Embed Size (px)
Citation preview

1.1 Inleiding
Groei en differentiatie zijn essentiële eigenschappen van levendeorganismen. Normale groei is gereguleerd. In het volwassen orga-nisme wordt onder fysiologische omstandigheden in de meesteweefsels geen groei meer waargenomen: de aanmaak van nieuwecellen is in evenwicht met het celverlies. Op celniveau gaat degroei in de meeste weefsels echter levenslang door: voortdurendworden oude cellen vervangen door nieuw aangemaakte. In som-mige weefsels, zoals beenmerg of darmslijmvlies, is de celgroeizeer actief. Deze weefsels worden gekenmerkt door de aanwezig-heid van stamcellen die ongedifferentieerd zijn en proliferatieveeigenschappen behouden. In andere, zoals hersen- of spierweef-sel, is celgroei (nagenoeg) afwezig. In weer andere weefsels kanonder bepaalde omstandigheden, zoals na weefselverlies (par-tiële leverresectie is hiervan een bekend voorbeeld), echter decelgroei worden geactiveerd. Deze weefsels hebben zogenaam-de facultatieve stamcellen, die zich uitsluitend manifesteren bijverhoogd celverlies. Na herstel van het celtekort treedt weer eenevenwicht in en keert de celproductie tot normale waardenterug. Bij tumoren is sprake van ontregeling van de celgroei en-differentiatie. Gezwellen zijn wat hun groei betreft in meer ofmindere mate autonoom en reageren niet adequaat op de groei-regulerende mechanismen waaraan de overige weefsels van dedrager van de tumor onderworpen zijn. Deze ontregeling van hetgroeiproces vinden we zowel bij goedaardige tumoren, hoewelin mindere mate, als bij kwaadaardige tumoren. Kanker is nietéén ziekte, maar moet beschouwd worden als een verzamel-naam voor een grote verscheidenheid aan ziekten, die perorgaan sterk verschillen van origine, celtype, biologisch gedrag,behandeling en prognose.
1.2 Kenmerken van gezwelgroei
1.2.1 Goedaardige en kwaadaardige gezwellen
Bij kanker denkt men meestal aan een tastbaar gezwel. Het isbelangrijk te beseffen dat sommige vormen van kanker zich nietals een tastbaar gezwel voordoen. Bloedkanker is daarvan hetmeest kenmerkende voorbeeld: bij leukemie worden meestalgeen tumorhaarden gevonden. Omgekeerd zijn niet alle tastbaregezwellen te beschouwen als kanker. Er zijn reactieve celwoe-keringen die zich als kanker kunnen voordoen, zoals inflamma-toire pseudo-tumoren en reactieve lymfekliervergrotingen. Erzijn goedaardige en kwaadaardige gezwellen, waarvan alleen de
laatste categorie met de term kanker wordt aangeduid. Tussengoedaardige en kwaadaardige gezwellen bestaan vrij karakteris-tieke verschillen, die het mogelijk maken ze goed van elkaar teonderscheiden. Deze verschillen worden samengevat in tabel1.1. Van deze kenmerken is histologisch infiltratieve groei hetmeest betrouwbaar voor een diagnose maligniteit. Van infiltra-tieve groei spreekt men als de tumorcellen penetreren buiten hetweefselcompartiment waarin ze zijn ontstaan. Epithelialetumorcellen in een coloncarcinoom, bijvoorbeeld, kunnen infil-treren in de submucosa en de muscularis mucosae van de darm-wand en zelfs de serosa bereiken. Infiltratieve groei verleent aantumorcellen het vermogen tot uitzaaien of metastaseren. Nietalle infiltratief groeiende tumoren metastaseren echter. Bekendeuitzonderingen zijn het basalecellencarcinoom van de huid ende gliomen in de hersenen. Soms is het moeilijk te differentië-ren tussen goedaardige en kwaadaardige laesies. Bij sommigeorganen wordt daarom een tussengroep onderscheiden die metde naam ‘borderline laesie’ wordt aangeduid. Men bedenke zichhierbij dat bij het ontstaan van sommige vormen van kanker ereen geleidelijke overgang is van een goedaardig voorstadiumnaar een kwaadaardig gezwel en dat in die situatie een scherponderscheid niet gemakkelijk kan worden gemaakt.
1.2.2 Architectuur van gezwellen
Gezwellen groeien niet als een ongestructureerde massa cellen,maar tonen vaak een karakteristieke microscopische architec-tuur die het mogelijk maakt de tumor te classificeren en daar-mee een voorspelling te doen over het te verwachten biologischegedrag. Belangrijk is te beseffen dat gezwellen niet alleen uittumorcellen bestaan, maar ook uit gastheercellen, zoals bij epi-theliale tumoren (carcinomen) het met de tumorcellen meegroei-
3
1 Fundamentele aspecten vankanker
F.T. Bosman, J.H.J.M. van Krieken
Tabel 1.1 Verschillen tussen goedaardige en kwaadaardige tumoren
kenmerk goedaardig kwaadaardig
begrenzing scherp onscherp, onregelmatig
(pseudo-)kapsel frequent zelden
groeiwijze expansief infiltratief
groeisnelheid laag hoog
necrose zelden frequent
differentiatie hoog matig tot slecht
cel/kernatypie gering sterk
mitotische activiteit gering hoog
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 3
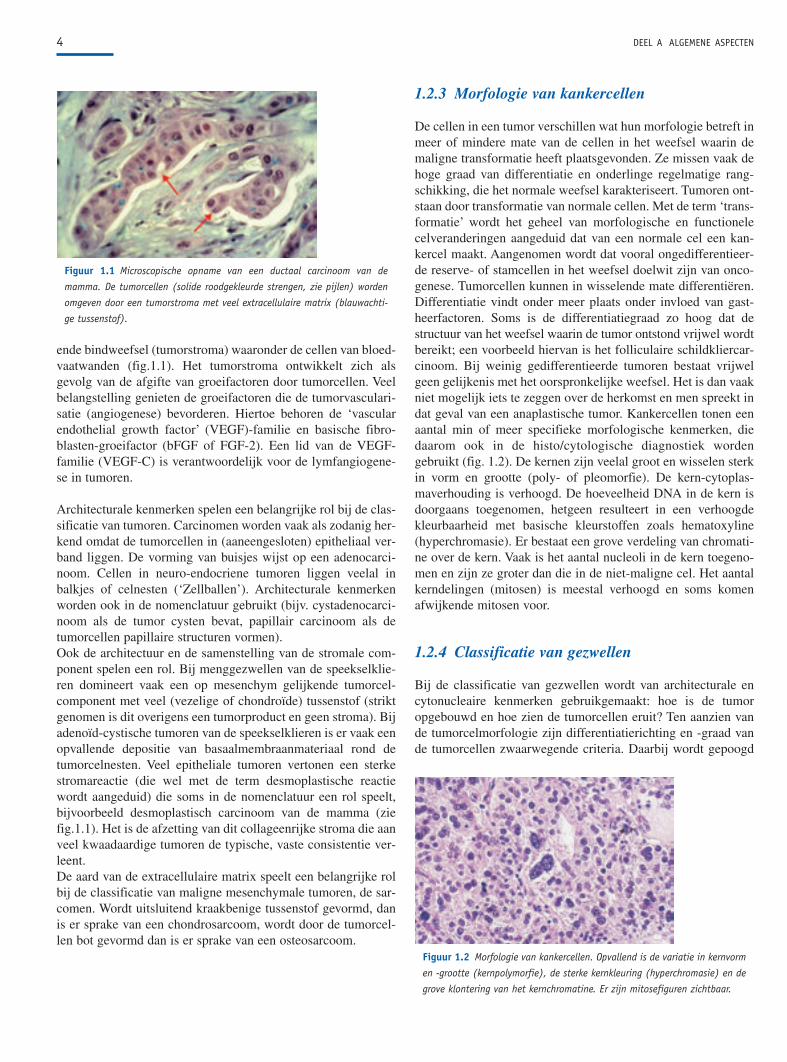
ende bindweefsel (tumorstroma) waaronder de cellen van bloed-vaatwanden (fig.1.1). Het tumorstroma ontwikkelt zich alsgevolg van de afgifte van groeifactoren door tumorcellen. Veelbelangstelling genieten de groeifactoren die de tumorvasculari-satie (angiogenese) bevorderen. Hiertoe behoren de ‘vascularendothelial growth factor’ (VEGF)-familie en basische fibro-blasten-groeifactor (bFGF of FGF-2). Een lid van de VEGF-familie (VEGF-C) is verantwoordelijk voor de lymfangiogene-se in tumoren.
Architecturale kenmerken spelen een belangrijke rol bij de clas-sificatie van tumoren. Carcinomen worden vaak als zodanig her-kend omdat de tumorcellen in (aaneengesloten) epitheliaal ver-band liggen. De vorming van buisjes wijst op een adenocarci-noom. Cellen in neuro-endocriene tumoren liggen veelal inbalkjes of celnesten (‘Zellballen’). Architecturale kenmerkenworden ook in de nomenclatuur gebruikt (bijv. cystadenocarci-noom als de tumor cysten bevat, papillair carcinoom als detumorcellen papillaire structuren vormen).Ook de architectuur en de samenstelling van de stromale com-ponent spelen een rol. Bij menggezwellen van de speekselklie-ren domineert vaak een op mesenchym gelijkende tumorcel-component met veel (vezelige of chondroïde) tussenstof (striktgenomen is dit overigens een tumorproduct en geen stroma). Bijadenoïd-cystische tumoren van de speekselklieren is er vaak eenopvallende depositie van basaalmembraanmateriaal rond detumorcelnesten. Veel epitheliale tumoren vertonen een sterkestromareactie (die wel met de term desmoplastische reactiewordt aangeduid) die soms in de nomenclatuur een rol speelt,bijvoorbeeld desmoplastisch carcinoom van de mamma (ziefig.1.1). Het is de afzetting van dit collageenrijke stroma die aanveel kwaadaardige tumoren de typische, vaste consistentie ver-leent.De aard van de extracellulaire matrix speelt een belangrijke rolbij de classificatie van maligne mesenchymale tumoren, de sar-comen. Wordt uitsluitend kraakbenige tussenstof gevormd, danis er sprake van een chondrosarcoom, wordt door de tumorcel-len bot gevormd dan is er sprake van een osteosarcoom.
1.2.3 Morfologie van kankercellen
De cellen in een tumor verschillen wat hun morfologie betreft inmeer of mindere mate van de cellen in het weefsel waarin demaligne transformatie heeft plaatsgevonden. Ze missen vaak dehoge graad van differentiatie en onderlinge regelmatige rang-schikking, die het normale weefsel karakteriseert. Tumoren ont-staan door transformatie van normale cellen. Met de term ‘trans-formatie’ wordt het geheel van morfologische en functionelecelveranderingen aangeduid dat van een normale cel een kan-kercel maakt. Aangenomen wordt dat vooral ongedifferentieer-de reserve- of stamcellen in het weefsel doelwit zijn van onco-genese. Tumorcellen kunnen in wisselende mate differentiëren.Differentiatie vindt onder meer plaats onder invloed van gast-heerfactoren. Soms is de differentiatiegraad zo hoog dat destructuur van het weefsel waarin de tumor ontstond vrijwel wordtbereikt; een voorbeeld hiervan is het folliculaire schildkliercar-cinoom. Bij weinig gedifferentieerde tumoren bestaat vrijwelgeen gelijkenis met het oorspronkelijke weefsel. Het is dan vaakniet mogelijk iets te zeggen over de herkomst en men spreekt indat geval van een anaplastische tumor. Kankercellen tonen eenaantal min of meer specifieke morfologische kenmerken, diedaarom ook in de histo/cytologische diagnostiek wordengebruikt (fig. 1.2). De kernen zijn veelal groot en wisselen sterkin vorm en grootte (poly- of pleomorfie). De kern-cytoplas-maverhouding is verhoogd. De hoeveelheid DNA in de kern isdoorgaans toegenomen, hetgeen resulteert in een verhoogdekleurbaarheid met basische kleurstoffen zoals hematoxyline(hyperchromasie). Er bestaat een grove verdeling van chromati-ne over de kern. Vaak is het aantal nucleoli in de kern toegeno-men en zijn ze groter dan die in de niet-maligne cel. Het aantalkerndelingen (mitosen) is meestal verhoogd en soms komenafwijkende mitosen voor.
1.2.4 Classificatie van gezwellen
Bij de classificatie van gezwellen wordt van architecturale encytonucleaire kenmerken gebruikgemaakt: hoe is de tumoropgebouwd en hoe zien de tumorcellen eruit? Ten aanzien vande tumorcelmorfologie zijn differentiatierichting en -graad vande tumorcellen zwaarwegende criteria. Daarbij wordt gepoogd
4 DEEL A ALGEMENE ASPECTEN
Figuur 1.1 Microscopische opname van een ductaal carcinoom van de
mamma. De tumorcellen (solide roodgekleurde strengen, zie pijlen) worden
omgeven door een tumorstroma met veel extracellulaire matrix (blauwachti-
ge tussenstof).
Figuur 1.2 Morfologie van kankercellen. Opvallend is de variatie in kernvorm
en -grootte (kernpolymorfie), de sterke kernkleuring (hyperchromasie) en de
grove klontering van het kernchromatine. Er zijn mitosefiguren zichtbaar.
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 4

