不同的 σ因子识别不同启动子
西安电子科技大学 生命科学技术学院
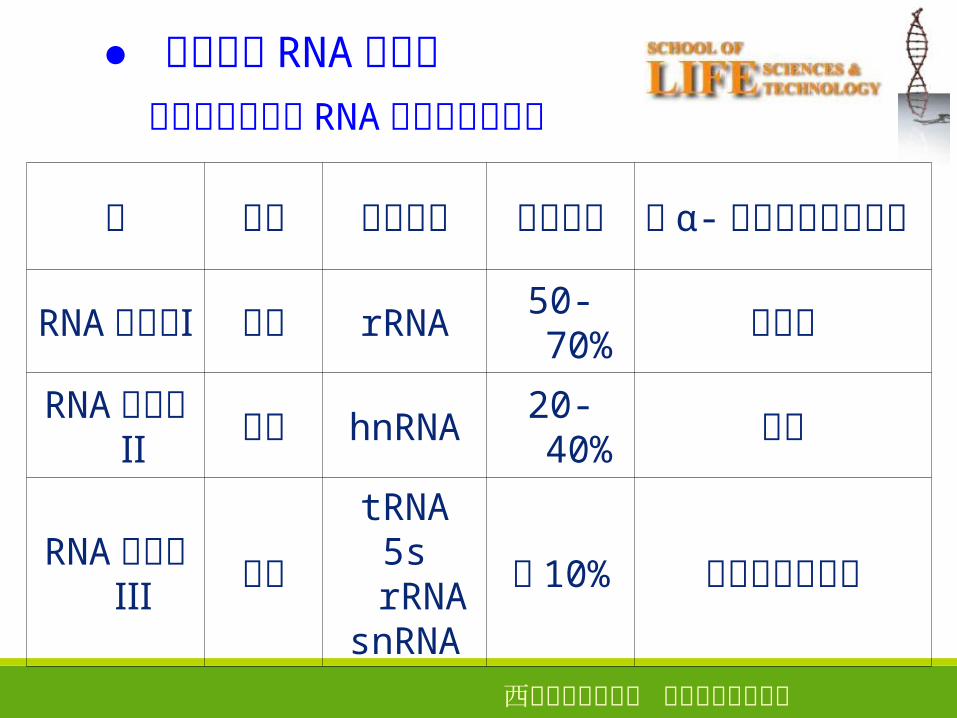
● 真核生物 RNA 聚合酶
酶 位置 转录产物
相对活性
对 α- 鹅膏蕈碱的敏感性
RNA 聚合酶Ⅰ
核仁 rRNA50-70%
不敏感
RNA 聚合酶Ⅱ
核质 hnRNA20-40%
敏感
RNA 聚合酶Ⅲ
核质tRNA5s rRNA
snRNA
约 10% 存在物种特异性
真核细胞的三种 RNA 聚合酶特征比较
西安电子科技大学 生命科学技术学院
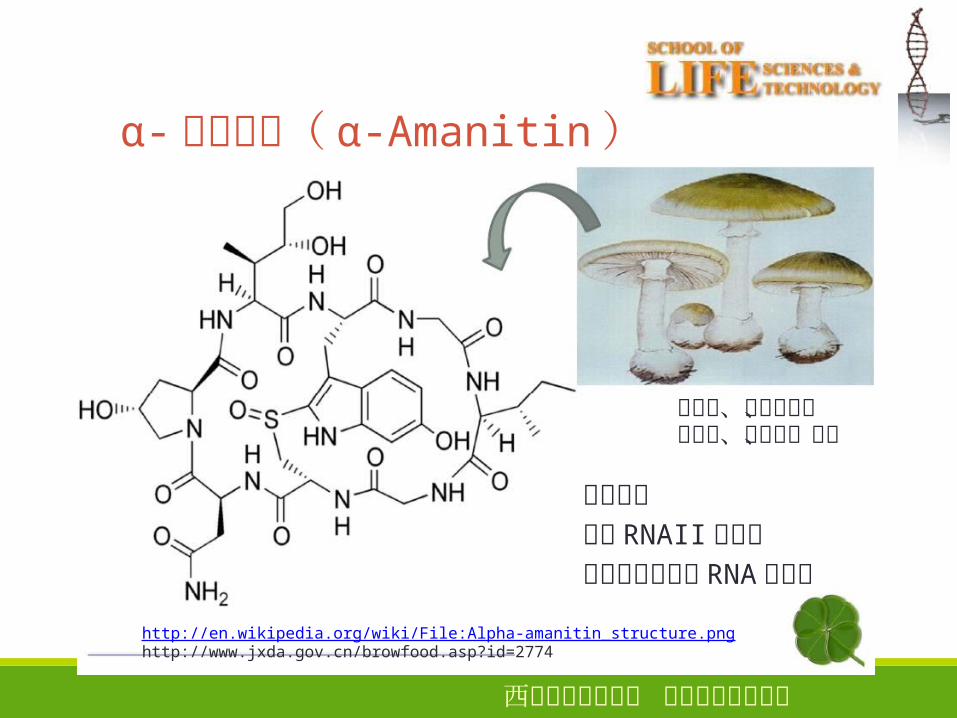
α-鹅膏覃碱( α-Amanitin)
28
绿帽菌、鬼笔鹅膏、蒜叶菌、高把菌、毒伞
二环八肽抑制 RNAII聚合酶不抑制原核生物 RNA聚合酶
http://en.wikipedia.org/wiki/File:Alpha-amanitin_structure.pnghttp://www.jxda.gov.cn/browfood.asp?id=2774
西安电子科技大学 生命科学技术学院
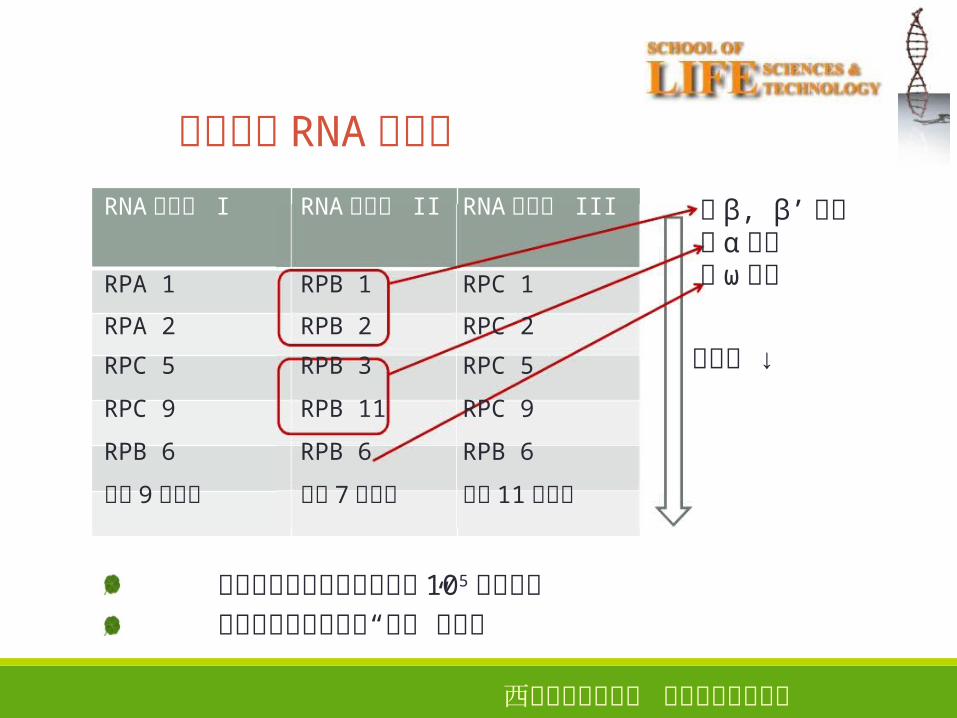
真核生物 RNA聚合酶
30
RNA聚合酶 I
RPA 1
RPA 2
RPC 5
RPC 9
RPB 6
其它 9个亚基
RNA聚合酶 II
RPB 1
RPB 2
RPB 3
RPB 11
RPB 6
其它 7个亚基
RNA聚合酶 III
RPC 1
RPC 2
RPC 5
RPC 9
RPB 6
其它 11个亚基
与 β, β’同源与 α同源与 ω同源
分子量 ↓
聚合酶中有两个分子量超过 105的大亚基“ ”同种生物三类聚合酶 共享 小亚基
西安电子科技大学 生命科学技术学院
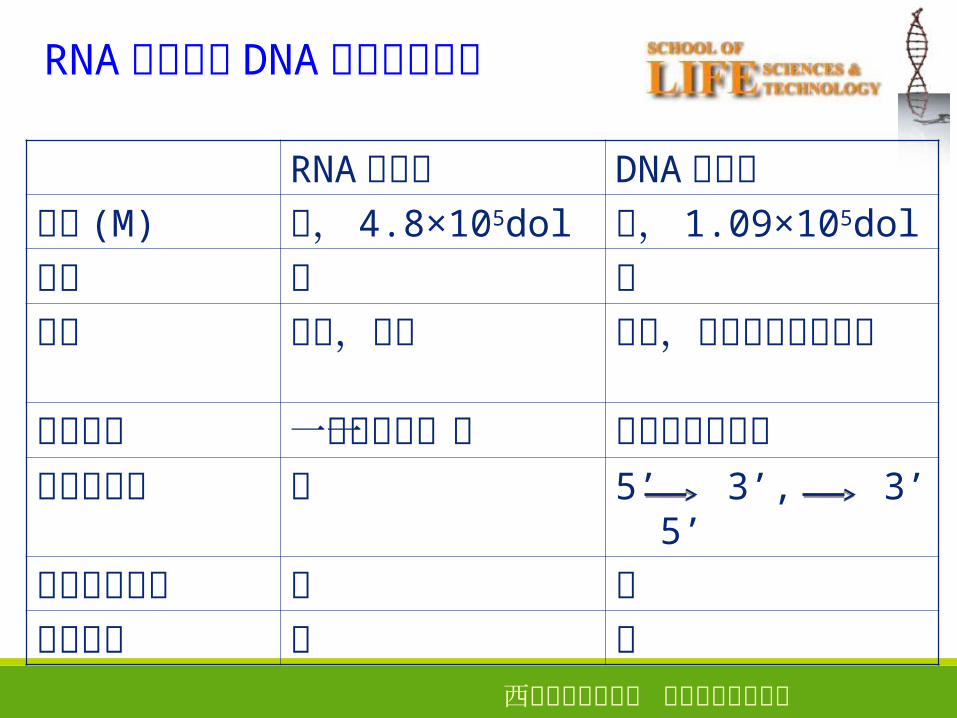
RNA 聚合酶与 DNA 聚合酶的区别
RNA 聚合酶 DNA 聚合酶大小 (M) 大, 4.8×105dol 小, 1.09×105do
l
引物 无 有产物 较短,游离 较长,与模板以氢
键相连作用方式 一条链的某一段 两条链同时进行外切酶活性 无 5’ 3’, 3’ 5’
校对合成能力 无 有修复能力 无 有

西安电子科技大学 生命科学技术学院
• 基本概念• RNA 的结构、分类和功能• 转录的基本过程• RNA 聚合酶、启动子• 原核生物与真核生物 mRNA 的特征比较• 转录后加工• RNA 合成与 DNA 合成异同点
Contents
西安电子科技大学 生命科学技术学院
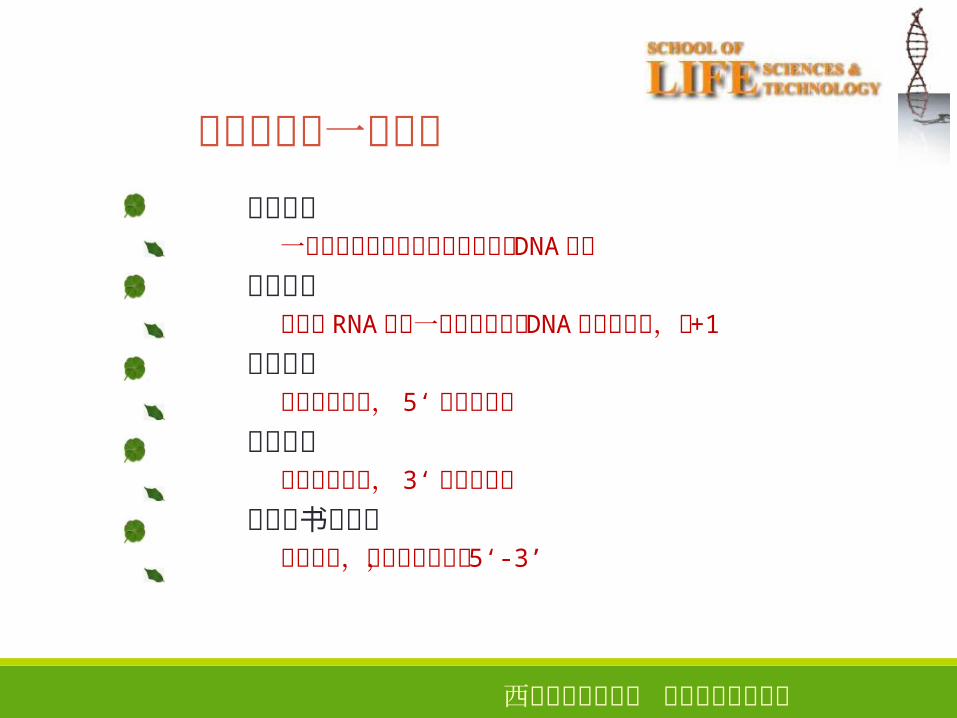
关于转录的一些概念
转录单位一段从启动子开始到终止子结束的 DNA序列
转录起点与新生 RNA链第一个核苷酸对应 DNA链上的碱基,为+1
上游序列转录起点前面, 5‘末端的序列
下游序列转录起点后面, 3‘末端的序列
序列的书写方向从左到右,从上游到下游, 5‘-3’
33

