Embed Size (px)
Citation preview

DNA Damage and Repair
DNADamage-InducibleGene,ReprimoFunctions asaTumorSuppressor and Is Suppressed by Promoter Methylation inGastric Cancer
Akira Ooki1,2, Keishi Yamashita1, Kensei Yamaguchi2, Anupom Mondal3, Hiroshi Nishimiya1, andMasahiko Watanabe1
AbstractIn several types of human cancer, the gene expression of Reprimo, a highly glycosylated protein, is
frequently silenced via methylation of its promoter. The aim of this study was to characterize the epigeneticinactivation of Reprimo and its biologic function and clinical relevance in gastric cancer. The correlationbetween Reprimo methylation and clinical relevance was assessed in 83 primary human gastric cancer tissues.The effects of Reprimo expression were also examined using in vitro and in vivo assays. Reprimo methylationwas cancer specific and frequently observed. In two gastric cancer cell lines without Reprimo methylation, weobserved faint or weak Reprimo expression under normal conditions and high expression under DNA-damaging conditions. In four gastric cancer cell lines with Reprimo methylation, however, Reprimoexpression remained faint even under DNA-damaging conditions, with expression being restored incombination with agents that induce demethylation. Enforced Reprimo expression robustly inhibited cellproliferation and anchorage-independent colony formation and enhanced DNA damage-induced apoptosis.Inverse effects were observed via siRNA-mediated knockdown of endogenous Reprimo. Reprimo expressioninhibited tumorigenesis in vivo. Reprimo methylation was also associated with a poor response in patientswith gastric cancer treated with chemotherapy (P1/4 0.028), and a poor prognosis in patients with advancedgastric cancer (P1/4 0.03). In conclusion, Reprimo expression is normally induced in response to DNAdamage, acting as a novel tumor suppressor in gastric cancer. However, Reprimo methylation abrogates itsexpression and effects. The clinical assessment of Reprimo promoter methylation may serve not only as apredictive marker for chemotherapy, but also as a marker for tumor aggressiveness. Mol Cancer Res; 11(11);1362–74. �2013 AACR.
IntroductionGastric cancer remains a major clinical challenge, being
both the fourthmost common cancer and the second leadingcause of cancer-related death worldwide (1). Globally, a totalof 989,600 new cases and 738,000 deaths are estimated tohave occurred in 2008. The case-fatality ratio of gastriccancer is higher than that of other common malignancies,such as colorectal, breast, or prostate cancer (2). However,although recent improvements in diagnostic tools and
methods have facilitated the detection of gastric cancer atan early stage, advanced gastric cancer is still associatedwith apoor clinical outcome due to the limited success of surgeryand chemotherapy.Accumulating evidence suggests that cancer is caused by
both epigenetic and genetic abnormalities (3). The epige-netic pathway involved in the development of cancer isdetermined by chromatin structure, including DNA meth-ylation, histone modifications, or noncoding regulatoryRNAs (4). DNA methylation occurs at the fifth carbon ofthe cytosine, termed a dinucleotide CpG, which precedes aguanine. When methylation occurs within dinucleiotideCpG-rich regions, "CpG islands," of a gene promoter, it isassociated with a compact chromatin structure and is accom-panied by transcriptional silencing of the affiliated gene,especially tumor-suppressor genes (5). DNA methylationoccurs more frequently than genetic mutations in gastriccancer and has recently opened the exciting doors fordeveloping cancer biomarkers and therapeutic targets (6–8).Reprimo is a highly glycosylated protein localized predom-
inantly in the cytoplasm, which was identified by differentialdisplay screening of genes distinctly expressed betweenX-ray–
Authors' Affiliations: 1Department of Surgery, Kitasato University Hos-pital, Kitasato, Sagamihara, Kanagawa; 2Department of Gastroenterology;and 3Research Institute for Clinical Oncology, Saitama Cancer Center,Komuro, Ina Machi, Kita-Adachi-gun, Saitama, Japan
Note: Supplementary data for this article are available at Molecular CancerResearch Online (http://mcr.aacrjournals.org/).
Corresponding Author: Akira Ooki, Department of Surgery, KitasatoUniversity School of Medicine, Kitasato 1-15-1, Sagamihara 228-8555,Kanagawa, Japan. Phone: 81-42-778-8735; Fax: 81-42-778-8735; E-mail:[email protected]
doi: 10.1158/1541-7786.MCR-13-0091
�2013 American Association for Cancer Research.
MolecularCancer
Research
Mol Cancer Res; 11(11) November 20131362
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091

irradiated wild-type and p53-deficient mouse embryonicfibroblast cells (9). Transiently enforced Reprimo expressioninduces G2 arrest of the cell cycle by inhibiting Cdc2 activityand nuclear translocation of the Cdc2–cyclin B1 complex invarious cell lines such as DLD1, HeLa, and MCF7 cells (9),whereas loss of Reprimo expression is associated with anincrease in uterine sarcoma incidence in mutant p53 trans-genic mice treated with the carcinogen 1,2-dimethylhydra-zine (10). These findings suggest that both the expression andfunction of Reprimo as a tumor-suppressor gene are regulatedin a p53-dependent manner. In contrast, Reprimo promotermethylation has been reported to be involved in an alternativemechanism that regulatesReprimo expression in various typesof tumors (11–13). Furthermore, the regulation of Reprimoexpression is independent of p53 status in neuronal cellstreated with copper, with neuronal damage being associatedwith Wilson disease (14). In gastric cancer, Reprimo pro-moter methylation is a frequent event as we and others havepreviously reported (11, 13, 15). The relationship betweenReprimo and gastric cancer, however, has never been exam-ined from a mechanistic and functional point of view.In the present study, we examined the clinical significance
of the Reprimo gene, especially for its promoter methylationin primary gastric cancer and assessed the functional changeassociated with its abnormality.
Materials and MethodsCell lines and tissue samplesThe gastric cancer cell line MKN7 was provided by the
Cell Resource Center for Biomedical Research Institute ofDevelopment, Aging and Cancer, Tohoku University inDecember 2006 (Miyagi, Japan). Seven other gastric cancercell lines (GCIY, AZ521, KatoIII, SH10, H111, MKN74,and NUGC4) were purchased from RIKEN BioResourceCenter in December 2006. All cell lines were grown inRPMI-1640 medium (GIBCO) supplemented with 10%FBS. We confirmed the authenticity of the cell lines usingshort tandem repeat profiling in June 2013. UV irradiation(10 mJ/cm2) was conducted without a medium using aStratalinker UVCrosslinker (Agilent Technologies) to causeDNA damage. The cells were then incubated in mediumwith 10% FBS for 24 hours and analyzed. The DNA-damaging agents cisplatin (CDDP) and 5-fluorouracil (5-FU) were purchased from Kyowa Hakko.Ten pairs of frozen primary human gastric cancer tissues
and corresponding normal mucosal tissues, obtained at least5 cm from the tumor edge, were analyzed as the preliminarydata set for expression and methylation status. Formalin-fixed, paraffin-embedded tissue samples were collected from83 patients with primary gastric cancer who underwent agastrectomy at the Kitasato University Hospital (Kanagawa,Japan) in accordance with the gastric cancer treatmentguidelines in Japan (16). Informed consent was obtainedfrom all patients before sample collection. All histopatho-logical findings were examined in accordance with the 14thedition of the Japanese Classification of Gastric Carcinoma(JCGC; ref. 17). Advanced gastric cancer is defined as cancer
invading the muscularis propria or deeper, irrespective of thepresence of lymph nodemetastases in JCGC. Tumor–node–metastasis classification was made in accordance with theseventh edition of the American Joint Committee on Can-cer/International Union Against Cancer (AJCC/UICC)staging system. Detailed information on the 83 patients issummarized in Table 1. This study was conducted withapproval of the Ethics Committee of Kitasato University.
Bisulfite treatment of DNA and sequencing analysisTissue sections from the tumor and the corresponding
normal mucosa were sharply dissected on hematoxylin andeosin-stained slides, and genomic DNA was subsequentlyextracted using a QIAamp DNA FFPE Kit (QIAGENSciences). Genomic DNA from cell lines and frozen tissueswere extracted using a QIAamp DNAMini Kit (QIAGEN).Bisulfite treatment was conducted using an EpiTect BisulfiteKit (QIAGEN) andwas subsequently amplified via PCR.ThePCR conditions and sequences of primers and probes used inthis study are shown in Supplementary Table S1. Primersequences for Reprimo promoter region were designed torecognize DNA alterations caused by bisulfite treatment. ThePCR products were purified and then inserted into a pCR4-TOPO vector using a TOPOTACloning Kit for sequencing(Invitrogen). Ten clones were selected for each sample andthen sequenced using a Big Dye Terminator v3.1 CycleSequencing Kit (Applied Biosystems). For analysis of p53mutation, genomic DNA was also sequenced using primersfor exons 4 to 9 of the p53 gene (18).
Quantitative methylation-specific PCRTo quantify the methylation level of Reprimo, quan-
titative methylation-specific PCR (Q-MSP) was con-ducted via an iQ Supermix (Bio-Rad Laboratories) intriplicate on the iCycler iQ Real-Time PCR Detectionsystem (Bio-Rad Laboratories). Serial dilutions of CpGe-nome universal methylated DNA (Chemicon Interna-tional, Inc.,) were used to construct the calibration curvefor each plate. CpGenome universal methylated DNAand unmethylated DNA (Chemicon) were used as amethylation positive and a negative control, respectively.The methylation value was defined by the ratio of fluo-rescence intensity emitted from the amplified bisulfiteDNA of the CpG islands of Reprimo gene divided by thatof b-actin and multiplied by 100 (Q-MSP value). Theoptimal cutoff value for Q-MSP was determined using areceiver operating characteristic (ROC) curve.
Quantitative reverse transcriptase PCRTotal RNA was isolated using TRIzol Reagent (Invitro-
gen) in accordance with the manufacturer's protocol andreverse transcribed with a SuperScript III Reverse Transcrip-tase kit (Invitrogen). Quantitative reverse transcriptase PCR(qRT-PCR) was conducted to quantify the expression levelof Reprimo gene using the iCycler iQ Real-Time PCRDetection System (Bio-Rad). Relative quantitative analysisnormalized to b-actin was conducted via the comparativecycle threshold method (19).