de tumorcelkenmerken te vergelijken met die van normale cel-len. Een eerste vraag is of de tumor goed- of kwaadaardig is. Dehierbij gebruikte morfologische criteria zijn in de vorige para-graaf behandeld (zie tabel 1.1). Soms tonen de tumorcellen indi-vidueel kenmerken die bij maligniteit worden gezien, zoalspolymorfie en hyperchromasie, maar nog geen infiltrerendegroei. Dan spreekt men van carcinoma in situ. Soms laat het his-tologisch onderzoek geen uitspraak toe over het te verwachtengedrag van de tumor. Men spreekt dan wel van een ‘border-line’-tumor. Er is een neiging om de generieke term ‘tumor’ tehanteren voor die gezwellen waarover op grond van het histolo-gisch onderzoek geen gefundeerde uitspraak kan wordengedaan ten aanzien van het te verwachten gedrag. Voorbeeldenzijn gastro-intestinale stromale tumor (GIST, zie hoofdstuk 18)of neuro-endocriene tumor (zie hoofdstuk 28).Een tweede vraag is van welk type tumor er sprake is. Daarbijwordt gelet op het celtype en op de onderlinge samenhang vande tumorcellen. Zijn de tumorcellen epitheliaal, dan wordt eengoedaardige vorm papilloom of adenoom genoemd en eenkwaadaardige vorm carcinoom. Een kwaadaardig bindweefsel-gezwel wordt sarcoom genoemd. Op grond van individuele cel-kenmerken kunnen deze categorieën verder worden onderver-deeld. Een carcinoom met plaveiselcellige differentiatie wordtplaveiselcelcarcinoom genoemd en een carcinoom met klier-buisdifferentiatie een adenocarcinoom. Naar de mate van diffe-rentiatie kan een verdere verdeling plaatsvinden (goed, matig,slecht gedifferentieerd).Sarcomen zijn onder te verdelen naar het celtype dat de tumorvormt, bijvoorbeeld vetcellen (liposarcoom), endotheel (angio-sarcoom), gladde spiercellen (leiomyosarcoom), enzovoort.Maligne lymfomen kunnen worden onderverdeeld naar B- of
T-cel-origine. Een samenvatting van de meest voorkomendetumortypen wordt gegeven in tabel 1.2. Sommige tumoren ver-tonen zowel epitheliale als mesenchymale kenmerken (mennoemt dit wel bifasisch). Hiervan zijn het synoviosarcoom enhet mesothelioom voorbeelden.
1.3 Klinische verschijnselen vangezwelgroei
Tumoren kunnen zich op heel verschillende manieren voordoen.Een overzicht van de belangrijkste symptomen wordt gegevenin tabel 1.3. De symptomen kunnen worden onderscheiden inlokale en systemische. Belangrijk is op te merken dat bij kwaad-aardige tumoren de metastasen zelf ook weer dergelijke symp-tomen veroorzaken. Bij goedaardige tumoren blijven de gevol-gen meestal beperkt tot compressie van omgevend weefsel. Ditkan belangrijke symptomen veroorzaken, zoals intermenstrueelbloedverlies door compressie van vaten bij een leiomyoma uteriof gezichtsvelduitval door compressie van de oogzenuwen bijeen hypofyseadenoom. Van goedaardige (d.w.z. niet infiltratief
1 FUNDAMENTELE ASPECTEN VAN KANKER 5
Tabel 1.2 Classificatie van gezwellen
differentiatierichting benigne maligne voorbeeld
epitheliaal
klierweefsel adenoom adenocarcinoom adenocarcinoom van de long
adenocarcinoom van de maag
plaveiselcel papilloom plaveiselcelcarcinoom plaveiselcelcarcinoom van de huid
overgangscel papilloom overgangscelcarcinoom overgangscelcarcinoom van de blaas
mesenchymaal
vetcel lipoom liposarcoom
fibroblast fibroom fibrosarcoom
skeletspiercel rabdomyoom rabdomyosarcoom
endotheelcel hemangioom hemangiosarcoom
bloedcel leukemie myeloïde leukemie
lymfatische leukemie
lymfocyt maligne lymfoom ziekte van Hodgkin
non-Hodgkin-lymfoom
(B-cellymfoom, T-cellymfoom)
neuro-ectodermaal
pigmentcel naevus melanoom
zenuwcel ganglioneuroom neuroblastoom
gliacel glioom glioblastoom astrocytoom
Tabel 1.3 Symptomen van gezwelgroei
lokaal zwelling
obstructie (van een bloedvat of hol orgaan)
ulceratie (van huid of slijmvlies)
bloedverlies (manifest of occult)
systemisch gewichtsverlies
paraneoplastische verschijnselen
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 5

groeiende en derhalve niet metastaserende) tumoren kan delokalisatie soms een ernstig probleem vormen bij de behande-ling (bijv. bij sommige meningiomen) waardoor de prognoseondanks het goedaardige karakter ongunstig kan zijn.
De lokale effecten van kwaadaardige gezwellen zijn meestal hetgevolg van ingroei in omliggende structuren of organen. Eenhuidcarcinoom of een carcinoom in de tractus digestivus zalzich vaak presenteren als een ulcus door necrose in de tumor.Uit een ulcus kan bloedverlies optreden. Ingroei in organen kanleiden tot pijn (bijv. botpijn bij primaire of metastatische tumo-ren in het skelet) of tot functieverlies (bijv. icterus bij uitgebrei-de tumorgroei in de lever).De systemische effecten van kwaadaardige gezwelgroei omvat-ten gewichtsverlies en de zogenaamde paraneoplastische syn-dromen. Deze laatste worden niet veroorzaakt door lokale effec-ten van tumorcelgroei maar door de afgifte van actieve stoffenaan de circulatie door de tumorcellen. Een typisch voorbeelddaarvan is hormoonproductie door tumorcellen. Op deze syn-dromen wordt in hoofdstuk 4 nader ingegaan. Het gewichtsver-lies kan veroorzaakt worden door lokale effecten van een tumor,bijvoorbeeld afsluiting van de oesophagus. Gewichtsverlies kanook het gevolg zijn van ineffectieve stofwisseling, bijvoorbeelddoor de productie in de tumor van stoffen die leiden tot een ver-hoogd katabolisme. Een voorbeeld van een dergelijke stof istumor necrosis factor (TNF-α).
1.4 Tumorcelgroei
Bij een volwassen persoon bestaat onder normale omstandighe-den in de meeste weefsels een evenwicht tussen celaanmaak encelverlies. Celverlies vindt vooral plaats door geprogrammeerdeceldood, ook wel apoptose genoemd. Celaanmaak vindt plaatsdoor celdeling (mitose). Bij weefsels zoals beenmerg, darm-slijmvlies en epidermis is er constant celverlies en derhalve ookconstante celaanmaak uit het compartiment ongedifferentieerdestamcellen. Sommige hematopoëtische stamcellen zijn pluripo-tent: ze zijn in staat te differentiëren in allerlei verschillenderichtingen (bijv. zenuwcellen en hartspiercellen). Sommige cel-len in het lichaam zijn zo ver gedifferentieerd dat ze niet meerkunnen delen (postmitotische cellen). Voorbeelden van postmi-totische cellen zijn zenuwcellen en spiercellen. Dergelijke cel-len kunnen niet regenereren. Andere cellen delen regelmatig enzijn dan of tussen twee delingen in (intermitotisch, in deG0-fase van de celcyclus) of in deling (in de celcyclus). Demeeste lichaamscellen zijn intermitotisch. Voor weer andere cel-typen geldt dat ze langdurig inactief zijn en alleen bij een ster-ke regeneratiebehoefte in deling gaan. Dat geldt bijvoorbeeldvoor levercellen.Bij tumorgroei is het evenwicht tussen celafbraak en celaan-maak gestoord; er worden meer cellen gevormd dan er te gron-de gaan. Dat kan zowel door vermeerderde celaanmaak als doorverminderde apoptose. De proliferatiesnelheid van tumorcellenis hoog, maar meestal lager dan die van normale foetale cellenen regenererende cellen. De volumetoename van een tumorblijkt bijna zonder uitzondering langzamer te verlopen dan ver-
wacht zou worden op grond van de delingsactiviteit van detumorcellen. Dat wordt verklaard door celverlies. Bij snelgroei-ende tumoren treedt veelal in het tumorcentrum door onvol-doende bloeddoorstroming weefselnecrose op. Belangrijker isvermoedelijk tumorcelverlies door apoptose. Deze fysiologi-sche vorm van celdood treedt in alle weefsels op en is normaalin evenwicht met celaanmaak. Van een apoptotische cel con-denseert de kern, wordt in een energiegebruikend specifiek pro-ces het DNA afgebroken en wordt het celrestant afgevoerd(veelal via fagocytose door macrofagen). Apoptose speelt eenbelangrijke rol in de tumorbiologie, enerzijds omdat verlies vanhet vermogen tot apoptose tot tumorgroei kan leiden en ander-zijds omdat het werkingsmechanisme van veel chemotherapeu-tica en van radiotherapie ten minste ten dele het aanschakelenvan apoptose omvat. Er zijn diverse mechanismen waarlangs intumorcellen apoptose kan worden geïnduceerd. Deze wordenschematisch samengevat in figuur 1.3.
De tijd die een tumor nodig heeft om zijn volume te verdubbelen,kan – afhankelijk van het tumortype en de omstandigheden –variëren van dagen tot jaren. Deze volumetoename is afhankelijkvan de tijd die verloopt tussen twee celdelingen, de groeifractie(het percentage cellen dat actief groeit) en het aantal cellen datin een bepaalde periode afsterft. Het tijdsverloop tussen tweeceldelingen is van groot belang. Opmerkelijk is dat de duur vande celcyclus zelf tamelijk constant is. Uit de G0-fase kan de celbij een adequate stimulus in de G1-fase terugkeren. Postmitoti-sche cellen keren niet terug naar de G1-fase. De G1-fase gaatover in de S-fase, waarin de cel DNA gaat synthetiseren ter ver-dubbeling van het genetische materiaal als voorbereiding op dedeling. Deze overgang is van groot belang omdat cellen als hetware een drempel over moeten om in de S-fase te raken; menspreekt wel van een restrictiepunt. Zijn de cellen hier voorbij,dan loopt de celcyclus verder geprogrammeerd af. Voordat decel zich gaat delen (mitose- of M-fase) is er een G2-fase; de kernbevat nu de dubbele hoeveelheid DNA. De minimumtijd van eencelcyclus, gemeten aan cellen in weefselkweek, bedraagt onge-veer 16 uur. In vivo is deze voor darmepitheel 12 uur, voor deepidermis 21 dagen en voor de lever 160 dagen. De duur van deS-fase en de G2-fase is vrij constant. De grote variabiliteit, ookvoor tumorcellen, ligt in de G0-fase en de G1-fase. De gediffe-rentieerde cellen met één specifieke functie (zoals zenuw- enspiercellen) blijven jarenlang of altijd in de G0-fase. Tumorcel-len vertonen soms ook deze eigenschap. Ze kunnen jarenlang inG0 in weefsels verborgen blijven (‘dormant’), om plotselingonder invloed van onbekende groeistimuli metastasen te vor-men. Late metastasen bij het mammacarcinoom zijn van dit ver-schijnsel een bekend voorbeeld. Vermeldenswaard is nog datgedurende de verschillende fasen van de celcyclus cellen eengeheel verschillende gevoeligheid hebben voor exogene invloe-den, zoals ioniserende stralen en cytostatica. Hiervan wordtgebruikgemaakt bij de ontwikkeling van doseringsschema’svoor chemo- en radiotherapie.De celcyclus en de belangrijkste regulatoren ervan zijn schema-tisch samengevat in figuur 1.4. De toegang van de cel tot de cel-cyclus (overgang van de G0- naar de G1-fase) en de voortgangvan de cel in de cyclus worden geregeld door een groep eiwit-
6 DEEL A ALGEMENE ASPECTEN
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 6

ten, cyclines genaamd. In de G1- en de S-fase spelen de cycli-nes D en E een rol. In de G2- en de S-fase is cycline A actief enin de G2- en de M-fase cycline B. De cyclines vormen een com-plex met de cycline-afhankelijke kinasen (CDK’s): voor elk vande fasen van de celcyclus is er een specifiek cycline/CDK-com-plex. Dit complex activeert door fosforylering transcriptiefacto-ren, die op hun beurt de voor de voortgang van de cel door decelcyclus essentiële genen activeren. De cycline/CDK-com-plexen worden geremd door CDK-remmers, zoals de eiwittenp21 en p27. Het evenwicht tussen het cycline/CDK-complex ende CDK-remmer bepaalt of de cel verdergaat in de celcyclus Inde M-fase spelen eiwitten een rol die de opbouw van de kern-spoel regelen en de condensatie van de chromosomen. Na decelcyclus worden al deze eiwitten verwijderd door binding aanhet eiwit ubiquitine, waarna ze in het proteasoom van de celworden afgebroken. Het belang van deze celcyclusregulatorenin de oncologie is groot: bij veel vormen van kanker is het even-wicht verstoord waardoor de cellen ongehinderd blijven proli-fereren. Veel nieuwe vormen van medicamenteuze kankerthera-pie zijn dan ook gericht op celcyclus regulerende eiwitten.
Omdat in de S-fase de cellen een verhoogd DNA-gehalte heb-ben (tussen de diploïde en tetraploïde hoeveelheid) kan doormeting van de hoeveelheid DNA via statische of flow-cytome-trie in de kern het aantal cellen in de S-fase van de celcyclusworden bepaald. Dit is een maat voor de delingsactiviteit in detumor. De prolifererende celfractie kan in weefselcoupes wor-den aangetoond met behulp van immunohistochemische tech-nieken. Daarbij wordt gebruikgemaakt van antilichamen dieantigenen herkennen die alleen in de kern van delende cellen tot
expressie komen. Bekend is vooral het Ki-67-antigeen, dat meteen monoklonaal antilichaam (MIB-1) ook in routinematigbewerkt (in formaline gefixeerd en in paraffine ingebed) weef-sel kan worden aangetoond (fig. 1.5). Het kunnen bepalen vande groeifractie in een tumor is van belang omdat deze prognos-
1 FUNDAMENTELE ASPECTEN VAN KANKER 7
p53-activatie
DNA-schade
DNA-fragmentatie
bestraling, vrije radicalen
activatoren
Bcl 2
Bxl x
andere
inhibitoren
Bax
Bad
andere
groeifactordepletie
interactie ligand-receptor bijv.TNFR-TNF, FasL-Fas
cytotoxischelymfocyten
adaptermoleculen vroege caspases
granzyme
afbraak celskelet
uitvoerende caspases
celkern
mito-chondrion
perforine
Figuur 1.3 Schematische weergave
van de mechanismen die apoptose
activeren in de cel, de intracellulaire
processen die apoptose inleiden en de
uiteindelijke afloop van het apoptose-
proces. Gebrek aan groeifactoren heeft
een effect op mitochondriaal niveau,
waardoor apoptoseactivatoren worden
vrijgemaakt. Apoptoseremmers bevin-
den zich eveneens op mitochondriaal
niveau. Receptoractivatie (met name
van de TNFR-familie) leidt via interme-
diaire stappen tot caspaseactivatie.
Granzyme-injectie in de cel door cyto-
toxische T-cellen activeert eveneens
caspases. Ook activatie van p53 kan
caspase activeren. In de effectorfase
van apoptose speelt afbraak van DNA
en celskeleteiwitten een rol.
nieuwe celin de cyclus
einddifferentiatiegeen verdere deling
G0
S
G1
M
G2
tuss
enfa
seceld
elin
g
Figuur 1.4 Schematische voorstelling van de celcyclus. De M(itose)-fase
omvat mitose en de feitelijke celdeling. De G(ap)1-fase gaat vooraf aan de
S(ynthese)-fase waarin de cel het DNA repliceert. Tussen S en M is er een G2-
fase. Na de M-fase kan een nieuwe celcyclus worden doorlopen; ook kunnen
cellen wat betreft deling in rust gaan (G0).
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 7