西安电子科技大学 生命科学技术学院
启动子 (promoter)
启动子定义:指能被 RNA 聚合酶识别、结合
并启动基因转录的一段 DNA 序列。
西安电子科技大学 生命科学技术学院
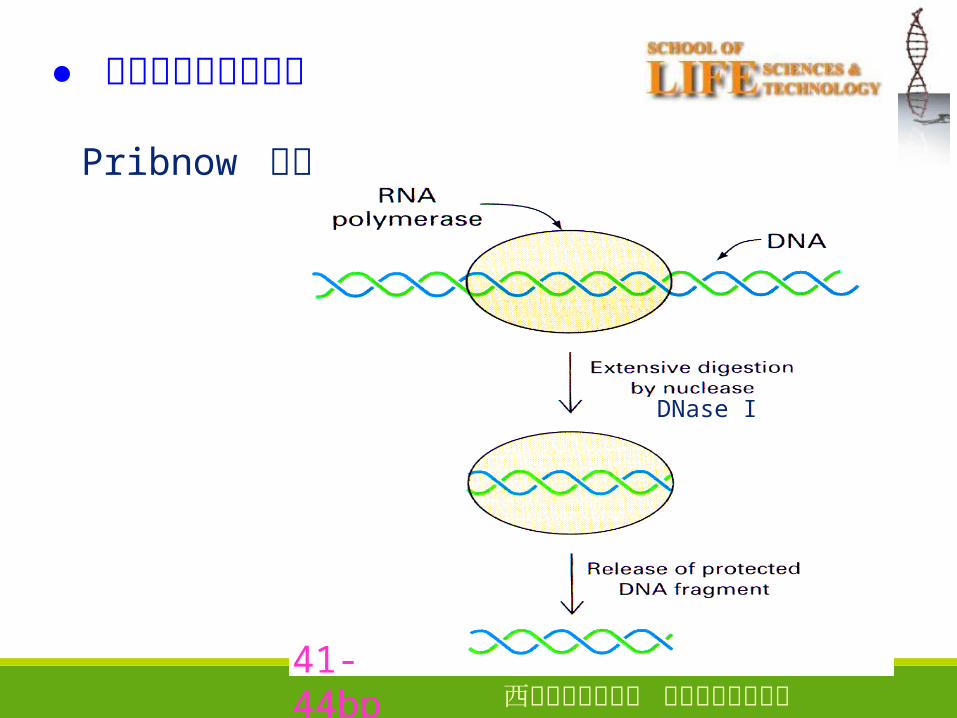
● 原核生物启动子结构
Pribnow 实验
41-44bp
DNase I

西安电子科技大学 生命科学技术学院
TTGACA 区(- 35 区):提供了 RNA 聚合酶全酶识别的信号
TATA 区(- 10 区, Pribnow 区):酶的紧密结合位点,富含 AT 碱基,利于双链打开。

西安电子科技大学 生命科学技术学院
模板链 AACTGT ATATTA
TTGACA TATAAT
3’
5’
DNA
转录起始点
Probnow盒子启动子
35 10 +1转录区
5’ 3’RNA
编码链

西安电子科技大学 生命科学技术学院
大肠杆菌 RNA 聚合酶全酶所识别的启动子区
T85T83G81A61C69A52 T89A89T50A65A100
- 10 区- 35 区
西安电子科技大学 生命科学技术学院
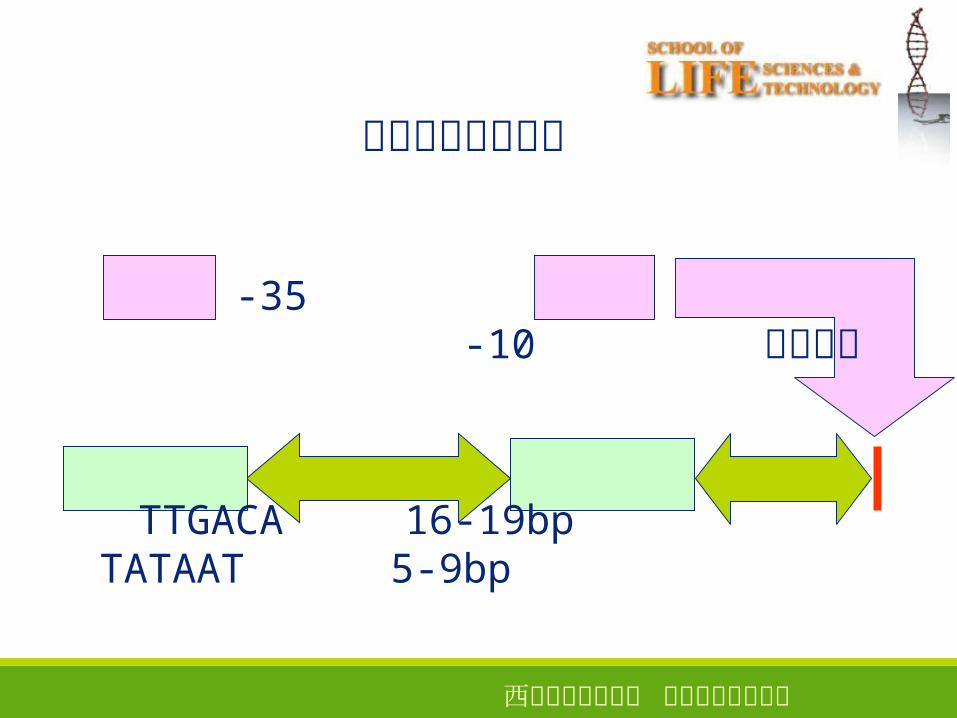
典型启动子的结构
-35 -10 转录起点
TTGACA 16-19bp TATAAT 5-9bp

西安电子科技大学 生命科学技术学院
-10区和 -35区的最佳间距
16~19bp�
增减 1bp� 会使两者产生的超螺旋的夹角变化 36°
细菌中的启动子突变下降突变:转录水平下降的突变, e.g.-10� 区的 TATAAT变成 AATAAT
上升突变:转录水平提高的突变, e.g.-10� 区的 TATGTT变成 TATATT
39
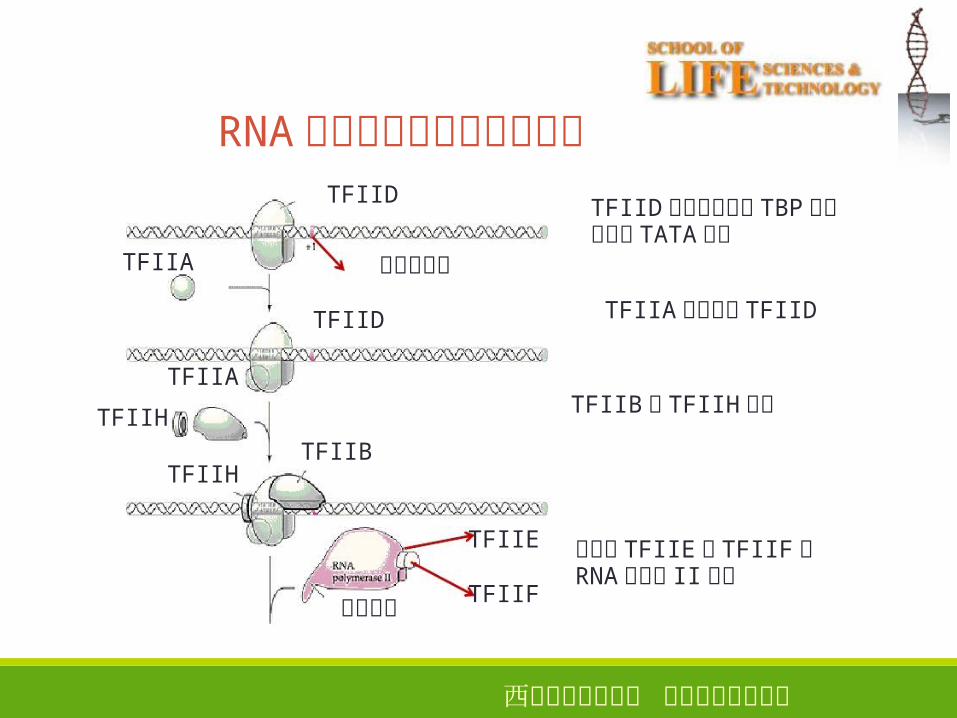
西安电子科技大学 生命科学技术学院
RNA聚合酶和启动子区的结合
37
TFIIA
TFIIHTFIIB
TFIIA
TFIIH
TFIID
转录起始点
TFIID
TFIID复合物通过其 TBP单元结合到 TATA区上
TFIIA帮助稳定 TFIID
TFIIB和 TFIIH加入
伴随了 TFIIE和 TFIIF的RNA聚合酶 II结合
羧端尾巴
TFIIE
TFIIF
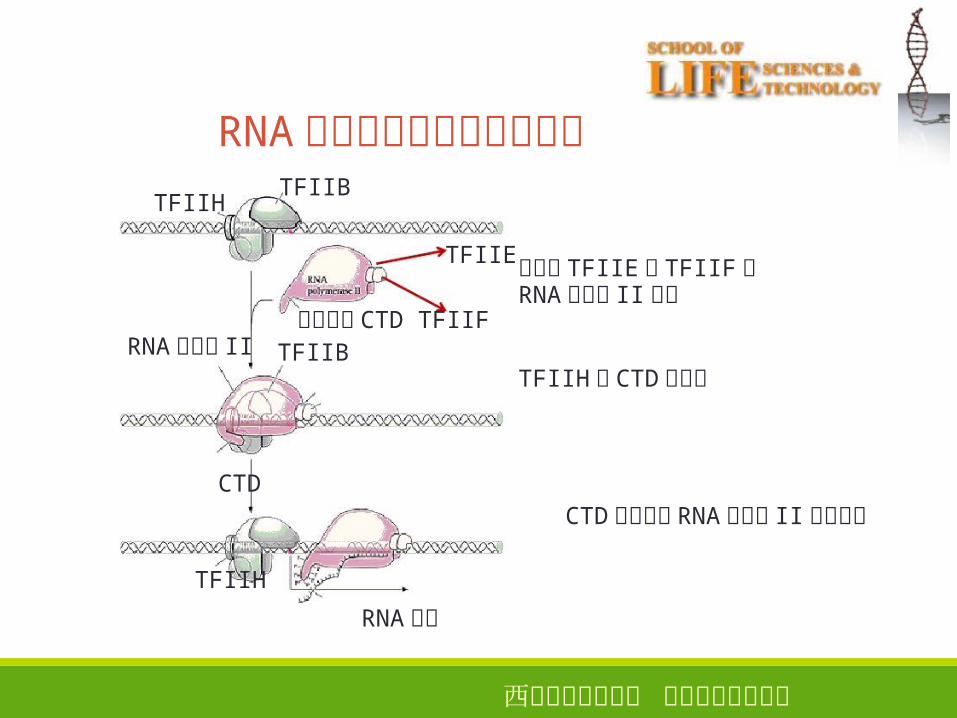
西安电子科技大学 生命科学技术学院
RNA聚合酶和启动子区的结合
38
TFIIBTFIIH
伴随了 TFIIE和 TFIIF的RNA聚合酶 II结合
TFIIH将 CTD磷酸化RNA聚合酶 II
TFIIE
羧端尾巴 CTD TFIIFTFIIB
CTDCTD磷酸化后 RNA聚合酶 II才能移动
TFIIH
RNA转录
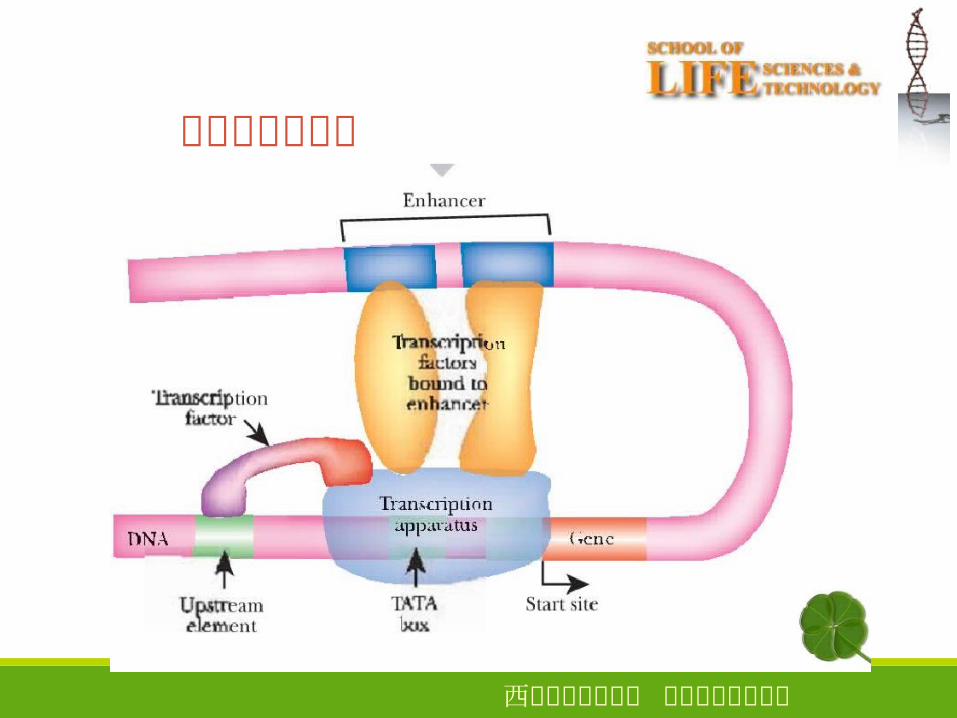
西安电子科技大学 生命科学技术学院
增强子及其功能
40
西安电子科技大学 生命科学技术学院
增强子及其功能
功能:强化转录的开始特点远距离效应 相距>10kb� 也能发挥作用无方向性 可以位于靶基因的上游、下游或内部顺式调节 只调节同一条染色体上的靶基因无物种和基因的特异性具有组织特异性 需要特定的蛋白质因子参与有相位性 与 DNA构象有关有的增强子可以对外部信号产生反应
41
西安电子科技大学 生命科学技术学院
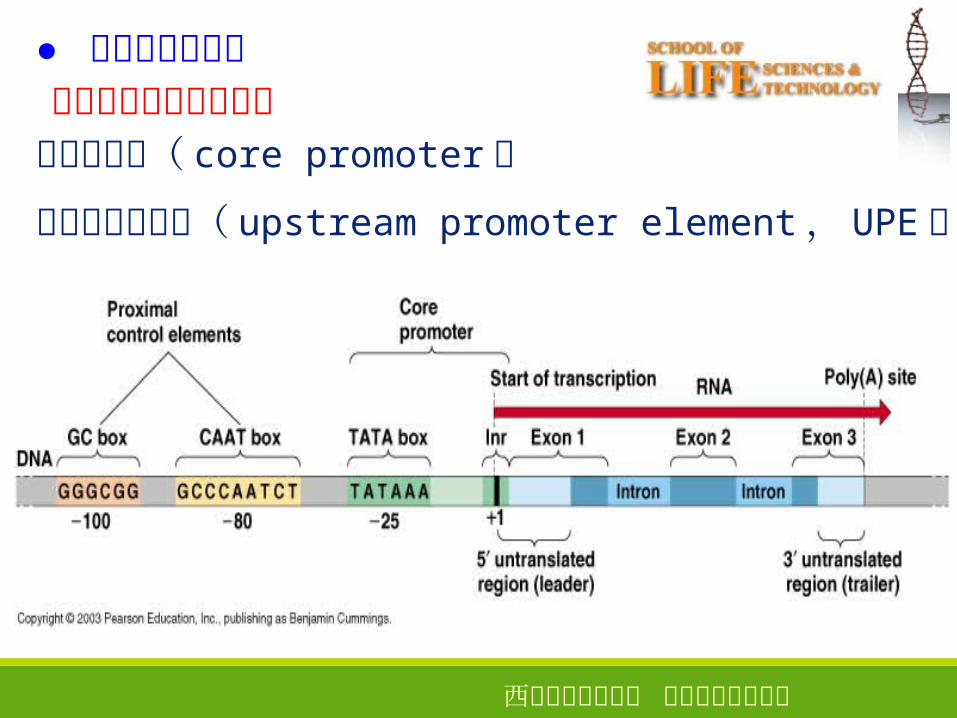
真核生物启动子的结构核心启动子( core promoter )
上游启动子元件( upstream promoter element, UPE )
● 真核生物启动子

