Embed Size (px)
Citation preview

!!!!!Study!on!the!role!of!Stx2!gene!in!spermatogenesis!
of!mice!!!�
�
�!
(Stx2������� ���������������
�
�
�
�
�
�
�
�
�
�
�
�
2013,!September!Yasuhiro!Fujiwara!
!!
Graduate!School!of!Natural!Science!and!Technology!!
Okayama!University!

! 1!
Contents
Chapter 1
General Introduction------------------------------------------------------- 3
Chapter 2
Histological and immunohistochemical analysis of the Stx2repro34 mice
Introduction---------------------------------------------------------------------------------- 24
Material and methods---------------------------------------------------------------------- 29
Results----------------------------------------------------------------------------------------- 37
Discussion------------------------------------------------------------------------------------- 42
Figures and tables--------------------------------------------------------------------------- 46
Chapter 3
Expression of STX2 in the seminiferous epithelium
Introduction---------------------------------------------------------------------------------- 60
Material and methods---------------------------------------------------------------------- 62
Results----------------------------------------------------------------------------------------- 65
Discussion------------------------------------------------------------------------------------ 67
Figures and tables-------------------------------------------------------------------------- 70

! 2!
Chapter 4
Aberrant localization of sulfoglycolipids lead to multinucleation of
the germ cells
Introduction---------------------------------------------------------------------------------- 78
Material and methods---------------------------------------------------------------------- 80
Results----------------------------------------------------------------------------------------- 81
Discussion------------------------------------------------------------------------------------ 83
Figures and tables--------------------------------------------------------------------------- 87
Chapter 5
General discussion and Conclusion------------------------------------------------------ 92
Acknowledgement--------------------------------------------------------------------------- 100
References------------------------------------------------------------------------------------ 101

! 3!
Chapter 1
General introduction

! 4!
Spermatogenesis
Spermatogenesis is a process to produce male gamates and a complex biological
process of cellular transformation. This process produces haploid gamates from
diploid spermatogonial stem cells, and the final maturation step in which an
elongated spermatid differentiate into a sperm is called spermiogenesis. During the
spermatogenesis, spermatogonial cells are transformed into spermatozoa over an
extended period of time, about 32 days in mice, within seminiferous epithelium
boundaries of the testis (Fig 1-1). The first cycle of spermatogenesis after the birth is
the synchronized process called the first-wave of spermatogenesis (Fig 1-2). This
precisely timed and highly organized spermatogenic cycle is essential for continuous
sperm production, which is dependent upon numerous factors; both intrinsic (Sertoli
and germ cell) and extrinsic (androgens, retinoic acids), as well as being species-
specific genetic background (1). The primary endocrine hormones are the
gonadotrophins, LH and FSH that are synthesized and secreted by the pituitary gland.
Gonadotropin receptors are expressed only by somatic cells within the testis. The LH
receptors are present on Leydig cells and the FSH receptors on Sertoli cells.
Synthesis of testosterone in the adult Leydig cells is stimulated by LH (2).
Russell, et al. (3) established a 12 staging method of the spermatogenic cycle
in the mouse following the Oakberg's 12-stage classification scheme (4). The
spermatogenic “cycle” is the sum of the stages, and the duration of the cycle is the
period taken for a single germ cell to pass through each of the stages. In rodents, the
first wave of the spermatogenetic process starts a few days after birth, when
gonocytes (prespermatogonia) resume proliferation and become undifferentiated
spermatogonia or spermatogonial stem cells (SSC). Once SSC are engaged in
differentiation process, the cells undergo six successive mitotic divisions, giving rise

! 5!
to meiotic spermatocytes. The spermatocytes go through meiosis which is composed
of two consecutive divisions, with a single round of DNA replication, to give rise to
haploid spermatids. The final step in spermatogenesis consists of the transformation
of spermatids into spermatozoa and this process is specifically called spermiogenesis.
Meiosis
Meiosis is a unique nucleic division to produce haploid nucleus in the germ cell. In
sexual reproduction, in order to inhibit the nucleus from doubling the number of their
chromosomes over generations, reproductive cells experience two meiotic divisions
during gametogenesis. Whereas meiotic division shares many molecular mechanisms
with mitotic division (5), meiotic progression accomplishes several specialized
mechanisms because of two chromosomal distributions without S phase. In many
animals and asexual cells with 2N chromosomes, these cells have each chromosome
containing a pair of two homologous chromosomes originated from their parents but
with subtle difference. On the other hand, in haploid germ cells, each chromosome
contains one homologous chromosome, and the cells differentiate into specialized
germ cells called gamete, such as egg or sperm.
Meiosis is characterized by an extended prophase, followed by two divisions
that produce gametes. In male mammal, a population of spermatogonial stem cells is
present at birth, and meiosis is initiated in maturing cohorts of spermatogonia,
resulting in the continuous production of sperm throughout the reproductive lifespan.
During the prophase of meiosis I, homologous chromosomes pair and synapse, which
is mediated by a meiotic framework called the synaptonemal complex (SC). The
substages of meiosis I prophase is defined by chromosome configurations and
structure (6).

! 6!
Meiotic prophase is classified into four substages, leptotene, zygotene,
pachytene, and diplotene, as shown in Fig 1-3. During leptotene stage, the
homologous chromosomes start to align, although they are not yet fully paired. Axial
element (AE) is a chromosomal scaffold, derived through the cohesin proteins, such
as REC8 and SMC1B, and SC-specific proteins, such as SYCP3 and SYCP2 (7, 8).
The chromatids experience genetically programmed double-strand DNA breaks
induced by SPO11. These DNA breaks are recognized by homologous recombination
repair machinery, such as phosphorylation of H2AX to form gammaH2AX by ataxia
telangiectasia mutated (ATM), which triggers binding of RECA-related proteins, such
as DMC1 and RAD51. These proteins co-localize to electron-dense structures called
recombination nodules (RN) along the developing AE (9). By zygotene stage,
homologous chromosomes start to pair each other and synapsis is initiated. This
initiation of synapsis facilitates the formation of the SC, and the AE begin to “zip”.
Through this process the AE become the lateral elements (LE) of the SC. Pachytene
stage is defined by completion of synapsis, at which point the central element (CE) of
the SC is apparent. CE is composed of proteins such as SYCP1, synaptonemal
complex central element protein 1 (SYCE1) and SYCE2 (10). The pachytene stage is
lengthy and includes maturation of a subset of meiotic recombination sites (<10%)
into crossovers marked by the mismatch repair proteins MutL protein homologue 1
(MLH1) and MLH3, which also co-localize to RN (11, 12). After recombination is
completed, chromosomes undergo desynapsis and condense in the final diplotene
substage. At this stage, the homologues are held together by the recombination sites
(crossovers), which are seen in cytological preparations as chiasmata (6).

! 7!
Mutagenesis
In recent years, several numbers of mouse models that show impaired
spermatogenesis have been made for genetic research. Whole-genome mutagenesis is
a powerful tool for this purpose. N-ethylnitrosourea (ENU) is one of frequently used
alkylating agents, which causes random mutations throughout the genome of all types
of cells, importantly germ cells (13). The Jackson Laboratory created a large number
of mouse infertility models by ENU-induced mutagenesis under the Reproductive
Genomics Program (14). Fig 1-4 shows the mating scheme to create infertile mutant
strains. High mutation load by ENU in treated (G0) males causes high rate of death
due to conditions, such as lymphoma. In order to reduce such conditions, mutated G0
males are bred with wild type females to generate a G1 generation, each of which are
considered to be a line founder during the relatively short period of the time. G1
males are then bred to produce the G2 and backcrossed to produce G3 generation that
will be homozygous for a series of mutations generated in the G0 mouse. Recessive
mutations affecting male infertility (assuming a 100% percentage rate) will be
detected in about 25% of G3 males within an affected line (15). G3 males and females
that fail to reproduce by natural mating to wild-type animals will be used for
secondary phenotype screening to assess gonad and accessory organ histology,
hormone production, gamete production and gamete function in fertilization. The
genetic transmission of the infertility trait in each family is confirmed and each
mutation is genetically mapped to a defined chromosome region, facilitating
identification of candidate genes from DNA sequence and expression database (14).

! 8!
repro34
repro34 is one of infertile mutant strains created by the Reproductive Genomics
Program using ENU at The Jackson Laboratory (14). repro34 males, but not in
females, show infertility due to defective spermatogenesis. Previous study using
linkage analysis and positional candidate approach identified Syntaxin2 (Stx2) gene,
also known as Epimorphin (Epim), on chromosome 5 as the gene responsible for
repro34 (16). A nonsense mutation of the Stx2 gene of repro34, a nucleotide
substitution of C to T in exon 3, led to a premature termination at codon 41. Because
of this non-sense mutation, the most of STX2 domains are expected to be lost in
repro34 (Fig 1-7). This mutation affected specifically male reproductive function
causing defective spermatogenesis with no apparent dysfunction in the other organs in
Stx2repro34. Homozygous females are normal in reproductive function (16).
Syntaxin2/Epimorphin
Epimorphin was initially identified on the surface of mesenchymal cells of mouse
embryonic tissues as the target of a monoclonal antibody that blocked hair follicle
morphogenesis in the dermal epithelium (17). Soon after the identification of
epimorphin, HPC1 (later renamed syntaxin), a neuronal cell-specific membrane
protein, was discovered (18, 19), and it was revealed that this syntaxin/HPC1 protein
was similar in structure to epimorphin but lacked a signal peptide at the N-terminus
(17). Epimorphin is now known to be involved in the morphogenesis and
development of many epithelial organs (20). Epimorphin is also known to mediate
vesicle trafficking (21) and localize throughout many epithelial organs including
intestine, pancreas, mammary gland, lung, gallbladder, liver, skin, kidney, and hair,

! 9!
where epimorphin is secreted from stromal cells (22) in the form of absence of N-
terminal signal peptide (23), playing a role in morphogenesis (Fig 1-8).
Syntaxin2 was known to be identical to nervous system-specific syntaxin1A in
rat, and thought to be important for calcium-regulated secretion during the docking of
synaptic vesicles with the presynaptic plasma membrane (18). Syntaxin2
(Epimorphin) consists of a three-helical N-terminal bundle (3-Hlx), namely SynN
domain, connected by a flexible linker to a C-terminal sequence containing the
SNARE and transmembrane (TM) domains (24), namely t-SNARE domain (Fig 1-6).
Sixty percent of the amino acid sequence of epimorphin/syntaxin2 is identical to
syntaxin/HPC1 (23). However, deletion analysis of epimorphin has revealed that the
morphogenic and vesicle-fusion functions can be separable, in which the C-terminal
SNARE and TM domains were essential for syntaxin-mediated vesicular fusion (24,
25), but they are dispensable for epimorphin-mediated morphogenic activity (23),
thereby N-terminal SynN domain containing three alpha helicles is required for
epimorphin activity (20) (Fig 1-6). In addition, for epimorphin to be functional, it
appeared to change its membrane orientation by unknown mechanism, and by loosing
its TM domain epimorphin would perform its extracelllular function (23) (Fig 1-6). It
has also been shown that membrane topology of proteins lacking N-terminal signal
peptides but containing hydrophobic signal-anchor domain (like epimorphin and
syntaxin family members) largely depends on the three-dimensional configuration of
N-terminal sequence (26, 27). This may indicate that intramolecullar interactions
within the epimorphin sequence allow a subpopulation to orient itself on the outer cell
surface (23). Furthermore, The expression and the subcellular localization of
epimorphin appeared to be highly regulated (23).

! 10!
In mammalian cells, Syntaxin2 is involved in cytokinesis through two possible
mechanisms. In the first possibility, Syntaxin2 may increase the surface of plasma
membrane during furrowing by mediating the fusion of vesicles with plasma
membrane close to the site of ingteression. On the other hand, in the second
possibility, syntaxin2 may have a direct involvement in the final abscission of the
midbody to completely separate into daughter cells (28).
In the testes, syntaxin2 is known to localize at acrosomal compartment,
playing a critical role in acrosomal reaction (29-31). Furthermore, Akiyama, et al.
(16) and Wang, et al. (32) have previously indicated its involvement in
spermatogenesis. Both Stx2repro34 and Stx2-knockout (KO) show a number of
multinucleated spermatocytes and no spermatid was observed in the seminiferous
tubules. Spermatogenic arrest in these mutants indicates syntaxin2 plays an important
role during spermatogensis, but its function has not yet been revealed.
The intercellular bridges of the germ cells
Mammalian germ cells are connected as syncytium through intercellular bridges that
exist between the spermatogenic cells at the same developing stage (33, 34). The
intercellular bridges are large open channels between clones of spermatogenic germ
cells (35, 36), and stable formation of the intercellular bridges is essential for
spermatogenesis and fertility (37). Disruption of the intercellular bridges prevents
normal formation of syncytium, which arrests spermatogenesis prior to meiotic
prophase, and resulted in male-restricted sterility (37). Although several roles of
intercellular bridges have been hypothesized (38), two hypothesizes are currently
accepted (5). The first hypothesis advocate that “cytoplasmic sharing” of essential
signals, such as transfer of mRNA, proteins, and ribosomes, between interconnected

! 11!
germ cells is required for the synchronous cell divisions (39-41). In the second
hypothesis, the communication between germ cells coordinates some of critical steps
of spermatogenesis, such as the entry of meiosis, and this intercellular communication
is controlled by the relatively rapid transfer of unknown signals or substances through
the intercellular bridges (42, 43). Although many components for intercellular bridges
have been reported, TEX14 is the only essential component (37). In addition, genes
essential for cytokinesis and formation of ring canals are not necessarily required for
ring canals or fertility in female (37, 44).
Formation of intercellular bridges shares many molecular mechanism with
cytokinetic division (5), although most aspects of the signaling process during
cytokinesis remain unknown (45). Proper formation of the intercellular bridge owes a
lot to retaining a midbody, a central part of the bridge which consists of various
molecules. In somatic cytokinesis, recruit of TSG101 and ALIX to the midbody and
their association with CEP55, a component of the midbody, trigger abscission of the
midbody and consequence to complete division of the daughter cells. This hypothesis
is proven by the appearance of multinucleated HeLa cells through siRNA inhibition
of CEP55 (46). In the case of male germ cells, TEX14 is speculated to disturb the
association of TSG101 and ALIX with CEP55 so that the midbody stays stable and
the intercellular bridge are maintained (47) (Fig. 1-8).
Glycolipids
In mammals, two major sulfoglycolipids are produced in the testis; sulfatide and
seminolipids, through catalyzation of alkylacylglycerol (EAG) by common enzymes,
cerebroside sulfotransferase (CST) (48, 49) and ceramide galactosyltransferase (CGT)
(50) one after another. Consequently alkylacylglycerol and ceramide are converted to

! 12!
seminolipids and galactocylsulfatide, respectively (48, 50, 51) (Fig. 1-9). Seminolipid
is synthesized in spermatocytes and maintained in the subsequent germ cell stages
(52). Significance of glycolipids in spermatogenesis has been evidenced with the
arrested spermatogenesis in CST and CGT deficient mice (48, 50). Zhang, et al. (53)
proved that synthesis of glycolipids in germ cells is essential for spermatogenesis,
however the precise function of glycolipids in spermatogenesis has not yet been
elucidated.
Glycans are expressed specifically in the mammalian testis. Seminolipids are
sulfated glycolipids and their distribution is restricted to the testis. Seminolipids
consist of 90% of glycolipids in the testis (54). The sugar chain of seminolipids shows
the same structure as sulfatides that are abundant in myelin sheath, but lipids chain of
seminolipids consists of glycelolipids that binds to ether. The sugar parts of
seminolipids and sulfatides are biosynthesized through CGT and CST. The
homologous deletion of CGT (50) and CST (53) resulted in male-restricted sterility.
In the testis of CGT KO mice, galactosyl-alkyl-acyl-glycerol and seminolipids are
depleted, and spermatogenesis was arrested prior to meiotic division (55, 56). On the
other hand, spermatogenesis of CST KO was arrested at metaphase of meiosis I (48).
These observations indicated that these two testis-specific glycolipids have similar but
distinct roles in mammalian spermatogenesis. Futhermore, transplantation of normal
spermatogonial germ cells into the seminiferous tubules of CST KO mice that
contained glycolipid-depleted somatic cells observed normal spermatozoon (53).
Taken together, glycolipids are required for germ cells themselves. Glycolipids seem
not to be required for the normal function of somatic cells in the seminiferous
epithelium,

! 13!
Several studies suggested that lipids play an important role in regulation of
cytokinesis as a component of lipid-order microdomains, termed lipid rafts (57), and
lipid rafts are thought to be platform of signaling molecules. Seminolipids are
produced in the Golgi membrane of spermatocytes (54) and transported to the plasma
membrane by vesicle transportation (52, 58). Glycolipids including seminolipids are
normally localized to the plasma membrane of spermatocytes or round spermatids,
but not on the spermatogonia (59), and testis-specific glycolipids are also thought to
be a component of lipid rafts on sperm (60). Several studies also suggested that
plasma membrane raft domains are formed in the furrows of dividing cells, indicating
an important role in cytokinesis (57). In addition, seminolipid in sperm is thought to
be a component of lipid rafts and to function for sperm to attach Zona pellucidae (ZP)
to penetrate during capacitation (60).