tische betekenis heeft: tumoren met een hoge S-fasefractie (zeeractief delend) hebben meestal een slechtere prognose dan tumo-ren met een lage S-fasefractie. Ze reageren daarentegen beter op(radio- of chemo)therapie.
De groeisnelheid van tumoren in vivo is niet eenvoudig uit degroeifractie en de celcyclusduur af te leiden omdat er belangrijkcelverlies is door necrose en/of apoptose. Van longmetastasen isde groeisnelheid onderzocht door metingen te verrichten aanopeenvolgende röntgenfoto’s. Daarbij bleek dat deze tumoreneen opvallend constante verdubbelingstijd vertonen. Men kanberekenen dat, uitgaande van één tumorcel met een gemiddeldvolume, voor het ontstaan van een tumor met een volume van 1 cm3 dertig verdubbelingen nodig zijn (fig. 1.6).
Indien in een bepaald geval de verdubbelingstijd honderd dagenbedraagt en constant blijft, vergt dit ongeveer acht jaar. Gedu-rende de volgende tien verdubbelingen zal het volume toenemen
met 1 cm3 tot 1000 cm3. Dat betekent dat tumoren vaak al jaren-lang aanwezig zijn, voordat we ze met de meest verfijnde dia-gnostische hulpmiddelen kunnen vaststellen. De hele periodevan de klinische observatie, diagnostiek en behandeling speeltzich in het algemeen af na de dertigste tumorcelverdubbeling.Screening en vroegdiagnostiek hebben hierop slechts eenbeperkte invloed.
1.5 Invasie en metastasering
Van kanker is een van de belangrijkste kenmerken het vermogentot infiltrerende groei in omliggende weefsels. Door dit vermo-gen kunnen kankercellen zich toegang verschaffen tot lymfeba-nen en versleept worden naar lymfeklieren. Ze kunnen ook dewanden van bloedvaten passeren en dan versleept worden naarandere organen. Uitgroei in lymfeklieren en organen op afstandwordt metastasering (lymfogeen resp. hematogeen) genoemd.Het vermogen tot infiltratieve groei kan ook leiden tot ingroei inaanliggende organen (uitbreiding per continuitatem) of inlichaamsholten, waar zich tevens metastasen kunnen vormen.Metastasering is klinisch het belangrijkste kenmerk van kwaad-aardige gezwelgroei, omdat de metastasen veelal onbehandel-baar zijn en in hoge mate de prognose bepalen.
1.5.1 Infiltratieve groei
Infiltratief groeiende kankercellen hebben het vermogen zichbuiten de grenzen te begeven van het weefselcompartimentwaarin ze zijn ontstaan. Het duidelijkst spreekt dat bij epithelia-le tumoren (carcinomen) waarbij de tumorcellen de epithelialebasaalmembraan kunnen passeren. Voor sommige normale cel-
8 DEEL A ALGEMENE ASPECTEN
Figuur 1.5 Ki-67-kleuring van een plaveiselcelcarcinoom van de huid (MIB-
1-antilichaam). Het merendeel van de tumorcellen toont kernkleuring, wat
een hoge delingsactiviteit van de tumorcellen impliceert.
1 mm3 celmassa
0,5 cm tumor, de onderste detectiegrensmet beeldvormende technieken
2 cm tumor, juist palpabel
tumor van 1kg
5 10 15 20 25 30 35 40
106
107
108
109
1010
aantal celdelingen >
aan
tal cellen
>
Figuur 1.6 Schematische
weergave van de groeisnel-
heid van tumoren als func-
tie van de delingsactiviteit
van tumorcellen. Op de X-as
het aantal doorgemaakte
celdelingen, op de Y-as het
daarmee bereikte tumor-
volume.
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 8

len is infiltratief vermogen fysiologisch: zonder deze eigen-schap zouden lymfocyten, macrofagen en granulocyten nietgoed kunnen functioneren. Gebleken is dat kankercellen bijinfiltratieve groei van dezelfde mechanismen gebruikmaken alsdeze normale cellen. Invasief en migrerend gedrag van cellen isals zodanig dus niet pathologisch; het is pathologisch als epi-theliale carcinoomcellen of mesenchymale sarcoomcellen datdoen. Bij infiltratie in de omgeving kunnen de volgende cellu-laire processen worden onderscheiden: losraken van de cel uithet weefselverband; proteolyse van extracellulaire matrixcom-ponenten; migratie van de infiltratieve kankercel. We zullenhierna deze processen wat meer in detail bespreken.
Losraken uit het weefselverbandEpitheliale cellen zijn in normale weefsels hecht met elkaar ver-bonden. Bij deze adhesie spelen cel-cel- en cel-matrix-adhesie-moleculen een rol. Van de celadhesiemoleculen zijn inmiddelszo’n vier verschillende families bekend. In dit kader zijn vanbelang E-cadherine (behorend tot de zgn. immunoglobulinesuperfamilie) en de integrinen.E-cadherine is een groot transmembraan glycoproteïne dathomotypische verbindingen vormt, dat wil zeggen dat de ex-tracellulaire domeinen van twee E-cadherinemoleculen eenhechte verbinding kunnen vormen. Bij het totstandkomen vancomplexe verbindingen tussen epitheelcellen (zoals desmoso-men) speelt E-cadherine een cruciale rol. Het in het cytoplasmagelegen einde van het molecuul is verbonden met het celskelet.Uit recent onderzoek is gebleken dat infiltratieve cellen verliesvan E-cadherine-expressie tonen (fig. 1.7). Sterk infiltratievetumorcellen met een totaalverlies van het vermogen tot het vor-men van een weefselverband, zoals het zegelringcelcarcinoomvan de maag of het lobulaircarcinoom van de mamma, vertonenfrequent mutaties in het E-cadherine-gen. Bij familiaire vormenvan het zegelringcelcarcinoom van de maag zijn kiembaanmu-taties in het E-cadherine-gen gevonden.
De integrinen zijn heterodimere (met een α- en een β-keten)transmembraanmoleculen die receptoren vormen voor extracel-lulaire matrixmoleculen. Specifieke heterodimeren hebben voorspecifieke matrixmoleculen hoge affiniteit (bijv. α6β1- en α 6β4-integrinen voor het basale membraaneiwit laminine). Bij infil-
tratieve groei raken de cellen los van hun basale membraan enhechten zich, om te kunnen migreren, aan interstitiële extracellu-laire matrixcomponenten. De cellen gebruiken hiervoor integri-nen. Bij infiltratieve groei zijn dan ook op tumorcellen kenmer-kende veranderingen in de expressie van integrinen gevonden.
Proteolyse van extracellulaire matrixcomponentenBij de afbraak van de extracellulaire matrix in het proces vaninfiltratieve groei zijn diverse enzymsystemen betrokken.Tumorcellen maar ook ontstekingscellen (zoals macrofagen)produceren niet-specifieke proteasen zoals trypsine en cathepsi-nen. Er worden echter ook specifieke proteasen geproduceerdwaaronder de matrix-metalloproteïnasen (MMP) en plasmino-geenactivatoren. Vooral MMP-2 en MMP-9 zijn van belangomdat die het collageen type IV van de basale membraan kun-nen afbreken. De activiteit van MMP’s wordt onder meer gere-guleerd door specifieke remmers, de ‘tissue inhibitors’ vanmetalloproteïnasen of TIMP’s. MMP’s worden niet alleen doortumorcellen geproduceerd maar ook door stromacellen. Tumor-cellen hebben receptoren op hun oppervlak waaraan dezeMMP’s binden. Bij de afbraak van de matrix door MMP’skomen groeifactoren vrij die in de matrix liggen opgeslagen.Naast het scheppen van ruimte voor migrerende tumorcellenbevorderen MMP’s derhalve ook de groei van infiltratievetumorcellen.Van de plasminogeenactivatoren (PA) is vooral het urokinase(uPA) uitvoerig onderzocht in het kader van infiltratieve groei.Ook daarbij is gebleken dat weliswaar soms tumorcellen uPAproduceren maar veelal juist de stromacellen. De tumorcellendragen dan receptoren voor uPA (uPAR) op hun celoppervlakwaardoor uPA wordt geconcentreerd en geactiveerd. PA zettenplasminogeen om in plasmine, dat betrokken is bij de bloedstol-ling maar ook bij de activatie van pro-enzymen door afsplitsingvan een peptide.Met al deze factoren in het spel is de afbraak van de extracellu-laire matrix bij invasieve groei een uiterst complex proces vaninteractie tussen tumorcellen en hun omgevende stroma. Veelaandacht bestaat er momenteel voor specifieke remmers vanmatrixproteasen als chemotherapeutica. Van sommige hiervan,zoals batimastat, is een specifiek tumorgroeiremmend effectvastgesteld.
1 FUNDAMENTELE ASPECTEN VAN KANKER 9
Figuur 1.7 E-cadherine kleuring van tumorcellen.
a Colonadenoom. Deze goed gedifferentieerde cellen tonen alle een celmembraan intens aankleurend voor dit celadhesie-eiwit.
b Coloncarcinoom. Rechts in het beeld een gebied met goed gedifferentieerde carcinoomcellen die sterk aankleuren. Links in het beeld diffuus infiltrerende weinig
gedifferentieerde cellen die hun onderlinge samenhang verloren hebben omdat E-cadherine niet langer tot expressie komt.
A B
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 9

MigratieGebleken is dat tumorcellen in vitro ten opzichte van normalecellen een verhoogde motiliteit bezitten. Invasief groeiendetumorcellen zijn in deze in-vitromodellen weer actiever danniet-invasieve tumorcellen. Bij deze verhoogde motiliteit is acti-vatie van het celskelet betrokken. Bij migratie door de ECMmaken tumorcellen gebruik van integrinen op hun celoppervlakvoor het (tijdelijk) hechten aan collagenen in de ECM. Er zijnverschillende factoren gevonden in tumorcellen die migratiebevorderen, waaronder de zogenaamde ‘autocrine motility’-fac-tor. Deze activeren het celskelet van de tumorcellen, die daar-door actief in beweging komen. Eenmaal infiltratief hebben dekankercellen het vermogen in lymfebanen en bloedvaten binnente dringen. Dit vormt het begin van metastasering.
1.5.2 Metastasering
Via lymfebanen worden tumorcellen versleept naar het eersteregionale lymfeklierstation. Via de afferente baan komen ze inde randsinus terecht vanwaar ze tot een metastatische tumorkunnen uitgroeien. Van daaruit kan een volgend lymfekliersta-tion worden bereikt dan wel een bloedvat. Ook kunnen uit lym-fekliermetastasen hematogene metastasen optreden.Hematogene metastasen ontstaan als de tumorcellen in debloedbaan terechtkomen. In veel gevallen bepaalt het gepene-treerde vat waar de metastase zal ontstaan, namelijk in het eerst-volgende capillairbed, waar de circulerende tumorcellen alstumorcelemboli vastlopen. Dit principe verklaart mede hetbestaan van specifieke metastaseringspatronen. Tumoren in debuikviscera zullen (via de v.portae) veelal levermetastasen vor-men. Tumoren uit huid, bewegingsapparaat en dergelijke zullenvia de v.cava veelal longmetastasen veroorzaken. Tumoren in delong zullen via de vv.pulmonales en de systemische arteriële cir-culatie metastasen in de perifere organen veroorzaken.Van een metastase wordt pas gesproken als de tumorcellen in het‘doelwit’orgaan een nieuwe tumor hebben gevormd. Bij lossetumorcellen in de randsinus van een lymfeklier of in een leversi-nus spreekt men dus niet van metastase. Pas wanneer de tumorcel-len de vaatwand penetreren, lokaal uitgroeien en een fibrovasculairstroma wordt gevormd, spreekt men van een metastatische haard.Sommige tumoren metastaseren in de eerste plaats hematogeen(sarcomen) en andere meer lymfogeen (carcinomen, melanoom).In het algemeen zullen tumorcellen via lymfevaten vanuit de pri-maire tumor worden getransporteerd naar het eerste lymfekliersta-tion. Vaak zullen deze lymfeklieren als filter functioneren. Vandeze filterfunctie wordt gebruikgemaakt bij het zoeken naar deschildwachtklier. Door in een tumor een radioactieve tracer of eenvitale kleurstof in te spuiten kan namelijk peroperatief de eerstelymfeklier die de lymfestroom bereikt worden geïdentificeerd. Indeze (schildwacht)lymfeklier vindt men de eerste lymfekliermeta-stasen. Als in deze lymfeklier geen metastasen worden gevonden,zijn er in het algemeen in andere lokale lymfeklieren ook geenmetastasen aanwezig en kan het lymfekliertoilet achterwege gela-ten worden. Bij de behandeling van het mammacarcinoom ensommige melanomen is de schildwachtlymfeklierprocedure stan-daard geworden. Dit wordt uitvoeriger besproken in latere hoofd-stukken.
De filterfunctie van lymfeklieren is niet zo effectief als men welgedacht had. Tumorcellen kunnen deze klieren namelijk passe-ren. De filterfunctie neemt af na lokale bestraling. Anderzijdszullen zeker niet alle tumorcellen die in het eerste lymfeklier-station worden gevangen, in staat zijn zich te handhaven en tevermeerderen. Als wel een lymfekliermetastase ontstaat, kan delymfeklier soms geheel worden vervangen door tumorweefsel,waarvan een retrograde lymfestroom het gevolg kan zijn. Vanuitlymfekliermetastasen kan niet alleen verdere lymfogene ver-spreiding optreden, maar kan ook hematogene disseminatie ont-staan.Het probleem van in het bloed circulerende tumorcellen heefteen tijd lang grote belangstelling genoten. Bij een groot aantalkankerpatiënten kunnen tumorcellen worden aangetoond in hetbloed van de venen die het tumorgebied draineren, en soms ookin bloed elders. Vooral met uiterst gevoelige moleculair-biologi-sche methoden (de polymerasekettingreactie of PCR), kunnentumorcellen in een groot aantal gevallen in de circulatie wordenaangetoond. Manipulatie van het tumorgebied, zoals palpatie,massage en curettage, verhoogt de kans op het voorkomen vandeze cellen in het bloed. Bij patiënten met een weinig gediffe-rentieerde tumor worden vaker tumorcellen in het bloed aange-troffen dan wanneer de primaire tumor een hoge differentiatie-graad bezit. De aanvankelijke verwachting dat het aantreffenvan tumorcellen in bloed een infauste prognose betekent, is nietbevestigd. Circulerende tumorcellen geven kennelijk niet steedsaanleiding tot aantoonbare metastasen. Ook op grond van expe-rimentele waarnemingen is het waarschijnlijk dat het merendeelvan deze in het bloed aanwezige cellen wordt vernietigd, het-geen echter niet betekent dat niet moet worden getracht deze uit-zaaiing te analyseren bij het diagnostisch en therapeutisch han-delen. Metastasering is alles bij elkaar een uiterst selectief pro-ces: weinig tumorcellen slagen erin. Echter, ondanks hetvermogen van het organisme om circulerende tumorcellen tevernietigen, zal een aantal nog vitale cellen in het capillaire enprecapillaire netwerk terechtkomen en daar worden vastgehou-den. De cellen ‘kleven’ vermoedelijk aan het endotheel en wor-den omgeven door een netwerk van fibrine, bloedplaatjes enleukocyten, voordat zij de vaatwand penetreren. Anticoagulantiaen fibrinolytische enzymen kunnen, na injectie van tumorcellenin de circulatie, het aantal metastasen verminderen.Of de tumorcellen die de vaatwand gepasseerd zijn, in hun nieu-we omgeving blijven leven en zich zullen vermeerderen, hangtin hoge mate af van de vascularisatie van het gebied waar zeterechtkomen en hun vermogen angiogene factoren te produce-ren. Tumorfragmenten, geïmplanteerd in de voorste oogkamervan een konijn, waar de vascularisatie minimaal is, gaan te gron-de. Dezelfde tumor, geënt in het gebied van de iris, met een rijkevascularisatie, groeit echter snel uit. Ook bij de mens kan hetbeloop van metastasen onverwacht zijn. De groeisnelheid kantijdelijk of permanent afnemen en spontane regressie is, hoewelzeldzaam, onder andere bij metastasen van niercarcinomen enmelanomen waargenomen. Anderzijds manifesteren metastasenzich soms vele jaren nadat een primaire tumor, voorzover is nate gaan, volledig is verwijderd, zoals bij mammacarcinoom.Men spreekt dan van ‘dormant’ metastasen. Wat hier preciesgebeurt is nog niet duidelijk. Het is denkbaar dat tumorcellenlangdurig in de G0-fase van de celcyclus blijven of dat trage
10 DEEL A ALGEMENE ASPECTEN
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 10

