Embed Size (px)
Citation preview

Abbiamo visto come p53 possa regolare a livello organismico sia la insorgenza di tumori che il mancato differenziamento di alcuni tessuti:
dunque il controllo della progressione del ciclo è legata al differenziamento ed alla trasformazione tumorale.
Abbiamo anche imparato il principio per cui una proteina ha moduli diversi che sono sfruttati da interattori (anche virali)
ORA CI CHIEDIAMO, PERCHE’ LE CELLULE HANNO UNA VITA PREDETERMINATA?

Ras slide

Fuga dalla Senescenza, una cellula può accumulare mutazioni e
diventare immortale

TELOMERI
Il dilemma
Dei Telomeri


Da qua

Cell death
Distinguere necrosi da
apoptosi

Dna laddering nell’apoptosi
TUNEL, teoria
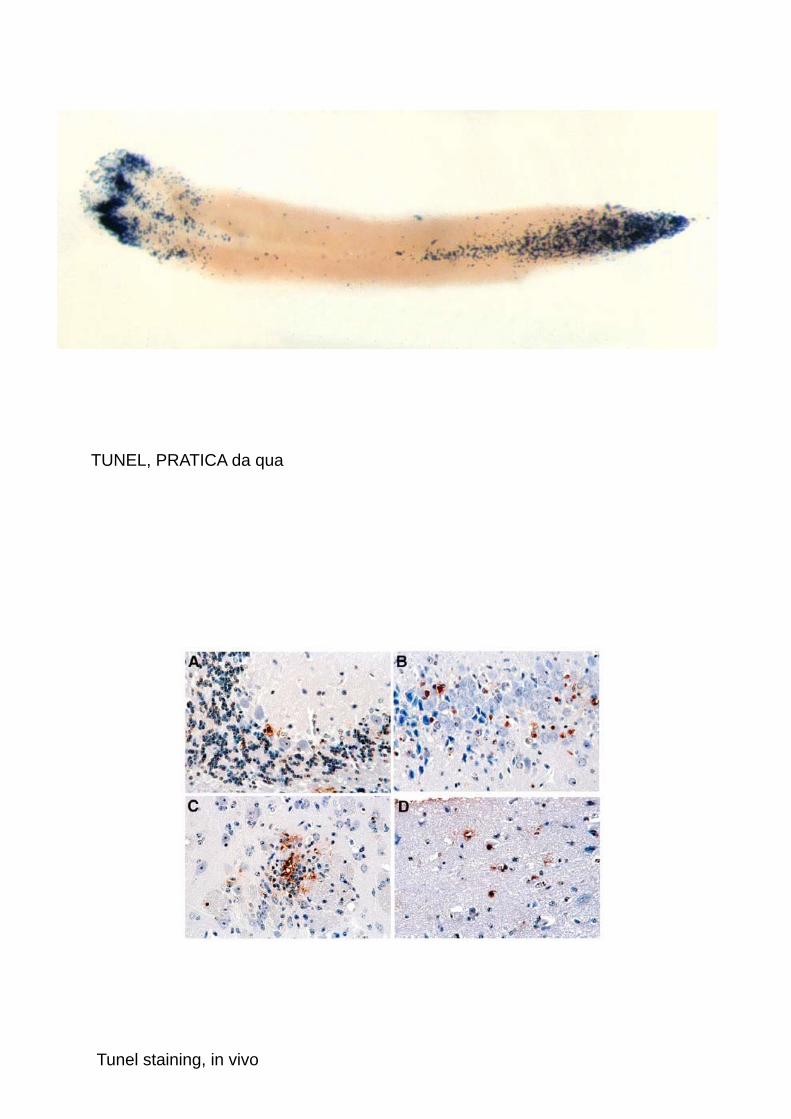
TUNEL, PRATICA da qua
Tunel staining, in vivo

Teoria delle neurotrofine
Desc sympath system
Da qui
Da Montalcini. UN SARCOMA (TU) STIMOLA LA CRESCITA DI FIBRE DEL
SIMPATICO

La storia del NGF
Impianto di NGF nel cervello attira fibre del sistema simpatico
Dipendenza dei neuroni da neurotrofine