西安电子科技大学 生命科学技术学院
1 、核心启动子
●定义:指保证 RNA 聚合酶Ⅱ转录正常起始所必需的、最少的 DNA 序列,包括转录起始位点及转录起始位点上游 TATA 区
●作用:选择正确的转录起始位点,保证精确起始
TATA 常在 -25bp左右,相当于原核的 -10 序列T85A97T93A85A63A83A50

西安电子科技大学 生命科学技术学院
2 、上游启动子元件
●包括CAAT盒( CCAAT )和 GC盒( GGGCGG )等
●作用:控制转录起始频率。
CAAT: -70 - -80bp
GGGCGG: -80 - -110bp
西安电子科技大学 生命科学技术学院
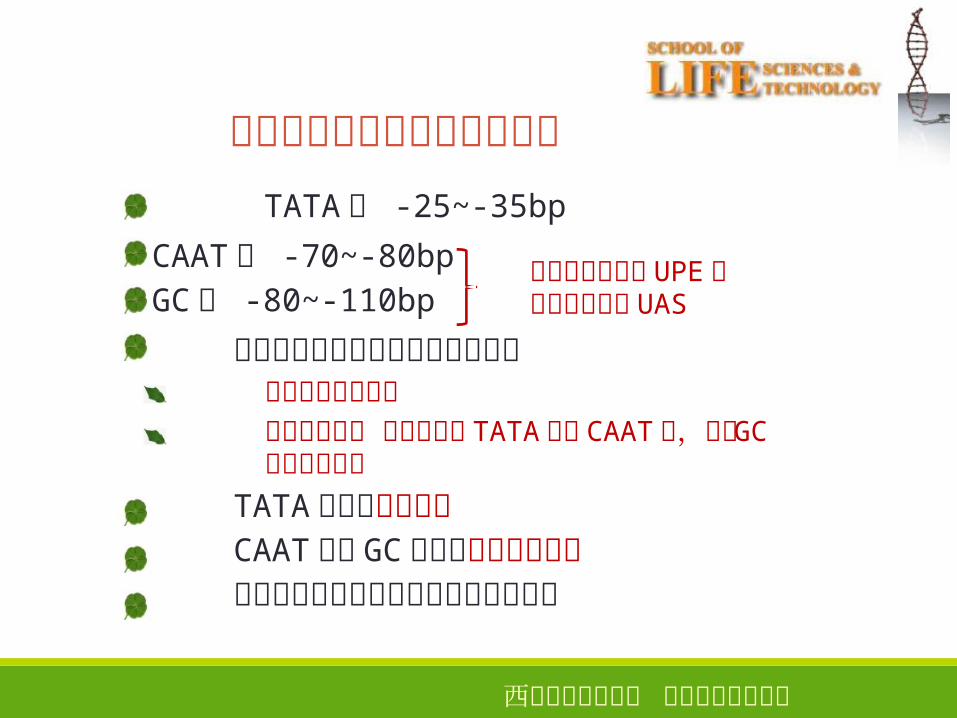
真核生物启动子对转录的影响
TATA区 -25~-35bp�CAAT区 -70~-80bp�GC区 -80~-110bp�真核生物启动区相对于原核的区别启动区的范围更大结合位点更多 除了对应的 TATA区和 CAAT区,还有 GC区和增强子区
TATA区确定起始位点CAAT区和 GC区控制转录起始频率不是所有的启动子区都含有这三种序列
42
上游启动子元件 UPE或上游激活序列 UAS
西安电子科技大学 生命科学技术学院
转录的抑制
DNA模板功能抑制剂与 DNA结合抑制模板的功能
RNA聚合酶抑制剂与 RNA聚合酶结合,抑制其活力
43

西安电子科技大学 生命科学技术学院
DNA模板功能抑制剂
放线菌素 D�与 DNA形成非共价复合物,抑制其作为模板的功能1mM �即可有效抑制转录过程,抗菌、抗癌烷化剂如氮芥,磺酸酯,氮丙啶等,使 DNA发生交联,抑制其模板功能,致癌如环磷酰胺,能选择地杀伤肿瘤细胞,治疗恶性肿瘤嵌入染料嵌入 DNA相邻碱基对之间,使得 DNA复制中缺失或增加一个核苷酸如 EB,与核酸结合后抑制其复制和转录
44
西安电子科技大学 生命科学技术学院
RNA聚合酶抑制物
利福霉素强烈抑制革兰阳性菌和结核杆菌,抗菌
利迪链霉素与细菌 RNA聚合酶的 β亚基结合,抑制转录的起始
α-鹅膏覃碱抑制真核生物 RNA聚合酶
45

西安电子科技大学 生命科学技术学院
• 基本概念• RNA 的结构、分类和功能• 转录的基本过程• RNA 聚合酶、启动子• 原核生物与真核生物 mRNA 的特征比较• 转录后加工• RNA 合成与 DNA 合成异同点
Contents
西安电子科技大学 生命科学技术学院
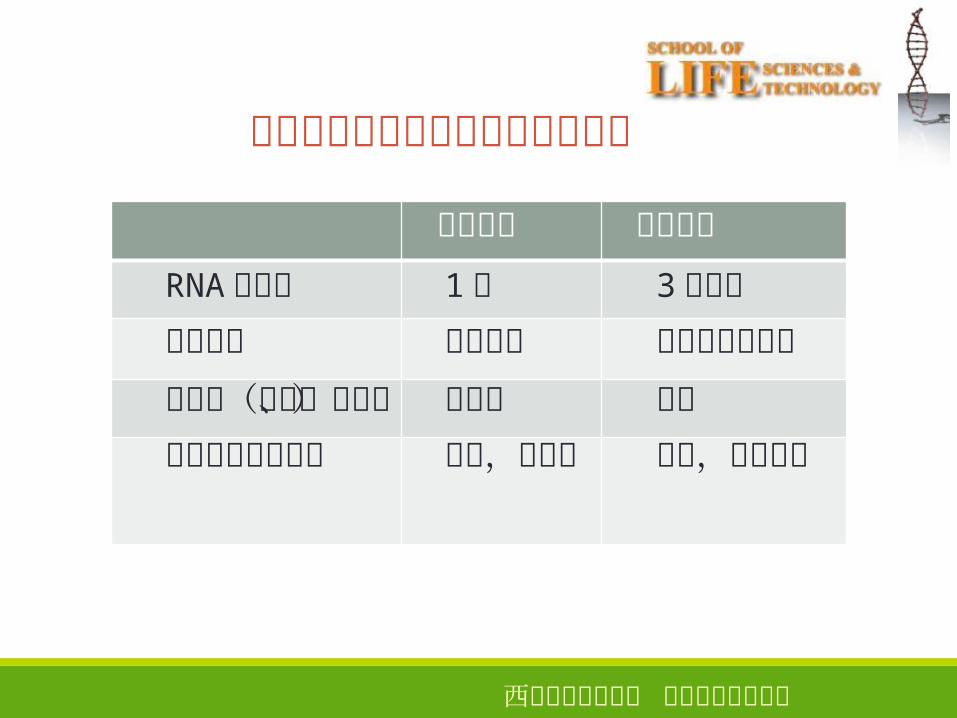
原核生物和真核生物转录产物比较
3
原核生物 真核生物
RNA聚合酶
转录产物
后加工(剪接、修饰)转录和翻译的时空
1种
编码序列
不需要
同时,同空间
3种以上
含有内含子序列
需要
先后,不同空间
西安电子科技大学 生命科学技术学院
原核生物与真核生物 mRNA 的特征比较
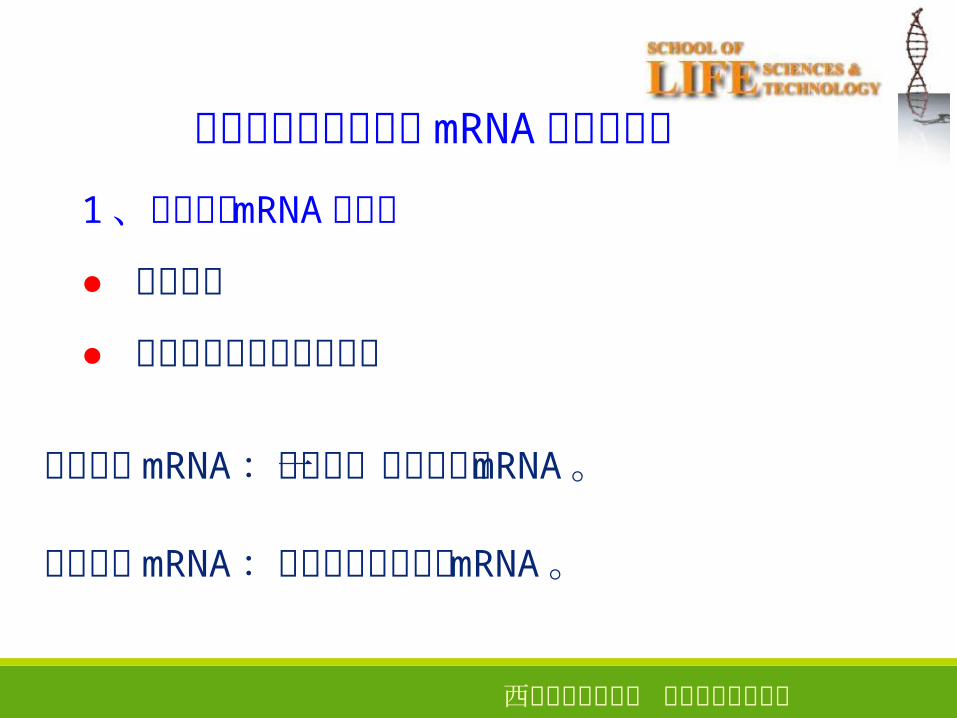
1 、原核生物 mRNA 的特征
● 半衰期短
● 多以多顺反子的形式存在
多顺反子 mRNA :编码多个蛋白质的 mRNA 。
单顺反子 mRNA :只编码一个蛋白质的 mRNA 。
西安电子科技大学 生命科学技术学院
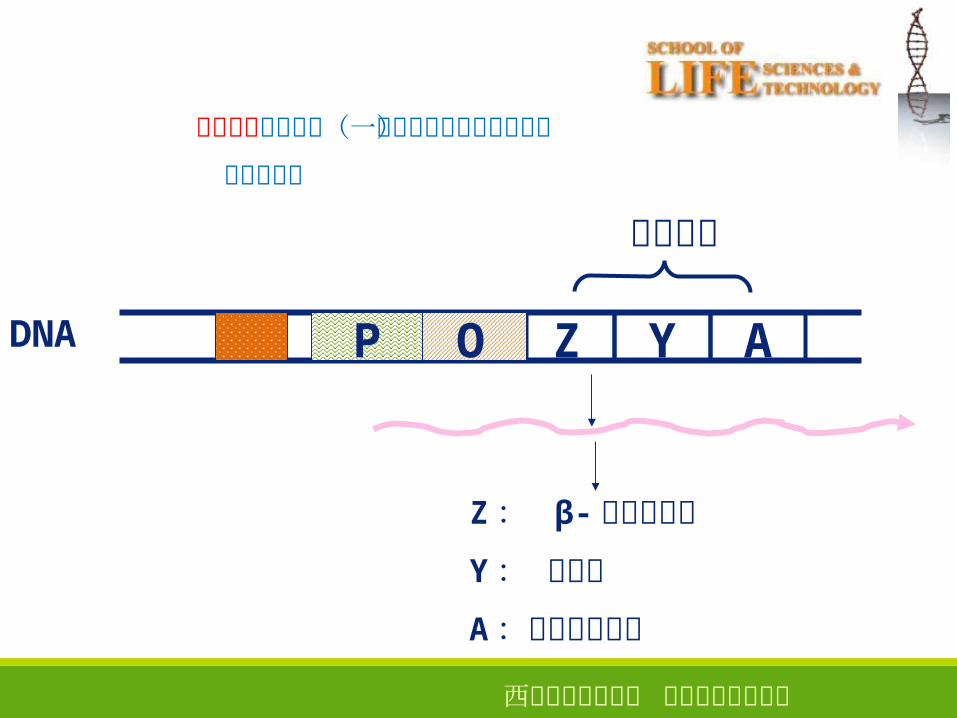
结构基因
Z: β-半乳糖苷酶
Y: 透过酶
A:乙酰基转移酶
Z Y AOPDNA
多顺反子是操纵子(一组相邻或相互重叠基因)
的转录产物
西安电子科技大学 生命科学技术学院
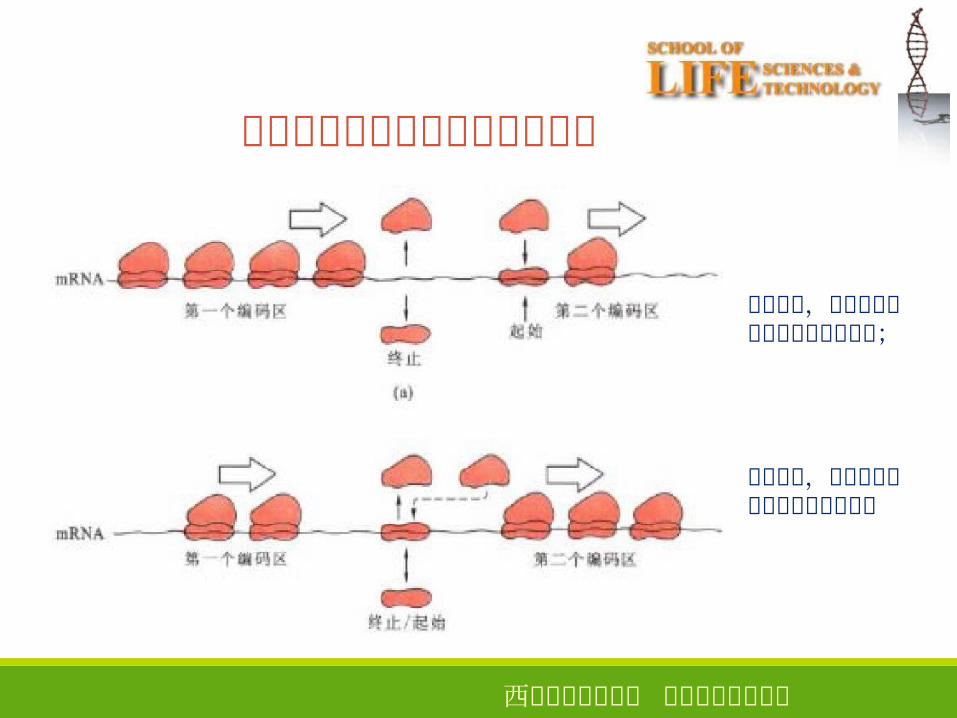
上游顺反子对下游顺反子的影响
7
相距较远,前后顺反子翻译终止和起始独立;
相距较近,前后顺反子翻译终止和起始衔接