proliferatie (geheel) wordt tenietgedaan door apoptose. Er ligt indat geval een lange periode tussen het ‘aanslaan’ van de ver-sleepte tumorcellen en het moment waarop ze tot een diagnosti-ceerbare tumorhaard zijn uitgegroeid.Zoals reeds is vermeld, is de lokalisatie van de eerste hemato-gene metastase vaak bepaald door de lokalisatie van het eerstecapillaire netwerk dat door de tumorcellen via de bloedbaanwordt bereikt (tabel 1.4). Indien de hematogene disseminatieplaatsvindt via het cava-systeem (o.a. nier-, testis-, en bottumo-ren), is de long het eerste station dat wordt bereikt(vena-cava-type). Bij vervoer via de vena portae (o.a. maag-,pancreas- en colontumoren) is dit de lever (vena-portae-type).Tumorcellen dia via de longvenen in het arteriële systeemterechtkomen (longtumoren), kunnen metastasen vormen in elkorgaan met een capillair netwerk, met als voorkeurslokalisatieshersenen, lever, botten en bijnieren (vena-pulmonalis-type); ditwordt samengevat in figuur 1.8. In de tweede fase van de meta-stasering bepaalt de lokalisatie van de eerste metastasen de ver-dere disseminatie.
1 FUNDAMENTELE ASPECTEN VAN KANKER 11
Tabel 1.4 Lokalisatie van hematogene metastasen*
tumortype long lever skelet hersenen
coloncarcinoom 25-40 60-70 05-10 < 1
cervixcarcinoom 20-30 20-35 10-20 < 5
ovariumcarcinoom 05-10 10-15 02-05 < 1
prostaatcarcinoom 15-50 05-10 50-70 < 1
niercarcinoom 50-75 30-40 25-50 5-10
Wilms-tumor (nier) 10 30-40 10 0
blaascarcinoom 25-30 30-50 0
Ewing-sarcoom (bot) 70 30 60 < 5
osteosarcoom 80-95 < 5 35-50 < 5
testis 70-80 50-80 20-25 < 10
bijnier
long 25-35 35-40 10-15 20-25
* Bij obductie, % van het totale aantal gevallen.
via vv. pulmonalis, linker hart,aorta, systemische circulatie
v. pulmonalis-type
v. cava-type
v. portae-type
via perifere venen, v. cavae, rechter hart,
longen
via v. portae
orofarynx mamma bot
blaas
prostaat
gonaden
schildklier
nier
buikorganen
pertifere organen
Figuur 1.8 Metastaseringspatronen. De organen waar men hematogene metastasen kan verwachten, worden in eerste instantie bepaald door de bloedstroom:
tumorcellen worden in veneus bloed afgevoerd en lopen vervolgens in het eerstvolgende capillaire vaatbed vast.
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 11

Hoewel de lokalisatie van de metastasen zich in veel gevallen,zoals te verwachten, manifesteert op grond van de eerderbesproken vaatverhoudingen, worden we zo vaak met lokalisa-ties geconfronteerd die hierdoor niet worden verklaard, dat meerfactoren een rol moeten spelen. Klinische, anatomische en expe-rimentele waarnemingen kunnen althans ten dele deze afwij-kende metastaseringspatronen verklaren. Tumorcellen kunneneen capillair netwerk passeren en worden dan dus niet vastge-houden in het eerste netwerk dat op hun weg komt, bijvoorbeeldde longen, waardoor de metastasen uiteindelijk niet wordengevonden in de longen maar in andere organen. De eerste aan-wijsbare metastasen van een prostaatcarcinoom bijvoorbeeld,worden niet zelden gevonden in de lumbale wervelkolom. Menheeft dit wel verklaard door een retrograde bloedstroom bij ver-hoging van de intra-abdominale druk. Wanneer dit gebeurt, kanbloed uit de plexus prostaticus, die anastomosen heeft met deveneuze plexus vertebralis, in deze laatste terechtkomen. Dezeverklaring is echter wat op de achtergrond geraakt. Momenteelwordt aangenomen dat het micromilieu in het ‘ontvangende’orgaan, bijvoorbeeld door de aanwezigheid van bepaalde groei-factoren, bepalend kan zijn voor het al dan niet uitgroeien vaneen in een bepaald capillairbed vastgelopen tumorcel.Dat metastaseringspatronen niet alleen verklaard worden doorde bloedstroom, blijkt ook uit het feit dat in sommige organenopvallend weinig metastasen worden aangetroffen. De hartspierontvangt in rust ongeveer 25% van het hartminuutvolume, maartumormetastasen in de hartspier zijn tamelijk ongewoon. Derge-lijke waarnemingen hebben aanleiding gegeven tot de zoge-noemde voedingsbodemtheorie (‘seed and soil’). Het lijkt waar-schijnlijk dat de productie van specifieke groeifactoren in hetontvangende orgaan hiervoor verantwoordelijk is. Dit wordtgeïllustreerd door onderzoek naar metastasering in de lever,waar in dierexperimenten meer metastasen worden aangetroffenna hemihepatectomie, die een sterke proliferatieve activiteit vanhepatocyten tot gevolg heeft door lokale afgifte van groeifacto-ren, die ook de uitgroei van metastasen bevorderen. Specifiekemetastaseringspatronen kunnen ook worden verklaard door spe-cifieke binding van tumorcellen aan endotheelcellen via interac-tie van glycoproteïnen op het tumorceloppervlak met selectinenop endotheelcellen.Niet alle maligne tumoren metastaseren, en het moment waaropen de mate waarin dit proces plaatsvindt kunnen sterk wisselen.Basalecellencarcinomen van de huid metastaseren nagenoegnooit. Glioblastomen in de hersenen metastaseren uiterst zelden,hoewel ze zeer sterke angiogenetische activiteit vertonen. Daar-entegen kunnen zeer kleine tumoren die nog geen enkele klachtveroorzaken, reeds aanleiding geven tot massale disseminatie,zodat de eerste symptomen worden veroorzaakt door de meta-stasen. Anderzijds kunnen metastasen ontbreken bij zeer groteprimaire tumoren. Vaak is het gedrag onverwacht en onver-klaard. In het algemeen metastaseren weinig gedifferentieerdetumoren vroeger en uitgebreider dan die met een hoge differen-tiatiegraad. Dit is echter geen vaste regel, want tumoren vanschildklier en bijnier met een structuur die nog in hoge mateovereenkomt met die van normaal schildklier- en bijnierweefsel,kunnen toch metastasen veroorzaken.
1.6 Ontstaan van kanker
Tumoren ontstaan als gevolg van celveranderingen, waardoor decellen zich kunnen onttrekken aan normale groeiregulerendemechanismen. Deze celveranderingen worden samengevat metde term ‘transformatie’. Aan het proces van transformatie liggenmultipele afwijkingen (mutaties) in het genoom van de getrans-formeerde cel ten grondslag. Kanker is derhalve te beschouwenals afwijking van het genoom van de cel. Het proces van het ont-staan van kwaadaardige tumoren wordt oncogenese genoemd.Het ontstaan van maligne epitheliale gezwellen wordt met determ carcinogenese aangeduid.Men gaat ervan uit dat de meeste tumoren ontstaan door omge-vingsfactoren. Een veel kleiner aantal is het gevolg van erfelij-ke afwijkingen in het genoom. Dit zijn de familiale tumorsyn-dromen (zoals retinoblastoom, familiaal coloncarcinoom offamiliaal mammacarcinoom). Erfelijke en omgevingsfactorenwerken samen bij de carcinogenese. De erfelijke factoren beper-ken zich niet tot de genen betrokken bij familiale kankersyndro-men. Ook het vermogen tot het afbreken van carcinogene stoffenof de reactie op (natuurlijke) bestraling verschilt per individu,waardoor de kans op de ontwikkeling van kanker sterk verschilttussen individuen. De omgevingsfactoren die een rol spelen bijhet ontstaan van tumoren worden carcinogene factorengenoemd. Omdat deze afwijkingen in het genoom induceren,zijn carcinogene factoren zonder uitzondering ook mutageen. Inhet algemeen zijn tumoren monoklonaal, dat wil zeggen dat decellen die een gezwel bevolken allemaal zijn ontstaan uit ééngetransformeerde tumor-‘stamcel’. Gebleken is dat niet eenenkele maar meer mutaties nodig zijn om een cel te transforme-ren en zo de tumorstamcel te doen ontstaan. Carcinogenese isderhalve een complex meerstapsproces. De klonale expansievan één getransformeerde cel tot een klinisch manifeste tumorwordt bovendien door gastheerfactoren (bijv. hormonale ofimmunologische factoren) gemoduleerd.
1.6.1 Oncogenese
Het ontstaan van kanker is vooral aan epitheelcellen onderzocht.We spitsen daarom de bespreking van de mechanismen van hetontstaan van kanker hierop toe: carcinogenese. Carcinogenese iseen zeer langdurig proces. De transformatie van een normale celtot kankercel duurt lang omdat daartoe diverse mutaties in dezich transformerende cel moeten accumuleren. Verder is er eengroot aantal celdelingen nodig om uit één enkele getransfor-meerde cel een klinisch manifeste tumor te laten ontstaan.Afhankelijk van de delingsfrequentie kan dat wel vijf tot tienjaar duren.
Initiatie en promotieHet concept van meerstapscarcinogenese werd voor het eerstexperimenteel bewezen bij onderzoek naar chemische carcinoge-nese in de huid van muizen (zie fig. 1.8). Werd de huid van mui-zen eenmalig aan een lage dosis carcinogeen (benzpyreen)blootgesteld, dan ontstond geen kanker. Werd vervolgens dehuid behandeld met de irriterende stof crotonolie, waarvan
12 DEEL A ALGEMENE ASPECTEN
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 12

bekend is dat deze alleen nooit huidkanker veroorzaakt, dan ont-stond er wel huidkanker. Deze waarneming suggereerde dat bijde eerste (niet-kankerverwekkende) behandeling wel essentiëleveranderingen in de cellen waren ontstaan; dit werd ‘initiatie’genoemd. De cellen hadden daarna kennelijk nog maar eenklein zetje nodig om kankercellen te worden, wat door de cro-tonoliebehandeling geschiedde. Deze laatste werking werd‘promotie’ genoemd. Het is waarschijnlijk dat initiatie en pro-motie ook bij het ontstaan van sommige tumoren bij de menseen rol spelen. Vaak zal er niet sprake zijn van twee stappen,maar van een hele reeks. Als initiator worden processen aange-duid die in het genoom van de cel specifieke mutaties induceren.Initiërende factoren, carcinogenen, zijn zonder uitzonderingmutageen. Promotoren stimuleren proliferatieve activiteit vande geïnitieerde cel en beïnvloeden het patroon van de gen-expressie, onder meer verband houdend met celdifferentiatie,waardoor het proces van carcinogenese wordt bespoedigd. In depraktijk is het vaak niet mogelijk initiatie en promotiescherp van elkaar te scheiden: veel initiatoren hebben ook eenpromotorwerking.
Meerstapscarcinogenese bij menselijke tumorenOok bij veel tumoren bij de mens is meerstapscarcinogenesewaarneembaar. In veel epithelia gaat aan het ontstaan van eengezwel een langdurige periode van abnormale groei en differen-tiatie vooraf. Vooral aan goed toegankelijke, bekledende epithe-lia is dit goed bestudeerd. Het meest treffende voorbeeld is decervix uteri, waar een plaveiselcelcarcinoom wordt voorafge-gaan door metaplasie, dysplasie en carcinoma in situ. Deze ont-wikkeling wordt schematisch samengevat in figuur 1.9.
Van metaplasie wordt gesproken als de differentiatierichting vaneen epitheel verandert. Voor de cervix uteri betekent dit dat decilindercellige bekleding verandert in – overigens morfologischgeheel normaal – plaveiselepitheel. Metaplasie is geheel rever-sibel en wordt als reactie gezien in diverse epithelia op een helescala van prikkels. Bij doorgaande prikkeling vertoont het meta-plastische epitheel toegenomen proliferatieve activiteit en afne-mende differentiatie. De celkernen worden groter, het chromati-ne verandert van textuur. Naar de ernst van de morfologischeafwijkingen wordt dysplasie gegradeerd in licht, matig en ern-stig. Ten slotte is het celbeeld zo atypisch dat de cellen eruitzienals kankercellen. Zolang echter geen sprake is van voor kwaad-aardige groei kenmerkende infiltratieve groei, wordt gesprokenvan carcinoma in situ. Andere voorbeelden van premaligneafwijkingen worden genoemd in tabel 1.5.
Tumorbiologisch onderzoek heeft inmiddels uitgewezen dat inde ontwikkelingslijn metaplasie, lichte–matige–ernstige dyspla-sie, carcinoma in situ en plaveiselcelcarcinoom er een voortdu-rende toename is van afwijkingen in het genoom en van degedragingen van de cel. Deze waarnemingen maken duidelijkdat het tweestapsconcept van initiatie en promotie voor car-cinogenese bij de mens te simpel is. In de paragrafen over deafwijkingen in het genoom bij kanker komen we daarop terug.
1.6.2 Carcinogene factoren
Het is vanzelfsprekend ondenkbaar mogelijke carcinogeneagentia op hun werking experimenteel te onderzoeken bij demens. Veel van de kennis omtrent de carcinogene effecten vanin ons leefmilieu voorkomende noxen is dan ook afkomstig uitepidemiologisch onderzoek. Directe causale verbanden kunnenevenwel moeilijk uit epidemiologische gegevens worden afge-leid. Daarom zijn additionele onderzoeksmethoden ontwikkeldom experimenteel bewijs te kunnen leveren voor de carcinoge-niciteit van bepaalde noxen. Daartoe behoren expositie vanproefdieren aan noxen, testen van mutageniciteit in bacteriecul-tures (zgn. Ames-test) en testen van transformerende eigen-schappen in celcultures in vitro.Carcinogene factoren omvatten chemische agentia, fysische fac-toren en biologische agentia.
Chemische carcinogenenReeds eeuwen was bekend dat het voorkomen van carcinomenvan het scrotum bij schoorsteenvegers vermoedelijk een gevolgwas van contact met roet. Dit vermoeden werd ondersteund doorde bevinding dat bij proefdieren huidcarcinoom geïnduceerdkan worden door de huid met teer te penselen. Chemisch onder-zoek toonde uiteindelijk aan, dat dibenzantraceen een van debelangrijkste carcinogenen is in koolteer.Het aantal stoffen dat in staat moet worden geacht malignetransformatie te bewerkstelligen, is zeer groot. Sommige onder-zoekers menen dat het merendeel van de bij de mens voorko-mende maligne tumoren mede een gevolg is van contact metdergelijke stoffen. Het zoeken naar carcinogenen die mogelijkuit voedsel en milieu geëlimineerd kunnen worden, is eenbelangrijk aspect van de kankerbestrijding.De meeste carcinogenen zijn zelf niet in staat kanker te verwek-ken; ze moeten in het organisme worden omgezet in de effectie-ve carcinogene stof. Deze stoffen worden aangeduid als ‘indi-recte carcinogenen’. Dat geldt bijvoorbeeld voor vinylchloride,dat wordt omgezet in een epoxide, dat covalent aan DNA kanbinden en daarmee mutagene (en carcinogene) eigenschappenheeft verworven. Vinylchloride is betrokken bij het ontstaan vanangiosarcomen in de lever. Bij de ‘directe carcinogenen’ isomzetting niet noodzakelijk; ze zijn direct werkzaam. De een-heid bij de grote verscheidenheid in chemische samenstellingvan de carcinogene stoffen, ligt in de aard van hun onder invloed
1 FUNDAMENTELE ASPECTEN VAN KANKER 13
Tabel 1.5 Voorbeelden van premaligne afwijkingen
orgaan afwijking predispositie voor
colon adenoom adenocarcinoom
(adenomateuze/
villeuze poliep)
cervix uteri dysplasie plaveiselcelcarcinoom
huid keratosis actinica plaveiselcelcarcinoom
blaas papilloom overgangsepitheelcarcinoom
orofarynx leukoplakie plaveiselcelcarcinoom
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 13