! 14!

! 15!

! 16!

! 17!

! 18!

! 19!

! 20!

! 21!

! 22!

! 23!
Chapter 2
Multinucleation of the germ cells in the
Stx2repro34 mice

! 24!
Introduction
In the previous study, infertile Stx2repro34 mutants showed spermatogenic failure
during meiosis, and this abnormal spermatogenesis was caused by a mutation on the
Stx2 gene (61). Homologous inactivation of Stx2 also showed similar spermatogenic
failure (62). These observations indicate the essential role of Stx2 in spermatogenesis.
However, the precise function of Stx2 in spermatogenesis is largely unknown.
Therefore, this study focused on further examination of the phenotype of aberrant
spermatogenesis in the Stx2repro34 mutant by histological and cytological analysis.
Firstly, in order to investigate the transition of spermatogenesis in the
Stx2repro34 mutant, the testis sections during the first-wave of spermatogenesis was
examined. When the spermatogenesis is initiated, the spermatogenic cells undergo
synchronized process within seminiferous tubules. During the first-wave of
spermatogenesis, primitive type A spermatogonia are first appeared on the day 6 after
the birth, and meiotic prophase I is initiated on the day 7 to 9 (63). Pachytene
spermatocytes first appeared on from day 12 to 15 (63, 64). Previous study revealed
that the expression of Stx2 start to increase at day 16 after the birth (P16) in the testis
(61), indicating that Stx2 plays an important role after pachytene stage.
Secondly, since the phenotype of spermatogenic defect in the Stx2repro34 was
characterized by remarked high frequency of multinucleation of the germ cells (61).
This study next examined the cytological effects of multinucleation by
immunohistrogical and immunocytrogical analysis using antibodies against several
maker proteins. GammaH2AX is a phosphorylated form of histone variant H2AX on
Serine 139 (65, 66). GammaH2AX marks chromatin domains with DNA damage,
including double strand breaks (DSBs) (66). During meiotic prophase, the
spermatocytes that are successful with repair of DSBs only exhibit restricted

! 25!
expression of γH2AX only in the chromatin of the XY body (sex chromosome) at
pachytene or diplotene stages (65, 67). By observing the pattern of γH2AX labeling in
spermatocytes, the extent of DNA repair machinery can be examined.
Multinucleation of germ cells often appears in the pathological testicular
conditions. One of ceuases for the formation of these abnormal cells is opening the
intercellular bridge, a stable cytoplasmic extension that interconnect a large number
of spermatogenic cells (3). Multinucleation of somatic cells is caused by two
mechanisms; cell-cell fusion and acytokinetic cell division (68). Hence, this study
thirdly examined the formation of the intercellular bridges in the Stx2repro34 by
observing immunohistochemical localization of a component of the intercellular
bridges. Although the intercellular bridges in germ cells have distinct functions and
are different from the interconnections in somatic cell cytokinesis, the molecular
mechanisms to form the intercellular bridges share the large part of those in somatic
cytokinesis (5). The intercellular bridges are large open channels between clones of
spermatogenic germ cells (35, 36), which continuously formed from the stage of
undifferentiated spermatogonia to that of elongated spermatids. Since TEX14 plays an
important role in stable formation of intercellular bridges of the germ cells, this study
observed localization of TEX14 in the multinucleated cells in the Stx2repro34 mutant.
In the Stx2repro34 mutant, multinucleation of germ cells can be observed in
pachynema. During pachytene stage, one of important events, meiotic synapsis, is
completed, which is essential for successful meiosis. Fourthly, in order to reveal the
status of synapsis during meiotic prophase I, surface spread chromosome preparation
was examined by immunocytrogical localization of some marker proteins including
SYCP3, SYCP1, and γH2AX. SYCP1 and SYCP3 are known to form during
prophase I as main components of two side rails of the synaptonemal complex (SC).

! 26!
SYCP1 functions in synaptonemal complex assembly, meiotic recombination, and
XY body formation (69), and SYCP3 is required for the formation of LEs as a main
structural element of the LE (70). Homologue juxtaposition during prophase I can be
divided into three structurally distinct steps, recognition, presynaptic alignment, and
synapsis by the SC (6). In many organisms, these three steps are distinguished by
their dependence on DNA DSBs (71, 72). Since γH2AX foci display the DNA DBSs
(66), fluorescenceimmunohistochemical staining of γH2AX indicates the status of
DNA integrity and chromatic organization.
As previously reported, the spermatogenesis in Stx2repro34 was arrested during
meiotic divisions since disrupted formation of metaphase plate (equatorial plate) and
many degenerating spermatocytes at metaphase were observed (61). In order to
complete a series of meiotic divisions, spermatogenic cells must undergo proper
assemble and segregation of chromosome contents. Abnormal assemble of
chromosome during metaphase resulted in Metaphase-specific cell death (73, 74).
This study fifthly conducted TdT-mediated dUTP nick end labeling (TUENL) assay
that detects DNA fragmentation of these denatured germ cells as an indicator of
programmed cell death, namely apoptosis (75).
In meiosis, a spermatocyte divides into four haploid daughter cells through
two meiotic divisions. During metaphase, homologous chromosomes align in the
middle of the cell before being separated into each of the two daughter cells.
Metaphase accounts for approximately 4% of the cell cycle's duration, and 60% to
70% of them are primary spermatocytes and the remaining is the secondary
spermatocytes (3). Microtubules formed during prophase attached themselves to
kinetochores during metaphase since prometaphase, followed by anaphase. The
centromeres of the chromosomes then are gathered on the metaphase plate in between

! 27!
two centrioles. This alignment is enabled by the counterbalance of the pulling power
generated by the opposing kinetochores (76). During metaphase I, SYCP3 protein co-
localize at the surface of contact region between sister chromatids, so-called
interchromatid domain or centromeres in bivalents (77). Hence, this study, next,
examined the cytological localization of SYCP3 in the spermatocytes at metaphase.
During prometaphase and metaphase, without all the chromosomes being
aligned at the metaphase plate, the cell would not be able to enter anaphase,
controlled by one of the cell cycle checkpoints, called spindle checkpoint (78). In the
case of meiosis, the consequences of error during meiotic division in spermatogenesis
would cause serious problems, such as aneuploid spermatozoa, embryonic lethality,
and developmental abnormalities (79). In mouse oocytes, failure of chromosome
segregation resulted in arrest of meiotic progression at metaphase I (80). In
mammalian cells, mitotic spindles are organized at the time of nuclear envelope
breakdown (NEBD), when two active microtubule-organizing centers located at
opposite poles of he nucleus start to emanate microtubules. A bipolar spindle is
organized when the plus ends of antiparallel microtubule arrays meet at the cell
equator (metaphase plate). During prometaphase, the chromosomes congress toward
the spindle equator where they ultimately attain an equilibrium position at metaphase.
Since a number of multinucleated spermatocytes at metaphase containing multiple
metaphase plates were observed in the Stx2repro34 (61), this study next performed
examination of structurally preserved chromosome preparation to visualize the
construction of spindle body.
Lastly, this study focused on the cellular fate of aberrant germ cells in the
Stx2repro34 mutant, since round spermatid-like germ cells were occasionally observed

! 28!
in the seminiferous epithelium. These abnormal cells expressed spermatid-specific
proteins, including IZUMO, which is required during fertilization (81, 82).

! 29!
Material and methods
Animals
The Stx2repro34 mice used were provided from the mouse mutant resource colony of
the Jackson Laboratory (JAX) (Bar Harbor, Maine, USA). The Stx2repro34 mutation
was induced in a C57BL/6J background and subsequently outcrossed to C3HeB/FeJ
inbred strain (CLEA Japan, Tokyo, Japan). A congenic line for the Stx2repro34 was
created on C3HeB/FeJ. For biological analyses, Stx2repro34 homozygotes and control
wild-type littermates at 10-12th weeks of age or at 1-3rd weeks of age for the sake of
first-wave of spermatogenesis were used. All the experiments using animals were
approved by the regulation of animal protection for the laboratory and animal
husbandry of Okayama University. Mice were kept in the animal facility of the
Okayama University, following the law of animal protection for the laboratory and
animal husbandry of Okayama University. The condition of the breeding-room was
maintained constant throughout the year, under 23-26°C temperature, 30-60%
humidity, and 12 hour light-dark cycle condition. Standard pellet diet (CE-2, CLEA
Japan) was fed to mice. Mice were euthanized by the painless method using carbon
dioxide (CO2) gas.
DNA extraction from tail
Genomic DNA was prepared from cut tails or fragments of ear of mice. These tissue
samples were collected at the weaning day as a part of identification. To break cell
structure and digest DNA-protecting proteins, the tissue samples were treated with 4
µl of ProteinaseK (10 mg/ml) in 150 µl of NP-40 detergent (0.14 M NaCl, 10 mM
EDTA, 100 mM Tris-HCl, 0.5% NP-40) and 150µL of 2X SDS (2% SDS, 20 mM
EDTA, 0.2 M NaCl) at 55°C overnight. Adding 4 µl of RNase A (10mg/ml) (nakarai

! 30!
tesque, Kyoto, Japan) to the mixture to digest RNA, the samples were incubated at
37°C for 2 hours. Protein residual substances and fragmented cellular debris were
separated by adding 300 µl of Phenol/Chloroform and 30 µl of 3 M sodium acetate
(pH 5.2). After gentle inverting for 5 minutes, the mixture was centrifuged at 12,000
rpm for 15 minutes at room temperature (RT). The supernatant was transferred to a
new tube containing 600 µl of anhydrous ethanol, and mixed well. Followed by
centrifuging at 12,000 rpm for 15 minutes at 4°C, the ethanol solution was replaced
with 100 µl of 70% ethanol, and centrifuged at 12,000 rpm for 15 minutes at 4°C. The
ethanol solution was removed to dry, and the collected DNA was dissolved in 50 µl of
TE buffer (10 mM Tris HCL and 1 mM EDTA in milli-Q water, pH 8.0).
Genotyping of Stx2repro34
For genotyping of Stx2repro34, template genomic DNA extracted from tail or ear, as
stated above, was used. The reaction mixture for PRC for each sample contained 0.2
µl of forward (5'-GTGGGATCACGAGTCACTCACT-3') and reverse (5'-
GAGTGAGTTCCACGACAGCCA-3') primers (10 µM), 1 µl of 10X PCR
amplification buffer, 1 µl of dNTP solution, 0.05 µl of Tap Polymerase (5 U/µl)
(Amersham Bioscience, Piscataway, NJ, USA), 1 µl of DNA sample (20 µg/ml) and
6.55 µl of autoclaved milli-Q water. Amplification of nucleic acids was processed by
denaturation, annealing, and polymerization cycles; first denaturation at 94°C for 5
minutes, followed by 40 cycles containing denaturation (94°C for 30 seconds),
annealing (58°C for 30 seconds), and polymerization (72°C for 30 seconds), finished
by the final polymerization step at 72°C for 5minutes. The PCR products were
digested with TaqαI restriction enzyme at 65°C for 16 hours, and thereafter analyzed
by electrophoresis through a 2.5% agarose gel. Wild-type and heterozygous mice

! 31!
showed digestion of amplified DNA fragment, creating 258 and 102 bp fragments,
while the amplified DNA fragment from homozygous mice remained undigested
(Fig1-6B).
Histological analysis by Mayer's HE staining
For histological analysis, mice were euthanized by CO2 gas, and testes and
epididymis were immediately extracted for fixation. The whole testes and epididymis
were fixed in Bouin's solution (picric acid: formaldehyde: acetic acid=15:5:1) for 12-
16 hours at 4°C. The fixed samples were then washed with ethanol from 70% x3,
80%, 90%, 95%, up to 99.5% for 1 hour each, and completely dehydrated in
anhydrous ethanol overnight. The tissue samples were placed in Xylene three times at
RT and paraffin three times at 60°C for 1 hour each, and embedded in paraffin block.
Testes samples were sliced by 5 µm using a microtome (RV-240, Yamato, Japan),
and pasted on slide glasses. After drying the slides on paraffin stretcher overnight, the
slides were de-paraffined in Xylene three times for 5 minutes each, and rehydrated
through ethanol at down graded concentrations from 100% twice, 95%, 90%, 80%, to
70% for 3-5 minutes each. Followed by staining in hematoxylin dye for 5 minutes, the
slides were washed by flowing tap water for about 10 minutes until the tissues were
decolorized to light staining color. The slides were then stained in eosin dye for 1
minute. After quick wash in tap water, the slides were decolorized in 70%, 80%, 90%,
95%, 100% ethanol for less than 30 seconds each to adjust the eosin color, and placed
in Xylene three times for 5 minutes each. The preparation slides were sealed using
Entellan New (Merck, USA). Stages of the seminiferous epithelium cycle were
identified according to established morphological criteria (3).