Reprimo in Gastric Cancer
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1363
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091
5-Aza-20-deoxycytidine treatmentCells (1� 106/T-75 flask) were treated with 5 mmol/L of
the demethylating agent 5-aza-20-deoxycytidine (5-Aza-dC;Sigma-Aldrich) dissolved in 50% acetic acid or were mocktreated with PBS including the same amount of acetic acidevery 24 hours for 5 days (20).
Western blot analysisWhole-cell lysates were extracted in radioimmunoprecipi-
tation assay buffer (Pierce) supplemented with 10 mL/mLHalt Protease Inhibitor Cocktail Kit (Pierce) and HaltPhosphatase Inhibitor Cocktail Kit (Pierce). The proteinconcentrations were determined using a Coomassie Plus-The
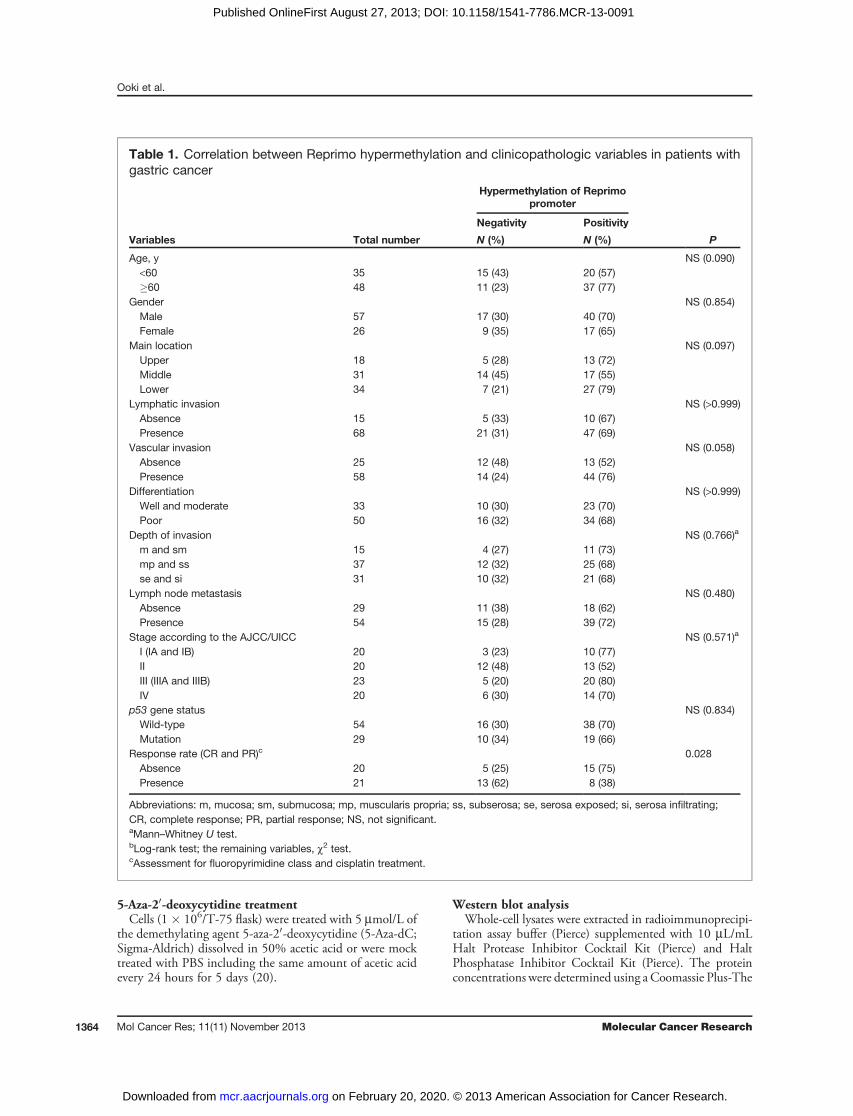
Table 1. Correlation between Reprimo hypermethylation and clinicopathologic variables in patients withgastric cancer
Hypermethylation of Reprimopromoter
Negativity Positivity
Variables Total number N (%) N (%) P
Age, y NS (0.090)<60 35 15 (43) 20 (57)�60 48 11 (23) 37 (77)
Gender NS (0.854)Male 57 17 (30) 40 (70)Female 26 9 (35) 17 (65)
Main location NS (0.097)Upper 18 5 (28) 13 (72)Middle 31 14 (45) 17 (55)Lower 34 7 (21) 27 (79)
Lymphatic invasion NS (>0.999)Absence 15 5 (33) 10 (67)Presence 68 21 (31) 47 (69)
Vascular invasion NS (0.058)Absence 25 12 (48) 13 (52)Presence 58 14 (24) 44 (76)
Differentiation NS (>0.999)Well and moderate 33 10 (30) 23 (70)Poor 50 16 (32) 34 (68)
Depth of invasion NS (0.766)a
m and sm 15 4 (27) 11 (73)mp and ss 37 12 (32) 25 (68)se and si 31 10 (32) 21 (68)
Lymph node metastasis NS (0.480)Absence 29 11 (38) 18 (62)Presence 54 15 (28) 39 (72)
Stage according to the AJCC/UICC NS (0.571)a
I (IA and IB) 20 3 (23) 10 (77)II 20 12 (48) 13 (52)III (IIIA and IIIB) 23 5 (20) 20 (80)IV 20 6 (30) 14 (70)
p53 gene status NS (0.834)Wild-type 54 16 (30) 38 (70)Mutation 29 10 (34) 19 (66)
Response rate (CR and PR)c 0.028Absence 20 5 (25) 15 (75)Presence 21 13 (62) 8 (38)
Abbreviations: m, mucosa; sm, submucosa; mp, muscularis propria; ss, subserosa; se, serosa exposed; si, serosa infiltrating;CR, complete response; PR, partial response; NS, not significant.aMann–Whitney U test.bLog-rank test; the remaining variables, c2 test.cAssessment for fluoropyrimidine class and cisplatin treatment.
Ooki et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1364
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091

Better Bradford Assay Kit (Pierce), separated on NuPAGE�
4% to 12% Bis-Tris Gel (Invitrogen) with anti-flag-V5mouse monoclonal antibody (Sigma-Aldrich) and anti-b-actin mouse monoclonal antibody (Sigma-Aldrich).
Plasmid transfection and siRNA transfectionA full-length complementary DNA (cDNA) of Reprimo
was isolated from GCIY cells using PCR with Platinum PfxDNA Polymerase (Invitrogen). The plasmid expressingReprimo (pcDNA 3.1-Reprimo) was obtained by cloningthe PCR products into the pcDNA 3.1D/V5-His-TOPOvector via a pcDNA 3.1 Directional TOPO Expression Kit(Invitrogen). The sequence of the cloned Reprimo cDNAwas verified by sequencing analysis, and an empty vectorwith self-ligation (pcDNA 3.1-mock) was used as a control.Cells were transfected using Lipofectamine 2000 Reagent(Invitrogen) in OPTI-MEM medium (Invitrogen). Stableclones with Reprimo or mock were established by G418(GIBCO) selection. For siRNA transfection, cells (2� 105)were seeded in 6-well plates 24 hours before transfection.The cells were transfected with 1 mmol/L Accell SMART-pool, siRNA-Reprimo (Thermo Fisher Scientific) mixedwith Accell siRNA Delivery Media (Thermo Fisher Scien-tific) according to the Thermo Scientific Dharmacon AccellsiRNA Delivery Protocol. The Accell Non-targeting Pool(siRNA-ctr) and Accell siRNA Delivery Media alone(siRNA-mock) were used as a control for nonsequence-specific effects and as a mock treatment, respectively.
Anchorage-independent colony formation assayAnchorage-independent cell growth was analyzed by plat-
ing 0.36% top agarose (Bacto Agar; Becton, Dickinson andCompany) containing 1 � 105 cells on a surface of 0.72%bottom agarose in the 6-well plates. Cells were fed weekly byoverlying fresh soft-agar solution, and colonies were photo-graphed after 2 weeks of incubation. The experiment wascarried out in triplicate.
Proliferation assay and invasion assayThe effects of Reprimo expression on cell proliferation and
viability (2� 103 cells/well) weremeasured using the PremixWST-1Cell ProliferationAssay System (Takara Bio, Co.,) in96-well plates. The invasion assay was conducted in the 24-well BD BioCoat Matrigel Invasion Chamber (BD Bios-ciences Discovery Labware). Forty-eight hours after trans-fection, the lower chamber was filled with 750 mL Dulbec-co's modified Eagle medium (DMEM; GIBCO) supple-mented with 10% FBS as a chemoattractant, and cells (5 �105/well) were then seeded into an upper chamber in 500mLof serum-free DMEM. After incubation for 22 hours, themembrane of the upper chamber was fixed and stained usingDiff-Quik reagent (Sysmex, Co.,). Cells that had invadedthrough the membrane were counted in four separated fieldsper well. Both experiments were carried out in triplicate.
Cell-cycle and apoptosis assaysCell-cycle and apoptosis assays were conducted using the
Guava PCA System (Guava Technologies, Inc.,). In the cell-
cycle assay, after synchronization for 48 hours in serum-freemedium, the cells were then cultured in complete medium.After 24 hours, the cells (2 � 105) were then fixed in 75%ethanol, and stained with propidium iodide (Guava CellCycle Reagent). In the apoptosis assay, the harvested cells(1� 105) were stained with Annexin V and 7-AAD (GuavaNexin Reagent) for discrimination of early and late apoptoticcells, respectively. Both experiments were carried out intriplicate and analyzed usingCytoSoft 2.1.5 software (GuavaTechnologies).