14 DEEL A ALGEMENE ASPECTEN
normaal
dysplasie
carcinoma in situ
invasief carcinoom
meerlagig plaveiselepitheel
bloed- en lymfevaten
verminderde stratificatieonrijpe cellen hoog in het epitheel
totaal verlies van stratificatieintacte basale lamina
doorbraak van de basale laminainvasieve cellen kunnen invaten groeien
via de lymfe- of bloedbaan kunnentumorcellen andere organen bereiken;in die organen (solide organen, skelet,lymfeklieren) kunnen metastasenontstaan
lymfekliermetastasen
Figuur 1.9 Evolutie van een invasief carcinoom
uit normaal plaveiselepitheel via dysplasie en
carcinoma in situ.
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 14

van enzymen gevormde reactieve groepen. De chemische reac-tie van deze groepen met DNA is waarschijnlijk een belangrijkestap bij de aanvang van de carcinogenese. Deze laatste veron-derstelling is in overeenstemming met de waarneming dat bijnaalle chemische carcinogenen tevens mutageen zijn. Hieropberust de eerdergenoemde Ames-test voor mogelijke carcinoge-ne eigenschappen van chemische verbindingen. De voor trans-formatie noodzakelijke concentratie van het carcinogeen is wis-selend en afhankelijk van de stof en van het individu. Sommigecarcinogenen zijn alleen werkzaam op de plaats van toediening,andere kunnen ook elders in het organisme tumoren verwekken,soms met een opvallende voorkeur voor bepaalde organen.Blaaskanker is hiervan een sprekend voorbeeld: de voor uro-theel carcinogene stoffen worden in de urine geconcentreerd.De belangrijkste carcinogene stoffen kunnen op grond van hunstructuur en werkzaamheid in enkele groepen worden onderge-bracht (tabel 1.6).
Directe carcinogenenTot de groep alkylerende agentia behoren enkele cytostatica,onder andere cyclofosfamide. Nu bepaalde vormen van kankerkunnen worden genezen door chemotherapie, dient rekening teworden gehouden met hun oncogene werking. Een tweedemaligniteit is van chemotherapie (maar ook van radiotherapie)inmiddels een bekende complicatie.
Indirecte carcinogenenTot de aromatische aminen behoren naftylamine, benzidine,acetylaminofluoreen en de azo-kleurstoffen. Deze worden doorhydroxylering in de lever omgezet in carcinogene metabolieten.Naftylamine is een van de belangrijkste oorzaken van blaaskan-ker. Het zijn vooral arbeiders betrokken bij de productie en ver-werking van anilinekleurstoffen, onder andere in de textielin-dustrie die hiermee in contact komen.
Tot de polycyclische koolwaterstoffen behoren verschillendederivaten van koolteer, zoals dimenthylbenzantraceen, benzpy-reen en methylcholantreen. Ze worden in de lever omgezet totepoxiden, die een hoge affiniteit voor DNA vertonen. Deze stof-fen kunnen zowel op basis van applicatie als – na absorptie – opafstand tumoren induceren. Benzpyreen speelt een belangrijkerol bij het ontstaan van bronchuscarcinoom bij zware rokers.
Hoewel men dit wel vermoedt, is de betekenis van nitrosaminenen nitrosamiden voor het ontstaan van tumoren bij de mens nogniet op overtuigende wijze aangetoond. Zij zijn van grote bete-kenis voor de experimentele oncologie. Het zijn typische indi-recte carcinogenen, die vaak een opvallende specificiteit tonenwat betreft de inductie van tumoren in bepaalde organen. Het totdeze groep behorende ethylnitrosamine kan bij zwangere rattende placenta passeren, hetgeen het voorkomen van hersentumo-ren bij de jongen tot gevolg heeft.
Asbest en enkele carcinogene metalen: vooral het in verhoogdefrequentie voorkomen van mesotheliomen en ook bronchuscar-cinomen bij arbeiders in de asbestverwerkende industrieënheeft, ook in ons land, sterk de aandacht getrokken. Beryllium,cadmium, kobalt, nikkel en lood zijn als ion elektrofiel, waar-door zij zich waarschijnlijk aan biologisch actieve moleculen inde cel kunnen binden en mogelijk zo transformatie van de celbewerkstelligen.
In tegenstelling tot de bovengenoemde carcinogenen zijnnatuurlijke carcinogenen metabole producten van cellen, vooralschimmels. Ze komen zo in het ‘natuurlijke’ milieu voor. Debekendste van deze mycotoxinen is het aflatoxine, een productvan Aspergillus flavus. Epidemiologische gegevens maken hetwaarschijnlijk dat deze toxinen, onder andere aanwezig inbeschimmelde aardnoten, een rol spelen bij het ontstaan vanlevercelcarcinomen bij de mens. Inmiddels is gebleken dat afla-toxine een specifieke mutatie in het p53-tumorsuppressorgeninduceert.
1.6.3 Fysische oorzaken van kanker
Het is reeds lang bekend dat ioniserende stralen kanker kunnenverwekken. Er bestaat een duidelijk verband tussen het contactmet deze stralensoort in het kader van diagnostische en thera-peutische handelingen in het verleden en het optreden van kan-ker. Bij de mens moet aan deze mogelijkheid worden gedachtonder andere bij leukemieën, schildkliercarcinomen, huidcarci-nomen en mesenchymale tumoren. Het frequent en langdurigblootstaan aan natuurlijk ultraviolet licht kan mede een oorzaakzijn van huidkanker, zowel van het basale als plaatepitheeltype.
1 FUNDAMENTELE ASPECTEN VAN KANKER 15
Tabel 1.6 Chemische carcinogenen
categorie carcinogene stof type kanker
polycyclische koolwaterstoffen teer (sigarettenrook), benzpyreen, huid, scrotale tumoren, kanker van de ademwegen en longen,
dibenzanthraceen blaaskanker
aromatische amines benzidine, 2-nafthylamine blaaskanker
aflatoxines hepatocellulair carcinoom
nitrosaminen oesophagus- en maagcarcinoom
chemotherapeutica cyclofosfamide, chloorambucil, thiotepa leukemie
zware metalen Ni, Chr, Ars, Cd huidkanker (Ars)
vinylchloride angiosarcoom van de lever
asbest mesothelioom, longkanker
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 15

Ook melanomen kunnen op deze wijze ontstaan. De wijze waar-op maligne transformatie totstandkomt, is nog steeds niet geheelduidelijk en waarschijnlijk niet steeds dezelfde. In ieder gevalspeelt de inductie van mutaties in het DNA een centrale rol.Ultraviolet licht leidt tot de vorming van pyrimidinedimeren(vooral van twee thymidinebasen) in het DNA en de daardoorontstane structuurverandering kan een mutatie tot gevolg heb-ben. Dat is vooral duidelijk geworden uit onderzoek bij patiën-ten met de erfelijke aandoening xeroderma pigmentosum bij wieop de aan zonlicht blootgestelde delen van de huid reeds opjeugdige leeftijd veelvuldig carcinomen ontstaan. Onderzoekheeft uitgewezen dat bij deze aandoening het mechanisme datde door UV geïnduceerde DNA-veranderingen herstelt (zgn.excision repair) defect is. Daardoor hopen mutaties zich op inhet genoom van de cellen, hetgeen de kans op de ontwikkelingvan gezwellen sterk verhoogt. Bij ioniserende straling kan eenmutatie ontstaan als gevolg van een direct effect van de stra-ling op DNA, maar ook via een reactie van DNA met zuurstof-radicalen, die door radiolyse van water worden gevormd. Opchromosomaal niveau zijn deze effecten zichtbaar als breuken inde individuele chromosomen.
1.6.4 Biologische oorzaken van kanker
Van diverse micro-organismen is een rol bij het ontstaan vankanker vastgesteld of ten minste aannemelijk. Sommige para-sieten zijn geassocieerd met kanker. Het bekendste voorbeeld isSchistosoma haematobium, die zich in de urineblaas nestelt endaar op de lange duur leidt tot plaveiselcelcarcinoom. Eenmomenteel sterk in de belangstelling staande bacterie is Helico-bacter pylori, waarvan gebleken is dat deze bij maligniteiten(carcinomen, lymfomen) van de maag een rol van betekenisspeelt. Het belangrijkst zijn echter de carcinogene virussen. Debiologische carcinogene agentia zijn samengevat in tabel 1.7.
Rous was de eerste die (al in 1910) ontdekte dat door inspuitingvan een celvrij extract van een kippensarcoom, het sarcoom bijeen kip kon worden opgewekt. Deze bevinding leidde tot de ver-onderstelling dat virussen een rol spelen bij het ontstaan vankanker. Vooral tussen 1970 en 1980 is zeer uitgebreid onderzoekgedaan naar deze mogelijkheid. Bij veel van dat onderzoekspeelde het Rous-sarcoomvirus (RSV) een cruciale rol. Ontrafe-ling van de structuur van het virale genoom leidde tot de her-kenning van het ras-gen, waarmee de transformatie kon wordenbewerkstelligd. Dit ras-gen is een van de eerst ontdekte oncoge-nen. De ontdekking van de oncogenen is dan ook het belang-rijkste effect geweest van dat massale onderzoek naar viraleoncogenese.Hoewel een virusinfectie niet de meest frequente oorzaak is vankanker, zijn er bepaalde typen frequente tumoren met een vira-le etiologie. Dit zijn vooral het levercelcarcinoom, waarbij hethepatitis-B- of hepatitis-C-virus een rol speelt, en het carcinoomvan de cervix uteri, met als belangrijke etiologische factor infec-tie met humaan papillomavirus. Gebleken is dat voor het optre-den van gezwelgroei het virale genoom dient te worden geïnte-greerd in het genoom van de gastheercel. Voor een DNA-virus
is dit geen probleem. Bij een RNA-virus, waartoe de meesteoncogene virussen blijken te behoren, dient het virale RNA eerstte worden geconverteerd in DNA. Daartoe bezit het virus eengen dat codeert voor een RNA-afhankelijk DNA-polymerase,ook wel reverse transcriptase genoemd (omdat bij transcriptienormaal RNA van DNA wordt afgelezen). Het virus brengt ditgen tot expressie in de gastheercel, deze schrijft het virale RNAom in DNA en dat kan dan in het gastheer-DNA integreren.Het is onwaarschijnlijk dat een virale infectie alleen bij de mensgezwelgroei kan opwekken. Er wordt van uitgegaan dat voor hetontstaan van een gezwel ten minste twee incidenten noodzakelijkzijn. Ten aanzien van de virale oncogenen wordt deze opvattingondersteund door het feit dat infecties met virussen geassocieerdmet kanker (zoals HBV of HCV en HPV) vaak voorkomen zon-der dat gezwelgroei optreedt.Voor de volgende gezwelsoorten is een rol van virussen bij deetiologie aannemelijk:– De associatie tussen humaan papillomavirus en preneoplasti-
sche en neoplastische veranderingen van de cervix uteri staatmomenteel sterk in de belangstelling, omdat de effectiviteitvan de (sedert meer dan 50 jaar cytologische) screening wel-licht via het aantonen van viraal DNA kan worden verhoogd.Bij een zeer groot deel van de patiënten met dysplasie, carci-noma in situ of invasief carcinoom van de cervix worden inde afwijkende cellen virale antigenen gevonden en metbehulp van moleculair biologische technieken is ook viraalDNA in de cellen aangetoond. Een probleem hierbij is echterdat HPV ook voorkomt in onschuldige genitale wratten (con-dylomata) en zelfs bij een aanzienlijk deel van de vrouwenzonder enige cervicale afwijking. Er bestaat echter een grootaantal (> 70) subtypen van HPV. Hiervan komen vooral detypen 16 en 18 voor in associatie met premaligne en maligne
16 DEEL A ALGEMENE ASPECTEN
Tabel 1.7 Biologische verwekkers van kanker
orgaan type tumor
parasieten
Schistosoma haematobium blaas plaveiselcel-
carcinoom
bacteriën
Helicobacter pylori maag carcinoom, lymfoom
virussen
humaan papillomavirus cervix uteri plaveiselcelcarcinoom
(type 16, 18)
Epstein-Barr-virus lymfatisch Burkitt-lymfoom
systeem
nasofarynx plaveiselcelcarcinoom
hepatitisvirus (B, C) lever levercelcarcinoom
humaan T-lymfocytvirus lymfatisch T-cellymfoom
systeem
humaan herpesvirus mesenchym Kaposi-sarcoom
(type 8) lymfatisch lichaamsholtelymfoom
systeem
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 16