! 32!
TUNEL assay (Fluorescent)
To distinguish apoptotic cells, TUNEL assay was performed using in situ Cell
Detection Kit (Cat#11684817910, Roche, Basel, Switzerland) according to the
manufacture’s instruction with some modifications. Sectioned testis slides were
treated in the same manner as histological analysis until the deparaffin treatment by
Xylene and hydrophilic treatment by ethanol. 25 µl of Proteinase K (20 µg/ml) in 10
mM Tris-HCl (pH 7.4-8.0) were applied to testes section samples circled by PAP pen
(Daido Sangyo, Tokyo, Japan), and incubated for 20 minutes at 37°C. Followed by
two time washes with PBS, one sample was treated with 25 µl of DNase (1 mg-100
µg/ml) as positive control or 10 minutes at RT. Tissue samples were incubated with
25 µl of TUNEL reaction mixture for 60 minutes at 37°C. After three times of wash,
the preparation slides were sealed with VECTASHIELD Mounting Medium with
DAPI (H-1000, Vector Laboratories. Inc. Burlingame, CA, USA), and observed using
ZEISS AXIO Imager. A1 upright microscope (Carl Zeiss AG, Oberkochen, Germany)
with DFC480 Digital Camera System (Leica Microsystems, Wetzlar, Germany).
TUNEL assay (DAB staining)
For non-fluorescent TUNE assay, DAB staining TUNEL assay was performed using
in situ Cell Detection Kit (Roche, USA) according to the manufacture’s instruction
with some modifications. The procedure was the same as fluorescent TUNEL assay
until the first PBS wash after deparaffin and hydrophilic treatment. To block inherent
peroxidase (POD), 20 µl of 3% H2O2 (nacalai tesque, Japan) in methanol was applied
to the tissue sections for 10 minutes at RT. After three time washes with PBS, the
tissue samples were incubated with 30 µl of Proteinase K (20 µg/ml) for 5 minutes at
RT. Followed by three time washes with PBS, the tissue samples were then reacted

! 33!
with TUNEL reaction mixture (TUNEL enzyme: buffer=1:9) for 20 minutes at 37°C.
The tissue samples were washed ten times with PBS, and incubated with 20 µl of
POD reaction mixture for 20 minutes at 37°C. Washing with PBS once, DAB mixture
(DAB substrate: buffer=1:50-100) was applied to the tissue samples for 2-3 seconds,
and immediately washed well with tap water to stop the reaction. The tissue samples
were conterstained with hematoxylin for 2 minutes, and the color of the dye was
adjusted by dipping through tap water for about 20 minutes. After dehydration
treatment with graded concentration of ethanol and xylene, the slide preparations
were sealed with Entellan new (Merck KGaA, Darmstadt, Germany).
Surface spread preparations and immunostaining
The surface spread preparation of testicular cells was prepared as previously
described with some modifications (83-85). Briefly, cells were collected in DMEM
medium (12100-046, Invitrogen, Carlsbad, CA, USA). After centrifugation at 1,800
rpm for 10 minutes at 4°C, pelleted cells were suspended in 0.5% NaCl solution then
spread on silanized slides. The preparation slides were soaked in 2% PFA containing
0.02% SDS in PBS (pH 7-8 was adjusted with 0.1 M Na2B4O7). After washing with
PBS containing 0.04% DRIWEL (Fuji photo film co., LTD, Tokyo, Japan) then dried,
preparation slides were stored in -80°C until use.
For immunostaining, after blocking with antibody dilution buffer (ADB; PBS
containing 2% BSA and 0.05% Triton-X 100), the slides were incubated with anti-
SYCP1, anti-SYCP3 (86) and anti-gammaH2AX in PBS containing 10% ADB
overnight at 4°C. Secondary antibodies, goat anti-mouse IgG-Alexa Flour 488 (A-
21121, Invitrogen) at 1:200 dilution and goat anti-guinea pig IgG-Alexa Fluor 594
(A11076, Invitrogen, Carlsbad, CA, USA) at 1:200 dilution, or bovine anti-rabbit

! 34!
IgG-FITC (SC-2365, Santa Cruz, CA, USA) at 1:400 dilution in PBS containing 10%
ADB, were used for 1 hour at RT. The slides were then mounted with
VECTASHIELD Mounting Medium (H-1200, Vector Laboratories Inc.) and
observed.
Structurally preserved chromosome preparation
Structurally preserved chromosome preparation was made following the previous
method (87) with some modifications. Testes were dissected out into test tubes
containing 1 ml of ice-cold Ringer solution (155 mM NaCl, 5.6 mM KCl, 2.2 mM
CaCl2, 2.4 mM NaHCO3) for 4 hours, and subsequently minced in the microtubule-
stabilizing buffer (100 mM PIPES (pH 6.8), 1 mM MgSO4, 1 mM EGTA, and 1%
Triton X-100). After centrifuging at 1,500 rpm for 5 minutes, collected suspension
containing germ cells was spread onto slides. The slide preparations were fixed in the
fixative (0.25% glutaraldehyde, 2% formaldehyde, in the microtubule-stabilizing
buffer without detergent) for 30 minutes. For further stabilization of cytoskeletons,
the slides were immersed in methanol at -20°C. The preparation was then treated with
NaBH4 (0.5 mg/ml in PBS), and washed three times in PBS containing 0.02% NaN3
and 0.1% Triton X-100 and once for 5 minutes with the same solution without the
detergent. For immunofluorescence reaction, the cells were reacted with anti-α-
TUBULIN antibody and secondary antibody anti-mouse IgG-Alexa Flour 488 (A-
11008, Invitorogen). After incubation, the slides were sealed with
VECTASHIELDMounting Medium with DAPI, and observed using KEYENCE
BIOREVO BZ-9000.

! 35!
Giemsa-staining air-dried chromosome preparation
Procedure and identification of cell stages were referred to the previously described
methods (88-91) with some modifications. The testes were quickly extracted from
live animals and collected samples were placed in 20 ml of 2.2% sodium citrate in a
laboratory dish. Tunica albuginea was then pealed off, and a cluster of seminiferous
tubules was disentangled using tweezers. These tubules were treated with 1% sodium
citrate solution for 30 minutes at room temperature. The hypotonic solution was then
changed with fixative (Methanol: Acetic acid=3:1) at 4°C for 10 minutes. Followed
by changing the old fixative with new one, the fixed tubules were then transferred to a
new tube containing 50% acetic acid, and waited until germ cells became suspended
due to the tubules being melted. After centrifugal separation at 1500 rpm for 10
minutes, the cell precipitation was resuspended in the same fixative for further use. A
few drops of cell suspension were dropped on a grass slide from the distance above.
The grass slide was placed on the hot plate whose temperature was 37°C to dry. The
chromosomes were stained with 5% Giemsa-solution (Sigma-aldrich, Missouri, USA)
for 5 minutes. The stained slide was gently washed with tap water, and left to dry for
a while. The slide preparation was sealed with Entellan new (Merck KGaA,
Germany).
Immunohistochemistry
Testes from mice and rat were fixed in Bouin's solution or 4% PFA fixative (4%
paraformaldehyde, 0.1 M phosphate buffer, pH 7.4). They were dehydrated then
embedded in paraffin. Sections at 5 µm thickness were cut and deparaffinized. For
immunostaining, deparaffinized sections were autoclaved in 0.1 M Sodium Citrate,
pH6.0, at 120°C for 20 minutes, or the sections were treated with 20 µg/ml

! 36!
ProteinaseK in 10 mM Tris-HCl (pH 7.4) at 37°C for 20 minutes for antigen retrieval,
when necessary. All the sections were incubated in 3% H2O2 in PBS for 15 minutes to
quench the endogenous peroxidase. They were then incubated in PBS containing 10%
BSA for 30 minutes at RT to reduce background. Sections were reacted with primary
antibodies diluted with PBS containing 1% BSA for 3 hours at 37°C. The antibodies
used and each conditions for staining were shown in Table 2-1. Subsequently HRP-
labeled secondary antibodies (Santa Cruz, CA, USA), were reacted at 1:400 dilution
for 1 hour at RT. Antigens were visualized using TSA Plus DNP (HRP) System
(Perkin Elmer, MA, USA) with NEL 747B Dako Liquid DAB+ Substrate Chromogen
System (20 µl/ml) (K3467, DAKO, Glostrup, Denmark). Sections were
counterstained with hematoxylin.

! 37!
Results
Affected spermatogenesis in the Stx2repro34 mutant
In the testis of adult Stx2repro34 mutant males, multinucleation of the germ cells was
observed in pachynema (Fig. 2-1A), diplonema (Fig. 2-1B), spermatocyte at
metaphase (Fig. 2-1C), and round spermatid-like germ cells (Fig. 2-1D). The round
spermatid-like germ cells expressed spermatid-specific protein IZUMO (Fig. 2-9D,E),
an acrosome-localized protein required for fertilization (81). Further histological
analysis observed that many degenerated germ cells were observed in the epididymis
of Stx2repro34 (Fig. 2-1F), while the epididymis of wild-type was filled with mature
spermatozoa (Fig 2-1E).
The postnatal Stx2repro34 mutant males were indistinguishable from the wild-
type males in growth (Fig. 2-2B,C), but the testis weight started to express
differences between two types of males P16 (Fig. 2-2D). The testicular size of
Stx2repro34 mutant is smaller than that of wild-type, although other reproductive organs
of male Stx2repro34 mutant appear to be normal (Fig. 2-2A). Histological examination
of the testes from the mice during first-wave of spermatogenesis revealed that the
abnormal formation of syncytial multinucleated germ cells was also seen in the
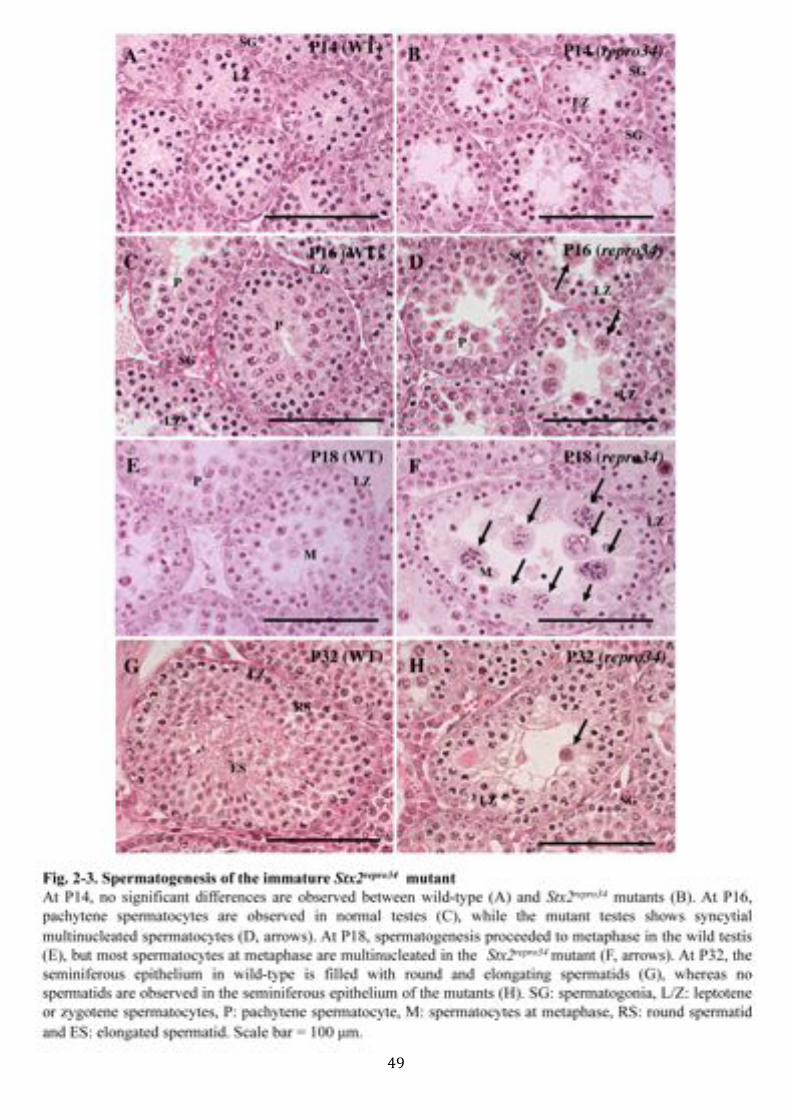
seminiferous epithelium of the Stx2repro34 mutant males. At 14, morphology of germ
cells were indistinguishable between wild-type (Fig. 2-3A) and Stx2repro34 mutant
(Fig. 2-3B). In contrast to the progress of meiosis in the wild-type males (Fig. 2-3C),
the Stx2repro34 mutant males at day 16 post birth (P16) showed the multinucleation of
spermatocytes in the seminiferous epithelium (Fig. 2-3D), which presumably
corresponded to mid to late pachytene stage in the normal testis (63). Subsequently, at
P18, multinucleated spermatocytes presumably at diplotene stage as well as at
metaphase were found in Stx2repro34 mutant males (Fig. 2-3F). At P32, the

! 38!
seminiferous tubules of the wild-type testis were filled in with round and elongated
spermatids, whereas the testis of Stx2repro34 mutant males included neither round nor
elongated spermatids, instead, multinucleated germ cells were observed in the inner
layer of the epithelium (Fig. 2-3H). Overall, the phenotypic features in
spermatogenesis of Stx2repro34 mutant males appeared to be the same in the immature
and adult mice. These observations exclude age-dependent regulation of Stx2 in
spermatogenesis.
Multinucleated germ cells
This study confirmed the arrest of spermatogenesis and multinucleation of germ cells
in the adult Stx2repro34 mutant males by immunohistochemistry. Stage-specific
expression patterns of gammaH2AX in the nucleus showed normal progress of
meiosis up to zygotene stage in the mutant. The expression of was restricted to XY
body in the nucleus of the pachynema in the wild-type male (Fig. 2-4C). The same
pattern of expression was observed in the pachytene nucleus in the mutant. In
addition, the positive reaction was observed in the syncytial germ cells (Fig. 2-4D).
The reactions were shown to be a part of the nucleus by staining with LAMIN-B1.
The nucleus of the multinucleated cells displayed positive reaction when
immunostained with a LaminB1 antibody, which indicates the integrity of the nuclear
membrane (Fig. 2-4A, inset).
This study then investigated the formation of the intercellular bridges in the
Stx2repro34 mutants with a TEX14 antibody. The intercellular bridges are large open
channels between clones of spermatogenic cells (35, 36) and TEX14 is one of the
specific proteins in the bridges (37). In the wild-type, the expressions were dotted
around the spermatocytes and spermatids, showing the connection of the germ cells

! 39!
with the bridges (Fig. 2-4E,G). The mutant testis exhibited the same expression in the
spermatocytes. When examined the multinucleated cells, patchy, not dotted
expression of TEX14 around the cells were observed (Fig. 2-4F,H). Although TEX14
shows ring-shape formation in normal mice (Fig. 2-4E,G), the expression of TEX14
was disrupted around many of multinucleated spermatocytes in the Stx2repro34 mutant
males (Fig. 2-4F,H). This abnormal formation of intercellular bridges was observed
during later prophase, which is consistent with the timing of STX2 protein and Stx2
gene expression (61) and multinucleation (Fig. 2-12). Hence, STX2 localized to the
intercellular bridges is in part required for maintenance of the intercellular bridges
especially during later prophase.
Arrest of spermatogenesis at pachytene stage often accompanied with aberrant
chromosome synapsis, therefore we performed immunostaining of spread
chromosome preparations using antibodies against SYCP3, SYCP1, and
gammaH2AX, maker proteins for meiotic synapsis. Labeling pattern of these maker
proteins indicate extent of synapsis. However, the labeling patterns of these proteins
were indistinguishable between wild-type and Stx2repro34 mutants (Fig. 2-5), indicating
that normal synapsis of homologous chromosomes in the Stx2repro34 mutant males.
These results indicated that STX2 is not involved in chromosomal synapsis.
As stated above, several types of multinucleated cells were observed in the
adult Stx2repro34 mutant males (Fig. 2-1A-D). This study, therefore, performed
immunocytochemistry of these syncytial germ cells obtained by means of squash
technique from the seminiferous tubular segment at stage VIII-I to unveil the
structural features. Expression pattern of SYCP3 at centromere appeared to be normal
in multinucleated spermatocytes at metaphase in the Stx2repro34 mutant males,
compared with that of spermatocytes at metaphase in the normal (77), but the number

! 40!
of the positive spots seemed twice or three times as many as those in the normal (Fig.
2-6A,B). Staining of alpha-TUBULIN revealed that multipolar formation of spindle
body in multinucleated metaphase spermatocytes (Fig. 2-6D). The aberrant
chromosomal contents were confirmed by Giemsa stained chromosome preparation
(Fig. 2-6G,H). More than 60% of metaphase spermatocytes were multiploid and
contained more than 20 bivalents or univalents in the Stx2repro34 mutant males, but
about 40% of metaphase spermatocytes contained 20 bivalents like wild type (Fig. 2-
7). Since secondary spermatocytes were barely observed in the Stx2repro34 mutants
(3.66%/N=86 in Stx2repro34, compared with 29.6%/N=145 in wild type), developing
germ cells were not capable of completion of meiotic division due to failure of
chromosome segregation. TUNEL assay further confirmed remarkably higher
frequency of apoptotic cells in the testis of Stx2repro34 mutant males compared with
wild-type (Fig. 2-1E,F), and stage-specific apoptosis at metaphase (Fig. 2-1G-I).
Meiotic arrest in round spermatid-like germ cells in Stx2repro34
In order to determine more precise timing of meiotic arrest in Stx2repro34 mutant, round
spermatid-like abnormal germ cells were labeled with some meiosis specific markers,
SYCP1 and SYCP3. SYCP1, a component of central element (CE), disappears at the
end of prophase I, and expression of SYCP3, a component of lateral element (LE),
can be observed at centromeres until metaphase I (77). In the tubules of wild-type at
XI, diplonema showed SYCP3 expression (Fig. 2-10A,C), and spermatocytes at MI
also showed SYCP3 expression (Fig. 2-6A). In contract, round spermatid-like
abnormal germ cells showed neither SYCP1 nor SYCP3 expression (Fig 2-10D-F).
As shown above, secondary spermatocytes were barely observed in Stx2repro34 mutants
(Fig. 2-6). These observations suggest that the meiotic progression of Stx2repro34

! 41!
mutants is arrested somewhere between anaphase I and prophase II, probably
interphase.