Mouse xenograft assayFemale BALB/cAJcl nude mice ages 5 weeks were pur-
chased from CLEA Japan Inc. Mice were subcutaneouslyinjected with AZ521 patient cells or AZ521 stable cellstransfected with pcDNA 3.1-Reprimo or pcDNA 3.1-mockon both flanks (2.5 � 106 cells per flank). In the treatmentcombinedwith CDDP and 5-FU,mice with tumors approx-imately 5 mm in diameter received intraperitoneal admin-istration of CDDP (0.1 mg/kg) and 5-FU (10 mg/kg) oncedaily for 7 days (21). Each group consisted of 3mice (n¼ 3).Tumor volumes were calculated from caliper measurementsof two orthogonal diameters [larger (x)] and smaller (y)diameters] using the following formula: volume ¼ xy2/2.All animal experiments were carried out in strict accordancewith the guidelines for animal experiments of the SaitamaCancer Center (Saitama, Japan).
Statistical analysisCategorical variables were analyzed via c2 test or Mann–
Whitney U test, and continuous variables were analyzed viaStudent t test. For continuous variables, the data areexpressed as mean� SD. The disease-specific survival (DSS)was measured from the date of surgery to the date of death orthe last follow-up. DSS was estimated via the Kaplan–Meiermethod, and values were compared using the log-rank test.P < 0.05 was considered to indicate statistical significance.All statistical analyses were conducted with the StatisticalAnalysis System (SAS) software package (SAS Institute).
ResultsCancer-specific Reprimo promoter methylation ingastric cancerThe location of CpG islands in the 50-flanking region of
Reprimo genomic sequencing is shown in Fig. 1A. TheReprimo promoter harbors CpG islands encompassing thetranscription start site, and we previously reported thatReprimo harbored its promoter methylation in 75% (6/8)of gastric cancer cell lines, 80% (8/10) of primary humangastric cancer tissues, and 10% (1/10) of correspondingnormal tissues in the preliminary dataset of 10matched pairsby bisulfite sequencing analysis (15). Q-MSP can achieve amuch more objective and specific assessment of a smallamount of genomic DNA through PCR amplification usingmethylation-specific primers and fluorescent probes (22).Q-MSP analysis was therefore conducted using the samesamples (15) to confirm the above findings on bisulfite
Reprimo in Gastric Cancer
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1365
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091
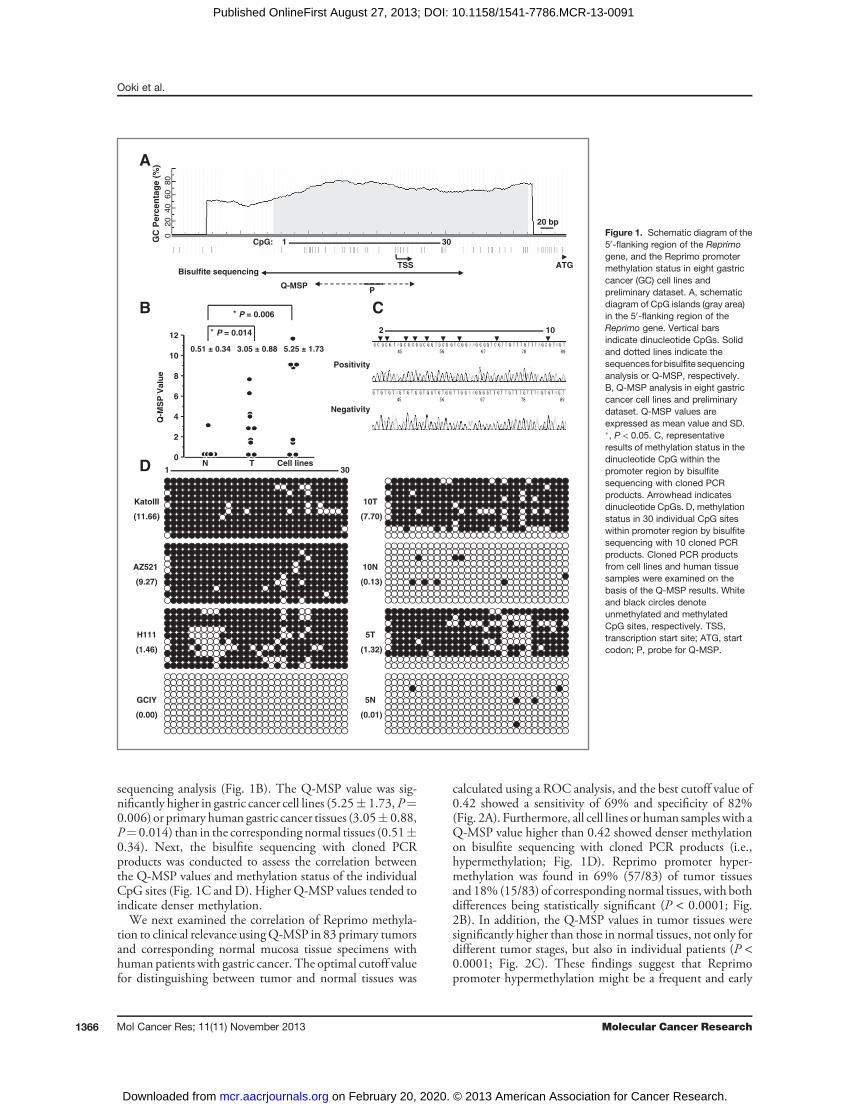
sequencing analysis (Fig. 1B). The Q-MSP value was sig-nificantly higher in gastric cancer cell lines (5.25� 1.73, P¼0.006) or primary human gastric cancer tissues (3.05� 0.88,P¼ 0.014) than in the corresponding normal tissues (0.51�0.34). Next, the bisulfite sequencing with cloned PCRproducts was conducted to assess the correlation betweenthe Q-MSP values and methylation status of the individualCpG sites (Fig. 1C andD). Higher Q-MSP values tended toindicate denser methylation.We next examined the correlation of Reprimo methyla-
tion to clinical relevance usingQ-MSP in 83 primary tumorsand corresponding normal mucosa tissue specimens withhuman patients with gastric cancer. The optimal cutoff valuefor distinguishing between tumor and normal tissues was
calculated using a ROC analysis, and the best cutoff value of0.42 showed a sensitivity of 69% and specificity of 82%(Fig. 2A). Furthermore, all cell lines or human samples with aQ-MSP value higher than 0.42 showed denser methylationon bisulfite sequencing with cloned PCR products (i.e.,hypermethylation; Fig. 1D). Reprimo promoter hyper-methylation was found in 69% (57/83) of tumor tissuesand 18% (15/83) of corresponding normal tissues, with bothdifferences being statistically significant (P < 0.0001; Fig.2B). In addition, the Q-MSP values in tumor tissues weresignificantly higher than those in normal tissues, not only fordifferent tumor stages, but also in individual patients (P <0.0001; Fig. 2C). These findings suggest that Reprimopromoter hypermethylation might be a frequent and early
20 bp
ATG
CpG: 1 30 GC
Pe
rce
nta
ge
(%
)
P
TSSBisulfite sequencing
Q-MSP
N T Cell lines
Q-M
SP
Valu
e
5N
(0.01)
10N
(0.13)
H111
(1.46)
AZ521
(9.27)
KatoIII
(11.66)
GCIY
(0.00)
5T
(1.32)
10T
(7.70)
A
D 1 30
Positivity
Negativity
2 10
CB
0.51 ± 0.34 3.05 ± 0.88 5.25 ± 1.73
* P = 0.014
* P = 0.006
0
2
4
6
8
10
12
020
4060
80
Figure 1. Schematic diagram of the50-flanking region of the Reprimogene, and the Reprimo promotermethylation status in eight gastriccancer (GC) cell lines andpreliminary dataset. A, schematicdiagram of CpG islands (gray area)in the 50-flanking region of theReprimo gene. Vertical barsindicate dinucleotide CpGs. Solidand dotted lines indicate thesequences for bisulfite sequencinganalysis or Q-MSP, respectively.B, Q-MSP analysis in eight gastriccancer cell lines and preliminarydataset. Q-MSP values areexpressed as mean value and SD.�, P < 0.05. C, representativeresults of methylation status in thedinucleotide CpG within thepromoter region by bisulfitesequencing with cloned PCRproducts. Arrowhead indicatesdinucleotide CpGs. D, methylationstatus in 30 individual CpG siteswithin promoter region by bisulfitesequencing with 10 cloned PCRproducts. Cloned PCR productsfrom cell lines and human tissuesamples were examined on thebasis of the Q-MSP results. Whiteand black circles denoteunmethylated and methylatedCpG sites, respectively. TSS,transcription start site; ATG, startcodon; P, probe for Q-MSP.