西安电子科技大学 生命科学技术学院
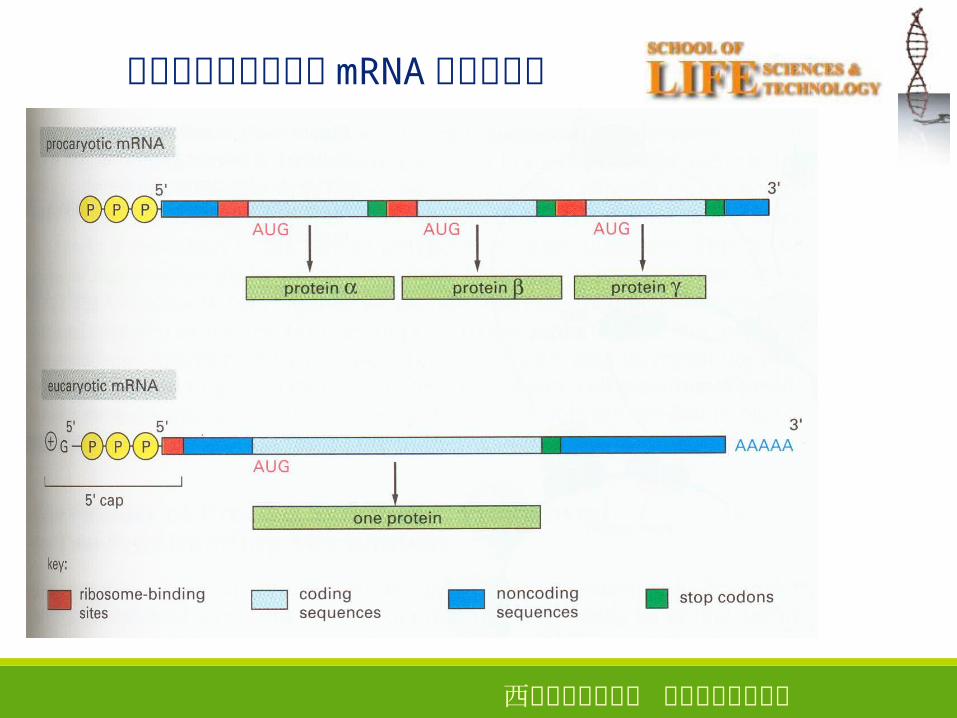
● 5’ 端无“帽子”结构, 3’ 端没有或只有较短的 poly( A )结构。
SD 序列: mRNA 中用于结合原核生物核糖体的序列。
•起始密码子 AUG上游 7~12个核苷酸• 与 16SrRNA� (核糖体中) 3’端反向互补•在核糖体与mRNA的结合过程中起作用

西安电子科技大学 生命科学技术学院
2 、真核生物 mRNA 的特征
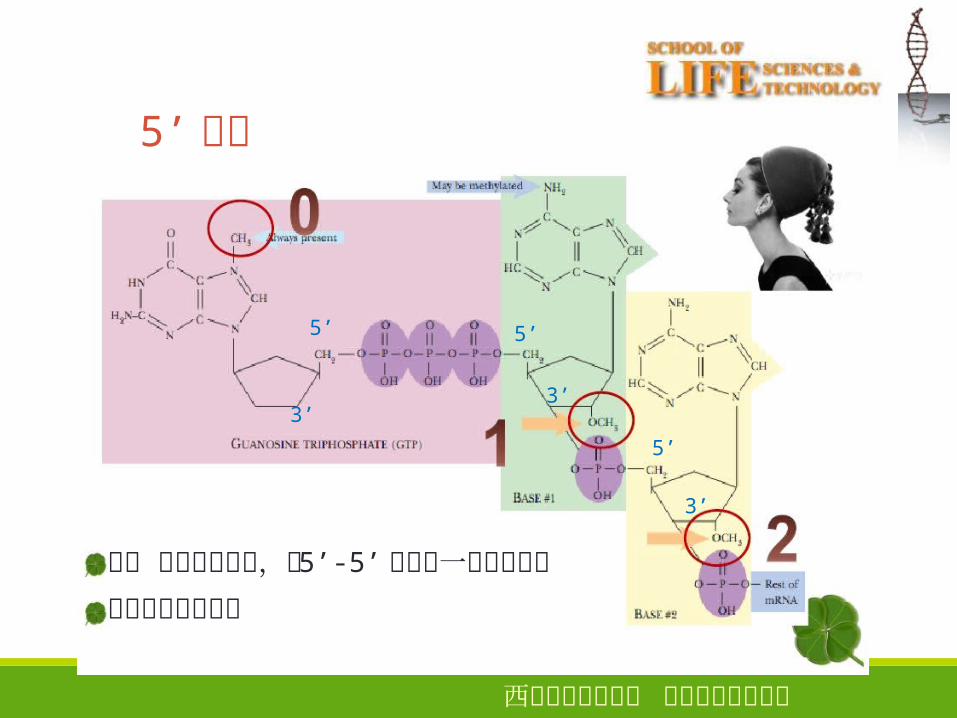
● 5’ 端存在“帽子”结构
●多数 mRNA 3’ 端具有 poly( A )尾巴(组蛋白除外)
●以单顺反子的形式存在
“ 基因”的分子生物学定义:产生一条多肽链或功能 RNA所必需的全部核甘酸序列。
西安电子科技大学 生命科学技术学院
3’
5’5’
3’
5’帽子
11
5’
3’
帽子 甲基化鸟嘌呤,是 5’-5’接到第一个核苷酸上
帽子常常被甲基化

西安电子科技大学 生命科学技术学院
帽子结构功能:
① 能被核糖体小亚基识别,促使 mRNA 和核糖体的结合;
②m7Gppp 结构能有效地封闭mRNA 5’末端,以保护mRNA免受 5’ 核酸外切酶的降解,增强mRNA 的稳定。
西安电子科技大学 生命科学技术学院
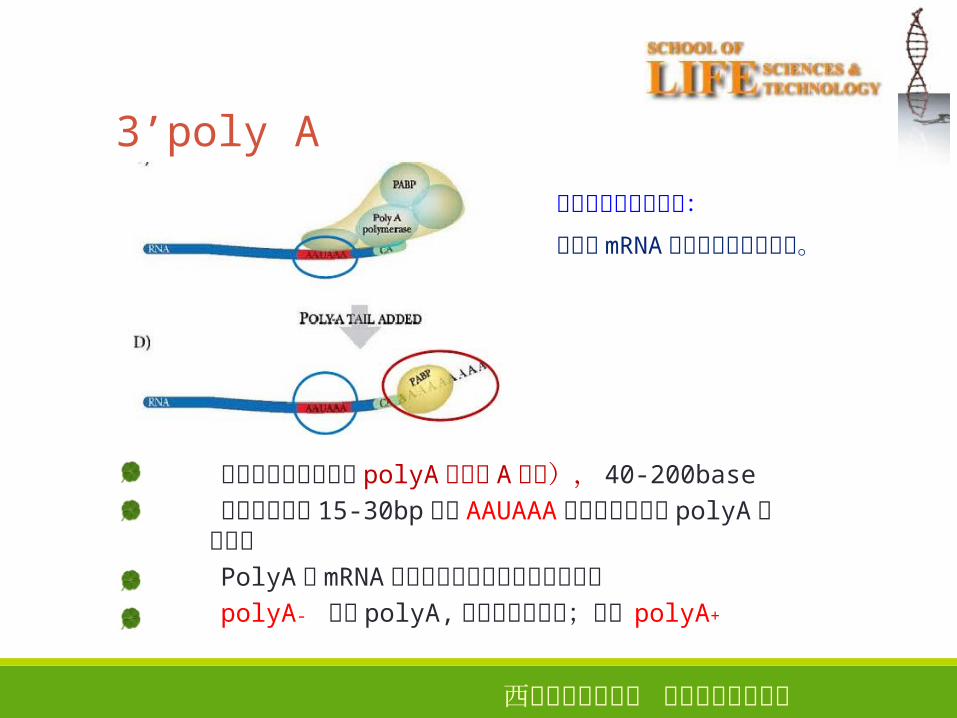
3’poly A
大多数真核生物都有 polyA� (多聚 A尾巴), 40-200base�终止位点上游 15-30bp� 处的 AAUAAA对于切割和加上 polyA� 是必需的PolyA� 是mRNA从细胞核进入细胞质的必需形式polyA- 没有 polyA,主要编码组蛋白;其它 polyA+
13
多聚腺苷酸尾巴功能:提高了 mRNA 在细胞质中的稳定性。
西安电子科技大学 生命科学技术学院
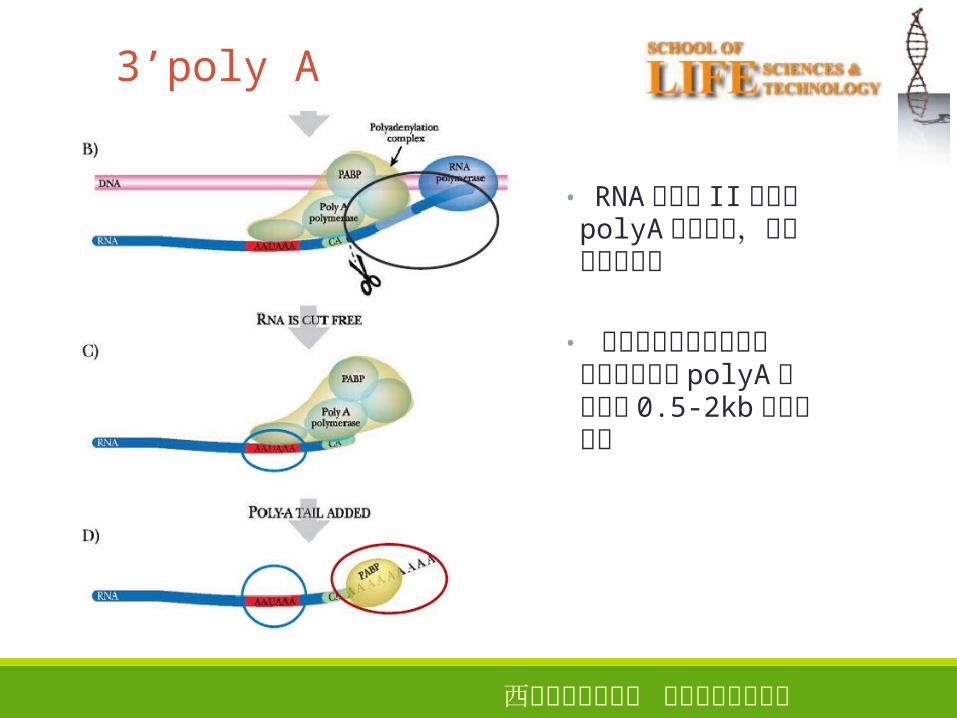
3’poly A 14
• RNA聚合酶 II并不在polyA位点终止,而往往继续转录
• 大部分已知基因的初级转录产物拥有 polyA位点下游 0.5-2kb核苷酸序列
西安电子科技大学 生命科学技术学院
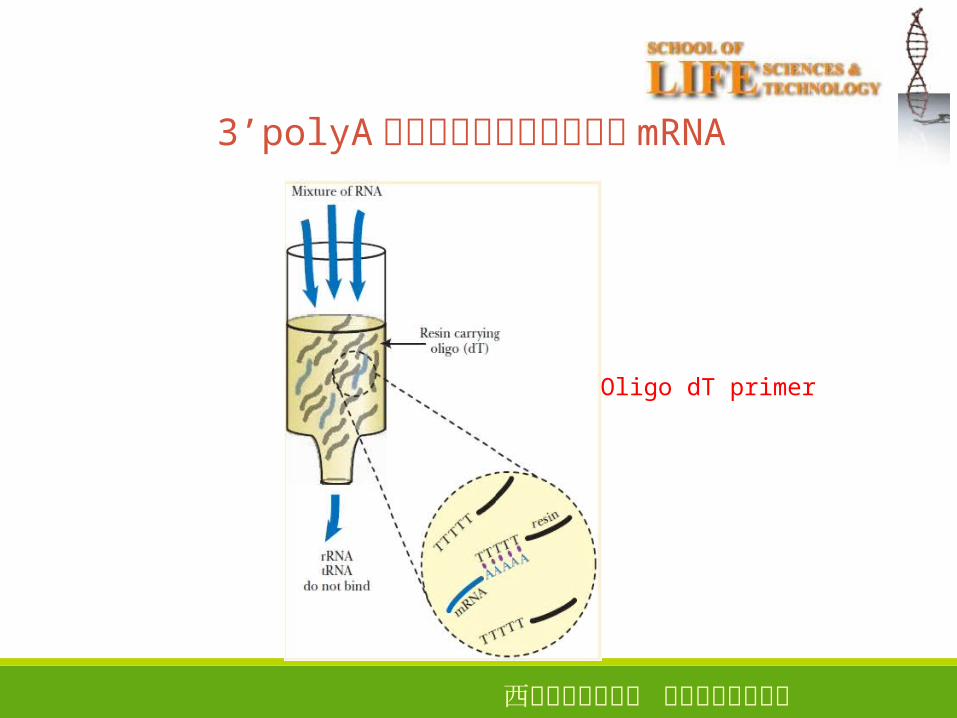
3’polyA� 的特征用于分离真核生物mRNA
15
Oligo dT primer
西安电子科技大学 生命科学技术学院
原核生物和真核生物 mRNA 结构的比较