! 42!
Discussion
Previous study showed gene disruption or a nonsence mutation of Stx2 gene caused
defective spermatogenesis, which lead to formation of syncytial multinucleated
spermatocytes (61, 62). This study further investigated the detail of the phenotype of
the Stx2 mutant repro34 to reveal the function of STX2.
Based on histological observation of the testis of both adult and immature
Stx2repro34 homozygotes, multinucleation of the germ cells was initiated during late
pachytene stage. It has been known that multinucleation of the cell can be caused by
two mechanisms: acytokinesis and cell-cell fusion (68), and in the germ cell,
multinucleation is often caused by abnormal opening of the intercellular bridges (3).
This study also revealed aberrant localization of TEX14, an intercellular bridge
specific protein (37), in the multinucleated germ cells, indicating that unstable
formation of the bridge caused opening of these bridges between syncytialy connected
germ cells. Therefore, it is hypothesized that the direct cause of formation of
multinucleated germ cells is unstable formation/maintenance of the intercellular
bridges in the Stx2repro34 mutant. This hypothesis is verified by the observation that the
nuclei within a giant cell are at the same developmental stage. Thus, the loss of STX2
probably affects the open channels of the intercellular bridges at specific stages,
resulting in the formation of abnormal syncytium.
However, the direct cause of multinucleation of germ cells in the Stx2repro34
remains unclear. Several mutant mice showed aberrant spermatogenesis with
multinucleation of the germ cells due to dysfunction of cytokinetic regulation (92),
dietary toxins (93, 94). In addition, the loss of adhesion between spermatids and
Sertoli cells resulted in multinucleation of symplastic giant germ cells that contained
multiple nuclei (95). Disruption of androgen signals also resulted in multinucleated

! 43!
germ cells (96, 97). Since Sertoli cell and Leydig cell play an important role in
differentiation of germ cells by controlling the movement of signal molecules or
maintaining the environmental surrounding of germ cells (98), the loss of STX2 might
affect the microenvironment of the testes. In several kinds of somatic cells, STX2 is
known to play an important role in vesicle fusion during exocytosis (28). Hence,
STX2 may be involved in paracrine communication between somatic cells and germ
cells in the testes, and loss of this function might result in abberant state of either
germ cells or somatic cells, which leaded to multinucleation.
As stated above, there is a possibility that testicular microenvironment is
affected since spermatogenic process is disrupted during later meiotic prophase.
Meiotic progression at pachytene is one of the most important phases for the
structural assemble of homologous chromosomes for recombination, which is
therefore highly regulated by several check point mechanisms during pachytene (99-
101). However, apoptotic cell death was barely observed in spermatocytes during
meiotic prophase (Fig. 2-1G). However, in the spermatocytes of Stx2repro34
homozygotes, in addition, pairing of homologous chromosomes or construction of
synaptonemal complex were apparently normal (Fig 2-5). Hence, multinucleation did
not affect the process of meiotic synapsis, and STX2 appear to have independent
functions from the meiotic synapsis.
In spite of a high frequency of apoptotic cell death in metaphase
spermatocytes due to spermatogenic arrest through spindle checkpoint, histological
analysis revealed the existence of multinucleated round spermatid-like germ cells (Fig
2-9D,E). It was therefore hypothesized that some spermatocytes were capable of
completion of meiotic progression. Immunostaining of the testes of Stx2repro34 with
two of marker proteins for meiotic prophase, SYCP1 and SYCP3, revealed that the

! 44!
nuclei of these round spermatid-like germ cells were at least completed meiotic
prophase (Fig 2-10). Other study examined haploidity of the nuclei in the syncytial
cells by means of reproductive engineering technique, round spermatid injection
(ROSI). Although the study did neither succeed to obtain siblings by ROSI using the
nucleus of the aberrant cells as donors nor confirm haploidity of the nuclei by
chromosome preparations, vestiges of the implantation of the transferred embryo were
detected. This might suggest that few of the nuclei in the syncytial cell completed
meiosis to have haploid genome. However, it was noteworthy that high rate of traces
of implantation was observed, and some recipient mice bred on the day 15 of
pregnancy. This may imply development of fetus to some extent, but abnormally.
Similar results, such as traces of implantations or abnormally developed fetuses, were
reported by microinjection of artificially made trisomy cell previously tested.
Aneuploidy also often leads to failure of normal development in transplanted embryo
(unpublished data). Chromosomal abnormalities due to aneuploidy in male are known
to cause several genetic diseases, such as several types of cancers (102), Alzheimer's
disease (103, 104), Down syndrome (105), or Turner's syndrome (106).
Contrary to the expectation of aneuploidy in the round spermatid-like germ
cell, no chromosome with aneuploidy was observed from the second trial of
microinjection. Since the nucleus of round spermatid-like cell was not a haploid but in
the status of spermatocytes, the two meiotic divisions in the Stx2repro34 were not
probably completed in full state. The chromosome of spermatocytes was rather
arrested during interphase, but morphologically "differentiate" into round spermatid-
like cells expressing some spermatid specific marker proteins including IZUMO (82)
(Fig. 2-9E) and TRA54 (data not shown).

! 45!
As a concluding remark of this chapter, apparent abnormality in spermatogenic
process was initially observed at late pachytene as a formation of multinucleated germ
cells, and the arrest of spermatogenic process in the Stx2repro34 mutant was probably a
secondary outcome owing to disrupted spindle assembly due to the multinucleation of
the germ cells. The multinucleation of the germ cells appeared to be caused by
disrupted intercellular bridges at specific stages in the Stx2repro34 mutant. Therefore, the
function of SXT2 seems to be related to the formation/maintenance of the intercellular
bridge. The later chapters will focus on this respect based on the observations of this
chapter.

! 46!

! 47!

! 48!
! 49!

! 50!

! 51!

! 52!

! 53!

! 54!

! 55!

! 56!

! 57!

! 58!

! 59!
Chapter 3
Expression of STX2 in the Seminiferous
Epithelium

! 60!
Introduction
The STX2 is one of SNARE (soluble N-ethylmaleimide-sensitive factor attachment
protein receptor) members, and many of SNARE family are involved in membrane
trafficking pathways during cytokinesis in a variety of organisms (107). In somatic
cytokinesis, since STX2 is essential for abscission of the midbody in HeLa cells (28),
it was assessed whether the protein localizes to the intercellular bridges of
spermatogenic cells. Further, STX2 is exhibited to localize in acrosomes of sperm to
induce membrane fusion during acrosomal reaction, resulting in release of acrosomal
components during fertilization (29, 30). Precise function of STX2 in the
spermatogenic cells remains to be addressed, but it is apparent that STX2 is one of
key factors for spermatogenesis.
Two Stx2 mutants shoswed an importnace of Stx2 in spermatogenesis (16, 32),
and previous chapter suggested that STX2 is required for maintenance of intercellular
bridges between germ cells. However, precise function of STX2 remains largely
unknown. To reveal the precise function of STX2 in spermatogenesis, this study
examined the expression and subcellular localization of secretory SNARE protein
STX2, and found that STX2 is localized to the Golgi apparatus in developing
spermatocytes and to the intercellular bridges between developing spermatogenic
cells. The SNARE hypothesis postulated that SNARE proteins are categorized into
two broad categories, v-SNAREs in transport vesicles and t-SNAREs in target
membranes, both of which pair specifically for membrane fusion during exocytosis
(108). STX2, t-SNARE protein, is involved in secretory granule fusion in pancreatic
cells (109), platelets (110), and alveolar cells (111), and is localized to the apical
plasma membrane in pancreatic acinar cells (112), and actively cycles between the
plasma membrane and endosomes under cultured cell (113). These data suggest that

! 61!
STX2 is involved in signaling transmission within a cell, but it has not been
elucidated which molecule is the target for the STX2.

! 62!
Material and methods
Animals
For biological analyses, Stx2repro34 homozygotes and control wild-type littermates at
10-12th weeks of age or at 1-3rd weeks of age for the sake of first-wave of
spermatogenesis were used. Rats of a Jcl:Wister closed colony rat at 9-10th weeks of
age were purchased from CLEA Japan (Tokyo, Japan). To obtain protein lysate for
western blotting analysis, immature Jcl:ICR closed colony mice (CLEA, Tokyo,
Japan) were used.
Western blotting
The testes removed from Stx2repro34 homozygous and normal mice were homogenized
in RIPA buffer (150 mM NaCl, 10 mM Tris, pH7.2, 0.1% SDS, 1.0% Triton X-100,
1% Deoxycholate, and 5 mM EDTA). The protein concentration of cell lysate was
measured using the bicinchoninic acid (BCA) protein assay kit (B9643, Sigma,
Missouri, USA), and 50 µg of total protein in each sample were electrophored on 10%
SDS-polyacrylamide gels and transferred onto Immobilon-P Transfer Membranes
(PVDF-Plus) (IPVH15150, Millipore, MA, USA). Immunoblots were incubated with
anti-Epimorphin antibody (BAF2568, R&D Systems, Minneapolis, USA) and anti-
βactin antibody (ab8227, abcam, Cambridge, UK) and subsequently with a
horseradish peroxidase-conjugated anti-goat or anti-rabbit IgG secondary antibodies
(1:20000, Santa Cruz Biotechnology). The membrane was developed using ECL
Advance Western Blotting Detection Kit (RPN2135, Amersham Biosciences,
Buckinghamshire, UK), and observed.

! 63!
Immunohistochemistry
Testes from mice and rat were fixed in Bouin's solution. They were dehydrated and
embedded in paraffin. Sections (5 µm) were deparaffinized and autoclaved in 0.1 M
Sodium Citrate, pH 6.0, at 120°C for 20 minutes, or the sections were treated with 20
µg/ml ProteinaseK in 10 mM Tris-HCl (pH 7.4) at 37°C for 20 minutes for antigen
retrieval. All the sections were incubated in 3% H2O2 in PBS for 15 minutes to
quench the endogenous peroxidase. They were then incubated in PBS containing 10%
BSA for 30 minutes at RT to reduce background. Sections were reacted with primary
antibody against STX2 (114) diluted at 1:100 in PBS containing 1% BSA for 3 hours
at 37°C. Subsequently HRP-labeled secondary antibodies (Santa Cruz, CA, USA),
were reacted at 1:400 dilution for 1 hour at RT. Antigens were visualized using TSA
Plus DNP (HRP) System (Perkin Elmer, MA, USA) with NEL 747B Dako Liquid
DAB+ Substrate Chromogen System (20 µl/ml) (K3467, DAKO). Sections were
counterstained with hematoxylin.
Co-localizations of STX2 and TEX14 were examined using paraffin sections
of the testis fixed in Bouin’s solution. Following the incubation with the first
antibodies (Table 2-1), sections were incubated with second antibodies, donkey anti-
goat IgG-DyLight 488 (705-485-003, Jackson Immuno Research) and donkey anti-
rabbit IgG-Cy3 (711-165-152, Jackson Immuno Reseach). The nuclei were
counterstained with DAPI then mounted using VECTASHIELDMounting Medium
(H-1200, Vector Laboratories Inc.), and observed as described above.
Squash preparation and immunocytochemistry
Squash preparation of germ cells was prepared based on the previously described
method with some modifications (115). Briefly, mouse testis was removed and

! 64!
decapsulated in PBS. Seminiferous tubules were segmented into 1-2 mm length, and
both side ends were cut to identify the stages of spermatogenesis by staining with
hematoxylin. The middle part of the tubular segments was applied for
immunocytochemistry, when the both side were identified at the same stage. The
segments at approximately 0.5 mm length were placed in 15 µl of 100 mM sucrose on
a glass slide. The cells were flown out by squashing with a coverslip. The slide was
quickly frozen in liquid nitrogen for 20 seconds, and the coverslip was removed using
a scalpel. The preparation slides were fixed with 4% PFA in PBS for 15 minutes at
RT, followed by treating with 0.1% Triton X-100 in PBS for 10 minutes at RT. After
washing twice in PBS and blocking with 5% skim milk in PBS, the preparation slides
were incubated with primary antibodies in 5% skim milk in PBS to examine co-
localizations of STX2 and TEX14. The second antibodies used were donkey anti-
rabbit IgG-Cy3 (711-165-152, Jackson Immuno Research) and donkey anti-goat IgG-
DyLight488 (711-485-033, Jackson Immuno Research. The nuclei were
counterstained with DAPI then mounted using VECTASHIELDMounting Medium
(H-1200, Vector Laboratories Inc.), and observed using BIOREVO BZ-9000
(KEYENCE).