Ooki et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1366
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091
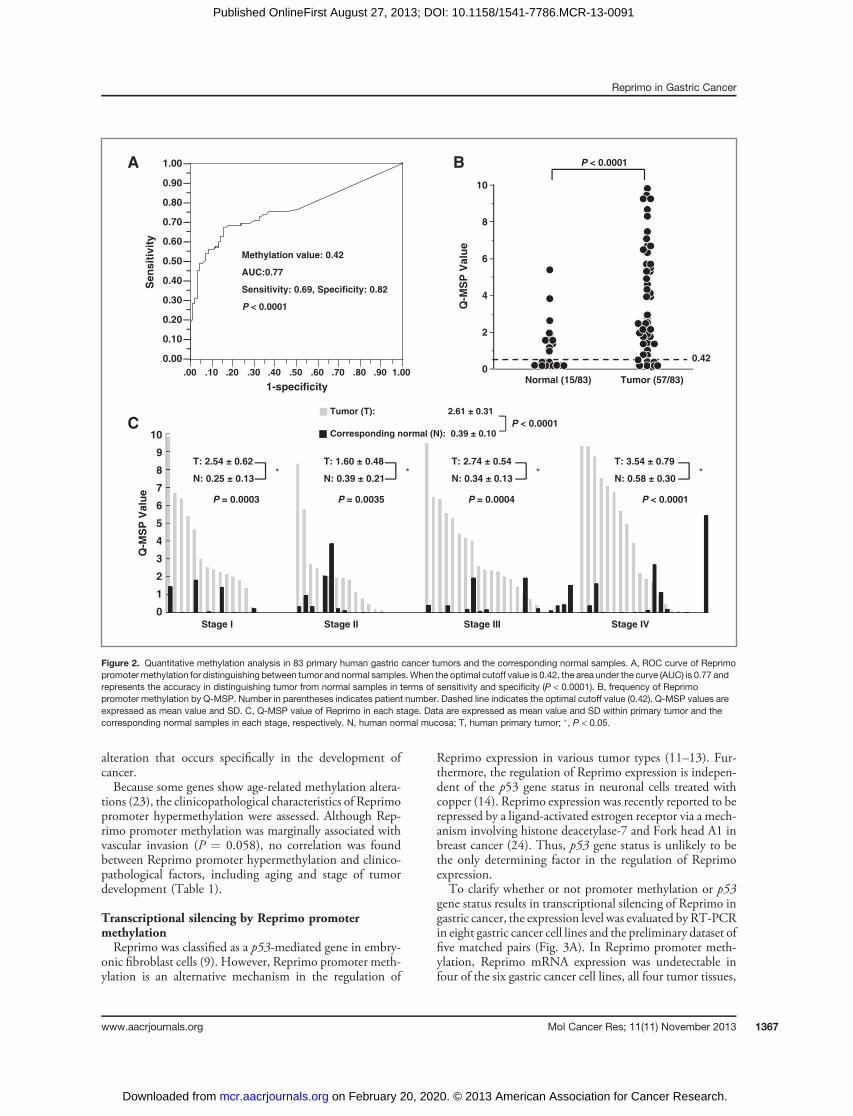
alteration that occurs specifically in the development ofcancer.Because some genes show age-related methylation altera-
tions (23), the clinicopathological characteristics of Reprimopromoter hypermethylation were assessed. Although Rep-rimo promoter methylation was marginally associated withvascular invasion (P ¼ 0.058), no correlation was foundbetween Reprimo promoter hypermethylation and clinico-pathological factors, including aging and stage of tumordevelopment (Table 1).
Transcriptional silencing by Reprimo promotermethylationReprimo was classified as a p53-mediated gene in embry-
onic fibroblast cells (9). However, Reprimo promoter meth-ylation is an alternative mechanism in the regulation of
Reprimo expression in various tumor types (11–13). Fur-thermore, the regulation of Reprimo expression is indepen-dent of the p53 gene status in neuronal cells treated withcopper (14). Reprimo expression was recently reported to berepressed by a ligand-activated estrogen receptor via a mech-anism involving histone deacetylase-7 and Fork head A1 inbreast cancer (24). Thus, p53 gene status is unlikely to bethe only determining factor in the regulation of Reprimoexpression.To clarify whether or not promoter methylation or p53
gene status results in transcriptional silencing of Reprimo ingastric cancer, the expression level was evaluated byRT-PCRin eight gastric cancer cell lines and the preliminary dataset offive matched pairs (Fig. 3A). In Reprimo promoter meth-ylation, Reprimo mRNA expression was undetectable infour of the six gastric cancer cell lines, all four tumor tissues,
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
.00 .10 .20 .30 .40 .50 .60 .70 .80 .90 1.00
Methylation value: 0.42
AUC:0.77
Sensitivity: 0.69, Specificity: 0.82
P < 0.0001
1-specificity
Se
ns
itiv
ity
BA
C
0.42
Normal (15/83) Tumor (57/83)
Q-M
SP
Va
lue
P < 0.0001
Stage IVStage IIIStage IIStage I
Q-M
SP
Valu
e
T: 2.54 ± 0.62
N: 0.25 ± 0.13
T: 3.54 ± 0.79
N: 0.58 ± 0.30
T: 2.74 ± 0.54
N: 0.34 ± 0.13
T: 1.60 ± 0.48
N: 0.39 ± 0.21
2.61 ± 0.31Tumor (T):
Corresponding normal (N): 0.39 ± 0.10P < 0.0001
0
2
4
6
8
10
P = 0.0003 P = 0.0004 P < 0.0001P = 0.0035
10
9
8
7
6
5
4
3
2
1
0
Figure 2. Quantitative methylation analysis in 83 primary human gastric cancer tumors and the corresponding normal samples. A, ROC curve of Reprimopromotermethylation for distinguishing between tumor and normal samples.When the optimal cutoff value is 0.42, the area under the curve (AUC) is 0.77 andrepresents the accuracy in distinguishing tumor from normal samples in terms of sensitivity and specificity (P < 0.0001). B, frequency of Reprimopromoter methylation by Q-MSP. Number in parentheses indicates patient number. Dashed line indicates the optimal cutoff value (0.42). Q-MSP values areexpressed as mean value and SD. C, Q-MSP value of Reprimo in each stage. Data are expressed as mean value and SD within primary tumor and thecorresponding normal samples in each stage, respectively. N, human normal mucosa; T, human primary tumor; �, P < 0.05.
Reprimo in Gastric Cancer
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1367
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091
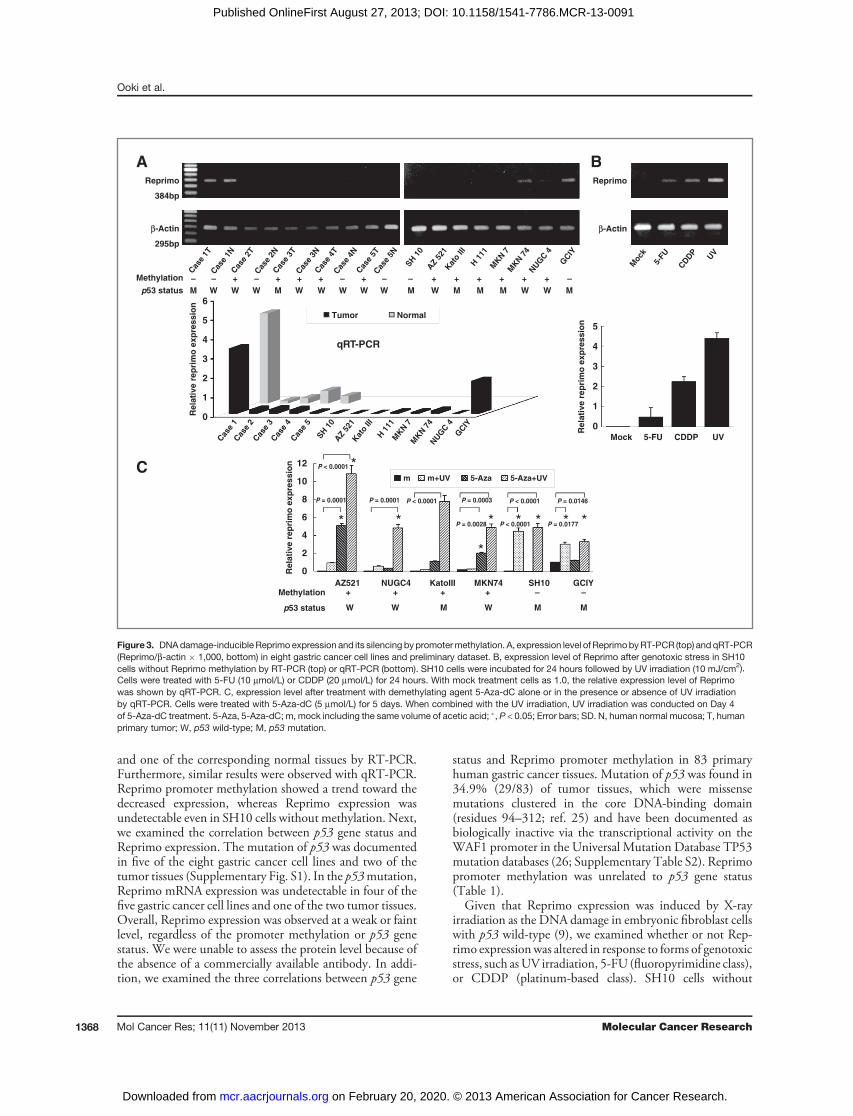
and one of the corresponding normal tissues by RT-PCR.Furthermore, similar results were observed with qRT-PCR.Reprimo promoter methylation showed a trend toward thedecreased expression, whereas Reprimo expression wasundetectable even in SH10 cells without methylation. Next,we examined the correlation between p53 gene status andReprimo expression. The mutation of p53 was documentedin five of the eight gastric cancer cell lines and two of thetumor tissues (Supplementary Fig. S1). In the p53mutation,Reprimo mRNA expression was undetectable in four of thefive gastric cancer cell lines and one of the two tumor tissues.Overall, Reprimo expression was observed at a weak or faintlevel, regardless of the promoter methylation or p53 genestatus. We were unable to assess the protein level because ofthe absence of a commercially available antibody. In addi-tion, we examined the three correlations between p53 gene
status and Reprimo promoter methylation in 83 primaryhuman gastric cancer tissues. Mutation of p53 was found in34.9% (29/83) of tumor tissues, which were missensemutations clustered in the core DNA-binding domain(residues 94–312; ref. 25) and have been documented asbiologically inactive via the transcriptional activity on theWAF1 promoter in the Universal Mutation Database TP53mutation databases (26; Supplementary Table S2). Reprimopromoter methylation was unrelated to p53 gene status(Table 1).Given that Reprimo expression was induced by X-ray
irradiation as the DNA damage in embryonic fibroblast cellswith p53 wild-type (9), we examined whether or not Rep-rimo expressionwas altered in response to forms of genotoxicstress, such asUV irradiation, 5-FU (fluoropyrimidine class),or CDDP (platinum-based class). SH10 cells without
m m+UV 5-Aza 5-Aza+UV
AZ521 NUGC4 KatoIII MKN74 SH10 GCIY –+ + + + –
W W M W M Mp53 status
Re
lati
ve
re
pri
mo
ex
pre
ss
ionC
*
*
* * * * * *
*
P = 0.0001
P < 0.0001
P = 0.0001 P < 0.0001
P = 0.0028
P = 0.0003 P < 0.0001
P < 0.0001
P = 0.0146
P = 0.0177
Tumor Normal
BAReprimo
384bp
β-Actin
295bp
Reprimo
β-Actin
Mock 5-FU CDDP UV
Re
lati
ve
re
pri
mo
ex
pre
ss
ion
– – – – – – –+ + + + + + + + + + +
M W W W M W W W W W M W M M M W W M
qRT-PCR
Cas
e 1T
Cas
e 2T
Cas
e 3T
Cas
e 4T
Cas
e 5T
SH 1
0
AZ
521
Kat
o III
H 1
11M
KN 7
MKN 7
4NUG
C 4
GCIY
Cas
e 1N
Cas
e 2N
Cas
e 3N
Cas
e 4N
Cas
e 5N
Mock
5-FU
CDDP
UV
Cas
e 1
Cas
e 2
Cas
e 3
Cas
e 4
Cas
e 5
SH
10
AZ
521
Kat
o III
H 1
11M
KN 7
MK
N 7
4NUG
C 4
GCIY
6
5
4
3
2
1
0
12
10
8
6
4
2
0
5
4
3
2
1
0
Re
lati
ve
re
pri
mo
ex
pre
ss
ion
p53 status
Methylation
Methylation
Figure3. DNAdamage-inducible Reprimo expression and its silencing bypromotermethylation. A, expression level of ReprimobyRT-PCR (top) andqRT-PCR(Reprimo/b-actin � 1,000, bottom) in eight gastric cancer cell lines and preliminary dataset. B, expression level of Reprimo after genotoxic stress in SH10cells without Reprimo methylation by RT-PCR (top) or qRT-PCR (bottom). SH10 cells were incubated for 24 hours followed by UV irradiation (10 mJ/cm2).Cells were treated with 5-FU (10 mmol/L) or CDDP (20 mmol/L) for 24 hours. With mock treatment cells as 1.0, the relative expression level of Reprimowas shown by qRT-PCR. C, expression level after treatment with demethylating agent 5-Aza-dC alone or in the presence or absence of UV irradiationby qRT-PCR. Cells were treated with 5-Aza-dC (5 mmol/L) for 5 days. When combined with the UV irradiation, UV irradiation was conducted on Day 4of 5-Aza-dC treatment. 5-Aza, 5-Aza-dC; m, mock including the same volume of acetic acid; �, P < 0.05; Error bars; SD. N, human normal mucosa; T, humanprimary tumor; W, p53 wild-type; M, p53 mutation.