西安电子科技大学 生命科学技术学院
• 基本概念• RNA 的结构、分类和功能• 转录的基本过程• RNA 聚合酶、启动子• 原核生物与真核生物 mRNA 的特征比较• 转录后加工• RNA 合成与 DNA 合成异同点
Contents
西安电子科技大学 生命科学技术学院
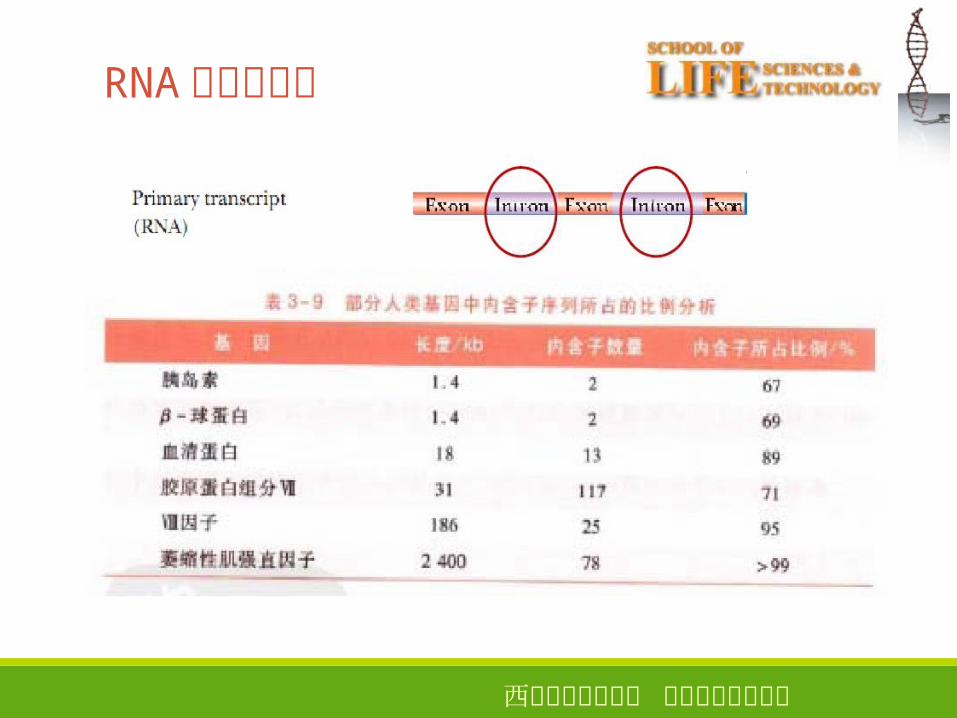
RNA中的内含子17

西安电子科技大学 生命科学技术学院
真核生物 tRNA前体的转录后加工
tRNA内含子的特点16-46个核苷酸位于反密码子的下游内含子和外显子之间没有保守序列
后加工过程内含子的剪接3’端添加 CCA
核苷酸修饰
18

西安电子科技大学 生命科学技术学院
真核生物 rRNA前体的转录后加工
大多数 rRNA基因无内含子
人 Hela细胞中 rRNA前体的剪切5’端切除非编码序列,生成 41S中间产物
41S RNA切成两段 32S 和 20S
32S RNA被切成 28S和 5.8S rRNA
20S RNA被切成 18S rRNA
19

西安电子科技大学 生命科学技术学院
真核生物mRNA剪接
Pre-mRNA的剪接
I类自剪接内含子
II类自剪接内含子
20

西安电子科技大学 生命科学技术学院
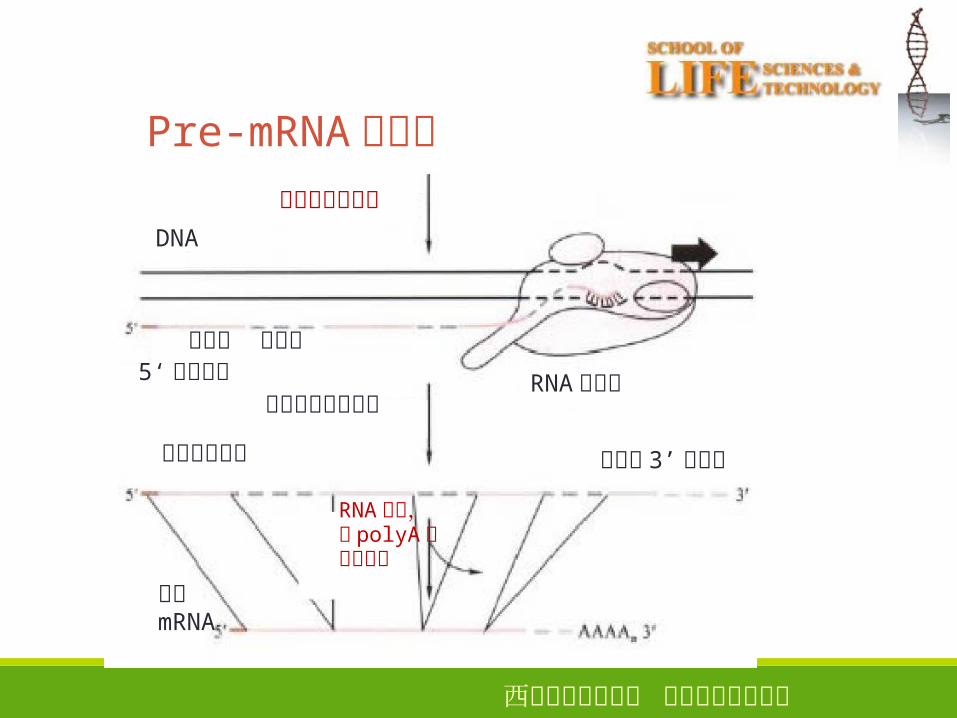
Pre-mRNA的剪接
西安电子科技大学 生命科学技术学院
21
Pre-mRNA的剪接转录和加帽反应
DNA
外显子 内含子5‘帽子结构
生成原始转录产物
原始转录产物
RNA切割,加 polyA和剪接过程
成熟mRNA
RNA聚合酶
非编码 3’端序列

西安电子科技大学 生命科学技术学院
RNA加工过程及其生理功能
22
加工过程
加帽子反应
推测的生理功能
mRNA从细胞核细胞质基质转运,翻译起始
加 polyA反应 转录终止,翻译起始,mRNA降解
RNA的剪接
RNA的切割
从mRNA, tRNA, rRNA分子中切除内含子
从前体 RNA中释放成熟 tRNA和 rRNA分子
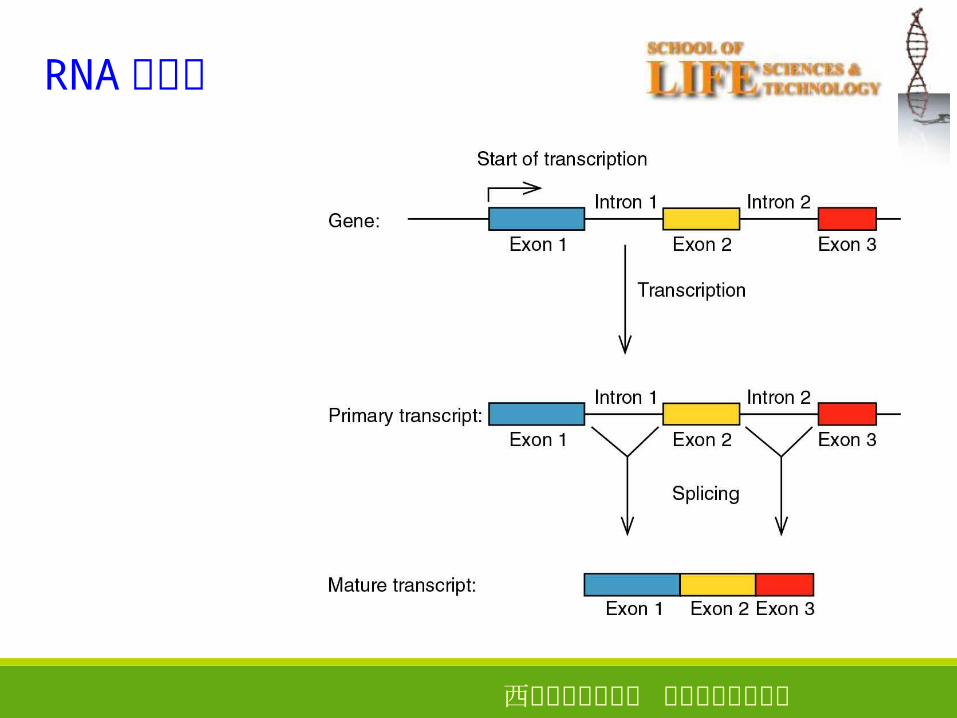
西安电子科技大学 生命科学技术学院
RNA 的剪接
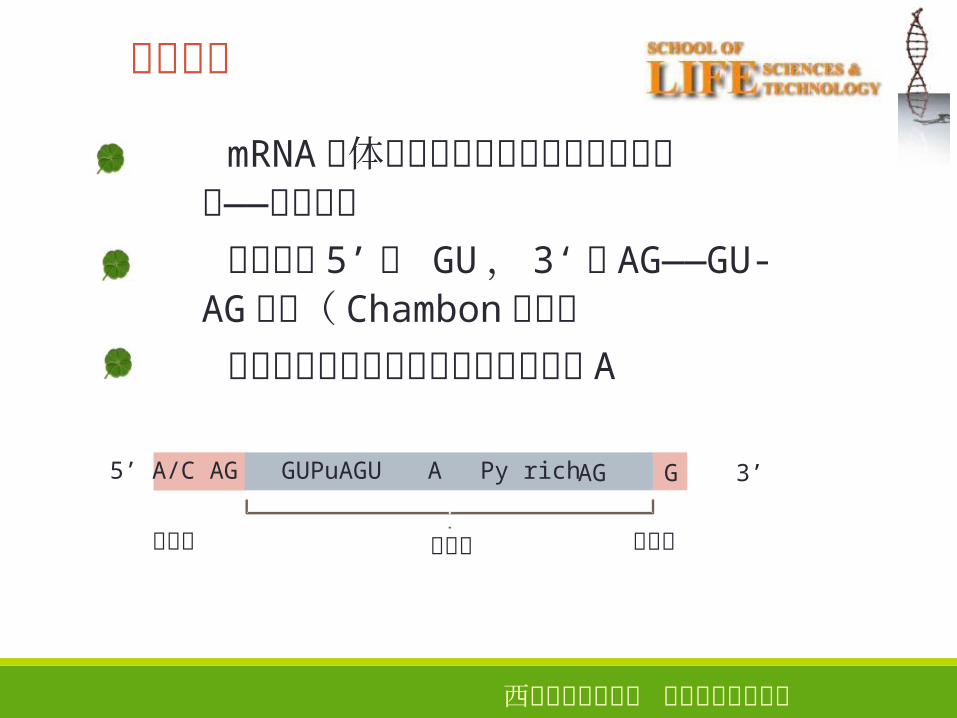
西安电子科技大学 生命科学技术学院
剪接信号
mRNA前体中内含子两端边界存在共同序——列 剪接信号内含子的 5’端 GU, 3‘端 AG——GU-
AG法则( Chambon法则)内含子的内部也可能参与,如分支点 A
23
Py rich5’ A
内含子
AG G 3’
外显子
A/C AG GUPuAGU
外显子
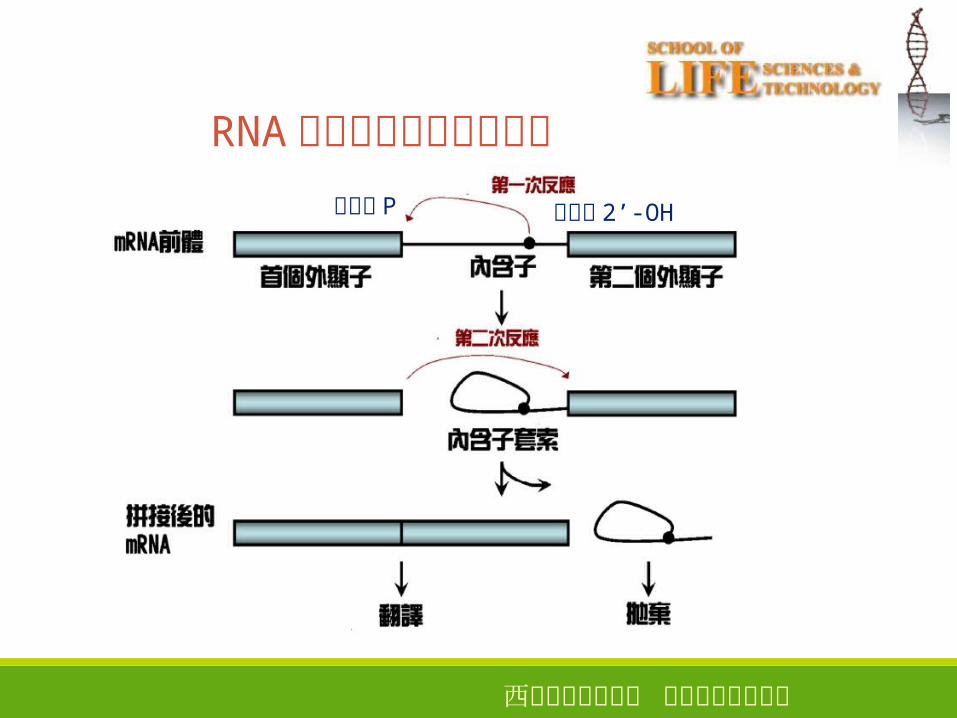
西安电子科技大学 生命科学技术学院
24
RNA剪接中的两步转酯反应
腺苷酸 2’-OH鸟苷酸 P