! 65!
Results
Expression and subcellular localizations of t-SNARE STX2: the Golgi apparatus
of spermatocytes and the intercellular bridges
This study examined the localization of STX2 in the mouse and rat testis. In the rat
testis, the cytoplasmic localization of STX2 was observed in the pachytene
spermatocyte at restricted stage of IX-X. On the other hand, localizations in the Golgi
apparatus in the pachytene and diplotene spermatocyte were seen at the stage VIII-
XIII. STX2 localizations were also detected in the pro-acrosome and acrosome
regions of the round and elongating spermatids in all the stages (Fig. 3-1A). Dividing
spermatocytes did not express STX2 in the rat testis section (Fig. 3-1B). The mouse
testis showed the results quite similar to those observed in the rat testis, but the signal
was not as clear as in the rat (Fig. 3-2C,D). It was revealed that STX2 was localized
in the Golgi apparatus, which confirmed with co-localization of Golgi maker, TRA54
in the immunofluorescence of the mouse testis (Fig. 3-2A,B). The Leydig cell and the
Sertoli cells did not firmly express STX2 in both the rat and mouse testes.
Since STX2 is shown to be an element of the midbody in the cytoplasmic
bridge (28), this study assessed whether the protein was localized in the intercellular
bridge of the germ cells. In the mouse testis, STX2 was present in the intercellular
bridges between both spermatocytes and spermatids (Fig. 3-3, 3-4). Localization of
TEX14, a marker protein of the bridge, was completely merged with the expression of
STX2 (Fig. 3-3A). The intercellular bridge increases its diameter as the germ cells
differentiate; 1-1.3 µm in spermatogonia, 1.4-1.7 µm spermatocytes, and 1.8 µm< in
spermatids (36). This study observed the tendency of an increase in the diameter of
the midbody ring. This study did not observe any consistent or definitively positive
reaction for STX2 in the testicular interstitial tissue. In immunostained germ cells

! 66!
from stage II-IV seminiferous tubule segments, STX2 was localized to bridges
between intermediate spermatogonia and between early pachytene-stage
spermatocytes (Fig. 3-3B). However, signal intensity for STX2 was not as strong in
these cells as it was in late pachytene spermatocytes and spermatids. To optimize
visualization of STX2 in bridges between spermatogonia and spermatocytes at early
stages of the meiotic prophase, this study utilized squash preparations of germ cells.
Examination of germ cells released from stage IX-X tubule segments revealed that
STX2 was localized to the perinuclear region (Fig. 3-4G). In mature spermatozoa,
STX2 is localized to the acrosomal region (30). In round and elongating spermatids,
STX2 was also localized to the pro-acrosome and acrosome regions, respectively
(Fig. 3-4H,I).
Western blotting confirmed that the expression of STX2 (37kD) throughout
the first-wave of spermatogenesis and was increased at P16 (Fig. 3-5). The increased
expression was consistent with the expression of Stx2 mRNA in the previous report
(61). There was no STX2 expression in the testes of adult Stx2repro34 mutants (Fig. 3-
5).

! 67!
Discussion
In reproductive cells, STX2 is known to co-localize with acrin 1, a 90kD acrosomal
protein, and functions in acrosomal reaction during fertilization (30). In somatic cell
cytokinetic division, STX2 and endobrevin colocalize laterally to the midbody matrix,
which is essential for abscission of the midbody in Hela cells. RNAi inhibition of
these proteins resulted in binucleated cells without any defects in nuclear envelope
and nuclear divisions (28). STX2 is target-SNARE, localized to the plasma
membrane, for vesicle-associated v-SNARE, such as endobrevin, localized to
signaling vesicle in somatic cells (28). This study revealed that STX2 is localized to
the Golgi complex of spermatocytes during later prophase. Hence, it was
hypothesized that molecules produced in the Golgi complex are transported to the
place needed via vesicular mechanism (52, 58) by STX2 as an important role for a
signaling receptor. Like mammalian Stx2repro34 mutants, previous reports also showed
that a Golgi-localized SNARE Syntaxin5 (Stx5) is required for ER-Golgi traffic in
yeast and Drosophila, and the loss of Stx5 lead to blockade of the ER-Golgi traffic for
secretory proteins, and failure of germ cells to complete cytokinesis which resulted in
the formation of multinucleated germ cells (116). The loss of genes expressing other
Golgi proteins that are required for vesicle trafficking also leads to multinucleation of
germ cells in Drosophila (117).
In the wild-type testis, STX2 was distinctively detected in the Golgi apparatus
of the spermatocyte and acrosomal vesicle/acrosome in the spermatid. Testicular
localization of STX2 was previously reported by Wang, et al. (32) and Roqueta-
Rivera, et al. (118), however, the results were not in accordance each other. Wang, et
al. (32) described Leydig cell-specific localization, whereas, Roqueta-Rivera, et al.
(118) reported acrosome specific localization in the spermatid and perinuclear

! 68!
localizations in the spermatocytes, speculating that the presence of STX2 in the
endoplasmic reticulum of spermatocytes. This study observed the localization of
STX2 commonly in mouse and rat testes using an antibody against the cytoplasmic
domains of rat STX2 (114). Although this study could not explain the discrepancy
among the studies, the presence of STX2 in the Golgi apparatus and in the acrosome
of the spermatid seems certain because the results are in accordance with the studies
that revealed localizations and functions of STX2 in acrosomes of the spermatozoa to
function in the membrane fusion in fertilization (29, 30).
This study also revealed localization of STX2 to the intercellular bridges of
the germ cells. STX2 is localized to the intercellular bridges of germ cells throughout
the spermatogenic process, while Golgi-localized STX2 is only found in the
spermatocytes during later prophase. These two distinct localizations of STX2
indicate Stx2 gene plays different roles in the different reproductive stages (Fig. 3-6).
A large part of molecular mechanism of the bridges in the germ cells share with that
of somatic cytokinesis (5). In somatic cell division, failed cytokinesis triggers
multinucleation. Binucleations were also observed when STX2 and endobrevin, a
ubiquitously expressed v-SNARE protein, were functionally impaired (28), indicating
that STX2 takes part in midbody abscission in somatic cell cytokinesis. Since almost
nothing is understood about the mechanism of enlargement of the cleavage furrow
following the failed abscission, it is also unknown whether the common mechanism
lies in the formation of multinucleated cells in the somatic and germ cell STX2
deficiency. The midbody in the intercellular bridge of the germ cells is kept by
TEX14, a germ cell intercellular bridge specific protein (47). The TEX14 deficient
testis exhibited lack of the intercellular bridge and frequent apoptosis of the germ
cells (37). Thus, these phenotypic differences indicate that direct molecular

! 69!
interaction between TEX14 and STX2 is less probable. The observation that TEX14
was abnormally labelled to the large region of the membrane of the multinucleated
cell in the Stx2repro34 mutant possibly reflects the structure of the midbody remained,
although its completeness could be lost. Therefore, STX2 rather seems to be required
for maintenance of the integrity of the intercellular bridges in germ cell, and STX2 is
not likely to function to promote midbody abscissions in the intercellular bridge of
germ cells, different from the role of STX2 in the somatic cell cytokinesis. STX2
seems to have opposite features in somatic cells and germcells. This might be because
STX2 has a different role from the one in somatic cell or because STX2 has different
target moelcule(s) as a member of SNAREs mechanism.

! 70!

! 71!

! 72!

! 73!

! 74!

! 75!

! 76!

! 77!
Chapter 4
Aberrant Localization of
Sulfoglycolipids Lead to
Multinucleation of the Germ Cells

! 78!
Introduction
Both Stx2 mutants (repro34 homozygous and knockout mice) exhibited arrested
spermatogenesis during meiotic divisions with the formation of multinucleated
spermatocytes, suggesting an essential role of the Stx2 during meiosis (61, 62). With
respect to the appearance of multinucleated cells in the affected spermatogenesis,
seminolipid deficient mice were also reported to show a similar phenotype to those
observed in the Stx2 mutants (multinucleation) (50, 53), which led us to speculate that
STX2-deficient mice might be impaired in sulfoglycolipid localization. Seminolipid is
sulfated glycolipid and is predominantly distributed in the testis. Seminolipid consists
of 90% of sulfoglycolipids in the testis (54), and their biosynthesis is responded by
ceramide galactosyltransferase (CGT) and cerebroside sulfotransferase (CST). The
loss of CGT (50) and CST (48) resulted in complete deficiency of seminolipid in the
testis and disruption of spermatogenesis. Moreover, Zhang, et al. (53) evidenced that
aberrant spermatogenesis in the CST deficiency was caused by impaired seminolipid
in germ cell components and that impaired seminolipid in the somatic components
could sustain spermatogenesis.
Multinucleation of the germ cells was common feature of the two kinds of
seminolipid deficiency (48, 50, 53). Multinucleated germ cells often appear in the
pathological testicular condition possibly due to opening of the intercellular bridges
(3), and this abnormal multinucleation is generally caused by two mechanisms; cell-
cell fusion or acytokinetic cell division (68). Although the function of the intercellular
bridges in the germ cells is distinct from that of somatic cell cytokinesis, the
molecular mechanism of formation of the intercellular bridges shares the large part of
that of somatic cytokinesis (5). Recent studies suggested that lipids play an important
role in regulation of cytokinesis as a component of lipid-order microdomains, termed

! 79!
lipid rafts (57). Seminolipid in sperm is a component of lipid rafts and functions for
sperm to attach Zona pellucidae (ZP) to penetrate during capacitation (60). These
biological and biochemical information directed to speculate that a common
molecular mechanism lies in the multinucleation of the germ cells in the STX2 and
seminolipid deficiencies, especially in breakage or instability of the intercellular
bridges.
In Chapter 2 and Chapter 3, it was suggested that t-SNARE STX2 is required
for maintenance of the intercellular bridges during spermatogenesis. To reveal the
regulation of testicular sulfoglycolipids through STX2 and the function of
sulfoglycolipids in spermatogenic process, this chapter shows testicular and
cytological localization of sulfoglycolipids in the germ cells of wild-type and the
Stx2repro34 mutant.

! 80!
Material and methods
Immunohistochemistry of sulfoglycolipids
Immunostaining of sulfoglycolipids was undertaken following the previously
described method (119). Briefly, testes samples were fixed in 4% PFA after
immobilization. Following substitution with sucrose in HBSS (137 mM NaCl, 5 mM
KCl, 0.44 mM KH2PO4, 5.6 mM Glucose, 0.34 mM Na2HPO4 anhydrous, 0.81 mM
MgSO4, 1.26 mM CaCl2, and 4.17 mM NaHCO3) with an increase gradient from 12
to 18%, tissue samples were embedded in OCT compound, and frozen. Frozen
sections (7 µm) were incubated with DI8 antibody (119) in PBS(-) solution containing
5% skim milk 4°C overnight. Following the first antibody incubation, the sections
were incubated with a secondary antibody, goat anti-mouse IgG-Alexa Flour 488
(A-21121, Invitrogen, CA, USA) at 1:200 dilution with the same buffer as the
primary antibody for 1 hour at RT. The nuclei were counterstained with DAPI, then
mounted using VECTASHIELDMounting Medium (H-1200, Vector Laboratories
Inc., Burlingame, CA, USA), and observed using KEYENCE BIOREVO BZ-9000
(KEYENCE, Osaka, Japan).
Squash preparation and immunocytochemistry
Squash preparation of germ cells was prepared and immunostained using anti-TEX14
and DI8 antibodies (Table 2-1) in the same manner as chapter 3. The secondary
antibodies for TEX14 and DI8 antibodies were goat anti-mouse IgG-Alexa Flour
488 (A-11001, Invitrogen) and donkey anti-goat IgG-Alexa Flour 568 (A-11057,
Invitrogen), respectively.

! 81!
Results
Affected intracellular localization of sulfoglycolipids in the Stx2repro34 mutant
germ cells
Considering the morphological similarity of affected spermatogenesis in the
sulfoglycolipids deficiency, it was speculated the relevance of these molecules with
STX2 in regulations of spermatogenesis. In order to address to this idea, this study
examined localization of sulfoglycolipids in the wild-type and the Stx2repro34 mutant
testes using the DI8 mouse monoclonal antibody against sulfoglycolipids, including
both sulfatide and seminolipids, that contain the terminal 3-O-sulfated galactose
structure (119).
In the wild-type testis, spermatogonia exhibited very little immunoreactivity
(Fig. 4-1A), while spermatocytes at preleptotene, leptotene/zygotene and early
pachytene stages exhibited a strong and characteristic immunoreaction of fine
granular distribution at peripheral plasma membrane (Fig. 4-1A-C). In late pachytene
and diplotene spermatocytes and spermatids, labeling in plasma membrane was quite
weak and entirely cytoplasmic (Fig. 4-1C). In Stx2repro34 mutant testes, spermatogenic
cells did not exhibit robust localization of sulfoglycolipids in peripheral plasma
membrane (Fig. 4-1D-F). In addition, an abnormal and patchy intercellular
localization was frequently observed at the periphery of mutant pachytene
spermatocytes or cytoplasm of Sertoli cells in stages I-IV of the seminiferous
epithelium (Fig. 4-1D). Cytoplasmic accumulation of sulfoglycolipids in mutant
spermatocytes at late pachytene and diplotene stages (Fig. 4-1E,F) was significantly
greater than that of that of wild-type (Fig. 4-1C).

! 82!
Localization of sulfoglycolipids to the intercellular bridges
Immunostaining of the wild-type germ cells obtained by squash preparation
also evidenced that sulfoglycolipids were localized to the plasma membrane of
spermatocytes (Fig. 4-2B) and spermatids. In addition, spermatocytes and spermatids
at any stages, but not spermatogonia, also exhibited ring-shaped sulfoglycolipid
localization, which was perfectly co-localized with TEX14 (Fig. 4-2A,B). This
reactivity was weak in preleptotene to early pachytene stages and stronger in
developing germ cell at later and post-pachytene stages. This confirms that
sulfoglycolipids are localized to intercellular bridges in specific developmental stages
of spermatogenesis (Fig. 4-3). Together, these observations suggest aberrant
localization and dispersal of sulfoglycolipids in mutant germ cells. Interestingly,
sulfoglycolipids were observed in the intercellular bridges of Stx2repro34 mutant germ
cells (Fig. 4-2C,D). The diameter of the intercellular bridges between mutant
spermatocytes was greater (Fig. 4-2C) than that of wild type (Fig. 4-2A,B), an
observation consistent with opening of the intercellular bridges in Stx2repro34 mutant
spermatocytes.

! 83!
Discussion
In this chapter, immunohistochemical staining of the testis of the Stx2repro34 mutant
revealed that sulfoglycolipids are aberrantly localized in the seminiferous epithelium.
Although this study did not perform experiments to show normal catalytic activity to
produce sulfoglycolipids, it is speculated that sulfoglycolipids are normally produced
but not properly localized in the Stx2repro34 mutant germ cells. In the wild-type testis,
STX2 was co-localized with the Golgi marker protein (Chapter 3), and Roqueta-
Rivera, et al. (118) reported perinuclear localizations of STX2 in the spermatocytes.
These observations suggested that STX2 is localized in the endoplasmic reticulum of
spermatocytes, most likely Golgi apparatus. Sulfoglycolipids are produced at the
outer membrane of the Golgi apparatus in pachytene spermatocytes and are present in
the plasma membrane of germ cells (52, 54, 58). This study also showed the
localization of sulfoglycolipids to the plasma membrane in the wild-type
spermatocyte. In contrast, in the Stx2repro34 mutant spermatocyte, sulfoglycolipids
were not markedly detected in the plasma membrane, and were abnormally
accumulated in the cytoplasm of later pachynema and diplonema. This may imply that
sulfoglycolipids were not guided to the plasma membrane or the intercellular bridges
and they accumulated in the cytoplasm. As a result, distinct expression of
sulfoglycolipids was not observed on the plasma membrane of spermatocyte during
later prophase in the normal testis (Fig. 4-4). Therefore, the results could implicate
that STX2 is required for transportation or distribution of sulfoglycolipids from the
Golgi complex to the site needed, especially to the cellular membrane in stage-
specific manner.
It was suggested that seminolipid is a component of lipid rafts in the germ
cells along with other lipid families (53). Lipid rafts are membrane domains which