Ooki et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1368
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091

Reprimo methylation increased Reprimo expression underDNA-damaging conditions despite the presence of the p53mutation (Fig. 3B), speculating that DNA damage-induc-ible Reprimo expression may be abrogated because of meth-ylation of the Reprimo promoter in a p53-independentmanner. To substantiate this hypothesis, six cell lines withdifferent p53 gene and Reprimo methylation statuses weretreated with the demethylating agent 5-Aza-dC in thepresence or absence of DNA-damaging agents (Fig. 3C).In GCIY cells without Reprimo promoter methylation,Reprimo expression was weak under normal conditions,but was induced by UV irradiation despite the presence ofp53 mutation, in line with findings in SH10 cells. On theother hand, in four cell lines (AZ521, NUGC, KatoIII, andMKN74 cells) with Reprimo promoter methylation, theexpression of Reprimo could be slightly restored by demeth-ylation alone, but not UV irradiation. Notably, in combi-nation with demethylation and UV irradiation, these celllines were restored regardless of p53 gene status. Similartrends were also observed in the treatment with 5-FU orCDDP (data not shown). We examined Reprimo muta-tions via direct sequencing analysis using primer sequencesincluding the entire open reading frame according to theprevious study (13) in eight gastric cancer cell lines and 83primary tumors, and no mutations were found (data notshown).
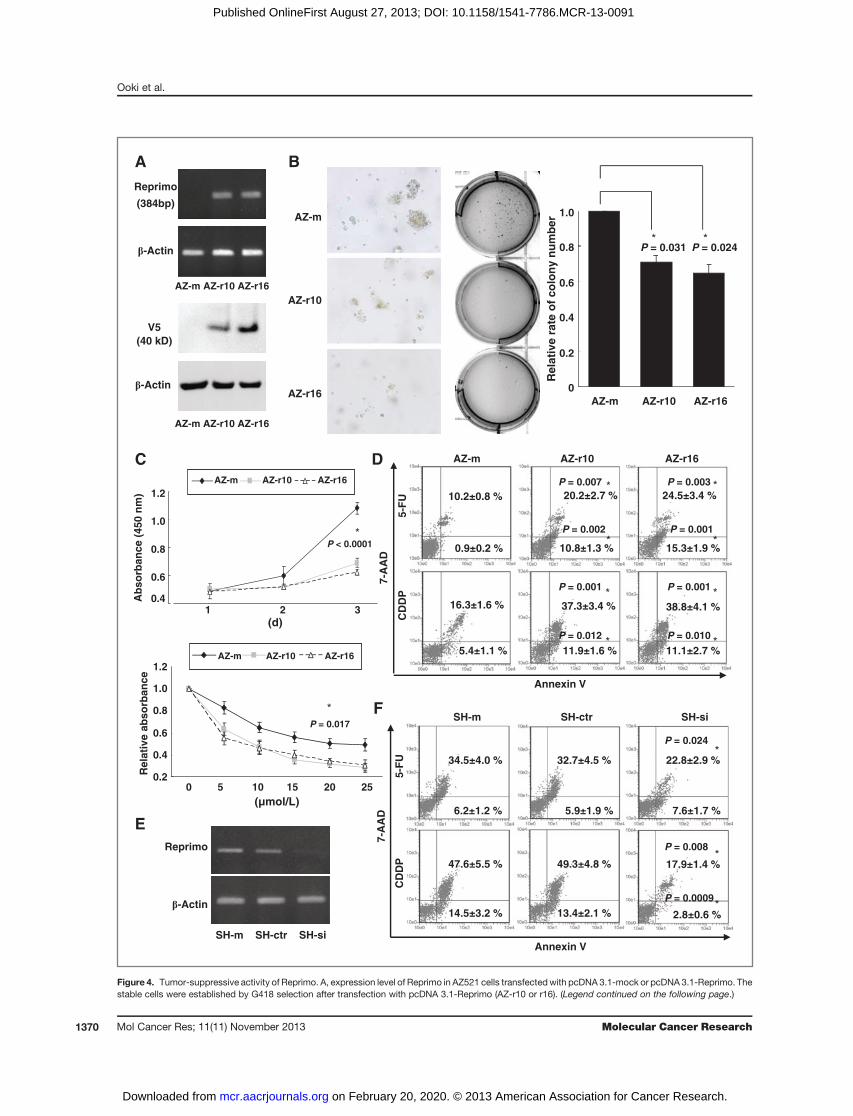
Tumor-suppressive activity after transfection of ReprimoTranscriptional silencing of Reprimo due to its frequent
promoter methylation prompted investigation into thepotential role of Reprimo as a tumor-suppressor gene ingastric cancer. AZ521 cells with methylation were stablytransfected with pcDNA 3.1-Reprimo containing the full-length cDNA of Reprimo gene (Fig. 4A). The AZ521 cellstransfected with pcDNA 3.1-Reprimo (AZ-r10 and AZ-r16cells) resulted in an increase of mRNA levels, unlike thosewith pcDNA 3.1-mock (AZ-mock cells). Reprimo proteinlevels were also confirmed byWestern blotting using an anti-flag-V5 antibody. An important characteristic in the growthof aggressive tumors is the ability of cancer cells to growunder anchorage-independent conditions. We thereforeexamined whether or not the exogenous expression ofReprimo could suppress the growth of cells in soft agar.The AZ-r10 and AZ-r16 cells showed significantly decreasedcell size and colony number (Fig. 4B), but no decrease inMatrigel invasive ability (Supplementary Fig. S2A) whencompared with AZ-m cells. We also observed a decrease incellular proliferation under normal conditions or CDDPexposure in AZ-r10 and AZ-r16 cells (Fig. 4C).Reprimo has been shown able to act as a G2 phase cell-
cycle break in cell lines such as DLD1, HeLa, and MCF7cells (9), but not in LbT2 and GH3 pituitary cells (27),indicating that it may have different functions depending onthe type of cell. To examine the role of Reprimo in regulatingcell growth in gastric cancer, a cell-cycle assay was conducted(Supplementary Fig. S2B). Analysis of the cell-cycle distri-bution showed a significantly increased sub-G1 populationalong with minimally affected S and G2–M populations in
AZ-r10 and AZ-r16 cells. H111 cells transfected withpcDNA 3.1-Reprimo also showed similar results. To verifythese results, an apoptosis assay was conducted under expo-sure to the DNA-damaging agents 5-FU or CDDP (Fig.4D). Both early and late apoptosis were significantlyenhanced in AZ-r10 and AZ-r16 cells compared withAZ-m cells.To further confirm the tumor-suppressive activity of
Reprimo, knockdown of endogenous Reprimo was con-ducted using siRNA. SH10 cells without methylation weretransfected with siRNA-Reprimo (SH-si cells) or siRNA-ctr(SH-ctr cells). SH-si cells, but not SH-ctr cells, resulted indecreased Reprimo expression induced by CDDP exposurewhen compared with mock treatment cells (Fig. 4E) andshowed a dramatic repression of 5-FU or CDDP-mediatedapoptosis (Fig. 4F). We observed similar results in GCIYcells (data not shown).