西安电子科技大学 生命科学技术学院
RNA剪接大多发生在剪接体上
剪接体: 150种蛋白质和 5种 RNA的大型复合体,大小与核糖体差不多
snRNA 核小 RNAsmallnuclearRNA� � �
snRNP 核小核糖核蛋白 smallnuclear� �ribonucleo-proteinparticle� �
25
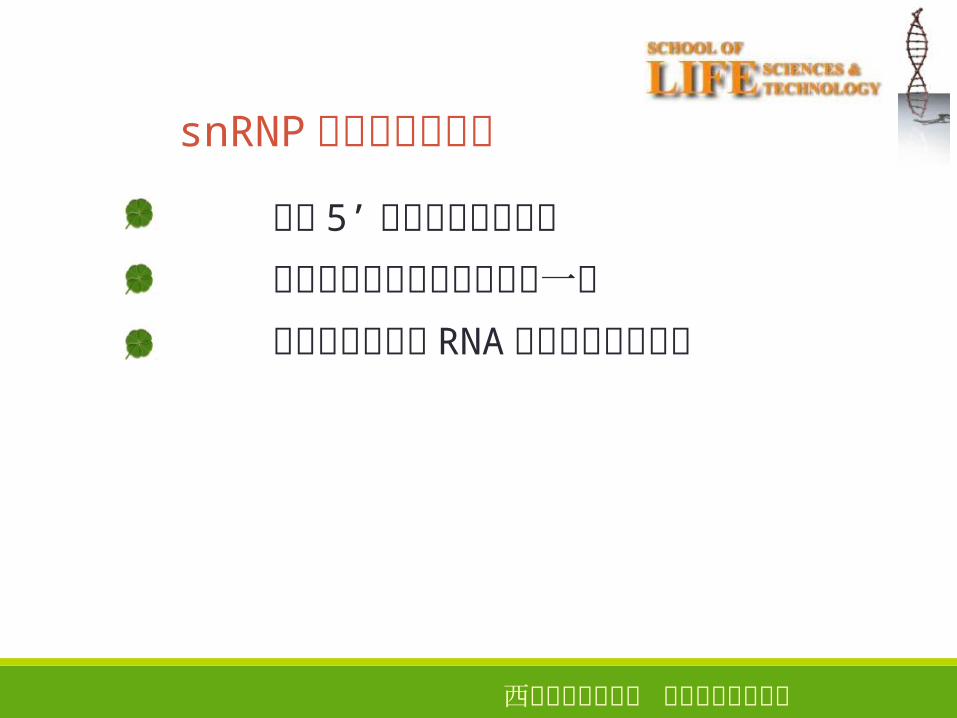
西安电子科技大学 生命科学技术学院
snRNP在剪接中的功能
识别 5’剪接位点和分支点
按需要把这两个位点集结到一起
催化或协助催化 RNA的剪接和连接反应
26
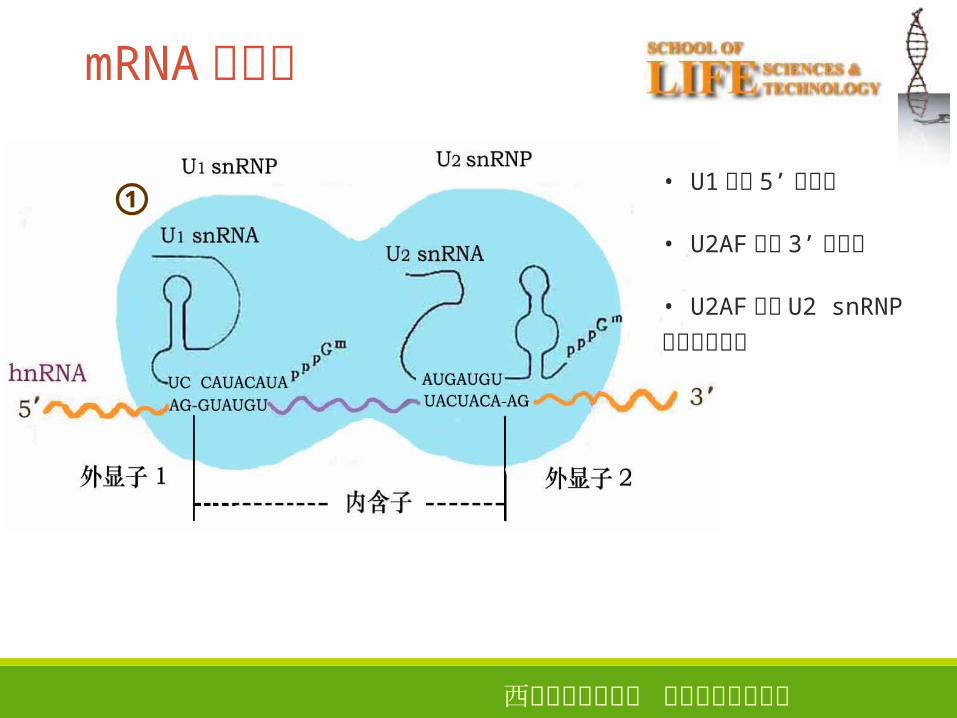
西安电子科技大学 生命科学技术学院
①
mRNA的剪接
• U1识别 5’剪接点
• U2AF识别 3’剪接点
• U2AF引导 U2 snRNP
与分支点结合
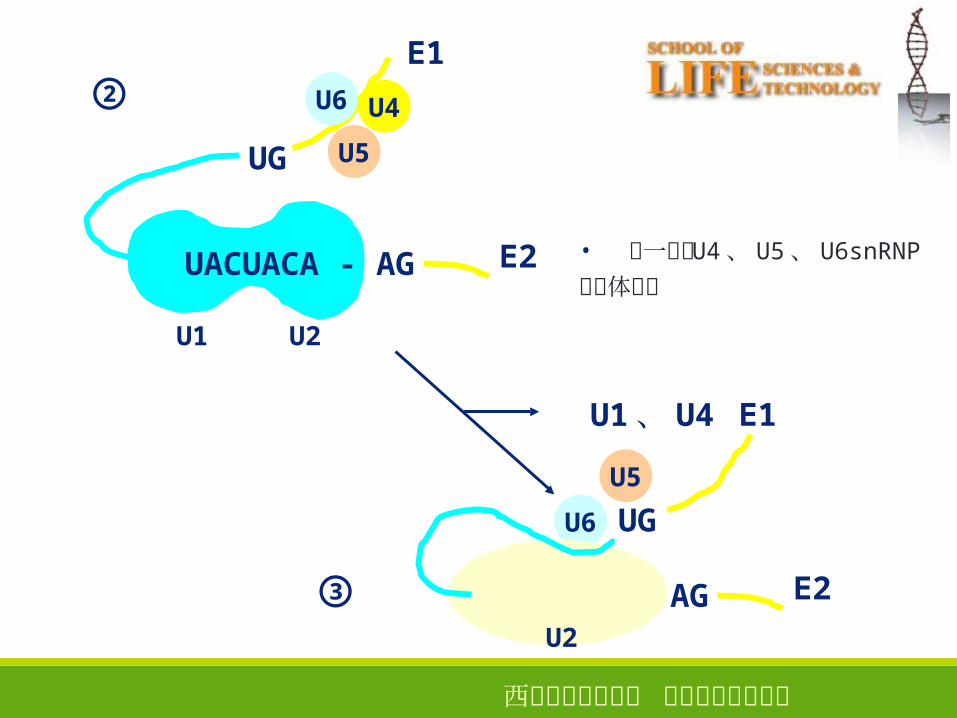
西安电子科技大学 生命科学技术学院
②
③
UACUACA - AG
UG
U4
U5
U6
E1
E2
U1 U2
UACUACA - AG
UGU6
E1
E2
U1、 U4
U5
U2
• 进一步与 U4、 U5、 U6snRNP�三聚体结合
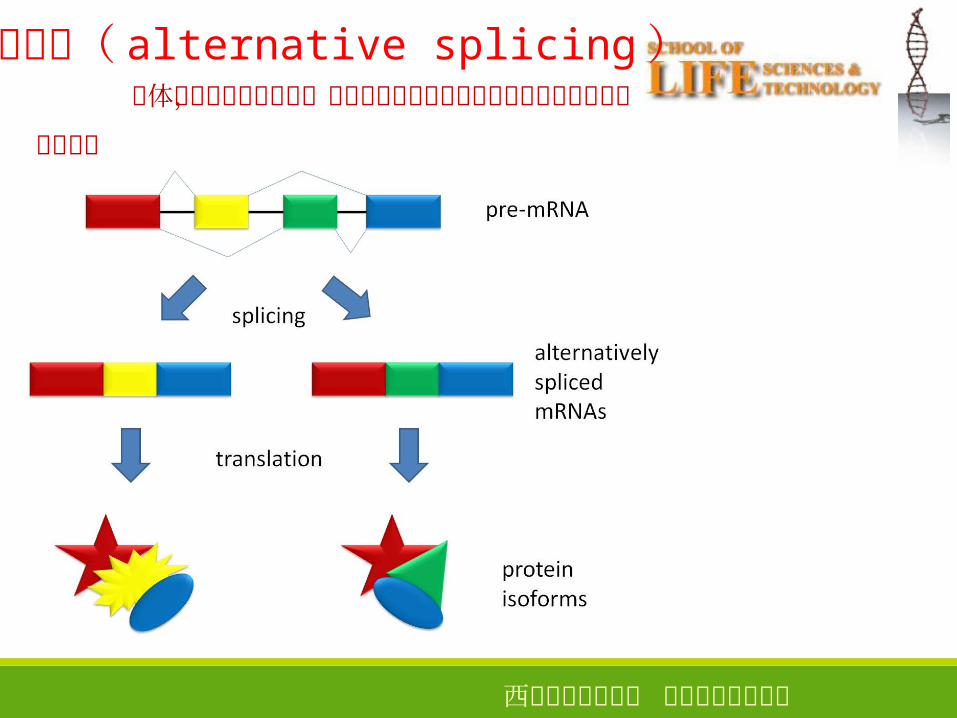
西安电子科技大学 生命科学技术学院
可变剪接( alternative splicing )个体发育或细胞分化时,选择性越过某些
外显子或某个剪接位点进行变位剪接
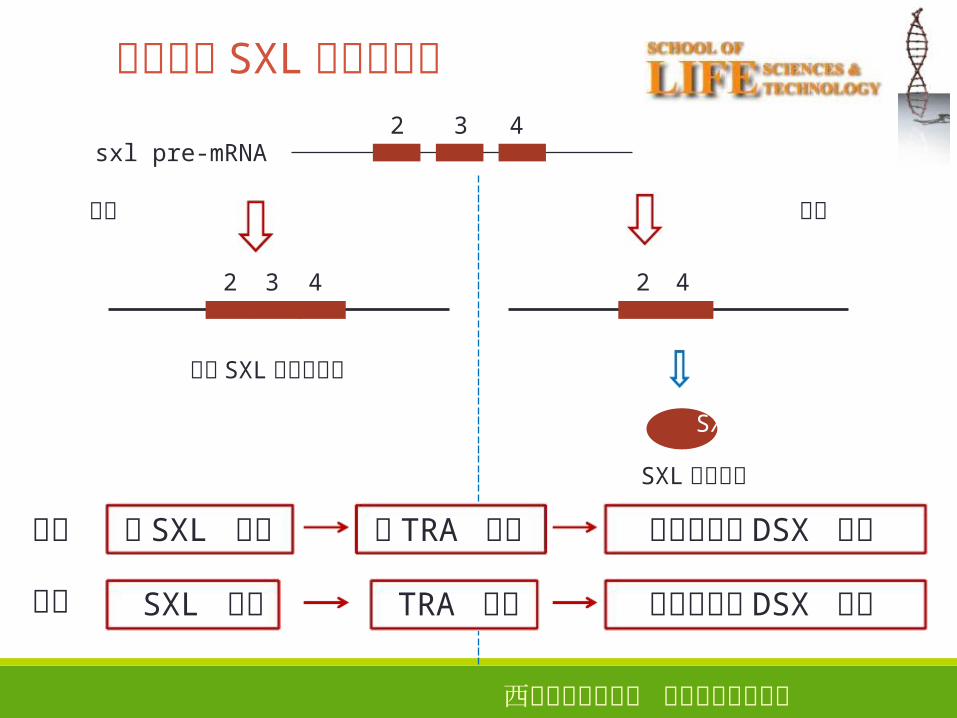
西安电子科技大学 生命科学技术学院
雌性果蝇 SXL � 蛋白的合成 32
2 3 4
2 3 4
sxl pre-mRNA
雄性 雌性
2 4
没有 SXL蛋白的合成
SXL
SXL蛋白合成
无 SXL 蛋白
SXL 蛋白
无 TRA 蛋白
TRA 蛋白
雄性特异性 DSX 蛋白
雌性特异性 DSX 蛋白
雄性
雌性
西安电子科技大学 生命科学技术学院
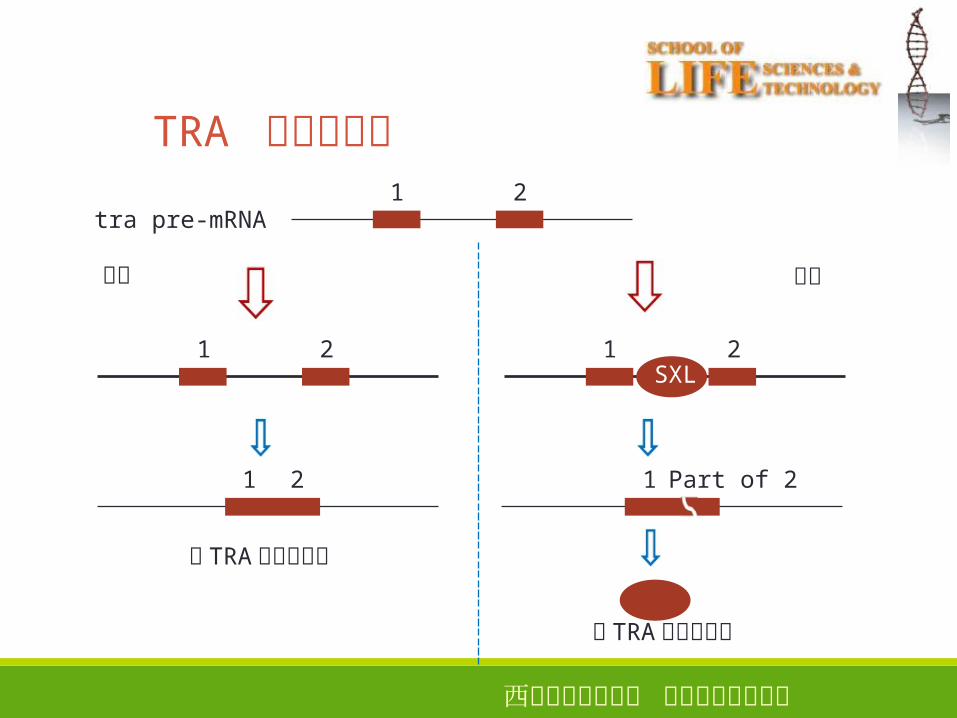
TRA 蛋白的合成
33
1 2
SXL
tra pre-mRNA
雄性 雌性
1 2
1 2
1 2
1 Part of 2
无 TRA蛋白的合成
TRA
有 TRA蛋白的合成
西安电子科技大学 生命科学技术学院
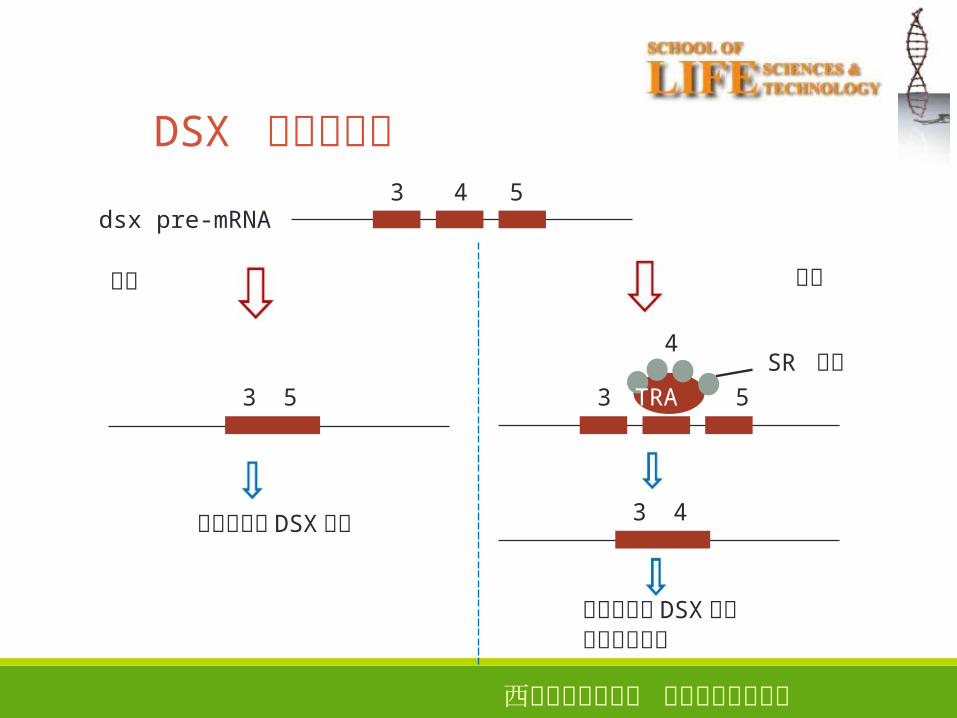
DSX 蛋白的合成
34
3 4 5
3 5
dsx pre-mRNA
雄性
雌性特异的 DSX蛋白参与性别分化
3
4
TRA 5
雌性
SR 蛋白
3 4雄性特异的 DSX蛋白
西安电子科技大学 生命科学技术学院
mRNA的剪接
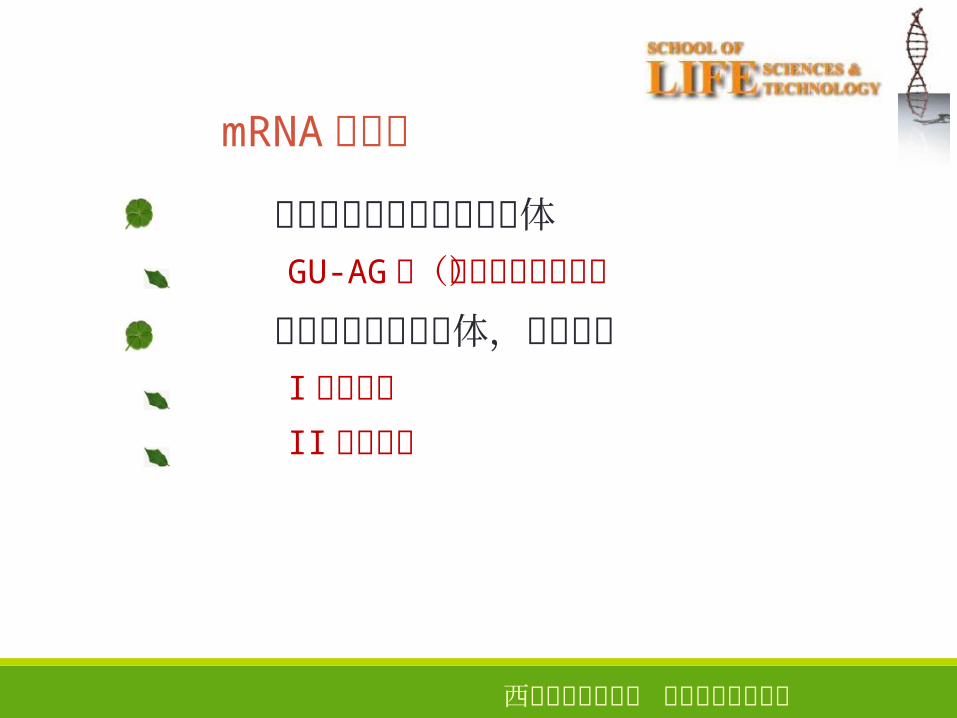
借助其它蛋白质形成剪接体GU-AG类(普通和变位剪接)
无需借助形成剪接体,自我剪接I类内含子
II类内含子
35
西安电子科技大学 生命科学技术学院
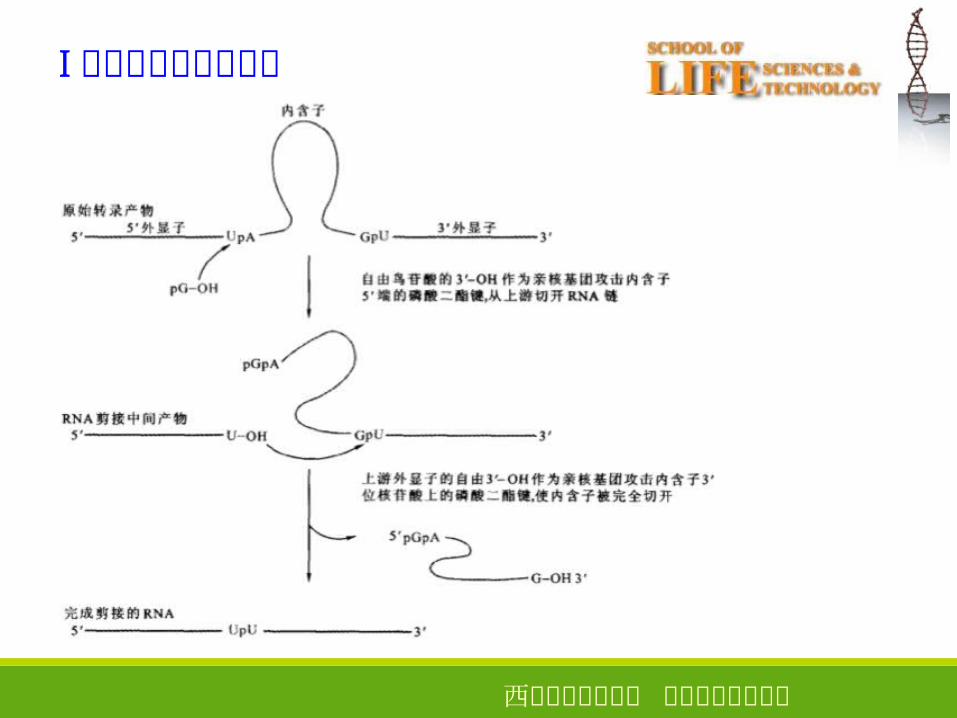
Ⅰ类内含子的自我剪接
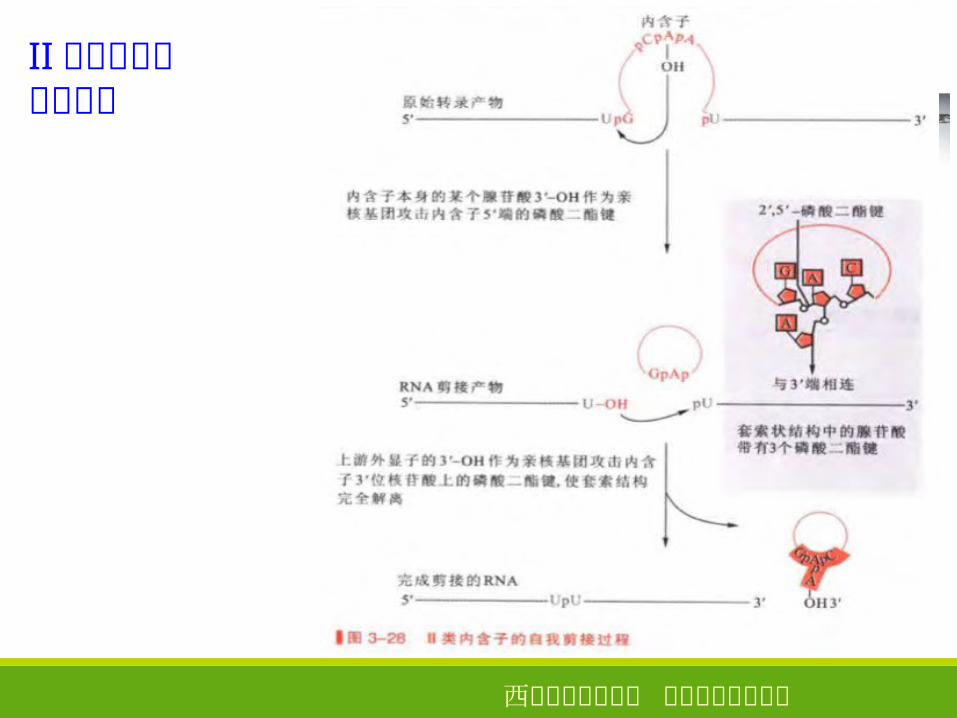
西安电子科技大学 生命科学技术学院
Ⅱ类内含子的自我剪接