LE TIROSINE CHINASI: pathway semplificato
Signaling del recettore trk

Effetto NGF sui neuroni

Effetto neurotrofine sulla morfologia dei neuroni

Table I. Families of structurally related neurotrophic factors and their receptorsNeurotrophic factor family Neurotrophic factor Preferred receptor(s)
--------------------------------------------------------------------------------
Neurotrophins Nerve growth factor (NGF) TrkA, p75NTR Brain-derived neurotrophic factor (BDNF) TrkB, p75NTR
Neurotropin-3 (NT3) TrkC, p75NTR Neurotrophin-4 (NT4) TrkB, p75NTR
GDNF family Glial cell-derived neurotrophic factor (GDNF) Ret, GFR-1 Neurturin Ret, GFR-2 Artemin Ret, GFR-3
Persephin Ret, GFR-4 Neurotrophic cytokines Ciliary neurotrophic factor (CNTF) gp130, LIFRß, CNTFR
Leukaemia-inhibitory factor (LIF) gp130, LIFRß Cardiotrophin-1 (CT-1) gp130, LIFRß Oncostatin-M (OSM) gp130, OSMRß
Interleukin-6 (IL-6) gp130, IL6R HGF family Hepatocyte growth factor (HGF) Met
Macrophage-stimulating protein (MSP) Ron
--------------------------------------------------------------------------------
Although several neurotrophic factors bind and activate more than one member of a family of receptors (e.g. NT3 activates TrkA and TrkB in addition to its preferred receptor tyrosine kinase TrkC), for simplicity, only the
preferred receptors for each factor are listed.
Ch!!!!
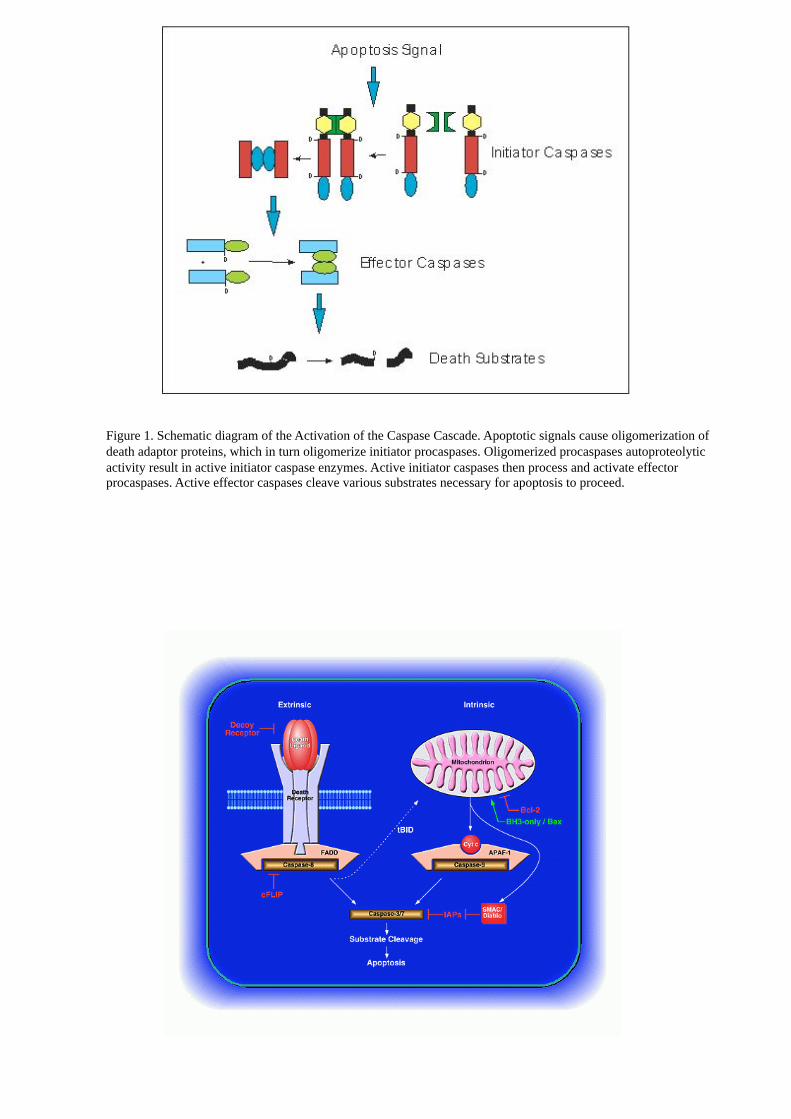
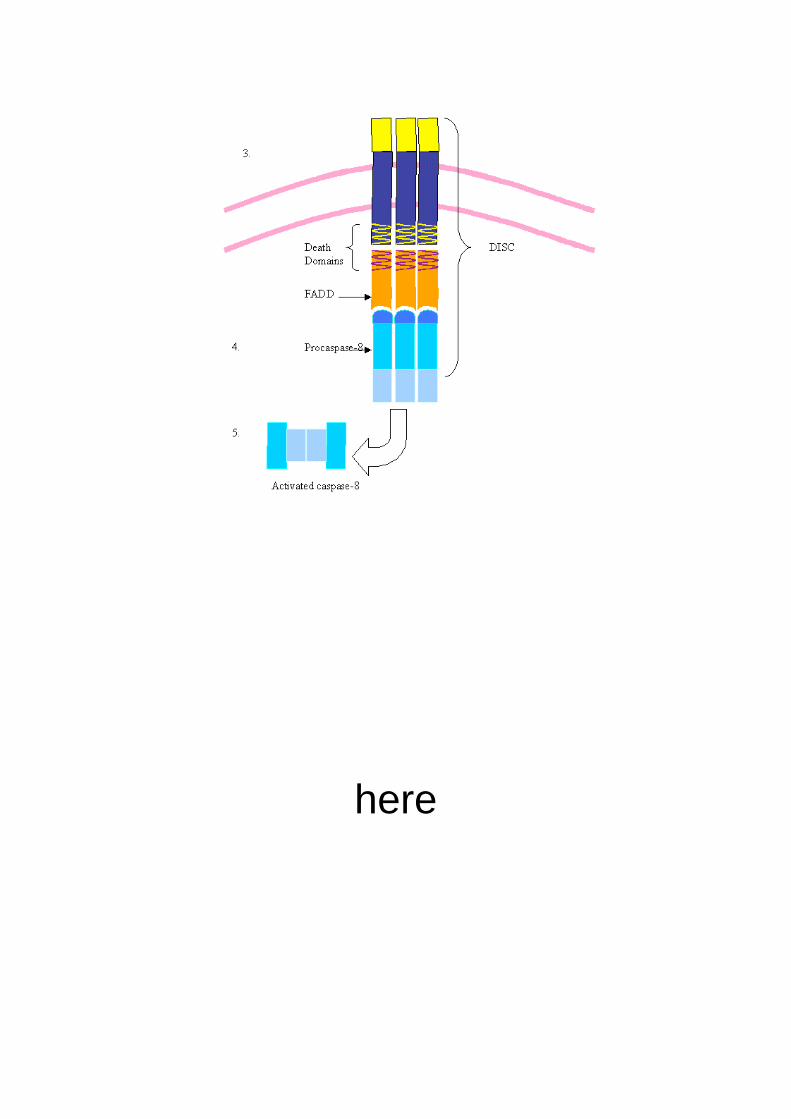
Figure 1. Schematic diagram of the Activation of the Caspase Cascade. Apoptotic signals cause oligomerization of
death adaptor proteins, which in turn oligomerize initiator procaspases. Oligomerized procaspases autoproteolytic
activity result in active initiator caspase enzymes. Active initiator caspases then process and activate effector
procaspases. Active effector caspases cleave various substrates necessary for apoptosis to proceed.