Tumor-suppressive activity of Reprimo in mousexenograft assayTo examine the effect of Reprimo on tumorigenesis in
vivo, AZ521 patient (AZ-p), AZ-m, AZ-r10, or AZ-r16 cellswere subcutaneously injected into both flanks of nude mice.A remarkable reduction of tumor volume was observed inmice injectedwith Reprimo-transfected cells (AZ-r10 or AZ-r16 cells) when compared with AZ-p or AZ-m cells. (Fig. 5Aand B). Tumor weight was also significantly decreased inmice injected with AZ-r10 or AZ-r16 cells (Fig. 5C). AsReprimo promoter methylation was marginally associatedwith vascular invasion in 83 patients with gastric cancer(Table 1), angiogenesis, visualized as amarked red color withthe formation of a dense surrounding vascular network, wasmore greatly observed in the tumors of mice injected withAZ-p or AZ-m cells than those injected with AZ-r10 or AZ-r16 cells (Fig. 5D).To examine the anticancer effect of treatment combined
with CDDP and 5-FU via Reprimo expression in vivo, micewith tumors approximately 5 mm in diameter receivedintraperitoneal administration of CDDP and 5-FU for 7days.No anticancer effect was observed inmice injectedwithAZ-p or AZ-m cells, but an anticancer effect was observed inthe mice injected with AZ-r10 or AZ-r16 cells (Supplemen-tary Fig. S3).
Potential use of Reprimo methylation as a predictivemarkerThemost promising candidate epigeneticmarker reported
thus far for the prediction of responses to chemotherapy isO6-methylguanine-methyltransferase (MGMT). MGMTreverses the addition of alkyl groups to the guanine base ofDNA and protects DNA damage caused by the alkylatingagents, such as temozolomide, thus generating chemoresis-tance. Therefore, themethylation ofMGMT is a predictor ofa favorable response to temozolomide in gliomas (28). In thepresent study, we found that Reprimo could enhance 5-FUor CDDP-mediated apoptosis in gastric cancer. The com-bination of CDDP and fluoropyrimidine class, including5-FU, capecitabine, or S-1,was themost standard regimen of
Reprimo in Gastric Cancer
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1369
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091
β-Actin
Reprimo
SH-m SH-ctr SH-si
5-F
UC
DD
P
Annexin V
7-A
AD
SH-m SH-ctr SH-si
47.6±5.5 %
14.5±3.2 %
49.3±4.8 %
13.4±2.1 %
17.9±1.4 %
2.8±0.6 %
34.5±4.0 %
6.2±1.2 %
32.7±4.5 %
5.9±1.9 %
22.8±2.9 %
7.6±1.7 %
A
AZ-m AZ-r10 AZ-r16
β-Actin
V5
(40 kD)
AZ-m AZ-r10 AZ-r16
β-Actin
Reprimo
(384bp)
B
Re
lati
ve
ra
te o
f c
olo
ny
nu
mb
erAZ-m
AZ-r10
AZ-r16AZ-m AZ-r10 AZ-r16
0
0.2
0.4
0.6
0.8
1.0
P = 0.024P = 0.031
1 2 3
C
0.4
0.6
0.8
1.0
1.2
AZ-m AZ-r10 AZ-r16
Ab
so
rban
ce (
450 n
m)
(d)
P < 0.0001
Rela
tive a
bso
rban
ce
0.2
0.4
0.6
0.8
1.0
1.2AZ-m AZ-r10 AZ-r16
0 5 10 15 20 25
(µmol/L)
5-F
U
Annexin V
7-A
AD
AZ-m AZ-r10 AZ-r16
16.3±1.6 %
5.4±1.1 %
37.3±3.4 %
11.9±1.6 %
38.8±4.1 %
11.1±2.7 %
10.2±0.8 %
0.9±0.2 %
20.2±2.7 % 24.5±3.4 %
10.8±1.3 % 15.3±1.9 %
CD
DP
P = 0.017
D
E
F
P = 0.007 P = 0.003
P = 0.002 P = 0.001
P = 0.001P = 0.001
P = 0.010P = 0.012
P = 0.024
P = 0.008
P = 0.0009
Figure 4. Tumor-suppressive activity of Reprimo. A, expression level of Reprimo in AZ521 cells transfectedwith pcDNA 3.1-mock or pcDNA 3.1-Reprimo. Thestable cells were established by G418 selection after transfection with pcDNA 3.1-Reprimo (AZ-r10 or r16). (Legend continued on the following page.)
Ooki et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1370
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091
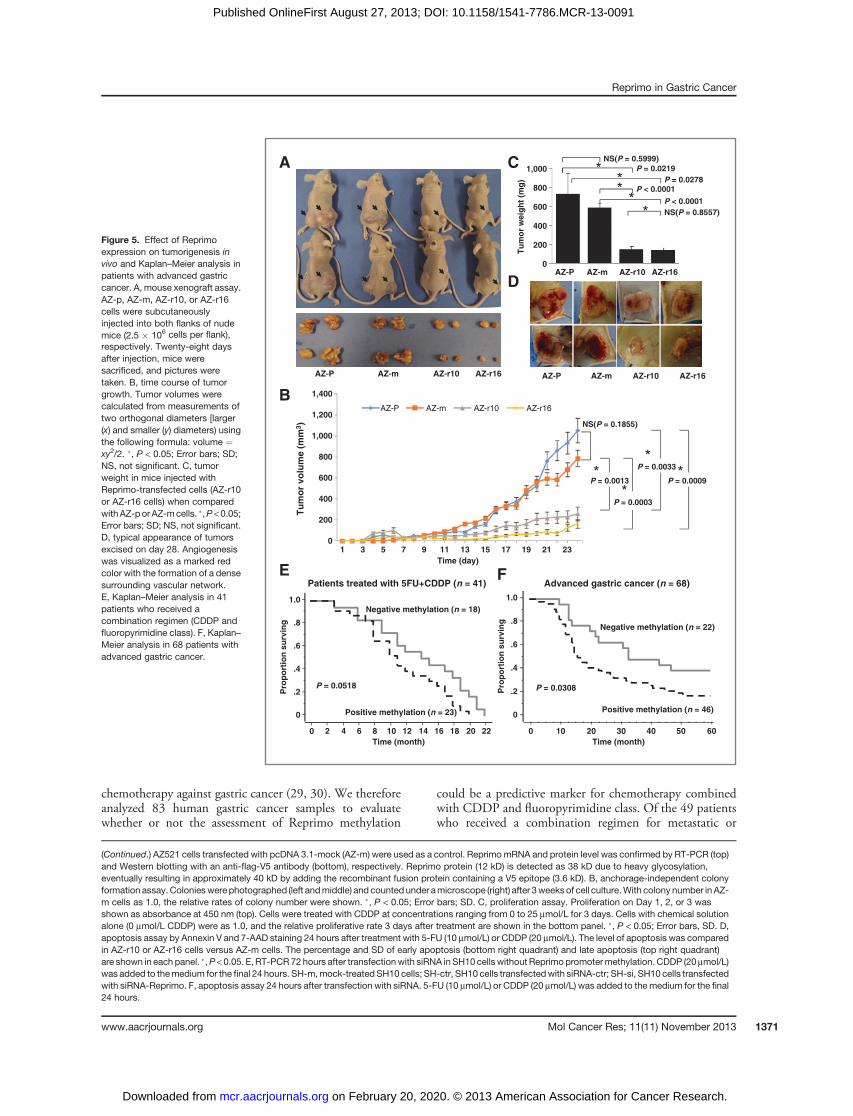
chemotherapy against gastric cancer (29, 30). We thereforeanalyzed 83 human gastric cancer samples to evaluatewhether or not the assessment of Reprimo methylation
could be a predictive marker for chemotherapy combinedwith CDDP and fluoropyrimidine class. Of the 49 patientswho received a combination regimen for metastatic or
CA
B
D
E F
0
.2
.4
.6
.8
1.0
0 10 20 30 40 50 60
Positive methylation (n = 46)
Negative methylation (n = 22)
P = 0.0308
Time (month)
Pro
po
rtio
n s
urv
ing
Advanced gastric cancer (n = 68)Patients treated with 5FU+CDDP (n = 41)
0
.2
.4
.6
.8
1.0
0 2 4 6 8 10 12 14 16 18 20 22
Positive methylation (n = 23)
Negative methylation (n = 18)
Time (month)
Pro
po
rtio
n s
urv
ing
P = 0.0518
NS(P = 0.5999)
P = 0.0219
P = 0.0278P < 0.0001
NS(P = 0.8557)
P < 0.0001
Tu
mo
r w
eig
ht
(mg
)
AZ-P AZ-m AZ-r10 AZ-r16
Tu
mo
r vo
lum
e (
mm
3)
Time (day)
1 3 5 7 9 11 13 15 17 19 21 23
AZ-P AZ-m AZ-r10 AZ-r16
P = 0.0009
NS(P = 0.1855)
P = 0.0033
P = 0.0013
P = 0.0003
1,000
800
600
400
200
0
1,400
1,200
1,000
800
600
400
200
0
AZ-P AZ-m AZ-r10 AZ-r16
AZ-P AZ-m AZ-r10 AZ-r16
Figure 5. Effect of Reprimoexpression on tumorigenesis invivo and Kaplan–Meier analysis inpatients with advanced gastriccancer. A, mouse xenograft assay.AZ-p, AZ-m, AZ-r10, or AZ-r16cells were subcutaneouslyinjected into both flanks of nudemice (2.5 � 106 cells per flank),respectively. Twenty-eight daysafter injection, mice weresacrificed, and pictures weretaken. B, time course of tumorgrowth. Tumor volumes werecalculated from measurements oftwo orthogonal diameters [larger(x) and smaller (y) diameters) usingthe following formula: volume ¼xy2/2. �, P < 0.05; Error bars; SD;NS, not significant. C, tumorweight in mice injected withReprimo-transfected cells (AZ-r10or AZ-r16 cells) when comparedwithAZ-por AZ-mcells. �,P<0.05;Error bars; SD; NS, not significant.D, typical appearance of tumorsexcised on day 28. Angiogenesiswas visualized as a marked redcolor with the formation of a densesurrounding vascular network.E, Kaplan–Meier analysis in 41patients who received acombination regimen (CDDP andfluoropyrimidine class). F, Kaplan–Meier analysis in 68 patients withadvanced gastric cancer.