西安电子科技大学 生命科学技术学院
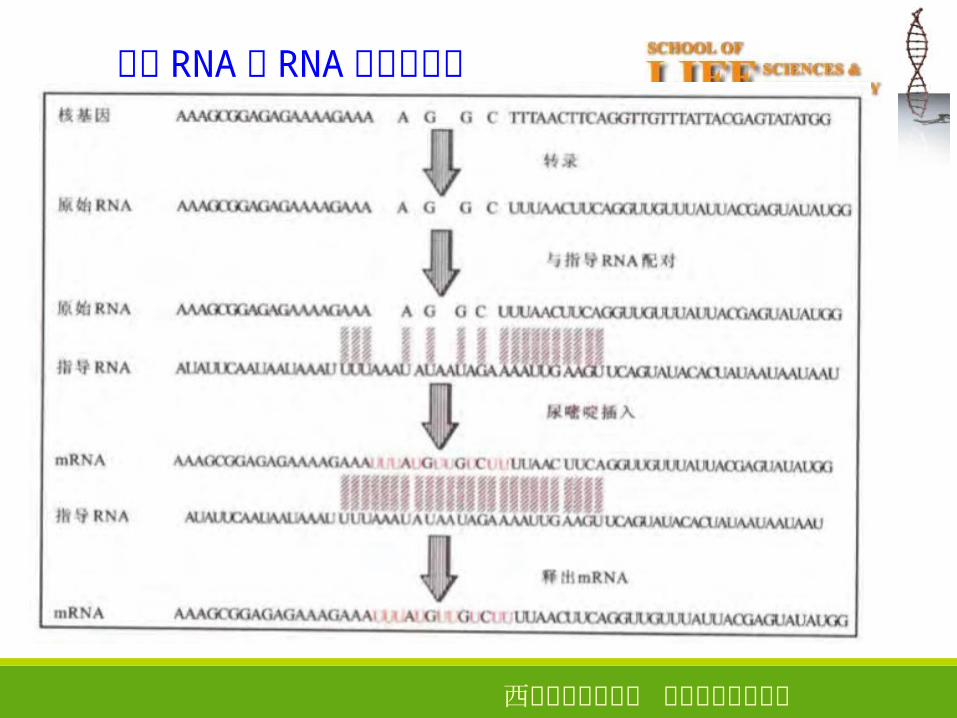
RNA 的编辑
• 编辑(editing )是指转录后的 RNA 在编码区发生碱基的突变、插入或删除
等现象。两种机制:
• 位点特异性脱氨基作用(C�→UA� → I)
• 引导 RNA 指导的尿嘧啶插入或删除
西安电子科技大学 生命科学技术学院
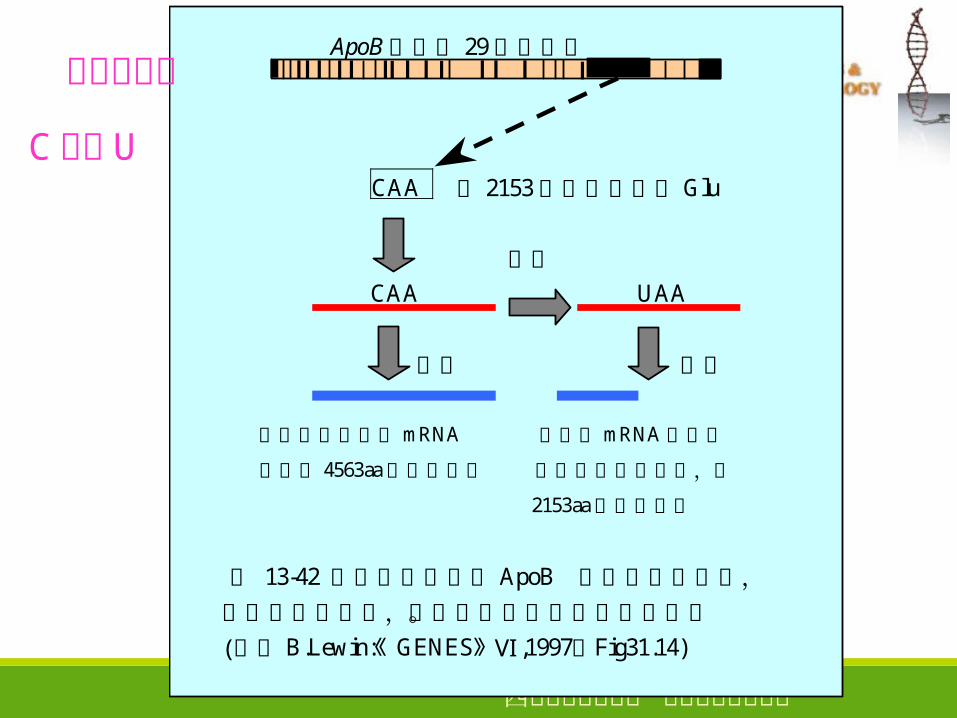
ApoB基因有 29个外显子
CAA 第 2153个密码子编码Glu
编辑 CAA UAA
翻译 翻译
在肝中剪接后的mRNA 肠中的mRNA经编辑
编码了 4563aa的载脂蛋白 产生了终止密码子,在
2153aa处终止合成
图 13-42 载脂蛋白的基因 ApoB 在肠中经过编辑, 引入终止密码子,不能翻译成完整的载脂蛋白。 (参考 B.Lewin:《GENES Ⅵ》 ,1997,Fig31.14)
C 变为 U
碱基的突变