! 84!
could function in membrane trafficking and/or remodeling. Considering the
accordance of subcellular localization of STX2 and sulfoglycolipids in the pachytene
spermatocyte, the same phenotype (multinucleation) in impairment of each molecule
might suggest that these molecules cooperate to prevent abscission of the midbody in
the intercellular bridge. Lipid rafts hypothesis postulates that sphingolipids and sterols
assemble to form a raft platform which contributes to membrane signaling and
trafficking (59). Since several studies suggested that plasma membrane raft domains
are formed in the furrows of dividing cells, indicating that lipid raft domains play an
important role in cytokinesis (120). The observation of this study suggested that the
loss of STX2 caused formation of multinucleated cells in Stx2repro34 mutant males. In
addition, this study showed the cytological localization of sulfoglycolipids to the
intercellular bridges. Hence, since the molecular mechanism of formation of
intercellular bridges in germ cells shares a large part of that in mitotic cytokinesis (5),
perhaps sulfoglycolipids play an important role in maintenance of intercellular
bridges in germ cells.
Interestingly, common morphological characteristics, abnormal fusion of
spermatogenic cells, in the testes of Stx2repro34 mutant males, CST (50) and CGT KOs
(53), was observed only at the late pachytene stage. This suggests that both
sulfoglycolipids and STX2 are required for maintenance of intercellular bridges. In
addition, MX, alpha-mannosidase IIx, is an enzyme to form intermediate asparagine-
linked carbohydrates (N-glycans), which plays an importnat role in germ cel-Sertoli
cell adhesion, and MX deficient mice also showed some multinucleated germ cells in
the seminiferous epithelium of the testis (121). It is, therefore, hypothesized that the
abnormal fusion of spermatocytes in Stx2repro34 and glycolipid-deficient mutants was
due to defective formation of intercellular bridges, since mammalian germ cells are

! 85!
connected as syncytium through intercellular bridges that exist between the
spermatogenic cells at the same developing stage (33, 34), and stable formation of the
intercellular bridges is essential for spermatogenesis and fertility (37). Disruption of
the intercellular bridges prevents normal formation of syncytium, which arrests
spermatogenesis prior to pachytene, and resulted in male-restricted sterility (37).
Moreover, glycolipid-deficient mutants also showed multinucleated spermatocytes
and male-restricted sterility, which resembles the phenotpe of repro34 (50, 53). In
addition, genes essential for cytokinesis and formation of male ring canals are not
necessarily required for female ring canals or fertility (37, 44).
During spermatogenesis, germ cells experience dynamic morphological
changes; late pachytene spermatocytes are larger than those in the previous stages (3),
while the diameter of intercellular bridges of spermatocytes remains the same (122). It
was postulated that both sulfoglycolipids and STX2 regulate stage-specific membrane
dynamics, and the observations on aberrant intracellular localization of
sulfoglycolipids in Stx2repro34 mutant testes are consistent with this hypothesis.
With respect to these phenotypes of several mutants, STX2 and
sulfoglycolipids appear to be required for maintenance of the intercellular bridges of
the germ cells, and the loss of either STX2 or sulfoglycolipids induced defect of the
intercellular bridges. Present data might indicate accurate localization of
sulfoglycolipids could be a key factor for spermatogenesis and STX2 could be
involved in the mechanisms that convey and settle them into the bridges between
pachynema.
The distinct STX2 localization to the Golgi apparatus we observed also could
implicate STX2 as interacting with target sulfoglycolipids, whose biosynthesis in the
Golgi membranes also involves CGT catalysis (54). Sulfoglycolipids form the

! 86!
platform of lipid raft domains that contribute to membrane signaling and trafficking
(59) and function in capacitation of the mouse and pig spermatozoa (60, 123). Mice
lacking complex gangliosides, other candidate components of lipid rafts, due to
disruption of ganglioside GM2/GD2 synthase also exhibited multinucleation of the
spermatids (124, 125) as do STX2-deficient mice. Taken together, these studies lead
to the idea that sulfoglycolipids may contribute to raft domains within germ-cell
plasma membrane and intercellular bridges. Further studies should identify molecules
interacting with STX2 and mediating transport of sulfoglycolipids and analyze
membrane domain function in maintaining the constricted intercellular bridges that
are a key characteristic of spermatogenesis.

! 87!

! 88!

! 89!

! 90!

! 91!
Chapter 5
General discussion and conclusion

! 92!
Conclusion
Expression and Subcellular localization of t-SNARE STX2 in the Golgi
Apparatus and Intercellular Bridges
This study evidenced the expression and subcellular localization of secretory SNARE
protein STX2 in both the Golgi apparatus and the intercellular bridges of
spermatocytes at meiotic prophase and of spermatids. Furthermore, this study
revealed that co-localization of sulfoglycolipids and STX2 to the intercellular bridges.
t-SNARE protein STX2 is one of two SNARE protein categories, and along with
reciprocal v-SNAREs in transport vesicles, STX2 function specifically for membrane
fusion during exocytosis (108). Although several studies suggested the functions of
STX2 in diverse signaling transmission in the cell, none of the target molecules in the
events have not been previously identified. Together with the facts of co-localization
of TEX14 and sulfoglycolipids to the intercellular bridges of the germ cells (Fig. 4-2),
disrupted distribution of glycolipids in the seminiferous epithelium of Stx2repro34 mice
(Fig. 4-1) and the resultant multinucleation of the germ cells (Fig. 2-1),
sulfoglycolipids are target molecular candidates for t-SNARE STX2. This
transportation system possibly functions in the maintenance of the intercellular bridge
in spermatogenesis. In the wild-type testis, STX2 was specifically detected in the
Golgi apparatus of the spermatocyte and acrosomal vesicle/acrosome in the
spermatid. This study observed the localization of STX2 commonly in mouse and rat
testis using an antibody against the cytoplasmic domains of rat STX2 (Fig. 1-5) (114).
Further, two distinct localizations of STX2 to the intercellular bridge and to the Golgi
apparatus indicate that the gene plays different roles in the different reproductive
stages.

! 93!
Abberant Localization of Sulfoglycolipids in Stx2repro34 Mutant Germ Cells
Immunochemical observations of sulfoglycolipids present in the cytoplasm
and dotted in the plasma membrane of the wild-type spermatocyte (Fig. 4-1, 4-2)
showed good coinsistency with the previous reports (52, 54, 58). Moreover, this study
revealed that sulfoglycolipids specifically localized to the intercellular bridge of these
cells in the squash preparation. In mammals, two major sulfoglycolipids, sulfatide and
galactosylsulfatide, are produced in the testis through catalyzation of
alkylacylglycerol and ceramide with cerebroside sulfotransferase (CST) (48, 49) and
ceramide galactosyltransferase (CGT) (50) one after another, consequently
alkylacylglycerol and ceramide are converted to seminolipids and sulfatide,
respectively (Fig. 1-9) (48, 50, 51). Significance of sulfoglycolipids in
spermatogenesis has been demonstrated with male specific infertility in CST and
CGT deficient mice (48, 50, 53). However, the precise roles of these molecules in
spermatogenesis are still to be clarified. The Stx2repro34 mutants exhibited defective
spermatogenesis with appearance of syncytial multinucleated germ cells during
meiotic divisions (61). This phenotype is quite similar to the glycolipid-deficient
mutants described above (50, 53). Here this study showed that sulfoglycolipids were
localized to the plasma membrane of normal pachynema but accumulated in the
cytoplasm of the mutant spermatocytes at late pachytene and diplotene stages (Fig. 4-
1). These results indicate that glycolipids could be a key factor for maintenance of the
intercellular bridges.
Multinicleated Germ Cell in Stx2repro34 mutants
Meiotic processes appear to proceed normally until the multinucleation
occurs. Immunocytochemical staining of chromosome preparation confirmed normal
chromosome synapsis (Fig. 2-5) and chiasma formation in the mutant testis (Fig. 2-6).

! 94!
Histological examination showed several types of the nuclei, including those of late
pachytene, diplotene and metaphase spermatocytes in the mutant syncytial germ cells
(Fig. 2-1). The multinucleated cells with the nuclei at metaphase suggested ongoing
meiosis after the defect of the intercellular bridge. Although this study did neither
succeed to obtain siblings by ROSI using the nucleus of the aberrant cells as donors
nor confirm haploidity of the nuclei by chromosome preparations, vestiges of the
implantation of the transferred embryo were detected (Fig. 2-10). This at least
suggested that some of the nuclei in the syncytial cell had haploid genome and that
multiploidy of the donor nuclei caused the arrest of embryo development through the
spindle checkpoint.
Absence of STX2 in the Golgi Apparatus and Intercellular Bridges Affect
Integrity of Intercellular Bridges
In somatic cell division, failed cytokinesis triggers multinucleation.
Completion of cytokinesis owes a lot to the abscission of the midbody, a central part
of the bridge that consists of various molecules. Recruitment of TSG101 and ALIX to
the midbody to interact with CEP55, a component of the midbody, triggers abscission
then consequently completes the division of the daughter cells. Once the abscission of
the midbody is failed, the mitotic cell results in formation of binucleated cells, as
observed when the CEP55-ALIX/TSG101 association was disrupted (46, 126). Also,
binucleations were observed when STX2 and endobrevin, a ubiquitously expressed v-
SNARE protein, were functionally impaired (28). Almost nothing is understood about
the mechanism in which the enlargement of the cleavage furrow following the
impaired abscission lies. It is also unknown whether the common molecular system
lies in the multinucleation in the STX2 deficiency of the Stx2repro34 mutant. The
midbody in the intercellular bridge of the germ cells is kept by TEX14, a germ cell

! 95!
intercellular bridge specific protein, which interacts with CEP55 instead of TSG101
and ALIX to disturb the midbody abscission (47). The TEX14 deficient testis
exhibited lack of the intercellular bridge and frequent apoptosis in the germ cells (37),
whereas the STX2 deficient Stx2repro34 mutant testis showed TEX14 expression and
normal formation of the intercellular bridges. Thus, these phenotypic differences
indicate that direct molecular interaction between TEX14 and STX2 is less probable.
STX2 rather seems to be required for maintenance of the integrity of the intercellular
bridges. Furthermore, since the structure of the midbody appeared to be remained in
the Stx2repro34, STX2 is not likely to function to promote midbody abscissions in the
intercellular bridge of the germ cells unlike the case of the somatic cell cytokinesis.
This controversy might come from the roles of the target molecule that the SNAREs
mechanism transports.
STX2 Is Implicated in Intracellular Transportation of Sulfoglycolipids
This study evidenced localizations of STX2 in the intercellular bridge of
spermatogonia, spermatocytes and spermatids (Fig. 3-3), however influences of the
STX2 deficiency in spermatogenesis appeared in the late pachytene stage with an
aberrant intercellular localization of sulfoglycolipids. From these results and the
common abnormal morphology (multinucleation) in the sulfoglycolipids deficiency, it
seems certain that both STX2 and sulfoglycolipids function in cooperation to maintain
the intercellular bridge. Interestingly, the mice lacking complex gangliosides by the
disruption of ganglioside GM2/GD2 synthase showed multinucleation of the
spermatids (124, 125). Gangliosides and choresterols are thought to form the lipid raft
platform which contributes to membrane signaling and trafficking (59), therefore the
lipid raft possibly plays roles to maintain the intercellular bridge of the spermatids.
On the other hand, sulfoglycolipids are shown to form the platform of the raft domain

! 96!
which functions in capacitation of the mouse and pig spermatozoa (60, 127). These
studies would lead to the idea that sulfoglycolipids lubricate the raft domain in the
spermatocytes, which are prerequisite for the intercellular bridge to stay stable.
Therefore, it is favored to speculate that STX2 transports sulfoglycolipids to the raft
with the SNARE membrane fusion mechanism. Further studies on the membrane
domain function for maintaining the furrowing of the intercellular bridge and
identification of the molecules which mediate STX2 transportation of sulfoglycolipids
to the membrane of the bridge are necessary.
Possible Function of STX2 in Sertoli cell
Xu, et al. (128) hypothesized that residual sulfoglycolipids that are observed
from the cytoplasm of elongated spermatids are “recycled” within the Sertoli cell to
convert them into a precursor for de novo synthesis of sulfoglycolipids through
arylsulfatase A (ARSA). In the Sertoli cells of the ARSA deficient mouse,
sulfoglycolipids remained its native status (sulfolipid) and abnormally accumulated in
the cytoplasm of the Sertoli cells (128). In the testis of the Stx2repro34 mutant,
abnormal granules of sulfoglycolipids were frequently observed presumably in the
cytoplasm of Sertoli cells (Fig. 4-1), which is similar to the phenotype of ARSA
deficient-mutant (128). Previous study showed active cycles of STX2 between the
plasma membrane and endosomes in cultured cell (113), and the localization of STX2
in cytoplasm of the Sertoli cell in vitro (129). Taken together, this study postulated
that t-SNARE STX2 is also involved in dynamic transportation of “recycled”
sulfoglycolipids in the Sertoli cells with stage specific-manner (Fig. 5-1). Future
studies need to be conducted to prove this hypothesis.

! 97!
Functions of Glycan in Reproduction
In the reproductive system, biological features of glycans have recently
attracted increasing attention. Although no human reproductive disorder related to
abnormality of glycans was previously reported, several studies have shown the
importance of carbohydrate moiety of zona pellucida (ZP) (130, 131) and endometrial
epithelial tissues (132, 133) in fertilization and implantation, respectively. During
capacitation, seminolipid in sperm is required for sperm to attach to ZP during
capacitation (60) and carbohydrate moiety of sulfoglycolipids on sperm plays an
important role in species-specificity of fertilization (134, 135). This study proposes
the indispensable role of sulfoglycolipids in male gametogenesis, specifically in
maintenance of stable intercellular bridges of germ cells. The loss of stable formation
of intercellular bridges resulted in sterility due to spermatogenic arrest prior to
meiosis (37). However, in Stx2repro34 mutant males, in contrast, the progression of
spermatogenesis and the formation of intercellular bridges were apparently affected
after mid to late pachytene. These observations suggested that cellular development
requires stage-specific mechanisms that regulate the maintenance of the intercellular
bridges at certain stages of spermatogenesis, and the results of this study suggested
that sulfoglycolipids could be one of key factors.
Conclusion
As a concluding remark of this study, STX2 is important for two distinctive
functions; exocytotic transportation of sulfoglycolipids from the Golgi complex and
subcellular guide to the intercellular bridges. However, since molecular mechanisms
of STX2 and functional roles of sulfoglycolipids during spermatogenesis remain
largely unknown, further studies must be conducted.

! 98!
The goal of whole genome, ramdom ENU mutagenesis is to identify novel
genes and functions. Infertile mouse models can provide useful findings of genes
essential for reproduction. Understanding the biology of reproduction is essential for
clinical application not only for human infertility but also diverse contraceptive
approaches. Contribution of this study is that repro34 mutant mice emphasized
importnace of intercellular bridges and sulfoglycolipids in spermatogenesis, and this
study showed a possible correlation between these two unrelated factors, although
precise functions of each factor are largely unknown. Therefore, how STX2 is
involved in these needs to be answered. This study using an ENU mutant took
advantage of foreward genetic approach. Hence, repro34 was found out to be an
importnat model mouse for reproduction, and findings of this study contribute to
reveal future directions that can be exlpoited in understandng of spermatogenesis.

! 99!

! 100!
Acknowledgement
I am grateful to Dr. Tetsuo Kunieda for his support of this study throughout
my academic study in Okayama University. I thank Dr. Junko Noguchi at National
Institute of Agrobiological Sciences, Drs. Atsuo Ogura, Kimiko Inoue and Ms.
Narumi Ogonuki at RIEKN Bioresource center, Dr. Mary Ann Handel at The Jackson
Laboratory for their experimental support and useful comments on my research. I
thank Drs. Takehito Tsuji, Takuro Oikawa, Katsunori Satoh, Hiroaki Funahashi and
members of the Animal Genetics Laboratory in Okayama University for their
comments and discussion on this study. I also thank Mr. Neil Allara for his
grammatical correction of early manuscript.
I would like to express my gratitude to my family and relatives for their
understanding and support.
Lastly, I am grateful to my wife Chizuru and my son Yuta for wholehearted
support.