intrinseca

MECCANISMI MOLECOLARI DELLA APOPTOSI:
CARATTERIZZAZIONE BIOCHIMICA, INTRINSECA ED ESTRINSECA
RUOLO DELLE CASPASI
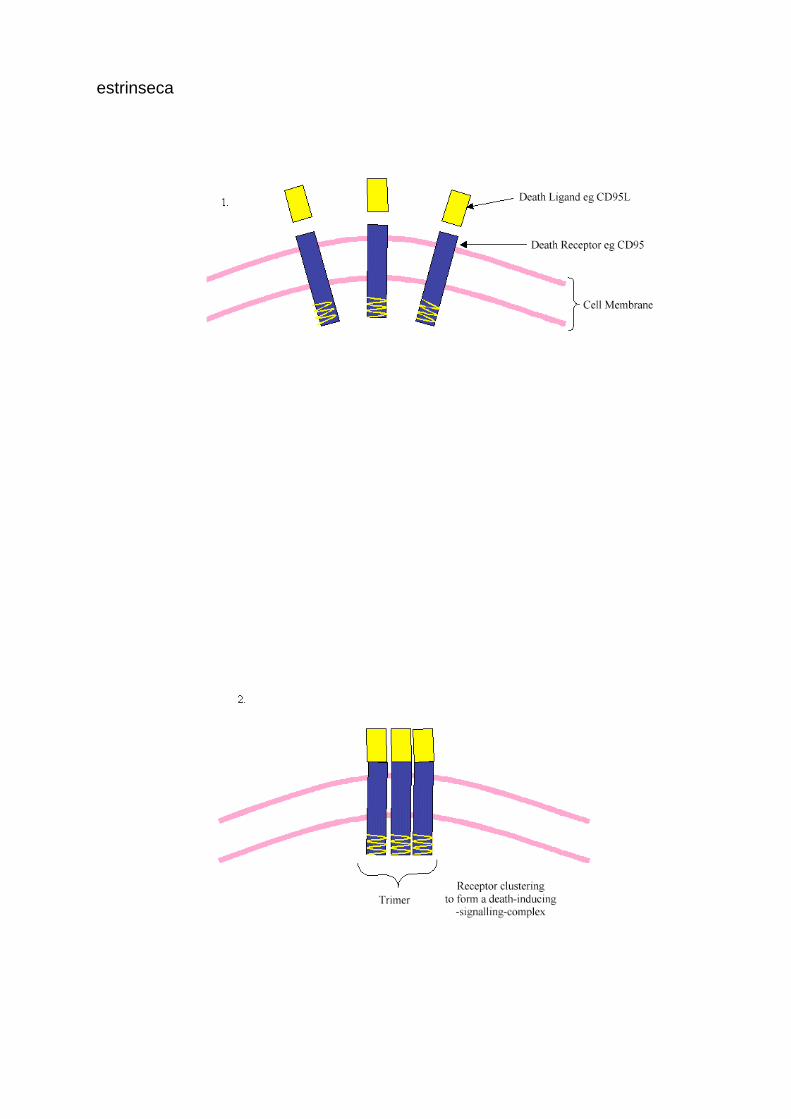
estrinseca
here


LINEAGE

LINEAGE
SEMPLI
FICATO
From here

The nuc-1 gene encodes a DNase II homolog similar to
mammalian and Drosophila DNaseII enzymes and is
required for DNA degradation during apoptosis as well
as for degradation of dietary DNA during normal
feeding; during apoptosis, NUC-1 functions in
apoptotic cells at an intermediate stage of DNA
degradation, after the killing step, but prior to cell-
corpse engulfment
Isolation of new ced mutations: Because living
animals with undegraded cell corpses can be
recognized
using Nomarski optics but appear normal in
general morphology and behavior (HEDGECOCK,
SULSTON
and THOMSON 1983), we used Nomarski microscopy
to screen for new ced mutants.