(Continued.) AZ521 cells transfected with pcDNA 3.1-mock (AZ-m) were used as a control. ReprimomRNA and protein level was confirmed by RT-PCR (top)and Western blotting with an anti-flag-V5 antibody (bottom), respectively. Reprimo protein (12 kD) is detected as 38 kD due to heavy glycosylation,eventually resulting in approximately 40 kD by adding the recombinant fusion protein containing a V5 epitope (3.6 kD). B, anchorage-independent colonyformation assay. Colonieswere photographed (left andmiddle) and counted under amicroscope (right) after 3weeksof cell culture.With colony number in AZ-m cells as 1.0, the relative rates of colony number were shown. �, P < 0.05; Error bars; SD. C, proliferation assay. Proliferation on Day 1, 2, or 3 wasshown as absorbance at 450 nm (top). Cells were treated with CDDP at concentrations ranging from 0 to 25 mmol/L for 3 days. Cells with chemical solutionalone (0 mmol/L CDDP) were as 1.0, and the relative proliferative rate 3 days after treatment are shown in the bottom panel. �, P < 0.05; Error bars, SD. D,apoptosis assay by Annexin V and 7-AAD staining 24 hours after treatment with 5-FU (10 mmol/L) or CDDP (20 mmol/L). The level of apoptosis was comparedin AZ-r10 or AZ-r16 cells versus AZ-m cells. The percentage and SD of early apoptosis (bottom right quadrant) and late apoptosis (top right quadrant)are shown in each panel. �,P < 0.05. E, RT-PCR72 hours after transfectionwith siRNA inSH10 cells without Reprimopromotermethylation. CDDP (20mmol/L)was added to themedium for the final 24 hours. SH-m,mock-treated SH10 cells; SH-ctr, SH10 cells transfectedwith siRNA-ctr; SH-si, SH10 cells transfectedwith siRNA-Reprimo. F, apoptosis assay 24 hours after transfection with siRNA. 5-FU (10 mmol/L) or CDDP (20 mmol/L) was added to the medium for the final24 hours.
Reprimo in Gastric Cancer
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1371
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091

recurrent gastric cancer, 41 had measurable lesions. Thetumor response was assessed according to Response Evalu-ation Criteria in Solid Tumors, version 1.1 (31). Reprimopromoter hypermethylation, previously defined as a valuemore than 0.42 onQ-MSP, was inversely associated with theresponse rate (P ¼ 0.028; Table 1), and showed a trendtoward a worsening prognosis, compared with absence ofReprimo promoter methylation (Fig. 5E).We next examined whether or not the assessment of
Reprimo hypermethylation could be used to predict patientprognosis and thereby serve as a marker for tumor aggres-siveness in 68 patients with advanced gastric cancer. Rep-rimo promoter hypermethylation had a significantly pooroutcome (P ¼ 0.031; Fig. 5F). In multivariate analysis,Reprimo promoter hypermethylation was independentlyassociated with a poor prognosis, with a HR of 2.148(P ¼ 0.026; Supplementary Table S3).
DiscussionWe previously reported that Reprimo promoter methyl-
ation was one of the most frequent cancer-specific altera-tions, using bisulfite sequencing analysis in primary gastriccancer tissues (15). Of note, many of the methylationalterations are tissue specific or associated with the field ofcancerization, but are not cancer specific (32). As such, weexamined the epigenetic inactivation, biologic function, andclinical relevance in gastric cancer in the present study.In gastric cancer cell lines without Reprimo promoter
methylation, we observed an unexpectedly faint or weakReprimo expression under normal conditions but a robustexpression under DNA-damaging conditions. In gastriccancer cell lines with Reprimo promoter methylation, how-ever, we observed faint expression even under DNA-dam-aging conditions, with expression being restored only incombination with the demethylating agent. In 83 humangastric cancer samples, Reprimo promotermethylation was acancer-specific alteration that frequently occurred at an earlystage in the development of cancer andwas not related to p53status. Thus, Reprimo promoter methylation may be thepredominant mechanism for loss of DNA damage-inducibleReprimo expression, regardless of p53 status.Although promoter methylation alterations are common
events in human cancers, most are merely passengers thathave no effect per se on the process of carcinogenesis. Incontrast, driver methylation mainly inactivates tumor-sup-pressive activity, leading to the processes of tumor initiation,progression, or metastasis. Thus, the identification of drivermethylation will be of key importance for cancer etiology,development of biomarkers, and therapeutic targets (33).Enforced Reprimo expression robustly inhibited tumor cellgrowth, including cell proliferation and anchorage-indepen-dent colony formation. In addition, the cell-cycle distribu-tion showed a significantly increased sub-G1 population, butnot G2 arrest that has been reported as a function of Reprimo(9). Our findings are in line with those in pituitary tumors(27), and Reprimo may therefore have differential rolesdepending on the type of tumor. Apoptosis assays also
showed that enforced Reprimo expression enhanced DNAdamage-induced apoptosis, indicating that Reprimo mightsuppress cell growth through the induction of apoptosis butnot cell-cycle arrest in gastric cancer. Inverse effects wereobserved on siRNA-mediated knockdown of endogenousReprimo. Furthermore, Reprimo expression inhibitedtumorigenesis in a mouse xenograft model. Taken together,these findings suggest that Reprimo possesses bona fidetumor-suppressive activity, and its methylation may be adriver methylation in gastric cancer. Further investigation ofthe Reprimo-regulated molecules is required to clarify themechanisms by which Reprimo exerts tumor-suppressiveactivity.Threemajor clinical oncology areas can potentially benefit
fromDNAmethylation-based biomarkers: cancer detection,tumor prognosis, and prediction of treatment responses(34). The cancer-specific Reprimo promoter methylationthat occurs at an early stage in disease development can serveas a potential biomarker for early detection in gastric cancer.Indeed, Reprimo promoter methylation was frequentlydetected in plasma from patients with gastric cancer (11).Furthermore, Reprimo promoter methylation is significant-ly lower in chemoradiotherapy responders than in nonre-sponders in esophageal cancer (35), and is predictive of apoor prognosis in pancreatic ductal carcinoma (36). Rep-rimo promoter methylation was inversely associated withresponse to the treatment combined with CDDP and thefluoropyrimidine class as a standard chemotherapy. In addi-tion, Reprimo expression enhanced CDDP or 5-FU–medi-ated apoptosis in vitro, and anticancer effect of treatmentcombined with CDDP and 5-FU in vivo.We were unable toassess any association between Reprimo protein level andresponse to chemotherapy due to lack of a commerciallyavailable antibody.However, Reprimo expression is inducedby DNA damage, and as such, potential functionality ofReprimo as tumor-suppressive activity may not be reflectedby expression level of mRNA or protein at diagnosis. Atpresent, the assessment of Reprimo promoter methylationmay be useful in predicting response to chemotherapy withCDDP and the fluoropyrimidine class. Furthermore, Rep-rimo promotermethylation in patients with advanced gastriccancer was an independent prognostic factor reflecting anaggressive tumor phenotype, indicating that Reprimo pro-moter methylation is one of the most critical alterations incancer progression and is also a potential marker for tumoraggressiveness.In the last decade, a number of landmark trials have shown
efficacy ofmultimodality therapy in resectable gastric cancer,such as perioperative chemotherapy (37), adjuvant chemor-adiotherapy (38), adjuvant chemotherapy (39, 40), andneoadjuvant chemoradiotherapy (41). However, the thera-peutic strategies by which clinicians distinguish between thevarious treatment options remain a contentious issue. Onepoint of clinical importance is that, although preoperativetherapy has the potential to improve the therapeutic out-come for responders (42), it runs the risk of shifting the statusof nonresponders from potentially resectable to unresect-able, despite that radical resection is the only curative
Ooki et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1372
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091

treatment modality, requiring the predictive marker fortreatment response. Therefore, information on Reprimopromoter methylation may provide much needed guidancein determining the therapeutic strategies, such as the selec-tion of the most suitable patients for preoperative therapy ingastric cancer.In conclusion, Reprimo expression is normally induced
in response to DNA damage, robustly inhibiting tumor-igenesis through enhancing apoptosis as a novel tumorsuppressor in gastric cancer. Methylation of the Reprimopromoter is a frequent and cancer-specific event thatabrogates its expression and functions in p53-independentmanner. In addition, clinical assessment of Reprimopromoter methylation may serve not only as a predictivemarker for chemotherapy consisting of CDDP and thefluoropyrimidine class, but also as a marker for tumoraggressiveness. Further research will be necessary tovalidate the clinical potential of Reprimo promoter meth-ylation as a biomarker in gastric cancer.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
DisclaimerThe funding agencies had no role in the design of the study, data collection,
analysis, interpretation of the results, preparation of the manuscript, or in the decisionto submit the manuscript for publication.
Authors' ContributionsConception and design: A. Ooki, K. Yamashita, M. WatanabeDevelopment of methodology: A. Ooki, K. Yamashita, M. WatanabeAcquisition of data (provided animals, acquired and managed patients, providedfacilities, etc.): A. Ooki, A. Mondal, H. Nishimiya, M. WatanabeAnalysis and interpretation of data (e.g., statistical analysis, biostatistics, compu-tational analysis): A. Ooki, K. Yamashita, M. WatanabeWriting, review, and/or revision of the manuscript: A. Ooki, K. Yamashita, K.Yamaguchi, M. WatanabeAdministrative, technical, or material support (i.e., reporting or organizing data,constructing databases): K. Yamashita, M. WatanabeStudy supervision: K. Yamashita, M. Watanabe
Grant SupportThis work was supported, in part, by the Grant-in-Aid for Cancer Research from
the Ministry of Health, Labour andWelfare of Japan and by the Japanese Foundationfor Multidisciplinary Treatment of Cancer.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be herebymarked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
Received February 21, 2013; revised July 8, 2013; accepted July 23, 2013;published OnlineFirst August 27, 2013.
References1. Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates
of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int JCancer 2010;127:2893–917.
2. Nagini S. Carcinoma of the stomach: a review of epidemiology,pathogenesis, molecular genetics and chemoprevention. WorldJ Gastrointest Oncol 2012;4:156–69.
3. Baylin SB, Jones PA. A decade of exploring the cancer epigenome -biological and translational implications. Nat Rev Cancer 2011;11:726–34.