! 101!
References
1. Hess RA & Renato de Franca L (2008) Spermatogenesis and cycle of the
seminiferous epithelium. Advances in Experimental Medicine and Biology
636:1-15.
2. Rolland AD, Jegou B, & Pineau C (2008) Testicular development and
spermatogenesis: harvesting the postgenomics bounty. Advances in
Experimental Medicine and Biology 636:16-41.
3. Russell LD, Rttlin RA, SinhaHikim AP, & Clegg ED (1990) Histological and
Histopathological Evaluation of the Testis (Cache River Press, USA) 1st Ed.
4. Oakberg EF (1956) Duration of spermatogenesis in the mouse and timing of
stages of the cycle of the seminiferous epithelium. American Journal of
Anatomy 99(3):507-516.
5. Greenbaum MP, Iwamori T, Buchold GM, & Matzuk MM (2011) Germ cell
intercellular bridges. Cold Spring Harbor Perspectives in Biology
3(8):a005850.
6. Zickler D & Kleckner N (1999) Meiotic chromosomes: integrating structure
and function. Annual Review of Genetics 33:603-754.
7. Eijpe M, Offenberg H, Jessberger R, Revenkova E, & Heyting C (2003)
Meiotic cohesin REC8 marks the axial elements of rat synaptonemal
complexes before cohesins SMC1beta and SMC3. J Cell Biol 160(5):657-670.
8. McDougall A, Elliott DJ, & Hunter N (2005) Pairing, connecting, exchanging,
pausing and pulling chromosomes. EMBO Rep 6(2):120-125.
9. Chicheportiche A, Bernardino-Sgherri J, de Massy B, & Dutrillaux B (2007)
Characterization of Spo11-dependent and independent phospho-H2AX foci
during meiotic prophase I in the male mouse. J Cell Sci 120(Pt 10):1733-1742.

! 102!
10. Costa Y & Cooke HJ (2007) Dissecting the mammalian synaptonemal
complex using targeted mutations. Chromosome Res 15(5):579-589.
11. Holloway JK, Booth J, Edelmann W, McGowan CH, & Cohen PE (2008)
MUS81 generates a subset of MLH1-MLH3-independent crossovers in
mammalian meiosis. PLoS Genet 4(9):e1000186.
12. Lipkin SM, Moens PB, Wang V, Lenzi M, Shanmugarajah D, Gilgeous A,
Thomas J, Cheng J, Touchman JW, Green ED, Schwartzberg P, Collins FS, &
Cohen PE (2002) Meiotic arrest and aneuploidy in MLH3-deficient mice. Nat
Genet 31(4):385-390.
13. Justice MJ, Noveroske JK, Weber JS, Zheng B, & Bradley A (1999) Mouse
ENU mutagenesis. Human Molecular Genetics 8(10):1955-1963.
14. Handel MA, Lessard C, Reinholdt L, Schimenti J, & Eppig JJ (2006)
Mutagenesis as an unbiased approach to identify novel contraceptive targets.
Mol Cell Endocrinol 250(1-2):201-205.
15. O'Bryan MK & de Kretser D (2006) Mouse models for genes involved in
impaired spermatogenesis. Int J Androl 29(1):76-89; discussion 105-108.
16. Akiyama K, Akimaru S, Asano Y, Khalaj M, Kiyosu C, Masoudi AA,
Takahashi S, Katayama K, Tsuji T, Noguchi J, & Kunieda T (2008) A new
ENU-induced mutant mouse with defective spermatogenesis caused by a
nonsense mutation of the syntaxin 2/epimorphin (Stx2/Epim) gene. The
Journal of Reproduction and Development 54(2):122-128.
17. Hirai Y, Takebe K, Takashina M, Kobayashi S, & Takeichi M (1992)
Epimorphin: a mesenchymal protein essential for epithelial morphogenesis.
Cell 69(3):471-481.

! 103!
18. Bennett MK, Calakos N, & Scheller RH (1992) Syntaxin: a synaptic protein
implicated in docking of synaptic vesicles at presynaptic active zones. Science
257(5067):255-259.
19. Inoue A, Obata K, & Akagawa K (1992) Cloning and sequence analysis of
cDNA for a neuronal cell membrane antigen, HPC-1. Journal of Biological
Chemistry 267(15):10613-10619.
20. Radisky DC, Hirai Y, & Bissell MJ (2003) Delivering the message:
epimorphin and mammary epithelial morphogenesis. Trends in Cell Biology
13(8):426-434.
21. Chen CS, Nelson CM, Khauv D, Bennett S, Radisky ES, Hirai Y, Bissell MJ,
& Radisky DC (2009) Homology with vesicle fusion mediator syntaxin-1a
predicts determinants of epimorphin/syntaxin-2 function in mammary
epithelial morphogenesis. Journal of Biological Chemistry 284(11):6877-
6884.
22. Hirai Y, Nelson CM, Yamazaki K, Takebe K, Przybylo J, Madden B, &
Radisky DC (2007) Non-classical export of epimorphin and its adhesion to
alphav-integrin in regulation of epithelial morphogenesis. Journal of Cell
Science 120(Pt 12):2032-2043.
23. Hirai Y (2001) Epimorphin as a morphogen: Does a protein for intracellular
vesicular targeting act as an extracellular signaling molecule? Cell Biology
International 25(3):193-195.
24. Chen YA & Scheller RH (2001) SNARE-mediated membrane fusion. Nature
Reviews: Molecular Cell Biology 2(2):98-106.

! 104!
25. Giraudo CG, Eng WS, Melia TJ, & Rothman JE (2006) A clamping
mechanism involved in SNARE-dependent exocytosis. Science
313(5787):676-680.
26. Denzer AJ, Nabholz CE, & Spiess M (1995) Transmembrane orientation of
signal-anchor proteins is affected by the folding state but not the size of the N-
terminal domain. EMBO Journal 14(24):6311-6317.
27. Spiess M (1995) Heads or tails--what determines the orientation of proteins in
the membrane. FEBS Letters 369(1):76-79.
28. Low SH, Li X, Miura M, Kudo N, Quinones B, & Weimbs T (2003) Syntaxin
2 and endobrevin are required for the terminal step of cytokinesis in
mammalian cells. Developmental Cell 4(5):753-759.
29. Hutt DM, Baltz JM, & Ngsee JK (2005) Synaptotagmin VI and VIII and
syntaxin 2 are essential for the mouse sperm acrosome reaction. Journal of
Biological Chemistry 280(21):20197-20203.
30. Katafuchi K, Mori T, Toshimori K, & Iida H (2000) Localization of a syntaxin
isoform, syntaxin 2, to the acrosomal region of rodent spermatozoa. Molecular
Reproduction and Development 57(4):375-383.
31. Ramalho-Santos J, Moreno RD, Wessel GM, Chan EK, & Schatten G (2001)
Membrane trafficking machinery components associated with the mammalian
acrosome during spermiogenesis. Exp Cell Res 267(1):45-60.
32. Wang Y, Wang L, Iordanov H, Swietlicki EA, Zheng Q, Jiang S, Tang Y,
Levin MS, & Rubin DC (2006) Epimorphin(-/-) mice have increased intestinal
growth, decreased susceptibility to dextran sodium sulfate colitis, and
impaired spermatogenesis. Journal of Clinical Investigation 116(6):1535-
1546.

! 105!
33. Burgos MH & Fawcett DW (1955) Studies on the fine structure of the
mammalian testis. I. Differentiation of the spermatids in the cat (Felis
domestica). The Journal of Biophysical and Biochemical Cytology 1(4):287-
300.
34. Fawcett DW, Ito S, & Slautterback D (1959) The occurrence of intercellular
bridges in groups of cells exhibiting synchronous differentiation. The Journal
of Biophysical and Biochemical Cytology 5(3):453-460.
35. Huckins C & Oakberg EF (1978) Morphological and quantitative analysis of
spermatogonia in mouse testes using whole mounted seminiferous tubules. II.
The irradiated testes. The Anatomical Record 192(4):529-542.
36. Weber JE & Russell LD (1987) A study of intercellular bridges during
spermatogenesis in the rat. American Journal of Anatomy 180(1):1-24.
37. Greenbaum MP, Yan W, Wu MH, Lin YN, Agno JE, Sharma M, Braun RE,
Rajkovic A, & Matzuk MM (2006) TEX14 is essential for intercellular
bridges and fertility in male mice. Proc Natl Acad Sci U S A 103(13):4982-
4987.
38. Guo GQ & Zheng GC (2004) Hypotheses for the functions of intercellular
bridges in male germ cell development and its cellular mechanisms. J Theor
Biol 229(1):139-146.
39. Caldwell KA & Handel MA (1991) Protamine transcript sharing among
postmeiotic spermatids. Proceedings of the National Academy of Sciences of
the United States of America 88(6):2407-2411.
40. Morales CR, Lefrancois S, Chennathukuzhi V, El-Alfy M, Wu X, Yang J,
Gerton GL, & Hecht NB (2002) A TB-RBP and Ter ATPase complex
accompanies specific mRNAs from nuclei through the nuclear pores and into

! 106!
intercellular bridges in mouse male germ cells. Developmental Biology
246(2):480-494.
41. Ventela S, Toppari J, & Parvinen M (2003) Intercellular organelle traffic
through cytoplasmic bridges in early spermatids of the rat: mechanisms of
haploid gene product sharing. Mol Biol Cell 14(7):2768-2780.
42. Robinson DN & Cooley L (1996) Stable intercellular bridges in development:
the cytoskeleton lining the tunnel. Trends in Cell Biology 6(12):474-479.
43. Stanley HP, Bowman JT, Romrell LJ, Reed SC, & Wilkinson RF (1972) Fine
structure of normal spermatid differentiation in Drosophila melanogaster.
Journal of Ultrastructure Research 41(5):433-466.
44. Brill JA, Hime GR, Scharer-Schuksz M, & Fuller MT (2000) A phospholipid
kinase regulates actin organization and intercellular bridge formation during
germline cytokinesis. Development 127(17):3855-3864.
45. Burgess DR & Chang F (2005) Site selection for the cleavage furrow at
cytokinesis. Trends Cell Biol 15(3):156-162.
46. Morita E, Sandrin V, Chung HY, Morham SG, Gygi SP, Rodesch CK, &
Sundquist WI (2007) Human ESCRT and ALIX proteins interact with proteins
of the midbody and function in cytokinesis. EMBO J 26(19):4215-4227.
47. Iwamori T, Iwamori N, Ma L, Edson MA, Greenbaum MP, & Matzuk MM
(2010) TEX14 interacts with CEP55 to block cell abscission. Mol Cell Biol
30(9):2280-2292.
48. Honke K, Hirahara Y, Dupree J, Suzuki K, Popko B, Fukushima K,
Fukushima J, Nagasawa T, Yoshida N, Wada Y, & Taniguchi N (2002)
Paranodal junction formation and spermatogenesis require sulfoglycolipids.
Proc Natl Acad Sci U S A 99(7):4227-4232.

! 107!
49. Honke K, Yamane M, Ishii A, Kobayashi T, & Makita A (1996) Purification
and characterization of 3'-phosphoadenosine-5'-phosphosulfate:GalCer
sulfotransferase from human renal cancer cells. The Journal of Biochemistry
119(3):421-427.
50. Fujimoto H, Tadano-Aritomi K, Tokumasu A, Ito K, Hikita T, Suzuki K, &
Ishizuka I (2000) Requirement of seminolipid in spermatogenesis revealed by
UDP-galactose: Ceramide galactosyltransferase-deficient mice. J Biol Chem
275(30):22623-22626.
51. Honke K, Tsuda M, Hirahara Y, Ishii A, Makita A, & Wada Y (1997)
Molecular cloning and expression of cDNA encoding human 3'-
phosphoadenylylsulfate:galactosylceramide 3'-sulfotransferase. Journal of
Biological Chemistry 272(8):4864-4868.
52. Vos JP, Lopes-Cardozo M, & Gadella BM (1994) Metabolic and functional
aspects of sulfogalactolipids. Biochimica et Biophysica Acta: Protein
Structure and Molecular Enzymology 1211(2):125-149.
53. Zhang Y, Hayashi Y, Cheng X, Watanabe T, Wang X, Taniguchi N, & Honke
K (2005) Testis-specific sulfoglycolipid, seminolipid, is essential for germ cell
function in spermatogenesis. Glycobiology 15(6):649-654.
54. Ishizuka I (1997) Chemistry and functional distribution of sulfoglycolipids.
Progress in Lipid Research 36(4):245-319.
55. Bosio A, Binczek E, & Stoffel W (1996) Functional breakdown of the lipid
bilayer of the myelin membrane in central and peripheral nervous system by
disrupted galactocerebroside synthesis. Proceedings of the National Academy
of Sciences of the United States of America 93(23):13280-13285.

! 108!
56. Coetzee T, Fujita N, Dupree J, Shi R, Blight A, Suzuki K, Suzuki K, & Popko
B (1996) Myelination in the absence of galactocerebroside and sulfatide:
normal structure with abnormal function and regional instability. Cell
86(2):209-219.
57. Neto H, Collins LL, & Gould GW (2011) Vesicle trafficking and membrane
remodelling in cytokinesis. Biochem J 437(1):13-24.
58. Koul O, Chou KH, & Jungalwala FB (1980) UDP-galactose-ceramide
galactosyltransferase in rat brain myelin subfractions during development.
Biochemical Journal 186(3):959-969.
59. Lingwood D & Simons K (2010) Lipid rafts as a membrane-organizing
principle. Science 327(5961):46-50.
60. Bou Khalil M, Chakrabandhu K, Xu H, Weerachatyanukul W, Buhr M,
Berger T, Carmona E, Vuong N, Kumarathasan P, Wong PT, Carrier D, &
Tanphaichitr N (2006) Sperm capacitation induces an increase in lipid rafts
having zona pellucida binding ability and containing
sulfogalactosylglycerolipid. Dev Biol 290(1):220-235.
61. Akiyama K, Akimaru S, Asano Y, Khalaj M, Kiyosu C, Masoudi AA,
Takahashi S, Katayama K, Tsuji T, Noguchi J, & Kunieda T (2008) A new
ENU-induced mutant mouse with defective spermatogenesis caused by a
nonsense mutation of the syntaxin 2/epimorphin (Stx2/Epim) gene. The
Journal of Reproduction and Development 54(2):122-128.
62. Wang Y, Wang L, Iordanov H, Swietlicki EA, Zheng Q, Jiang S, Tang Y,
Levin MS, & Rubin DC (2006) Epimorphin(-/-) mice have increased intestinal
growth, decreased susceptibility to dextran sodium sulfate colitis, and

! 109!
impaired spermatogenesis. Journal of Clinical Investigation 116(6):1535-
1546.
63. Bellve AR, Cavicchia JC, Millette CF, O'Brien DA, Bhatnagar YM, & Dym
M (1977) Spermatogenic cells of the prepuberal mouse. Isolation and
morphological characterization. Journal of Cell Biology 74(1):68-85.
64. Mu X, Lee YF, Liu NC, Chen YT, Kim E, Shyr CR, & Chang C (2004)
Targeted inactivation of testicular nuclear orphan receptor 4 delays and
disrupts late meiotic prophase and subsequent meiotic divisions of
spermatogenesis. Mol Cell Biol 24(13):5887-5899.
65. Hamer G, Roepers-Gajadien HL, van Duyn-Goedhart A, Gademan IS, Kal
HB, van Buul PP, & de Rooij DG (2003) DNA double-strand breaks and
gamma-H2AX signaling in the testis. Biology of Reproduction 68(2):628-634.
66. Rogakou EP, Boon C, Redon C, & Bonner WM (1999) Megabase chromatin
domains involved in DNA double-strand breaks in vivo. Journal of Cell
Biology 146(5):905-916.
67. Mahadevaiah SK, Turner JM, Baudat F, Rogakou EP, de Boer P, Blanco-
Rodriguez J, Jasin M, Keeney S, Bonner WM, & Burgoyne PS (2001)
Recombinational DNA double-strand breaks in mice precede synapsis. Nature
Genetics 27(3):271-276.
68. Chambers TJ (1978) Multinucleate giant cells. Journal of Pathology
126(3):125-148.
69. de Vries FA, de Boer E, van den Bosch M, Baarends WM, Ooms M, Yuan L,
Liu JG, van Zeeland AA, Heyting C, & Pastink A (2005) Mouse Sycp1
functions in synaptonemal complex assembly, meiotic recombination, and XY
body formation. Genes & Development 19(11):1376-1389.