veranderingen en de typen 6 en 11 in associatie met goedaar-dige aandoeningen. Opmerkelijk is dat bij afwijkingen zon-der maligne potentie (condyloma acuminatum) hetHPV-genoom niet in het gastheer-DNA wordt geïntegreerden feitelijk een normale productieve infectiecyclus door-maakt. Bij cervixcarcinoom daarentegen wordt hetHPV-genoom geïntegreerd in het gastheer-DNA. Recentelijkis opgehelderd hoe expressie van virale eiwitten in cellengeïnfecteerd met HPV 16 of 18 leidt tot chromosoominstabi-liteit, mutaties in DNA en gestoorde groeiregulatie. De eiwit-ten E6 en E7 van het HPV kunnen zich binden aan het p53-eiwit of aan het Rb-eiwit, beide betrokken bij de regulatie vande celcyclus of van apoptose. De gevormde complexen inac-tiveren p53 en Rb, waardoor onder meer de celcyclus wordtontregeld.
– Bij het ontstaan van levercelcarcinoom spelen het hepati-tis-B-virus (HBV) en het hepatitis-C-virus (HCV) mogelijkeen belangrijke rol. Hiervoor zijn belangrijke epidemiologi-sche aanwijzingen. De frequentie van levercelcarcinoom ishoog in landen met een hoge besmettingsgraad met HBV ofHCV. China is daarvan een goed voorbeeld: levercelcarci-noom is daar een van de meest voorkomende gezwelsoorten.Ook hier geldt weer dat de virusinfectie niet de enige oorzaakkan zijn. Niet in alle gevallen van levercelcarcinoom wordtHBV of HCV gevonden en omgekeerd krijgt slechts eenbeperkt deel van de HBV- of HCV-geïnfecteerden een lever-celcarcinoom.
– Een zeldzame groep tumorgeassocieerde virussen vormen dehumane T-lymfocytvirussen (HTLV). Het HTLV-I is geasso-cieerd met lymfomen van het T-celtype. Bekender is hethumaan immunodeficiëntievirus (HIV), de veroorzaker vanaids. Kenmerkend voor deze aandoening is het optreden vanvaattumoren, het zogenaamde Kaposi-sarcoom. Inmiddels isduidelijk dat een herpesvirus, het humane herpesvirus type 8(HHV-8) een rol speelt bij het ontstaan van het Kaposi-sar-coom. HHV-8 is ook gevonden in het zeldzame lymfoom vande lichaamsholten, dat bij aids-patiënten voorkomt.
– Patiënten die lijden aan de oorspronkelijk in Afrika doorBurkitt beschreven lymfomen, tonen verhoogde antistoftiterstegen antigenen van het Epstein-Barr-virus (EBV), een DNA-virus van het herpestype. Zowel EBV-DNA als nucleaire anti-genen van EBV kunnen in tumorcellen worden aangetoond. Innasofarynxcarcinomen, met name van het lymfo-epithelialetype, in een kleine subgroep van maagcarcinoom en bij ver-schillende vormen van maligne lymfoom kan eveneens EBVworden aangetoond. Welke rol EBV bij de pathogenese vandeze tumoren speelt is slechts ten dele opgehelderd. Eenbelangrijke rol speelt het immuunsysteem: EBV-gerelateerdevormen van kanker komen vooral bij patiënten met immuun-insufficiëntie voor.
1.7 Moleculair-genetische aspecten vancarcinogenese
De laatste vijfentwintig jaar is veel duidelijk geworden over dewijze waarop genmutaties de transformatie van een normale celtot kankercel kunnen bewerkstelligen. Langzamerhand begintdeze kennis ook invloed uit te oefenen op de diagnostiek en demedicamenteuze behandeling van kanker. Van ten minste viergroepen genen is het aannemelijk dat ze een rol spelen bij hetontstaan van kanker: oncogenen, tumorsuppressorgenen, genendie zijn betrokken bij DNA-herstelprocessen, en telomerase.
1.7.1 Oncogenen
Het verhaal van de oncogenen begint met het onderzoek naar demogelijkheid dat virussen een rol zouden kunnen spelen bij hetontstaan van kanker. Bij dit onderzoek werd gebruikgemaaktvan virussen waarvan bekend was dat ze bij bepaalde diersoor-ten kanker konden verwekken (zoals sarcoom bij kippen, leuke-mie bij katten). Ontrafeling van het genoom van de betreffendevirussen maakte het mogelijk die gensequenties te identificerendie voor de maligne transformatie verantwoordelijk waren.Deze genen werden oncogenen genoemd. Bishop en Varmuswaren de eersten die in het midden van de jaren zeventig ont-dekten dat identieke of sterk gelijkende gensequenties voorko-men in normale, niet-getransformeerde cellen. Voor deze ont-dekking ontvingen zij in 1989 de Nobelprijs. Nader onderzoekwees uit dat deze genen zijn betrokken bij regulatie van celgroeien cellulaire differentiatie. De normale, aan oncogenen verwan-te, cellulaire genen worden proto-oncogenen genoemd. De pro-ducten van deze genen spelen een rol als groeifactor (bijv. het c-sis-gen dat homoloog is aan plaatjes-groeifactor PDGF), alsreceptor voor groeifactor (bijv. het c-erb-B2-gen dat homoloogis aan het gen dat codeert voor de receptor voor epidermalegroeifactor EGF), als doorgever van een signaal van het celop-pervlak naar de celkern (bijv. het c-H-ras gen) of als gentrans-criptieregulerende factor (bijv. c-fos). Dit wordt schematischsamengevat in figuur 1.10. De bekendste oncogenen wordensamengevat in tabel 1.8.
Mutaties van proto-oncogenen zijn dominant, dat wil zeggen datde expressie van een abnormaal eiwit door één gemuteerd allelvan het betreffende gen of van abnormale hoeveelheden van eennormaal oncoproteïne al transformerende werking heeft. Deveranderingen die cellulaire proto-oncogenen ondergaan bijactivatie tot oncogen zijn steeds activerend, dat wil zeggen zestimuleren een celfunctie die leidt tot ontregelde celgroei en -differentiatie. Voor de ontregeling van de expressie van onco-genen zijn ten minste vier mechanismen geïdentificeerd.
GenamplificatieHet blijkt dat van bepaalde oncogenen in kankercellen in plaatsvan één kopie per chromosoom een sterk verhoogd aantal voor-komt. Bekende voorbeelden hiervan zijn het c-erb-B2 (ook welneu)-oncogen dat bij een subgroep van het mammacarcinoom isgeamplificeerd en het N-myc-gen dat bij neuroblastoom geam-
1 FUNDAMENTELE ASPECTEN VAN KANKER 17
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 17

plificeerd kan zijn. Van de amplificatie van het c-erb-B2 wordttherapeutisch gebruikgemaakt bij het mammacarcinoom: her-ceptin is een anti-c-erb-B2 (neu)-antilichaam met celdodendeeigenschappen. Hoe dergelijke amplificaties totstandkomen isnog onduidelijk, maar wel is aangetoond dat dit toegenomengenetische materiaal leidt tot verhoogde aanmaak van het des-betreffende genproduct. Dit beïnvloedt het groeigedrag van decel: zo zijn neuroblastomen met een sterk geamplificeerdN-myc-gen agressiever dan neuroblastomen zonder of metslechts geringe N-myc-amplificatie.
Chromosomale translocatieBij sommige vormen van kanker komen karakteristieke chro-mosomale veranderingen voor. Een typisch voorbeeld daarvanis het Philadelphia-chromosoom bij chronische myeloïde leuke-mie, waarbij in feite sprake is van een translocatie van een stukjevan chromosoom 9 naar chromosoom 22. Op het stukje chro-
mosoom 9 ligt het c-abl-oncogen dat door deze locatie gefu-seerd is met het bcr-gen op chromosoom 22. Het nieuwe fusie-gen leidt tot de productie van een fusie-eiwit (bcr-abl), dat veelactiever is dan het oorspronkelijke c-abl-eiwit. Het fusie-eiwitheeft tyrosinekinaseactiviteit, dat met een specifieke remmer(Glivec) kan worden geblokkeerd. Dit heeft de resultaten van debehandeling van chronische myeloïde leukemie, maar ook vanandere tumoren met afwijkende tyrosinekinasereceptoren zoalsgastro-intestinale stromatumoren (ook bekend als GIST), sterkverbeterd.
PuntmutatieIn een proto-oncogen kan, zoals in elk ander gen, een puntmu-tatie optreden. Van veel van de inmiddels geïdentificeerde punt-mutaties is (nog) onduidelijk hoe ze precies ontstaan. Wel iszeker dat mutagene invloeden uit ons leefmilieu daarvoor ver-antwoordelijk zijn, met als duidelijke voorbeelden ioniserendestraling en chemische carcinogenen. Van deze laatste is inbepaalde gevallen vastgesteld welke mutaties zij veroorzaken.Zo is uit recent onderzoek gebleken dat benzpyrenen uit siga-rettenrook bijvoorbeeld in het ras-proto-oncogen vooral adduc-ten vormen met guanosine en dat deze in het DNA muteren naaradenosine (G- → A-substitutie). De bij coloncarcinomen betrok-ken carcinogenen veroorzaken juist G-T-substitutie. Door dezemutaties verandert de genetische code, wordt een eiwit gepro-duceerd met een andere aminozuurvolgorde en als dit actiever isdan het niet-gemuteerde eiwit kan de mutatie een oncogeeneffect hebben.
Virale infectieDoor middel van infectie met een (oncogeen) virus kan nieuwDNA in het genoom van de cel worden geïncorporeerd. Viralegenen zijn meestal vergezeld van promotor-DNA-sequenties dieleiden tot actieve transcriptie van het geïncorporeerde viraleDNA. Als het resulterende eiwit transformerende eigenschap-pen heeft, kan een gezwel zich ontwikkelen. Transformerendegenen van oncogene virussen interfereren vaak met de cellulai-re regelsystemen van de gastheercel. Het transformerende eiwitvan SV40-virus bijvoorbeeld (het zgn. large T-antigen) bindtaan het cellulair p53-eiwit dat een rol speelt bij de regulatie vande celcyclus ten behoeve van DNA-herstelprocessen in de cel-kern. Het E7-eiwit van het humaan papillomavirus bindt, zoalseerder reeds gemeld, aan het cellulaire Rb-eiwit dat eveneenseen regulerende functie heeft in de celcyclus.
1.7.2 Tumorsuppressorgenen
Zoals in de vorige paragraaf werd besproken, leidt abnormaleexpressie van een oncogen tot ontregeling van de cellulairegroei en/of differentiatie en daarmee tot de transformatie totkankercel. De laatste jaren is duidelijk geworden dat er ookgenen bestaan waarvan de expressie het ontstaan van kankercel-len onderdrukt. Kanker kan optreden als deze genen niet totexpressie komen of door een puntmutatie een abnormaal func-tioneel inactief eiwit maken. Deze genen worden tumorsuppres-sorgenen genoemd. Het bestaan van tumorsuppressorgenen is
18 DEEL A ALGEMENE ASPECTEN
translatie
kernmRNA
RNA
DNA
IV
III
III
celmembraan
ras
GAP
abl
transciptie
Figuur 1.10 Regelmechanismen in de cel, waarbij afwijkingen aan oncoge-
nen ontregeld betrokken kunnen zijn.
I Groeifactoren (bijv. PDGF).
II Groeifactor- en hormoonreceptoren (bijv. EGFR, neu).
III Intracellulaire signaaltransductie-eiwitten (bijv. ras, abl).
IV Transcriptieregulerende eiwitten (bijv. myc, fos, jun).
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 18

waarschijnlijk geworden door onderzoek naar erfelijke tumoren.Bekende voorbeelden hiervan zijn het retinoblastoom, familiairepolyposis coli en het familiair mammacarcinoom. Retinoblas-toom is een kwaadaardig gezwel van de pigmentcellaag van hetoog, voorkomend bij kinderen. Dit tumortype komt sporadischvoor, maar is in 30% van de gevallen familiair (en is dan door-gaans dubbelzijdig). Bij de familiaire vorm bestaat een kenmer-kende deletie in chromosoom 13. Deze bevinding suggereerdedat afwezigheid van een gen bij de pathogenese een rol zou kun-nen spelen. Uitgebreid moleculair genetisch onderzoek heeftuitgewezen dat er inderdaad een gen bestaat dat met retinoblas-toom is geassocieerd. Dit gen (Rb-1) is enkele jaren geledengeïsoleerd en blijkt te coderen voor een membraangebondenglycoproteïne dat een rol speelt in de regulatie van de celcyclus.
Ook het voor familiaire polyposis coli verantwoordelijke tumor-suppressorgen is recentelijk geïsoleerd. De tot nu toe geïdentifi-ceerde tumorsuppressorgenen worden samengevat in tabel 1.9.
Afwijkingen aan tumorsuppressorgenen zijn recessief, dat wilzeggen dat pas als beide allelen afwijkend zijn of ontbreken,tumorgroei ontstaat. Voor familiair retinoblastoom betekent datbijvoorbeeld dat niet de deletie van chromosoom 13 de tumorveroorzaakt. Deze treedt pas op als door een mutatie in het welaanwezige allel het normale genproduct niet langer tot expressiekomt. Deze werkhypothese (de zogenaante ‘double hit’-hypo-these) werd door Knudson geformuleerd op grond van zijnonderzoek van retinoblastoompatiënten. Dit wordt schematischgeïllustreerd in figuur 1.11. Belangwekkend is de vinding dat
1 FUNDAMENTELE ASPECTEN VAN KANKER 19
Tabel 1.8 Oncogenen, activatiemechanismen en de tumoren waarbij ze een rol spelen
categorie proto-oncogen activatiemechanisme type tumor
groeifactor
PDGF-B sis overexpressie glioom, osteosarcoom
fibroblastgroeifactor int-1 overexpressie mammacarcinoom
groeifactorreceptor
EGF-R erb-B1 amplificatie glioom, vele adenocarcinomen
EGF-like-R erb-B2 (neu) amplificatie mammacarcinoom,
maagcarcinoom
signaaltransductiefactoren
gtp-bindend eiwit ras puntmutaties long-, colon-, pancreascarcinoom
tyrosinekinase abl translocatie chronische myeloïde leukemie
transcriptiefactoren myc translocatie Burkitt-lymfoom
N-myc amplificatie neuroblastoom
kleincellig longcarcinoom
L-myc amplificatie kleincellig longcarcinoom
fos puntmutatie nier-, colon-, longcarcinoom
mitochondriaal eiwit
apoptoseremmer bcl-2 translocatie B-cellymfoom (folliculair)
schildklier-, coloncarcinoom
celcyclus-geassocieerd eiwit
cycline cycline D1 amplificatie oesophagus-, maagcarcinoom
translocatie mantelcellymfoom
Tabel 1.9 Tumorsuppressorgenen, hun functie en de tumoren waarbij ze een rol spelen
categorie tumorsuppressorgen functie type tumor
celmembraaneiwit DCC cel-cel-communicatie coloncarcinoom
APC celmembraan-celskeletinteractie, signaaltransductie coloncarcinoom
modulator signaaltransductie NF-1 GTP-ase-activerend eiwit neurofibromatose type 1
transcriptiefactor Rb-1 celcyclusbarrière (G1- → S-fase) retinoblastoom
p53 celcyclusbarrière, apoptose-inductie talrijke carcinomen
WT-1 onbekend Wilms-tumor
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 19