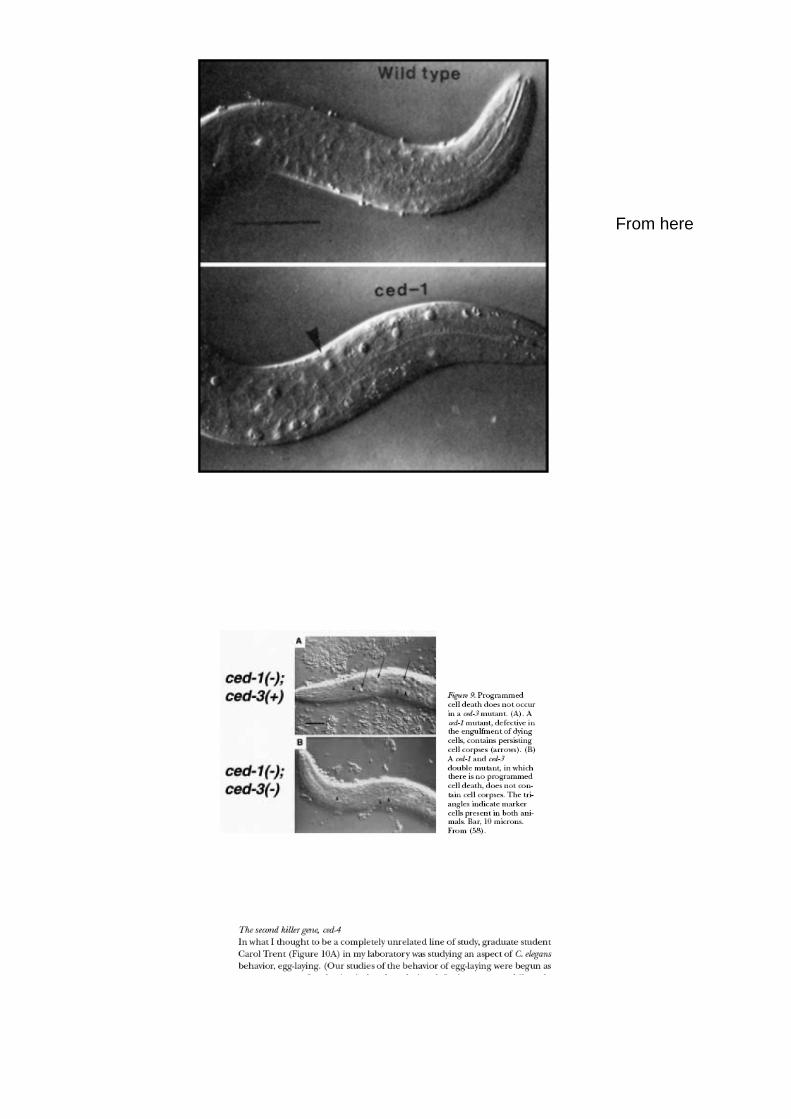
Nel mutante Ced-1, le Cellule non sono fagocitate dopo la morte,
I cadaveri rimangono
Ced-3, Ced-1 double mutant, NO CADAVERI
= Primo pathway apoptotico determinato geneticamente
The Caenorhabditis elegans gene ced-9 prevents cells from undergoing programmed cell death and
encodes a protein similar to the mammalian cell-death inhibitor Bcl-2.
We have cloned the C. elegans cell death gene ced-3. A ced-3 transcript is most abundant
during embryogenesis, the stage during which most programmed cell deaths occur. The
predicted CED-3 protein shows similarity to human and murine interleukin-1 beta-
converting enzyme and to the product of the mouse nedd-2 gene, which is expressed in the
embryonic brain. The sequences of 12 ced-3 mutations as well as the sequences of ced-3 genes
from two related nematode species identify sites of potential functional importance. We
propose that the CED-3 protein acts as a cysteine protease
Here we identify a new gene, dark, which encodes a Drosophila homologue of mammalian Apaf-1 and Caenorhabditis elegans CED-4, cell-
death proteins. Like Apaf-1, but in contrast to CED-4, Dark contains a carboxy-terminal WD-repeat domain necessary for interactions with
the mitochondrial protein cytochrome c. Dark selectively associates with another protein involved in apoptosis, the fly apical caspase,
Dredd. Dark-induced cell killing is suppressed by caspase-inhibitory peptides and by a dominant-negative mutant Dredd protein, and
enhanced by removal of the WD domain.
In the nematode C. elegans, genetic studies led to the discovery of 15 genes that function in
programmed cell death 1 (Fig. 1). These 15 genes have been divided into four groups based on
the order of their activity during the process of programmed cell death: (1) those involved in
the decision making (ces-1 and ces-2); (2) in the process of execution (ced-3, ced-4, ced-9 and
egl-1); (3) in the engulfment of dying cells by engulfing cells (ced-1, ced-2, ced-5, ced-6, ced-7,
ced-10, ced-12); and (4) those in the degradation of cell corpses within engulfing cells (nuc-1).
Da Horvitz, articolo originale