4. You JS, Jones PA. Cancer genetics and epigenetics: two sides of thesame coin? Cancer Cell 2012;22:9–20.
5. Baylin SB, OhmJE. Epigenetic gene silencing in cancer - amechanismfor early oncogenic pathway addiction? Nat Rev Cancer 2006;6:107–16.
6. Sapari NS, Loh M, Vaithilingam A, Soong R. Clinical potential of DNAmethylation in gastric cancer: a meta-analysis. PLoS One 2012;7:e36275.
7. Ushijima T, Sasako M. Focus on gastric cancer. Cancer Cell 2004;5:121–5.
8. Zouridis H, Deng N, Ivanova T, Zhu Y, Wong B, Huang D, et al.Methylation subtypes and large-scale epigenetic alterations in gastriccancer. Sci Transl Med 2012;4:156ra140.
9. Ohki R, Nemoto J, Murasawa H, Oda E, Inazawa J, Tanaka N, et al.Reprimo, a new candidate mediator of the p53-mediated cell cyclearrest at the G2 phase. J Biol Chem 2000;275:22627–30.
10. Zhang Z, Li J, Lantry LE, Wang Y, Wiseman RW, Lubet RA, et al. p53transgenic mice are highly susceptible to 1, 2-dimethylhydrazine-induced uterine sarcomas. Cancer Res 2002;62:3024–9.
11. Bernal C, Aguayo F, Villarroel C, Vargas M, Diaz I, Ossandon FJ, et al.Reprimo as a potential biomarker for early detection in gastric cancer.Clin Cancer Res 2008;14:6264–9.
12. Sato N, Goggins M. The role of epigenetic alterations in pancreaticcancer. J Hepatobiliary Pancreat Surg 2006;13:286–95.
13. Takahashi T, Suzuki M, Shigematsu H, Shivapurkar N, Echebiri C,Nomura M, et al. Aberrant methylation of Reprimo in human malig-nancies. Int J Cancer 2005;115:503–10.
14. Vanlandingham JW, Tassabehji NM, Somers RC, Levenson CW.Expression profiling of p53-target genes in copper-mediated neuronalapoptosis. Neuromolecular Med 2005;7:311–24.
15. Ooki A, Yamashita K, Kikuchi S, Sakuramoto S, Katada N, Kokubo K,et al. Potential utility of HOP homeobox gene promoter methylation asa marker of tumor aggressiveness in gastric cancer. Oncogene2010;29:3263–75.
16. Japanese Gastric Cancer Association. Japanese gastric cancer treat-ment guidelines 2010 (ver. 3). Gastric Cancer 2011;14:113–23.
17. Japanese Gastric Cancer Association. Japanese classification of gas-tric carcinoma: 3rd English edition. Gastric Cancer 2011;14:101–12.
18. Fenoglio-Preiser CM, Wang J, Stemmermann GN, Noffsinger A. TP53and gastric carcinoma: a review. Hum Mutat 2003;21:258–70.
19. Livak KJ, Schmittgen TD. Analysis of relative gene expression datausing real-time quantitaitve PCR and the 2(-Delta Delata C(T)) Method.Methods 2001;25:402–8.
20. Mandelker DL, Yamashita K, Tokumaru Y, Mimori K, Howard DL,Tanaka Y, et al. PGP9.5 promoter methylation is an independentprognostic factor for esophageal squamous cell carcinoma. CancerRes 2005;65:4963–8.
21. Tanaka F, Yamaguchi H, Ohta M, Mashino K, Sonoda H, Sadanaga N,et al. Intratumoral injection of dendritic cells after treatment of anti-cancer drugs induces tumor-specific antitumor effect in vivo. Int JCancer 2002;101:265–9.
22. Brena RM, Huang TH, Plass C. Quantitative assessment of DNA meth-ylation: potential applications for disease diagnosis, classification, andprognosis in clinical settings. J Mol Med (Berl) 2006;84:365–77.
23. Maegawa S, Hinkal G, Kim HS, Shen L, Zhang L, Zhang J, et al.Widespread and tissue specific age-related DNAmethylation changesin mice. Genome Res 2010;20:332–40.
24. Malik S, Jiang S, Garee JP, Verdin E, Lee AV, O'Malley BW, et al.Histone deacetylase 7 and FoxA1 in estrogen-mediated repression ofRPRM. Mol Cell Biol 2010;30:399–412.
25. Kato S, Han SY, Liu W, Otsuka K, Shibata H, Kanamaru R, et al.Understanding the function-structure and function-mutation rela-tionships of p53 tumor suppressor protein by high-resolution mis-sense mutation analysis. Proc Natl Acad Sci U S A 2003;100:8424–9.
26. Edlund K, Larsson O, Ameur A, Bunikis I, Gyllensten U, Leroy B, et al.Data-driven unbiased curation of the TP53 tumor suppressor genemutation database and validation by ultradeep sequencing of humantumors. Proc Natl Acad Sci U S A 2012;109:9551–6.
Reprimo in Gastric Cancer
www.aacrjournals.org Mol Cancer Res; 11(11) November 2013 1373
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091

27. Xu M, Knox AJ, Michaelis KA, Kiseljak-Vassiliades K, Kleinschmidt-DeMasters BK, Lillehei KO, et al. Reprimo (RPRM) is a novel tumorsuppressor in pituitary tumors and regulates survival, proliferation, andtumorigenicity. Endocrinology 2012;153:2963–73.
28. Hegi ME, Diserens AC, Gorlia T, Hamou MF, de Tribolet N, Weller M,et al. MGMT gene silencing and benefit from temozolomide in glio-blastoma. N Engl J Med 2005;352:997–1003.
29. Allum WH, Blazeby JM, Griffin SM, Cunningham D, Jankowski JA,Wong R. Guidelines for the management of oesophageal and gastriccancer. Gut 2011;60:1449–72.
30. Koizumi W, Narahara H, Hara T, Takagane A, Akiya T, Takagi M,et al. S-1 plus cisplatin versus S-1 alone for first-line treatment ofadvanced gastric cancer (SPIRITS trial): a phase III trial. LancetOncol 2008;9:215–21.
31. Eisenhauer EA, Therasse P, Bogaerts J, Schwartz LH, Sargent D, FordR, et al. New response evaluation criteria in solid tumours: revisedRECIST guideline (version 1.1). Eur J Cancer 2009;45:228–47.
32. Ushijima T. Epigenetic field for cancerization. J Biochem Mol Biol2007;40:142–50.
33. Kalari S, Pfeifer GP. Identification of driver and passenger DNA meth-ylation in cancer byepigenomicanalysis. AdvGenet 2010;70:277–308.
34. Rodriguez-Paredes M, Esteller M. Cancer epigenetics reaches main-stream oncology. Nat Med 2011;17:330–9.
35. Hamilton JP, Sato F, Greenwald BD, Suntharalingam M, Krasna MJ,Edelman MJ, et al. Promoter methylation and response to chemother-apy and radiation in esophageal cancer. Clin Gastroenterol Hepatol2006;4:701–8.
36. Sato N, Fukushima N, Matsubayashi H, Iacobuzio-Donahue CA, YeoCJ, Goggins M. Aberrant methylation of Reprimo correlates withgenetic instability and predicts poor prognosis in pancreatic ductaladenocarcinoma. Cancer 2006;107:251–7.
37. Cunningham D, Allum WH, Stenning SP, Thompson JN, Van de VeldeCJ, NicolsonM, et al. Perioperative chemotherapy versus surgery alonefor resectable gastroesophageal cancer. N Engl JMed 2006;355:11–20.
38. Macdonald JS, Smalley SR, Benedetti J, Hundahl SA, Estes NC,Stemmermann GN, et al. Chemoradiotherapy after surgery comparedwith surgery alone for adenocarcinoma of the stomach or gastro-esophageal junction. N Engl J Med 2001;345:725–30.
39. Bang YJ, Kim YW, Yang HK, Chung HC, Park YK, Lee KH. Adjuvantcapecitabine and oxaliplatin for gastric cancer after D2 gastrectomy(CLASSIC): a phase 3 open-label, randomised controlled trial. Lancet2012;379:315–21.
40. Sasako M, Sakuramoto S, Katai H, Kinoshita T, Furukawa H, Yama-guchi T. Five-year outcomes of a randomized phase III trial comparingadjuvant chemotherapy with S-1 versus surgery alone in stage II or IIIgastric cancer. J Clin Oncol 2011;29:4387–93.
41. Ajani JA, Winter K, Okawara GS, Donohue JH, Pisters PW, Crane CH,et al. Phase II trial of preoperative chemoradiation in patients withlocalized gastric adenocarcinoma (RTOG 9904): quality of combinedmodality therapyandpathologicresponse.JClinOncol2006;24:3953–8.
42. Mansour JC, Tang L, ShahM, BentremD, Klimstra DS, GonenM, et al.Does graded histologic response after neoadjuvant chemotherapypredict survival for completely resected gastric cancer? Ann SurgOncol 2007;14:3412–8.
Ooki et al.
Mol Cancer Res; 11(11) November 2013 Molecular Cancer Research1374
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091

2013;11:1362-1374. Published OnlineFirst August 27, 2013.Mol Cancer Res Akira Ooki, Keishi Yamashita, Kensei Yamaguchi, et al. CancerSuppressor and Is Suppressed by Promoter Methylation in Gastric DNA Damage-Inducible Gene, Reprimo Functions as a Tumor
Updated version
10.1158/1541-7786.MCR-13-0091doi:
Access the most recent version of this article at:
Material
Supplementary
http://mcr.aacrjournals.org/content/suppl/2013/09/04/1541-7786.MCR-13-0091.DC1
Access the most recent supplemental material at:
Cited articles
http://mcr.aacrjournals.org/content/11/11/1362.full#ref-list-1
This article cites 42 articles, 12 of which you can access for free at:
Citing articles
http://mcr.aacrjournals.org/content/11/11/1362.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mcr.aacrjournals.org/content/11/11/1362To request permission to re-use all or part of this article, use this link
on February 20, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 27, 2013; DOI: 10.1158/1541-7786.MCR-13-0091