tumorsuppressorgenen niet alleen door mutatie of door genver-lies worden uitgeschakeld. Methylering van de genpromotor isin de laatste jaren geïdentificeerd als een belangrijk aanvullendmechanisme, dat epigenetisch wordt genoemd omdat het nietgaat om permanente irreversibele genafwijkingen. Van veeltumorsuppressorgenen is inmiddels duidelijk dat promotor-methylering een belangrijke rol speelt bij hun inactivering. Welkmechanisme voor abnormale promotor-methylering verant-woordelijk is, is nog niet duidelijk.
De moleculair-genetische bevindingen zijn bij familiaire vor-men van kanker van grote betekenis, omdat de genetische afwij-king kan worden vastgesteld voordat de tumor zich heeft ont-wikkeld. Voor ons inzicht in de oorzaken van kanker is dit typeonderzoek eveneens van doorslaggevend belang, omdat hiermeeeen geheel nieuw mechanisme van regulatie van celgroei is ont-dekt en een geheel nieuwe categorie van cellulaire regelgenen.
1.7.3 Genen betrokken bij herstelprocessen
Er is nog een aantal aandoeningen waarbij een sterk verhoogdefrequentie van bepaalde vormen van kanker voorkomt. Ditbetreft onder meer de zogenaamde Fanconi-anemie, waarbijleukemie voorkomt; ataxia teleangiectasia, waarbij malignelymfomen voorkomen; het syndroom van Bloom, waarbij leu-kemie voorkomt; en xeroderma pigmentosum, waarbij zich inde aan de zon blootgestelde delen van de huid in hoge frequen-tie plaveiselcelcarcinomen, basaliomen en melanomen ontwik-kelen. Gebleken is dat de cel een complex systeem heeft omfouten in DNA, ontstaan bij de replicatie of door chemische of
fysische mutagene invloeden, ter herstellen. De mutaties geïn-duceerd door (chemische of fysische) mutagene factoren wor-den in het proces van de zogenaamde excision repair hersteld.Hierbij zijn eiwitten betrokken die de mutatie herkennen, ver-wijderen uit DNA (vandaar ‘excision’) en het defect met de cor-recte sequentie herstellen. De ERCC-gen-familie codeert voordeze eiwitten en mutaties hierin leiden tot onder meer xeroder-ma pigmentosum.Fouten die ontstaan bij de DNA-replicatie (replication errors)worden hersteld door het zogenaamde ‘mismatch repair’-sys-teem. Defecten in dit systeem leiden tot instabiliteit van hetmicrosatelliet DNA. Dit zijn niet-coderende DNA , opgebouwduit repetitieve bi-, tri- of complexere nucleotidensequenties.Deze microsatellietinstabiliteit (MSI) komt bij een klein per-centage sporadische tumoren voor maar is een essentieelkenmerk van de tumoren bij het syndroom van Lynch, ook welhereditair niet-poliep-geassocieerd coloncarcinoom (HNPCC)genoemd. Genen waarvan mutaties verantwoordelijk zijn voordit syndroom zijn inmiddels gekarakteriseerd (MLH-1, MSH-2,MSH-6, PMS-2). Mutaties in DNA-herstelgenen zijn niet obli-gaat carcinogeen. Ze leiden echter tot instabiliteit in het genoomen de begeleidende verhoogde mutatiefrequentie speelt in detumorontwikkeling een rol. Vogelstein noemt deze genen ‘care-takers’, de verzorgers van de integriteit van het genoom. Degenen die de delingsactiviteit van cellen regelen, en waarvanmutaties wel obligaat betrokken zijn bij de carcinogenese,noemt hij ‘gatekeepers’, toezichthouders op de snelheid van deceldeling.
20 DEEL A ALGEMENE ASPECTEN
‘double hit’-hypothese van Knudson
A. sporadische tumor
B. erfelijke tumor
eerste hit: somatischemutatie in één allel
tweede hit: deletie vanhet tweede allel
sporadische tumor:enkelvoudig, oplatere leeftijd
kiembaanmutatiein één allel
tweede hit: deletie vanhet tweede allel
erfelijke tumor:meerdere tumoren, opjonge leeftijd
wild type (normaal) tumorsuppressorgen
gemuteerd tumorsuppressorgen
Figuur 1.11 Schematische weergave van de ‘double hit’-hypothese van Knudson, die postuleert dat in een cel beide allelen van een tumorsuppressorgen moeten
worden geïnactiveerd alvorens een tumorcel kan ontstaan. De eerste stap is bij familiale vormen van kanker al in de kiemlijn aanwezig. Tumoren ontstaan in der-
gelijke families derhalve al op jonge leeftijd en zijn vaak multipel. De beide allelen moeten in een zich ontwikkelende kankercel worden uitgeschakeld door twee
genetische incidenten (‘hits’). Sporadische vormen van kanker doen zich daarom later voor en de tumoren zijn meestal enkelvoudig.
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 20

1.7.4 Telomerase
De laatste jaren staat de vraag hoe tumorcellen een onbeperktdelingsvermogen hebben verworven sterk in de belangstelling.Normale lichaamscellen hebben een beperkt replicatief vermo-gen. Huidfibroblasten in vitro, bijvoorbeeld, verliezen dit ver-mogen na zo’n 60-70 verdubbelingen. Aangenomen wordt dathierbij verlies van DNA aan de chromosomale uiteinden, detelomeren, een rol speelt. Experimenteel onderzoek heeft aan-getoond dat telomeerverlies inderdaad leidt tot verlies vandelend vermogen. Recentelijk is ontdekt dat sommige lichaams-cellen, met name spermatogonia, hematopoëtische stamcellenen stamcellen in andere sneldelende weefsels (bijvoorbeelddarmslijmvlies) een enzymsysteem hebben dat telomeren oplengte houdt. Dit telomerase is een enzym dat op basis van eenRNA-matrijs, die deel uitmaakt van het enzymcomplex, het ver-loren gegane telomere DNA aanvult. Dit telomere DNA is repe-titief, dat wil zeggen het bestaat uit herhalingen van het motiefTTAGGG. Telomerase is in bijna alle carcinoomcellen gevon-den en in een deel van de sarcomen. De verrassende specificiteitvan telomeraseactivatie voor maligne tumorcellen heeft geleidtot hoge verwachtingen ten aanzien van de mogelijkheid telo-meraseactiviteit in de diagnostiek te gebruiken. Voorts zoudentelomeraseremmers theoretisch vrij specifieke chemotherapeuti-ca opleveren. Onderzoek zal moeten uitwijzen hoe waardevoltelomerase als diagnostisch en therapeutisch doelwit is.
1.7.5 Grove chromosomale veranderingen
Zoals boven beschreven is het hoogst waarschijnlijk dat discre-te genetische veranderingen (zoals puntmutaties of genamplifi-caties) een rol spelen bij het ontstaan van kanker. De meestekankercellen zijn genetisch onstabiel, hetzij door instabiliteitvan chromosomen hetzij door defect DNA-herstel. Deze zijndus niet de oorzaak maar het gevolg van de transformatie tot
kankercel. Deze genetische instabiliteit, die kan leiden tot ver-anderingen in de eigenschappen van de getransformeerde cel,speelt vermoedelijk een rol bij de progressie van kanker, hetproces dat uiteindelijk leidt tot een invasief groeiend en meta-staserend gezwel. Het complexe proces van de carcinogenesewordt schematisch samengevat in figuur 1.12.
Bij chromosomaal onderzoek van kankercellen worden afwij-kingen in het aantal chromosomen gevonden (monosomie bijverlies van een chromosoom of polysomie bij een teveel). Ookkomen structurele veranderingen voor, zoals translocaties endeleties. Bij sommige soorten kanker komen karakteristiekeveranderingen voor, zoals bij Burkitt-lymfoom (8;14-transloca-tie), chronische myeloïde leukemie (9;22-translocatie) en reti-noblastoom (deletie chromosoom 13). Wekedelensarcomenhebben translocaties die voor sommige tumortypen bijna net zospecifiek zijn als het histologische beeld, een eigenschap waar-van bij de moleculaire kankerdiagnostiek in toenemende mategebruik wordt gemaakt. De chromosomale veranderingen leidenveelal tot veranderingen in de hoeveelheid DNA in de celkern.Als globale maat voor de ernst van deze veranderingen kan dehoeveelheid DNA per celkern worden gemeten met behulp vanstatische of doorstroomcytometrie. Van deze techniek wordt inde tumordiagnostiek in toenemende mate gebruikgemaakt. Eenabnormaal DNA-gehalte (aneuploïdie) komt nagenoeg alleen bijkankercellen voor en voorts blijken aneuploïde kankercellenzich meestal agressiever te gedragen dan euploïde. Dit wordtgeïllustreerd in figuur 1.13.
1.8 Stromareactie, angiogenese
Als in de ontspoorde cel eenmaal een zodanig aantal verande-ringen in de groeiregulerende genen is opgetreden dat deze zichaan fysiologische groeiregulatie onttrekt, is er in principe spra-ke van een kankercel. Men gaat ervan uit dat een grote meer-
1 FUNDAMENTELE ASPECTEN VAN KANKER 21
initiatie
promotie
carcinogeen agens (chemisch, fysisch, biologisch)
binding aan of modificatievan DNA
geïnitieerde cellen
klonale expansie
tumorcelkloon
metabolische activatie
elektrofiel stofwisselingsproduct
apoptose
DNA-herstel
tumor
stromale reactie, angiogenese
Figuur 1.12 Samenvatting van de meerstapscarcinogenese
met als model expositie aan een chemisch carcinogeen.
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 21

derheid van alle goed- en kwaadaardige gezwellen ontstaat uitéén cel; tumoren zijn derhalve monoklonale celpopulaties. Voordeze veronderstelling bestaat een grote hoeveelheid experimen-teel bewijs. Van deze eigenschap van tumoren wordt in deimmunohistochemische en moleculaire diagnostiek steeds meergebruikgemaakt.Eén cel is echter nog geen tumor. Klinisch manifest wordentumoren meestal pas als ze een minimumvolume van 0,5-1 cm3
hebben bereikt. Aanvankelijk vermeerdert het celklompje zichzonder duidelijke reactie van de omgeving. Bij een volume vanongeveer 1 mm3 (dit is experimenteel onderzocht) induceren detumorcellen proliferatie van (myo)fibroblasten en endotheel-spruitjes. Dit proces is vergelijkbaar met de weefselherstelreac-tie bij wondgenezing en wordt gemedieerd door dezelfde groei-factoren. Met name de ontwikkeling van de vascularisatie vantumoren, de zogenaamde angiogenese, is momenteel sterk in debelangstelling. Angiogenese blijkt een delicaat gereguleerd pro-ces waarbij groeifactoren, onder invloed van hypoxie geprodu-ceerd door tumorcellen (‘vascular endothelial growth’-factor ofVEGF, fibroblastengroeifactor of bFGF en angiopoëtines), maarook remmers van vaatgroei (angiostatine en endostatine) een rolspelen.Veel onderzoek wordt momenteel gewijd aan mogelijk-heden angiogenese te remmen en daarmee verdere groei van detumor te voorkomen. Verder is gebleken dat de vaatdichtheid intumoren een belangrijke prognostische factor is.Hoewel alle cellen in een maligne tumor uit één cel afkomstigzijn (tumorstamcel), blijken toch niet alle cellen in zo’n gezwelidentiek te zijn. De cellen in één tumor kunnen verschillen ver-tonen in morfologie, differentiatiegraad, vermogen tot infiltra-tieve groei en metastaserende eigenschappen en in hun genoom.Voor een deel berusten deze verschillen op invloeden uit deomgeving van de tumorcellen: stromale componenten kunnenbijvoorbeeld differentiatie in tumorcellen induceren. Zoalsgezegd worden voor een belangrijk deel deze verschillen ver-oorzaakt door genetische instabiliteit van maligne cellen. Doordeze eigenschap ontstaan in een kankercel steeds weer nieuwegenetische veranderingen die van betekenis kunnen zijn voor hetgedrag van de kankercel. Nieuwe afwijkingen die de cel een
groeivoordeel zullen verschaffen, zullen leiden tot versneldetumorgroei. Dit verschijnsel, waarbij steeds nieuwe genetischeafwijkingen in een micro-evolutionair proces het ontstaan vansteeds agressievere kankercellen tot gevolg hebben, wordttumorprogressie genoemd. Tumorprogressie wordt verantwoor-delijk geacht voor veel klinische verschijnselen van kanker: hetresistent worden tegen hormonale en chemotherapie, het agres-sievere groeigedrag van metastasen ten opzichte van de primairetumor en het agressievere groeigedrag van lokale tumorrecidie-ven.
1.9 Factoren die het ontstaan van kankerbeïnvloeden
1.9.1 Hormonale factoren
Reeds in 1895 nam Beatson in Glasgow waar dat de verwijde-ring van de ovaria bij een vrouw met borstkanker regressie vanhet proces tot gevolg had. De rol van de hormonen bij het ont-staan en de ontwikkeling van sommige vormen van kanker issindsdien door dierexperimenten en klinische waarnemingenoverduidelijk aangetoond. Hormonen werken echter op eenandere wijze dan de eerdergenoemde exogene factoren, diewaarschijnlijk direct het DNA in de celkern beïnvloeden. Hor-monen maken de cel gevoeliger voor de invloed van bijvoor-beeld virussen of chemische carcinogenen. Mogelijk is er ooksprake van een ‘promoting effect’, dat wil zeggen dat de celreeds veranderingen heeft ondergaan die niet hebben geleid toteen autonome groei, maar dat onder invloed van het hormoondat de gen-expressie van de doelwitcel beïnvloedt, de tumor-groei manifest wordt.Klinische waarnemingen bij de mens maken het zeer aanneme-lijk dat het ontstaan en het gedrag van het mamma-, endometri-um- en prostaatcarcinoom mede afhankelijk zijn van endogeneof exogene hormonen. Een voorbeeld hiervan is de zeer lage fre-quentie van prostaatkanker bij castraten. Van de groeireguleren-
22 DEEL A ALGEMENE ASPECTEN
aan
tal cellen
>
a b c
G0,1
G2,M
S
G0,1G0,1T
G0,1G0,1T1
G0,1T2
Figuur 1.13 DNA-flowcytometrische analyse van tumorcellen.
a Een normale (delende) celpopulatie. De diploïde G0- en G1-fasecellen hebben een diploïde DNA-hoeveelheid. Gedurende de S-fase neemt deze gestaag toe en bij
de celdeling (na de G2- en M-fase waarin er een dubbele hoeveelheid is) ontstaan weer twee diploïde cellen.
b Als er bovendien aneuploïde tumorcellen zijn, wordt een G1-celpopulatie gezien met een hoeveelheid DNA groter dan van de diploïde cellen (G1T). De tumor-
cellen in de G2/M-fase (G2/MT) hebben een eigen piek in het histogram.
c Bij sommige tumoren bestaan verscheidene aneuploïde celpopulaties, die met verschillende DNA-hoeveelheden in het DNA-histogram vertegenwoordigd zijn (G1T1,
G1T2).
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 22