! 110!
70. Yuan L, Liu JG, Zhao J, Brundell E, Daneholt B, & Hoog C (2000) The
murine SCP3 gene is required for synaptonemal complex assembly,
chromosome synapsis, and male fertility. Molecular Cell 5(1):73-83.
71. Henderson KA & Keeney S (2004) Tying synaptonemal complex initiation to
the formation and programmed repair of DNA double-strand breaks.
Proceedings of the National Academy of Sciences of the United States of
America 101(13):4519-4524.
72. Tesse S, Storlazzi A, Kleckner N, Gargano S, & Zickler D (2003) Localization
and roles of Ski8p protein in Sordaria meiosis and delineation of three
mechanistically distinct steps of meiotic homolog juxtaposition. Proceedings
of the National Academy of Sciences of the United States of America
100(22):12865-12870.
73. Kon Y (2005) Morphogenetic investigation of metaphase-specific cell death in
meiotic spermatocytes in mice. Anat Sci Int 80(3):141-152.
74. Kon Y, Horikoshi H, & Endoh D (1999) Metaphase-specific cell death in
meiotic spermatocytes in mice. Cell and Tissue Research 296(2):359-369.
75. Gavrieli Y, Sherman Y, & Ben-Sasson SA (1992) Identification of
programmed cell death in situ via specific labeling of nuclear DNA
fragmentation. Journal of Cell Biology 119(3):493-501.
76. Gadde S & Heald R (2004) Mechanisms and molecules of the mitotic spindle.
Current Biology 14(18):R797-805.
77. Parra MT, Viera A, Gomez R, Page J, Benavente R, Santos JL, Rufas JS, &
Suja JA (2004) Involvement of the cohesin Rad21 and SCP3 in monopolar
attachment of sister kinetochores during mouse meiosis I. J Cell Sci 117(Pt
7):1221-1234.

! 111!
78. Wittmann T, Hyman A, & Desai A (2001) The spindle: a dynamic assembly
of microtubules and motors. Nature Cell Biology 3(1):E28-34.
79. Handel MA, Cobb J, & Eaker S (1999) What are the spermatocyte's
requirements for successful meiotic division? J Exp Zool 285(3):243-250.
80. Hupalowska A, Kalaszczynska I, Hoffmann S, Tsurumi C, Kubiak JZ,
Polanski Z, & Ciemerych MA (2008) Metaphase I arrest in LT/Sv mouse
oocytes involves the spindle assembly checkpoint. Biology of Reproduction
79(6):1102-1110.
81. Inoue N, Ikawa M, Isotani A, & Okabe M (2005) The immunoglobulin
superfamily protein Izumo is required for sperm to fuse with eggs. Nature
434(7030):234-238.
82. Inoue N, Ikawa M, & Okabe M (2008) Putative sperm fusion protein IZUMO
and the role of N-glycosylation. Biochem Biophys Res Commun 377(3):910-
914.
83. Cobb J, Reddy RK, Park C, & Handel MA (1997) Analysis of expression and
function of topoisomerase I and II during meiosis in male mice. Molecular
Reproduction and Development 46(4):489-498.
84. Dobson MJ, Pearlman RE, Karaiskakis A, Spyropoulos B, & Moens PB
(1994) Synaptonemal complex proteins: occurrence, epitope mapping and
chromosome disjunction. Journal of Cell Science 107 (Pt 10):2749-2760.
85. Dresser ME & Moses MJ (1980) Synaptonemal complex karyotyping in
spermatocytes of the Chinese hamster (Cricetulus griseus). IV. Light and
electron microscopy of synapsis and nucleolar development by silver staining.
Chromosoma 76(1):1-22.

! 112!
86. Grey C, Baudat F, & de Massy B (2009) Genome-wide control of the
distribution of meiotic recombination. PLoS Biol 7(2):e35.
87. Wolf KW & Joshi HC (1996) Microtubule organization and distribution of
gamma-tubulin in male meiosis of lepidoptera. Molecular Reproduction and
Development 45(4):547-559.
88. Evans EP, Breckon G, & Ford CE (1964) An Air-Drying Method for Meiotic
Preparations from Mammalian Testes. Cytogenetics 15:289-294.
89. Fernandez R, Barragan MJ, Bullejos M, Marchal JA, Diaz De La Guardia R,
& Sanchez A (2002) New C-band protocol by heat denaturation in the
presence of formamide. Hereditas 137(2):145-148.
90. Klasterska I, Natarajan AT, & Ramel C (1976) New observations on
mammalian male meiosis. I. Laboratory mouse (Mus musculus) and Rhesus
monkey (Macaca mulatta). Hereditas 83(2):203-214.
91. Stock AD, Burnham DB, & Hsu TC (1972) Giemsa banding of meiotic
chromosomes with description of a procedure for cytological preparations
from solid tissues. Cytogenetics 11(6):534-539.
92. Cunto FD (2002) Essential role of citron kinase in cytokinesis of
spermatogenic precursors. Journal of Cell Science 115(24):4819-4826.
93. Faisal K, Faridha A, & Akbarsha MA (2008) Induction of meiotic micronuclei
in spermatocytes in vivo by aflatoxin B1: light and transmission electron
microscopic study in Swiss mouse. Reprod Toxicol 26(3-4):303-309.
94. Singh SK & Chakravarty S (2001) Effect of nitrofurazone on the reproductive
organs in adult male mice. Asian J Androl 3(1):39-44.

! 113!
95. Obholz KL, Akopyan A, Waymire KG, & MacGregor GR (2006) FNDC3A is
required for adhesion between spermatids and Sertoli cells. Dev Biol
298(2):498-513.
96. Leal MC, Pinheiro SV, Ferreira AJ, Santos RA, Bordoni LS, Alenina N, Bader
M, & Franca LR (2009) The role of angiotensin-(1-7) receptor Mas in
spermatogenesis in mice and rats. J Anat 214(5):736-743.
97. Scott HM, Hutchison GR, Mahood IK, Hallmark N, Welsh M, De Gendt K,
Verhoeven G, O'Shaughnessy P, & Sharpe RM (2007) Role of androgens in
fetal testis development and dysgenesis. Endocrinology 148(5):2027-2036.
98. Wakayama T & Iseki S (2009) Role of the spermatogenic-Sertoli cell
interaction through cell adhesion molecule-1 (CADM1) in spermatogenesis.
Anat Sci Int 84(3):112-121.
99. Ashley T, Westphal C, Plug-de Maggio A, & de Rooij DG (2004) The
mammalian mid-pachytene checkpoint: meiotic arrest in spermatocytes with a
mutation in Atm alone or in combination with a Trp53 (p53) or Cdkn1a
(p21/cip1) mutation. Cytogenet Genome Res 107(3-4):256-262.
100. Meier B & Gartner A (2006) Meiosis: checking chromosomes pair up
properly. Current Biology 16(7):R249-251.
101. Roeder GS & Bailis JM (2000) The pachytene checkpoint. Trends in Genetics
16(9):395-403.
102. Lengauer C, Kinzler KW, & Vogelstein B (1998) Genetic instabilities in
human cancers. Nature 396(6712):643-649.
103. Buckton KE, Whalley LJ, Lee M, & Christie JE (1983) Chromosome changes
in Alzheimer's presenile dementia. Journal of Medical Genetics 20(1):46-51.

! 114!
104. Moorhead PS & Heyman A (1983) Chromosome studies of patients with
Alzheimer disease. Am J Med Genet 14(3):545-556.
105. Modi D, Berde P, & Bhartiya D (2003) Down syndrome: a study of
chromosomal mosaicism. Reprod Biomed Online 6(4):499-503.
106. Palmer CG & Reichmann A (1976) Chromosomal and clinical findings in 110
females with Turner syndrome. Human Genetics 35(1):35-49.
107. Albertson R, Riggs B, & Sullivan W (2005) Membrane traffic: a driving force
in cytokinesis. Trends Cell Biol 15(2):92-101.
108. Rothman JE (1994) Mechanisms of intracellular protein transport. Nature
372(6501):55-63.
109. Hansen NJ, Antonin W, & Edwardson JM (1999) Identification of SNAREs
involved in regulated exocytosis in the pancreatic acinar cell. Journal of
Biological Chemistry 274(32):22871-22876.
110. Chen D, Lemons PP, Schraw T, & Whiteheart SW (2000) Molecular
mechanisms of platelet exocytosis: role of SNAP-23 and syntaxin 2 and 4 in
lysosome release. Blood 96(5):1782-1788.
111. Abonyo BO, Gou D, Wang P, Narasaraju T, Wang Z, & Liu L (2004)
Syntaxin 2 and SNAP-23 are required for regulated surfactant secretion.
Biochemistry 43(12):3499-3506.
112. Gaisano HY, Ghai M, Malkus PN, Sheu L, Bouquillon A, Bennett MK, &
Trimble WS (1996) Distinct cellular locations of the syntaxin family of
proteins in rat pancreatic acinar cells. Molecular Biology of the Cell
7(12):2019-2027.

! 115!
113. Band AM & Kuismanen E (2005) Localization of plasma membrane t-
SNAREs syntaxin 2 and 3 in intracellular compartments. BMC Cell Biology
6(1):26.
114. Low SH, Miura M, Roche PA, Valdez AC, Mostov KE, & Weimbs T (2000)
Intracellular redirection of plasma membrane trafficking after loss of epithelial
cell polarity. Mol Biol Cell 11(9):3045-3060.
115. Kotaja N, Kimmins S, Brancorsini S, Hentsch D, Vonesch JL, Davidson I,
Parvinen M, & Sassone-Corsi P (2004) Preparation, isolation and
characterization of stage-specific spermatogenic cells for cellular and
molecular analysis. Nature Methods 1(3):249-254.
116. Xu H, Brill JA, Hsien J, McBride R, Boulianne GL, & Trimble WS (2002)
Syntaxin 5 Is Required for Cytokinesis and Spermatid Differentiation in
Drosophila. Developmental Biology 251(2):294-306.
117. Farkas RM, Giansanti MG, Gatti M, & Fuller MT (2003) The Drosophila
Cog5 homologue is required for cytokinesis, cell elongation, and assembly of
specialized Golgi architecture during spermatogenesis. Mol Biol Cell
14(1):190-200.
118. Roqueta-Rivera M, Abbott TL, Sivaguru M, Hess RA, & Nakamura MT
(2011) Deficiency in the omega-3 fatty acid pathway results in failure of
acrosome biogenesis in mice. Biol Reprod 85(4):721-732.
119. Cheng X, Zhang Y, Kotani N, Watanabe T, Lee S, Wang X, Kawashima I, Tai
T, Taniguchi N, & Honke K (2005) Production of a recombinant single-chain
variable-fragment (scFv) antibody against sulfoglycolipid. J Biochem
137(3):415-421.

! 116!
120. Ng MM, Chang F, & Burgess DR (2005) Movement of membrane domains
and requirement of membrane signaling molecules for cytokinesis. Dev Cell
9(6):781-790.
121. Akama TO, Nakagawa H, Sugihara K, Narisawa S, Ohyama C, Nishimura S,
O'Brien DA, Moremen KW, Millan JL, & Fukuda MN (2002) Germ cell
survival through carbohydrate-mediated interaction with Sertoli cells. Science
295(5552):124-127.
122. Weber JE & Russell LD (1987) A study of intercellular bridges during
spermatogenesis in the rat. American Journal of Anatomy 180(1):1-24.
123. Nixon B, Bielanowicz A, McLaughlin EA, Tanphaichitr N, Ensslin MA, &
Aitken RJ (2009) Composition and significance of detergent resistant
membranes in mouse spermatozoa. Journal of Cellular Physiology
218(1):122-134.
124. Sandhoff R, Geyer R, Jennemann R, Paret C, Kiss E, Yamashita T, Gorgas K,
Sijmonsma TP, Iwamori M, Finaz C, Proia RL, Wiegandt H, & Grone HJ
(2005) Novel class of glycosphingolipids involved in male fertility. J Biol
Chem 280(29):27310-27318.
125. Takamiya K, Yamamoto A, Furukawa K, Zhao J, Fukumoto S, Yamashiro S,
Okada M, Haraguchi M, Shin M, Kishikawa M, Shiku H, Aizawa S, &
Furukawa K (1998) Complex gangliosides are essential in spermatogenesis of
mice: possible roles in the transport of testosterone. Proceedings of the
National Academy of Sciences of the United States of America 95(21):12147-
12152.

! 117!
126. Carlton JG & Martin-Serrano J (2007) Parallels between cytokinesis and
retroviral budding: a role for the ESCRT machinery. Science 316(5833):1908-
1912.
127. Nixon B, Bielanowicz A, McLaughlin EA, Tanphaichitr N, Ensslin MA, &
Aitken RJ (2009) Composition and significance of detergent resistant
membranes in mouse spermatozoa. J Cell Physiol 218(1):122-134.
128. Xu H, Kongmanas K, Kadunganattil S, Smith CE, Rupar T, Goto-Inoue N,
Hermo L, Faull KF, & Tanphaichitr N (2011) Arylsulfatase A deficiency
causes seminolipid accumulation and a lysosomal storage disorder in Sertoli
cells. J Lipid Res 52(12):2187-2197.
129. Best D, Sahlender DA, Walther N, Peden AA, & Adams IR (2008) Sdmg1 is a
conserved transmembrane protein associated with germ cell sex determination
and germline-soma interactions in mice. Development 135(8):1415-1425.
130. Rankin T, Familari M, Lee E, Ginsberg A, Dwyer N, Blanchette-Mackie J,
Drago J, Westphal H, & Dean J (1996) Mice homozygous for an insertional
mutation in the Zp3 gene lack a zona pellucida and are infertile. Development
122(9):2903-2910.
131. Wassarman PM (1990) Profile of a mammalian sperm receptor. Development
108(1):1-17.
132. Dominguez F, Yanez-Mo M, Sanchez-Madrid F, & Simon C (2005)
Embryonic implantation and leukocyte transendothelial migration: different
processes with similar players? FASEB Journal 19(9):1056-1060.
133. Genbacev OD, Prakobphol A, Foulk RA, Krtolica AR, Ilic D, Singer MS,
Yang ZQ, Kiessling LL, Rosen SD, & Fisher SJ (2003) Trophoblast L-

! 118!
selectin-mediated adhesion at the maternal-fetal interface. Science
299(5605):405-408.
134. Dell A, Chalabi S, Easton RL, Haslam SM, Sutton-Smith M, Patankar MS,
Lattanzio F, Panico M, Morris HR, & Clark GF (2003) Murine and human
zona pellucida 3 derived from mouse eggs express identical O-glycans. Proc
Natl Acad Sci U S A 100(26):15631-15636.
135. Yonezawa N, Kudo K, Terauchi H, Kanai S, Yoda N, Tanokura M, Ito K,
Miura K, Katsumata T, & Nakano M (2005) Recombinant porcine zona
pellucida glycoproteins expressed in Sf9 cells bind to bovine sperm but not to
porcine sperm. J Biol Chem 280(21):20189-20196.





