西安电子科技大学 生命科学技术学院
尿苷酸的缺失和添加
• 1986.R.Benne 在研究锥虫线粒体 mRNA 转录加工时发现mRNA 的多个编码位置上加入或丢失尿苷酸, 1990年在高等动物和病毒中也发现了编辑现象。
西安电子科技大学 生命科学技术学院
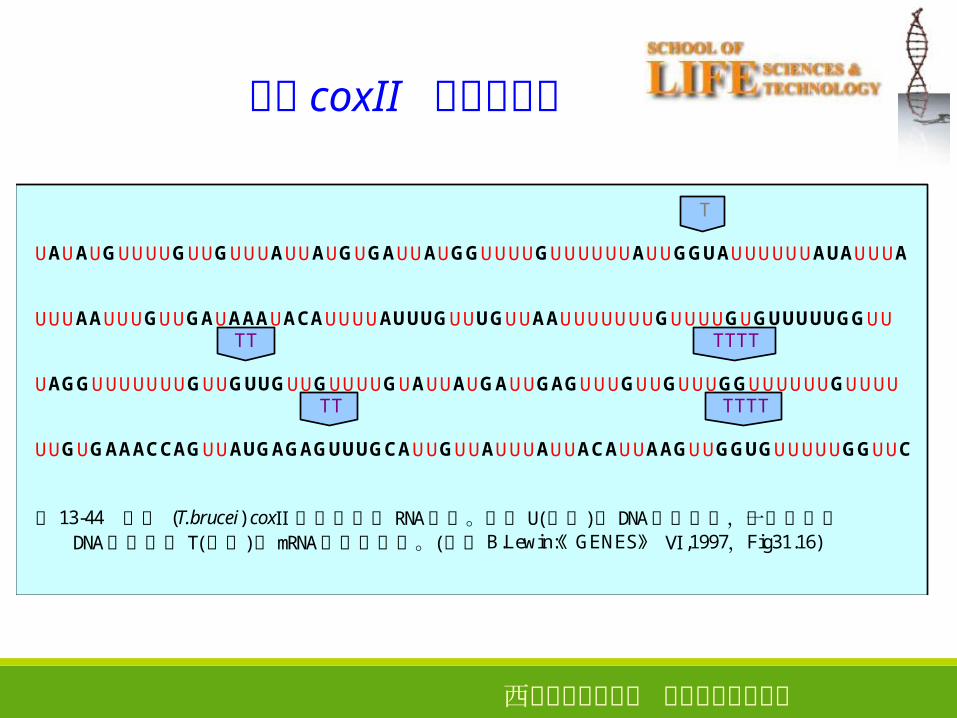
锥虫 coxII 基因的编辑
T
UAUAUGUUUUGUUGUUUAUUAUGUGAUUAUGGUUUUGUUUUUUAUUGGUAUUUUUUAUAUUUA
UUUAAUUUGUUGAUAAAUACAUUUUAUUUGUUUGUUAAUUUUUUUGUUUUGUGUUUUUGGUU TT TTTT
UAGGUUUUUUUGUUGUUGUUGUUUUGUAUUAUGAUUGAGUUUGUUGUUUGGUUUUUUGUUUU TT TTTT
UUGUGAAACCAGUUAUGAGAGUUUGCAUUGUUAUUUAUUACAUUAAGUUGGUGUUUUUGGUUC
图 13-44 锥虫 (T.brucei) coxⅡ 基因的部分 RNA顺序。很多 U(红色)在 DNA中未编码,而另一些在 DNA中编码的 T(紫色)在mRNA中被删除了。(参考 B.Lewin:《GENES》 Ⅵ ,1997,Fig31.16)
西安电子科技大学 生命科学技术学院
指导 RNA和 RNA 的编辑机制

西安电子科技大学 生命科学技术学院
RNA 编辑的功能
• 校正作用
• 调控翻译
• 扩充遗传信息
西安电子科技大学 生命科学技术学院
RNA的再编码
RNA的再编码是指 RNA编码和读码方式的改变
表现方式
+1/-1 � 移位
核糖体跳跃 甚至可能跳过 50个核苷酸
终止子通读 硒代半胱氨酸和吡咯赖氨酸
意义产生许多相互关联却不同的蛋白质
43
西安电子科技大学 生命科学技术学院
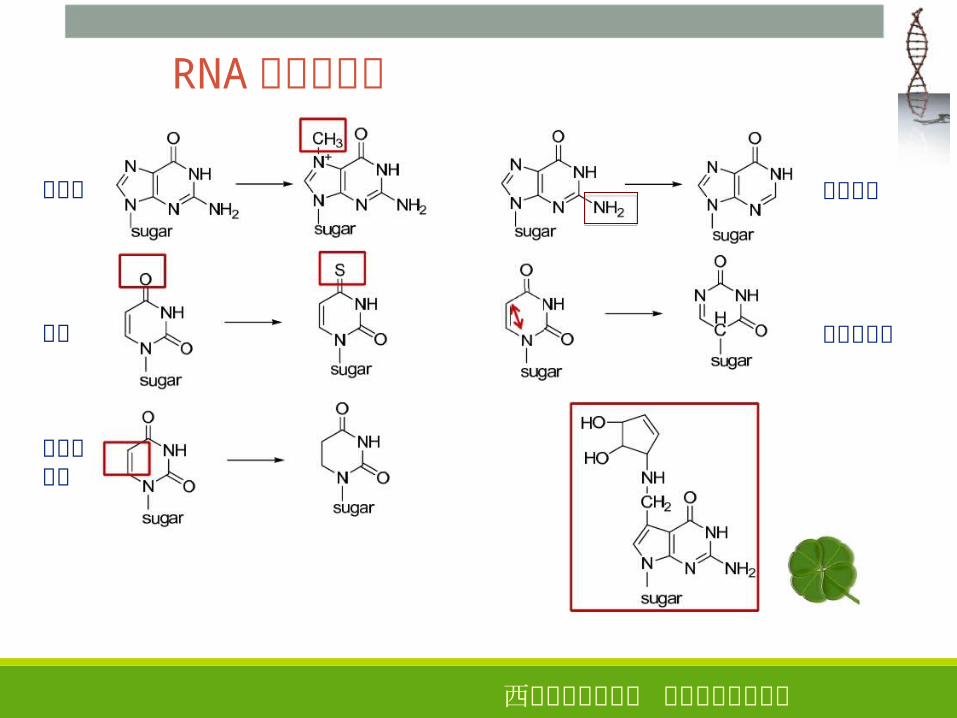
RNA� 的化学修饰
甲基化
硫代
二价键饱和
去氨基化
同分异构化
西安电子科技大学 生命科学技术学院
UAA14
A13
G12
A9
G8
N7
A6
G5
U4
C3
H
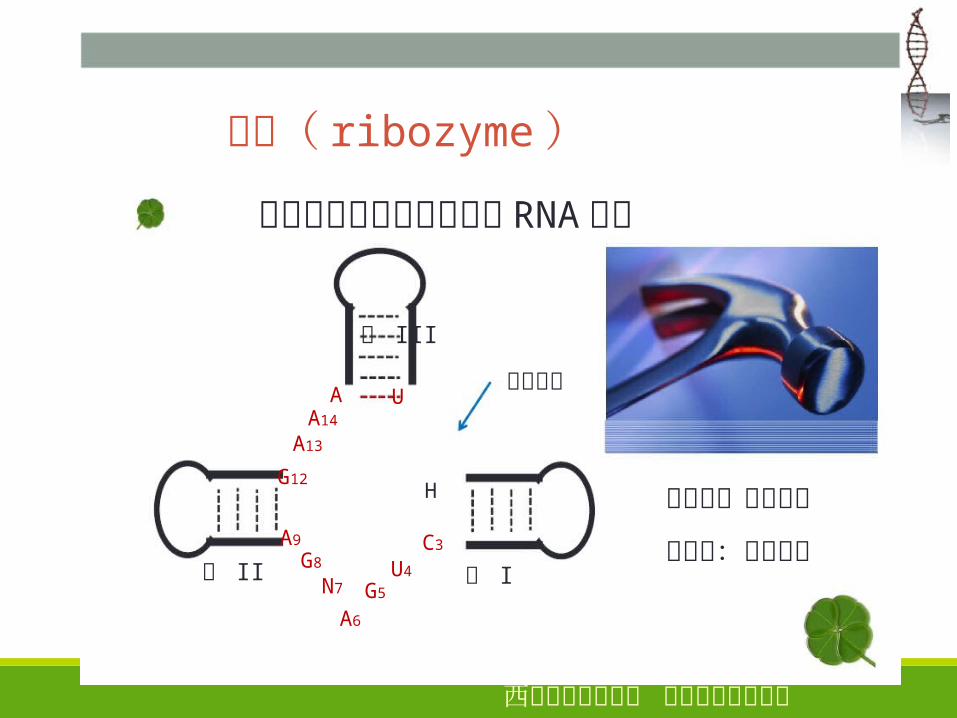
� 核酶( ribozyme )
是指一类具有催化功能的 RNA分子
茎 III
茎 II 茎 I
切割位点
剪切型:只剪不接
剪接型:既剪又接
西安电子科技大学 生命科学技术学院
RNA� 在生物进化中的地位
RNA在基因表达过程中有一定的离散性,再反转录影响后代的基因型
RNA是获得性遗传的分子基础
46

西安电子科技大学 生命科学技术学院
• 基本概念• RNA 的结构、分类和功能• 转录的基本过程• RNA 聚合酶、启动子• 原核生物与真核生物 mRNA 的特征比较• 转录后加工• RNA 合成与 DNA 合成异同点
Contents

西安电子科技大学 生命科学技术学院
RNA 合成与 DNA 合成异同点
相同点:
1 、都以 DNA 链作为模板
2 、合成的方向均为 5’→3’
3 、聚合反应均是通过核苷酸之间形成的 3’,5’-磷酸二酯键,使核苷酸链延长。
西安电子科技大学 生命科学技术学院
不同点:
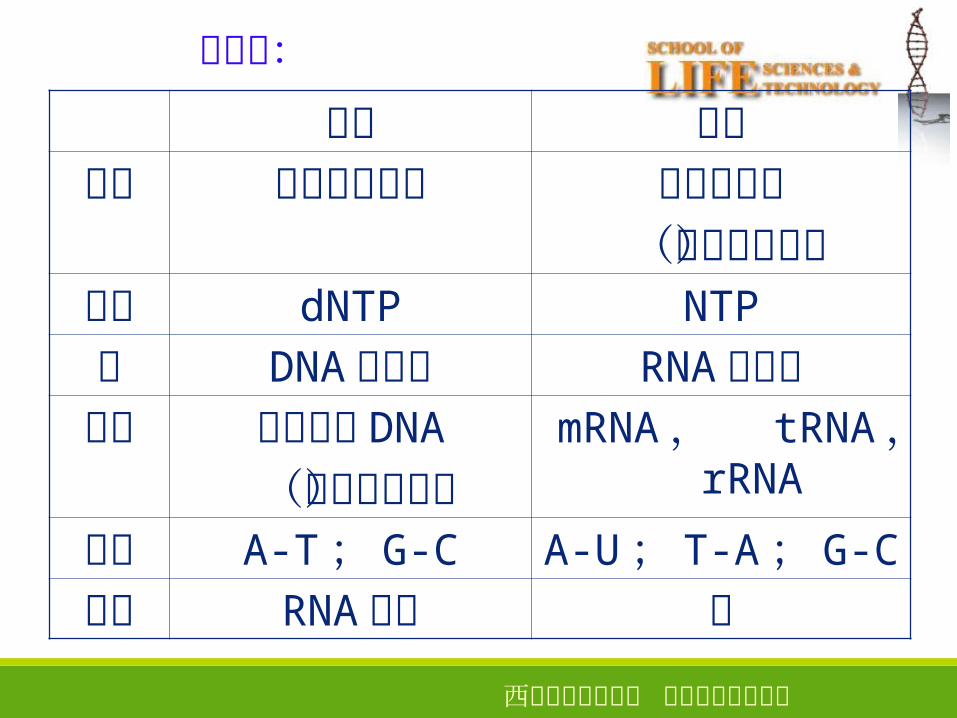
复制 转录模板 两条链均复制 模板链转录
(不对称转录)原料 dNTP NTP
酶 DNA 聚合酶 RNA 聚合酶产物 子代双链 DNA
(半保留复制)mRNA , tRNA ,
rRNA
配对 A-T; G-C A-U; T-A; G-C
引物 RNA 引物 无
西安电子科技大学 生命科学技术学院
47
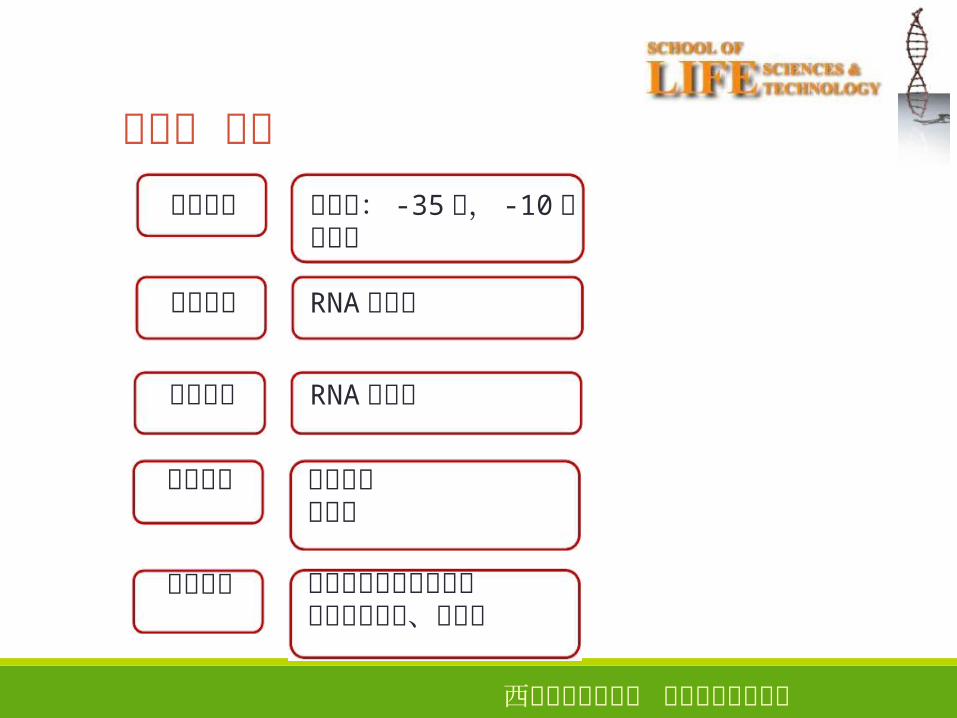
第三章 小结
模板识别
转录起始
转录延伸
转录终止
后续加工
启动子: -35区, -10区增强子
RNA聚合酶
RNA聚合酶
终止方式抗终止
真核生物的加帽和加尾内含子的剪接、编辑等