de (stimulerende, maar ook remmende) werking wordt gebruik-gemaakt bij de hormonale behandeling van kanker.Het is bekend dat sommige maligne tumoren, zoals het pros-taatcarcinoom, het mamma- en mogelijk het endometriumcar-cinoom, nog een duidelijke hormoongevoeligheid kunnen tonenen beïnvloedbaar zijn in hun groei door de hormonale status tewijzigen. Prostaatcarcinomen reageren vaak goed op castratie oftoediening van oestrogenen. Sommige mammacarcinomen kun-nen in gunstige zin worden beïnvloed door toediening van hor-monen, castratie en adrenalectomie. Het tegenovergesteldeeffect zien we soms gedurende een zwangerschap; een bestaandmammacarcinoom kan dan sneller gaan groeien. De gevoelig-heid voor deze hormonen wordt bepaald door de aanwezigheidvan receptoren voor steroïdhormonen.
1.9.2 Immunologische factoren
Tumorcellen zijn in het organisme zelf (autochtone gastheer) uiteigen cellen ontstaan. Men zou dus verwachten dat zij als‘eigen’ worden herkend, door het ontbreken van antigene deter-minanten op de cellen waarvoor het organisme niet tolerant is.Dit in tegenstelling tot de situatie bij binnengedrongenmicro-organismen, die wel vreemde antigenen hebben en dusals lichaamsvreemd kunnen worden herkend. Uit vroege tumor-transplantatie-experimenten is echter gebleken dat tumortrans-plantaten frequent worden afgestoten. Deze bevinding leidde totde conclusie dat er tumorgeassocieerde transplantatie-antigenen(TATA) moeten zijn. Inderdaad is het bestaan van TATA in expe-rimentele modellen bevestigd. In het bijzonder kon worden aan-getoond dat sommige chemisch geïnduceerde carcinomen insyngene (genetisch homogene) muizenstammen wel degelijkeen immuunreactie oproepen.Bij experimenteel opgewekte tumoren is het ontstaan van dezeantigenen afhankelijk van het agens dat de tumorgroei heeftgeïnduceerd. Cellen uit door virussen geïnduceerde tumorenzijn in het bezit van oppervlakteantigenen die zijn gecodeerddoor het virale genoom. Vaccins gemaakt van dergelijke tumor-cellen zijn in sommige gevallen in staat andere dieren tebeschermen tegen de oncogene activiteit van dat virus. Dat isanders bij tumoren, geïnduceerd door chemische of fysischecarcinogenen. Tumorcellen die zijn ontstaan na een dergelijkecarcinogene prikkel, hebben niet steeds dezelfde specifiekeantigenen op het celoppervlak. Indien specifieke antigenen ont-staan, wisselen deze sterk binnen de populatie van de tumorcel-len. De antigeniciteit van tumoren, die door chemische of fysi-sche carcinogenese in een dier zijn opgewekt, kan van tumor tottumor sterk wisselen. Voor het ontstaan van dergelijke antigenenop tumorcellen zijn verschillende verklaringen aangevoerd.Denkbaar is dat door het carcinogene agens genetische infor-matie tot expressie komt die normaal niet tot expressie komt.Een andere mogelijkheid is dat door veranderde synthese van dekoolhydraatcomponent van celoppervlakgebonden glycoproteï-nen afwijkende antigene structuren ontstaan. Ten slotte is hetdenkbaar dat antigenen die normaal binnen de cel blijven, doordefecte celfunctie of celnecrose op het celoppervlak terechtko-men.
Voor het bestaan van TATA in menselijke tumoren zijn slechtsindirecte aanwijzingen gevonden. Bij het Burkitt-lymfoom bij-voorbeeld, dat geassocieerd is met het Epstein-Barr-virus,komen afwijkende oppervlakteantigenen voor. Een treffendvoorbeeld is verder het choriocarcinoom bij de vrouw, dat uit deplacenta ontstaat en derhalve lichaamsvreemde HLA-antigenenbezit. Mogelijk is het optreden van een immuunreactie tegen detumorcellen een bijkomende reden waarom chemotherapie vanchoriocarcinoom bij de vrouw zo succesvol is gebleken.Bij een immuunreactie tegen tumorcellen spelen lymfocyten eendoorslaggevende rol. Een belangrijk bewijs hiervoor is dat in hetdiermodel lymfocyten van een tumordragend dier immuniteittegen de tumor overdragen naar een gezonde gastheer. VooralT-lymfocyten zijn belangrijk. T-helpercellen herkennen hettumorantigeen en produceren cytokinen (zoals interleukine-2)die de cytotoxische T-cellen aanzetten tot interactie met detumorcel, leidend tot celdood. Van de inductie van cytotoxischeeigenschappen door cytokinen wordt gebruikgemaakt bij som-mige vormen van immunotherapie. Daarbij worden de patiënteigen, in vitro door cytokinen geactiveerde killer-cellen (lymfo-kine activated killer of LAK-cellen) toegediend. Een anderecategorie lymfocyten, de zogenoemde ‘natural killer’ (NK)-cel-len, speelt eveneens een belangrijke rol. De celdodende eigen-schappen van NK-cellen worden niet geïnduceerd door eenspecifieke immunologische reactie. Waarom tumorcellen gevoe-liger zijn voor NK-celactiviteit dan normale cellen is nog goed-deels onbekend. Ook antilichaamafhankelijke ‘killer’-cellenhebben een belangrijke rol. Dit zijn killerlymfocyten (misschienook monocyten) die een receptor hebben voor het Fc-deel vanIgG en cellen herkennen die aan hun oppervlakteantigenen IgGdragen. Specifieke immunotherapie heeft lange tijd veel belangstellinggehad, zoals blijkt uit onderzoek naar de effectiviteit van behan-deling met LAK- en TIL-cellen. Echte doorbraken zijn nog nietbereikt. Een andere vorm van specifieke immunotherapie isimmunisatie van een patiënt met een preparaat gemaakt uiteigen tumorcellen. Momenteel wordt actieve immunisatie metpeptiden die voorkomen op de tumorcellen van de patiënt of metDNA, coderend voor dergelijke peptiden onderzocht. Ook kun-nen dendritische cellen uit lymfatisch weefsel van de patiëntworden geïsoleerd, blootgesteld aan antigene peptiden en aldusspecifiek geactiveerd aan de patiënt worden teruggegeven. Der-gelijke vormen van manipulatie van het immuunsysteem zijnechter nog experimenteel.Niet-specifieke immunotherapie is eveneens uitvoerig onder-zocht. Het meest toegepast is immunisatie met BCG of Coryne-bacterium parvum, die een niet-specifieke immuunreactie indu-ceert waarmee ook de specifieke reactie tegen de tumorcellenzou kunnen worden gestimuleerd. Voor sommige vormen vankanker is daarmee enig resultaat verkregen. In opkomst is pas-sieve immunotherapie met monoklonale antistoffen, gerichttegen tumorcellen. Deze benadering heeft, zoals eerder reedsgemeld, enig succes bij mammacarcinoom, waarbij antistoffentegen het her-2-neu-oncogen (herceptine) worden toegepast.Het moge duidelijk zijn dat de beperkte kwantitatieve verschil-len, meer dan kwalitatieve antigene verschillen tussen normalecellen en tumorcellen, effectieve immunotherapie niet gemak-
1 FUNDAMENTELE ASPECTEN VAN KANKER 23
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 23

kelijker maken. De verschillen zijn er echter en verwacht magworden dat immunotherapie een plaats zal verwerven in het toe-komstige therapeutische arsenaal.
1.10 Conclusies
Het kankeronderzoek heeft in het laatste decennium grote vor-deringen gemaakt, vooral door de toepassing van de modernemoleculair-biologische technieken. Moleculair-genetisch onder-zoek leidde tot de ontdekking van de kankergeassocieerdegenen. Het concept dat kanker een aandoening is van hetgenoom of van ontregelde genexpressie, met name van de genendie celgroei, apoptose en celdifferentiatie regelen, heeft alge-meen ingang gevonden. De vraag rijst wat al deze kennis bete-kent in klinisch perspectief. Afgezien van de mogelijkheid metbehulp van moleculair-genetische technieken familiale vormenvan kanker op te sporen lijkt het verdiepte inzicht in de ont-staanswijze van kanker slechts geringe invloed te hebben op dediagnostiek en de behandeling van de meest frequente vormenvan kanker. Het is belangrijk hier op te merken dat veel van hetfundamentele kankeronderzoek zich heeft gericht op de rol vanéén of hoogstens enkele genen bij het ontstaan van één bepaal-de vorm van kanker. Deze reductionistische benadering heeftzijn beperkingen. Het zou voor de hand liggen bij een aandoe-ning die veroorzaakt wordt door een samenspel van een veelheidvan factoren een wat globalere analyse te doen van afwijkingenin genexpressie. De beperkingen van de beschikbare analy-setechnieken zijn er verantwoordelijk voor dat dit tot voor kortzo weinig gebeurde. Terugblikkend is het niet moeilijk te begrij-pen waarom de studie van één enkel of hooguit enkele genen ineen bepaald type kanker geen afdoende antwoorden opleverdeop de belangrijkste vragen. Het is de combinatie van verschil-lende genafwijkingen en afwijkende expressie van crucialegenen die bepalend is voor het type tumor dat zich ontwikkelten het te verwachten gedrag van de tumor. Twee ontwikkelingen zijn in dit perspectief essentieel. In deeerste plaats betreft dat het ‘pathway’ concept. Het zijn niet deindividuele ontregelde genen maar het is de combinatie van ver-anderingen in een tumor die bepalend is voor het tumorgedrag.Individuele genveranderingen treffen steeds voor de regulatievan celgroei en -differentiatie essentiële signaaltransductieme-chanismen (pathways). Voorbeelden hiervan zijn de Wnt-path-way, essentieel in de colorectale carcinogenese en met alsbelangrijke vertegenwoordigers APC, β-catenine, Lef/Tcf, deRb-pathway met p16, CDK enzovoort die bijvoorbeeld een rolspelen bij het ontstaan van melanoom en de TGF-β-pathwaymet de Smad-genen die bij colon- en pancreascarcinoom een rolspelen.In de tweede plaats is er de ontwikkeling van de microarray-technieken voor de grootschalige analyse van eventuele muta-ties of de expressie van duizenden genen in slechts enkele expe-rimenten. Hiermee is een beter beeld verkregen van het geheelvan afwijkingen op moleculair niveau in kankercellen. Dergelij-ke technieken zullen het op redelijk korte termijn mogelijkmaken een betrouwbaarder voorspelling te doen ten aanzien vanhet tumorgedrag en de reactie op bepaalde vormen van therapie.
Het heeft lang geduurd voordat het fundamenteel kankeronder-zoek invloed begon te krijgen op de dagelijkse diagnostiek enbehandeling van kanker. Daarin komt snel verandering.
1.11 Samenvatting
Kanker is niet één ziekte, maar een uiterst heterogene groepaandoeningen die als gemeenschappelijk kenmerk hebben datde regulatie van de celgroei en/of celdood is gestoord, waardooreen pathologische vermeerdering van cellen optreedt. In combi-natie met de stromale reactie, die de groeiende tumorcellenoproepen in het weefsel waarin ze ontstaan, leidt dit tot detumor. Van kwaadaardige tumoren, in het dagelijkse spraakge-bruik met de term ‘kanker’ aangeduid, is het belangrijkste ken-merk dat ze uitzaaiingen (metastasen) kunnen veroorzaken.Kanker ontstaat door afwijkingen in de genen die celgroei enceldood regelen.Een correcte classificatie van tumoren is belangrijk voor ade-quate therapie. De klassieke morfologische diagnostiek is in delaatste decennia verrijkt met geavanceerde diagnostische hulp-middelen, zoals immunohistochemie en cytometrie, waarmeetumormerkstoffen en afwijkingen in het genoom van de cel kun-nen worden aangetoond. In de laatste jaren neemt het gebruikvan moleculair biologische diagnostiek van kanker sterk toe.Deze aanpak is gebaseerd op een sterk verbeterd inzicht in demoleculair genetische processen die aan het ontstaan van kankerten grondslag liggen. Deze technieken maken het bijvoorbeeldmogelijk de reactie van individuele tumoren op bepaalde behan-delingsvormen te voorspellen.
Kernpunten
• Tumoren bestaan niet alleen uit kankercellen maar ook uittumorstroma, waarin tumorvaten worden gevormd doorangiogenese.
• Tumoren ontstaan niet alleen door overmatige productievan cellen (verhoogde celproliferatie), maar ook dooronvoldoende celversterf (verminderde apoptose). Versto-ring van de functie van celcyclusregulatoren speelt eenbelangrijke rol bij het ontstaan van kanker.
• Metastasering is het belangrijkste kenmerk van kwaadaar-dige gezwelgroei, omdat de metastasen in hoge mate deprognose bepalen.
• De plaats van het ontstaan van tumormetastasen wordtbepaald door de bloedstroom (tumorcellen nestelen zichvooral in het eerstvolgende capillaire vaatbed), specifie-ke binding tussen tumorcellen en endotheelcellen, engroeifactoren uit het ontvangende weefsel.
• De schildwachtklier is de eerste lymfeklier die door een ineen lymfebaan migrerende kankercel wordt bereikt.
• Kanker ontstaat door een combinatie van afwijkingen inhet genoom van de cel.
24 DEEL A ALGEMENE ASPECTEN
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 24

• Carcinogenese is een complex proces dat verschillend ver-loopt voor verschillende tumoren; daarbij kunnen zowelchemische als fysische en biologische carcinogene noxenzijn betrokken.
• Vier groepen van genen spelen een rol bij het ontstaanvan kanker: oncogenen, tumorsuppressorgenen, DNA-her-stelgenen en telomerase.
• Tumorprogressie, het steeds agressiever worden vantumorcellen, is een micro-evolutionair proces waaringenetische instabiliteit van tumorcellen een centrale rolspeelt.
1 FUNDAMENTELE ASPECTEN VAN KANKER 25
Literatuur
Blume-Jensen P, Hunter T. Oncogenic kinase signalling. Nature 2001; 411: 355.Cao Y. Antiangiogenic cancer therapy. Semin Cancer Biol 2004; 14: 139.Cohen SM. Cell proliferation and carcinogenesis. Drug Metab Rev 1998; 30: 339-57.Fearon ER. Human cancer syndromes: Clues to the origin and nature of cancer. Science 1997; 278: 1043.Helman LJ, Meltzer PS. Mechanisms of sarcoma development. Nat Rev Cancer 2003; 3: 685.Hoeijmakers JH. Genome maintenance mechanisms for preventing cancer. Nature 2001; 411: 366.Kim R, Emi M, Tanabe K, et al. The role of Fas ligand and transforming growth factor beta in tumor progression: molecular
mechanisms of immune privilege via Fas-mediated apoptosis and potential targets for cancer therapy. Cancer 2004; 100: 2281-91.Kirkin V, Joos S, Zornig M. The role of Bcl-2 family members in tumorigenesis. Biochim Biophys Acta 2004; 1644: 229.Lengauer C, Kinzler KW, Vogelstein B. Genetic instabilities in human cancers. Nature 1999; 396: 643.Levine AJ. p53, the cellular gatekeeper for growth and division. Cell 1997; 88: 323.Melief CJ, Toes RE, Medema JP, et al. Strategies for immunotherapy of cancer. Adv Immunol 2000; 75: 235.Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherin pathways. Science 2004; 303: 1483.Oving IM, Clevers HC. Molecular causes of colon cancer. Eur J Clin Invest 2002; 32: 448.Pamies RJ, Crawford DR. Tumor markers: an update. Med Clin North Am 1996; 80: 185.Perera FP. Environment and cancer: who are susceptible? Science 1997; 278: 1068.Rauth AM. Radiation carcinogenesis. In: Tannock IF, Hill RP (eds). The basic science of oncology (2nd ed.). New York:
McGraw-Hill, 1992: 119.Rusciano D, Burger M. Why do cancer cells metastasize into particular organs. Bioessays 1992; 14: 185.Sheer D, Squire J. Clinical application of genetic rearrangements in cancer. Semin Cancer Biol 1996; 7: 25.Sherr CJ, McCormick F. The RB and p53 pathways in cancer. Cancer Cell 2002; 2: 103.Tennant R, et al. Chemical carcinogenesis. In: Franks LM, Teich NM (eds). An introduction to the cellular and molecular biology
of cancer (3rd ed.). Oxford: Oxford University Press, 1997: 106.Vogelstein B, Kinzler KW. Cancer genes and the pathways they control. Nat Med 2004; 10: 789-99.Yamasaki L. Role of the RB tumor suppressor in cancer. Cancer Treat Res 2003; 115: 209.Young LS, Murray PG. Epstein-Barr virus and oncogenesis: from latent genes to tumours. Oncogene 2003; 22: 5108.Zink D, Fische AH, Nickerson JA. Nuclear structure in cancer cells. Nat Rev Cancer 2004; 4: 677.
01-Oncologie-H01 t/m H05 21-06-2005 14:25 Pagina 25





























